Область техники
Настоящее изобретение предлагает метод получения неистощенных незрелых человеческих дендритных клеток (ДК), полученных из моноцитов.
Уровень техники
Традиционные методы лечения рака, такие как хирургия, лучевая и химиотерапия, часто оказываются не эффективными в лечении больных и, как правило, вызывают тяжелые побочные эффекты. Есть надежда, что иммунотерапия станет альтернативным методом лечения с меньшими нежелательными побочными эффектами.
В настоящее время установлено, что иммунная система включает клетки, в частности CD8+ цитотоксические Т-лимфоциты (Цтл), которые способны распознавать и, в потенциале, убивать опухолевые клетки. Тем не менее, серьезная проблема заключается в том, что способность уничтожать раковые клетки у этих Т-клеток в организме онкологических больных либо вообще не индуцируются, либо лишь слабо индуцируются. Одна из возможностей заключается том, что имеет место неадекватная презантация опухолевых антигенов и ко-стимуляция дендритными клетками (ДК), "природными адъювантами" для инициации функционального и опухолево-специфичного Т-клеточного иммунитета у онкологических больных.
Существующие стратегии иммунотерапии рака в основном сосредоточены на нагруженных антигенами аутологичных, взятых у пациента ДК, которые дифференцируют и нагружают антигенами ex vivo. Основной предпосылкой этого подхода является то, что эффективность и контроль, обеспечиваемые ex vivo манипуляцями с ДК, дают ДК с высокой способностью к презентации антигенов и костимуляции. Качество Т-клеточного ответа зависит от способности этих аутологичных ДК к ГКГС-ограниченному презентированию опухолевых антигенов Т-клеткам (ДК и Т-клетки должны быть от одного и того же индивидуума) в дренирующих лимфатических узлах и, соответственно, созданию опухолевоспецифического Т-клеточного ответа.
Аутологичные ДК, полученные из моноцитов, являются наиболее широко используемыми в экспериментальных исследованиях ДК, так как существует возможность получать миллиарды моноцитов из лейкоцитов периферической крови, собранных в ходе лейкафереза - трудоемкого и длительного процесса, в котором лейкоциты выделяют из циркулирующей крови. Существует несколько методов для последующего обогащения моноцитов, включая: элютриацию и выделение с использованием антител/гранул, которые могут выполняться в соответствии с принципами надлежащей производственной практики (GMP).
Затем моноциты культивируют в среде с добавлением ГМ-КСФ и IL-4 в течение 4-7 дней, что приводит к их дифференцировке в незрелые ДК, которые характеризуются выдающейся способностью продуцировать большое количество провоспалительных хемокинов и цитокинов при последующей стимуляции некоторыми типами активирующих факторов (Sallusto et al, Eur J Immunol, 1999. 29:1617; Napolitani et al, Natuer Immunology 2005. 6:769). Стимулированные ДК обычно предварительно нагружают соответствующими антигеном (-ами) опухоли и активируют в течение 1-2 дней перед вакцинацией. Тем не менее, иммунные ответы на такие ДК-вакцины часто слабы и клинические результаты редко бывают полными и продолжительными.
Мало что было известно о судьбе и функции получаемых ex vivo аутологичных ДК после введения. Недавно исследовали миграцию образца введенной ДК-вакцины in vivo и обнаружили, что менее 5% введенной ДК достигало дренирующих лимфатических узлов, а большинство ДК оставались в месте инъекции. Эти локально запертые ДК-вакцины быстро теряли свою жизнеспособность и впоследствии удалялись рекрутированными антигенпрезентирующими клетками.
Теперь получены данные, что введенные в составе вакцины ДК, которые активировали ex vivo в течение ограниченного периода времени (т.е. от 6 до 18 ч), становятся провоспалительными ДК, которые способны опосредованно активировать собственные CD8+ Т-клетки in vivo, действуя в качестве местного иммунного адъюванта. Эта адъювантная функция введенных провоспалительных ДК сильно зависит от секретирования ими определенных рекрутирующих ДК и НК-клети хемокинов в момент введения (после удаления факторов активации). Такие провоспалительные ДК также экспрессируют/секретируют факторы, которые вызывают активацию рекрутированных эндогенных НК-клеток и ДК в месте вакцинации. В отличие от провоспалительных ДК, ДК, активируемые в течение длительного времени (т.е. >24 ч) широко используются в клинических испытаниях и характеризуются «истощенным» состоянием (Langenkamp et al 2000), и, следовательно, не в состоянии вырабатывать желаемые хемокины и ДК-активирующие факторы во время введения.
Наконец, провоспалительные ДК могут действовать не только как прямые стимуляторы ГКГС-совместимых аутологичных Т-клеток, но также в качестве адъюванта продуцирования большого количества провоспалительных хемокинов и цитокинов в момент введения. Местное введение провоспалительных ДК приведет к рекрутингу и активации других иммунных клеток, в том числе циркулирующих НК-клеток и ДК-предшественников. Если введенные провоспалительные ДК были предварительно нагружены соответствующими опухолевыми антигенами или введены непосредственно в существующий опухолевой очаг, рекрутированные эндогенные ДК будут поглощать умирающие клетки вакцины, экспрессирующие соответствующие опухолевые антигены, или умирающие антиген-экспрессирующие опухолевые клетки, соответственно. После активации эти рекрутированные, а затем нагруженные антигенами эндогенные ДК будут мигрировать в дренирующие лимфатические узлы, где они активируют опухолевые Т-клетки по механизму, ограниченному ГКГС (Liu et al, 2008). Этот вывод подтверждается данными из нескольких недавних доклинических исследований, в которых рост опухоли значительно сокращался при применении терапевтических вакцин с неистощенными ГКГС-несовместимыми аллогенными провоспалительными ДК (Alder et al 2008, Siders et al 2009, Edlich et al 2010).
Сильная адъювантная функция провоспалительных ДК не нуждается в ГКГС-совместимости между провоспалительными ДК и Т-клетками пациента, что дает возможность использования предварительно подготовленных и хранящихся в замороженном состоянии ГКГС-несовместимых аллогенных провоспалительных ДК, как "готовых к использованию" вакцин, представляющих собой действенную, реальную альтернативу используемым в настоящее время изготавливаемым индивидуально пациент-специфическим ДК вакцинам. Использование таких ГКГС-несовместимых аллогенных провоспалительных ДК описано в ЕР 1509244 В1 и WO 2011/098516.
По этическим соображениям крупномасштабная заготовка моноцитов из нормальных доноров крови с помощью лейкафереза с единственной целью производства вакцин коммерческого масштаба для клинического использования не представляется возможной. В силу вышесказанного, на практике доступное сырье, то есть моноциты для производства провоспалительных ДК, ограничено моноцитами, полученными из побочных продуктов (лейкоцитарных пленок и/или использованных лейкоцитарных фильтров) в ходе выделения ненужных лейкоцитов из различных компонентов цельной крови, или моноцитами, полученными из лейкоцитарной пленки в банках крови.
Однако общее количество моноцитов, которые могут быть выделены из каждой лейкоцитарной пленки или из каждого лейкоцитарного фильтра из контейнера для крови обычно меньше 200 миллионов (Ebner S et al., Generation of large numbers of human dendritic cells from whole blood passaged through leukocyte removal filters: an alternative to standard buffy coats J Immunol Methods 252 (2001), что обуславливает неприемлемо высокую стоимость обогащения и последующей ДК-дифференцировки отдельных партий моноцитов (каждая партия получена от одного донора) методами, соответствующими принципам надлежащей производственной практики (GMP).
Таким образом, существует потребность в техническом способе для крупномасштабного и рентабельного производства неистощенных, незрелых дендритных клеток клинического качества из содержащих моноциты побочных продуктов, полученных из банков крови.
Краткое описание изобретения
Соответственно, настоящее изобретение направлено на уменьшение, смягчение, исключение или преодоление одного или более из вышеуказанных потенциальных сложностей в данной области и недостатков по отдельности, или в любой комбинации, за счет способа получения неистощенных незрелых дендритных клеток (ДК), полученных от по меньшей мере двух различных, аллогенных доноров. В способе используется смесь аллогенных лейкоцитов, которые были получены от по меньшей мере двух различных, аллогенных доноров. Затем аллогенные моноциты выделяют из смеси аллогенных лейкоцитов, в результате чего получают обогащенные моноцитами аллогенные лейкоциты. После этого из обогащенных моноцитами аллогенных лейкоцитов получают неистощенные незрелые ДК путем совместного культивирования в течение от 2 до 7 дней в водной культуральной среде без примесей сыворотки нечеловеческого происхождения с добавлением интерлейкина-4 (IL-4) и гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ).
В противоположность сложившемуся мнению получаемые моноцитарные незрелые дендритные клетки являются неистощенными и, соответственно, способны стабильно продуцировать значительное количество провоспалительных хемокинов и провоспалительных цитокинов после активации в провоспалительные ДК. Таким образом, данный способ представляет собой экономически эффективный способ крупномасштабного получения неистощенных незрелых дендритных клеток клинического качества.
Еще один аспект настоящего изобретения относится к смеси аллогенных неистощенных незрелых дендритных клеток (ДК), полученных от по меньшей мере двух различных, аллогенных доноров. Такая смесь может быть получена описанным способом.
Следующий аспект настоящего изобретения относится к способу получения провоспалительных ДК. В результате активации неистощенных незрелых ДК получают провоспалительные ДК. С помощью такого способа можно получить смесь аллогенных провоспалительных дендритных клеток, происходящих от по меньшей мере двух различных, аллогенных доноров. Смесь может быть приготовлена в виде фармацевтической композиции, дополнительно содержащей по меньшей мере один фармацевтически приемлемый носитель. Смесь и фармацевтическая композиция, соответственно, могут применяться в лечении рака.
Другие предпочтительные признаки изобретения определены в соответствующих пунктах формулы изобретения. Кроме того, предпочтительные признаки изобретения подробно описаны в вариантах реализации.
Подробное описание предпочтительных вариантов реализации
Авторы настоящего изобретения предполагали, что содержащие моноциты популяции лейкоцитов, присутствующие в лейкоцитарных пленках или задержанные лейкоцитарными фильтрами, используемыми для удаления лейкоцитов из цельной крови (используются для обогащения эритроцитов) или лейкоцитарными фильтрами, используемыми для удаления лейкоцитов из пула лейкоцитарных пленок (используются для обогащения тромбоцитов), потенциально могут быть использованы для крупномасштабного и экономически эффективного производства неистощенных незрелых дендритных клеток.
Как было показано ранее, лейкоциты, задержанные различными типами лейкоцитарных фильтров, могут быть выделены с помощью обратной промывки в подходящей среде с последующим обогащением моноцитами (Ebner S et al., Generation of large numbers of human dendritic cells from whole blood passaged through leukocyte removal filters: an alternative to standard buffy coats J Immunol Methods 252 (2001) 93-104; и Meyer T P H et al., Filter Buffy Coats (FBC): A source of peripheral blood leukocytes recovered from leukocyte depletion filters. J Immunol Methods 307 (2005) 150-166).
Лейкоцитарная пленка представляет собой фракцию образца крови с антикоагулянтом, которая содержит большое количество лейкоцитов, в том числе нейтрофилов, базофилов, эозинофилов, моноцитов, лимфоцитов, а также тромбоцитов, следующих в градиенте плотности цельной крови после центрифугирования. Лейкоцитарная пленка, как правило, используется в качестве сырья для получения тромбоцитов. В ходе этого процесса происходит уменьшение количества лейкоцитов с помощью лейкоцитарных фильтров, и его обычно проводят после объединения от 4 до 8 лейкоцитарных пленок. Как правило, для выделения тромбоцитов из лейкоцитарных пленок используется оборудование TACSI или система OrbiSac.
Оборудование TACSI для получения тромбоцитов из объединенных лейкоцитарных пленок содержит системный блок (закрепленный на роторе) и вставки, которая может быть удалена для установки комплекта TACSI. Каждый аппарат снабжен системой прессования, которая управляется и контролируется отдельным микропроцессором. На первом этапе внутри накопительных контейнеров, пулы лейкоцитарных пленок осаждают центрифугированием в вертикальном положении. На следующем этапе, слой супернатанта лейкоцитарных пленок, обогащенного тромбоцитами (также содержащий значительное количество лейкоцитов, включая моноциты и лимфоциты) переносят в контейнер для хранения путемактивации системы прессования в каждом боксе. Кроме того, в комплект TACSI включен фильтр для отделения лейкоцитов, находящийся между камерой обработки и конечным контейнером для хранения. И лейкоцитарный фильтр, и остаток лейкоцитарной пленки из контейнера для пула лейкоцитарных пленок, содержащие значительное количество моноцитов, затем выбрасывают.
В альтернативной системе автоматического обогащения тромбоцитов OrbiSac контейнер для пула лейкоцитарных пленок имеет форму кольца. После центрифугирования обогащенную тромбоцитами центральную часть супернатанта переносят в сосуд, расположенный в центре центрифуги. Перенос осуществляется с использованием интегрированного лейкоцитарного фильтра. И лейкоцитарный фильтр, и остаток лейкоцитарной пленки из контейнера для пула лейкоцитарных пленок, содержащие значительное количество моноцитов, которые, затем выбрасывают.
Таким образом, методы в области обогащения тромбоцитов из лейкоцитарных пленок обеспечивают два возможных источника моноцитов, а именно остатки лейкоцитарной пленки, обедненной тромбоцитами, и лейкоцитарные фильтры. Смесь лейкоцитов, содержащаяся в обедненной тромбоцитами лейкоцитарной пленке или отработанном лейкоцитарном фильтре, содержит до 1 миллиарда моноцитов. Тем не менее, эти моноциты аллогенны по отношению друг к другу, так как они происходят от различных аллогенных доноров, поскольку лейкоцитарные пленки объединяют перед удалением тромбоцитов.
Необъединенные лейкоцитарные пленки, содержащие тромбоциты, или фильтры, полученные после выделения лейкоцитов из цельной крови, могут максимально обеспечивать лишь от 100 до 200 миллионов моноцитов с каждой лейкоцитарной пленки или фильтра. Поэтому их необходимо объединять для получения количестве, достаточного для рентабельного производства неистощенных незрелых ДК, соответствующего требованиям GMP.
Подобно лейкоцитам, полученным в ходе производства тромбоцитов, объединенные лейкоциты из лейкоцитарных пленок, содержащие тромбоциты, или из фильтров, используемых для выделения лейкоцитов из цельной крови, будут также состоять из смешанной популяции клеток, полученной от различных аллогенных доноров.
Объединение лейкоцитов из по меньшей мере 5-10 лейкоцитарных пленок или объединение лейкоцитов, элюированных из по меньшей мере 5-10 лейкоцитарных фильтров для цельной крови, могли бы, по меньшей мере теоретически, решить проблему обеспечения достаточного количества лейкоцитов для крупномасштабного и экономически эффективного производства неистощенных незрелых ДК клинического качества.
Как было показано ранее, лейкоциты, задерживаемые лейкоцитарными фильтрами, могут быть извлечены путем обратной промывки подходящей средой с последующим обогащением моноцитов (Ebner S et al., Generation of large numbers of human dendritic cells from whole blood passaged through leukocyte removal filters: an alternative to standard buffy coats J Immunol Methods 252 (2001) 93-104; и Meyer T P H et al., Filter Buffy Coats (FBC): A source of peripheral blood leukocytes recovered from leukocyte depletion filters. J Immunol Methods 307 (2005) 150-166).
Однако, предполагаемая проблема, связанная с последующим совместным культивированием лейкоцитов, обогащенных моноцитами, которые были получены от разных, аллогенных доноров, заключается в том, что ожидается, что несовместимость по антигенам класса I и II главного комплекса гистосовместимости (ГКГС), приведет к преждевременной активации моноцитов/незрелых ДК от одного донора аллореактивными Т-клетками и/или НК-клетками от другого донора.
Как известно, совместное культивирование в стандартной среде для культивирования клеток, такой как RPMI-1640 (RPMI = Институт Розуэлла Парка, в котором Moore с соавт. разработали оригинальную среду) с фетальной бычьей сывороткой, или в бессывороточной среде для культивирования клеток, такой как X-VIVO 15, из мононуклеарных клеток, включая моноциты, лимфоциты и НК-клетки от двух аллогенных доноров индуцирует продукцию известных ДК-активирующих факторов, в том числе ФНО-альфа (Laurin et al, Transplantation 2004; 77:267; Wallgren et al, Scand J Immunol 2005; 62:234). Последующее добавление экстрактов данных культур в стандартную среду RPMI-1640 содержащую фетальную бычью сыворотку, или же в стандартную бессывороточную среду для аллогеничных мононуклеарных клеток вызывает индукцию активации/созревания незрелых мононуклеарных ДК (Laurin et al, Transplantation 2004; 77:267; Wallgren et al, Scand J Immunol 2005; 62:234). Кроме того, было показано что добавление ФНО-альфа в стандартную среду RPMI-1640 с добавлением ГМ-КСФ и IL-4, используемую для дифференцировки моноцитов в незрелые ДК, вызывает преждевременную активацию и последующее истощение (толерантность) дифференцированных ДК (Rieser С et al., Differential Deactivation of Human Dendritic Cells by Endotoxin Desensitization: Role of Tumor Necrosis Factor-α and Prostaglandin E2. Blood 91 (1998) 3112-3117).
