Область техники, к которой относится изобретение
Изобретение в основном относится к новым и усовершенствованным нуклеотидным последовательностям, кодирующим полипептид или белок, обладающие фитазной ферментативной активностью, и способам эффективной экспрессии таких ферментов в клетке-хозяине.
В первом аспекте данное изобретение относится к выделенной молекуле нуклеиновой кислоты i) содержащей нуклеотидную последовательность в соответствии с SEQ ID NO: 1, кодирующую белок с фитазной активностью, или ii) имеющей процентное сходство по меньшей мере 78% с нуклеотидной последовательностью в соответствии с SEQ ID NO: 1, кодирующей белок с фитазной активностью.
Настоящее изобретение также относится к экспрессирующей конструкции или к вектору, включающим вышеупомянутые выделенные молекулы нуклеиновой кислоты. Настоящее изобретение дополнительно относится к способу производства белка, имеющего фитазную ферментативную активность, который включает этапы:
a) введение в клетку-хозяина вектора: i) содержащего элементы, регулирующие экспрессию, которые функционируют в клетке-хозяине; и ii) функционально связанного с молекулой нуклеиновой кислоты, описанной в настоящем документе;
b) культивирование клеток-хозяев, полученных на этапе a) в условиях, подходящих для экспрессии белка, и, при необходимости,
с) выделение белка, произведенного на этапе b), из культуры клеток.
Настоящее изобретение дополнительно относится к белкам, полученным способом соответственно изобретению, и к его применению для изготовления добавок в корм для животных.
УРОВЕНЬ ТЕХНИКИ
Изобретение, в основном, относится к новым молекулам нуклеиновой кислоты, кодирующим полипептид или белок, имеющий фитазную ферментативную активность, и способу эффективного производства этого белка с фитазной активностью в клетке-хозяине. Изобретение также относится к полипептиду или белку, полученному способом в соответствии с настоящим изобретением, и его применению в качестве активного ингредиента изготовления добавок в корм для животных и/или для высвобождения фосфата органических веществ почвы.
Элементарный или чистый фосфор (P) достаточно редок в природе, и обычно содержится в молекулах, включающих фосфатную группу (PО4³-), присоединенную к органической группе, имеющей в основе углерод животных и растительных тканей.
Однако фосфор (P) является важным питательным веществом для растений и животных. Фосфор представляет собой важный макроэлемент растений, составляющий около 0.2% сухого веса растений. Это компонент таких ключевых молекул, как нуклеиновые кислоты, фосфолипиды и АТФ, и, следовательно, растения не могут расти без надежного снабжения этим элементом. Неорганический фосфор является единственной формой фосфора, способной поглощаться и усваиваться растениями. Из-за отсутствия в почве неорганического фосфора, фермеры вынуждены обеспечить поступление макроэлемента P.
Кроме того, животным также требуется фосфор в качестве питательного вещества, которое обычно предоставляется либо в виде неорганического фосфора, например в рационе питания, либо в виде продуктов деградации различных фосфор-содержащих органических ингредиентов, составляющих рацион питания.
Фитиновая кислота, или фитат, представляет собой основную форму хранения фосфора в таких культурах, как зерновые, бобовые, масличные, составляющую от 1 до 5% по массе. Фитиновая кислота, также известная как инозитгексафосфат (IP6), или ее соль "фитат", является насыщенной циклической кислотой. Кроме того, фитиновая кислота, как правило, хелатирует и таким образом делает неусвояемыми некоторые важные микроэлементы, такие как цинк и железо, а также, в меньшей степени, такие макроэлементы, как кальций и магний. Термин "фитин" относится конкретно к кальциевой или магниевой солям фитиновой кислоты. Продукты распада фитиновой кислоты называются низшими инозитолфосфатами. Примерами являются инозитолпентафосфат (IP5), инозитолтетрафосфат (IP4) и инозитолтрифосфат (IP3). Более 2/3 фосфора в зерновых и бобовых содержится в форме фитата.
Фитат не усваивается людьми или нежвачными (моногастричными) животными, и поэтому при непосредственном употреблении в пищу он не является надлежащим источником ни инозитола, ни фосфата. В частности, фосфор и инозитол, присутствующие в форме фитата, не имеют биологически доступной формы для нежвачных животных, поскольку у последних отсутствует пищеварительный фермент фитаза, требующийся для удаления фосфата из инозитола в молекуле фитата. Фосфор, представленный в форме фитата, составляет от 20 до 40% фосфора растительного происхождения, потребляемого одножелудочными животными.
Поскольку фитат не усваивается нежвачными животными, фитат обычно проходит через желудочно-кишечный тракт, повышая количество фосфора в навозе. Избыточное содержание фосфора может привести к экологическим проблемам, таким, как эвтрофикация. Загрязнение почвы фосфором представляет собой серьезную проблему в сельском хозяйстве, особенно в районах развитого сельского хозяйства с использованием одножелудочных животных.
Кроме того, хотя некоторые микроорганизмы в микрофлоре тонкой кишки способны гидролизовать фитиновую кислоту, фосфор усваивается лишь в малых количествах. С другой стороны, избыток фитата приводит к снижению питательности, поскольку фитат способен хелатировать ионы таких металлов, как кальций, медь или цинк, которые необходимы для питания животных, что снижает его питательную ценность.
Во избежание вышеупомянутых недостатков корма животных обычно обогащаются неорганическим фосфором, что в свою очередь повышает стоимость производства животноводческой продукции. Еще один подход, который в последнее время был направлен на устранение этих недостатков, заключается в непосредственном добавлении фитазы, которая катализирует гидролиз фитата, для высвобождения органического фосфора и перевода его в доступную форму.
Фитаза бактериального происхождения в настоящее время разрешена к использованию в кормах для животных с целью повышения доступности фосфора из фитиновой кислоты. Например, применение 6-фитазы, производимой Aspergillus oryzae (DSM 14223), разрешено для кур на откорме, кур-несушек, индеек на откорме, свиней на откорме и свиноматок Регламентом (ЕС) № 255/2005 Европейской Комиссии. Эти фитазы являются крайне слабыми и нестабильными, так как имеют максимальную активность в диапазоне pH от 5.0 до 7.5, ввиду чего активность существенно снижена и ограничена из-за низких значений pH в желудке (pH 2-3). Кроме того, известно, что активность фитазы сильно ингибируется избытком субстрата (фитата) и продукта (неорганического фосфора) и высокими температурами (Power and Khon, 1993).
Синтетические фитазы представляют собой фосфомоноэстеразы, способные гидролизовать фитиновую кислоту до неорганического ортофосфата и свободного миоинозитола с низким выходом фосфорных эфиров и рядом фосфатов от пента- до монофосфата в качестве промежуточных продуктов (Nayini and Markakis, 1986; Lasztity and Lasztity, 1988; Harland and Morris, 1995). Номенклатура IUPAC-IUB (1976) признает 3-фитазу (EC 3.1.3.8), выделенную из животных и микроорганизмов (Reddy and col., 1982; Lasztity and Lasztity, 1988).
Экзогенные фитазы были найдены в таких микроорганизмах, как грибы, в том числе дрожжи (Saccharomyces cerevisae и Aspergillus sp), а также бактерии (Bacillus subtilis, Pseudomonas). Фитазы, выделенные из Aspergillus sp, следуют определенному порядку гидролиза молекулы фитата, при котором после отщепления фосфатной группы из позиции 3 продолжается отщепление групп в следующем порядке: 4, 5, 6 и 2 (Venekamp et al., 1995). Эти ферменты обладают различной активностью при значениях pH 2.5 и pH 5.5. Активность при рН 2.5 на 40% меньше, чем при pH 5.5, что имеет важное значение с учетом того, что усвоение фосфора происходит в основном в тонком кишечнике, имеющем рН 5.5 (Power and Khon, 1993).
Фитаза, произведенная с использованием Aspergillus ficuum, является очищенным гликопротеином, ферментативная активность которого зависит от температуры и рН. Оптимальная температура составляет от 60 до 70°С. Однако при инкубации в течение 10 минут при 68°С отмечается 60%-ное снижение активности (Nasi, 1990). Термостойкие ферменты Aspergillus niger устойчивы к осаждению центрифугированием, и его активность составляет не менее 5000 ед. фитазной активности/г. (Nasi, 1990).
Патент WO 2011/141613 A2 описывает новый идентифицированный ген Serratia odorifera, который кодирует белок или полипептид, имеющий фитазную активность, процесс получения такового в клетках дрожжей, таких как Pichia pastoris, и использование этого фермента в качестве активного ингредиента в производстве добавок в корм для животных. В патенте WO 2011/141613 описывается фитаза, нечувствительная к разнице pH между желудком (рН 3.5) и кишечным трактом (pH 7.0) и сохраняющая свою активность в широком диапазоне pH. Было показано, что фитаза, описанная в WO 2011/1416, имеет улучшенную термостойкость.
Однако невысокий уровень экспрессии этой фитазы дикого типа в Serratia по-прежнему является узким местом для крупномасштабного производства белка. По-прежнему ощущается настоятельная необходимость в определении факторов и условий, способствующих улучшению и повышению эффективности экспрессии белка фитазы в клетке-хозяине. Передовые технологии по-прежнему не располагают средствами и способами, позволяющими эффективно производить больше белка с фитазной активностью в той же единице объема и за тот же промежуток времени. Таким образом, по-прежнему существует настоятельная необходимость в улучшении условий промышленного производства, с тем чтобы обеспечить эффективный и надежный способ.
Сущность изобретения
Для достижения вышеуказанной цели используются способы и средства в соответствии с настоящим изобретением, описанные и заявленные в нем. Настоящее изобретение направлено на решение этих потребностей и представляет собой новую выделенную молекулу нуклеиновой кислот из Serratia odorifera, кодирующую белок или полипептид с фитазной активностью. Настоящее изобретение дополнительно предоставляет способы эффективной экспрессии и последующей очистки отдельных генов в соответствии с изобретением в клетке-хозяине.
В первом аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, i) содержащей нуклеотидную последовательность в соответствии с SEQ ID NO: 1, кодирующую белок с фитазной активностью, или ii) имеющей процентное сходство по меньшей мере 83% с нуклеотидной последовательностью в соответствии с SEQ ID NO: 1, кодирующей белок с фитазной активностью.
Дополнительно, в более предпочтительном варианте осуществления молекула выделенной нуклеиновой кислоты имеет процентное сходство по меньшей мере 90% с последовательностью нуклеотидов в соответствии с SEQ ID NO: 1, кодирующей белок с фитазной активностью.
В дополнительном более предпочтительном варианте осуществления молекула нуклеиновой кислоты не содержит нуклеотидную последовательность, кодирующую N-концевой сигнальный пептид.
В дополнительном предпочтительном варианте осуществления выделенная молекула нуклеиновой кислоты дополнительно содержит кодон-оптимизирующие мутации, учитывающие особенности организма клетки-хозяина.
В дополнительном предпочтительном варианте осуществления организмом клетки-хозяина является Pichia pastoris.
В дополнительном предпочтительном варианте осуществления настоящего изобретения молекула нуклеиновой кислоты отличается от соответствующей кодирующей последовательности нуклеотидов дикого типа наличием не менее 50 кодон-оптимизирующих мутаций.
В дополнительном предпочтительном варианте осуществления не менее 50% кодонов соответствующей кодирующей последовательности нуклеотидов дикого типа заменены на кодоны, в наибольшей степени предпочитаемые организмом клетки-хозяина.
В одном из предпочтительных вариантах осуществления все кодоны в выделенной молекуле нуклеиновой кислоты заменены на кодоны, в наибольшей степени предпочитаемые организмом клетки-хозяина.
В дополнительных предпочтительных вариантах осуществления выделенная молекула нуклеиновой кислоты кодирует аминокислотную последовательность белка с фитазной активностью в соответствии с SEQ ID NO: 3.
Во втором аспекте настоящее изобретение относится к экспрессирующей конструкции, включающей вышеуказанную выделенную молекулу нуклеиновой кислоты, функционально связанную с элементами, регулирующими экспрессию нуклеотидной последовательности.
В дополнительном предпочтительном варианте осуществления указанной экспрессирующей конструкции элементы, регулирующие экспрессию, включают в себя промотор, функционирующий в клетке-хозяине, и, при необходимости, терминирующую последовательность.
В дополнительном предпочтительном варианте осуществления, клетка-хозяин является клеткой гриба.
В дополнительном предпочтительном варианте осуществления клетка гриба является клеткой Pichia pastoris.
В третьем аспекте изобретение относится к вектору, содержащему указанную молекулу нуклеиновой кислоты или экспрессирующую конструкцию.
В четвертом аспекте настоящее изобретение относится к способу производства белка, обладающего ферментативной активностью фитазы, включающему этапы:
a) введение в клетку-хозяина вектора, i) содержащего элементы, регулирующие экспрессию, которые функционируют в клетке-хозяине; и ii) функционально связанного в ней с молекулой нуклеиновой кислоты, которая кодирует белок с фитазной активностью, как определено в настоящем документе;
b) культивирование клеток-хозяев, полученных на этапе a) в условиях, подходящих для экспрессии белка, и, при необходимости,
с) выделение белка, произведенного на этапе b), из культуры клеток.
В дополнительном предпочтительном варианте осуществления указанной экспрессирующей конструкции элементы, регулирующие экспрессию, включают в себя промотор, функционирующий в клетке-хозяине, и, при необходимости, терминирующую последовательность.
