ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к адъюванту, полученному из лимфоцитов человека. Этот адъювант может быть использован в комбинации с традиционными вакцинами или в раковой иммунотерапии для усиления ответа иммунной системы пациента на вакцину или другой иммунотерапевтический агент.
УРОВЕНЬ ТЕХНИКИ
Иммунологические адъюванты используют в комбинации с вакцинами для увеличения иммунного ответа на антиген. Одним из способов функционирования иммунологического адъюванта является аттрактирование макрофагов к антигену, так что эти макрофаги могут презентировать антиген регионарным лимфатическим узлам и инициировать эффективный антигенный ответ. Адъюванты могут также сами действовать в качестве носителей для антигена или могут влиять на иммунный ответ другими механизмами, такими как создание депо-эффекта, индукция цитокинов, активация комплемента, рекрутинг различных клеточных популяций иммунологической системы, доставка антигена к различным антигенпрезентирующим клеткам, регуляция экспрессии молекул HLA класса I или класса II и стимуляция продуцирования различных подтипов антител. Многие более новые вакцины являются только слабо иммуногенными и, следовательно, требуют присутствия адъювантов.
Материалы, имеющие адъювантную активность, хорошо известны. Квасцы (Al(ОН)3 и сходные гели алюминия являются адъювантами, лицензированными для использования для человека. Адъювантная активность квасцов была впервые открыта в 1926 году Glenny (Chemistry and Industry, Jun.15, 1926; J.Path. Bacteriol, 34, 267). Гидроксид алюминия и фосфат алюминия (в целом обычно называемые квасцами) рутинным образом используются в качестве адъювантов в применяемых для человека и ветеринарных вакцинах. Эффективность квасцов в увеличении ответов в виде антител на дифтерийный и столбнячный токсоиды хорошо установлена, и более недавно вакцина HBsAg использовала квасцы в качестве адъюванта.
Известно, что другие материалы также имеют адъювантную активность, и они включают полный адъювант Фрейнда, эмульсию типа вода-в-минеральном масле, которая содержит убитые высушенные микобактерии в масляной фазе; неполный адъювант Фрейнда, более мягкая композиция без микобактерии; сапонин, мембраноактивный глюкозид, экстрагированный из дерева Quilla saponaria; неионогенные блок-сополимерные поверхностно-активные вещества, неметаболизируемые синтетические молекулы, которые имеют тенденцию связывать белки на клеточных поверхностях; ISCOM, липидные мицеллы, включающие Quil А (сапонин), который имитирует, в физическом смысле, инфекционные частицы; и мурамилдипептид, стимулирующую лейкоциты молекулу, которая является одним из наиболее активных компонентов убитых микобактерий. Известным адъювантом в раковой терапии является бацилла Кальметта-Герена (Bacillus Calmette-Guerin, BCG), которую используют в комбинации с различными противораковыми вакцинными стратегиями. Было обнаружено, что GM-CSF также является эффективным адъювантом при использовании в комбинации с аутологичными опухолевыми клетками.
Со всеми этими агентами токсичность, неприемлемые хронические реакции и/или низкая активность (в случае BCG) являются характерными чертами, которые ограничивают в настоящее время их применение в качестве потенциальных адъювантов. Таким образом, существует растущая и актуальная потребность в новых адъювантах для усиления иммунного ответа человека на вакцины, как в раковой терапии, так и при лечении других заболеваний.
Одна линия исследования в развитии адъювантов была направлена на исследование дендритных клеток. Дендритные клетки (ДК) являются профессиональными антигенпрезентирующими клетками (АПК), которые имеют уникальную способность инициировать первичные иммунные ответы in vivo и in vitro [1-3]. Они происходят из миелоидных (ДК1) или лимфоидных (ДК2) предшественников и распределены в их незрелой форме по всему телу в тканях, которые обычно сталкиваются с патогенами окружающей среды (коже, слизистых мембранах, эпителии кишечника и т.д.) [1, 2, 4-7]. В то время как ДК1 и ДК2 составляют небольшой процент от общего числа мононуклеарных клеток в периферическом кровотоке, предшественники ДК1 в форме CD14+/CD11с+/HLA-DR+ моноцитов являются относительно обильными, составляя приблизительно 10%-15% мононуклеарных клеток крови [11-15].
Незрелые ДК экспрессируют множество поверхностных структур, которые участвуют в приобретении антигена, активации/созревании ДК и презентации антигена [1, 2, 8]. Как только ДК сталкиваются с антигеном, они подвергаются процессу созревания, характеризующемуся увеличением экспрессии молекул HLA класса I и II, а также костимуляторных молекул и взаимодействуют с родственными рецепторами на Т- и В-лимфоцитах, приводя к генерированию антигенспецифических клеточных и гуморальных иммунных ответов [1, 2, 9, 10].
ДК считают первичными АПК в иммунной системе. Возможность выделения этих клеток и/или их предшественников и исследования их in vitro добавили значительный объем знания их роли во врожденном и приобретенном иммунитете [1, 2]. Классическим способом генерирования ДК человека in vitro является выделение и обогащение СD14+-моноцитов из периферической крови и культивирование их в течение различных периодов времени в GM-CSF и IL-4 с последующим конечным созреванием с рядом цитокинов, в том числе IL-2, IL-6, IL-7, IL-13, IL-15, TNFα, IL-1β [16, 36] или с различными другими агентами, в том числе липополисахаридами, PGE2, интерферонами типа 1 или двухцепочечной РНК [20, 24].
Многочисленные исследователи показали, что эти генерируемые in vitro, происходящие из моноцитов ДК, являются сильными антигенпрезентирующими клетками (АПК), способными инициировать первичный и вторичный антигенспецифический CD4+ и CD8+ Т-клеточные ответы [27-30]. Недавние исследования in vitro создали довольно большой объем информации в отношении биологии ДК1 и пролили свет на процессы, посредством которых антигенспецифические иммунные ответы генерируются in vivo [1-2]. В периферических тканях незрелые ДК получают антигенные материалы в контексте сигналов об опасности, инициирующих сложное окружение из цитокинов/хемокинов, которое генерируется ДК и другими типами клеток поблизости [31]. Растворимые медиаторы, продуцируемые ДК, могут действовать аутокринным или паракринным образом. Т-клетки продуцируют дополнительные цитокины и хемокины после взаимодействия с вооруженными антигеном ДК, как это делают и другие иммунные клетки, которые активируются высвобождаемыми цитокинами [32-35]. Эта сложная сеть взаимодействий может, в свою очередь, создавать окружающую среду, которая ускоряет генерирование ДК из их моноцитарных предшественников.
Несколько исследователей описали применение различных бесклеточных культуральных супернатантов, также называемых «кондиционированными средами», в качестве агентов созревания ДК. Эти среды содержат более или менее определенные смеси цитокинов [12, 25, 26]. Было показано, что кондиционированные моноцитами среды (МСМ), содержащие IFNα, IL-1β, IL-6 и TNFα, индуцируют экспрессию CD83 и р55, поверхностных молекул, которые являются характеристиками зрелых ДК [26]. Однако при добавлении комбинаций этих цитокинов к незрелым ДК в концентрациях, сравнимых с концентрациями, обнаруживаемыми в кондиционируемых средах, они были менее эффективными в созревании ДК по сравнению с МСМ. Эти результаты предполагают, что требовались дополнительные компоненты для осуществления полного созревания незрелых ДК.
В одном исследовании Kato и др. получали кондиционированные среды (названные ТССМ) культивированием выделенных Т-клеток с моноклональными антителами к CD3, которые были прикреплены к пластиковым поверхностям [25]. Эта среда была способна вызывать созревание ДК, которые были генерированы из моноцитов, культивируемых в GM-CSF и IL-4. Интересно, что различные клоны антител к CD3 индуцировали различные количества растворимого лиганда CD40 и IFNγ, и эти различия были отражены в способности этой среды вызывать созревание ДК.
В то время как МСМ и ТССМ являются очень эффективными медиаторами конечного созревания ДК, о их способности дифференцировать моноциты в незрелые ДК не сообщалась. Авторам настоящего изобретения известно только одно сообщение, в котором эту активность наблюдали со средами от РВМС, стимулированных олигонуклеотидами CpG-A [33]. Хорошо установлено, что CpG-A индуцирует продуцирование интерферонов типа 1 (IFNα/IFNβ) плазмацитоидными ДК, минорным клеточным компонентом в РВМС [6, 37-39]. В цитируемом исследовании антитела к IFNα уменьшали, но не аннулировали активность в этой культуральной среде, что предполагает, что дополнительные цитокины могут участвовать в индукции дифференцировки моноцитов. Это определенно является возможным, так как известно, что IFNa индуцирует продуцирование цитокинов в других типах клеток (в том числе Т-клетках), что может влиять на дифференцировку моноцитов [6, 38, 39].
Предполагается, что соединения или композиции, которые ускоряют созревание дендритных клеток при введении в комбинации с вакцинным антигеном, будут приводить к большему количеству антигенпрезентирующих клеток, презентирующих вакцинный антиген Т-лимфоцитам и В-клеткам, поддерживая посредством этого иммунный ответ на вакцинный антиген.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение решает вышеуказанную потребность обеспечением новых адъювантов, основанных на продуктах лимфоцитов человека, которые обеспечивают иммунологическое потенциирование и увеличение количества и качества иммунного ответа на вакцинные антигены.
В одном аспекте данного изобретения этот адъювант получают из супернатантного материала, собранного из in vitro стимулированных культивируемых мононуклеарных клеток периферической крови человека. "Необученные" Т-клетки активируются во время этого процесса культивирования. Этот адъювант действует с усилением способности вакцины к инициации, созданию, усилению и/или поддержанию иммунного ответа на агент у людей и других видов животных или растений.
Данное изобретение обеспечивает адъювант, основанный на смеси цитокинов и хемокинов, полученных из мононуклеарных клеток периферической крови, стимулированных гранулами, покрытыми антителами к CD3/CD28. В применении здесь термин "кондиционированная лимфоцитами среда (LCM)" будет использоваться для названия этого адъюванта. Было обнаружено, что LCM является высокоэффективной кондиционированной средой со способностью вызывать созревание происходящих из моноцитов ДК и превращать моноциты в сильные АПК. Этот адъювант может обеспечивать быстрый, экономичный и, возможно, более «физиологический» способ получения больших количеств ДК1 из клеток-предшественников in vitro, и, следовательно, LCM может функционировать в качестве эффективного адъюванта для вакцины. Авторы считают, что произведенные из РВМС продукты могут обеспечивать окружение цитокинов, необходимое для быстрого генерирования ДК1 из их предшественников после инициации иммунного ответа in vivo.
Известно, что цитокины и хемокины, идентифицированные в препаратах LCM, участвуют в генерировании иммунных ответов посредством их аутокринного или паракринного действия на АПК и отвечающие Т- и В-клетки. Концентрации цитокинов, обнаруженных в LCM, являются значительно более низкими, чем концентрации цитокинов, которые обычно используют для дифференцировки моноцитов в незрелые ДК или для созревания ДК in vitro [16, 22]. LCM содержит цитокины (IFNγ, IL-12) и растворимый лиганд CD40, которые, как известно, поляризуют Т-клетки в отношении ответа ТН1, а также цитокины (IL-4 и IL-10), которые поляризуют Т-клетки в направлении ответов ТН2 [5, 40, 41]. Эти последние цитокины могут также индуцировать анергию (толерантность) Т-клеток, когда они присутствуют в культурах антигенпрезентирующих незрелых клеток ДК и Т-клеток. Однако присутствие IL-10 и небольших количеств IL-4 в LCM не аннулирует Т-клеточный вторичный ответ на ТТ; скорее эти Т-клеточные ответы были увеличенными, ясно демонстрируя, что эффект цитокинов ТН1 доминирует.
