Область техники, к которой относится изобретение
Изобретение относится к новым последовательностям ДНК, которые кодируют полипептиды, обладающие фосфолипазной активностью, преимущественно без липазной активности. Изобретение далее относится к новым полипептидам, обладающим фосфолипазной активностью, преимущественно без липазной активности. Эти полипептиды представляют собой кислые фосфолипазы с низкой молекулярной массой, высокой термостабильностью и устойчивостью к высокой температуре. Кроме того, эти полипептиды являются активными в широком интервале рН. Более того, изобретение также относится к применению этих фосфолипаз для снижения содержащих фосфор соединений, например, в производстве пищевых масел, а также к применению этих фосфолипаз в качестве улучшителя хлебобулочных изделий, кормовой добавки для животных, добавки в производстве текстильных сырьевых материалов и т.д.
Уровень техники
Сырые растительные масла содержат сопутствующие материалы (такие как свободные жирные кислоты, фосфолипиды, тяжелые металлы, красящие вещества…), которые влияют на качество и срок хранения масла и осложняют его дальнейшую переработку из-за гидролитической или окислительной модификации липидов во время хранения. Таким образом, необходимо очищать сырые растительные масла после их экстрагирования для того, чтобы удалить примеси. В производстве пищевых масел высокого качества процесс очистки включает стадии дегуммирования, отбеливания и деодорирования.
Первой стадией в процессе очистки масел является дегуммирование. В ходе процесса дегуммирования удаляются смолистые вещества, в первую очередь фосфолипиды, которые негативно влияют на вкус масла и которые мешают дальнейшим стадиям процесса очистка масла.
Так как содержание фосфора является показателем степени дегуммирования, качество полученного дегуммированного масла может быть оценено путем определения содержания остаточного фосфора. Для дальнейших стадий процесса очистки требуется дегуммированное масло с содержанием остаточного фосфора менее чем 10 м.д.
Фосфолипиды представляют собой сложные содержащие фосфор липиды. Фосфолипиды, такие как фосфатидилхолин или лецитин, состоят из глицериновой структуры, которая этерифицирована жирными кислотами в положениях sn-1 и sn-2, и связанной со сложным эфиром фосфатной группой в положении sn-3. Фосфатная группа сама по себе может быть этерифицирована, например, первичной спиртовой группой. В положениях sn-1 и sn-2 природные фосфолипиды содержат цепи различных жирных кислот, которые в случае растений, в основном, представляют собой полиненасыщенные ацильные цепи. Существуют два вида фосфолипидов: гидратируемые и негидратируемые. Для того чтобы удалить фосфолипиды, используются различные способы.
Наиболее простым способом является водное дегуммирование. В ходе этого процесса гидратируемые фосфолипиды могут быть вымыты при помощи воды и, таким образом, удалены из масла. В зависимости от типа и качества сырого масла дегуммированное масло после этого процесса по-прежнему содержит от 80 до 200 м.д. фосфора.
В процессе кислотного дегуммирования масло обрабатывается кислотой. Кислота превращает негидратируемые фосфолипиды в гидратируемые фосфолипиды. Гидратируемые фосфолипиды становятся нерастворимыми в масле. Образуется нерастворимая в масле масса, которая удаляется из масла посредством центрифугирования или фильтрования. После этого процесса содержание остаточного фосфора в масле составляет приблизительно от 25 до 100 м.д.
Ферментативное дегуммирование предлагает эффективный, рентабельный и экологически безопасный процесс для аккуратного удаления фосфолипидов из пищевого масла.
Фосфолипазы представляют собой ферменты, которые расщепляют фосфолипиды и подразделяются в соответствии с их сайтами ферментативного расщепления в фосфолипиде на ацилгидролазы (фосфолипаза А1, А2 и В) и фосфодиэстеразы (фосфолипаза С и D). Фосфолипаза А1 (КФ 3.1.1.32) гидролизует жирную кислоту в положении фосфолипидных молекул sn-1, и фосфолипаза А2 (КФ 3.1.1.4) специфично отщепляет сложноэфирную связь sn-2 от фосфолипида. В качестве продуктов реакции образуются лизофосфолипид и свободная жирная кислота. Фосфолипаза В (КФ 3.1.1.5) неспецифично расщепляет жирную кислоту как в положении sn-1, так и в положении sn-2. Фосфолипаза С (КФ 3.1.4.10) гидролизует фосфатную сложноэфирную связь между глицерином и фосфатной группой с образованием моноэфира фосфата и диацилглицерина. Фосфолипаза D (КФ 3.1.4.4) катализирует гидролиз концевой фосфодиэфирной связи с образованием расщепленных продуктов фосфатидной кислоты и холина.
Уже известна экстракция фосфолипазы А2 из бычьей и свиной поджелудочных желез и из токсина медоносных пчел или нескольких видов змей. Фосфолипаза также может быть экстрагирована из микроорганизмов, таких как бактерии и грибы, и может быть получена с достаточным выходом посредством рекомбинантых методик.
В патенте ЕР 0513709 впервые представлен процесс эффективного ферментативного дегуммирования. В новом процессе впервые используется фермент фосфолипаза А2, которая расщепляет жирную кислоту в положении sn-2 фосфолипидов. Процесс был протестирован с соевым и рапсовым маслом, имеющими содержание фосфора от 72 до 110 м.д. Реакционная загрузка содержала вплоть до 5 масс.% (масс./масс.) воды по отношению к маслу и была выдержана при значении рН от 5.0 до 5.5 при 40°С или, как вариант, при 60°С в течение вплоть до 5 часов. Образовавшийся лизофосфолипид может быть удален из масла посредством центрифугирования. После процесса содержание остаточного фосфора в дегуммированном масле составляет менее чем 5 м.д.
В ходе процесса дегуммирования масло смешивается с определенным количеством воды. Затем содержащую фермент водную фазу и масляную фазу смешивают для того, чтобы дать возможность ферменту подействовать. Количество воды должно быть при этом настолько малым, насколько возможно. Использование больших количеств воды приводит к увеличенному потреблению энергии и увеличенным расходам по утилизации. Таким образом, процесс ферментативного дегуммирования с низким содержанием воды (2%) уже является предпочтительным в этом отношении.
В US 2007/134777 утверждается, что ферментативное дегуммирование растительного масла при помощи фосфолипазы А1 проводится при значении рН между 4.0 и 5.0. Оптимальным значением рН для проведения этого процесса является значение между 4.5 и 5.0. В этом интервале рН высвободившиеся ионы кальция и/или магния могут соединяться с другими химическими веществами (анионами) реакционного буфера с образованием трудно растворимых солей, которые откладываются на поверхности реакторов и, таким образом, загрязняют аппарат. Удаление таких загрязнителей и очистка аппарата является трудоемкой. Для того чтобы снизить загрязнение внутренней части аппарата, реакция предпочтительно проводится при рН приблизительно 4. Однако фермент становится менее активным или функционально неактивным, если значение рН реакции в дальнейшем снижается.
В ЕР 0904357 описывается, что в Aspergillus niger была обнаружена фосфолипаза А для дегуммирования пищевого масла.
В WO 2008/040466 описывается, что фосфолипаза, имеющая молекулярную массу 65 кДа, может быть использована в Aspergillus fumigatus для дегуммирования пищевого масла с содержанием воды 5% при температурах вплоть до 65°С.
В ЕР 1788080 описывается использование фосфолипазы С из Bacillus cereus для дегуммирования масла в течение 6 часов при 60°С с содержанием воды 15% по отношению к маслу. После процесса содержание остаточного фосфора составляет менее чем 5 м.д.
В WO 2008/094847 описано, что время реакции процесса дегуммирования масла может быть снижено до 30 мин при реакции фосфолипазы А1 или А2 с фосфолипазой С соответственно.
Фосфолипазы также многообразно используются в пищевой промышленности и промышленности кормов для животных, например, для приготовления теста, для приготовления хлебобулочных изделий, для увеличения выхода производства сыра и т.д. Таким образом, существует необходимость в фосфолипазах, которые могут быть многосторонне использованы в технологии.
В биотехнологии фосфолипазы используются в качестве биокатализаторов для экстракции фосфолипидов. Фосфолипиды представляют собой полярные липиды и выступают в качестве эмульгаторов благодаря их липофильным и гидрофильным структурным характеристикам. Примерами являются применение фосфолипаз в производстве модифицированных лецитинов, в качестве пищевых эмульгаторов в производстве соусов, майонеза и заправок для салатов, в производстве быстрорастворимых порошков, таких как порошки молока, какао и кофе, в качестве улучшителей текучести в производстве шоколада и в качестве пищевых добавок. В фармацевтической и косметической промышленности фосфолипиды и лизофосфолипиды используются для производства кремов, лосьонов, гелей и липосомных составов.
Лецитин также требуется для производства лаков, красок, магнитных лент, специализированных бумаг, кожи и текстиля. Фосфолипазы также используются в текстильной промышленности для «биоотварки» с целью очистки растительного волокна перед проведением дальнейших стадий процесса, таких как окрашивание. Здесь также может быть использована смесь фосфолипаз вместе с другими ферментами. Другие ферменты могут быть выбраны из группы целлюлаз, гемицеллюлаз, пектиназ, протеаз и оксидоредуктаз.
Таким образом, существует постоянная необходимость в фосфолипазах, имеющих возможно большую и оптимальную область применения в данной области техники.
Раскрытие изобретения
Вследствие этого основывается на задаче обеспечения белками или полипептидами с улучшенными фосфолипазными свойствами. Особенно, новые фосфолипазы не должны проявлять липазную активность, относящуюся к технологическим процессам. В особенности, белки, обладающие фосфолипазной активностью должны быть активными в большом интервале рН и должны быть крайне устойчивыми к температуре.
Более того, производство белков, обладающих фосфолипазной активностью должно быть простым, рентабельным и промышленным. Кроме того, должны обеспечиваться экспрессионные конструкты по изобретению, которые являются подходящими для производства белков, обладающих фосфолипазной активностью.
Указанные выше объекты решаются посредством последовательности ДНК, которая кодирует полипептид, обладающий фосфолипазной активностью, преимущественно без липазной активности, характеризующейся тем, что последовательность ДНК выбирается из а) последовательностей ДНК, которые содержат нуклеотидную последовательность, соответствующую SEQ ID NO:1, b) последовательностей ДНК, которые содержат кодирующую последовательность, соответствующую SEQ ID NO:1, с) последовательностей ДНК, которые кодируют белковую последовательность, соответствующую SEQ ID NO:2, d) последовательностей ДНК, которые кодируются плазмидой pPL3949-Topo2.5 с рестрикционной картой в соответствии с фигурой 7, которая помещена под номером доступа DSM 22741, е) последовательностей ДНК, которые гибридизуются при жестких условиях с одной из последовательностей ДНК, соответствующей а), b), с) или d), f) последовательностей ДНК, которые связаны с одной из последовательностей, соответствующей а), b), с), d) или е) из-за вырожденности генетического кода и g) комплементарных нитей к последовательностям, соответствующим от а) до f).
Изобретение далее относится к полипептиду, обладающему фосфолипазной активностью, преимущественно без липазной активности, выбранному из а) полипептида, который кодируется путем кодирующей части последовательности ДНК, как определено выше, b) полипептида, имеющего последовательность, соответствующую SEQ ID NO:2, или последовательность, образованную из этой, которая может быть получена путем замещения, добавления, удаления одной или более аминокислот, с) полипептида, имеющего последовательность, которая имеет по крайней мере 83% идентичности с аминокислотами от 1 до 299 из SEQ ID NO:2, d) полипептида, который кодируется нуклеиновокислотной последовательностью, которая гибридизуется при жестких условиях с (i) нуклеотидами от 55 до 1106 из SEQ ID NO:1, (ii) последовательностью кДНК, содержащейся в нуклеотидах от 55 до 1106 из SEQ ID NO:1, (iii) частичной последовательностью (i) или (ii), составленной из по крайней мере 100 нуклеотидов или (iv) комплементарной нити (i), (ii) или (iii), е) варианта полипептида, имеющего SEQ ID NO:2, содержащую замещение, удаление и/или введение одной или более аминокислот, f) аллельных вариантов к аминокислотным последовательностям от а) до е).
Кроме того, изобретение относится к экспрессионным конструктам или хозяевам, которые способны экспрессировать полипептиды, обладающие фосфолипазной активностью по изобретению. Более того, изобретение также относится к соответствующим экспрессионным плазмидам и векторам. Кроме того, изобретение относится к процессам дегуммирования растительного масла с использованием полипептидов по изобретению, а также к применению полипептидов по изобретению в области пищевой технологии, в особенности для производства теста, хлебобулочных изделий или молочных продуктов, или в питании животных и в производстве текстильных сырьевых материалов, так называемой отварке или биоотварке.
