По данной заявке испрашивается приоритет на основании предварительной патентной заявки США № 62/184955, поданной 26 июня 2015 г., и предварительной патентной заявки США № 62/341568, поданной 25 мая 2016 г., каждая из которых включена в настоящий документ в полном объеме посредством ссылки.
Настоящая заявка содержит список последовательностей, который был подан в электронном виде в формате ASCII и который включен в настоящий документ в полном объеме посредством ссылки. Указанная копия в формате ASCII, созданная 23 июня 2016 г., имеет название «PAT056955-WO-PCT_SL.txt» и размер 45685 байтов.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Тромбозом называют образование тромбов в кровеносных сосудах как следствие сочетания наследственных и приобретенных факторов риска, известное как тромбофилия или гиперкоагуляционные состояния. Повреждение стенок сосудов, застой крови, повышенная реакционная способность тромбоцитов и активация факторов свертывания крови являются некоторыми из основных признаков тромбоза. Тромбоз может иметь место в системе как венозного, так и артериального кровообращения, и может приводить к развитию тромбоза глубоких вен (ТГВ), легочной эмболии и инсульта. Если тромбоз имеет место в системе артериального кровообращения, в последующих зонах может возникать ишемия, приводящая к острым коронарным синдромам (ОКС), ишемическому инсульту и острой ишемии конечностей. Образование тромбов в системе венозного кровообращения, как правило, приводит к тромбозу глубоких вен, легочной эмболии и хронической тромбоэмболической легочной гипертензии. Сгустки крови также могут образовываться в придатке левого предсердия у пациентов с фибрилляцией предсердий (ФП), и оторвавшиеся тромбы могут приводить к потенциально очень тяжелым осложнениям, то есть, тромбоэмболическому инсульту и системной эмболии. Использование всех известных в настоящее время антитромботических препаратов, включая низкомолекулярный гепарин (НМГ), ингибиторы тромбина и ингибиторы фактора Xa (FXa), связано со значительным риском кровотечений (Weitz J.I. (2010) Thromb. Haemost. 103, 62). Разработка антитромботического средства, не влияющего на гемостаз, и, следовательно, не приводящего к таким осложнениям, как кровотечения, была бы крайне желательной.
Современные антикоагулянты либо вводят инъекцией, либо принимают перорально. Инъекционный антикоагулянт НМГ широко используется и обладает улучшенным терапевтическим профилем по сравнению с применяемым ранее не фракционированным гепарином. В течение нескольких последних десятилетий наиболее часто используемым пероральным антикоагулянтом был варфарин. Варфарин имеет узкое терапевтическое окно, требующее частого мониторинга коагуляционного статуса, и по-разному взаимодействует с другими лекарственными средствами. В последнее время на рынок антикоагулянтов поступили и находят все большее применение принимаемые перорально прямые ингибиторы FXa и тромбина.
Все препараты НМГ, ингибиторов FXa и ингибиторов тромбина эффективны для предотвращения послеоперационного венозного тромбоэмболического заболевания, для лечения спонтанного ТГВ и легочной эмболии, а также для предотвращения инсульта при фибрилляции предсердий. Однако эти антикоагулянты также вызывают осложнения в виде кровотечения, как правило, аналогичные тем, которые случались при использовании старых лекарственных средств варфарина и не фракционированного гепарина. В клиническом испытании ADVANCE-2 ингибитор FXa апиксабан (эликвис) сравнивали с НМГ эноксапарином у пациентов после полной замены коленного сустава. При том, что в краткосрочной терапии апиксабан был более эффективен для предотвращения венозного тромбоэмболического заболевания, чем эноксапарин, оба средства были связаны со значительным риском кровотечения. Клинически значимое кровотечение имело место у 4% пациентов, получавших апиксабан, и у 5% пациентов, получавших эноксапарин (Lassen, M.R., et al. (2009) N. Engl. J. Med. 361, 594).
В клиническом испытании RE-LY прямой ингибитор тромбина дабигатран (прадакса) сравнивали с варфарином у пациентов, имеющих фибрилляцию предсердий с риском инсульта (Connolly, S.J., et al. (2009) N. Engl. J. Med. 361, 1139). Хроническая терапия дабигатраном была связана со значительно более низким риском инсульта или системной эмболии. Однако осложнения в виде сильных кровотечений имели место у 3,1% пациентов, получавших 150 мг в сутки дабигатрана, и у 3,4% пациентов, получавших варфарин (p=0,31).
Фибрилляция предсердий (ФП) остается наиболее распространенным видом сердечной аритмии в клинической практике, являясь причиной примерно трети случаев госпитализации в связи с нарушениями сердечного ритма. В настоящее время, по оценкам, от нее страдают более 6 миллионов пациентов в Европе и примерно 2,3 миллиона в США, и эти показатели продолжают быстро расти из-за увеличения доли стареющего населения. По оценкам, у примерно 5% людей в возрасте старше 65 лет и у 10% людей в возрасте старше 80 лет разовьется ФП, однако увеличение частоты случаев ФП выходит за пределы того, что можно объяснить только возрастными причинами. Частота встречаемости факторов риска для ФП, таких как гипертензия, застойная сердечная недостаточность, гипертрофия левого желудочка, болезнь коронарных артерий и сахарный диабет, а также обструктивное апноэ во сне, также возрастает. Как следствие, число людей, страдающих ФП, по прогнозам, увеличится в два-три раза в течение следующих трех десятилетий среди населения западных стран. (Kannel and Benjamin (2008) Med Clin North Am. 2008; 92: 17-40; Bunch, et al. (2012) J Innovations of Card Rhythm Manag 2012; 3: 855-63).
Принципиальный риск ФП возрастает в четыре-пять раз при эмболическом инсульте. Характерный риск инсульта, связанного с ФП, резко возрастает с возрастом, составляя 23,5% в возрасте 80-89 лет. ФП связана с удвоением показателей смертности как у мужчин, так и у женщин (Kannel and Benjamin, 2008). ФП также независимо связана со снижением когнитивной функции и всеми формами деменции (Marzona, et al. (2012) CMAJ 2012; 184: 329-36; Geita et al. 2013; Bunch et al. 2012).
Большинству пациентов с ФП требуется пожизненная антикоагулянтная терапия для профилактики кардиоэмболического инсульта и системной эмболии. Показатель риска в баллах CHA2DS2-VASc является проверенным и широко используемым инструментом стратификации для прогнозирования риска тромбоэмболии у пациентов с фибрилляцией предсердий и для идентификации пациентов, которым будет полезна антикоагулянтная терапия (LIP 2011; Camm, et al. (2012) Eur Heart J 2012; 33: 2719-2747); накопленные данные показывают, что CHA2DS2-VASc является по меньшей мере таким же точным или, возможно, более точным, чем такие показатели, как CHADS2, для идентификации пациентов, у которых разовьется инсульт и тромбоэмболия, и определенно более точным для идентификации пациентов с ФП, имеющих «действительно низкий уровень риска». По оценкам, для 85-90% пациентов с ФП будет необходима антикоагулянтная терапия.
В метаанализе, включающем результаты 6 клинических испытаний, в которых оценивали эффект антагонистов витамина K (АВК) на снижение вероятности инсульта и системной эмболии, наблюдали значительное снижение риска возникновения инсульта (относительное снижение риска для инсульта 67%). Показатели смертности по любой причине были значительно снижены (26%) относительно контроля при приеме скорректированной дозы АВК (Hart, Pearce, and Aguilar (2007) Ann Intern Med 2007; 146: 857-867). Целевое значение международного нормализованного отношения (МНО), составляющее от 2 до 3, связано с наилучшим соотношением пользы и риска (Hylek et al. (2003) N Engl J Med; 349: 1019-1026) и повсеместно принято для использования в международных и национальных руководствах.
В последние годы новые пероральные антикоагулянты (НПАК), также называемые прямыми пероральными антикоагулянтами (ППАК), были одобрены для применения и введены в клиническую практику. Эти лекарственные средства являются по меньшей мере такими же, или даже более, эффективными, чем варфарин, для снижения вероятности тромбоэмболического заболевания (Connolly, et al. (2009) N Engl J Med; 361: 1139-51; Connolly, et al. (2011) N Engl J Med; 364: 806-17; Patel, et al. (2011) N Engl J Med 2011; 365: 883-91). При приеме НПАК также сильно снижается количество наиболее опасных осложнений, связанных с приемом варфарина, а именно, геморрагического инсульта и интракраниального кровотечения. Случаи обширного кровотечения имели место в аналогичном или несколько меньшем количестве, чем при проведении терапии варфарином. Кроме того, НПАК отличаются меньшей степенью взаимодействия с другими лекарственными средствами, чем варфарин, и могут быть использованы без регулярного мониторинга; ожидается, что это облегчит их использование в повседневной медицинской практике.
Несмотря на недавние усовершенствования, риск кровотечений остается высоким при использовании антикоагулянтов. Например, ежегодная частота случаев обширных и клинически значимых необширных кровотечений составляла 14,9%, и ежегодная частота случаев обширных кровотечений составляла 3,6% у пациентов, получавших ривароксабан, в исследовании ROCKET (Patel et al. 2011). Ежегодная частота случаев обширных кровотечений составляла >5% у пациентов с высоким риском кровотечений, определяемым как показатель риска кровотечений HAS ≥3 (Gallego, et al. (2012) Carc Arrhythm Electrophysiol.; 5: 312-318). Обширное кровотечение является особенно релевантным клиническим результатом; например, в исследовании ROCKET после эпизодов обширного кровотечения показатель смертности по любой причине составлял 20,4% в группе приема ривароксабана и 26,1% в группе приема варфарина. После эпизодов обширных кровотечений инсульт и системная эмболия имели место у 4,7% и 5,4% пациентов в группах приема ривароксабана и варфарина, соответственно (Piccini, et al. (2014) Eur Heart J; 35: 1873-80). Эпизоды обширного кровотечения также сильно влияют на продолжительность госпитализации, необходимость переливания препаратов крови и использование имеющихся ресурсов. Риск кровотечений также является основной причиной отказа от приема антикоагулянтов у пациентов, нуждающихся в них. Согласно Европейскому кардиологическому исследованию фибрилляции предсердий, включающему данные из 182 больниц в 35 странах от 5333 амбулаторных и госпитализированных пациентов с ФП, лишь 67% пациентов, нуждающихся в пероральном антикоагулянте, принимали его при выписке (Nieuwlaat, et al. (2005) Eur Heart J; 26, 2422-2434).
Таким образом, существует высокая неудовлетворенная медицинская потребность в более безопасных методах лечения, которые могут приводить к уменьшению тромбоэмболических осложнений ФП, таких как инсульт, системная эмболия, снижение когнитивной функции и смертность, с эффективностью, сопоставимой с эффективностью существующих методов лечения, но с меньшей вероятностью кровотечений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к моноклональным антителам, связывающимся с человеческими факторами свертывания крови XI и XIa (активированный фактор XI) (далее в настоящем документе иногда использованы сокращения «FXI», «FXIa» и аналогичные термины), и содержащим их фармацевтическим композициям, а также к способам лечения, включающим их введение. Разработка противотромботического средства, которое эффективно для предотвращения и лечения тромбоза или тромбоэмболического заболевания/нарушения (например, тромбозного инсульта, фибрилляции предсердий, предотвращения инсульта при фибрилляции предсердий (ПИФП), тромбоза глубоких вен, венозной тромбоэмболии, легочной эмболии, острых коронарных синдромов (ОКС), ишемического инсульта, острой ишемии конечностей, хронической тромбоэмболической легочной гипертензии, системной эмболии), но не несет, или несет лишь минимальный, риск кровотечений, позволила бы удовлетворить имеющуюся существенную медицинскую потребность.
В конкретных аспектах антитела (например, человеческие, химерные, гуманизированные моноклональные антитела), предложенные в настоящем документе, связывают с аналогичной высокой аффинностью каталитический домен (КД) человеческих факторов FXIa и FXI, и вызывают образование неактивной конформации домена протеазы в FXIa.
Выделенные анти-FXI и/или анти-FXIa антитела, описанные в настоящем документе, например, полноразмерные IgG с двумя связывающими сайтами, описанные в настоящем документе, связывают FXI и/или FXIa с равновесной константой диссоциации (KD) менее или равной 100 пМ. Например, выделенные антитела, описанные в настоящем документе, могут связывать человеческий FXI и/или FXIa с KD менее или равной 100 пМ, менее или равной 50 пМ, менее или равной 45 пМ, менее или равной 40 пМ, менее или равной 35 пМ, менее или равной 20 пМ, или менее или равной 10 пМ. Более конкретно, выделенные антитела, описанные в настоящем документе, также могут связывать человеческий FXI и/или FXIa с KD менее или равной 34 пМ при измерении методом поверхностного плазмонного резонанса (ППР), например, в анализе BIACORE™, либо менее или равной 4 пМ при измерении в анализе титрования равновесного раствора (ТРР); и также могут связывать FXI и/или FXIa яванского макака с KD менее или равной 53 пМ при измерении в анализе BIACORE™ или менее или равной 4 пМ при измерении в анализе ТРР. В конкретных аспектах выделенные антитела, описанные в настоящем документе (например, NOV1401), связывают человеческие FXI и FXIa с кажущейся KD менее или равной примерно 5 пМ (например, 4,7 пМ) и 2 пМ (например, 1,3 пМ), соответственно, например, при измерении в анализе титрования равновесного раствора (ТРР). В конкретных вариантах осуществления анти-FXI/FXIa антитела, описанные в настоящем документе, связывают FXI/FXIa яванского макака с кажущейся KD примерно 12,5 (±6,6) пМ для FXIa и примерно 5,0 (±0,7) пМ при измерении в анализе ТРР (смотри, например, пример 2). В конкретных вариантах осуществления анти-FXI/FXIa антитела, описанные в настоящем документе, связывают FXI и/или FXIa кролика с KD примерно 20 (±2) нМ. В конкретных аспектах анти-FXI/FXIa антитела, описанные в настоящем документе, связывают FXI и/или FXIa человека, яванского макака и кролика, но не связывают специфически мышиный или крысиный FXI.
Выделенные анти-FXI и/или анти-FXIa антигенсвязывающие фрагменты, описанные в настоящем документе, например, Fab-фрагменты и другие фрагменты, содержащие один связывающий сайт, связывают FXI и/или FXIa с равновесной константой диссоциации (KD) менее или равной 10 нМ. Например, выделенные антигенсвязывающие фрагменты, описанные в настоящем документе, могут связывать человеческий FXI и/или FXIa с KD менее или равной 10 нМ, менее или равной 5 нМ, менее или равной 1 нМ, менее или равной 500 пМ, менее или равной 305 пМ, менее или равной 62 пМ. Более конкретно, выделенные антигенсвязывающие фрагменты, описанные в настоящем документе, также могут связывать человеческий FXI и/или FXIa с KD менее или равной 305 пМ.
Настоящее изобретение относится к выделенному антителу, или его антигенсвязывающим фрагментам, которое связывает FXIa человека, кролика и яванского макака. Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, которое связывается внутри каталитического домена FXI и/или FXIa, в частности, с поверхностью области активного центра.
Настоящее изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, которое связывает FXI и/или FXIa и также конкурирует за связывание с антителом, приведенным в таблице 1 (например, NOV1401). Используемый в настоящем документе термин «конкуренция» между антителами и/или их антигенсвязывающими фрагментами означает, что оба антитела (или их связывающих фрагмента) связывают один и тот же, или перекрывающийся, эпитоп FXI и/или FXIa (например, как определяют в анализе конкурентного связывания любыми методами, хорошо известными специалистам в данной области). При описании в настоящем документе антитело, или его антигенсвязывающий фрагмент, не «конкурирует» с антителом, или антигенсвязывающим фрагментом, по изобретению против FXI и/или FXIa (например, NOV1401 или NOV1090), если только указанное конкурирующее антитело, или его антигенсвязывающий фрагмент, не связывает тот же самый эпитоп FXI и/или FXIa, или перекрывающийся эпитоп FXI и/или FXIa, что и антитело, или антигенсвязывающий фрагмент, по изобретению. При описании в настоящем документе конкурирующее антитело, или его антигенсвязывающий фрагмент, не включает то, которое (i) стерически блокирует антитело, или антигенсвязывающий фрагмент, по изобретению от связывания с его мишенью (например, если указанное конкурирующее антитело связывается с соседним, не перекрывающимся эпитопом FXI и/или FXIa и физически препятствует связыванию антитела, или антигенсвязывающего фрагмента, по изобретению с его мишенью); и/или (ii) связывает другой, не перекрывающийся эпитоп FXI и/или FXIa и вызывает конформационное изменение белка FXI и/или FXIa таким образом, что указанный белок больше не может быть связан антителом, или антигенсвязывающим фрагментом, по изобретению против FXI и/или FXIa так, как в случае отсутствия указанного конформационного изменения.
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают FXI и/или FXIa и также конкурируют с антителом, приведенным в таблице 1, за связывание с большинством аминокислот в эпитопе(ах), который связывает указанное антитело из таблицы 1. В другом варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают FXI и/или FXIa и также конкурируют с антителом, приведенным в таблице 1, за связывание со всем эпитопом(ами), который связывает указанное антитело из таблицы 1.
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают активный FXI (FXIa), и при связывании с каталитическим доменом активного FXI (FXIa) вызывают изменение конформации FXIa в неактивную конформацию. В другом варианте осуществления указанные выделенные антитела, или их антигенсвязывающие фрагменты, дополнительно вызывают изменение, при котором 4 N-концевых остатка, петли 145, 188 и 220 указанной неактивной конформации смещены и/или являются неупорядоченными по сравнению с активной конформацией.
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают FXI (например, FXI человека) и при связывании с FXI предотвращают принятие каталитическим доменом FXI активной конформации, при которой петли 145, 188 и 220 упорядочены, как в структуре каталитического домена FXIa.
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают FXI и при связывании FXI предотвращают принятие каталитическим доменом FXI активной конформации, при которой 4 N-концевых остатка, петли 145, 188 и 220 упорядочены, как в структуре каталитического домена FXIa.
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают FXI и при связывании FXI предотвращают принятие каталитическим доменом FXI активной конформации, вызывая конформационные изменения в структуре зимогена, приводящие к ингибированной конформации FXI, близкой к той, которая наблюдается при связывании с FXIa.
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают FXI и/или FXIa, и при связывании с FXI и/или FXIa и образовании комплекса антитело: антиген с каталитическим доменом FXI и/или FXIa вызывают смещение и/или неупорядоченность петель 145, 188 и 220 по сравнению со структурой каталитического домена не находящегося в комплексе активного фактора XI (FXIa).
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают FXI и/или FXIa, и при связывании с FXI и/или FXIa и образовании комплекса антитело: антиген с каталитическим доменом FXI и/или FXIa вызывают смещение и/или неупорядоченность 4 N-концевых остатков, петель 145, 188 и 220 по сравнению со структурой каталитического домена не находящегося в комплексе активного фактора XI (FXIa).
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают активный FXI (FXIa) и вызывают изменение конформации каталитического домена FXI (FXIa) в неактивную конформацию, при которой петли 145, 188 и 220 смещены и/или являются неупорядоченными по сравнению с активной конформацией.
В одном варианте осуществления выделенные антитела, или их антигенсвязывающие фрагменты, связывают FXI и предотвращают принятие каталитическим доменом активной конформации, вызывая конформационные изменения в структуре зимогена, тем самым приводя к ингибированной конформации FXI, близкой к той, которая наблюдается при связывании с FXIa.
Настоящее изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, которое связывает тот же эпитоп, что и антитело, приведенное в таблице 1 (например, NOV1401).
Аффинность связывания выделенных антител и антигенсвязывающих фрагментов, описанных в настоящем документе, можно определять методом титрования равновесного раствора (ТРР). Методы ТРР известны в данной области и более подробно описаны ниже. Альтернативно, аффинность связывания выделенных антител, или фрагментов, описанных в настоящем документе, можно определять методом поверхностного плазмонного резонанса, например, в анализах BIACORE™. Методы анализа кинетики BIACORE™ известны в данной области и более подробно описаны ниже.
Выделенные анти-FXI и/или FXIa антитела и антигенсвязывающие фрагменты, описанные в настоящем документе, могут быть использованы для ингибирования прямой или непрямой активации фактора IX (также известного как FIX), фактора X (FX) и/или тромбина, и/или связывания с рецепторами тромбоцитов, и таким образом, могут предотвращать активацию внутреннего и/или общего путей активации свертывания крови.
Выделенные анти-FXI и/или анти-FXIa антитела и антигенсвязывающие фрагменты, описанные в настоящем документе, могут быть использованы для ингибирования прямой или непрямой активации фактора IX (также известного как FIX), фактора X (FX) и/или тромбина с величиной IC50 менее или равной 100 нМ, менее или равной 50 нМ, менее или равной 35 нМ, менее или равной 25 нМ, менее или равной 10 нМ, или менее или равной 5,2 нМ. Более конкретно, выделенное антитело, или его антигенсвязывающие фрагменты, описанное в настоящем документе, может ингибировать прямую или непрямую активацию фактора IX (также известного как FIX), фактора X (FX) и/или тромбина с величиной IC50 менее или равной 100 нМ, менее или равной 50 нМ, менее или равной 35 нМ, менее или равной 25 нМ, менее или равной 10 нМ, или менее или равной 5,2 нМ. Более конкретно, выделенное антитело, или его антигенсвязывающие фрагменты, описанное в настоящем документе, может ингибировать прямую или непрямую активацию фактора IX (также известного как FIX), фактора X (FX) и/или тромбина с величиной IC50 менее или равной 100 нМ, менее или равной 50 нМ, менее или равной 35 нМ, менее или равной 25 нМ, менее или равной 20 нМ, или менее или равной 18 нМ. Более конкретно, выделенное антитело, или его антигенсвязывающие фрагменты, описанное в настоящем документе, может ингибировать прямую или непрямую активацию фактора IX (также известного как FIX), фактора X (FX) и/или тромбина с величиной IC50 менее или равной 100 нМ, менее или равной 50 нМ, менее или равной 35 нМ, менее или равной 25 нМ, менее или равной 10 нМ, или менее или равной 5 нМ. В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе, или его антигенсвязывающий фрагмент, ингибирует FXIa-опосредованную активацию его естественного субстрата FIX с величиной IC50 менее или равной 2 нМ, например, 1,8 нМ.
Выделенные анти-FXI и/или анти-FXIa антитела, или их антигенсвязывающие фрагменты, могут быть использованы для ингибирования (например, блокирования активации) внутреннего и/или общего путей активации свертывания крови, например, путем ингибирования FXI и/или FXIa-опосредованной активации FIX. Выделенные анти-FXI/FXIa антитела, или их антигенсвязывающие фрагменты, таким образом, могут быть использованы для предотвращения свертывания крови или распространения свертывания крови. Выделенные антитела, или их антигенсвязывающие фрагменты, могут быть использованы для предотвращения, лечения или ослабления таких заболеваний свертывания крови, как тромбоз глубоких вен и инсульт (например, ишемический инсульт), за счет ингибирования FXI-опосредованной активации FIX.
В конкретных вариантах осуществления анти-FXI и/или анти-FXIa антитела, или их антигенсвязывающие фрагменты, способны увеличивать время свертывания (например, время до начала образования сгустков крови) в человеческой плазме зависимым от концентрации образом, что определяют в анализе аЧТВ, например, как описано в разделе «Примеры». В конкретном варианте осуществления время свертывания крови (аЧТВ) удваивается по сравнению с исходным уровнем при общей концентрации анти-FXI антитела (например, NOV1401) в диапазоне от 10 нМ до 20 нМ, например, примерно 14 нМ или 15 нМ, при определении в анализе аЧТВ. В конкретных вариантах осуществления анти-FXI и/или анти-FXIa антитела, или их антигенсвязывающие фрагменты, способны увеличивать время свертывания в человеческой плазме зависимым от концентрации образом с величиной IC50 в диапазоне от 5 нМ до 20 нМ, например, примерно 13 нМ, при определении в анализе аЧТВ, например, как описано в разделе «Примеры».
В конкретных вариантах осуществления анти-FXI и/или анти-FXIa антитела, описанные в настоящем документе, или их антигенсвязывающие фрагменты, способны увеличивать время свертывания (например, время до начала образования сгустков крови) в человеческой плазме по меньшей мере в 1,1 раза, 1,2 раза, 1,3 раза, 1,4 раза, 1,5 раза, 1,6 раза, 1,7 раза, 1,8 раза, 1,9 раза или 2 раза, например, зависимым от концентрации образом, при определении в анализе аЧТВ, например, как описано в разделе «Примеры». В конкретных вариантах осуществления анти-FXI и/или анти-FXIa антитела, описанные в настоящем документе, или их антигенсвязывающие фрагменты, способны увеличивать время свертывания (например, время до начала образования сгустков крови) в человеческой плазме по меньшей мере в 1,4 раза, 1,5 раза, 1,6 раза или 1,7 раза, при определении в анализе аЧТВ, например, как описано в разделе «Примеры».
В конкретных аспектах анти-FXI и/или анти-FXIa антитела, или их антигенсвязывающие фрагменты, описанные в настоящем документе, способны приводить к уменьшению количества тромбина зависимым от концентрации образом в анализе образования тромбина (АОТ) в человеческой плазме, в котором измеряют эффект ингибирования FXIa на петлю положительной обратной связи тромбин→FXIa в присутствии очень низких концентраций тканевого фактора (TF). В конкретных вариантах осуществления анти-FXI и/или анти-FXIa антитела, или их антигенсвязывающие фрагменты, описанные в настоящем документе, способны приводить к уменьшению количества тромбина в анализе образования тромбина (АОТ) в человеческой плазме с величиной IC50 в диапазоне от 10 нМ до 30 нМ, например, примерно 20 нМ или 24 нМ, и остаточной концентрацией тромбина примерно 159 нМ.
В конкретных аспектах настоящего изобретения предложены антитела (например, антитела в таблице 1, такие как NOV1401, или антитела, содержащие области HCDR 1-3 и области LCDR 1-3 из NOV1401), или их антигенсвязывающие фрагменты, которые специфически связывают каталитический домен человеческого FXI и/или FXIa, и которые имеют терминальный период полувыведения (t½) всего антитела в организме яванских макак примерно 14-15 дней. В конкретных вариантах осуществления такие анти-FXI/FXIa антитела демонстрируют абсолютную биодоступность при подкожном (п/к) введении примерно 61-66%.
В конкретном варианте осуществления антитело, или его антигенсвязывающий фрагмент, по настоящему изобретению (например, антитело, приведенное в таблице 1, такое как NOV1401), которое специфически связывает человеческий FXI и/или FXIa, имеет одну или более (например, две или три, или четыре, или пять, или шесть, или семь), или все, из следующих характеристик:
(i) специфически связывает каталитический домен (КД) человеческого FXI и FXIa, например, с кажущейся KD примерно 1-2 пМ и 4-5 пМ, соответственно;
(ii) увеличивает время свертывания крови при оценке в анализе активированного частичного тромбопластинового времени (аЧТВ);
(iii) ингибирует образование тромбина в человеческой плазме за счет ингибирования активации FXI активированным фактором XII (FXIIa) и тромбином, соответственно;
(iv) проявляет антитромботическую и антикоагулянтную активность у мышей FXI-/- с введенным человеческим FXI;
(v) уменьшает или пролонгирует уменьшение уровней свободного FXI (FXIс), например, у яванских макак;
(vi) имеет терминальный период полувыведения всего антитела примерно 14-15 дней, например, у яванских макак;
(vii) специфически связывает человеческий и обезьяний FXI и/или FXIa, но не связывает специфически мышиный или крысиный FXI и/или FXIa; и
(viii) контактирует с одним или более (например, двумя, тремя, четырьмя, пятью, шестью или семью, или более), или некоторыми, или всеми, из следующих остатков человеческого FXI (нумерация Swissprot): Pro410, Arg413, Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472-Glu476, Tyr521-Lys527, Arg548, His552, Ser575, Ser594-Glu597 и Arg602-Arg604.
Выделенные анти-FXI и/или анти-FXIa антитела, или их антигенсвязывающие фрагменты, описанные в настоящем документе, могут представлять собой моноклональные антитела, человеческие или гуманизированные антитела, химерные антитела, одноцепочечные антитела, Fab-фрагменты, Fv-фрагменты, F(ab')2-фрагменты или scFv-фрагменты, и/или изотипы IgG (например, IgG1, такие как человеческие IgG1). В конкретных вариантах осуществления анти-FXI и/или анти-FXIa антитела, описанные в настоящем документе, представляют собой рекомбинантные человеческие антитела. В конкретных вариантах осуществления анти-FXI и/или анти-FXIa антитела, описанные в настоящем документе, представляют собой человеческие IgG1/лямбда (λ) антитела. В конкретных вариантах осуществления анти-FXI и/или анти-FXIa антитела, описанные в настоящем документе, представляют собой человеческие IgG1/лямбда (λ) антитела, содержащие Fc-домен, сконструированный для уменьшения возможности эффекторной функции (например, ADCC и/или CDC), например, человеческий Fc-домен, содержащий замены D265A и/или P329A.
Выделенные анти-FXI и/или анти-FXIa антитела, или их антигенсвязывающие фрагменты, описанные в настоящем документе, также могут содержать каркас, в котором имеет место аминокислотная замена по сравнению с соответствующими человеческими последовательностями VH или VL зародышевой линии.
Другой аспект изобретения относится к выделенному антителу, или его антигенсвязывающим фрагментам, имеющему полноразмерные последовательности тяжелой и легкой цепей Fab-фрагментов, приведенные в таблице 1. Более конкретно, выделенное антитело, или его антигенсвязывающие фрагменты, может иметь последовательности тяжелой и легкой цепей из NOV1090 и NOV1401.
Следующий аспект изобретения относится к выделенному антителу, или его антигенсвязывающим фрагментам, имеющему последовательности вариабельных доменов тяжелой и легкой цепей Fab-фрагментов, приведенные в таблице 1. Более конкретно, выделенное антитело, или его антигенсвязывающий фрагмент, может иметь последовательности вариабельных доменов тяжелой и легкой цепей из NOV1090 и NOV1401.
Следующий аспект изобретения относится к выделенному антителу, или его антигенсвязывающим фрагментам, имеющему последовательности CDR вариабельного домена тяжелой цепи (то есть, HCDR1, HCDR2 и HCDR3) и CDR вариабельного домена легкой цепи (то есть, LCDR1, LCDR2 и LCDR3) из антител, приведенных в таблице 1, такие как CDR по системе Kabat, CDR по системе IMGT, CDR по системе Chothia или комбинированные CDR. Более конкретно, выделенное антитело, или его антигенсвязывающий фрагмент, может иметь последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 из NOV1090 и NOV1401, например, приведенные в таблице 1, такие как CDR по системе Kabat, CDR по системе IMGT, CDR по системе Chothia или комбинированные CDR.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, содержащему CDR1 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NOs: 3 и 23; CDR2 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NOs: 4 и 24; и CDR3 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NOs: 5 и 25, при этом выделенное антитело, или его антигенсвязывающие фрагменты, связывает человеческий FXI и/или FXIa. В другом аспекте такое выделенное антитело, или его антигенсвязывающие фрагменты, дополнительно содержит CDR1 легкой цепи, выбранную из группы, состоящей из SEQ ID NOs: 13 и 33; CDR2 легкой цепи, выбранную из группы, состоящей из SEQ ID NOs: 14 и 34; и CDR3 легкой цепи, выбранную из группы, состоящей из SEQ ID NOs: 15 и 35.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, содержащему CDR1 легкой цепи, выбранную из группы, состоящей из SEQ ID NOs: 13 и 33; CDR2 легкой цепи, выбранную из группы, состоящей из SEQ ID NOs: 14 и 34; и CDR3 легкой цепи, выбранную из группы, состоящей из SEQ ID NOs: 15 и 35, при этом выделенное антитело, или его антигенсвязывающие фрагменты, связывает человеческий FXI и/или FXIa.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, связывающему FXI и/или FXIa, которое имеет HCDR1, HCDR2 и HCDR3, а также LCDR1, LCDR2 и LCDR3, при этом HCDR1, HCDR2 и HCDR3 содержат SEQ ID NOs: 3, 4 и 5, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NOs: 13, 14 и 15; или HCDR1, HCDR2 и HCDR3 содержат SEQ ID NOs: 23, 24 и 25, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NOs: 33, 34 и 35.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, связывающему FXI и/или FXIa, которое имеет HCDR1, HCDR2 и HCDR3, а также LCDR1, LCDR2 и LCDR3, при этом HCDR1, HCDR2 и HCDR3 содержат SEQ ID NOs: 43, 44 и 45, соответственно, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NOs: 47, 37 и 15, соответственно.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, связывающему FXI и/или FXIa, которое имеет HCDR1, HCDR2 и HCDR3, а также LCDR1, LCDR2 и LCDR3, при этом HCDR1, HCDR2 и HCDR3 содержат SEQ ID NOs: 46, 4 и 5, соответственно, и LCDR1, LCDR2, LCDR3 содержат SEQ ID NOs: 33, 14 и 15, соответственно.
Изобретение также относится к антителу, или антигенсвязывающему фрагменту, имеющему HCDR1, HCDR2 и HCDR3 вариабельного домена тяжелой цепи с SEQ ID NOs: 9 и 29, и LCDR1, LCDR2 и LCDR3 вариабельного домена легкой цепи с SEQ ID NOs: 19 и 39 по определению Chothia. В другом аспекте изобретения антитело, или антигенсвязывающий фрагмент, может иметь HCDR1, HCDR2 и HCDR3 вариабельного домена тяжелой цепи с последовательностями SEQ ID NOs: 9 и 29, и LCDR1, LCDR2 и LCDR3 вариабельного домена легкой цепи с последовательностями SEQ ID NOs: 19 и 39 по определению Kabat.
Изобретение также относится к антителу, или антигенсвязывающему фрагменту, имеющему HCDR1, HCDR2 и HCDR3 вариабельного домена тяжелой цепи с SEQ ID NOs: 9 и 29 и LCDR1, LCDR2 и LCDR3 вариабельного домена легкой цепи с SEQ ID NOs: 19 и 39 по определению IMGT. В другом аспекте изобретения антитело, или антигенсвязывающий фрагмент, может иметь HCDR1, HCDR2 и HCDR3 вариабельного домена тяжелой цепи с последовательностями SEQ ID NOs: 9 и 29 и LCDR1, LCDR2 и LCDR3 вариабельного домена легкой цепи с последовательностями SEQ ID NOs: 19 и 39 по определению комбинированной системы.
В одном аспекте изобретения выделенное антитело, или его антигенсвязывающие фрагменты, содержит последовательность вариабельного домена тяжелой цепи, выбранную из группы, состоящей из SEQ ID NOs: 9 и 29. Выделенное антитело, или антигенсвязывающий фрагмент, также может содержать последовательность вариабельного домена легкой цепи, при этом вариабельный домен тяжелой цепи и вариабельный домен легкой цепи в совокупности образуют антигенсвязывающий сайт для FXIa. В частности, последовательность вариабельного домена легкой цепи может быть выбрана из SEQ ID NOs: 19 и 39, при этом указанное выделенное антитело, или его антигенсвязывающие фрагменты, связывает FXI и/или FXIa.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, содержащему последовательность вариабельного домена легкой цепи, выбранную из группы, состоящей из SEQ ID NOs: 19 и 39, при этом указанное выделенное антитело, или его антигенсвязывающие фрагменты, связывает человеческий FXI и/или FXIa. Выделенное антитело, или антигенсвязывающий фрагмент, может дополнительно содержать последовательность вариабельного домена тяжелой цепи, при этом вариабельный домен легкой цепи и вариабельный домен тяжелой цепи в совокупности образуют антигенсвязывающий сайт для FXI и/или FXIa.
В частности, выделенное антитело, или его антигенсвязывающие фрагменты, которое связывает FXI и/или FXIa, может иметь вариабельные домены тяжелой и легкой цепей, содержащие последовательности SEQ ID NOs: 9 и 19; или 19 и 39, соответственно.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, содержащему вариабельный домен тяжелой цепи, имеющий по меньшей мере 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 9 и 29, при этом указанное антитело связывает FXI и/или FXIa. В одном аспекте выделенное антитело, или его антигенсвязывающие фрагменты, также содержит вариабельный домен легкой цепи, имеющий по меньшей мере 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 19 и 39. В следующем аспекте изобретения выделенное антитело, или антигенсвязывающий фрагмент, имеет HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 в соответствии с определением Kabat, приведенные в таблице 1. В конкретном варианте осуществления выделенное антитело, или антигенсвязывающий фрагмент, имеет HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 в соответствии с определением Chothia, IMGT или комбинированной системы, приведенные в таблице 1.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, которое имеет вариабельный домен легкой цепи, имеющий по меньшей мере 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 19 и 39, при этом указанное антитело связывает FXI и/или FXIa.
В другом аспекте изобретения выделенное антитело, или его антигенсвязывающие фрагменты, которое связывает FXI и/или FXIa, может иметь тяжелую цепь, содержащую последовательность SEQ ID NOs: 11 или 31. Выделенное антитело также может иметь легкую цепь, которая может в сочетании с тяжелой цепью образовывать антигенсвязывающий сайт для человеческого FXI и/или FXIa. В частности, легкая цепь может иметь последовательность, содержащую SEQ ID NOs: 21 или 41. В частности, выделенное антитело, или его антигенсвязывающие фрагменты, которое связывает FXI и/или FXIa, может иметь тяжелую цепь и легкую цепь, содержащие последовательности SEQ ID NOs: 11 и 21; или 31 и 41, соответственно.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, которое содержит тяжелую цепь, имеющую по меньшей мере 90% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 11 или 31, при этом указанное антитело связывает FXI и/или FXIa. В одном аспекте выделенное антитело, или его антигенсвязывающие фрагменты, также содержит легкую цепь, имеющую по меньшей мере 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 21 или 41.
Изобретение также относится к выделенному антителу, или его антигенсвязывающим фрагментам, содержащему легкую цепь, имеющую по меньшей мере 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 21 или 41, при этом указанное антитело связывает FXI и/или FXIa.
Изобретение также относится к композициям, содержащим выделенное антитело, или его антигенсвязывающие фрагменты, описанное в настоящем документе, а также к композициям антитела в сочетании с фармацевтически приемлемым носителем. В частности, изобретение также относится к фармацевтическим композициям, содержащим антитело, или его антигенсвязывающие фрагменты, из таблицы 1, такое как, например, антитело NOV1090 и NOV1401. Изобретение также относится к фармацевтическим композициям, содержащим сочетание двух или более выделенных антител, или их антигенсвязывающих фрагментов, из таблицы 1.
Изобретение также относится к последовательности выделенной нуклеиновой кислоты, кодирующей вариабельный домен тяжелой цепи, имеющий последовательность, выбранную из SEQ ID NOs: 9 и 29. В частности, последовательность нуклеиновой кислоты имеет по меньшей мере 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 10 и 30. В следующем аспекте изобретения последовательность представляет собой SEQ ID NOs: 10 или 30.
Изобретение также относится к последовательности выделенной нуклеиновой кислоты, кодирующей вариабельный домен легкой цепи, имеющий последовательность, выбранную из SEQ ID NOs: 20 и 40. В частности, последовательность нуклеиновой кислоты имеет по меньшей мере 80%, 85%, 90%, 95%, 97%, 98% или 99% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 20 и 40. В следующем аспекте изобретения последовательность представляет собой SEQ ID NOs: 20 и 40.
Изобретение также относится к выделенной нуклеиновой кислоте, содержащей последовательность, кодирующую полипептид, содержащий вариабельный домен легкой цепи, имеющую по меньшей мере 90% идентичности последовательности с последовательностью, выбранной из группы, состоящей из SEQ ID NOs: 20 и 40.
Изобретение также относится к вектору, содержащему одну или более молекул нуклеиновой кислоты, описанных в настоящем документе.
Изобретение также относится к выделенной клетке-хозяину, содержащей последовательность рекомбинантной ДНК, кодирующую тяжелую цепь антитела, описанного выше, и вторую последовательность рекомбинантной ДНК, кодирующую легкую цепь антитела, описанного выше, при этом указанные последовательности ДНК функционально связаны с промотором и способны экспрессироваться в клетке-хозяине. Предусмотрено, что антитело может представлять собой человеческое моноклональное антитело. Также предусмотрено, что клетка-хозяин представляет собой клетку млекопитающего, отличного от человека.
Изобретение также относится к способу уменьшения экспрессии FXI и/или FXIa, и/или активации внутреннего и/или общего путей активации свертывания крови, включающему этап создания контакта клетки с эффективным количеством композиции, содержащей выделенное антитело, или его антигенсвязывающие фрагменты, описанное в настоящем документе.
Изобретение также относится к способу ингибирования связывания FXI и/или FXIa с FIX, включающему этап создания контакта клетки с эффективным количеством композиции, содержащей выделенное антитело, или его антигенсвязывающие фрагменты, описанное в настоящем документе.
Предусмотрено, что клетка представляет собой человеческую клетку. Также предусмотрено, что клетка находится в организме субъекта. В одном варианте осуществления предусмотрено, что клетка представляет собой тромбоцит. Также предусмотрено, что субъект является человеком.
Изобретение также относится к способу лечения, ослабления или предотвращения тромбоэмболического заболевания у субъекта, включающему этап введения субъекту эффективного количества композиции, содержащей антитело, или его антигенсвязывающие фрагменты, описанное в настоящем документе. В одном аспекте тромбоэмболическое заболевание представляет собой тромботическое заболевание (например, тромбоз, тромбозный инсульт, фибрилляцию предсердий, предотвращение инсульта при фибрилляции предсердий (ПИФП), тромбоз глубоких вен, венозную тромбоэмболию и легочную эмболию). Предусмотрено, что субъект является человеком.
Любое из вышеупомянутых выделенных антител, или их антигенсвязывающих фрагментов, может представлять собой моноклональное антитело, или его антигенсвязывающие фрагменты.
Неограничивающие варианты осуществления изобретения описаны в следующих пунктах:
1. Выделенное анти-FXI и/или анти-FXIa антитело, или его фрагмент, которое связывается внутри каталитического домена FXI и/или FXIa.
2. Выделенное антитело, или его фрагмент, которое связывает один или более эпитопов анти-FXI и/или FXIa, при этом эпитоп содержит два или более аминокислотных остатков из Pro410, Arg413, Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472, Lys473, Met474, Ala475, Glu476, Tyr521, Arg522, Lys523, Leu524, Arg525, Asp526, Lys527, Arg548, His552, Ser575, Ser594, Trp595, Gly596, Glu597, Arg602, Glu603 и Arg604.
3. Выделенное антитело, или фрагмент, по п. 2, при этом эпитоп содержит четыре или более аминокислотных остатков из Pro410, Arg413, Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472, Lys473, Met474, Ala475, Glu476, Tyr521, Arg522, Lys523, Leu524, Arg525, Asp526, Lys527, Arg548, His552, Ser575, Ser594, Trp595, Gly596, Glu597, Arg602, Glu603 и Arg604.
4. Выделенное антитело, или фрагмент, по п. 2, при этом эпитоп содержит шесть или более аминокислотных остатков из Pro410, Arg413, Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472, Lys473, Met474, Ala475, Glu476, Tyr521, Arg522, Lys523, Leu524, Arg525, Asp526, Lys527, Arg548, His552, Ser575, Ser594, Trp595, Gly596, Glu597, Arg602, Glu603 и Arg604.
5. Выделенное антитело, или фрагмент, по п. 2, при этом эпитоп содержит восемь или более аминокислотных остатков из Pro410, Arg413, Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472, Lys473, Met474, Ala475, Glu476, Tyr521, Arg522, Lys523, Leu524, Arg525, Asp526, Lys527, Arg548, His552, Ser575, Ser594, Trp595, Gly596, Glu597, Arg602, Glu603 и Arg604.
6. Выделенное антитело, или фрагмент, по п. 2, при этом эпитоп содержит остатки Pro410, Arg413, Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472, Lys473, Met474, Ala475, Glu476, Tyr521, Arg522, Lys523, Leu524, Arg525, Asp526, Lys527, Arg548, His552, Ser575, Ser594, Trp595, Gly596, Glu597, Arg602, Glu603 и Arg604.
7. Выделенное антитело, или фрагмент, по п. 2, при этом эпитоп содержит аминокислотные остатки Pro410, Arg413, Lys527 и один или более аминокислотных остатков из Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472, Lys473, Met474, Ala475, Glu476, Tyr521, Arg522, Lys523, Leu524, Arg525, Asp526, Arg548, His552, Ser575, Ser594, Trp595, Gly596, Glu597, Arg602, Glu603 и Arg604.
8. Выделенное антитело, или фрагмент, по п. 2, при этом эпитоп содержит аминокислотные остатки Pro410, Arg413, Lys527 и четыре или более аминокислотных остатков из Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472, Lys473, Met474, Ala475, Glu476, Tyr521, Arg522, Lys523, Leu524, Arg525, Asp526, Arg548, His552, Ser575, Ser594, Trp595, Gly596, Glu597, Arg602, Glu603 и Arg604.
9. Выделенное антитело, или фрагмент, по п. 2, при этом эпитоп содержит аминокислотные остатки Pro410, Arg413, Lys527 и шесть или более аминокислотных остатков из Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472, Lys473, Met474, Ala475, Glu476, Tyr521, Arg522, Lys523, Leu524, Arg525, Asp526, Arg548, His552, Ser575, Ser594, Trp595, Gly596, Glu597, Arg602, Glu603 и Arg604.