Кроме провоспалительных цитокинов в LCM были обнаружены высокие концентрации хемокинов. Эти хемокины продуцируются лимфоидными клетками, а также нелимфоидными клетками в контексте воспалительного процесса [35, 42]. В качестве примера RANTES продуцируется СD8+-Т-клетками и он, в свою очередь, индуцирует генерирование других цитокинов и хемокинов (МIР1β, IL-2, IL-6 и интерферонов типа 1), которые активируют Т-клетки, а также и моноциты [43]. Индукция этих цитокинов и хемокинов может быть характерным показателем того, что происходит при активации Т-клеток АПК in vivo в контексте с презентацией антигена. После активации Т-клеток через рецептор Т-клеток и лигирование CD28 эти Т-клетки высвобождают цитокины и хемокины, которые, как известно, влияют на дифференцировку моноцитов в незрелые ДК, а также на их миграцию в регионарные лимфоидные органы. Эти растворимые факторы могут также аттрактировать ДК-предшественники и другие АПК в среду начального столкновения с антигеном (к сигналу опасности). Можно было бы ожидать, что вместе эти цитокины и хемокины, продуцируемые активированными Т-клетками и далее по ходу процесса клетками-«свидетелями» (bystander cells), увеличивают каскад иммунного ответа, что является желаемым свойством адъюванта. Увеличивается осознание того, что различные цитокины способны действовать в качестве адъювантов для вакцин, в частности, GM-CSF и IL-2.