В соответствии с дальнейшим воплощением изобретение относится к полипептиду, обладающему фосфолипазной активностью, преимущественно без липазной активности, характеризующемуся тем, что имеет молекулярную массу в интервале от 28 до 30 кДа и предпочтительно приблизительно 28.6 кДа, широкий оптимум рН и устойчивость к высокой температуре и тем, что он может быть выделен из организма рода Aspergillus.
Устойчивость к высокой температуре в этом контексте означает, что после 6 часов при температуре 60°С в условиях процесса дегуммирования масла с низким 2% содержанием воды фермент сохраняет свою активность в промышленно используемом объеме. Фермент сохраняет свою активность в промышленно используемом объеме, если он производит масла, у которых содержание остаточного фосфора является технологически незначительным, если можно так сказать. Предпочтительно содержание остаточного фосфора в ферментативно дегуммированном масле составляет менее 10 м.д., более предпочтительно менее 5 м.д.
Неожиданно было обнаружено, что последовательность ДНК, которая кодирует полипептид, обладающий фосфолипазной активностью, преимущественно без липазной активности, который имеет низкую молекулярную массу, а также устойчивость к высокой температуре, может быть выделена из штамма рода Aspergillus fumigatus. Эта фосфолипаза представляет собой кислую фосфолипазу, полученную из нитчатого гриба, с рассчитанной молекулярной массой приблизительно 28.6 кДа, которая способна гидролизовать по крайней мере одну из двух жирных кислот из лецитина.
В противоположность полипептидам, обладающим фосфолипазной активностью, известным из существующего уровня техники, фосфолипазы по изобретению обладают устойчивостью к высокой температуре (при 60°С) и могут, таким образом, быть с пользой использованы в процессах ферментативного дегуммирования с низким 2% содержанием воды (по отношению к маслу) и при значении рН 4.0. Это представляет особенный экономический интерес, поскольку температура масла не должна вначале понижаться при процессах дегуммирования для осуществления ферментативного дегуммирования без возможной инактивации фермента, и впоследствии температура масла опять должна быть увеличена для того, чтобы снизить вязкость масла для стадии центрифугирования, используемой для отделения масляных фаз от водных фаз. Устойчивость полипептидов к температуре, обладающих фосфолипазной активностью по изобретению, является также предпочтительным для других применений в области пищевой технологии, и питании животных, и в текстильном производстве, соответственно.
Фосфолипазы, известные из существующего уровня техники, исключены из объема изобретения.
Таким образом, особенно выгодным является то, что фосфолипазы, выделенные из А. fumigatus по изобретению, являются активными в большом интервале рН от 3 до 5 или показывают широкий оптимум активности в этом диапазоне. Таким образом, фосфолипазы по изобретению не только имеют выгоду в том, что они преимущественно не имеют липазной активности. Они дополнительно обнаруживают ферментную активность в большом интервале рН и, тем самым, могут быть использованы в большом интервале рН.
Ферменты, обладающие фосфолипазной активностью (фосфолипаза А, В, С или D), из Aspergillus fumigatus до настоящего времени уже упоминались в нескольких публикациях (Birch et al., Comparison of extracellular phospholipase activities in clinical and environmental Aspergillus fumigatus isolates, 2004, Med Mycol 42(1): 81-86; Rementeria et al., Genes and molecules involved in Aspergillus fumigatus virulence, 2005, Rev Iberoam Micol 22(1): 1-23), но конкретно не были охарактеризованы. Из геномной последовательности Aspergillus fumigatus (Nierman et al., Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus, 2005, Nature, 438(7071): 1151-6) было выделено несколько гипотетических генов фосфолипазы (А, В, С, D) и лизофосфолипазы под термином «концептуальная трансляция». Таким образом, например, были представлены одна гипотетическая фосфолипаза А, имеющая 241 аминокислоту, и три лизофосфолипазы Plb1, Plb2 и Plb3, имеющие высокую молекулярную массу. Кроме того, база данных показывает небольшую гипотетическую внеклеточную липазу, имеющую 299 аминокислот и рассчитанную молекулярную массу приблизительно 28.5 кДа (GenBank EAL86100), а также несколько липаз, имеющих от 409 до 587 аминокислот.
Shen et al. (Characterisation and expression of phospholipase В from the opportunistic fungus Aspergillus fumigatus, 2004, FEMS Microbol Lett 239(1): 87-93) добились успеха в клонировании и описании 3 фосфолипазных генов из Aspergillus fumigatus. Секретированные белки AfPL1, имеющие 633 аминокислоты, и AfPL3, имеющие 630 аминокислот, имеют молекулярную массу приблизительно 68 кДа. Белок AfPL2 представляет собой цитоплазматический белок, имеющий 588 аминокислот, и имеет молекулярную массу приблизительно 63 кДа.
Более того, фосфолипаза, имеющая 633 аминокислоты (WO 2008/040466), и лизофосфолипаза, имеющая 611 аминокислот (WO 2008/040465), были выделены из Aspergillus fumigatus RH3949.
Кроме того, кислая фосфолипаза (pl 4.1), имеющая низкую молекулярную массу в интервале от 28 до 30 кДа, была обнаружена в геноме Aspergillus fumigatus RH3949.
В процессе секвенирования белка этой «маленькой» фосфолипазы первые 16 аминокислот показали 93%-ую идентичность в N-конце с N-терминальной последовательностью гипотетической внеклеточной липазы, имеющей 299 аминокислот (GenBank EAL86100), из Aspergillus fumigatus Af293. Идентичность зрелой (АА 30-299) последовательности «маленькой фосфолипазы» из RH3949 с последовательностью гипотетической внеклеточной липазы составляет, однако, только приблизительно 82%. Это было особенно неожиданным, поскольку на сопоставимом сайте в геноме дополнительного штамма Aspergillus fumigatus (АН 63) была обнаружена последовательность, имеющая значительно более высокую идентичность с гипотетической липазой из Af293 (GenBank EDP1054) (Fedorova et al., Genomic Islands in the Pathogenic Filamentous Fungus Aspergillus fumigatus, 2008, PloS Genet.4. e1000046).
В противоположность этому не существует аналогии между двумя N-терминальными фосфолипазными последовательностями из Aspergillus fumigatus Af293 (фосфолипаза А, имеющая 241 аминокислоту, GenBank EAL85761) и Aspergillus fumigatus RH3949 (фосфолипаза, имеющая pl 4.1).
Следовательно, фосфолипаза по изобретению отличается от известных фосфолипаз из Aspergillus fumigatus, а также от последовательностей, аннотированных как липазы, близкородственные штаммам Aspergillus fumigatus, таким как Af293.
В соответствии с изобретением некоторые олигонуклеотидные праймеры были получены и синтезированы из данных по последовательности ДНК (GenBank AAHF01000011) для гипотетической внеклеточной липазы (GenBank EAL86100).
Неожиданно было обнаружено, что последовательность ДНК по изобретению может быть выделена из штамма RH3949 Aspergillus посредством праймеров, полученных из геномной последовательности штамма Af293 (см. пример 4). Это было неожиданным, поскольку штамм RH3949 (выделенный из окружающей среды), использованный для амплификации, отличается от Af293 (клинически выделенный) фенотипически и, следовательно, наиболее вероятно также генотипически (последовательностью). Таким образом, было невозможно в равной степени амплифицировать фосфолипазный ген из RH3949 посредством Другой праймерной пары, имеющей связанные сайты, расположенные рядом с N2-3948 и ApaI-3949 на геномной ДНК штамма Af293.
Амплификация гена была проведена с помощью полимеразной цепной реакции (ПЦР) из геномной ДНК Aspergillus fumigatus RH3949.
Устойчивость к температуре и широкий оптимум рН фосфолипазы по изобретению были неожиданными и неожидаемыми на основании фосфолипаз, описанных в уровне техники. Ни одна из встречающихся в природе фосфолипаз, полученных из нитчатых грибов, как описано в уровне техники, не обладает этими свойствами, которые были описаны или просто предложены.
Фосфолипазная последовательность в соответствии с SEQ ID NO:2 по изобретению была сравнена с фосфолипазными последовательностями из уровня техники. Была обнаружена частичная аналогия аминокислотной последовательности с известными аминокислотными последовательностями из других штаммов Aspergillus, то есть аналогия на 60% с липазой из Aspergillus tubingensis (WO 98/45453) или с лизофосфолипазой из Aspergillus foetidus (EP 0808903), 59% с фосфолипазой Aspergillus niger (WO 03/097825, WO 98/31790).
Последовательность SEQ ID NO:2 проявляет наибольшую идентичность в 82% с гипотетической последовательностью для внеклеточной липазы (GenBank EAL86100) из Aspergillus fumigatus Af293 (Nierman et al., Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus, 2005, Nature, 438(7071): 1151-6).
Неожиданно, при условиях ферментативного дегуммирования пищевого масла фосфолипаза, выделенная из Aspergillus fumigatus RH3949 по изобретению, не проявляет какой-либо липазной активности, соответствующей этому процессу. Кроме того, этот фермент имеет заметно широкий оптимум рН и устойчивость к высокой температуре в противоположность фосфолипазам из других штаммов Aspergillus, известных к настоящему времени. Таким образом, фермент по изобретению может быть использован предпочтительно в процессе ферментативного дегуммирования пищевых масел, поскольку он не гидролизует любые или совсем незначительные части триглицеридных связей в масле.
Более того, изобретение также относится к полипептидам, обладающим фосфолипазной активностью, преимущественно без липазной активности, с последовательностью, которая имеет идентичность по крайней мере 83% с последовательностью в соответствии с SEQ ID NO:2. Предпочтительно изобретение относится к полипептиду, обладающему фосфолипазной активностью, с последовательностью, которая имеет идентичность по крайней мере 83% с аминокислотами от 1 до 299 SEQ ID NO:2. Предпочтительно степень идентичности с аминокислотами от 1 до 299 SEQ ID NO:2 составляет по крайней мере 90%, более предпочтительно по крайней мере 95%, даже более предпочтительно по крайней мере 97% и особенно предпочтительно по крайней мере 98% при условии, что соответствующие последовательности обладают фосфолипазной активностью, преимущественно без липазной активности.
Полипептиды, обладающие фосфолипазной активностью по изобретению, не обладают какой-либо значительной липазной активностью или точнее являются преимущественно без липазной активности. Полипептиды по изобретению преимущественно не обладают какой-либо липазной активностью, неблагоприятной для промышленных процессов дегуммирования масла, то есть полипептиды по изобретению преимущественно не проявляют какую-либо активность против липолитически расщепляемых соединений в масле, которое необходимо дегуммировать. Это означает, что при условиях ферментативного дегуммирования пищевого масла фосфолипазы по изобретению не обладают какой-либо липазной активностью, имеющей отношение к этому процессу. Говоря технологически, это означает, что полипептиды, обладающие фосфолипазной активностью по изобретению, гидролизуют п-нитрофенилпальмитат как липазный субстрат только до незначительной и/или необнаруживаемой степени. Для полипептидов, обладающих фосфолипазной активностью по изобретению, соотношение фосфолипазной активности к липазной активности составляет предпочтительно >1000:1, более предпочтительно от 5000:1 до 10000:1, даже более предпочтительно 7000:1 и наиболее предпочтительно 7500:1.
Степень идентичности последовательностей предпочтительно определяется таким образом, что определяется число остатков более короткой последовательности, которая вовлечена в сравнение и имеет «соответствующий» эквивалент в другой последовательности. Для этих целей настоящего изобретения идентичность предпочтительно определяется обычной методикой с использованием обычных алгоритмов. В соответствии с изобретением для сравнения используются только кДНК или аминокислоты соответствующих зрелых белков. Подобно этому предпочтительно идентичные эквиваленты последовательности были определены по изобретению как гомологичные последовательности посредством известных компьютерных программ. Примером такой программы является программа Clone Manager Suite, которая включает часть программы Align Plus и поставляется Scientific & Educational Software, Durham, NC, U.S.A. Сравнение двух последовательностей ДНК или аминокислотных последовательностей, как определено выше, вследствие этого проводится с помощью опции локального выравнивания, либо в соответствии со способом FastScan-MaxScore, либо в соответствии со способом Needleman-Wunsch, сохраняя данные по умолчанию. Версия программы "Clone Manager 7 Align Plus 5" с функциями "Compare Two Sequences/Local Fast Scan-Max Score/Compare DNA sequences" или для аминокислот "Compare Two Sequences/Global/Compare sequences as Amino Acids" предпочтительно использовалась для расчета идентичности в соответствии с изобретением. Вследствие этого использовались алгоритмы, ставшие доступными из следующих источников: Hirschberg, D.S. 1975. A linear space algorithm for computing maximal common subsequences. Commun Assoc Comput Mach 18:341-343; Myers, E.W. and W.Miller. 1988. Optimal alignments in linear space. CABIOS 4:1, 11-17; Chao, K-M, W.R.Pearson and W.Miller. 1992. Aligning two sequences within a specified diagonal band. CABIOS 8:5, 481-487.
