Уровень техники
Настоящее изобретение относится к биспецифическим антителам, которые функционально используют вместо кофакторов, усиливающих ферментативную реакцию, и фармацевтическим композициям, содержащим это антитело в качестве действующего ингредиента.
Предшествующий уровень техники
Антителам в качестве лекарственного средства уделяют много внимания благодаря их высокой стабильности в крови и низкой антигенности. Среди них - биспецифические антитела, которые могут одновременно распознавать два типа антигенов. За последнее время были предложены биспецифические антитела; однако сообщалось только об антителах, которые просто связывают два типа антигенов, например, такие, которые переносят NK-клетки, макрофаги и Т-клетки (см. непатентный документ 7). Например, MDX-210, в настоящее время проходящее клиническое испытание, представляет собой биспецифическое антитело, которое просто перенацеливается на FcγRI-экспрессирующие моноциты, а также на раковые клетки, экспрессирующие HER-2/neu. Таким образом, до сих пор не было примеров применения биспецифического антитела в качестве альтернативного средства для функционального замещения кофактора, который усиливает ферментативную реакцию.
Примерами кофакторов являются тканевой фактор (TF), фактор свертывания крови V (F.V), активированный фактор свертывания крови V (F.Va), фактор свертывания крови VIII (F.VIII), активированный фактор свертывания крови VIII (F.VIIIa), тромбомодулин (ТМ), белок S (PS), белок Z (PZ), гепарин, комплемент С4b, регуляторный фактор Н системы комплемента, мембранный кофакторный белок (МСР) и рецептор комплемента 1 (CR1).
Из числа вышеперечисленного F.VIII/F.VIIIa представляет собой кофактор, необходимый для достаточно эффективной экспрессии активированного фактора свертывания крови IX (F.IXa). Scheiflinger F. et al. в результате хромогенного исследования выявили, что определенное анти-F.IX/F.IXa антитело действует, способствуя активированию фактора свертывания крови Х (F.X) посредством F.IXa (патентный документ 1). Однако в исследовании способности к восстановлению коагуляции плазмы с дефицитом фактора F.VIII способность восстановления коагуляции наблюдали только при добавлении внешнего фактора F.IXa, но не в случае использования одного только антитела.
Для фактора F.VIIIa известно, что он взаимодействует не только с фактором F.IXa, но также и с F.X (см. непатентные документы 5 и 6). В этом отношении антитело, описанное Scheiflinger F. et al., не может быть названо как достаточно эффективное при использовании вместо функции F.VIII/F.VIIIa, и его активность также представляется недостаточной.
В результате специализированного научного исследования авторы настоящего изобретения преуспели в получении биспецифических антител, функционально замещающих кофакторы, которые усиливают ферментативную активность, и таким образом осуществили данное изобретение.
[Патентный документ 1] WO 01/19992
[Патентный документ 2] Патент США № 4474893
[Патентный документ 3] EP 404097
[Патентный документ 4] WO 93/11161
[Патентный документ 5] Японская патентная заявка № 2002-112369
[Патентный документ 6] Японская патентная заявка № 2003-012648
[Патентный документ 7] Японская патентная заявка Kokai Publication № (JP-A) H5-304992 (не рассмотренная, опубликованная Японская патентная заявка)
[Патентный документ 8] JP-A H2-145187
[Патентный документ 9] JP-A H5-213775
[Патентный документ 10] JP-A H10-165184
[Патентный документ 11] JP-A H11-71288
[Патентный документ 12] JP-A 2002-518041
[Патентный документ 13] JP-A H11-506310
[Патентный документ 14] JP-A H5-199894
[Патентный документ 15] JP-A H10-511085
[Патентный документ 16] JP-A H5-184383
[Непатентный документ 1] Nilsson IM et al., "J. Intern. Med." 1992, Vol.235, p.25-32
[Непатентный документ 2] Lofqvist T et al., "J. Intern. Med" 1997, Vol.241, p.395-400 (фамилия Lofqvist написана с двумя точками над гласной «о»)
[Непатентный документ 3] 24th Meeting of The Japanese Society on Thrombosis and Hematosis, Special Committee on Examining Hemophilia Standardization, Mini-symposium, 2001, http://www.jsth.org
[Непатентный документ 4] Medical Bulletin #193 1994
[Непатентный документ 5] Mertens K et al., "Thromb. Haemost." 1999, Vol.82, p.209-217
[Непатентный документ 6] Lapan KA et al., "Thromb. Haemost." 1998, Vol.80, p.418-422
[Непатентный документ 7] Segal DM et al., "Journal of Immunological Methods" 2001, Vol.248, p.1-6
[Непатентный документ 8] Bos R and Nieuwenhuitzen W, "Hybridoma" 1992, Vol.11, № 1, p.41-51
[Непатентный документ 9] Brennan M et al., "Science" 1985, Vol.229, № 1708, p.81-3
[Непатентный документ 10] Karpovsky B et al., "J. Exp. Med." 1984, Vol.160, № 6, p.1686-701
[Непатентный документ 11] Suresh MR et al., "Methods Enzymol." 1986, Vol.121, p.210-28
[Непатентный документ 12] Massimo YS et al., "J. Immunol. Methods" 1997, Vol.201, p.57-66
[Непатентный документ 13] Brennan M et al., "Science" 1985, Vol.229, p.81
[Непатентный документ 14] Shalaby MR et al., "J. Exp. Med." 1992, Vol.175, p.217-25
[Непатентный документ 15] Holliner P et al., "Proc. Natl. Acad. Sci. USA" 1993, Vol.90, p.6444-8
[Непатентный документ 16] Ridgway JB et al., "Protein Eng." 1996, Vol.9, p.617-21
[Непатентный документ 17] Hammerling U et al., "J. Exp. Med." 1968, Vol.128, p.1461-73
[Непатентный документ 18] Kurokawa T et al., "Bio/Technology" 1989, Vol.7, p.1163
[Непатентный документ 19] Link BK et al., "Blood" 1993, Vol.81, p.3343
[Непатентный документ 20] Nitta T et al., "Lancet" 1990, Vol.335, p.368-71
[Непатентный документ 21] deLeij L et al., "Foundation Nationale de Transfusion Sanguine, Les Ulis France" 1990, p.249-53
[Непатентный документ 22] Le Doussal JM et al., "J. Nucl. Med." 1993, Vol.34, p.1662-71
[Непатентный документ 23] Stickney DR et al., "Cancer Res." 1991, Vol.51, p.6650-5
[Непатентный документ 24] Weiner LM et al., "Cancer Res." 1993, Vol.53, p.94-100
[Непатентный документ 25] Kroesen BJ et al., "Br. J. Cancer" 1994, Vol.70, p.652-61
[Непатентный документ 26] Weiner GJ et al., "J. Immunol." 1994, Vol.152, p.2385
[Непатентный документ 27] Suresh MR et al., "Proc. Natl. Acad. Sci. USA" 1986, Vol.83, p.7989-93
[Непатентный документ 28] Milstein C and Cuello AC, "Nature" 1983, Vol.305, p.537
[Непатентный документ 29] Xiang J et al., "Mol. Immunol." 1990, Vol.27, p.809
[Непатентный документ 30] Bebbington CR et al., "Bio/Technology" 1992, Vol.10, p.169
[Непатентный документ 31] Huse WD et al., "Science" 1989, Vol.246, p.1275
[Непатентный документ 32] McCafferty J et al., "Nature" 1990, Vol.348, p.552
[Непатентный документ 33] Kang AS et al., "Proc. Natl. Acad. Sci. USA" 1991, Vol.88, p.4363
Изложение сущности изобретения
Целью настоящего изобретения является обеспечение биспецифических антител, функционально замещающих кофакторы, которые усиливают ферментативную реакцию.
В результате специализированного научного исследования авторы настоящего изобретения преуспели в изобретении биспецифических антител, которые, в частности, связывают и F.IX/F.IXa, и F.X, и функционально замещают кофактор F.VIIIa (то есть действие способствует активации F.X под действием F.IXa). Значит, авторы настоящего изобретения преуспели в получении биспецифических антител, которые распознают и фермент, и его субстрат и функционально замещают кофакторы фермента.
Настоящее изобретение относится к биспецифическим антителам, которые функционально применяют вместо кофакторов, которые усиливают ферментативную реакцию, и, более конкретно, к:
[1] Антителу, распознающему и фермент, и его субстрат, где указанное антитело представляет собой биспецифическое антитело, которое функционально применяют вместо кофактора, который усиливает ферментативную реакцию.
[2] Антителу по п.[1], где указанный фермент является протеолитическим ферментом.
[3] Антителу по п.[2], где указанные протеолитический фермент, субстрат и кофактор представляют собой факторы, ассоциированные со свертыванием/фибринолизом крови.
[4] Антителу по п.[3], где фермент фактора, ассоциированного со свертыванием/фибринолизом крови, представляет собой фактор свертывания крови IX и/или активированный фактор свертывания крови IX; субстрат представляет собой фактор свертывания крови X; и кофактор представляет собой фактор свертывания крови VIII и/или активированный фактор свертывания крови VIII.
[5] Антителу по п.п.[1]-[4], где указанное антитело включает в себя гипервариабельный участок, включающий в себя аминокислотную последовательность CDR3 антитела против фактора свертывания крови IX/IXa следующих пунктов (а1) или (а2), или гипервариабельный участок, функционально эквивалентный этому, и гипервариабельный участок, включающий в себя аминокислотную последовательность CDR3 антитела против фактора свертывания крови X, описанную в одном из следующих пунктов (b1)-(b9), или гипервариабельный участок, функционально эквивалентный этому:
(а1) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:16;
(а2) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:20;
(b1) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:24;
(b2) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:28;
(b3) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:32;
(b4) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:36;
(b5) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:40;
(b6) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:44;
(b7) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:48;
(b8) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:52;
(b9) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:56.
[6] Антителу по п.п.[1]-[4], где указанное антитело включает в себя гипервариабельный участок, включающий в себя аминокислотную последовательность CDR антитела против фактора свертывания крови IX/IXa согласно следующим п.п.(а1) или (а2), или гипервариабельный участок, функционально эквивалентный этому, и гипервариабельный участок, включающий в себя аминокислотную последовательность CDR антитела против фактора свертывания крови X, описанную в одном из следующих пунктов (b1)-(b9), или гипервариабельный участок, функционально эквивалентный этому:
(a1) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:14, 15 и 16, соответственно;
(a2) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:18, 19 и 20, соответственно;
(b1) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:22, 23 и 24, соответственно;
(b2) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:26, 27 и 28, соответственно;
(b3) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:30, 31 и 32, соответственно;
(b4) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:34, 35 и 36, соответственно;
(b5) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:38, 39 и 40, соответственно;
(b6) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:42, 43 и 44, соответственно;
(b7) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:46, 47 и 48, соответственно;
(b8) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:50, 51 и 52, соответственно;
(b9) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:54, 55 и 56, соответственно.
[7] Композиции, включающей в себя антитело по любому из п.п.[1]-[6] и фармацевтически подходящий носитель.
[8] Композиции по п.[7], где указанная композиция представляет собой фармацевтическую композицию, используемую для профилактики и/или лечения кровотечения, заболевания, сопровождающегося кровотечением, или заболевания, вызванного кровотечением.
[9] Композиции по п.[8], где кровотечение, заболевание, сопровождающееся кровотечением, или заболевание, вызванное кровотечением, представляет собой заболевание, которое возникает и/или прогрессирует в результате снижения активности или дефицита фактора свертывания крови VIII и/или активированного фактора свертывания крови VIII.
[10] Композиции по п.[9], где заболевание, которое возникает и/или прогрессирует в результате снижения активности или дефицита фактора свертывания крови VIII и/или активированного фактора свертывания крови VIII, представляет собой гемофилию А.
[11] Композиции по п.[9], где заболевание, которое возникает и/или прогрессирует в результате снижения активности или дефицита фактора свертывания крови VIII и/или активированного фактора свертывания крови VIII, представляет собой заболевание, при котором вырабатывается ингибитор фактора свертывания крови VIII и/или активированного фактора свертывания крови VIII.
[12] Композиции по п.[9], где заболевание, которое возникает и/или прогрессирует в результате снижения активности или дефицита фактора свертывания крови VIII и/или активированного фактора свертывания крови VIII, представляет собой приобретенную гемофилию.
[13] Композиции по п.[9], где заболевание, которое возникает и/или прогрессирует в результате снижения активности или дефицита фактора свертывания крови VIII и/или активированного фактора свертывания крови VIII, представляет собой болезнь Виллебранда-Юргенса.
[14] Способу профилактики и/или лечения кровотечения, заболевания, сопровождающегося кровотечением, или заболевания, вызванного кровотечением, где указанный способ включает в себя стадию введения антитела по любому из п.п.[1]-[6] или композиции по любому из п.п.[7]-[13].
[15] Применению антитела по любому из п.п.[1]-[6] для получения композиции по любому из п.п.[7]-[13].
[16] Набору, используемому в способе профилактики и/или лечения заболеваний по п.[14], где указанный набор включает в себя по меньшей мере антитело по любому из п.п.[1]-[6] или композицию по п.[7].
[17] Способу профилактики и/или лечения кровотечения, заболевания, сопровождающегося кровотечением или заболевания, вызванного кровотечением, где указанный способ включает в себя стадию введения антитела по любому из п.п.[4]-[6] или композиции по любому из п.п.[7]-[13] в комбинации с фактором свертывания крови VIII.
[18] Набору, используемому в способе профилактики и/или лечения кровотечения, заболевания, сопровождающегося кровотечением, или заболевания, вызванного кровотечением, по п.[17], где указанный набор включает в себя по меньшей мере антитело по любому из п.п.[4]-[6] или композицию по п.[7] и фактор свертывания крови VIII.
Краткое описание чертежей
На фиг.1 изображена область вставки pcDNA4-g4H.
На фиг.2 изображены области вставок pcDNA4-g4L и pIND-g4L.
На фиг.3 изображена область вставки pIND-g4H.
На фиг.4 отображены результаты измерения F.VIIIa-подобного действия анти-F.IXa/анти-F.X биспецифического антитела, генерированного из анти-F.IXa антитела XB12 и анти-F.X антитела SB04, SB21, SB42, SB38, SB30, SB07, SB05, SB06, или SB34. Концентрация растворов антител составляла 10 мкг/мл (конечная концентрация 1 мкг/мл). Результатом являются девять типов биспецифических антител, которые показывают повышение F.VIIIa-подобной активности: XB12/SB04, XB12/SB21, XB12/SB42, XB12/SB38, XB12/SB30, XB12/SB07, XB12/SB05, XB12/SB06 и XB12/SB34, в порядке интенсивности действия.
На фиг.5 отображены результаты измерения F.VIIIa-подобной активности анти-F.IXa/анти-F.X биспецифического антитела, генерированного из анти-F.IXa антитела XT04 и анти-F.X антитела SB04, SB21, SB42, SB38, SB30, SB07, SB05, SB06 или SB34. Концентрация растворов антител составляла 10 мкг/мл (конечная концентрация 1 мкг/мл). В результате XT04/SB04, XT04/SB21, XT04/SB42, XT04/SB38, XT04/SB30, XT04/SB07, XT04/SB05, XT04/SB06 и XT04/SB34 показывали повышение F.VIIIa-подобной активности.
На фиг.6 отображены результаты измерения F.VIIIa-подобной активности различных концентраций XB12/SB04, для которого на фиг.4 продемонстрирована наивысшая активность. В результате для XB12/SB04 показано зависимое от концентрации повышение F.VIIIa-подобной активности.
На фиг.7 отображены результаты измерения время свертывания плазмы (APTT) в присутствии XB12/SB04, XB12/SB21, XB12/SB42, XB12/SB38, XB12/SB30, XB12/SB07, XB12/SB05, XB12/SB06 или XB12/SB34. Концентрация растворов антитела, смешанных с плазмой с дефицитом фактора F.VIII, составляла 1,7 мкг/мл для XB12/SB06 и 10 мкг/мл для остальных антител. В результате для XB12/SB04, XB12/SB21, XB12/SB42, XB12/SB38, XB12/SB30, XB12/SB07, XB12/SB05, XB12/SB06 и XB12/SB34 показан эффект укорочения времени свертывания по сравнению с отсутствием антител.
На фиг.8 отображены результаты измерения времени коагуляции плазмы (APTT) в присутствии XT04/SB04, XT04/SB21, XT04/SB42, XT04/SB38, XT04/SB30, XT04/SB07, XT04/SB05, XT04/SB06 и XT04/SB34. Концентрация растворов антител, смешанных с плазмой с дефицитом фактора F.VIII, составляла 5 мкг/мл для XT04/SB06 и 10 мкг/мл для остальных. В результате для XT04/SB04, XT04/SB21, XT04/SB42, XT04/SB38, XT04/SB30, XT04/SB07, XT04/SB05 и XT04/SB06 показан эффект укорочения времени коагуляции по сравнению с отсутствием антител. Для XT04/SB34 не выявлен эффект укорочения времени коагуляции.
На фиг.9 отображены результаты измерения времени коагуляции в присутствии различных концентраций XB12/SB04, для которого на фиг.7 и 8 показан наибольший эффект укорочения времени коагуляции (APTT). В результате для XB12/SB04 продемонстрирован зависимый от концентрации эффект укорочения времени коагуляции. Концентрация антитела на фигуре демонстрирует значения для раствора антитела, смешанного с плазмой с дефицитом фактора F.VIII.
На фиг.10 отображены результаты GST-AP вестерн-блоттинга антител SB04 или SB06, где 1), 2) и 3) представляют собой результаты реагирования считанного GST-AP с SB04, SB06 и образцом, не содержащим антитело, соответственно. Результаты свидетельствуют об обнаружении только реакции связывания SB04 с GST-AP.
На фиг.11 изображен вектор pELBGlacI. ColE1ori: область начала репликации ColE1 плазмидного ряда; f1ori: область начала репликации фага f1; lacI: кодирующая область лактозного репрессорного белка; Plac: лактозный промотор; pelBss: сигнальная последовательность белка PelB E. coli; scFv: кодирующая область одноцепочечной молекулы антитела; ген III (ген3): кодирующая область белка гена III фага f1; Ampr: ген устойчивости к ампициллину; и Sfi I: участок расщепления ферментом рестрикции Sfi I.
На фиг.12 отображены результаты измерения F.VIIIa-подобной активности с использованием культуральных супернатантов экспрессированных биспецифических антител, которые представляют собой комбинации анти-F.IXa антитела (A19, A25, A31, A38, A39, A40, A41, A44, A50, A69 или XB12) и анти-F.X антитела (B2, B5, B9, B10, B11, B12, B13, B14, B15, B16, B18, B19, B20, B21, B23, B25, B26, B27, B31, B34-1, B34-2, B35, B36, B38, B42, SB04, SB15 или SB27). Знаком "+" обозначены случаи, где F.VIIIa-подобная активность равна 0,1 или более.
На фиг.13 отображены результаты анализа коагулирующей активности плазмы, выполненного с использованием очищенных препаратов экспрессированных биспецифических антител, которые представляют собой комбинацию анти-F.IXa антитела (A19, A25, A31, A38, A39, A40, A41, A44, A50, A69 или XB12) и анти-F.X антитела (B2, B5, B9, B10, B11, B12, B13, B14, B15, B16, B18, B19, B20, B21, B23, B25, B26, B27, B31, B34-1, B34-2, B35, B36, B38, B42, SB04, SB15 или SB27). Время свертывания крови укорачивалось на 10-20 секунд ("+"), 20-40 секунд ("++"), 40-50 секунд ("+++") или 50 секунд ("++++") или более при добавлении антитела по сравнению с отсутствием антител.
На фиг.14 отображены результаты измерения времени коагуляции при различных концентрациях A44/B26, имеющего наибольший эффект укорочения времени коагуляции (APTT), что показано на фиг.13. Без добавления антитела время коагуляции составляло 113 секунд. В результате для A44/B26 продемонстрирован зависимый от концентрации эффект укорочения времени коагуляции. Концентрация антитела на фигуре демонстрирует значения для раствора антитела, смешанного с плазмой с дефицитом фактора F.VIII.
На фиг.15 отображены результаты измерения времени коагуляции при различных концентрациях A69/B26, имеющего наибольший эффект укорочения времени коагуляции (APTT), что показано на фиг.13. Без добавления антитела время коагуляции составляло 109,6 секунд. В результате для A69/B26 продемонстрирован зависимый от концентрации эффект укорочения времени коагуляции. Концентрация антитела на фигуре демонстрирует значения для раствора антитела, смешанного с плазмой с дефицитом фактора F.VIII.
На фиг.16 отображены результаты измерения времени коагуляции (APTT) при совместном присутствии A44/B26 или XB12/SB04 с фактором F.VIII. В результате при сравнении с одиночным фактором F.VIII для смешанного раствора A44/B26 или XB12/SB04 с фактором F.VIII показан эффект укорочения времени коагуляции.
На фиг.17 отображены результаты измерения времени коагуляции (APTT) в ингибирующей плазме в присутствии A44/B26 или XB12/SB04. В результате по сравнению с отсутствием антител при наличии A44/B26 или XB12/SB04 продемонстрирован эффект укорочения времени коагуляции.