В дополнительном предпочтительном варианте осуществления настоящего изобретения клетка-хозяин представляет собой дрожжевую клетку.
В дополнительном частном варианте осуществления дрожжевая клетка является клеткой Pichia pastoris.
В дополнительном предпочтительном варианте осуществления настоящего изобретения экспрессия белка осуществляется с использованием метанольной системы экспрессии.
В дополнительном предпочтительном варианте осуществления белок, выделенный на шаге c), обладает фитазной активностью по крайней мере 1000 единиц/л.
В дополнительном предпочтительном варианте осуществления указанный шаг выделения включает в себя i) отделение секретируемого белка от компонентов среды и/или ii) отделение белка от компонентов клетки-хозяина.
В пятом аспекте нынешнее изобретение относится к белку, полученному способом в соответствии с изобретением.
В дополнительном аспекте настоящее изобретение относится к применению белка в соответствии с настоящим изобретением или полученного способом, описанным в настоящем документе, для производства добавок в корм для животных.
В еще одном аспекте настоящее изобретение относится к добавкам в корм для животных, содержащим описанный в настоящем документе белок, или содержащим белок, произведенный способом, описанным в настоящем документе.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Следующие рисунки иллюстрируют осуществление изобретения и не предусматривают ограничения сферы применения изобретения, которая определяется формулой изобретения.
Фиг.1 Определение молекулярной массы белка фитазы AppAs-r Оптима путем денатурирующего электрофореза в акриламидном геле (SDS-PAGE): Среда культивирования без метанольной индукции (Дорожка 1); экспрессия фитазы AppAs-r в среде культивирования с метанолом (Дорожка 2); экспрессия фитазы AppAs-r Оптима в среде культивирования с метанолом (Дорожка 3); маркер молекулярной массы (Дорожка 4)
Фиг.2 Определение ферментативной активности (%) белка фитазы AppAs-r Оптима в зависимости от температуры (°С) и его термостойкость
Фиг.3 Определение ферментативной активности белка фитазы AppAs-r Оптима в зависимости от pH
Фиг.4 Действие протеаз пепсина (закрашенные ромбы) и трипсина (закрашенные квадраты) на ферментативную активность AppAs-r Оптима
Фиг.5 Поддержание фитазной активности AppAs-r Оптима во времени
Фиг.6А Оригинальная нуклеотидная последовательность фитазы Serratia odorifera дикого типа (SEQ ID NO: 2) и последовательность нуклеотидов Serratia odorifera без последовательности нуклеотидов, кодирующей N-концевой сигнальный пептид (SEQ ID NO: 8). Нуклеотидная последовательность, кодирующая N-концевой сигнальный пептид (SEQ ID NO: 4) выделена серым цветом
Фиг.6B Нуклеотидная последовательность оптимизированной/усовершенствованной фитазы из Serratia odorifera (AppAs-r Оптима/фитаза Оптима) (SEQ ID NO: 1)
Фиг.6С Аминокислотная последовательность белка-фитазы AppAs-r Оптима из Serratia odorifera (SEQ ID NO: 3)
Фиг.6D Сравнение аминокислотных последовательностей, транслируемых с нуклеотидных последовательностей на Фиг.6А (верхняя последовательность) и Фиг.6B (нижняя последовательность)
Фиг.6E: Сравнение нуклеотидной последовательности гена на Фиг.6А (верхняя последовательность) и нуклеотидной последовательности гена на Фиг.6B (нижняя последовательность)
Фиг.7 Профили экспрессии фитазы дикого типа и улучшенной фитазы (фитазы Оптима)
Фиг.8 График, отображающий временную (часы) динамику производства фитазы (единицы/л) дикого типа по сравнению с улучшенной фитазой (фитазой Оптима)
ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Изобретение в основном относится к новым и усовершенствованным нуклеотидным последовательностям, кодирующим полипептид или белок, обладающие фитазной ферментативной активностью, и способам эффективной экспрессии таких ферментов в клетке-хозяине.
Хотя настоящее изобретение будет описано в отношении конкретных вариантов осуществления, это описание не должно толковаться в ограничительном смысле.
Перед подробным описанием примеров вариантов осуществления настоящего изобретения приводятся определения, имеющие важное значение для понимания настоящего изобретения.
В данной спецификации, а также в прилагаемой формуле изобретения, формы единственного числа также подразумевают соответствующие формы множественного числа, если контекст не подразумевает иного явным образом.
В контексте настоящего изобретения термины "примерно" и "приблизительно" отмечают интервал точности, который должен пониматься специалистом в данной области техники как интервал, в пределах которого гарантируется технический эффект данной характеристики. Этот термин обычно указывает на отклонение от указанного числового значения на ± 20%, предпочтительно на ± 15%, более предпочтительно на ± 10% и еще более предпочтительно на ± 5%.
Следует понимать, что термин "содержащий" не применяется в ограничительном смысле. Для целей настоящего изобретения термин «состоящий из» считается предпочтительным вариантом термина «в составе». Если далее в настоящем документе группа определена как содержащая по крайней мере определенное число вариантов осуществления, то это также относится к группе, которая предпочтительно состоит только из этих вариантов осуществлений.
Кроме того, термины «первый», «второй», «третий», или «(a)», «(b)», «(c)», «(d)» и т.д., и подобные им в описании изобретения и в формуле изобретения используются для проведения различия между схожими элементами и не обязательно подразумевают последовательный или хронологический порядок. Следует понимать, что используемые таким образом термины являются взаимозаменяемыми в соответствующих обстоятельствах и что осуществление изобретения, описанного в настоящем документе, возможно и в других последовательностях, нежели описано или проиллюстрировано в настоящем документе.
В случае, если термины «первый», «второй», «третий», или «(a)», «(b)», «(c)», «(d)», «i», «ii» и им подобные относятся к этапам способа или осуществлению процедуры, между этапами и интервалами между ними не подразумевается согласования во времени, т.е. этапы могут выполняться одновременно или с интервалами времени порядка секунд, минут, часов, дней, недель, месяцев или даже лет между этапами, если иное не указано выше или ниже в настоящем документе.
Следует понимать, что настоящее изобретение не ограничивается конкретной методологией, протоколами, реагентами и так далее, описанными в настоящем документе, поскольку они могут варьировать. Следует также понимать, что используемая в настоящем документе терминология предназначена только для описания конкретных вариантов осуществления и не подразумевает ограничения сферы применения настоящего изобретения, которая будет ограничена только формулой изобретения. Если не определено иное, все технические и научные термины, используемые в настоящем документе, обозначают то же, что обычно понимается под ними специалистом в данной области техники.
Для достижения вышеуказанной цели используются способы и средства в соответствии с настоящим изобретением, описанные и заявленные в нем. Настоящее изобретение направлено на решение этих потребностей и предоставляет средства и способы получения выделенного гена Serratia odorifera, кодирующего белок или полипептид с фитазной активностью. Настоящее изобретение дополнительно предоставляет способы эффективной экспрессии и последующей очистки отдельных генов в соответствии с изобретением в клетке-хозяине.
Как принято в настоящем документе, термин «ген» относится к определенному участку, который расположен в геноме и который может включать в себя регуляторные последовательности нуклеотидов, ответственные за контроль экспрессии, т.е. транскрипцию и трансляцию кодирующего фрагмента. Ген может также включать в себя иные 5'- и 3'-концевые нетранслируемые последовательности и терминирующие последовательности. Дополнительно могут присутствовать иные элементы, например, интроны.
"Выделенная молекула нуклеиновой кислоты", кодирующая белок с фитазной активностью, в соответствии с настоящим изобретением относится к молекуле нуклеиновой кислоты, идентифицированной и очищенной по меньшей мере от одной примесной молекулы нуклеиновой кислоты, с которой она обычно ассоциирована в естественном источнике полипептид-кодирующей нуклеиновой кислоты. Выделенная полипептид-кодирующая молекула нуклеиновой кислоты имеет формы и параметры, отличные от тех, которые присущи ей в в природе.
Поэтому выделенные молекулы нуклеиновой кислоты можно отличить от других специфических полипептид-кодирующих молекул нуклеиновой кислоты, существующих в клетках естественным образом. Однако понятие выделенной молекулы нуклеиновой кислоты включает в себя полипептид-кодирующие молекулы нуклеиновой кислоты, содержащиеся в клетках, которые обычно экспрессируют данный полипептид, если, например, молекула нуклеиновой кислоты находится в локусе хромосомы, отличном от локуса, занимаемого ею в естественных клетках.
Как принято в настоящем документе, термин "мутация" означает любое изменение в полипептиде или молекуле нуклеиновой кислоты относительно полипептидов или нуклеиновых кислот дикого типа, изменением которых получен "мутант", и может, например, подразумевать однократные или множественные замены аминокислот или нуклеотидов, или замены как нуклеотидов, так и аминокислот, включая точечные мутации, нулевые мутации, мутации, приводящие к сдвигу рамки считывания, а также может подразумевать делеции, или вставки, или замены одной или нескольких нуклеиновых кислот или аминокислот, которые могут включать в себя нуклеотиды или аминокислоты или их аналоги, встречающиеся и не встречающиеся естественным образом.
Термины "нуклеиновая кислота" или "молекула нуклеиновой кислоты", как принято в настоящем документе, в основном относятся к дезоксирибонуклеотидам и рибонуклеотидам, а также их полимерам в форме одинарной, двойной или тройной спирали. Под этим термином могут подразумеваться нуклеиновые кислоты, содержащие известные аналоги естественных нуклеотидов, имеющие параметры химической связи, сходные с референсной нуклеиновой кислотой. Конкретная последовательность нуклеиновой кислоты может также неявно подразумевать консервативно модифицированные варианты (например, вырожденные замены кодонов) и комплементарные последовательности. Термины "нуклеиновая кислота", "последовательность нуклеиновой кислоты" или "полинуклеотид", могут также быть использованы взаимозаменяемо с геном, кДНК и мРНК, закодированными в гене.
Термины "изобретенная(ые) нуклеиновая(ые) кислота(ы)" или "изобретенная(ые) молекула(ы) нуклеиновой кислоты", используемые в настоящем документе, определяют последовательность нуклеотидов, которая может использоваться как таковая, или в комбинации со способами, описанными в настоящем документе. Эти термины относятся ко всей кодирующей последовательности белка или полипептида из Serratia odorifera с фитазной активностью в соответствии с SEQ ID NO: 2, указанной в настоящем документе. Кроме того, эти термины также обозначают нуклеиновые кислоты, кодирующие функциональные фрагменты белка, векторы, содержащие кодирующие последовательности или функциональные фрагменты вышеуказанных белков, а также производные от нуклеиновых кислот, указанных в настоящем документе, которые имеют модификации, т.е. делеции, вставки, инверсии, и т.п., одного или нескольких нуклеотидов, например, от 1 до 50, от 1 до 40, от 1 до 30, от 1 до 20, от 1 до 10, от 1 до 5, от 1 до 3, 2 или 1 нуклеотид(ов), и которые тем не менее кодируют полипептиды, имеющие фитазную активность, сравнимую с активностью фитазы Serratia odorifera в соответствии с SEQ ID NO: 2.
Функциональные производные изобретенной нуклеиновой кислоты кодируют фитазные полипептиды, которые имеют фитазную активность не менее 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или более по сравнению с полипептидом дикого типа.
Кроме того, как указано выше, "изобретенная(ые) нуклеиновая(ые) кислота(ы)" или "изобретенная(ые) молекула(ы) нуклеиновой кислоты", определяют также векторы, содержащие нуклеиновые кислоты, описанные в настоящем документе. Эти векторы могут содержать регуляторные последовательности, обеспечивающие эффективную транскрипцию и трансляцию описанных в настоящем документе нуклеиновых кислот.
Термины «полипептид», «пептид» и «белок» могут также обозначать полимеры, содержащие такие модификации, как гликозилирование, присоединение липидов, сульфатирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и ADP-рибозилирование, но не ограничивающиеся ими.
Термин "выделенные полипептид", примененный по отношению к различным полипептидам, описанным в настоящем документе, означает полипептид, идентифицированный и отделенный от примесей и/или выделенный из компонента его естественной среды. Как правило, однако, очистка выделенного полипептида включает по меньшей мере один этап.
В первом аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, i) включающей нуклеотидную последовательность в соответствии с SEQ ID NO: 1, кодирующую белок с фитазной активностью, или ii) имеющей процентное сходство по меньшей мере 78% с нуклеотидной последовательностью в соответствии с SEQ ID NO: 1, кодирующей белок с фитазной активностью.
Изобретение представляет собой выделенную, синтетическую или рекомбинантную молекулу нуклеиновой кислоты, включающую (a) нуклеиновую кислоту (нуклеотид), кодирующую по меньшей мере один полипептид, имеющий фитазную активность, причем нуклеиновая кислота включает последовательность, имеющую процентное сходство по меньшей мере 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более, или полное (100%), с нуклеотидной последовательностью (полинуклеотидом) в соответствии с SEQ ID NO: 1. В дополнительном предпочтительном варианте выделенная молекула нуклеиновой кислоты имеет процентное сходство по меньшей мере 90% с последовательностью нуклеотидов в соответствии с SEQ ID NO: 1.