Изобретение также относится к молекулам с добавлением и/или удалению молекул с делениями указанных выше полипептидов, обладающих фосфолипазной активностью. Таким образом, полипептид, обладающий фосфолипазной активностью, модифицированный в соответствии с изобретением, может быть удлинен путем добавления дополнительных последовательностей к N- и/или С- концу, в результате чего полученные таким образом аминокислотные последовательности по-прежнему должны проявлять фосфолипазную активность преимущественно без липазной активности. В результате этого могут быть получены гибридные молекулы, которые имеют дополнительные преимущественные свойства. Например, суспензия белков или их природных предшественных форм может быть добавлена к секретированным в значительной степени белкам, которая в дальнейшем увеличивает эффективность секретирования. Более того, активные сегменты последовательности других ферментов могут быть добавлены для получения ферментов с множественной специфичностью. Кроме того, полярные и неполярные последовательности могут быть добавлены для специфического влияния на свойства растворимости или подвижность мембраны полученных таким образом белков.
Сегменты последовательности полипептида, обладающего фосфолипазной активностью, могут также быть удалены в соответствии с изобретением, с сохранением фосфолипазной активности, преимущественно без липазной активности. Мутации, удлинения и укорачивания могут быть проведены путем, известным per se, и с помощью способов, хорошо известных в существующем уровне техники. Укороченные полипептиды часто характеризуются повышенной степенью секреции по сравнению с непроцессированными полипептидами. Она также могут проявлять более высокие термостабильности по сравнению с непроцессированным полипептидом, поскольку они содержат только «упакованное ядро».
Получение таких вариантов обычно известно из существующего уровня техники. Например, варианты аминокислотной последовательности полипептидов могут быть получены путем мутации в ДНК. Процессы для мутагенеза и изменений в нуклеотидной последовательности являются хорошо известными в существующем уровне техники (см., например, Tomic et al. NAR, 18:1656 (1990), Giebel and Sprtiz NAR, 18:4947 (1990)).
Детали по подходящим замещениям аминокислоты, которые не влияют негативно на биологическую активность целевого белка, могут быть найдены в модели Dayhoff et al., Atlas of Protein Sequence and Structure, Natl. Biomed. Res. Found., Washington, D.C. (1978). Предпочтительными являются традиционные замещения, такие как замещение аминокислоты другой аминокислотой с похожими свойствами. Эти замещения могут быть разделены на две главные группы в общей сложности с четырьмя подгруппами, и замещение в каждой группе называется традиционным замещением, которое предпочтительно не влияет на активность или фолдинг белка.
Таблица 1
Выражения «белок», «пептид» и «полипептид» преимущественно используются взаимозаменяемо. Полипептид или фермент, обладающий фосфолипазной активностью, или фосфолипаза относится к ферменту, который катализирует высвобождение жирных кислот из фосфолипидов, например, лецитинов. Фосфолипазная активность может быть определена посредством использования любого измерительного способа, известного per se, в котором используется один из этих субстратов.
В отношении полипептидов по изобретению выражения «фосфолипаза» или фосфолипаза А относятся к ферментам, обладающими фосфолипазной А1 активностью, а также фосфолипазной А2 активностью. Фосфолипаза А1 или А2 в результате этого определяется в соответствии со стандартной классификацией ферментов КФ как КФ 3.1.1.32 или 3.1.1.4 соответственно.
Фосфолипаза В или лизофосфолипаза являются полипептидами в соответствии со стандартной КФ классификацией ферментов КФ 3.1.1.5.
Изобретение также относится к последовательностям ДНК, которые кодируют полипептид, обладающий фосфолипазной активностью, включая мутации, модификации или вариации последовательности в соответствии с SEQ ID NO:1. Кроме того, изобретение также относится к последовательностям, которые гибридизуются с вышеупомянутыми последовательностями при мягких или жестких условиях. Следующие условия считаются жесткими: гибридизация при 65°С 18 ч в растворе сульфата декстрана (GenescreenPlus, DuPont), последующее промывание фильтра в течение 30 мин соответственно сначала 6×SSC, дважды 2×SSC, дважды 2×SSC, 0.1% ДСН и в конце 0.2×SSC при 65°С (способы перемещения и удаления мембраны, Amersham).
Кроме того, изобретение также относится к последовательностям ДНК, которые относятся к вышеупомянутым по изобретению из-за вырожденности генетического кода, а также аллельным вариантам указанных. Вырожденность генетического кода может в связи с этим являться результатом природной вырожденности или специально выбранного применения кодона. Встречающиеся в природе аллельные варианты могут быть идентифицированы посредством хорошо известных методик молекулярной биологии, таких как полимеразная цепная реакция (ПЦР), и методик гибридизации.
Изобретение также относится к процессу получения полипептида, обладающего фосфолипазной активностью, с использованием рекомбинантных методик, включающих выращивание рекомбинантных прокариотических и/или эукариотических клеток-хозяев, которые содержат последовательность ДНК по изобретению при условиях, которые промотируют экспрессию фермента, а также последующую экстракцию фермента. Изобретение также относится к применению полинуклеотидных последовательностей по изобретению для получения зондов для обнаружения подобных последовательностей, которые кодируют соответствующие ферменты в других организмах, а также для трансформации клеток-хозяев.
Последовательность ДНК, которая кодирует полипептид по изобретению, может быть использована для трансформации любых клеток-хозяев, таких как клетки грибов, дрожжей, бактерий, растений или млекопитающих. Клетки, трансформированные таким образом, характеризуются секретированием фосфолипазы по изобретению. Полученный таким образом фосфолипазный фермент влияет на эффективный гидролиз жирных кислот из фосфолипидов.
Изобретение также относится к экспрессионным кассетам, которые могут быть использованы для введения последовательности ДНК, кодирующей фосфолипазу по изобретению, или открытой рамки считывания в клетку-хозяина. Они предпочтительно включают транскрипционный инициирующий участок, который связан с открытой рамкой считывания. Такая экспрессионная кассета может содержать разнообразие рестрикционных расщепляющих сайтов для включения открытой рамки считывания и/или других ДНК, например, транскрипционного регуляторного участка и/или селектируемых маркерных генов. Транскрипционная кассета содержит в направлении транскрипции 5'→3' транскрипционный инициирующий участок и трансляционный инициирующий участок, целевую последовательность ДНК и транскрипционный завершающий участок и трансляционный завершающий участок, который является функциональным в микробной клетке. Участок терминации может быть нативным в отношении транскрипционного инициирующего участка, может быть нативным в отношении целевой последовательности ДНК или может быть получен из любого другого источника.
Выражение «открытая рамка считывания» (ОРС) относится к аминокислотной последовательности, которая закодирована между трансляционными инициирующими кодонами и трансляционными завершающими кодонами кодирующей последовательности. Выражения «инициирующий кодон» и «завершающий кодон» относятся к объединению трех соседних нуклеотидов (кодонов) в кодирующей последовательности, которые определяют начало цепи и завершение цепи белкового синтеза (трансляция мРНК).
Применительно к нуклеиновой кислоте «функциональное сцепление» относится к соединению как части той же молекулы нуклеиновой кислоты в подходящем положении и ориентации по отношению к точке начала транскрипции промотора. ДНК в функциональной связи с промотором расположена под контролем инициации транскрипции промотора. Кодирующие последовательность могут быть функционально связаны с регуляторной последовательностью в смысловой ориентации или антисмысловой ориентации. Что касается полипептидов, функциональное сцепление относится к соединению как части того же полипептида, то есть посредством пептидильных связей.
В соответствии с изобретением может быть использован любой промотор. Промотор обычно относится к нукпеотидной последовательности, расположенной против хода (5') к кодирующей последовательности, и контролирует экспрессию кодирующей последовательности путем обеспечения распознавания РНК-полимеразы и других факторов, которые необходимы для правильной транскрипции. Промотор, использованный в соответствии с изобретением, может содержать минимальный промотор, то есть короткую последовательность ДНК из ТАТА-бокса, и другие последовательности, которые определяют транскрипционный инициирующий сайт, к которому регуляторные элементы прикреплены для контролирования экспрессии.
Промотор, использованный в соответствии с изобретением, может также содержать нуклеотидную последовательность, которая содержит минимальный промотор и регуляторные элементы, и может контролировать экспрессию кодирующей последовательности или функциональной РНК. Этот тип промоторной последовательности состоит из проксимальных и дистальных элементов, расположенных против хода, в соответствии с чем элементы, упомянутые последними, часто называются энхансерами. Следовательно, энхансер представляет собой последовательность ДНК, которая может стимулировать активность промотора и может быть элементом, присущим промотору, или внедренным гетерологичным элементом для улучшения степени экспрессии или тканевой специфичности промотора. Он может работать в обеих ориентациях и может даже работать, если он расположен против хода или по ходу транскрипции относительно промотора. Не только энхансеры, но также другие расположенные против хода элементы промотора специфично в отношении последовательности связывают ДНК-связанные белки, опосредуя их эффекты. Промоторы могут быть выделены из нативного гена в своей целостности или могут быть составлены из различных элементов, полученных из различных встречающихся в природе промоторов, или могут даже быть составлены из сегментов синтетической ДНК. Промотор может также содержать последовательности ДНК, которые вовлечены в связывание белковых факторов, которые контролируют эффективность инициации транскрипции как ответа на физиологические или связанные с развитием условия.
Элементы промотора, особенно ТАТА-элементы, которые являются неактивными или обладают сильно сниженной промоторной активностью в отсутствие активации против хода, называются минимальными промоторами или коревыми промоторами. В присутствии подходящего транскрипционного фактора или подходящих транскрипционных факторов функцией минимального промотора является предоставление возможности транскрипции. Таким образом, минимальный промотор или коровый промотор состоит только из всех основных элементов, которые необходимы для инициации транскрипции, то есть ТАТА-бокса и/или инициатора.
Изобретение также относится к векторным конструктам, содержащим последовательности ДНК по изобретению. Эти векторные конструкты содержат любую плазмиду, космиду, фаг или другой вектор в двухцепочечной или одноцепочечной, линейной или кольцевой форме, которые могут быть передаваемыми или мобилизуемыми сами по себе и могут либо трансформировать прокариотического или эукариотического хозяина посредством интеграции в клеточный геном, или присутствовать экстрахромосомно (например, автономно реплицирующиеся плазмиды с репликационным происхождением).
Векторы, плазмиды, космиды, искусственные дрожжевые хромосомы (ИДХ), искусственные бактериальные хромосомы (ИБХ) и сегменты ДНК подлежат использованию для трансформации клеток, обычно включающих ДНК, которая кодирует фосфолипазу в соответствии с изобретением, а также другую ДНК, такую как кДНК, ген или гены, который/ые должен/жны быть введен(ы) в клетки. Эти конструкты ДНК могут содержать дополнительные структуры, такие как промоторы, энхансеры, полилинкеры или также регуляторные гены, если необходимо. Один из сегментов ДНК или генов, который/ые был(и) выбран(ы) для клеточного введения условно кодирует/кодируют белок, который экспрессируется в полученных таким образом трансформированных (рекомбинантных) клетках, что приводит к наблюдаемому или произвольному свойству и/или обеспечивает трансформированные клетки улучшенным фенотипом.
Конструкция векторов, которая может быть использована в соответствии с изобретением, известна специалисту в данной области техники вследствие вышеупомянутого раскрытия и общих знаний специалиста (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual (2nd ed., Cold Spring Harbor Laboratory Press, Plainview, N.Y. (1989))).
Экспрессионная кассета по изобретению может содержать один или несколько рестрикционных сайт(ов) для помещения полинуклеотида, кодирующего фосфолипазу, под контроль регуляторной последовательности. Экспрессионная кассета может также включать сигнал терминации в функциональной связи с полинуклеотидом, а также регуляторные последовательности, которые требуются для правильной трансляции полинуклеотида. Экспрессионная кассета, которая содержит полинуклеотид по изобретению, может быть химерной, то есть по крайней мере один из ее компонентов является гетерологичным по отношению к по крайней мере одному из других компонентов. Экспрессия полинуклеотида в экспрессионной кассете может контролироваться конститутивным промотором, индуцируемым промотором, регулируемым промотором, вирусным промотором или синтетическим промотором.