Генерирование происходящей из РВМС кондиционированной среды делает возможным генерирование больших количеств незрелых ДК из их предшественников и созревание полученных из моноцитов ДК. Этот быстрый и экономичный способ может играть важную роль в развитии будущих вакцин и в применении в качестве адъюванта. Кроме того, большой диапазон цитокинов и хемокинов, содержащихся в LCM, предполагает более физиологический стимул, обеспечивающий окружение цитокинов, сходное с окружением, которое может быть обнаружено in vivo после контактирования Т-клеток с антигеном.
Данное изобретение обеспечивает способ применения адъюванта как на основе супернатанта, так и на основе клеток, в комбинации с вакцинным антигеном, для обеспечения усиленного иммунного ответа на эту вакцину.
Таким образом, одной целью данного изобретения является обеспечение адъюванта для вакцины, способного усиливать иммуногенный ответ на вакцину.
Другой целью данного изобретения является обеспечение адъюванта для вакцины, полученного из лимфоцитов человека.
Следующей целью данного изобретения является обеспечение адъюванта для вакцины, полученного из супернатанта, собранного из стимулированных культивируемых лимфоцитов человека.
Дополнительной целью данного изобретения является обеспечение способа применения адъюванта для вакцины введением этого адъюванта животному-хозяину в комбинации с вакциной.
Эти и другие цели данного изобретения станут более очевидными из следующих описания, графического материала и прилагаемой формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ФИГУР
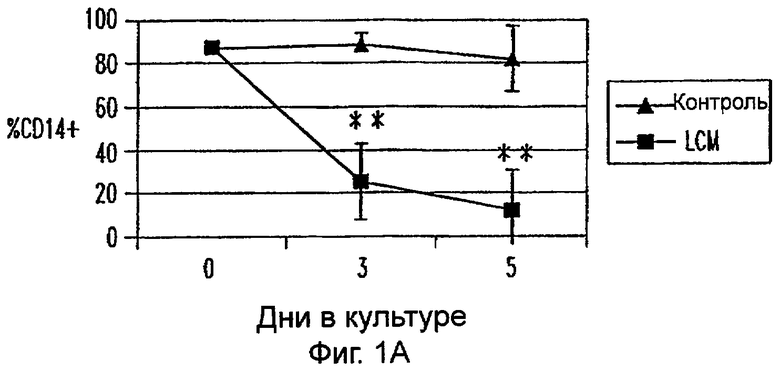
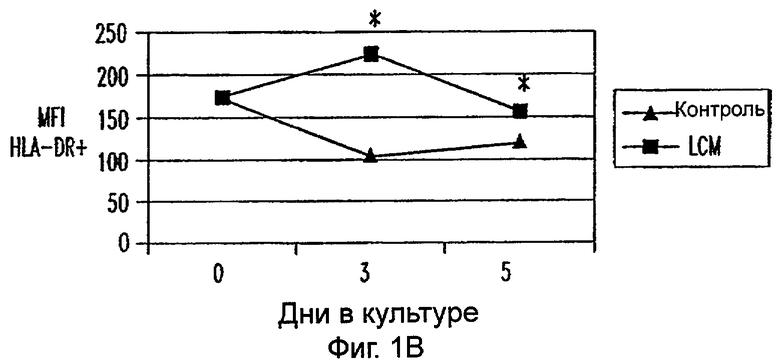
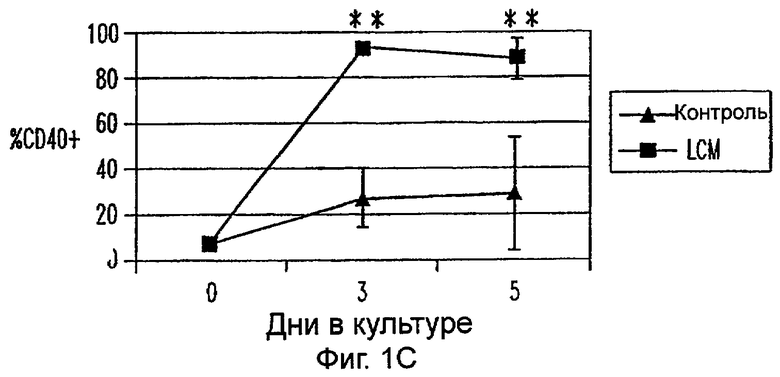