10. Выделенное анти-FXI и/или анти-FXIa антитело, или его фрагмент, которое связывается внутри каталитического домена FXI и/или FXIa, при этом указанное антитело, или фрагмент, блокирует связывание FXI и/или FXIa с одним или более из фактора IX, фактора XIIa и тромбина.
11. Выделенное антитело, или фрагмент, по п. 10, при этом указанное антитело, или фрагмент, блокирует связывание FXI и/или FXIa с одним или более из фактора IX, фактора XIIa или тромбина, и другими компонентами пути активации свертывания крови.
12. Выделенное антитело, или фрагмент, по п. 1, при этом указанное антитело, или фрагмент, блокирует связывание одного или более из FIX, FXI и FXIa с рецепторами тромбоцитов.
13. Выделенное антитело, или фрагмент, по п. 1, при этом указанное антитело, или фрагмент, предотвращает активацию внутреннего или общего путей активации свертывания крови.
14. Выделенное антитело, или его фрагмент, которое связывает человеческий белок FXI и/или FXIa с KD менее или равной 34 нМ при измерении в анализе BIACORE™, или менее или равной 4 пМ при измерении в анализе титрования равновесного раствора (ТРР).
15. Выделенное антитело, или фрагмент, по п. 1, при этом указанное антитело, или фрагмент, содержит по меньшей мере одну определяющую комплементарность область, имеющую по меньшей мере 90% идентичности с по меньшей мере одной из областей CDR, приведенных в таблице 1.
16. Выделенное антитело, или фрагмент, по п. 1, при этом указанное антитело, или фрагмент, содержит CDR1, CDR2 и CDR3 из таблицы 1.
17. Выделенный вариант антитела, или фрагмента, по п. 1, при этом указанное антитело, или фрагмент, содержит CDR1, CDR2 и CDR3 из таблицы 1, и при этом вариант имеет по меньшей мере одно-четыре аминокислотных изменений в одной из областей CDR1, CDR2 или CDR3.
18. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит CDR3 тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 5 и 25.
20. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит VH, выбранную из группы, состоящей из SEQ ID NO: 9 и 29, или аминокислотную последовательность, имеющую 90% идентичности с ней; и VL, выбранную из группы, состоящей из SEQ ID NO: 19 и 39, или аминокислотную последовательность, имеющую 90% идентичности с ней.
21. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит VH, выбранную из группы, состоящей из SEQ ID NO: 9 и 29, или аминокислотную последовательность, имеющую 95% идентичности с ней; и VL, выбранную из группы, состоящей из SEQ ID NO: 19 и 39, или аминокислотную последовательность, имеющую 95% идентичности с ней.
22. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит VH, выбранную из группы, состоящей из SEQ ID NO: 9 и 29, или аминокислотную последовательность, имеющую 97% идентичности с ней; и VL, выбранную из группы, состоящей из SEQ ID NO: 19 и 39, или аминокислотную последовательность, имеющую 97% идентичности с ней.
23. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит последовательность вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 9 и 29.
24. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит последовательность вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 19 и 39.
25. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит последовательность вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 9 и 29; и последовательность вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 19 и 39.
26. Выделенное антитело, или фрагмент, по п. 1, которое выбирают из группы, состоящей из антитела, или фрагмента, содержащего последовательность вариабельной области тяжелой цепи SEQ ID NO: 9 и последовательность вариабельной области легкой цепи SEQ ID NO: 19, и антитела, или фрагмента, содержащего последовательность вариабельной области тяжелой цепи SEQ ID NO: 29 и последовательность вариабельной области легкой цепи SEQ ID NO: 39.
27. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит CDR1 вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 46; CDR2, выбранную из группы, состоящей из SEQ ID NO: 4; CDR3, выбранную из группы, состоящей из SEQ ID NO: 5; CDR1 вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 33; CDR2, выбранную из группы, состоящей из SEQ ID NO: 14; и CDR3, выбранную из группы, состоящей из SEQ ID NO: 15.
28. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит CDR1 вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 3 и 23; CDR2, выбранную из группы, состоящей из SEQ ID NO: 4 и 24; CDR3, выбранную из группы, состоящей из 5 и 25; CDR1 вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 13 и 33; CDR2, выбранную из группы, состоящей из SEQ ID NO: 14 и 34; и CDR3, выбранную из группы, состоящей из SEQ ID NO: 15 и 35.
29. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит CDR1 вариабельной области тяжелой цепи, выбранную из группы, состоящей из SEQ ID NO: 6 и 26; CDR2, выбранную из группы, состоящей из SEQ ID NO: 7 и 27; CDR3, выбранную из группы, состоящей из 8 и 28; CDR1 вариабельной области легкой цепи, выбранную из группы, состоящей из SEQ ID NO: 16 и 36; CDR2, выбранную из группы, состоящей из SEQ ID NO: 17 и 37; и CDR3, выбранную из группы, состоящей из SEQ ID NO: 18 и 38.
30. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит CDR1 вариабельной области тяжелой цепи SEQ ID NO: 3; CDR2 вариабельной области тяжелой цепи SEQ ID NO: 4; CDR3 вариабельной области тяжелой цепи SEQ ID NO: 5; CDR1 вариабельной области легкой цепи SEQ ID NO: 13; CDR2 вариабельной области легкой цепи SEQ ID NO: 14 и CDR3 вариабельной области легкой цепи SEQ ID NO: 15.
31. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит CDR1 вариабельной области тяжелой цепи SEQ ID NO: 23; CDR2 вариабельной области тяжелой цепи SEQ ID NO: 24; CDR3 вариабельной области тяжелой цепи SEQ ID NO: 25; CDR1 вариабельной области легкой цепи SEQ ID NO: 33; CDR2 вариабельной области легкой цепи SEQ ID NO: 34 и CDR3 вариабельной области легкой цепи SEQ ID NO: 35.
32. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит CDR1 вариабельной области тяжелой цепи SEQ ID NO: 6; CDR2 вариабельной области тяжелой цепи SEQ ID NO: 7; CDR3 вариабельной области тяжелой цепи SEQ ID NO: 8; CDR1 вариабельной области легкой цепи SEQ ID NO: 16; CDR2 вариабельной области легкой цепи SEQ ID NO: 17 и CDR3 вариабельной области легкой цепи SEQ ID NO: 18.
33. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, содержит CDR1 вариабельной области тяжелой цепи SEQ ID NO: 26; CDR2 вариабельной области тяжелой цепи SEQ ID NO: 27; CDR3 вариабельной области тяжелой цепи SEQ ID NO: 28; CDR1 вариабельной области легкой цепи SEQ ID NO: 36; CDR2 вариабельной области легкой цепи SEQ ID NO: 37 и CDR3 вариабельной области легкой цепи SEQ ID NO: 38.
34. Фармацевтическая композиция, содержащая антитело, или его фрагмент, по одному из предшествующих пунктов и фармацевтически приемлемый носитель.
35. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, связывает тот же эпитоп, что и выделенное антитело, или фрагмент, по любому из предшествующих пунктов.
36. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, конкурирует за связывание человеческого белка FXI и/или FXIa с выделенным антителом, или фрагментом по любому из предшествующих пунктов.
37. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, выбирают из группы, состоящей из NOV1090 и NOV1401.
38. Способ лечения тромбоэмболического заболевания, включающий введение субъекту, страдающему тромбоэмболическим заболеванием, эффективного количества фармацевтической композиции, содержащей антитело, или фрагмент, по любому из предшествующих пунктов.
39. Способ по п. 38, отличающийся тем, что субъект страдает одним или более из ишемического инсульта, связанного с фибрилляцией предсердий, и тромбоза глубоких вен.
40. Способ по п. 38, отличающийся тем, что субъект страдает ишемическим инсультом, связанным с фибрилляцией предсердий.
41. Способ лечения тромбоэмболического заболевания, включающий введение субъекту, страдающему тромбоэмболическим заболеванием, эффективного количества фармацевтической композиции, содержащей антитело, или фрагмент, по любому из предшествующих пунктов в сочетании с терапией статинами.
42. Лекарственное средство, содержащее антитело по любому из предшествующих пунктов.
43. Нуклеиновая кислота, кодирующая одно или более из антител по любому из предшествующих пунктов.
44. Вектор, содержащий нуклеиновую кислоту по п. 43.
45. Клетка-хозяин, содержащая вектор по п. 44.
46. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, при связывании с каталитическим доменом активного FXI (FXIa) приводит к изменению конформации FXIa в неактивную конформацию, при которой 4 N-концевых остатки, петли 145, 188 и 220 смещены и/или являются неупорядоченными по сравнению с активной конформацией.
47. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, при связывании с FXI предотвращает принятие каталитическим доменом FXI активной конформации, при которой петли 145, 188 и 220 упорядочены, как в структуре каталитического домена FXIa.
48. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, при связывании с FXI предотвращает принятие каталитическим доменом FXI активной конформации, при которой 4 N-концевых остатка, петли 145, 188 и 220 упорядочены, как в структуре каталитического домена FXIa.
49. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, при связывании с FXI предотвращает принятие каталитическим доменом FXI активной конформации, вызывая конформационные изменения в структуре зимогена, впоследствии приводящие к ингибированной конформации FXI, близкой к той, которая наблюдается при связывании с FXIa.
50. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, при связывании с FXI и/или FXIa и образовании комплекса антитело: антиген с каталитическим доменом FXI и/или FXIa вызывает смещение и/или неупорядоченность петель 145, 188 и 220 по сравнению со структурой каталитического домена не находящегося в комплексе активного фактора XI (FXIa).
51. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, при связывании с FXI и/или FXIa и образовании комплекса антитело: антиген с каталитическим доменом FXI и/или FXIa вызывает смещение и/или неупорядоченность 4 N-концевых остатков, петель 145, 188 и 220 по сравнению со структурой каталитического домена не находящегося в комплексе активного фактора XI (FXIa).
52. Выделенное антитело, или фрагмент, по п. 1, при этом антитело, или фрагмент, связывает активный FXI (FXIa) и вызывает изменение конформации каталитического домена FXI (FXIa) в неактивную конформацию, при которой петли 145, 188 и 220 смещены и/или являются неупорядоченными по сравнению с активной конформацией.
ОПРЕДЕЛЕНИЯ
Если нет иных указаний, все технические и научные термины, используемые в настоящем документе, имеют то значение, которое им обычно придают специалисты в области, к которой относится данное изобретение.
Термины «белок FXI», «антиген FXI» и «FXI» используются взаимозаменяемо и относятся к белку фактора XI у разных биологических видов. Фактор XI представляет собой фактор свертывания крови XI в плазме млекопитающих, гликопротеин, присутствующий в плазме человека в концентрации 25-30 нМ в виде зимогена, который затем в результате ограниченного протеолиза превращается в активную сериновую протеазу и участвует во внутреннем или общем путях активации свертывания крови.
Термины «белок FXIa», «антиген FXIa» и «FXIa» используются взаимозаменяемо и относятся к активированному белку FXI у разных биологических видов. Зимоген фактор XI превращается в его активную форму, фактор свертывания крови XIa (FXIa), либо через контактную фазу свертывания крови, либо через опосредованную тромбином активацию на поверхности тромбоцитов. В процессе этой активации фактора XI внутренняя пептидная связь расщепляется в каждой из двух цепей, в результате чего образуется активированный фактор XIa, сериновая протеаза, состоящая из двух тяжелых и двух легких цепей, связанных дисульфидными связями. Эта сериновая протеаза FXIa превращает фактор свертывания крови IX в IXa, который впоследствии активирует фактор свертывания крови X (Xa). Xa затем может опосредовать активацию фактора свертывания крови II/тромбина. Например, FXI человека имеет последовательность, приведенную в таблице 1 (SEQ ID NO:1), и был описан в предыдущих докладах и в литературе (Mandle RJ Jr, et al. (1979) Blood; 54(4): 850; эталонная последовательность в NCBI: AAA51985).
В контексте данного изобретения термины «FXI» и «FXIa» (и тому подобные) включают мутанты и варианты природного белка FXI и FXIa, соответственно, которые имеют по существу ту же аминокислотную последовательность, что и природная первичная структура (аминокислотная последовательность), приведенная в вышеупомянутых литературных источниках.
Используемые в настоящем документе термины «каталитический домен», «каталитический домен сериновой протеазы» и аналогичные термины означают аминокислоты от Ile370 до Val607, считая от остатка Glu1 на N-конце зрелого белка, присутствующего в системе кровообращения. Их также можно описывать, как остатки 388-625 на C-конце FXI. Используемый в настоящем документе термин «активный центр» означает каталитическую триаду, состоящую из аминокислот His413, Asp462 и Se557. (Bane and Gailani (2014) Drug Disc. 19(9)).
Термин «примерно» применительно к численному значению x означает, например, x ± 10%.
Используемый в настоящем документе термин «антитело» означает целое антитело и любой антигенсвязывающий фрагмент (то есть, «антигенсвязывающую часть»), либо их одиночную цепь. Целое антитело представляет собой гликопротеин, содержащий по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (обозначенной в настоящем документе VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (обозначенной в настоящем документе VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL можно дополнительно подразделять на области гипервариабельности, называемые определяющими комплементарность областями (CDR), которые перемежаются с более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбоксильному концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3 FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Используемый в настоящем документе термин «антигенсвязывающая часть» или «антигенсвязывающий фрагмент» антитела означает один или более фрагментов интактного антитела, которые сохраняют способность специфически связывать конкретный антиген (например, фактор XIa (FXIa)). Антигенсвязывающие функции антитела могут выполняться фрагментами интактного антитела. Примеры связывающих фрагментов, охваченных термином «антигенсвязывающая часть» или «антигенсвязывающий фрагмент» антитела, включают Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; F(ab)2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, связанные дисульфидным мостом в шарнирной области; Fd-фрагмент, состоящий из доменов VH и CH1; Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела; однодоменное антитело (dAb) (Ward et al., 1989 Nature 341: 544-546), которое состоит из домена VH или домена VL, а также выделенную определяющую комплементарность область (CDR).
Кроме того, хотя два домена Fv-фрагмента, VL и VH, закодированы разными генами, они могут быть соединены рекомбинантными методами при помощи искусственного пептидного линкера, который позволяет им быть полученными в виде одной белковой цепи, в которой области VL и VH спариваются, с образованием одновалентной молекулы (известной, как одноцепочечная Fv (scFv); смотри, например, Bird et al., 1988 Science 242: 423-426; и Huston et al., 1988 Proc. Natl. Acad. Sci. 85: 5879-5883). Такие одноцепочечные антитела содержат одну или более антигенсвязывающих частей, или фрагментов, антитела. Эти фрагменты антитела получают с использованием общепринятых методов, известных специалистам в данной области, и проводят скрининг фрагментов на пригодность таким же образом, как и интактных антител.
Антигенсвязывающие фрагменты также можно встраивать в однодоменные антитела, макситела, минитела, интратела, диатела, триатела, тетратела, v-NAR и бис-scFv (смотри, например, Hollinger and Hudson, 2005, Nature Biotechnology, 23, 9, 1126-1136). Антигенсвязывающие части антител можно прививать на каркасы на основе полипептидов, таких как фибронектин III типа (Fn3) (смотри патент США № 6703199, в котором описаны монотела на основе полипептида фибронектина).
Антигенсвязывающие фрагменты можно встраивать в одноцепочечные молекулы, содержащие пару тандемных Fv-сегментов (VH-CH1-VH-CH1), которые, совместно с комплементарными полипептидами легкой цепи, образуют пару антигенсвязывающих областей (Zapata et al., 1995 Protein Eng. 8(10): 1057-1062; и патент США № 5641870).
Используемый в настоящем документе термин «аффинность» означает силу взаимодействия между антителом и антигеном в отдельных антигенных сайтах. В каждом антигенном сайте вариабельная область «плеча» антитела взаимодействует за счет слабых нековалентных сил с антигеном в многочисленных точках; чем больше взаимодействий, тем сильнее аффинность. Используемый в настоящем документе термин «высокая аффинность» применительно к антителу или его антигенсвязывающим фрагментам (например, Fab-фрагменту), как правило, относится к антителу, или антигенсвязывающему фрагменту, имеющему KD 10-9 M или менее (например, KD 10-10 M или менее, KD 10-11 M или менее, KD 10-12 M или менее, KD 10-13 M или менее, KD 10-14 M или менее и так далее).
Термин «аминокислота» относится к природным и синтетическим аминокислотам, а также аминокислотным аналогам и аминокислотным миметикам, которые действуют аналогично природным аминокислотам. Природными аминокислотами являются аминокислоты, кодируемые генетическим кодом, а также аминокислоты, которые впоследствии были модифицированы, например, гидроксипролин, γ-карбоксиглутамат и O-фосфосерин. Аминокислотные аналоги представляют собой соединения, имеющие ту же основную химическую структуру, что и природная аминокислота, то есть, альфа-атом углерода, связанный с атомом водорода, карбоксильную группу, аминогруппу и R-группу, например, гомосерин, норлейцин, метионин-сульфоксид, метионин-метил-сульфоний. Такие аналоги имеют модифицированные R-группы (например, норлейцин) или модифицированные пептидные каркасы, но сохраняют ту же основную химическую структуру, что и природные аминокислоты. Аминокислотные миметики представляют собой химические соединения, имеющие структуру, которая отличается от общей химической структуры аминокислот, но которые действуют аналогично природным аминокислотам.
Используемый в настоящем документе термин «специфичность связывания» означает способность отдельного связывающего сайта антитела взаимодействовать только с одной антигенной детерминантой.
Выражение «специфически (или избирательно) связывает» применительно к антителу (например, FXI и/или FXIa-связывающему антителу) означает реакцию связывания, которая обусловлена присутствием узнаваемого антигена (например, FXI и/или FXIa человека или FXI и/или FXIa яванского макака) в гетерогенной популяции белков и других биологических молекул. Выражения «антитело, узнающее антиген» и «антитело, специфичное для антигена» в настоящем документе используют взаимозаменяемо с выражением «антитело, которое специфически связывает антиген».
Термин «опосредуемый FXI и/или FXIa» относится к тому факту, что FXI и/или FXIa опосредует внутренний и/или общий пути активации свертывания крови за счет прямой или непрямой активации фактора IX (также известного как FIX), фактора X (FX) и/или тромбина, и/или за счет связывания с рецепторами тромбоцитов.
Термин «гемостаз» относится к основным механизмам остановки потока крови в участках повреждения и восстановления проходимости сосудов во время заживления ран, соответственно. При нормальном гемостазе и патологическом тромбозе три механизма активируются одновременно: первичный гемостаз, представляющий собой взаимодействие активированных тромбоцитов с сосудистой стенкой, образование фибрина и процесс, называемый фибринолизом.
Термины «свертывание крови и каскад свертывания крови», «каскадная модель свертывания крови», и тому подобные, относятся к белковой системе, которая служит для стабилизации сгустка, образующегося для закупоривания раны. Путь активации свертывания крови представляет собой протеолитический каскад. Каждый фермент в данном пути присутствует в плазме в виде зимогена (в неактивной форме), который при активации подвергается протеолитическому расщеплению, высвобождая активный фактор из молекулы-предшественника. Каскад свертывания крови действует в виде серии петель положительной и отрицательной обратной связи, контролирующих процесс активации. Конечной целью пути активации является образование тромбина, который затем может превращать растворимый фибриноген в фибрин, образующий сгусток.
Процесс образования тромбина можно разделить на три фазы: внутренний и внешний пути, которые обеспечивают альтернативные пути образования активного фактора свертывания крови: FXa (активированный фактор-X), и завершающий общий путь, который приводит к образованию тромбина (Hoffman M.M. and Monroe D.M. (2005) Curr Hematol Rep. 4: 391-396; Johne J, et al. (2006) Biol Chem. 387: 173-178).
«Агрегация тромбоцитов» означает процесс, за счет которого при возникновении разрыва кровеносного сосуда высвобождаются вещества, обычно не контактирующие непосредственно с кровотоком. Эти вещества (главным образом, коллаген и фактор фон Виллебранда) позволяют тромбоцитам прикрепляться к поврежденной поверхности. После прикрепления тромбоцита к поверхности он высвобождает химические вещества, которые привлекают дополнительные тромбоциты к области повреждения, это явление называют агрегацией тромбоцитов. Эти два процесса являются первыми ответами, направленными на остановку кровотечения.
Используемый в настоящем документе термин «тромбоэмболическое заболевание» или аналогичные термины означают любые состояния или заболевания, при которых внутренний и/или общий пути активации свертывания крови аберрантно активированы или не являются естественным образом дезактивированными (например, без применения терапевтических средств). Эти состояния включают, но не ограничиваются ими, тромбозный инсульт, фибрилляцию предсердий, предотвращение инсульта при фибрилляции предсердий (ПИФП), тромбоз глубоких вен, венозную тромбоэмболию и легочную эмболию. Они также могут включать состояния, связанные с введением катетера (например, катетера Хикмана у онкологических пациентов), при которых катетеры забиваются тромбами, и с экстракорпоральной мембранной оксигенацией (ЭКМО), при которой в трубках образуются сгустки.
Используемый в настоящем документе термин «тромбоэмболические» или аналогичные термины также могут относится к любому количеству из перечисленных ниже состояний, для предотвращения или лечения которых можно использовать анти-FXI и/или анти-FXIa Ат, или их антигенсвязывающие фрагменты, по изобретению:
- тромбоэмболия у субъектов с предполагаемой или подтвержденной сердечной аритмией, такой как пароксизмальная, персистентная или перманентная фибрилляция предсердий или трепетание предсердий;
- предотвращение инсульта при фибрилляции предсердий (ПИФП), одной из подгрупп пациентов являются пациенты с ФП, подвергаемые чрескожному коронарному вмешательству (ЧКВ);
- лечение острых венозных тромбоэмболических событий (ВТС) и расширенная вторичная профилактика ВТС у пациентов с высоким риском кровотечений;
- церебральные и сердечно-сосудистые события во вторичной профилактике после транзиторной ишемической атаки (ТИА) или не инвалидизирующего инсульта и предотвращение тромбоэмболических событий при сердечной недостаточности с синусовым ритмом;
- образование сгустка в левом предсердии и тромбоэмболия у субъектов, подвергаемых кардиоверсии в связи с сердечной аритмией;
- тромбоз до, в процессе и после процедуры абляции в связи с сердечной аритмией;
- венозный тромбоз, сюда относятся, но без ограничения, лечение и вторичная профилактика тромбоза глубоких и поверхностных вен в нижних конечностях или верхних конечностях, тромбоз в брюшных и грудных венах, синус-тромбоз и тромбоз яремных вен;
- тромбоз на любой искусственной поверхности в венах, такой как катетер или провода кардиостимулятора;
- легочная эмболия у пациентов с венозным тромбозом или без него;
- хроническая тромбоэмболическая легочная гипертензия (ХТЭЛГ);
- артериальный тромбоз на месте оторвавшейся атеросклеротической бляшки, тромбоз на внутриартериальном протезе или катетере и тромбоз в кажущихся нормальными артериях, включая, но без ограничения, острые коронарные синдромы, инфаркт миокарда с повышением ST-сегмента, инфаркт миокарда без повышения ST-сегмента, нестабильную стенокардию, тромбоз стентов, тромбоз любой искусственной поверхности в системе артериального кровообращения и тромбоз легочных артерий у субъектов с легочной гипертензией и без нее;
- тромбоз и тромбоэмболия у пациентов, подвергаемых чрескожному коронарному вмешательству (ЧКВ);
- кардиоэмболические и криптогенные инсульты;
- тромбоз у пациентов с инвазивными и неинвазивными злокачественными новообразованиями;
- тромбоз в полостном катетере;
- тромбоз и тромбоэмболия у пациентов в тяжелом состоянии;
- сердечный тромбоз и тромбоэмболия, включая, но без ограничения, сердечный тромбоз после инфаркта миокарда, сердечный тромбоз, связанный с таким состоянием, как сердечная аневризма, миокардиальный фиброз, увеличение сердца и сердечная недостаточность, миокардит и искусственные поверхности в сердце;
- тромбоэмболия у пациентов с болезнью клапанов сердца, с фибрилляцией или без фибрилляции предсердий;
- тромбоэмболия в механических или биологических протезах клапанов;
- тромбоэмболия у пациентов, имеющих естественные или искусственные заплаты в сердце, артериальные или венозные проводящие трубки после операции на сердце по поводу простых или сложных пороков сердца;
- венозный тромбоз и тромбоэмболия после операции по замене коленного сустава, операции по замене тазобедренного сустава, а также ортопедической операции, торакальной или абдоминальной хирургической операции;
- артериальный или венозный тромбоз после нейрохирургической операции, включая интракраниальные операции и операции на спинном мозге;
- врожденная или приобретенная тромбофилия, включая, но без ограничения, мутацию фактора V Лейдена, мутацию протромбина, дефицит антитромбина III, белка C и белка S, мутацию фактора XIII, семейную дисфибриногенемию, врожденный дефицит плазминогена, повышенные уровни фактора XI, серповидно-клеточное заболевание, антифосфолипидный синдром, аутоиммунное заболевание, хроническое заболевание кишечника, нефротический синдром, гемолитическую уремию, миелопролиферативное заболевание, диссеминированное внутрисосудистое свертывание крови, пароксизмальную ночную гемоглобинурию и вызванную гепарином тромбопению;
- тромбоз и тромбоэмболия при хронической почечной недостаточности; и
- тромбоз и тромбоэмболия у пациентов, подвергаемых гемодиализу, и у пациентов, подвергаемых экстракорпоральной мембранной оксигенации.
Термин «химерное антитело» означает молекулу антитела, в котором (a) константная область, или ее часть, изменена, замещена или заменена таким образом, что антигенсвязывающий сайт (вариабельная область) связан с константной областью другого или измененного класса, с другой эффекторной функцией и/или из другого биологического вида, или полностью иной молекулой, которая придает новые свойства химерному антителу, например, ферментом, токсином, гормоном, фактором роста, лекарственным средством и так далее; или (b) вариабельная область, или ее часть, изменена, замещена или заменена вариабельной областью, имеющей другую или измененную антигенную специфичность. Например, мышиное антитело может быть модифицировано путем замены его константной области на константную область из человеческого иммуноглобулина. Вследствие замены на человеческую константную область химерное антитело может сохранять его специфичность узнавания антигена и при этом иметь меньшую антигенность в организме человека по сравнению с исходным мышиным антителом.
Термин «консервативно модифицированный вариант» применим как к аминокислотным, так и к нуклеотидным последовательностям. Применительно к конкретным последовательностям нуклеиновой кислоты, консервативно модифицированными вариантами являются такие нуклеиновые кислоты, которые кодируют идентичные или по существу идентичные аминокислотные последовательности, или, если нуклеиновая кислота не кодирует аминокислотную последовательность, то по существу идентичные последовательности. Из-за вырожденности генетического кода очень многие функционально идентичные нуклеиновые кислоты кодируют какой-либо конкретный белок. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, где аланин определяется кодоном, кодон может быть заменен на любой из соответствующих приведенных кодонов без изменения закодированного полипептида. Такие вариации нуклеиновой кислоты являются «молчащими вариациями», которые представляют собой один из видов консервативно модифицированных вариаций. В настоящем документе каждая нуклеотидная последовательность, кодирующая полипептид, также включает любую возможную молчащую вариацию нуклеиновой кислоты. Специалист в данной области понимает, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно является единственным кодоном для метионина, и TGG, который обычно является единственным кодоном для триптофана) может быть изменен, с получением функционально идентичной молекулы. Соответственно, каждая молчащая вариация нуклеиновой кислоты, которая кодирует полипептид, подразумевается для каждой описанной последовательности.
В случае полипептидных последовательностей «консервативно модифицированные варианты» включают отдельные замены, делеции или добавления в полипептидную последовательность, приводящие к замене аминокислоты на химически аналогичную аминокислоту. Таблицы консервативных замен, представляющие функционально аналогичные аминокислоты, хорошо известны в данной области. Такие консервативно модифицированные варианты дополняют, но не исключают, полиморфные варианты, межвидовые гомологи и аллели по изобретению. Следующие восемь групп включают аминокислоты, которые являются консервативными заменами друг для друга: 1) аланин (A), глицин (G); 2) аспарагиновая кислота (D), глутаминовая кислота (E); 3) аспарагин (N), глутамин (Q); 4) аргинин (R), лизин (K); 5) изолейцин (I), лейцин (L), метионин (M), валин (V); 6) фенилаланин (F), тирозин (Y), триптофан (W); 7) серин (S), треонин (T) и 8) цистеин (C), метионин (M) (смотри, например, Creighton, Proteins (1984)). В некоторых вариантах осуществления термин «консервативные модификации последовательности» используют для обозначения аминокислотных модификаций, которые существенно не влияют или не изменяют характеристики связывания антитела, содержащего аминокислотную последовательность.
Термин «эпитоп» означает белковую детерминанту, способную специфически связываться с антителом. Эпитопы, как правило, состоят из химически активных поверхностных групп молекул, таких как аминокислоты или сахарные боковые цепи, и, как правило, имеют специфические трехмерные структурные характеристики, а также специфические зарядные характеристики. Конформационные и не конформационные эпитопы отличаются тем, что связывание с первыми, но не с последними, теряется в присутствии денатурирующих растворителей. Говорят, что два антитела «конкурируют», если показано, что одно антитело связывает тот же эпитоп, что и второе антитело, в конкурентном анализе связывания любым из методов, хорошо известных специалистам в данной области.
Используемый в настоящем документе термин «человеческое антитело» включает антитела, имеющие вариабельные области, в которых как каркасные, так и CDR, области происходят из последовательностей человеческих антител. Кроме того, если антитело содержит константную область, константная область также происходит из таких человеческих последовательностей, например, человеческих последовательностей зародышевой линии, или мутантных вариантов человеческих последовательностей зародышевой линии. Человеческие антитела по изобретению могут включать аминокислотные остатки, не кодируемые человеческими последовательностями (например, мутации, внесенные за счет случайного или сайт-специфического мутагенеза in vitro, или за счет соматической мутации in vivo).
Термин «человеческое моноклональное антитело» относится к антителам, проявляющим одну специфичность связывания, в которых вариабельные области как в каркасных, так и в CDR, областях происходят из человеческих последовательностей. В одном варианте осуществления человеческие моноклональные антитела получают с использованием методов фагового дисплея для скрининга библиотек генов человеческих иммуноглобулинов.
«Гуманизированное» антитело представляет собой антитело, которое сохраняет реакционную способность антитела, не принадлежащего человеку, но при этом является менее иммуногенным в организме человека. Это может быть достигнуто, например, путем сохранения не принадлежащих человеку областей CDR и замены оставшихся частей антитела их аналогами из иммуноглобулинов человека (то есть, константной области, а также каркасных фрагментов вариабельной области). Смотри, например, Morrison et al., Proc. Natl. Acad. Sci. USA, 81: 6851-6855, 1984; Morrison and Oi, Adv. Immunol., 44: 65-92, 1988; Verhoeyen et al., Science, 239: 1534-1536, 1988; Padlan, Molec. Immun., 28: 489-498, 1991; и Padlan, Molec. Immun., 31: 169-217, 1994. Другие примеры технологий генетических модификаций человеческих антител включают, но без ограничения, технологию Xoma, раскрытую в US 5766886.
Термины «идентичные» или процент «идентичности» в контексте двух или более нуклеотидных, или полипептидных, последовательностей относятся к двум или более последовательностям, или подпоследовательностям, которые являются одинаковыми. Две последовательности являются «по существу идентичными», если две последовательности имеют определенную процентную долю аминокислотных остатков или нуклеотидов, которые являются одинаковыми (то есть, 60% идентичности, необязательно, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентичности на протяжении конкретной области или, если не указано, на протяжении всей последовательности) при сравнении и выравнивании для максимального соответствия в окне сравнения, или определенной области, что измеряют с использованием одного из следующих алгоритмов сравнения последовательностей или путем выравнивания вручную и визуальной инспекции. Необязательно, идентичность существует на протяжении области, которая имеет длину по меньшей мере примерно 50 нуклеотидов (или 10 аминокислот) или более, предпочтительно на протяжении области, которая имеет длину 100-500 или 1000, или более нуклеотидов (или 20, 50, 200 или более аминокислот).
При сравнении последовательностей, как правило, одна последовательность выступает в роли эталонной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей информацию о тестируемых и эталонных последовательностях вводят в компьютер, определяют координаты подпоследовательностей, при необходимости, и определяют параметры программы алгоритма сравнения последовательностей. Можно использовать параметры по умолчанию программы, или можно устанавливать альтернативные параметры. Затем алгоритм сравнения последовательностей рассчитывает процент идентичности последовательностей для тестируемых последовательностей относительно эталонной последовательности, на основе параметров программы.
Используемый в настоящем документе термин «окно сравнения» означает сегмент из любого числа смежных положений, выбранного из группы, состоящей из 20-600, как правило, от примерно 50 до примерно 200, чаще от примерно 100 до примерно 150, в котором последовательность может сравниваться с эталонной последовательностью из такого же числа смежных положений после оптимального выравнивания двух последовательностей. Способы выравнивания последовательностей для целей сравнения хорошо известны в данной области. Оптимальное выравнивание последовательностей для целей сравнения можно осуществлять, например, с помощью алгоритма локальной гомологии Смита-Уотермана (1970) Adv. Appl. Math. 2: 482c, с помощью алгоритма выравнивания областей гомологии Нидлмана-Вунша, J. Mol. Biol. 48: 443, 1970, методом поиска подобия Пирсона-Липмана, Proc. Nat'l. Acad. Sci. USA 85: 2444, 1988, с помощью компьютеризированной реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программ Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI), или путем выравнивания вручную и визуальной инспекции (смотри, например, Brent et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (Ringbou ed., 2003)).
Двумя примерами алгоритмов, которые подходят для определения процента идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al., (1977) Nuc. Acids Res. 25: 3389-3402; и Altschul et al., (1990) J. Mol. Biol. 215: 403-410, соответственно. Программа для выполнения анализов BLAST является общедоступной через Национальный центр биотехнологической информации. Этот алгоритм включает прежде всего идентификацию пар последовательностей с высоким показателем сходства (HSP) путем выявления коротких слов с длиной W в искомой последовательности, которые либо совпадают, либо удовлетворяют некоторому положительному пороговому значению T, при выравнивании со словом той же длины в базе данных для последовательностей. T называют порогом показателя сходства слов (Altschul et al., выше). Эти начальные удачно найденные слова, хиты, используют в качестве затравки для начала поиска более длинных содержащих их HSP. Хиты слов продлевают в обоих направлениях вдоль каждой последовательности до тех пор, пока кумулятивный показатель выравнивания может быть увеличен. Кумулятивные показатели рассчитывают с использованием, в случае нуклеотидных последовательностей, параметров M (премия за совпадение пары остатков; всегда >0) и N (штраф за несовпадение остатков; всегда <0). В случае аминокислотных последовательностей используют матрицу замен для расчета кумулятивного показателя. Продление хитов слов в каждом направлении прекращают, когда: кумулятивный показатель выравнивания падает на величину X от его максимального достигнутого значения; кумулятивный показатель доходит до нуля или ниже из-за накопления одного или более отрицательных показателей выравнивания остатков; или достигают конца любой из последовательностей. Параметры W, T и X алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) используют по умолчанию длину слова (W) 11, ожидание (E) 10, M=5, N=-4 и сравнение обеих цепей. Для аминокислотных последовательностей в программе BLASTP используют по умолчанию длину слова 3 и ожидание (E) 10, а также матрицу замен BLOSUM62 (смотри Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89: 10915, 1989), выравнивания (B) 50, ожидание (E) 10, M=5, N=-4 и сравнение обеих цепей.
Алгоритм BLAST также выполняет статистический анализ сходства между двумя последовательностями (смотри, например, Karlin and Altschul, Proc. Natl. Acad. Sci. USA 90: 5873-5787, 1993). Одной мерой сходства, предоставляемой алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), которая является показателем вероятности того, что совпадение между двумя нуклеотидными или аминокислотными последовательностями произошло бы случайно. Например, нуклеиновая кислота считается сходной с эталонной последовательностью, если наименьшая суммарная вероятность при сравнении тестируемой нуклеиновой кислоты с эталонной нуклеиновой кислотой составляет менее примерно 0,2, более предпочтительно менее примерно 0,01 и наиболее предпочтительно менее примерно 0,001.
Процент идентичности двух аминокислотных последовательностей также можно определять с использованием алгоритма Маерса-Миллера (Comput. Appl. Biosci., 4: 11-17, 1988), который включен в программу ALIGN (версия 2.0), с использованием таблицы весов замен остатков PAM120, штрафа за удлинение делеции 12 и штрафа за внесение делеции 4. Кроме того, процент идентичности двух аминокислотных последовательностей можно определять с использованием алгоритма Нидлмана-Вунша (J. Mol, Biol. 48: 444-453, 1970), который включен в программу GAP в пакете программ GCG (доступном в сети интернет по сетевому адресу gcg.com), с использованием либо матрицы Blossom 62, либо матрицы PAM250, штрафа за внесение делеции 16, 14, 12, 10, 8, 6, или 4 и штрафа за удлинение делеции 1, 2, 3, 4, 5 или 6.
Помимо процента идентичности последовательностей, описанного выше, другим указанием на то, что последовательности двух нуклеиновых кислот или полипептидов по существу идентичны, является то, что с полипептидом, кодируемым первой нуклеиновой кислотой, иммунологически перекрестно реагируют антитела, выработанные против полипептида, кодируемого второй нуклеиновой кислотой, как описано ниже. Таким образом, полипептид, как правило, является по существу идентичным второму полипептиду, например, если два пептида отличаются только консервативными заменами. Другим указанием на то, что две нуклеотидные последовательности по существу идентичны, является то, что две молекулы или их комплементы гибридизуются друг с другом в строгих условиях, как описано ниже. Еще одним указанием на то, что две нуклеотидные последовательности по существу идентичны, является то, что одни и те же праймеры можно использовать для амплификации последовательностей.
Термин «выделенное антитело» означает антитело, которое по существу свободно от других антител, имеющих другие антигенные специфичности (например, выделенное антитело, которое специфически связывает FXI и/или FXIa, по существу свободно от антител, которые специфически связывают антигены, отличные от FXI и/или FXIa). Выделенное антитело, специфически связывающее FXI и/или FXIa, может, однако, иметь перекрестную реакционную способность в отношении других антигенов. Кроме того, выделенное антитело может быть по существу свободным от другого клеточного материала и/или химических реагентов.
Термин «изотип» означает класс антитела (например, IgM, IgE, IgG, такое как IgG1 или IgG4), который определяется генами константной области тяжелой цепи. Изотип также включает модифицированные варианты одного из этих классов, в которых модификации были внесены для изменения функции Fc-области, например, для усиления или ослабления эффекторных функций, или связывания с Fc-рецепторами.
Используемый в настоящем документе термин «kасс» или «ka», означает скорость ассоциации в конкретном взаимодействии антитело-антиген, в то время как используемый в настоящем документе термин «kдисс» или «kd» означает скорость диссоциации в конкретном взаимодействии антитело-антиген. Используемый в настоящем документе термин «KD» означает константу диссоциации, которую определяют на основании отношения kd к ka (то есть, kd/ka) и выражают в виде молярной концентрации (M). Величины KD для антител можно определять методами, хорошо известными в данной области. Методы определения KD антитела включают измерение методом поверхностного плазмонного резонанса с использованием биосенсорной системы, такой как система BIACORE™, или измерение аффинности в растворе методом титрования равновесного раствора (ТРР).
Используемые в настоящем документе термины «моноклональное антитело» или «композиция моноклонального антитела» означают препарат молекул антитела одного молекулярного состава. Композиция моноклонального антитела проявляет одну специфичность связывания и аффинность для конкретного эпитопа.
Термин «нуклеиновая кислота» в настоящем документе используется взаимозаменяемо с термином «полинуклеотид» и означает дезоксирибонуклеотиды или рибонуклеотиды, а также их полимеры, в одно- или двухцепочечной форме. Термин охватывает нуклеиновые кислоты, содержащие известные аналоги нуклеотидов или модифицированные каркасные остатки или связи, являющиеся синтетическими, природными и неприродными, которые имеют связывающие свойства, аналогичные свойствам эталонной нуклеиновой кислоты, и которые метаболизируются аналогично эталонным нуклеотидам. Примеры таких аналогов включают, без ограничения, фосфоротиоаты, фосфорамидаты, метилфосфонаты, хирал-метилфосфонаты, 2-O-метилрибонуклеотиды, пептид-нуклеиновые кислоты (ПНК).
Если нет иных указаний, подразумевается, что конкретная нуклеотидная последовательность также включает ее консервативно модифицированные варианты (например, замены вырожденными кодонами) и комплементарные последовательности, а также специально указанную последовательность. В частности, как подробно описано ниже, замены вырожденными кодонами можно осуществлять, создавая последовательности, в которых в третьем положении одного или более выбранных (или всех) кодонов имеет место замена на смешанные основания и/или остатки дезоксиинозина (Batzer et al., Nucleic Acid Res. 19: 5081, 1991; Ohtsuka et al., J. Biol. Chem. 260: 2605-2608, 1985; и Rossolini et al., Mol. Cell. Probes 8: 91-98, 1994).
Термин «функционально связанные» означает наличие функциональной связи между двумя или более полинуклеотидными сегментами (например, ДНК). Как правило, термин означает функциональную связь транскрипционной регуляторной последовательности с транскрибируемой последовательностью. Например, последовательность промотора или энхансера функционально связана с кодирующей последовательностью, если она стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке-хозяине или другой системе экспрессии. Как правило, транскрипционные регуляторные последовательности промотора, которые функционально связаны с транскрибируемой последовательностью, физически примыкают к транскрибируемой последовательности, то есть, они действуют в цис-положении. Однако некоторые транскрипционные регуляторные последовательности, такие как энхансеры, не обязательно должны физически примыкать или находиться в непосредственной близости к кодирующим последовательностям, транскрипцию которых они усиливают.
Используемый в настоящем документе термин «оптимизированная» означает, что нуклеотидная последовательность была изменена для кодирования аминокислотной последовательности с использованием кодонов, которые являются предпочтительными в продуцирующей клетке или организме, как правило, эукариотической клетке, например, клетке пичиа, клетке яичника китайского хомяка (CHO) или клетке человека. Оптимизированная нуклеотидная последовательность сконструирована для полного или максимально возможного сохранения аминокислотной последовательности, кодируемой исходной нуклеотидной последовательностью, которую также называют «родительской» последовательностью. Описанные в настоящем документе оптимизированные последовательности были сконструированы для содержания кодонов, предпочтительных в клетках млекопитающих. Однако в настоящем документе также предусмотрена оптимизированная экспрессия этих последовательностей в других эукариотических клетках или прокариотических клетках. Аминокислотные последовательности, закодированные оптимизированными нуклеотидными последовательностями, также называют оптимизированными.
В настоящем документе термины «полипептид» и «белок» используются взаимозаменяемо и означают полимер из аминокислотных остатков. Термины применимы к аминокислотным полимерам, в которых один или более аминокислотных остатков представляют собой искусственные химические миметики соответствующих природных аминокислот, а также к полимерам природных аминокислот и полимерам неприродных аминокислот. Если нет иных указаний, подразумевается, что конкретная полипептидная последовательность также включает ее консервативно модифицированные варианты.
Используемый в настоящем документе термин «рекомбинантное человеческое антитело» включает все человеческие антитела, которые получены, экспрессированы, созданы или выделены рекомбинантными методами, например, антитела, полученные от животного (например, мыши), которое является трансгенным или трансхромосомным по генам человеческих иммуноглобулинов, или выделенные из гибридомы, полученной из его клеток; антитела, выделенные из клетки-хозяина, трансформированной для экспрессии человеческого антитела, например, из трансфектомы; антитела, выделенные из рекомбинантной, комбинаторной библиотеки человеческих антител, а также антитела, полученные, экспрессированные, созданные или выделенные любыми другими методами, которые включают сплайсинг всей или части последовательности гена человеческого иммуноглобулина с другими последовательностями ДНК. Такие рекомбинантные человеческие антитела имеют вариабельные области, в которых каркасные и CDR области получены из последовательностей человеческого иммуноглобулина зародышевой линии. Однако в конкретных вариантах осуществления такие рекомбинантные человеческие антитела могут быть подвергнуты in vitro мутагенезу (или, если используют животное, трансгенное по последовательностям человеческого Ig, in vivo соматическому мутагенезу) и, таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител представляют собой последовательности, которые, хотя и получены из, и являются родственными для, человеческих последовательностей VH и VL зародышевой линии, могут не присутствовать естественным образом в репертуаре человеческих антител зародышевой линии in vivo.
Термин «рекомбинантная клетка-хозяин» (или просто «клетка-хозяин») означает клетку, в которую был введен рекомбинантный экспрессионный вектор. Следует понимать, что такие термины должны означать не только конкретную клетку субъекта, но и потомство такой клетки. Поскольку некоторые модификации могут возникать в последующих поколениях вследствие либо мутации, либо влияния факторов внешней среды, такое потомство фактически может не быть идентичным родительской клетке, но оно все еще охвачено используемым в настоящем документе термином «клетка-хозяин».
Термин «субъект» включает людей и животных, отличных от людей. Отличные от людей животные включают всех позвоночных (например, млекопитающих и не млекопитающих), таких как приматы, кроме людей (например, яванские макаки), овцы, кролики, собаки, коровы, куры, амфибии и рептилии. Если нет иных указаний, термины «пациент» или «субъект» в настоящем документе используют взаимозаменяемо. Используемые в настоящем документе термины «яванский макак» или «макак-крабоед» относятся к яванскому макаку (Macaca fascicularis). В конкретных аспектах пациент или субъект является человеком.