Векторы уже могут содержать регуляторные элементы, например, промоторы, или последовательности ДНК по изобретению могут быть обработаны таким путем, что они содержат такие элементы. Подходящие промоторные элементы, которые могут быть использованы, известны в существующем уровне техники и представляют собой, например, для Trichoderma reesei промотор cbh1 или промотор cbh2, для Aspergillus oryzae промотор amy, для Aspergillus niger промотор xyl, промотор glaA, промотор aleA, промотор aphA, промотор tpiA, промотор gpdA, промотор sucl и промотор pkiA. Подходящие промоторные элементы, которые могут быть использованы для экспрессии в дрожжах, известны из существующего уровня техники и представляют собой, например, промотор pho5 или промотор gap для экспрессии в Saccharomyces cerevisiae и для Pichia pastoris, например, промотор aoxl, или промотор fmd, или промотор mox для Н. polymorpha.
ДНК, которая является подходящей для введения в клетки, может также содержать кроме ДНК по изобретению ДНК, которая получена или выделена из любого источника. Примером полученной ДНК является последовательность ДНК, которая была идентифицирована в данном организме как полезный фрагмент и затем химически синтезирована в основном в очищенной форме. Примером такой ДНК является подходящая последовательность ДНК, которая была, например, получена при использовании рестрикционных эндонуклеаз так, что она могла быть в дальнейшем обработана, например, амплифицирована в соответствии с изобретением. Ген amdS из Aspergillus nidulans, который может быть использован в качестве маркерного гена, и его регуляторные последовательности, а также полилинкеры применяются среди прочих inter alia.
Такая ДНК обычно называется рекомбинантной ДНК. Таким образом, подходящая ДНК содержит полностью синтетическую ДНК, полусинтетическую ДНК, ДНК, выделенную из биологических источников, и ДНК, полученную из введенной РНК. Обычно введенная ДНК не является исходной частью генотипа реципиентной ДНК, однако, в соответствии с изобретением ген может также быть выделен из данного генотипа и необязательно изменен, и впоследствии множественные копии гена могут быть введены в тот же самый генотип, например, для увеличения образования данного генного продукта.
Введенная ДНК содержит без ограничений ДНК из генов, таких как из бактерий, дрожжей, грибов или вирусов. Введенная ДНК может содержать модифицированные или синтетические гены, части генов или химерные гены, включая гены такого же или отличающегося генотипа. Она также может включать, например, ДНК плазмид pUC18, pUC19.
ДНК, использованная в соответствии с изобретением для трансформации, может быть кольцевой или линейной, двухцепочечной или одноцепочечной. В общем, ДНК является химерной ДНК, такой как плазмида ДНК, которая также содержит кодирующие участки, которые фланкированы с помощью регуляторных последовательностей и поддерживают экспрессию рекомбинатной ДНК, присутствующей в трансформированной клетке. Например, ДНК сама по себе может содержать или состоять из промотора, который является активным в клетке, который получен из источника, отличающегося от клетки, или промотора, который уже присутствует в клетке, то есть может быть использована трансформационная клетка-мишень.
В общем, введенная ДНК является относительно маленькой, менее чем около 30 т.п.н., для того чтобы минимизировать чувствительность к физическому, химическому или ферментативному разрушению, которое увеличивается с размером ДНК.
Выбор подходящего экспрессионного вектора зависит от клетки-хозяина. Экспрессионные векторы дрожжей или экспрессионные векторы грибов могут содержать начало репликации, подходящий промотор и энхансер, а также любые необходимые сайты связывания рибосом, сайты полиаденилирования, донорные сайты сплайсинга и акцепторные сайты сплайсинга, транскрипционные терминирующие последовательности и нетранскрибируемые 5'-фланкирующие последовательности.
Примерами подходящих клеток-хозяев являются клетки грибов рода Aspergillus, Rhizopus, Trichoderma, Neurospora, Mucor, Penicillium и т.д., такие как дрожжи видов Kluyveromyces, Saccharomyces, Schizosaccharomyces, Trichosporon, Schwanniomyces, Hansenula, Pichia и подобные. Подходящими системами хозяев являются, например, грибы, такие как Aspergilli, например, Aspergillus niger (ATCC 9142), или Aspergillus ficuum (NRLL 3135), или Trichoderma (например, Trichodenna reesei QM6a), и дрожжи, такие как Saccharomyces, например, Saccharomyces cerevisiae, или Pichia, такие как, например, Pichia pastoris, или Hansenula, например, Н. polymorpha (DSMZ 70277). Такие микроорганизмы могут быть получены из утвержденных депозитарных учреждений, например, the American Type Culture Collection (ATCC), the Centraalbureau voor Schimmelcultures (CBS), или the Deutsche Sammlung fur Mikroorganismen und Zeilkulturen GmbH (DSMZ), или любого другого депозитарного учреждения.
Экспрессионная кассета может включать в направлении транскрипции 5'-3' участок инициации транскрипции и участок инициации трансляции полинуклеотида по изобретению и участок транскрипции и участок терминации, которые являются функциональными in vivo или in vitro. Терминационный участок может быть нативным по отношению к транскрипционному инициирующему участку или может быть нативным или другого происхождения по отношению к полинуклеотиду. Регуляторные последовательности могут быть расположены против хода (5' некодирующие последовательности), внутри (интроны) или по ходу транскрипции (3'-некодирующие последовательности) кодирующей последовательности и могут влиять на транскрипцию, процессинг РНК или стабильность и/или трансляцию связанной кодирующей последовательности. Регуляторные последовательности могут содержать без ограничения энхансеры, промоторы, репрессорные связывающие сайты, трансляционные лидирующие последовательности, интроны или сигнальные последовательности полиаденилирования. Они могут содержать природные и синтетические последовательности, а также последовательности, которые являются комбинацией синтетических и природных последовательностей.
Вектор, использованный по изобретению, может также содержать подходящие последовательности для амплификации экспрессии.
Примерами промоторов, которые могут быть использованы по изобретению, являются промоторы, о которых известно, что они контролируют экспрессию в эукариотических клетках. Может быть использован любой промотор со способностью экспрессировать в нитчатых грибах. Примерами являются промоторы, которые сильно индуцируются крахмалом или целлюлозой, например, промотор глюкоамилазы или α-амилазы из рода Aspergillus, или промотор целлюлазы (целлобиогидролазы) из рода Trichoderma, промотор ферментов гликолитического метаболического пути, таких как фосфоглицераткиназа (ФГК) и глицеринальдегид-3-фосфатдегидрогеназа (ГФД) и т.д. Предпочтительным является промотор целлобиогидролазы-I, промотор целлобиогидролазы-II, промотор амилазы, промотор глюкоамилазы, промотор ксиланазы или промотор энолазы.
В дополнение к использованию специального промотора другие типы элементов могут влиять на экспрессию трансгенов. В особенности было продемонстрировано, что интроны имеют возможность амплифицировать трансгенную экспрессию.
Экспрессионная кассета может содержать дополнительные элементы, например, элементы, которые могут регулироваться эндогенными или экзогенными элементами, такими как белки «цинковые пальцы», включающие встречающиеся в природе белки «цинковые пальцы», или химерные белки «цинковые пальцы».
Экспрессионная кассета, использованная в соответствии с изобретением, может также содержать энхансерные элементы или расположенные против хода промоторные элементы.
Векторы для использования в соответствии с изобретением могут быть созданы таким путем, что они содержат энхансерный элемент. Таким образом, конструкты по изобретению содержат целевой ген вместе с 3'-последовательностью ДНК, которая выступает в качестве сигнала для терминации транскрипции и позволяет полиаденилирование полученной таким образом мРНК. Может быть использована любая сигнальная последовательность, которая делает возможным секретирование из выбранного организма-хозяина. Предпочтительной сигнальной последовательностью является фосфолипазная сигнальная последовательность из Aspergillus fumigatus или сигнальная последовательность, полученная на основании этого для секретирования из нитчатых грибов.
Также может быть использована специальная лидирующая последовательность, поскольку последовательность ДНК между транскрипционным инициирующим сайтом и началом кодирующей последовательности, то есть нетранслируемая лидерная последовательность, может влиять на экспрессию генов. Предпочтительная лидерная последовательность содержит последовательности, которые контролируют оптимальную экспрессию сцепленного гена, то есть они содержат предпочтительную консенсусную лидерная последовательность, которая увеличивает или сохраняет стабильность мРНК и предотвращает неподходящую инициацию трансляции. Выбор таких последовательностей хорошо известен специалисту в данной области техники.
Для того чтобы улучшить возможность идентификации трансформантов, в экспрессионную кассету может быть включен выбираемый или обнаруживаемый маркерный ген. Такие маркерные гены хорошо известны специалисту в данной области техники.
Экспрессионная кассета или векторный конструкт, который содержит экспрессионную кассету, вводится в клетку-хозяина. Разнообразие методик доступно и хорошо известно квалифицированному специалисту в области техники по введению конструктов в клетку-хозяина. Трансформация микробных клеток может быть проведена посредством полиэтиленгликоля, хлорида кальция, вирусной инфекции, диэтиламиноэтилдекстрана (ДЭАЭ декстрана), фаговых инфекций, электропорации и других способов, известных в существующем уровне техники. Трансформация грибов может быть проведена в соответствии с Penttila et al., Gene 61:155-164, 1987. Введение рекомбинантного вектора в дрожжи может быть проведено в соответствии со способами, известными per se, включающими электропорацию, использование сферопластов, ацетата лития и подобными.
Как только получена экспрессионная кассета или последовательность ДНК по изобретению, она может быть введена в векторы в соответствии со способами, известными per se, для того чтобы избыточно экспрессировать закодированный полипептид в подходящих системах-хозяевах. Однако последовательности ДНК как таковые также могут быть использованы для трансформации подходящих систем-хозяев по изобретению для того, чтобы получить избыточную экспрессию закодированного полипептида.
Как только последовательность ДНК в соответствии с изобретением экспрессирована в подходящей клетке-хозяине в подходящей среде, закодированная фосфолипаза может быть сконцентрирована и/или выделена в соответствии с процессами, известными per se, либо из среды, если фосфолипаза выделена в среду, или из организма-хозяина, если фосфолипаза присутствует внутри клетки, например, в периплазматическом пространстве. Известные процессы для выделения нерастворимых частей культуральной среды и биомассы с последующими процессами концентрирования фосфолипазы могут быть использованы для получения концентрированных растворов фосфолипазы или для подготовки высушивания фосфолипазы. Например, процессы фильтрования или процессы центрифугирования могут быть использованы для удаления нерастворимых компонентов с последующими процессами ультрафильтрации для концентрирования или используются процессы фильтрации в тангенциальном потоке. Высушивание может быть проведено посредством замораживания или распылительной сушки, процессов гранулирования, экструзии или других процессов. Могут быть использованы известные процессы очистки белков для выделения фосфолипаз в соответствии с изобретением. Например, различные хроматографические или гель-хроматографические процессы могут быть использованы по отдельности или в комбинации. В зависимости от использованной клетки-хозяина в процессе рекомбинантного получения фермент по изобретению может быть или может не быть ковалентно модифицирован посредством гликозилирования. В эукариотических клетках гликозилирование секретированных белков обеспечивает основу для модуляции фолдинга белка, конформационной стабильности, термической стабильности и устойчивости к протеолизу. Что касается специфического применения фосфолипазы, гликозилированный вариант фермента может быть предпочтительным перед негликозилированным вариантом.
Изобретение также относится к композициям выделенной или по существу очищенной нуклеиновой кислоты и композициям белка. Выделенный и очищенный полинуклеотид/полипептид или сегмент указанного относится к полинуклеотиду, или полипептиду, или сегменту указанного, который выделен из нативной среды и присутствует в очищенной форме для дальнейшего применения. Выделенный сегмент полинуклеиновой кислоты или полипептид может присутствовать в очищенной форме или может присутствовать в неестественной среде, такой как трансгенная клетка-хозяин. Например, выделенный или очищенный сегмент полинуклеотида, или белок, или биологически активная часть указанного, в основном, не содержит дополнительного клеточного материала или культуральной среды, если получен в соответствии с рекомбинантными методиками или, в основном, не содержит химических предшественников или других химических соединений. Выделенный полинуклеотид предпочтительно не содержит последовательностей (предпочтительно кодирующих белки последовательностей), которые естественным образом фланкируют нуклеиновую кислоту (то есть последовательности, которые расположены на конце 5' и конце 3' нуклеиновой кислоты) в геномной ДНК организма, из которой нуклеиновая кислота получена. Например, в соответствии с различными воплощениями молекула выделенной нуклеиновой кислоты может содержать менее чем приблизительно 5 т.п.н., 4 т.п.н., 3 т.п.н., 2 т.п.н., 1 т.п.н., 0.5 т.п.н. или 0.1 т.п.н. нуклеотидных последовательностей, которые естественным образом фланкируют молекулу нуклеиновой кислоты в геномной ДНК клетки, из которой получена нуклеиновая кислота. Белок, который в основном не содержит клеточный материал, содержит композиции белка и полипептида с менее чем приблизительно 70%, 50%, 30%, 20%, 10%, 5% (по отношению к сухой массе) загрязняющего белка. Если белок по изобретению или биологически активный фрагмент указанного образуется рекомбинантным путем, культуральная среда предпочтительно содержит менее чем приблизительно 70%, 50%, 30%, 20%, 10%, 5% (по отношению к сухой массе) химических предшественников или не похожих не белок химических субстанций.