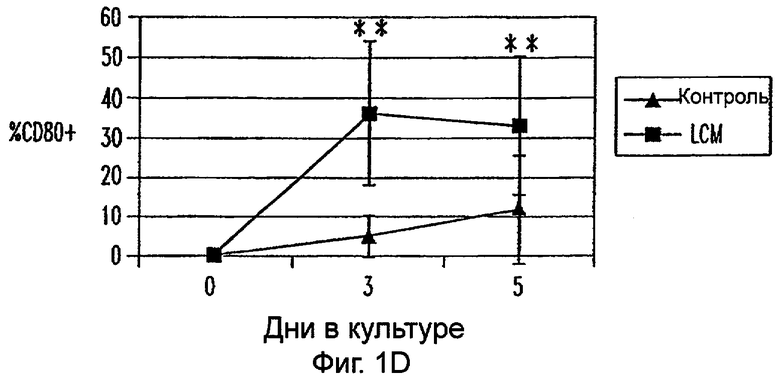
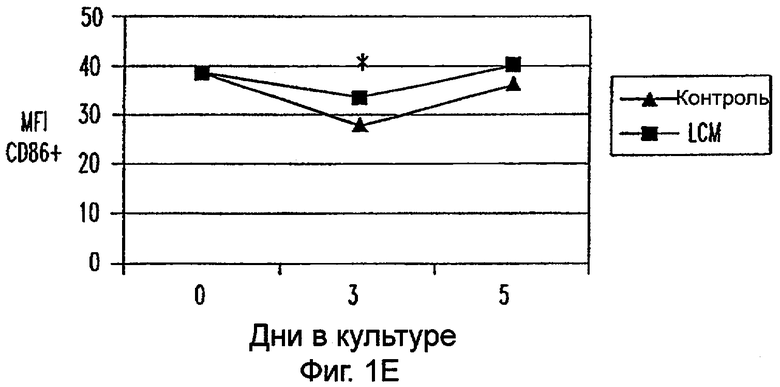
Фиг.1А-1Е: LCM индуцирует ДК-подобный фенотип в моноцитах, культивируемых только с LCM. Экспрессию CD14 (фиг.1А), HLA-DR (фиг.1В), CD40 (фиг.1С), CD80 (фиг.1D) и CD86 (фиг.1Е) анализировали в различных временных точках. Данные представляют средние величины ±SEM из 9 экспериментов. * указывает р<0,05, и ** указывает р<0,005.
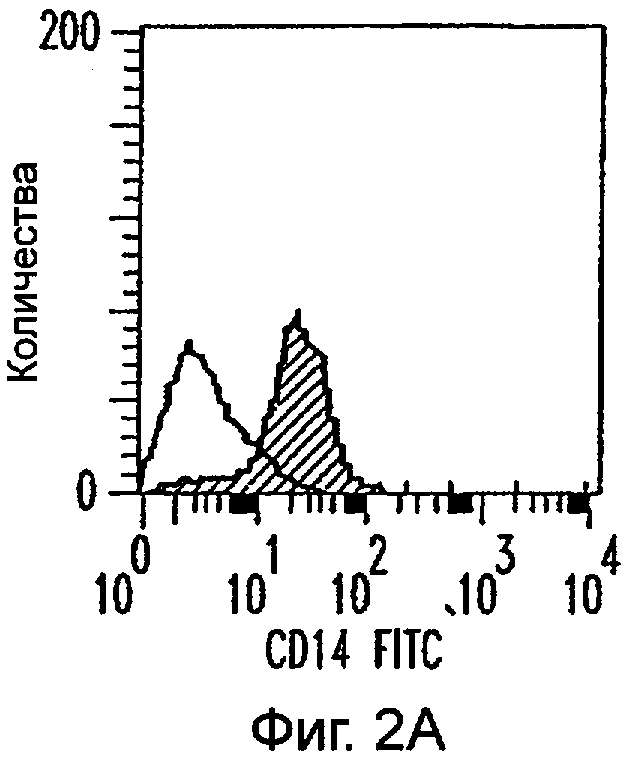
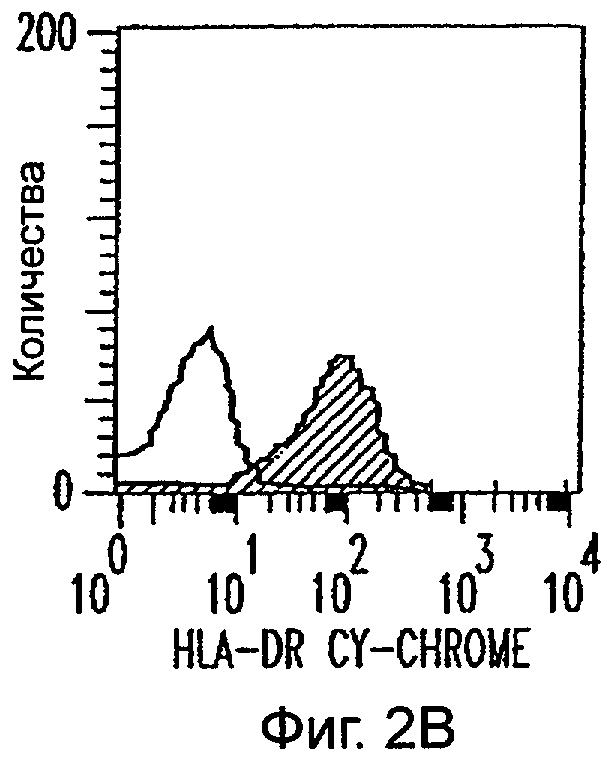
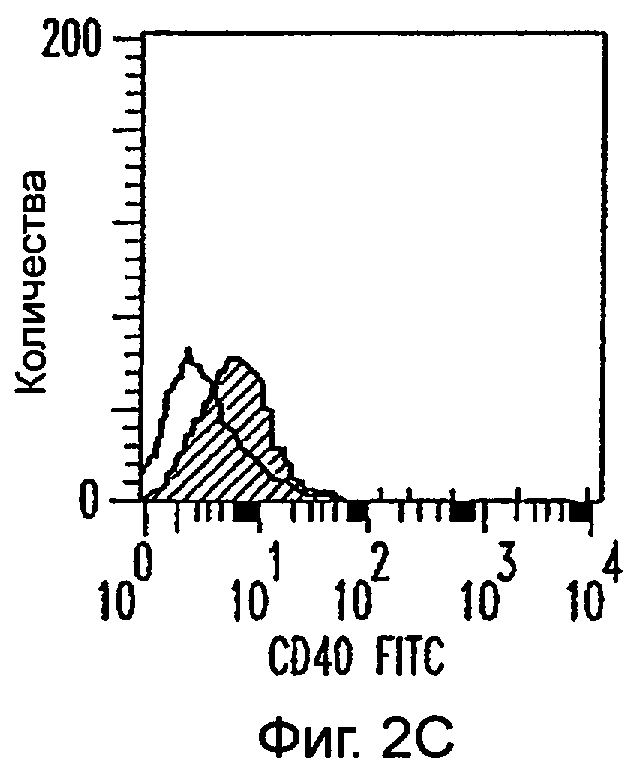
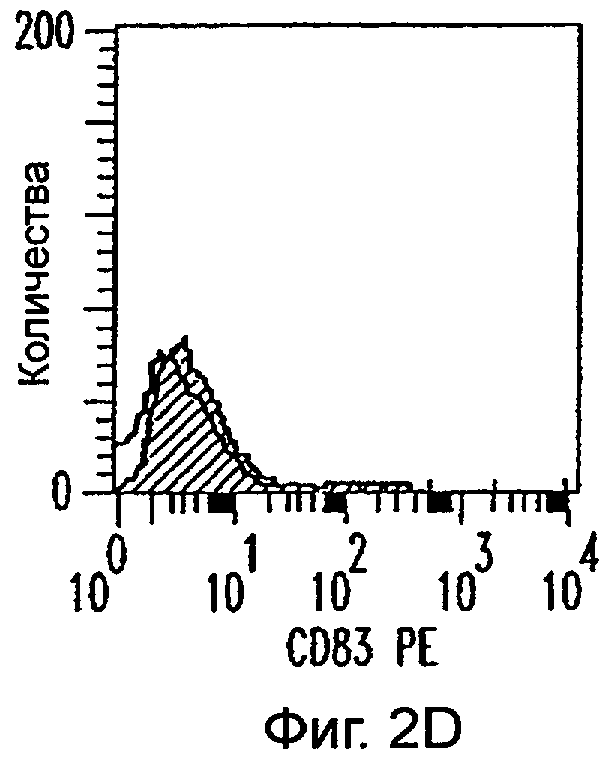
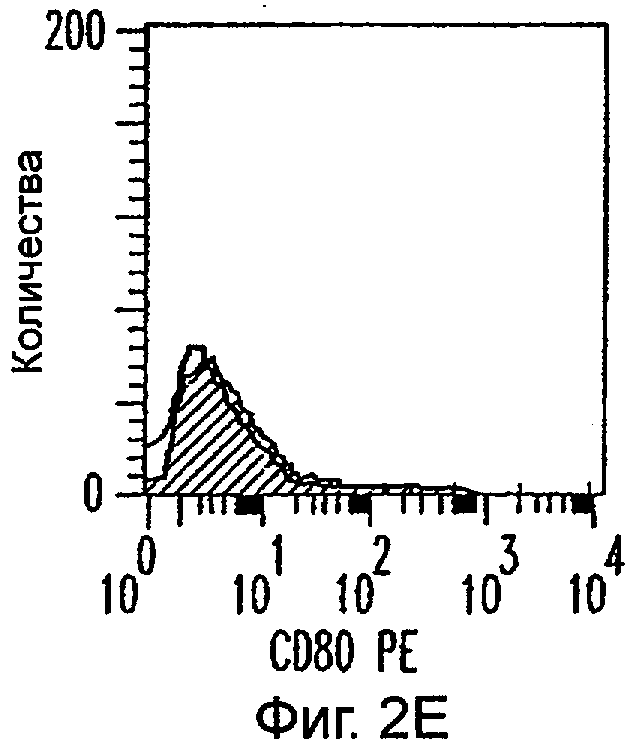
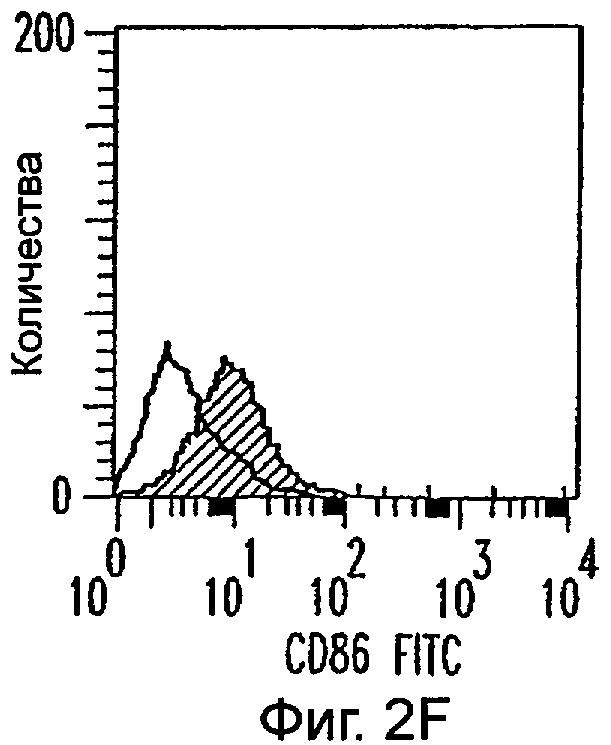
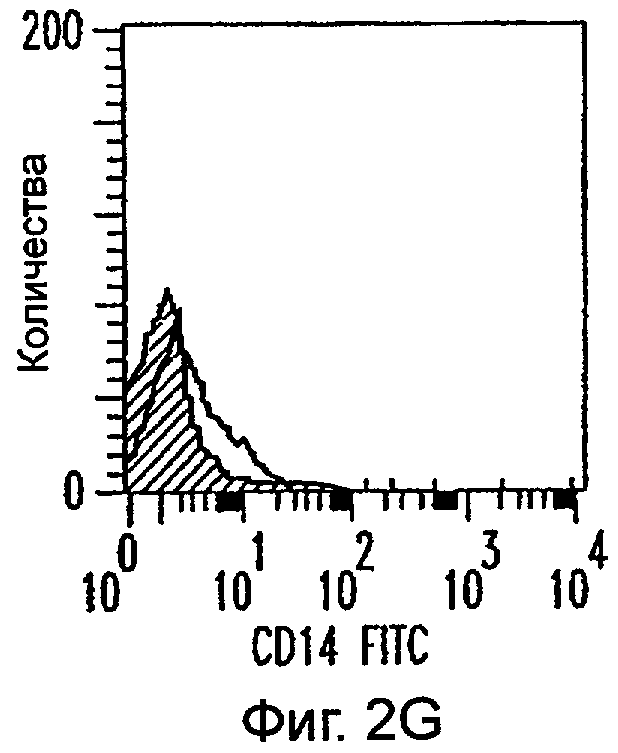
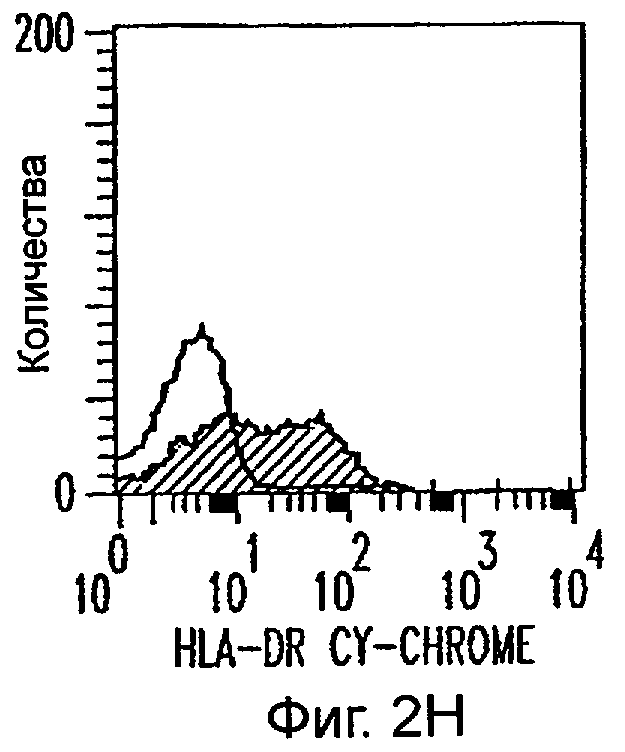
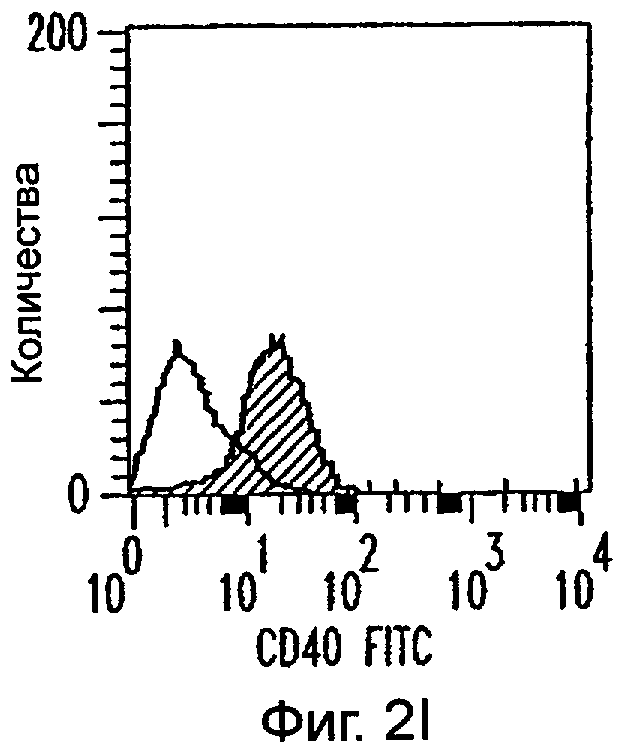
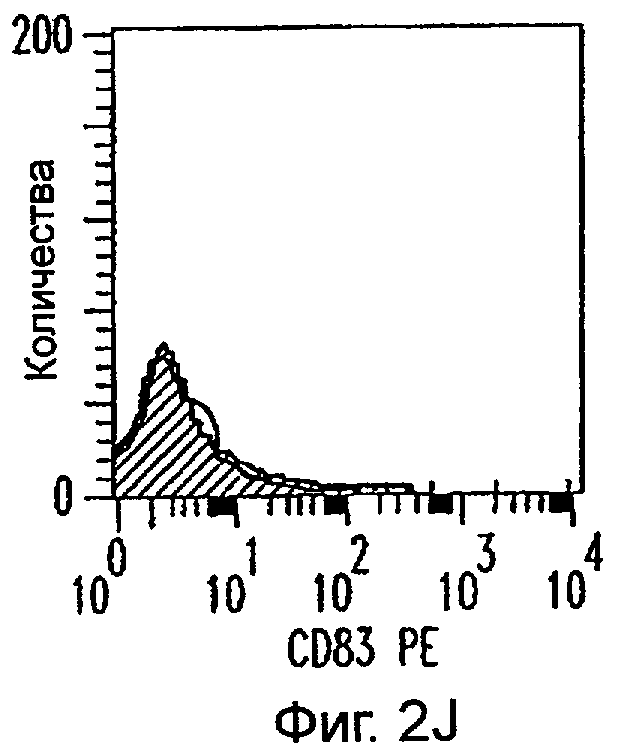
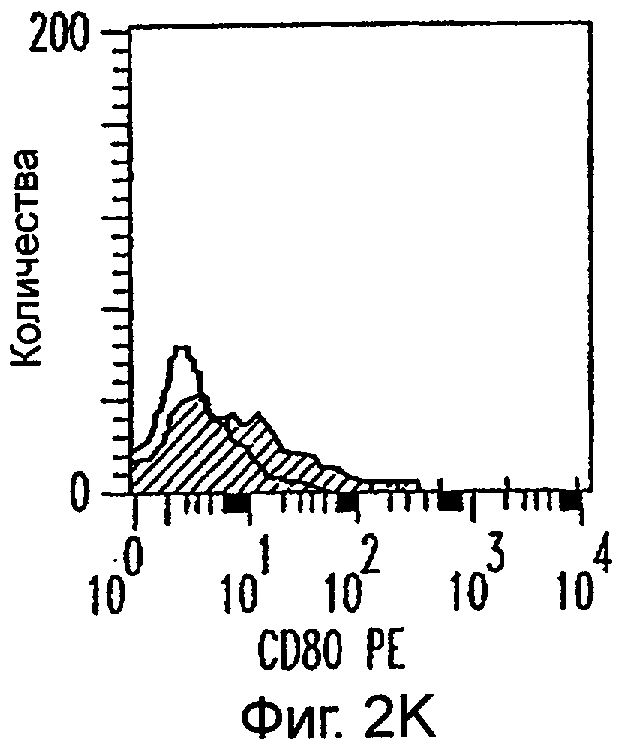
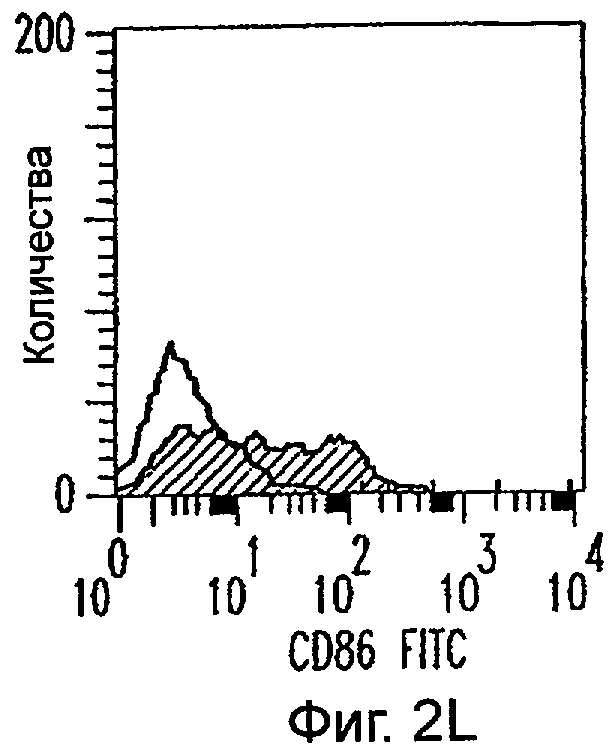
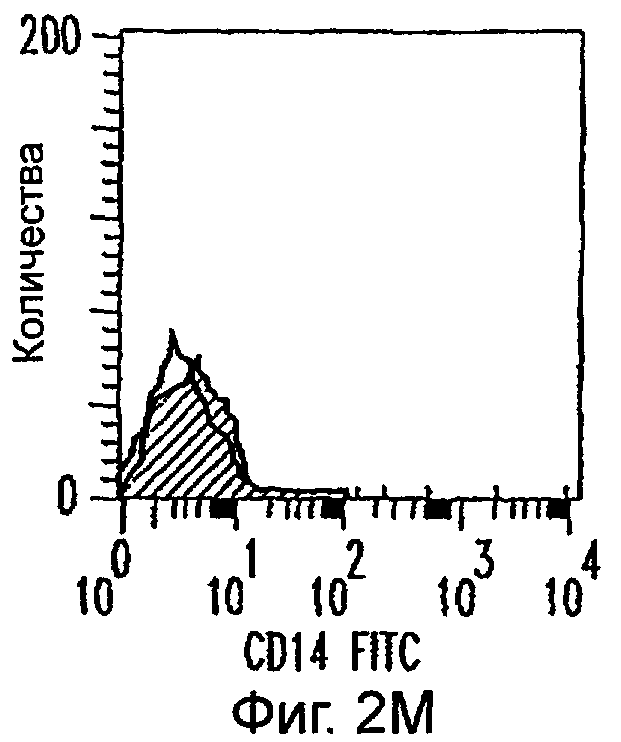
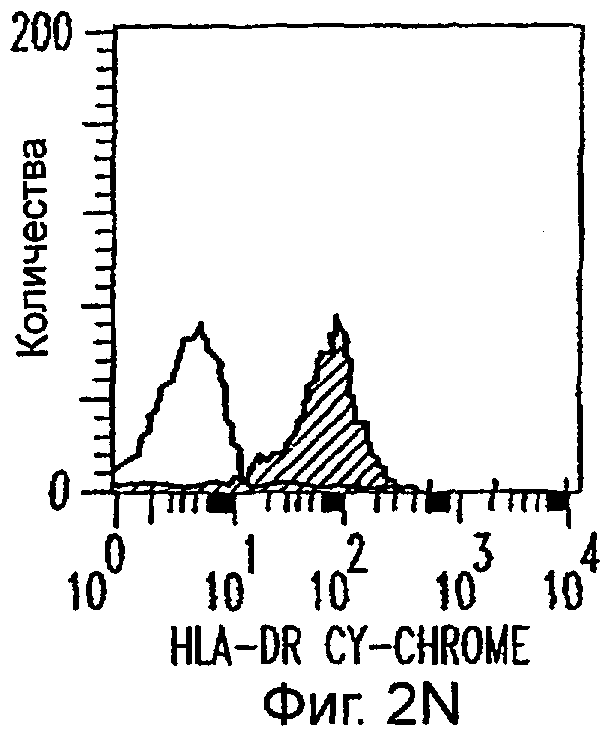
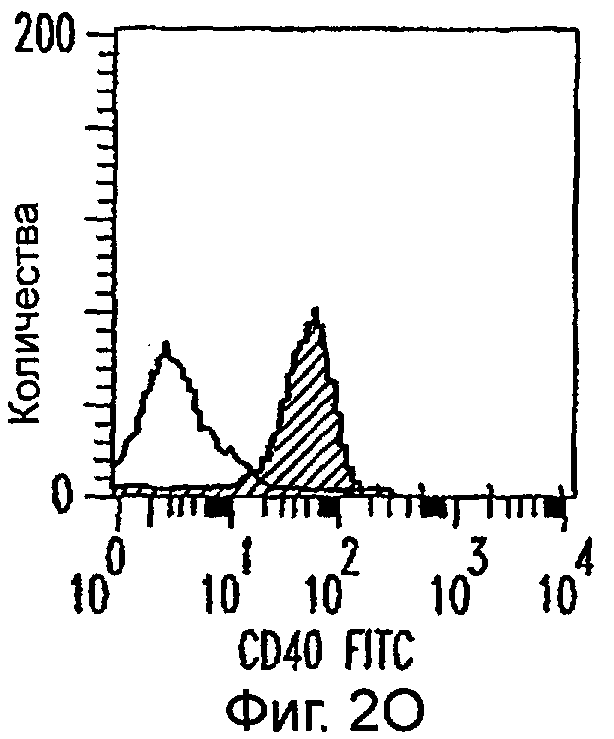
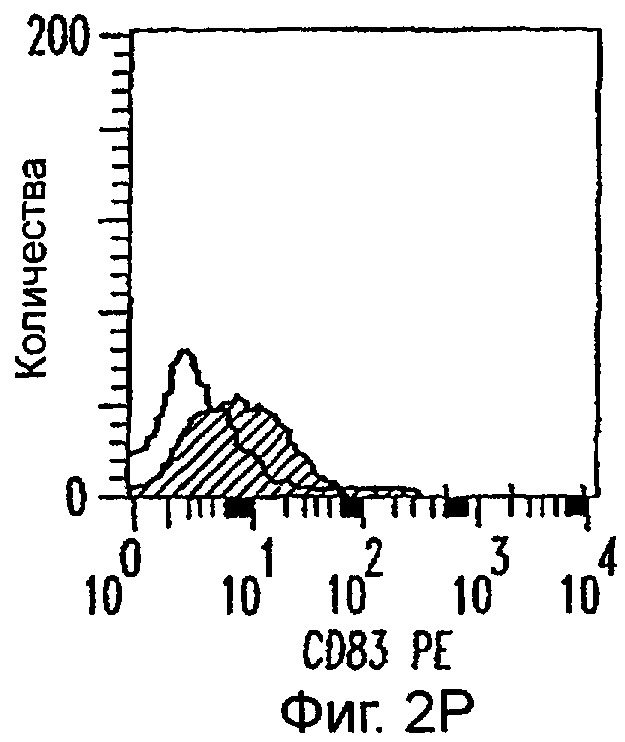
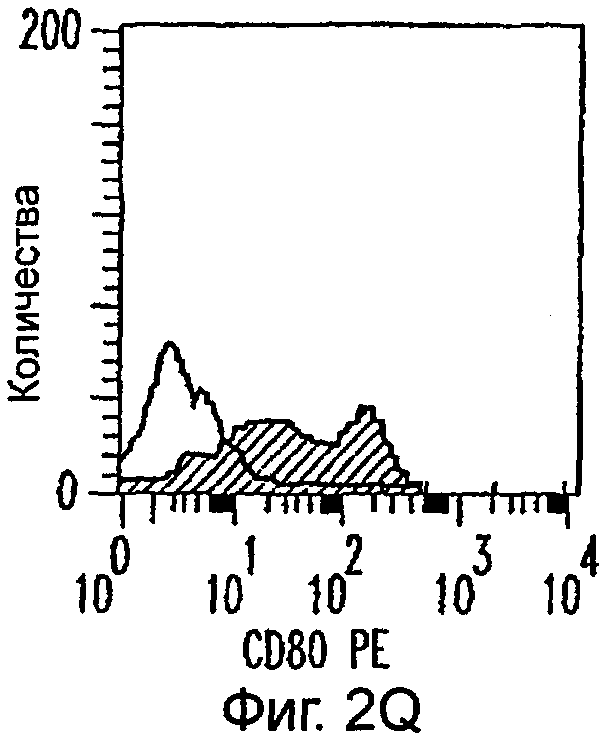
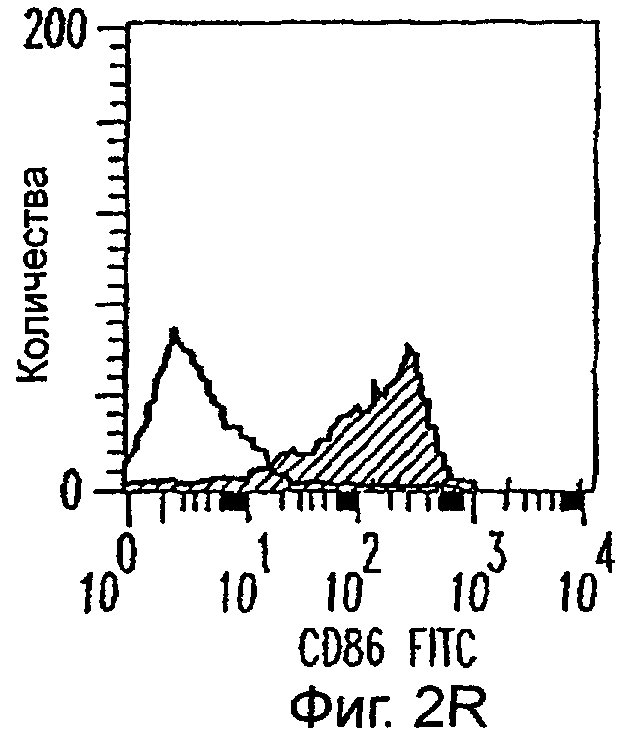
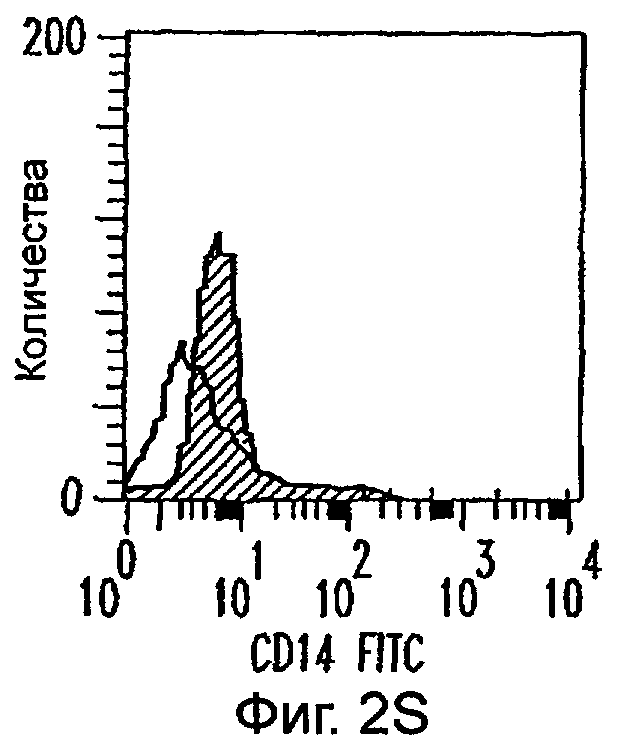
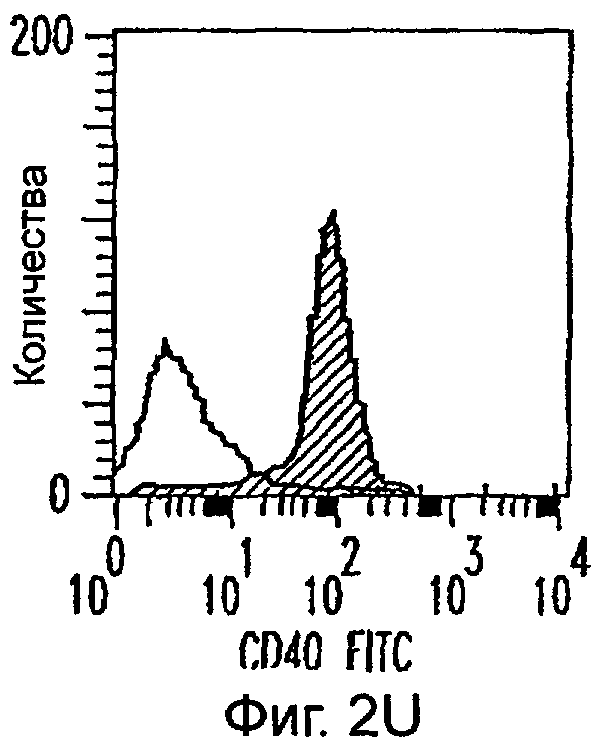
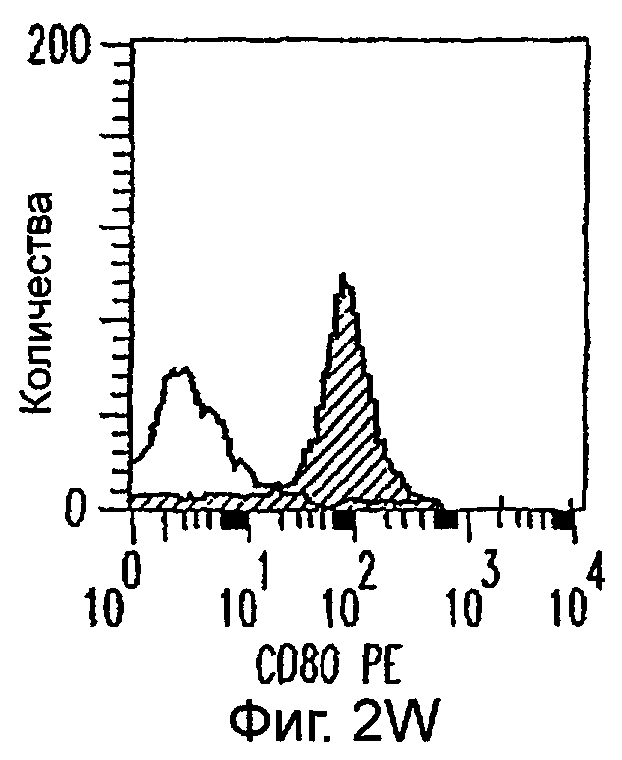
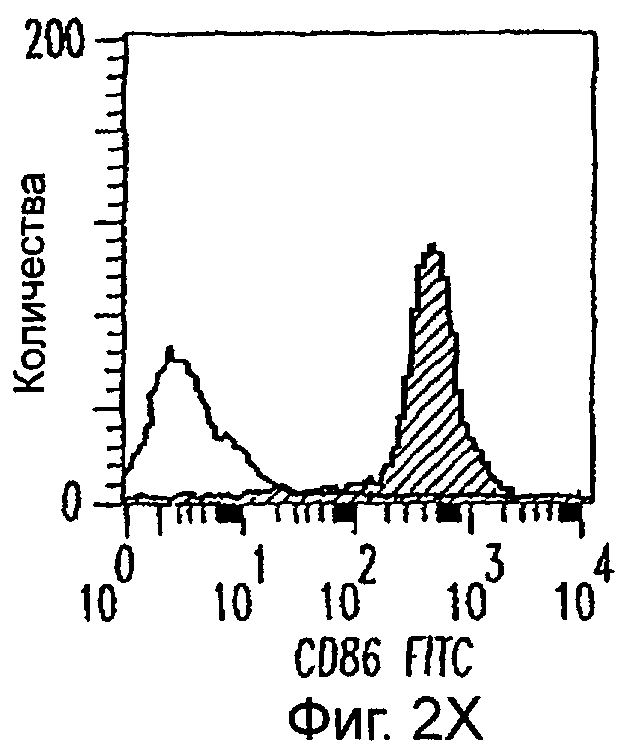