Проблема загрязняющих Т-клеток и НК-клеток является актуальной даже после обогащения в соответствии по протоколам GMP (элютриация и выделение с использованием антител/гранул) моноцитов из объединенных лейкоцитарных пленок или из лейкоцитарных фильтров (Schwanke et al, Journal of Clinical Apheresis 21: 153-157 (2006); Meyer et al, Journal of Immunological Methods 307 (2005) 150-166) из-за трудностей в подготовке популяций клеток моноцитов без примесей Т-клеток и НК-клеток.
Кроме того, важно, что не только совместное культивирование с аллогенными лимфоцитами в стандартной среде, но и совместное культивирование моноцитов с аллогенными нейтрофилами в среде RPMI-1640 с добавлением фетальной бычьей сыворотки приводит к повышенной регуляции CD40, CD86 мембран и человеческого лейкоцитарного антигена (HLA)-DR на ДК, т.е. преждевременной активации, как было показано Meggiovanni и др (см. Journal of Leukocyte Biology, 2006; 79; 977-988). Было показано, что существенное удаление нейтрофилов из моноцитов в объединенных лейкоцитарных пленках или лейкоцитах, элюированных с лейкоцитарных фильтров, с использованием элютриации (Schwanke et al, Journal of Clinical Apheresis 21: 153-157 (2006) или выделения с использованием антител/гранул (Meyer et al, Journal of Immunological Methods 307 (2005) 150-166) трудноосуществимо. Обычно такой обогащенный моноцитами продукт содержит значительное количество (т.е. 25-40% от общего числа присутствующих клеток) нейтрофилов. Однако, с точки зрения безопасности загрязнение нейтрофилами не является проблемой.
Более того, даже если бы было возможно приготовить 100% чистую популяцию клеток моноцитов, это бы не исключало риск преждевременной активации в результате активных взаимодействий между моноцитами от различных аллогенных доноров. В недавней обзорной статье под названием "Происхождение и биология аллогенного ответа" («Origin and biology of the allogenic response»), уважаемыми и известными иммунологами Fadi G. Lakkis и Robert I. Lechler (cf. Cold Spring Harbor perspectives in medicine, Vol. 3, No. 8, 2013) был сделан вывод, что на самом деле существуют врожденные механизмы аллогенного распознавания. Авторы утверждают, что: "отторжение трансплантата не ограничивается позвоночными животными, наделенными адаптивными иммунными системами, но является общим для многих беспозвоночных организмов, которые эволюционно возникли до развития адаптивного иммунитета (животные, которые не имеют Т- и Б-лимфоцитов, НК-клеток, соматических генов перестройки ферментов и ГКГС)".
Также важно отметить, что аллогенные ответы наблюдаются также у мышей, лишенных лимфоидных клеток. Активация моноцитов зависит от различий в антигенах, не относящихся к ГКГС, между моноцитами реципиента и введенными аллогенными лейкоцитами донора, в том числе аллогенными моноцитами (Zecher D et al., An Innate Response to Allogeneic Nonself Mediated by Monocytes. J Immunol 83 (2009) 7810-7816). Zecher и др., показали, что введение аллогенных лейкоцитов в ушную раковину мыши RAG2/2 не имеющей Т- и В-лимфоцитов, вызывает значительно большую опухоль и инфильтрацию кожи миелоидными клетками хозяина, чем введение сингенных лейкоцитов. Ответ на аллогенные лейкоциты возникал независимо от НК-клеток и осуществлялся моноцитами. Кроме того, ответ моноцитов аллоантигенную детерминанту не был связана с ГКГС.
В недавней статье (Zeng Q, et al. "Innate recognition of allogeneic non-self induces monocyte differentiation to mature dendritic cells in vivo." Am J Transplant 12: 148-148, 2012), авторы показали, что аллотрансплантанты сердца, пересаженные мышам GC2/2RAG2/2 не имеющим Т-, В-, и НК-клеток, быстро инфильтрировались моноцитами хозяина, которые дифференцировались в зрелые IL-12-экспрессирующие дендритные клетки (ДК). Однако детерминанты в аллогенных клетках, вызывающие созревание моноцитов хозяина, и возможные рецепторы моноцитов, которые распознают их, пока не известны.
Таким образом, очевидно, что моноциты млекопитающих непосредственно реагируют на не связанные с ГКГС детерминанты на аллогенных клетках независимо от Т-, В-, и НК-клеток. Следовательно, в соответствии со сложившимся общим представлением, аллореактивность считается общим свойством моноцитов.
Совместно эти данные указывают на то, явно ожидается, что совместное культивирование клеточных популяций, обогащенных моноцитами, полученных от различных аллогенных доноров всегда приводит к предварительной активации и последующему истощению моноцитов в ходе их дифференцировки в моноцитарные ДК. Таким образом, ДК не могут стать провоспалительными ДК, стабильно продуцирующими необходимые адьювантные факторы при повторном стимулировании соответствующими активирующими факторами.
Это ожидаемое индуцируемое активацией истощение аналогично известному истощению моноцитов, макрофагов и ДК, индуцированное преждевременной активацией воспалительными агентами, такими как ФНО-α (Park et al, Nat Immunol. 2012; 12: 607-615) или микробными липополисахаридами (ЛПС) (Rieser С et al., Differential Deactivation of Human Dendritic Cells by Endotoxin Desensitization: Role of Tumor Necrosis Factor-α and Prostaglandin E2. Blood 91 (1998) 3112-3117; Langenkamp A et al., Kinetics of dendritic cell activation: impact on priming TH1, TH2 and nonpolarized T cells. Nature Immunol. 1 (2000) 311-316;).
Однако авторы настоящего изобретения неожиданно обнаружили, что неистощенные незрелые ДК на самом деле могут быть получены из исходной клеточной популяции, состоящей из смеси обогащенных моноцитами аллогенных лейкоцитов от различных аллогенных доноров, способом, сходным с тем, который используется для размножения неистощенных незрелых ДК из обогащенных моноцитов, которые получены от одного донора (см WO 2011/098516), т.е. с использованием водной культуральной среды без примесей сыворотки нечеловеческого происхождения, но с добавлением интерлейкина-4 (IL-4) и гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ).
Вопреки превалирующему мнению неожиданно было показано, что получение незрелых ДК из смеси обогащенных моноцитов из разных, аллогенных доноров, в определенных условиях неожиданно показало, не приводит к преждевременной активации и последующему истощению ДК. Эти условия включают в себя совместное культивирование в водной культуральной среде без примесей сыворотки нечеловеческого происхождения и с добавлением ГМ-КСФ и IL-4.
Однако, совместное культивирование в водной культуральной среде без примесей сыворотки нечеловеческого происхождения и без добавления интерлейкина-4 (IL-4) и гранулоцитарно-макрофагального колониестимулирующего фактора, так же как и совместное культивирование в водной культуральной среде, содержащей сыворотку нечеловеческого происхождения, например, фетальную бычью сыворотку, с добавлением интерлейкина-4 (IL-4) и гранулоцитарно-макрофагального колониестимулирующего фактора же, как и ожидалось на основании общих знаний, приводит к преждевременной активации и последующему истощению ДК.
Таким образом, моноциты могут быть выделены из объединенных популяций лейкоцитов от разных аллогенных доноров, что делает возможным проведения крупномасштабного и экономичного соответствующего требованиям GMP обогащения моноцитов, такими способами как элютриация (Elutra) или выделение с использованием антител/гранул (CliniMacs). Затем из выделенных обогащенных моноцитами аллогенных лейкоцитов могут быть получены неистощенные незрелые ДК без преждевременной активации.
Такие неистощенные незрелые ДК могут применяться получения провоспалительных ДК, которые могут действовать в качестве противоопухолевой вакцины при введении опухоль (см WO 2011/098516). Кроме того, неистощенные незрелые ДК от разных аллогенных доноров могут быть нагружены антигеном (-ами) опухоли перед активацией с получением "готовой" клеточной аллогенной вакцины против рака (см. ЕР 1509244 В1), которую можно инъецировать в различные области, включая внутриопухолевые, подкожные, кожные, внутримышечные и/или внутривенные области.
Таким образом, один из вариантов реализации изобретения относится к способу получения неистощенных незрелых ДК из смеси обогащенных моноцитами аллогенных лейкоцитов. В этом методе используется смесь аллогенных лейкоцитов, полученная от по меньшей мере двух различных аллогенных доноров. В соответствии с одним из вариантов реализации изобретения «два разных аллогенных, донора» означает, что два индивидуума, являющиеся донорами лейкоцитов, относятся к одному виду, различаются по составу генов, т.е. различаются по антигенам. Как уже было описано, аллогенные лейкоциты могут быть получены из объединенных лейкоцитарных пленок или элюированием лейкоцитов из использованных лейкоцитарных фильтров. Затем аллогенные моноциты выделяют из смеси аллогенных лейкоцитов. Затем из выделенных обогащенных моноцитами аллогенных лейкоцитов получают неистощенные незрелые ДК.
Помимо возможности осуществления крупномасштабного и экономичного обогащения моноцитами в соответствии с требованиями GMP, еще одним преимуществом неистощенных незрелых ДК, получаемых из смеси аллогенных моноцитов, полученных от по меньшей мере двух различных аллогенных доноров, является то, что будет снижена нормальная биологическая вариация при получении различных провоспалительных факторов при активации, как известно, существующей между провоспалительными ДК от разных доноров.
В предпочтительном варианте реализации изобретения аллогенные лейкоциты получают путем объединения по меньшей мере двух лейкоцитарных пленок, содержащих лейкоциты. Объединенные лейкоцитарные пленки получают от по меньшей мере двух различных аллогенных доноров. Объединенные лейкоцитарные пленки могут как содержать тромбоциты, так и быть лишены их.
Аллогенные лейкоциты могут быть также получены элюированием лейкоцитов из по меньшей мере двух лейкоцитарных фильтров, которые, соответственно, ранее были использованы для отделения лейкоцитов от цельной крови по меньшей мере двух различных аллогенных доноров. После элюирования полученные лейкоциты собирают с получением смеси аллогенных лейкоцитов. Очевидно, хотя этот вариант менее предпочтителен, также возможно объединение цельной крови перед удалением лейкоцитов. Процедура элюирования из лейкоцитарных фильтров, которые ранее были использованы для выделения лейкоцитов из цельной крови, описана Ebner и др. (см. Journal of Immunological Methods 252 (2001) 93-104).
Аналогичным образом, аллогенные лейкоциты могут быть также получены элюированием лейкоцитов из лейкоцитарного фильтра, который был использован для выделения лейкоцитов из объединенных лейкоцитарных пленок, причем объединенные лейкоцитарные пленки получены от по меньшей мере двух различных аллогенных доноров. Процедура элюирования лейкоцитов из использованного фильтра, использовавшегося для выделения лейкоцитов из лейкоцитарной пленки, описана Meyer с соавт. (см. Journal of Immunological Methods 307 (2005) 150-166).
Хотя такие аллогенные лейкоциты, полученные из лейкоцитарного фильтра, также могут быть использованы для получения неистощенных незрелых ДК, по-видимому, предпочтительнее использовать аллогенные лейкоциты, полученные объединением по меньшей мере двух лейкоцитарных пленок, полученных от по меньшей мере двух различных аллогенных доноров. Аллогенные лейкоциты, элюированные из лейкоцитарных фильтров, могут после созревания продуцировать несколько сниженное количество хемокинов, за исключением MIG и цитокинов по сравнению с аллогенными моноцитами, полученными непосредственно из объединенных образцов периферической крови или из объединенных лейкоцитарных пленок.
Выделение моноцитов из смеси различных лейкоцитов хорошо известно в данной области. В соответствии с одним из вариантов реализации изобретения, моноциты выделяют из смеси аллогенных лейкоцитов, полученных известными методами производства в соответствии с принципами GMP. Таким образом, моноциты могут быть выделены смеси аллогенных, полученной лейкоцитов путем элютриации или путем выделения с использованием антител/гранул.
Элютриация является методом, в котором промывка в непрерывном противопотоке разделяется клетки на несколько фракций. В общем виде, постоянная центробежная сила, которая разделяет клетки по плотности, противостоит постепенно увеличивающемуся потоку среды, проходящей через осадок, разделяя клетки по размеру. Соответственно, самые маленькие/легкие частицы выходят первыми, а самые крупные/тяжелые - последними, поток среды разделяет клетки на несколько продуктов.
Выделение моноцитов с использованием антител/гранул осуществляется путем (иммуно)-магнитно-активированной сортировки клеток (MACS). MACS является широко используемым методом селективного выделения клеток из цельной крови и лейкоцитарных пленок. В общем виде, несущие CD-специфические антитела, несущие ферромагнитные гранулы на своих Fc-концах связываются с целевыми (положительная селекция) или нежелательными (отрицательная селекция) клетками. Обработанные таким образом клетки могут быть задерживаться пористой колонком с металлическим покрытием при воздействии сильного магнитного поля.
После выделения моноциты дифференцируют в незрелые ДК, то есть получают незрелые ДК. Незрелые ДК получают путем совместного культивирования аллогенных моноцитов в водной культуральной среде, не содержащей клеток сыворотки нечеловеческого происхождения и с добавлением гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ) в сочетании с интерлейкином-4 (IL-4) в течение от 2 до 7 дней, например, примерно 5 дней, что приводит к дифференцировке моноцитов в незрелые ДК.
Поскольку в клеточной культуральной среде, содержащей фетальную бычью сыворотку, было отмечена преждевременная активация несмотря на добавление ГМ-КСФ и IL-4, важно чтобы используемая среда не содержала сыворотки нечеловеческого происхождения.
Незрелые ДК могут также быть получены путем культивирования аллогенных моноцитов в водной среде, содержащей ГМ-КСФ в сочетании с интерлейкином-2 (IL-2), интерлейкином-15 (IL-15) или интерфероном-альфа в течение от 2 до 7 дней, например, 5 дней, и, в результате чего происходит дифференцировка моноцитов в незрелые ДК. Использование ГМ-КСФ в сочетании с IL-4, однако, предпочтительнее, поскольку он, как было показано, предотвращает аллореактивность и преждевременную активацию при использовании в комбинации со средой, не содержащей сыворотки нечеловеческого происхождения.
Специалисты в данной области знакомы с культуральной средой для выращивания клеток, а также с ее компонентами. Как правило, используемая культуральная среда включает:
по меньшей мере одну соль, например NaCl, KCl, MgSO4 и/или Ca(NO3)2;
по меньшей мере один сахар, например, глюкоза;
и одну или несколько аминокислот, например, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин, L-валин, L-аргинин, L-аспарагин L-аспарагиновой, L-цистин, L-глутамин, L-глутаминовую кислоту, глицин, L-гистидин, L-гидроксипролин, L-изолейцин, L-лейцин и/или L-лизин;
один или несколько витаминов и других важных питательных веществ, таких как: глутатион, биотин, витамин В12, D-пантотенат кальция, холина хлорид, фолиевая кислота, миоинозит, никотинамид, п-аминобензойная кислота, пиридоксин, рибофлавин и/или тиамин и по меньшей мере один буфер, такой как фосфатная соль (например, NaHPO4) и/или карбонат соли (например, NaHCO3).
В соответствии с одним из вариантов реализации изобретения, культуральная среда содержит по меньшей мере одну соль, например NaCl, по меньшей мере один сахар, такой как глюкоза, одну или несколько аминокислот, один или несколько витаминов, и буфер, такой как фосфатная соль (например, NaHPO4) и/или карбонатная соль (например, NaHCO3).
Кроме того, хотя клеточная культуральная среда дополнительно не содержит сыворотки нечеловеческого происхождения, она обычно содержит по меньшей мере человеческий полипептид. В соответствии с одним из вариантов реализации изобретения клеточная культуральная среда содержит по меньшей мере один человеческий полипептид, выбранный из группы, состоящей из трансферрина, альбумина и инсулина; предпочтительная среда для культивирования клеток содержит все три из них. Человеческий полипептид может быть получен из человеческой плазмы. Кроме того, они могут быть получены рекомбинантным путем. В качестве примера, инсулин может быть получен рекомбинантным путем в клетках дрожжей.
В качестве примера, клеточная культуральная среда, не содержащая сыворотки нечеловеческого происхождения может представлять собой CellGro®, которая является бессывороточной средой для дендритных клеток (ДК), соответствующей требованиям GMP, поставляемой CellGenix GmbH. В США эта среда продается под торговой маркой CellGenix™.
В соответствии с принятым в данной области пониманием и приведенным здесь объяснением термин "выделен" не обязательно относится к 100%-ной чистоте, а относится к моноцитам, полученным в процессе выделения, селективном в отношении моноцитов. Моноциты, полученные таким способом, могут быть отнесены к обогащенным моноцитами аллогенным лейкоцитам, поскольку помимо моноцитов будут также содержать другие лейкоциты.
В соответствии с одним из вариантов реализации изобретения аллогенные моноциты получают путем обогащения смеси аллогенных лейкоцитов. Обогащенные моноцитами аллогенные лейкоциты, помимо моноцитов обычно содержат также аллогенные нейтрофилы. Кроме того, они могут содержать и другие гранулоциты.