На фиг.18 отображены результаты измерения времени коагуляции при различных концентрациях XB12/SB04 и гуманизированного XB12/гуманизированного SB04. Без добавления антитела время коагуляции составляло 111,3 секунд. В результате измерения для гуманизированного XB12/гуманизированного SB04 антитела выявлен эффект укорочения времени коагуляции, сходный с таковым для XB12/SB04. Концентрация антитела на фигуре демонстрирует значения для раствора антитела, смешанного с плазмой с дефицитом фактора F.VIII.
Наилучший вариант осуществления изобретения
Биспецифическое антитело по настоящему изобретению представляет собой молекулу, содержащую в себе два типа антител или фрагменты антител, имеющие специфичность для различных антигенов. Биспецифическое антитело, особым образом не ограничено, но предпочтительно является моноклональным.
Биспецифические антитела по настоящему изобретению являются предпочтительно рекомбинантными антителами, созданными с применением технологий рекомбинантных генов (см., например, Borrebaeck CAK and Larrick JW, THERAPEUTIC MONOCLONAL ANTIBODIES, Published in the United Kingdom by MACMILLAN PUBLISHERS LTD, 1990). Рекомбинантное антитело может быть получено в результате клонирования ДНК, кодирующей антитело, выделенной из клеток, продуцирующих антитело, как, например, гибридомы или сенсибилизированные лимфоциты, встраивания ДНК в соответствующий вектор и введения вектора в организм-хозяина для продукции антитела.
Кроме того, антитела по настоящему изобретению могут быть фрагментами антител или модифицированными антителами. Фрагменты антител включают в себя диатело (Db), линейное антитело, молекулы одноцепочечного антитела (далее также называемое scFv) и тому подобное. В этом документе фрагмент "Fv" означает наименьший фрагмент антитела, включающий в себя полностью сайт распознавания антигена и сайт связывания. Фрагмент "Fv" представляет собой димер (димер VH-VL), в котором вариабельная область (VH) тяжелой цепи (H) и вариабельная область (VL) легкой цепи (L) являются прочно связанными посредством нековалентной связи. Три гипервариабельных участка (CDRs) каждой вариабельной области взаимодействуют для образования рецепторной зоны на поверхности димера VH-VL. Шесть CDRs образуют рецепторную зону антитела. Однако даже одна вариабельная область (или половина Fv, которая содержит только три антиген-специфических CDR) способна распознавать антиген и связывать его, хотя ее аффинность ниже, чем аффинность цельной рецепторной зоны.
Кроме того, фрагмент Fab (также обозначаемый как (F(ab)) дополнительно содержит константную область L-цепи и константную область H-цепи (CH1). Фрагмент Fab' отличается от фрагмента Fab тем, что содержит несколько дополнительных остатков, полученных из карбоксильного конца СН1 области Н-цепи, которые включают в себя один или несколько остатков цистеина из шарнирной области антитела. Фрагмент Fab'-SH относится к фрагменту Fab', имеющему свободную тиоловую группу в одном или нескольких остатках цистеина констатной области. Фрагменты F(ab') получают расщеплением дисульфидной связи в остатках цистеина в шарнирнирном участке F(ab')2 пепсинового гидролизата. Прочие химически связанные фрагменты антитела также известны специалисту в области уровня техники.
Диатело означает фрагмент полного антитела, полученный в результате слияния генов (Holliger P et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993); EP 404,097; WO 93/11161 и т.д.). Диатело представляет собой димер, включающий в себя две пептидные цепи; в каждой полипептидной цепи вариабельная область (VL) L-цепи соединена с вариабельной областью (VH) Н-цепи той же полипептидной цепи с помощью линкера, который является слишком коротким для того, чтобы допустить спаривание между двумя областями (например, около 5 остатков). VL и VH,кодируемые на одной цепи полипептида, образуют димер, потому что они не могут образовать одноцепочечный фрагмент вариабельной области из-за наличия между ними короткого линкера. Таким образом, диатело оканчивается двумя рецепторными зонами.
Одноцепочечное антитело, или фрагмент scFv, содержит VH и VL области антитела, и эти области находятся на одиночной полипептидной цепи. В целом, полипептид Fv дополнительно содержит полипептидный линкер между областями VH и VL, так что фрагмент scFv способен формировать структуру, необходимую для связывания антигена (см. Pluckthun "The Pharmacology of Monoclonal Antibodies" Vol.113 (Rosenburg and Moore ed (Springer Verlag, New York) pp.269-315, 1994 для общих сведений по scFv). Линкеры по настоящему изобретению специальным образом не ограничены, поскольку они не ингибируют экспрессию вариабельных областей антитела, связанных с обоими концами линкера.
Биспецифическое антитело типа IgG может быть синтезировано межвидовой гибридомой (квадрогибридома), образуемой в результате слияния двух типов гибридом, которые продуцируют антитела типа IgG (Milstein C et al., Nature 1983, 305:537-540). Такое антитело также может быть синтезировано в результате введения для совместной экспрессии в клетки генов L-цепей и H-цепей, которые образуют два типа представляющих интерес IgG (всего четыре вида генов).
Однако теоретически в иммуноглобулинах G, полученных таким способом, существуют не менее десяти комбинаций H-цепей и L-цепей. Трудно очистить IgG, содержащий желаемую комбинацию H и L-цепей, от десяти различных типов IgG. Кроме того, теоретически объем представляющей интерес комбинации резко снижен, и, таким образом, необходимо использовать крупномасштабную клеточную культуру, что приводит к дополнительному увеличению себестоимости способа получения.
В этом случае в результате соответствующей замены аминокислоты (аминокислот) в области CH3 H-цепи возможно избирательно синтезировать IgG, которые имеют гетерологичную комбинацию H-цепей (Ridgway, JB et al. Protein Engineering 1996, 9:617-621, Merchant, AM et al. Nature Biotechnology 1998, 16:677-681).
Что касается L-цепей, то вариабельная область L-цепи по сравнению с вариабельной областью Н-цепи является менее разнообразной; следовательно, можно рассчитывать на получение общей L-цепи, что обеспечивает связывающие активности с двумя H-цепями. Эффективная экспрессия биспецифического IgG становится возможной в результате введения генов этой общей L-цепи и обеих Н-цепей в клетки для экспрессии IgG (Nature Biotechnology. 1998, 16, 677-681). Однако возможность того, что два типа антитела, выбранные случайным образом, будут содержать одну и ту же L-цепь, является низкой; следовательно, сложно осуществить вышеупомянутый способ на практике. В связи с этим был предложен способ выбора общей L-цепи, адаптирующий любые различные Н-цепи к проявлению высокой связывающей активности (WO 2004/065611). Н-цепь, имеющая ранее описанный вариант CH3 (Nature Biotechnology. 1998, 16, 677-681), исключительно редко секретируется в отсутствие другой Н-цепи. Используя эту особенность для стимулирования первой экспрессии правого плеча L-цепи и H-цепи и остановки экспрессии, а затем для стимулирования экспрессии левого плеча L-цепи и H-цепи, можно увеличить пропорцию экспрессированных IgG в представляющей интерес комбинации (PCT/JP2004/008585).
Биспецифическое антитело также может быть получено посредством использования химически сшитых Fab'. Биспецифический F(ab')2 может быть получен, например, посредством малеимидирования Fab', полученного из одного антитела вместе с o-PDM (орто-фениленди-малеимид), и взаимодействия продукта с Fab', полученным из другого антитела, чтобы сшитые Fab' происходили из разных антител (Keler T et al. Cancer Research 1997, 57:4008-4014). Кроме того, также известен способ химического соединения фрагментов антител, как, например, производных Fab'-тионитробензойной кислоты (TNB) и Fab'-тиола (SH) (Brennan M et al. Science. 1985, 229:81-83).
Вместо перекрестной сшивки можно использовать лейциновые застежки, полученные из Fos и Jun или вроде этого. Хотя Fos и Jun также образуют гомодимер, предпочтительно используют их гетеродимерное образование. Для получения экспрессировали Fab', соединенный с Fos лейциновыми застежками, и второй Fab', соединенный с Jun лейциновыми застежками. Посредством смешивания и взаимодействия мономерных Fab'-Fos и Fab'-Jun, преобразованных при мягких условиях, можно сформировать биспецифический F(ab')2 (Kostelny SA et al. J. of Immunology, 1992, 148:1547-53). Этот способ не ограничивается Fab' и также может быть применен с scFv, Fv и прочим.
Биспецифическое антитело также может быть получено в виде диатела. Биспецифическое диатело представляет собой гетеродимер, включающий в себя два перекрестных фрагмента scFv. Значит, биспецифическое диатело может быть получено посредством построения гетеродимера с использованием VH(A)-VL(B) и VH(B)-VL(A), которые образованы посредством соединения VH и VL, происходящих из двух типов антител: A и B, с относительно коротким линкером, длиной приблизительно 5 аминокислотных остатков (Holliger P et al. Proc. of the National Academy of Sciences of the USA. 1993, 90:6444-6448).
В этом случае конструирование представляющего интерес биспецифического диатела может быть простимулировано посредством осуществления соответствующих аминокислотных замен («ключ-замок»: Zhu Z et al. Protein Science. 1997, 6:781-788), для того, чтобы соединить два типа scFv с гибким и относительно длинным линкером, продолжительностью около 15 аминокислотных остатков (одноцепочечное диатело: Kipriyanov SM et al. J. of Molecular Biology. 1999, 293:41-56).
sc(Fv)2, который может быть получен посредством соединения двух типов scFv с гибким и относительно длинным линкером, продолжительностью около 15 аминокислотных остатков, также может стать биспецифическим антителом (Mallender WD et al. J. of Biological Chemistry, 1994, 269:199-206).
Модифицированное антитело может представлять собой, например, антитело, которое связывает разные молекулы, как, например, полиэтиленгликоль (PEG). Для модифицированных антител по настоящему изобретению связываемые вещества не ограничены. Такие модифицированные антитела могут быть получены посредством химического изменения полученных антител. Эти способы уже являются общепризнанными в данной области.
Антитела по настоящему изобретению включают в себя человеческие антитела, мышиные антитела, крысиные антитела и тому подобное, без каких-либо ограничений их происхождения, и могут представлять собой генетически измененные антитела, как, например, химерное антитело и гуманизированное антитело.
Способы получения человеческих антител являются известными, и представляющее интерес человеческое антитело может быть получено, например, посредством иммунизирования представляющим интерес антигеном трансгенных животных, имеющих весь спектр генов человеческих антител (см. WO 93/12227, WO 92/03918, WO 94/02602, WO 94/25585, WO 96/34096, WO 96/33735).
Генетически модифицированные антитела могут быть получены с помощью известных способов. В частности, например, химерное антитело включает в себя различные вариабельные области H- и L-цепей антитела иммунизированных животных и константные области H- и L-цепей человеческого антитела. Химерное антитело может быть получено посредством сшивки ДНК, кодирующей вариабельную область антитела, полученного от иммунизированных животных, с ДНК, кодирующей константную область человеческого антитела, вставки полученной ДНК в экспрессирующий вектор и введения этого рекомбинантного вектора в организм хозяина для выработки антитела.
Гуманизированное антитело представляет собой модифицированное антитело, также обозначаемое как реконструированное человеческое антитело. Гуманизированное антитело сконструировано посредством трансплантации гипервариабельного участка (CDR) антитела, полученного от иммунизированных животных, в CDR человеческого антитела. Общие технологии генной инженерии также являются известными.
В частности, последовательность ДНК, предназначенная для сшивания CDR мышиного антитела с каркасной областью (FR) человеческого антитела, синтезирована посредством ПЦР с использованием нескольких олигонуклеотидов, имеющих перекрывающиеся участки в их концевых областях. После сшивания полученной ДНК с ДНК, кодирующей константную область человеческого антитела, полученную в результате этого ДНК встраивают в вектор экспрессии и вводят в организм хозяина для выработки гуманизированного антитела (см. EP 239400 и WO 96/02576). В качестве FR человеческого антитела, связанного через CDR, выбирают антитело, способное к образованию антигенсвязывающего сайта с соответствующим гипервариабельным участком. При необходимости могут быть заменены аминокислоты каркасной области вариабельного участка антитела, для того чтобы гипервариабельный участок реконструированного человеческого антитела образовал соответствующий антигенсвязывающий центр антитела (Sato K et al, Cancer Research 1993, 53:851-856). Каркасная область может быть заменена на каркасные области, полученные из разнообразных человеческих антител (см. WO 99/51743).
Настоящее изобретение обеспечивает биспецифические антитела, функционально замещающие кофакторы, которые распознают и фермент, и его субстрат.
Кофакторы по настоящему изобретению особым образом не ограничены, при условии, что они способны воздействовать на фермент для усиления ферментативной реакции. Например, кофактор по настоящему изобретению представляет собой кофактор протеолитического фермента. Конкретными примерами кофактора протеолитического фермента являются кофакторы факторов свертывания крови и фибринолиза (F.VIII/F.VIIIa, F.V/F.Va, PZ, TM, система TM/PS), кофакторы реакций комплемента (C4b, MCP, CR1, фактор H) и тому подобные.
Следующие комбинации могут быть перечислены в качестве конкретных примеров фермента и ферментного субстрата, так же как кофакторов фермента.
(а) Кофактор фактора свертывания крови и фибринолиза (Пример 1)
Фермент: F.IXa
Субстрат: F.X
Кофактор: F.VIII/F.VIIIa
Кофактор F.VIIIa присоединяется и к F.IXa и F.X и усиливает активирование F.X посредством F.IXa. Среди биспецифических антител, которые распознают и вышеуказанный фермент F.IXa, и субстрат F.X, некоторые имеют усиливающий эффект на активирование F.X. Предполагают, что некоторые из этих антител оказывают действие, замещающее функцию кофактора F.VIII/F.VIIIa.
(b) Кофактор фактора свертывания крови и фибринолиза (пример 2)
Фермент: ZPI
Субстрат: F.X/F.Xa
Кофактор: PZ
Кофактор PZ связан с ZPI семейства серпина и активированным фактором свертывания крови X (F.Xa), для усиления игибирующего действия ZPI на F.Xa. В частности, некоторые биспецифические антитела распознают и ZPI, и F.X/F.Xa, и предполагается, что они оказывают действие, замещающее функцию PZ.
(c) Кофактор фактора свертывания крови и фибринолиза (пример 3)
Фермент: тромбин
Субстрат: TAFI
Кофактор: TM
Кофактор TM усиливает активирование TAFI посредством тромбина. В частности, некоторые биспецифические антитела, которые распознают и тромбин и TAFI, предположительно оказывают действие, замещающее функцию ТМ.
(d) Кофакторы фактора свертывания крови и фибринолиза (пример 4)
Фермент: тромбин
Субстрат: PC
Кофакторы: TM/PS
Система TM/PS усиливает активирование PC посредством тромбина. В частности, некоторые биспецифические антитела, которые распознают и тромбин, и РС, предположительно функционально замещают систему TM/PS.
(e) Кофактор фактора свертывания крови и фибринолиза (пример 5)
Фермент: F.Xa
Субстрат: протромбин
Кофактор: F.V/F.Va
Кофактор F.Va связывает и F.Xa, и протромбин для усиления активации протромбина посредством F.Xa. Среди биспецифических антител, которые распознают и вышеуказанный фермент F.Xa, и его субстрат протромбин, некоторые имеют усиливающее действие на активацию протромбина. Некоторые из этих антител предположительно имеют функцию, замещающую функцию кофактора F.V/F.Va.
(f) Кофактор реакции комплемента (пример 1)
Фермент: C1s
Субстрат: C2
Кофактор: C4b
C4b имеет стимулирующее действие C1s на расщепление C2. В частности, некоторые биспецифические антитела, которые распознают и C1s, и C2, предположительно функционально замещают C4b.
(g) Кофакторы реакции комплемента (пример 2)
Фермент: Регуляторный фактор I системы комплемента
Субстрат: C3b
Кофакторы: регуляторный фактор Н системы комплемента
Мембранный кофакторный белок (MCP), и
Рецептор комплемента 1 (CR1)
Регуляторные факторы H, MCP и CR1 системы комплемента оказывают стимулирующее действие регуляторного фактора 1 системы комплемента на деградацию C3b. В частности, среди биспецифических антител, которые распознают и регуляторный фактор 1 системы комплемента, и C3b, некоторые предположительно функционально замещают регуляторные факторы H, MCP и CR1 системы комплемента.
Среди вышеуказанных кофакторов, в частности, предпочтителен F.VIII/F.VIIIa. Не смотря на то, что F.VIII/F.VIIIa претерпевает ограниченный протеолиз такими протеолитическими ферментами, как тромбин, пока он имеет активность F.VIII/F.VIIIa, его форма не имеет значения. Более того, варианты F.VIII/F.VIIIa и F.VIII/F.VIIIa, которые были искусственно модифицированы посредством применения методик генной рекомбинации, также относятся к F.VIII/F.VIIIa, до тех пор пока они сохраняют активность кофактора F.VIII/F.VIIIa.
Способы получения биспецифических антител, которые функционально замещают кофакторы по настоящему изобретению, особым образом не ограничены и могут быть получены посредством любых способов. Например, при получении биспецифического антитела, функционально замещающего фермент А и субстрат В, и ферментом А, и субстратом B иммунизируют животное для получения антифермента А антитела и антисубстрата В антитела. Затем получают биспецифическое антитело, включающее в себя H- и L-цепи антифермента А антитела и H- и L-цепи антисубстрата В антитела. Здесь желательно получение нескольких типов каждого из антител к ферменту А и антитела к субстрату В, таким образом, чтобы эти антитела предпочтительно могли быть использованы для получения всех возможных комбинаций биспецифических антител. После получения биспецифических антител отбирают антитела с действием, замещающим функцию кофактора.
Антитела против фермента или субстрата могут быть получены с использованием способов, известных специалисту в уровне техники. Например, антитела могут быть получены в результате иммунизации животных антигенами. Антигенами для иммунизирования животных являются, например, полные антигены, имеющие иммуногенность и неполные антигены (включая гаптен), не имеющие иммуногенность. В настоящем изобретении в качестве вышеуказанного антигена (иммуногенного вещества) используют фермент, чей кофактор может быть функционально замещен антителом по настоящему изобретению, действующий как кофактор, или ферментный субстрат. В качестве животных для иммунизирования можно использовать, например, мышь, крысу, хомяка, морскую свинку, кролика, цыпленка, макаку-резус и тому подобное. Иммунизирование этих животных антигенами можно провести любыми способами, известными специалисту в уровне техники. В настоящем изобретении вариабельные области L-цепи и H-цепи антитела предпочтительно получены от иммунизированных животных или из их клеток. Эта процедура может быть выполнена специалистом в уровне техники посредством применения общеизвестных способов. Антиген-иммунизированные животные вырабатывают антитела против антигена, в частности в клетках селезенки. Таким образом, например, мРНК может быть выделена из клеток селезенки иммунизированного животного, и вариабельные области L-цепи и Н-цепи могут быть восстановлены посредством ревертазо-полимеразной цепной реакции (RT-PCR) с использованием праймеров к вариабельным областям животного.
В частности, животных иммунизируют ферментом или субстратом. Фермент и субстрат, используемые в качестве иммуногенных веществ, могут быть цельными белками или их частичными пептидами. Кроме того, в зависимости от условий в качестве иммуногенов для иммунизирования животных можно использовать кандидатный антиген, связанный с другой молекулой для получения растворимого антигена, или его фрагменты.
Клетки селезенки изолируют из селезенки иммунизированных мышей и соединяют с клетками мышиной миеломы для получения гибридом. После отбора гибридом, которые связывают соответствующие антигены, вариабельные области L-цепи и Н-цепи восстанавливают посредством RT-PCR с использованием, например, праймеров, соответствующих вариабельным областям. Также можно использовать праймеры к CDR, праймеры к каркасным областям, которые менее разнообразны CDR, или праймеры к сигнальным последовательностям и CH1 или константной области L-цепи (CL).
В качестве варианта мРНК выделяют из клеток селезенки иммунизированных животных и кДНК вариабельных областей L-цепи и Н-цепи восстанавливают посредством RT-PCR, используя праймеры к области, близлежащей к вариабельным областям. Кроме того, лимфоциты также могут быть иммунизированы in vitro и использованы для конструирования представительных библиотек scFv или Fab. Вариабельные области могут быть получены посредством концентрирования и клонирования антигенсвязывающего клона антитела, используя «пэннинг»-метод. В этом случае также может быть проведен скрининг с использованием аналогичных библиотек, сконструированных из мРНК, выделенных из моноцитов периферической крови, селезенки, миндалины и тому подобного человека или неиммунизированных животных.
Затем используют различные области для получения векторов, экспрессирующих антитело. Биспецифические антитела могут быть получены посредством введения в одну и ту же клетку вектора экспрессии антифермента антитела вектора экспрессии антисубстрата антитела и экспрессирования антител.
Антитела, проявляющие действие, замещающее функцию кофактора, могут быть отобраны, например, с использованием способов, описанных ниже.