Используемый в настоящем документе термин «терапия» или «лечение» какого-либо заболевания или нарушения (например, тромбоэмболического заболевания) в одном варианте осуществления означает ослабление заболевания или нарушения (то есть, замедление или остановку, или ослабление развития заболевания, или по меньшей мере одного из его клинических симптомов). В другом варианте осуществления «терапия» или «лечение» означает ослабление или уменьшение по меньшей мере одного физического параметра, включая те, которые могут быть незаметны для пациента. В другом варианте осуществления «терапия» или «лечение» означает модулирование заболевания или нарушения, либо физическое (например, стабилизацию заметного симптома), либо физиологическое (например, стабилизацию физического параметра), либо и то, и другое. В другом варианте осуществления «терапия» или «лечение» означает предотвращение, либо отсрочку начала или развития, или прогрессирования заболевания или нарушения.
«Предотвращение» применительно к состояниям, описанным в настоящем документе, включая, например, тромбоэмболическое заболевание, означает любое действие, которое предотвращает или замедляет ухудшение, например, параметров тромбоэмболического заболевания, как описано ниже, у пациента, имеющего риск такого ухудшения.
Термин «вектор» означает молекулу полинуклеотида, способную переносить другой полинуклеотид, с которым она связана. Одним из видов вектора является «плазмида», представляющая собой кольцевую двухцепочечную ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим видом вектора является вирусный вектор, такой как аденоассоциированный вирусный вектор (AAV или AAV2), при этом в вирусный геном могут быть лигированы дополнительные сегменты ДНК. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную точку начала репликации, и эписомные векторы млекопитающих). Другие векторы (например, не эписомные векторы млекопитающих) могут интегрироваться в геном клетки-хозяина при введении их в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны управлять экспрессией генов, с которыми они функционально связаны. В настоящем документе такие векторы называют «рекомбинантными экспрессионными векторами» (или просто «экспрессионными векторами»). Как правило, экспрессионные векторы, используемые в методах рекомбинантной ДНК, часто имеют форму плазмид. В настоящем описании термины «плазмида» и «вектор» могут использоваться взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора. Однако изобретение включает использование и других форм экспрессионных векторов, таких как вирусные векторы (например, репликативно-дефектные ретровирусы, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигуры 1A-C показывают эффект NOV1401 на индуцированный FeCl3 тромбоз у мышей FXI-/- с введенным человеческим белком FXI. NOV1401 ингибировало тромбоз зависимым от дозы образом. Антитело приводило к пролонгированию аЧТВ до такой же степени, как у не подвергнутых воздействию мышей FXI-/-.
Фигуры 2A-B показывают эффект нескольких внутривенных (в/в) (A; N=2) или подкожных (п/к) (B; N=2) доз 3 мг/кг, 10 мг/кг и 30 мг/кг NOV1401 на аЧТВ (ромбы) и связь с общими уровнями в плазме NOV1401 (квадраты) у яванских макак. Одна доза 3 мг/кг приводила к примерно 2-кратному увеличению аЧТВ, которое сохранялось в течение 5-6 недель. Все протестированные дозы вызывали пролонгирование аЧТВ в сходной степени, и более высокие протестированные дозы, очевидно, не приводили к большему пролонгированию аЧТВ, чем в случае дозы 3 мг/кг.
Фигуры 3A-B показывают эффект нескольких в/в (A; N=2) или п/к (B; N=2) доз 3 мг/кг, 10 мг/кг и 30 мг/кг NOV1401 на содержание в плазме свободного FXI (квадраты) и связь с аЧТВ (ромбы) у яванских макак. Одна доза 3 мг/кг приводила к уменьшению содержания свободного FXI примерно на 90% в течение 5-6 недель. Все протестированные дозы вызывали снижение содержания свободного FXI в сходной степени, и более высокие протестированные дозы, очевидно, не приводили к большему снижению содержания свободного FXI, чем в случае дозы 3 мг/кг.
Фигуры 4A-B показывают рентгеновскую структуру Fab-фрагмента антитела NOV1401 по изобретению, связанного с FXI. Фигура 4A показывает рентгеновскую структуру комплекса NOV1401 Fab-FXI КД. Каталитический домен FXI изображен в виде серой поверхности, Fab изображен в виде ленты светло-серого (легкая цепь) и темно-серого (тяжелая цепь) цвета. Фигура 4B показывает рентгеновскую структуру комплекса NOV1401 Fab-FXI КД с наложением на структуру зимогена FXI. Каталитический домен FXI изображен в виде ленты серого цвета. Вариабельные домены Fab изображены в виде ленты светло-серого (VL) и темно-серого (VH) цвета. Наложено изображение структуры зимогена, включая четыре яблоковидных домена в виде темно-серой ленты у основания структуры (PDB 2F83). Указан сайт активирующего расщепления (Ile370).
Фигуры 5A-B показывают структурные изменения FXIa при связывании Fab-фрагмента NOV1401. На Фигуре 5A приведено изображение активного центра FXIa до связывания Fab. FXIa изображен в виде ленты с прозрачной поверхностью. Секции структуры, меняющие конформацию при связывании с Fab, отмечены (петля 145, петля 188 и петля 220). Указаны подкарманы S1 и S1'. На Фигуре 5B показана неактивная конформация FXI в комплексе с Fab (Fab не показан).
Фигуры 6A-C показывают кривые зависимости ответа от концентрации соединения для анти-FXI/FXIa антитела. На Фигуре 6A показано ингибирование активности фактора XIa за счет NOV1401. Репрезентативная кривая зависимости ингибирования ферментативной активности полноразмерного человеческого FXIa от концентрации антитела NOV1401. В анализе измеряли расщепление флуоресцентно меченого пептида, как описано в примере 3. Для подгонки нелинейной кривой использовали модель логистической кривой [y=A2+(A1-A2)/(1+(x/IC50)^p), где y представляет собой % ингибирования при концентрации ингибитора x. A1 представляет собой наименьшее значение ингибирования и A2 максимальное значение ингибирования. Экспонента, p, представляет собой коэффициент Хилла для этого репрезентативного набора данных, на основании чего было получено значение IC50 160 пМ. На Фигуре 6B показана кривая зависимости аЧТВ от концентрации антитела. Репрезентативная кривая зависимости увеличения времени свертывания крови в анализе аЧТВ от концентрации антитела NOV1401 с использованием объединенной человеческой плазмы. В анализе измеряли время до свертывания крови после запуска внутреннего каскада свертывания крови в присутствии NOV1401 в разных концентрациях, как описано в примере 4. Черная линия представляет подгонку с использованием модели логистической кривой. Пунктирная линия соответствует исходному времени свертывания крови для объединенной человеческой плазмы в отсутствие NOV1401. Исходное время свертывания крови составляет 32,3 секунд, и показано серой пунктирной линией на графике. Линия из серых точек показывает концентрацию антитела, при которой время свертывания крови удваивается по сравнению с исходным, то есть, достигает значения 2x аЧТВ, эта концентрация составляет 14 нМ. На Фигуре 6C показана кривая зависимости от концентрации антитела в анализе АОТ. Показана репрезентативная кривая зависимости ингибирования образования тромбина в анализе АОТ от концентрации антитела NOV1401 с использованием объединенной человеческой плазмы. В анализе измеряли эффекты разных концентраций NOV1401 на FXI-зависимое образование тромбина через так называемую петлю положительной обратной связи тромбин→FXIa, которое может быть запущено очень низкими концентрациями тканевого фактора (TF), как описано в примере 4. Черная линия представляет собой подгонку с использованием четырехпараметрической модели кривой «доза-ответ». Линия из точек представляет остаточную концентрацию тромбина из-за образования тромбина, индуцированного небольшими количествами TF. Значение IC50 24 нМ и остаточная концентрация тромбина 159 нМ (линия из точек) рассчитаны на основании этой кривой «доза-ответ».
Фигуры 7A-B показывают эффект еженедельного п/к введения доз 10 мг/кг (N=3) и 100 мг/кг (N=5) NOV1401 в течение 13 недель (14 доз) или в/в введения дозы 50 мг/кг (N=3) в течение 4 недель (5 доз) на аЧТВ и активность FXI (FXI:C). Фигура 7A показывает эффект на аЧТВ, измеренный в дни исследования 2, 23 и 79. аЧТВ увеличивалось в 2,1-3 раза у всех животных, получающих NOV1401, и оставалось повышенным на протяжении фазы дозирования исследования. Зависимость от дозы отсутствовала, и не было отмечено зависимости от пола мышей. Фигура 7B показывает эффект на FXI:C, измеренный в дни исследования 2, 23 и 79 и выраженный в виде процента активности FXI в плазме. Величина FXI:C снижалась у всех животных, получающих NOV1401, до уровней 5-12% и оставалась на этих уровнях на протяжении фазы дозирования исследования. Зависимость от дозы отсутствовала, и не было отмечено зависимости от пола мышей.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение частично основано на открытии молекул антител, которые специфически связывают FXIa и ингибируют его биологическую активность. Изобретение относится как к IgG антителам полноразмерного формата, так и к их антигенсвязывающим фрагментам, таким как Fab-фрагменты (например, антител NOV1090 и NOV1401).
Соответственно, настоящее изобретение относится к антителам, которые специфически связывают FXI и/или FXIa (например, FXI и/или FXIa человека, кролика и яванского макака), фармацевтическим композициям, способам получения и способам применения таких антител и композиций.
Фактор XI
FXI играет важную роль как во внутреннем, так и во внешнем путях активации свертывания крови, а также в запуске фаз инициации и амплификации гемостаза плазмы. И фактор XIIa, и тромбин, могут активировать FXI, что приводит к постоянному образованию тромбина и ингибированию фибринолиза. FXI играет незначительную роль в нормальном гемостазе в условиях высокого содержания тканевого фактора «после повреждения сосудов», в то же время он, очевидно, играет ключевую роль в возникновении тромбоза. Тяжелый дефицит фактора XI связан с пониженной частотой возникновения ишемического инсульта и венозных тромбоэмболических событий (Salomon et al. 2008; Salomon, et al. (2011) Thromb Haemost.; 105: 269-73). Кровотечения у субъектов с тяжелым дефицитом фактора XI случаются редко, в основном носят умеренный характер, вызываются повреждениями и затрагивают преимущественно ткани с повышенной фибринолитической активностью, такие как слизистая оболочка ротовой полости, слизистая оболочка носовой полости и мочевыводящих путей (Salomon et al., 2011). Кровотечение в критических органах если и случается, то чрезвычайно редко.
Свертывание крови является последовательным процессом, во время которого факторы свертывания крови взаимодействуют и активируются, что в конечном итоге приводит к образованию фибрина и формированию сгустка. В классической каскадной модели свертывания крови процесс образования фибрина может быть инициирован двумя различными путями, то есть, внутренним и внешним путями, соответственно (Mackman, 2008).
Во внешнем пути повреждение сосудов дает возможность внесосудистому тканевому фактору (TF) взаимодействовать с фактором VII (FVII) и активировать его, что впоследствии приводит к активации фактора X и протромбина. Активный тромбин в конечном итоге превращает растворимый фибриноген в фибрин. Внешний путь является основным для гемостаза, препятствование действию факторов свертывания крови в этом пути связано с риском кровотечения.
Во внутреннем пути фактор XII может в некоторых случаях активироваться в процессе так называемой контактной активации. Появление активированного фактора XIIa приводит к последующей активации фактора XI и фактора IX. Поскольку фактор IXa активирует фактор X, внешний и внутренний пути на этой стадии сходятся (в общий путь). Активность тромбина усиливается за счет увеличения его образования через петлю положительной обратной связи, в которой тромбин активирует фактор XI независимо от фактора XII. Эта петля положительной обратной связи способствует постоянному росту тромбов, но лишь минимально участвует в гемостазе, поскольку сильная активация внесосудистым тканевым фактором является достаточной для образования сгустка. Таким образом, внутренний путь незначительно вовлечен в гемостаз (Gailani and Renné (2007) Arterioscler Thromb Vasc Biol. 2007, 27(12): 2507-13, Müller, Gailiani, and Renné 2011).
Доклинические исследования с использованием различных подходов к ингибированию FXI или FXIa у разных биологических видов внесли свой вклад в подтверждение важности этой терапевтической мишени. Мыши FXI-/- устойчивы к экспериментальному венозному (Wang, et al. (2006) J Thromb Haemost; 4: 1982-8) и артериальному (Wang, et al. (2005) J Thromb Haemost; 3: 695-702) тромбозу. Введение мышам антитела (Ат 14E11), которое блокирует активацию FXI за счет FXIIa, приводило к ингибированию экспериментального тромбоза (Cheng, et al. (2010) Blood, 116: 3981-9) и способствовало уменьшению размера зоны церебрального инфаркта в мышиной модели ишемического инсульта (Leung, et al. (2012) Transl Stroke Res 2012; 3: 381-9). У павианов, которым вводили анти-FXI Ат, блокирующее связывание и активацию FIX фактором FXIa, наблюдали уменьшение роста богатых тромбоцитами тромбов на покрытых коллагеном сосудистых имплантатах (Tucker, et al. (2009) Blood 2009; 113: 936-44), и аналогичные результаты были получены с антителом 14E11 в данной модели (Cheng, 2010). Ни в одном из этих исследований не наблюдали чрезмерного кровотечения.
Блокирование синтеза FXI с помощью антисмысловых олигонуклеотидов у мышей (Zhang, et al. (2010) Blood, 2010; 116: 4684-92), яванских макак (Younis, et al. (2012) Blood 2012; 119: 2401-8) и павианов (Crosby, et al. (2013) Arterioscler Thromb Vasc Biol 2013; 33: 1670-8) приводило к антитромботическим и антикоагулянтным эффектам без чрезмерного кровотечения. Кроме того, аналогичные эффекты были получены при блокировании FXIa низкомолекулярными ингибиторами в моделях венозного и артериального тромбоза на крысах (Schumacher, et al. (2007) Eur J Pharmacol 2007; 570: 167-74) и кроликах (Wong, et al. (2011) J Thromb Thrombolysis 2011; 32: 129-37).
Пациенты с тяжелым дефицитом FXI редко испытывают спонтанные кровотечения и у них случается лишь умеренное вызванное травмой кровотечение, за исключением тканей с высокой фибринoлитической активностью. Редкость случаев тяжелого дефицита FXI приводит к необходимости проведения популяционных исследований для выяснения тромботического профиля этих пациентов по сравнению с населением в целом. Примечательно, что в отчетах о таких исследованиях отмечается снижение частоты случаев ишемического инсульта (Salomon, 2008) и тромбоза глубоких вен (ТГВ) (Salomon, et al. (2011) Blood 2008; 111: 4113-17) у таких пациентов. Так, количество ишемических инсультов (N=1) у 115 пациентов с тяжелым дефицитом FXI было ниже (p <0,003), чем ожидаемая частота встречаемости (N=8,6) у населения Израиля в целом, в то же время частота ТГВ (N=0) была ниже (p <0,019) у пациентов с тяжелым дефицитом FXI, чем ожидаемая частота в контрольной популяции (N=4,7). И наоборот, люди с уровнями FXI выше 90-го процентиля имеют в два раза более высокий риск развития ТГВ (Meijers, et al. (2000) N Engl J Med. 2000; 342: 696-701).
Недавно к пациентам, подвергаемым операции по полной замене коленного сустава, процедуре, которая предрасполагает к ТГВ, применяли терапию антисмысловыми препаратами против FXI или стандартное лечение (эноксапарин). В группе пациентов, получавших антисмысловые препараты (300 мг), снижалась в 7 раз частота случаев венозного тромбоза и уменьшалось (незначительно) количество эпизодов кровотечения по сравнению с пациентами, получавшими стандартное лечение (Büller et al., (2014) N Engl J Med. 372(3): 232-40. doi: 10.1056/NEJMoa1405760. Epub 2014 Dec 7).
В совокупности, вышеописанные результаты убедительно свидетельствуют в пользу FXI как как важной мишени для антитромботической терапии.
Анти-FXIa антитела и антигенсвязывающие фрагменты
Настоящее изобретение относится к антителам, которые специфически связывают FXI и/или FXIa. В некоторых вариантах осуществления настоящее изобретение относится к антителам, которые специфически связывают FXI и/или FXIa человека, кролика и яванского макака. Антитела по изобретению включают, но не ограничиваются ими, человеческие моноклональные антитела и Fab-фрагменты, выделенные, как описано в разделе «Примеры».
Настоящее изобретение относится к антителам, которые специфически связывают белок FXI и/или FXIa (например, FXI и/или FXIa человека, кролика и яванского макака), при этом антитела содержат домен VH, имеющий аминокислотную последовательность SEQ ID NOs: 9 и 29. Настоящее изобретение также относится к антителам, которые специфически связывают белок FXI и/или FXIa, при этом антитела содержат VH CDR, имеющую аминокислотную последовательность любой из VH CDR, приведенных в таблице 1, ниже. В частности, изобретение относится к антителам, которые специфически связывают белок FXI и/или FXIa (например, FXI и/или FXIa человека, кролика и яванского макака), при этом антитела содержат (или, альтернативно, состоят из) одну, две, три или более VH CDR, имеющих аминокислотную последовательность любой из VH CDR, приведенных в таблице 1, ниже.
Настоящее изобретение относится к антителам, которые специфически связывают белок FXIa, при этом указанные антитела содержат домен VL, имеющий аминокислотную последовательность SEQ ID NOs: 19 или 39. Настоящее изобретение также относится к антителам, которые специфически связывают белок FXI и/или FXIa (например, FXI и/или FXIa человека, кролика и яванского макака), при этом антитела содержат VL CDR, имеющую аминокислотную последовательность любой из VL CDR, приведенных в таблице 1, ниже. В частности, изобретение относится к антителам, которые специфически связывают белок FXI и/или FXIa (например, FXI и/или FXIa человека, кролика и яванского макака), при этом антитела содержат (или, альтернативно, состоят из) одну, две, три или более VL CDR, имеющих аминокислотную последовательность любой из VL CDR, приведенных в таблице 1, ниже.
Другие антитела по изобретению содержат аминокислотные последовательности, которые были мутированы, но все-еще имеют по меньшей мере 60, 70, 80, 85, 90 или 95 процентов идентичности областей CDR с областями CDR, последовательности которых приведены в таблице 1. В некоторых вариантах осуществления предложены мутантные аминокислотные последовательности, в которых не более 1, 2, 3, 4 или 5 аминокислот были мутированы в областях CDR по сравнению с областями CDR, последовательности которых приведены в таблице 1.
Настоящее изобретение также относится к нуклеотидным последовательностям, кодирующим VH, VL, полноразмерную тяжелую цепь и полноразмерную легкую цепь антител, которые специфически связывают белок FXI и/или FXIa (например, FXI и/или FXIa человека, кролика и яванского макака). Такие нуклеотидные последовательности могут быть оптимизированы для экспрессии в клетках млекопитающих (например, в таблице 1 приведены оптимизированные нуклеотидные последовательности для тяжелой цепи и легкой цепи антител по изобретению).
Таблица 1. Примеры анти-FXIa антител, Fab-фрагментов и белков FXIa.
kginynssva ksaqecqerc tddvhchfft yatrqfpsle hrnicllkht qtgtptritk ldkvvsgfsl kscalsnlac irdifpntvf adsnidsvma pdafvsgric thhpgclfft
ffsqewpkes qrnlcllkts esglpstrik kskalsgfsl qscrhsipvf chssfyhdtd flgeeldiva aksheacqkl ctnavrcqff tytpaqascn egkgkcylkl ssngsptkil
hgrggisgyt lrlckmdnec ttkikprivg gtasvrgewp wqvtlhttsp tqrhlcggsi ignqwiltaa hcfygvespk ilrvysgiln qseikedtsf fgvqeiiihd qykmaesgyd
iallklettv nytdsqrpic lpskgdrnvi ytdcwvtgwg yrklrdkiqn tlqkakiplv tneecqkryr ghkithkmic agyreggkda ckgdsggpls ckhnevwhlv gitswgegca
qrerpgvytn vveyvdwile ktqav
tatttctgac atactaagat tttaacgact ttcacaaata tgctgtactg agagagaatg ttacataaca ttgagaacta gtacaagtaa atattaaagt gaagtgacca tttcctacac
aagctcattc agaggaggat gaagaccatt ttggaggaag aaaagcaccc ttattaagaa ttgcagcaag taagccaaca aggtcttttc aggatgattt tcttatatca agtggtacat
ttcattttat ttacttcagt ttctggtgaa tgtgtgactc agttgttgaa ggacacctgc tttgaaggag gggacattac tacggtcttc acaccaagcg ccaagtactg ccaggtagtc
tgcacttacc acccaagatg tttactcttc actttcacgg cggaatcacc atctgaggat cccacccgat ggtttacttg tgtcctgaaa gacagtgtta cagaaacact gccaagagtg
aataggacag cagcgatttc tgggtattct ttcaagcaat gctcacacca aataagcgct tgcaacaaag acatttatgt ggacctagac atgaagggca taaactataa cagctcagtt
gccaagagtg ctcaagaatg ccaagaaaga tgcacggatg acgtccactg ccactttttc acgtacgcca caaggcagtt tcccagcctg gagcatcgta acatttgtct actgaagcac
acccaaacag ggacaccaac cagaataacg aagctcgata aagtggtgtc tggattttca ctgaaatcct gtgcactttc taatctggct tgtattaggg acattttccc taatacggtg
tttgcagaca gcaacatcga cagtgtcatg gctcccgatg cttttgtctg tggccgaatc tgcactcatc atcccggttg cttgtttttt accttctttt cccaggaatg gcccaaagaa
tctcaaagaa atctttgtct ccttaaaaca tctgagagtg gattgcccag tacacgcatt aaaaagagca aagctctttc tggtttcagt ctacaaagct gcaggcacag catcccagtg
ttctgccatt cttcatttta ccatgacact gatttcttgg gagaagaact ggatattgtt gctgcaaaaa gtcacgaggc ctgccagaaa ctgtgcacca atgccgtccg ctgccagttt
tttacctata ccccagccca agcatcctgc aacgaaggga agggcaagtg ttacttaaag ctttcttcaa acggatctcc aactaaaata cttcacggga gaggaggcat ctctggatac
acattaaggt tgtgtaaaat ggataatgag tgtaccacca aaatcaagcc caggatcgtt ggaggaactg cgtctgttcg tggtgagtgg ccgtggcagg tgaccctgca cacaacctca
cccactcaga gacacctgtg tggaggctcc atcattggaa accagtggat attaacagcc gctcactgtt tctatggggt agagtcacct aagattttgc gtgtctacag tggcatttta
aatcaatctg aaataaaaga ggacacatct ttctttgggg ttcaagaaat aataatccat gatcagtata aaatggcaga aagcgggtat gatattgcct tgttgaaact ggaaaccaca
gtgaattaca cagattctca acgacccata tgcctgcctt ccaaaggaga tagaaatgta atatacactg attgctgggt gactggatgg gggtacagaa aactaagaga caaaatacaa
aatactctcc agaaagccaa gataccctta gtgaccaacg aagagtgcca gaagagatac agaggacata aaataaccca taagatgatc tgtgccggct acagggaagg agggaaggac
gcttgcaagg gagattcggg aggccctctg tcctgcaaac acaatgaggt ctggcatctg gtaggcatca cgagctgggg cgaaggctgt gctcaaaggg agcggccagg tgtttacacc
aacgtggtcg agtacgtgga ctggattctg gagaaaactc aagcagtgtg aatgggttcc caggggccat tggagtccct gaaggaccca ggatttgctg ggagagggtg ttgagttcac
tgtgccagca tgcttcctcc acagtaacac gctgaagggg cttggtgttt gtaagaaaat gctagaagaa aacaaactgt cacaagttgt tatgtccaaa actcccgttc tatgatcgtt
gtagtttgtt tgagcattca gtctctttgt ttttgatcac gcttctatgg agtccaagaa ttaccataag gcaatatttc tgaagattac tatataggca gatatagcag aaaataacca
agtagtggca gtggggatca ggcagaagaa ctggtaaaag aagccaccat aaatagattt gttcgatgaa agatgaaaac tggaagaaag gagaacaaag acagtcttca ccattttgca
ggaatctaca ctctgcctat gtgaacacat ttcttttgta aagaaagaaa ttgattgcat ttaatggcag attttcagaa tagtcaggaa ttcttgtcat ttccatttta aaatatatat
taaaaaaaat cagttcgagt agacacgagc taagagtgaa tgtgaagata acagaatttc tgtgtggaag aggattacaa gcagcaattt acctggaagt gataccttag gggcaatctt
gaagatacac tttcctgaaa aatgatttgt gatggattgt atatttattt aaaatatctt gggaggggag gctgatggag atagggagca tgctcaaacc tccctaagac aagctgctgc
tgtgactatg ggctcccaaa gagctagatc gtatatttat ttgacaaaaa tcaccataga ctgcatccat actacagaga aaaaacaatt agggcgcaaa tggatagtta cagtaaagtc
ttcagcaagc agctgcctgt attctaagca ctgggatttt ctgtttcgtg caaatattta tctcattatt gttgtgatct agttcaataa cctagaattt gaattgtcac cacatagctt
tcaatctgtg ccaacaacta tacaattcat caagtgtg
Другие антитела по изобретению включают те, в которых аминокислотные последовательности или нуклеотидные последовательности, кодирующие аминокислотные последовательности, были мутированы, но все-еще имеют по меньшей мере 60, 65, 70, 75, 80, 85, 90 или 95 процентов идентичности с последовательностями, приведенными в таблице 1. Некоторые варианты осуществления включают мутантные аминокислотные последовательности, в которых не более 1, 2, 3, 4 или 5 аминокислот были мутированы в вариабельных областях по сравнению последовательностями вариабельных областей, приведенными в таблице 1, но при этом они сохраняют по существу ту же самую антигенсвязывающую активность.
Поскольку каждое из этих антител может связывать FXI и/или FXIa, последовательности VH, VL, полноразмерной легкой цепи и полноразмерной тяжелой цепи (аминокислотные последовательности и нуклеотидные последовательности, кодирующие аминокислотные последовательности) можно «сочетать и комбинировать» для создания других связывающих FXI и/или FXIa антител по изобретению. Такие полученные путем «сочетания и комбинирования» связывающие FXI и/или FXIa антитела можно тестировать в анализах связывания, известных в данной области (например, ELISA и других анализах, описанных в разделе «Примеры»). При сочетании и комбинировании этих цепей последовательность VH из конкретной пары VH/VL должна быть заменена структурно сходной последовательностью VH. Аналогично, последовательность полноразмерной тяжелой цепи из конкретной пары полноразмерная тяжелая цепь/полноразмерная легкая цепь должна быть заменена структурно сходной последовательностью полноразмерной тяжелой цепи. Аналогично, последовательность VL из конкретной пары VH/VL должна быть заменена структурно сходной последовательностью VL. Аналогично, последовательность полноразмерной легкой цепи из конкретной пары полноразмерная тяжелая цепь/полноразмерная легкая цепь должна быть заменена структурно сходной последовательностью полноразмерной легкой цепи.
Соответственно, в одном аспекте изобретение относится к выделенному антителу, или его антигенсвязывающей области, имеющему: вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 9 и 29, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 19 и 39, при этом антитело специфически связывает FXI и/или FXIa (например, FXIa человека, кролика и яванского макака).
Более конкретно, в некоторых аспектах изобретение относится к выделенному антителу, или его антигенсвязывающей области, имеющему вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, содержащие аминокислотные последовательности, выбранные из SEQ ID NOs: 9 и 29; или 19 и 39, соответственно.
В конкретном варианте осуществления антитело, или его антигенсвязывающий фрагмент, предложенное в настоящем документе, которое специфически связывает FXI и/или FXIa человека, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 9, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 19.
В конкретном варианте осуществления антитело, или его антигенсвязывающий фрагмент, предложенное в настоящем документе, которое специфически связывает FXI и/или FXIa человека, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 29, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 39.
В другом аспекте изобретения предложено (i) выделенное антитело, имеющее: полноразмерную тяжелую цепь, содержащую аминокислотную последовательность, которая была оптимизирована для экспрессии в клетках млекопитающих, выбранную из группы, состоящей из SEQ ID NOs: 11 или 31, и полноразмерную легкую цепь, содержащую аминокислотную последовательность, которая была оптимизирована для экспрессии в клетках млекопитающих, выбранную из группы, состоящей из SEQ ID NOs: 21 или 41; или (ii) функциональный белок, содержащий его антигенсвязывающий фрагмент. Более конкретно, в некоторых аспектах изобретение относится к выделенному антителу, или его антигенсвязывающей области, имеющему тяжелую цепь и легкую цепь, содержащие аминокислотные последовательности, выбранные из SEQ ID NOs: 11 и 31; или 19 и 39, соответственно.
В конкретном варианте осуществления антитело, или его антигенсвязывающий фрагмент, предложенное в настоящем документе, которое специфически связывает FXI и/или FXIa человека, содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 21.
В конкретном варианте осуществления антитело, или его антигенсвязывающий фрагмент, предложенное в настоящем документе, которое специфически связывает FXI и/или FXIa человека, содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 31, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 41.
Используемые в настоящем документе термины «определяющая комплементарность область» и «CDR» означают аминокислотные последовательности в вариабельных областях антитела, которые придают антигенную специфичность и аффинность связывания. Как правило, существуют три области CDR в каждой вариабельной области тяжелой цепи (HCDR1, HCDR2, HCDR3) и три области CDR в каждой вариабельной области легкой цепи (LCDR1, LCDR2, LCDR3).
Точные границы аминокислотной последовательности конкретной CDR могут быть легко определены с использованием любой из хорошо известных систем, включая те, которые описаны в: Kabat et al. (1991), «Sequences of Proteins of Immunological Interest», 5-е издание, Public Health Service, National Institutes of Health, Bethesda, MD (система нумерации «Kabat»), Al-Lazikani et al., (1997) JMB 273, 927-948 (система нумерации «Chothia»), Lefranc et al., (2003) Dev. Comp. Immunol., 27, 55-77 (система нумерации «IMGT»), или «комбинированную» систему.
Например, в соответствии с системой Kabat аминокислотные остатки CDR антитела NOV1090 в вариабельном домене тяжелой цепи (VH) имеют порядковые номера 31-35 (HCDR1), 50-66 (HCDR2) и 99-111 (HCDR3); и аминокислотные остатки CDR в вариабельном домене легкой цепи (VL) имеют порядковые номера 22-35 (LCDR1), 51-57 (LCDR2) и 90-100 (LCDR3). В соответствии с системой Chothia аминокислоты CDR в VH имеют порядковые номера 26-32 (HCDR1), 52-57 (HCDR2) и 99-111 (HCDR3); и аминокислотные остатки в VL имеют порядковые номера 25-33 (LCDR1), 51-53 (LCDR2) и 92-99 (LCDR3). При комбинировании определений CDR по системе Kabat и системе Chothia области CDR состоят из аминокислотных остатков 26-35 (HCDR1), 50-66 (HCDR2) и 99-111 (HCDR3) в человеческой области VH и аминокислотных остатков 22-35 (LCDR1), 51-57 (LCDR2) и 90-100 (LCDR3) в человеческой области VL. При комбинировании определений CDR по системе Kabat и системе Chothia «комбинированные» области CDR состоят из аминокислотных остатков 26-35 (HCDR1), 50-66 (HCDR2) и 99-108 (HCDR3) в человеческой области VH и аминокислотных остатков 24-38 (LCDR1), 54-60 (LCDR2) и 93-101 (LCDR3) в человеческой области VL. В качестве другого примера, в соответствии с системой IMGT аминокислотные остатки CDR в вариабельном домене тяжелой цепи (VH) имеют порядковые номера 26-33 (HCDR1), 51-58 (HCDR2) и 97-108 (HCDR3); и аминокислотные остатки CDR в вариабельном домене легкой цепи (VL) имеют порядковые номера 27-36 (LCDR1), 54-56 (LCDR2) и 93-101 (LCDR3). В таблице 1 приведены иллюстративные последовательности по системам Kabat, Chothia, комбинированной и IMGT для HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 анти-FXI/FXIa антител, например, NOV1090 и NOV1401. В другом аспекте настоящее изобретение относится к связывающим FXIa антителам, содержащим области CDR1, CDR2 и CDR3 тяжелой цепи и легкой цепи, приведенные в таблице 1, или их сочетание. Аминокислотные последовательности VH CDR1 антител приведены в SEQ ID NOs: 3 и 23. Аминокислотные последовательности VH CDR2 антител приведены в SEQ ID NOs: 4 и 24. Аминокислотные последовательности VH CDR3 антител приведены в SEQ ID NOs: 5 и 25. Аминокислотные последовательности VL CDR1 антител приведены в SEQ ID NOs: 13 и 33. Аминокислотные последовательности VL CDR2 антител приведены в SEQ ID NOs: 14 и 34. Аминокислотные последовательности VL CDR3 антител приведены в SEQ ID NOs: 15 и 35. Эти области CDR определены в соответствии с системой Kabat.
Альтернативно, при определении в соответствии с системой Chothia (Al-Lazikani et al., (1997) JMB 273, 927-948) аминокислотные последовательности VH CDR1 антител приведены в SEQ ID NOs: 6 и 26. Аминокислотные последовательности VH CDR2 антител приведены в SEQ ID NOs: 7 и 27. Аминокислотные последовательности VH CDR3 антител приведены в SEQ ID NOs: 8 и 28. Аминокислотные последовательности VL CDR1 антител приведены в SEQ ID NOs: 16 и 36. Аминокислотные последовательности VL CDR2 антител приведены в SEQ ID NOs: 17 и 37. Аминокислотные последовательности VL CDR3 антител приведены в SEQ ID NOs: 18 и 38.
Альтернативно, при определении в соответствии с комбинированной системой аминокислотные последовательности VH CDR1 антител приведены в SEQ ID NO: 46. Аминокислотные последовательности VH CDR2 антител приведены в SEQ ID NO: 4. Аминокислотные последовательности VH CDR3 антител приведены в SEQ ID NO: 5. Аминокислотные последовательности VL CDR1 антител приведены в SEQ ID NO: 33. Аминокислотные последовательности VL CDR2 антител приведены в SEQ ID NO: 14. Аминокислотные последовательности VL CDR3 антител приведены в SEQ ID NO: 15.
Альтернативно, при определении в соответствии с системой нумерации IMGT аминокислотные последовательности VH CDR1 антител приведены в SEQ ID NO: 43. Аминокислотные последовательности VH CDR2 антител приведены в SEQ ID NO: 44. Аминокислотные последовательности VH CDR3 антител приведены в SEQ ID NO: 45. Аминокислотные последовательности VL CDR1 антител приведены в SEQ ID NO: 47. Аминокислотные последовательности VL CDR2 антител приведены в SEQ ID NO: 37. Аминокислотные последовательности VL CDR3 антител приведены в SEQ ID NO: 15.
Поскольку каждое из этих антител может связывать FXI и/или FXIa и антигенсвязывающая специфичность определяется в основном областями CDR1, 2 и 3, последовательности VH CDR1, 2 и 3 и последовательности VL CDR1, 2 и 3 можно «сочетать и комбинировать» (то есть, CDR из разных антител можно сочетать и комбинировать, хотя каждое антитело предпочтительно содержит VH CDR1, 2 и 3 и VL CDR1, 2 и 3, для создания других связывающих FXI и/или FXIa молекул по изобретению). Такие полученные путем «сочетания и комбинирования» связывающие FXI и/или FXIa антитела можно тестировать в анализах связывания, которые известны в данной области и которые описаны в разделе «Примеры» (например, анализах ELISA, SET, BIACORE™). При сочетании и комбинировании последовательностей VH CDR последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VH должна быть заменена структурно сходной последовательностью(ми) CDR. Аналогично, при сочетании и комбинировании последовательностей VL CDR последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VL должна быть заменена структурно сходной последовательностью(ми) CDR. Специалистам в данной области понятно, что новые последовательности VH и VL можно создавать путем замены одной или более последовательностей областей CDR VH и/или VL структурно сходными последовательностями из последовательностей CDR, приведенных в настоящем документе для моноклональных антител по настоящему изобретению. В одном варианте осуществления помимо вышеуказанных, антигенсвязывающие фрагменты антител, описанных в настоящем документе, могут содержать VH CDR1, 2 и 3 или VL CDR 1, 2 и 3, при этом фрагмент связывает FXI и/или FXIa, как один вариабельный домен.
В конкретных вариантах осуществления изобретения антитела, или их антигенсвязывающие фрагменты, могут иметь последовательности тяжелой и легкой цепей из Fab, приведенные в таблице 1. Более конкретно, антитело, или его антигенсвязывающие фрагменты, может иметь последовательность тяжелой и легкой цепей из NOV1090 и NOV1401.
В других вариантах осуществления изобретения антитело, или антигенсвязывающий фрагмент, которое специфически связывает FXI и/или FXIa, содержит CDR1 вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи, CDR3 вариабельной области тяжелой цепи, CDR1 вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи и CDR3 вариабельной области легкой цепи, определенные в соответствии с системой Kabat и приведенные в таблице 1. В других вариантах осуществления изобретения антитело, или антигенсвязывающий фрагмент, которое специфически связывает FXI и/или FXIa, содержит CDR1 вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи, CDR3 вариабельной области тяжелой цепи, CDR1 вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи и CDR3 вариабельной области легкой цепи, определенные в соответствии с системой Chothia и приведенные в таблице 1. В других вариантах осуществления антитело, или антигенсвязывающий фрагмент, которое специфически связывает FXI и/или FXIa, содержит CDR1 вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи, CDR3 вариабельной области тяжелой цепи, CDR1 вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи и CDR3 вариабельной области легкой цепи, определенные в соответствии с комбинированной системой и приведенные в таблице 1. В других вариантах осуществления изобретения антитело, или антигенсвязывающий фрагмент, которое специфически связывает FXI и/или FXIa, содержит CDR1 вариабельной области тяжелой цепи, CDR2 вариабельной области тяжелой цепи, CDR3 вариабельной области тяжелой цепи, CDR1 вариабельной области легкой цепи, CDR2 вариабельной области легкой цепи и CDR3 вариабельной области легкой цепи, определенные в соответствии с системой IMGT и приведенные в таблице 1.
В конкретном варианте осуществления изобретение относится к антителу, специфически связывающему FXI и/или FXIa, которое содержит CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 3; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 4; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 5; CDR1 вариабельной области легкой цепи с SEQ ID NO: 13; CDR2 вариабельной области легкой цепи с SEQ ID NO: 14 и CDR3 вариабельной области легкой цепи с SEQ ID NO: 15.
В конкретном варианте осуществления изобретение относится к антителу, специфически связывающему FXI и/или FXIa, которое содержит CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 23; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 24; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 25; CDR1 вариабельной области легкой цепи с SEQ ID NO: 33; CDR2 вариабельной области легкой цепи с SEQ ID NO: 34 и CDR3 вариабельной области легкой цепи с SEQ ID NO: 35.
В конкретном варианте осуществления изобретение относится к антителу, специфически связывающему FXI и/или FXIa, которое содержит CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 6; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 7; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 8; CDR1 вариабельной области легкой цепи с SEQ ID NO: 16; CDR2 вариабельной области легкой цепи с SEQ ID NO: 17 и CDR3 вариабельной области легкой цепи с SEQ ID NO: 18.
В конкретном варианте осуществления изобретение относится к антителу, специфически связывающему FXI и/или FXIa, которое содержит CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 26; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 27; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 28; CDR1 вариабельной области легкой цепи с SEQ ID NO: 36; CDR2 вариабельной области легкой цепи с SEQ ID NO: 37 и CDR3 вариабельной области легкой цепи с SEQ ID NO: 38.
В конкретном варианте осуществления изобретение относится к антителу, специфически связывающему FXI и/или FXIa, которое содержит CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 43; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 44; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 45; CDR1 вариабельной области легкой цепи с SEQ ID NO: 47; CDR2 вариабельной области легкой цепи с SEQ ID NO: 37 и CDR3 вариабельной области легкой цепи с SEQ ID NO: 15.
В конкретном варианте осуществления изобретение относится к антителу, специфически связывающему FXI и/или FXIa, которое содержит CDR1 вариабельной области тяжелой цепи с SEQ ID NO: 46; CDR2 вариабельной области тяжелой цепи с SEQ ID NO: 4; CDR3 вариабельной области тяжелой цепи с SEQ ID NO: 5; CDR1 вариабельной области легкой цепи с SEQ ID NO: 33; CDR2 вариабельной области легкой цепи с SEQ ID NO: 14 и CDR3 вариабельной области легкой цепи с SEQ ID NO: 15.
В конкретных вариантах осуществления изобретение относится к антителам, или антигенсвязывающим фрагментам, специфически связывающим FXI и/или FXIa, которые приведены в таблице 1. В предпочтительном варианте осуществления антитело, или антигенсвязывающий фрагмент, которое связывает FXI и/или FXIa, представляет собой NOV1090 и NOV1401.
Как описано в настоящем документе, человеческое антитело содержит вариабельные области тяжелой или легкой цепи, или полноразмерные тяжелые или легкие цепи, которые являются «продуктом» или «происходят из» конкретной последовательности зародышевой линии, если вариабельные области или полноразмерные цепи антитела получены из системы, в которой используются человеческие гены иммуноглобулинов зародышевой линии. Такие системы включают иммунизацию трансгенной мыши, несущей человеческие гены иммуноглобулинов, интересующим антигеном или скрининг библиотеки человеческих генов иммуноглобулинов, экспонированной на фаге, с помощью интересующего антигена. Человеческое антитело, которое является «продуктом» или «происходит из» человеческой последовательности иммуноглобулина зародышевой линии, можно идентифицировать, как таковое, путем сравнения аминокислотной последовательности человеческого антитела с аминокислотными последовательностями человеческих иммуноглобулинов зародышевой линии и выбора человеческой последовательности иммуноглобулина зародышевой линии, которая наиболее близка (то есть, имеет наибольший % идентичности) с последовательностью человеческого антитела.
Человеческое антитело, которое является «продуктом» или «происходит из» конкретной человеческой последовательности иммуноглобулина зародышевой линии, может иметь аминокислотные отличия по сравнению с последовательностью зародышевой линии, вследствие, например, естественных соматических мутаций или намеренного внесения сайт-специфических мутаций. Однако в каркасных областях VH или VL выбранное человеческое антитело, как правило, по меньшей мере на 90% идентично по аминокислотной последовательности с аминокислотной последовательностью, кодируемой человеческим геном иммуноглобулина зародышевой линии, и содержит аминокислотные остатки, которые определяют человеческое антитело именно как человеческое, при сравнении с аминокислотными последовательностями иммуноглобулинов зародышевой линии из других биологических видов (например, мышиными последовательностями иммуноглобулинов зародышевой линии). В некоторых случаях человеческое антитело может быть по меньшей мере на 60%, 70%, 80%, 90%, или по меньшей мере на 95%, или даже по меньшей мере на 96%, 97%, 98% или 99% идентичным по аминокислотной последовательности с аминокислотной последовательностью, кодируемой геном иммуноглобулина зародышевой линии.
Как правило, рекомбинантное человеческое антитело будет иметь не более 10 аминокислотных отличий от аминокислотной последовательности, кодируемой человеческим геном иммуноглобулина зародышевой линии, в каркасных областях VH или VL. В некоторых случаях человеческое антитело может иметь не более 5, или даже не более 4, 3, 2 или 1 аминокислотного отличия от аминокислотной последовательности, кодируемой геном иммуноглобулина зародышевой линии. Примеры человеческих генов иммуноглобулинов зародышевой линии включают, но не ограничиваются ими, фрагменты вариабельного домена зародышевой линии, описанные ниже, а также DP47 и DPK9.
Гомологичные антитела
В другом варианте осуществления настоящее изобретение относится к антителу, или его антигенсвязывающему фрагменту, содержащему аминокислотные последовательности, которые гомологичны последовательностям, приведенным в таблице 1 (например, SEQ ID NOs: 29, 31, 39 или 41), при этом антитело связывает белок FXI и/или FXIa (например, FXIa человека, кролика и яванского макака) и сохраняет желательные функциональные свойства антител, приведенных в таблице 1, таких как NOV1090 и NOV1401. В конкретных аспектах такие гомологичные антитела сохраняют аминокислотные последовательности CDR, приведенные в таблице 1 (например, CDR по системе Kabat, CDR по системе Chothia, CDR по системе IMGT или комбинированные CDR).