Вопреки сложившемуся мнению, не наблюдалось никаких признаков преждевременной активации при совместном культивировании аллогенных моноцитов в водной культуральной среде, не содержащей сыворотки нечеловеческого происхождения и с добавлением ГМ-КСФ/IL-4, несмотря на тот факт, что незрелые ДК были получены из смеси аллогенных лейкоцитов. Соответственно, такие незрелые ДК являются неистощенными, и таким образом, в состоянии стабильно продуцировать значительное количество, например, более 2000, 5000, или 7 500 пг/мл, провоспалительных хемокинов, в том числе MIP-1 альфа, MIP-1 бета, RANTES и MIG, и существенное количество, например, 500, 1500 или 3000 пг/мл, провоспалительных цитокинов, в том числе IL-12p70 и ФНО-альфа после удаления активирующих факторов.
В соответствии с одним из вариантов реализации изобретения термин «незрелые» обозначает ДК, которые экспрессируют только низкие уровни маркеров созревания ДК, CD83 и CD86, и которые способны продуцировать большое количество провоспалительных хемокинов и цитокинов после активации. Низкие уровни, в соответствии с одним из вариантов реализации изобретения, следует понимать в том смысле, что после активации наблюдаются увеличение CD83-экспрессии по меньшей мере в 3-5 раз, и что после активации наблюдается увеличение CD86-экспрессии по меньшей мере в 5-8 раз.
Поскольку ожидалась преждевременная активация, в некоторых экспериментах добавляли ингибитор циклооксигеназы-2 NS-398 - известного фактора, препятствующего опосредуемому простагландином Е2 (PGE2) истощению активированных ДК. Тем не менее, присутствие NS-398 в ходе культивирования моноцитов в ДК не увеличивало, а уменьшило индуцированную активацией продукцию MIG и IL-12p70. Таким образом, нет никаких признаков опосредуемого PGE2 истощения дифференцированных незрелых ДК из совместных культур смешанных аллогенных моноцитов.
Как известно в данной области техники (см, например, ЕР 1509244 В1 и WO 2011/098516), неистощенные незрелые дендритные клетки (ДК) можно применять в производстве фармацевтических композиций для лечения рака. Таким образом, один из вариантов реализации изобретения относится к смеси аллогенных неистощенных незрелых дендритных клеток (ДК), полученных от по меньшей мере двух различных аллогенных доноров. Такие дендритные клетки (ДК) могут быть получены описанным здесь способом. После дифференцировки в незрелые ДК незрелые ДК могут быть активированы, в результате чего они станут провоспалительными ДК. Активация может быть достигнута несколькими способами. Было показано, что многие сигналы вызывают по меньшей мере некоторые аспекты активации ДК. К наиболее мощным из них относятся микробные и вирусные продукты (патоген-ассоциированные молекулярные паттерны (РАМО), которые непосредственно распознаются паттерн-распознающими рецепторами (PRR), в том числе рецепторами из группы Toll-подобных (TLR)). PRR контролируют экспрессию множества генов врожденного ответа и могут непосредственно вызывать активацию ДК. Кроме того, передача сигналов рецепторов PRR в иммунных и не иммунных клетках часто приводит к синтезу провоспалительных цитокинов, таких как фактор некроза опухоли (ФНО) и интерлейкин 1 (IL-1), которые также могут способствовать активации ДК. Таким образом, добавление воспалительных цитокинов может также способствовать активации незрелых ДК.
В соответствии с одним из вариантов реализации изобретения для получения клеточной аллогенной противораковой вакцины незрелые ДК загружаются антигенами до или одновременно с активацией нагружают антигенами. Нагрузка антигенами хорошо известна в данной области (см, например, ЕР 1509244 В1) и может осуществляться с использованием таких методов, как пульсация, трансфекция, инфекция или слияние. В качестве примера, антигены могут быть получены из опухоли; обычно используется тот же тип опухоли, против которой должна быть направлена вакцина. При получении антигенов, как правило, используется образец интересующего типа рака.
В соответствии с предпочтительным вариантом реализации изобретения активация незрелых ДК осуществляется в соответствии со способом, описанным в WO 2011/098516. Созревание, таким образом, может быть обеспечено добавлением лиганда Toll-подобного рецептора 3 (TLR3) поли (I:С), лиганда TLR7/8, такого как R848 (Резиквимод) и цитокин интерферон гамма (ИФН-γ). Лиганд поли(I:С) Toll-подобного рецептора 3 (TLR3) представляет собой синтетический аналог дцРНК, содержащий цепь поли (I), соединенную с цепью поли(С). Размер нити может быть различным. Размер может составлять от 200 пар оснований до 8000 пар оснований, например от 200 до 1500 или от 1500 до 8000 пар оснований. В данной области лиганд TLR7/8 R848 также называется "Резиквимод". В качестве альтернативы Резиквимоду в качестве лиганда TLR7/8 могут использоваться Гардиквимод или Имиквимод. Как правило, незрелые ДК подвергают воздействию факторов активации в течение от 8 до 24 часов, например 18 часов.
Активация может дополнительно включать добавление по меньшей мере одного вещества, выбранного из группы, состоящей из лигандов TLR2, лигандов TLR4, таких как бактериальный липополисахарид и монофосфориллипид А, лигандов TLR9, таких как последовательности олигонуклеотидов CpG (ODN), которые отличают микробные ДНК от ДНК млекопитающих, интерферон альфа (ИФН-α), интерлейкин 1β (IL-1β) и фактор некроза опухоли альфа (ФНО-α). Кроме того, активация предпочтительно не включают добавление простагландина Е2 (PGE2) для предотвращения превращения зрелых ДК в мигрирующие ДК, которые быстро покинут место инъекции (опухоли), что было бы невыгодно в контексте настоящего изобретения.
После активации полученные провоспалительные ДК могут быть промыты для по существу полного удаления факторов активации. Таким образом, перед использованием провоспалительных ДК в качестве вакцины факторы активации обычно отмывают. Удаление факторов активации позволяет избежать совместного введения факторов активации (направленных на активацию провоспалительных ДК ex vivo). Совместное введение факторов активации скорее всего приведет к сильной и стойкой активации рекрутированных внутрь опухоли незрелых ДК, что приведет к их дифференцировке в провоспалительные зрелые ДК, а не к целевой дифференцировке в мигрирующие зрелые ДК.
Как уже было описано (см. WO 2011/098516), провоспалительные дендритные клетки могут применяться в лечении рака, так как они могут активировать собственные ДК пациента и развиваться в нагруженные опухолевыми антигенами мигрирующие ДК. Соответственно, один из вариантов реализации изобретения относится к смеси аллогенных провоспалительных дендритных клеток, полученных от по меньшей мере двух различных аллогенных доноров. Такие аллогенные провоспалительных ДК могут быть получены способом, описанным здесь. Замораживание провоспалительных ДК после активации обеспечивает возможность их хранения. Обычно провоспалительные ДК замораживают в среде, содержащей диметилсульфоксид (ДМСО) и сыворотку или плазму. Перед использованием замороженные клетки размораживают, а ДМСО отмывают.
В лечении рака такие провоспалительные дендритные клетки можно применять в виде фармацевтической композиции. Фармацевтическая композиция может содержать по меньшей мере один фармацевтически приемлемый носитель, такой как фосфатно-буферный солевой раствор, вода, и эмульсии, такие как эмульсия масло/вода или вода/масло, а также различные типы смачивающих агентов. Кроме того, она может содержать фармацевтически приемлемые вспомогательные вещества, наполнители, консерванты, стабилизаторы и/или другие компоненты, известные в данной области. В качестве примера, носителем может быть физиологический раствор, содержащий человеческий сывороточный альбумин.
Другой вариант реализации изобретения относится к такой смеси аллогенных провоспалительных ДК, или к такой композиции, содержащей аллогенные провоспалительне ДК, для применения в лечении рака. Аналогичным образом, один из вариантов реализации изобретения относится к применению смеси аллогенных провоспалительных ДК для применения в изготовлении лекарственного средства для лечения рака. Еще один вариант реализации относится к способу лечения рака, в котором смесь аллогенных провоспалительных ДК вводят пациенту, нуждающемуся в таком лечении, в дозе, достаточной для активации собственных ДК пациента с получением нагруженных опухолевыми антигенами мигрирующих ДК.
Не вдаваясь в дальнейшие детали, считается, что специалист в данной области может использовать предшествующее описание и следующую далее экспериментальную часть для применения данного изобретения в его полном объеме. Предпочтительные конкретные варианты реализации, описанные в данном документе, должны пониматься исключительно в качестве иллюстрации, а не ограничения остальной части описания. Кроме того, хотя настоящее изобретение описано выше со ссылкой на конкретные варианты реализации, не предполагается, что оно ограничивается какой-либо конкретной формой, приведенной в данном документе. Наоборот, настоящее изобретение ограничено только прилагаемой формулой изобретения, и варианты реализации изобретения, не описанные выше, например, отличающиеся от описанных выше, в равной степени возможны в пределах объема прилагаемой формулы изобретения.
В формуле изобретения термин «содержит/содержащий» не исключает присутствия других элементов или этапов. Кроме того, хотя отдельные признаки могут быть включены в различные пункты формулы изобретения, возможны их комбинации, которые могут быть предпочтительными, и включение в различные пункты формулы изобретения не подразумевает, что комбинация признаков не представляется возможной, и/или преимущественной.
Кроме того, описание в единственном числе не исключает множественного. Термины "первый", "второй" и т.д., например, не исключают множества.
Краткое описание графических материалов
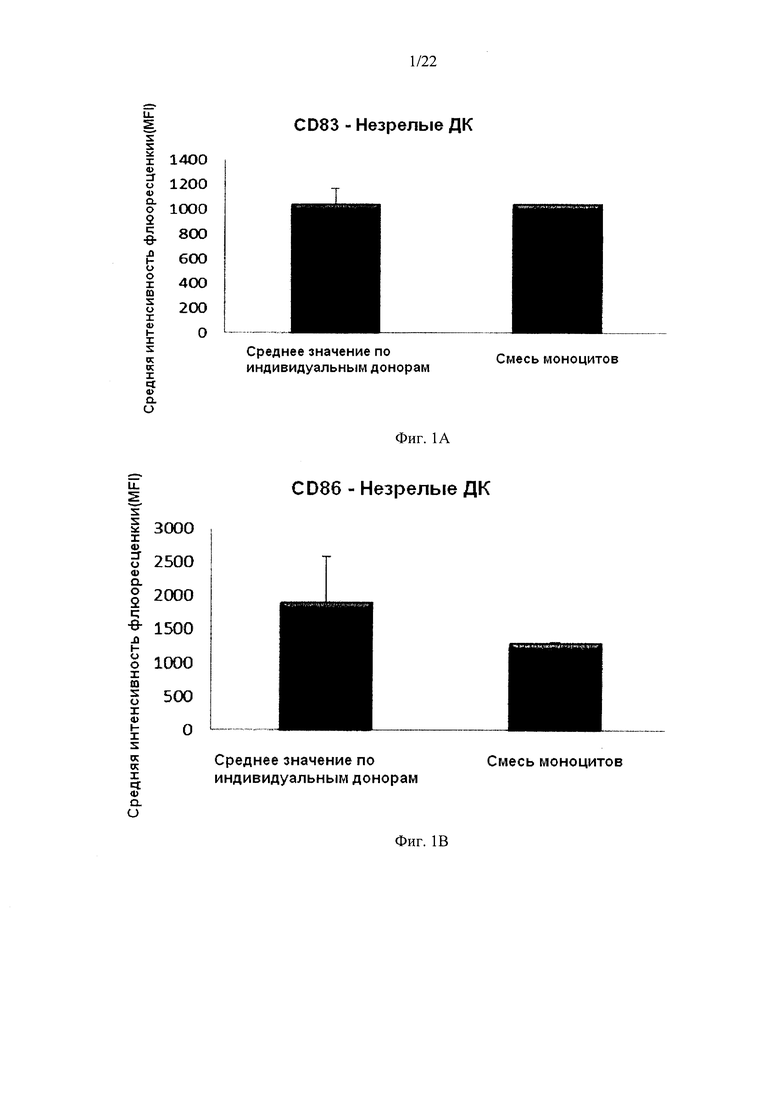
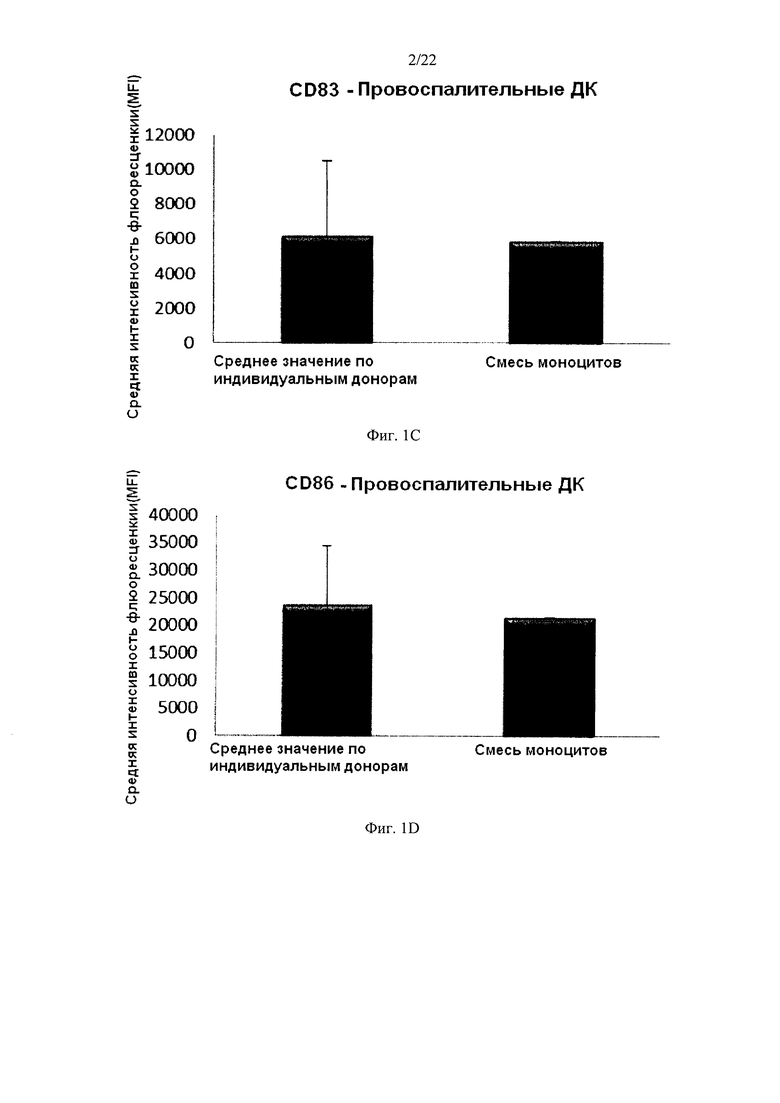
На Фиг. 1 показана экспрессия маркеров активации/созревания CD86 и CD83 на незрелых ДК и провоспалительных ДК, полученных из индивидуальных или смешанных культур моноцитов периферической крови.