(1) В реакционной системе, включающей в себя фермент и субстрат, отбор осуществляется с использованием в качестве показателя повышения ферментативной активности (способность расщеплять субстрат), где повышение ферментативной активности происходит в результате добавления антитела.
(2) В системе для определения или моделирования биологических функций, в которые вовлечены фермент, субстрат и кофактор (например, система для определения коагуляции плазмы), отбор осуществляется с использованием в качестве показателя интенсивности восстановления функции, где интенсивность восстановления функции является результатом добавления антитела в отсутствие кофактора.
Антитело, полученное таким образом, может быть очищено до однородности. Разделение и очистка антитела может быть осуществлена посредством способов разделения и очистки, используемых для обычных белков. Например, антитела могут быть разделены и очищены посредством соответствующим образом подобранных и комбинированных колонок для хроматографии, как, например, аффинной хроматографии, фильтрации, ультрацентрифугирования, высаливания, диализа, электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия, изоэлектрического электрофореза и прочего (Antibodies: A Laboratory Manual. Ed Harlow and David Lane, Cold Spring Harbor Laboratory, 1988), но способы этим не ограничиваются. Колонка, используемая при аффинной хроматографии, например, представляет собой протеин А-колонку, протеин G-колонку и тому подобное.
Например, когда F.VIII/F.VIIIa является заменяемым кофактором, то есть когда комбинация фермента и субстрата представляет собой факторы коагуляции плазмы и фибринолиза F.IXa и F.X, биспецифическое антитело по настоящему изобретению имеет структуру, включающую в себя вариабельную область анти-F.IXa антитела и вариабельную область анти-F.X антитела.
Биспецифические антитела по настоящему изобретению, функционально замещающие F.VIII/F.VIIIa, были получены с использованием следующего способа. Мыши были подкожно иммунизированы коммерческим фактором F.IXa или F.X. Клетки селезенки изолировали из селезенок иммунизированных мышей с повышенным титром антител и соединяли с клетками мышиной миеломы для образования гибридом. Отбирали гибридомы, которые связывали антиген F.IXa или F.X, и восстанавливали вариабельные области L-цепи и H-цепи посредством RT-PCR, с использованием праймеров к вариабельным областям. Вариабельную область L-цепи встраивали в экспрессирующий L-цепь вектор, содержащий CL, а вариабельную область Н-цепи встраивали в экспрессирующий Н-цепь вектор, содержащий константную область Н-цепи. Кроме того, мРНК выделяли из селезенок иммунизированных мышей и каждую кДНК вариабельных областей L-цепи и Н-цепи восстанавливали посредством RT-PCR, используя праймеры к соответствующей вариабельной области. Используя эти вариабельные области, конструировали scFv-представительную библиотеку фага. Клоны антигенсвязывающих антител концентрировали и клонировали, используя «пэннинг»-метод, и, используя их вариабельные области, создавали векторы экспрессии антител. Для экспрессии антитела векторы экспрессии анти-F.IXa антитела (H-цепь, L-цепь) и векторы экспрессии анти-F.X антитела (H-цепь, L-цепь) вводили в одну и ту же клетку и получали биспецифические антитела.
Биспецифические антитела, полученные таким образом, оценивали для определения их действий, для функционального замещения F.VIII/F.VIIIa (кофакторы для активирования F.X посредством F.IXa) в аналитической системе, включающей в себя F.XIa (F.IX аминоацил-тРНК-синтетаза), F.IX, F.X, синтетический субстрат (S-2222) для F.Xa и фосфолипид. По существу, основываясь на результатах, полученных в этом исследовании, в качестве биспецифического антитела, имеющего действие, функционально замещающее F.VIII/F.VIIIa, были отобраны биспецифические антитела, проявляющие в этой аналитической системе F.VIIIa-подобную активность 0,1 или более. F.VIIIa-подобная активность, упомянутая здесь, представляет собой значение, полученное в течение 30 или 60 минут, посредством вычитания значения изменения абсорбции сольвента или супернатанта культуры, которая не экспрессирует антитело, из значения изменения абсорбции раствора антитела или супернатанта культуры, экспрессирующей антитело.
Биспецифические антитела, отобранные выше, или биспецифические антитела, близкородственные им, оценивали по их способности восстанавливать коагуляцию в аналитической системе для исследования времени свертывания крови, в которой использовали человеческую плазму с дефицитом фактора F.VIII. В результате были получены биспецифические антитела, способные укорачивать время свертывания крови по сравнению тем, когда их не добавляют. Время свертывания крови, как упоминается здесь, представляет собой, как показано в примере 7, активированное частичное тромбопластиновое время, определенное с использованием человеческой плазмы с дефицитом фактора F.VIII. Среди этих биспецифических антител предпочтительные биспецифические антитела способны укорачивать время свертывания крови на 10 секунд или более, более предпочтительно на 20 секунд или более, еще более предпочтительно на 40 секунд или более и наиболее предпочтительно на 50 секунд или более.
CDR3 H-цепи антител по настоящему изобретению особым образом не ограничены, но, в частности, имеют гипервариабельную область, включающую в себя последовательность аминокислот, описанную в любой из последовательностей CDR3 H-цепи (SEQ ID NO:16, 20, 60, 64, 68, 72, 76, 80, 84, 88, 92 и 96) XB12, XT04, A19, A25, A31, A38, A39, A40, A41, A44, A50 и A69, описанных в примерах, приведенных ниже, или их функциональных аналогов, и гипервариабельную область, включающую в себя аминокислотную последовательность, описанную в любой из последовательностей CDR3 H-цепи (SEQ ID NO:24, 28, 32, 36, 40, 44, 48, 52, 56, 100, 104, 108, 112, 116, 120, 124, 128, 132, 136, 140, 144, 148, 152, 156, 160, 164, 168, 172, 176, 180, 184, 188, 192, 196, 200 и 204) в SB04, SB05, SB06, SB07, SB21, SB30, SB34, SB38, SB42, B2, B5, B9, B10, B11, B12, B13, B14, B15, B16, B18, B19, B20, B21, B23, B25, B26, B27, B31, B34-1, B34-2, B35, B36, B38, B42, SB15 и SB27, соответственно, или их функциональных эквивалентов.
Более того, конкретный пример вышеупомянутых антител предпочтительно комбинируется из антитела, имеющего гипервариабельный участок, включающий в себя любую из аминокислотных последовательностей CDR Н-цепи ХВ12, ХТ04, А19, А25, А31, А38, А39, А40, А41, А44, А50 и А69 (SEQ ID NO:14-16, 18-20, 58-60, 62-64, 66-68, 70-72, 74-76, 78-80, 82-84, 86-88, 90-92 и 94-96) или гипервариабельный участок функционально эквивалентный ему, и антитела, имеющего гипервариабельный участок, включающий в себя любую из аминокислотных последовательностей CDR Н-цепи (SEQ ID NO:22-24, 26-28, 30-32, 34-36, 38-40, 42-44, 46-48, 50-52, 54-56, 98-100, 102-104, 106-108, 110-112, 114-116, 118-120, 122-124, 126-128, 130-132, 134-136, 138-140, 142-144, 146-148, 150-152, 154-156, 158-160, 162-164, 166-168, 170-172, 174-176, 178-180, 182-184, 186-188, 190-192, 194-196, 198-200 и 202-204) в SB04, SB05, SB06, SB07, SB21, SB30, SB34, SB38, SB42, В2, В5, В9, В10, ВЦ, В12, В13, В14, В15, В16, В18, В19, В20, В21, В23, В25, В26, В27, В31, В34-1, В34-2, В35, В36, В38, В42, SB15 и SB27) или гипервариабельный участок, функционально эквивалентный ему.
Аминокислотные последовательности вариабельных областей Н-цепи ХВ12, ХТ04, А19, А25, А31, А38, А39, А40, А41, А44, А50, А69, SB04, SB05, SB06, SB07, SB21, SB30, SB34, SB38, SB42, В2, В5, В9, В10, В11, В12, В13, В14, В15, В16, В18, В19, В20, В21, В23, В25, В26, В27, В31, В34-1, В34-2, В35, В36, В38, В42, SB15 и SB27, раскрытые в настоящем изобретении, обозначены как SEQ ID N0:13, 17, 57, 61, 65, 69, 73, 77, 81, 85, 89, 93, 21, 25, 29, 33, 37, 41, 45, 49, 53, 97, 101, 105, 109, 113, 117, 121, 125, 129, 133, 137, 141, 145, 149, 153, 157, 161, 165, 169, 173, 177, 181, 185, 189, 193, 197 и 201.
Аминокислотные последовательности вариабельных областей L-цепи A44, B26, XB12 и SB04, раскрытые в настоящем изобретении, обозначены как SEQ ID NO:205, 209, 213 и 217. Последовательности CDR L-цепи A44, B26, XB12 и SB04 обозначены как SEQ ID NO:206-208, 210-212, 214-216 и 218-220. Нуклеотидные последовательности CDR H-цепи XB12, SB04, A44, и B26 обозначены как SEQ ID NO:221 (222), 223 (224), 225 (226), 233 (234), 235 (236), 237 (238), 245 (246), 247 (248), 249 (250), 257 (258), 259 (260) и 261 (262) (последовательности в скобках представляют собой аминокислотные последовательности, кодируемые соответствующими аминокислотами), а нуклеотидные последовательности CDR их L-цепи обозначены как SEQ ID NO:227 (228), 229 (230), 231 (232), 239 (240), 241 (242), 243 (244), 251 (252), 253 (254), 255 (256), 263 (264), 265 (266) и 267 (268).
Последовательности SEQ ID NO:58, 62, 66, 70, 74, 78, 82, 86, 90, 94, 98, 102, 106, 110, 114, 118, 122, 126, 130, 134, 138, 142, 146, 150, 154, 158, 162, 166, 170, 174, 178, 182, 186, 190, 194, 198, 202, 206, 210, 214, 218, 222, 228, 234, 240, 246, 252, 258 и 264 представляют CDR1.
Последовательности SEQ ID NO:59, 63, 67, 71, 75, 79, 83, 87, 91, 95, 99, 103, 107, 111, 115, 119, 123, 127, 131, 135, 139, 143, 147, 151, 155, 159, 163, 167, 171, 175, 179, 183, 187, 191, 195, 199, 203, 207, 211, 215, 219, 224, 230, 236, 242, 248, 254, 260 и 266 представляют CDR2.
Последовательности SEQ ID NO:60, 64, 68, 72, 76, 80, 84, 88, 92, 96, 100, 104, 108, 112, 116, 120, 124, 128, 132, 136, 140, 144, 148, 152, 156, 160, 164, 168, 172, 176, 180, 184, 188, 192, 196, 200, 204, 208, 212, 216, 220, 226, 232, 238, 244, 250, 256, 262 и 268 представляют CDR3.
Антитела по настоящему изобретению не ограничиваются, но предпочтительно представляют собой биспецифические антитела, объединяющие в себя антифактор IXa антитело и антифактор X антитело, которые имеют такие же эпитопы, как и вышеупомянутые антитела, или эпитопы, близкородственные им. Антитела, имеющие такой же или близкородственный эпитоп, означают здесь, например, те, которые конкурируют друг с другом за связывание антигена в конкурентном иммуноферментном анализе и тому подобном. Без ограничения, в этом компетентном иммуноферментом анализе, фактор IX/IXa или фактор X фиксируют на 96-луночном планшете MicroWell, одновременно добавляют соответствующим образом меченное антитело и определяемое антитело и определяют связанное антитело, используя метку. Эта метка не лимитирована и включает в себя щелочно-фосфатазную метку, пероксидазную метку, биотин меченый-стрептавидин связывающий фермент (щелочная фосфатаза, пероксидаза и тому подобное), флуоресцинизотиоцианат (FITC) и тому подобное. Существует перекрывание эпитопов, если наблюдается по меньшей мере 50% конкуренция, когда антитело присутствует в концентрации вплоть до 100000-кратного превышения концентрации оцениваемого антитела.
При получении полноразмерного антитела с использованием вариабельных областей, раскрытых в настоящем изобретении, константные области антитела не являются частично лимитированными и известны специалисту в уровне техники, например, можно использовать описанные в "Sequences of proteins of immunological interest, (1991), U.S. Department of Health and Human Services. Public Health Service National Institutes of Health" and "An efficient route to human bispecific IgG, (1998). Nature Biotechnology vol.16, 677-681" и тому подобное.
В одном варианте воплощения антител по настоящему изобретению антитела проявляют эффект, функционально замещающий кофакторы, и, как ожидается, становятся эффективными лекарственными средствами при лечении заболеваний, развившихся в результате снижения активности (функции) этих кофакторов. В случаях, когда кофактор, функционально замещенный антителом по настоящему изобретению, является фактором свертывания крови или фибринолиза, вышеуказанные заболевания представляют собой, например, кровотечение, заболевания, сопровождающиеся кровотечением, заболевания, развившиеся в результате кровотечения, и тому подобное. В частности известно, что снижение функции и дефицит факторов F.VIII/F.VIIIa, F.IX/F.IXa и F.XI/F.XIa являются причиной патологического кровотечения, называемого гемофилией.
Среди гемофилий патологическое кровотечение в результате наследственной гипофункции F.VIII/F.VIIIa или дефицита F.VIII/F.VIIIa называется гемофилией А. Когда у больного гемофилией А начинается кровотечение, проводят заместительную терапию препаратом фактора F.VIII. Кроме того, может осуществляться профилактический прием препарата фактора F.VIII (см. непатентные документы 1 и 2) в день силовой нагрузки или пешеходной экскурсии, когда часто случаются внутрисуставные кровотечения, или если больной классифицируется, как имеющий тяжелое течение гемофилии. Поскольку это профилактическое применение препарата фактора F.VIII заметно сокращает эпизоды кровотечения у больных гемофилией А, в последнее время оно стало широко известным. Сокращение эпизодов кровотечения не только сокращает риски смертельных и не смертельных кровотечений и сопровождающей агонии, но также предотвращает гемофилическую артропатию, являющуюся результатом частых внутрисуставных кровотечений. В результате это значительно способствует улучшению качества жизни больных гемофилией А.
Время полужизни препарата фактора F.VIII в кровотоке находится в пределах от 12 до 16 часов. Поэтому для непрерывной профилактики необходимо применять препарат фактора F.VIII приблизительно три раза в неделю. Это эквивалентно поддержанию приблизительно 1% или более активности F.VIII (см. непатентные документы 3 и 4). Также при проведении заместительных терапий в случаях кровотечения необходимо периодически принимать препараты, усиливающие фактор F.VIII какой-то период времени, за исключением тех случаев, когда кровотечение является умеренным, для профилактики повторных кровотечений и установления полной остановки кровотечения.
Кроме того, препараты фактора F.VIII вводят в организм внутривенно. Существуют технические сложности при выполнении внутривенного введения, и это еще более сложно при внутривенном введении больным молодого возраста, чьи вены тонкие.
При вышеописанном профилактическом введении препарата фактора F.VIII и его экстренном введении в случае кровотечения в большинстве случаев применяется домашнее лечение и самостоятельные инъекции препарата. Необходимость частого введения и встречающиеся технические трудности не только причиняют боль больным, но также препятствуют распространению домашнего лечения и проведению самостоятельных инъекций больным.
В этой связи существует высокий спрос по сравнению с существующими в настоящее время препаратами фактора свертывания крови VIII на лекарственные средства, имеющие более длительные интервалы между введениями, и лекарственные средства, которые могут быть легко введены в организм.
Далее, анти-F.VIII антитела, называемые ингибиторами, могут быть выработаны у больных гемофилией А, в частности у больных тяжелой формой гемофилии А. Если ингибитор выработан, эффекты препарата фактора F.VIII замедляются ингибитором. В результате больному становится очень трудно контролировать гемостаз.
При кровотечении у такого больного с выработанным ингибитором обычно осуществляют компенсаторную терапию, используя введение больших доз препарата фактора F.VIII, или альтернативную терапию, используя комплексный концентрат или препарат фактора F.VIIa. Однако применение компенсаторной терапии, введение больших доз препарата фактора F.VIII, может негативно увеличить титр ингибитора (анти-F.VIII антитело). Кроме того, относительно короткое время полужизни (около 2-8 часов) комплексных концентратов и препаратов фактора F.VIIa делает проблематичной альтернативную терапию. Более того, поскольку механизмы их действия независимы от функции F.VIII/F.VIIIa, то есть функции катализировать активирование фактора F.X посредством фактора F.IXa, механизм гемостаза может функционировать неэффективно и стать безответным. По этой причине нередко у больных гемофилией А, у которых вырабатывается ингибитор, не получены удовлетворительные гемостатические эффекты, по сравнению с больными гемофилией А, у которых ингибитор не вырабатывается.
Поэтому существует большой спрос на лекарственные средства, которые не зависят от наличия ингибиторов и которые могут функционально заместить F.VIII/F.VIIIa.
Кроме гемофилии и гемофилии, вызванной анти-F.VIII аутоантителом, также в качестве заболевания с патологическим кровотечением, ассоциированным с F.VIII/F.VIIIa, известна болезнь Виллебранда-Юргенса, развивающаяся в результате функциональной патологии или дефицита vWF. vWF необходим не только для нормальной адгезии тромбоцитов на субэндотелиальные ткани в участках повреждения сосудистой стенки, но также для образования комплексов с фактором F.VIII для поддержания нормального уровня F.VIII в плазме. У пациентов с болезнью Виллебранда-Юргенса эти функции снижены, и наблюдается патологический гомеостаз.
Принимая во внимание вышеописанное, могут быть рассмотрены способы применения антител для создания лекарственных средств, которые (i) имеют длинные интервалы введения, (ii) легко применимы, (iii) не зависят от наличия ингибиторов и (iv) могут функционально замещать факторы F.VIII/F.VIIIa в фактор F.VIII/F.VIIIa-независимой манере. В целом время полужизни антител в кровотоке относительно длинное - от нескольких дней до нескольких недель. Кроме того, известно, что антитела мигрируют в кровоток после подкожного введения. То есть лекарственные средства, содержащие антитела, в основном соответствуют описанным выше требованиям (i) и (ii).
Настоящее изобретение обеспечивает фармацевтические композиции, включающие в себя в качестве действующего ингредиента антитело по настоящему изобретению. Например, когда антитело по настоящему изобретению является одним из антител, которые распознают и F.IX/F.IXa, и F.X, и может функционально замещать F.VIIIa, это антитело, как ожидается, становится фармацевтическим препаратом (фармацевтической композицией) или лекарственным средством для профилактики или лечения кровотечения, заболеваний, сопровождающихся кровотечением, или заболеваний, вызванных кровотечением. Кроме того, когда антитело по настоящему изобретению является одним из антител, которые распознают и F.X/F.Xa, и протромбин, и может функционально замещать F.Va, это антитело, как ожидается, становится фармацевтическим препаратом (фармацевтической композицией) или лекарственным средством для профилактики или лечения кровотечения, заболеваний, сопровождающихся кровотечением, или заболеваний, вызванных кровотечением.
В то же время предполагается, что антитело, которое связывает ZPI и F.X и функционально замещает PZ, становится фармацевтическим препаратом (фармацевтической композицией) или лекарственным средством с антитромботическим действием, антитело, которое связывает тромбин и TAFI и функционально замещающее TM, становится фармацевтическим препаратом (фармацевтической композицией) или лекарственным средством со стимулирующим гемостаз эффектом, и антитело, которое связывает тромбин и PC и функционально замещающее систему PS/TM, становится фармацевтическим препаратом (фармацевтической композицией) или лекарственным средством с модифицирующим коагуляцию эффектом.
Кроме того, поскольку дефицит комплемента С4 является причиной системной красной волчанки (SLE), антитело, которое функционально замещает C4b, предположительно становится фармацевтическим препаратом (фармацевтической композицией) или лекарственным средством с эффектом, подавляющим возникновение SLE. Поскольку дефицит фактора H является причиной гнойной инфекции и аутоиммунного гломерулонефрита, антитело, функционально замещающее фактор Н, предположительно становится фармацевтическим препаратом (фармацевтической композицией) или лекарственным средством с эффектом, подавляющим начало этих заболеваний.
Для разработки фармацевтического препарата фармацевтические композиции, включающие в себя в качестве действующего ингредиента антитело по настоящему изобретению, применяемые для лечения или профилактики, при необходимости могут быть смешаны с соответствующим фармацевтически подходящим носителем, субстратом и тому подобным, которые являются инертными. Например, могут быть упомянуты стерилизованная вода или физиологический солевой раствор, антикоагулянт, наполнитель, антиоксидант (аскорбиновая кислота и тому подобное), буфер (фосфорная кислота, лимонная кислота, другие органические кислоты и тому подобное), антисептик, поверхностно-активное соединение (PEG, Tween и тому подобное), хелатообразующий агент (EDTA и тому подобное), связывающий агент и тому подобное. Фармацевтические композиции могут также содержать другие низкомолекулярные полипептиды, белки как, например, сывороточный альбумин, желатин и иммуноглобулин, аминокислоты как, например, глицин, глутамин, аспарагин, аргинин и лизин, сахара как, например, полисахарид и моносахарид и углеводы, и спирты сахаров, как, например, маннит и сорбит. При приготовлении водных растворов для инъекции, например, солюбилизирующие агенты, включая физиологический солевой раствор, изотонические растворы, содержащие глюкозу и другие добавочные агенты, как, например, D-сорбит, D-маннозу, D-маннит и хлорид натрия, могут быть использованы в комбинации с соответствующими солюбилизирующими агентами, как, например, спирт (этиловый и другие), многоатомный спирт (пропиленгликоль, PEG и другие), и неионогенное поверхностно-активное вещество (полисорбат 80, HCO-50 и другие).