Например, изобретение относится к выделенному антителу, или его функциональному антигенсвязывающему фрагменту, содержащему вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, при этом вариабельный домен тяжелой цепи содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 9 и 29; вариабельный домен легкой цепи содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 19 и 39; и антитело специфически связывает FXI и/или FXIa (например, FXIa человека, кролика и яванского макака). В одном варианте осуществления выделенное антитело, или его функциональный антигенсвязывающий фрагмент, содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, при этом вариабельный домен тяжелой цепи содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности SEQ ID NO: 9; вариабельный домен легкой цепи содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности SEQ ID NO: 19; и антитело специфически связывает FXI и/или FXIa (например, FXIa человека, кролика и яванского макака). В одном варианте осуществления выделенное антитело, или его функциональный антигенсвязывающий фрагмент, содержит вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, при этом вариабельный домен тяжелой цепи содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности SEQ ID NO: 29; вариабельный домен легкой цепи содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности SEQ ID NO: 39; и антитело специфически связывает FXI и/или FXIa (например, FXIa человека, кролика и яванского макака). В некоторых аспектах изобретения последовательности тяжелой и легкой цепи дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные в соответствии с системой Kabat, например, SEQ ID NOs: 3, 4, 5, 13, 14 и 15, соответственно. В некоторых других аспектах изобретения последовательности тяжелой и легкой цепи дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные в соответствии с системой Chothia, например, SEQ ID NOs: 6, 7, 8, 16, 17 и 18, соответственно. В некоторых других аспектах последовательности тяжелой и легкой цепи дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные в соответствии с комбинированной системой, например, SEQ ID NOs: 46, 4, 5, 33, 14 и 15, соответственно. В некоторых других аспектах последовательности тяжелой и легкой цепи дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные в соответствии с системой IMGT, например, SEQ ID NOs: 43, 44, 45, 47, 37 и 15, соответственно.
В других вариантах осуществления аминокислотные последовательности VH и/или VL могут быть на 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, приведенным в таблице 1. В других вариантах осуществления аминокислотные последовательности VH и/или VL могут быть идентичны за исключением аминокислотной замены не более чем в 1, 2, 3, 4 или 5 аминокислотных положениях. Антитело, имеющее области VH и VL с высокой степенью (то есть, 80% или более) идентичности с областями VH и VL антител, приведенных в таблице 1, можно получать путем мутагенеза (например, сайт-направленного или ПЦР-опосредованного мутагенеза) кодирующих молекул нуклеиновой кислоты, SEQ ID NOs: 10 или 30 и SEQ ID NOs: 20 и 40, соответственно, с последующим тестированием закодированного измененного антитела на сохранение функции с использованием функциональных анализов, описанных в настоящем документе.
В других вариантах осуществления аминокислотные последовательности полноразмерной тяжелой цепи и/или полноразмерной легкой цепи могут быть на 50% 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, приведенным в таблице 1 (например, SEQ ID NOs: 11 и/или 21, или 31 и/или 41). Антитело, имеющее полноразмерную тяжелую цепь и полноразмерную легкую цепь с высокой степенью (то есть, 80% или более) идентичности с полноразмерными тяжелыми цепями, имеющими любую из SEQ ID NOs: 11 или 31, и полноразмерными легкими цепями, имеющими любую из SEQ ID NOs: 21 или 41, можно получать путем мутагенеза (например, сайт-направленного или ПЦР-опосредованного мутагенеза) молекул нуклеиновой кислоты, кодирующих такие полипептиды, с последующим тестированием закодированного измененного антитела на сохранение функции с использованием функциональных анализов, описанных в настоящем документе.
В одном аспекте настоящего изобретения предложено выделенное антитело, или его функциональный антигенсвязывающий фрагмент, содержащее тяжелую цепь и легкую цепь, при этом тяжелая цепь содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 11 и 31; легкая цепь содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 21 и 41, и антитело специфически связывает FXI и/или FXIa (например, FXIa человека, кролика и яванского макака). В одном варианте осуществления выделенное антитело, или его функциональный антигенсвязывающий фрагмент, содержит тяжелую цепь и легкую цепь, при этом тяжелая цепь содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности SEQ ID NO: 11; легкая цепь содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности SEQ ID NO: 21, и антитело специфически связывает FXI и/или FXIa (например, FXIa человека, кролика и яванского макака). В одном варианте осуществления выделенное антитело, или его функциональный антигенсвязывающий фрагмент, содержит тяжелую цепь и легкую цепь, при этом тяжелая цепь содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности SEQ ID NO: 31; легкая цепь содержит аминокислотную последовательность, которая на по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентична аминокислотной последовательности SEQ ID NO: 41, и антитело специфически связывает FXI и/или FXIa (например, FXIa человека, кролика и яванского макака). В некоторых аспектах изобретения последовательности тяжелой и легкой цепи дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные в соответствии с системой Kabat, например, SEQ ID NOs: 3, 4, 5, 13, 14 и 15, соответственно. В некоторых других аспектах изобретения последовательности тяжелой и легкой цепи дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные в соответствии с системой Chothia, например, SEQ ID NOs: 6, 7, 8, 16, 17 и 18, соответственно. В некоторых других аспектах последовательности тяжелой и легкой цепи дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные в соответствии с комбинированной системой, например, SEQ ID NOs: 46, 4, 5, 33, 14 и 15, соответственно. В некоторых других аспектах последовательности тяжелой и легкой цепи дополнительно содержат последовательности HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, определенные в соответствии с системой IMGT, например, SEQ ID NOs: 43, 44, 45, 47, 37 и 15, соответственно.
В других вариантах осуществления нуклеотидные последовательности полноразмерной тяжелой цепи и/или полноразмерной легкой цепи могут быть на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, приведенным в таблице 1 (например, SEQ ID NOs: 12 и/или 22, или 32 и/или 42).
В других вариантах осуществления нуклеотидные последовательности вариабельных областей тяжелой цепи и/или вариабельных областей легкой цепи могут быть на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, приведенным в таблице 1 (например, SEQ ID NOs: 10 и/или 20, или 30 и/или 40).
При использовании в настоящем документе, процент идентичности между двумя последовательностями зависит от числа идентичных положений в обеих последовательностях (то есть, % идентичности равен числу идентичных положений/общее число положений x 100), с учетом количества делеций и длины каждой делеции, которые необходимо вносить для оптимального выравнивания двух последовательностей. Сравнение последовательностей и определение процента идентичности между двумя последовательностями можно осуществлять с использованием математического алгоритма, как описано в неограничивающих примерах, ниже.
Дополнительно или альтернативно, белковые последовательности по настоящему изобретению также можно использовать в качестве «искомой последовательности» для проведения поиска в общедоступных базах данных, например, для идентификации родственных последовательностей. Например, такой поиск можно проводить с использованием программы BLAST (версия 2.0), описанной в Altschul, et al., 1990 J. Mol. Biol. 215: 403-10.
Антитела с консервативными модификациями
В конкретных вариантах осуществления антитело по изобретению содержит вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, при этом одна или более из этих последовательностей CDR имеют конкретные аминокислотные последовательности антител, описанных в настоящем документе, или их консервативные модификации, и при этом антитела сохраняют желательные функциональные свойства FXIa-связывающих антител по изобретению.
Соответственно, изобретение относится к выделенному антителу, или его антигенсвязывающему фрагменту, состоящему из вариабельной области тяжелой цепи, содержащей последовательности CDR1, CDR2 и CDR3, и вариабельной области легкой цепи, содержащей последовательности CDR1, CDR2 и CDR3, при этом: аминокислотные последовательности CDR1 вариабельной области тяжелой цепи выбирают из группы, состоящей из SEQ ID NOs: 3 и 23, а также их консервативных модификаций; аминокислотные последовательности CDR2 вариабельной области тяжелой цепи выбирают из группы, состоящей из SEQ ID NOs: 4 и 24, а также их консервативных модификаций; аминокислотные последовательности CDR3 вариабельной области тяжелой цепи выбирают из группы, состоящей из SEQ ID NOs: 5 и 25, а также их консервативных модификаций; аминокислотные последовательности CDR1 вариабельной области легкой цепи выбирают из группы, состоящей из SEQ ID NOs: 13 и 33, а также их консервативных модификаций; аминокислотные последовательности CDR2 вариабельной области легкой цепи выбирают из группы, состоящей из SEQ ID NOs: 14 и 34, а также их консервативных модификаций; аминокислотные последовательности CDR3 вариабельной области легкой цепи выбирают из группы, состоящей из SEQ ID NOs: 15 и 35, а также их консервативных модификаций; при этом антитело, или его антигенсвязывающие фрагменты, специфически связывает FXIa.
В одном аспекте настоящее изобретение относится к выделенному антителу, или его антигенсвязывающему фрагменту, состоящему из вариабельной области тяжелой цепи, содержащей последовательности CDR1, CDR2 и CDR3, и вариабельной области легкой цепи, содержащей последовательности CDR1, CDR2 и CDR3, при этом: аминокислотные последовательности CDR1 вариабельной области тяжелой цепи выбирают из группы, состоящей из последовательностей, приведенных в таблице 1, а также их консервативных модификаций; аминокислотные последовательности CDR2 вариабельной области тяжелой цепи выбирают из группы, состоящей из последовательностей, приведенных в таблице 1, а также их консервативных модификаций; аминокислотные последовательности CDR3 вариабельной области тяжелой цепи выбирают из группы, состоящей из последовательностей, приведенных в таблице 1, а также их консервативных модификаций; аминокислотные последовательности CDR1 вариабельной области легкой цепи выбирают из группы, состоящей из последовательностей, приведенных в таблице 1, а также их консервативных модификаций; аминокислотные последовательности CDR2 вариабельной области легкой цепи выбирают из группы, состоящей из последовательностей, приведенных в таблице 1, а также их консервативных модификаций; аминокислотные последовательности CDR3 вариабельной области легкой цепи выбирают из группы, состоящей из последовательностей, приведенных в таблице 1, а также их консервативных модификаций; при этом антитело, или его антигенсвязывающие фрагменты, специфически связывает FXIa.
В других вариантах осуществления антитело по изобретению оптимизировано для экспрессии в клетке млекопитающего и имеет последовательность полноразмерной тяжелой цепи и последовательность полноразмерной легкой цепи, при этом одна или более из этих последовательностей содержат конкретные аминокислотные последовательности антител, описанных в настоящем документе, или их консервативные модификации, и при этом антитела сохраняют желательные функциональные свойства FXIa-связывающих антител по изобретению. Соответственно, изобретение относится к выделенному антителу, оптимизированному для экспрессии в клетке млекопитающего, состоящему из полноразмерной тяжелой цепи и полноразмерной легкой цепи, при этом полноразмерная тяжелая цепь содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NOs: 11 или 31, а также их консервативных модификаций; и полноразмерная легкая цепь содержит аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NOs: 21 или 41, а также их консервативных модификаций; и антитело специфически связывает FXI и/или FXIa (например, FXIa человека, кролика и яванского макака).
Антитела, которые связывают тот же эпитоп
Настоящее изобретение относится к антителам, которые связывают тот же эпитоп, что и связывающие FXI и/или FXIa антитела, приведенные в таблице 1. Таким образом, дополнительные антитела можно идентифицировать на основании их способности конкурировать (например, конкурентно ингибировать связывание, в статистически значимой степени, за счет связывания с тем же или перекрывающимся эпитопом) с другими антителами по изобретению в анализах связывания FXI и/или FXIa (таких как те, которые описаны в разделе «Примеры»). Способность тестируемого антитела ингибировать связывание антител по настоящему изобретению с белком FXI и/или FXIa демонстрирует, что тестируемое антитело может конкурировать с данным антителом за связывание с FXI и/или FXIa; такое антитело может, в соответствии с неограничивающей теорией, связывать тот же или родственный (например, структурно сходный или пространственно проксимальный) эпитоп на белке FXI и/или FXIa, что и антитело, с которым оно конкурирует. В конкретном варианте осуществления антитело, которое связывает тот же эпитоп на FXI и/или FXIa, что и антитела по настоящему изобретению, представляет собой человеческое моноклональное антитело. Такие моноклональные антитела могут быть получены и выделены, как описано в настоящем документе.
При описании в настоящем документе, антитело «конкурирует» за связывание, когда конкурирующее антитело связывает тот же эпитоп FXI и/или FXIa, что и антитело, или антигенсвязывающий фрагмент, по изобретению (например, NOV1401 или NOV1090) и ингибирует связывание FXI и/или FXIa антителом, или антигенсвязывающим фрагментом, по изобретению более чем на 50% (например, 80%, 85%, 90%, 95%, 98% или 99%) в присутствии эквимолярной концентрации конкурирующего антитела. Это можно определять, например, в конкурентном анализе связывания любыми методами, хорошо известными специалистам в данной области.
При описании в настоящем документе, антитело, или его антигенсвязывающий фрагмент, не «конкурирует» с анти-FXI и/или анти-FXIa антителом, или антигенсвязывающим фрагментом, по изобретению (например, NOV1401 или NOV1090), если только указанное конкурирующее антитело, или его антигенсвязывающий фрагмент, не связывает тот же эпитоп FXI и/или FXIa, или перекрывающийся эпитоп FXI и/или FXIa, что и антитело, или антигенсвязывающий фрагмент, по изобретению. При описании в настоящем документе, конкурирующее антитело, или его антигенсвязывающий фрагмент, не включает антитело, которое (i) стерически блокирует антитело, или антигенсвязывающий фрагмент, по изобретению от связывания с его мишенью (например, если указанное конкурирующее антитело связывает расположенный вблизи, не перекрывающийся эпитоп FXI и/или FXIa и физически препятствует связыванию антитела, или антигенсвязывающего фрагмента, по изобретению с его мишенью); и/или (ii) связывает другой, не перекрывающийся эпитоп FXI и/или FXIa и индуцирует конформационное изменение белка FXI и/или FXIa таким образом, что указанный белок больше не может быть связан анти-FXI и/или FXIa антителом, или антигенсвязывающим фрагментом, по изобретению так, как это происходило бы в отсутствие указанного конформационного изменения.
Сконструированные и модифицированные антитела
Антитело по изобретению также можно получать с использованием антитела, содержащего одну или более из последовательностей VH и/или VL, приведенных в настоящем документе, в качестве исходного материала для конструирования модифицированного антитела, при этом модифицированное антитело может иметь измененные свойства по сравнению с исходным антителом. Антитело может быть сконструировано путем модификации одного или более остатков в одной или обеих вариабельных областях (то есть, VH и/или VL), например, в одной или более областях CDR и/или в одной или более каркасных областях. Дополнительно или альтернативно, антитело может быть сконструировано путем модификации остатков в константной области(ях), например, для изменения эффекторной функции(ий) антитела.
Одним из видов конструирования вариабельной области, которое можно выполнять, является прививание CDR. Антитела взаимодействуют с целевыми антигенами преимущественно через аминокислотные остатки, которые расположены в шести определяющих комплементарность областях (CDR) тяжелой и легкой цепи. Вследствие этого, аминокислотные последовательности в областях CDR в большей степени различаются у отдельных антител, чем последовательности за пределами областей CDR. Поскольку последовательности CDR отвечают за большинство взаимодействий антитело-антиген, можно осуществлять экспрессию рекомбинантных антител, которые имитируют свойства конкретных природных антител, путем конструирования экспрессионных векторов, содержащих последовательности CDR из конкретного природного антитела, привитые на каркасные последовательности из другого антитела с иными свойствами (смотри, например, Riechmann, L. et al., 1998 Nature 332: 323-327; Jones, P. et al., 1986 Nature 321: 522-525; Queen, C. et al., 1989 Proc. Natl. Acad., U.S.A. 86: 10029-10033; патент США № 5225539 автора Winter, и патенты США № 5530101; 5585089; 5693762 и 6180370 авторов Queen et al.)
Соответственно, другой вариант осуществления изобретения относится к выделенному антителу, или его антигенсвязывающему фрагменту, содержащему вариабельную область тяжелой цепи, содержащую последовательности CDR1, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 3 и 23; последовательности CDR2, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 4 и 24; последовательности CDR3, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 5 и 25, соответственно; и вариабельную область легкой цепи, содержащую последовательности CDR1, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 13 и 33; последовательности CDR2, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 14 и 34; и последовательности CDR3, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 15 и 35, соответственно. Таким образом, такие антитела содержат последовательности CDR из VH и VL моноклональных антител, хотя могут содержать каркасные последовательности, отличные от последовательностей из этих антител.
Такие каркасные последовательности можно получать из общедоступных баз данных ДНК или опубликованных источников, которые содержат последовательности генов антител зародышевой линии. Например, последовательности ДНК зародышевой линии для человеческих генов вариабельных областей тяжелой и легкой цепей можно найти в базе данных «VBase» последовательностей человеческих генов зародышевой линии (доступной в сети интернет по сетевому адресу mrc-cpe.cam.ac.uk/vbase), а также в публикациях Kabat, E. A., et al., 1991 Sequences of Proteins of Immunological Interest, пятое издание, U.S. Department of Health and Human Services, NIH Publication No. 91-3242; Tomlinson, I. M., et al., 1992 J. Mol. Biol. 227: 776-798; и Cox, J. P. L. et al., 1994 Eur. J Immunol. 24: 827-836; содержание каждой из которых включено в настоящий документ посредством ссылки в полном объеме.
Примером каркасных последовательностей для использования в антителах по изобретению являются те, которые имеют структурное сходство с каркасными последовательностями, используемыми в выбранных антителах по изобретению, например, консенсусными последовательностями и/или каркасными последовательностями, используемыми в моноклональных антителах по изобретению. Последовательности VH CDR1, 2 и 3, и VL CDR1, 2 и 3 могут быть привиты на каркасные области, имеющие последовательность, идентичную последовательности гена иммуноглобулина зародышевой линии, из которой происходит каркасная последовательность, или последовательности CDR могут быть привиты на каркасные области, имеющие одну или более мутаций по сравнению с последовательностями зародышевой линии. Например, было установлено, что в некоторых случаях полезно осуществлять мутации остатков в каркасных областях для сохранения или усиления антигенсвязывающей способности антитела (смотри, например, патенты США №№ 5530101; 5585089; 5693762 и 6180370 авторов Queen et al.). Каркасы, которые могут быть использованы в качестве остовов, на которых можно создавать антитела и антигенсвязывающие фрагменты, описанные в настоящем документе, включают, но не ограничиваются ими, VH1A, VH1B, VH3, Vk1, Vl2 и Vk2. Дополнительные каркасы известны в данной области и могут быть найдены, например, в базе данных vBase в сети интернет по сетевому адресу vbase.mrc-cpe.cam.ac.uk/index.php?& MMN_position=1:1.
Соответственно, один из вариантов осуществления изобретения относится к выделенным FXIa-связывающим антителам, или их антигенсвязывающим фрагментам, содержащим вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 9 и 29, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений в каркасной области таких последовательностей, и также содержащим вариабельную область легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 19 или 39, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений в каркасной области таких последовательностей.
Другой вид модификации вариабельной области представляет собой внесение мутаций в аминокислотные остатки в областях VH и/или VL CDR1, CDR2 и/или CDR3 для усовершенствования одного или более свойств связывания (например, аффинности) интересующего антитела, известное как «созревание аффинности». Можно выполнять сайт-направленный мутагенез или ПЦР-опосредованный мутагенез для внесения мутации(ий), и эффект на связывание антитела, или другое интересующее функциональное свойство, можно оценивать в in vitro или in vivo анализах, как описано в настоящем документе в разделе «Примеры». Можно выполнять консервативные модификации (как описано выше). Мутации могут представлять собой аминокислотные замены, добавления или делеции. Кроме того, как правило, изменяют не более одного, двух, трех, четырех или пяти остатков в области CDR.
Соответственно, в другом варианте осуществления изобретение относится к выделенным FXIa-связывающим антителам, или их антигенсвязывающим фрагментам, состоящим из вариабельной области тяжелой цепи, содержащей область VH CDR1, состоящую из аминокислотной последовательности, выбранной из группы, включающей SEQ ID NOs: 3 и 23, или аминокислотной последовательности, имеющей одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 3 и 23; область VH CDR2, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 4 и 24, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 4 и 24; область VH CDR3, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 5 и 25, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 5 и 25; область VL CDR1, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 13 и 33, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 13 и 33; область VL CDR2, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 14 и 34, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 14 и 34; и область VL CDR3, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 15 и 35, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 15 и 35.
Соответственно, в другом варианте осуществления изобретение относится к выделенным FXIa-связывающим антителам, или их антигенсвязывающим фрагментам, состоящим из вариабельной области тяжелой цепи, содержащей область VH CDR1, состоящую из аминокислотной последовательности, выбранной из группы, включающей SEQ ID NOs: 6 и 26, или аминокислотной последовательности, имеющей одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению SEQ ID NOs: 6 и 26; область VH CDR2, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 7 и 27, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 7 и 27; область VH CDR3, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 8 и 28, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 8 и 28; область VL CDR1, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 16 и 36, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 16 и 36; область VL CDR2, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 17 и 37, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 17 и 37; и область VL CDR3, имеющую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 18 и 38, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений по сравнению с SEQ ID NOs: 18 и 38.
Прививание антигенсвязывающих доменов на альтернативные каркасы или остовы
Можно использовать самые разнообразные каркасы или остовы антитела/иммуноглобулина при условии, что полученный полипептид будет содержать по меньшей мере одну связывающую область, которая специфически связывает FXIa. Такие каркасы или остовы включают 5 основных идиотипов человеческих иммуноглобулинов, или их фрагментов, и включают иммуноглобулины других видов животных, предпочтительно имеющих гуманизированные аспекты. Антитела с одной тяжелой цепью, такие как те, которые обнаружены у верблюдовых, представляют особый интерес в этом отношении. Специалисты в данной области продолжают открывать и разрабатывать новые каркасы, остовы и фрагменты антител.
В одном аспекте изобретение относится к получению антител не на основе иммуноглобулинов с использованием не иммуноглобулиновых каркасов, на которые могут быть привиты области CDR по изобретению. Можно использовать известные или разработанные в будущем не иммуноглобулиновые каркасы и остовы при условии, что они содержат связывающую область, специфичную для целевого белка FXI и/или FXIa. Известные не иммуноглобулиновые каркасы или остовы включают, но не ограничиваются ими, фибронектин (Compound Therapeutics, Inc., Waltham, MA), анкирин (Molecular Partners AG, Zurich, Switzerland), доменные антитела (Domantis, Ltd., Cambridge, MA, и Ablynx nv, Zwijnaarde, Belgium), липокалин (Pieris Proteolab AG, Freising, Germany), малые модульные иммунофармацевтические средства (Trubion Pharmaceuticals Inc., Seattle, WA), макситела (Avidia, Inc., Mountain View, CA), белок A (Affibody AG, Sweden) и аффилин (гамма-кристаллин или убиквитин) (Scil Proteins GmbH, Halle, Germany).
Фибронектиновые каркасы основаны на домене фибронектина III типа (например, десятом модуле фибронектина III типа (10 домене Fn3)). Домен фибронектина III типа имеет 7 или 8 бета-тяжей, распределенных между двумя бета-слоями, которые сами упакованы друг против друга, образуя ядро белка, и также содержат петли (аналогичные областям CDR), которые соединяют бета-тяжи друг с другом и экспонированы для растворителя. Имеется по меньшей мере три такие петли на каждом краю сэндвича из бета-листов, где край является границей белка, перпендикулярной к направлению бета-тяжей (смотри US 6818418). Эти каркасы на основе фибронектина не являются иммуноглобулинами, хотя, в целом, их укладка обладает большим сходством с таковой у наименьшего функционального фрагмента антитела, вариабельной области тяжелой цепи, которая содержит полный антиген-узнающий модуль в IgG у верблюдов и лам. Благодаря такой структуре не иммуноглобулиновое антитело имитирует антигенсвязывающие свойства, которые по характеру и аффинности сходны со свойствами антител. Такие каркасы можно использовать в стратегии рандомизации и перетасовки петель in vitro, что аналогично процессу созревания аффинности антител in vivo. Эти молекулы на основе фибронектина можно использовать в качестве каркасов, где области петель молекулы могут быть заменены областями CDR по изобретению с использованием стандартных методов клонирования.
Технология анкиринов основана на использовании белков c повторяющимися модулями анкирина в качестве каркасов для несения вариабельных областей, которые могут быть использованы для связывания разных мишеней. Повторяющийся модуль анкирина представляет собой 33-аминокислотный полипептид, состоящий из двух антипараллельных α-спиралей и β-поворота. Связывание вариабельных областей в основном оптимизируют с использованием рибосомного дисплея.
Авимеры получают из природного содержащего A-домен белка, такого как LRP-1. Эти домены используются в природе для белок-белковых взаимодействий, и у человека свыше 250 белков имеют в структурной основе A-домены. Авимеры состоят из ряда разных мономеров «A-домена» (2-10), связанных через аминокислотные линкеры. Можно создавать авимеры, которые способны связывать целевой антиген, с использованием методологии, описанной, например, в публикациях патентных заявок США №№ 20040175756, 20050053973, 20050048512 и 20060008844.
Аффинные лиганды аффитела представляют собой небольшие простые белки, состоящие из трехспирального пучка на основе каркаса одного из IgG-связывающих доменов белка A. Белок A представляет собой поверхностный белок бактерии Staphylococcus aureus. Этот каркасный домен состоит из 58 аминокислот, 13 из которых рандомизируют для получения библиотек аффител с большим числом вариантов лигандов (смотри, например, US 5831012). Молекулы аффител имитируют антитела, они имеют молекулярную массу 6 кДа, в сравнении с молекулярной массой антител, которая составляет 150 кДа. Несмотря на небольшой размер, связывающий сайт молекул аффител аналогичен таковому у антител.
Антикалины являются продуктами, разработанными компанией Pieris ProteoLab AG. Они получены из липокаинов, широко распространенной группы небольших и устойчивых белков, которые, как правило, вовлечены в физиологический транспорт или хранение химически чувствительных или нерастворимых соединений. Несколько видов природных липокаинов присутствуют в человеческих тканях или жидкостях организма. Структура белков напоминает структуру иммуноглобулинов, с гипервариабельными петлями на вершине жесткого каркаса. Однако в отличие от антител или их рекомбинантных фрагментов липокаины состоят из одной полипептидной цепи с 160-180 аминокислотными остатками, являясь лишь ненамного крупнее одного домена иммуноглобулинов. Набор из четырех петель, образующих связывающий карман, проявляет выраженную структурную пластичность и допускает присутствие различных боковых цепей. Сайт связывания, таким образом, может быть преобразован патентованным методом для узнавания запланированных целевых молекул разной формы с высокой аффинностью и специфичностью. Один из белков семейства липокаинов, билин-связывающий белок (BBP) из Pieris brassicae, был использован для разработки антикалинов путем мутагенеза набора из четырех петель. Одним из примеров патентных заявок, в которых описаны антикалины, является PCT публикация № WO 199916873.
Молекулы аффилинов представляют собой небольшие белки, не являющиеся иммуноглобулинами, которые сконструированы для обладания специфической аффинностью в отношении белков и малых молекул. Новые молекулы аффилинов можно очень быстро выбирать из двух библиотек, каждая из которых основана на разных человеческих каркасных белках. Молекулы аффилинов не обладают какой-либо структурной гомологией с белками иммуноглобулинов. В настоящее время используют два аффилиновых каркаса, одним из которых является гамма-кристаллин, человеческий структурный белок глазного хрусталика, а другой является представителем суперсемейства белков «убиквитинов». Оба каркасных белка человека имеют очень небольшие размеры, демонстрируют высокую температурную устойчивость и почти полную устойчивость к изменениям pH и денатурирующим средствам. Такая высокая стабильность в основном является результатом протяженной бета-складчатой структуры белков. Примеры белков, полученных из гамма-кристаллина, описаны в WO 200104144 и примеры «убиквитин-подобных» белков описаны в WO 2004106368.
Миметики белковых эпитопов (PEM) представляют собой циклические пептид-подобные молекулы среднего размера (ММ 1-2 кДа), имитирующие бета-шпилечные вторичные структуры белков, основные вторичные структуры, участвующие в белок-белковых взаимодействиях.
Настоящее изобретение относится к полностью человеческим антителам, которые специфически связывают белок FXIa. В сравнении с химерными или гуманизированными антителами, человеческие FXIa-связывающие антитела по изобретению имеют еще более сниженную антигенность при введении субъектам-людям.
Антитела верблюдовых
Белки антител, полученные от представителей семейства верблюдовых двугорбых и одногорбых верблюдов (Camelus bactrianus и Calelus dromaderius), включая представителей семейства из Нового света, таких как ламы (Lama paccos, Lama glama и Lama vicugna), были охарактеризованы с точки зрения размера, структурной сложности и антигенности для субъектов-людей. Некоторые IgG антитела представителей этого семейства млекопитающих, как установлено, естественным образом лишены легких цепей и, таким образом, их структура отличаются от типичной четырехцепочечной четвертичной структуры с двумя тяжелыми и двумя легкими цепями антител других животных. Смотри PCT/EP93/02214 (WO 94/04678, дата публикации 3 марта 1994 г.).
Область антитела верблюдовых, которая представляет собой небольшой одиночный вариабельный домен, идентифицированный как VHH, можно получать методами генетической инженерии в виде небольшого белка с высокой аффинностью для мишени, представляющего собой низкомолекулярный полученный из антитела белок, известный как «нанотело верблюдовых». Смотри патент США № 5759808, выданный 2 июня, 1998 г.; смотри также Stijlemans, B. et al., 2004 J Biol Chem 279: 1256-1261; Dumoulin, M. et al., 2003 Nature 424: 783-788; Pleschberger, M. et al. 2003 Bioconjugate Chem 14: 440-448; Cortez-Retamozo, V. et al. 2002 Int J Cancer 89: 456-62; и Lauwereys, M. et al. 1998 EMBO J 17: 3512-3520. Сконструированные библиотеки антител и фрагментов антител верблюдовых коммерчески доступны, например, от компании Ablynx, Ghent, Belgium. Как и в случае других антител, отличных от человеческих, аминокислотная последовательность антитела верблюдовых может быть изменена рекомбинантными методами, с получением последовательности, которая более близко напоминает человеческую последовательность, то есть, нанотело может быть «гуманизированным». Таким образом, естественная низкая антигенность антител верблюдовых для человека может быть дополнительно снижена.
Нанотело верблюдовых имеет молекулярную массу, составляющую примерно одну десятую часть молекулярной массы молекулы человеческого IgG, и белок имеет физический диаметр всего несколько нанометров. Одним из следствий небольшого размера является способность нанотел верблюдовых связываться с антигенными сайтами, которые являются функционально невидимыми для более крупных белков антител, то есть, нанотела верблюдовых полезны в качестве реагентов для обнаружения антигенов, которые остались бы незаметными при использовании классических иммунологических методов, а также в качестве возможных терапевтических средств. Таким образом, еще одним следствием небольшого размера является то, что нанотело верблюдовых может ингибировать за счет связывания с конкретным сайтом в бороздке или узкой щели целевого белка и, таким образом, может выполнять роль, которая больше напоминает функцию классического низкомолекулярного лекарственного средства, чем функцию классического антитела.
Низкая молекулярная масса и компактный размер также приводят к тому, что нанотела верблюдовых являются чрезвычайно термоустойчивыми, устойчивыми к экстремальным значениям pH и к протеолитическому расщеплению, а также слабо антигенными. Другим следствием является то, что нанотела верблюдовых легко перемещаются из системы кровообращения в ткани, и даже пересекают гематоэнцефалический барьер и могут лечить заболевания, поражающие нервную ткань. Нанотела также могут способствовать переносу лекарственного средства через гематоэнцефалический барьер. Смотри патентную заявку США 20040161738, опубликованную 19 августа 2004 г. Эти особенности в сочетании с низкой антигенностью для людей свидетельствуют об огромном терапевтическом потенциале. Кроме того, эти молекулы могут полностью экспрессироваться в прокариотических клетках, таких как E. coli, экспрессируются в виде слитых белков с бактериофагом и являются функциональными.
Соответственно, одним из признаков по настоящему изобретению является антитело или нанотело верблюдовых, имеющее высокую аффинность для FXI и/или FXIa. В конкретных вариантах осуществления настоящего изобретения антитело или нанотело верблюдовых получено естественным образом в организме представителя верблюдовых, то есть, получено в организме представителя верблюдовых после иммунизации FXI и/или FXIa, или их пептидным фрагментом, с использованием методов, описанных в настоящем документе для других антител. Альтернативно, связывающее FXI и/или FXIa нанотело верблюдовых является сконструированным, то есть, полученным путем селекции, например, из библиотеки фагового дисплея соответствующим образом мутированных белков нанотел верблюдовых с использованием методов пэннинга с FXI и/или FXIa, и/или их доменами, и/или пептидными фрагментами, в качестве мишени, как описано в примерах, приведенных в настоящем документе. Сконструированные нанотела могут быть дополнительно модифицированы методами генной инженерии, чтобы иметь период полувыведения в организме субъекта-реципиента от 45 минут до двух недель. В конкретном варианте осуществления антитело или нанотело верблюдовых получают путем прививания последовательностей CDR тяжелой или легкой цепи человеческих антител по изобретению на каркасные последовательности нанотела или однодоменного антитела, как описано, например, в PCT/EP93/02214.
Биспецифические молекулы и мультивалентные антитела
В другом аспекте настоящее изобретение относится к биспецифическим или мультиспецифическим молекулам, содержащим связывающее FXI и/или FXIa антитело, или его фрагмент, по изобретению. Антитело по изобретению, или его антигенсвязывающие фрагменты, может быть дериватизировано или связано с другой функциональной молекулой, например, другим пептидом или белком (например, другим антителом или лигандом для рецептора) для получения биспецифической молекулы, которая связывает по меньшей мере два разных сайта связывания или две молекулы-мишени. Фактически, антитело по изобретению может быть дериватизировано или связано с более чем одной другой функциональной молекулой для получения мультиспецифических молекул, которые связывают более двух разных сайтов связывания и/или молекул-мишеней; такие мультиспецифические молекулы также охвачены используемым в настоящем документе термином «биспецифическая молекула». Для создания биспецифической молекулы по изобретению антитело по изобретению может быть функционально связано (например, за счет химического связывания, генетического слияния, нековалентной ассоциации или иным образом) с одной или более другими связывающими молекулами, такими как другое антитело, фрагмент антитела, пептид или связывающий миметик, с получением биспецифической молекулы.
Соответственно, настоящее изобретение включает биспецифические молекулы, имеющие по меньшей мере одну первую специфичность связывания для FXI и/или FXIa и вторую специфичность связывания для второго целевого эпитопа. Например, второй целевой эпитоп представляет собой другой эпитоп FXI и/или FXIa, отличный от первого целевого эпитопа.
Кроме того, в варианте осуществления изобретения, в котором биспецифическая молекула является мультиспецифической, молекула может дополнительно иметь третью специфичность связывания для дополнительного эпитопа, помимо первого и второго целевых эпитопов.
В одном варианте осуществления биспецифические молекулы по изобретению содержат в качестве специфичности связывания по меньшей мере одно антитело, или его фрагмент, включая, например, Fab, Fab', F(ab')2, Fv или одноцепочечный Fv-фрагмент. Антитело также может представлять собой димер легкой цепи или тяжелой цепи, или их любой минимальный фрагмент, такой как Fv, или одноцепочечный конструкт, как описано в Ladner et al. патент США № 4946778.
Диатела представляют собой двухвалентные, биспецифические молекулы, в которых домены VH и VL экспрессированы на одной полипептидной цепи и связаны посредством линкера, который является слишком коротким, чтобы допускать спаривание между двумя доменами на одной и той же цепи. Домены VH и VL образуют пары с комплементарными доменами другой цепи, в результате чего образуются два антигенсвязывающих сайта (смотри, например, Holliger et al., 1993 Proc. Natl. Acad. Sci. USA 90: 6444-6448; Poljak et al., 1994 Structure 2: 1121-1123). Диатела могут быть получены путем экспрессии двух полипептидных цепей со структурой либо VHA-VLB и VHB-VLA (конфигурация VH-VL), либо VLA-VHB и VLB-VHA (конфигурация VL-VH), в одной и той же клетке. Большинство из них могут экспрессироваться в растворимой форме в бактериях. Одноцепочечные диатела (scDb) получают путем соединения двух образующих диатело полипептидных цепей линкером из примерно 15 аминокислотных остатков (смотри Holliger and Winter, 1997 Cancer Immunol. Immunother., 45(3-4): 128-30; Wu et al., 1996 Immunotechnology, 2(1): 21-36). scDb могут экспрессироваться в бактериях в растворимой активной мономерной форме (смотри Holliger and Winter, 1997 Cancer Immunol. Immunother., 45(34): 128-30; Wu et al., 1996 Immunotechnology, 2(1): 21-36; Pluckthun and Pack, 1997 Immunotechnology, 3(2): 83-105; Ridgway et al., 1996 Protein Eng., 9(7): 617-21). Диатело может быть слито с Fc-фрагментом для получения «ди-диатела» (смотри Lu et al., 2004 J. Biol. Chem., 279(4): 2856-65).
Другие антитела, которые можно использовать в биспецифических молекулах по изобретению, представляют собой мышиные, химерные и гуманизированные моноклональные антитела.
Биспецифические молекулы можно получать путем конъюгации составляющих специфичностей связывания методами, известными в данной области. Например, каждую специфичность связывания биспецифической молекулы можно создавать отдельно, а затем конъюгировать друг с другом. Когда специфичности связывания представляют собой белки или пептиды, можно использовать различные связывающие или сшивающие реагенты для ковалентной конъюгации. Примеры сшивающих реагентов включают белок A, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 5,5'-дитиобис(2-нитробензойную кислоту) (DTNB), o-фенилендималеинимид (oPDM), N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) и сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC) (смотри, например, Karpovsky et al., 1984 J. Exp. Med. 160: 1686; Liu, MA et al., 1985 Proc. Natl. Acad. Sci. USA 82: 8648). Другие методы включают те, которые описаны в Paulus, 1985 Behring Ins. Mitt. No. 78, 118-132; Brennan et al., 1985 Science 229: 81-83 и Glennie et al., 1987 J. Immunol. 139: 2367-2375. Конъюгирующими реагентами являются SATA и сульфо-SMCC, оба доступны от компании Pierce Chemical Co. (Rockford, IL).
Когда специфичности связывания представляет собой антитела, они могут быть конъюгированы путем сульфгидрильного связывания C-концевых шарнирных областей двух тяжелых цепей. В конкретном варианте осуществления шарнирную область модифицируют для содержания нечетного числа сульфгидрильных остатков, например, одного, перед конъюгацией.
Альтернативно, обе специфичности связывания могут быть закодированы в одном и том же векторе, и экспрессироваться и собираться в одной и той же клетке-хозяине. Этот способ особенно полезен, когда биспецифическая молекула представляет собой слитый белок мАт x мАт, мАт x Fab, Fab x F(ab')2 или лиганд x Fab. Биспецифическая молекула по изобретению может представлять собой одноцепочечную молекулу, содержащую одно одноцепочечное антитело и связывающую детерминанту, или одноцепочечную биспецифическую молекулу, содержащую две связывающие детерминанты. Биспецифические молекулы могут содержать по меньшей мере две одноцепочечные молекулы. Способы получения биспецифических молекул описаны, например, в патенте США номер 5260203; патенте США номер 5455030; патенте США номер 4881175; патенте США номер 5132405; патенте США номер 5091513; патенте США номер 5476786; патенте США номер 5013653; патенте США номер 5258498 и патенте США номер 5482858.
Связывание биспецифических молекул с их специфическими мишенями можно подтверждать, например, иммуноферментным анализом (ELISA), радиоиммуноанализом (REA), FACS-анализом, биоанализом (например, ингибированием роста) или вестерн-блот анализом. В каждом из этих анализов, как правило, определяют наличие интересующих комплексов белок-антитело с использованием меченого реагента (например, антитела), специфичного для интересующего комплекса.
В другом аспекте настоящее изобретение относится к мультивалентным соединениям, содержащим по меньшей мере два идентичных или разных антигенсвязывающих фрагмента антител по изобретению, связывающих FXIa. Антигенсвязывающие фрагменты могут быть связаны вместе за счет слияния белков, либо за счет ковалентной или нековалентной связи. Альтернативно, методы связывания были описаны для биспецифических молекул. Тетравалентные соединения могут быть получены, например, путем сшивания антител по изобретению с антителом, которое связывает константные области антител по изобретению, например, Fc или шарнирную область.
Тримеризированные домены описаны, например, в Borean патенте EP 1 012 280B1. Пентамеризированные модули описаны, например, в PCT/EP97/05897.
Антитела с увеличенным периодом полувыведения
Настоящее изобретение относится к антителам, специфически связывающим белок FXIa, которые имеют увеличенный период полувыведения in vivo.
Многие факторы могут влиять на период полувыведения белка in vivo. Например, фильтрование в почках, метаболизм в печени, расщепление протеолитическими ферментами (протеазами) и иммунные ответы (например, нейтрализация белка антителами и поглощение макрофагами и дендритными клетками). Различные стратегии можно использовать для продления периода полувыведения антител по настоящему изобретению. Например, химическое связывание с полиэтиленгликолем (PEG), reCODE PEG, каркасом антител, полисиаловой кислотой (PSA), гидроксиэтилкрахмалом (HES), альбумин-связывающими лигандами и углеводными защитными группами; генетическое слияние с белками, связывающимися с сывороточными белками, такими как альбумин, IgG, FcRn и трансферрин; связывание (генетическое или химическое) с другими связывающими фрагментами, которые связывают сывороточные белки, такими как нанотела, Fab-фрагменты, дарпины, авимеры, аффитела и антикалины; генетическое слияние с rPEG, альбумином, доменом альбумина, альбумин-связывающими белками и Fc-фрагментом; или заключение в наноносители, препараты с замедленным высвобождением или медицинские устройства.
Для продления циркуляции антител в сыворотке in vivo инертные полимерные молекулы, такие как высокомолекулярный PEG, можно присоединять к антителу, или его фрагменту, с добавлением или без добавления мультифункционального линкера, либо посредством сайт-специфической конъюгации PEG к N- или C-концу антител, либо через эпсилон-аминогруппы, находящиеся на остатках лизина. Для пегилирования антитела, как правило, проводят реакцию антитела, или его фрагмента, с полиэтиленгликолем (PEG), например, реакционноспособным сложноэфирным или альдегидным производным PEG, в условиях, при которых одна или более групп PEG присоединяются к антителу, или фрагменту антитела. Пегилирование можно проводить путем реакции ацилирования или реакции алкилирования с реакционноспособной молекулой PEG (или аналогичным реакционноспособным водорастворимым полимером). Используемый в настоящем документе термин «полиэтиленгликоль» охватывает любую из форм PEG, используемую для дериватизации других белков, например, моно(C1-C10) алкокси- или арилоксиполиэтиленгликоль или полиэтиленгликоль-малеинимид. В конкретных вариантах осуществления пегилируемое антитело представляет собой aгликозилированное антитело. Используют дериватизацию линейным или разветвленным полимером, что приводит к минимальной потере биологической активности. Степень конъюгации можно тщательно контролировать методом SDS-ПААГ и масс-спектрометрии для обеспечения надлежащей конъюгации молекул PEG с антителами. Непрореагировавший PEG можно отделять от конъюгатов антитело-PEG эксклюзионной или ионообменной хроматографией. Дериватизированные PEG антитела можно тестировать на связывающую способность, а также на in vivo эффективность методами, хорошо известными специалистам в данной области, например, с использованием иммуноанализов, описанных в настоящем документе. Методы пегилирования белков известны в данной области и могут быть применены к антителам по изобретению. Смотри, например, EP 0 154 316 авторов Nishimura et al. и EP 0 401 384 авторов Ishikawa et al.
Другие технологии модифицирующего пегилирования включают технологию восстановительной химически ортогонально направленной инженерии (ReCODE PEG), которая позволяет включать химически точно определенные боковые цепи в биосинтетические белки посредством восстановленной системы, включающей тРНК-синтетазу и тРНК. Эта технология позволяет включать более 30 новых аминокислот в биосинтетические белки в E. coli, дрожжах и клетках млекопитающих. тРНК включает «неродную» аминокислоту в любом месте, где расположен амбер-кодон, превращая амбер-кодон из стоп-кодона в кодон, который дает сигнал о включении химически точно определенной аминокислоты.
Технологию рекомбинантного пегилирования (rPEG) также можно использовать для увеличения периода полувыведения в сыворотке. Данная технология включает генетическое слияние неструктурированного белкового «хвоста» из 300-600 аминокислот с существующим фармацевтическим белком. Поскольку кажущаяся молекулярная масса такой неструктурированной белковой цепи примерно в 15-раз больше, чем ее фактическая молекулярная масса, период полувыведения белка в сыворотке намного возрастает. В отличие от традиционного пегилирования, для которого необходима химическая конъюгация и повторная очистка, производственный процесс сильно упрощается и продукт является гомогенным.
Полисиалилирование является другой технологией, в которой используют природный полимер полисиаловую кислоту (PSA) для продления активного состояния и повышения стабильности терапевтических пептидов и белков. PSA представляет собой полимер сиаловой кислоты (сахара). При использовании для доставки белкового и пептидного терапевтического средства полисиаловая кислота обеспечивает защитное микроокружение за счет конъюгации. Это продлевает активное состояние терапевтического белка в системе циркуляции и предотвращает его узнавание иммунной системой. Полимер PSA естественным образом присутствует в теле человека. Некоторые бактерии за миллионы лет эволюционировали таким образом, что их стенки покрыты этим полимером. Такие естественным образом полисиалилированные бактерии стали способны, в силу молекулярной мимикрии, обманывать защитную систему организма. PSA, основа природной технологии создания предельной незаметности, может быть легко получена из таких бактерий в больших количествах и с заранее определенными физическими характеристиками. Бактериальная PSA является абсолютно не иммуногенной, даже при связывании с белками, поскольку она химически идентична PSA в теле человека.
Другая технология включает использование гидроксиэтилкрахмальных («HES») производных, связанных с антителами. HES представляет собой модифицированный природный полимер из крахмала восковой кукурузы и может метаболизироваться ферментами тела. Растворы HES, как правило, вводят для восполнения недостатка объема крови и для улучшения реологических свойств крови. HESилирование антитела позволяет увеличивать период полувыведения в системе циркуляции за счет повышения стабильности молекулы, а также за счет уменьшения почечного клиренса, что приводит к увеличению биологической активности. Изменяя разные параметры, такие как молекулярная масса HES, можно получать самые разные конъюгаты HES с антителом, адаптированные для конкретных задач.