Изобретение также относится к композициям фосфолипазы, которые содержат полипептид по изобретению. Фосфолипазные композиции обычно являются жидкими или сухими. Жидкие композиции предпочтительно содержат фосфолипазный фермент в очищенной или обогащенной форме. Однако могут быть добавлены вспомогательные вещества, такие как стабилизатор и/или глицерин, сорбитол или монопропиленгликоль, аддитивы, такие как соли, сахар, консерванты, агенты для установления значения рН и белки. Типичные жидкие композиции являются водными или масляными суспензиями.
Сухие композиции могут быть высушенными замораживанием, высушенными распылением, гранулированными или экструдированными композициями, которые могут включать только фермент. Сухие композиции могут быть гранулятами, которые могут быть легко смешаны с, например, пищевыми или кормовыми компонентами, или которые предпочтительно образуют компонент премикса. Предпочтительно, размер частиц гранулята фермента является сравнимым с размером другого компонента смеси. Это позволяет для безопасных и целевых агентов вводить ферменты в обработанную пищу, премиксы или корм для животных, например.
Сухие композиции могут также содержать другие аддитивы, такие как соли, особенно фосфатные соли и их ангидридные формы, и стабилизаторы, такие как поли(винилпирролидон) и т.д., для регулирования конкретных условий, таких как значение рН при применении.
Пищевая добавка в соответствии с этим воплощением настоящего изобретения может быть соединена с другими пищевыми компонентами подобным образом, посредством чего образуются обработанные пищевые продукты. Такие другие пищевые компоненты содержат одну или более ферментную добавку, витамины, минералы или микроэлементы. Полученная таким образом диетическая добавка может быть затем смешана с другими пищевыми компонентами, такими как зерно или растительные белки, в подходящем количестве с получением обработанной пищи. Обработка этих компонентов для обработанной пищи может быть проведена посредством обрабатывающих устройств, известных per se.
В предпочтительном воплощении фосфолипазные композиции по изобретению дополнительно содержат эффективное количество одного иди более фермента(ов) для пищи или корма для животных, или для применения на предварительных стадиях для получения пищи или корма для животных, или для применения в текстильной промышленности, предпочтительно выбранных из альфа-галактозидаз, бета-галактозидаз, лакказ, других фосфолипаз, фосфатаз, эндоглюканаз, особенно эндо-бета-1,4-глюканаз, эндо-бета-1,3(4)-глюканаз, эндо-1,2-бета-глюканаз и эндо-1,3-альфа-глюканаз, целлюлаз, ксилозидаз, галактаназ, особенно арабиногалактан-эндо-1,4-бета-галактозидаз и арабиногалактан-эндо-1,3-бета-галактозидаз, разрушающих пектин ферментов, особенно пектиназ, пектинестераз, пектинлиаз, полигалактуроназ, арабананаз, рамногалактуроназ, рамногалактуронанацетилестераз, рамногалактуронан-альфа-рамнозидаз, пектатлиаз и альфа-галактуронидаз, маннаназ, бета-маннозидаз, маннанацетилэстераз, ксиланацетилэстераз, протеаз, ксиланаз, арабиноксиланаз, липолитических ферментов, таких как липазы, дигалактозид-диглицеринэстеразы и кутиназы, и других ферментов, таких как лакказы и трансглутаминазы.
Фосфолипазы по изобретению могут быть использованы для разнообразных применений. Примерами являются применения в пекарном деле и в кормлении животных, а также в производстве топлив из возобновляемых источников энергии, таких как рапс или в производстве текстильных сырьевых материалов.
Предпочтительным применением является использование полипептидов, обладающих фосфолипазной активностью по изобретению, в процессах дегуммирования растительного масла. Пищевое масло, которое необходимо дегуммировать, например, обрабатывается полипептидом по изобретению, при этом большинство фосфолипидов гидролизуется, и затем водная фаза, содержащая гидролизованные фосфолипиды, отделяется от масла. Такой процесс является особенно подходящим для очистки пищевых масел, содержащих фосфолипиды, например, растительных масел, таких как соевое масло, рапсовое масло и подсолнечное масло.
До обработки фосфолипазой масло предпочтительно предварительно обрабатывается для удаления смолистых веществ, например, путем влажного рафинирования. Как правило, масло содержит от 50 до 850 м.д. фосфора в виде фосфолипида в начале обработки фосфолипазой по изобретению. После обработки содержание фосфора составляет, как правило, между 2 и 10 м.д.
Обработка фосфолипазой обычно проводится таким образом, что фосфолипаза диспергируется в водном растворе, предпочтительно в форме каплей со средним диаметром <10 мкм. Количество воды предпочтительно составляет от 0.5 до 5 масс.% (масс./масс.) по отношению к маслу. Необязательно может быть добавлен эмульгатор. Он может быть механически перемешан для сохранения эмульсии. Обработка фосфолипазой может быть проведена при значении рН в интервале от приблизительно 3.5 до приблизительно 5.0. Значение рН процесса может быть в интервале от приблизительно 3.5 до приблизительно 5, предпочтительно от 3.8 до 4.5 и наиболее предпочтительно от 4.0 до 4.2 для того чтобы максимально увеличить действие фермента. Значение рН может быть установлено путем, например, добавления лимонной кислоты, цитратного буфера, фосфорной кислоты или соляной кислоты. Подходящей температурой является обычно 30°-70°С, предпочтительно 45°-65°С и наиболее предпочтительно 55°-62°С. Время реакции, как правило, составляет от 1 до 12 часов, предпочтительно от 2 до 6 часов. Подходящая дозировка фермента составляет обычно от 120 до 3,000 единиц на кг масла, предпочтительно от 250 до 2,000 и наиболее предпочтительно от 750 до 1,500 единиц на кг масла.
Обработка фосфолипазой может быть проведена периодически, например, в реакторе с мешалкой, или может быть непрерывной, например, в последовательно соединенных проточных реакторах с мешалкой.
За обработкой фосфолипазой следует разделение на водную фазу и масляную фазу. Разделение может быть проведено путем традиционных средств, например, центрифугированием. Водный раствор содержит фосфолипазы, и фермент может быть снова использован для увеличения экономичности процесса.
Обработка может быть проведена посредством процессов, известных per se.
Преимущественно, фосфолипаза по изобретению может быть также использована для приготовления теста и хлебобулочных изделий, при этом в тесто вводится эффективное количество полипептида по изобретению. При добавлении полипептида, обладающего фосфолипазной активностью по изобретению, одно или несколько свойств(о) теста или хлебобулочного изделия, приготовленного с помощью теста, может быть улучшено по сравнению с тестом или хлебобулочным изделием, к которому не добавлялся полипептид, обладающий фосфолипазной активностью по изобретению.
При приготовлении теста с помощью фосфолипазы по изобретению фосфолипаза может быть добавлена в само тесто, к любому ингредиенту, из которого готовится тесто, и/или к смеси ингредиентов для теста, из которой готовится тесто. Полипептид, обладающий фосфолипазной активностью по изобретению, может таким образом быть добавлен как таковой на любой стадии приготовления теста или может быть добавлен на одной, двух или более стадии(ях). Здесь эффективное количество относится к количеству фосфолипазы, которое является достаточным для того, чтобы вызвать поддающееся измерению влияние на по крайней мере одно целевое свойство теста и/или хлебобулочного изделия.
Выражение «улучшенное свойство» определяется в настоящей заявке как любое свойство теста и/или продукта, который получен из теста, особенно хлебобулочного изделия, которое было улучшено благодаря влиянию фосфолипазы по отношению к тесту или продукту, к которому фосфолипаза по изобретению не была добавлена. Улучшенное свойство может включать, например, улучшенную упругость теста, улучшенную эластичность теста, улучшенную стабильность теста, пониженную липкость теста, улучшенную способность к растяжению теста, улучшенную обрабатываемость теста, увеличенный объем хлебобулочного изделия, улучшенную зернистую структуру хлебобулочного изделия, улучшенную мягкость хлебобулочного изделия, улучшенный аромат хлебобулочного изделия и/или замедленное очерствение хлебобулочного изделия. Процессы для определения этих свойств хорошо известны в существующем уровне техники.
Тесто определяется в настоящей заявке как смесь муки и других ингредиентов, которая достаточно твердая для замешивания или раскатывания. Тесто может быть свежим, замороженным, предварительно подготовленным или предварительно запеченным.
Выражение «хлебобулочное изделие» в настоящей заявке относится к любому продукту, который готовится из теста и является либо мягким, либо хрустящим. Примерами хлебобулочных изделий, которые могут быть приготовлены с помощью фосфолипазы по изобретению, являются, например, хлеб (особенно белый хлеб, цельномолотый хлеб или ржаной хлеб), обычно в форме буханок или французского хлеба типа французского багета, паста, пита, тортилья, тако, кексы, блины, печенье и пирожные, запеченный хлеб, дважды запеченный хлеб и подобные.
В приготовлении этих хлебобулочных изделий может быть добавлен полипептид, обладающий фосфолипазной активностью по изобретению, и/или один или более дополнительных фермент(ов) в любом составе, который является подходящим для соответствующего применения, например, в сухой форме, в качестве жидкости или в качестве премикса. Кроме того, один или более дополнительных фермент(ов) могут быть добавлены в тесто. Эти дополнительные ферменты могут быть любого происхождения и могут быть получены, например, из млекопитающих и растений. Предпочтительно они имеют микробное происхождение и особенно предпочтительными являются полученные из бактерий или грибов.
В соответствии с предпочтительным воплощением дополнительные ферменты могут быть амилазами, такими как α-амилаза (подходящая для получения Сахаров, которые ферментируются дрожжами, и для замедленного очерствения) или β-амилаза, циклодекстринглюканотрансферазой, пептидазой, особенно экзопептидазой (подходящей для усиления аромата), трансглутамазой, липазой (подходящей для модифицирования липидов, присутствующих в тесте или в составляющих теста, для того, чтобы сделать тесто мягче), фосфолипазой (пригодной для модифицирования липидов, присутствующих в тесте или в составляющих теста, для того, чтобы сделать тесто мягче и для улучшения газоудерживающей способности теста), целлюлазой, гемицеллюлазой, особенно пентозаназой, такой как ксиланаза (пригодная для частичного гидролиза пентозанов, которые улучшают растяжимость теста), протеазой (пригодной для смягчения глютена, особенно если используется мука из твердой пшеницы), протеиндисульфидизомеразой (например, протеиндисульфидизомеразой, раскрытой в WO 95/00636), гликозилтрансферазой, пероксидазой (пригодной для улучшения консистенции теста), лакказой или оксидазой, например, альзозоксидазой, глюкозоксидазой, пиранозоксидазой, липоксигеназой или L-аминокислотной оксидазой (пригодной для улучшения консистенции теста).
Этот/Эти необязательный(ые) дополнительный(ые) фермент/ферменты может/могут быть необязательно добавлен(ы) отдельно или вместе в полипептидом, обладающим фосфолипазной активностью по изобретению, в качестве компонентов агентов для выпекания или добавок к тесту. Изобретение также относится к приготовлению такого теста, а также к приготовлению соответствующих хлебобулочных изделий, сделанных из этого теста.
Изобретение также относится к премиксу, например, в форме мучной композиции для приготовления теста и/или хлебобулочных изделий, сделанных из теста, при этом этот премикс содержит полипептиды, обладающие фосфолипазной активностью по изобретению.
Полипептиды, обладающие фосфолипазной активностью по изобретению, могут также быть использованы в качестве добавок к кормам для животных. При добавлении фосфолипаз к кормам для животных, увеличивается эффективность использования кормов для животных. Таким образом, улучшается рост животных, которые питаются таким кормом. Фосфолипаза по изобретению может при этом быть добавлена сама по себе или в качестве кормового концентрата. Кроме того, фосфолипаза может также быть добавлена к корму для животных посредством трансгенных растений, в которых фосфолипаза была синтезирована путем экспрессии гетерологичного гена. Процессы производства таких трансгенных растений раскрыты в ЕР 0449376.
Полипептиды, обладающие фосфолипазной активностью по изобретению, могут также быть использованы в процессе выварки при производстве текстильных сырьевых материалов, таких как хлопковые волокна, для облегчения дальнейшей обработки волокон. Полученные улучшения при выварке влияют на эксплуатационные свойства в ходе окрашивания, а также на дальнейшую механическую и ферментную обработку волокна и ткани, полученной из него.