Кроме того, при необходимости антитела по настоящему изобретению могут быть инкапсулированы в микрокапсулы (микрокапсулы сделаны из гидроксиметил целлюлозы, желатина, полиметилметакрилат и тому подобного) или включены в коллоидальную систему доставки лекарственного вещества к участку действия (липосома, альбуминовая микросфера, микроэмульсия, наночастица и нанокапсула и тому подобное) (см. "Remington's Pharmaceutical Science 16th edition", Oslo Ed. (1980) и тому подобное). Способы формулирования лекарств замедленного действия также известны и могут быть применены к антителам по настоящему изобретению (Langer et al., J. Biomed. Mater. Res. 15:267-277 (1981); Langer, Chemtech. 12:98-105 (1982); Патент США № 3773919; Европейская патентная заявка № (EP) 58,481; Sidman et al., Biopolymers 22:547-556 (1983); EP133988).
Антитела или фармацевтические композиции по настоящему изобретению могут быть использованы в комбинации с фактором свертывания крови VIII. Антитела или фармацевтические композиции по настоящему изобретению могут быть введены в организм одновременно с фактором свертывания крови VIII или с некоторым интервалом между их введениями. Прием лекарственного средства может осуществляться в наборе, который объединяет антитело или фармацевтическую композицию по настоящему изобретению с фактором свертывания крови VIII. Когда антитело или фармацевтическую композицию по настоящему изобретению используют в комбинации с фактором свертывания крови VIII, если желательно, также возможно применять более низкие дозы, чем когда они используются по отдельности.
Хотя дозировка фармацевтических композиций по настоящему изобретению соответствующим образом определена доктором с учетом типа композиции, способа введения, возраста и веса больных, имеющихся у больных симптомов, типа и развития болезни и тому подобного, обычно дозировки для взрослых от 0,1 до 2000 мг/день могут быть разделены на один или несколько приемов. Предпочтительны дозировки от 1 до 1000 мг/день, более предпочтительны от 5 до 500 мг/день и наиболее предпочтительны от 100 до 300 мг/день. Хотя дозировки варьируют в соответствии с весом и возрастом больных, способами введения и тому подобным, специалист в уровне техники может подобрать подходящую дозировку. Предпочтительно период дозированного введения лекарственного средства также соответствующим образом определен на основании, например, процесса излечения больного.
Кроме того, также возможно провести генотерапию посредством вставки гена, кодирующего антитело по настоящему изобретению, в генотерапевтические векторы. В качестве способа введения, помимо прямого введения в «голые» плазмиды, гены также могут быть введены посредством упаковки в липосому и тому подобное, или вставки в различные вирусные векторы, как, например, вектор ретровируса, векторы аденовирусов, векторы вируса коровьей оспы, векторы поксовируса, векторы аденоассоциированного вируса и векторы HVJ (см. Adolph "Virus Genome Method" CRC Press, Florid (1996)), или посредством напыления на сферические частицы носителя, как, например, коллоидальная золотая корпускула (WO93/17706 и другие). Тем не менее, ген может быть введен в организм посредством любых способов, пока антитело может быть экспрессировано in vivo для того, чтобы проявлять свое действие. Предпочтительно достаточная доза вводится в организм через соответствующий парентеральный путь, как, например, внутривенная, внутрибрюшинная, подкожная, внутрикожная, внутрь жировой ткани, интрамаммарная и внутримышечная инъекция и инфузия, ингаляция, способ газ-индуцированной бомбардировки частицами (с помощью электронной пушки и тому подобного), или путь введения через слизистые с использованием капель в нос. Гены, кодирующие антитело по настоящему изобретению, могут быть введены в организм посредством введения гена в клетки крови, клетки, полученные из спинного мозга и тому подобное, с использованием трансфекции липосомы ex vivo, способа бомбардировки частицами (патент США № 4945050) или вирусной инфекции, и повторным введением этих клеток в организм животных. В генотерапии может быть использован любой ген, кодирующий антитело по настоящему изобретению, например гены, включающие в себя нуклеотидные последовательности CDR вышеописанных XB12, SB04, A44 и B26.
Настоящее изобретение также обеспечивает способы профилактики и/или лечения кровотечения, заболеваний, сопровождающихся кровотечением или заболеваний, вызванных кровотечением, включающие в себя этапы применения антитела или композиции по настоящему изобретению. Антитела или композиции могут быть введены в организм, например, вышеупомянутыми способами.
Настоящее изобретение также относится к применению антител по этому изобретению для промышленного производства (фармацевтических) композиций по настоящему изобретению.
Кроме того, настоящее изобретение обеспечивает наборы, содержащие в себе, по меньшей мере, антитело или композицию по настоящему изобретению, которые используются в вышеописанных способах. Стеклянный шприц, игла для инъекции, фармацевтически подходящая среда, ватка, пропитанная спиртом, бинт, инструкция, в которой описан способ применения, и тому подобное также могут быть при желании упакованы в эти наборы.
Все документы, касающиеся предшествующего уровня техники, цитируемые в настоящей спецификации, были объединены здесь в качестве ссылок.
Далее настоящее изобретение будет специфически описано с ссылками на Примеры, но это не должно рассматриваться как ограничение изобретения.
[Пример 1] Получение не нейтрализующего антитела против фактора IXa (F.IXa)
1-1. Иммунизация и получение гибридом
Восемь мышей BALB/c (самцы, возраст на момент начала иммунизации 6 недель (Charles River, Япония)) и пять мышей MRL/lpr (самцы, возраст на момент начала иммунизации 6 недель (Charles River, Япония)) были иммунизированы человеческим фактором Ixaβ (Enzyme Research Laboratories, Inc.), как описано ниже. Первичную вакцинацию фактором IXaβ (40 мкг/особь), эмульгированным с FCA (полный адъювант Фрейнда H37 Ra (Difco laboratories)), проводили подкожно. Через две недели подкожно вводили фактор IXaβ (40 мкг/особь), эмульгированный с FIA (неполный адъювант Фрейнда H37 Ra (Difco laboratories)). Впоследствии были проведены от трех до семи бустерных иммунизаций с однонедельными интервалами. После того как с помощью метода ELISA (твердофазный иммуноферментный анализ), описанного в 1-2, было подтверждено увеличение в плазме титра антител против фактора IXaβ, в качестве окончательной вакцинации внутривенно вводили фактор IXaβ (40 мкг/особь), разведенный в PBS(-) (забуференный фосфатом физиологический раствор, не содержащий ионов кальция и ионов магния). Через три дня после проведения окончательной вакцинации у мышей удаляли селезенки. Наряду с использованием части селезенок в примере 10-2 оставшиеся клетки селезенки объединяли с клетками мышиной миеломы P3X63Ag8U.1 (называемые как P3U1, ATCC CRL-1597) посредством стандартного способа с использованием PEG1500 (Roche Diagnosticks). Объединенные клетки, суспендированные в среде RPMI1640 (Invitrogen), содержащей 10% FBS (Invitrogen) (далее называемый здесь как 10%FBS/RPMI1640), высевали в 96-луночный культуральный планшет и через 1, 2, 3 и 5 дней после слияния среду заменяли на селекционную среду HAT (10% FBS/RPMI1640/2% HAT 50-кратный концентрат (Dainippon Pharmaceutical Co. Ltd)/5% BM-Condimed H1 (Roche Diagnostics) для селективного культивирования гибридом. Используя супернатанты, собранные на восьмой или девятый день после слияния, посредством метода ELISA, описанного в 1-2, измеряли фактор IXa-связывающую активность с целью отбора гибридом, имеющих фактор IXa-связывающую активность. Впоследствии действие, нейтрализующее ферментативную активность фактора IXa, определяли с использованием способа, описанного в 1-3 с целью отбора гибридом, которые не проявляют действия, нейтрализующего фактор IXa. Гибридомы дважды клонировали посредством осуществления предельного разведения, в результате которого в каждую лунку 96-луночного культурального планшета высевали одну клетку. Одиночные клеточные колонии, подтвержденные посредством проведения микроскопического исследования, подвергали твердофазному иммуноферментному анализу и осуществляли анализ нейтрализационной активности, описанный в 1-2 и 1-3, для отбора клонов. С помощью способа, описанного в 1-4, были получены перитонеальные выпоты, содержащие клонированное антитело, и антитело очищали от перитонеальных выпотов. Очищенное антитело было неспособно удлинять АРТТ (активированное частичное тромбопластиновое время), что было подтверждено способом, описанным в 1-5.
1-2. Иммуноферментный анализ (ELISA) фактора IXa
Фактор IXaβ разводили до концентрации 1 мкг/мл буфером для сенсибилизации поверхностей (100 мМ гидрокарбонат натрия, pH 9,6, 0,02% азид натрия) и распределяли в планшет Nunc-Immuno (Nunc-ImmunoTM 96 MicroWellTM планшеты MaxiSorpTM (Nalge Nunc International)) по 100 мкл в лунку. Затем планшет инкубировали в течение ночи при 4°C. После тройной отмывки планшета PBS(-), содержащим Tween® 20, блокировали буфером для разведения (50 мМ Tris-HCl, pH 8,1, 1% бычий сывороточный альбумин, 1 мМ MgCl2, 0,15 M NaCl, 0,05% Tween® 20, 0,02% азид натрия) при комнатной температуре в течение 2 часов. После удаления буфера добавляли по 100 мкл/лунку мышиной иммунной сыворотки, разведенной буфером для разведения, или супернатанта культуры гибридомы, и инкубировали при комнатной температуре в течение 1 часа. После тройной отмывки планшета добавляли по 100 мкл/лунку меченного щелочной фосфотазой антимышиного IgG козла (H+L) (Zymed Laboratories), который был разведен до 1/2000 буфером для разведения, и инкубировали при комнатной температуре в течение 1 часа. После шестикратной отмывки планшета добавляли по 100 мкл/лунку колориметрического субстрата Blue-PhosTM Phosphate (Kirkegaad & Perry Laboratories) и инкубировали при комнатной температуре 20 минут. После добавления стоп-раствора Blue-PhosTM (Kirkegaad & Perry Laboratories) (100 мкл/лунку) измеряли оптическую плотность при 595 нм с помощью аппарата для прочтения микропланшетов Model 3550 (Bio-Rad Laboratories).
1-3. Измерение нейтрализационной активности относительно фактора IXa
Для получения раствора фосфолипида (400 мкг/мл) фосфолипид (Sigma-Aldrich) разводили в дистиллированной воде для инъекций и подвергали ультразвуковой обработке. Tris-забуференный физиологический раствор, содержащий 0,1% бычий сывороточный альбумин (далее здесь сокращаемый как TBSB) (40 мкл), 30 нг/мл фактора IXaβ (Enzyme Research Laboratories) (10 мкл), 400 мкг/мл раствор фосфолипида (5 мкл), TBSB, содержащий 100 мМ CaCl2 и20 мМ MgCl2 (5 мкл), и супернатант культуры гибридомы (10 мкл), смешивали в 96-луночном планшете и инкубировали при комнатной температуре в течение 1 часа. К этому смешанному раствору добавляли 50 мкг/мл фактор X (Enzyme Research Laboratories) (20 мкл) и 3 Ед./мл фактор VIIIa (American diagnostica) (10 мкл) и проводили реакцию при комнатной температуре в течение 30 минут. Реакцию останавливали посредством добавления 0,5 M EDTA (10 мкл). После добавления раствора S-2222 (50 мкл; Chromogenix) и инкубирования при комнатной температуре в течение 30 минут измеряли оптическую плотность при длине волны 405 нм и при эталонной длине волны 655 нм с помощью аппарата для прочтения микропланшетов Model 3550 (Bio-Rad Laboratories, Inc.).
1-4. Получение перитонеальных выпотов и очищение антитела
Перитонеальные выпоты созданных гибридом получали соответственно стандартным процедурам. А именно, гибридому культивировали in vitro (2×106) и трансплантировали в перитонеальную полость мышей BALB/c (самцы, возраст на момент начала эксперимента от 5 до 7 недель, Japan Charles River) или «голых» мышей BALB/c (самки, возраст на момент начала эксперимента от 5 до 6 недель, Japan Charles River и Japan CLEA), которым предварительно дважды внутрибрюшинно вводили пристан (2,6,10,14-тетраметилпентадекан, WAKO Pure Chemical Industries). Через одну-четыре недели после трансплантации собирали перитонеальные выпоты у мышей со вздутым животом.
Антитело очищали из перитонеальных выпотов с использованием колонки Protein G SepharoseTM 4 Fast Flow (Amersham Biosciences). Перитонеальные выпоты разводили в 2 раза буфером для связывания (20 мМ ацетат натрия, pH 5,0) и наносили на колонку, отмытую 10 объемами колонки буфера для связывания. Антитело элюировали пятью объемами колонки буфера для элюции (0,1 M глицин-HCl, pH 2,5) и нейтрализовали буфером для нейтрализации (1 M Tris-HCl, pH 9,0). Полученный раствор концентрировали, используя CentriprepTM 10 (Millipore), и сольвент заменяли на TBS (50 мМ Tris-забуференный физиологический раствор). Концентрацию антитела высчитывали исходя из значения оптической плотности при 280 нм со значением A (1%, 1 см)=13,5. Оптическую плотность определяли, используя DU-650 (Beckman Coulter).
1-5. Определение APTT (активированное частичное тромбопластиновое время)
APTT определяли, используя CR-A (Amelung)-связанный KC10A (Amelung). Смесь раствора антитела, разбавленного TBSB (50 мкл), стандартной плазмы крови человека (Dade Behring) (50 мкл) и реагента APTT (Dade Behring) (50 мкл) прогревали при 3°C в течение 3 минут. К этой смеси добавляли 20 мМ CaCl2 (Dade Behring) (50 мкл) для начала реакции коагуляции и измеряли время коагуляции.
[Пример 2] Получение кофактора X (F.X)-нейтрализующего антитела
2-1. Иммунизация и получение гибридом
Восемь мышей BALB/c (самцы, возраст на момент начала иммунизации 6 недель (Charles River, Япония)) и пять мышей MRL/lpr (самцы, возраст на момент начала иммунизации 6 недель (Charles River, Япония)) были иммунизированы человеческим фактором X (Enzyme Research Laboratories, Inc.), как описано ниже. Первичную вакцинацию фактором X (40 мкг/особь), эмульгированным с FCA, проводили подкожно. Через две недели подкожно вводили фактор X (20 или 40 мкг/особь), эмульгированный с FIA. Впоследствии были проведены от трех до семи бустерных иммунизаций с однонедельными интервалами. После того как с помощью метода ELISA, описанного в 2-2, было подтверждено увеличение в плазме титра антител против фактора X, в качестве окончательной вакцинации внутривенно вводили фактор X (20 или 40 мкг/особь), разведенный в PBS(-). Через три дня после проведения окончательной вакцинации у мышей удаляли селезенки. Наряду с использованием части селезенок в примере 10-2 оставшиеся клетки селезенки объединяли с клетками мышиной миеломы P3U1 посредством стандартного способа с использованием PEG1500. Объединенные клетки, суспендированные в среде 10% FBS/RPMI1640, высевали в 96-луночный культуральный планшет и через 1, 2, 3 и 5 дней после слияния среду заменяли на селекционную среду HAT для селективного культивирования гибридом. Используя супернатанты, собранные на восьмой или девятый день после слияния, посредством метода ELISA, описанного в 1-2, измеряли связывающую активность относительно фактора X. Отбирали гибридомы, имеющие связывающую активность относительно фактора Х, и, используя способ, описанный в 2-3, измеряли их способность нейтрализовать ферментную активность фактора Xa. Гибридомы, не имеющие нейтрализующей активности относительно фактора Ха, клонировали посредством выполнения дважды лимитирующего разведения. С помощью способа, описанного в 1-4, получали перитонеальные выпоты, содержащие клонированное антитело, и антитело очищали из перитонеальных выпотов. Очищенное антитело было неспособно продлевать APTT, что было подтверждено посредством способа, описанного в 1-5.
2-2. Иммуноферментный анализ (ELISA) фактора Х.
Фактор X разводили до концентрации 1 мкг/мл буфером для сенсибилизации поверхностей и распределяли в планшет Nunc-Immuno по 100 мкл в лунку. Затем планшет инкубировали в течение ночи при 4°C. После тройной отмывки планшета PBS(-), содержащим Tween® 20, блокировали буфером для разведения при комнатной температуре в течение 2 часов. После удаления буфера добавляли по 100 мкл/лунку мышиной иммунной сыворотки, разведенной буфером для разведения, или супернатанта культуры гибридомы и инкубировали при комнатной температуре в течение 1 часа. После тройной отмывки планшета добавляли по 100 мкл/лунку меченного щелочной фосфотазой анти-мышиного IgG козла (H+L), который был разведен до 1/2000 буфером для разведения, и инкубировали при комнатной температуре в течение 1 часа. После шестикратной отмывки планшета добавляли по 100 мкл/лунку колориметрического субстрата Blue-PhosTM Phosphate (Kirkegaad & Perry Laboratories) и инкубировали при комнатной температуре 20 минут. После добавления стоп-раствора Blue-PhosTM (Kirkegaad & Perry Laboratories) (100 мкл/лунку) измеряли оптическую плотность при 595 нм с помощью аппарата для прочтения микропланшетов Model 3550 (Bio-Rad Laboratories).
2-3. Измерение нейтрализационной активности относительно фактора Xa
Супернатант культуры гибридомы, разбавленный TBSB до 1/5 (10 мкл), смешивали с 40 мкл TBCP (TBSB, содержащий 2,78 мМ CaCl2 и 22,2 мкМ фосфолипиды (фосфатидилхолин:фосфатидилсерин =75:25, Sigma-Aldrich), содержащим 250 пг/мл фактора Xa (Enzyme Research Laboratories), и инкубировали при комнатной температуре в течение 1 часа. К этому смешанному раствору добавляли TBCP (50 мкл), содержащий протромбин (Enzyme Research Laboratories) (20 мкг/мл) и 100 нг/мл активированного фактора свертывания крови V (фактор Va (Haematologic Technologies)), и проводили реакцию при комнатной температуре в течение 10 минут. Реакцию останавливали посредством добавления 0,5 M EDTA (10 мкл). К этому реакционному раствору добавляли 1 мМ раствор S-2238 (Chromogenix) (50 мкл) и после инкубирования при комнатной температуре в течение 30 минут измеряли оптическую плотность при 405 нм с помощью аппарата для прочтения микропланшетов Model 3550 (Bio-Rad Laboratories, Inc.).
[Пример 3] Конструирование вектора экспрессии химерного биспецифического антитела
3-1. Получение из гибридом фрагментов ДНК, кодирующих вариабельную область антитела
Используя набор для выделения QIAGEN® RNeasy® Mini (QIAGEN), в соответствии со способом, описанным в инструкции по эксплуатации, общую РНК выделяли из гибридом, которые продуцируют анти-F.IXa антитело или анти-F.X антитело. Общую РНК растворяли в стерилизованной воде (40 мкл). В соответствии со способом, описанным в инструкции по эксплуатации, однонитевую кДНК синтезировали посредством RT-PCR, с использованием кДНК синтезирующей системы SuperScript (Invitrogen) с очищенной РНК (от 1 до 2 мкг) в качестве матрицы.
3-2. ПЦР амплификация вариабельной области Н-цепи антитела и секвенирование
В качестве праймеров для амплификации кДНК вариабельной области Н-цепи (VH) антитела, создавали смесь праймеров HB и смесь праймеров HF, описанные в сообщении Krebber et al. (J. Immunol. Methods 1997; 201:35-55). Используя по 0,5 мкл каждой 100 мМ смеси праймеров HB и 100 мМ смеси праймеров HF, готовили реакционный раствор (25 мкл) (раствор кДНК, полученный в 3-1 (2,5 мкл), буфер KOD-плюс (TOYOBO), 0,2 мМ dNTPs, 1,5 мМ MgCl2, 0,75 единиц ДНК-полимеразы KOD-плюс (TOYOBO)). Используя термоциклер GeneAmp PCR system 9700 (Parkin Elmer), проводили ПЦР в соответствии с продуктивностью амплификации фрагментов кДНК либо при условиях А (нагрев до 98°C - 3 минуты с последующими 32 циклами реакции (один цикл состоит: 98°C - 20 секунд 58°C - 20 секунд, и 72°C - 30 секунд)), либо при условиях B (нагрев до 94°C - 3 минуты с последующими 5 циклами реакции (один цикл состоит: 94°C - 20 секунд, 46°C - 20 секунд и 68°C - 30 секунд) и 30 циклами реакции (один цикл состоит: 94°C - 20 секунд, 58°C - 20 секунд и 72°C - 30 секунд)). После ПЦР реакционный раствор подвергали электрофорезу в 1% агарозном геле. Амплифицированные фрагменты необходимого размера (около 400 пн) очищали, используя набор QIAquick Gel Extraction (QIAGEN), в соответствии со способами, описанными в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (30 мкл). Последовательности нуклеотидов фрагментов ДНК определяли с помощью ДНК-секвенатора ABI PRISM 3100 Genetic Analyzer (Applied Biosystems), с использованием набора BigDye Terminator Cycle Sequencing (Applied Biosystems), в соответствии со способом, описанным в прилагаемой инструкции по эксплуатации. Участки последовательности, определенные этим способом, сравнительно анализировали, используя аналитическое программное обеспечение GENETYX-SV/RC Версия 6.1 (Genetyx) и отбирали ДНК с различными последовательностями.