SEQ ID NO: 1 относится к нуклеотидной последовательности молекулы нуклеиновой кислоты, кодирующей белок или полипептид из Serratia odorifera, обладающий фитазной активностью. Последовательность нуклеотидов этой усовершенствованной или оптимизированной нуклеотидной последовательности фитазы в настоящем документе обозначается как ''AppAs-r Оптима'' или ''фитаза Оптима'' (SEQ ID NO: 1) и показана на Фиг.6B:
SEQ ID NO: 1
gagcctagatacgttttggaaaaggtggttgaggtctctcgacacggcgttagaccaccaacctcaggtaacagacaggctatgcaagcgggaaccggaagggagtggccacaatggctgactcgtgacggagaactaactggacacggttatgcagccgcaacacttaaaggaagatacgaagccgattattatagacgtcagggcctattggcaaacggttgtccatctgctggggctgtttacgtctgggctagtcctctacaaagaacaagggccaccgcacaggctttgatggacggtgcatttcccggatgcggggtcgccattcatgctgcagccactgaacaagatcccttgtttcaagcagataagatgggtcttgttccactcgatgccgaaagggctagaactgcaataaggcaggcaatgggtggctcagccgagcaagtgaagacacgttttagtgctgacattaggcgtctgcaagctgctgtttgtttgcctcaacaggcttgtcctgcctttgaacaaccttgggaaattactcaagagcatgacggtagattcagcatcaatggtctaggaacattgtccaatatggctgaatctattagacttgcctactctgaaaaccagccaacggctcaagtcgcatttggacacggtgtcaacgcatccgccgtcgctcctttgttaccactgctaaccgccagatatgactttaccaatgatgtgccctatatcgctcaaagaggtggctccgttctgttaaaccaaattgctttggctcttgccgcagataggacttctgccggggcgccacctgctgctcgttggttgttatttgtcgctcatgacaccaatatcgcttatttaagaactctgcttggtttttcttggcaacagggactttacccaagaggtaatattccccctgctggaagtttggttttcgaaagatggcgtgatagacaaacaggtcaaaggttcttacgtctgtacttccaggctcaatcgttggatcaaatcagacagttgtcaccactttctacattatccccacctttaaaaaccgagttctctcgtcctggttgcaggcagttgtcacttggcgttctctgtccctggactgagtccatgcaaagaatgagagctgctatcgacccaactgcgttgcctacagtgcagtacagaccataa
Одним из аспектов настоящего изобретения является определение и введение подходящих мутаций в последовательность гена дикого типа Serratia odorifera, с тем чтобы усилить и улучшить экспрессию белка в клетке-хозяине. Неизмененная или немодифицированная нуклеотидная последовательность дикого типа описана в заявке WO 2011/141613 A2 (включенной посредством ссылки) и в настоящем документе обозначается как SEQ ID NO: 2 (см. Фиг.6А):
SEQ ID NO: 2
atgttgctattgcaaaaggactggtcgcgtctgttatttgccgtcacgctgggtatgatttccagcgtagcccaggctgagccgcgctacgtattggaaaaggtggttgaggtcagccgccacggcgtacgcccgccgacctcaggcaaccggcaggcgatgcaggcgggaaccggccgagagtggccacaatggctgacgcgcgacggcgaactcactggccacggttatgccgccgccacgctgaaaggacgctatgaagccgactattatcgccgtcagggcctattggccaacggctgtccgagcgcgggggcggtgtatgtctgggccagtccgctacagcgcacgcgagccaccgcacaggcgttgatggacggcgcatttcccggctgcggggtcgccattcatgcggccgccaccgaacaggaccccctgtttcaggcagataaaatgggcctggtgccgctcgatgccgaacgggctcgcacggcaataaggcaggcaatgggcggcagcgccgagcaggtgaaaacgcgctttagcgctgacattcggcgtctgcaagcggcggtctgcctgccgcaacaggcttgcccggcctttgaacaaccgtgggaaatcactcaggagcacgacggccgcttcagcatcaacggtctggggacgttgtccaacatggcggaaagcattcgcctggcctacagcgaaaaccagccgacggcgcaggtcgcctttggccacggtgtcaacgcatcggccgtcgcgccgttgctgcccctgctcaccgcccgctatgactttaccaatgacgtgccctatatcgcgcaacgcggtggctcggtgctgttaaaccaaatcgcgctggcgctggccgccgatcgaacctctgccggggcgccaccggcggcgcgctggttgctgtttgtcgcgcatgacaccaatatcgcttatctgcgcaccctgcttggctttagctggcaacaggggctttacccacgcggcaatattcccccggctggcagtctggtattcgaacgctggcgcgatcggcaaacgggccagcgcttcctgcgtctgtacttccaggcgcaatcgctggatcaaatccgccagttgtcaccgctgagcacgctgtcgccaccgttaaaaaccgagttcagccgtcctggctgccggcagttgtcactgggcgtactctgtccctggactgagtcgatgcaacggatgcgcgcggctatcgacccgacggcgctgcctacggtgcagtaccggccataa
Оптимизированная нуклеотидная последовательность фитазы Serratia odorifera в соответствии с SEQ ID NO: 1 отличается от нуклеотидной последовательности дикого типа в соответствии с SEQ ID NO: 2 наличием 211 точечных мутаций или модификаций в нуклеотидной последовательности.
Оптимизированная нуклеотидная последовательность фитазы Serratia odorifera в соответствии с SEQ ID NO: 1 не включает нуклеотидную последовательность, кодирующую N-концевой сигнальный пептид в последовательности дикого типа в соответствии с SEQ ID NO: 4. Последовательность сигнального пептида в вышеуказанной нуклеотидной последовательности фитазы в соответствии с SEQ ID NO: 2 выделена серым цветом.
Кодирующая последовательность дикого типа без последовательности сигнального пептида в настоящем документе обозачается как SEQ ID NO: 8.
Сравнение последовательностей SEQ ID NO: 1 с SEQ ID NO: 2 показано на Фиг.6Е. Каждое несоответствие в выравнивании последовательностей соответствует мутированной или модифицированной позиции, в которой нуклеотид по отношению к исходной последовательности заменен на другой нуклеотид.
Согласно выравниванию последовательностей, представленному на Фиг.6, нуклеотидная последовательность оптимизированной фитазы из Serratia odorifera в соответствии с SEQ ID NO: 1 отличается от последовательности нуклеотидов дикого типа в соответствии с SEQ ID NO: 8 наличием 211 точечных мутаций или модификаций в последовательности нуклеотидов.
Экспрессия последовательности нуклеотидов в соответствии с SEQ ID NO: 1 приводит к полипептидной последовательности фитазы из Serratia odorifera, как показано на Фиг.6С (SEQ ID NO: 3). SEQ ID NO: 3 соответствует полипептидной последовательности зрелого белка фитазы. Как видно из выравнивания последовательностей на Фиг.6С, транслируемые полипептидные последовательности SEQ ID NO: 1 и SEQ ID NO: 2 на 100% сходны, т.е. ни одна из 211 мутаций, содержащихся в SEQ ID NO: 1, не привела к замене одной из закодированных аминокислот.
Предпочтительно, термин «модификация» или «мутация» по смыслу нынешнего изобретения относится к изменению нуклеотидной позиции в последовательности дикого типа, которое приводит не к замене закодированной аминокислоты, а к изменению кодона данной аминокислоты, т.е., к кодон-подстраивающим или к кодон-оптимизирующим нуклеотидным заменам.
Как будет отмечено, кодон-оптимизация в основном учитывает частоту использования кодона в конкретном организме-хозяине, в котором данная нуклеотидная последовательность экспрессируется, т.е. транслируется в соответствующий белок. Кодон-оптимизация нуклеиновой кислоты, кодирующей фитазный белок Serratia odorifera, описанная в настоящем документе, учитывает относительные частоты встречаемости кодонов, оптимизированное содержание GC и нестабильность мРНК.
В основном, «процентное сходство» (в %) относится к количественному измерению сходства между двумя последовательностями (ДНК, аминокислот или иных). Ожидается, что близкородственные виды имеют более высокое процентное сходство для данной последовательности, чем более отдаленные виды, и, таким образом, процентное сходство в известной степени отражает родство. Процентное сходство вычисляется путем умножения числа совпадений в паре последовательностей на 100 и деления на длину выровненной области, включая пропуски. При оценке сходства подсчитываются только идеальные совпадения, и не учитывается степень сходства аминокислот друг с другом. Для расчета в длину включаются только пропуски внутри последовательности, но не пропуски на концах последовательности.
Процентное сходство=(Число совпадений x 100)/Длина выровненной области (с пропусками)
"процентное (%) сходство нуклеотидной последовательности" по отношению к последовательности нуклеиновых кислот в соответствии с SEQ ID NO: 1, кодирующей белок с фитазной активностью, приводимое в настоящем документе, определяется как процентная доля нуклеотидов в последовательности-кандидате, идентичных нуклеотидам в нуклеотидной последовательности в соответствии с SEQ ID NO: 1, после выравнивания последовательностей и введения пропусков, если это необходимо, для достижения максимального значения процентного сходства. Выравнивание с целью определения процентного сходства нуклеотидных последовательностей может быть достигнуто различными путями в пределах уровня техники с использованием общедоступного программного обеспечения, такого как программы BLAST, BLAST-2, ALIGN или Megalign (DNASTAR).
Например, процентное сходство нуклеотидной последовательности может быть определено с помощью программы сравнения последовательностей NCBI-BLAST2 (Altschul et al., Nucleic Acids Res. 25: 3389-3402 (1997). Программа сравнения последовательностей NCBI-BLAST2 может быть загружена с сайта http://www.NCBI.nlm.nih.gov или иным образом получена из Национального института здравоохранения, Бетесда, Мэриленд. NCBI-BLAST2 использует несколько параметров поиска, причем для всех параметров поиска заданы значения по умолчанию, включая, например, unmask=yes, strand=all, expected occurrences=10, minimum low complexity length=15/5, multi-pass e-value=0.01, constant for multi-pass=25, dropoff for final gapped alignment=25 and scoring matrix=BLOSUM62.
В случае, если длина данной нуклеотидной последовательности, т.е. последовательности AppAs-r Оптима (SEQ ID NO: 1) не равна длине нуклеотидной последовательности сравнения, процентное сходство вычисляется следующим образом:
1. Последовательность AppAs-r Оптима (SEQ ID NO: 1) короче, чем ДНК сравнения:
AppAs-r Оптима NNNNNNNNNNNNNN (длина=14 нуклеотидов)
ДНК сравнения NNNNNNXXXXXXXXXX (длина=16 нуклеотидов)
(Длины последовательностей приведены для примера)
Процентное сходство нуклеотидных последовательностей=(число совпадающих нуклеотидов между двумя последовательностями нуклеиновых кислот, определенное, например, программой NCBI-BLAST2) x 100, разделенное на (общее количество нуклеотидов в нуклеотидной последовательности AppAs-r Оптима)=6×100/14=42.9%
2. Последовательность AppAs-r Оптима (SEQ ID NO: 1) длиннее ДНК сравнения:
AppAs-r Оптима NNNNNNNNNNNNNN (длина=12 нуклеотидов)
ДНК сравнения NNNNNNXXXYY (длина=9 нуклеотидов)
(Длины последовательностей приведены для примера)
Процентное сходство нуклеотидных последовательностей=(число совпадающих нуклеотидов между двумя последовательностями нуклеиновых кислот, определенное, например, программой NCBI-BLAST2) x 100, разделенное на (общее количество нуклеотидов в нуклеотидной последовательности AppAs-r Оптима)=4×100/12=33.3%
Таким образом, длина референсной последовательности (нуклеотидная последовательность AppAs-r Оптима) определяет длину выровненного участка. Следует отметить, что этот принцип применим и для сравнения полипептидных последовательностей.
Выражения "белок с фитазной активностью" или "фитаза", используемые в настоящем документе, относятся к мезо-инозитгексафосфат-фосфогидролазам, катализирующим диссоциацию фосфатных групп от фитиновой кислоты (IP6), или фитата. Результатом этой ферментативной реакции является свободная фосфатная группа и эфир инозитолфосфата (IP5-IP1). В основном, фитазы можно классифицировать на две группы: бактериальные фитазы или фитазы грибов (E.C. 3.1.3.8), называемые 3-фитазами, которые начинают гидролиз с фосфорной группы, находящейся в положении C1 или C3 инозитольного кольца, и фитазы растений (E.C. 3.1.3.26), или 6-фитазы, которые начинают гидролиз, предпочтительно, с фосфатной группы, расположенной в положении C6 инозитольного кольца.
Соответственно "фитазная активность", используемая в настоящем документе, означает ферментативную реакцию гидролиза фосфатных групп от фитиновой кислоты (IP6), или фитата. В одном из вариантов осуществления ферментативная фитазная активность является фитазной активностью, описанной для белка Serratia odorifera в соответствии с SEQ ID NO: 2.
Настоящее изобретение также предусматривает измерение фитазной активности. Например, фитазная активность может быть измерена с помощью анализа, описанного в заявке WO 2011/141613 A2 или описанного в настоящем документе в Примере 3, в котором специфическая активность оценивалась путем измерения концентрации свободного фосфата или фосфора, отщепленного фитазой, с использованием молибдат-ванадатного способа.