Фиг.2А-2Х: Действие LCM на созревание незрелых полученных из моноцитов ДК. Подвергнутые элютриации моноциты культивировали с GM-CSF/IL-4 в течение 3-4 дней с последующим добавлением только среды (фиг.2G-2L), LCM (фиг.2M-2R) или Смеси для Созревания (фиг.2S-2X) в течение 48 часов. Моноциты, культивируемые только в cRPMI (фиг.2A-2F), использовали в качестве отрицательного контроля. СD11 с+-ДК испытывали на поверхностную экспрессию CD14, HLA-DR, CD40, CD83, CD80 и CD86 проточной цитометрией. Открытые гистограммы представляют окрашивание ДК с использованием mAb изотипа-контроля, а затененные гистограммы представляют окрашивание ДК специфическим mAb.
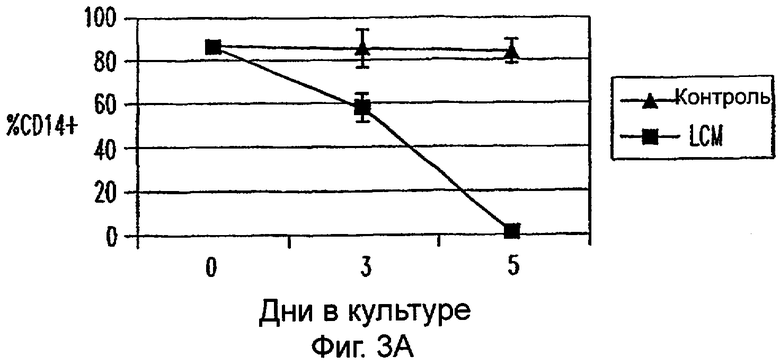
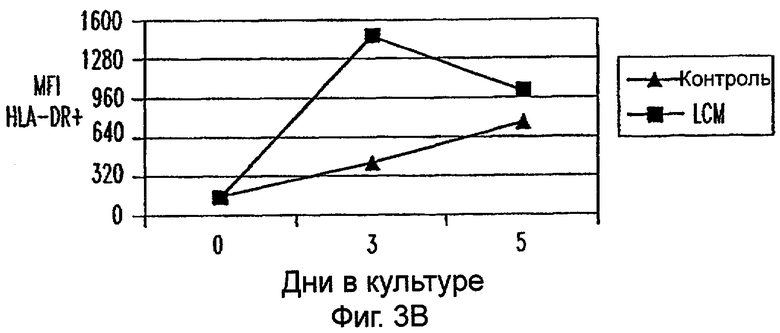
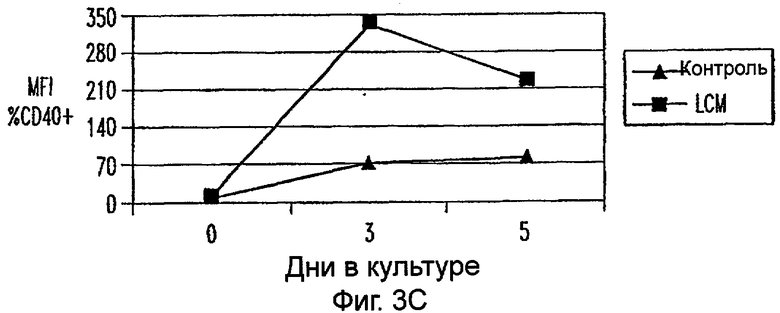
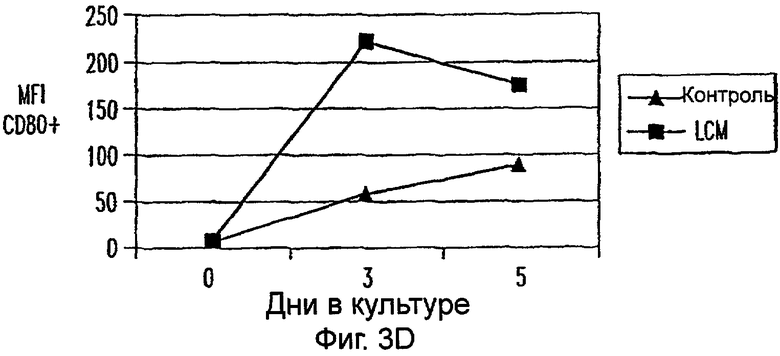
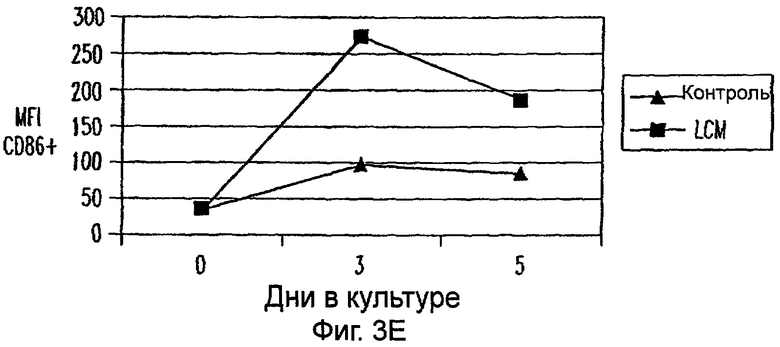
Фиг.3А-3Е: LCM дифференцирует моноциты до ДК-подобного фенотипа при добавлении всех популяций PBMC. CD11c+ клетки анализировали на экспрессию CD14 (фиг.3А), HLA-DR (фиг.3В), CD40 (фиг.3С), CD80 (фиг.3D) и CD85 (фиг.3Е) при днях 0, 3 и 5. Данные представляют среднее из двух экспериментов.
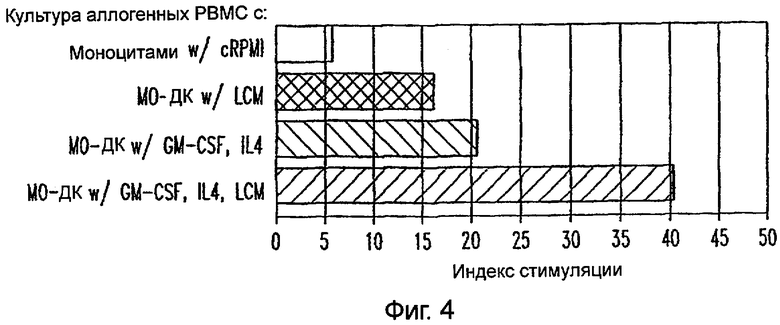
Фиг.4: ДК, генерированные из подвергнутых элютриации моноцитов, культивируемых в присутствии LCM, являются превосходящими в их способности стимулировать аллогенные реакции PBMC, чем ДК из моноцитов, культивируемых в отсутствие LCM. Аллогенные PBMC (1×105 клеток) культивировали в 96-луночных U-донных культуральных планшетах с 1×104 стимуляторными клетками, как указано, в трех повторностях. Реакция PBMC выражена в виде индекса стимуляции (отношения среднего СРМ (имп/мин) индивидуальных MILR к среднему СРМ PBMC, культивируемых только в контрольной среде).