3-3. Получение фрагментов ДНК вариабельной области антитела для клонирования
Следующую процедуру осуществляли для добавления участков расщепления рестрикционного фермента для клонирования на обоих концах амплифицированных фрагментов вариабельной области антитела.
Для амплификации фрагментов VH, дополненных участком для расщепления Sfi I (Sfi I-VH), синтезировали праймер (праймер VH-5'), в котором (Gly4Ser)2-линкерная последовательность праймера HB заменена на последовательность, содержащую в себе участок расщепления для рестриктазы Sfi I (SEQ ID NO:5). Используя по 0,5 мкл каждого 10 мкМ сиквенс-специфического праймера VH-5' и 10 мкМ праймера scfor (J. Immunol. Methods 1997; 201:35-55), готовили реакционный раствор (20 мкл) (очищенный раствор амплифицированного фрагмента кДНК VH, полученный в 3-2 (1 мкл), буфер KOD-плюс (TOYOBO), 0,2 мМ dNTPs, 1,5 мМ MgCl2, 0,5 единиц ДНК-полимеразы KOD-плюс (TOYOBO)). Используя термоциклер GeneAmp PCR system 9700 (Parkin Elmer), проводили ПЦР в соответствии с продуктивностью амплификации фрагментов кДНК либо при условиях А (нагрев до 98°C - 3 минуты с последующими 32 циклами реакции (один цикл состоит: 98°C - 20 секунд, 58°C - 20 секунд и 72°C - 30 секунд)), либо при условиях B (нагрев до 94°C - 3 минуты с последующими 5 циклами реакции (один цикл состоит: 94°C - 20 секунд, 46°C - 20 секунд и 68°C - 30 секунд) и 30 циклами реакции (один цикл состоит: 94°C - 20 секунд, 58°C - 20 секунд и 72°C - 30 секунд)). После ПЦР реакционный раствор подвергали электрофорезу в 1% агарозном геле. Амплифицированные фрагменты необходимого размера (около 400 пн) очищали, используя набор QIAquick Gel Extraction (QIAGEN), в соответствии со способами, описанными в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (30 мкл).
Для амплификации фрагментов ДНК вариабельной области L-цепи (VL) мышиного антитела сначала использовали по 0,5 мкл каждой 100 мкМ смеси праймеров LB и 100 мкМ смеси праймеров LF, описанных в сообщении Krebber et al. (J. Immunol. Methods 1997; 201: 35-55), и готовили реакционный раствор (25 мкл) (раствор кДНК, полученный в 3-1 (2,5 мкл), буфер KOD-плюс (TOYOBO), 0,2 мМ dNTPs, 1,5 мМ MgCl2, 0,75 единиц ДНК-полимеразы KOD-плюс (TOYOBO)). Используя термоциклер GeneAmp PCR system 9700 (Parkin Elmer), проводили ПЦР в соответствии с продуктивностью амплификации фрагментов кДНК, при условиях: нагрев до 94°C - 3 минуты с последующими 5 циклами реакции (один цикл состоит: 94°C - 20 секунд, 46°C - 20 секунд и 68°C - 30 секунд) и 30 циклами реакции (один цикл состоит: 94°C - 20 секунд, 58°C - 20 секунд и 72°C - 30 секунд)). После ПЦР реакционный раствор подвергали электрофорезу в 1% агарозном геле. Амплифицированные фрагменты необходимого размера (около 400 пн) очищали, используя набор QIAquick Gel Extraction (QIAGEN), в соответствии со способами, описанными в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (30 мкл). Фрагменты находятся в таком состоянии, при котором (Gly4Ser)3-линкерная последовательность, производная праймера LF, добавлена к их С-концам. Для того чтобы добавить участок расщепления Sfi I к С-концу фрагментов, был синтезирован праймер (праймер VL-3'), в котором (Gly4Ser)3-линкерная последовательность праймера LF заменена на последовательность, имеющую участок расщепления Sfi I (SEQ ID NO:6). Для амплификации фрагментов VL, дополненных участком для расщепления Sfi I (Sfi I-VL), использовали по 0,5 мкл каждой 10 мкМ смеси праймеров VL-3' 10 мкМ праймера scback и готовили реакционную смесь (20 мкл) (очищенный раствор амплифицированного фрагмента кДНК VL (1 мкл), буфер KOD-плюс (TOYOBO), 0,2 мМ dNTPs, 1,5 мМ MgCl2, 0,5 единиц ДНК-полимеразы KOD-плюс (TOYOBO)). Используя термоциклер GeneAmp PCR system 9700 (Parkin Elmer), проводили ПЦР в соответствии с продуктивностью амплификации фрагментов кДНК, при условиях: нагрев до 94°C - 3 минуты с последующими 5 циклами реакции (один цикл состоит: 94°C - 20 секунд, 46°C - 20 секунд и 68°C - 30 секунд) и 30 циклами реакции (один цикл состоит: 94°C - 20 секунд, 58°C - 20 секунд и 72°C - 30 секунд)). После ПЦР реакционный раствор подвергали электрофорезу в 1% агарозном геле. Амплифицированные фрагменты необходимого размера (около 400 пн) очищали, используя набор QIAquick Gel Extraction (QIAGEN), в соответствии со способами, описанными в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (30 мкл).
Очищенные фрагменты Sfi I-VH и Sfi I-VL подвергали расщеплению рестриктазой Sfi I (Takara Bio) в реакционном растворе, приготовленном в соответствие со способом, описанным в прилагаемой инструкции по эксплуатации, при 50°C в течение ночи. Затем реакционный раствор очищали, используя набор QIAquick PCR Purification (QIAGEN), посредством способа, описанного в прилагаемой инструкции по эксплуатации, и элюировали буфером EB (30 мкл), входящим в набор.
3-4. Плазмида, экспрессирующая биспецифическое IgG антитело
Для получения представляющего интерес биспецифического IgG антитела технология «ключ-замок» IgG1 (Ridgway et al., Protein Eng. 1996; 9:617-621) относилась к технологиям, которые использовали при получении IgG4 с замещенным СН3 участком аминокислоты для образования гетеромолекул для каждой Н-цепи. В типе а (IgG4γa) произведены замены Y349C и T366W, а в типе b (IgG4γb) произведены замены E356C, T366S, L368A и Y407V. Далее замена (-ppcpScp- → -ppcpPcp-) также была введена в шарнирные области обоих типов. С использованием этой методики почти все H-цепи превращаются в гетеромолекулы; однако нет необходимости применять ее к L-цепям, и образование излишних молекул антител может влиять на последующие измерения активности. Следовательно, для раздельной экспрессии плечей каждой молекулы антитела (называемой HL молекулой), которые имеют различную специфичность и эффективно образуют тип представляющего интерес биспецифического IgG антитела в клетках, в качестве векторов экспрессии для каждой HL молекулы использовали те, которые индуцируются под действием различных лекарственных средств.
В качестве вектора экспрессии для плеча молекулы антитела (для удобства называемого правым плечом HL молекулы) получали pcDNA4-g4H или pcDNA4-g4L (фиг.1 или фиг.2), в которых соответствующий участок Н-цепи или L-цепи, то есть соответствующую вариабельную область мышиного антитела (VH или VL) и константную область человеческого IgG4γa (SEQ ID NO:7) или константная область κ (SEQ ID NO:8), встраивали в вектор pcDNA4 (Invitrogen) тетрациклин-индуцибельного типа ниже сигнальной последовательности (IL3ss) для клеток животных (Proc. Natl. Acad. Sci. USA. 1984; 81:1075). Сначала использовали Eco RV и Not I (Takara Bio) для расщепления pcDNA4 в сайтах расщепления ферментами рестрикции, которые находятся в его участке мультиклонирования. Правое плечо единицы экспрессии H-цепи или L-цепи (около 1,6 т.п.н. или около 1,0 т.п.н. соответственно) химерного биспецифического антитела, имеющего соответствующие вариабельные области антитела, расщепляли с помощью Xho I (Takara Bio). Затем его очищали с использованием набора QIAquick PCR Purification (QIAGEN), с помощью способа, описанного в прилагаемой инструкции по эксплуатации, и проводили реакцию с ДНК-полимеразой KOD (TOYOBO) при 72°C в течение 10 минут в реакционном растворе, описанном в прилагаемой инструкции по эксплуатации, для образования слепых концов. Фрагменты со слепыми концами очищали с использованием набора QIAquick PCR Purification (QIAGEN) посредством способа, описанного в прилагаемой инструкции по эксплуатации, расщепляли Not I (Takara Bio). Not I/слепые концы фрагменты (около 1,6 т.п.н. или 1,0 т.п.н., соответственно) и pcDNA4, расщепленные Eco RV/Not I, подвергали реакции лигирования с использованием Ligation High (TOYOBO) в соответствии со способом, описанным в прилагаемой инструкции по эксплуатации. Штамм DH5α E. coli (Competent high DH5α (TOYOBO)) трансформировали посредством вышеописанного реакционного раствора. Из полученных таким образом клонов, устойчивых к ампициллину, изолировали соответствующие плазмидные ДНК с использованием набора QIAprep Spin Miniprep (QIAGEN).
В качестве вектора экспрессии другого плеча молекулы антитела (для удобства называемое левым плечом HL молекулы) получали pIND-g4H или pIND-g4L (фиг.2 или фиг.3) в соответствии с вышеописанным способом, в которых соответствующую область H-цепи или L-цепи, то есть соответствующую вариабельную область мышиного антитела (VH или VL) и константную область человеческого IgG4γb (SEQ ID NO:9) или константную область κ (SEQ ID NO:8), встраивали в индуцируемый аналогом экдизона тип вектора pIND (Invitrogen) ниже сигнальной последовательности (IL3ss) для клеток животного (EMBO. J. 1987; 6:2939) и изолировали соответствующие плазмидные ДНК
3-5. Конструирование вектора экспрессии биспецифического антитела
Тетрациклин-индуцибельный тип экспрессионной плазмиды, полученный в 3-4 (pcDNA4-g4H или pcDNA4-g4L), расщепляли рестриктазой Sfi I, а затем подвергали электрофорезу в 1% агарозном геле. Фрагменты (около 5 т.п.н.), лишенные характерной части вариабельной области антитела (VH или VL (см. фиг.1 или фиг.2)), очищали, используя набор QIAquick Gel Extraction (QIAGEN), посредством способа, описанного в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (30 мкл). Эти фрагменты и соответствующий фрагмент Sfi I-VH или Sfi-VL, полученный из обработанного рестриктазой Sfi I анти-F.IXa антитела, полученного в 3-3, подвергали лигированию, используя набор Quick Ligation (New England Biolabs), в соответствии со способом, описанным в прилагаемой инструкции по эксплуатации. Штамм DH5α E. coli (Competent high DH5α (TOYOBO)) был трасформирован вышеописанным реакционным раствором. Далее фрагменты, полученные путем удаления участка вариабельной области антитела посредством схожей описанной выше технологии (VH или VL (см. фиг.2 или фиг.33)) из расщепленного Sfi I индуцированного аналогом экдизона типа экспрессионной плазмиды (pIND-g4H или pIND-4GL), полученного в 3-4, и соответствующий фрагмент Sfi I-VH или Sfi I-VL, полученный в результате обработки рестриктазой Sfi I анти-F.X антитела, объединяли посредством схожего способа.
В каждом из полученных таким способом трансформантов, устойчивых к ампициллину, вставку представляющего интерес фрагмента подтверждали посредством способа ПЦР колоний с использованием праймеров, которые образуют сэндвич-структуру с вставленным фрагментом. Сначала для вектора экспрессии химерной H-цепи или L-цепи анти-F.IXa антитела синтезировали 21-мерный праймер CMVF (SEQ ID NO:10), который отжигается на прямом участке прайминга CMV выше участка вставки, и 18-мерный праймер BGHR (SEQ ID NO:11), который отжигается на обратном участке прайминга BGH ниже участка вставки (Sigma Genoys). Для вектора экспрессии химерной H-цепи или L-цепи анти-F.X антитела синтезировали 24-мерный праймер EcdF (SEQ ID NO:12), который отжигается выше участка вставки, и 18-мерный праймер BGHR(SEQ ID NO:11), который отжигается на обратном участке прайминга BGH ниже участка вставки (Sigma Genoys). Для ПЦР колоний готовили реакционный раствор (20 мкл) (0,2 мкл праймера (10 мкМ), буфер KOD dash (TOYOBO), 0,2 мМ dNTPs и 0,75 единиц ДНК-полимеразы KOD dash) (TOYOBO)). В этот реакционный раствор добавляли клетки трансформированного штамма в соответствующем количестве и проводили ПЦР. ПЦР проводили с использованием термоциклера GeneAmp PCR system 9700 (Parkin Elmer) при следующих условиях: нагревание при 96°C - 1 минута, с последующими 30 циклами реакции (один цикл состоит: 96°C - 10 секунд, 55°C - 10 секунд и 72°C - 30 секунд). После ПЦР реакционный раствор подвергали электрофорезу в 1% агарозном геле и отбирали клоны амплифицированных фрагментов нужного размера. ПЦР-продукт обрабатывали ExoSAP-IT (Amersham Biosciences) для инактивирования избытка праймеров и dNTPs в соответствии с прилагаемой инструкцией по эксплуатации. Последовательности нуклеотидов фрагментов ДНК определяли посредством ДНК-секвенатора ABI PRISM 3100 Genetic Analyzer (Applied Biosystems) с использованием набора BigDye Terminator Cycle Sequencing (Applied Biosystems) в соответствии со способом, описанным в прилагаемой инструкции по эксплуатации. Участки последовательности, определенные этим способом, сравнительно анализировали, используя аналитическое программное обеспечение GENETYX-SV/RC Версия 6.1 (Genetyx). Для VH были отобраны представляющие интерес клоны, не имеющие вставок, делеций или мутаций. Для VL, отличной от используемого в гибридомах P3U1-производного псевдо VL гена, были отобраны представляющие интерес клоны, не имеющие вставки, делеции или мутации.
Из представляющих интерес клонов соответствующие плазмидные ДНК были выделены с использованием набора QIAprep Spin Miniprep (QIAGEN) и затем растворены в стерилизованной воде (100 мкл). Вектор экспрессии химерной Н-цепи анти-F.IXa антитела, вектор экспрессии химерной L-цепи анти-F.IXa антитела, вектор экспрессии химерной Н-цепи анти-F.X антитела, вектор экспрессии химерной L-цепи анти-F.X антитела были обозначены как pcDNA4-g4IXaHn, pcDNA4-g4IXaLn, pIND-g4XHn и pIND-g4XLn соответственно. Каждый раствор плазмиды до применения хранили при 4°C.
[Пример 4] Экспрессия химер биспецифических антител в клетках животного
4-1. Приготовление растворов ДНК
Экспрессию векторов экспрессии правого плеча молекулы HL (pcDNA4-g4IXaHn и pcDNA4-g4IXaLn) индуцировали тетрациклином. В отсутствие тетрациклина Tet-репрессор кодирующая плазмида pcDNA6/TR (Invitrogen) необходима для полной супрессии их экспрессии. Далее экспрессию векторов экспрессии левого плеча молекулы HL (pIND-g4XHn и pIND-g4XLn) индуцировали аналогом экдизона насекомого (понастерон А). Для этого необходима плазмида pVgRXR (Invitrogen), которая кодирует рецептор экдизона и ретиноид Х-рецептор, которые, реагируя с понастероном А, индуцируют экспрессию. В связи с этим для трансфекции в клетки животного была приготовлена смесь в совокупности шести типов плазмидных ДНК. На 1 мл клеточной культуры использовали pcDNA4-g4IXaHn, pcDNA4-g4IXaLn, pIND-g4XHn и pIND-g4XLn (218,8 нг каждой), а также pcDNA6/TR и pVgRXR (1312,5 нг каждой).
4-2. Трансфекция в клетки животного
Штамм HEK293H клеток эмбриональной почечной карциномы человека (Invitrogen) суспендировали в среде для культивирования DMEM (Invitrogen), содержащей 10% FCS (MOREGATE), и по 1 мл этой суспензии при плотности клеток 5×105 клеток/мл высевали в каждую лунку 12-луночного планшета для адгезии клеток (CORNING) и культивировали в CO2-инкубаторе (37°C, 5% CO2). Смесь плазмидных ДНК, полученную в 4-1, добавляли к смеси реагентов для трансфекции, Lipofectaine 2000 (Invitrogen) (7 мкл) и среде Opti-MEM I (Invitrogen) (250 мкл) и оставляли при комнатной температуре в течение 20 минут. Полученную смесь добавляли к клеткам в каждую лунку и инкубировали от 4 до 5 часов в CO2-инкубаторе (37°C, 5% CO2).
4-3. Индукция экспрессии биспецифического IgG антитела
Среду для культивирования удаляли путем аспирации из трансфицированной клеточной культуры, описанной выше, а затем добавляли 1 мл среды CHO-S-SFM-II (Invitrogen), содержащей 1 мкг/мл тетрациклина (Wako Pure Chemical Industries). Полученную смесь инкубировали в течение одного дня в CO2-инкубаторе (37°C, 5% CO2) для индукции первичной экспрессии правого плеча HL молекулы антитела. Затем после удаления среды посредством аспирации, отмывки 1 мл среды CHO-S-SFM-II и добавления 1 мл среды CHO-S-SFM-II, содержащей 5 мкМ понастерона А (Invitrogen), смесь инкубировали в CO2-инкубаторе (37°C, 5% CO2) в течение 2-3 дней, и была осуществлена индукция вторичной экспрессии левого плеча HL молекулы антитела, таким образом биспецифическое IgG антитело было выделено в среду. Культуральный супернатант собирали и центрифугировали (около 2000 г, 5 минут, комнатная температура) для удаления клеток и, если необходимо, концентрировали, используя Microcon® YM-50 (Millipore). Образец хранили при 4°C до использования.
[Пример 5] Количественное определение концентрации человеческого IgG
Концентрацию очищенного аффинного антитела козла к человеческому IgG Fc (Cappel) доводили буфером для сенсибилизации поверхностей до 1 мкг/мл и фиксировали на планшете Nunc-Immuno. После блокирования буфером для разбавления (D.B.) добавляли образец культурального супернатанта, соответствующим образом разбавленного буфером для разбавления. Далее в качестве стандарта для подсчета концентрации антитела аналогичным образом добавляли человеческий IgG4 (гуманизированное анти-TF антитело, см. WO 99/51743), разбавленный буфером для разбавления в серии двукратного разбавления до 11 стадий, начинающихся с 1000 нг/мл. После трехкратной отмывки щелочная фосфатаза козла против человеческого IgG (Biosource International) вступала в реакцию. После отмывки 5 раз, используя в качестве субстрата Sigma 104® фосфатазный субстрат (Sigma-Aldrich), в планшете сформировали цвет и измеряли оптическую плотность при 405 нм с помощью аппарата для прочтения микропланшетов Model 3550 (Bio-Rad Laboratories, Inc.) с эталонной длиной волны 655 нм. Используя программное обеспечение Microplate Manager III (Bio-Rad Laboratories), на основании калибровочной кривой подсчитывали концентрацию человеческого IgG в культуральном супернатанте.
[Пример 6] Исследование фактор F.VIIIa (активированный фактор свертывания крови VIII)-подобной активности
F.VIIIa-подобную активность биспецифического антитела определяли посредством следующего ферментативного анализа. Все следующие реакции проводили при комнатной температуре. Смесь 40 мкл фактора IX (3,75 мкг/мл; Enzyme Research Laboratories) и 10 мкл раствора антитела инкубировали в 96-луночном планшете в течение 1 часа. Затем для инициации ферментативной реакции добавляли 10 мкл фактора XIa (10 нг/мл; Enzyme Research Laboratories), 20 мкл фактора X (50 мкг/мл; Enzyme Research Laboratories), 5 мкл фосфолипида (400 мкг/мл; см. примеры 1-3) и 15 мкл TBSB, содержащего 5 мМ CaCl2 и 1 мМ MgCl2 (далее сокращенный здесь как TBSB-S). Через 30 минут эту реакцию останавливали посредством добавления 10 мкл 0,5 М EDTA.