Антитела, имеющие увеличенный период полувыведения in vivo, также можно получать путем внесения одной или более аминокислотных модификаций (то есть, замен, вставок или делеций) в константный домен IgG или его FcRn связывающий фрагмент (предпочтительно Fc-домен или шарнирный участок Fc-домена). Смотри, например, международную публикацию № WO 98/23289; международную публикацию № WO 97/34631 и патент США № 6277375.
Кроме того, антитела можно конъюгировать с альбумином (например, человеческим сывороточным альбумином; ЧСА) для придания антителу, или фрагменту антитела, большей стабильности in vivo или увеличения периода полувыведения in vivo. Данные методы хорошо известны в данной области, смотри, например, международные публикации №№ WO 93/15199, WO 93/15200 и WO 01/77137; и Европейский патент № EP 413622. Кроме того, в контексте биспецифического антитела, описанного выше, специфичности антитела можно конструировать таким образом, что один связывающий домен антитела связывает FXIa, в то время как второй связывающий домен антитела связывает сывороточный альбумин, предпочтительно ЧСА.
Стратегии увеличения периода полувыведения особенно полезны в случае нанотел, связывающих соединений на основе фибронектина и других антител или белков, для которых желательно увеличение периода полувыведения in vivo.
Конъюгаты антитела
Настоящее изобретение относится к антителам, или их фрагментам, специфически связывающим белок FXIa, которые рекомбинантно слиты или химически конъюгированы (включая как ковалентную, так и не ковалентную конъюгацию) с гетерологичным белком или полипептидом (или его фрагментом, предпочтительно с полипептидом из по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислот) для получения слитых белков. В частности, изобретение относится к слитым белкам, содержащим антигенсвязывающий фрагмент антитела, описанного в настоящем документе (например, Fab-фрагмент, Fd-фрагмент, Fv-фрагмент, F(ab)2-фрагмент, домен VH, VH CDR, домен VL или VL CDR), и гетерологичный белок, полипептид или пептид. Способы слияния или конъюгации белков, полипептидов или пептидов с антителом, или фрагментом антитела, известны в данной области. Смотри, например, патенты США №№ 5336603, 5622929, 5359046, 5349053, 5447851 и 5112946; Европейские патенты №№ EP 307434 и EP 367166; международные публикации патентов №№ WO 96/04388 и WO 91/06570; Ashkenazi et al., 1991, Proc. Natl. Acad. Sci. USA 88: 10535-10539; Zheng et al., 1995, J. Immunol. 154: 5590-5600; и Vil et al., 1992, Proc. Natl. Acad. Sci. USA 89: 11337-11341.
Дополнительные слитые белки могут быть получены с использованием методов перетасовки генов, перетасовки мотивов, перетасовки экзонов и/или перетасовки кодонов (носящих общее название «перетасовка ДНК»). Перетасовку ДНК можно использовать для изменения активностей антител, или их фрагментов, по изобретению (например, для получения антител, или их фрагментов, с более высокими показателями аффинности и более низкими скоростями диссоциации). Смотри, в основном, патенты США №№ 5605793, 5811238, 5830721, 5834252 и 5837458; Patten et al., 1997, Curr. Opinion Biotechnol. 8: 724-33; Harayama, 1998, Trends Biotechnol. 16(2): 76-82; Hansson, et al., 1999, J. Mol. Biol. 287: 265-76 и Lorenzo and Blasco, 1998, Biotechniques 24(2): 308-313 (полное содержание всех этих патентов и публикаций включено в настоящий документ посредством ссылки). Антитела или их фрагменты, или закодированные антитела или их фрагменты, можно изменять, подвергая их случайному мутагенезу методом ПЦР с пониженной точностью, случайной вставки нуклеотидов или другими методами перед рекомбинацией. Можно проводить рекомбинацию полинуклеотида, кодирующего антитело, или его фрагмент, которое специфически связывает белок FXIa, с одним или более компонентами, мотивами, секциями, частями, доменами, фрагментами, и так далее, одной или более гетерологичных молекул.
Кроме того, можно проводить слияние антител, или их фрагментов, с маркерными последовательностями, например, с пептидом, облегчающим очистку. В предпочтительных вариантах осуществления маркерная аминокислотная последовательность представляет собой гексагистидиновый пептид (SEQ ID NO: 48), такой как тег, предоставляемый в векторе pQE (QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, CA, 91311), в числе прочих, многие из таких последовательностей коммерчески доступны. Как описано в публикации Gentz et al., 1989, Proc. Natl. Acad. Sci. USA 86: 821-824, например, гексагистидин (SEQ ID NO: 48) обеспечивает удобную очистку слитого белка. Другие пептидные теги, полезные для очистки, включают, но не ограничиваются ими, гемагглютининовый («HA») тег, который соответствует эпитопу, получаемому из белка гемагглютинина вируса гриппа (Wilson et al., 1984, Cell 37: 767), и тег «flag».
В других вариантах осуществления антитела по настоящему изобретению, или их фрагменты, конъюгированы с диагностическим или детектируемым средством. Такие антитела могут быть полезны для мониторинга или прогнозирования начала развития, прогрессирования и/или степени тяжести заболевания или нарушения в виде части клинической процедуры тестирования, такой как определение эффективности конкретной терапии. Такое диагностирование и детекцию можно осуществлять путем связывания антитела с детектируемыми средствами, включая, но без ограничения, различные ферменты, такие как, но без ограничения, пероксидаза хрена, щелочная фосфатаза, бета-галактозидаза или ацетилхолинэстераза; простетические группы, такие как, но без ограничения, стрептавидин/биотин и авидин/биотин; флуоресцентные материалы, такие как, но без ограничения, умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансил хлорид или фикоэритрин; люминесцентные материалы, такие как, но без ограничения, люминол; биолюминесцентные материалы, такие как, но без ограничения, люцифераза, люциферин и экворин; радиоактивные материалы, такие как, но без ограничения, йод (131I, 125I, 123I и 121I), углерод (14C), сера (35S), тритий (3H), индий (115In, 113In, 112In и 111In), технеций (99Tc), таллий (201Ti), галлий (68Ga, 67Ga), палладий (103Pd), молибден (99Mo), ксенон (133Xe), фтор (18F), 153Sm, 177Lu, 159Gd, 149Pm, 140La, 175Yb, 166Ho, 90Y, 47Sc, 186Re, 188Re, 142Pr, 105Rh, 97Ru, 68Ge, 57Co, 65Zn, 85Sr, 32P, 153Gd, 169Yb, 51Cr, 54Mn, 75Se, 113Sn и 117Tin; и позитронно-активные металлы для использования в различных методах позитрон-эмиссионной томографии, а также нерадиоактивные парамагнитные ионы металлов.
Настоящее изобретение также включает применение антител, или их фрагментов, конъюгированных с терапевтическим средством. Антитело, или его фрагмент, можно конъюгировать с таким терапевтическим фрагментом, как цитотоксин, например, цитостатическое или цитоцидное средство, терапевтическое средство или ион радиоактивного металла, например, излучатели альфа-частиц. Цитотоксин, или цитотоксическое средство, включает любое средство, губительное для клеток.
Кроме того, антитело, или его фрагмент, можно конъюгировать с терапевтическим средством или лекарственным средством, которое модифицирует конкретный биологический ответ. Терапевтические средства или лекарственные средства не следует понимать, как ограниченные классическими химическими лекарственными средствами. Например, лекарственное средство может представлять собой белок, пептид или полипептид, обладающий желательной биологической активностью. Такие белки могут включать, например, токсин, такой как абрин, рицин A, экзотоксин псевдомонад, холерный токсин или дифтерийный токсин; белок, такой как фактор некроза опухолей, α-интерферон, β-интерферон, фактор роста нервов, тромбоцитарный фактор роста, тканевой активатор плазминогена, апоптотическое средство, антиапоптотическое средство или модификатор биологического ответа, такой как, например, лимфокин.
Кроме того, антитело можно конъюгировать с терапевтическими средствами, такими как ион радиоактивного металла, например, излучатели альфа-частиц, такие как 213Bi, или макроциклическими хелаторами, полезными для конъюгации ионов радиоактивных металлов, включая, но без ограничения, 131In, 131LU, 131Y, 131Ho, 131Sm, с полипептидами. В конкретных вариантах осуществления макроциклический хелатор представляет собой 1,4,7,10-тетраазациклододекан-N,N',N'',N'''-тетрауксусную кислоту (DOTA), которая может быть присоединена к антителу через молекулу линкера. Такие молекулы линкеров общеизвестны в данной области и описаны в публикациях Denardo et al., 1998, Clin Cancer Res. 4(10): 2483-90; Peterson et al., 1999, Bioconjug. Chem. 10(4): 553-7; и Zimmerman et al., 1999, Nucl. Med. Biol. 26(8): 943-50, полное содержание каждой из которых включено в настоящий документ посредством ссылки.
Методы конъюгации терапевтических средств с антителами хорошо известны, смотри, например, Arnon et al., «Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy», в: Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., «Antibodies For Drug Delivery», в: Controlled Drug Delivery (2-е издание), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review», в: Monoclonal Antibodies 84: Biological And Clinical Applications, Pinchera et al. (eds.), pp. 475-506 (1985); «Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy», в: Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), и Thorpe et al., 1982, Immunol. Rev. 62: 119-58.
Антитела также можно связывать с твердыми подложками, которые особенно полезны для проведения иммуноанализов или очистки целевого антигена. Такие твердые подложки включают, но не ограничиваются ими, стекло, целлюлозу, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен.
Способы получения антител
Нуклеиновые кислоты, кодирующие антитела
Изобретение относится к по существу очищенным молекулам нуклеиновой кислоты, кодирующим полипептиды, содержащие сегменты или домены цепей FXIa-связывающего антитела, описанного выше. Некоторые из нуклеиновых кислот по изобретению содержат нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи, приведенную в SEQ ID NO: 10 или 30, и/или нуклеотидную последовательность, кодирующую вариабельную область легкой цепи, приведенную в SEQ ID NO: 20 или 40. В конкретном варианте осуществления молекулы нуклеиновой кислоты являются теми, которые приведены в таблице 1. Некоторые другие молекулы нуклеиновой кислоты по изобретению содержат нуклеотидные последовательности, которые по существу идентичны (например, по меньшей мере на 65%, 80%, 95% или 99%) нуклеотидным последовательностям, приведенным в таблице 1. При экспрессии с соответствующих экспрессионных векторов полипептиды, кодируемые этими полинуклеотидами, способны проявлять способность к связыванию антигена FXI и/или FXIa.
Изобретение также относится к полинуклеотидам, кодирующим по меньшей мере одну область CDR и, как правило, все три области CDR, тяжелой или легкой цепи FXIa-связывающего антитела, описанного выше. Некоторые другие полинуклеотиды кодируют всю или практически всю последовательность вариабельной области тяжелой цепи и/или легкой цепи FXIa-связывающего антитела, описанного выше. Вследствие вырожденности генетического кода, разные нуклеотидные последовательности будут кодировать каждую из аминокислотных последовательностей иммуноглобулина.
Молекулы нуклеиновой кислоты по изобретению могут кодировать как вариабельную область, так и константную область антитела. Некоторые из нуклеотидных последовательностей по изобретению содержат нуклеотиды, кодирующие последовательность тяжелой цепи, которая по существу идентична (например, по меньшей мере на 80%, 90% или 99%) последовательности тяжелой цепи, приведенной в SEQ ID NO: 11 или 31. Некоторые другие нуклеотидные последовательности содержат нуклеотиды, кодирующие последовательность легкой цепи, которая по существу идентична (например, по меньшей мере на 80%, 90% или 99%) последовательности легкой цепи, приведенной в SEQ ID NO: 21 или 41.
Полинуклеотидные последовательности могут быть получены de novo твердофазным синтезом ДНК или ПЦР-мутагенезом существующей последовательности (например, последовательностей, описанных в разделе «Примеры», ниже), кодирующей FXIa-связывающее антитело или его связывающий фрагмент. Прямой химический синтез нуклеиновых кислот можно выполнять методами, известными в данной области, такими как фосфотриэфирный метод, описанный в Narang et al., 1979, Meth. Enzymol. 68: 90; фосфодиэфирный метод, описанный в Brown et al., Meth. Enzymol. 68: 109, 1979; диэтилфосфорамидитный метод, описанный в Beaucage et al., Tetra. Lett., 22: 1859, 1981; и метод с твердой подложкой, описанный в патенте США № 4458066. Внесение мутаций в полинуклеотидную последовательность методом ПЦР можно выполнять, как описано, например, в PCR Technology: Principles and Applications for DNA Amplification, H.A. Erlich (Ed.), Freeman Press, NY, NY, 1992; PCR Protocols: A Guide to Methods and Applications, Innis et al. (Ed.), Academic Press, San Diego, CA, 1990; Mattila et al., Nucleic Acids Res. 19: 967, 1991; и Eckert et al., PCR Methods and Applications 1: 17, 1991.
Изобретение также относится к экспрессионным векторам и клеткам-хозяевам для продуцирования связывающих FXI и/или FXIa антител, описанных выше. Можно использовать различные экспрессионные векторы для экспрессии полинуклеотидов, кодирующих цепи FXIa-связывающего антитела, или связывающих фрагментов. Можно использовать как экспрессионные векторы на основе вирусов, так и невирусные экспрессионные векторы, для продуцирования антител в клетке-хозяине млекопитающего. Невирусные векторы и системы включают плазмиды, эписомные векторы, как правило, с экспрессионной кассетой для экспрессии белка или РНК, и человеческие искусственные хромосомы (смотри, например, Harrington et al., Nat Genet 15: 345, 1997). Например, невирусные векторы, полезные для экспрессии полинуклеотидов FXIa-связывающих полипептидов в клетках млекопитающих (например, человека), включают pThioHis A, B и C, pcDNA3.1/His, pEBVHis A, B и C, (Invitrogen, San Diego, CA), векторы MPSV, а также многие другие векторы, известные в данной области, для экспрессии других белков. Полезные вирусные векторы включают векторы на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, вирусов герпеса, векторы на основе SV40, вируса папилломы, вируса Эпштейна-Барр HBP, вируса осповакцины и вируса леса Семлики (SFV). Смотри, Brent et al., выше; Smith, Annu. Rev. Microbiol. 49: 807, 1995; и Rosenfeld et al., Cell 68: 143, 1992.
Выбор экспрессионного вектора зависит от планируемых клеток-хозяев, в которых вектор будет экспрессироваться. Как правило, экспрессионные векторы содержат промотор и другие регуляторные последовательности (например, энхансеры), которые функционально связаны с полинуклеотидами, кодирующими цепь или фрагмент FXIa-связывающего антитела. В некоторых вариантах осуществления используют индуцируемый промотор для предотвращения экспрессии вставленных последовательностей в отсутствие индуцирующих условий. Индуцируемые промоторы включают, например, арабинозный, lacZ, металлотионеиновый промотор или промотор белка теплового шока. Культуры трансформированных микроорганизмов можно наращивать в не индуцирующих условиях без оказания влияния на популяцию кодирующих последовательностей, продукты экспрессии которых лучше переносятся клетками-хозяевами. Помимо промоторов, для эффективной экспрессии цепи или фрагмента FXIa-связывающего антитела также могут быть необходимы или желательны и другие регуляторные элементы. Эти элементы, как правило, включают инициирующий кодон ATG и соседний участок связывания рибосомы или другие последовательности. Кроме того, эффективность экспрессии можно увеличивать за счет включения энхансеров, соответствующих используемой клеточной системе (смотри, например, Scharf et al., Results Probl. Cell Differ. 20: 125, 1994; и Bittner et al., Meth. Enzymol., 153: 516, 1987). Например, для увеличения экспрессии в клетках-хозяевах млекопитающих можно использовать энхансер SV40 или энхансер CMV.
Экспрессионные векторы также могут предоставлять положение сигнальной последовательности секреции для образования слитого белка с полипептидами, кодируемыми вставленными последовательностями FXIa-связывающего антитела. Чаще, вставленные последовательности связывающего FXI и/или FXIa антитела связывают с сигнальными последовательностями до включения в вектор. Векторы, используемые для приема последовательностей, кодирующих вариабельные домены легкой и тяжелой цепей связывающего FXI и/или FXIa антитела, иногда также кодируют константные области или их части. Такие векторы делают возможной экспрессию вариабельных областей в виде слитых белков с константными областями, результатом чего является продуцирование интактных антител или их фрагментов. Как правило, такие константные области являются человеческими.
Клетки-хозяева для содержания и экспрессии последовательностей цепей связывающего FXI и/или FXIa антитела могут быть либо прокариотическими, либо эукариотическими. E. coli является одним из прокариотических хозяев, полезных для клонирования и экспрессии полинуклеотидов по настоящему изобретению. Другие подходящие для использования микробные хозяева включают бациллы, такие как Bacillus subtilis, и другие энтеробактерии, такие как Salmonella, Serratia, а также различные виды Pseudomonas. В этих прокариотических хозяевах также могут присутствовать экспрессионные векторы, которые, как правило, содержат последовательности контроля экспрессии, совместимые с клеткой-хозяином (например, точку начала репликации). Кроме того, там будет содержаться любое число различных хорошо известных промоторов, таких как лактозная промоторная система, триптофановая (trp) промоторная система, бета-лактамазная промоторная система или промоторная система из фага лямбда. Промоторы, как правило, контролируют экспрессию, необязательно, с последовательностью оператора, и также присутствуют последовательности сайта связывания рибосомы, и тому подобное, для инициации и завершения транскрипции и трансляции. Другие микроорганизмы, такие как дрожжи, также можно использовать для экспрессии FXIa-связывающих полипептидов по изобретению. Также можно использовать клетки насекомых в сочетании с бакуловирусными векторами.
В некоторых предпочтительных вариантах осуществления для экспрессии и продуцирования связывающих FXI и/или FXIa полипептидов по настоящему изобретению используют клетки-хозяева млекопитающих. К ним относятся любые нормальные смертные, либо нормальные или аномальные иммортализованные клетки животных или человека. Например, был разработан ряд подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины, включая линии клеток CHO, различные линии клеток Cos, клетки HeLa, линии клеток миеломы и трансформированные B-клетки. Использование культур клеток млекопитающих для экспрессии полипептидов, в основном, описано, например, в сборнике Winnacker, FROM GENES TO CLONES, VCH Publishers, N.Y., N.Y., 1987. Экспрессионные векторы для клеток-хозяев млекопитающих могут содержать последовательности контроля экспрессии, такие как точка начала репликации, промотор и энхансер (смотри, например, Queen, et al., Immunol. Rev. 89: 49-68, 1986), а также необходимые сайты процессинга информации, такие как сайты связывания рибосомы, сайты сплайсинга РНК, сайты полиаденилирования и последовательности терминации транскрипции.
Эти экспрессионные векторы, как правило, содержат промоторы, полученные из генов млекопитающих или из вирусов млекопитающих. Подходящие промоторы могут быть конститутивными, специфичными для типа клеток, специфичными для стадии развития и/или модулируемыми или регулируемыми. Полезные промоторы включают, но не ограничиваются ими, металлотионеиновый промотор, конститутивный аденовирусный главный поздний промотор, дексаметазон-индуцируемый промотор MMTV, промотор SV40, промотор MRP polIII, конститутивный промотор MPSV, тетрациклин-индуцируемый промотор CMV (такой как человеческий предранний промотор CMV), конститутивный промотор CMV и сочетания промотор-энхансер, известные в данной области.
Методы введения экспрессионных векторов, содержащих интересующие полинуклеотидные последовательности, варьируются в зависимости от типа клетки-хозяина. Например, трансфекцию с хлоридом кальция обычно используют для прокариотических клеток, в то время как обработку фосфатом кальция или электропорацию можно использовать для других клеток-хозяев. (Смотри, в основном, Sambrook, et al., выше). Другие методы включают, например, электропорацию, обработку фосфатом кальция, опосредованную липосомами трансформацию, инъекцию и микроинъекцию, баллистические методы, виросомы, иммунолипосомы, конъюгаты поликатион:нуклеиновая кислота, «голую» ДНК, искусственные вирионы, слияние со структурным белком VP22 вируса герпеса (Elliot and O'Hare, Cell 88: 223, 1997), усиленное специальными средствами поглощение ДНК и ex vivo трансдукцию. Для долгосрочного высокопродуктивного производства рекомбинантных белков часто бывает нужна стабильная экспрессия. Например, линии клеток, стабильно экспрессирующих цепи или связывающие фрагменты FXIa-связывающего антитела, можно получать с использованием экспрессионных векторов по изобретению, которые содержат вирусную точку начала репликации или эндогенные экспрессионные элементы и ген селективного маркера. После введения вектора клетки можно оставлять расти в течение 1-2 дней в обогащенной среде перед тем, как перенести их в селективную среду. Селективный маркер должен подтверждать устойчивость к селекции, и его присутствие позволяет расти в селективной среде клеткам, которые успешно экспрессируют введенные последовательности. Устойчивые, стабильно трансфицированные клетки можно размножать с использованием методов культивирования, подходящих для конкретного типа клеток.
Конструирование каркаса или Fc-области
Сконструированные антитела по изобретению включают те, каркасные остатки в VH и/или VL которых были подвергнуты модификациям, например, для улучшения свойств антитела. Как правило, такие модификации каркаса производят для уменьшения иммуногенности антитела. Например, одним из подходов является «обратная мутация» одного или более каркасных остатков в соответствующую последовательность зародышевой линии. Более конкретно, антитело, которое подверглось соматической мутации, может содержать каркасные остатки, отличающиеся от остатков в последовательности зародышевой линии, из которой получено антитело. Такие остатки можно идентифицировать путем сравнения каркасных последовательностей антитела с последовательностями зародышевой линии, из которых было получено антитело. Для возвращения последовательностей каркасной области к их конфигурации зародышевой линии можно проводить «обратную мутацию» соматических мутаций в последовательность зародышевой линии, например, с помощью сайт-направленного мутагенеза. Такие «обратно мутировавшие» антитела также входят в объем изобретения.
Другой вид модификаций каркаса включает осуществление мутации одного или более остатков в каркасной области, или даже в одной или более областях CDR, для удаления T-клеточных эпитопов с целью уменьшения потенциальной иммуногенности антитела. Такой подход также называют «деиммунизацией», и он описан более подробно в публикации патента США № 20030153043 авторов Carr et al.
Дополнительно или альтернативно модификациям, производимым в областях каркаса и CDR, антитела по изобретению могут быть сконструированы для включения модификаций в Fc-области, как правило, для изменения одного или более функциональных свойств антитела, таких как период полувыведения в сыворотке, фиксация комплемента, связывание Fc-рецептора и/или антигензависимая клеточная цитотоксичность. Кроме того, антитело по изобретению может быть химически модифицировано (например, один или более химических фрагментов могут быть присоединены к антителу) или быть модифицировано для изменения его гликозилирования, опять-таки для изменения одного или более функциональных свойств антитела. Каждый из таких вариантов осуществления описан более подробно ниже. Нумерация остатков в Fc-области соответствует EU-индексу Kabat.
В одном варианте осуществления шарнирная область CH1 модифицирована таким образом, что число остатков цистеина в шарнирной области изменено, например, увеличено или уменьшено. Этот подход дополнительно описан в патенте США № 5677425 авторов Bodmer et al. Число остатков цистеина в шарнирной области CH1 изменено, например, для облегчения сборки легкой и тяжелой цепей, либо для увеличения или уменьшения стабильности антитела.
В другом варианте осуществления шарнирная область Fc антитела подвергнута мутации для уменьшения биологического периода полувыведения антитела. Более конкретно, одну или более аминокислотных мутаций вносят в граничную область доменов CH2-CH3 Fc-шарнирного фрагмента так, что у антитела нарушается связывание с белком A стафилококков (SpA) по сравнению со связыванием SpA нативным Fc-шарнирным доменом. Такой подход описан более подробно в патенте США № 6165745 авторов Ward et al.
В другом варианте осуществления антитело модифицировано для увеличения его биологического периода полувыведения. Можно использовать разные подходы. Например, можно использовать одну или более из мутаций, описанных в патенте США № 6277375 автора Ward. Альтернативно, для увеличения биологического периода полувыведения антитело можно изменять в области CH1 или CL для содержания эпитопа связывания рецептора реутилизации, взятого из двух петель домена CH2 Fc-области IgG, как описано в патентах США №№ 5869046 и 6121022 авторов Presta et al.
В других вариантах осуществления Fc-область модифицируют путем замены по меньшей мере одного аминокислотного остатка другим аминокислотным остатком для изменения эффекторных функций антитела. Например, одну или более аминокислот можно заменять другим аминокислотным остатком так, что антитело имеет измененную аффинность связывания для эффекторного лиганда, но сохраняет способность исходного антитела к связыванию антигена. Эффекторный лиганд, аффинность к которому изменяют, может представлять собой, например, Fc-рецептор или компонент C1 комплемента. Такой подход описан более подробно в патентах США №№ 5624821 и 5648260 оба из которых выданы Winter et al.
В другом варианте осуществления одну или более аминокислот, выбранных из аминокислотных остатков, можно заменять другим аминокислотным остатком так, что у антитела изменяется способность к связыванию C1q и/или уменьшается или исчезает комплементзависимая цитотоксичность (CDC). Такой подход описан более подробно в патенте США № 6194551 авторов Idusogie et al.
В другом варианте осуществления один или более аминокислотных остатков изменяют для изменения способности антитела к фиксации комплемента. Такой подход описан более подробно в PCT публикации WO 94/29351 авторов Bodmer et al.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, содержащее VL CDR и VH CDR из NOV1401), содержит Fc-область человеческого IgG (например, IgG1), имеющую две аминокислотные замены, D265A и P329A, для снижения вероятности ADCC или CDC, вызываемых любым связанным с поверхностью FXI. Показано, что такие замены на остатки аланина приводят к снижению ADCC и CDC (смотри, например, Idosugie et al., J. Immunol. 164: 4178-4184, 2000; Shields et al., J. Biol. Chem. 276: 6591-6604, 2001).
В другом варианте осуществления Fc-область модифицируют для увеличения способности антитела опосредовать антителозависимую клеточную цитотоксичность (ADCC) и/или для увеличения аффинности антитела к Fcγ-рецептору путем изменения одной или более аминокислот. Такой подход описан более подробно в PCT публикации WO 00/42072 автора Presta. Кроме того, были картированы сайты связывания для FcγRI, FcγRII, FcγRIII и FcRn на IgG1 человека и описаны варианты с улучшенными параметрами связывания (смотри Shields, R.L. et al., 2001 J. Biol. Chen. 276: 6591-6604).
В другом варианте осуществления модифицируют гликозилирование антитела. Например, можно получать агликозилированное антитело (то есть, антитело без гликозилирования). Гликозилирование можно изменять, например, для увеличения аффинности антитела к «антигену». Такие углеводные модификации можно осуществлять, например, путем изменения одного или более сайтов гликозилирования в последовательности антитела. Например, можно производить замены одной или более аминокислот, что приводит к элиминации одного или более сайтов гликозилирования в каркасе вариабельной области, для устранения, таким образом, гликозилирования в этом сайте. Такое aгликозилирование может приводить к увеличению аффинности антитела в отношении антигена. Такой подход описан более подробно в патентах США №№ 5714350 и 6350861, выданных Co et al.
Дополнительно или альтернативно, можно получать антитело с измененным типом гликозилирования, например, гипофукозилированное антитело, содержащее меньшее количество фукозильных остатков, или антитело, содержащее больше структур бисектного GlcNac. Показано, что такие измененные схемы гликозилирования увеличивают ADCC активность антител. Такие углеводные модификации можно осуществлять, например, за счет экспрессии антитела в клетке-хозяине с измененным аппаратом гликозилирования. Клетки с измененным аппаратом гликозилирования описаны в данной области и могут быть использованы в качестве клеток-хозяев для экспрессии рекомбинантных антител по изобретению с целью получения антител с измененной картиной гликозилирования. Например, в патенте EP 1176195 авторов Hang et al. описана линия клеток с функциональным нарушением гена FUT8, кодирующего фукозилтрансферазу, в результате чего антитела, экспрессируемые в клетках такой линии, отличаются гипофукозилированием. В PCT публикации WO 03/035835 автора Presta описана вариантная линия клеток CHO, клетки Lecl3, с пониженной способностью присоединять фукозу к Asn(297)-связанным углеводам, что также приводит к гипофукозилированию антител, экспрессируемых в такой клетке-хозяине (смотри также Shields, R.L. et al., 2002 J. Biol. Chem. 277: 26733-26740). В PCT публикации WO 99/54342 авторов Umana et al. описаны линии клеток, сконструированных для экспрессии модифицирующих гликопротеины гликозилтрансфераз (например, бета(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII)), в результате чего антитела, экспрессируемые в клетках этой линии, демонстрируют наличие увеличенного количества структур бисектного GlcNac, что приводит к увеличению ADCC активности антител (смотри также Umana et al., 1999 Nat. Biotech. 17: 176-180).
Способы конструирования измененных антител
Как описано выше, FXIa-связывающие антитела, имеющие последовательности VH и VL или полноразмерные последовательности тяжелых и легких цепей, описанные в настоящем документе, можно использовать для создания новых FXIa-связывающих антител путем изменения полноразмерных последовательностей тяжелой цепи и/или легкой цепи, последовательностей VH и/или VL, или последовательностей присоединенной к ним константной области(ей). Таким образом, в другом аспекте изобретения структурные особенности FXIa-связывающего антитела по изобретению используют для создания структурно родственных FXIa-связывающих антител, сохраняющих по меньшей мере одно функциональное свойство антител по изобретению, такое как связывание человеческого FXIa, а также ингибирование одного или более функциональных свойств FXIa (например, ингибирование связывания FXIa с рецептором FXIa, ингибирование FXIa-зависимой клеточной пролиферации).
Например, одну или более областей CDR антител по настоящему изобретению, или их мутантные формы, можно комбинировать рекомбинантными методами с известными каркасными областями и/или другими областями CDR для создания дополнительных, сконструированных рекомбинантными методами, FXIa-связывающих антител по изобретению, как описано выше. Другие виды модификаций включают те, которые описаны в предыдущем разделе. Исходным материалом для конструирования является одна или более из последовательностей VH и/или VL, приведенных в настоящем документе, или одна или более из их областей CDR. Для создания сконструированного антитела нет необходимости фактически получать (то есть, экспрессировать в виде белка) антитело, имеющее одну или более из последовательностей VH и/или VL, приведенных в настоящем документе, или одну или более из их областей CDR. Скорее, информацию, содержащуюся в последовательности(ях), используют в качестве исходного материала для создания последовательности(ей) «второго поколения», производных от оригинальной последовательности(ей), и затем последовательность(и) «второго поколения» получают и экспрессируют в виде белка.
Соответственно, в другом варианте осуществления изобретение относится к способу получения FXIa-связывающего антитела, состоящего из последовательности вариабельной области тяжелой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NOs: 3 и 23, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NOs: 4 и 24, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NOs: 5 и 25; и последовательности вариабельной области легкой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NOs: 13 и 33, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NOs: 14 и 34, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NOs: 15 и 35; изменения по меньшей мере одного аминокислотного остатка в последовательности вариабельной области тяжелой цепи антитела и/или последовательности вариабельной области легкой цепи антитела для создания по меньшей мере одной измененной последовательности антитела; и экспрессии измененной последовательности антитела в виде белка.
Соответственно, в другом варианте осуществления изобретение относится к способу получения FXIa-связывающего антитела, состоящего из последовательности вариабельной области тяжелой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NOs: 6 и 26, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NOs: 7 и 27, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NOs: 8 и 28; и последовательности вариабельной области легкой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NOs: 16 и 36, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NOs: 17 и 37, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NOs: 18 и 38; изменения по меньшей мере одного аминокислотного остатка в последовательности вариабельной области тяжелой цепи антитела и/или последовательности вариабельной области легкой цепи антитела для создания по меньшей мере одной измененной последовательности антитела; и экспрессии измененной последовательности антитела в виде белка.
Соответственно, в другом варианте осуществления изобретение относится к способу получения FXIa-связывающего антитела, оптимизированного для экспрессии в клетке млекопитающего, состоящего из: полноразмерной последовательности тяжелой цепи антитела, имеющей последовательность, выбранную из группы, состоящей из SEQ ID NOs: 11 или 31; и полноразмерной последовательности легкой цепи антитела, имеющей последовательность, выбранную из группы, состоящей из SEQ ID NOs: 21 или 41; изменения по меньшей мере одного аминокислотного остатка в полноразмерной последовательности тяжелой цепи антитела и/или полноразмерной последовательности легкой цепи антитела для создания по меньшей мере одной измененной последовательности антитела; и экспрессии измененной последовательности антитела в виде белка. В одном варианте осуществления изменение в тяжелой или легкой цепи имеет место в каркасной области тяжелой или легкой цепи.
Измененную последовательность антитела также можно получать путем скрининга библиотек антител, имеющих фиксированные последовательности CDR3 или минимальные ключевые связывающие детерминанты, как описано в US2005/0255552, и различные последовательности CDR1 и CDR2. Скрининг можно выполнять любым методом, подходящим для скрининга антител из библиотек антител, например, с использованием технологии фагового дисплея.
Можно использовать стандартные методы молекулярной биологии для получения и экспрессии измененной последовательности антитела. Антитело, кодируемое измененной последовательностью(ми) антитела, представляет собой антитело, которое сохраняет одно, несколько или все из функциональных свойств FXIa-связывающих антител, описанных в настоящем документе, при этом функциональные свойства включают, но не ограничиваются ими, специфическое связывание с FXIa человека, яванского макака, крысы и/или мыши; и ингибирование антителом FXIa-зависимой клеточной пролиферации в анализе клеточной пролиферации с использованием F36E и/или Ba/F3-FXIaR.
В конкретных вариантах осуществления способов конструирования антител по изобретению мутации можно вносить случайным или избирательным образом на протяжении всей или части кодирующей последовательности FXIa-связывающего антитела, и можно проводить скрининг полученных модифицированных FXIa-связывающих антител на связывающую активность и/или другие функциональные свойства, как описано в настоящем документе. Методы осуществления мутаций описаны в данной области. Например, в PCT публикации WO 02/092780 автора Short описаны способы создания и скрининга мутантных антител с использованием насыщающего мутагенеза, синтетической сборки лигированием или их сочетания. Альтернативно, в PCT публикации WO 03/074679 авторов Lazar et al. описано использование компьютерных методов скрининга для оптимизации физико-химических свойств антител.
В конкретных вариантах осуществления изобретения антитела были сконструированы для удаления сайтов дезамидирования. Известно, что дезамидирование вызывает структурные и функциональные изменения в пептиде или белке. Дезамидирование может приводить к снижению биологической активности, а также к изменениям фармакокинетики и антигенности белкового фармацевтического средства (Anal Chem. 2005 Mar 1; 77(5): 1432-9).
В конкретных вариантах осуществления изобретения антитела были сконструированы для увеличения значения pI и улучшения их свойств в качестве лекарственных средств. Значение pI белка является ключевым фактором, определяющим общие биофизические свойства молекулы. Известно, что антитела, имеющие низкие значения pI, менее растворимы, менее стабильны и имеют тенденцию к агрегации. Кроме того, очистка антител с низким значением pI является сложной и может быть проблематичной, особенно при крупномасштабном производстве для клинического применения. Увеличение значения pI анти-FXI/FXIa антител, или Fab-фрагментов, по изобретению приводит к улучшению их растворимости, позволяя формулировать антитела в более высоких концентрациях (>100 мг/мл). Формулирование антител в высоких концентрациях (например, >100 мг/мл) имеет то преимущество, что позволяет вводить более высокие дозы антител, что, в свою очередь, позволяет снижать частоту дозирования, это важное преимущество при лечении хронических заболеваний, включая тромботические и/или тромбоэмболические заболевания. Более высокие значения pI также могут способствовать увеличению FcRn-опосредованного рециклинга варианта IgG антитела, таким образом, позволяя лекарственному средству оставаться в организме в течение более длительного времени, как следствие, потребуется меньше инъекций. И наконец, общая стабильность антител значительно повышается благодаря высоким значениям pI, что приводит к увеличению срока хранения и биологической активности in vivo. Предпочтительно, значение pI превышает или равно 8,2.
Функциональные свойства измененных антител можно оценивать с использованием стандартных анализов, доступных в данной области и/или описанных в настоящем документе, таких как те, которые приведены в разделе «Примеры» (например, ELISA).
Профилактическое и терапевтическое применение
Антитела, связывающие FXI и/или FXIa, которые описаны в настоящем документе (например, антитела, приведенные в таблице 1, такие как анти-FXI/FXIa антитела, содержащие VL CDR и VH CDR из NOV1401), могут быть использованы в терапевтически полезной концентрации для лечения тромбоэмболического заболевания или нарушения (например, тромбозного инсульта, фибрилляции предсердий, предотвращения инсульта при фибрилляции предсердий (ПИФП), тромбоза глубоких вен, венозной тромбоэмболии, легочной эмболии, острых коронарных синдромов (ОКС), ишемического инсульта, острой ишемии конечностей, хронической тромбоэмболической легочной гипертензии или системной эмболии) путем введения субъекту, который нуждается в этом, эффективного количества антител, или антигенсвязывающих фрагментов, по изобретению. Настоящее изобретение относится к способу лечения тромбоэмболического заболевания (например, тромботических заболеваний) путем введения субъекту, который нуждается в этом, эффективного количества антител по изобретению. Настоящее изобретение относится к способу лечения тромбоэмболических заболеваний (например, тромбозного инсульта, фибрилляции предсердий, предотвращения инсульта при фибрилляции предсердий (ПИФП), тромбоза глубоких вен, венозной тромбоэмболии, легочной эмболии, острых коронарных синдромов (ОКС), ишемического инсульта, острой ишемии конечностей, хронической тромбоэмболической легочной гипертензии или системной эмболии) путем введения субъекту, который нуждается в этом, эффективного количества антител по изобретению.
Антитела, описанные в настоящем документе (например, антитела, приведенные в таблице 1, такие как NOV1401, или анти-FXI/FXIa антитела, содержащие VL CDR и VH CDR из NOV1401), могут быть использованы, помимо прочего, для предотвращения, лечения или ослабления тромбоэмболических состояний или заболеваний, включая, но без ограничения, тромботические заболевания, описанные более подробно в настоящем документе.
Антитела, предложенные в настоящем документе (например, антитела, приведенные в таблице 1, такие как анти-FXI/FXIa антитела, содержащие VL CDR и VH CDR из NOV1401), также можно использовать в сочетании с другими средствами для предотвращения, лечения или ослабления тромбоэмболических заболеваний. Например, терапию статинами можно использовать в сочетании с анти-FXIa антителами и антигенсвязывающими фрагментами по изобретению для лечения пациентов с тромботическими и/или тромбоэмболическими заболеваниями.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения или предотвращения инсульта у пациента с фибрилляцией предсердий, включающий введение пациенту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ контроля или предотвращения рисков или состояний, связанных с фибрилляцией предсердий (ФП), таких как эмболический инсульт и системная эмболия, у пациента с фибрилляцией предсердий, включающий введение пациенту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения состояний, связанных с фибрилляцией предсердий (ФП), таких как эмболический инсульт и системная эмболия, у пациента с фибрилляцией предсердий, включающий введение пациенту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401. В конкретных вариантах осуществления, пациент с ФП имеет высокий риск кровотечений.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения тромбоза глубоких вен или связанных с ним состояний у субъекта (например, субъекта, страдающего тромбозом глубоких вен или имеющего риск его развития), включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения венозной тромбоэмболии (ВТЭ) или связанных с ней состояний у субъекта (например, субъекта, страдающего венозной тромбоэмболией или имеющего риск ее развития), включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401. В конкретных вариантах осуществления субъекты, получающие лечение анти-FXI/FXIa антителом, предложенным в настоящем документе, испытали 1) первичную неспровоцированную ВТЭ с низким риском кровотечения, 2) рецидив неспровоцированной ВТЭ или 3) ВТЭ, ассоциированную с тромбофилией, включая онкологических пациентов.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения легочной эмболии или связанных с ней состояний у субъекта (например, субъекта, страдающего легочной эмболией или имеющего риск ее развития), включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения острых коронарных синдромов (ОКС) или связанных с ними состояний у субъекта, включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения ишемического инсульта у субъекта (например, субъекта, страдающего ишемическим инсультом или имеющего риск его развития), включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения острой ишемии конечностей у субъекта, включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения хронической тромбоэмболической легочной гипертензии у субъекта, включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения системной эмболии у субъекта (например, субъекта, страдающего системной эмболией или имеющего риск ее развития), включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретном варианте осуществления настоящего изобретения предложен способ лечения, контроля или предотвращения тромбоэмболических состояний, связанных с введением катетера (например, катетера Хикмана у онкологических пациентов), при которых катетеры забиваются тромбами, и экстракорпоральной мембранной оксигенацией (ЭКМО), при которой в трубках образуются сгустки, включающий введение субъекту, который нуждается в этом, эффективного количества анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401.
В конкретных вариантах осуществления субъекты, которые нуждаются в лечении анти-FXI/FXIa антителом, описанным в настоящем документе, например, анти-FXI/FXIa антителом, приведенным в таблице 1, таким как NOV1401, или анти-FXI/FXIa антителами, содержащими VL CDR и VH CDR из NOV1401, могут включать:
- субъектов с показаниями для хронической антикоагулянтной терапии (например, в случае ФП, тромба в левом желудочке, ранее перенесенного кардиоэмболического инсульта)
- субъектов со средним или высоким риском обширного кровотечения;
- субъектов, подвергаемых элективному или первичному чрескожному коронарному вмешательству (ЧКВ) с установлением стентов, которым может потребоваться двойная антитромбоцитарная терапия (аспирин и антагонисты рецептора P2Y12) для предотвращения тромбоза стентов.
В конкретных вариантах осуществления одно из следующих состояний можно лечить или контролировать при помощи анти-FXI/FXIa антитела, описанного в настоящем документе, например, анти-FXI/FXIa антитела, приведенного в таблице 1, такого как NOV1401, или анти-FXI/FXIa антител, содержащих VL CDR и VH CDR из NOV1401:
- тромбоэмболия у субъектов с предполагаемой или подтвержденной сердечной аритмией, такой как пароксизмальная, персистентная или перманентная фибрилляция предсердий или трепетание предсердий;
- предотвращение инсульта при фибрилляции предсердий (ПИФП), одной из подгрупп пациентов являются пациенты с ФП, подвергаемые чрескожному коронарному вмешательству (ЧКВ);
- лечение острых венозных тромбоэмболических событий (ВТС) и расширенная вторичная профилактика ВТС у пациентов с высоким риском кровотечений;
- церебральные и сердечно-сосудистые события во вторичной профилактике после транзиторной ишемической атаки (ТИА) или не инвалидизирующего инсульта и предотвращение тромбоэмболических событий при сердечной недостаточности с синусовым ритмом;
- образование сгустка в левом предсердии и тромбоэмболия у субъектов, подвергаемых кардиоверсии в связи с сердечной аритмией;
- тромбоз до, в процессе и после процедуры абляции в связи с сердечной аритмией;
- венозный тромбоз, сюда относятся, но без ограничения, лечение и вторичная профилактика тромбоза глубоких и поверхностных вен в нижних конечностях или верхних конечностях, тромбоз в брюшных и грудных венах, синус-тромбоз и тромбоз яремных вен;
- тромбоз на любой искусственной поверхности в венах, такой как катетер или провода кардиостимулятора;
- легочная эмболия у пациентов с венозным тромбозом или без него;
- хроническая тромбоэмболическая легочная гипертензия (ХТЭЛГ);
- артериальный тромбоз на месте оторвавшейся атеросклеротической бляшки, тромбоз на внутриартериальном протезе или катетере и тромбоз в кажущихся нормальными артериях, включая, но без ограничения, острые коронарные синдромы, инфаркт миокарда с повышением ST-сегмента, инфаркт миокарда без повышения ST-сегмента, нестабильную стенокардию, тромбоз стентов, тромбоз любой искусственной поверхности в системе артериального кровообращения и тромбоз легочных артерий у субъектов с легочной гипертензией и без нее;
- тромбоз и тромбоэмболия у пациентов, подвергаемых чрескожному коронарному вмешательству (ЧКВ);
- кардиоэмболические и криптогенные инсульты;
- тромбоз у пациентов с инвазивными и неинвазивными злокачественными новообразованиями;
- тромбоз в полостном катетере;
- тромбоз и тромбоэмболия у пациентов в тяжелом состоянии;
- сердечный тромбоз и тромбоэмболия, включая, но без ограничения, сердечный тромбоз после инфаркта миокарда, сердечный тромбоз, связанный с таким состоянием, как сердечная аневризма, миокардиальный фиброз, увеличение сердца и сердечная недостаточность, миокардит и искусственные поверхности в сердце;
- тромбоэмболия у пациентов с болезнью клапанов сердца, с фибрилляцией или без фибрилляции предсердий;
- тромбоэмболия в механических или биологических протезах клапанов;
- тромбоэмболия у пациентов, имеющих естественные или искусственные заплаты в сердце, артериальные или венозные проводящие трубки после операции на сердце по поводу простых или сложных пороков сердца;
- венозный тромбоз и тромбоэмболия после операции по замене коленного сустава, операции по замене тазобедренного сустава, а также ортопедической операции, торакальной или абдоминальной хирургической операции;
- артериальный или венозный тромбоз после нейрохирургической операции, включая интракраниальные операции и операции на спинном мозге;
- врожденная или приобретенная тромбофилия, включая, но без ограничения, мутацию фактора V Лейдена, мутацию протромбина, дефицит антитромбина III, белка C и белка S, мутацию фактора XIII, семейную дисфибриногенемию, врожденный дефицит плазминогена, повышенные уровни фактора XI, серповидно-клеточное заболевание, антифосфолипидный синдром, аутоиммунное заболевание, хроническое заболевание кишечника, нефротический синдром, гемолитическую уремию, миелопролиферативное заболевание, диссеминированное внутрисосудистое свертывание крови, пароксизмальную ночную гемоглобинурию и вызванную гепарином тромбопению;
- тромбоз и тромбоэмболия при хронической почечной недостаточности; и
- тромбоз и тромбоэмболия у пациентов, подвергаемых гемодиализу, и у пациентов, подвергаемых экстракорпоральной мембранной оксигенации.