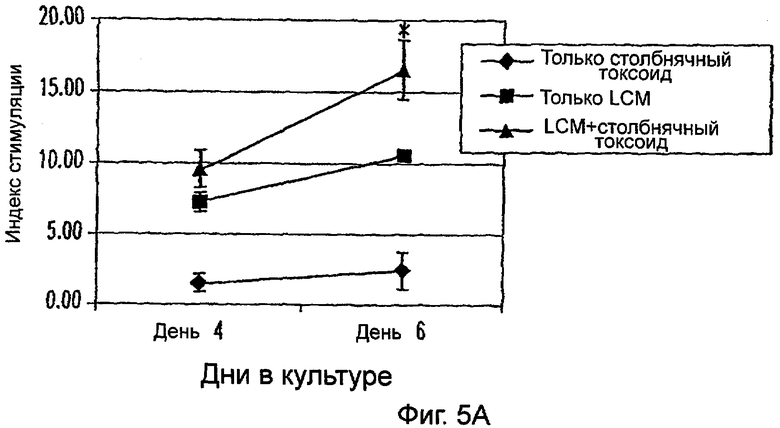
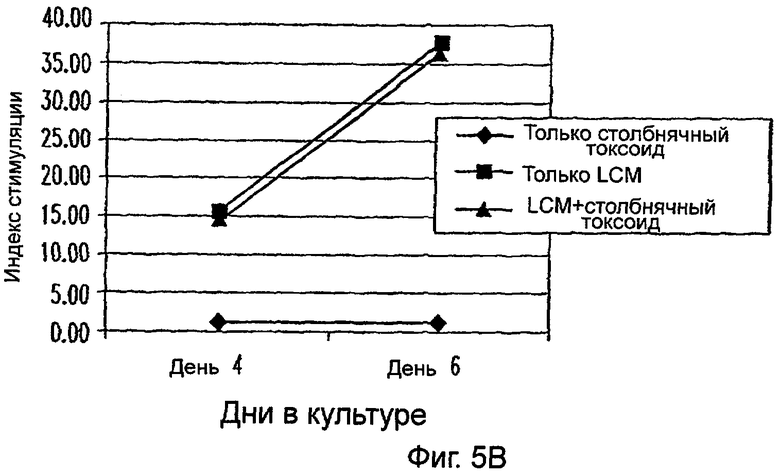
Фиг.5А-5В: LCM увеличивает пролиферативный ответ всех PBMC на столбнячный токсоид. Все PBMC или PBMC, лишенные моноцитов, культивировали в 96-луночных U-донных культуральных планшетах в LCM с 10 мкг/мл белка столбнячного токсоида или без него и в cRPMI с антигеном. PBMC, культивируемые в cRPMT, содержащей 10 мкг/мл Конканавалина А, служили в качестве положительного контроля (данные не показаны). Ответ выражали в виде индекса стимуляции (SI) (отношения среднего СРМ РВМС, культивируемых с ТТ, LCM или TT+LCM (фиг.5А) к среднему СРМ РВМС, культивируемых только в cRPM (фиг.5В)). Данные представляют среднее из двух экспериментов. * указывает р<0,05.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ
В одном аспекте данное изобретение обеспечивает способ усиления иммунного ответа на вакцинный антиген у млекопитающего-хозяина, предусматривающий введение кондиционированной лимфоцитами среды, супернатанта, полученного из активированных клеток-лимфоцитов человека, культивируемых со средой для выращивания, в комбинации с вакцинным антигеном. Предпочтительно этим млекопитающим является человек. Способы и протокол культивирования являются стандартными и известными в данной области. Мононуклеарные клетки периферической крови человека (или другого млекопитающего в зависимости от подлежащего лечению млекопитающего), получаемые из любого источника, разводят в коммерчески доступной среде для культуры ткани. Эти клетки инкубируют с активирующим агентом, состоящим из гранул, покрытых антителами к CD3/CD28. Приблизительно в день 3 клетки и гранулы отделяют от культуральной среды, и эти клетки и гранулы ресуспендируют в дополнительной среде для выращивания, по мере необходимости. Для сбора этих клеток их ресуспендируют в центрифужных пробирках и осаждают, после чего супернатант может быть удален пипеткой и может храниться для более позднего использования.
В данном контексте термин «супернатант» относится к жидкости, удаленной от культивируемых клеток описанным выше способом. «Кондиционированная лимфоцитами среда» и «супернатант» используются здесь взаимозаменяемо и относятся к жидкости, удаленной из культивируемых клеток. Исследования проводили для характеристики этого супернатанта, и было обнаружено, что он содержит молекулы, имеющие средние молекулярные массы менее приблизительно 100000 дальтон.
Введение выполняют известными способами, используемыми для вакцинации, и подходящие способы доставки включают, но не ограничиваются ими, внутримышечную, внутрикожную и подкожную инъекцию. Обычно приблизительно 10 мкг - приблизительно 500 мкг вводят в комбинации с антигеном. Введение может выполняться один раз в неделю, один раз в две недели, один раз в месяц или один раз в год в зависимости от антигена и от уровня желаемого иммунного ответа. Усиление иммунного ответа можно наблюдать, например, проведением стандартных анализов, известных в данной области, которые оценивают клеточный иммунитет (например, пролиферацию Т-клеток) и измеряют титры антител после иммунизации.
Данное изобретение пригодно для использования с большим разнообразием вакцин, в том числе, но не только, вакцин против кори, эпидемического паротита, краснухи, гриппа, вакцин против Haemophilus influanzae типа В, дифтерии, столбняка, коклюша, пневмококковых полисахаридных вакцин, менингококковых полисахаридных вакцин, вакцин против Staphylococcus aureus, респираторного синтициального вируса, стрептококка, парагриппа, Mycoplasma pneumoniae, Mycobacterium leprae, нокардий, легионел, псевдомонас, холерных вакцин, вакцин против брюшного тифа, полиовируса, вакцины против гепатита А, вакцины против ротавируса, Escherichia coli, Shigella, гепатита Е, Listeria, Giardia lamblia, токсокароза, трихиуреаза, аскаридоза, амебиоза, цистицеркоза, рекомбинантных и полученных из плазмы вакцин против гепатита В, вакцин против ВИЧ-1 и ВИЧ-2; Т-лимфотропного вируса типа I человека и Т-лимфотропного вирса типа II человека, вируса Эпстейна-Барра, гепатита С, герпеса В, папиломавируса человека, простого герпеса типа 1 и 2, хламидии, гонореи, трепонемы (сифилиса), сибирской язвы, бешенства, шистосомоза, бляшки Холленхорста, вакцин против желтой лихорадки, японского энцефалита и вакцин против переносимого клещами энцефалита, малярии, лейшманиоза, болезни Лайма, лимфатического филяриатоза и онхоцеркоза, трипаносомоза и болезни Шагаса, рикеттсий и сыпного тифа, лихорадки Денге, аденовирусных вакцин, вакцин против ветряной оспы, цитомегаловируса, коронавирусов и риниовирусов, Streptobacillus, аллергической пептидной вакцины, пептидной вакцины против инфекционного заболевания, раковой пептидной вакцины, аутоиммунной пептидной вакцины и раковой вакцины, использующей антиген, пептид, ДНК-фрагменты и/или другие молекулярные частицы на поверхности или внутри раковой клетки.
ПРИМЕРЫ
Следующие примеры предназначены для иллюстрации данного изобретения и не должны рассматриваться как ограничивающие каким-либо образом данное изобретение. Эксперименты проводили с использованием следующих материалов и способов:
Клетки
Мононуклеарные клетки периферической крови человека (РВМС), используемые для приготовления кондиционированных сред, выделяли из продуктов лейкафереза нормальных здоровых доноров центрифугированием в градиенте плотности в Среде для Разделения Лимфоцитов (ICN Biomedicals Inc., Aurora, ОН). Эти клетки замораживали без утраты жизнеспособности в среде RPMI-1640 (Invitrogen Corp., Grand Island, NY), содержащей 20% АВ-сыворотку человека (Gemini Bio-Products, Woodland, CA) и 10% ДМСО (Sigma, St. Louis, МО), с использованием автоматического клеточного морозильника (Gordinier Electronics, Rosevilie, MI) и хранили в паровой фазе жидкого азота до использования. Моноциты выделяли из РВМС противоточной центрифужной элютриацией и использовали немедленно или замораживали с сохранением жизнеспособности в фетальной телячьей сыворотке (Summit Biotechnology), содержащей 10% ДМСО и 5% глюкозу (Sigma), для более позднего использования.
Получение LCM
Криосохраненные РВМС оттаивали в среде RPMI-1640, дополненной 20% АВ сывороткой человека (hAB), промывали дважды средой RPMI-1640, содержащей 10% hAB. Клетки ресуспендировали в cRPMI [RPMI-1640, дополненной 10% hAB, 2 мМ L-глутамином (Invitrogen), 1% раствором Пенициллина/Стрептомицина (Invitrogen), 20 мМ HEPES-буфером (Invitrogen)], или X-Vivo 15 (BioWhittaker, Walkersville), MD), дополненной 2 мМ L-глутамином, раствором 1% Пенициллина/Стрептомицина, 20 мМ HEPES-буфером. К этим клеткам добавляли CD3/CD28-Dynabeads (Dynal, Lake Success, NY) при 75 мкл гранул на каждые 1×106 РВМС, и эти культуры инкубировали в течение 3 дней при 37°С в 5% СО2. Затем бесклеточные супернатанты собирали и хранили при 2-8°С перед использованием. LCM использовали при разведении 1:1 в cRPMI.
Культивирование подвергнутых элютриации моноцитов
Подвергнутые элютриации моноциты промывали cRPMI, ресуспендировали в равных объемах LCM и cRPMI в концентрации 5×105 клеток/мл и культивировали в 24-луночных планшетах (Denville Scientific Inc., Metuchen, NJ) при 37°С в 5% СО2 в течение 5-6 дней. Альтернативно моноциты культивировали в cRPMI, содержащей 750 Е/мл GM-CSF ((R&D Systems, Minneapolis, MN) и 720 Е/мл IL-4 (R&D Systems), в течение 3-4 дней с последующим добавлением кондиционированной среды или комбинации цитокинов [10 нг/мл IL-1β, 10 нг/мл TNFα, 0,91 нг/мл IL-6 (R&D Systems), 1 мкг/мл PGE2 (Sigma)] (Смесь для Созревания) в течение дополнительных 48 часов. Смесь для Созревания служила в качестве положительного контроля, а моноциты, культивируемые только в cRPMI, использовали в качестве отрицательного контроля.
Проточная цитометрия
Клетки промывали один раз в cRPMI, ресуспендировали в 1х ЗФР (Invitrogen), содержащем 5% hAB, и инкубировали в течение 15 минуй при комнатной температуре (22-25°С) для блокирования Fc-рецепторов. Затем клетки промывали, ресуспендировали в промывочном буфере (1Х ЗФР с 1% hAB и 0,1% NaN3) и метили конъюгированными с флуорохромом антителами (BD BioSciences, San Diego, СА) против CD14, CD11c, CD40, CD83, CD80, CD86 и контрольными антителами подходящего изотипа. Спустя 30 минут при 4°С эти клетки промывали буфером и фиксировали в 1Х ЗФР с 1% параформальдегидом. Данные проточной цитометрии получали с использованием проточного цитометра FACScan и анализировали с использованием программного обеспечения CellQuest (Becton Dickinson, SanJose, CA). Окна возбуждения флуоресценции устанавливали в соответствии с пробами контрольного изотипа.