После добавления в каждую лунку раствора колориметрического субстрата (50 мкл) измеряли оптическую плотность при 405 нм (эталонная длина волны 655 нм) на 0 минуте и на 30 минуте с помощью аппарата для прочтения микропланшетов Model 3550 (Bio-Rad Laboratories). F.VIIIa-подобная активность представлена значением, полученным в результате вычитания значения изменения оптической плотности через 30 минут без добавления антитела из значения изменения оптической плотности через 30 минут с добавлением антитела (см. фиг.4 и фиг.5).
В качестве сольвента для фосфолипидов использовали TBSB, в то время как TBSB-S использовали в качестве сольвента для фактора XIa, фактора IX и фактора X. Раствор колориметрического субстрата представлял собой смесь 1:1 колориметрического субстрата S-2222 "Tesutochimu" (Chromogenix), разведенного в соответствии с прилагаемой инструкцией по эксплуатации, и раствора полибрена (0,6 мг/л гексадиметрин-бромид (Sigma)).
Далее измеряли концентрационную зависимость F.VIIIa-подобной активности XB12/SB04, которая была самой высокой (фиг.6).
[Пример 7] Анализ коагулирующей активности плазмы
Для выявления того, корректирует ли биспецифическое антитело коагулирующую способность при гемофилии А, исследовали действие биспецифического антитела на активированное частичное тромбопластиновое время (APTT), используя плазму с дефицитом фактора F.VIII. Смешанный раствор, включающий в себя раствор антитела в различных концентрациях (50 мкл), плазму с дефицитом фактора F.VIII (50 мкл; Biomerieux) и реагент APTT (50 мкл; Dade Behring), прогревали при 37°C в течение 3 минут. Реакцию коагуляции инициировали посредством добавления к вышеописанной смеси 20 мМ CaCl2 (50 мкл; Dade Behring). Время, необходимое для коагуляции, определяли с помощью CR-A (Amelung)-связанного KC10A (Amelung) (фиг.7 и фиг.8).
Далее для XB12/SB04, чья активность, уменьшающая время коагуляции, была наибольшей, определяли его концентрационную зависимость (фиг.9).
[Пример 8] Очищение антитела
Культуральный супернатант (10 мл), полученный способом, описанным в примере 4, концентрировали до 1 мл, используя Centricon® YM-50 (Millipore). К этому концентрату добавляли 10% BSA (10 мкл), 1% Tween® 20 (10 мкл) и rProtein A SepharoseTM Fast Flow (Amersham Biosciences) (100 мкл) и этот раствор смешивали путем переворачивания в течение ночи при 4°C. Раствор переносили в 0,22 мкм чашку фильтра Ultrafree®-MC (Millipore) и после тройной отмывки TBS, содержащим 0,01% Tween® 20 (500 мкл), смолу rProtein A SepharoseTM суспендировали в 100 мкл 10 мМ HCl/0,01% Tween® 20 (pH 2,0) и оставляли стоять в течение 3 минут. Затем элюировали антитело, и элюат был незамедлительно нейтрализован добавлением 5 мкл 1 M Tris-HCl, pH 8,0. Используя программное обеспечение Microplate Manager III (Bio-Rad Laboratories), на основании калибровочной кривой подсчитывали концентрацию человеческого IgG. Концентрацию антитела количественно определяли в соответствии с примером 5.
[Пример 9] GST-AP вестерн-блоттинг анти-F.X антитела
Конструировали рекомбинантную E. coli, экспрессирующую пептидилированный белок активированного пептида F.X (AP) с глутатион-S-трансферазой (GST). кДНК, охватывающая всю область трансляции человеческого F.X, была амплифицирована из кДНК печени человека Marathon-Ready (Clontech). Эту кДНК затем использовали в качестве матрицы для амплификации посредством ПЦР области, кодирующей область AP (Leytus et al., Biochemistry 1986; 25:5098), которая была субклонирована в вектор pGEM-T (Promega) для получения GST-AP-кодирующего pGEX-F10AP. E. coli, трансформированную этой плазмидой, культивировали и при OD=0,8 1 мМ IPTG добавляли для индукции экспрессии GST-AP. После центрифугирования культурального раствора (3000×g, 30 минут, 4°C) клетки собирали и хранили при -20°C до использования.
После ресуспендирования дебриса в объеме PBS, составляющем 1/20 объема культуры, на каждый 0,1 мл суспензии добавляли 2,4 мл буфера для образца при SDS-PAGE (IWAKI) и полученную смесь кипятили при 95°C в течение 5 минут. Этот реакционный раствор (10 мкл) добавляли в каждую лунку 14% SDS-PAGE мини-геля (Asahi Technoglass) и подвергали электрофорезу. После электрофореза гель переносили на мембрану Immobilon-PTM Transfer Membrane (MILLIPORE), используя полусухой блоттер (BIO-RAD), и блокировали посредством BT-PBS (PBS, содержащий 2% BSA и 0,05% Tween® 20). После полного блокирования в течение 1 часа проводили реакцию с мышиным анти-F.X антителом SB04 или SB06, очищенным в примерах 1-4, и разбавляли BT-PBS до 2 мкг/мл. После отмывка PBS, содержащим 0,05% Tween® 20, мембрану в течение 1 часа подвергали взаимодействию с меченным щелочной фосфатазой козла антимышиным IgG (H+L) (Zymed Laboratories), разбавленным в 2000 раз BT-PBS. После отмывки PBS, содержащим 0,05% Tween® 20, на мембране сформировали цвет посредством колориметрического субстрата BCIP/NBT Phosphatase (Kirkegaad & Perry Laboratories) (см. фиг.10).
[Пример 10] Получение биспецифического антитела из scFv библиотеки из селезенок иммунизированных мышей
10-1. Антиген и иммунизация
Три мыши BALB/c (самцы, возраст на момент начала иммунизации 6 недель (Japan Charles River)), 3 мыши MRL/lpr (самцы, возраст на момент начала иммунизации 6 недель (Japan Charles River)) и 3 мыши C57BL/6N (самцы, возраст на момент начала иммунизации 6 недель (Japan Charles River)) были иммунизированы антигенным фактором IXaβ (Enzyme Research Laboratories, Inc.) или фактором X (Enzyme Research Laboratories, Inc.), как описано ниже. В качестве первичной иммунизации подкожно вводили антиген (40 мкг/особь), эмульгированный с FCA (полный адъювант Фрейнда H37 Ra; Difco laboratories)). Через две недели подкожно вводили антиген (40 мкг/особь), эмульгированный с FIA (неполный адъювант Фрейнда; Difco laboratories)). Впоследствии с интервалом одну неделю проводили три бустерные иммунизации и через 8 дней после окончательной иммунизации мышам удалили селезенки.
10-2. Конструирование фаговой библиотеки
Часть селезенок, удаленных у иммунизированных мышей, полученных в примерах 1-1 и 2-1, и селезенки, удаленные у иммунизированных мышей, полученных в примере 10-1, помещали в тризоловый реагент (Invitrogen) (50 мг селезенки/мл реагента) и гомогенизировали, используя стеклянный гомогенизатор. Затем выделяли общую РНК, используя способ, описанный в прилагаемой инструкции по эксплуатации. polyA(+)РНК выделяли из экстрагирующего раствора, используя набор PolyATract System 1000 (Promega), в соответствии со способом, описанным в прилагаемой инструкции по эксплуатации. кДНК синтезировали посредством RT-PCR (SuperScript III First-Strand Synthesis, Invitrogen) и хранили до использования при -20°C.
В качестве праймеров для амплификации кДНК вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL) мышиного антитела готовили смесь праймеров HB, смесь праймеров HF, смесь праймеров LB и смесь праймеров LF, использованных в примерах 3-2 и 3-3. Для амплификации VH готовили 50 мкл реакционного раствора (2,5 мкл раствора кДНК, буфер KOD-плюс (TOYOBO), 0,2 мМ dNTPs, 1,5 мМ MgCl2, 3,75 единиц ДНК-полимеразы KOD-плюс (TOYOBO)), используя по 1 мкл каждой 100 мкМ смеси праймеров HB и 100 мкМ смеси праймеров HF. Далее для амплификации VL готовили 50 мкл реакционного раствора такого же состава, как описано выше, используя по 1 мкл каждой 100 мкМ смеси праймеров LB и 100 мкМ смеси праймеров LF. ПЦР осуществляли, используя термоциклер GeneAmp PCR system 9700 (Parkin Elmer), посредством нагревания при 98°C в течение 3 минут с последующими 32 циклами реакции (один цикл: 98°C - 20 секунд, 58°C - 20 секунд и 72°C - 30 секунд). После ПЦР реакционный раствор подвергали электрофорезу в 2% агарозном геле. Амплифицированные фрагменты представляющего интерес размера (около 400 п.н.) очищали, используя набор QIAquick Gel Extraction (QIAGEN), посредством способа, описанного в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (50 мкл). Далее для амплификации фрагментов scFv подготавливали десять пробирок со 100 мкл реакционного раствора (3 мкл раствора фрагмента VH, 3 мкл раствора фрагмента VL, буфер KOD-плюс (TOYOBO), 0,2 мМ dNTPs, 1 мМ MgCl2, 5 единиц ДНК-полимеразы KOD-плюс (TOYOBO)). После первой ПЦР (3-минутное нагревание при 94°C с последующими 7 циклами реакции (один цикл: 94°C - 1 минута и 63°C - 4 минуты)) в каждую пробирку с поддерживающейся температурой 63°C добавляли 10 мкМ праймер scfor и 10 мкМ праймер scback (по 2,5 мкл каждого), а затем осуществляли вторую ПЦР (35-секундное нагревание при 94°C с последующими 30 циклами реакции (один цикл: 94°C - 2 минуты и 63°C - 2 минуты)). После ПЦР реакционный раствор очищали, используя набор QIAquick PCR purification (QIAGEN), и очищенные продукты подвергали расщеплению рестриктазой Sfi I (Takara Bio) при 50°C в течение ночи. После подвергания рестриктов электрофорезу в 2% агарозном геле амплифицированные фрагменты с представляющим интерес размером (около 800 п.н.) были очищены с использованием набора QIAquick Gel Extraction (QIAGEN) с помощью способа, описанного в прилагаемой инструкции по эксплуатации, и элюированы соответствующим объемом стерилизованной воды. Для обеспечения scFv на белке гена III фага в качестве фагемидного вектора использовали pELBGlacI (см. фиг.11). После расщепления вектора (10 мкг) рестриктазой Sfi I (Takara Bio) при 50°C в течение ночи фрагменты рестрикции представляющего интерес размера (около 5 т.п.н.) были очищены с использованием набора QIAquick Gel Extraction (QIAGEN) с помощью способа, описанного в прилагаемой инструкции по эксплуатации, и элюированы соответствующим объемом стерилизованной воды. Очищенный ПЦР-продукт и очищенный фрагмент вектора подвергали лигированию при 16°C в течение ночи, используя Ligation High (TOYOBO), в соответствии со способом, описанным в прилагаемой инструкции по эксплуатации. Электрокомпетентные голубые клетки E. coli XL1 (Stratagene) или электромакс клетки DH12s (Invitrogen) были трансформированы с использованием реакционного раствора посредством способа электропорации, описанного в прилагаемой инструкции по эксплуатации. Все полученные таким образом трансформанты, устойчивые к ампициллину, были собраны и хранились в качестве рекомбинантной библиотеки при -20°C до использования.
Библиотеку E. coli (2×109 кое) инокулировали в 50 мл 2x YTAG (2x TY, содержащий 100 мкг/мл ампициллина и 2% глюкозу) и культивировали при 37°C до тех пор, пока оптическая плотность OD 600 не достигла 0,4-0,5. В культуру добавляли 4×1011 фаг-помощник VCSM13 (Stratagene) и оставляли стоять при 37°C в течение 15 минут для инфицирования клеток. Инфицированные клетки культивировали при 30°C в течение 10 часов с последующим добавлением 450 мл 2x YTAK (2x TY, содержащий 100 мкг/мл ампициллина и 25 мкг/мл канамицина) и 25 мкл 1 моль/л IPTG. Культуральный супернатант собирали центрифугированием, смешивали со 100 мл раствора PEG-NaCl (10% полиэтиленгликоль 8000, 2,5 моль/л NaCl) и оставляли отстояться при 4°C в течение 60 минут. Фаг осаждали центрифугированием при 10800×g в течение 30 минут и осадок суспендировали в 40 мл воды, смешивали с 8 мл раствора PEG-NaCl, оставляли стоять при 4°C в течение 1 часа. Фаг осаждали центрифугированием при 10800×g в течение 30 минут и суспендировали в 5 мл PBS для получения фаговой библиотеки. Фаг хранили при 4°C до использования.
10-3. Концентрирование связанного фага посредством пэннинга
Фактор IXaβ или фактор X метили биотиновой меткой, используя микропробирки No-Weigh Premeasured NHS-PEO4-Biotin (Pierce). Биотин-меченый фактор IXaβ или фактор X (100 пмоль) добавляли к раствору фаговой библиотеки, полученному в 10-2 (600 мкл), и осуществляли взаимодействие с антигеном в течение 60 минут. Добавляли Dynabeads M-280 Streptavidin (600 мкл; DYNAL), отмытые 5% M-PBS (PBS, содержащий 5% вес/объем снятого латекса), для осуществления связывания в течение 15 минут. Фаг с намагниченных бус отмывали несколько раз с помощью PBST (PBS, содержащий 0,1% Tween-20; 1 мл), а затем отмывали PBS. Бусы суспендировали в 0,8 мл 0,1 моль/л глицин/HCl (pH 2,2) в течение 5 для элюирования фага.
Альтернативно, фаговую библиотеку (80 мкл/лунку × 5), которую инкубировали с 2,5% вес/объем снятым латексом в течение 15 минут, добавляли к фактору IXaβ или фактору X (10 мкг/лунку × 5), фиксированному на иммунопланшете (MaxiSorp, Nunc), и осуществляли взаимодействие с антигеном в течение 60 минут. Антиген-связанный фаг отмывали несколько раз PBST (PBS, содержащий 0,1% Tween-20; 1 мл), а затем - PBS. Связанный фаг инкубировали с 0,8 мл 0,1 мл/л глицин/HCl (pH 2,2) в течение 5 минут для элюирования фага.
Собранный таким образом раствор фага нейтрализовали посредством добавления 2 моль/л Tris (45 мкл), добавляли к 10 мл голубых клеток XL1 в фазе логарифмического роста (OD 600=0,4-0,5) и оставляли стоять в течение 30 минут при 37°C для инфицирования клеток. Смесь распределяли в плашке 2xYTAG и культивировали при 30°C. Собирали колонии, инокулировали в 2x YTAG и культивировали при 37°C до достижения оптической плотности OD 600=0,4-0,5. IPTG (1 моль/л; 5 мкл) и фаг-помощник VCSM13 (1011 кое) добавляли в культуральный раствор (10 мл) и смесь оставляли стоять при 37°C в течение 30 минут. Клетки собирали центрифугированием, ресуспендировали в 2x YTAK (100 мл) и культивировали при 30°C в течение 10 часов. Культуральный супернатант восстанавливали центрифугированием, смешивали с 10% PEG-5 моль/л NaCl растворе (20 мл) и оставляли стоять при 4°C в течение 20 минут. Фаг осаждали центрифугированием при 10800×g в течение 30 минут и суспендировали в PBS (2 мл), и предназначены для последующего пэннинга.
10-4. Иммуноферментный анализ (ELISA) фага
Вышеописанная единичная колония была инокулирована в 2x YTAG (100 мкл) и культивирована при 30°C в течение ночи. Затем 5 мкл этой культуры инокулировали в 2x YTAG (500 мкл) и культивировали при 37°C в течение 5 часов, добавляли фаг-помощник (2×108 кое) и затем культуру оставляли стоять при 37°C в течение 30 минут. Далее после 30-минутного культивирования с встряхиванием при 37°C добавляли 2xYTAK, содержащий 0,5 мМ IPTG (120 мкл). После культивирования в течение ночи при 30°C супернатант, собранный центрифугированием, подвергали ELISA. Для ELISA клонов, полученных посредством пэннинга биотин-меченых антигенов, использовали 96-луночный микротитровальный планшет StreptaWell 96 (Roche), покрытый 1,0 мкг/мл биотин-меченого антигена. Кроме того, для ELISA клонов, полученных посредством пэннинга нативных антигенов, использовали иммунопланшет (MaxiSorp, Nunc), фиксированный 1,0 мкг/мл нативным антигеном. После отмывки PBST для удаления антигена реакцию блокировали 200 мкл 2% M-PBS или 2% BSA-PBS (PBS, содержащий 2% вес/объем BSA) в качестве блокирующего буфера в течение 1 часа при комнатной температуре. После удаления буфера культуральный супернатант добавляли в планшет и оставляли стоять в течение 60 минут для связывания фага. После отмывки связанный фаг выявляли с помощью HRP-связанного анти-M13 антитела (Amersham Pharmacia Biotech) и субстрата TMB (Zymed). Реакцию останавливали посредством добавления 1 моль/л H2SO4 и измеряли значение A450 с помощью планшет-ридера.
10-5. Определение последовательности и селекция клонов
Используя культуральную среду 2x YTAG ELISA-положительного клона рекомбинантной E. coli, определяли нуклеотидную последовательность области scFv посредством ПЦР амплификации с праймерами PBG3-F1 (5'- CAGCTATGAAATACCTATTGCC -3'/SEQ ID NO:1) и PBG3-R1 (5'- CTTTTCATAATCAAAATCACCGG -3'/SEQ ID NO:2). 15 мкл ПЦР-раствора, содержащего 1 мкл культуральной среды, 1,5 мкл 10x буфера KOD Dash, по 0,2 мкл каждого 10 мкмоль/л праймера, и 0,3 мкл полимеразы KOD Dash (TOYOBO, 2,5 Ед./мкл), подвергали 30 циклам амплификации (96°C-10 секунд, 55°C-10 секунд и 72°C-30 секунд), используя термоциклер GeneAmp PCR system 9700 (Perkin Elmer). После ПЦР добавляли 3 мкл ExoSAP-IT (Amersham) к 5 мкл реакционного раствора и температуру смеси поддерживали на 37°C в течение 15 минут с последующим инкубированием при 80°C в течение 15 минут. Реакцию этого образца проводили, используя набор BigDye Terminator Cycle Sequencing (Applied Biosystems) и праймеры PBG3-F2 (5'- ATTGCCTACGGCAGCCGCT -3'/SEQ ID NO:3) или PBG3-R2 (5'- AAATCACCGGAACCAGAGCC -3'/SEQ ID NO:4), и продукты подвергали электрофорезу, используя ДНК-секвенатор Applied Biosystems PRISM 3700. В результате клоны, имеющие различные аминокислотные последовательности CDR3, определенные по нуклеотидным последовательностям, были отсортированы на 52 клона как антифактор IXa и 33 клона как антифактор-Х.
10-6. Конструирование вектора экспрессии биспецифического IgG антитела
Для экспрессии антитела scFv в качестве типа IgG вариабельные области антитела (VH, VL) клонировали в индуцируемый тип векторов экспрессии посредством схожих способов, описанных в примерах 3-3, 3-4 и 3-5. Вариабельные области анти-F.IXa антитела (VH и VL) по отдельности встраивали в тетрациклин-индуцируемый вектор (pcDNA4-g4H и pcDNA4-g4L соответственно). Вариабельные области анти-F.X антитела (VH и VL) по отдельности встраивали в индуцируемый аналогом экдизона тип вектора (pIND-g4H и pcDNA4-g4L соответственно). Из клонов, представляющих интерес, изолировали соответствующие плазмидные ДНК, используя набор QIAprep Spin Miniprep (QIAGEN), и разводили в стерилизованной воде (100 мкл).
10-7. Экспрессия химеры биспецифического антитела в животных клетках
Используя раствор ДНК, полученный посредством способа, схожего с описанным в примере 4-1, ДНК вводили в животные клетки посредством способов, описанных в примерах 4-2 и 4-3, и собирали культуральный супернатант. Образцы хранили при 4°C до использования.
[Пример 11] Очистка антитела
К 10 мл культурального супернатанта, полученного посредством способа, описанного в примере 10-7, добавляли 100 мкл rProtein A SepharoseTM Fast Flow (Amersham Biosciences) и смешивали переворачиванием при 4°C в течение ночи. Раствор переносили в 0,22 мкм чашку фильтра Ultrafree®-MC (Millipore) и после троекратной отмывки 500 мкл TBS, содержащим 0,01% Tween® 20, смолу rProtein A SepharoseTM суспендировали в 100 мкл 10 мМ HCl/0,01% Tween® 20 (pH 2,0), и оставляли стоять в течение 3 минут. Затем антитело элюировали и элюат незамедлительно нейтрализовали добавлением 5 мкл 1 M Tris-HCl, pH 8,0. Используя программное обеспечение Microplate Manager III (Bio-Rad Laboratories), на основании калибровочной кривой человеческого IgG4 вычисляли концентрацию человеческого IgG в культуральном супернатанте (гуманизированное анти-TF антитело, смотри WO 99/51743). Концентрацию антитела количественно определяли соответственно примеру 5.