В конкретном аспекте настоящего изобретения предложены способы контроля кровотечения у пациента, который получает, или которому вводят, анти-FXI/FXIa антитело, предложенное в настоящем документе (например, антитело, приведенное в таблице 1, такое как анти-FXI/FXIa антитело, содержащее VL CDR и VH CDR из NOV1401), например, кровотечения, связанного с травмой, хирургической операцией, менструацией или родами, включающие обращение вспять антикоагулянтного эффекта. Дефицит FXI редко связан со спонтанными эпизодами кровотечения; в конкретных аспектах кровотечение чаще всего связано с травмой, хирургической операцией, менструацией или родами. Продолжительное кровотечение может иметь место после обширной травмы или после хирургической операции на органах с тканями, имеющими повышенную фибринолитическую активность, такими как слизистая оболочка ротовой полости, слизистая оболочка носовой полости, слизистая оболочка половых или мочевыводящих путей. Удаление зубов, тонзилэктомия, а также абляция матки или предстательной железы, являются примерами хирургических операций, которые влекут за собой высокий риск кровотечений. Люди с данным заболеванием также имеют сильную тенденцию к носовым кровотечениям и подкожным кровоизлияниям, и, реже, маточному или кишечному кровотечению. Частота спонтанных кровотечений в мышцах или суставах и интракраниального кровотечения не увеличена у пациентов с дефицитом FXI. Прокол вен, как правило, не связан с продолжительным кровотечением. Другие генетические мутации, связанные с дефицитом FXI, могут вносить вклад в склонность к гетерогенным и непредсказуемым кровотечениям у пациентов с тяжелым дефицитом FXI. Одновременный прием антитромбоцитарных препаратов, других антикоагулянтов и фибринолитических средств может увеличивать риск кровотечений.
В конкретных вариантах осуществления настоящего изобретения предложен способ контроля кровотечений у пациента, получающего лечение анти-FXI/FXIa антителом, предложенным в настоящем документе (например, антителом, приведенным в таблице 1, таким как анти-FXI/FXIa антитело, содержащее VL CDR и VH CDR из NOV1401), включающий временное обращение вспять антикоагулянтного эффекта на время, достаточное для прекращения кровотечения. В конкретных вариантах осуществления этап обращения вспять антикоагулянтного эффекта включает (i) инфузионную терапию с использованием коллоидов, кристаллоидов, человеческой плазмы или белков плазмы, таких как альбумин; или (ii) переливание эритроцитарной массы или цельной крови. В конкретном варианте осуществления терапевтические средства для обращения вспять эффекта антикоагулянтов, например, в чрезвычайной ситуации, включают, но не ограничиваются ими, прогемостатические компоненты крови, такие как свежезамороженная плазма (СЗП), концентраты протромбинового комплекса (КПК) и активированные КПК [(АКПК); например, ингибитор обходной активности фактора VIII (ИОАФВ)], а также рекомбинантный активированный фактор VII (rFVIIa). В одном варианте осуществления схема, включающая введение rFVIIa в дозе 30 мкг/кг, с последующим введением rFVIIa в дозе 15-30 мкг/кг каждые 2-4 часа в течение 24-48 часов в дополнение к приему 1 г транексамовой кислоты каждые 6 часов в течение 5-7 дней, потенциально может восстанавливать гемостаз и останавливать кровотечение у субъектов, получающих лечение анти-FXI/FXIa антителом, предложенным в настоящем документе (например, NOV1401 или антителом, содержащим VL CDR и VH CDR из NOV1401), подвергаемых серьезной операции, и у пациентов с недоступной зоной активного кровотечения. Например, Riddell et al. сообщали о 4 пациентах с тяжелым дефицитом FXI без ингибитора, подвергаемых хирургической операции (Riddell et al., 2011, Thromb. Haemost.; 106: 521-527); пациентам вводили rFVIIa в дозе 30 мкг/кг и 1 г транексамовой кислоты в/в при индукции анестезии. Последующие болюсные дозы 15-30 мкг/кг rFVIIa вводили с интервалами в 2-4 часа, руководствуясь результатами ротационной тромбоэластометрии (ROTEM). Пациенты получали rFVIIa в вышеуказанных дозах в течение 24-48 часов. Введение 1 г транексамовой кислоты каждые шесть часов продолжали в течение пяти дней. В этой небольшой группе пациентов введение rFVIIa в дозах всего лишь 15-30 мкг/кг в сочетании с транексамовой кислотой было безопасным и эффективным для коррекции дефекта гемостаза у пациентов с тяжелым дефицитом FXI в данном исследовании. В другом исследовании с участием 4 пациентов с тяжелым дефицитом FXI с ингибитором (аутологичные нейтрализующие анти-FXI антитела, как правило, приобретенные после переливания или введения препаратов крови пациентам с тяжелым дефицитом FXI), перенесших 5 операций, авторы (Livnat et al., 2009, Thromb. Haemost.; 102: 487-492) использовали следующий протокол: 1 г транексамовой кислоты перорально за два часа до операции, затем пациенты получали непосредственно перед хирургическим вмешательством еще 1 г транексамовой кислоты в/в. Рекомбинантный FVIIa в диапазоне доз от 15 до 30 мкг/кг вводили инфузией при завершении операции. Затем пациенты принимали перорально 1 г транексамовой кислоты каждые 6 часов в течение по меньшей мере 7 дней. Фибриновый клей наносили распылением на ложе удаленного желчного пузыря у одного пациента. Этот протокол обеспечивал нормальный гемостаз у пациентов с тяжелым дефицитом FXI с ингибитором.
В одном аспекте фибриновый клей можно использовать для восстановления локального гемостаза во время стоматологической операции у пациентов с дефицитом FXI (Bolton-Maggs (2000) Haemophilia; 6 (S1): 100-9). В конкретном варианте осуществления, относящемся к способам контроля кровотечения у пациентов, получающих лечение анти-FXI/FXIa антителом, предложенным в настоящем документе (например, NOV1401), схема, включающая прием 1 г транексамовой кислоты каждые 6 часов в течение 5-7 дней в сочетании с использованием фибринового клея, может быть использована для создания локального гемостаза у субъектов, подвергаемых небольшой хирургической операции, и у субъектов с доступной зоной кровотечения, включая кровотечения в ротовой и носовой полости.
Фармацевтические композиции
Изобретение относится к фармацевтическим композициям, содержащим FXIa-связывающие антитела (интактные антитела или связывающие фрагменты), сформулированные с фармацевтически приемлемым носителем. Кроме того, композиции могут содержать одно или более других терапевтических средств, которые подходят для лечения или предотвращения, например, тромбоэмболических заболеваний (например, тромботических заболеваний). Фармацевтически приемлемые носители усиливают действие или стабилизируют композицию, или могут быть использованы для облегчения приготовления композиции. Фармацевтически приемлемые носители включают растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие абсорбцию средства, и тому подобное, которые являются физиологически совместимыми.
Фармацевтическую композицию по настоящему изобретению можно вводить разными способами, известными в данной области. Путь и/или способ введения варьируется в зависимости от нужных результатов. Предпочтительным является внутривенное (в/в), внутримышечное (в/м), внутрибрюшинное (в/б) или подкожное (п/к) введение, либо введение вблизи от целевой области. Фармацевтически приемлемый носитель должен быть подходящим для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от пути введения активное соединение, то есть, антитело, биспецифическая и мультиспецифическая молекула, может быть покрыто материалом для защиты соединения от действия кислот и других природных условий, которые могут инактивировать соединение.
В конкретных аспектах анти-FXI/FXIa антитела, описанные в настоящем документе (например, антитела, приведенные в таблице 1, такие как NOV1401, или антитела, содержащие области LCDR и HCDR из NOV1401), сформулированы в концентрации от примерно 75 мг/1 мл до примерно 200 мг/1 мл, во флаконах с жидкостью для подкожных инъекций. В конкретных вариантах осуществления фармацевтическая композиция содержит фармацевтический носитель или эксципиент, например, сахарозу и полисорбат 20. В конкретных вариантах осуществления фармацевтическая композиция содержит L-гистидин и/или гистидин-HCl моногидрат. В конкретных вариантах осуществления фармацевтическая композиция имеет значение pH примерно 4-7 или 5-6.
В конкретных аспектах анти-FXI/FXIa антитела, описанные в настоящем документе (например, антитела, приведенные в таблице 1, такие как NOV1401, или антитела, содержащие области LCDR и HCDR из NOV1401), сформулированы в концентрации 150 мг/1 мл во флаконах с жидкостью для подкожных инъекций. В одном варианте осуществления жидкий препарат с концентрацией 150 мг/мл содержит 150 мг анти-FXI/FXIa антитела, L-гистидин, гистидин-HCl моногидрат, сахарозу и полисорбат 20, и имеет значение pH=5,5 ± 0,5. Композиция должна быть стерильной и жидкой. Надлежащую текучесть можно поддерживать, например, за счет использования такого покрытия, как лецитин, за счет поддержания необходимого размера частиц в случае дисперсии и за счет использования сурфактантов. Во многих случаях в композицию предпочтительно включать изотонические средства, например, сахара, полиспирты, такие как маннит или сорбит, и хлорид натрия. Долгосрочной абсорбции инъекционных композиций можно добиваться за счет включения в композицию средства, замедляющего абсорбцию, например, алюминия моностеарата или желатина.
Фармацевтические композиции по изобретению можно получать методами, хорошо известными и обычно практикуемыми в данной области. Смотри, например, Remington: The Science and Practice of Pharmacy, Mack Publishing Co., 20-е издание, 2000; и Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978. Фармацевтические композиции предпочтительно производят в соответствии с требованиями GMP. Как правило, в фармацевтических композициях по изобретению используют терапевтически эффективную дозу или эффективную дозу FXIa-связывающего антитела. FXIa-связывающие антитела формулируют в фармацевтически приемлемых лекарственных формах общепринятыми методами, известными специалистам в данной области. Схемы дозирования корректируют для обеспечения оптимального желательного ответа (например, терапевтического ответа). Например, можно вводить однократный болюс, можно вводить несколько разделенных доз на протяжении некоторого времени, или дозу можно пропорционально уменьшать или увеличивать в зависимости от терапевтической ситуации. Особенно предпочтительно формулировать парентеральные композиции в стандартной лекарственной форме для легкости введения и единообразия доз. Используемый в настоящем документе термин «стандартная лекарственная форма» означает физически дискретные единицы, подходящие в качестве однократных доз для получающих лечение субъектов; каждая единица содержит заранее определенное количество активного соединения, рассчитанное для достижения желаемого терапевтического эффекта, в сочетании с необходимым фармацевтическим носителем.
Фактические уровни доз активных ингредиентов в фармацевтических композициях по настоящему изобретению можно варьировать, чтобы добиться количества активного ингредиента, которое эффективно для достижения желательного терапевтического ответа для конкретного пациента, композиции и способа введения, без токсичности для пациента. Выбранный уровень доз зависит от различных фармакокинетических факторов, включая активность конкретных используемых композиций по настоящему изобретению, пути их введения, время их введения, скорость выведения конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, используемые в сочетании с конкретными используемыми композициями, возраст, пол, массу тела, патологическое состояние, общее состояние здоровья и предшествующую медицинскую историю пациента, получающего лечение, и тому подобные факторы.
Врач может начинать вводить дозы антител по изобретению, используемых в фармацевтической композиции, на более низких уровнях, чем необходимо для достижения желательного терапевтического эффекта, и постепенно увеличивать дозы до достижения желательного эффекта. Как правило, эффективные дозы композиций по настоящему изобретению для лечения тромботических и/или тромбоэмболических заболеваний, описанных в настоящем документе, варьируются в зависимости от множества разных факторов, включая способы введения, целевую зону, физиологическое состояние пациента, другие вводимые препараты, а также от того, является ли применение профилактическим или терапевтическим. Лечебные дозы необходимо титровать для оптимизации безопасности и эффективности. Для системного введения антитела дозы находятся в диапазоне примерно от 0,01 до 15 мг/кг массы тела субъекта. Для введения (например, подкожного введения) антитела доза может находиться в диапазоне от 0,1 мг до 5 мг или от 1 мг до 600 мг. Например, анти-FXI/FXIa антитело, описанное в настоящем документе, можно вводить в дозе 0,1 мг/кг, 0,2 мг/кг, 0,3 мг/кг, 0,4 мг/кг, 0,5 мг/кг, 0,6 мг/кг, 0,7 мг/кг, 0,8 мг/кг, 0,9 мг/кг, 1,0 мг/кг, 1,1 мг/кг, 1,2 мг/кг, 1,3 мг/кг, 1,4 мг/кг, 1,5 мг/кг, 1,6 мг/кг, 1,7 мг/кг, 1,8 мг/кг, 1,9 мг/кг, 2,0 мг/кг, 2,1 мг/кг, 2,2 мг/кг, 2,3 мг/кг, 2,4 мг/кг, 2,5 мг/кг, 2,6 мг/кг, 2,7 мг/кг, 2,8 мг/кг, 2,9 мг/кг, 3,0 мг/кг, 3,1 мг/кг, 3,2 мг/кг, 3,3 мг/кг, 3,4 мг/кг, 3,5 мг/кг, 3,6 мг/кг, 3,7 мг/кг, 3,8 мг/кг, 3,9 мг/кг, 4,0 мг/кг, 4,1 мг/кг, 4,2 мг/кг, 4,3 мг/кг, 4,4 мг/кг, 4,5 мг/кг, 4,6 мг/кг, 4,7 мг/кг, 4,8 мг/кг, 4,9 мг/кг или 5,0 мг/кг. Иллюстративная схема лечения включает системное введение один раз каждые две недели или один раз в месяц, или один раз каждые 3-6 месяцев. Иллюстративная схема лечения включает системное введение один раз в неделю, один раз каждые две недели, один раз каждые три недели, один раз в месяц или один раз каждые 3-6 месяцев, или по мере необходимости (PRN).
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе 3 мг/кг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе 10 мг/кг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе 30 мг/кг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе 50 мг/кг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе 100 мг/кг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе, находящейся в диапазоне от 5 мг до 600 мг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе примерно 5 мг, 10 мг, 15 мг, 20 мг, 30 мг, 40 мг, 50 мг, 60 мг, 90 мг, 100 мг, 120 мг, 150 мг, 180 мг, 200 мг, 210 мг, 240 мг, 250 мг, 270 мг, 300 мг, 330 мг, 350 мг, 360 мг, 390 мг, 400 мг, 420 мг, 450 мг, 480 мг, 500 мг, 510 мг, 540 мг, 550 мг, 570 мг или 600 мг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, п/к в дозе 5 мг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, п/к в дозе 15 мг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, п/к в дозе 50 мг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, п/к в дозе 150 мг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, п/к в дозе 300 мг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, п/к в дозе 600 мг.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе, достаточной для достижения пролонгации средней продолжительности аЧТВ в 2 раза или более на период времени не более 30 дней, 35 дней, 36 дней, 37 дней, 38 дней, 39 дней, 40 дней, 41 дня, 42 дней, 43 дней, 44 дней, 45 дней или 50 дней.
В конкретном варианте осуществления анти-FXI/FXIa антитело, описанное в настоящем документе (например, антитело, приведенное в таблице 1, такое как NOV1401, или антитело, содержащее VL CDR и VH CDR из NOV1401), вводят, например, в/в или п/к в дозе, достаточной для достижения пролонгации средней продолжительности аЧТВ в 2 раза или более на период времени не более 42 дней.
Антитело, как правило, вводят несколько раз. Интервалы между отдельными дозами могут составлять неделю, две недели, месяц или год. Интервалы также могут быть нерегулярными, определяемыми результатами измерения уровней связывающего FXI и/или FXIa антитела в крови у пациента. Кроме того, альтернативные интервалы дозирования может определять врач и вводить дозы ежемесячно или по мере необходимости для обеспечения эффективности. В некоторых методах системного введения дозы корректируют для достижения в плазме концентрации антитела 1-1000 мкг/мл или 1-1200 мкг/мл, и в некоторых методах 25-500 мкг/мл. Альтернативно, антитело можно вводить в виде препарата с замедленным высвобождением, в этом случае необходимо менее частое введение. Дозы и частота введения варьируются в зависимости от периода полувыведения антитела и его мишени в организме пациента. Как правило, человеческие и гуманизированные антитела имеют более длительный период полувыведения у человека, чем химерные антитела и антитела, отличные от человеческих. Дозы и частота введения могут варьироваться в зависимости от того, является ли применение профилактическим или терапевтическим. В случаях профилактического применения относительно низкую дозу вводят сравнительно редко на протяжении длительного периода времени. Некоторые пациенты продолжают получать препарат до конца своей жизни. В случаях терапевтического применения иногда необходимо вводить относительно высокую дозу со сравнительно короткими интервалами до тех пор, пока прогрессирование заболевания не замедлится или не прекратится, и предпочтительно до тех пор, пока у пациента не будет наблюдаться частичное или полное уменьшение симптомов заболевания. Затем пациенту можно вводить препарат в профилактическом режиме.
ПРИМЕРЫ
Следующие примеры приведены для дополнительной иллюстрации изобретения, но не для ограничения его объема. Другие варианты осуществления изобретения будут очевидны для специалиста в данной области и входят в объем прилагаемой формулы изобретения.
Пример 1
Пэннинг фаговой библиотеки Fab человека
Для отбора антител, узнающих человеческий фактор XI, использовали несколько стратегий пэннинга. Терапевтические антитела против разных вариантов белка каталитического домена человеческого фактора XI и кроличьего фактора XIa получали путем отбора клонов антител, связывающих фактор XI, с использованием в качестве источника антител коммерчески доступную библиотеку фагового дисплея, библиотеку Morphosys HuCAL PLATINUM®. Фагмидная библиотека основана на концепции HuCAL® (Knappiketal.,2000,JMolBiol296:57-86), и в ней используется технология CysDisplay™ для экспонирования Fab-фрагментов на поверхности фага (WO01/05950). Для выделения антител против фактора XI использовали стратегии жидкофазного пэннинга.
Анализ перекрестной реактивности
Очищенные Fab-фрагменты тестировали в анализе ELISA на связывание с разными вариантами биотинилированных белков каталитических доменов человеческого фактора XI (каталитического домена фактора XI, фактора XIa и фактора XIa) и кроличьего фактора XIa. Для этой цели 384-луночные планшеты Maxisorp™ (Nunc) покрывали нейтравидином в концентрации 10 мкг/мл в PBS в течение ночи при 4°C. Проводили связывание антигенов с нейтравидином через биотин при комнатной температуре (RT) в течение 30 минут. Связывание Fab-фрагментов при разных концентрациях обнаруживали при помощи специфичных для F(ab)2 антител козы против IgG человека, конъюгированных со щелочной фосфатазой (разведение 1:5000), используя флуоресцентный субстрат Attophos (Roche, каталожный № 11681982001). Флуоресцентное излучение при 535 нм регистрировали с возбуждением при 430 нм.
Модификация IgG и экспрессия IgG
Для экспрессии полноразмерного IgG в клетках CAP-T фрагменты вариабельных доменов тяжелой (VH) и легкой (VL) цепей субклонировали из экспрессионных векторов Fab в соответствующие векторы pMorph®_hIg для человеческих IgG1. Осуществляли две аминокислотные замены (D265A и P329A) в Fc-области для уменьшения вероятности ADCC или CDC, вызываемой каким-либо связанным с поверхностью FXI. Показано, что эти замены на остатки аланина уменьшают вероятность ADCC и CDC (смотри, например, Idosugie et al., J. Immunol. 164: 4178-4184, 2000; Shields et al., J. Biol. Chem. 276: 6591-6604, 2001). Супернатант клеточных культур собирали через 7 дней после трансфекции. После стерилизации фильтрованием раствор подвергали аффинной хроматографии с белком A, используя автоматизированную рабочую станцию дозирования жидкостей. Образцы элюировали в 50 нМ цитрате, 140 нМ NaOH, pH доводили до нейтрального значения 1 M трис-буфером и проводили стерилизацию фильтрованием (размер пор 0,2 мкм). Концентрации белка определяли методом УФ-спектрофотометрии при 280 нм и чистоту IgG анализировали методом SDS-ПААГ в денатурирующих восстанавливающих условиях.
Пример 2
Данные связывания
Анализ методом поверхностного плазмонного резонанса (ППР) для каталитического домена FXI.
Измерения методом ППР выполняли с использованием оптического биосенсора на основе системы поверхностного плазмонного резонанса BIACORE™ T200 (BIACORE™, GE Healthcare, Uppsala). Сенсорные чипы серии S (CM5), наборы для иммобилизации и регенерирующий буфер приобретали у компании GE Healthcare (Uppsala). Выполняли два различных варианта анализа в зависимости от формата лиганда, IgG или Fab. Во-первых, поверхность активировали N-гидроксисукцинимидом (NHS) и N-(3-диметиламинопропил)-N-этилкарбодиимид гидрохлоридом (EDC). NOV1401-Fab ковалентно присоединяли к активированной декстрановой матрице на чипе CM5 стандартным методом связывания аминогрупп (GE Healthcare, Uppsala). В случае NOV1401-IgG проводили анализ методом захвата, и антитело козы против IgG-Fc человека (JIR) иммобилизовали на чипе в количестве 14000 ЕО. Оставшиеся активные поверхностные группы инактивировали этаноламином (EA). Готовили эталонную ячейку без иммобилизованного лиганда, и систему уравновешивали 1x буфером HBS-EP+ (10 мМ HEPES, 150 мМ NaCl, 3 мМ ЭДТА, 0,05% P20, pH 7,4, Teknova H8022).
Все эксперименты по связыванию проводили при 25°C со скоростью потока 50 мкл/мин, используя буфер HBS-EP+. Для анализа с захватом проводили захват NOV1401-IgG до достижения уровня ЕО, составляющего 80. Для исследований кинетики использовали серию разведений каталитического домена FXI с диапазоном концентраций 0-200 нМ в буфере HBS-EP+. Время ассоциации составляло 120 с, и время диссоциации составляло 180 с. Поверхность регенерировали однократной инъекцией 10 мМ глицина, pH 1,5 (время контакта 60 с, время стабилизации 120 с). Обработку данных, а также определение kon, koff и KD выполняли с помощью программы T200 BiaEvaluation версии 1.0. Использовали двойной контроль (вычитание эталонной и пустой пробы) для коррекции объемных эффектов и других артефактов системы. Подгонку сенсограмм выполняли с использованием модели связывания 1:1 (Rmax устанавливали на «глобальный»).
Титрование равновесного раствора (ТРР) для FXI и FXIa
22 серийных 1,6-кратных разведения антигенов готовили в буфере для образцов (PBS, pH 7,4, содержащий 0,5% БСА и 0,02% Tween 20), и антитело NOV1401-Fab (200 пМ для huFXI и 500 пМ для huFXIa) или NOV1401 (10 пМ для huFXI и huFXIa) в постоянной концентрации добавляли к антигену в каждой концентрации. Начальные концентрации для серии разведений антигенов составляли 100 нМ в случае huFXIa и 20 нМ в случае huFXI (анализ Fab) или 1 нМ (анализ IgG). По 60 мкл/лунку смеси каждого разведения добавляли в двойном повторе в лунки 384-луночного полипропиленового планшета для микротитрования (MTP). Буфер для образцов служил в качестве отрицательного контроля, и образец, не содержащий антиген, служил в качестве положительного контроля (Bmax). Планшеты герметично закрывали и инкубировали в течение ночи при RT на шейкере. Стандартный 384-луночный MTP (MSD) покрывали по 30 мкл/лунку huFXIa в концентрации 0,1 мкг/мл (для huFXIa и huFXI), разведенным в PBS, герметично закрывали и инкубировали в течение ночи при 4°C.
После инкубации и трехкратного промывания TBST (TBS, содержащий 0,05% Tween 20) покрытый антигеном MSD планшет блокировали путем внесения по 50 мкл/лунку блокирующего буфера (PBS, содержащий 5% БСА) и инкубировали в течение 1 ч при RT на шейкере. Этап промывания повторяли, препарат Fab-/IgG-антиген в количестве 30 мкл/лунку переносили из полипропиленового MTP в покрытый антигеном MSD планшет и инкубировали в течение 20 мин при RT на шейкере. После дополнительного этапа промывания 30 мкл ECL-меченого обнаруживающего антитела козы против IgG/Fab человека (MSD) в концентрации 0,5 мкг/мл, разведенного в буфере для образцов, добавляли в каждую лунку и инкубировали в течение 30 мин при RT со встряхиванием. После повторного трехкратного промывания планшета 35 мкл буфера считывания (MSD) добавляли в каждую лунку. Сигналы электрохемолюминесценции (ECL) получали и считывали с использованием устройства MSD Sector Imager 6000.
Среднее значение ECL-сигналов рассчитывали на основании результатов измерения дублированных лунок в каждом анализе. Данные корректировали на исходные значения путем вычитания наименьшего значения из всех точек данных и строили график зависимости от соответствующих концентраций антигена. Значения KD определяли путем построения графика в соответствии с моделью связывания 1:1 (для Fab) или 1:2 (для IgG) (согласно Piehler et al., 1997).
Результаты
Результаты суммированы в таблицах 3 и 4. В случае обоих форматов NOV1401, Fab и IgG, значения KD примерно 20 нМ были получены для каталитического домена FXI при определении с использованием BIACORE™. Значения аффинности Fab в отношении как активированного, так и зимогена, FXI находились в пМ диапазоне и были в 66 и 300 раз выше, чем аффинность к каталитическому домену, соответственно. Учитывая их высокую аффинность, для данных взаимодействий проводили измерения в анализах ТРР. NOV1401-Fab демонстрировал в пять раз более высокую аффинность в отношении зимогена FXI (62 пМ), чем в отношении активированного FXI (305 пМ). Значения аффинности NOV1401-IgG в отношении как димерного зимогена, так и активированного FXI, помечены как значения кажущейся KD, поскольку на взаимодействие могли влиять эффекты авидности.
Для подтверждения того, что NOV2401 также связывает FXI яванского макака, проводили эксперименты ТРР с активированным FXI яванского макака и зимогеном FXI яванского макака, в которых были получены значения кажущейся KD 12,5 ± 6,6 пМ (N=2) и 5,0 ± 0,7 пМ (N=2), соответственно. Таким образом, значения аффинности NOV1401 в отношении белков FXI яванского макака (активной формы и зимогена) были сопоставимы с таковыми для связывания с человеческим FXI (таблица 3).
Таблица 2. Значения KD и кинетические параметры связывания NOV1401-Fab/IgG в отношении каталитического домена FXI человека при определении с использованием BIACORE™.
Таблица 3. Значения KD NOV1401-Fab/IgG для активированного и зимогена FXI, определенные методом титрования равновесного раствора (ТРР). *Приведены только значения кажущейся KD, поскольку на взаимодействие могли влиять эффекты авидности.
Литература: Piehler et al. Assessment of affinity constants by rapid solid phase detection of equilibrium binding in a flow system, J.Immunol. Meth. 1997, 189-206.
Пример 3
Биохимический анализ: Ингибирование FXIa в анализе активности с использованием флуоресцентного пептида в качестве субстрата
Активность человеческого FXIa (Kordia Life Science NL, каталожный номер HFXIa 1111a) определяли путем мониторинга расщепления флуоресцентно меченого пептида с последовательностью D-Leu-Pro-Arg*Rh110-D-Pro (каталожный номер BS-2494; Biosyntan GmbH, Berlin, Germany). В последовательности субстрата, приведенной выше, * указывает на неустойчивую химическую связь, D-Leu: D-лейцин, Pro: пролин, Arg: аргинин, Rh110: родамин 110, D-Pro: D-пролин). Опосредованное FXIa расщепление неустойчивой связи пептидного субстрата приводит к увеличению интенсивности флуоресценции родамина 110 при использовании длин волн возбуждения и излучения 485 nm и 535 nm, соответственно. Интенсивность флуоресценции непрерывно измеряли на планшетном ридере Safire2 (TECAN, Maennedorf, Switzerland) при комнатной температуре (RT). Аналитический буфер содержал 50 мМ HEPES с pH 7,4, 125 мМ NaCl, 5 мМ CaCl2 и 0,05% (масс/об) CHAPS. В конечном анализе активности человеческий FXIa и субстрат BS-2494 имели аналитические концентрации 0,1 нМ и 0,5 мкМ, соответственно. В этих условиях зависимость увеличения интенсивности флуоресценции от времени была линейной в течение по меньшей мере 60 минут.
Для тестирования ингибирующей активности антител готовили серийные разведения антител в буфере PBS (137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,8 мМ KH2PO4), содержащем 0,05% (масс/об) CHAPS. Два мкл раствора антитела преинкубировали с 10 мкл раствора FXIa (в аналитическом буфере) в течение 60 минут при RT. После этапа преинкубации добавляли 10 мкл субстрата BS-2494 (разведенного в аналитическом буфере) и оставляли для протекания ферментативной реакции на 60 минут, после чего измеряли интенсивность флуоресценции. Значения интенсивности флуоресценции преобразовывали в процент ингибирования с использованием контрольных реакций (сигнал реакций без ингибирования эквивалентен 0% ингибирования и сигнал в реакционной смеси без фермента эквивалентен 100% ингибирования) и следующей формулы для преобразования значений:
y=100% - [FI(x)-FI(мин)]/[FI(макс)-FI(мин)],
where y представляет собой % ингибирования при концентрации x антитела, FI(x) представляет собой интенсивность флуоресценции, измеренную при концентрации x антитела, FI(мин) представляет собой интенсивность флуоресценции, измеренную в контрольной реакции в отсутствие антитела и FI(макс) представляет собой интенсивность флуоресценции, измеренную в контрольной реакции без ингибирования. Данные анализировали с использованием программы Origin 7.5SR6 (OriginLab Corporation, USA). Значения IC50 из усредненных данных рассчитывали с использованием логистической функции:
y=A2+(A1-A2)/(1+(x/IC50)^p),
где y представляет собой % ингибирования при концентрации x антитела, A1 представляет собой наименьшее значение ингибирования и A2 представляет собой максимальное значение ингибирования. Экспонента, p, представляет собой коэффициент Хилла.
На фигуре 6A приведена репрезентативная кривая зависимости ответа от концентрации соединения для ингибирования антителом NOV1401 ферментативной активности полноразмерного человеческого FXIa. Результаты показывают, что NOV1401 ингибирует ферментативную активность человеческого полноразмерного FXIa зависимым от концентрации образом (фигура 6A). При подгонке с использованием модели логистической кривой было получено значение IC50 примерно 160 пМ.
Пример 4
Антикоагулянтная активность анти-FXIa Ат
Антитромботическую активность антител NOV1401 и NOV1090 тестировали в анализе определения активированного частичного тромбопластинового времени (аЧТВ) и анализе образования тромбина (АОТ).
Анализ аЧТВ:
Лиофилизированную нормальную человеческую плазму «Coagulation Control N» (каталожный № 5020050) приобретали у компании Technoclone GmbH (Vienna, Austria). Она представляла собой объединенную плазму от выбранных здоровых доноров с добавлением цитрата натрия. Время свертывания крови, определенное для этой нормальной плазмы, отражает нормальные концентрации факторов свертывания крови, участвующих в образовании сгустка. Лиофилизированную плазму хранили при 4°C. Перед использованием плазму ресуспендировали в 1 мл дистиллированной воды, осторожно переворачивая пробирку, а затем выдерживали ее в течение 10 минут при RT.
Реагент, инициирующий внутренний путь активации свертывания крови, «aPTT-s» (каталожный № TE0350) приобретали у компании SYCOmed (Lemgo, Germany), он содержал фосфолипид и силикат (коллоидный) в буферном растворе (хлорид натрия, полиэтиленгликоль 20000; сахароза, азид натрия). Раствор хранили при 4°C.
Хлорид кальция (каталожный № C1016-500G; Sigma-Aldrich Chemie GmbH, Steinheim, Germany) готовили в бидистиллированной воде в маточной концентрации 25 мМ.
Трис/HCl буфер с pH 7,5, UltraPure (каталожный № 15567-027; Life Technologies Corporation, NY, USA) и фосфатно-солевой буфер (PBS, каталожный № P4417-100TAB; Sigma-Aldrich Chemie GmbH, Steinheim, Germany) использовали для растворения соединений.
3-[(3-холамидопропил)диметилaммоний]-1-пропансульфонат гидрат (CHAPS, каталожный № C3023-25G) и безводный диметилсульфоксид (DMSO, каталожный № 276855-100 мл) приобретали у компании Sigma-Aldrich Chemie GmbH (Steinheim, Germany).
Измерения времени свертывания крови проводили в коагулометре с шариками Amelung модели KC4A (приобретено у компании SYCOmed, Lemgo, Germany), который представляет собой полуавтоматическую механическую систему обнаружения сгустков. В системе используется специальная кювета (каталожный № AI4000; SYCOmed), в которую помещают шарик из нержавеющей стали (каталожный № AI5000; SYCOmed).
Образец добавляют в кювету. После соответствующего периода инкубации кювету помещают в измерительную ячейку коагулометра. Измерительная ячейка медленно вращается, вызывая вращение кюветы вдоль ее продольной оси. Поскольку кювета расположена под небольшим углом, то из-за гравитации и инерции шарик всегда находится в самой нижней точке кюветы. Точно напротив занимаемого шариком положения находится магнитный датчик. При добавлении инициирующего реагента запускают таймер. По мере того, как происходит свертывание крови, в реакционной смеси образуются нити фибрина. Нити фибрина оттягивают шарик от его инерционного положения, что вызывает импульс в магнитном датчике. Этот импульс электронным образом останавливает таймер. Последовательность действий была следующей (Таблица 4a):
Таблица 4a
объем [мкл]
Измерение образцов выполняли в двойном повторе при температуре 37°C в коагулометре с шариками Amelung.
На фигуре 6B приведена репрезентативная кривая зависимости ответа от концентрации соединения для антитела NOV1401, приводящего к зависимому от концентрации пролонгированию времени свертывания крови в анализе аЧТВ. Результаты свидетельствуют о том, что NOV1401 приводит к пролонгированию времени свертывания крови в анализе аЧТВ человеческой плазмы зависимым от концентрации образом. Время свертывания крови в анализе аЧТВ удваивалось по сравнению с исходным значением при концентрации NOV1401 примерно 14 нМ. Рассчитанное значение IC50 составляло примерно 13 нМ.
Анализ образования тромбина (АОТ):
Для анализа АОТ лиофилизированную нормальную человеческую плазму (Coagulation Control N) приобретали у компании Technoclone GmbH, (каталожный номер 5020040, лот № 1P37B00) и восстанавливали в дистиллированной воде в объеме, предлагаемом производителем.
Раствор субстрата готовили с использованием флуорогенного субстрата Z-Gly-Gly-Arg-AMC от компании Technoclone GmbH (каталожный номер 5006230, лот № 8F41B00). Аликвоты лиофилизированного субстрата хранили при 4°C. Субстрат заново растворяли в объеме дистиллированной воды, указанном на флаконе, за 20 минут до использования в анализе. Восстановленный раствор субстрата содержал флуорогенный пептид в концентрации 1 мМ и CaCl2 в концентрации 15 мМ.
Два разных реагента «TGA RD» (каталожный № 500622) и «TGA RC low» (каталожный № 5006213) для инициации внутреннего и внешнего пути активации свертывания крови, соответственно, приобретали у компании Technoclone GmbH (Vienna, Austria). Инициирующий реагент «platelet poor plasma (PPP)-reagent low» приобретали у компании Thrombinoscope (TS31.00, лот № PPL1409/01) и восстанавливали в объеме дистиллированной воды, указанном на флаконе. «PPP-reagent low» содержит смесь фосфолипидов и тканевого фактора в очень низкой концентрации. Реагент разводили в 8 раз в 80 мМ трис/HCl, pH 7,4, 0,05% (масс/об) CHAPS непосредственно перед использованием.
Образцы разделяли на аликвоты и проводили измерения в 96-луночных черных планшетах с прозрачным дном, приобретенных у компании Costar (каталожный номер 3603). Для автоматического переноса образцы помещали в 96-луночный планшет с V-образным дном (Costar, 3894) и переносили с использованием автоматизированной системы CyBio (Analytik Jena US, Woburn, MA, USA).
Восстановленную плазму крови человека, инициирующий реагент «PPP-reagent low» и субстрат предварительно нагревали в течение 10 минут в водяной бане при 37°C. Готовили серийные разведения 1:3 антитела в PBS в 96-луночном планшете, начиная с концентрации NOV1401 5 мкМ (5x наивысшая конечная концентрация от 1 мкМ), в общей сложности 8 разведений. 222 мкл инициирующего реагента смешивали с 1108 мкл раствора субстрата, получая смесь 10+50 инициирующего реагента с субстратом. Добавляли по 80 мкл в лунку 96-луночного планшета с V-образным дном для последующего переноса с помощью автоматизированной системы. Планшет хранили при 37°C. Реагенты добавляли в соответствии со схемой, приведенной в таблице 4b.
Таблица 4b
Смеси инициатор/субстрат переносили с использованием автоматизированной системы. После добавления смесей немедленно регистрировали показатели при длинах волн возбуждения и излучения 360 нм и 460 нм, соответственно, на приборе Synergy Neo (BioTek Instrument Inc., Winooski, VT, USA). Измерения для образцов выполняли в двойном повторе при температуре 37°C в планшетном ридере в течение 90 минут с интервалами 55 секунд.
Для получения пиковых значений концентрации тромбина данные обрабатывали с использованием файла оценочной программы для АОТ, предоставленного компанией Technoclone. Для построения графиков зависимости пиковой концентрации тромбина от концентрации антитела выполняли подгонку данных с использованием программы GraphPad. Эти данные были подогнаны к модели нелинейной регрессии с помощью программы GraphPad Prism5 (GraphPad Software Inc., La Jolla, CA, USA). Величину IC50 определяли с использованием встроенного четырехпараметрического уравнения кривой зависимости «доза-ответ» (изменяемый наклон): y = низшая + (высшая - низшая) / (1 + 10^((LogIC50 - x) * наклон)), где y представляет собой максимальную концентрацию тромбина, образованного при концентрации x ингибитора; а высшая и низшая представляют собой концентрации тромбина без ингибитора и при наивысшей концентрации ингибитора, соответственно.
На фигуре 6C приведена репрезентативная кривая зависимости ответа от концентрации соединения для антитела NOV1401, демонстрирующая зависимое от концентрации ингибирование образования тромбина в анализе АОТ. Значение IC50, составляющее 24 нМ, и остаточная концентрация тромбина, составляющая 159 нМ (пунктирная линия), были рассчитаны на основании данной кривой зависимости ответа от концентрации соединения.
Пример 5
Экспрессия белка, образование комплекса, кристаллизация и определение структуры комплекса NOV1401 (Fab) -FXI (каталитический домен)
Структуру Fab-фрагмента антитела NOV1401, полученного в результате расщепления папаином, в комплексе с каталитическим доменом фактора XI определяли путем совместной кристаллизации с разрешением 2,04Е.
Экспрессия белка :
Экспрессионный конструкт для каталитического домена FXI состоял из аминокислотных остатков 388-625 (Swissprot P03951) с неспаренным остатком цистеина, C500, мутированным в цистеин, с N-концевым удлинением, состоящим из аминокислот MGSS (SEQ ID NO: 49), октагистидиновым тегом (SEQ ID NO: 50), сайтом расщепления PreScission™, за которым следовал сайт расщепления энтерокиназой. Конструкт собирали методом генного синтеза, клонировали в экспрессионный вектор pET24a и экспрессировали в виде телец включения в клетках E. coli штамма BL21 (DE3), растущих в LB-среде. Тельца включения солюбилизировали в смеси 50 мМ трис/HCl, pH 8,0, 6,0 M гуанидин хлорида, 50 мМ DTT в течение 2 часов, полностью денатурируя рекомбинантный белок. Буфер для рефолдинга (0,5 M трис, pH 8,0, 0,9 M аргинин-HCl, 5 мМ GSH, 0,5 мМ GSSG, 1 мМ ЭДТА) в большом избытке быстро добавляли в раствор IB, получая конечную концентрацию белка 50 мкг/мл, и инкубировали при 4°C в течение 5 дней. Рефолдинг и образование дисульфидных мостов осуществляли путем диализа с буфером A (50 мМ трис, pH 8,0) в течение трех дней.
Белок в восстановленной конформации наносили на колонку для анионообменной хроматографии, содержащую Q-Sepharose® FF (GE Healthcare), уравновешенную и промытую буфером A. Не связавшийся рекомбинантный белок собирали в протекшем через колонку буфере и фракциях промывки. Значение pH доводили до 7,4 путем диализа с 50 мМ трис-буфером, pH 7,4, перед удалением N-концевой последовательности тега с использованием энтерокиназы (соотношение энтерокиназа: рекомбинантный белок 1:100, время инкубации 2,5 ч). Реакцию расщепления останавливали, нанося образец на бензамидиновую колонку для аффинной хроматографии, содержащую бензамидин-Sepharose 4 FF (high sub) (GE Healthcare), уравновешенную и промытую буфером B (50 мМ трис, pH 7,4, 0,5 M NaCl), и элюировали буфером C (буфер B, содержащий 50 мМ бензамидин). Активный каталитический домен FXI наносили на XK 26/600 Superdex 75 колонку для эксклюзионной хроматографии (GE Healthcare), уравновешенную 20 мМ Na-ацетатным буфером, pH 5,3, с 75 мМ NaCl. Конечная концентрация белка составляла 1,07 мг/мл.
Fab-фрагмент NOV1401 получали путем расщепления IgG папаином. Конечная концентрация Fab в PBS составляла 11 мг/мл. Расщепление проводили в течение ночи при 37°C с использованием папаина (Roche Diagnostics 108 014; 10 мг/мл), добавленного к антителу в соотношении 1:100 (по массе), в присутствии 1 мМ цистеина (добавленного к исходному раствору IgG). Расщепление останавливали добавлением 50 мкМ специфического ингибитора папаина E64, (N-[N-(L-3-транс-карбоксиран-2-карбонил)-L-лейцил]агматина, и продукты расщепления пропускали через небольшую колонку с белком A (5 мл) для удаления Fc-фрагмента. Fab-фрагменты собирали в протекшем через колонку буфере, диализовали против PBS, концентрировали до их конечной концентрации ультрафильтрацией и стерилизовали фильтрованием (0,22 мкм).
Образование комплекса, кристаллизация и определение структуры:
Каталитический домен FXI и Fab смешивали в эквимолярном соотношении и концентрировали до конечной концентрации 9 мг/мл.
Кристаллы, используемые для сбора данных, получали при 277K с использованием диффузии паров в сидячей капле, смешивая 0,3 мкл резервуарного раствора (0,2 M хлорид аммония, 20% PEG 3350), 0,2 мкл комплекса Fab-FXI и 0,1 мкл кристалла-затравки из кристаллов, полученных в первом раунде скрининга кристаллов.
Для сбора данных кристаллы быстро замораживали в жидком азоте. Данные получали на канале синхронного излучения Swiss Light Source X10SA при длине волны 1,00002 Е с использованием пиксельного детектора Pilatus (Dectris) при 100K. Обработку и масштабирование данных выполняли с использованием XDS и XSCALE (Kabsch, W. (2010) Acta Cryst. D66, 125-132). Кристалл дифрагировал до разрешения 2,04 Е с размерами элементарной ячейки a=191,27, b=53,22, c=65,164, альфа=90,0, бета=94,56, гамма=90,0 (пространственная группа C2) с одной копией комплекса на ассиметричную единицу.
Структуру комплекса определяли путем молекулярной замены с использованием структур каталитического домена FXI и укороченного Fab-фрагмента, ранее определенных в собственной лаборатории, в качестве моделей поиска, используя PHASER (McCoy, A.J. et al. (2007) J. Appl. Cryst. 40, 658-674). Проводили чередующиеся циклы уточнения и реконструкции с использованием buster и coot rsp. (Bricogne, G. et al. (2010) BUSTER версия 2.9. Cambridge, United Kingdom: Global Phasing Ltd.; Emsley, P. and Cowtan, K. (2004). Acta Crystallogr. D60, 2126-2132). Полученные данные и статистика уточнений приведены в таблице 5.
Таблица 5. Совокупность данных и статистика уточнений
*Значения в скобках соответствуют области наивысшего разрешения.