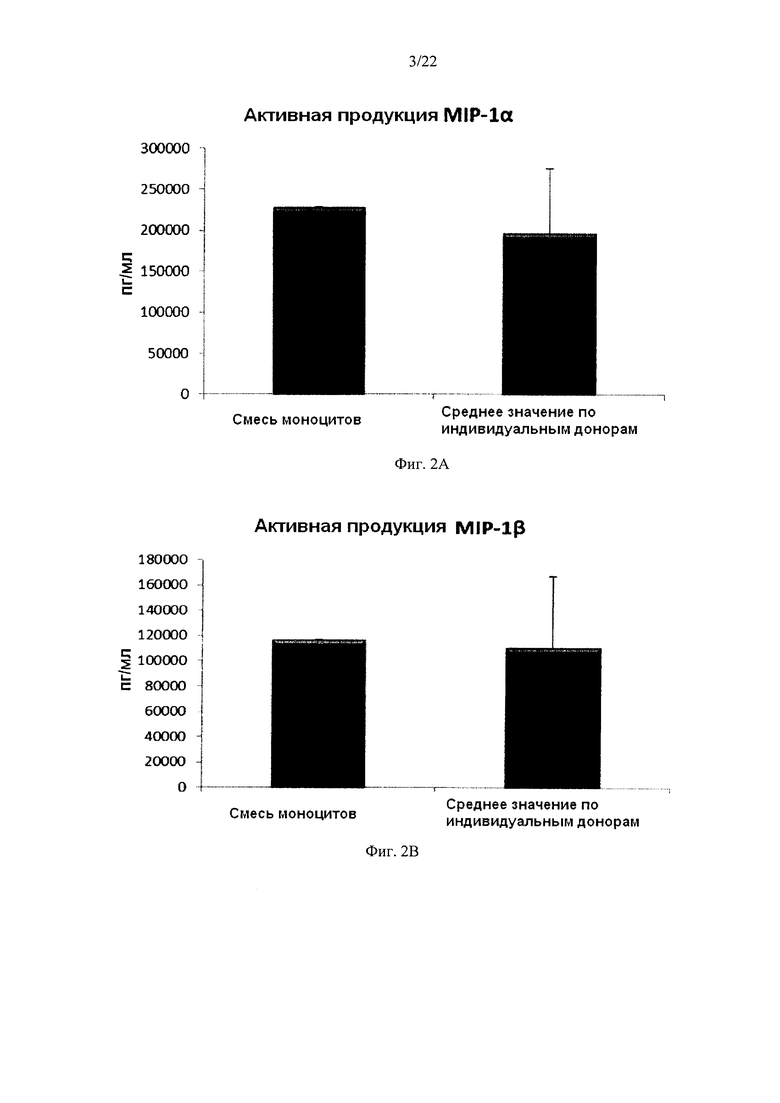
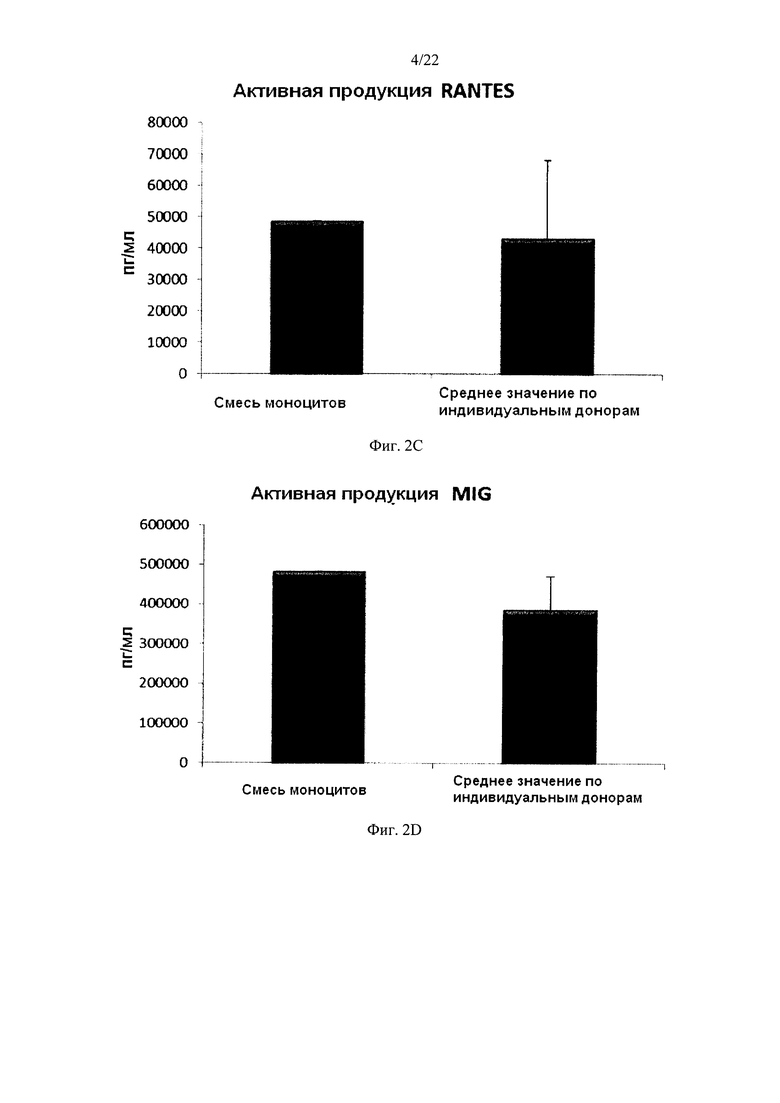
На Фиг. 2 показана продукция провоспалительных хемокинов незрелыми ДК (полученными из индивидуальной или смешанной культуры моноцитов периферических крови) в течение 18 часов непрерывной стимуляции в присутствии факторов активации ("Активная продукция").
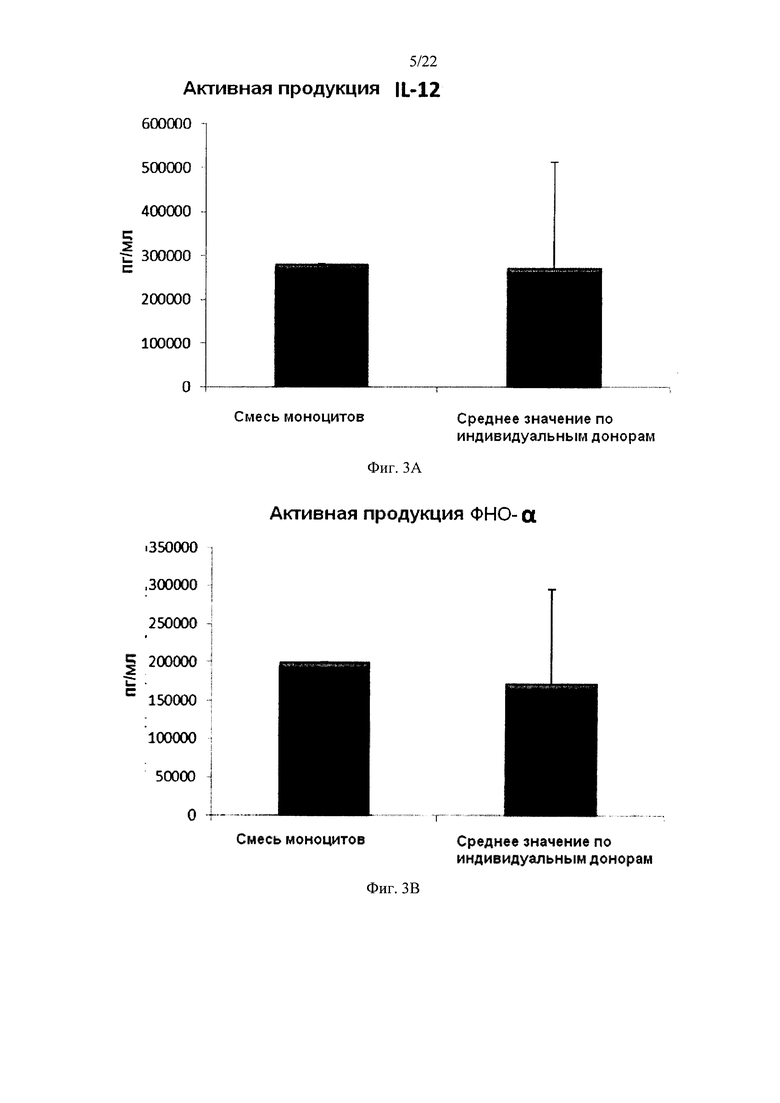
На Фиг. 3 показана продукция провоспалительных хемокинов незрелыми ДК (полученными из индивидуальной или смешанной культуры или моноцитов периферических крови) в течение 18 часов непрерывной стимуляции в присутствии факторов активации ("Активная продукция ").
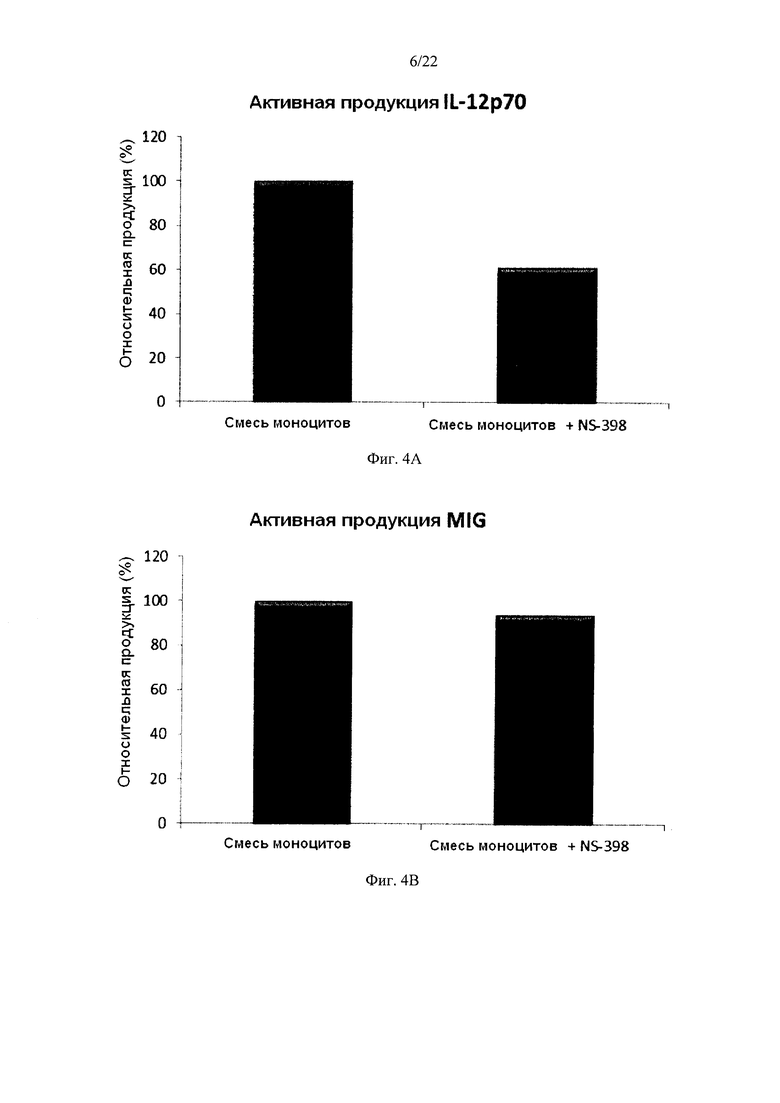
На Фиг. 4 показана продукция провоспалительных цитокинов незрелыми ДК (полученными из индивидуальной культуры или смешанной культур моноцитов периферической крови) в течение 18 часов непрерывной стимуляции в присутствии факторов активации +/- добавление ингибитора циклооксигеназы-2 (ЦОГ-2) NS-398.
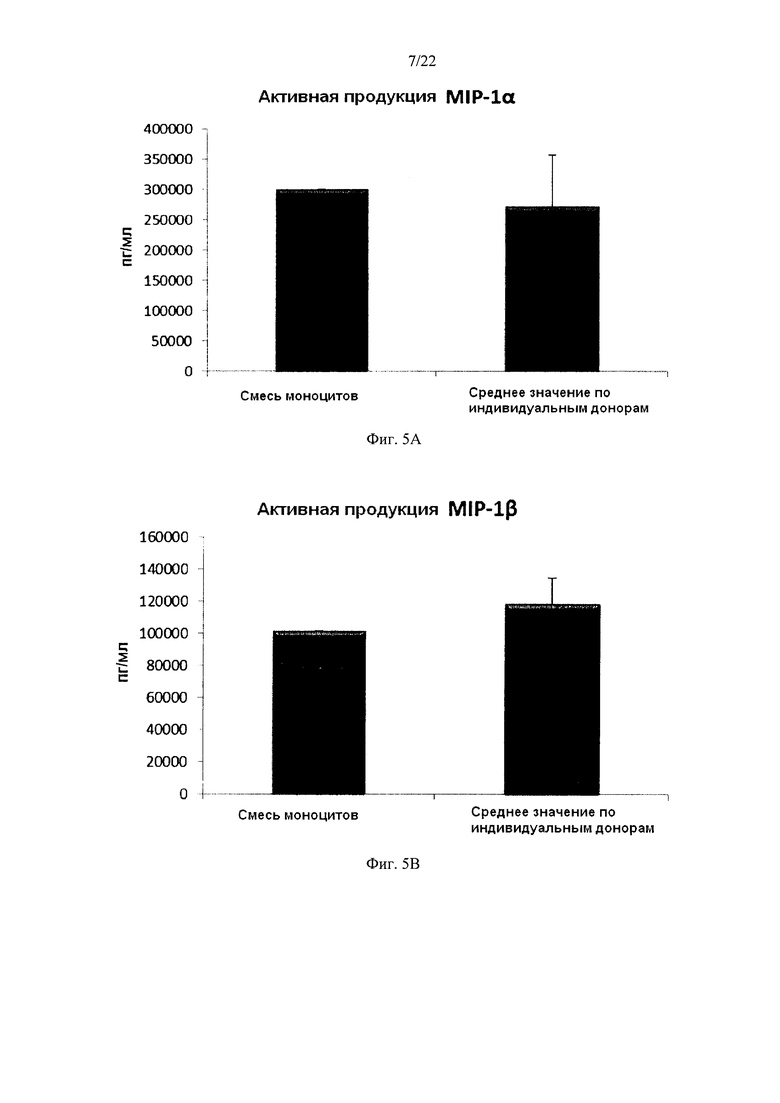
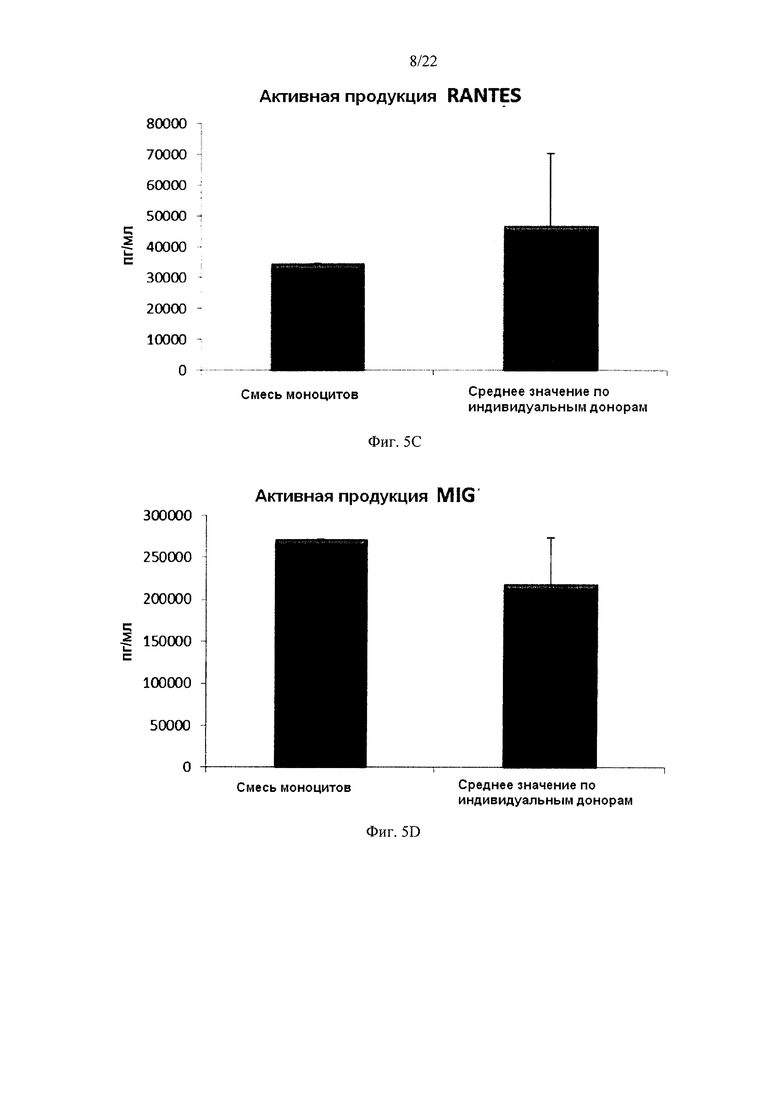
На Фиг. 5 показана продукция провоспалительных хемокинов незрелыми ДК (полученными из индивидуальной культуры или смешанной культуры моноцитов лейкоцитарных пленок) в течение 18 часов непрерывной стимуляции в присутствии факторов активации ("Активное продуцирование").