Ген для фосфолипазы, который был выделен из микроорганизма Aspergillus fumigates, депонирован в плазмиде pPL3949-Topo2.5 под номером доступа DSM 22741 в Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ), Inhoffenstraue 7B, D-38124 Braunschweig 2 июля 2009 в соответствии с положениями Будапештского договора.
Краткий перечень фигур
Изобретение далее описывается на основании приложенных фигур. Оно показано на:
Фигуре 1: IEF гель очищенной фосфолипазы из Aspegillus fumigatus.
Полоса 1: маркерный белок из Набора Калибровки Изоэлектрического Фокусирования, рН 2.5-6.5,
Полоса 2-3: Бэнд фосфолипазы при pl приблизит.4.1, указанный стрелкой.
Фигуре 2: Кривая оптимума Т для рекомбинантной фосфолипазы, экспрессированной в Trichoderma reesei RH32664.
Фигуре 3: Кривая оптимума рН для рекомбинантной фосфолипазы, экспрессированной в Trichoderma reesei RH32664.
Фигуре 4: Нуклеотидная последовательность и аминокислотная последовательность, полученные из хромосомного фосфолипазного гена из Aspergillus fumigatus RH3949. Интроны напечатаны курсивом и аминокислотные последовательности -жирным шрифтом (SEQ ID NO:1).
Фигуре 5: Нуклеотидная последовательность хромосомного фосфолипазного гена из Aspergillus fumigatus RH3949 (SEQ ID NO:1).
Фигуре 6: Аминокислотная последовательность фосфолипазного гена из Aspergillus fumigatus RH3949 (SEQ ID NO:2).
Фигуре 7: Рестрикционная карта вектора pPL3949-Topo2.5
Фигуре 8: Рестрикционная карта экспрессионного вектора pAB500-PL3949.
Осуществление изобретения
Справочный пример 1
Определение фосфолипазной активности
1 единица фосфолипазы (PLU) соответствует количеству фермента, который высвобождает 1 мкмоль жирной кислоты в минуту из фосфатидилхолина при стандартных условиях.
Реагенты:
Эмульсия субстрата:
1 г Epikuron 200 (очищенный фосфатидилхолин, полученный из сои, компании Lucas Meyer, в настоящее время доступный у Cargill), 100 мл деионизированной воды и 5 мл 0.32 М раствора CaCl2 гомогенизировали с помощью Ultra Turrax в течение 2 мин при 24,000 об/мин. Эмульсия субстрата является стабильной при 4°-8°С в течение 3-4 д.
Другие растворы:
0.32 раствор MgCl2, свежий раствор 3.3. мМ моногидрата лимонной кислоты, 10 мМ раствора КОН, 1% раствор Тритон Х100 (компания Fluka) в деминерализованной воде.
Раствор фермента:
Составы фермента растворяют в деионизированной воде. Концентрация фермента в партии может не превышать 2.5 ед./г
Процедура:
Основные пробы
10 мл эмульсии субстрата
10 мл 1% раствора Тритон Х100.
5 мл 3.3 мМ раствора моногидрата лимонной кислоты отмеряли пипеткой в 25-мл колбу Эрленмейера с широким горлом и выдерживали при 40°С в течение 10 мин. Значение рН устанавливается приблизительно 3.3.-3.5. После добавления 0.1 мл раствора фермента анализируемую партию инкубировали в течение 10 мин при 40°С. Когда время инкубации закончилось, раствор титровали 10 мМ КОН до рН 10.0, при этом первые 5 мл КОН добавляют быстро (продолжительность около 1 мин). Записывают расход КОН.
Холостые пробы
Исходный раствор фермента нагревают в течение 15 мин при 95°С и таким образом деактивируют. После охлаждения до комнатной температуры дальнейшую обработку проводят как для основных проб.
Инкубация холостых проб не является необходимым.
Оценка:

Справочный пример 2
Быстрый тест для определения фосфолипазы при рН 3.5
Реагенты:
Эмульсия субстрата:
1 г Epikuron 200 смешивают со 100 мг воды Milli Q и 5 мл 0.32 М раствора хлорида кальция и гомогенизируют с помощью Ultra-Turrax (в течение около 1-2 мин при 24000 об/мин).
Для анализа партии 10 мл этой эмульсии субстрата смешивают с 10 мл 1% раствора Тритон Х-100 и 5 мл 3.3 мМ раствора моногидрата лимонной кислоты.
Свободные жирные кислоты, полу-микротест (Boehringer)
(Полу-микротест содержит реагенты для реакционной смеси А, реакционной смеси В и N-этилмалеимид).
Процедура:
Сначала 5 мкл разбавленного раствора фермента переносят в микротитрационный планшет и смешивают с 0.1 мл эмульсии субстрата.
Партии субстрат/фермент инкубируют на водяной бане в течение 10 мин при 40°С.
После этого 5 мкл партии переносят пипеткой во второй микротитрационный планшет к 100 мкл реакционной смеси А и инкубируют в течение 5 мин при 40°С.
После того как время инкубации заканчивается, добавляют 5 мкл смеси реакционной смеси В и раствора N-этилмалеимида в соотношении 1:1 и снова инкубируют в течение 5 мин при 40°С.
Фосфолипазная активность присутствует, если реакционная партия становится красного цвета.
Справочный пример 3
Определение содержания свободных жирных кислот
Реагенты:
Этанол (96% раствор), толуол.
Этанол и толуол смешивают в соотношении 1:1 (об./об.).
0.1 н. спиртовой КОН.
1% раствор фенолфталеина в этаноле.
Процедура:
Приблизительно 3 г безводного масла взвешивают в колбе Эрленмейера с точностью до десятого знака, растворяют в 20 мл смеси этанол-толуол, смешивают с 2-3 каплями фенолфталеина и титруют 0.1 н. раствором КОН до тех пор, пока оно имеет устойчивый красный цвет.
Оценка:
Кислотное число - это индикатор содержания свободных жирных кислот. Кислотное число относится к количеству гидроксида калия в г, необходимому для нейтрализации свободных жирных кислот, содержащихся в 1 кг масла. Кислотное число (КЧ) рассчитывается в соответствии со следующим уравнением:
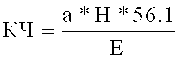
а - количество использованного раствора КОН в мл
Н - нормальность КОН
Е - количество взвешенного жира в г
56.1 - молярная масса КОН в г/моль.
Кислотное число может быть использовано для расчета содержания свободных жирных кислот (СЖК) в процентах:

282 - молярная масса олеиновой кислоты.
Справочный пример 4
Определение липазы
Количественное определение липазы на оливковом масле-агаре проводится аналогично в соответствии со способом Kouker and Jaeger (Applied Environ. Microbiol., 59: 211-213 (1987)).
Для определения липазной активности используют агаровые пластинки, сделанные из агара с трибутирином (Fluka 91015), которые были получены с использованием 1% оливкового масла. Значения рН устанавливается 5.5.
Определение липазной активности проводится фотометрически с эмульгированным п-нитрофенилпальмитатом (Sigma N2752) в качестве субстрата в 0.5 М цитрат/фосфатном буфере, рН 5.1 в соответствии с Winkler and Stuckmann (1979) (J. Вас., 138:663-670 (1979)).
Пример 1
Экстракция фосфолипазы из штамма RH3949 Aspergillus fumigatus
Aspergillus fumigates выращивали в 200 мл плоскодонных колбах с 50 мл среды при 28°С, 200 об/мин более 5 д. Среда состояла из 0.5% Epicuron 200 (Lucas Meyer), 0.5% порошка кукурузного экстракта, 0.2% NH4NO3, 100 мМ KH2O4 и 0.1% Тритон Х100. Значение рН устанавливали до рН 6 перед стерилизацией. Среду инокулировали суспензией спор. После 5 дней культуральный супернатант отделяли от мицелия посредством фильтрования и измеряли фосфолипазную активность в жидкости.
Пример 2
Очистка фосфолипазы из штамма RH3949 Aspergillus fumigates
Стадия 1: Анионообменник, Macro Prep Q
Для того чтобы очистить фосфолипазу, сначала проводили выделение белков на анионообменнике Macro Prep Q.
С этой целью концентрированный культуральный супернатант из культур по примеру 1 разбавляли полностью обессоленной водой до тех пор, пока раствор белка имел такую же проводимость, что и буфер А. После этого образец белка довели до рН 7 с помощью 1 М NaOH и загрузили на колонку, уравновешенную буфером А. После промывки колонки буфером А фосфолипазу разбавили линейно увеличивающимся градиентом NaCl 0-1 М. Фракции, обладающие фосфолипазной активностью, были соединены и далее очищены.
Буфер А: 5 мМ CaCl2+20 мМ Tris-HCl, рН 7.0
Буфер В: 5 mM СаС2+20 тМ Tris-HCl, рН 7.0+1 М NaCl
Стадия 2: Хроматография с гидрофобным взаимодействием, низкое замещение в быстром потоке фенилсефарозой 6
Образец белка, обладающий фосфолипазной активностью согласно стадии 1, смешали с 3.4 М раствором сульфата аммония в соотношении 1:1 и установили рН 7.0 с помощью 1 М раствора NaOH. После нанесения образца на фенилсефарозную колонку, которую также уравновешивали буфером А, фосфолипазу разбавляли уменьшающимся градиентом сульфата аммония.
Буфер А: 5 мМ CaCl2+20 мМ Tris-HCl, рН 7.0+1.7 М сульфата аммония
Буфер В: 5 мМ CaCl2+20 мМ Tris-HCl, рН 7.0
Стадия 3: Гель-фильтрация, Супероза 12 HR 10/30
В качестве конечной стадии очистки белки выделяли в гель-фильтрационной колонке. Для этого образец фосфолипазы согласно стадии 2 диализировали против полностью обессоленной воды в диализной трубке (Naturin protein farce) в течение 1.5 ч и затем лиофилизовали. Лиофилизат абсорбировали в 500 мкл полностью обессоленной воды. 250 мкл вводили в колонку в две стадии соответственно и разбавляли буфером А.
Буфер А: 5 мМ CaCl2+20 мМ Tris-HCl, рН 7.0
Очищенную фосфолипазу вносили в гель IEF. Результаты представлены на Фигуре 1.
Бэнды были отрезаны для идентифицирования и исследования фосфолипазной активности в соответствии с описанным способом анализа.
Пример 3
N-концевое секвенирование белка
После конечной стадии очистки с помощью суперозы очищенный белок выделяли на нативном геле. Белковые бэнды, обладающие фосфолипазной активностью, были отрезаны и опять нанесены на гель ДСН для того, чтобы определить молекулярную массу. Для определения N-концевых аминокислот белковые бэнды были перемещены из нативного геля на ПВДФ мембрану (Fluotrans Transfer Membrane, Pall), и определяли N-концевые аминокислотные последовательности в аминокислотном секвенаторе (Applied Biosystems Model 470A) в соответствии с процессом окрашивания Кумасси. Они представляют собой:
1DVSAS VLQKL SLFAQ Y16 (SEQ ID NO:3)
Сравнение последовательностей показывает, что N-концевая аминокислотная последовательность фосфолипазного гена имеет высокую схожесть последовательности с геном внеклеточной липазы из штамма Af293 Aspergillus fumigatus (GenBank EAL86100).
Пример 4
Клонирование хромосомного фосфолипазного гена из штамма RH3949 Aspergillus fumigatus посредством полимеразной цепной реакции (ПЦР)
Различные олигомеры для амплификации фосфолипазы ДНК были получены из данных последовательности хромосомной ДНК липазы Aspergillus fumigatus. Приготовление хромосомной ДНК проводили в соответствии с измененными инструкциями Hynes, M.J et al., (1983) Mol. Cell. Biol. 3, 1430-1439. Амплификацию фосфолипазного гена проводили посредством способа ПЦР. Продукты ПЦР клонировали в плазмиду pCR2.1-TOPO и секвенировали. Здесь показано, что праймерная пара N2-3948 и 3949-ApaI приводит к гену, обладающему фосфолипазной ДНК по изобретению.
N2-3948 5'-AGAGTCTGCC TATATTCTCT CTGAAAGG-3' (SEQ ID NO. 4)
3949-ApaI 5'-TATGACAATTTCCGTGATTACTG-3' (SEQ ID NO:5)
Реакционная партия в 100 мкл содержала: 10 мкл 10×буфера (200 мМ Tris/HCl, pH 8.4, 500 мМ KCl), 3 мкл 50 мМ MgCl2, 2 мкл 10 мМ дНТФ, 50 пмоль каждого олигопраймера (N2-3948 и 3949-ApaI), приблизит. 10 нг хромосомной ДНК, 5U ДНК-полимеразы Taq (Invitrogen). Партию обрабатывали для денатурирования при 95°С/5 мин 45 циклов (95°С/1 мин, 45°С/1 мин, 72°С/1 мин) и затем проводили удлинение при 72°С /10 мин.