Описание структуры:
Структурные данные показали, что связывающий эпитоп антитела NOV1401 связывается с поверхностью активного центра, при этом петля CDR3 тяжелой цепи накрывает части подцентров S3, S2, S1-beta и S1'. Соседние петли CDR1 и CDR2 тяжелой цепи вызывают конформационные изменения в петлях 145 и 220 FXI (химотрипсиновая нумерация). Кроме того, четыре N-концевых остатка FXI, а также остатки, окружающие Asp189, становятся неупорядоченными; обе эти области являются ключевыми областями для каталитической активности FXI. Конформационное изменение петли 145 приводит к закрытию кармана S1 остатком Arg144 и подцентра S2' остатком Tyr143. Таким образом, связывание антитела приводит к принятию FXI ингибированной конформации за счет нескольких механизмов.
Наблюдаемая ингибированная форма имеет особенности, аналогичные тем, которые описаны для полноразмерной формы зимогена FXI (PDB 2F83). Части каталитического домена FXI, которые изменили конформацию и стали неупорядоченными вследствие связывания антитела, в зимогене также являются неупорядоченными. Это объясняет сильное связывание NOV1401 также и с FXI в форме зимогена.
Данные авторов изобретения, указывающие на то, что NOV1401 не ингибирует активацию зимогена FXI, объясняются расстоянием между связывающим эпитопом и сайтом расщепления для активации зимогена FXIa. NOV1401 связывает как FXI, так и FXIa. Рентгеноструктурный анализ комплекса Fab-FXI КД позволил выявить уникальный способ связывания и механизм ингибирования FXIa. NOV1401 связывается с активным центром FXIa (фигура 4) и вызывает конформационные изменения четырех N-концевых остатков и петель каталитического домена, приводящие к возникновению неактивной конформации. Эта неактивная конформация имеет сходные особенности со структурой неактивного каталитического домена в зимогене (фигура 5), это объясняет, каким образом и FXI, и FXIa, могут связываться с высокой аффинностью антителом NOV1401. Например, три петли каталитического центра (например, петля 220, петля 188 и петля 145), которые являются неупорядоченными в структуре зимогена, также являются неупорядоченными или смещенными в структуре комплекса NOV1401 Fab-FXI КД, и N-концевой солевой мост, наблюдаемый в активной конформации, отсутствует в структурах как зимогена, так и комплекса NOV1401 Fab-FXI КД (таблица 6). Таким образом, NOV1401, судя по всему, вызывает конформационные изменения в КД, приводящие к возникновению неактивной, зимоген-подобной конформации.
Таблица 6: Структурные характеристики FXIa КД, FXIa КД в комплексе с NOV1401 и зимогена FXI (КД):
Пример 6
Картирование эпитопов на основании рентгеноструктурного анализа
Остатки FXI, контактирующие с Fab, проанализированные с использованием AREAIMOL (Briggs, P.J. (2000) CCP4 Newsletter No. 38) путем определения различий в площади поверхности остатков при расчете без связанного Fab и в комплексе с Fab, приведены в таблице 7 и таблице 8a (нумерация Swissprot):
Таблица 7: Эпитоп FXI
Эпитоп (подчеркнуты: контакты легкой цепи и тяжелой цепи):
Pro410
Arg413
His431
Tyr434
Gly435
Glu437
Tyr472
Lys473
Met474
Glu476
Tyr521
Leu524
Arg525
Asp526
His552
Leu415
Cys416
His431
Cys432
Tyr434
Tyr472
Met474
Ala475
Glu476
Tyr521
Arg522
Lys523
Leu524
Arg525
Asp526
Lys527
Arg548
Ser575
Ser594
Trp595
Gly596
Glu597
Arg602
Glu603
Arg604
Эпитоп FXI образован следующими остатками:
Pro410, Arg413, Leu415, Cys416, His431, Cys432, Tyr434, Gly435, Glu437, Tyr472-Glu476, Tyr521-Lys527, Arg548, His552, Ser575, Ser594-Glu597, Arg602-Arg604.
Таблица 8a: Остатки FXI, контактирующие с NOV1401 (эпитоп)
Эпитоп, картированный методом рентгеноструктурного анализа на последовательности каталитического домена (остатки, образующие эпитоп, выделены жирным шрифтом и подчеркнуты):

В таблице 8b приведены остатки антитела, контактирующие с FXI (паратоп).
Таблица 8b: Остатки NOV1401, контактирующие с FXI (паратоп).
L, легкая цепь; H, тяжелая цепь
Пример 7
Эффект анти-FXI антитела на индуцированный FeCl3 тромбоз у мышей
Мыши с дефицитом FXI (мыши FXI-/-) на фоновом генотипе C57Bl были получены от компании Novartis (E. Hanover, NJ) и использованы для оценки антитромботической эффективности NOV1401. После внутривенного введения человеческого FXI (чFXI) эти мыши приобретают тромбофилический фенотип дикого типа под воздействием тромбогенного стимула. В исследованиях, описанных в настоящем документе, тромбоз индуцировали в сонной артерии путем нанесения на поверхность артерии хлорида железа (FeCl3).
NOV1401 вводили болюсной инъекцией через яремную вену анестезированным мышам за 15 минут до индукции тромбоза. Дозы антитела находились в диапазоне 0,24 мг/кг - 0,47 мг/кг. У мышей FXI-/- восстанавливали дефицит за счет человеческого FXI, вводя инъекцией 0,47 мг/кг человеческого FXI через яремную вену за 10 минут до стимуляции FeCl3. Два фрагмента фильтровальной бумаги размером 1 мм x 1,5 мм, насыщенные 3,5% раствором FeCl3, наносили на противоположные стороны сонной артерии, в контакте с ее адвентициальной поверхностью, и удаляли через 3 минуты, с последующим тщательным промыванием солевым раствором. Поток крови через сонную артерию измеряли с помощью датчика измерения потока от компании Transonic. Исходный поток крови определяли в течение 5 минут перед нанесением FeCl3, а затем в течение 30 минут после нанесения FeCl3 (то есть, на протяжении тромбогенного периода). В конце эксперимента образец крови отбирали из полой вены в шприц, содержащий 3,8% раствор цитрата натрия, получали плазму и проводили анализ аЧТВ.
На фигуре 1A показан эффект NOV1401 на индуцированный FeCl3 тромбоз у мышей FXI-/- с введенным человеческим FXI (мышиная модель с гуманизированным FXI). На фигуре 1B показан эффект NOV1401 на аЧТВ в той же мышиной модели. На фигуре 1C показано пролонгирование аЧТВ у мышей FXI-/- по сравнению с мышами дикого типа.
Антитело NOV1401 полностью ингибировало индуцированное FeCl3 образование тромба у мышей FXI-/- с введенным чFXI (фигура 1A), начиная с концентрации 0,24 мг/кг. Наблюдали очень сильную реакцию на введенную дозу, по всей вероятности, отражающую стехиометрический антитромботический ответ типа «все или ничего». Продолжительность аЧТВ была увеличена в 1,6 раза в группе высокой дозы относительно контролей с растворителем (фигура 1 B), что соответствовало такому же уровню пролонгации, что и в случае генетического дефицита FXI (фигура 1C), то есть, бы достигнут максимальный эффект. Эти результаты показывают, что NOV1401 обладает антитромботической активностью в мышиной модели индуцированного FeCl3 тромбоза.
Пример 8
Эффекты анти-FXI антитела на уровень свободного FXI и аЧТВ у яванских макак
Для оценки фармакокинетического (ФК) профиля и фармакологических эффектов анти-FXI/FXIa антитела, такого как NOV1401, антитело вводили подкожными (п/к) или внутривенными (в/в) инъекциями яванским макакам в исследовании с повышением дозы.
Антикоагулянтный эффект NOV1401 изучали на яванских макаках, тестируя способность антитела пролонгировать аЧТВ и снижать уровни свободного FXI (FXIс) после одной внутривенной (N=2) или подкожной (N=2) инъекции дозы 3 мг/кг. Вторую дозу 10 мг/кг вводили всем животным, с последующим введением третьей дозы 30 мг/кг для определения того, могут ли эффекты, наблюдаемые в случае дозы 3 мг/кг, быть потенцированы при более высоких дозах. Полученные результаты показывают, что антитело NOV1401 обладает устойчивой антикоагулянтной активностью у яванских макак. Фармакодинамику (ФД) NOV1401, характеризуемую его антикоагулянтным эффектом, определяемым на основании аЧТВ и уровней FXIс, затем сопоставляли с ФК профилем. Сравнение показало, что имеет место хорошая корреляция ФК/ФД.
Животным вводили либо в/в (N=2), либо п/к (N=2), инъекцией NOV1401 в день исследования 1 в дозе 3 мг/кг, день 85 в дозе 10 мг/кг и день 114 в дозе 30 мг/кг. Образцы крови собирали в пробирки с цитратом натрия через 15 минут и 2 часа после введения доз у животных в случае в/в введения и у всех животных до начала исследования, через 6, 24, 48, 72 и 96 часов после введения доз (при введении в дни 1, 85 и 114) и через 8, 11, 15, 18, 22, 25, 29, 32, 36, 39, 43, 46, 50, 53, 57, 60, 64, 66, 71, 75 и 78 дней после введения доз (только при введении в день 1). Кровь также собирали в дни 92, 95, 99, 102, 107, 110, 121, 124, 128 и перед дозированием в день 114. Все образцы крови центрифугировали, получали образцы плазмы и замораживали при температуре примерно -70 C или ниже.
Общие концентрации NOV1401 в плазме измеряли в стандартном анализе ELISA для обнаружения человеческих IgG, используя сэндвич-метод иммуноферментного анализа с моноклональными антителами мыши против IgG человека в качестве захватывающего антитела и мечеными HRP антителами козы против IgG человека в качестве обнаруживающего антитела.
Для измерений свободного FXI в образцах плазмы, содержащих как FXI, так и NOV1401, несвязанный FXI захватывали с помощью иммобилизованного NOV1401 и FXI, уже находящийся в комплексе с NOV1401, отмывали. Затем связанный с планшетом FXI обнаруживали с помощью содержащего Fc мышиного антитела 14E11, моноклонального антитела, которое связывает домен A2 в FXI и описано в литературе (Cheng, et al. Blood, 116: 3981-3989, 2010). Очень высокая аффинность NOV1401 в отношении как FXI, так и FXIa, а также разные сайты связывания для NOV1401 и обнаруживающего антитела 14E11 позволили произвести точные измерения свободного FXI.
Планшеты для ELISA (384-луночные LUMITRAC™ 600 HB) покрывали NOV1401 (5 мкг/мл в PBS) для связывания свободного FXI. После блокирования (молочный блокирующий раствор: KPL № 50-82-01, разведение 1:20) и промывания планшетов промывочным буфером (PBS; 0,05% Tween 20), инкубировали образцы плазмы, разведенные 1:40 в аналитическом буфере (50 мМ HEPES, pH 7,4, 125 мМ NaCl, 5 мМ CaCl2, 5 мМ ЭДТА и 0,05% (масс/об) CHAPS), при RT в течение 30 минут и промывали 3 раза промывочным буфером. Обнаруживающее антитело 14E11 добавляли в концентрации 1 мкг/мл в буфере разведения (1,7 мМ одноосновный фосфат натрия, 8,1 мМ двухосновный фосфат натрия гептагидрат, 0,15 M хлорид натрия, 0,7% Triton X-100 и 0,1% азид натрия, pH 7), содержащем 0,7% казеина. После промывания планшетов промывочным буфером добавляли вторичное обнаруживающее антитело, меченое пероксидазой антитело против IgG мыши (Sigma № A5278), в концентрации 0,5 мкг/мл в буфере разведения, содержащем 0,4% казеина. После промывания планшетов промывочным буфером добавляли 50 мкл раствора хемилюминесцентного субстрата пероксидазы (LumiGLO, KPL № 54-61-01), и люминесцентный сигнал немедленно считывали на многорежимном планшетном ридере (SPECTRAMAX M5E). Концентрацию свободного FXI в каждом образце определяли при помощи стандартной кривой, построенной с использованием человеческого FXI (зимогена) от компании Enzyme Research Laboratories (каталожный № HFXI 1111), начиная с 100 нМ FXI с фактором разведения 2 и 22 шагами разведения. Нижний предел количественной оценки (LLOQ) составлял 0,24 нМ, с учетом разведения 1:40 до проведения измерений.
Образцы плазмы, собранные во всех временных точках, подвергали анализу аЧТВ, и результаты анализа аЧТВ сопоставляли с общей концентрацией в плазме NOV1401 и уровнями свободного FXI. На фигурах 2A и 2B показаны изменения времени свертывания крови в анализе аЧТВ относительно общих уровней в плазме NOV1401 для животных, которым вводили дозы в/в и п/к. На фигурах 3A и 3B показаны изменения времени свертывания крови в анализе аЧТВ относительно уровней свободного FXI для животных, которым вводили дозы в/в и п/к.
В случае в/в вводимого NOV1401 максимальные общие уровни в плазме NOV1401 наблюдали через 15 минут после введения доз (фигура 2A). В это время аЧТВ было примерно удвоенным относительно исходного уровня у обоих животных и оставалось на этом уровне в среднем в течение 5-6 недель. Среднее пролонгирование аЧТВ от временной точки 15 минут после введения дозы на протяжении периода измерений до снижения к исходному уровню составляло 2,0 ± 0,02 раза и 1,9 ± 0,03 раза для каждого животного.
К дню 85, перед введением второй дозы, аЧТВ достигало исходных уровней и концентрации в плазме NOV1401 падали ниже 10 нМ. Вторую дозу 10 мг/кг вводили в день 85, что приводило к увеличению общей концентрации в плазме NOV1401 по меньшей мере примерно в 3 раза и вызывало пролонгирование аЧТВ, аналогичное тому, которое наблюдали после введения первой дозы. Третью дозу 30 мг/кг вводили в день 114, при том, что аЧТВ все еще было пролонгированным, и она не приводила к какому-либо существенному дополнительному пролонгированию аЧТВ, несмотря на еще одно по меньшей мере 3-кратное увеличение общей концентрации в плазме NOV1401 (фигура 2A). Таким образом, дозы NOV1401, превышающие 3 мг/кг, приводили к пролонгированию аЧТВ, сопоставимому с пролонгированием в случае дозы 3 мг/кг, и по-видимому не вызывали увеличение степени пролонгирования аЧТВ. Как и ожидалось, п/к введение NOV1401 приводило к более медленному возрастанию аЧТВ, чем в случае в/в введения, однако степень пролонгирования была сопоставима с таковой в группе в/в введения (фигура 2B). У двух животных аЧТВ было пролонгировано относительно исходного значения в среднем в течение 5-6 недель. Средняя кратность пролонгирования аЧТВ была аналогичной той, которую наблюдали у животных, получавших в/в дозу: 2,0 ± 0,03 и 1,8 ± 0,02 от временной точки 6 часов после введения дозы на протяжении периода измерений до снижения к исходному уровню. Как и при в/в введении, введение более высоких доз не приводило к более высоким показателям аЧТВ, несмотря на более высокую экспозицию NOV1401 в плазме.
Результаты на фигурах 2A-2B показывают, что NOV1401 приводит к пролонгированию аЧТВ у яванских макак.
Средняя исходная концентрация в плазме FXIс составляла 10,9 ± 0,3 нМ в группе в/в введения и быстро падала (за 15 мин) после введения NOV1401 (фигура 3A). Уровни в плазме FXIс оставались низкими до того, как общие уровни в плазме NOV1401 падали до значений 15 нМ - 25 нМ (фигура 2A, фигура 3A). В группе п/к введения средняя исходная концентрация FXIс составляла 14,3 ± 1,0 нМ. Концентрация FXIс была резко снижена относительно исходной концентрации через 6 часов после введения дозы (фигура 3B) и оставалась низкой до того, как уровни в плазме NOV1401 снижались до значений 15 нМ - 25 нМ (фигура 2B, фигура 3B). Содержание FXIс вновь резко падало после второй дозы 10 мг/кг у всех животных и оставалось низким до конца исследования. Две более высокие дозы не приводили к дополнительному снижению уровня FXIс относительно исходных значений.
У всех подвергнутых воздействию животных падение и восстановление уровней FXIс было временным и обратно пропорциональным индуцированному NOV1401 пролонгированию аЧТВ, это подтверждало, что NOV1401 ингибирует функцию внутреннего пути активации свертывания крови (пролонгирует аЧТВ) за счет снижения уровня FXIс.
Данные результаты (например, фигуры 3A и 3B) демонстрируют, что NOV1401 вызывает снижение уровней в плазме FXIс у яванских макак. В исследованиях на яванских макаках не наблюдали признаков избыточного кровотечения в местах прокола вен или при патологоанатомическом исследовании. Кроме того, во время исследования в стуле не была обнаружена скрытая кровь.
Устойчивый антикоагулянтный эффект NOV1401 также наблюдали у яванских макак в исследовании токсичности при повторном введении дозы, которое продолжалось 13 недель в случае п/к введения или 4 недель в случае в/в введения. В данном исследовании NOV1401 вводили еженедельно в дозах 10 мг/кг (N=3, самцы и самки вместе) и 100 мг/кг (N=5, самцы и самки вместе) п/к в течение 13 недель (14 доз) или в дозе 50 мг/кг (N=3, самцы и самки вместе) в/в в течение 4 недель (5 доз). Контрольная группа (N=5, самцы и самки вместе) получала растворитель в течение 13 и 4 недель п/к и в/в, соответственно. FXI:C оценивали путем измерения времени свертывания крови в образцах плазмы яванских макак в присутствии дефицитной по FXI человеческой плазмы (одностадийное аЧТВ). аЧТВ и FXI:C измеряли в дни исследования 2, 23 и 79 в группах п/к введения, и в дни 2 и 23 в группе в/в введения. У всех животных и во всех группах введения наблюдали пролонгирование аЧТВ в 2,1-3 раза (фигура 7A). Эффект сохранялся на протяжении фазы дозирования исследования и никакой зависимости от дозы не наблюдали так же, как и в предшествующем исследовании с повышением дозы. Значение FXI:C уменьшалось у животных во всех группах введения на 88-95% и оставалось на этих уровнях на протяжении фазы дозирования исследования (фигура 7B). Эффект на FXI:C не зависел от дозы в данном диапазоне доз.
Никаких макроскопических или микроскопических признаков кровотечения, включая избыточное кровотечение, не наблюдали в местах прокола вен (включая зоны п/к и в/в инъекций, а также зоны отбора образцов крови) или при патологоанатомическом исследовании. Более того, скрытую кровь не обнаруживали в стуле в конце исследования. Кроме того, отсутствовали случаи гибели животных и отсутствовали связанные с тестируемым веществом эффекты на клинические признаки, массу тела, потребление пищи, офтальмологические и электрокардиографические параметры, показатели гематологии, клинической химии или результаты анализа мочи. Не были идентифицированы никакие органы-мишени токсичности.
Увеличение массы щитовидной железы наблюдали у самцов в группе п/к введения дозы 100 мг/кг. Однако токсикологическая значимость этих результатов является неубедительной, поскольку не было никаких гистологических корреляций. У животных имела место сильная вариабельность массы щитовидной железы, и аномальные показатели наблюдали только у животных одного пола. При микроскопическом исследовании наблюдали зависимый от дозы фиброз в местах п/к инъекций как у самцов, так и у самок, в группе п/к введения в дозах 10 и 100 мг/кг/неделю. Эти результаты не считались неблагоприятными.
Никаких существенных признаков токсичности не наблюдали в исследовании однократных возрастающих доз или исследовании общей токсичности при повторном введении доз у яванских макак в течение вплоть до 13 недель. Вследствие этого, наивысший уровень п/к вводимой дозы в 13-недельном GLP исследовании токсичности (100 мг/кг/неделю) определяли, как NOAEL.
Пример 9
Фармакокинетика у яванских макак - однократная доза
Яванским макакам (самкам, N=2) вводили одну дозу 3 мг/кг NOV1401 либо в/в, либо п/к, и наблюдали до тех пор, пока концентрации в плазме FXIс и аЧТВ не возвращались к уровню до введения дозы. Затем животным вводили одну дозу 10 мг/кг NOV1401 либо в/в, либо п/к, через 2 недели после которой вводили дозу 30 мг/кг либо в/в, либо п/к, с последующим дополнительным 2-недельным периодом наблюдения. ФК NOV1401 оценивали путем измерения общего уровня NOV1401. Показатели экспозиции общего NOV1401, определяемые либо по максимальной наблюдаемой общей концентрации NOV1401 (Cmax), либо по площади под кривой зависимости общей концентрации NOV1401 от времени (AUC0-14д), были сопоставимы у отдельных животных в каждой группе. Экспозиция (Cmax или AUC0-14д) была примерно зависимой от дозы при каждом пути введения (таблица 9). Значение Cmax было примерно в 3 раза выше в группе в/в введения, чем в группе п/к введения. Однако общие концентрации в плазме NOV1401 были сходными в обеих группах после начальной фазы распределения. Терминальный период полувыведения (t1/2) оценивали для каждого животного с использованием двухкомпонентной модели после введения дозы 3 мг/кг. Период t1/2 составлял 14-15 дней (N=2). Абсолютная биодоступность после п/к инъекции составляла 61-66% (3 уровня дозы). Ни у одного животного не были обнаружены анти-NOV1401 антитела ни после в/в, ни после п/к введения.
Таблица 9: Средние фармакокинетические параметры после введения одной (возрастающей) дозы у самок яванских макак
* tmax представлено в виде среднего значения.
Пример 10
Токсикокинетика у яванских макак - повторное введение доз
Яванским макакам вводили еженедельно дозы 10 или 100 мг/кг NOV1401 п/к в течение 13 недель (14 доз) или дозы 50 мг/кг NOV1401 в/в в течение 4 недель (5 доз). Животные, получавшие NOV1401, были подвергнуты воздействию NOV1401 в течение фазы дозирования исследования; у контрольных животных воздействие отсутствовало. Не наблюдали никаких связанных с полом различий экспозиции в плазме общего NOV1401. Увеличение экспозиции (как Cmax, так и AUC0-7д) было зависимым от дозы как у самцов, так и у самок (таблица 10). Анти-NOV1401 антитела были обнаружены у 5 из 6 животных в группе 10 мг/кг/неделю п/к, у 1 из 10 животных в группе 100 мг/кг/неделю п/к и у 1 из 6 животных в группе 50 мг/кг/неделю в/в. Экспозиция общего NOV1401 не была аномальной ни в одной из групп п/к введения. Только одно животное, положительное по антителам против лекарственного средства (ADA), имело значение AUC0-7д в день исследования 22, которое было ниже, чем у других животных в той же группе (50 мг/кг/неделю в/в). У данного животного не было обнаружено влияния на пролонгирование аЧТВ и не наблюдали никакой токсичности.
Таблица 10: Средние токсикокинетические параметры для предпоследней дозы (день исследования 85 для групп п/к введения, день исследования 22 для группы в/в введения) в 13-недельном/4-недельном GLP исследовании токсичности у яванских макак (самцов+самок вместе)
* tmax представлено в виде диапазона наблюдаемых значений.
Пример 11
Изучение эффекта повышения дозы у человека
Исследования с участием людей проводят для оценки безопасности и переносимости анти-FXI/FXIa антител, таких как NOV1401, после однократного введения дозы здоровым субъектам. В общей сложности примерно 60 здоровых мужчин, а также женщин в постменопаузальном возрасте/после хирургической стерилизации, в возрасте 18-55 лет принимают участие в данном исследовании. Хорошее состояние здоровья определяют на основании анамнеза, медицинского осмотра, неврологического обследования, определения жизненно важных показателей, электрокардиограммы (ЭКГ) и результатов лабораторных анализов при скрининге. Выбранные субъекты имеют массу тела по меньшей мере 50 кг и индекс массы тела (ИМТ) в пределах 18-35 кг/м2. ИМТ=масса тела (кг)/[рост (м)]2.
Шесть уровней п/к вводимых доз 5, 15, 50, 150, 300 и 600 мг будут протестированы в исследовании с участием людей при условии, что прогнозируемая средняя продолжительность пролонгирования аЧТВ ≥ 2-кратной не сохраняется на период ≥ 42 дней для любой тестируемой дозы. Проводят два промежуточных анализа (ПА) для подтверждения выбора дозы для 2 наиболее высоких уровней дозы. Если прогнозируемая с помощью модели средняя продолжительность пролонгирования аЧТВ будет ≥ 2-кратной в течение периода более 42 дней при дозе 300 мг или дозе 600 мг, дозу можно снижать, например, на основании имитационной модели, для обеспечения того, чтобы средняя продолжительность пролонгирования аЧТВ не превышала 2-кратную в течение ≥ 42 дней. Неограничивающие иллюстративные корректировки дозы могут включать снижение дозы с использованием шагов понижения дозы 10 мг, 20 мг, 30 мг, 40 мг или 50 мг.
Первые три этапа повышения дозы выполняют с шагами повышения дозы ≈1/2 log. Последние 2 этапа повышения дозы составляют ≤2-кратных повышений для снижения риска продолжительной насыщенности мишени и увеличения пролонгирования аЧТВ.
Максимальную продолжительность 2-кратного пролонгирования аЧТВ в течение определенного количества дней (например, 30 дней, 35 дней, 40 дней, 42 дня и так далее) с терапией, направленной на FXI, можно оценивать на основании генетических данных, демонстрирующих фенотип с умеренными кровотечениями у пациентов с тяжелым дефицитом FXI, данных от пациентов с дефицитом FXI с приобретенным ингибитором, а также результатов исследований с участием людей, например, FXI-ASO (смотри, например, Liu et al., (2011) «ISIS-FXIRx, a novel and specific antisense inhibitor of factor XI, caused significant reduction in FXI antigen and activity and increased aPTT without causing bleeding in healthy volunteers». Представлено на 53-м ежегодном собрании и выставке Американского гематологического общества, San Diego, California. Blood; 118: Abstract 209), когда введение нескольких доз FXI-ASO на протяжении 6 недель приводило к сильному и устойчивому истощению FXI в течение > 6 недель (42 дней) без эпизодов кровотечения. В конкретных вариантах осуществления анализ модели позволяет прогнозировать, что максимальное пролонгирование аЧТВ, равное ≈ 2,7-кратному (относительно состояния до введения дозы) может быть временно достигнуто при п/к введении дозы 50 мг NOV1401 (60-кг субъекту). В конкретных вариантах осуществления прогнозируется, что более высокие дозы будут приводить к увеличению продолжительности этого максимального пролонгирования аЧТВ в 2,7 раза.
На протяжении исследования проводят мониторинг субъектов в отношении параметров безопасности и/или конечных точек исследования, например, медицинский осмотр, неврологическое обследование, определение жизненно важных показателей, электрокардиографическое обследование (ЭКГ), лабораторные анализы и выявление нежелательных явлений (НЯ), включая серьезные НЯ (СНЯ), вплоть до, и включительно, дня 106 после введения дозы.
Эффект анти-FXI/FXIa антитела (например, NOV1401) на аЧТВ оценивают на основании относительного изменения от исходных значений. Общие концентрации в плазме анти-FXI/FXIa антитела (например, NOV1401) измеряют для оценки ФК однократных доз у этих субъектов.
Для оценки иммуногенности (ИГ) анти-FXI/FXIa антител (например, NOV1401) проводят скрининг и подтверждение для ADA.
Содержание свободного и общего FXI, а также коагуляционную активность FXI (FXI:C) измеряют для оценки эффектов анти-FXI/FXIa антител (например, NOV1401) на связывание мишени и относящиеся к мишени ФД параметры.
Содержание D-димера, протромбиновых фрагментов 1,2 (F1,2) и протромбин-антитромбинового комплекса (ТАТ) оценивают для определения эффектов анти-FXI/FXIa антител (например, NOV1401) на параметры тромбогенеза.
Для изучения эффектов анти-FXI/FXIa антител (например, NOV1401) на другие параметры свертывания крови у субъектов можно оценивать следующие показатели: протромбиновое время (ПВ), тромбиновое время (ТВ) и диагностические лабораторные показатели свертывания крови, такие как активируемый тромбином ингибитор фибринолиза, фибриноген, тканевой активатор плазминогена (tPA) и АОТ.
Исследуемые биомаркеры могут включать, но не ограничиваются ими: D-димер, активность FXI, ПВ/МНО, ТВ, F1,2, фибриноген, АОТ, активность АТИФ, ТАТ, антиген ИАП-1, активность ИПТФ, активность ТАП и активность ффВ.
Включение посредством ссылки
Все литературные источники, цитированные в настоящем документе, включая патенты, патентные заявки, статьи, публикации, руководства и так далее, а также приведенные в них ссылки, даже если они не упомянуты самостоятельно, включены в настоящий документ посредством ссылки в полном объеме.
Эквиваленты
Приведенная выше письменная спецификация считается достаточной для того, чтобы специалист в данной области мог осуществлять на практике настоящее изобретение. В приведенной выше спецификации и примерах подробно описаны некоторые предпочтительные варианты осуществления изобретения и описаны наилучшие варианты осуществления, предусмотренные авторами изобретения. Однако следует понимать, что, как бы подробно изобретение не было описано, его можно осуществлять на практике множеством способов, и изобретение должно толковаться в соответствии с прилагаемой формулой изобретения и любыми ее эквивалентами.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> NOVARTIS AG
<120> АНТИТЕЛА К ФАКТОРУ XI И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> PAT056955-WO-PCT
<140>
<141>
<150> 62/341,568
<151> 2016-05-25
<150> 62/184,955
<151> 2015-06-26
<160> 51
<170> PatentIn version 3.5
<210> 1
<211> 625
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Ile Phe Leu Tyr Gln Val Val His Phe Ile Leu Phe Thr Ser Val
1. 5 10 15
Ser Gly Glu Cys Val Thr Gln Leu Leu Lys Asp Thr Cys Phe Glu Gly
20 25 30
Gly Asp Ile Thr Thr Val Phe Thr Pro Ser Ala Lys Tyr Cys Gln Val
35 40 45
Val Cys Thr Tyr His Pro Arg Cys Leu Leu Phe Thr Phe Thr Ala Glu
50 55 60
Ser Pro Ser Glu Asp Pro Thr Arg Trp Phe Thr Cys Val Leu Lys Asp
65 70 75 80
Ser Val Thr Glu Thr Leu Pro Arg Val Asn Arg Thr Ala Ala Ile Ser
85 90 95
Gly Tyr Ser Phe Lys Gln Cys Ser His Gln Ile Ser Ala Cys Asn Lys
100 105 110
Asp Ile Tyr Val Asp Leu Asp Met Lys Gly Ile Asn Tyr Asn Ser Ser
115 120 125
Val Ala Lys Ser Ala Gln Glu Cys Gln Glu Arg Cys Thr Asp Asp Val
130 135 140
His Cys His Phe Phe Thr Tyr Ala Thr Arg Gln Phe Pro Ser Leu Glu
145 150 155 160
His Arg Asn Ile Cys Leu Leu Lys His Thr Gln Thr Gly Thr Pro Thr
165 170 175