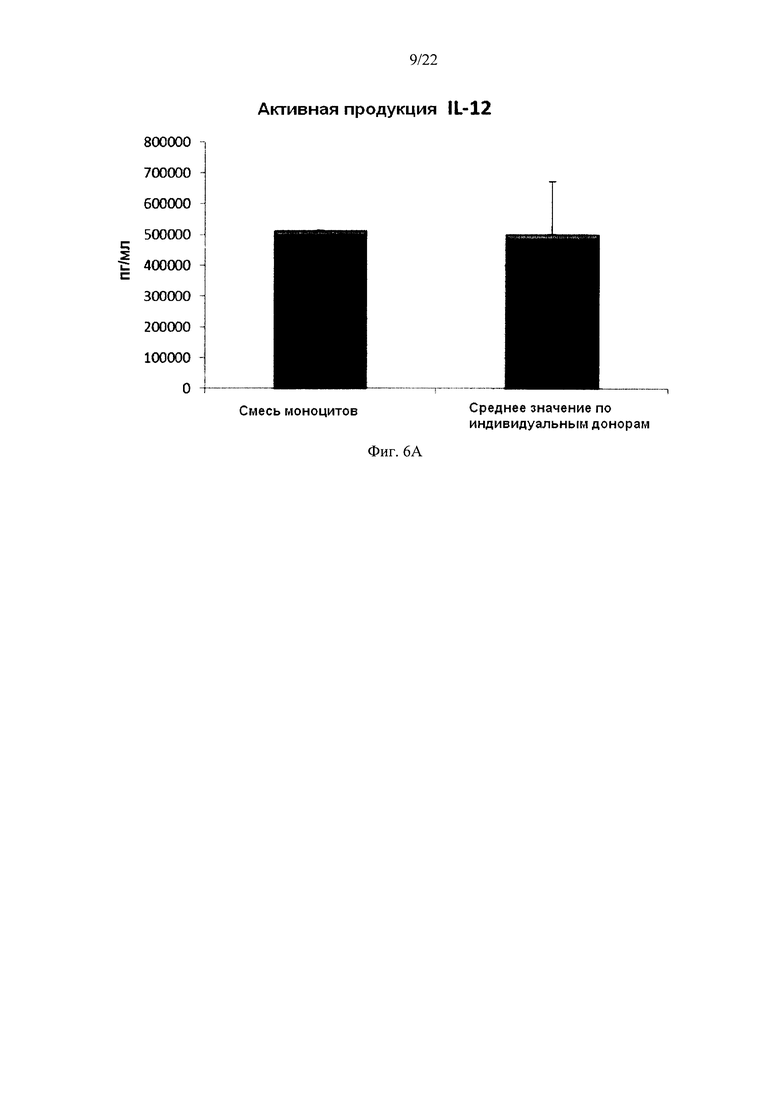
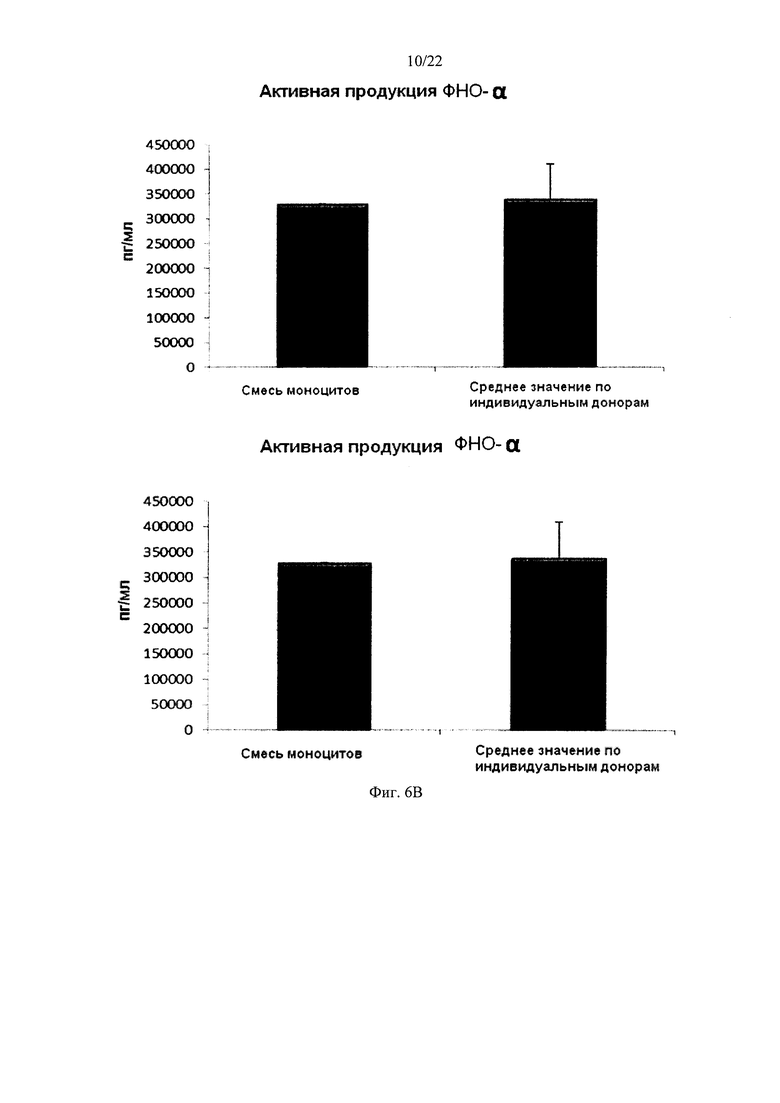
На Фиг. 6 показана продукция провоспалительных цитокинов незрелыми ДК (полученными из индивидуальной культуры или смешанной культуры моноцитов лейкоцитарных пленок) в течение 18 часов непрерывной стимуляции в присутствии факторов активации ("Активное продуцирование").
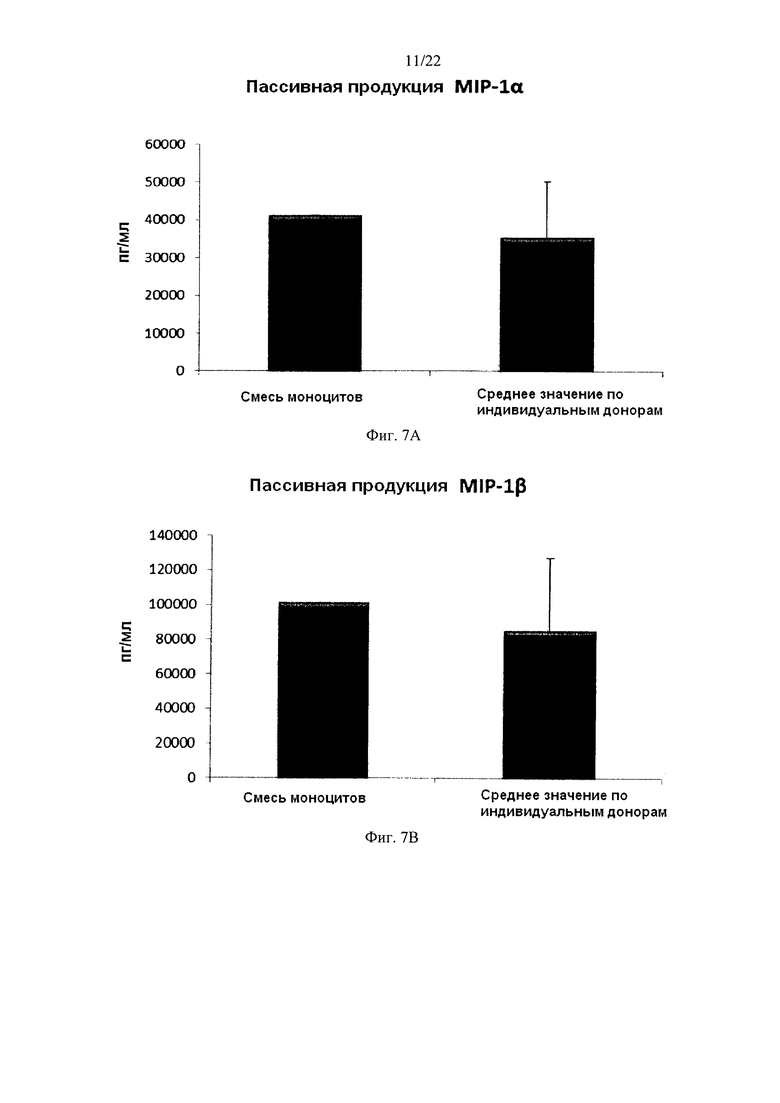
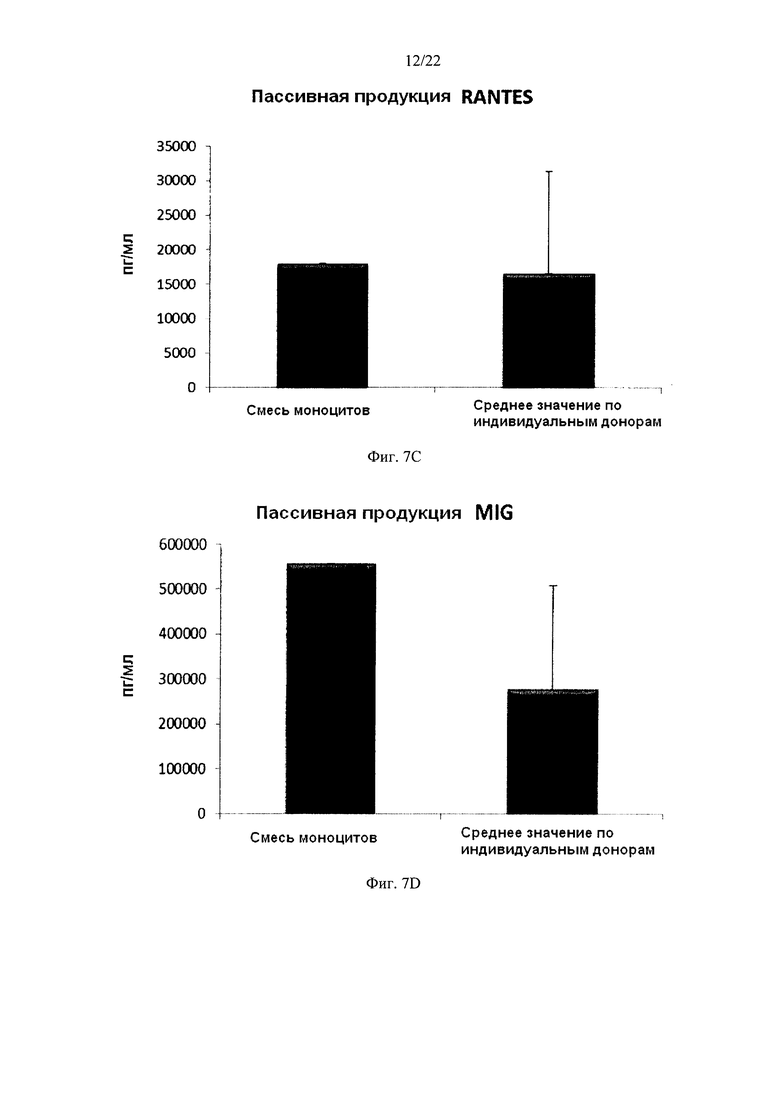
На Фиг. 7 показана продукция провоспалительных хемокинов провоспалительными ДК (полученными из индивидуальной культуры или смешанной культуры моноцитов периферических крови). Эти провоспалительные ДК были промыты после стимуляции в присутствии факторов активации в течение 18 часов, после чего их снова культивировали в течение 24 часов без добавления факторов активации ("пассивная продукция").
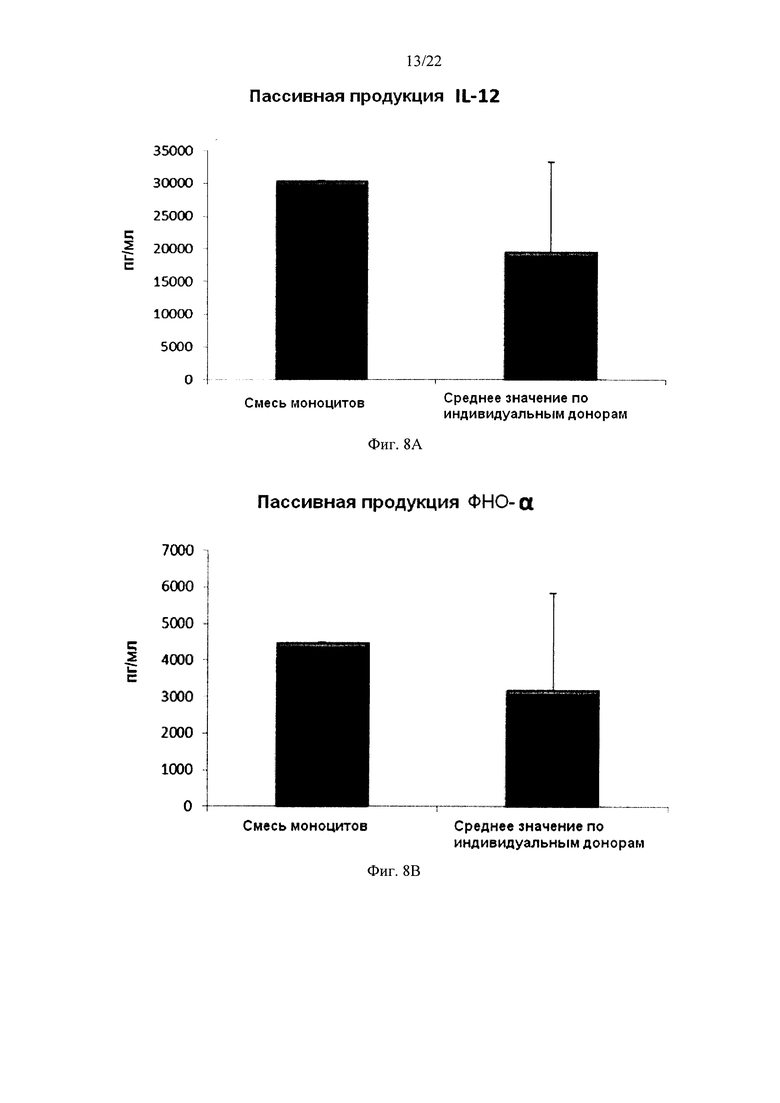
На Фиг. 8 показана продукция провоспалительных цитокинов провоспалительными ДК (полученными из индивидуальной культуры или смешанной культуры моноцитов периферических крови). Эти провоспалительные ДК были промыты после стимуляции в присутствии факторов активации в течение 18 часов, после чего их снова культивировали в течение 24 часов без добавления факторов активации ("пассивная продукция").
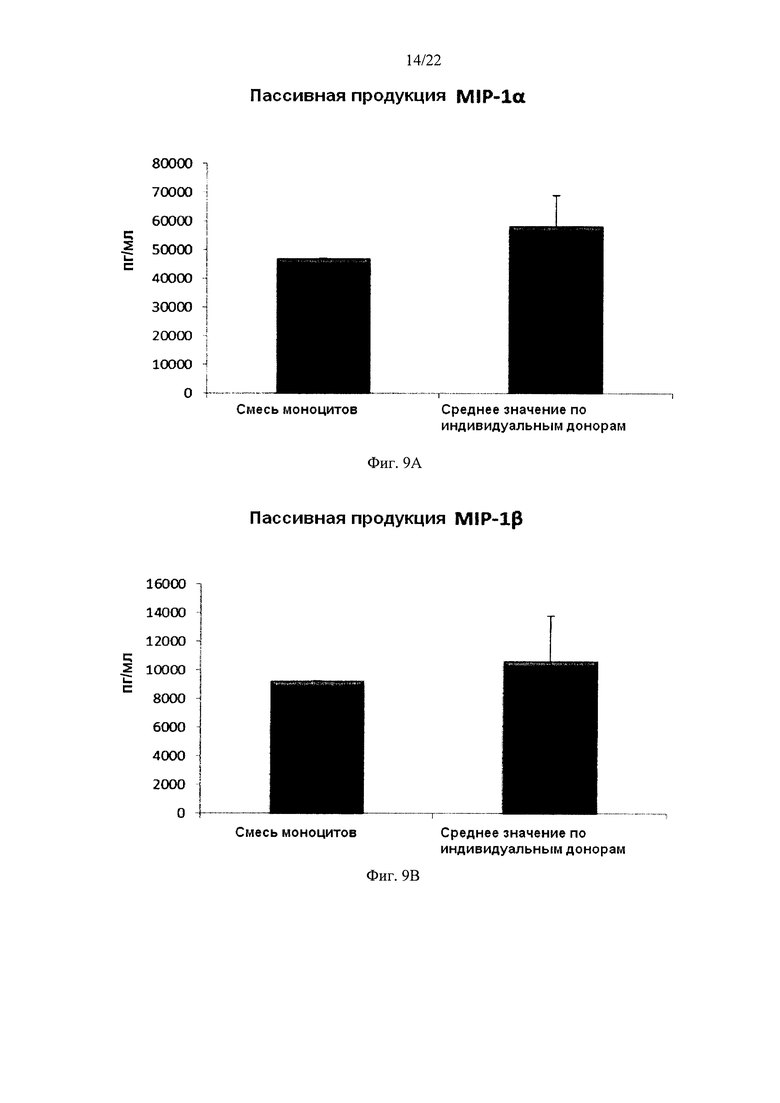
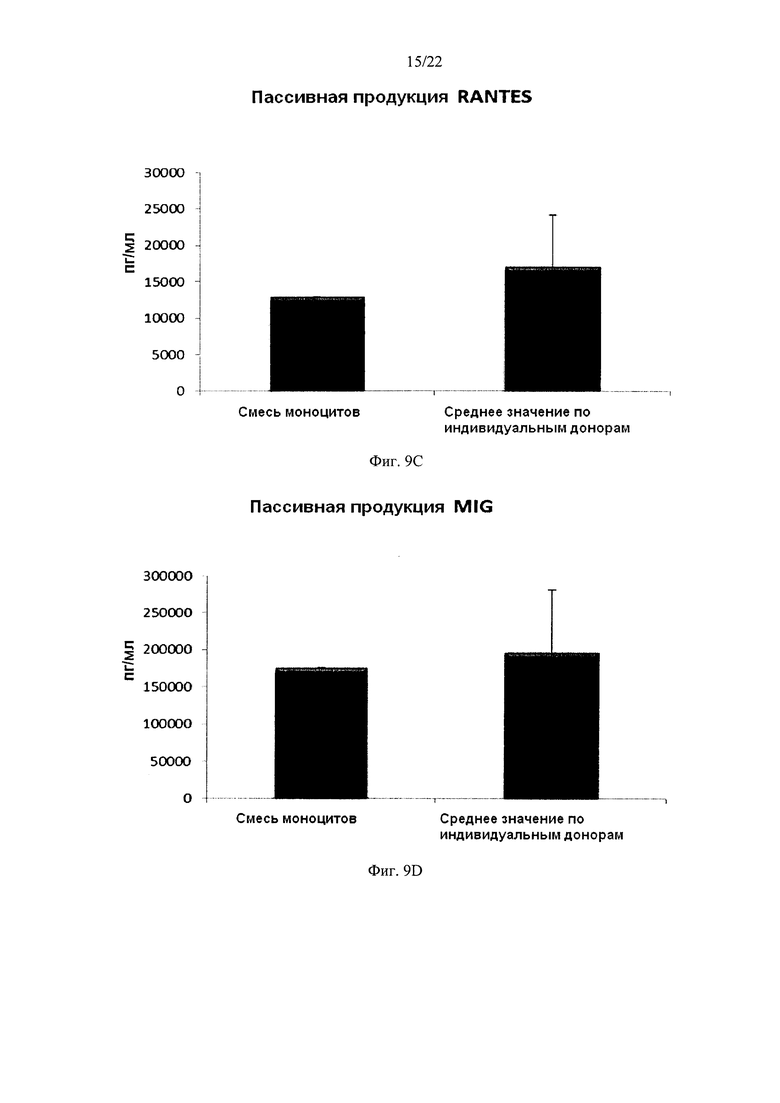
На Фиг. 9 показана продукция провоспалительных хемокинов провоспалительными ДК (полученными из индивидуальной культуры или смешанной культуры моноцитов лейкоцитарных пленок). Эти провоспалительные ДК были промыты после стимуляции в присутствии факторов активации в течение 18 часов, после чего их снова культивировали в течение 24 часов без добавления факторов активации ("пассивная продукция").