Этот метод основан на стандартном способе "4500-P C" Американской ассоциации охраны общественного здоровья. В кислотах ортофосфат-ионы реагируют с молибдатом аммония и ванадатом аммония, образуя желтый фосфорованадомолибдат аммония, который может быть фотометрически проанализирован при длине волны 415 нм. Интенсивность желтого цвета пропорциональна концентрации фосфатов. Однако нынешнее изобретение также предусматривает другие способы или анализы для измерения или определения фитазной активности, рутинно выполняемые специалистом в данной области техники. Альтернативным способом является способ молибденовой сини. В кислой среде ортофосфаты связываются с молибдатом аммония, образуя фосфорномолибденовую кислоту. Под действием восстанавливающего агента она формирует фосфорномолибденовую синь. Фотометрическое измерение интенсивности красителя может быть произведено на 880 Нм.
В конкретном аспекте изобретение относится к нуклеотидной последовательности, в которой отсутствует N-концевая сигнальная последовательность и/или начальный метионин, и которая закодирована последовательностью нуклеотидов, кодирующей аминокислотную последовательность, описанную выше.
Последовательность N-терминального сигнального пептида гена Serratia odorifera дикого типа определяется нуклеотидной последовательностью в соответствии с SEQ ID NO: 4, и аминокислотной последовательностью в соответствии с SEQ ID NO: 5.
SEQ ID NO: 4:
atgttgctattgcaaaaggactggtcgcgtctgttatttgccgtcacgctgggtatgatttccagcgtagcccaggct
SEQ ID NO: 5:
Mlllqkdwsrllfavtlgmissvaqa
Авторы настоящего изобретения наблюдали, что экспрессия полноразмерной версии гена фитазы дикого типа из Serratia odorifera, показанной на Фиг.6A (SEQ ID NO: 2), содержащего сигнальный пептид, состоящий из 26 аминокислот, приводит к неустойчивому белку, который после экспрессии быстро исчезает. Авторы настоящего изобретения также наблюдали, что пропуск 26-звенного сигнального пептида приводит к усилению экспрессии (см. Пример 1).
В конкретном варианте осуществления изобретение представляет собой оптимизированную последовательность AppAs-r для экспрессии в подходящей для клетки-хозяина системе экспрессии. В оптимизированной последовательности учитывается изменение частоты использования кодонов в организме клетки-хозяина. В одном из вариантов осуществления используются наиболее предпочтительные или наиболее частые кодоны для каждой аминокислоты в заданной нуклеотидной последовательности, кодирующей белок с фитазным действием, как описано в настоящем документе. В альтернативном варианте осуществления может быть достаточно частично оптимизированной последовательности, т.е. последовательности, в которой лишь часть кодонов нуклеотидной последовательности заменены на наиболее часто используемые кодоны. В некоторых вариантах осуществления достаточно оптимизировать лишь некоторые определенные аминокислоты в последовательности AppAs-r.
Предпочтительный вариант осуществления настоящего изобретения относится к нуклеотидной последовательности, в которой по меньшей мере 5%, 10%, 20%, 25%, 30%, 35%, 40%, 45%, 50% кодонов нуклеотидной последовательности AppAs-r дикого типа заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В более предпочтительных вариантах осуществления по меньшей мере 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58% или 59% кодонов в нуклеотидной последовательности заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В еще более предпочтительных вариантах осуществления по меньшей мере 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68% или 69% кодонов в нуклеотидной последовательности дикого типа заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В еще более предпочтительных вариантах осуществления по меньшей мере 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78% или 79% кодонов в нуклеотидной последовательности дикого типа заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В еще более предпочтительных вариантах осуществления по меньшей мере 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88% или 89% кодонов в нуклеотидной последовательности заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В еще более предпочтительных вариантах осуществления по меньшей мере 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% кодонов в нуклеотидной последовательности дикого типа заменены на кодоны, наиболее часто используемые организмом клетки-хозяина.
В предпочтительных вариантах осуществления выделенная молекула нуклеиновой кислоты отличается от соответствующей нуклеотидной последовательности фитазы дикого типа в соответствии с SEQ ID NO: 8 наличием кодон-оптимизирующих мутаций.
Термин "кодон-оптимизирующие мутации" в соответствии с настоящим изобретением относится к молчащим мутациям кодирующей последовательности, которые не приводят к изменению аминокислотной последовательности. Использование новой нуклеотидной последовательности фитазы, включающей кодон-оптимизирующие мутации в соответствии с настоящим изобретением, имеет значительное техническое преимущество по сравнению с существующим уровнем техники экспрессии фитазы, состоящее в том, что улучшенная экспрессия в данном организме клетки-хозяина возможна без негативно воздействующего на фитазную активность изменения аминокислотной последовательности, таким образом, структура или функция результирующего фитазного белка сохраняется.
Дополнительно, предпочтительные варианты осуществления настоящего изобретения относятся к последовательностям нуклеотидов, в которых по меньшей мере 5%, 10%, 20%, 25%, 30%, 35%, 40%, 45%, 50% от нуклеотидной последовательности дикого типа, кодирующей фитазу, в соответствии с SEQ ID NO: 8 заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В более предпочтительных вариантах осуществления 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58% или 59% кодонов нуклеотидной последовательности дикого типа, кодирующей фитазу, в соответствии с SEQ ID NO: 8 заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В более предпочтительных вариантах осуществления 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68% или 69% кодонов нуклеотидной последовательности дикого типа, кодирующей фитазу, в соответствии с SEQ ID NO: 8 заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В более предпочтительных вариантах осуществления 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78% или 79% кодонов нуклеотидной последовательности дикого типа, кодирующей фитазу, в соответствии с SEQ ID NO: 8 заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В более предпочтительных вариантах осуществления 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88% или 89% кодонов нуклеотидной последовательности дикого типа, кодирующей фитазу, в соответствии с SEQ ID NO: 8 заменены на кодоны, наиболее часто используемые организмом клетки-хозяина. В более предпочтительных вариантах осуществления 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, или 99% кодонов нуклеотидной последовательности дикого типа, кодирующей фитазу, в соответствии с SEQ ID NO: 8 заменены на кодоны, наиболее часто используемые организмом клетки-хозяина.
В некоторых из вариантов осуществления последовательность выделенной нуклеиновой кислоты включает по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 кодон-оптимизирующих мутаций по сравнению с организмом клетки-хозяина.
В дополнительном варианте осуществления настоящего изобретения последовательность выделенной нуклеиновой кислоты включает по меньшей мере 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 кодон-оптимизирующих мутаций по сравнению с организмом клетки-хозяина.
В более предпочтительном варианте осуществления настоящего изобретения последовательность выделенной нуклеиновой кислоты включает по меньшей мере 110, 120, 130, 140, 150, 160, 170, 180, 190, 200 или 210 кодон-оптимизирующих мутаций по сравнению с организмом клетки-хозяина.
В наиболее предпочтительном варианте осуществления все кодоны последовательности выделенной нуклеиновой кислоты кодон-оптимизированы по сравнению с организмом клетки-хозяина.
В некоторых вариантах осуществления в 99, 98, 97, 96, 95, 94, 93, 92, 91 и 90% всех аминокислотных положений используется наиболее предпочтительный кодон клетки-хозяина.
В дополнительных конкретных вариантах осуществления нуклеотидная последовательность оптимизирована с учетом частоты использования кодонов в метилотрофных дрожжах Pichia pastoris. В конкретных вариантах осуществления, как далее описано в примере 5 ниже, оригинальные кодоны последовательно анализировали и заменяли на наиболее часто используемые кодоны для каждой аминокислоты в P. pastoris. Смещение частоты использования кодонов в P. pastoris было ранее описано в работах Bai et al. (Bai J., Swartz D.J., Protasevich I.I., Brouillette C.G., Harrell P.M., Hidebrandt E., Gasser B., Mattanovich D., Ward A., Chang G. and Urbatsch I.L. (2011) A gene optimization strategy that enhances production of fully functional P-glycoprotein in Pichia pastoris. PLosOne 6:1-15), Sinclair et al. (Sinclair G. and Choy F.Y.M. (2002) Synonymous codon usage bias and the expression of human glucocerebrosidase in the methylotrophic yeast Pichia pastoris. Protein Expression and Purification 26:96-105) и Huang et al. (Huang H, Yang P, Luo H, Tang H, Shao N, et al. (2008) High-level expression of a truncated 1,3-1,4-beta-D-glucanase from Fibrobacter succinogenes in Pichia pastoris by optimization of codons and fermentation. Appl Microbiol Biotechnol. 78: 95-103.).
Термины "экспрессирующая конструкция" или "экспрессирующая кассета" взаимозаменяемы и относятся к нуклеотидной последовательности, способной приводить к экспрессии определенной последовательности нуклеотидов в соответствующей клетке-хозяине, и включающей промотор, функционально связанный с интересующей последовательностью нуклеотидов, которая, в свою очередь, связана с сигналами терминации. Экспрессирующая конструкция обычно содержит последовательности, необходимые для правильной трансляции последовательности нуклеотидов. Кодирующий участок обычно кодирует интересующий белок. Экспрессирующая кассета, включающая интересующую последовательность нуклеотидов, может быть химерной, что означает, что по крайней мере один из ее компонентов является гетерологичным по крайней мере к одному из других ее компонентов. Экспрессирующая кассета может встречаться в природе, но при этом быть получена в рекомбинантной форме, подходящей для гетерологичной экспрессии. Однако, как правило, экспрессирующая кассета гетерологична по отношению к организму-хозяину, т.е. конкретная последовательность ДНК в этой экспрессирующей кассете не встречается естественным образом в клетке-хозяине и должна быть введена в клетку-хозяина или предшественника клетки-хозяина с помощью трансформации. Экспрессия нуклеотидной последовательности в экспрессирующей кассете может находиться под контролем конститутивного промотора или же индуцируемого промотора, который инициирует транскрипцию только когда клетка-хозяин подвергается действию некоторого внешнего стимула. В одном из вариантов осуществления экспрессирующий вектор включает в себя репортерный ген, который для экспрессии функционально связан с нуклеотидной последовательностью, кодирующей полипептид с фитазной активностью.
Термин "гетерологичный" в настоящем документе используется в значении "имеющий иное естественное происхождение" или обозначает неестественное состояние. Например, если в клетку-хозяина путем трансформации вводится нуклеотидная последовательность, полученная из другого организма, в особенности из другого вида, то данная нуклеотидная последовательность является гетерологичной по отношению к этой клетке-хозяину, а также по отношению к потомкам клетки-хозяина, несущим в себе эту нуклеотидную последовательность. Аналогичным образом, термин "гетерологичный" относится к нуклеотидной последовательности, полученной и вводимой в один и тот же естественный, оригинальный тип клетки, но находящейся в неестественном состоянии, например, в другом количестве копий или под контролем других регуляторных элементов.
Термин "регуляторные элементы" в основном относится к последовательностям, участвующих в обеспечении экспрессии нуклеотидной последовательности. Регуляторные элементы включают в себя, как правило, промотор, функционально связанный с интересующей нуклеотидной последовательностью и сигналами терминации. К ним также обычно относятся последовательности, необходимые для правильной трансляции нуклеотидной последовательности. "Регуляторные элементы" в случае прокариот, например, включают в себя промотор, при необходимости, операторный участок, и сайт связывания рибосомы. Эукариотические клетки, как известно, используют промотеры, полиаденилирование и энхансеры.
Термин "функционально связанные", в настоящем документе означает, что одна нуклеотидная последовательность имеет функциональное отношение к другой последовательности нуклеиновых кислот. Например, ДНК предпоследовательности или лидерного пептида секретируемого белка функционально связана с полипептидом, если она экспрессируется в виде белка-предшественника, участвующего в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, он влияет на транскрипцию последовательности; так же, сайт связывания рибосомы функционально связан с кодирующей последовательностью, если его расположение способствует трансляции. В основном, "функционально связанный" означает, что последовательности ДНК следуют друг за другом подряд, и, в случае лидерным пептидом, они следуют подряд и без смещения рамки считывания. Энхансеры не обязательно следуют подряд. Связь осуществляется посредством лигирования в удобных сайтах узнавания рестриктаз. Если таких сайтов не имеется, то, в соответствии с обычной практикой, используются синтетические олигонуклеотидные адаптеры или линкеры.
Подходящие "векторы экспрессии" или "плазмиды экспрессии" для экспрессии оптимизированного гена фитазы в соответствии с настоящим изобретением в различных системах экспрессии не выходят за пределы существующего уровня техники. Подходящие векторы экспрессии для экспрессии в дрожжах Pichia, например, включают в себя, серию векторов pPIC, но не ограничиваются ими. Эти векторы используют промотор AOX1, индуцируемый метанолом. Плазмиды экспрессии могут содержать элементы для включения инородной ДНК в геном дрожжей и сигнальную последовательность для секреции экспрессированного белка.