Продукты ПЦР очищали на колонке Qiaquick и клонировали в плазмиду pCR2.1-TOPO. После секвенирования один трансформант содержал фосфолипазную последовательность ДНК по изобретению (Фигура 5, SEQ ID NO:1) и назывался pPL3949-Торо2.5 (Фигура 7).
Открытая рамка считывания, которая кодирует фосфолипазу, содержит 1052 нуклеотидов, содержащих 299 аминокислот. Фосфолипазный ген содержит 3 интрона.
Полученная N-концевая аминокислотная последовательность соответствует пептидным последовательностям, определенным посредством белкового секвенирования (Пример 3, SEQ ID NO:3).
Полученная молекулярная масса приблизит. 28.6 кДа соответствует приблизит. 29 кДа, которые были определены посредством SDS-PAGE (Пример 8).
Определение сигнальной последовательности проводили с помощью компьютерной программы (PSORT) Nakai and Kanehisa (1992, Genomics 14, 897-911). В соответствии с этой программой фосфолипазный ген имеет сигнальную последовательность из 21 аминокислоты и пропептид из 8 аминокислот.
Пример 5
Сборка экспрессионного вектора pAB500-PL3949
В экспрессионном векторе pAB500-PL3949 фосфолипазный ген Т. reesei контролируется промотором cbhl и терминатором cbhl.
Для сборки плазмиды pAB500-PL3949 ген, кодирующий фосфолипазу, был амплифицирован из плазмиды pPL3949.Topo2.5 посредством ПЦР. Продукт ПЦР гидролизовали ферментами AvrII/PacI и затем инсертировали в рестрикционные сайты SpeI и PacI после промотора Т. reesei cbh1 в плазмиде рАВ500. Полученная плазмида называется pAB500-PL3949.
Сборку плазмиды рАВ500 проводили в соответствии со следующими стадиями:
Путем введения дополнительных рестрикционных сайтов (SpeI и PacI в сайт SacII между промотором cbhl и терминатором cbhl) была получена плазмида рАВ487 из плазмиды pALK487 (WO 94/28117). Рестрикционные сайты SpeI-PacI используются для прямого клонирования фосфолипазного гена.
Ген amdS, включающий его промотор и его терминатор, был амплифицирован из плазмиды p3SR2 (GenBank 16371) посредством ПЦР. Продукт ПЦР был отрезан с помощью AscI и NruI и вставлен в расщепляющий сайт AscI/StuI рАВ487, тем самым была получена плазмида рАВ500.
Пример 6
Трансформация Т. reesei
Методики, использованные для трансформации и обработки Т. reesei, являлись теми в соответствии с Penttila et al. (1987, Gene 61: 155-164).
Т. reesei RH32439 трансформировали линеаризованной экспрессионной кассетой, выделенной из плазмиды pAB500-PL3949.
Траснформанты выделили и очистили посредством однократного выделения спор. Из всех трансформантов были выбраны те, которые имели наибольшую способность к секретированию, и использованы в Примере 7 для получения ферментного материала.
Пример 7
Получение ферментных растворов посредством ферментирования в плоскодонных колбах
Трансформанты, которые несут экспрессионную кассету из Примера 6, культивировали в индуцированной целлюлазой среде в плоскодонных колбах. Полученные культуральные фильтраты после 6 дней выращивания использовали для определения характеристик фермента (Пример 8) и для анализа процесса дегуммирования масла (Пример 9).
Пример 8
Определение характеристик рекомбинантных фосфолипаз
При определении молекулярной массы посредством электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE) получили молекулярную массу для фосфолипазы по изобретению приблизит. 29 кДа.
N-терминальное секвенирование рекомбинантной фосфолипазы проводилось компанией Chromatec (Германия). Аминокислотная последовательность, полученная посредством секвенирования, соответствует N-концевой последовательности фосфолипазы (Пример 3).
Идентификация рекомбинантного белка посредством MALDI-MS проводилось компанией Protagen (Германия). Результат подтверждает, что последовательность фосфолипазы по изобретению является правильной.
Температурную зависимость ферментной активности определяли при различных температурах посредством способа определения, как описано выше. Температурный оптимум фосфолипазы составляет 50°С (см. Фигуру 2).
Оптимальное значение рН ферментной активности определяли при различных значениях рН посредством метода определения, как описано выше. С этой целью значение рН в реакционной массе устанавливали с помощью лимонной кислоты.
Фермент является активным в широком интервале рН от 3 до 5 (см. Фигуру 3).
Липазную активность определяли посредством метода определения, как описано выше. Фосфолипаза A. fumigatus по изобретению обладает очень низкой липазной активностью. В соответствии с расчетами отношение фосфолипазной активности к липазной активности составило 7480:1 (с пределом экспериментальных вариаций ±10%).
Пример 9
Дегуммирование масла посредством фермента из культуральных супернатантов рекомбинантных штаммов Trichoderma reesei, имеющих ген из A. fumigatus RH3949
200 г пищевого масла (соевое масло 1 с содержанием фосфора 163.6 м.д.; рапсовое масло с содержанием фосфора 161.6 м.д.; соевое масло 2 с содержанием фосфора 592.8 м.д. и соевое масло 3 с содержанием фосфора 81.1 м.д.) смешивали с 0.42 мл 46% раствора лимонной кислоты и водой в 400 мл лабораторном стакане. При этом общее содержание воды не должно превышать 2%. Значение рН реакционной смеси устанавливали до рН 4.0 путем добавления 7% раствора NaOH (0.6 мл). После добавления раствора фермента по изобретению (50U) реакционную партию смешивали с помощью Ultra Turrax в течение 2 мин при 24000 об/мин. Затем смесь переливали в трехгорлую круглодонную колбу и выдерживали при 55°С или 60°С при медленном перемешивании (200 об/мин).
Образец в 20 мл отбирали каждые 120 мин после добавления раствора фермента. Образцы центрифугировали в течение 5 мин при 4300×g и фотометрически определяли содержание фосфолипида, выраженное в м.д. фосфора, как фосфорно-молибдатный комплекс при 830 нм в масле после сжигания при 850°С при добавлении оксида магния.
Рекомбинантный штамм Trichoderma reesei, который включает плазмиду pAB500-PL3949 по изобретению, называется RH32664.
Результаты, полученные в процессе дегуммирования пищевого масла при 55°С или 60°С с помощью фосфолипазы, полученной из рекомбинантного штамма RH3949 Aspergillus fumigates, показаны в Таблице 2.
Результаты показывают четкий эффект дегуммирования в процессе дегуммирования с использованием фермента по изобретению по сравнению с процессом водного дегуммирования с использованием лимонной кислоты. После только 4 часов содержание остаточного фосфора составляет менее чем 10 м.д.
Таблица 2: Дегуммирование соевого масла и рапсового масла посредством фосфолипазных ферментов по изобретению (250 U на кг пищевого масла) из культуральных супернатантов рекомбинантного штамма Trichoderma reesei, имеющего содержание воды менее 2% при рН 4.0.
Пример 10
Содержание свободных жирных кислот в процессе дегуммирования масла.
Определение содержания свободных жирных кислот в процессе дегуммирования масла проводили, как описано в справочном примере 3. Пищевое масло смешивали с фосфолипазой, как описано в Примере 9 и выдерживали в течение 6 часов при 57°С. Ферментный гидролиз фосфолипидов вызывали фосфолипазами, которые расщепляют жирные кислоты. Для целей сравнения был проведен тот же анализ с чистым пищевым маслом в качестве холостой пробы.
В противоположность холостой пробе (ХП) содержание свободных жирных кислот (СЖК) в образцах увеличивалось только немного, и разница (ΔСЖК) составляет 0.18% после обработки пищевого масла фосфолипазой при температуре 57°С (Таблица 3).
| название | год | авторы | номер документа |
|---|---|---|---|
| Мутант фосфолипазы С с высокой ферментативной активностью | 2019 |
|
RU2818353C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВАРИАНТА ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА И ЛИПОЛИТИЧЕСКИЙ ФЕРМЕНТ (ВАРИАНТЫ) | 1999 |
|
RU2235775C2 |
| ПОЛИПЕПТИД С АКТИВНОСТЬЮ ФОСФОЛИПАЗЫ И ЕГО ПРИМЕНЕНИЕ | 2004 |
|
RU2363731C2 |
| ЛИПОЛИТИЧЕСКИЕ ФЕРМЕНТЫ, ИХ ПРИМЕНЕНИЕ ПРИ ИСПОЛЬЗОВАНИИ ПИЩЕВЫХ ПРОДУКТОВ | 2010 |
|
RU2538144C2 |
| ПРИМЕНЕНИЕ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА В ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ | 2005 |
|
RU2406759C2 |
| БЕЛОК | 2005 |
|
RU2380414C2 |
| СПОСОБ | 2004 |
|
RU2376868C2 |
| ПОЛУЧЕНИЕ ЛИЗОГЛИКОЛИПИДА И БИОКОНВЕРСИЯ ГЛИКОЛИПИДОВ С ИСПОЛЬЗОВАНИЕМ ЛИПОЛИТИЧЕСКОГО ФЕРМЕНТА | 2005 |
|
RU2548805C2 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| ЛИПОЛИТИЧЕСКИЙ ФЕРМЕНТ ДЛЯ ПРИМЕНЕНИЯ В ХЛЕБОПЕЧЕНИИ | 2018 |
|
RU2763469C2 |




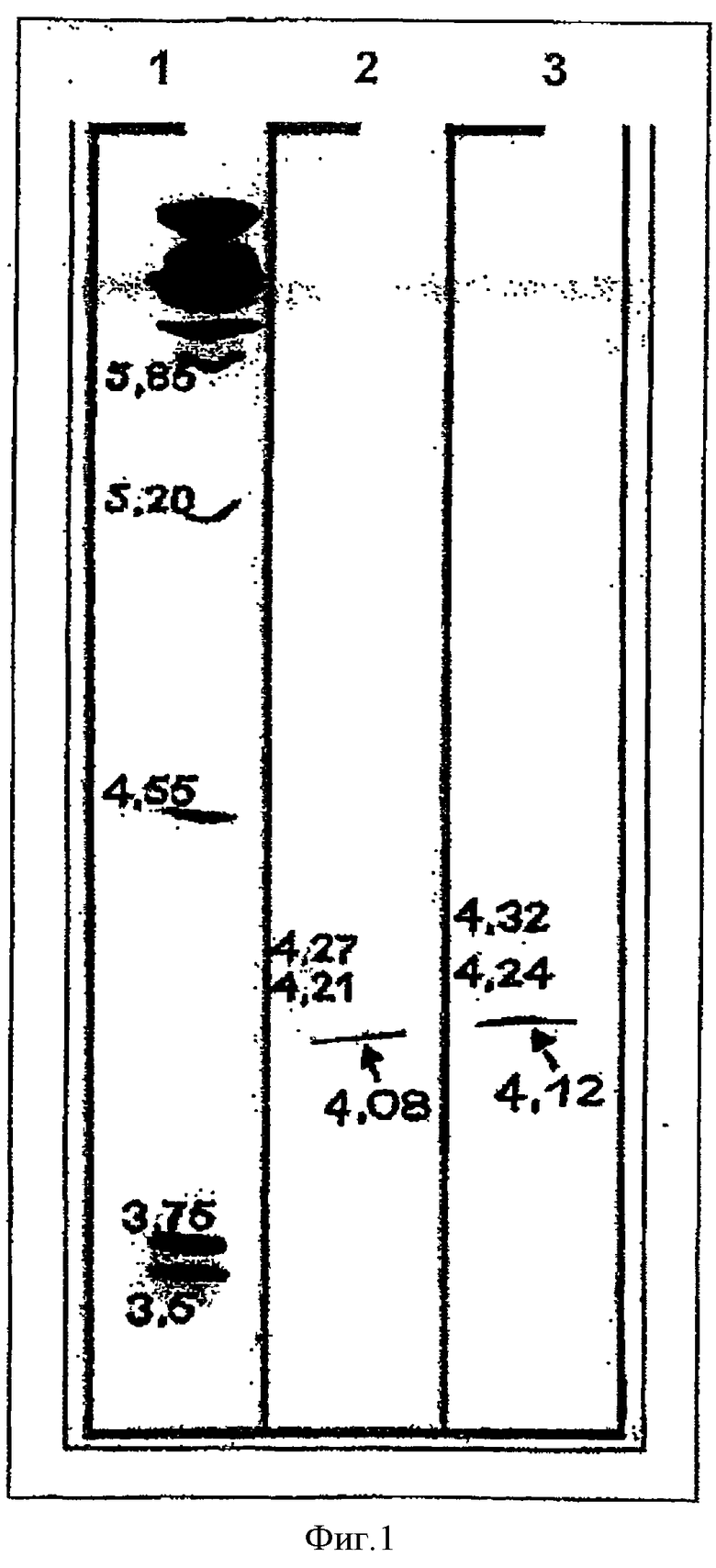
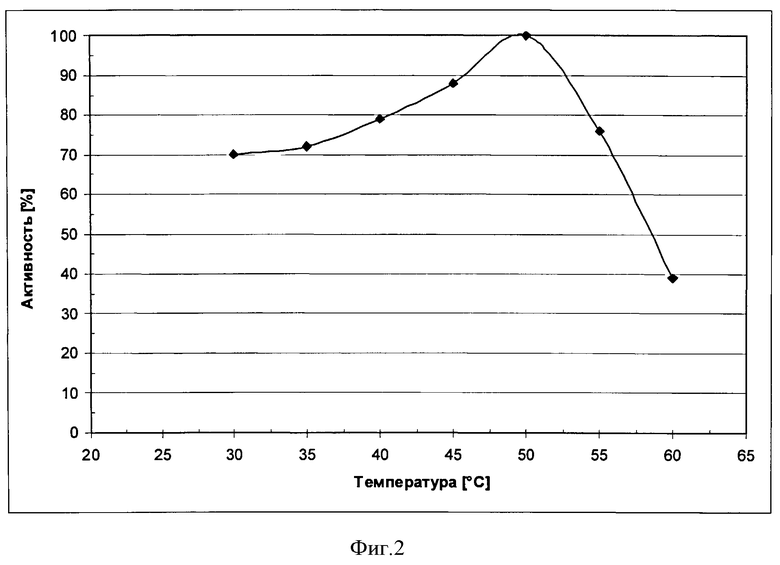





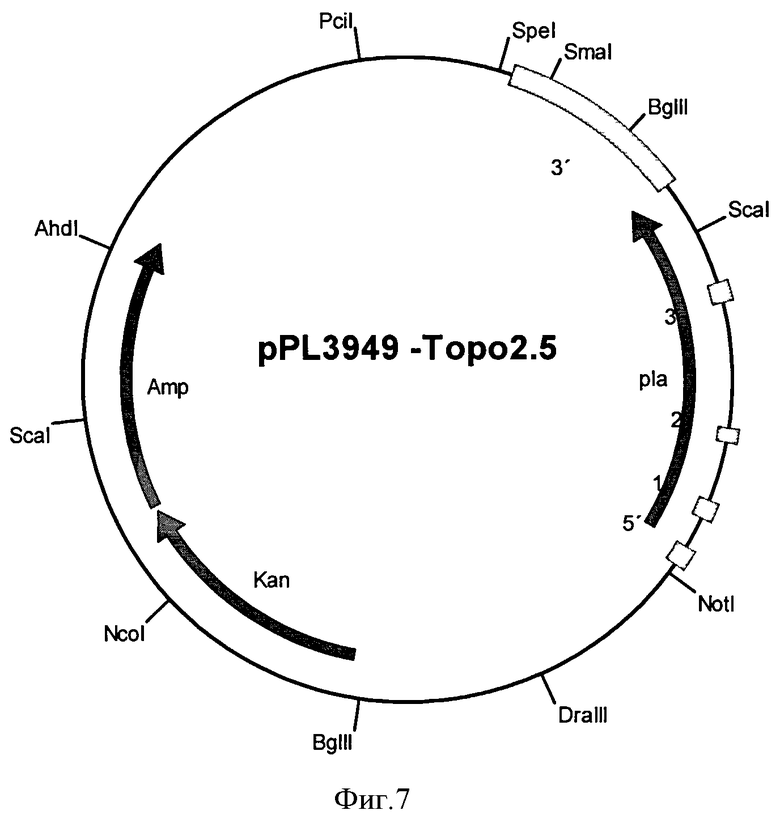