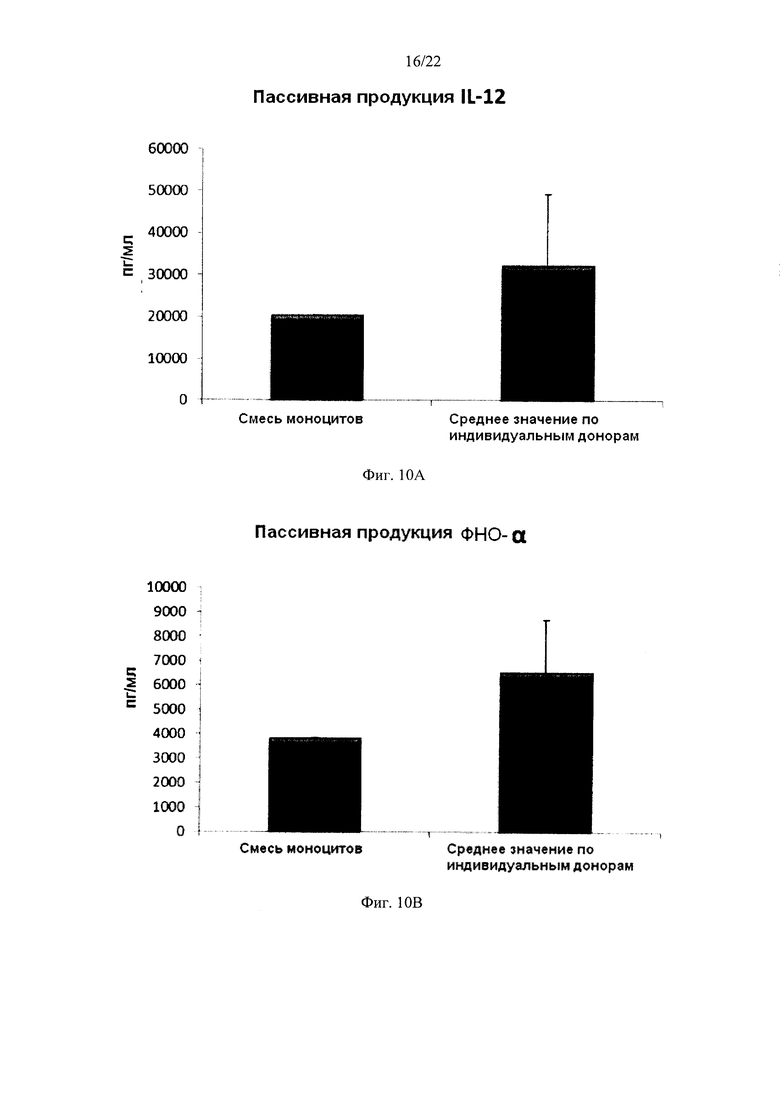
На Фиг. 10 показана продукция провоспалительных хемокинов провоспалительными ДК (полученными из индивидуальной культуры или смешанной культуры моноцитов лейкоцитарных пленок). Эти провоспалительные ДК были промыты после стимуляции в присутствии факторов активации в течение 18 часов, после чего их снова культивировали в течение 24 часов без добавления факторов активации ("пассивная продукция").
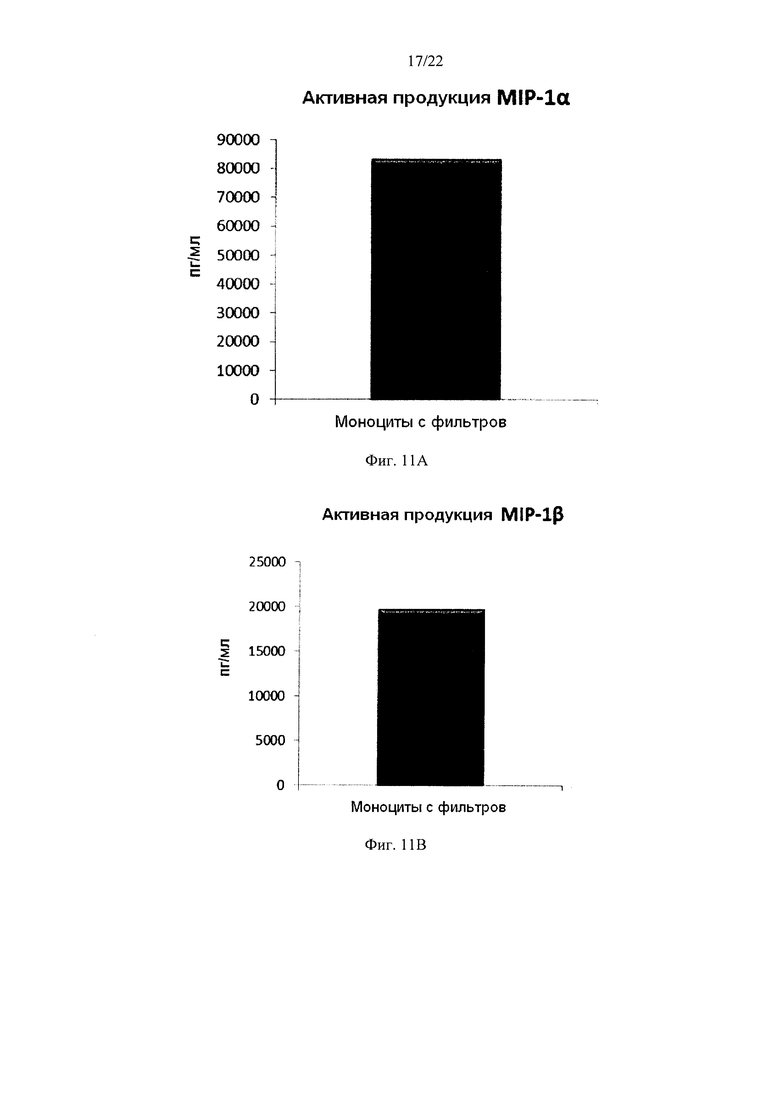
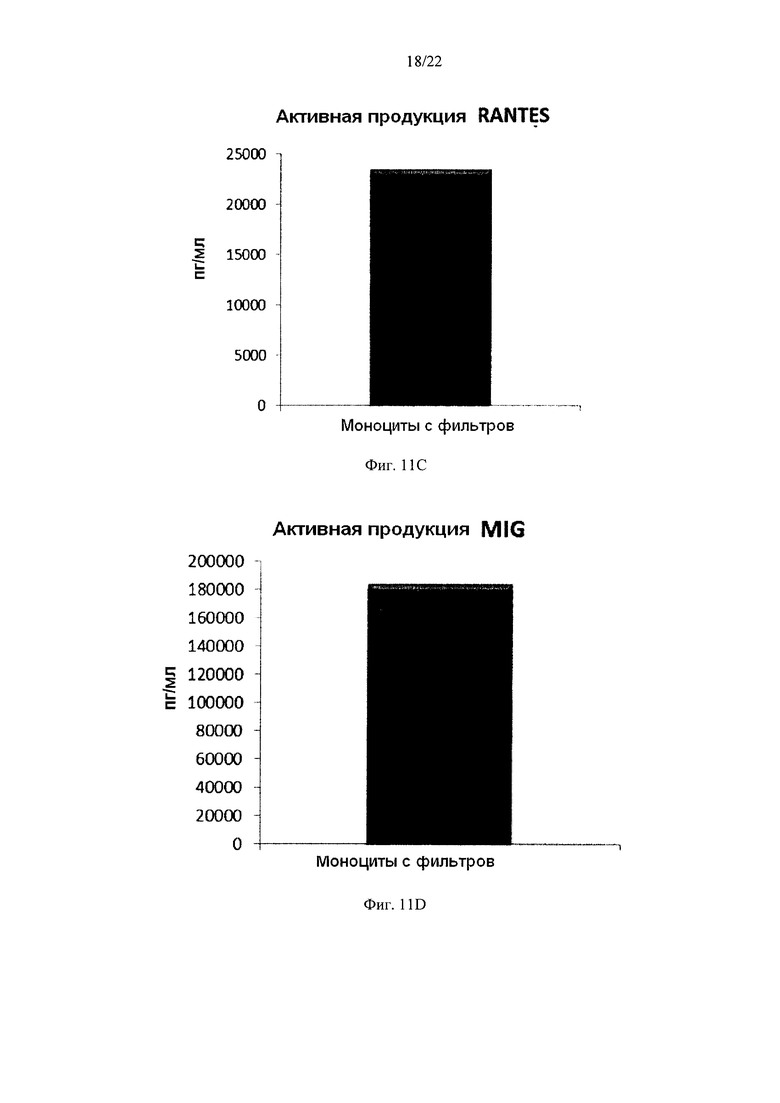
Фиг. 11 иллюстрирует, что смешанные незрелые ДК, полученные из моноцитов с фильтров, продуцируют значительные количества провоспалительных хемокинов в течение 18 часов непрерывной стимуляции факторами активации ("активное продуцирование")
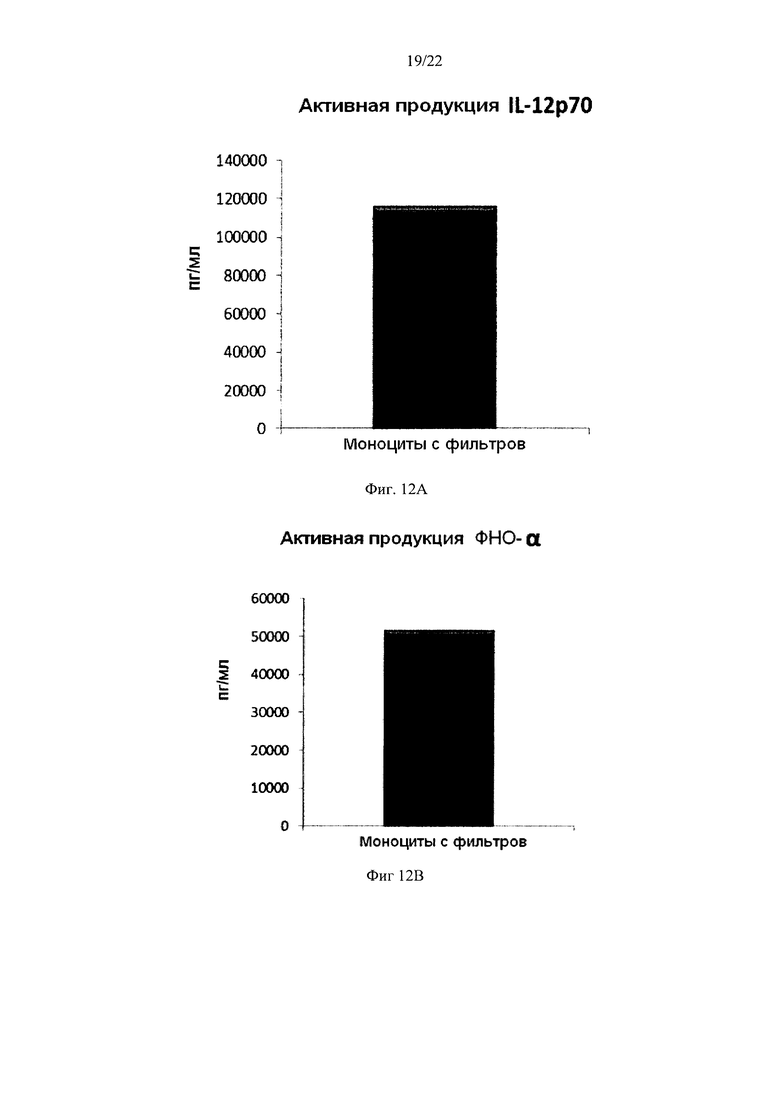
Фиг. 12 иллюстрирует, что смешанные незрелые ДК, полученные из моноцитов с фильтров, продуцируют значительные количества провоспалительных хемокинов в течение 18 часов непрерывной стимуляции в присутствии факторов активации ("активное продуцирование")
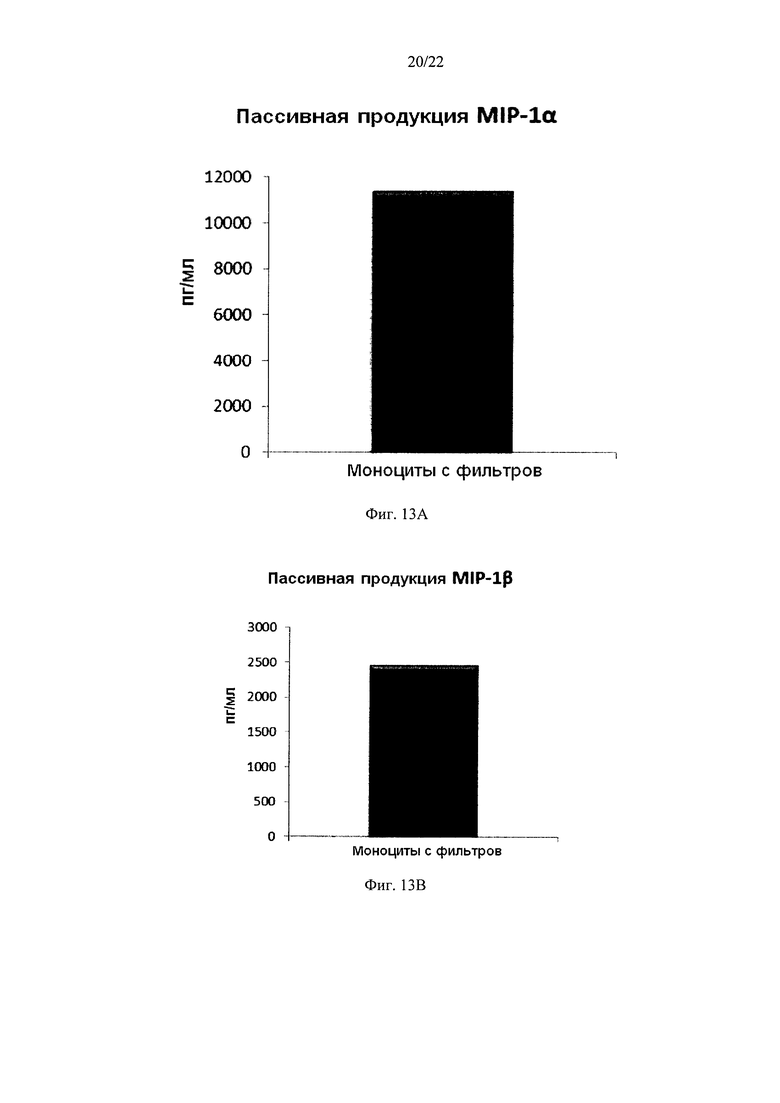
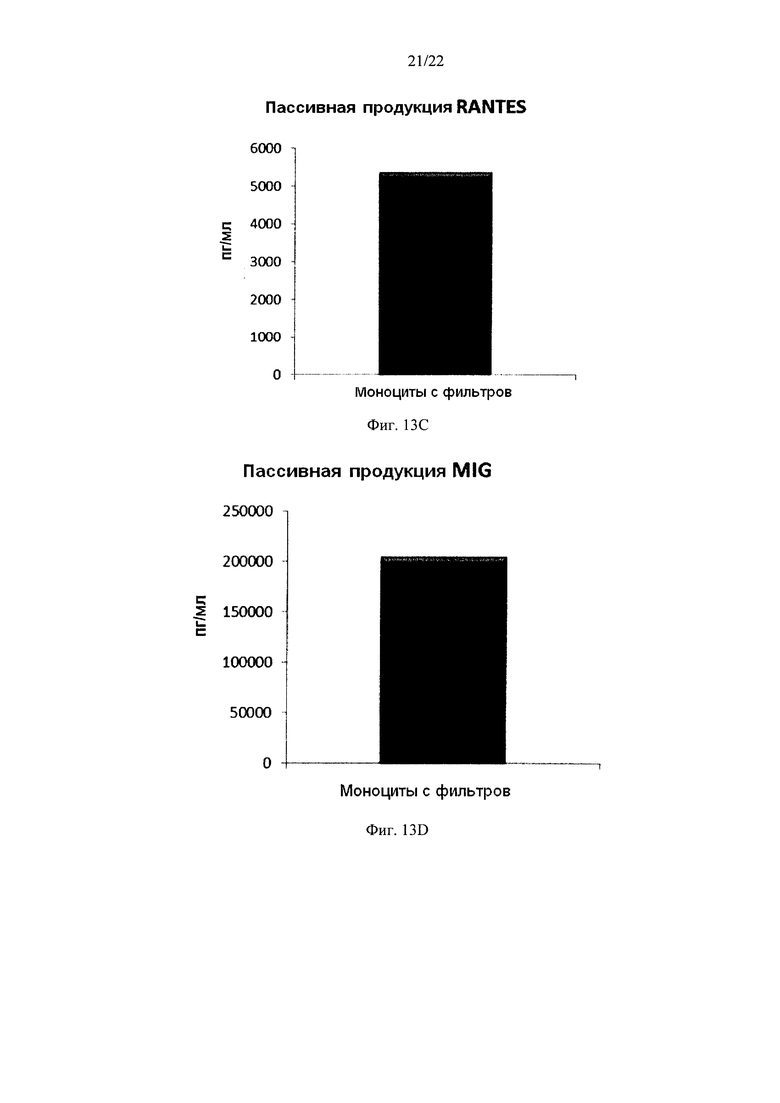
Фиг. 13 иллюстрирует, что смешанные провоспалительные ДК, моноцитов с фильтров, демонстрируют значительную продукцию провоспалительных хемокинов после извлечения активирующих факторов. Эти провоспалительные ДК были промыты после стимуляции в присутствии факторов активации в течение 18 часов, после чего их снова культивировали в течение 24 часов без добавления факторов активации ("пассивная продукция").