Оптимизированный белок фитазы в соответствии с настоящим изобретением может быть экспрессирован в системах экспрессии белка на основе любых подходящих клеток-хозяев. Рутинно используемые системы экспрессии белка имеют в своей основе бактериальные или дрожжевые клетки, клетки насекомых, зараженные бакуловирусом, клетки млекопитающих, а также нитчатых грибов, таких как гриб Myceliophthora thermophile, имеющий коммерческую значимость. Подходящие бактериальные системы включают Escherichia coli, Corynebacterium, или Pseudomonas fluorescens, но не ограничиваются ими. Эукариотические системы экспрессии включают дрожжевые системы, такие как Saccharomyces cerevisiae, Pichia pastoris или Kluyveromyces lactis. В настоящем изобретении также предусматривается использование других грибных систем экспрессии, таких как нитчатые грибы Aspergillus, Trichoderma и Myceliophthora thermophila, C1, которые недавно описывались для производства различных промышленных ферментов. Дополнительные эукариотические системы экспрессии включают клетки, зараженные бакуловирусом, например, зараженные клетки насекомых, такие как клоны Sf9, Sf21, High Five, или клетки млекопитающих, такие как HeLa и HEK 293, позволяющие также экспрессировать гликозилированные белки, которые не могут быть экспрессированы с помощью дрожжей или прокариотических клеток (например, E. coli). Это очень полезная система, позволяющая экспрессировать белки в больших количествах. Дополнительные подходящие эукариотические системы также включают в себя нелитическую систему экспрессии в клетках насекомых, представляющую собой альтернативу литическим бакуловирусным системам экспрессии, а именно, систему экспрессии, использующую непатогенный штамм простейших Leishmania tarentolae, а также растительные системы (например, табак) и клетки млекопитающих, такие как Bos primigenius (клетки быка), Mus musculus (мыши), клетки яичников китайского хомячка, эмбриональные клетки почек человека и клетки почки новорожденного хомяка.
В еще более предпочтительном варианте осуществления используется клетка гриба Pichia pastoris (см. Cregg JM, Cereghino JL, Shi J, Higgins DR (September 2000). "Recombinant protein expression in Pichia pastoris". Mol. Biotechnol. 16 (1): 23-52). Подходящие для экспрессии белка штаммы Pichia pastoris, как описано в настоящем документе, включают в себя GS 115, X33 или KH71H (Invitrogen, 1600 Faraday Avenue, Carlsbad, California, 92008, USA). В предпочтительном варианте осуществления используется штамм Pichia pastoris KH71H.
Для экспрессии новой нуклеотидной последовательности фитазы, представленной в настоящем изобретении, ген клонируется в подходящий экспрессирующий вектор, включающий в себя указанную молекулу нуклеиновых кислот или экспрессирующую конструкцию, как описано в приведенных ниже примерах.
Настоящее изобретение относится к способу производства белка, имеющего фитазную ферментативную активность, который включает следующие этапы:
a) введение в клетку-хозяина экспрессирующего вектора, i) содержащего элементы, регулирующие экспрессию и функционирующие в клетке-хозяине; и ii) функционально связанного с молекулой нуклеиновой кислоты, как описано в настоящем документе;
b) культивирование клеток-хозяев, полученных на этапе a) в условиях, подходящих для экспрессии белка, и, при необходимости,
с) выделение белка, произведенного на этапе b), из культуры клеток.
Этап внедрения экспрессирующего вектора общеизвестен под названием "трансформации", которая определяется как генетическое изменение клетки в результате прямого захвата и встраивания внешнего генетического материала (внешнего ДНК) из окружающей среды через клеточную мембрану (мембраны). Способы трансформации клетки-хозяина с помощью экспрессирующего вектора или экспрессирующей конструкции, а также оптимальные условия в основном зависят от клетки-хозяина и находятся в пределах уровня техники. В соответствии с этим, квалифицированным специалистам хорошо известны конкретные условия индукции экспрессии в клетке-хозяине и культивирования клетки-хозяина в соответствующей среде культивирования.
Термины "выделение" и "очистка" могут использоваться в настоящем документе взаимозаменяемо и относятся к процессу отделения экспрессированного полипептида от компонентов клеточной культуры с целью получить чистый или в значительной степени чистый препарат полипептида, т.е. свободный от присутствия других белков и/или липидов и/или нуклеиновых кислот и/или компонентов клетки-хозяина в культуре. По смыслу нынешнего изобретения указанный этап выделения включает i) отделение секретируемого белка от компонентов среды и/или ii) отделение белка от компонентов клетки-хозяина.
В конкретном варианте осуществления оптимизированная фитаза, описанная в настоящем изобретении, экспрессируется в дрожжах Pichia pastoris. Производство рекомбинантного белка в дрожжах Pichia pastoris имеет несколько преимуществ по сравнению с другими эукариотическими и прокариотическими системами экспрессии, а именно: высокая скорость роста, высокая плотность клеток, продуктивность в практически безбелковой среде; отсутствие заражения эндотоксинами и бактериофагами; разнообразные посттрансляционные модификации, включающие в себя фолдинг полипептидов, гликозилирование, метилирование, ацилирование, протеолитическую обработку и распределение по субклеточным компартментам; а также простая очистка от среды культивирования.
Большинство систем экспрессии с использованием P. pastoris основаны на метанол-индуцированном промоторе алкоголь-оксидазы (AOX1) (см. Romanos, M. A., Scorer, C. A., & Clare, J. J. (1992). Yeast, 8, 423-488.) После индукции метанолом доля общего растворимого белка, состоящего из алкоголь-оксидазы, может, как правило, возрасти до 30%. Широко используемые экспрессирующие векторы P. pastoris включают следующие элементы: (1) 5′-AOX1 (алкоголь-оксидазный промотор, вышележащий по отношению к интересующему гену); (2) SIG (последовательность сигнала секреции); 3) MCS (сайт множественного клонирования); 4) TT (сайт терминации транскрипции); 5) HIS4 (маркер, необходимый для распознавания гидроксигистидиназой); 6) Ampr (для отбора по ампицилиновому тесту); и 7) ColB1 (репликационный элемент для распространения плазмид в E.coli) (см. Li, P. Z., Gao, X.-G., Arellano, R. O., & Renugopalakrishnan, V. (2001). Protein Expression and Purification, 22, 369-380.)
Дополнительные подходящие коммерчески доступные векторные системы для экспрессии P. pastoris включают векторы pPICZα A, B и C (Pichia Expression Kit K1710-01 или EasySelect™ Pichia Expression Kit no. К1740-01, продаваемый Invitrogen), основанный на селекции рекомбинантного белка с помощью зеоцина (Zeocin®).
В дополнительном аспекте настоящее изобретение касается белка, полученного способом в соответствии с изобретением. Описанная в настоящем документе методология позволяет производить кодон-оптимизированный фитазный белок из Serratia odorifera в соответствии с полипептидной последовательностью SEQ ID NO: 2. Экспрессированный генный продукт сконструированного и оптимизированного гена Serratia odorifera соответствует зрелому белку, в котором отсутствуют первые 26 аминокислот, т.е. после отщепления сигнального пептида в клетке, который имеет молекулярный вес около 45-46 кДа, как видно из результатов гель-электрофореза в SDS на Фиг.1, который также показывает, что в одних и тех же условиях и при одной и той же загрузке препарата, уровень экспрессии гена AppAs-r Оптима значительно выше, чем гена AppAs-r.
Как показано в примере 5, а также на Фиг.7 и 8, использование кодон-оптимизированной последовательности SEQ ID NO: 1 приводит к усиленной экспрессии по сравнению с экспрессией последовательности фитазы соответствующего дикого типа в соответствии с SEQ ID NO: 2 или SEQ ID NO: 8. В этом контексте Пример 5 показывает, что экспрессия соответствующей последовательности фитазы дикого типа не превышает 143 единиц/литр (ед/л), в то время как уровень экспрессии фитазы Оптима может достигать 29973 единиц/литр. Пример 5 демонстрирует, что мутации, внесенные в последовательность Оптима, с учетом частоты использования кодонов в соответствующей дрожжевой клетке-хозяине Pichia pastoris в значительной степени способствует усиленной и эффективной экспрессии белка, при которой увеличивается как количество экспрессированного белка, так и скорость экспрессии. Таким образом, экспрессия оптимизированной фитазной последовательности Serratia odorifera приводит к существенному усилению экспрессии белка.
В конкретных предпочтительных вариантах осуществления экспрессия выделенной нуклеотидной последовательности осуществляется в системе экспрессии P. pastoris, как описано в настоящем документе.
В более предпочтительном варианте осуществления настоящего изобретения, экспрессия белка осуществляется с использованием метанольной системы экспрессии, как описано в примерах 1 или 5, и фитазная активность измеряется в ед/л с помощью молибдат-ванадатного способа в кислой среде (ISO 300024:2009 (E)), как описано в Примере 3.
В предпочтительных вариантах осуществления, способ в соответствии с изобретением приводит к усиленной экспрессии, при которой белок имеет фитазную активность по меньшей мере 250, 500, 750 или по меньшей мере 1000 ед/л, как определено в описанных здесь способах.
В предпочтительных вариантах осуществления, способ в соответствии с изобретением приводит к усиленной экспрессии, при которой белок имеет фитазную активность по меньшей мере 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000, 5500, 6000, 6500, 7000, 8500, 8000, 9500 или по меньшей мере 10000 ед/л, как определено в описанных здесь способах. В более предпочтительных вариантах осуществления, способ в соответствии с изобретением приводит к усиленной экспрессии, при которой белок имеет фитазную активность по меньшей мере 11000, 12000, 13000, 14000, 15000, 16000, 17000, 18000, 19000 или по меньшей мере 20000 ед/л, как определено в описанных здесь способах.
Кроме того, в Примере 3 оптимизированная фитазная последовательность Serratia odorifera была проанализирована в отношении фитазной активности в зависимости от температуры, pH, присутствия протеаз, а также сохранения фитазной активности во времени. Результаты, приведенные в Примере 3, удивительным образом показывают, что кодон-оптимизированный фитазный белок из Serratia odorifera, полученный способом в соответствии с настоящим изобретением, приводит к специфически устойчивому белку с повышенной фитазной активностью.
В настоящем изобретении также предусматривается применение белка в соответствии с настоящим изобретением или полученным способом, описанным в настоящем документе, для изготовления добавок в корм для животных.
Оптимизированный фитазный белок, экспрессированный способами, описанными в настоящем документе, имеет следующие характеристики:
a) Молекулярная масса от 45 до 47 кДа, предпочтительно приблизительно 46 кДа;
b) оптимум pH в диапазоне от 2.0 до 9, предпочтительно от 2.5 до 8, еще более предпочтительно от 3.0 до 7, и наиболее предпочтительно от 3.3 и 5.8.
c) ферментативная активность наблюдается в диапазоне от 40 до 55°С, предпочтительно при температуре 50°С;
d) высокая удельная активность, составляющая более чем 500 ед/мг, предпочтительно более 600, 700, 800, 900 или 1000 ед/мг, более предпочтительно около 1120 +/- 150 ед/мг
e) высокая устойчивость к действию протеаз, предпочтительно устойчивость к действию пепсина и трипсина
f) Изоэлектрическая точка при 9.3.
В еще одном аспекте настоящее изобретение относится к добавкам в корм для животных, содержащим белок, описанный в настоящем документе, или содержащим белок, полученный описанным в настоящем документе способом. Добавка может быть надлежащим образом изготовлена в твердом или жидком виде. Добавки могут дополнительно включать другие ферментативные препараты, обычно используемые для приготовления твердых или жидких добавок или обычно используемые при подготовке кормов для животных. Добавки в корм для животных могут включать в себя эффективное количество фитазы, достаточное для ассимиляции и ферментативного отщепления свободной фосфорной кислоты из фитата.
ПРИМЕРЫ
Пример 1: Экспрессия в Pichia pastoris
1. Конструкция экспрессирующего вектора.
Для изоляции области, кодирующей зрелый белок, использовались следующие олигонуклеотиды:
SerrF: GCGCGCGAATTCGAGCCGCGCTACGTATTGG (SEQ ID NO: 6)
SerrR: GCGCGCAAGCTTGTCTAGACGTGGCCGGTACTGCACCG (SEQ ID NO: 7)
Область, кодирующую зрелый белок, амплифицировали с помощью олигонуклеотидов SerrF и SerrR. Продукт амплификации анализировали на агарозном геле, соответствующую ожидаемому размеру полосу вырезали, и ДНК извлекли из геля с помощью NucleoSpin Extract II Kit (Macherey-Nagel).
Очищенную ДНК вставили в вектор pPICZα A (Invitrogen, Сан-Диего, Калифорния), используя сайты рестрикции EcoRI and Xbal, содержащиеся в олигонуклеотидах. Сайт рестрикции EcoRI в SerrF и сайт рестрикции XbaI в SerrR выделены жирным шрифтом. Конструкцию со вставленной очищенной ДНК трансформировали в клетки DH5α, которые впоследствии высевали на чашки Петри в среде средой Лурии-Бертани, содержащей 25 мкг/мл зеоцин. Положительные колонии отбирали, и выделенные векторы секвенировали. Положительный клон с правильной последовательностью культивировали в больших объемах с целью получения ДНК для трансформации в дрожжи.
2. Трансформация в дрожжи и экспрессия белка
Штамм Pichia pastoris KH71H (Invitrogen) культивировали на среде YPD и подготовили к трансформации. 10 мкг плазмиды pPICZα A AppAs-r линеаризовали с помощью рестриктазы Pmel и ввели в клетки дрожжей с помощью литий/полиэтиленового шока.
Трансформированные дрожжи высевали на среду YPDZ, содержащую на 100 мкг/мл зеоцин, что позволяло отобрать колонии, интегрировавшие ген Sh ble (который придает устойчивость к зеоцину) в хромосомную ДНК хозяина. Лишь те трансформанты, которые интегрировали ген Sh ble, были способны расти на среде YPDZ. Через два дня трансформанты инокулировали и выращивали на минимальной среде, содержащей глицерин (BMGY) в течение 24 часов, после чего дрожжи отделяли центрифугированием (2500g, 3 мин) и ресуспендировали в среде с 0.5% метанола (BMMY) для индукции экспрессии гена фитазы.