Arg Ile Thr Lys Leu Asp Lys Val Val Ser Gly Phe Ser Leu Lys Ser
180 185 190
Cys Ala Leu Ser Asn Leu Ala Cys Ile Arg Asp Ile Phe Pro Asn Thr
195 200 205
Val Phe Ala Asp Ser Asn Ile Asp Ser Val Met Ala Pro Asp Ala Phe
210 215 220
Val Ser Gly Arg Ile Cys Thr His His Pro Gly Cys Leu Phe Phe Thr
225 230 235 240
Phe Phe Ser Gln Glu Trp Pro Lys Glu Ser Gln Arg Asn Leu Cys Leu
245 250 255
Leu Lys Thr Ser Glu Ser Gly Leu Pro Ser Thr Arg Ile Lys Lys Ser
260 265 270
Lys Ala Leu Ser Gly Phe Ser Leu Gln Ser Cys Arg His Ser Ile Pro
275 280 285
Val Phe Cys His Ser Ser Phe Tyr His Asp Thr Asp Phe Leu Gly Glu
290 295 300
Glu Leu Asp Ile Val Ala Ala Lys Ser His Glu Ala Cys Gln Lys Leu
305 310 315 320
Cys Thr Asn Ala Val Arg Cys Gln Phe Phe Thr Tyr Thr Pro Ala Gln
325 330 335
Ala Ser Cys Asn Glu Gly Lys Gly Lys Cys Tyr Leu Lys Leu Ser Ser
340 345 350
Asn Gly Ser Pro Thr Lys Ile Leu His Gly Arg Gly Gly Ile Ser Gly
355 360 365
Tyr Thr Leu Arg Leu Cys Lys Met Asp Asn Glu Cys Thr Thr Lys Ile
370 375 380
Lys Pro Arg Ile Val Gly Gly Thr Ala Ser Val Arg Gly Glu Trp Pro
385 390 395 400
Trp Gln Val Thr Leu His Thr Thr Ser Pro Thr Gln Arg His Leu Cys
405 410 415
Gly Gly Ser Ile Ile Gly Asn Gln Trp Ile Leu Thr Ala Ala His Cys
420 425 430
Phe Tyr Gly Val Glu Ser Pro Lys Ile Leu Arg Val Tyr Ser Gly Ile
435 440 445
Leu Asn Gln Ser Glu Ile Lys Glu Asp Thr Ser Phe Phe Gly Val Gln
450 455 460
Glu Ile Ile Ile His Asp Gln Tyr Lys Met Ala Glu Ser Gly Tyr Asp
465 470 475 480
Ile Ala Leu Leu Lys Leu Glu Thr Thr Val Asn Tyr Thr Asp Ser Gln
485 490 495
Arg Pro Ile Cys Leu Pro Ser Lys Gly Asp Arg Asn Val Ile Tyr Thr
500 505 510
Asp Cys Trp Val Thr Gly Trp Gly Tyr Arg Lys Leu Arg Asp Lys Ile
515 520 525
Gln Asn Thr Leu Gln Lys Ala Lys Ile Pro Leu Val Thr Asn Glu Glu
530 535 540
Cys Gln Lys Arg Tyr Arg Gly His Lys Ile Thr His Lys Met Ile Cys
545 550 555 560
Ala Gly Tyr Arg Glu Gly Gly Lys Asp Ala Cys Lys Gly Asp Ser Gly
565 570 575
Gly Pro Leu Ser Cys Lys His Asn Glu Val Trp His Leu Val Gly Ile
580 585 590
Thr Ser Trp Gly Glu Gly Cys Ala Gln Arg Glu Arg Pro Gly Val Tyr
595 600 605
Thr Asn Val Val Glu Tyr Val Asp Trp Ile Leu Glu Lys Thr Gln Ala
610 615 620
Val
625
<210> 2
<211> 3278
<212> ДНК
<213> Homo sapiens
<400> 2
aggcacacag gcaaaatcaa gttctacatc tgtccctgtg tatgtcactt gtttgaatac 60
gaaataaaat taaaaaaata aattcagtgt attgagaaag caagcaattc tctcaaggta 120
tatttctgac atactaagat tttaacgact ttcacaaata tgctgtactg agagagaatg 180
ttacataaca ttgagaacta gtacaagtaa atattaaagt gaagtgacca tttcctacac 240
aagctcattc agaggaggat gaagaccatt ttggaggaag aaaagcaccc ttattaagaa 300
ttgcagcaag taagccaaca aggtcttttc aggatgattt tcttatatca agtggtacat 360
ttcattttat ttacttcagt ttctggtgaa tgtgtgactc agttgttgaa ggacacctgc 420
tttgaaggag gggacattac tacggtcttc acaccaagcg ccaagtactg ccaggtagtc 480
tgcacttacc acccaagatg tttactcttc actttcacgg cggaatcacc atctgaggat 540
cccacccgat ggtttacttg tgtcctgaaa gacagtgtta cagaaacact gccaagagtg 600
aataggacag cagcgatttc tgggtattct ttcaagcaat gctcacacca aataagcgct 660
tgcaacaaag acatttatgt ggacctagac atgaagggca taaactataa cagctcagtt 720
gccaagagtg ctcaagaatg ccaagaaaga tgcacggatg acgtccactg ccactttttc 780
acgtacgcca caaggcagtt tcccagcctg gagcatcgta acatttgtct actgaagcac 840
acccaaacag ggacaccaac cagaataacg aagctcgata aagtggtgtc tggattttca 900
ctgaaatcct gtgcactttc taatctggct tgtattaggg acattttccc taatacggtg 960
tttgcagaca gcaacatcga cagtgtcatg gctcccgatg cttttgtctg tggccgaatc 1020
tgcactcatc atcccggttg cttgtttttt accttctttt cccaggaatg gcccaaagaa 1080
tctcaaagaa atctttgtct ccttaaaaca tctgagagtg gattgcccag tacacgcatt 1140
aaaaagagca aagctctttc tggtttcagt ctacaaagct gcaggcacag catcccagtg 1200
ttctgccatt cttcatttta ccatgacact gatttcttgg gagaagaact ggatattgtt 1260
gctgcaaaaa gtcacgaggc ctgccagaaa ctgtgcacca atgccgtccg ctgccagttt 1320
tttacctata ccccagccca agcatcctgc aacgaaggga agggcaagtg ttacttaaag 1380
ctttcttcaa acggatctcc aactaaaata cttcacggga gaggaggcat ctctggatac 1440
acattaaggt tgtgtaaaat ggataatgag tgtaccacca aaatcaagcc caggatcgtt 1500
ggaggaactg cgtctgttcg tggtgagtgg ccgtggcagg tgaccctgca cacaacctca 1560
cccactcaga gacacctgtg tggaggctcc atcattggaa accagtggat attaacagcc 1620
gctcactgtt tctatggggt agagtcacct aagattttgc gtgtctacag tggcatttta 1680
aatcaatctg aaataaaaga ggacacatct ttctttgggg ttcaagaaat aataatccat 1740
gatcagtata aaatggcaga aagcgggtat gatattgcct tgttgaaact ggaaaccaca 1800
gtgaattaca cagattctca acgacccata tgcctgcctt ccaaaggaga tagaaatgta 1860
atatacactg attgctgggt gactggatgg gggtacagaa aactaagaga caaaatacaa 1920
aatactctcc agaaagccaa gataccctta gtgaccaacg aagagtgcca gaagagatac 1980
agaggacata aaataaccca taagatgatc tgtgccggct acagggaagg agggaaggac 2040
gcttgcaagg gagattcggg aggccctctg tcctgcaaac acaatgaggt ctggcatctg 2100
gtaggcatca cgagctgggg cgaaggctgt gctcaaaggg agcggccagg tgtttacacc 2160
aacgtggtcg agtacgtgga ctggattctg gagaaaactc aagcagtgtg aatgggttcc 2220
caggggccat tggagtccct gaaggaccca ggatttgctg ggagagggtg ttgagttcac 2280
tgtgccagca tgcttcctcc acagtaacac gctgaagggg cttggtgttt gtaagaaaat 2340
gctagaagaa aacaaactgt cacaagttgt tatgtccaaa actcccgttc tatgatcgtt 2400
gtagtttgtt tgagcattca gtctctttgt ttttgatcac gcttctatgg agtccaagaa 2460
ttaccataag gcaatatttc tgaagattac tatataggca gatatagcag aaaataacca 2520
agtagtggca gtggggatca ggcagaagaa ctggtaaaag aagccaccat aaatagattt 2580
gttcgatgaa agatgaaaac tggaagaaag gagaacaaag acagtcttca ccattttgca 2640
ggaatctaca ctctgcctat gtgaacacat ttcttttgta aagaaagaaa ttgattgcat 2700
ttaatggcag attttcagaa tagtcaggaa ttcttgtcat ttccatttta aaatatatat 2760
taaaaaaaat cagttcgagt agacacgagc taagagtgaa tgtgaagata acagaatttc 2820
tgtgtggaag aggattacaa gcagcaattt acctggaagt gataccttag gggcaatctt 2880
gaagatacac tttcctgaaa aatgatttgt gatggattgt atatttattt aaaatatctt 2940
gggaggggag gctgatggag atagggagca tgctcaaacc tccctaagac aagctgctgc 3000
tgtgactatg ggctcccaaa gagctagatc gtatatttat ttgacaaaaa tcaccataga 3060
ctgcatccat actacagaga aaaaacaatt agggcgcaaa tggatagtta cagtaaagtc 3120
ttcagcaagc agctgcctgt attctaagca ctgggatttt ctgtttcgtg caaatattta 3180
tctcattatt gttgtgatct agttcaataa cctagaattt gaattgtcac cacatagctt 3240
tcaatctgtg ccaacaacta tacaattcat caagtgtg 3278
<210> 3
<211> 5
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 3
Thr Ala Ala Met Ser
1. 5
<210> 4
<211> 17
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 4
Gly Ile Ser Gly Ser Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1. 5 10 15
Gly
<210> 5
<211> 13
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 5
Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr
1. 5 10
<210> 6
<211> 7
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 6
Gly Phe Thr Phe Ser Thr Ala
1. 5
<210> 7
<211> 6
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 7
Ser Gly Ser Gly Ser Ser
1. 5
<210> 8
<211> 13
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 8
Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr
1. 5 10
<210> 9
<211> 122
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 9
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Ala
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Ser Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 10
<211> 366
<212> ДНК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИНУКЛЕОТИД"
<400> 10
caggtgcaat tgctggaaag cggcggtggc ctggtgcagc cgggtggcag cctgcgtctg 60
agctgcgcgg cgtccggatt caccttttct actgctgcta tgtcttgggt gcgccaggcc 120
ccgggcaaag gtctcgagtg ggtttccggt atctctggtt ctggttcttc tacctactat 180
gcggatagcg tgaaaggccg ctttaccatc agccgcgata attcgaaaaa caccctgtat 240
ctgcaaatga acagcctgcg tgcggaagat acggccgtgt attattgcgc gcgtgaactg 300
tcttacctgt actctggtta ctacttcgat tactggggcc aaggcaccct ggtgactgtt 360
agctca 366
<210> 11
<211> 452
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 11
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Ala
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Ser Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 12
<211> 1356
<212> ДНК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИНУКЛЕОТИД"
<400> 12
caggtgcaat tgctggaaag cggcggtggc ctggtgcagc cgggtggcag cctgcgtctg 60
agctgcgcgg cgtccggatt caccttttct actgctgcta tgtcttgggt gcgccaggcc 120
ccgggcaaag gtctcgagtg ggtttccggt atctctggtt ctggttcttc tacctactat 180
gcggatagcg tgaaaggccg ctttaccatc agccgcgata attcgaaaaa caccctgtat 240
ctgcaaatga acagcctgcg tgcggaagat acggccgtgt attattgcgc gcgtgaactg 300
tcttacctgt actctggtta ctacttcgat tactggggcc aaggcaccct ggtgactgtt 360
agctcagcct ccaccaaggg tccatcggtc ttccccctgg caccctcctc caagagcacc 420
tctgggggca cagcggccct gggctgcctg gtcaaggact acttccccga accggtgacg 480
gtgtcgtgga actcaggcgc cctgaccagc ggcgtgcaca ccttcccggc tgtcctacag 540
tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcag cttgggcacc 600
cagacctaca tctgcaacgt gaatcacaag cccagcaaca ccaaggtgga caagagagtt 660
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaagcagcg 720
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 780
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 840
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 900
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 960
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 1020
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 1080
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 1140
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1200
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1260
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1320
tacacgcaga agagcctctc cctgtctccg ggtaaa 1356
<210> 13
<211> 13
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 13
Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Asp Val Ser
1. 5 10
<210> 14
<211> 7
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 14
Lys Asn Tyr Asn Arg Pro Ser
1. 5
<210> 15
<211> 11
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 15
Ser Ala Trp Asp Gln Arg Gln Phe Asp Val Val
1. 5 10
<210> 16
<211> 9
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 16
Ser Ser Ser Asn Ile Gly Ser Asn Asp
1. 5
<210> 17
<211> 3
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 17
Lys Asn Tyr
1
<210> 18
<211> 8
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 18
Trp Asp Gln Arg Gln Phe Asp Val
1. 5
<210> 19
<211> 110
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 19
Asp Ile Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1. 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Asp Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Lys Asn Tyr Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ala Trp Asp Gln Arg Gln
85 90 95
Phe Asp Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 20
<211> 330
<212> ДНК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИНУКЛЕОТИД"
<400> 20
gatatcgtgc tgacccagcc gccgagcgtg agcggtgcac cgggccagcg cgtgaccatt 60
agctgtagcg gcagcagcag caacattggt tctaacgacg tgtcttggta ccagcagctg 120
ccgggcacgg cgccgaaact gctgatctac aaaaactaca accgcccgag cggcgtgccg 180
gatcgcttta gcggatccaa aagcggcacc agcgccagcc tggcgattac cggcctgcaa 240
gcagaagacg aagcggatta ttactgctct gcttgggacc agcgtcagtt cgacgttgtg 300
tttggcggcg gcacgaagtt aaccgtccta 330
<210> 21
<211> 216
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 21
Asp Ile Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1. 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Asp Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Lys Asn Tyr Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ala Trp Asp Gln Arg Gln
85 90 95
Phe Asp Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 22
<211> 648
<212> ДНК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИНУКЛЕОТИД"
<400> 22
gatatcgtgc tgacccagcc gccgagcgtg agcggtgcac cgggccagcg cgtgaccatt 60
agctgtagcg gcagcagcag caacattggt tctaacgacg tgtcttggta ccagcagctg 120
ccgggcacgg cgccgaaact gctgatctac aaaaactaca accgcccgag cggcgtgccg 180
gatcgcttta gcggatccaa aagcggcacc agcgccagcc tggcgattac cggcctgcaa 240
gcagaagacg aagcggatta ttactgctct gcttgggacc agcgtcagtt cgacgttgtg 300
tttggcggcg gcacgaagtt aaccgtccta ggtcagccca aggctgcccc ctcggtcact 360
ctgttcccgc cctcctctga ggagcttcaa gccaacaagg ccacactggt gtgtctcata 420
agtgacttct acccgggagc cgtgacagtg gcctggaagg cagatagcag ccccgtcaag 480
gcgggagtgg agaccaccac accctccaaa caaagcaaca acaagtacgc ggccagcagc 540
tatctgagcc tgacgcctga gcagtggaag tcccacagaa gctacagctg ccaggtcacg 600
catgaaggga gcaccgtgga gaagacagtg gcccctacag aatgttca 648
<210> 23
<211> 5
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 23
Thr Ala Ala Met Ser
1. 5
<210> 24
<211> 17
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 24
Gly Ile Ser Gly Ser Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1. 5 10 15
Gly
<210> 25
<211> 13
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 25
Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr
1. 5 10
<210> 26
<211> 7
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 26
Gly Phe Thr Phe Ser Thr Ala
1. 5
<210> 27
<211> 6
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 27
Ser Gly Ser Gly Ser Ser
1. 5
<210> 28
<211> 13
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 28
Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr
1. 5 10
<210> 29
<211> 122
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 29
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Ala
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Ser Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 30
<211> 366
<212> ДНК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИНУКЛЕОТИД"
<400> 30
caggtgcagc tgctggaatc aggcggcgga ctggtgcagc ctggcggtag cctgagactg 60
agctgcgctg ctagtggctt cacctttagc accgccgcta tgagctgggt tcgacaggcc 120
ccagggaaag gcctcgagtg ggtctcaggg attagcggta gcggctctag cacctactac 180
gccgatagcg tgaagggccg gttcactatc tctagggata actctaagaa caccctgtac 240
ctgcagatga atagcctgag agccgaggac accgccgtct actactgcgc tagagagctg 300
agctacctgt atagcggcta ctacttcgac tactggggtc aaggcaccct ggtcaccgtg 360
tctagc 366
<210> 31
<211> 452
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 31
Gln Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Ala
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Ser Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 32
<211> 1356
<212> ДНК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИНУКЛЕОТИД"
<400> 32
caggtgcagc tgctggaatc aggcggcgga ctggtgcagc ctggcggtag cctgagactg 60
agctgcgctg ctagtggctt cacctttagc accgccgcta tgagctgggt tcgacaggcc 120
ccagggaaag gcctcgagtg ggtctcaggg attagcggta gcggctctag cacctactac 180
gccgatagcg tgaagggccg gttcactatc tctagggata actctaagaa caccctgtac 240
ctgcagatga atagcctgag agccgaggac accgccgtct actactgcgc tagagagctg 300
agctacctgt atagcggcta ctacttcgac tactggggtc aaggcaccct ggtcaccgtg 360
tctagcgcta gcactaaggg cccctccgtg ttccctctgg ccccttccag caagtctacc 420
tccggcggca cagctgctct gggctgcctg gtcaaggact acttccctga gcctgtgaca 480
gtgtcctgga actctggcgc cctgacctct ggcgtgcaca ccttccctgc cgtgctgcag 540
tcctccggcc tgtactccct gtcctccgtg gtcacagtgc cttcaagcag cctgggcacc 600
cagacctata tctgcaacgt gaaccacaag ccttccaaca ccaaggtgga caagcgggtg 660
gagcctaagt cctgcgacaa gacccacacc tgtcctccct gccctgctcc tgaactgctg 720
ggcggccctt ctgtgttcct gttccctcca aagcccaagg acaccctgat gatctcccgg 780
acccctgaag tgacctgcgt ggtggtggcc gtgtcccacg aggatcctga agtgaagttc 840
aattggtacg tggacggcgt ggaggtgcac aacgccaaga ccaagcctcg ggaggaacag 900
tacaactcca cctaccgggt ggtgtccgtg ctgaccgtgc tgcaccagga ctggctgaac 960
ggcaaagagt acaagtgcaa agtctccaac aaggccctgg ccgcccctat cgaaaagaca 1020
atctccaagg ccaagggcca gcctagggaa ccccaggtgt acaccctgcc acccagccgg 1080
gaggaaatga ccaagaacca ggtgtccctg acctgtctgg tcaagggctt ctacccttcc 1140
gatatcgccg tggagtggga gtctaacggc cagcctgaga acaactacaa gaccacccct 1200
cctgtgctgg actccgacgg ctccttcttc ctgtactcca aactgaccgt ggacaagtcc 1260
cggtggcagc agggcaacgt gttctcctgc tccgtgatgc acgaggccct gcacaaccac 1320
tacacccaga agtccctgtc cctgtctccc ggcaag 1356
<210> 33
<211> 13
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 33
Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Asp Val Ser
1. 5 10
<210> 34
<211> 7
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 34
Lys Asn Tyr Asn Arg Pro Ser
1. 5
<210> 35
<211> 11
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 35
Ser Ala Trp Asp Gln Arg Gln Phe Asp Val Val
1. 5 10
<210> 36
<211> 9
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 36
Ser Ser Ser Asn Ile Gly Ser Asn Asp
1. 5
<210> 37
<211> 3
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 37
Lys Asn Tyr
1
<210> 38
<211> 8
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 38
Trp Asp Gln Arg Gln Phe Asp Val
1. 5
<210> 39
<211> 110
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 39
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1. 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Asp Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Lys Asn Tyr Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ala Trp Asp Gln Arg Gln
85 90 95
Phe Asp Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 40
<211> 330
<212> ДНК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИНУКЛЕОТИД"
<400> 40
cagtcagtcc tgactcagcc ccctagcgct agtggcaccc ctggtcaaag agtgactatt 60
agctgtagcg gctctagctc taatatcggc tctaacgacg tcagctggta tcagcagctg 120
cccggcaccg cccctaagct gctgatctat aagaactata ataggcctag cggcgtgccc 180
gataggttta gcggatctaa atcagggact tctgctagtc tggctattag cggcctgcag 240
tcagaggacg aggccgacta ctactgtagc gcctgggatc agcgtcagtt cgacgtggtg 300
ttcggcggag gcactaagct gaccgtgctg 330
<210> 41
<211> 216
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 41
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1. 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Asp Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Lys Asn Tyr Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ala Trp Asp Gln Arg Gln
85 90 95
Phe Asp Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 42
<211> 648
<212> ДНК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИНУКЛЕОТИД"
<400> 42
cagtcagtcc tgactcagcc ccctagcgct agtggcaccc ctggtcaaag agtgactatt 60
agctgtagcg gctctagctc taatatcggc tctaacgacg tcagctggta tcagcagctg 120
cccggcaccg cccctaagct gctgatctat aagaactata ataggcctag cggcgtgccc 180
gataggttta gcggatctaa atcagggact tctgctagtc tggctattag cggcctgcag 240
tcagaggacg aggccgacta ctactgtagc gcctgggatc agcgtcagtt cgacgtggtg 300
ttcggcggag gcactaagct gaccgtgctg ggtcaaccta aggctgcccc cagcgtgacc 360
ctgttccccc ccagcagcga ggagctgcag gccaacaagg ccaccctggt gtgcctgatc 420
agcgacttct acccaggcgc cgtgaccgtg gcctggaagg ccgacagcag ccccgtgaag 480
gccggcgtgg agaccaccac ccccagcaag cagagcaaca acaagtacgc cgccagcagc 540
tacctgagcc tgacccccga gcagtggaag agccacaggt cctacagctg ccaggtgacc 600
cacgagggca gcaccgtgga aaagaccgtg gccccaaccg agtgcagc 648
<210> 43
<211> 8
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 43
Gly Phe Thr Phe Ser Thr Ala Ala
1. 5
<210> 44
<211> 8
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 44
Ile Ser Gly Ser Gly Ser Ser Thr
1. 5
<210> 45
<211> 15
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 45
Ala Arg Glu Leu Ser Tyr Leu Tyr Ser Gly Tyr Tyr Phe Asp Tyr
1. 5 10 15
<210> 46
<211> 10
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 46
Gly Phe Thr Phe Ser Thr Ala Ala Met Ser
1. 5 10
<210> 47
<211> 8
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 47
Ser Ser Asn Ile Gly Ser Asn Asp
1. 5
<210> 48
<211> 6
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
6xHis tag"
<400> 48
His His His His His His
1. 5
<210> 49
<211> 4
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПЕПТИД"
<400> 49
Met Gly Ser Ser
1
<210> 50
<211> 8
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
8xHis tag"
<400> 50
His His His His His His His His
1. 5
<210> 51
<211> 238
<212> БЕЛОК
<213> ИСКУССТВЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ
<220>
<221> ИСТОЧНИК
<223> /ПРИМЕЧАНИЕ="ОПИСАНИЕ ИСКУССТВЕННОЙ ПОСЛЕДОВАТЕЛЬНОСТИ: СИНТЕТИЧЕСКИЙ
ПОЛИПЕПТИД"
<400> 51
Ile Val Gly Gly Thr Ala Ser Val Arg Gly Glu Trp Pro Trp Gln Val
1. 5 10 15
Thr Leu His Thr Thr Ser Pro Thr Gln Arg His Leu Cys Gly Gly Ser
20 25 30
Ile Ile Gly Asn Gln Trp Ile Leu Thr Ala Ala His Cys Phe Tyr Gly
35 40 45
Val Glu Ser Pro Lys Ile Leu Arg Val Tyr Ser Gly Ile Leu Asn Gln
50 55 60
Ser Glu Ile Lys Glu Asp Thr Ser Phe Phe Gly Val Gln Glu Ile Ile
65 70 75 80
Ile His Asp Gln Tyr Lys Met Ala Glu Ser Gly Tyr Asp Ile Ala Leu
85 90 95
Leu Lys Leu Glu Thr Thr Val Asn Tyr Thr Asp Ser Gln Arg Pro Ile
100 105 110
Cys Leu Pro Ser Lys Gly Asp Arg Asn Val Ile Tyr Thr Asp Cys Trp
115 120 125
Val Thr Gly Trp Gly Tyr Arg Lys Leu Arg Asp Lys Ile Gln Asn Thr
130 135 140
Leu Gln Lys Ala Lys Ile Pro Leu Val Thr Asn Glu Glu Cys Gln Lys
145 150 155 160
Arg Tyr Arg Gly His Lys Ile Thr His Lys Met Ile Cys Ala Gly Tyr
165 170 175
Arg Glu Gly Gly Lys Asp Ala Cys Lys Gly Asp Ser Gly Gly Pro Leu
180 185 190
Ser Cys Lys His Asn Glu Val Trp His Leu Val Gly Ile Thr Ser Trp
195 200 205
Gly Glu Gly Cys Ala Gln Arg Glu Arg Pro Gly Val Tyr Thr Asn Val
210 215 220
Val Glu Tyr Val Asp Trp Ile Leu Glu Lys Thr Gln Ala Val
225 230 235
2
1
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Антитело против FXI/FXIA, его антигенсвязывающий фрагмент и фармацевтическое применение | 2021 |
|
RU2838408C1 |
| НОВЫЕ АНТИТЕЛА ПРОТИВ ФАКТОРА XI И ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2758160C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА СВЕРТЫВАНИЯ XI | 2017 |
|
RU2757314C2 |
| НОВЫЕ ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К ФАКТОРУ XI С АНТИТРОМБОТИЧЕСКИМ И ПРОТИВОВОСПАЛИТЕЛЬНЫМ ДЕЙСТВИЕМ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2817219C2 |
| АНТИТЕЛА, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ В-КЛЕТОК, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2766094C2 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| АНТИ-DR5-АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2735956C1 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ ИНТЕРЛЕЙКИНУ-2 И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2745451C1 |
| НОВЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1(PD-1) | 2016 |
|
RU2757316C2 |
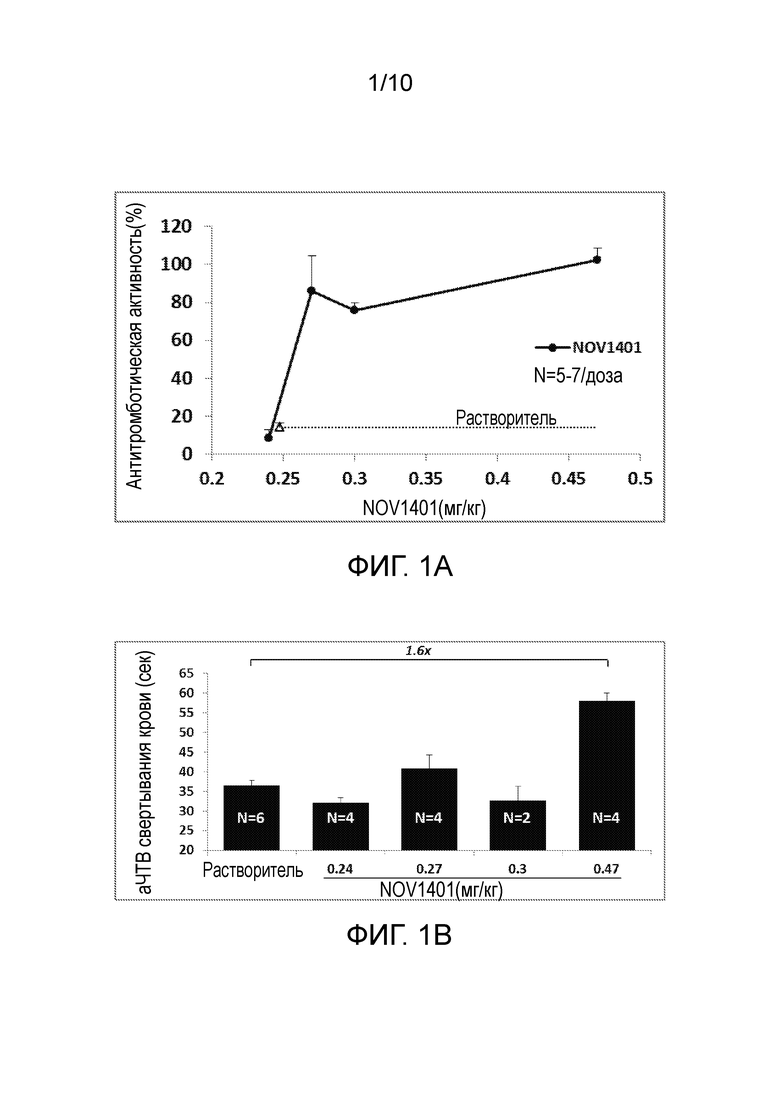
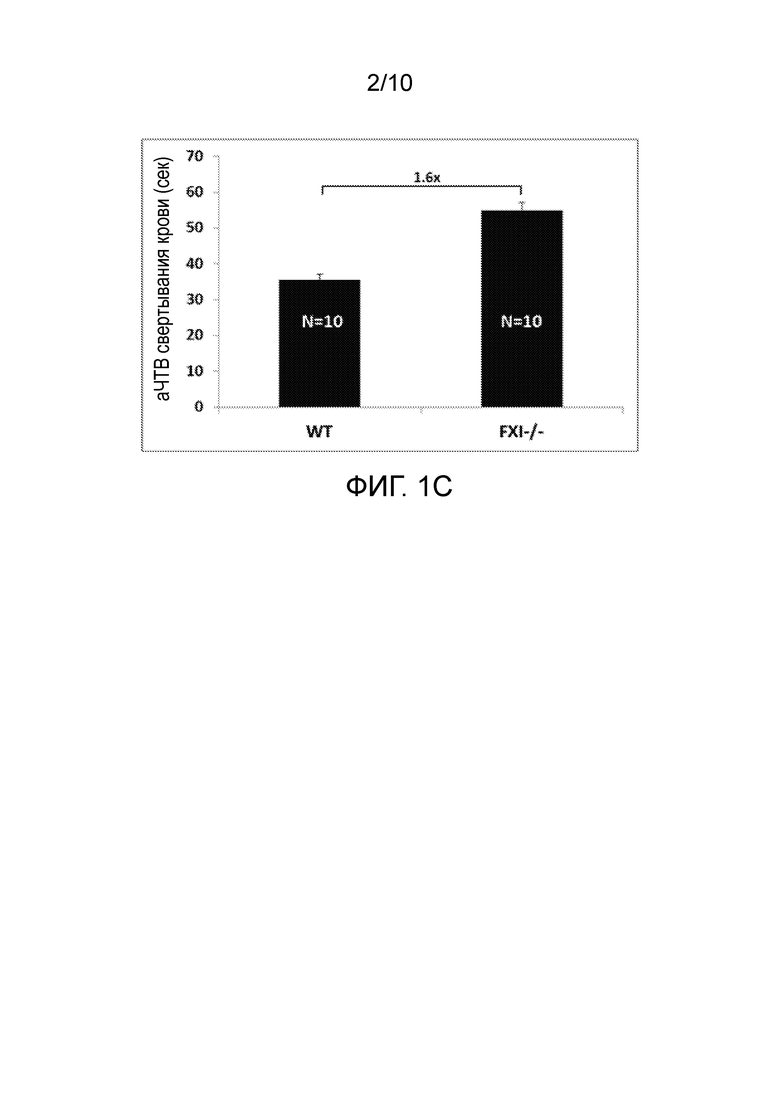
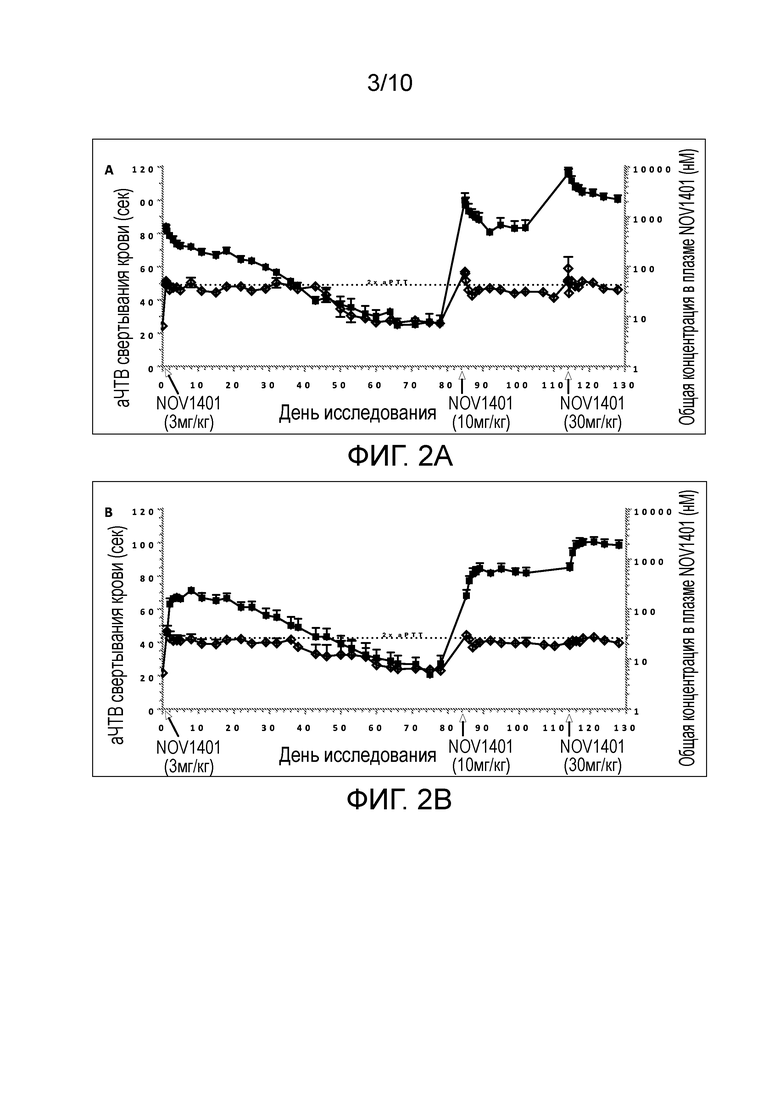
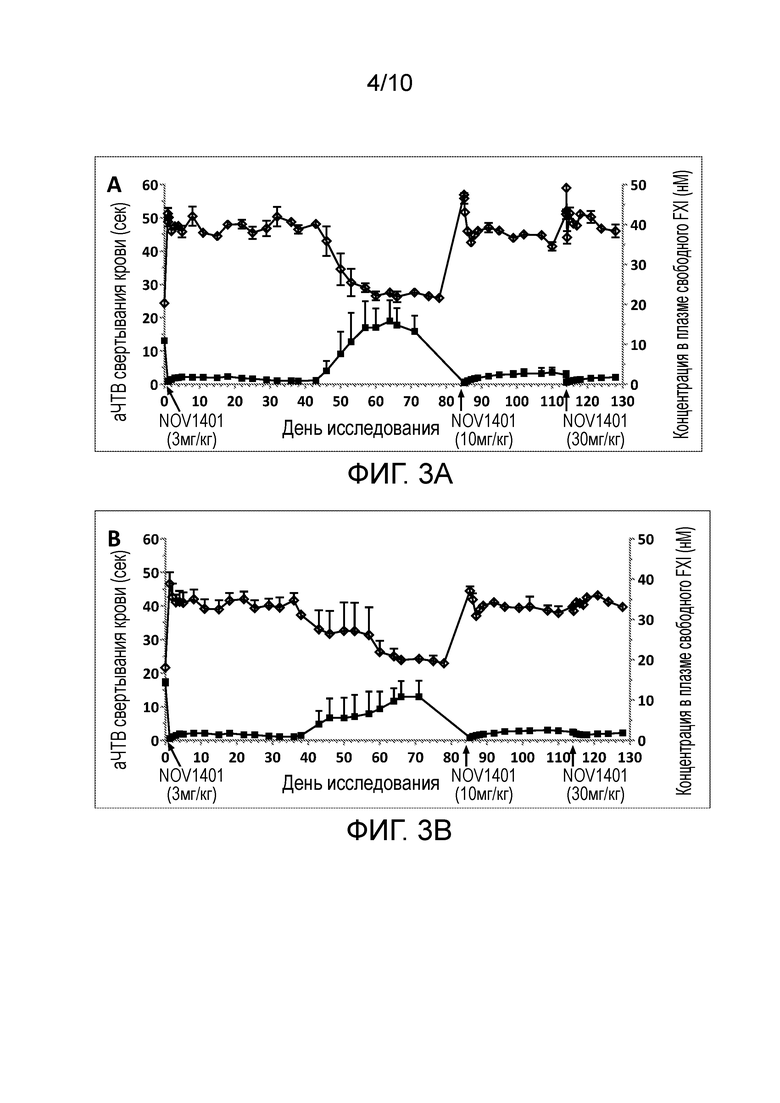
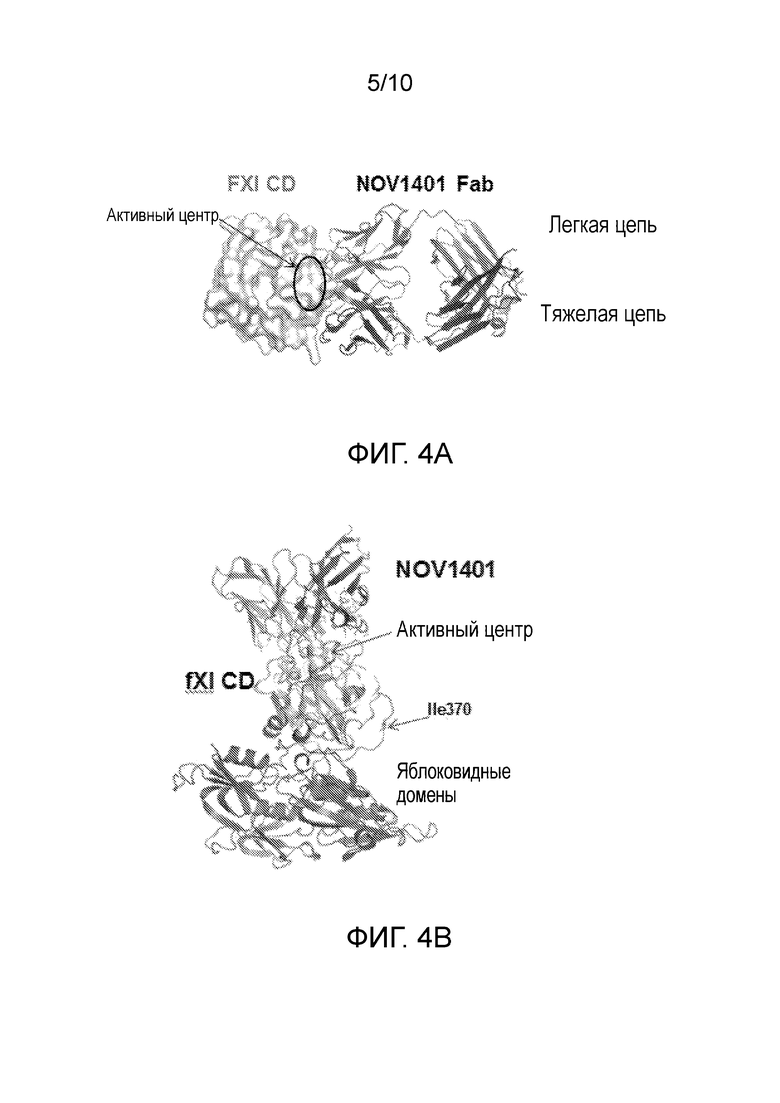
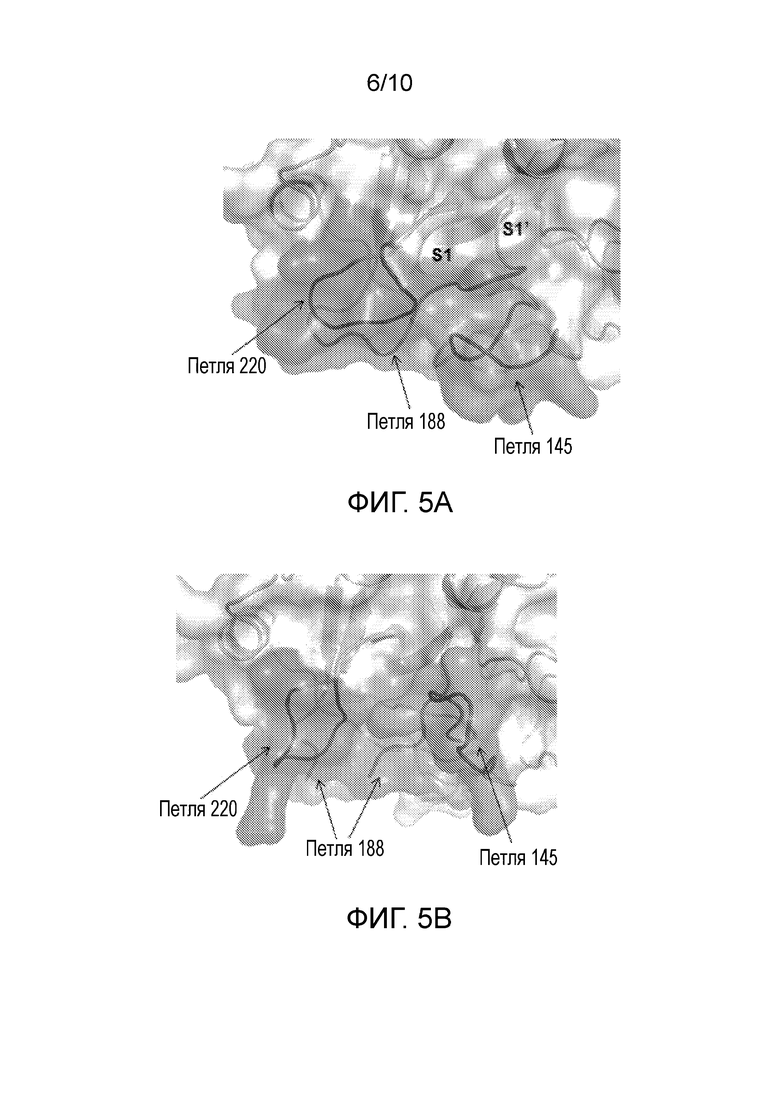
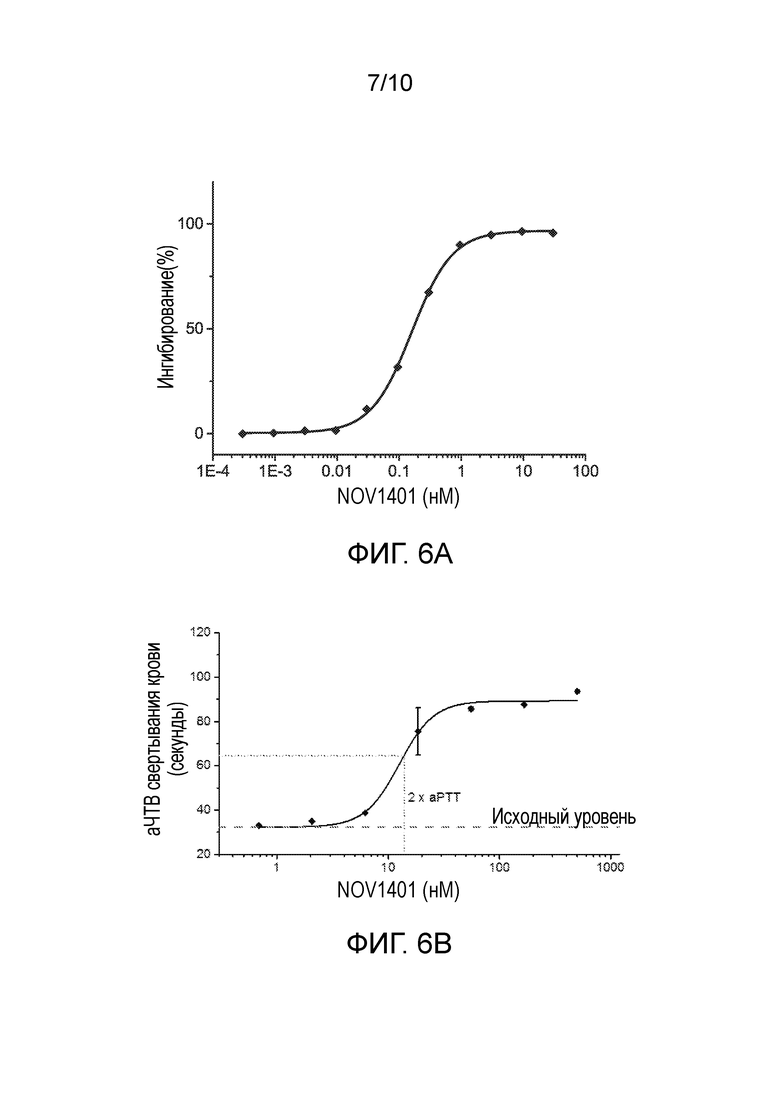
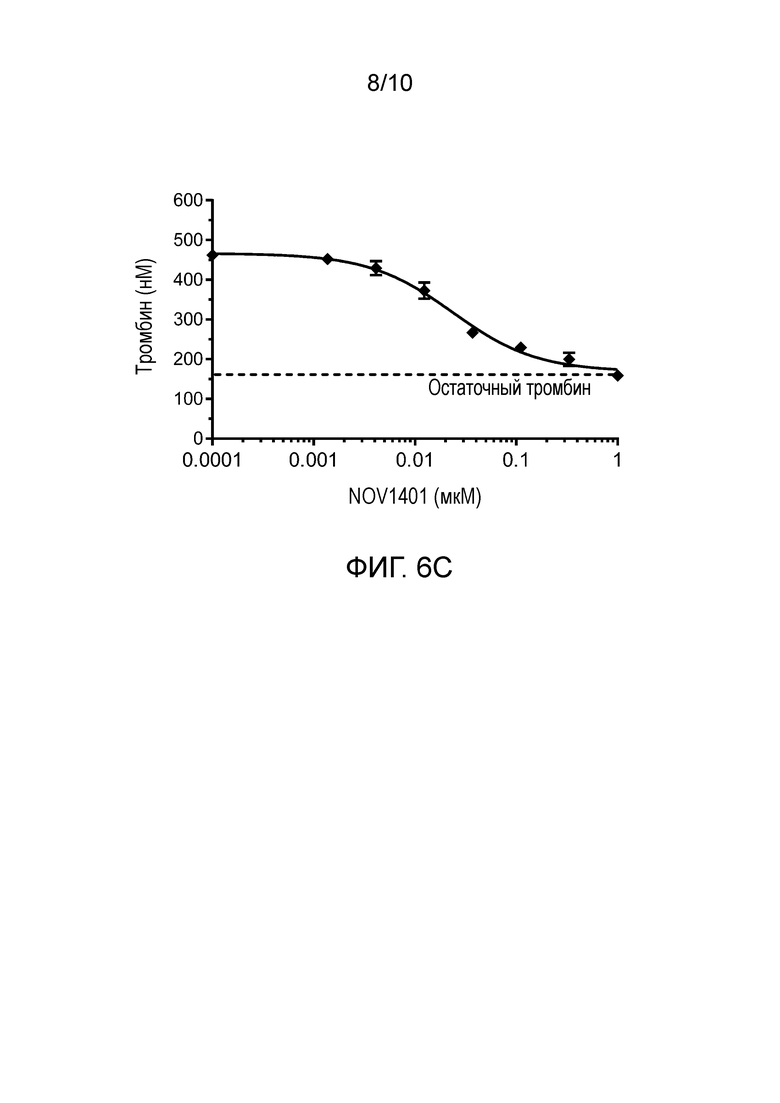
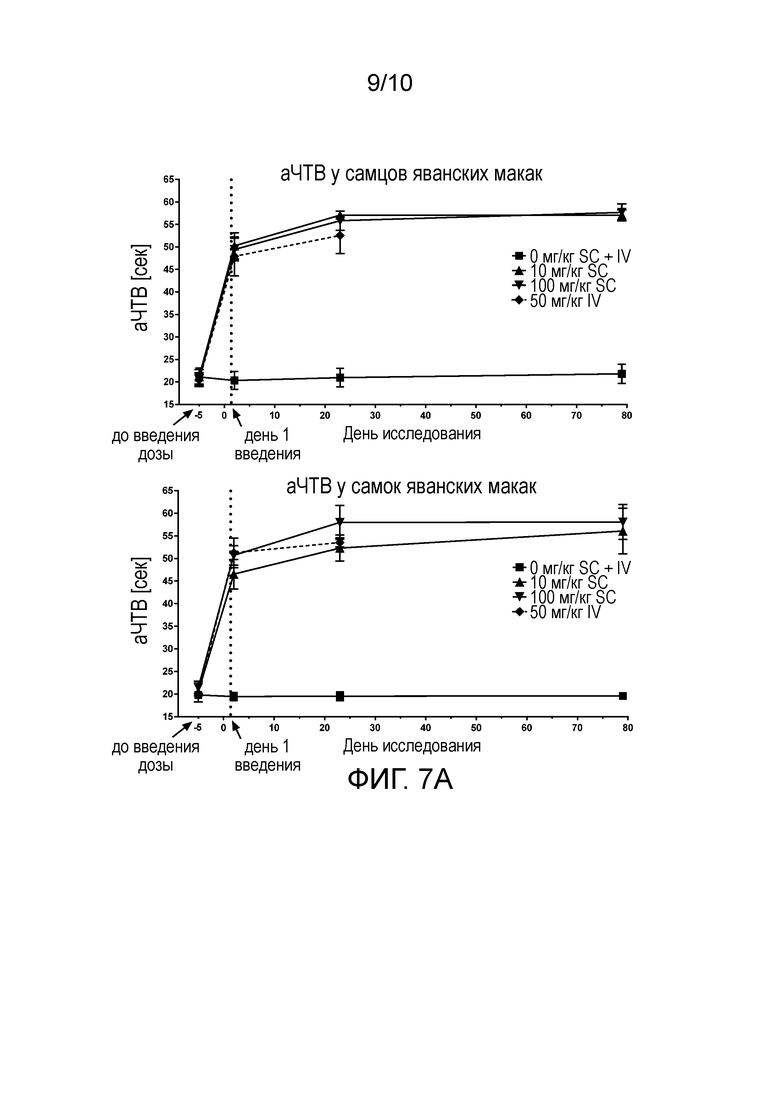
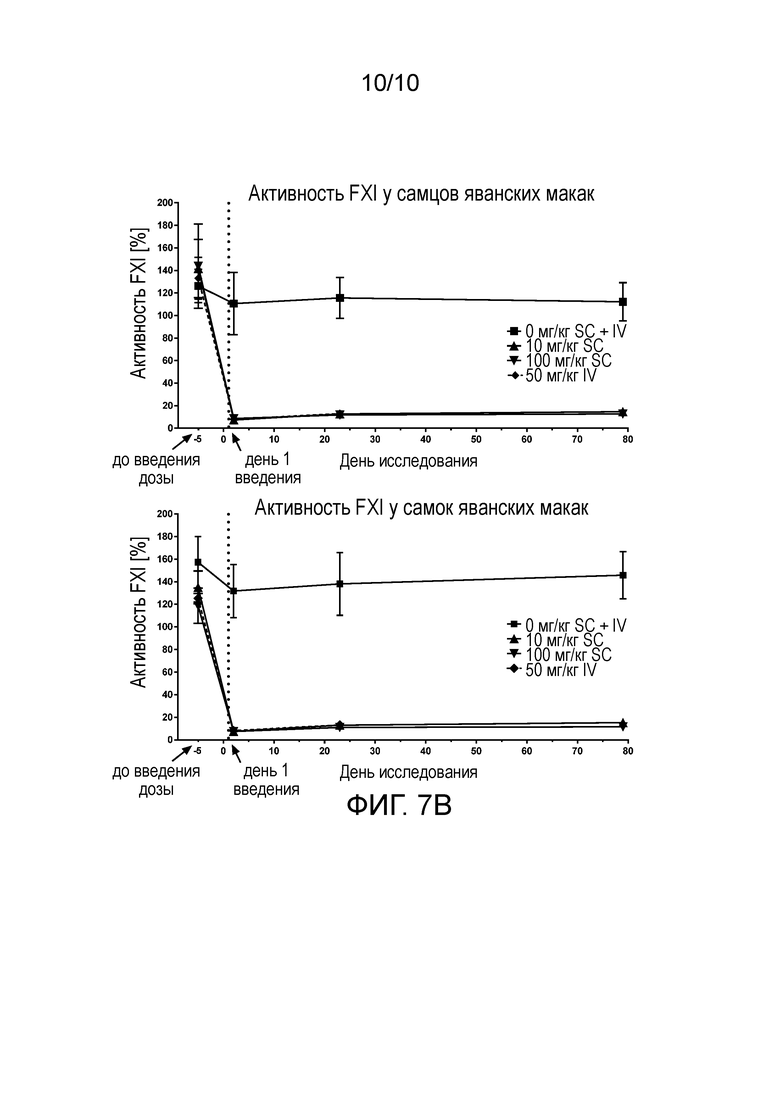
Изобретение относится к области биотехнологии. Предложены моноклональное антитело и его антигенсвязывающий фрагмент, которые связывают человеческий фактор XI. Также предложены кодирующая указанное антитело нуклеиновая кислота, экспрессионный вектор, клетка-хозяин для продуцирования антитела. Кроме того, изобретение относится к фармацевтическим композициям и способам лечения тромбоэмболического заболевания с применением анти-фактор XI антитела. Изобретение может быть использовано для эффективного лечения тромбоэмболического заболевания с уменьшенным риском кровотечений. 10 н. и 10 з.п. ф-лы, 16 ил., 12 табл., 11 пр.
1. Выделенное анти-фактор XI (FXI) антитело или его антигенсвязывающий фрагмент, при этом антитело или антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи (VH), содержащую определяющие комплементарность области HCDR1, HCDR2 и HCDR3, из SEQ ID NO: 9 или 29; и вариабельную область легкой цепи (VL), содержащую определяющие комплементарность области LCDR1, LCDR2, LCDR3, из SEQ ID NO: 19 или 39.
2. Выделенное антитело или его антигенсвязывающий фрагмент по п. 1, при этом антитело или антигенсвязывающий фрагмент содержат:
i. CDR1 вариабельной области тяжелой цепи SEQ ID NO: 23; CDR2 вариабельной области тяжелой цепи SEQ ID NO: 24; CDR3 вариабельной области тяжелой цепи SEQ ID NO: 25; CDR1 вариабельной области легкой цепи SEQ ID NO: 33; CDR2 вариабельной области легкой цепи SEQ ID NO: 34 и CDR3 вариабельной области легкой цепи SEQ ID NO: 35;
ii. CDR1 вариабельной области тяжелой цепи SEQ ID NO: 26; CDR2 вариабельной области тяжелой цепи SEQ ID NO: 27; CDR3 вариабельной области тяжелой цепи SEQ ID NO: 28; CDR1 вариабельной области легкой цепи SEQ ID NO: 36; CDR2 вариабельной области легкой цепи SEQ ID NO: 37 и CDR3 вариабельной области легкой цепи SEQ ID NO: 38;
iii. CDR1 вариабельной области тяжелой цепи SEQ ID NO: 43; CDR2 вариабельной области тяжелой цепи SEQ ID NO: 44; CDR3 вариабельной области тяжелой цепи SEQ ID NO: 45; CDR1 вариабельной области легкой цепи SEQ ID NO: 47; CDR2 вариабельной области легкой цепи SEQ ID NO: 37 и CDR3 вариабельной области легкой цепи SEQ ID NO: 15; или
iv. CDR1 вариабельной области тяжелой цепи SEQ ID NO: 46; CDR2 вариабельной области тяжелой цепи SEQ ID NO: 4; CDR3 вариабельной области тяжелой цепи SEQ ID NO: 5; CDR1 вариабельной области легкой цепи SEQ ID NO: 33; CDR2 вариабельной области легкой цепи SEQ ID NO: 14 и CDR3 вариабельной области легкой цепи SEQ ID NO: 15.
3. Выделенное антитело или антигенсвязывающий фрагмент по п. 1, при этом антитело или антигенсвязывающий фрагмент содержит последовательность вариабельной области тяжелой цепи (VH), выбранную из группы, состоящей из SEQ ID NO: 9 и 29; и последовательность вариабельной области легкой цепи (VL), выбранную из группы, состоящей из SEQ ID NO: 19 и 39.
4. Выделенное антитело или антигенсвязывающий фрагмент по п. 1, при этом антитело или антигенсвязывающий фрагмент содержат последовательность вариабельной области тяжелой цепи SEQ ID NO: 9 и последовательность вариабельной области легкой цепи SEQ ID NO: 19.
5. Выделенное антитело или антигенсвязывающий фрагмент по п. 1, где антитело или антигенсвязывающий фрагмент содержат последовательность вариабельной области тяжелой цепи SEQ ID NO: 29 и последовательность вариабельной области легкой цепи SEQ ID NO: 39.
6. Выделенное антитело или антигенсвязывающий фрагмент по п. 1, где антитело содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 31, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 41.
7. Выделенное антитело или антигенсвязывающий фрагмент по п. 1, где антитело содержит тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 11, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 21.
8. Выделенное антитело или антигенсвязывающий фрагмент по п. 1, при этом антитело представляет собой моноклональное антитело человека.
9. Выделенное антитело или антигенсвязывающий фрагмент по п. 1, при этом антитело представляет собой человеческое или гуманизированное антитело изотипа IgG1.
10. Фармацевтическая композиция для лечения тромбоэмболического заболевания, содержащая эффективное количество антитела или его антигенсвязывающего фрагмента по любому из предшествующих пп. 1-9 и фармацевтически приемлемый носитель.
11. Способ лечения тромбоэмболического заболевания, включающий введение субъекту, страдающему тромбоэмболическим заболеванием, эффективного количества фармацевтической композиции, содержащей антитело или его антигенсвязывающий фрагмент по любому из пп. 1-9.
12. Способ по п. 11, отличающийся тем, что субъект страдает одним или более из ишемического инсульта, связанного с фибрилляцией предсердий, и тромбоза глубоких вен.
13. Способ по п. 11, отличающийся тем, что субъект страдает ишемическим инсультом, связанным с фибрилляцией предсердий.
14. Способ лечения тромбоэмболического заболевания, включающий введение субъекту, страдающему тромбоэмболическим заболеванием, эффективного количества фармацевтической композиции, содержащей антитело или фрагмент по любому из пп. 1-9, в сочетании с терапией статинами.
15. Лекарственное средство для лечения тромбоэмболического заболевания, содержащее антитело по любому из пп. 1-9.
16. Нуклеиновая кислота, кодирующая антитело по любому из пп. 1-9.
17. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 16.
18. Клетка-хозяин для продуцирования антитела, содержащая экспрессионный вектор по п. 17.
19. Применение антитела или антигенсвязывающего фрагмента по любому из пп. 1-9 для производства лекарственного средства для лечения тромбоэмболического заболевания.
20. Применение антитела или антигенсвязывающего фрагмента по любому из пп. 1-9 для производства лекарственного средства для лечения тромбоэмболического заболевания в сочетании с терапией статинами.
| WO 2013167669 A1, 14.11.2013 | |||
| WO 2010080623 A2, 15.07.2010 | |||
| WO 2009067660 A2, 28.05.2009 | |||
| MATAFONOV A | |||
| et al., Evidence for factor IX-independent roles for factor XIa in blood coagulation, JOURNAL OF THROMBOSIS AND HAEMOSTASIS, 2013, vol.11, no.12, pp.2118-2127 | |||
| КОЛЯДКО В.Н | |||
| и др., Молекулярные механизмы тромбоза | |||
| Фундаментальные и прикладные |