[Пример 12] Исследование F.VIIIa (активированный фактор свертывания VIII)-подобной активности
F.VIIIa-подобную активность биспецифического антитела определяли посредством следующего ферментативного анализа. Все следующие реакции проводили при комнатной температуре. Смешанный раствор 10 мкл фактора IX (15 мкг/мл; Enzyme Research Laboratories), 5 мкл TBSB, содержащего 100 мМ CaCl2 и 20 мМ MgCl2, и 50 мкл культурального супернатанта, полученного посредством способа, описанного в примере 10-7, инкубировали в 96-луночном планшете в течение 1 часа. Затем для инициации ферментативной реакции добавляли 10 мкл фактора XIa (10 нг/мл; Enzyme Research Laboratories), 20 мкл фактора X (50 мкг/мл; Enzyme Research Laboratories) и 5 мкл фосфолипидов (400 мкг/мл). Через 30 минут реакцию останавливали добавлением 10 мкл 0,5 M EDTA.
После добавления в каждую лунку по 50 мкл раствора колориметрического субстрата измеряли оптическую плотность на 0 и 30 минуте при длине волны 405 нм (эталонная длина волны 655 нм) с помощью аппарата для прочтения микропланшетов Model 3550 (Bio-Rad Laboratories). F.VIIIa-подобная активность выражалась как значение, полученное в результате вычитания значения изменения оптической плотности в культуральном супернатанте, неэкспрессирующем антитело, из значения изменения оптической плотности в культуральном супернатанте, экспрессирующем антитело (см. фиг.12).
В качестве сольвента для фосфолипида, фактора XIa, фактора IX и фактора X использовали TBSB. Раствор колориметрического субстрата представлял собой 1:1 смесь колориметрического субстрата S-2222 "Tesutochimu" (Chromogenix), разведенного в соответствии с прилагаемой инструкцией по эксплуатации, и раствора полибрена (0,6 мг/л гексадиметрин-бромид; Sigma).
[Пример 13] Анализ коагулирующей активности плазмы
Для выявления того, корректирует ли биспецифическое антитело, полученное в соответствии со способом по примеру 11, коагулирующую способность при гемофилии А, исследовали действие биспецифического антитела на активированное частичное тромбопластиновое время (APTT), используя плазму с дефицитом фактора F.VIII, посредством способа, схожего с описанным в примере 7 (см. фиг.13). Далее определяли концентрационную зависимость A44/B26 и A69/B26, которые наиболее эффективны в укорачивании времени коагуляции (см. фиг.14 и 15).
[Пример 14] Оценка совместного применения биспецифического антитела и F.VIII
Совместное применение биспецифического антитела и F.VIII оценивали при следующих аналитических условиях коагуляции плазмы. Смесь 40 мкл раствора антитела (25 мкг/мл) и 50 мкл плазмы с дефицитом фактора F.VIII (Biomerieux) инкубировали при комнатной температуре в течение 30 минут. К этой смеси добавляли 10 мкл состава рекомбинантного фактора свертывания крови VIII Kogenate® FS (BAYER) и 50 мкл реагентов APTT (Dade Behring) и нагревали при 37°C в течение 3 минут. Реакцию коагуляции инициировали добавлением 50 мкл 20 мМ CaCl2 (Dade Behring). Время, необходимое для коагуляции, измеряли, используя CR-A (Amelung)-связанный KC10A (Amelung) (см. фиг.16).
[Пример 15] Эффекты биспецифического IgG антитела в ингибирующей плазме
Эффекты биспецифического IgG антитела в ингибирующей плазме оценивали при следующих условиях анализа коагулирующей активности плазмы. Смесь 50 мкл плазмы с дефицитом F.VIII (Biomerieux) и 10 мкл нейтрализующего антитела против человеческого фактора F.VIII (100 мкг/мл; каталожный номер: MAB3440, CHEMICON) инкубировали при комнатной температуре в течение 30 минут. Эту плазму использовали в качестве ингибирующей плазмы. К этой ингибирующей плазме добавляли 40 мкл раствора антитела (25 мкг/мл) и 50 мкл реагентов APTT (Dade Behring) и нагревали при 37°C в течение 3 минут. Реакцию коагуляции инициировали добавлением 50 мкл 20 мМ CaCl2 (Dade Behring). Время, необходимое для коагуляции, измеряли, используя CR-A (Amelung)-связанный KC10A (Amelung) (см. фиг.17).
[Пример 16] Гуманизация биспецифического антитела
Из числа биспецифических антител, полученных в примерах 1-7, XB12 (анти-фактор IXa антитело мыши)/SB04 (анти-фактор X антитело мыши), являющееся наиболее эффективным при укорачивании времени коагуляции, подвергали гуманизации, как указано далее.
16-1. Поиск гомологии человеческих антител
База данных была создана на основе аминокислотных последовательностей человеческих антител, полученных из базы данных Kabat (ftp://ftp.ebi.ac.uk/pub/databases/kabat/) и базы данных IMGT (http://imgt.cines.fr/), доступных публично, поиск гомологии осуществляли по отдельности для вариабельной области Н-цепи XB12 мыши, вариабельной области L-цепи XB12 мыши, вариабельной области Н-цепи SB04 мыши и вариабельной области L-цепи SB04 мыши. Результаты подтвердили, что они имеют высокую степень гомологии с последовательностями человеческих антител, указанных ниже, и таким образом было решено, что их можно использовать в качестве каркасной области (далее здесь сокращенно как FR) гуманизированных антител.
(1) Вариабельная область Н-цепи XB12: KABATID-020619 (база данных Kabat) (Mariette et al., Arthritis Rheum. 1993; 36:1315-1324).
(2) Вариабельная область L-цепи XB12: EMBL Accession № X61642 (база данных IMGT) Mark et al., J Mol Biol. 1991; 222:581-597).
(3) Вариабельная область Н-цепи SB04-: KABATID-025255 (база данных Kabat) (Demaison et al., Immunogetetics 1995; 42:342-352).
(4) Вариабельная область L-цепи SB04: EMBL Accession № AB064111 (база данных IMGT) (неопубликованные данные).
Для получения гуманизироанных антител гипервариабельные участки каждого антитела мыши (далее здесь сокращенно как CDR) трансплантировали в FRs человеческих антител (1)-(4).
Также использовали доступный публично Web-сайт NCBI (http://www.ncbi.nlm.nih.gov/BLAST/) для поиска секреторных сигнальных последовательностей человеческого антитела, которые имеют высокую гомологию с человеческими антителами (1)-(4). Использовали следующие секреторные сигнальные последовательности, полученные посредством поиска гомологии.
(1) Вариабельная область Н-цепи XB12: GenBank Accession № AF062120.
(2) Вариабельная область L-цепи XB12: GenBank Accession № M74019.
(3) Вариабельная область Н-цепи SB04: GenBank Accession № BC019337.
(4) Вариабельная область L-цепи SB04: GenBank Accession № AY204756.
16-2. Конструирование вектора экспрессии гена гуманизированного антитела.
Двенадцать синтетических олигонуклеотидов длиной приблизительно 50 оснований получали, исходя из нуклеотидной последовательности, кодирующей аминокислотную последовательность секреторной сигнальной последовательности для вариабельной области антитела, так что около 20 оснований их 3'-концов гибридизуются друг с другом. Далее синтезировали праймер, гибридизирующийся с 5'-концом вариабельной области гена антитела и имеющий последовательность, расщепляющуюся XhoI, и праймер, гибридизирующийся с 3'-концом вариабельной области гена антитела и имеющий последовательность, расщепляющуюся SfiI.
Смешивали полученные синтетические олигонуклеотиды (2,5 мкМ, 1 мкл каждого) и добавляли 1x TaKaRa Ex Taq буфер, 0,4 мМ dNTPs и 0,5 единиц TaKaRa Ex Taq (все производства компании Takara Shuzo) для получения 48 мкл реакционного раствора. После нагревания смеси в течение 5 минут при 94°C проводили 2 цикла реакции (94°C - 2 минуты, 55°C - 2 минуты и 72°C - 2 минуты) для сборки и удлинения каждой из синтетических олиго ДНК. Далее добавляли праймер, гибридизирующийся с 5'-концом, и праймер, гибридизирующийся с 3'-концом, гена антитела (10 мкМ, 1 мкл каждый), и вариабельные области гена антитела амплифицировали посредством 35 циклов реакции (94°C - 30 секунд, 55°C - 30 секунд и 72°C - 1 минута) и 5 минут реакции при 75°C. После ПЦР реакционный раствор полностью подвергали электорофорезу в 1% агарозном геле. Амплифицированные фрагменты предполагаемого размера (около 400 п.н.) очищали, используя набор QIAquick Gel Extraction (QIAGEN), посредством способа, описанного в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (30 мкл). Фрагменты клонировали, используя систему pGEM-T Easy Vector System (Promega), посредством способа, описанного в прилагаемой инструкции по эксплуатации. Нуклеотидные последовательности фрагментов ДНК определяли, используя набор BigDye Terminator Cycle Sequencing (Applied Biosystems) на ДНК-секвенаторе ABI PRISM 3700 (Applied Biosystems), в соответствии со способом, описанным в прилагаемой инструкции по эксплуатации.