Анализ цитокинов и хемокинов
Цитокины и хемокины в LCM определяли количественно с использованием коммерчески доступных твердофазных иммуноферментных анализов (ELISA; R&D Systems, Minneapolis, MN) в соответствии с рекомендациями изготовителя. Концентрацию PGE2 также измеряли при помощи ELISA (Cayman Chemical Co., Ann. Arbor, Ml). Все определения выполняли в двух повторностях.
Аллогенная MLR
Дендритные клетки собирали из культур, промывали дважды в cRPVI и высевали в 96-луночные U-донные культуральные планшеты (Denville Scientific, Inc.) при 104, 103 и 102 клеток на лунку. РВМС аллогенного респондера добавляли в каждую лунку при 1×105 клеток на лунку в общем объеме 200 мкл. Все условия высевали в трех повторностях. Клетки инкубировали в течение 3 дней при 37°С в 5% СО2, импульсно обрабатывали 1,0 мкКм тритированного тимидина (Perkin Elmer, Boston, MA) в течение 16 часов и собирали с использованием автоматизированного многолуночного устройства для сбора клеток (Tomtec, Orange, CT). Количество тритированного тимидина, включенного в клетки-респондеры, определяли с помощью жидкостного сцинтилляционного счетчика MicroBeta TriLux (Wallac, Turku, Finland).
Анализ антигенной стимуляции
Непраймированные нормальные РВМС человека (1×105/100 мкл cRPMI) высевали в 96-луночные U-донные культуральные планшеты, к которым добавляли 100 мкл LCM или 100 мкл cRPMI с 10 мкг/мл столбнячного токсоида (Accurate Chemical and Scientific Corp., Westbury, NY) или без него. РВМС культивировали в cRPMI с 10 мкг/мл конканавалина A (Sigma, St. Louis, МО), служившего в качестве положительного контроля. Все условия высевали в трех повторностях, культивировали при 37°С в 5% CO2 в течение 3 и 5 дней, импульсно обрабатывали 1,0 мкКм тритированного тимидина в течение 16 часов и собирали с использованием автоматизированного многолуночного устройства для сбора клеток. Количество тритированного тимидина, включенного в клетки-респондеры, определяли с помощью жидкостного сцинтилляционного счетчика MicroBeta TriLux.
LCM дифференцирует моноциты в ДК в отсутствие дополнительных цитокинов
Пытаясь генерировать высокоэффективную смесь цитокинов для генерирования и созревания больших количеств ДК человека из их предшественников in vitro, авторы настоящего изобретения получили культуральные супернатанты от РВМС, стимулированных антителами к CD3/CD28, (LCM), как описано в Материалах и Методах, и исследовали их действие на высокоочищенные моноциты человека, полученные из РВМС противоточным центрифугированием. Моноциты культивировали в LCM в отсутствие GM-CSF и IL-4, и экспрессию маркеров клеточной поверхности, которые характеризуют дифференцированные и зрелые ДК, проверяли с помощью проточной цитометрии. До культивирования моноциты в существенной степени экспрессировали CD11c, а также CD14 и HLA-DR. Культивирование в LCM приводило к значительному уменьшению процента клеток, экспрессирующих CD14, к дню 3, и впоследствии оставалось только несколько клеток, экспрессирующих этот маркер (фиг.1А). Относительные уровни экспрессии (mfi) HLA-DR (фиг.1В), CD40 (фиг.1С), CD80 (фиг.1D) и CD86 (фиг.1Е) были устойчиво увеличены на клетках, культивируемых в LCM, по сравнению с клетками, культивируемыми только на среде. В 4 из 9 экспериментов наблюдали небольшой процент СD83+-ДК; однако эти результаты не были значимыми. В целом LCM дифференцирует моноциты в фенотипически незрелые ДК, стадию развития, которая является оптимальной для получения антигена.
LCM вызывает созревание моноцитов, культивируемых в GM-CSF/IL-4
Оценивали способность LCM вызывать созревание незрелых ДК, которые были генерированы in vitro из моноцитов в присутствии GM-CSF и IL-4. Моноциты, полученные из РВМС элютриацией, культивировали в течение 3-4 дней в среде, содержащей GM-CSF и IL-4, с последующим добавлением LCM или стандартной Смеси для Созревания, содержащей IL-1β, TNFα, IL-6 и PGE2 (см. Материалы и методы). Спустя 48 часов выполняли проточную цитометрию. Как показано на фиг.2 и в таблице 1, как LCM, так и Смесь для Созревания индуцировали экспрессию HLA-DR, костимуляторных молекул и CD83, общепринятые сигнатуры для зрелых ДК. Эти результаты указывают на то, что LCM может облегчать конечное созревание незрелых полученных из моноцитов ДК. Активность созревания ДК посредством LCM, полученных в целом из четырех доноров, была очень согласующаяся (данные не показаны).
Экспрессия поверхностных маркеров на моноцитах, культивируемых в GM-CSF и IL-4 с LCM или без LCM
Таблица 1. Статистическая оценка MFI и экспрессии в процентах CD40, CD83, CD80 и CD86 на моноцитах, культивируемых с GM-CSF/IL-3 с добавлением (+LCM) или без добавления (-LCM) LCM в течение последних 48 часов. Данные выражены в виде средних величин ±SEM из 8 экспериментов и статистическую значимость определяли парным двусторонним t-критерием Стьюдента.
Моноциты в РВМС экспрессируют вспомогательные молекулы при культивировании cLCM.
Для исследования действия LCM на распределение и фенотип различных популяций клеток, содержащихся в периферической крови in vivo, все РВМС культивировали в LCM в течение 3 или 5 дней. В этих двух временных точках не были обнаружены различия в процентах CD3+, CD4+, CD8+, CD56+ и CD19+ клетках (данные не показаны). Кроме того, проценты CD3+ клеток, коэкспрессирующих CD25, маркер для активации, не увеличивались на протяжении 5-дневного периода культивирования (данные не показаны). Однако экспрессиия CD14 на моноцитах, идентифицированная по их экспрессии CD11c, уменьшалась в день 3 и почти полностью прекращалась ко дню 5 (фиг.3А). В отличие от этого экспрессия HLA-DR, CD40, CD80 и CD86 на CD11c+ клетках увеличивалась (фиг.3). Таким образом, LCM дифференцирует высокоочищенные моноциты, а также моноциты в целых РВМС в клетки с ДК-подобным фенотипом.
Концентрации цитокинов и хемокинов в LCM.
Концентрации цитокинов и хемокинов в LCM (таблица 2) определяли стандартными способами ELISA и сравнивали с концентрациями цитокинов, обычно используемых для генерирования или созревания ДК (таблица 3). Была идентифицирована целая батарея растворимых медиаторов, включающая GM-CSF и IL-4, воспалительные цитокины (IL-1β, IL-6, PGE2, TNFα, IFNγ), хемокины (МСР-1, МIР1, RANTES) и SCD40L (таблица 2). Удивительно, что концентрации GM-CSF и IL-4 в LCM были значимо более низкими, чем концентрации этих двух цитокинов, используемых в стандартных протоколах для генерирования незрелых ДК из моноцитов (таблица 3). Одним из действий IL-4 на моноциты во время дифференцировки in vitro в незрелые ДК является уменьшение экспрессии CD14. Низкий уровень IL-4 в LCM может быть ответственным за относительно низкую экспрессию CD14 на моноцитах, культивируемых в LCM (фиг.1А, 3А). Были также более низкие концентрации воспалительных цитокинов в LCM по сравнению со Смесью для Созревания, даже хотя оба агента созревания оказывали сравнимые действия на незрелые ДК.
Количественное определение цитокинов и хемокинов, содержащихся в LCM
Таблица 2: Средняя концентрация цитокинов или хемокинов в четырех различных препаратах LCM, определенная при помощи ELISA.
Таблица 3: Количества GM-CSF и IL-4, содержащиеся в LCM, сравнивали с их количествами, используемыми в стандартных протоколах для генерирования полученных из моноцитов ДК, а также с концентрациями выбранных провоспалительных цитокинов в LCM и Смеси для Созревания. * Следует обратить внимание на то, что концентрации цитокинов для LCM, приведенные в этой таблице, составляют половину концентраций в таблице 2: так как LCM добавляют к клеткам при разведении 1:1 в cRPMI.
Моноциты, культивируемые в LCM, стимулируют аллогенные РВМС
Функциональной характеристикой полученных из моноцитов ДК является их способность эффективно индуцировать аллогенные ответы in vitro. Определяли аллогенные ответы РВМС на моноциты, которые культивировали в течение пяти дней в отсутствие или в присутствии LCM или которые культивировали в средах, содержащих GM-CSF/IL-4, с последующим добавлением cRPVI или LCM. Как показано на фиг.4, индекс стимуляции (SI), генерируемый моноцитами, культивируемыми только в LCM, был приблизительно в 3 раза большим, чем индекс стимуляции, индуцируемый моноцитами, которые культивировали только в cRPMI. Однако SI был наивысшим в случае моноцитов, которые культивировались в GM-CSF/IL-4 и подвергались созреванию с LCM. Эти результаты показывают, что LCM превращает моноциты и незрелые ДК в более эффективные антигенпрезентирующие клетки.
LCM усиливает ответы РВМС на столбнячный токсоид
Кроме того, исследовали пролиферативные ответы РВМС на вторичный антиген столбнячного токсоида (ТТ), опять в присутствии или в отсутствие LCM (фиг.5А). В отсутствие LCM и других цитокинов РВМС обнаруживали низкие уровни ответов на ТТ; однако с добавлением LCM ответ на ТТ значимо увеличивался в день 6 (фиг.5а). Важно указать на то, что только LCM индуцировал синтез ДНК в РВМС даже в отсутствие специфического антигена; тем не менее ответ на специфический антиген был значимо увеличенным. Для определения был ли наблюдаемый ответ на ТТ опосредован моноцитами, эти эксперименты повторяли с использованием РВМС, которые были истощены в отношении CD14+ клеток иммуномагнитными гранулами. Как показано на фиг.5В, не было ответа на ТТ с добавлением или без добавления LCM. Более высокий SI в экспериментах, где моноциты удаляли (фиг.5В), может быть ответственным за относительное увеличение количеств других типов клеток, в частности, CD3+ клеток (данные не показаны). Эти результаты ясно указывают на то, что LCM усиливает способность РВМС отвечать на ТТ, и эта реакция опосредована моноцитами.
Описанные выше открытия указывают на то, что LCM превращает высокоочищенные моноциты и моноциты в целых РВМС в ДК-подобный фенотип. Кроме того, незрелые полученные из моноцитов ДК развивали фенотип зрелых ДК после культивирования в LCM. Функционально эти полученные из LCM ДК обнаруживали увеличенную способность стимулировать аллогенные РВМС по сравнению с их моноцитарными предшественниками и значимо увеличивали ответ на ТТ. Наблюдаемые эффекты LCM могут быть ответственными за батарею провоспалительных цитокинов и хемокинов, идентифицированных в этой кондиционированной среде.
В то время как конкретные варианты осуществления данного изобретения были описаны выше для целей иллюстрации, специалистам с квалификацией в данной области будет очевидно, что многочисленные вариации деталей данного изобретения могут быть произведены без отклонения от данного изобретения, определенного в прилагаемой формуле изобретения.
Источники информации
1. Banchereau J, Briere F, Caux C, et al. Immunobiology of dendritic cells. Annu Rev Immunol JID - 8309206 2000; 18:767-811.
2. Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature JID 0410462 1998; 392:245-252.
3. Steinman RM. The dendritic cell system and its role in inmunogenicity. Annu Rev Immunol JID - 8309206 1991; 9:271-296.
4. Hart DN, McKenzie JL. Interstitial dendritic cells. Int Rev Immunol JID - 8712260 1990; 6:127-138.
5. Liu YJ. Dendritic cell subsets and lineages, and their functions in innate and adaptive immunity. Cell JID - 0413066 2001; 106:259-262.