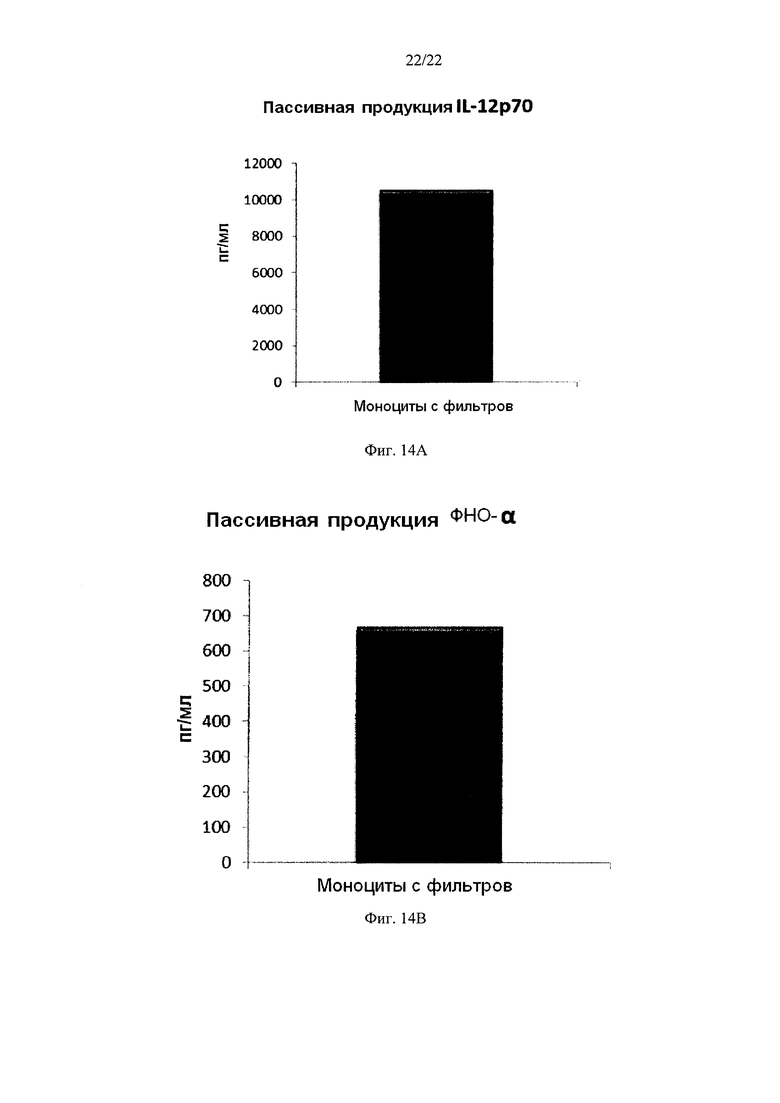
Фиг. 14 иллюстрирует, что смешанные провоспалительные ДК, моноцитов с фильтров, демонстрируют значительную продукцию провоспалительных цитокинов после удаления активирующих факторов. Эти провоспалительные ДК были промыты после стимуляции в присутствии факторов активации в течение 18 часов, после чего их снова культивировали в течение 24 часов без добавления факторов активации ("пассивное продуцирование").
Экспериментальная часть
Приведенные ниже примеры являются просто примерами, и из не следует истолковывать как ограничение объема изобретения. Напротив, изобретение ограничено только прилагаемой формулой изобретения.
Лейкоциты различного происхождения
Выделение лейкоцитов из лейкоцитарных фильтров (фильтры TACSI)
Лейкоцитарные фильтры (лейкоцитарные фильтры, TACSI используемые для стандартного выделения лейкоцитов, из 4 объединенных лейкоцитарных пленок из процесса производства тромбоцитов) были собраны в лаборатории компонентов на отделении трансфузионной медицины больницы Университета Sahlgrenska, Гетеборг, и доставлены в лабораторию (Кафедра клинической иммунологии, университетская больница Sahlgrenska) во льду.
В лаборатории шприц (Terumo) наполняли 50 миллилитрами буфера ФБР/ЭДТА (CliniMACS) и присоединяли к фильтру TACSI через фитинг с фиксатором Люэра. Фильтр трижды промывали обратным потоком в стерильную стеклянную колбу (150 мл буфера ФБР/ЭДТА в общей сложности). В конце элюированную клеточную суспензию разбавляли буфером ФБР (РАА, Fisher Scientific) в отношении 1:2 в пробирке Falcon (бренд Fisher, Fisher Scientific).
Лейкоцитарные пленки
Лейкоцитарные пленки здоровых доноров крови были собраны на кафедре трансфузионной медицины и доставлены в лабораторию при комнатной температуре.
Периферическая кровь
Периферическая кровь здоровых доноров была собрана на кафедре трансфузионной медицины и доставлена в лабораторию при комнатной температуре. В лаборатории кровь смешали с ФБР при комнатной температуре в отношении 1:2 в пробирке Falcon.
Выделение мононуклеарных клеток периферической крови (МКПК)
Периферическая кровь здоровых доноров была собрана на кафедре трансфузионной медицины и доставлена в лабораторию при комнатной температуре. В лаборатории кровь смешали с ФБР при комнатной температуре в отношении 1:2 в пробирке Falcon. Суспензию клеток осторожно перенесли в 10 мл центрифужные пробирки (Nunc), содержащие 3 мл Lymphoprep (Axis-Shield). В каждую пробирку перенесли по 5-6 мл, после чего их центрифугировали при 2000 оборотах в минуту непрерывно в течение 20 минут при комнатной температуре. Выделенные МКПК перенесли в предварительно охлажденные пробирки объемом 10 мл. Клетки дважды промывали путем заполнения трубки холодным ФБР с последующим центрифугированием при 1450 оборотах в минуту, 10 минут при 4°С. Супернатант выбросили, а осадок ресуспендировали в 1 мл холодного ФБР. Затем в каждую пробирку добавляли еще 9 мл.
Выделение моноцитов
Фракции элюированных лейкоцитов с фильтров / их лейкоцитарных пленное объемом по 5 мл или выделенных из 10-20 мл цельной периферической крови МКПК центрифугировали в пробирках при 1450 оборотах в минуту в течение 10 минут при 4°С. Супернатант полностью удаляли, а клеточный осадок ресуспендировали в ФБР/ЭДТА (Miltenyi), 80 мкл на каждые 107 клеток. Добавляли 20 мкл микрогранул CD14 (Miltenyi) на каждые 107 клеток. Клетки перемешивали и инкубировали в течение 15 минут при 4°С, а затем промывали добавлением 1-2 мл ФБР/ЭДТА, после чего центрифугировали при 300×g в течение 10 минут. Супернатант полностью удаляли, а оставшиеся клетки ресуспендировали в 500 мкл ФБР/ЭДТА.
Сепараторы MidiMACS (Miltenyi) помещали в магнитные стойки (Miltenyi) и промывали 3 мл ФБР/ЭДТА. Клеточные суспензии помещали в сепараторы MidiMACS, позволяя клеткам пройти сквозь сепараторы. Сепараторы MidiMACS промывали три раза с использованием 3 мл ФБР/ЭДТА. Выходящую фракцию с немечеными клетками отбрасывали. Сепараторы MidiMACS удалили из магнитной стойки и поместили на пробирку Falcon. Затем пипеткой внесли на колонку 5 мл ФБР/ЭДТАбуфера и сразу протолкнули протолкнули поршнем.
Концентрацию клеток была определяли в камере Бюркера. Суспензии клетоксодержащие моноциты центрифугировали при 1450 оборотах в минуту, 10 минут при 4°С. Супернатант выбрасывали, клетки ресуспендировали в среде для ДК Cellgro (CellGenix). Чистота CD14 + моноцитов во всех клеточных культурах была >80%, по результатам FACS-анализа, см. ниже.
Получение незрелых ДК
Лейкоциты, полученные из фильтров TACSI, ресуспендировали в среде для ДК Cellgro без примесей человеческой сыворотки в концентрации 300000 клеток/мл и высевали в 24-луночные планшеты (1 мл на лунку). Обогащенные моноцитами лейкоциты из лейкоцитарных пленок и периферической крови сначала ресуспендировали до концентрации 5×105 моноцитов/мл в среде Cellgro. 400 мкл среды Cellgro (без клеток) добавляли в 12 лункок (A1-6, B1-3, С1-3) в 24-луночном планшете. 600 мкл обогащенной моноцитами клеточной суспензии донора А (или лейкоцитарной пленки периферической крови, соответственно) переносили в лунки А1-3. 600 мкл обогащенной моноцитами клеточной суспензии донора В (или лейкоцитарной пленки периферической крови) переносили в лунки В1-3. 600 мкл обогащенной моноцитами клеточной суспензии донора С переносили в лунки С1-3 (или лейкоцитарной пленки периферической крови). В лунки А4-6 переносили 200 мкл обогащенной моноцитами клеточной суспензии из всех трех доноров (или лейкоцитарной пленки периферической крови). Окончательное количество клеток во всех лунках составляло 300000 клеток (в объеме 1 мл Cellgro среды на лунку).
Для того чтобы дифференцировать моноциты в незрелые ДК культуральную среду была дополняли 1000 ед/мл рекомбинантного человеческого IL-4 и 1000 ед/мл рекомбинантного человеческого ГМ-КСФ (все из CellGenix, Фрайбург, Германия), затем клетки культивировали в течение 5 дней.
Активация/созревание незрелых ДК
После 5 дней культивирования в среде Cellgro, дополненной IL-4 и ГМ-КСФ, индуцировали активацию/созревание незрелых ДК добавлением 20 мкг/мл поли I:С (Sigma, Штейнгейм, Германия), иммуностимулятора специфичного в отношении рецептора TLR-3, также известного как полиинозиновая : полицитидиловая кислота или натриевая соль полиинозиновой : полицитидиловой кислоты, 2,5 мкг/мл R848 (Sigma, Штейнгейм, Германия), лиганда toll-подобнго рецептора TLR 7/8, также известного как резиквимод, и 1000 ед/мл интерферона гамма (ИФН-γ, R&D Systems, Миннеаполис, США). После 18 часов инкубации клетки трижды промывали и затем инкубировали в свежей среде AIM-V (без добавления экзогенных факторов активации) в течение 24 часов. Супернатант культуры собирали в соответствии с протоколами, хорошо известными специалисту в данной области техники.
Для анализа уровней провоспалительных хемокинов и провоспалительных цитокинов проводили анализ супернатантаов методом ELISA как описано ниже.
Оценка уровней провоспалительных хемокинов и провоспалительных цитокинов методом ELISA
Провоспалительные хемокины CCL3/MIP-1α, CCL4/MIP-1β, CCL5/RANTES и CXCL9/MIG и провоспалительные цитокины IL-12p70 и ФНО-α измеряли методом твердофазного иммуноферментного анализа (ELISA) с использованием набора Duo Set ELISA Development System (R&D systems, Миннеаполис, США) в соответствии с инструкциями изготовителя.
Фенотипическое исследование методом проточной цитометрии
Моноциты и моноцитарные ДК были получены как описано выше. Количество CD14 + моноцитов после выделения оценивали окрашиванием клеток конъюгатом FITC-антититело к человеческому CD14. После 5 дней инкубации в среде Cellgro с добавлением IL-4 и ГМ-КСФ незрелые ДК промывали, и затем окрашивали конъюгатом РЕ-антитело к человеческому CD86 в комбинации с конъюгатом FITC-антититело к человеческому CD83. Незрелые ДК, которые впоследствии активировали в течение 18 часов в присутствии факторов активации, также окрашивали конъюгатом РЕ-антитело к человеческому CD86 в комбинации конъюгатом FITC- антититело к человеческому CD83. Мышинные IgG1 и IgG2, окрашенные FITCh РЕ использовали в качестве контроля изотипов (все из BD Biosciences, Калифорния, США). Образцы исследовали методом проточной цитометрии (FACS) с использованием программного обеспечения Cell Quest (BD Bioscience, Калифорния, США).
Результаты
Ниже приведены комментарии к результатам, полученным в экспериментальной части.
ДК, полученные из совместных культур обогащенных моноцитами аллогенных лейкоцитов, фенотипически не активировались/созревали при совместном культивировании в водной культуральной среде без примесей сыворотки нечеловеческого происхождения и с добавлением ГМ-КСФ и IL-4
Культивирование моноцитов от одного донора крови в культуре среде без примесей сыворотки нечеловеческого происхождения и с добавлением ГМ-КСФ и IL-4 в течение 4-7 дней привело к получению неистощенных ДК с типичным "незрелым" фенотипом, в том числе низким уровнем экспрессии маркера созревания CD83 и низкой экспрессией костимулирующей молекулы CD86. Как видно на Фиг. 1a и b, средняя экспрессия CD83 и CD86 у 3 различных "индивидуальных" ДК была близка к экспрессии CD83 (Фиг. 1а) и CD86 (Фиг. 1b) у ДК, полученных из смеси от всех трех доноров. Как видно на Фигурах lend, сильно повышенная средняя экспрессия маркеров активации/созревания CD83 и CD86 у "индивидуальных" ДК (исследовали ДК от 3 различных доноров периферической крови) после культивирования в водной культуральной среде, без примесей сыворотки нечеловеческого происхождения, с добавлением ГМ-КСФ и IL-4, в течение 4 дней и последующей непрерывной активации в присутствии стимулирующих факторов в течение 18 часов, была близка к экспрессии CD83 (фиг. 1с) и CD86 (Fig. 1d) экспрессии на активированных ДК, полученных из смеси аллогенных обогащенных моноцитами лейкоцитов от всех трех доноров.
В совокупности эти данные показывают, что из ДК, полученные из моноцитов из смешанной аллогенной популяции, являются незрелыми после культивирования в ГМ-КСФ и IL-4 в течение 5 дней и, следовательно, они не подверглись активации/созреванию во время дифференцировки из моноцитов в незрелые ДК. Кроме того, незрелые ДК из аллогенной смешанной популяции моноцитов по меньшей мере фенотипически не истощены, поскольку они выражено реагируют фенотипическим созреванием на стимуляцию факторами активации.
Данные, полученные методом проточной цитометрии. Соответствующая Y-ось показывает среднюю интенсивность флуоресценции (MFI) для CD83 и CD86 до и после непрерывной стимуляции в присутствии факторов активации в течение 18 часов. По оси X показаны различные измеренные комбинации.
Незрелые ДК, полученные из совместных культур смешанных аллогенных моноцитов периферической крови, функционально не истощены.
Культивирование моноцитов (от одного донора) в культуральной среде, содержащей ГМ-КСФ и IL-4, в течение 4-7 дней, как известно, в результате дает неистощенные ДК, которые на стимуляцию определенными факторами активации, отвечают интенсивным продуцированием провоспалительных хемокинов (MIP-1 альфа, MIP-1 бета, RANTES и MIG) и провоспалительных цитокинов (IL-12р70 и ФНО-альфа) Как показано на фиг. 2, высокие средние уровни MIP-1 альфа (Фиг. 2а), MIP-1 бета (фиг. 2b), RANTES (фиг. 2с), MIG (фиг. 2d), полученные от "индивидуальных" ДК (исследовали ДК от трех различных доноров периферической крови) при непрерывной активации в присутствии стимулирующих факторов в течение 18 часов, были близки к уровням ДК, полученных из смеси моноцитов от всех трех доноров. Следует отметить, что имеют место значительные различия в индуцированной активацией продукции хемокинов между различными отдельными донорами ДК. Как видно на Фиг. 4, высокие средние уровни продукции IL-12р70 (фиг. 3а) и ФНО-альфа (фиг. 3b) активированными "индивидуальными" ДК были близки к уровням ДК, полученным из смеси моноцитов от всех трех доноров. Следует отметить, что имеют место значительные различия в продукции IL-12p70 и ФНО-альфа между ДК от различных индивидуальных доноров.