После обработки рестриктазами XhoI и SfiI плазмид, для которых подтверждено, что они содержат соответствующую последовательность вариабельной области гена гуманизированного антитела, реакционный раствор подвергали электрофорезу в 1% агарозном геле. Фрагменты ДНК предполагаемого размера (около 400 п.н.) очищали, используя набор QIAquick Gel Extraction (QIAGEN) посредством способа, описанного в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (30 мкл). Далее после расщепления рестриктазами Xhol и SfiI экспрессионных плазмид, индуцируемых тетрациклином (pcDNA4-g4H, pcDNA4-g4L), и плазмид, индуцируемых аналогом экдизона (pIND-g4H, pIND-g4L), полученных в примере 3-4, фрагменты, содержащие константную область антитела (около 5 т.п.н.), очищали, используя набор QIAquick Gel Extraction (QIAGEN), посредством способа, описанного в прилагаемой инструкции по эксплуатации, и элюировали стерилизованной водой (30 мкл). Фрагмент гена гуманизированного антитела XB12 (вариабельая область Н-цепи (далее здесь VH) или вариабельная область L-цепи (далее здесь VL)) обрабатывали рестриктазами XhoI и SfiI, а экспрессионные плазмиды, индуцируемые тетрациклином (pcDNA4-g4H, pcDNA4-g4L), обработанные рестриктазами XhoI и SfiI, подвергали реакции лигирования, используя набор Rapid DNA Ligation (Roche Diagnotics), посредством способа, описанного в прилагаемой инструкции по эксплуатации. Дополнительно фрагмент гена гуманизированного антитела SB04, расщепленный рестриктазами XhoI и SfiI (вариабельная область H-цепи или вариабельная область L-цепи), и экспрессирующую плазмиду, индуцируемую аналогом экдизона, расщепленную рестриктазами XhoI и SfiI (pIND-g4H, pIND-g4L), подвергали реакции лигирования, используя набор Rapid DNA Ligation (Roche Diagnostics), посредством способа, описанного в прилагаемой инструкции по эксплуатации. Для трансформации DH5α штамма E. coli (TOYOBO) использовали часть каждой реакционной смеси.
16-3. Получение гуманизированного биспецифического антитела
Гены трансфицировались и экспрессировались в HEK293H посредством способов, описанных в примерах 4-2 и 4-3, используя четыре типа векторов экспрессии гуманизированных антител, а также и pcDNA6/TR, и pVgRXR. Далее очистку антитела и количественное определение концентрации антитела проводили посредством способов, описанных в примерах 8 и 5.
16-4. Оценка активности гуманизированного биспецифического антитела и модификация последовательности антитела
Для оценки способности таким образом полученных гуманизированных биспецифических антител и химеры биспецифического антитела XB12/SB04 коагулировать плазму, используя плазму с дефицитом фактора F.VIII, проверяли влияние антител на APTT. Аминокислоты FR человеческого антитела модифицировали с целью повышения активности гуманизированных биспецифических антител, чья способность коагулировать кровь была снижена. Кроме того, остатки цистеина в CDR3 тяжелой цепи антитела XB12 были изменены на аланин, рассматривая возможное снижение его термостабильности. В частности, мутации вводили в вектор экспрессии гуманизированного антитела, используя набор QuikChange Site-Directed Mutagenesis (Stratagene), посредством способа, описанного в прилагаемой инструкции по эксплуатации. В результате повторения модификации аминокислот FR последовательности и оценки способности коагуляции крови было получено гуманизированное биспецифическое антитело (гуманизированное антитело XB12 (VH:hXB12f-A, VL:hXBVL)/гуманизированное антитело SB04 (VH:hSB04e, VL:hSBVL-F3f))(фиг.18).
Промышленная применимость
Настоящее изобретение обеспечивает биспецифические антитела, которые распознают и фермент, и его субстрат и которые функционально замещают кофактор, который усиливает ферментативную активность.
Биспецифические антитела в соответствии с настоящим изобретением предположительно имеют высокую стабильность в крови и низкую антигенность. Таким образом, весьма ожидаемо, что они станут лекарственными препаратами.
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТИСПЕЦИФИЧЕСКАЯ СВЯЗЫВАЮЩАЯ АНТИГЕН МОЛЕКУЛА, КОТОРАЯ ОБЛАДАЕТ АЛЬТЕРНАТИВНОЙ ФУНКЦИЕЙ К ФУНКЦИИ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ VIII | 2013 |
|
RU2534347C1 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ СВЯЗЫВАЮЩАЯ АНТИГЕН МОЛЕКУЛА, КОТОРАЯ ОБЛАДАЕТ АЛЬТЕРНАТИВНОЙ ФУНКЦИЕЙ К ФУНКЦИИ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ VIII | 2013 |
|
RU2534564C1 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ СВЯЗЫВАЮЩАЯ АНТИГЕН МОЛЕКУЛА, КОТОРАЯ ОБЛАДАЕТ АЛЬТЕРНАТИВНОЙ ФУНКЦИЕЙ К ФУНКЦИИ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ VIII | 2011 |
|
RU2620071C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО, ОБЛАДАЮЩЕЕ ПОВЫШЕННОЙ АКТИВНОСТЬЮ, АЛЬТЕРНАТИВНОЙ ФУНКЦИИ КОФАКТОРА FVIII | 2017 |
|
RU2821642C2 |
| АНТИТЕЛО, КОТОРОЕ ОБЛАДАЕТ СПОСОБНОСТЬЮ НЕЙТРАЛИЗОВАТЬ СУБСТАНЦИЮ, ОБЛАДАЮЩУЮ АКТИВНОСТЬЮ, АЛЬТЕРНАТИВНОЙ ФУНКЦИИ ФАКТОРА СВЕРТЫВАНИЯ VIII (FVIII) | 2015 |
|
RU2737145C2 |
| АНТИ-СD3-АНТИТЕЛО И МОЛЕКУЛА, СОДЕРЖАЩАЯ ЭТО АНТИТЕЛО | 2017 |
|
RU2790326C2 |
| АНТИТЕЛА К ФАКТОРУ XI И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2739946C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ РАКА ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ | 2012 |
|
RU2630638C2 |
| СПОСОБ ИЗМЕРЕНИЯ РЕАКТИВНОСТИ FVIII | 2015 |
|
RU2752595C2 |
| ИНГИБИРУЮЩИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ ФАКТОРА XII/XIIA И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2660370C2 |










































































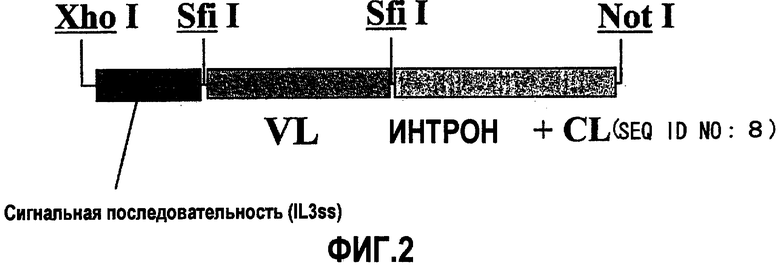
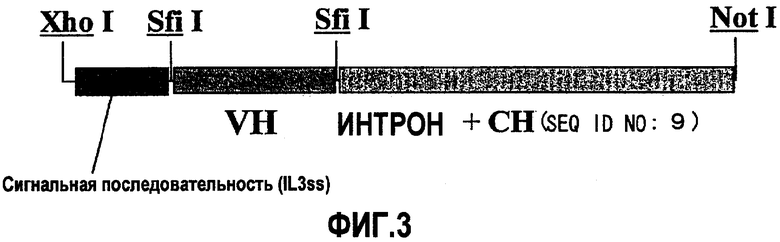
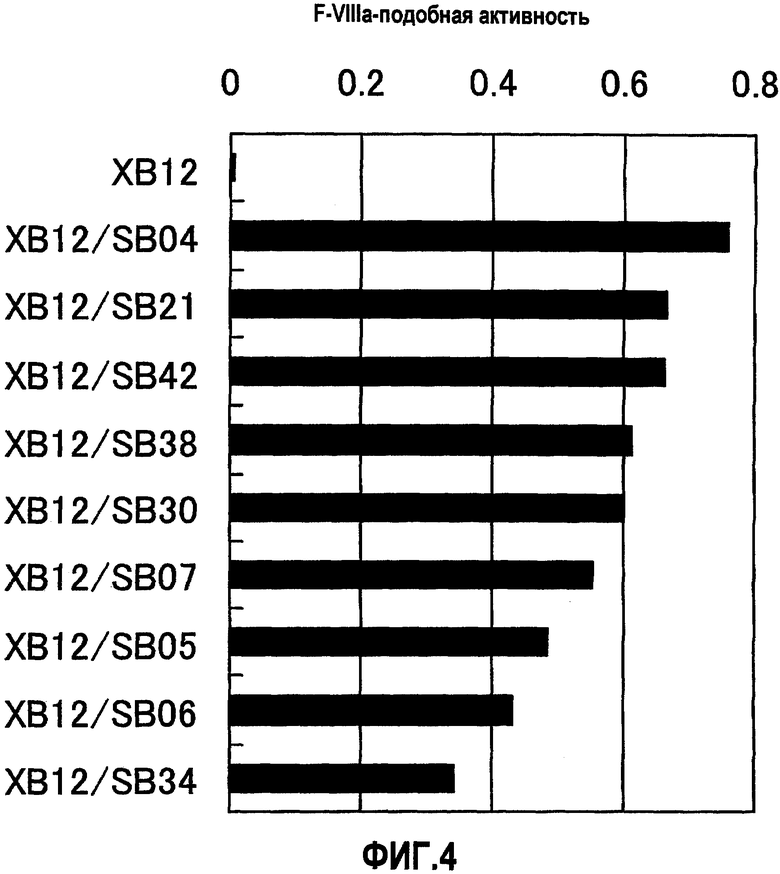
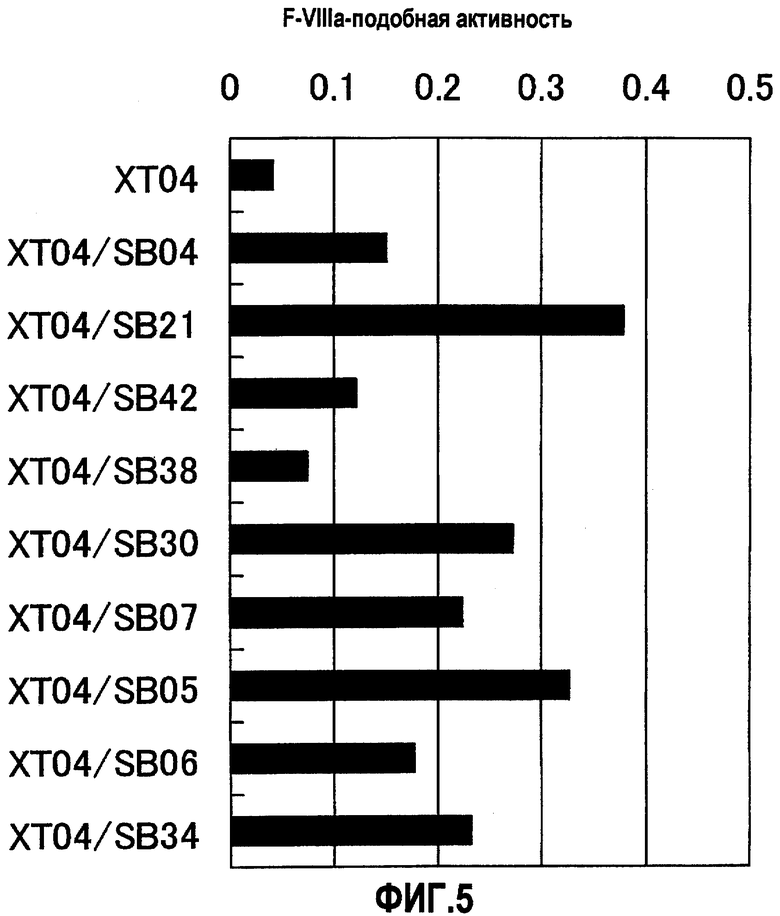
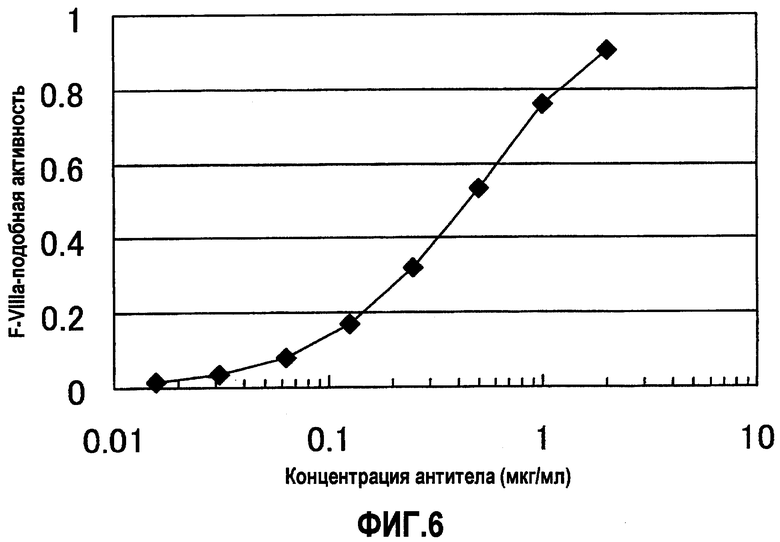
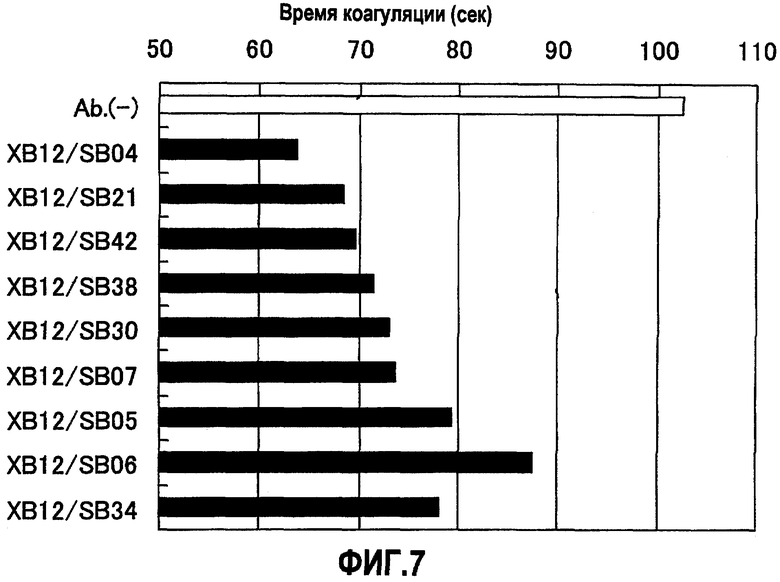
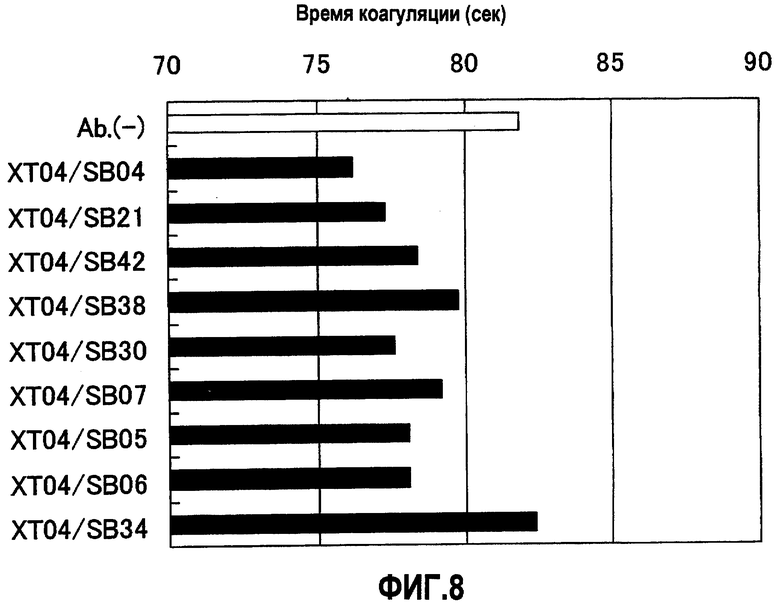
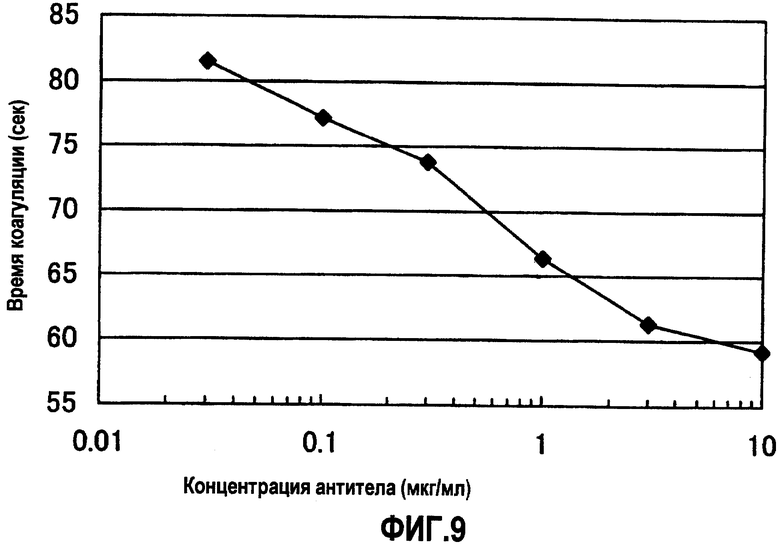
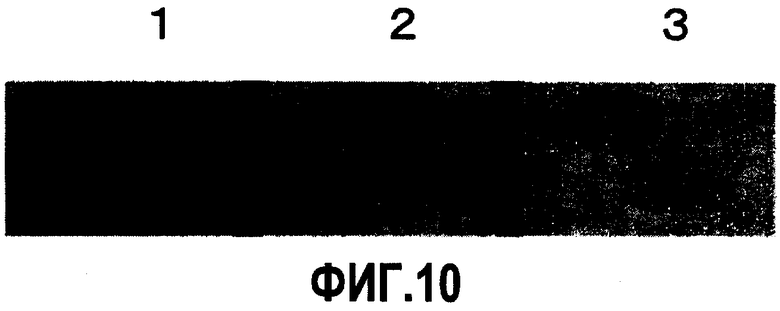
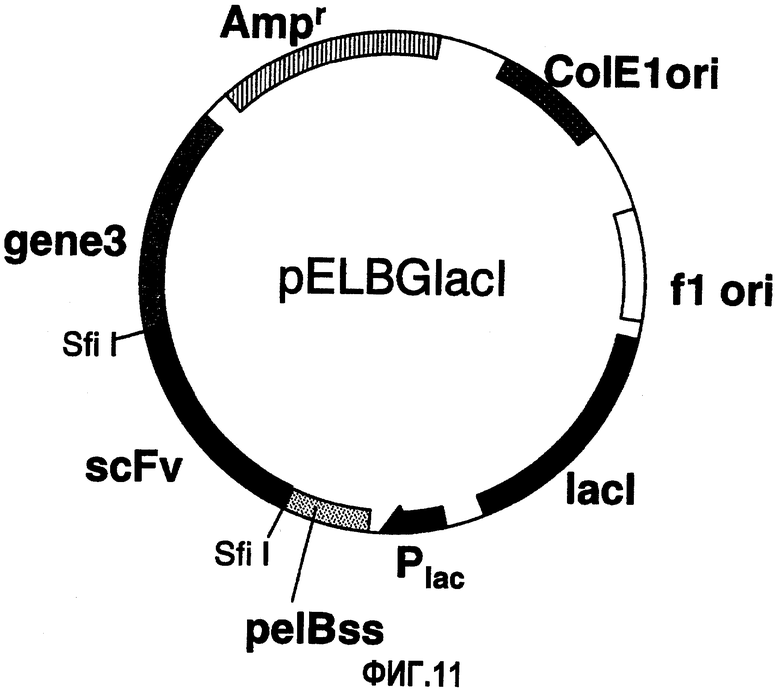
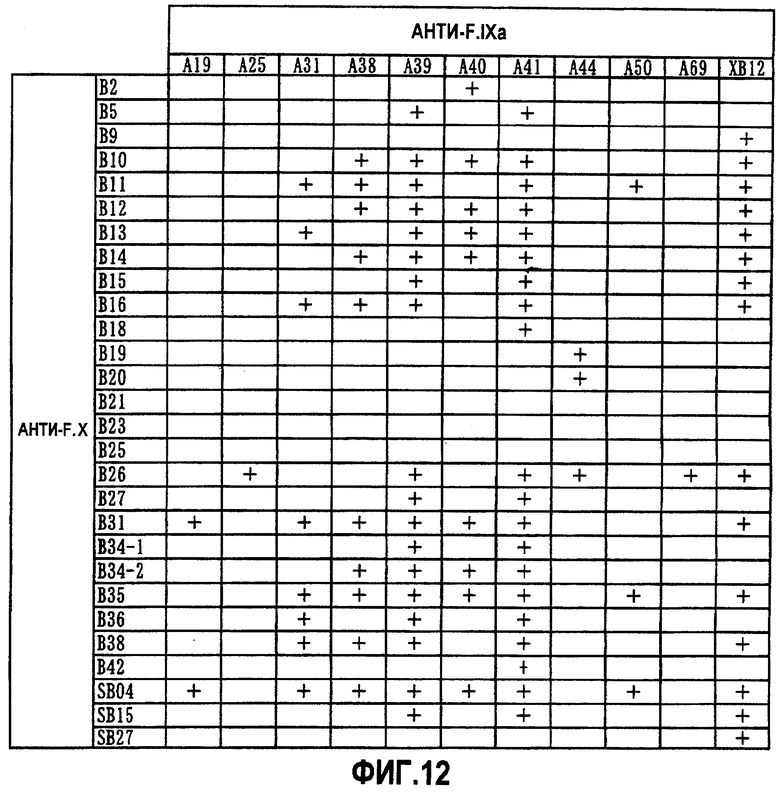
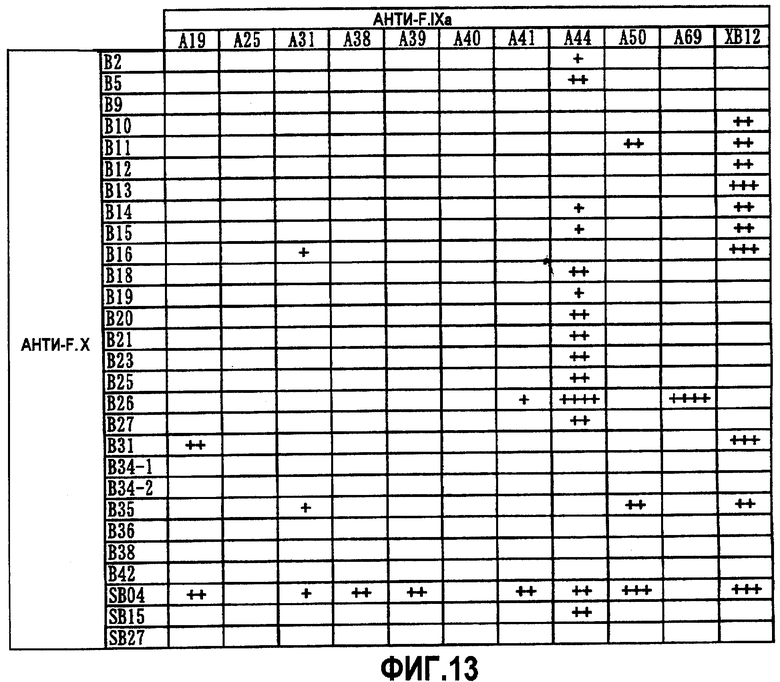
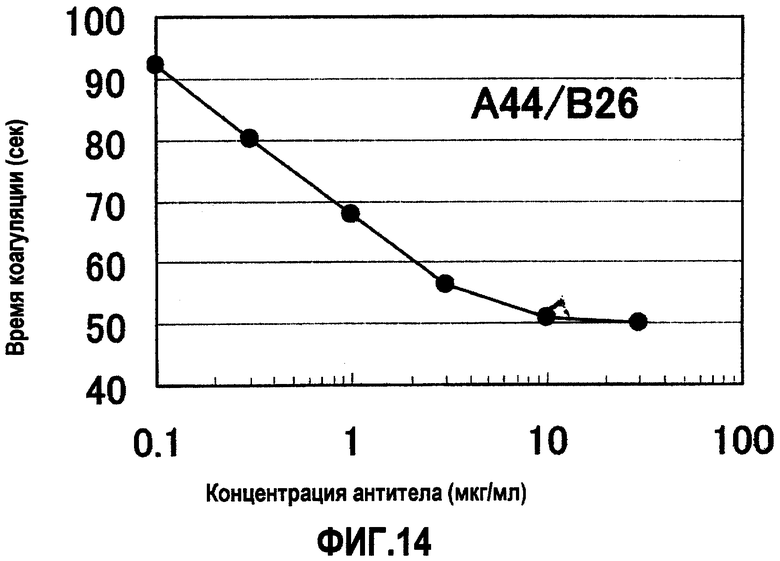
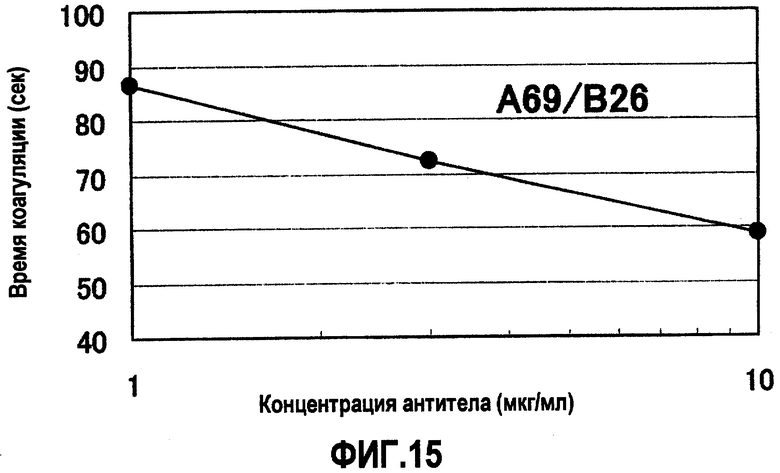
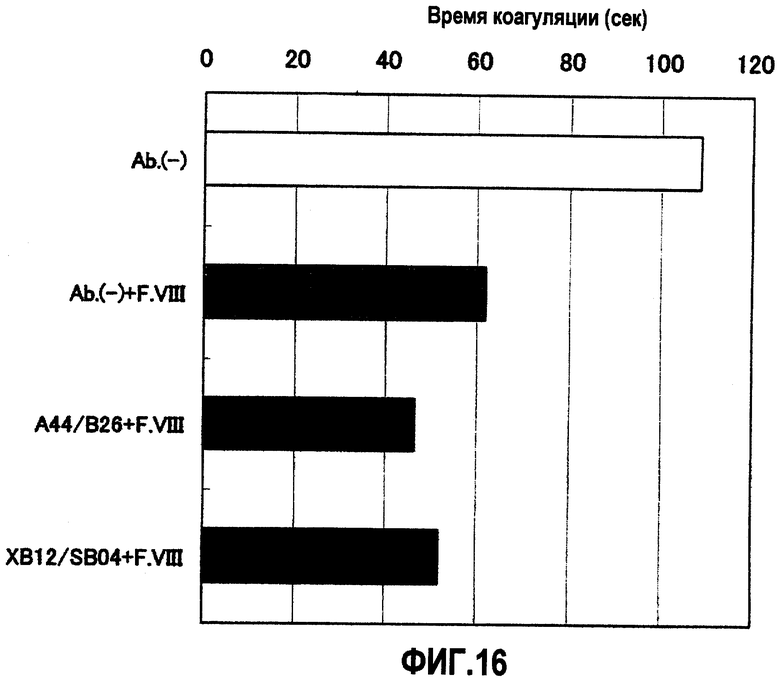
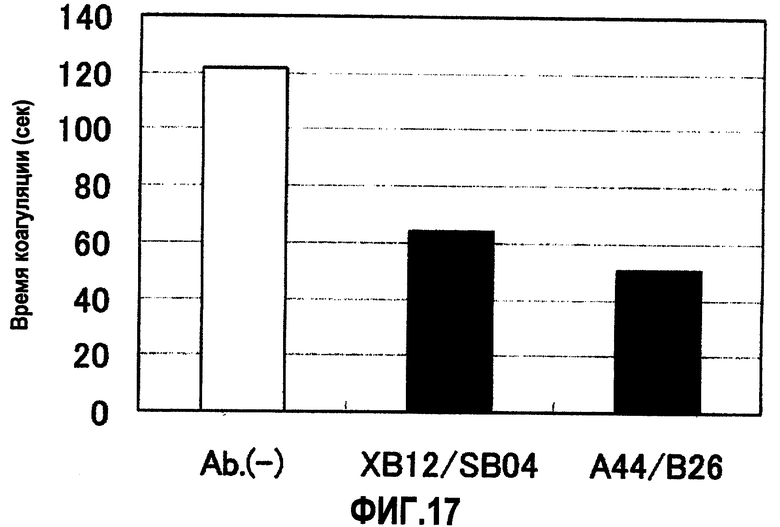
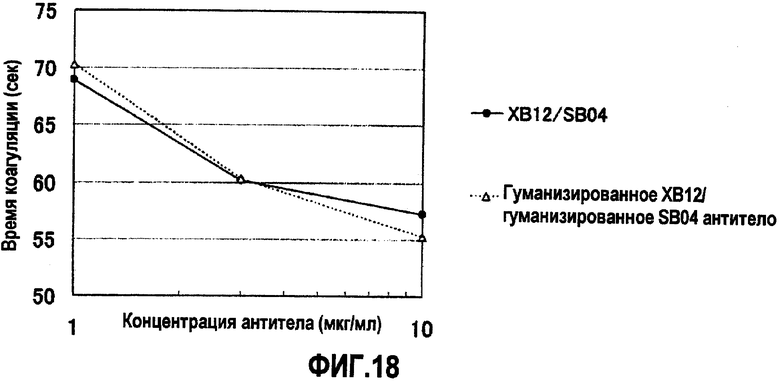
Изобретение относится к биотехнологии. Описано биспецифическое антитело, которое связывает и фактор свертывания крови IX или активированный фактор свертывания крови IX, и фактор свертывания крови X и функционально замещает фактор свертывания крови VIII или активированный фактор свертывания крови VIII, который усиливает ферментативную реакцию. Раскрыта фармацевтическая композиция, содержащая описанное антитело. Настоящее изобретение может использоваться в качестве альтернативного средства для функционального замещения кофактора, который усиливает ферментативную реакцию. 5 н. и 9 з.п. ф-лы, 18 ил.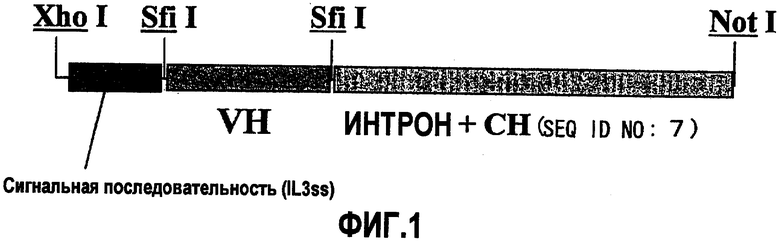
(а1) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:16;
(а2) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:20;
(b1) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:24;
(b2) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:28;
(b3) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:32;
(b4) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:36;
(b5) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:40;
(b6) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:44;
(b7) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:48;
(b8) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:52;
(b9) аминокислотная последовательность CDR3 Н-цепи, описанная в SEQ ID NO:56.
(а1) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:14, 15 и 16, соответственно;
(а2) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:18, 19 и 20, соответственно;
(b1) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:22, 23 и 24, соответственно;
(b2) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:26, 27 и 28, соответственно;
(b3) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:30, 31 и 32, соответственно;
(b4) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:34, 35 и 36, соответственно;
(b5) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:38, 39 и 40, соответственно;
(b6) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:42, 43 и 44, соответственно;
(b7) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:46, 47 и 48, соответственно;
(b8) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:50, 51 и 52, соответственно;
(b9) аминокислотные последовательности CDR 1, 2, и 3 Н-цепи, описанные в SEQ ID NO:54, 55 и 56, соответственно.
Приоритет по пунктам:
| DATA BASE, GenPept, AC: AAC26541 | |||
| http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=Protein&list_uids=3322235&dopt=GenPept, 01-08-2001 | |||
| WO 9108770 A1, 27.06.1991 | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