3. Количественная оценка фитазной активности трансформантов
В общей сложности была проанализирована и количественно оценена фитазная активность 61 трансформанта с помощью молибдат-ванадатного способа в кислой среде (ISO 300024:2009 (E)).
1.160 мкл раствора субстрата (5 мМ фитат натрия 250 мМ натрий-ацетатном буфере, pH 5.5) добавили к 40 мкл разбавленного раствора фермента. Реакцию проводили при 37 °С в течение 30 минут. Далее для прекращения реакции к раствору добавили 800 мкл STOP-раствора (14% азотной кислоты; 3.3% четырехводный гептамолибдат аммония, 0.078% ванадат). В контрольном эксперименте STOP-раствор добавили к разбавленному раствору фермента для его инактивации, после чего добавили раствор субстрата. Через 10 минут измерили интенсивность желтого цвета при 415 Нм, и определили активность ферментного раствора и соответствующего контрольного раствора. Одна единица фитазной активности определяется как количество фермента, который отщепляет 1 мкмоль фосфора в минуту. Через два дня после индукции метанолом 6 из 61 трансформанта имели активность от 10 до 55 ед/мл в среде культивирования. Полученный ген представляет собой новый ген, который кодирует полипептид с фитазной активностью. Полипептид с фитазной активностью обозначается как AppAs-r.
Пример 2: Очистка полипептида AppAs-r, экспрессированного в Pichia pastoris
Для очистки фитазы, наработанной в Pichia pastoris, трансформанты с активностью 55 ед/мл культивировали и индуцировали в них экспрессию в оптимальных условиях. Через четыре дня после индукции метанолом, фитазную активность измеряли с использованием того же способа, что и в предыдущих примерах, который показал активность в супернатанте 163ед/л. Супернатант получали центрифугированием при 14100g в течение 5 минут. Осевшие клетки отбрасывали.
Супернатант получали центрифугированием через концентратор VIVASPIN 20 (размер поры 50 кДа (Sartorius Studium)). Концентрированный раствор затем диализовали в натрий-ацетатном буфере (0.25 М, рН 5.5). Наконец, фитазу очищали с помощью гель-фильтрации (Sephacryl S-200) с тем же буфером, что использовался для диализа.
Пример 3: Характеристика AppAs-r Оптима, экспрессированной в Pichia pastoris
1. Определение молекулярной массы
Молекулярную массу фитазы AppAs-r Оптима охарактеризовали с использованием денатурирующего акриламидного геля (SDS-PAGE). На Фиг.1 показан результат SDS-PAGE:
1) Первая дорожка соответствует контрольной пробе (среда культивирования без индукции метанолом),
2) вторая дорожка соответствует экспрессии фитазы AppAs-r в среде культивирования с метанолом,
3) третья полоса соответствует экспрессии фитазы AppAs-r Оптима в среде культивирования с метанолом,
4) четвертая полоса соответствует маркеру молекулярной массы.
В дорожки 1, 2 и 3 был загружен один и тот же объем образца.
Как видно из Фиг.1, молекулярная масса AppAs-r Оптима составляет около 46 кДа, что соответствует предполагаемой молекулярной массе полипептидной последовательности, соответствующей клонированной последовательности ДНК. Кроме того, из SDS-PAGE можно увидеть, что в одних и тех же условиях и при загрузке одного и того же количества образца уровень экспрессии гена AppAs-r Оптима значительно выше, чем гена AppAs-r.
2. Определение ферментативной активности AppAs-r Оптима в зависимости от температуры
Как показано на Фиг.2, активность фермента меняется с температурой. Оптимальную температуру определяли при 5.5 pH, что является показателем pH, требуемым ISO 300024:2009 (Е). Активность испытывали в диапазоне температур от 35°С до 85°С. Согласно измерениям, максимальная активность наблюдалась при температуре 50°С. Как показано на Фиг.2, после 10 минут при 60°С фермент по-прежнему сохраняет 40% своей активности.
3. Определение ферментативной активности AppAs-r Оптима в зависимости от pH
Влияние рН на фитазную активность испытывали, используя следующие буферы: глицин-HCl при рН 1.5-3.5; натрий-ацетатный при рН 3.5-6.0; Трис-HCl при рН 6.0-8.5; глицин-NaOH при рН 8.5-10.0. Все буферы содержали 0.05% IGPAL.
Как показано на Фиг.2, ферментативная активность меняется с pH. Максимальная активность наблюдалась при 5.0 pH. Активность 70% по-прежнему сохранялась при 4.0 рН, и более 80% активности сохранялось при pH 5.5.
4. Влияние протеаз на ферментативную активность
Чтобы определить устойчивость к действию протеаз, очищенную фитазу AppAs-r Оптима (1.6 мг/мл) инкубировали с 5 мг/мл пепсина или 50 мг/мл трипсина при 37°С. После инкубации в течение различных периодов времени (5, 10, 15, 30 и 60 минут) количественно оценивали остаточную фитазную активность в образце. Как видно из Фиг.4, через час после инкубации с пепсином фитаза AppAs-r сохраняла более 70% (73%) своей первоначальной активности и более 90% (95%) своей первоначальной активности через час после инкубации с трипсином. Это свидетельствует о том, что испытываемая фитаза является высокостойкой к действию протеаз, что может быть очень выгодно при использовании в кормах для животных.
5. Сохранение фитазной активности с течением времени
Для определения активности фитазы AppAs-r с течением времени специфическую активность в минуту определяли в различные моменты времени после добавления субстрата. Как следует из Фиг.5, AppAs-r отличается от большинства фитаз тем, что ее специфическая активность увеличивается с течением времени. AppAs-r по-прежнему активна по меньшей мере через 8 часов после добавления субстрата и удваивает свою первоначальную специфическую активность после 4.5 часов работы (Фиг.5). Подобное увеличение и поддержание активности с течением времени не была показана ни для каких других фитаз. Это очень важное обстоятельство, так как пищеварение животных обычно занимает более 30 минут, и поэтому необходимо, чтобы фермент оставался активным как можно дольше. Возрастающая и устойчивая во времени активность может также быть полезной при создании жидких кормов. Таким образом, это позволяет освободить наибольшую часть фосфора, присутствующего в форме фитата в корме, используя меньшее количество фермента.
6. Определение специфической активности
Для определения специфической активности концентрацию сконцентрированной и очищенной AppAs-r фитазы измеряли с использованием метода Лоури, и специфическую активность оценивали с применением молибдованадиевой кислоты при 37°С и рН 4.75. Результирующая специфическая активность очищенной AppAs-r составила 1.123 ± 112 ед/мг.
Пример 4: Высвобождение фосфата из органического материала почвы
В связи с длительным периодом активности фитазы важно определить ее способность высвобождать неорганический фосфор из органического материала в почве. Результаты этого анализа были сочтены полезными для приготовления добавок для удобрений. Для этого анализа взяли 5 образцов почвы. 2.5 г каждого образца взвесили и ресуспендировали в ацетатном буфере, и конечную концентрацию 0.25 г образца/мл рассчитали и скорректировали. Образцы инкубировали в течение 20 минут при постоянном перемешивании и температуре окружающей среды. После десятикратного разбавления образцов ацетатным буфером, эквивалент 1ед фитазы AppAs-r на 100 г почвы добавили к 1.2 мл каждого образца и инкубировали в течение 24 часов при температуре 25°С. Результаты свидетельствуют о том, что высвобождение фосфора составило 53.6 ± 27.8 мг фосфора/100 г почвы, таким образом повысив свободный фосфат почвы на 23.4 ± 18.1%.
Пример 5: Сравнительная экспрессия гена фитазы дикого типа и модифицированного гена фитазы (фитазы Оптима)
Авторы настоящего изобретения заметили, что экспрессия полноразмерной версии гена фитазы дикого типа из Serratia odorifera, показанной на Фиг.6A (SEQ ID NO: 2), содержащего сигнальный пептид, состоящий из 26 аминокислот, приводит к неустойчивому белку, который после экспрессии быстро исчезает. Авторы нынешнего изобретения также наблюдали, что пропуск 26-звенного сигнального пептида (SEQ ID NO: 8) приводил к усилению экспрессии (см. Пример 1).
С момента этого наблюдения, произведенного для вектора pPICZα, содержащего нуклеотидную последовательность, кодирующую зрелую фитазу дикого типа Serratia odorifera, как описано в примере 1, было оценено, как можно изменить последовательность нуклеотидов с точки зрения улучшения экспрессии белка фитазы в клетке-хозяине.
Чтобы получить улучшенную последовательность AppAs-r, оригинальные кодоны последовательно анализировали и заменяли на наиболее часто используемые кодоны для каждой аминокислоты в P. pastoris. Смещение частоты использования кодонов в P. pastoris было ранее описано в работах Bai et al. (Bai J., Swartz D.J., Protasevich I.I., Brouillette C.G., Harrell P.M., Hidebrandt E., Gasser B., Mattanovich D., Ward A., Chang G. and Urbatsch I.L. (2011) A gene optimization strategy that enhances production of fully functional P-glycoprotein in Pichia pastoris. PLosOne 6:1-15), Sinclair et al. (Sinclair G. and Choy F.Y.M. (2002) Synonymous codon usage bias and the expression of human glucocerebrosidase in the methylotrophic yeast Pichia pastoris. Protein Expression and Purification 26:96-105) и Huang et al. (Huang H, Yang P, Luo H, Tang H, Shao N, et al. (2008) High-level expression of a truncated 1,3-1,4-beta-D-glucanase from Fibrobacter succinogenes in Pichia pastoris by optimization of codons and fermentation. Appl Microbiol Biotechnol. 78: 95-103.).
В последовательности SEQ ID NO: 1, наиболее часто используемые кодоны в P. pastoris были проанализированы, идентифицированы и применены. В последовательности SEQ ID NO: 1 были оптимизированы все 211 кодонов кодирующей последовательности дикого типа без последовательности, кодирующей N-концевой сигнальный пептид (SEQ ID NO: 8). В оценках также учитывается, что частота использования кодонов, описанная в литературе, в определенной степени была сохранена в некоторых генах P. pastoris, кодирующих белки с каталитической активностью, такие как фосфатазы и киназы.
Модифицированный полинуклеотид фитазы, включающий оптимизированный кодон, как показано в SEQ ID NO: 1 был получен с помощью синтеза генов. Модифицированный полинуклеотид, кодирующий фитазу, был куплен у компании GenScript (Испания) и вставлен в вектор pPICZα A (Invitrogen, Сан-Диего, Калифорния) через рестрикционные сайты EcoRI и Xbal.
Результирующая плазмида, содержащая ген AppAs-rOp, была линеаризована воздействием рестриктазы Pmel и трансформирована в клетки дрожжей с помощью литий/полиэтиленового шока, как в примере 1.
Экспрессированный генный продукт сконструированного и оптимизированного гена Serratia odorifera соответствует зрелому белку, в котором отсутствуют первые 26 аминокислот, т.е. после отщепления сигнального пептида в клетке, который имеет молекулярный вес около 45-46 кДа, как видно из результатов гель-электрофореза в SDS на Фиг.1.
Начиная с вектора pPICZα, содержащего нуклеотидную последовательность, кодирующую зрелую фитазу Serratia odorifera, как описано в примере 1, изобретатели настоящего изобретения оценили и испытали точечные мутации в этой последовательности нуклеотидов, которые могли бы привести к усиленной экспрессии белка фитазы в клетке-хозяине. Было отмечено, что 211 точечных мутаций, показанных на Фиг.6B (SEQ ID NO: 1), по сравнению с последовательностью фитазы дикого типа (SEQ ID NO: 2) способствовали эффективной и устойчивой экспрессии в дрожжах Pichia pastoris. На Фиг.6С показано выравнивание последовательности фитазы Оптима (SEQ ID NO: 1) с последовательностью фитазы дикого типа (SEQ ID NO: 2). Несоответствия указывают на точечные мутации, сделанные в исходной последовательности генов. Точечные мутации приводят к кодонам, наиболее часто используемым в дрожжевой клетке-хозяине Pichia pastoris. Как видно из Фиг.6D, на котором показано выравнивание транслированных зрелых белков SEQ ID NO: 1 и SEQ ID NO: 2, все 211 мутаций являются молчащими, т.е. ни одна из этих мутаций не приводит к замене аминокислоты в фитазном полипептиде.
Чтобы определить, действительно ли мутации, внесенные в SEQ ID NO: 1 влияют на экспрессию белка в дрожжах, модифицированную последовательность нуклеотида экспрессировали в тех же условиях, что и в примере 1. Чтобы определить профили экспрессии, образцы были отобраны в моменты времени 0, 19, 31, 43, 55, 67, 79 и 91 час после индукции экспрессии метанолом. Фитазную активность измеряли с использованием молибдат-ванадатного способа в кислой среде (ISO 300024:2009 (E)), как это описано в примере 3. В качестве контроля использовался AppAs-r из примера 3.
Результаты суммированы в Таблице 1. Данные этого контрольного примера представлены в логарифмической шкале на графике, показанном на Фиг.8 и отображающем активность, выраженную в единицах/л в зависимости от времени (часы).
На Фиг.8 профиль экспрессии фитазы дикого типа AppsAs-r (контроль) показан синим цветом (закрашенные ромбы), а профиль экспрессии в последовательности Оптима показан красным цветом (закрашенные квадраты). Как видно из графика, экспрессия генов фитазы дикого типа не является оптимальной. Первые поддающиеся измерению продукты появляются через 40 часов после индукции. Через 91 час кривая насыщения показывает максимальную активность, составляющую 143 единицы/л. Примечательно, что экспрессия модифицированной генной последовательности фитазы (последовательность Оптима) показывает детектируемый продукт гена фитазы непосредственно в первые часы экспрессии белка, и достигает насыщения после 20-30 часов с момента индукции экспрессии гена. По истечении 90 часов производство фитазы превышало контроль более чем в 200 раз. Это увеличение производства было неожиданным.
Пример 5, таким образом, показывает, что мутации, осуществленные в последовательности Оптима и учитывающие частоту использования кодона в соответствующей дрожжевой клетке-хозяине Pichia pastoris, действительно способствует повышенной и эффективной экспрессии белка.
Таблица 1: Профиль экспрессии фитазы дикого типа по сравнению с фитазой Оптима
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Фертинагро Нутриентес, ООО
<120> ИННОВАЦИОННАЯ ФИТАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ИСПОЛЬЗОВАНИЯ
<130> FERNU.0010 WO
<160> 8
<170> PatentIn version 3.5
<210> 1
<211> 1203
<212> ДНК
<213> Serratia odorifera
<400> 1
gagcctagat acgttttgga aaaggtggtt gaggtctctc gacacggcgt tagaccacca 60
acctcaggta acagacaggc tatgcaagcg ggaaccggaa gggagtggcc acaatggctg 120
actcgtgacg gagaactaac tggacacggt tatgcagccg caacacttaa aggaagatac 180
gaagccgatt attatagacg tcagggccta ttggcaaacg gttgtccatc tgctggggct 240
gtttacgtct gggctagtcc tctacaaaga acaagggcca ccgcacaggc tttgatggac 300
ggtgcatttc ccggatgcgg ggtcgccatt catgctgcag ccactgaaca agatcccttg 360
tttcaagcag ataagatggg tcttgttcca ctcgatgccg aaagggctag aactgcaata 420
aggcaggcaa tgggtggctc agccgagcaa gtgaagacac gttttagtgc tgacattagg 480
cgtctgcaag ctgctgtttg tttgcctcaa caggcttgtc ctgcctttga acaaccttgg 540
gaaattactc aagagcatga cggtagattc agcatcaatg gtctaggaac attgtccaat 600
atggctgaat ctattagact tgcctactct gaaaaccagc caacggctca agtcgcattt 660
ggacacggtg tcaacgcatc cgccgtcgct cctttgttac cactgctaac cgccagatat 720
gactttacca atgatgtgcc ctatatcgct caaagaggtg gctccgttct gttaaaccaa 780
attgctttgg ctcttgccgc agataggact tctgccgggg cgccacctgc tgctcgttgg 840
ttgttatttg tcgctcatga caccaatatc gcttatttaa gaactctgct tggtttttct 900
tggcaacagg gactttaccc aagaggtaat attccccctg ctggaagttt ggttttcgaa 960
agatggcgtg atagacaaac aggtcaaagg ttcttacgtc tgtacttcca ggctcaatcg 1020
ttggatcaaa tcagacagtt gtcaccactt tctacattat ccccaccttt aaaaaccgag 1080
ttctctcgtc ctggttgcag gcagttgtca cttggcgttc tctgtccctg gactgagtcc 1140
atgcaaagaa tgagagctgc tatcgaccca actgcgttgc ctacagtgca gtacagacca 1200
taa 1203
<210> 2
<211> 1281
<212> ДНК
<213> Serratia odorifera
<400> 2
atgttgctat tgcaaaagga ctggtcgcgt ctgttatttg ccgtcacgct gggtatgatt 60
tccagcgtag cccaggctga gccgcgctac gtattggaaa aggtggttga ggtcagccgc 120
cacggcgtac gcccgccgac ctcaggcaac cggcaggcga tgcaggcggg aaccggccga 180
gagtggccac aatggctgac gcgcgacggc gaactcactg gccacggtta tgccgccgcc 240
acgctgaaag gacgctatga agccgactat tatcgccgtc agggcctatt ggccaacggc 300
tgtccgagcg cgggggcggt gtatgtctgg gccagtccgc tacagcgcac gcgagccacc 360
gcacaggcgt tgatggacgg cgcatttccc ggctgcgggg tcgccattca tgcggccgcc 420
accgaacagg accccctgtt tcaggcagat aaaatgggcc tggtgccgct cgatgccgaa 480
cgggctcgca cggcaataag gcaggcaatg ggcggcagcg ccgagcaggt gaaaacgcgc 540
tttagcgctg acattcggcg tctgcaagcg gcggtctgcc tgccgcaaca ggcttgcccg 600
gcctttgaac aaccgtggga aatcactcag gagcacgacg gccgcttcag catcaacggt 660
ctggggacgt tgtccaacat ggcggaaagc attcgcctgg cctacagcga aaaccagccg 720
acggcgcagg tcgcctttgg ccacggtgtc aacgcatcgg ccgtcgcgcc gttgctgccc 780
ctgctcaccg cccgctatga ctttaccaat gacgtgccct atatcgcgca acgcggtggc 840
tcggtgctgt taaaccaaat cgcgctggcg ctggccgccg atcgaacctc tgccggggcg 900
ccaccggcgg cgcgctggtt gctgtttgtc gcgcatgaca ccaatatcgc ttatctgcgc 960
accctgcttg gctttagctg gcaacagggg ctttacccac gcggcaatat tcccccggct 1020
ggcagtctgg tattcgaacg ctggcgcgat cggcaaacgg gccagcgctt cctgcgtctg 1080
tacttccagg cgcaatcgct ggatcaaatc cgccagttgt caccgctgag cacgctgtcg 1140
ccaccgttaa aaaccgagtt cagccgtcct ggctgccggc agttgtcact gggcgtactc 1200
tgtccctgga ctgagtcgat gcaacggatg cgcgcggcta tcgacccgac ggcgctgcct 1260
acggtgcagt accggccata a 1281
<210> 3
<211> 400
<212> БЕЛОК
<213> Serratia odorifera
<400> 3
Glu Pro Arg Tyr Val Leu Glu Lys Val Val Glu Val Ser Arg His Gly
1 5 10 15
Val Arg Pro Pro Thr Ser Gly Asn Arg Gln Ala Met Gln Ala Gly Thr
20 25 30
Gly Arg Glu Trp Pro Gln Trp Leu Thr Arg Asp Gly Glu Leu Thr Gly
35 40 45
His Gly Tyr Ala Ala Ala Thr Leu Lys Gly Arg Tyr Glu Ala Asp Tyr
50 55 60
Tyr Arg Arg Gln Gly Leu Leu Ala Asn Gly Cys Pro Ser Ala Gly Ala
65 70 75 80
Val Tyr Val Trp Ala Ser Pro Leu Gln Arg Thr Arg Ala Thr Ala Gln
85 90 95
Ala Leu Met Asp Gly Ala Phe Pro Gly Cys Gly Val Ala Ile His Ala
100 105 110
Ala Ala Thr Glu Gln Asp Pro Leu Phe Gln Ala Asp Lys Met Gly Leu
115 120 125
Val Pro Leu Asp Ala Glu Arg Ala Arg Thr Ala Ile Arg Gln Ala Met
130 135 140
Gly Gly Ser Ala Glu Gln Val Lys Thr Arg Phe Ser Ala Asp Ile Arg
145 150 155 160
Arg Leu Gln Ala Ala Val Cys Leu Pro Gln Gln Ala Cys Pro Ala Phe
165 170 175
Glu Gln Pro Trp Glu Ile Thr Gln Glu His Asp Gly Arg Phe Ser Ile
180 185 190
Asn Gly Leu Gly Thr Leu Ser Asn Met Ala Glu Ser Ile Arg Leu Ala
195 200 205
Tyr Ser Glu Asn Gln Pro Thr Ala Gln Val Ala Phe Gly His Gly Val
210 215 220
Asn Ala Ser Ala Val Ala Pro Leu Leu Pro Leu Leu Thr Ala Arg Tyr
225 230 235 240
Asp Phe Thr Asn Asp Val Pro Tyr Ile Ala Gln Arg Gly Gly Ser Val
245 250 255
Leu Leu Asn Gln Ile Ala Leu Ala Leu Ala Ala Asp Arg Thr Ser Ala
260 265 270
Gly Ala Pro Pro Ala Ala Arg Trp Leu Leu Phe Val Ala His Asp Thr
275 280 285
Asn Ile Ala Tyr Leu Arg Thr Leu Leu Gly Phe Ser Trp Gln Gln Gly
290 295 300
Leu Tyr Pro Arg Gly Asn Ile Pro Pro Ala Gly Ser Leu Val Phe Glu
305 310 315 320
Arg Trp Arg Asp Arg Gln Thr Gly Gln Arg Phe Leu Arg Leu Tyr Phe
325 330 335
Gln Ala Gln Ser Leu Asp Gln Ile Arg Gln Leu Ser Pro Leu Ser Thr
340 345 350
Leu Ser Pro Pro Leu Lys Thr Glu Phe Ser Arg Pro Gly Cys Arg Gln
355 360 365
Leu Ser Leu Gly Val Leu Cys Pro Trp Thr Glu Ser Met Gln Arg Met
370 375 380
Arg Ala Ala Ile Asp Pro Thr Ala Leu Pro Thr Val Gln Tyr Arg Pro
385 390 395 400
<210> 4
<211> 78
<212> ДНК
<213> Serratia odorifera
<400> 4
atgttgctat tgcaaaagga ctggtcgcgt ctgttatttg ccgtcacgct gggtatgatt 60
tccagcgtag cccaggct 78
<210> 5
<211> 26
<212> БЕЛОК
<213> Serratia odorifera
<400> 5
Met Leu Leu Leu Gln Lys Asp Trp Ser Arg Leu Leu Phe Ala Val Thr
1 5 10 15
Leu Gly Met Ile Ser Ser Val Ala Gln Ala
20 25
<210> 6
<211> 31
<212> ДНК
<213> Serratia odorifera
<400> 6
gcgcgcgaat tcgagccgcg ctacgtattg g 31
<210> 7
<211> 38
<212> ДНК
<213> Serratia odorifera
<400> 7
gcgcgcaagc ttgtctagac gtggccggta ctgcaccg 38
<210> 8
<211> 1203
<212> ДНК
<213> Serratia odorifera
<400> 8
gagccgcgct acgtattgga aaaggtggtt gaggtcagcc gccacggcgt acgcccgccg 60
acctcaggca accggcaggc gatgcaggcg ggaaccggcc gagagtggcc acaatggctg 120
acgcgcgacg gcgaactcac tggccacggt tatgccgccg ccacgctgaa aggacgctat 180
gaagccgact attatcgccg tcagggccta ttggccaacg gctgtccgag cgcgggggcg 240
gtgtatgtct gggccagtcc gctacagcgc acgcgagcca ccgcacaggc gttgatggac 300
ggcgcatttc ccggctgcgg ggtcgccatt catgcggccg ccaccgaaca ggaccccctg 360
tttcaggcag ataaaatggg cctggtgccg ctcgatgccg aacgggctcg cacggcaata 420
aggcaggcaa tgggcggcag cgccgagcag gtgaaaacgc gctttagcgc tgacattcgg 480
cgtctgcaag cggcggtctg cctgccgcaa caggcttgcc cggcctttga acaaccgtgg 540
gaaatcactc aggagcacga cggccgcttc agcatcaacg gtctggggac gttgtccaac 600
atggcggaaa gcattcgcct ggcctacagc gaaaaccagc cgacggcgca ggtcgccttt 660
ggccacggtg tcaacgcatc ggccgtcgcg ccgttgctgc ccctgctcac cgcccgctat 720
gactttacca atgacgtgcc ctatatcgcg caacgcggtg gctcggtgct gttaaaccaa 780
atcgcgctgg cgctggccgc cgatcgaacc tctgccgggg cgccaccggc ggcgcgctgg 840
ttgctgtttg tcgcgcatga caccaatatc gcttatctgc gcaccctgct tggctttagc 900
tggcaacagg ggctttaccc acgcggcaat attcccccgg ctggcagtct ggtattcgaa 960
cgctggcgcg atcggcaaac gggccagcgc ttcctgcgtc tgtacttcca ggcgcaatcg 1020
ctggatcaaa tccgccagtt gtcaccgctg agcacgctgt cgccaccgtt aaaaaccgag 1080
ttcagccgtc ctggctgccg gcagttgtca ctgggcgtac tctgtccctg gactgagtcg 1140
atgcaacgga tgcgcgcggc tatcgacccg acggcgctgc ctacggtgca gtaccggcca 1200
taa 1203
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТНЫЕ ФИТАЗНЫЕ ФЕРМЕНТЫ | 2005 |
|
RU2421520C2 |
| НОВЫЕ ТЕРМОСТАБИЛЬНЫЕ ФИТАЗЫ С ВЫСОКОЙ КАТАЛИТИЧЕСКОЙ ЭФФЕКТИВНОСТЬЮ | 2018 |
|
RU2758271C2 |
| Мутант фосфолипазы С с высокой ферментативной активностью | 2019 |
|
RU2818353C2 |
| МУТАНТНАЯ РЕКОМБИНАНТНАЯ ТЕРМОСТАБИЛЬНАЯ ФИТАЗА (ВАРИАНТЫ), ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ УКАЗАННУЮ ФИТАЗУ (ВАРИАНТЫ), ШТАММ Pichia pastoris - ПРОДУЦЕНТ УКАЗАННОЙ ФИТАЗЫ (ВАРИАНТЫ) | 2009 |
|
RU2472855C2 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок SMN1, и ее применение | 2020 |
|
RU2742837C1 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ, ВЕКТОРЫ, КЛЕТКИ ХОЗЯИНА И СПОСОБЫ ПОЛУЧЕНИЯ БЕТА-ФРУКТОФУРАНОЗИДАЗЫ ИЗ ASPERGILLUS NIGER | 2020 |
|
RU2840101C1 |
| МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, КОДИРУЮЩАЯ HPV31 L1, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ВИРУСОПОДОБНАЯ ЧАСТИЦА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ВАКЦИНА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБЫ С ИХ ИСПОЛЬЗОВАНИЕМ | 2004 |
|
RU2356943C2 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ, ВЕКТОРЫ, КЛЕТКИ ХОЗЯИНА И СПОСОБЫ ПОЛУЧЕНИЯ ФРУКТОЗИЛТРАНСФЕРАЗЫ ИЗ ASPERGILLUS JAPONICUS | 2020 |
|
RU2837819C1 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок фактора свёртывания крови VIII c делетированным B доменом, и ее применение | 2022 |
|
RU2808564C2 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок фактора свёртывания крови IX, и ее применение | 2021 |
|
RU2831751C2 |
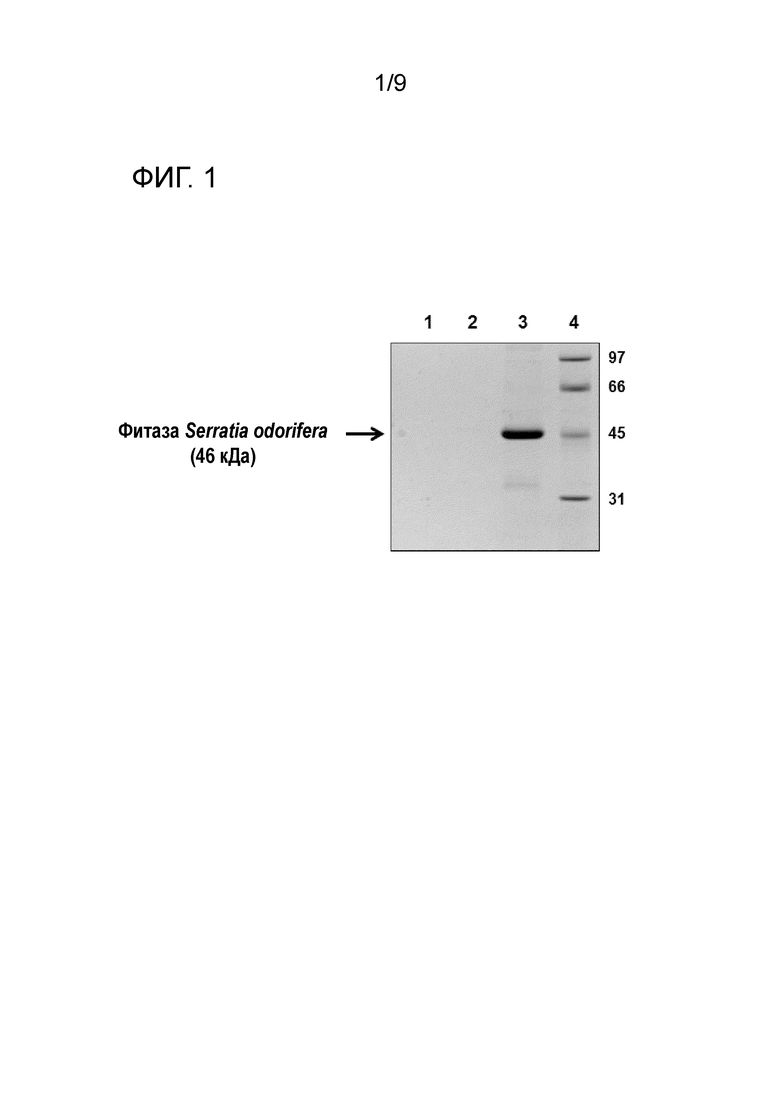

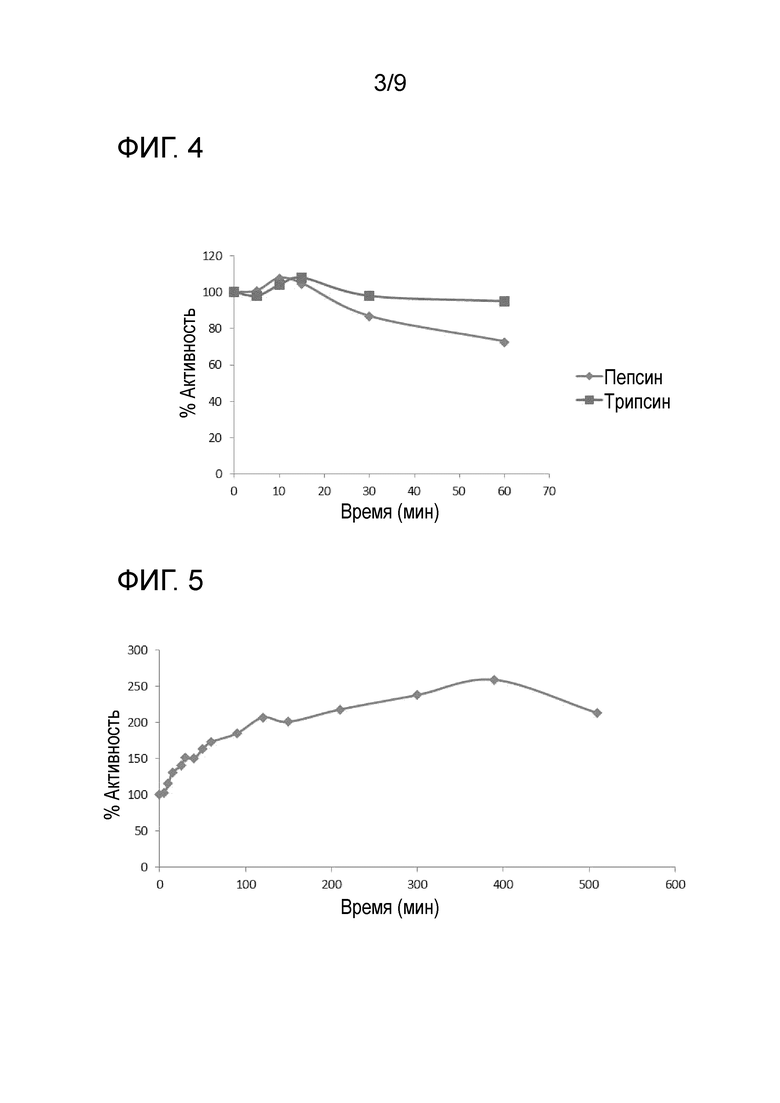





Изобретение относится к области биотехнологии. Предложена выделенная молекула нуклеиновой кислоты, кодирующая белок с фитазной активностью. Молекула нуклеиновой кислоты содержит кодон-оптимизирующие мутации, учитывающие особенности организма клетки-хозяина, представляющего собой Pichia pastoris. Изобретение также относится к экспрессирующей конструкции и к экспрессирующему вектору, включающим вышеупомянутые выделенные молекулы нуклеиновой кислоты, а также к способу получения белка, имеющего фитазную ферментативную активность. Изобретение обеспечивает усиленную экспрессию модифицированной фитазы по сравнению с фитазой дикого типа и может применяться для изготовления добавок в корм для животных. 4 н. и 9 з.п. ф-лы, 12 ил., 1 табл., 5 пр.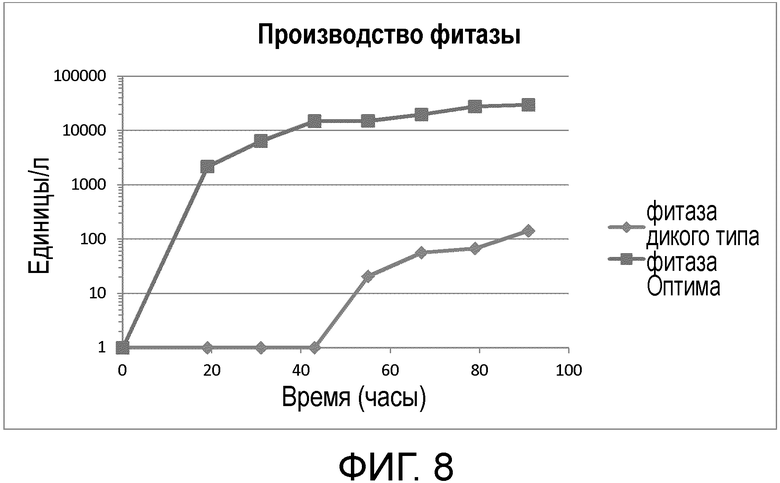
1. Выделенная молекула нуклеиновой кислоты
i) содержащая нуклеотидную последовательность, показанную в SEQ ID NO: 1, кодирующую белок с фитазной активностью, или
ii) обладающая идентичностью последовательности по меньшей мере 86% с нуклеотидной последовательностью, показанной в SEQ ID NO: 1, кодирующей белок с фитазной активностью,
причем выделенная молекула нуклеиновой кислоты содержит кодон-оптимизирующие мутации, учитывающие особенности организма клетки-хозяина,
причем организм клетки-хозяина представляет собой Pichia pastoris, и
причем молекула нуклеиновой кислоты отличается от соответствующей кодирующей нуклеотидной последовательности дикого типа согласно SEQ ID NO: 8 наличием по меньшей мере 50 кодон-оптимизирующих мутаций.
2. Выделенная молекула нуклеиновой кислоты по п. 1, отличающаяся тем, что молекула нуклеиновой кислоты не включает нуклеотидную последовательность, кодирующую N-концевой сигнальный пептид.
3. Выделенная молекула нуклеиновой кислоты по любому из предшествующих пунктов, отличающаяся тем, что по меньшей мере 50% кодонов соответствующей нуклеотидной последовательности дикого типа заменены на кодоны, наиболее часто используемые организмом клетки-хозяина.
4. Выделенная молекула нуклеиновой кислоты по любому из предшествующих пунктов, отличающаяся тем, что все кодоны выделенной молекулы нуклеиновой кислоты заменены на кодоны, наиболее часто используемые организмом клетки-хозяина.
5. Выделенная молекула нуклеиновой кислоты по любому из предшествующих пунктов, кодирующая аминокислотную последовательность белка с фитазной активностью в соответствии с SEQ ID NO: 3.
6. Экспрессирующая конструкция, содержащая выделенную молекулу нуклеиновой кислоты по любому из пп. 1-5, функционально связанную с элементами, регулирующими экспрессию нуклеотидной последовательности в клетке-хозяине, причем организм клетки-хозяина представляет собой Pichia pastoris.
7. Экспрессирующая конструкция по п. 6, отличающаяся тем, что элементы, регулирующие экспрессию, включают промотор, функционирующий в клетке-хозяине и, необязательно, терминирующую последовательность.
8. Экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты по пп. 1-5 или экспрессирующую конструкцию по любому из пп. 6-7.
9. Способ получения белка, имеющего фитазную ферментативную активность, включающий этапы:
a) введение в клетку-хозяина вектора, i) содержащего элементы, регулирующие экспрессию, функционирующие в клетке-хозяине; и ii) функционально связанного с молекулой нуклеиновой кислоты по любому из пп. 1 или 2;
b) культивирование клеток-хозяев, полученных на этапе a) в условиях, подходящих для экспрессии белка, и, при необходимости,
c) выделение белка, полученного на этапе b), из культуры клеток,
причем организм клетки-хозяина представляет собой Pichia pastoris.
10. Способ по п. 9, отличающийся тем, что элементы, регулирующие экспрессию, включают в себя промотор, функционирующий в клетке-хозяине и, при необходимости, терминирующую последовательность.
11. Способ по п. 9 или 10, в котором экспрессия белка осуществляется с использованием метанольной системы экспрессии.
12. Способ по любому из пп. 9-11, отличающийся тем, что белок, выделенный на этапе c), обладает фитазной активностью по меньшей мере 1000 единиц/л.
13. Способ по любому из пп. 9-12, отличающийся тем, что этап выделения включает a) отделение секретируемого белка от компонентов среды и/или b) отделение белка от компонентов клетки-хозяина.
| WO 2011141613 A2, 17.11.2011 | |||
| MIMI YANG et al | |||
| Enhancement of alkaline phytase production in Pichia pastoris: Influence of gene dosage, sequence optimization and expression temperature, Protein Expression and Purification, 2012, vol | |||
| Способ приготовления сернистого красителя защитного цвета | 1921 |
|
SU84A1 |
| RUI ZHANG et al | |||
| Two Types of Phytases (Histidine Acid Phytase and β-Propeller Phytase) | |||