Изобретение относится к биотехнологии и представляет собой молекулу ДНК, которая кодирует полипептид, обладающий фосфолипазной активностью, по существу без липазной активности, причем последовательность молекулы ДНК предпочтительно происходит из Aspergillus и более предпочтительно из Aspergillus fumigatus. Изобретение относится также к полипептиду, кодируемому указанной молекулой ДНК, экспрессионному конструкту, клетке-хозяине, плазмиде, содержащим такую молекулу ДНК. Изобретение позволяет эффективно использовать полипептид согласно изобретению для дегуммирования растительного масла, для приготовления теста и/или хлебобулочных изделий, молочных продуктов, в качестве добавки к корму и для обработки текстильных материалов. 14 н. и 5 з.п. ф-лы, 8 ил., 3 табл., 9 пр.
1. Молекула ДНК, которая кодирует полипептид, обладающий фосфолипазной активностью, по существу без липазной активности, характеризующаяся тем, что последовательность молекулы ДНК выбирается из:
a) последовательности ДНК, которая содержит нуклеотидную последовательность SEQ ID NO: 1,
b) последовательности ДНК, которая содержит кодирующую последовательность в соответствии с SEQ ID NO: 1,
c) последовательности ДНК, которая кодирует аминокислотную последовательность SEQ ID NO: 2,
d) последовательности ДНК, которая гибридизуется в жестких условиях с одной из последовательностей ДНК согласно а), b) или с),
e) последовательности ДНК, которая отличается от последовательностей ДНК согласно а), b), с) или d) вследствие вырожденности генетического кода, и
f) последовательности ДНК, которая является комплементарной нитью к одной из последовательностей согласно а)-е),
причем последовательность молекулы ДНК предпочтительно происходит из Aspergillus и более предпочтительно из Aspergillus fumigatus.
2. Молекула нуклеиновой кислоты, которая кодирует полипептид, обладающий фосфолипазной активностью, по существу без липазной активности, характеризующаяся тем, что последовательность молекулы нуклеиновой кислоты выбирается из:
a) последовательности ДНК, которая содержит нуклеотидную последовательность SEQ ID NO: 1, или последовательности, которая обладает идентичностью по меньшей мере 88% с указанной последовательностью;
b) последовательности ДНК, которая гибридизуется с нуклеотидной последовательностью SEQ ID NO: 1 в жестких условиях;
c) последовательности ДНК, которая является комплементарной нитью к нуклеотидной последовательности SEQ ID NO: 1;
d) последовательности ДНК, которая содержит кодирующую последовательность в соответствии с SEQ ID NO: 1, или последовательности, которая обладает идентичностью по меньшей мере 88% с указанной ДНК последовательностью;
e) последовательности ДНК, которая гибридизуется в жестких условиях с последовательностью ДНК, которая содержит кодирующую последовательность в соответствии с SEQ ID NO: 1;
f) последовательности ДНК, которая является комплементарной нитью к последовательности ДНК, которая содержит кодирующую последовательность в соответствии с SEQ ID NO: 1;
g) последовательности ДНК, которая кодирует аминокислотную последовательность SEQ ID NO: 2, или последовательности, которая обладает идентичностью по меньшей мере 88% с указанной ДНК последовательностью; h) последовательности ДНК, которая гибридизуется в жестких условиях с последовательностью ДНК, которая кодирует аминокислотную последовательность SEQ ID NO: 2;
и
i) последовательности ДНК, которая является комплементарной нитью к последовательности ДНК, которая кодирует аминокислотную последовательность SEQ ID NO: 2.
3. Экспрессионный конструкт, который содержит последовательность по одному из пп. 1 и 2 в функциональном сцеплении с одной или более последовательностью, используемой для направления экспрессии полипептида, обладающего фосфолипазной активностью в подходящей клетке-хозяине, где последовательность, используемая для направления экспрессии полипептида, является предпочтительно промотором, выбранным из промотора глюкоамилазы или промотора α-амилазы рода Aspergillus, промотора целлюлазы (целлобиогидролазы) рода Trichoderma, промотора фермента в гликолитическом метаболическом пути, такого как фосфоглицераткиназа или глицеральдегид-3-фосфатдегидрогеназа, промотора ксиланазы или промотора энолазы, и который необязательно дополнительно содержит секретирующую лидерную последовательность.
4. Рекомбинантная клетка-хозяин, экспрессирующая полипептид, обладающий фосфолипазной активностью, по существу без липазной активности, характеризующаяся тем, что она трансформирована посредством экспрессионного конструкта по пункту 3, который содержит нуклеиновую кислоту по п. 1 или 2.
5. Рекомбинантная клетка-хозяин по п. 4, характеризующаяся тем, что она происходит из грибной клетки рода Aspergillus, Rhizopus, Trichoderma, Neurospora, Mucor или Penicillium или из дрожжевой клетки рода Kluyveromyces, Saccharomyces, Schizosaccharomyces, Trichosporon, Schwanniomyces, Hansenula или Pichia.
6. Плазмида pPL3949-Topo2.5 для депонирования фосфолипазного гена из Aspergillus fumigates RH3949 (SEQ ID NO: 1), причем плазмида имеет рестрикционную карту в соответствии с Фигурой 7 и депонирована под номером доступа DSM 22741.
7. Полипептид, обладающий фосфолипазной активностью, по существу без липазной активности, выбранный из
a) полипептида, который кодируется кодирующей частью последовательности ДНК по любому из пп. 1 и 2,
b) полипептида, имеющего последовательность в соответствии с SEQ ID NO: 2,
c) полипептида, имеющего последовательность, которая обладает идентичностью по меньшей мере 90% с аминокислотами от 1 до 299 из SEQ ID NO: 2,
d) полипептида, который кодируется нуклеотидной последовательностью, которая гибридизуется в жестких условиях с
(i) нуклеотидами от 55 до 1106 из SEQ ID NO: 1,
(ii) последовательностью кДНК, содержащейся в нуклеотидах от 55 до 1106 из SEQ ID NO: 1, или
(iii) комплементарной нитью (i) или (ii).
8. Полипептид по п. 7, характеризующийся тем, что
- имеет молекулярную массу в интервале от 28 до 30 кДа;
- может гидролизовать по меньшей мере одну из двух жирных кислот лецитина
- по существу не проявляет липазной активности;
- имеет устойчивость к высокой температуре;
- может быть выделен из организма рода Aspergillus.
9. Полипептид по п. 8, характеризующийся тем, что он может быть выделен из Aspergillus fumigatus.
10. Полипептид по п. 9, характеризующийся тем, что он является иммунологически реакционно-способным с антителом против последовательности SEQ ID NO: 2.
11. Полипептид по любому одному из пп. 8-10, характеризующийся тем, что он содержит последовательность в соответствии с SEQ ID NO: 2.
12. Фосфолипазная композиция, характеризующаяся тем, что она содержит полипептид по любому из пп. 7-11 вместе с вспомогательными веществами.
13. Применение полипептида по любому из пп. 7-11 или фосфолипазной композиции по п. 12 для дегуммирования растительного масла.
14. Применение полипептида по любому из пп. 7-11 или фосфолипазной композиции по п. 12 для приготовления теста и/или хлебобулочных изделий.
15. Применение полипептида по любому из пп. 7-11 или фосфолипазной композиции по п. 12 для приготовления молочных продуктов.
16. Применение полипептида по любому из пп. 7-11 или фосфолипазной композиции по п. 12 в качестве добавки к корму для животных.
17. Применение полипептида по любому из пп. 7-11 или фосфолипазной композиции по п. 12 для обработки текстильных сырьевых материалов.
18. Способ получения полипептида, обладающего фосфолипазной активностью по любому из пп. 7-11, характеризующийся тем, что клетку-хозяина по любому из
пп. 4 или 5 выращивают в условиях, которые поддерживают экспрессию полипептида, и затем экстрагируют полипептид.
19. Способ дегуммирования растительного масла, характеризующийся тем, что
a) растительное масло, которое должно быть обработано, необязательно предварительно обрабатывают для удаления смолистых веществ, например путем влажного рафинирования,
b) водный раствор, содержащий полипептид по любому из пп. 7-11 или фосфолипазную композицию по п. 12, добавляют к маслу, необязательно предварительно обработанному согласно а),
c) реакционную партию в соответствии с b) нагревают от 1 до 12 ч до температуры между 30°C и 70°C и
d) разделяют водную и масляную фазы.
| WO 2008040466 A1, 10.04.2008.NCBI Reference Sequence: XP_748138.1, 19.02.2008, найдено в Интернет по адресу http://www.ncbi.nlm.nih.gov./protein/XP_748138.EA 8607 В1, 29.06.2007 | |||
| Приспособление для регулирования подачи топлива и воздуха в топках с цепной колосниковой решеткой | 1928 |
|
SU10903A1 |