Данные были получены методом твердофазного иммуноферментного анализа (ELISA). Результаты представляют собой средние значения ± SD (стандартное отклонение) по трем индивидуумам, и значения, полученные для смеси от всех трех доноров. Ось Y показывает количество соответствующего продуцируемого вещества в пг/мл/1×106 клеток, в течение 18 часов непрерывной стимуляции/активации. По оси X показаны различные измеренные комбинации.
Предположили, что простагландин Е2 (PGE2) играет центральную роль в вызванном активацией истощении незрелых ДК (Rieser С et al., Differential Deactivation of Human Dendritic Cells by Endotoxin Desensitization: Role of Tumor Necrosis Factor-α and Prostaglandin E2. Blood 91 (1998) 3112-3117). Поэтому мы исследовали, будет ли добавление ингибитора ЦОГ-2 NS-398 (с целью препятствовать потенциальному продуцированию PGE2) во время совместного культивирования аллогенных моноцитов увеличивать продуцирование провоспалительных хемокинов (MIG) или провоспалительных цитокинов (IL12p70) при последующей активации. Как видно на фиг. 4, присутствие ингибитора ЦОГ-2 NS-398, в ходе превращения моноцитов в ДК, не увеличивало, а снижало индуцируемую активацией продукцию MIG и IL-12p70. Таким образом, нет никаких признаков опосредуемого PGE2 истощения дифференцированных незрелых ДК из совместных культур смешанных аллогенных моноцитов.
Данные были получены методом твердофазного иммуноферментного анализа (ELISA). Показаны результаты одного эксперимента, где была использована смесь от всех трех доноров. Ось Y показывает количество соответствующего продуцируемого вещества в пг/мл / 1×106 клеток, в течение 18 часов непрерывной стимуляции/активации. Ось X показывает различные измеренные комбинации.
Незрелые ДК, полученные из совместных культур смешанных обогащенных моноцитами аллогенных лейкоцитарных пленок функционально не истощены.
Как видно на Фиг. 5, высокие средние уровни индуцированных активацией провоспалительных хемокинов MIP-1 альфа (фиг. 5а), MIP-1 бета (фиг. 5b), RANTES (фиг. 5, с), МIG (фиг. 5d), продуцируемые «индивидуальными» ДК (ДК от трех разных доноров лейкоцитарных пленок) во время непрерыной активации в присутствии факторов активации в течение 18 часов, были близки к уровням для ДК, полученных из смеси моноцитов от всех трех доноров. При этом имеют место значительные различия в продукции хемокинов между различными ДК от индивидуальных доноров.
Как видно на фиг. 6, высокие индуцированные активацией средние уровни IL-12р70 (фиг. 6а) и ФНО-альфа (фиг. 6b), продуцируемые «индивидуальными» ДК, были близки к уровням для ДК, полученных из обогащенной моноцитами лейкоцитной смеси от всех трех доноров. При этом имеют место значительные различия в продукции IL-12p70 и TNF-альфа между различными ДК от индивидуальных доноров.
Данные были получены методом твердофазного иммуноферментного анализа (ELISA). Результаты представляют собой средние значения ± SD по трем донорам, и значения, полученные для смеси от всех трех доноров. Ось Y показывает количество соответствующего продуцируемого вещества в пг/мл / 1×106 клеток, в течение 18 часов непрерывной стимуляции/активации. По оси X показаны различные измеренные комбинации.
Провоспалительные ДК, полученные из совместных культур смешанных аллогенных моноцитов периферической крови, демонстрируют устойчивое продуцирование провоспалительных хемокинов и цитокинов
Для того чтобы ввести активированные провоспалительные ДК в организм пациента, их перед введением обычно следует промыть. В противном случае может произойти нежелательная побочная реакция, вызванная одновременным применением стимулирующих агентов (направленных на индукцию провоспалительных ДК ex vivo). Поэтому незрелые ДК должны быть активированы в провоспалительные ДК, стабильно продуцирующие необходимые факторы и после удаления индуцирующих активацию факторов. Как видно на фиг. 7, средние уровни продукции MIP-1 альфа (фиг. 7а), MIP-1 бета (фиг. 7, b), RANTES (фиг. 7с), MIG (фиг. d) "индивидуальными" провоспалительными ДК после удаления факторов активации (исследовали провоспалительные ДК из моноцитов периферической крови трех различных доноров) были близки к провоспалительным ДК, полученным из смеси моноцитов всех трех доноров периферической крови. При этом имеют место значительные различия в продукции хемокинов между провоспалительными ДК от разных доноров после удаления факторов активации. Средние значения продукции IL-12p70 (фиг. 8a) и ФНО-альфа (фиг. 8b), провоспалительными ДК после удаления факторов активации также были близки к уровням у промытых провоспалительных ДК, полученных из смеси моноцитов от всех трех доноров. При этом имеют место значительные различия в продукции цитокинов между провоспалительными ДК от разных индивидуальных доноров после удаления факторов активации.
Данные были получены методом твердофазного иммуноферментного анализа (ELISA). Результаты представляют собой средние значения ± SD по трем донорам, и значения, полученные для смеси от всех трех доноров. Ось Y показывает количество соответствующего продуцируемого вещества в пг/мл / 1×106 клеток, в течение 24 часов после удаления факторов активации. По оси X показаны различные измеренные комбинации.
Провоспалительные ДК, полученные из совместных культур смешанных аллогенных, обогащенных моноцитами, периферических лейкоцитарных пленок, демонстрируют устойчивую высокую продукцию провоспалительных хемокинов и цитокинов
Как видно на фиг. 9, средние уровни MIP-1 альфа (фиг. 9а), MIP-1 бета (фиг. 9b), RANTES (фиг. 9 с), MIG (фиг 9d), продуцируемые "индивидуальными" провоспалительными ДК после удаления факторов активации (исследовали провоспалительные ДК из моноцитов лейкоцитарной пленки от трех различных доноров) были близки к уровням у провоспалительных ДК, полученных из смеси моноцитов от всех трех доноров лейкоцитарной пленки. При этом имеют место значительные различия в продукции хемокинов между провоспалительными ДК от различных доноров. Среднее значение продукции IL-12p70 (фиг. 10а) и ФНО-альфа (фиг. 10b) "индивидуальными" провоспалительными ДК после удаления факторов активации также близко к уровням, обеспечиваемы промытыми провоспалительными ДК, полученными из смеси моноцитов от всех трех доноров лейкоцитарной пленки. При этом имеют место значительные различия в продукции цитокинов между провоспалительными ДК от различных доноров.
Данные были получены методом твердофазного иммуноферментного анализа (ELISA). Результаты представляют собой средние значения ± SD по трем донорам, и значения, полученные для смеси от всех трех доноров. Ось Y показывает количество соответствующего продуцируемого вещества в пг/мл / 1×106 клеток, в течение 24 часов после отмены факторов активации. По оси X показаны различные измеренные комбинации.
Смешанные незрелые ДК, полученные из обогащенных моноцитами лейкоцитов с фильтров, при активации продуцируют количества объемы провоспалительных хемокинов и цитокинов.
Как видно на Фиг. 11, активированные смешанные ДК, полученные из обогащенных моноцитами лейкоцитов с фильтров (исходную популяцию лейкоцитов элюировали из с лейкоцитарного фильтра, использованного для очистки 4 лейкоцитарных пленок), продуцировали значительное количество MIP-1 альфа (Фиг. 11a), MIP-1 бета (Фиг. 11b), RANTES (Фиг. 11с), МИГ (фиг. 11d). Как видно на Фиг. 12, также продуцировалось значительное количество IL-12р70 (Фиг. 12а) и ФНО-альфа (Фиг. 12b).
Данные были получены методом твердофазного иммуноферментного анализа (ELISA). Результаты представляют собой средние значения ±SD по трем донорам, и значения, полученные для смеси от всех трех доноров. Ось Y показывает количество соответствующего продуцируемого вещества в пг/мл / 1×106 клеток, в течение 18 часов непрерывной стимуляции/активации.
Смешанные провоспалительные ДК, полученные из обогащенных моноцитами лейкоцитов, после удаления активирующих факторов демонстрируют значительную продукцию провоспалительных хемокинов и цитокинов.
Как видно на фиг. 13, активированные смешанные ДК, полученные из моноцитов с фильтров (исходную популяцию лейкоцитов элюировали из с лейкоцитарного фильтра, использованного для очистки 4 лейкоцитарных пленок), продуцировали значительное количество MIP-1 альфа (Фиг. 13а), MIP-1 бета (Фиг. 13с), RANTES (Фиг. 13с), MIG (Фиг. 13d) после удаления активирующих факторов. Как видно на Аиг.14, также продуцировалось значительное количество количество IL-12p70 (Фиг. 14а) и ФНО-альфа (Фиг. 14и).
Данные были получены методом твердофазного иммуноферментного анализа (ELISA). Результаты представляют собой средние значения ±SD по трем донорам, и значения, полученные для смеси от всех трех доноров. Ось Y показывает количество соответствующего продуцируемого вещества в пг/мл / 1×106 клеток, в течение 24 часов после удаления факторов активации.
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕННАЯ КОМПОЗИЦИЯ ДЛЯ ИНГИБИРОВАНИЯ ПРОЛИФЕРАЦИИ ОПУХОЛЕВЫХ КЛЕТОК | 2011 |
|
RU2565542C2 |
| СПОСОБ ГЕНЕРАЦИИ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК С ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ | 2011 |
|
RU2458985C1 |
| ГЕНЕРИРОВАНИЕ ДЕНДРИТНЫХ КЛЕТОК ИЗ МОНОЦИТАРНЫХ ПРЕДШЕСТВЕННИКОВ ДЕНДРИТНЫХ КЛЕТОК С ПОМОЩЬЮ GM-CSF В ОТСУТСТВИЕ ДОПОЛНИТЕЛЬНЫХ ЦИТОКИНОВ | 2004 |
|
RU2364625C2 |
| СПОСОБ НАГРУЗКИ ДЕНДРИТНЫХ КЛЕТОК АНТИГЕНОМ ИНФЕКЦИОННОГО ПРОИСХОЖДЕНИЯ Opisthorchis felineus | 2011 |
|
RU2486238C1 |
| СПОСОБ ПОЛУЧЕНИЯ КЛЕТОЧНОГО ПРЕПАРАТА НА ОСНОВЕ АКТИВИРОВАННЫХ ЛИМФОЦИТОВ ИЗ ЛЕЙКОЦИТАРНЫХ ФИЛЬТРОВ | 2024 |
|
RU2835686C2 |
| СПОСОБ ГЕНЕРАЦИИ АНТИГЕН-СПЕЦИФИЧЕСКИХ ПРОТИВОТУБЕРКУЛЕЗНЫХ КЛЕТОК | 2008 |
|
RU2378373C2 |
| СПОСОБ ИММУНОТЕРАПИИ ХРОНИЧЕСКОГО ВИРУСНОГО ГЕПАТИТА С | 2016 |
|
RU2637631C2 |
| СПОСОБ ГЕНЕРАЦИИ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК С АКТИВНОСТЬЮ ПРОТИВ КЛЕТОК, ИНФИЦИРОВАННЫХ ВИРУСОМ ГЕПАТИТА В | 2008 |
|
RU2366707C1 |
| АУТОЛОГИЧНАЯ ВАКЦИНА ДЛЯ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2008 |
|
RU2392946C2 |
| СПОСОБ ГЕНЕРАЦИИ СПЕЦИФИЧЕСКОГО ИММУННОГО ОТВЕТА ПРОТИВ АНТИГЕНОВ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА | 2011 |
|
RU2465324C1 |
Изобретение относится к области биотехнологии, конкретно к способу получения неистощенных незрелых дендритных клеток (ДК), происходящих из двух различных аллогенных доноров, с их последующей активацией и получением провоспалительных ДК. Изобретение позволяет получать провоспалительные ДК для терапии рака. 5 н. и 14 з.п. ф-лы, 42 ил., 1 пр.
1. Способ получения неистощенных незрелых дендритных клеток (ДК), включающий следующие этапы:
- получение смеси аллогенных лейкоцитов, где указанные аллогенные лейкоциты получены от по меньшей мере двух различных аллогенных доноров;
- выделение аллогенных моноцитов из указанной смеси аллогенных лейкоцитов с получением обогащенных моноцитами аллогенных лейкоцитов; и
- получение неистощенных незрелых ДК из указанных обогащенных моноцитами аллогенных лейкоцитов, причем получение неистощенных незрелых дендритных клеток (ДК) осуществляют путем совместного культивирования указанных обогащенных моноцитами аллогенных лейкоцитов в течение от 2 до 7 дней в водной клеточной культуральной среде без примесей сыворотки нечеловеческого происхождения с добавлением интерлейкина-4 (IL-4) и гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ).
2. Способ по п. 1, отличающийся тем, что указанная клеточная культуральная среда содержит по меньшей мере один человеческий полипептид.
3. Способ по п. 2, отличающийся тем, что указанный человеческий полипептид выбран из группы, состоящей из трансферрина, альбумина и инсулина.
4. Способ по п. 1, отличающийся тем, что указанные обогащенные моноцитами аллогенные лейкоциты включают аллогенные нейтрофилы.
5. Способ по п. 1, отличающийся тем, что указанную смесь аллогенных лейкоцитов получают объединением по меньшей мере двух лейкоцитарных пленок, содержащих лейкоциты, причем указанные объединяемые лейкоцитарные пленки получены от по меньшей мере двух различных аллогенных доноров.
6. Способ по п. 5, отличающийся тем, что указанные объединяемые лейкоцитарные пленки содержат тромбоциты или характеризуются сниженным количеством тромбоцитов.
7. Способ по п. 1, отличающийся тем, что указанную смесь аллогенных лейкоцитов получают путем:
- элюирования лейкоцитов из по меньшей мере двух лейкоцитарных фильтров, соответственно, которые ранее были использованы для выделения лейкоцитов из цельной крови, причем указанная цельная кровь была получена от по меньшей мере двух различных, аллогенных доноров; и
- объединения полученных лейкоцитов с получением указанной смеси аллогенных лейкоцитов;
или путем
- элюирования лейкоцитов из лейкоцитарного фильтра, который был использован для выделения лейкоцитов из объединенных лейкоцитарных пленок, которые были получены от двух разных аллогенных доноров.
8. Способ по п. 1, отличающийся тем, что указанные аллогенные моноциты выделены путем элютриации или выделения с использованием антител/гранул.
9. Способ по п. 1, отличающийся тем, что совместное культивирование проводят в течение приблизительно 5 дней.
10. Способ по п. 1, дополнительно включающий стадию нагрузки неистощенных незрелых ДК антигеном.
11. Способ получения провоспалительных ДК, включающий этапы:
- получения неистощенных незрелых ДК по любому из пп. 1-10;
- активации неистощенных незрелых ДК путем добавления фактора активации с получением провоспалительных ДК.
12. Способ по п. 11, отличающийся тем, что указанную активацию осуществляют путем добавления лиганда Toll-подобного рецептора 3 (TLR3) поли(I):поли(С), лиганда TLR7 / 8 и гамма-интерферона (ИФН-γ), что приводит к индукции активации.
13. Способ по п. 12, отличающийся тем, что указанный лиганд TLR7 / 8 выбран из резиквимода, гардиквимода и имиквимода.
14. Способ по п. 12, отличающийся тем, что активация дополнительно включает добавление по меньшей мере одного вещества, выбранного из группы, состоящей из лигандов TLR2, лигандов TLR4, лигандов TLR9, интерферона-альфа (ИФН-α), интерлейкина 1β (IL-1β) и фактора некроза опухоли альфа (ФНО-α), что приводит к индукции активации.
15. Способ по п. 12, в котором указанная активация не включает добавление простагландина Е2 (PGE2).
16. Способ по п. 12, отличающийся тем, что указанные незрелые ДК подвергаются воздействию факторов активации в течение от 8 до 24 часов, после чего факторы активации по существу полностью вымывают.
17. Применение смеси аллогенных провоспалительных дендритных клеток, полученных по меньшей мере от двух различных аллогенных доноров для лечения рака, причем указанные дендритные клетки получают способом по любому из пп. 11-16.
18. Фармацевтическая композиция для лечения рака, содержащая смесь аллогенных провоспалительных дендритных клеток, полученных способом по любому из пп. 11-16, в дозе, достаточной для активации собственных ДК пациента с получением нагруженных опухолевыми антигенами мигрирующих ДК, и по меньшей мере один фармацевтически приемлемый носитель.
19. Способ лечения рака, включающий введение смеси аллогенных провоспалительных дендритных клеток, полученных способом по любому из пп. 11-16, или композиции по п. 18 пациенту, нуждающемуся в таком лечении.
| WO 2011098516, 18.08.2011 | |||
| RU 2012137173, 10.02.2011 | |||
| LAURIN D | |||
| et al., Allogenetic reaction induces dendritic cell maturation through proinflammatory cytokine secretion, Transplantation, 2004, V | |||
| Спускная труба при плотине | 0 |
|
SU77A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Тепловой измеритель силы тока | 1921 |
|
SU267A1 |
| JONULEIT H | |||
| et al., Pro ‐ inflammatory cytokines and prostaglandins induce maturation of potent immunostimulatory dendritic cells under fetal calf serum ‐ free conditions, European journal of immunology, 1997, V | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Способ приготовления каучукового клея | 1916 |
|
SU3135A1 |
| WO 2003010292, 06.02.2003. | |||