6. Liu YJ, Kanzler H, Soumelis V, Gilliet M. Dendritic cell lineage, plasticity and cross-regulation. Nat Immunol JID - 100941354 2001; 2:585-589.
7. Palucka К, Banchereau J. How dendritic cells and microbes interact to elicit or subvert protective immune responses. Curr Opin Immunol JID - 8900118 2002; 14:420-431.
8. Sallusto F, Cella M, Danieli C, Lanzavecchia A. Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolccules in the major histocompalibility complex class II compartment: downrcgulalion by cytokines and bacterial products. J Exp Med JID - 2985109R 1995; 182:389-400.
9. Bottomly К. Т cells and dendritic cells get intimate. Science JID - 0404511 1999; 283:1124-1125.
10. Reid SD, Penna G, Adorini L. The control of Т cell responses by dendritic cell subsets. Curr Opin Inimunol JBD - 8900118 2000; 12:114-121.
11. O'Doherty U, Peng M, Gezelter S, et al. Human blood contains two subsets of dendritic cells, one immunologically mature and the other immature. Immunology JID - 0374672 1994; 82:487-493.
12. O'Doherty U, Steinman RM, Peng M, et al. Dendritic cells freshly isolated from human blood express CD4 and mature into typical immunostimuiatory dendritic cells after culture in monocyte-conditioned medium. J Exp Med JID - 2985109R 1993; 178:1067-1076.
13. Randolph GJ, Inaba K, Robbiani DF, Sleinman RM, Muller WA. Differentiation of phagocytic monocytes into lymph node dendritic cells in vivo. Immunity JID - 9432918 1999; 11:753-761.
14. Robinson SP, Patterson S, English N, Davies D, Knight SC, Reid CD. Human peripheral blood contains two distinct lineages of dendritic cells. Eur J Immunol JID - 1273201 1999; 29:2769-2778.
15. Zhou LJ, Tedder TF. CD14 + blood monocytes can differentiate into functionally mature CD83 + dendritic cells. Proc Nail Acad Sci USA JID - 7505876 1996; 93:2588-2592.
16. Dauer M, Obermaier B, Herten J, et al. Mature dendritic cells derived from human monocytes within 48 hours: a novel strategy for dendritic cell differentiation from blood precursors. J Immunol JVD - 2985117R 2003; 170:4069-4076.
17. Kiertscher SM, Roth MD. Human CD14 + leukocytes acquire the phenotype and function of antigen-presenting dendritic cells when cultured in GM-CSF and IL-4. J Leukoc Biol JID - 8405628 1996; 59:208-218.
18. Pickl WF, Majdic O, Kohl P. et al. Molecular and functional characteristics of dendritic cells generated from highly purified CD14 + peripheral blood monocytes. J Immunol JID - 2985117R 1996; 157:3850-3859.
19. Sallusto F, Lanzavecchia A. Efficient presentation of soluble antigen by cultured human dendritic cells is maintained by granulocyte/macrophage colony-stimulating factor plus interleukin 4 and downregulated by tumor necrosis factor alpha. J Exp Med JID - 2985109R 1994; 179:1109-1118.
20. Cella M, Salio M, Sakakibara Y, Langen H, Julkunen I, Lanzavecchia A. Maturation, activation, and protection of dendritic cells induced by double-stranded RNA. J Exp Med JID - 2985109R 1999; 189:821-829.
21. Chomarat P, Dantin С, Bennett L, Banchereau J, Palucka AK. TNF skews monocyte differentiation from macrophages to dendritic cells. J Immunol JID - 2985117R 2003; 171:2262-2269.
22. Jonuleit H, Kuhn U, Muller G, et al. Pro-inflammatory cytokines and prostaglandins induce maturation of potent immunostimulatory dendritic cells under fetal calf serum-free conditions. Eur J Immunol JID - 1273201 1997; 27:3135-3142.
23. Verdijk RM, Mutis T, Esendam B, et al. Polyriboinosinic polyribocytidylic acid (poly(I:C)) induces stable maturation of functionally active human dendritic cells. J Immunol JID - 2985117R 1999; 163:57-61.
24. Zou GM, Tam YK. Cytokines in the generation and maturation of dendritic cells: recent advances. Eur Cytokine Netw JID - 9100879 2002; 13:186-199.
25. Kato К, Takaue Y, Wakasugi H. T-cell-conditioncd medium efficiently induces the maturation and function of human dendritic cells. J Leukoc Biol JID - 8405628 2001; 70:941-949.
26. Reddy A, Sapp M, Feldman M, Subklewe M, Bliardwaj N. A monocyte conditioned medium is more effective than defined cytokines in mediating the terminal maturation of human dendritic cells. Blood JID - 7603509 1997; 90:3640-3646.
27. Guermonprez P, Valladeau J, Zitvogel L, Thery C, Amigorena S. Antigen presentation and Т cell stimulation by dendritic cells. Annu Rev Immunol JTD - 8309206 1903; 20:621-667.
28. Inaba K, Metlay JP, Crowley MT, Steinman RM. Dendritic cells pulsed with protein antigens in vitro can prime antigen-specific, MHC-restricted Т cells in situ. J Exp Med JID - 2985109R 1990; 172:631-640.
29. Toujas L, Delcros JG, Diez E, et al. Human monocyte-derived macrophages and dendritic cells are comparably effective in vitro in presenting HLA class I-restricted exogenous peptides. Immunology JID-0374672 1997; 91:635-642.
30. Weissman D, Ni H, Scales D, et al. HIV gag mRNA transfection of dendritic cells (DC) delivers encoded antigen to MHC class I and II molecules, causes DC maturation, and induces a potent human in vitro primary immune response. J Immunol JID - 2985117R 2000; 165:4710-4717.
31. Gallucci S, Lolkema M, Matzinger P. Natural adjuvants: endogenous activators of dendritic cells. Nat Med JID - 9502015 1999; 5:1249-1255.
32. Dieu MC, Vanbervliet B, Vicari A, et al. Selective recruitment of immature and mature dendritic cells by distinet chemokines expressed in different anatomic sites. J Exp Med JID - 2985109R 1998; 188:373-386.
33. Krug A, Rothenfusser S, Selinger S, et al. CpG-A oligonucleotides induce a monocyte-derived dendritic cell-like phenotype that preferentially activates CD8 Т cells. J Immunol JID - 2985117R 2003; 170:3468-3477.
34. Luft T, Jefford M, Luetjens P. et al. Functionally distinct dendritic cell (DC) populations induced by physiologic stimuli: prostaglandin E(2) regulates the migratory capacity of specific DC subsets. Blood JID -7603509 2002; 100:1362-1372.
35. Sallusto F, Palermo B, Lenig D, et al. Distinet patterns and kinetics of chemokine production regulate dendritic cell function. Eur J Immunol JID - 1273201 1999; 29:1617-1625.
36. Thumer B, Roder С, Dieckmann D, et al. Generation of large numbers of fully mature and stable dendritic cells from leukapheresis products for clinical application. J Immunol Methods HD - 1305440 1999; 223:1-15.
37. Hartmann G, Weiner GJ, Krieg AM. CpG DNA: a potent signal for growth, activation, and maturation of human dendritic cells. Proc Natl Acad Sci USA JID - 7505876 1999; 96:9305-9310.
38. Kadowaki N, Liu YJ. Natural type I interferon-producing cells as a link between innate and adaptive immunity. Hum Immunol JID - 8010936 2002; 63:1126-1132.
39. Rothenfusser S, Tuma E, Endres S, Hartmann G. Plasmacytoid dendritic cells: the key to CpG(1). Hum Immunol JID - 8010936 2002; 63:1111-1119.
40. O'Garra A. Cytokines induce the development of functionally heterogeneous Т helper cell subsets. Immunity JID - 9432918 1998; 8:275-283.
41. Pulendran B, Smith JL, Caspary G, et al. Distinct dendritic cell subsets differentially regulate the class of immune response in vivo. Proc Nail Acad Sci USA JID - 7505876 1999; 96:1036-1041.
42. Sallusto F, Schaerli P, Loetscher P, et al. Rapid and coordinated switch in chemokine receptor expression during dendritic cell maturation. Eur J Immunol JID - 1273201 1998; 28:2760-2769.
43. Appay V, Rowland-Jones SL. RANTES: a versatile and controversial chemokine. Trends Immunol JID - 100966032 2001; 22:83-87.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИММУНОТЕРАПИИ ХРОНИЧЕСКОГО ВИРУСНОГО ГЕПАТИТА С | 2016 |
|
RU2637631C2 |
| КЛЕТОЧНЫЙ ПРОДУКТ ДЛЯ НАГРУЗКИ И АКТИВАЦИИ ДЕНДРИТНЫХ КЛЕТОК ЧЕЛОВЕКА | 2019 |
|
RU2714208C1 |
| ГЕНЕРИРОВАНИЕ ДЕНДРИТНЫХ КЛЕТОК ИЗ МОНОЦИТАРНЫХ ПРЕДШЕСТВЕННИКОВ ДЕНДРИТНЫХ КЛЕТОК С ПОМОЩЬЮ GM-CSF В ОТСУТСТВИЕ ДОПОЛНИТЕЛЬНЫХ ЦИТОКИНОВ | 2004 |
|
RU2364625C2 |
| СОВМЕСТНАЯ ДИФФЕРЕНЦИРОВКА МОНОЦИТОВ ОТ АЛЛОГЕННЫХ ДОНОРОВ | 2013 |
|
RU2668816C2 |
| СПОСОБ ИНДУКЦИИ ПРОТИВООПУХОЛЕВОГО ЦИТОТОКСИЧЕСКОГО ИММУННОГО ОТВЕТА in vitro С ПОМОЩЬЮ ДЕНДРИТНЫХ КЛЕТОК, ТРАНСФИЦИРОВАННЫХ РНК ОПУХОЛЕВЫХ КЛЕТОК, ПРОТИВ КЛЕТОК НЕМЕЛКОКЛЕТОЧНОГО РАКА ЛЕГКОГО | 2015 |
|
RU2578008C1 |
| КОМБИНИРОВАННЫЙ КЛЕТОЧНЫЙ ТРАНСПЛАНТАТ НА ОСНОВЕ ЛИМФОКИНАКТИВИРОВАННЫХ КИЛЛЕРОВ И ДЕНДРИТНЫХ КЛЕТОК, СПОСОБ ЕГО ПОЛУЧЕНИЯ И СПОСОБ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ ОНКОЛОГИЧЕСКИХ, ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ И ИММУНОДЕФИЦИТНЫХ СОСТОЯНИЙ | 2006 |
|
RU2309753C1 |
| СПОСОБ ГЕНЕРАЦИИ ЦИТОТОКСИЧЕСКИХ КЛЕТОК С АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК НЕМЕЛКОКЛЕТОЧНОГО РАКА ЛЕГКОГО | 2014 |
|
RU2577992C2 |
| Способ иммунотерапии рака молочной железы с помощью антиген-активированных дендритных клеток | 2016 |
|
RU2645464C1 |
| Способ стимуляции презентирующей активности дендритных клеток | 2019 |
|
RU2728592C1 |
| АУТОЛОГИЧНАЯ ВАКЦИНА ДЛЯ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2008 |
|
RU2392946C2 |


Изобретение относится к иммунологии и медицине. Предложенный способ предусматривает введение млекопитающему кондиционированной лимфоцитами среды, содержащей «необученные» Т-клетки, культивируемые с покрытыми антителами к CD3/CD28 гранулами, в комбинации с антигеном. Изобретение позволяет усилить ответ иммунной системы пациента на вакцину или другой иммунотерапевтический агент. Способ может быть использован в медицине. 9 з.п. ф-лы, 5 ил., 3 табл.
| BRICE G.T | |||
| et al., "Development of an animal model for autotransfusion therapy: in vitro characterization and analysis of anti-CD3/CD28 expanded cells", J Acquir Immune Defic Syndr Hum Retrovirol | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |

