Изобретение относится к молекулам антител, которые обладают способностью специфически распознавать две области пептида β-А4, где первая область содержит аминокислотную последовательность AEFRHDSGY, представленную в SEQ ID NO:1, или ее фрагмент, и где вторая область содержит аминокислотную последовательность VHHQKLVFFAEDVG, представленную в SEQ ID NO:2, или ее фрагмент. Кроме того, в изобретении описаны молекулы нуклеиновых кислот, кодирующие молекулы антител, и векторы и хозяева, которые содержат эти молекулы нуклеиновых кислот. Кроме того, настоящее изобретение относится к композициям, предпочтительно фармацевтическим или диагностическим композициям, которые содержат соединения, предлагаемые в изобретении, а также к конкретному применению молекул антител, молекул нуклеиновых кислот, векторов или хозяев, предлагаемых в изобретении.
В описании процитирован ряд документов. Каждый из процитированных в описании документов (включая любые спецификации, инструкции и т.д. производителей) включен в настоящее описание в качестве ссылки.
Примерно в 70% всех случаев деменция является следствием болезни Альцгеймера, которая связана с избирательным поражением областей головного мозга и нервных цепей, имеющих решающее значение для умственной способности. Болезнь Альцгеймера характеризуется наличием нейрофибриллярных сплетений в пирамидальных нейронах гиппокампа и многочисленных амилоидных бляшек, состоящих в основном из плотного ядра отложений амилоида и неслитых оболочек.
Внеклеточные невритные бляшки содержат большие количества в основном фибриллярного пептида, обозначаемого как «амилоид β», «А-бета», «Aβ4», «β-А4» или «Аβ» (см. у Selkoe, Ann. Rev. Cell Biol. 10, 1004, сс.373-403, Коо, PNAS т. 96, 1999, сс.9989-9990, US 4666829 или Glenner, BBRC 12, 1984, сс.1131). Этот амилоид β образуется из «протеина-предшественника Альцгеймера /протеина-предшественника β-амилоида» (АРР). АРР - общее название связанных с мембраной гликопротеинов (см. у Sisodia, PNAS, т. 89, 1992, сс.6075), и они чувствительны к эндопротеолитическому расщеплению в Аβ-последовательности протеазой плазматической мембраны, α-секретазой (см. Sisodia (1992), loc. cit.). Кроме того, другие виды секретазной активности, в частности, β-секретазная и γ-секретазная активность, приводят к внеклеточному высвобождению β-амилоида (Аβ), который содержит либо 39 аминокислот (Аβ39), либо 40 аминокислот (Аβ40), либо 42 аминокислоты (Аβ42), либо 43 аминокислоты (Aβ43); см. у Sinha, PNAS 96, 1999, cc. 11094-1053; Price, Science 282, 1998. сс.1078-1083; WO 00/72880 или Hardy, TINS 20, 1997, с. 154.
Установлено, что Аβ существует в виде нескольких встречающихся в естественном состоянии форм, при этом встречающиеся в организме человека формы обозначают, как указано выше, т.е. Аβ39, Aβ40, Aβ41, Aβ42 и Аβ43. Наиболее известная форма Аβ42 имеет следующую аминокислотную последовательность (начиная с N-конца):
DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVVIA (SEQ ID NO:27).
В Аβ41, Аβ40, Аβ39 отсутствуют С-концевые аминокислоты A, IA и VIA соответственно. В Аβ43-форме на С конце присутствует остаток треонина, дополнительный по сравнению с С-концом описанной выше последовательности (SEQ ID NO:27).
Установлено, что время, необходимое для образования ядра Аβ40-фибрилл, существенно превышает время, необходимое для образования ядра Aβ42-фибрилл; см. у Коо, loc. cit. и Harper, Ann. Rev. Biochem. 66, 1997, cc. 385-407. В обзоре Wagner, J. Clin. Invest. 104, 1999, cc.1239-1332 указано, что Аβ42-форма чаще других связана с невритными бляшками и считается наиболее фибриллогенной in vitro. Существует также мнение, что Аβ42 служит в качестве «затравки» для связанной с нуклеацией полимеризацией упорядоченных некристаллических Аβ-пептидов; Jarrett, Cell 93, 1993, cc.1055-1058.
Следует подчеркнуть, что процессинг модифицированного АРР и/или образование внеклеточных бляшек, содержащих напоминающие белок отложения, известно не только в патологии, связанной с болезнью Альцгеймера, но обнаружено также у пациентов, которые страдают другими неврологическими и/или нейродегенеративными нарушениями. Эти нарушения включают среди прочего синдром Дауна, наследственное внутримозговое кровоизлияние с амилоидозом Дутча-типа, болезнь Перкинсона, ALS (амиотрофический боковой склероз), болезнь Крейтцфельдта-Якоба, связанную с ВИЧ деменцию и моторную невропатию.
Для предупреждения, лечения и/или облегчения нарушений и/или заболеваний, связанных с патологическим отложением амилоидных бляшек, разработаны средства и методы, которые или влияют на образование β-амилоидной бляшки, обладают способностью предупреждать агрегацию Аβ и/или участвуют в деполяризации уже сформировавшихся амилоидных отложений агрегатов амилоида-β.
Таким образом, с учетом серьезных поражений, связанных с изменением и/или патологическими биологическими особенностями амилоида, существует необходимость в создании средств и методов лечения связанных с амилоидом нарушений. В частности, требуются лекарственные средства, оказывающие воздействие на патологическую агрегацию амилоида или обладающие способностью участвовать в деполимеризации агрегированного Аβ. Кроме того, существует необходимость в диагностических методах, предназначенных для выявления среди прочего амилоидных бляшек.
Таким образом, техническая задача, положенная в основу настоящего изобретения, связана с решением указанных выше проблем.
Таким образом, настоящее изобретение относится к молекуле антитела, которая обладает способностью специфически распознавать две области пептида β-А4/Аβ4, где первая область содержит аминокислотную последовательность AEFRHDSGY (SEQ ID NO:1) или ее фрагмент, а вторая область содержит аминокислотную последовательность VHHQKLVFFAEDVG (SEQ ID NO:2) или ее фрагмент.
В контексте настоящего описания понятие «молекула антитела» относится к полным молекулам иммуноглобулинов, предпочтительно IgM, IgD, IgE, IgA или IgG, более предпочтительно IgG1, IgG2, IgG2b, IgG3 или IgG4, а также к фрагментам таких молекул иммуноглобулина, таким как Fab-фрагменты или VL-, VH- или CDR-участки. Кроме того, понятие относится к модифицированным и/или измененным молекулам антител, типа химерных или гуманизированных антител. Понятие относится также к модифицированным или измененным моноклональным или поликлональным антителам, а также к антителам, полученным с помощью рекомбинации или с помощью методов синтеза/синтезированным антителам. Понятие включает также интактные антитела и фрагменты/участки антител, типа разделенных легкой и тяжелой цепей. Fab, Fab/c, Fv, Fab', F(ab')2. Понятие «молекула антитела» относится также к производным антител, гетеровалентным антителам и конструкциям антител, типа одноцепочечных Fv-фрагментов (scFv) или слитых с протеинами антител. Более подробно понятие «молекула антитела» будет объяснено ниже в описании.
Понятие «специфическое распознавание» в контексте настоящего описания обозначает, что молекула антитела обладает способностью специфически взаимодействовать и/или связываться по меньшей мере с двумя аминокислотами каждой из двух областей β-А4, указанных в настоящем описании. Это понятие относится к специфичности молекулы антитела, т.е. ее способности различать специфические указанные выше области пептида β-А4 и другие области, не связанные с пептидом β-A4, или другой, не связанный с АРР протеин /пептид/(не связанный) тест-пептид. Таким образом, специфичность можно определять экспериментально с помощью методов, известных в данной области, и методов, представленных в настоящем описании. Такие методы включают (но, не ограничиваясь ими) Вестерн-блоттинг, анализы ELISA (твердофазный иммуноферментный анализ), РИА (радиоиммунный анализ), ECL, ИРМА (иммунорадиометрический анализ) и пептидное сканирование. Такие методы включают также определение значений KD, что проиллюстрировано также ниже в примерах. Пептидное сканирование (pep spot-анализ), как правило, применяют для картирования линейных эпитопов в полипептиде антигена. Первичную последовательность полипептида синтезируют последовательно на активированной целлюлозе с пептидами, перекрывающими друг друга. Распознавание определенных пептидов антителом, которое тестируют в отношении способности обнаруживать или распознавать специфический антиген/эпитоп, оценивают стандартным методом, основанным на появлении цветного окрашивания (используют вторичное антитело, конъюгированное с пероксидазой из хрена, и 4-хлорнафтол и перекись водорода), с помощью реакции хемолюминесценции или с помощью аналогичных методов, известных в данной области. При применении, в частности, реакций хемолюминесценции реакцию можно оценивать количественно. Если антитело взаимодействует с определенным набором перекрывающихся пептидов, то можно выводить минимальную последовательность аминокислот, необходимую для реакции; см. приведенный в качестве иллюстрации метод, описанный в примере 6, и прилагаемую таблицу 2.
Этот же анализ позволяет обнаруживать два различных кластера реактивных пептидов, что свидетельствует о распознавании прерывистого, т.е. конформационного эпитопа в антигенном полипептиде (Geysen, Mol. Immunol. 23, 1986, сс.709-715).
Помимо pepspot-анализа можно осуществлять также стандартный метод ELISA. Как продемонстрировано ниже в примерах, небольшие гексапептиды можно сливать с протеином и сенсибилизировать планшет для иммунного анализа и подвергать взаимодействию с антителами. Оценку можно осуществлять с помощью стандартного метода, основанного на появлении цветного окрашивания (например, с помощью вторичного антитела, конъюгированного с пероксидазой из хрена, и тетраметилбензина с перекисью водорода). Реакцию в определенных лунках оценивают по оптической плотности (ОП), например, при длине волны 450 нм. Типичный фоновый уровень (соответствует отрицательной реакции) может составлять 0,1 ОП, типичный для положительной реакции уровень может составлять 1 ОП. Это означает, что различие (соотношение) положительной/отрицательной реакции может быть более чем 10-кратным. Дополнительные детали приведены ниже в примерах. Другие количественные методы определения специфичности и способности «специфически распознавать» указанные две области пептида β-А4 представлены ниже в настоящем описании.
Понятие «две области пептида β-А4» относится к двум областям, которые характеризуются аминокислотными последовательностями, представленными в SEQ ID NO:1 и 2, которые относятся к N-концевым аминокислотам 2-10 и центральным аминокислотам 12-25 пептида β-А4. Понятие «пептид β-А4» в контексте настоящего описания относится к указанным выше формам Аβ39, Аβ41, Аβ43, предпочтительно Аβ40 и Аβ42. Аβ42-форма представлена также ниже как последовательность SEQ ID NO:27. Следует отметить, что понятие «две области пептида β-А4» относится также к «эпитопу» и/или «антигенной детерминанте», которые содержат две указанные выше области пептида β-А4 или их фрагменты. Согласно настоящему изобретению эти две области пептида β-А4 разделены (на уровне аминокислотной последовательности) в первичной структуре пептида β-А4 по меньшей мере одной аминокислотой, предпочтительно по меньшей мере двумя аминокислотами, более предпочтительно по меньшей мере тремя аминокислотами, более предпочтительно по меньшей мере четырьмя аминокислотами, более предпочтительно по меньшей мере пятью аминокислотами, более предпочтительно по меньшей мере шестью аминокислотами, более предпочтительно по меньшей мере девятью аминокислотами и наиболее предпочтительно по меньшей мере двенадцатью аминокислотами. Как указано в настоящем описании и подтверждено ниже в примерах, антитела/молекул антител, предлагаемые в изобретении, обнаруживают/взаимодействуют и/или связываются с двумя указанными выше областями пептида β-А4, где эти две области разделены (на уровне первичной структуры аминокислотной последовательности) по меньшей мере одной аминокислотой и где последовательность, разделяющая две области /«эпитопы», может содержать более десяти аминокислот, предпочтительно 14 аминокислот, более предпочтительно 15 аминокислот или 16 аминокислот. Например, Fab-фрагмент MSR-3 (в качестве молекулы антитела, предлагаемой в изобретении) распознает, обнаруживает /взаимодействует с двумя областями пептида β-А4, где первая область содержит аминокислоты 3 и 4 (EF), и вторая область содержит аминокислоты 18-23 (DVFFAED). Таким образом, разделяющая последовательность между областью/эпитопами, которые подлежат обнаружению/распознаванию, состоит из 13 аминокислот в первичной структуре аминокислотной последовательности. Аналогично этому IgGI MSR. №3.4H7, оптимизированные и зрелые молекулы антител, полученные из MSR-3 и входящие в состав каркасного участка IgGI, обнаруживают/взаимодействуют/связываются с двумя эпитопами/областями β-А4, которые находятся в первой области в положениях 1-4 (DAEF) и во второй области в положениях 19-24 (FFAEDV) указанных выше областей β-А4. Таким образом, IgG1 MSR №3.4Н7 распознает/обнаруживает/взаимодействует/связывается с двумя эпитопами/областями, которые на уровне первичной аминокислотной последовательности разделены 14 аминокислотами. Как будет более подробно описано ниже в примерах, созревание аффинности и превращение моновалентных Fab-фрагментов, предлагаемых в изобретении, в полноразмерные антитела в виде IgGI может приводить к определенному расширению эпитопов/областей, обнаруживаемых с помощью pepspot-анализа, ELISA и т.п. Таким образом, молекулы антител, предлагаемые в изобретении, обладают способностью одновременно и независимо распознавать две области пептида β-А4/Aβ4, где указанные области содержат аминокислотную последовательность, представленную в SEQ ID NO:1 (или ее фрагменты), и аминокислотную последовательность, представленную в SEQ ID NO:2 (или ее фрагмент(ы)). Из-за возможного расширения эпитопов, что более подробно будет описано ниже, существует также вероятность того, что могут обнаруживаться/распознаваться аминокислоты, близко примыкающие к последовательностям SEQ ID NO:1 и 2, т.е. дополнительные аминокислоты являются фрагментами двух областей, подлежащих обнаружению/распознаванию. Таким образом, можно также предположить, что, например, первая аминокислота пептида Аβ (1-42), как он определен выше, т.е. D (аспарагиновая кислота), в фрагменте одного эпитопа будет обнаружена/распознана, или что аминокислоты, локализованные после области Аβ (1-42), соответствующей SEQ ID NO:2, будут обнаружены/распознаны. Эта дополнительная аминокислота может, например, представлять собой аминокислоту в положении 26 SEQ ID NO:27 (βА4/Аβ (1-42)), т.е. S (серин).
Понятие может относиться также к конформационному эпитопу, структурному эпитопу или к прерывистому эпитопу, состоящему из указанных двух областей или их фрагментов; см. также у Geysen (1986), loc. cit. В контексте настоящего описания конформационный эпитоп определен двумя или большим количеством дискретных аминокислотных последовательностей, разделенных на первичной последовательности, которые появляются вместе на поверхности, когда происходит укладка полипептида с образованием нативного протеина (Sela, Science 166, 1996, с.1365 и Laver, Cell 61, 1990, сс.553-556). Молекулы антител, предлагаемые в настоящем изобретении, вероятно, специфически связываются/взаимодействуют с конформационным(ыми)/структурным(и) эпитопом(ами), который(е) состоят из и/или содержат две описанные выше области β-А4 или их фрагменты, что будет подробнее описано ниже. Считается, что «молекулы антител», предлагаемые в настоящем изобретении, обладают одновременно и независимо двойной специфичностью по отношению к (а) аминокислотному участку, содержащему аминокислоты 2-10 (или его фрагменту(ам)) пептида β-А4, и (б) аминокислотному участку, содержащему аминокислоты 12-25 (или его фрагменту(ам)) пептида β-А4 (SEQ ID NO.27). Фрагменты или части этих участков содержат по меньшей мере две, более предпочтительно по меньшей мере три аминокислоты. Предпочтительные фрагменты или части расположены в первой области/участке SEQ ID NO:27 и имеют аминокислотные последовательности AEFRHD, EF, EFR, FR, EFRHDSG, EFRHD или HDSG и во второй области/участке SEQ ID NO:27 и имеют аминокислотные последовательности HHQKL, LV, LVFFAE, VFFAED VFFA или FFAEDV. Как отмечалось выше, эти фрагменты могут содержать также дополнительные аминокислоты или могут представлять собой части указанных выше фрагментов. Их конкретными примерами являются DAE, DAEF, FRH или RHDSG.
В данной области описан целый ряд антител, которые специфически распознают Аβ-пептиды. Эти антитела главным образом получают, осуществляя иммунизацию животных Аβ 1-40 или Аβ 1-42 или их фрагментами с использованием стандартных методик. Согласно опубликованным данным моноклональные антитела, которые получают путем иммунизации с использованием полного Аβ-пептида (1-40 или 1-42), распознают исключительно эпитоп, примыкающий к N-концу Аβ. Кроме того, примерами антител являются антитела ВАР-1 и ВАР-2 (Brockhaus, неопубликованные данные), которые получают иммунизацией мышей с помощью Аβ1-40 и которые распознают аминокислоты 4-6 в контексте более крупных Аβ-пептидов; см. ниже пример 7, таблица 2 и пример 12, таблица 7. Антитела, которые распознают среднюю часть Аβ, получают иммунизацией пептидами меньшего размера. Например, антитело 4G8, полученное иммунизацией Аβ-пептидом 1-24, распознает исключительно последовательность 17-24 (Kim, Neuroscience Research Communications 2, 1988, сс.121-130). Целый ряд других моноклональных антител был получен иммунизацией мышей выведенными из Ар фрагментами, и антитела, которые распознают С-конец Аβ1-40 и Аβ1-42, широко применяют для выявления и количественной оценки соответствующих Аβ-пептидов в биологических жидкостях и тканях с помощью ELISA, Вестерн-блоттинга и иммуногистохимических анализов (Ida и др., J. Biol. Chem. 271, 1996, сс.22908-22914; Johnson-Wood и др., Proc. Natl. Acad. Sci. USA 1994, 1997, сс.1550-1555; Suzuki и др. Science 264, 1994, сс.1336-1340; Brockhaus Neuro Rep. 9, 1998, сс.1481-1486). ВАР-17 представляет собой мышиное моноклональное антитело, полученное иммунизацией мышей Аβ-фрагментом 35-40. Оно специфически распознает С-конец Aβ1-40 (Brockhaus Neuroreport 9, 1998, сс.1481-1486).
Вероятно, для иммунизации зависящими от Т-клеток антигенами (которые часто являются слабыми иммунгенами) требуется протеолитическое расщепление антигена в эндосомах антигенпредставляющих клеток. Отбор in vivo высокоаффинных антител после иммунизации обеспечивается контактом Т-клеток-хелперов с антигенпредставляющими клетками. Антиген презентирующие клетки презентируют только короткие пептиды и не представляют полипептиды более крупного размера. Таким образом, эти клетки имеют сложный (но хорошо известный) механизм, предназначенный для эндоцитоза антигена(ов), расщепления антигена(ов) в эндосомах, объединения отобранных пептидов с пригодными молекулами МНС класса II и экспорта комплекса пептид-МНС на клеточную поверхность. Это имеет место, когда происходит специфическое распознавание антигена Т-клетками, что обеспечивает помощь созревающим В-клеткам. В-клетки, которые получили наибольшую помощь от Т-клеток, имеют предпочтительный шанс к превращению в секретирующие антитела клетки и к пролиферации. Этот факт свидетельствует о том, что процессинг антигенов с помощью протеолиза является важной стадией для генерации высокоаффинного гуморального иммунного ответа in vivo и может объяснять доминирование N-концевого Аβ-эпитопа для известных из существующего уровня техники моноклональных и поликлональных антител, полученных иммунизацией.
В противоположность этому отбор антител/молекул антител, предлагаемых в настоящем изобретении, обусловливается физической адгезией экспрессирующих Fab-фрагменты фагов с антигеном. В этом процессе отбора in vitro отсутствует стадия расщепления антигена. Фаги, экспрессирующие Fab-фрагмент, который обладает наиболее высокой аффинностью к антигену, отбирают и размножают. Синтетическая библиотека, применяемая в приведенных ниже примерах для отбора специфических молекул антител, предлагаемых в настоящем изобретении, наиболее пригодна для исключения любых стандартных ошибок, связанных с индивидуальными непрерывными эпитопами, которые часто встречаются в библиотеках, выведенных из иммунизированных В-клеток.
Следует отметить, что в известном уровне техники не описаны молекулы антител, распознающие две независимые области Аβ4, которые специфически распознают (а) прерывистый(ые)/структурный(ые)/конформационный(ые) эпитоп(ы) и/или которые обладают способностью одновременно и независимо распознавать две(два) области/эпитопа Aβ4.
Вакцинация трансгенных мышей, для которых характерна сверхэкспрессия мутантного человеческого APPV717F (PDAPP-мыши), с использованием Аβ1-42 приводит к практически полному предотвращению отложения амилоида в головном мозге, если лечению начинают подвергать молодых животных, т.е. до появления нейропатологий, в то время как у более старых животных наблюдается уменьшение уже образовавшихся бляшек, что позволяет предположить наличие опосредуемого антителом клиренса бляшек (Schenk и др., Nature, 400, 1999, сс.173-177). Антитела, полученные с помощью такой процедуры иммунизации, обладают реактивностью в отношении N-конца Аβ4, простирающегося на эпитоп вокруг аминокислот 3-7 (Schenk и др., (1999), loc. cit.; WO 00/72880). Активная иммунизация с использованием Aβ1-42 также снижала нарушение поведения и потерю памяти в различных трансгенных моделях болезни Альцгеймера (Janus и др. Nature 408, (2000), сс.979-982; Morgan и др., Nature 408, (2000), сс.982-985). Последующие исследования с использованием периферического введения антител, т.е. пассивной иммунизации, подтвердили, что антитела могут проникать в центральную нервную систему, «метить» бляшки и индуцировать клиренс уже существующих амилоидных бляшек у трансгенных экспрессирующих АРР мышей (PDAPP-мышей) (Bard и др., Nat. Med. 6, (2000), сс.916-919; WO 00/72880). В этих исследованиях моноклональные антитела, обладающие наиболее высокой эффективностью in vivo и ex vivo (стимулирующие фагоцитоз в экзогенных микроглиальных клетках), представляли собой антитела, которые распознавали N-концевые эпитопы 1-5 Аβ4 (МАт 3D6, IgG2b) или 3-6 (МАт 10D5, IgGI). Аналогично этому, поликлональные антитела, выделенные из организма мышей, кроликов или обезьян после иммунизации Aβ1-42, характеризовались аналогичной специфичностью в отношении N-концевого эпитопа и также обладали способностью стимулировать фагоцитоз и клиренс бляшек in vivo. В противоположность этому специфические для С-конца антитела, связывающиеся с высокой аффинностью с Аβ1-40 или Aβ1-42, не индуцировали фагоцитоз ех vivo и не обладали эффективностью in vivo (WO 00/72880). Было получено моноклональное антитело m266 (WO 00/72880), которое вырабатывалось при обработке Ар 13-28 (центральный домен Аβ), и картирование эпитопа подтвердило специфичность антитела в отношении аминокислот 16-24 в последовательности Аβ. Это антитело плохо связывается с агрегированным Аβ и амилоидными отложениями и взаимодействует только с растворимым (мономерным) Аβ, т.е. обладает свойствами, аналогичными свойствам другого хорошо известного и поступающего в продажу моноклонального антитела (4G8; Kim, Neuroscience Research Communications 2, 1988, сс.121-130; поступающего в продажу от фирмы Signet Laboratories Inc. Дедхам, шт. Миннесота, США), который распознает этот же эпитоп.
В настоящее время в опытах in vivo установлено, что антитело m266 после периферического введения в значительной степени снижает отложение Аβ в организме PDAPP-мышей (DeMattos, Proc. Nail. Acad. Sci. USA, 98, 2001, сс.8850-8855). Однако m266 в противоположность специфическим для N-конца антител не распознает амилоидных бляшек in vivo, и поэтому можно предположить, что уровень Аβ в головном мозге снижается в результате индуцированного антителом сдвига в равновесии между содержанием Аβ в ЦНС и плазме, что приводит к накоплению полученного из головного мозга Ар на периферии в виде устойчивого комплекса с m266 (DeMattos, (2001), loc. cit.).
Антитела/молекулы антител, предлагаемые в настоящем изобретении, в результате одновременного связывания (например, с структурным/конформационным эпитопом, образованным N-концевой и центральной областью βА4, представленным в настоящем описании) и независимого связывания (например, как установлено с помощью pepspot-анлизов, описанных в приведенном ниже экспериментальном разделе) с N-концевым и центральным эпитопами объединяют в одной молекуле свойства специфического для N-конца антитела и антитела, специфического для расположенного в центральной области эпитопа. Антитела со специфичностью к двум эпитопам, которые описаны в настоящем изобретении, вероятно, должны обладать большей эффективностью in vivo, в частности при использовании в медицинских и диагностических процедурах, например, для снижения массы амилоидных бляшек или амилоидогенеза или для уменьшения отложений амилоидов в бляшках. Так как индивидуальные специфические для N-конца и специфические для расположенного в центральной области эпитопа антитела, как было установлено, снижают уровень амилоидных бляшек в трансгенных мышах, антитела, специфические для двух эпитопов, которые описаны в настоящем изобретении, могут обладать более высокой эффективностью in vivo. Хорошо известно, что в процессе агрегации Аβ4 и отложения амилоида происходят конформационные изменения, и если находящийся в центральной области эпитоп легко доступен в растворимом Аβ4, вероятно, он становится спрятанным и менее реактивным в агрегированном или фибриллярном Аβ4. Тот факт, что специфическое в отношении центрального/срединного эпитопа антитело m266 обладает эффективностью in vivo, свидетельствует о том, что нейтрализация растворимого Аβ4 может также представлять собой имеющий решающее значение параметр. Антитела/молекулы антител, предлагаемые в настоящем изобретении, благодаря их специфичности в отношении двух эпитопов, могут связываться как с фибриллярным, так и с растворимым Аβ4 с одинаковой эффективностью, что обеспечивает как взаимодействие с амилоидными бляшками, так и нейтрализацию растворимого Аβ4. Понятие «одновременно и независимо связывается с N-концевыми и центральными/срединными эпитопами β-А4» в контексте настоящего описания применительно к молекулам антител относится к тому факту, что антитела/молекулы антител, представленные в настоящем описании, могут обнаруживать и/или связываться одновременно с обоими эпитопами, т.е. в одно и то же время (например, с конформационными/структурными эпитопами, образованными N-концевым эпитопом (или его фрагментом(ами) и центральными эпитопами (или его фрагментом(ами)) βА4, как указано выше, и что одни и те же молекулы антител при этом могут также обнаруживать/связываться с каждым из указанных эпитопов независимым образом, что среди прочего продемонстрировано с помощью описанного в примерах pepspot-анализа.
Клиренс амилоидных бляшек in vivo из организма PDAPP-мышей после непосредственной обработки головного мозга антителами не зависит от подтипа IgG и может включать также механизм, не опосредуемый Fc, т.е. при котором активированная микроглия не вовлечена в клиренс бляшек (Bacskai, Abstract Society for Neuroscience 31ый Annual Meeting, ноябрь 10-15, 2001, Сан-Диего). Эти данные не согласуются с выводами, постулированными в более раннем исследовании Bard (2000), loc. cit.
В другом исследовании обнаружено, что антитела к пептидам Аβ1-28 и Аβ1-16 являются эффективными в отношении дисагрегации Аβ-фибрилл in vitro, в то время как антитело, специфическое в отношении Ар 13-28, оказалось менее активным в этом опыте (Solomon, Proc. Natl. Acad. Sci. USA, 94, 1997, сс.4109-4112). Также известны данные о предотвращении агрегации Аβ с помощью антитела к Aβ1-28 (AMY-33) (Solomon, Proc. Natl. Acad. Sci. USA, 93, 1996, сс.452-455). В этом же исследовании установлено, что образовавшееся антитело 6F/3D к Аβ-фрагменту 8-17 оказывало слабое воздействие на индуцируемую Zn2+ агрегацию Аβ и совсем не оказывало действия на самоагрегацию, индуцируемую другими индукторами агрегации.
В этих опытах установлено, что эффективность различных антител in vitro коррелирует с доступностью для них эпитопов в агрегатах Аβ4. N-конец является доступным, и специфические для N-конца антитела выраженно индуцируют деполимеризацию, в то время как центральная область и С-конец спрятаны и не являются легкодоступными, и поэтому антитела к этим эпитопам обладают существенно меньшей эффективностью.
Исследования, касающиеся доступности эпитопов для антител, позволили установить, что в агрегированном Аβ N-концевой эпитоп доступен и вступает в реакцию с антителом ВАР-1, в то время как срединный или центральный эпитоп в действительности остается скрытым, т.е. не связывается с антителом 4G8. Однако в мономерном Аβ оба эпитопа открыты и одинаково распознаются обоими антителами, которые известны из существующего уровня техники.
В противоположность вышесказанному при создании настоящего изобретения неожиданно было установлено, что предлагаемые в изобретении молекулы антител распознают две прерывистые аминокислотные последовательности, например, конформационный/структурный эпитоп, в пептиде Аβ. Понятие две «прерывистые аминокислотные последовательности» в контексте настоящего описания обозначает, что две указанные аминокислотные последовательности, образующие N-конец и центральный/срединный эпитопы соответственно, разделены в первичной структуре β-А4 по меньшей мере двумя аминокислотами, которые не являются частью любого эпитопа.
Антигенсвязвающий центр Fab-фрагмента антитела (паратоп) занимает на поверхности молекулы участок размером примерно 30×30  (Laver, Cell, 61 1990, сс.553-556). Это достаточно для контакта с 15-22 аминокислотными остатками, которые могут присутствовать на поверхности нескольких петель. Прерывистый эпитоп, распознаваемый молекулами антител, предлагаемыми в изобретении, напоминает конформацию, при которой N-конец (остатки 2-10 или их фрагменты) и срединные пептидные последовательности Аβ (остатки 12-25 или их фрагменты) находятся в пространственной близости. Только в рамках такой конформации можно получить максимальное количество контактов антиген-антитело и состояние с наиболее низким значением свободной энергии.
(Laver, Cell, 61 1990, сс.553-556). Это достаточно для контакта с 15-22 аминокислотными остатками, которые могут присутствовать на поверхности нескольких петель. Прерывистый эпитоп, распознаваемый молекулами антител, предлагаемыми в изобретении, напоминает конформацию, при которой N-конец (остатки 2-10 или их фрагменты) и срединные пептидные последовательности Аβ (остатки 12-25 или их фрагменты) находятся в пространственной близости. Только в рамках такой конформации можно получить максимальное количество контактов антиген-антитело и состояние с наиболее низким значением свободной энергии.
На основе энергетических расчетов было высказано предположение о том, что меньший по размеру поднабор, состоящий из 5-6 остатков, которые не организованы в линейную последовательность, а разбросаны по поверхности эпитопа, вносят наибольший вклад в энергию связывания, в то время как окружающие их остатки могут представлять собой только дополнительные элементы (Laver (1990) loc. cit.).
Антитела/молекулы антител, предлагаемые в изобретении, обладают способностью связываться с агрегированным Аβ и дают сильную реакцию с амилоидными бляшками в головном мозге страдающих болезнью Альцгеймера (AD) пациентов (что продемонстрировано ниже в примерах). Кроме того, они обладают способностью деполимеризировать/дезинтегрировать амилоидные агрегаты.
Не вдаваясь в теорию, можно предполагать, что конформационный эпитоп (состоящий из двух областей Аβ4 или их фрагмента(ов), как описано выше) частично доступен в агрегированном Аβ. Однако известно, что большая часть срединного(ой)/вторичного(ой) эпитопа/области сама по себе не является легко доступной в этих агрегатах Аβ (предположение основано на низкой реактивности специфических для срединных эпитопов антител 4G8 и m266). С другой стороны, и в контексте рассматриваемых ранее положений вероятно, что один или несколько остатков срединной области являются компонентами конформационного эпитопа, и в сочетании с остатками N-концевой области они доступны для антител, предлагаемых в настоящем изобретении, вследствие чего они вносят существенный вклад в энергию связывания взаимодействия антитело-Аβ4. Таким образом, реактивность молекул антител, предлагаемых в изобретении, с конформационным эпитопом в агрегированном Аβ является уникальной и отличной от реактивности антител к α-Аβ4, известных из существующего уровня техники. Кроме того, как отмечалось выше, еще одной уникальной особенностью антител/молекул антител, предлагаемых в изобретении, является их способность одновременно и независимо связываться/ распознавать два различных эпитопа в β-А4, как они описаны выше и в прилагаемых примерах.
Согласно предпочтительному варианту осуществления изобретения молекула антитела, предлагаемая в изобретении, представляет собой молекулу антитела, где по меньшей мере две области β-А4, специфически распознаваемые этим антителом, образуют конформационный эпитоп/структурный эпитоп или прерывистый эпитоп; см. у Geysen (1986), loc. cit.; Ghoshal, J. Neurochem. 77, 2001, сс.1372-1385; Hochleitner, J. Imm. 164, 2000, сс.4156-4161; Laver (1990), loc. cit. Понятие «прерывистый эпитоп» в контексте настоящего описания обозначает нелинейные эпитопы, состоящие из остатков из отдаленных друг от друга участков полипептидной цепи. Эти остатки появляются вместе на поверхности при укладке полипептидной цепи в трехмерную структуру для образования конформационного/структурного эпитопа. Настоящее изобретение относится к предпочтительным неизвестным ранее эпитопам в β-А4, которые обеспечивают секрецию согласно изобретению специфических молекул антител, обладающих способностью взаимодействовать с этими эпитопами. Эти предлагаемые в изобретении антитела/молекулы антител являются основой для повышения эффективности и обладают пониженной способностью оказывать побочные действия. Как отмечалось выше, предлагаемые в изобретении антитела при этом обладают также способностью независимо взаимодействовать с каждой(ым) из указанных двух областей/эпитопов β-А4, например, что установлено с помощью pepspot-анализов, описанных ниже в примерах.
Таким образом, настоящее изобретение относится к уникальным «инструментам», которые можно применять для деполимеризации агрегированных Аβ-фибрилл in vivo и in vitro, и/или которые обладают способностью стабилизировать и/или нейтрализовать конформационный эпитоп мономерного Аβ и тем самым могут предупреждать патологическую агрегацию Аβ.
Кроме того, можно предполагать, что предлагаемые в изобретениие антитела связываются с отложениями на краю Аβ-амилоидных бляшек в том числе среди прочего в головном мозге страдающих болезнью Альцгеймера пациентов и эффективно растворяют протофибриллы и фибриллы, с которыми связана патология.
Согласно предпочтительному варианту осуществления изобретения молекула антитела, предлагаемая в изобретении, распознает по меньшей мере две последовательные аминокислоты, расположенные в двух областях Aβ4, как они определены выше, более предпочтительно эта молекула антитела распознает в первой области аминокислотную последовательность, которая содержит аминокислоты: AEFRHD, EF, EFR, FR, EFRHDSG, EFRHD или HDSG, и во второй области аминокислотную последовательность, которая содержит аминокислоты: HHQKL, LV, LVFFAE, VFFAED, VFFA или FFAEDV. Кроме того, фрагменты или расширенные участки содержат: DAE, DAEF, FRH или RHDSG.
Особенно предпочтительно, когда молекула антитела, предлагаемая в изобретении, содержит вариабельную VH-область, кодируемую молекулой нуклеиновой кислоты, представленной в SEQ ID NO:3, 5 или 7, или вариабельную VH-область, которая имеет аминокислотные последовательности, представленные в SEQ ID NO:4, 6 или 8. Последовательности, представленные в SEQ ID NO:3 и 4, обозначают кодирующую область и аминокислотную последовательность соответственно VH-области предлагаемого в изобретении родительского антитела MSR-3 (MS-Roche 3), последовательности, представленные в SEQ ID NO:5 и 6, обозначают кодирующую область и аминокислотную последовательность соответственно VH-области предлагаемого в изобретении родительского антитела MSR-7 (MS-Roche 7), и SEQ ID NO:7 и 8 обозначают кодирующую область и аминокислотную последовательность соответственно VH-области предлагаемого в изобретении родительского антитела MSR-8 (MS-Roche 8). Таким образом, изобретение относится также к молекулам антител, которые содержат вариабельную VL-область, кодируемую молекулой нуклеиновой кислоты, которая имеет последовательность (SEQ ID NO), выбранную из ряда, включающего SEQ ID NO:9, 11 или 13, или вариабельную VL-область, имеющую аминокислотную последовательность, которая представлена в SEQ ID NO:10, 12 или 14. SEQ ID NO:9 и 10 соответствуют VL-области MSR-3, SEQ ID NO:11 и 12 соответствуют VL-области MSR-7 и SEQ ID NO:13 и 14 соответствуют VL-области MSR-8. Как описано в приведенных ниже примерах, родительские антитела MSR-3, -7 и -8 применяют для последующего создания оптимизированных молекул антител, которые обладают либо улучшенными свойствами и/или сродством к связыванию. Некоторые из соответствующих и возможных стратегий представлены в настоящем описании и приведены ниже в примерах.
Применение стратегии оптимизации, как это проиллюстрировано ниже в примерах, позволяет получать множество предлагаемых в изобретении оптимизированных антител. Эти оптимизированные антитела несут такой же как у родительских антител CDR-3-участок VH-области. Поскольку исходный каркасный участок (представленный ниже на фиг.1) остается без изменений, то в созревших/оптимизированных молекулах антител изменяют CDR1, CDR2 и/или CDR3-участки VL-области. Приведенные в качестве иллюстрации модифицированные последовательности мотивов оптимизированных молекул антител представлены ниже в таблице 1. Таким образом, под объем настоящего изобретения подпадают также оптимизированные молекулы антител, которые получают из описанных антител MSR-3, -7 и -8 и которые обладают способностью специфически реагировать на/специфически распознавать две указанные области пептида β-А4. В частности CDR-участки, предпочтительно CDR1, более предпочтительно CDR1- и CDR2-, наиболее предпочтительно CDR1-, CDR2- и CDR3-участки, как они обозначены в описании, можно применять для создания дополнительных антител/молекул антител, предлагаемых в настоящем изобретении, среди прочего с помощью методов трансплантации CDR, известных в данной области; см. у Jones, Nature 321, 1986, сс.522-515 или у Riechmann, Nature 332, 1988, сс.323-327. Наиболее предпочтительно антитела/молекулы антител, предлагаемые в настоящем изобретении, а также фрагменты или производные антител получают из указанных родительских антител, и они несут, как указано выше, CDR-3-участок VH-области, совпадающий с соответствующим участком по меньшей мере одного из указанных родительских антител. Как проиллюстрировано ниже, также рассматривается вопрос о том, чтобы с помощью перекрестного клонирования получать антитела, которые следует рассматривать как оптимизированные/созревшие антитела/молекулы антител, предлагаемые в настоящем изобретении. Таким образом, предпочтительные молекулы антител могут содержать также или их можно получать также из антител/молекул антител, которые характеризуются VH-областями, представленными в любой из последовательностей SEQ ID NO:32-45, VL-областями, представленными в любой из последовательностей SEQ ID NO:46- 59, или которые могут содержать CDR-3-участок, представленный в любой из SEQ ID NO:60-87. Согласно конкретному предпочтительному варианту осуществления изобретения оптимизированная молекула антитела, предлагаемая в настоящем изобретении, содержит VH-области и VL-области, представленные в SEQ ID NO:88/89 и 90/91 соответственно или их фрагменты. Кроме того, они могут представлять собой CDR-участок(H), предпочтительно CDR3-участок(H). Наиболее предпочтительные молекулы антител оптимизированного типа несут H-CDR3, который представлен в SEQ ID NO:92 или 93, и/или L-CDR3, который представлен в SEQ ID NO:94 или 95. Предпочтительно, чтобы антитела/молекулы антител, предлагаемые в изобретении, отличались по специфической реактивности в отношении β-А4 и/или пептидов-производных β-А4. Например, можно определять оптические плотности с помощью тестов на основе ELISA, как описано ниже в примерах, и применять соотношение оптических плотностей для оценки специфической реактивности родительских или оптимизированных антител. Таким образом, предпочтительное антитело, предлагаемое в изобретении, представляет собой антитело, которое при оценке с помощью ELISA при оптической плотности, измеренной при 450 нм, дает реакцию с β-А4, превышающую в 10 раз оптическую плотность, измеренную без β-А4, т.е. превышающую в 10 раз фоновый уровень. Предпочтительно измерение оптической плотности осуществляют в течение нескольких минут (например, 1, 2, 3, 4, 5, 6 или 7 мин) после инициации реакции, характеризующейся появлением окрашивания, с участием антитела для того, чтобы оптимизировать соотношение сигнала и фонового уровня.
Согласно особенно предпочтительному варианту осуществления изобретения молекула антитела, предлагаемая в изобретении, содержит по меньшей мере один CDR3-участок VL-области, кодируемый молекулой нуклеиновой кислоты, которая представлена в SEQ ID NO:15, 17 или 19, или по меньшей мере один CDR3-участок аминокислотной последовательности VL-области, представленной в SEQ ID NO:16, 18 или 20, и/или указанная молекула антитела содержит по меньшей мере один СDR3-участок VH-области, кодируемый молекулой нуклеиновой кислоты, которая представлена в SEQ ID NO:21, 23 или 25, или по меньшей мере один СDR3-участок аминокислотной последовательности VH-области, представленный в SEQ ID NO:22, 24 или 26. Наиболее предпочтительными являются антитела, которые содержат по меньшей мере один СDR3-участок VH-области, представленный в настоящем описании. Указанные выше CDR3-участки родственны участкам предлагаемых в настоящем изобретении, приведенных в качестве иллюстрации родительских молекул антител MSR-3, -7 или -8. Однако, как проиллюстрировано ниже в таблицах 1, 8 или 10, созревшие и/или оптимизированные молекулы антител, получаемые с помощью методов, которые описаны ниже в примерах, могут содержать модифицированные CDR1-, CDR2- и СDR3-участки VH-, VL-областей. Таким образом, молекулу антитела, предлагаемую в изобретении, предпочтительно выбирают из ряда, включающего MSR-3, -7 и -8, или версию с созревшей аффинностью MSR-3, -7 или -8. Версии с созревшей аффинностью MSR-3, -7 и -8 содержат среди прочего молекулы антител, которые несут CDR1-, CDR2- и/или CDR3-участки, представленные в таблице 1 или 8, или которые характеризуются любой из последовательностей SEQ ID NO:15-20, 21- 26, 60-74, 75-87, 92 и 93 или 94 и 95, а также SEQ ID NO:354-413. Наиболее предпочтительно антитело, предлагаемое в изобретении, содержит по меньшей мере один CDR, предпочтительно CDR1, более предпочтительно CDR2, наиболее предпочтительно CDR3, которые представлены ниже в таблицах 1, 8 или в таблице 10.
Следует отметить, что методы созревания аффинности хорошо известны в данной области, и они описаны ниже в примерах и среди прочего у Knappik, J. Mol. Biol. 296, 2000, с. 55; Krebs, J. Imm. Meth. 254, 2000, сс.67-84; WO 01/87337; WO 01/87338; US 6300064; ЕР 96929278.8, а также в процитированных ниже ссылках.
Согласно более предпочтительному варианту осуществления изобретения молекула антитела представляет собой полноразмерное антитело (иммуноглобулин, типа IgG1, IgG2, IgG2b, IgG3, IgG4, IgA, IgM, IgD или IgE), F(ab)-, Fabc-, Fv-, Fab'-, Р(ab')2-фрагмент, одноцепочечное антитело, химерное антитело, антитело с трансплантированным CDR-участком, конструкцию бивалентного антитела, антитело, слитое с протеином, полученное путем перекрестного клонирования антитело или синтетическое антитело. Под объем изобретения подпадают также генетические варианты генов иммуноглобулинов. Генетические варианты, например, тяжелой цепи иммуноглобулина G подкласса 1 (IgG1), могут нести аллотипические маркеры Glm(17) или Glm(3) в СН1-домене или аллотипические маркеры Glm(1) или Glm(non-1) в СН3-домене. Молекула антитела, предлагаемая в изобретении, может содержать также модифицированные или мутантные антитела, типа мутанта IgG с усиленной или ослабленной способностью к связыванию с Fc-рецептором или активации комплемента. Предусмотрено также, что антитела, предлагаемые в изобретении, получают общепринятыми методами, например, для получения специфических моноклональных антител используют иммунизацию млекопитающих, предпочтительно мышей, пептидом, который содержит указанные выше две области βА4, например, N-концевую(ой) и центральную(ый) область/эпитоп, которые содержат (а) аминокислотный участок 2-10 (или его фрагмент(ы)) β-А4 и (б) аминокислотный участок, содержащий аминокислоты 12-25 (или его фрагмент(ы)) β-А4 (SEQ ID NO:27). Таким образом, специалист в данной области может получать моноклональные антитела к указанному пептиду и может осуществлять скрининг полученных антител на способность одновременно и независимо связываться/реагировать с N-концевой(ым) и центральной(ым) областью/эпитопом βА4, как они определены в настоящем описании. Соответствующие методы скрининга описаны ниже в примерах.
Как проиллюстрировано ниже в примерах, антитела/молекулы антител, предлагаемые в изобретении, можно легко конструировать с помощью метода рекомбинации и экспрессировать. Предпочтительно молекула антитела, предлагаемая в изобретении, содержит по меньшей мере один, более предпочтительно по меньшей мере два, предпочтительно по меньшей мере три, более предпочтительно по меньшей мере четыре, еще более предпочтительно по меньшей мере пять и наиболее предпочтительно по меньшей мере шесть CDR-участков представленных в описании родительских антител MSR-3, MSR-7 или MSR-8 или с созревшей аффинностью/оптимизированных антител, выведенных из указанных родительских антител. Следует отметить, что полученные с помощью метода рекомбинации антитела, предлагаемые в изобретении, могут содержать более шести CDR-участков. Специалист в данной области легко может с помощью информации, приведенной ниже примерах, определить CDR, соответствующие родительским, а также антитела с оптимизированной аффинностью. Примеры оптимизированных антител, которые получают с помощью созревания/оптимизации родительских антител, представлены среди прочего в приведенной ниже таблице 1. Созревшая/оптимизированная молекула антитела, предлагаемая в изобретении, представляет собой, например, антитело MSR 7.9H7, которое характеризуется также последовательностями, приведенными в настоящем описании, такими как SEQ ID NO:88-95, и которые соответствуют VH-области MSR 7.9H7 (SEQ ID NO:88 и 89), VL-области MSR 7.9H7 (SEQ ID NO:90 и 91), H-CDR3 антитела MSR 7.9H7 (SEQ ID NO:92 и 93), а также L-CDR3 антитела MSR 7.9H7 (SEQ ID NO:94 и 95). Приведенную в качестве примера молекулу антитела 7.9H7 получают из родительского антитела MSR7, и она является особенно предпочтительным примером оптимизированной/созревшей молекулы антитела, предлагаемой в настоящем изобретении. Эту молекулу антитела можно дополнительно модифицировать согласно настоящему изобретению, например, с помощью перекрестного клонирования, см. ниже и в прилагаемых примерах.
Как продемонстрировано в прилагаемых примерах, антитела, предлагаемые в изобретении, могут представлять собой также полученные перекрестным клонированием антитела, т.е. антитела, которые содержат различные области антител (например, CDR-участки) одного или нескольких указанных родительских антител или антител с оптимизированной аффинностью. Эти полученные перекрестным клонированием антитела могут представлять собой антитела с несколькими различными каркасными участками, причем наиболее предпочтительным каркасным участком является каркасный участок IgG, еще более предпочтительно каркасный участок IgG1, IgG2a или IgG2b. Особенно предпочтительным каркасным участком антитела является каркасный участок млекопитающего, наиболее предпочтительно человеческий каркасный участок. Домены на легкой и тяжелой цепях имеют одинаковое общее строение и каждый домен содержит четыре каркасных участка, последовательности которых являются относительно консервативными, связанные с тремя гипервариабельными участками, которые называют определяющими комплементарность участками (CDR1-3).
В контексте настоящего описания понятие «человеческий каркасный участок» относится к каркасному участку, который практически идентичен (примерно на 85% или более, обычно на 90-95% или более) каркасному участку встречающегося в естественных условиях человеческого иммуноглобулина. Каркасный участок антитела, который представляет собой объединенные каркасные участки, входящие в легкую и тяжелую цепи, служит для позиционирования и выстраивания в требуемой последовательности CDR-участков. CDR прежде всего ответственны за связывание с эпитопом антигена. Следует отметить, что в каркасный участок иммуноглобулина можно интродуцировать не только предпочтительные (человеческие) каркасные участки полученных перекрестным клонированием антител, которые представлены в настоящем описании, но также и молекулы, содержащие CDR, полученные среди прочего из указанных родительских антител MSR-3, -7 или 8 или созревших антител, выведенных из указанных родительских антител. Предпочтительными каркасными участками являются каркасные участки IgG1, IgG2a или IgG2b. Наиболее предпочтительными являются человеческие каркасные участки и каркасные участки человеческого IgG1.
Как описано ниже в примерах, среди прочего можно с помощью методов генной инженерии, известных в данной области, переносить также полные легкие цепи из оптимизированного клона-донора в оптимизированный клон-реципиент. Примером оптимизированного клона-донора является, например, L-CDR1 (L1), а примером оптимизированного клона-реципиента является H-CDR2 (Н2). Можно достигать консервативной эпитопспецифичности путем объединения клонов, которые несут одинаковые H-CDR-3-участки. Дополнительные детали проиллюстрированы в примере 13.
Предпочтительные полученные перекрестным клонированием молекулы антител, предлагаемые в изобретении, выбирают из группы, включающей MS-R №3.3H1×3.4L1, MS-R №3.3H1×3.4L9, MS-R №3.4H1×3.4L9, MS-R №3.4H3×3.4L7, MS-R №3.4H3×3.4L9, MS-R №3.4H7×3.4L9, MS-R №3.4H7×3.4L7, MS-R №3.6H5×3.6L1, MS-R №3.6H5×3.6L2, MS-R №3.6.H8×3.6.L2, MS-R №7.2H2×7.2L1, MS-R №7.4H2×7.2Ll, MS-R №7.4Н2×7.12L2, MS-R №7.9Н2×7.2L1(L1), MS-R №7.9Н2×7.12L1, MS-R 7.9H2×7.12L2, MS-R №7.9Н2×7.12L2(L1+2), MS-R №7.9Н4×7.11.L2, MS-R №7.11H1×7.2L1, MS-R №7.11Н1×7.11L1. MS-R №7.11H2×7.2L1(L1), MS-R №7.11Н2×7.9L1 (L1) или MS-R №8.1H1×8.2L1.
Получение антител перекрестным клонированием проиллюстрировано также в приведенных ниже примерах. Перечисленные выше предпочтительные полученные перекрестным клонированием антитела/молекулы антител представляют собой оптимизированные/созревшие молекулы антител, выведенные из описанных родительских антител, в частности из MSR-3 и MSR-7. Кроме того, характерные CDR-последовательности и V-области полученных перекрестным клонированием молекул антител/антител представлены ниже в SEQ ID NO:32, 33, 46 и 47 (MSR 3.6H5×3.6.L2; VH-, VL-области); 34, 35, 48 и 49 (MSR 3.6H8×3.6.L2; VH-, VL-области); 36, 37, 50 и 51 (MSR 7.4H2×7.2.L1; VH-, VL-области); 38, 39, 52 и 53 (MSR 7.9H2×7.12.L2; VH-, VL-области); 40, 41, 54 и 55 (MSR №7-9H4×7.12.L2; VH-, VL-области); 42, 43, 56 и 57 (MSR №7.11Н1×7.11.L1; VH-, VL-области) и 44, 45, 58 и 59 (MSR №7.11H1×7.2.L1; VH-, VL-области). Соответствующие CDR3-участки этих особенно предпочтительных полученных перекрестным клонированием молекул антител представлены в SEQ ID NO:60-87. Для других молекул MSR-антител CDR-участки VH-, VL-области можно обнаружить ниже в таблицах 8 или 10 и в прилагаемом перечне последовательностей, в частности SEQ ID NO:32-95 для MS-R- антител/молекул антител №3.6Н5×3.6L2, №3.6Н8×3.6L2, №7.4Н2×7.2L1, №7.9Н2×7.12L2, №7.9Н4×7.12L2, №7.11Н1×7.11L1, №7.11Н1×7.2L1 и №7.9Н7 или SEQ ID NO:294-413 для MSR-R-антител/молекул антител №3.3Н1×3.4L1, №3.4Н1×3.4L9, №3.4Н3×3.4L7, №3.4Н3×3.4L9, №3.4Н7×3.4L9, №3.4Н7×3.4L7, №3.6Н5×3.6L1, №7.2Н2×7.2L1, №7.4Н2×7.12L2, №7.9Н2x7.2L1, №7.9Н2×7.12L1, №7.11Н2×7.2L1, №7.11Н2×7.9L1, №7.11Н2×7.12L1 или №8.1Н1×8.2L1. Таким образом, помимо указанных выше VH-областей предпочтительные молекулы антител, предлагаемые в изобретении, могут содержать VH-области, указанные в любой из SEQ ID NO:294-323. Аналогично этому, в SEQ ID NO:324-353 представлены VL-области, которые помимо указанных выше VL-областей, могут нести молекулы антител, предлагаемые в изобретении. Соответствующие CDR-участки указаны выше, а также могут быть представлены последовательностями, указанными в SEQ ID NO:354-413.
Молекулы антител, предлагаемые в изобретении, можно легко получать в достаточных количествах в том числе с помощью методов рекомбинации, известных в данной области, см., например, у Bentley, Hybridoma 17, 1998, сс.559-567; Racher, Appl. Microbiol. Biotechnol. 40, 1994, сс.851-856; Samuelsson, Eur. J. Immunol. 26, 1996, сс.3029-3034.
Теоретически в растворимом βА4 (мономерном/олигомерном) как N-концевой, так и срединный эпитопы доступны для связи с антителом, и молекулы антител, предлагаемые в настоящем изобретении, могут связываться по отдельности либо с N-концевым, либо со срединным эпитопом, но в таких условиях не может быть достигнута максимальная аффинность. Однако более вероятно, что оптимальный контакт с паратопом антитела будет достигаться при одновременном связывании с обоими эпитопами, т.е. аналогично связыванию с агрегированным β-А4. Таким образом, антитела, предлагаемые в настоящем изобретении, являются уникальными антителами к Аβ, поскольку они связываются с агрегированным β-А4 (посредством взаимодействия с N-концевым и срединным эпитопом) и при этом могут также стабилизировать и нейтрализовать конформационный эпитоп в растворимом β-А4. Эти антитела отличаются от известных из протипов антител.
Наиболее предпочтительными являются молекулы антител, предлагаемые в изобретении, аффинность которых к Аβ или его определенным фрагментам характеризуется значением KD, составляющим менее 2000 нМ, предпочтительно менее 100 нМ, более предпочтительно менее 10 нМ, наиболее предпочтительно менее 1 нМ. Оценку аффинности/аффинностей можно осуществлять методами, которые проиллюстрированы в примерах и известны в данной области. Такие методы включают (но, не ограничиваясь ими) BIACORETM-анализы (www.biacore.com; Malmquist, Biochem. Soc. Trans, 27, 1999, сс.335-340) и твердофазные анализы с использованием меченых антител или меченого Аβ.
Предпочтительно молекула антитела, предлагаемая в изобретении, обладает способностью выявлять /реагировать/связываться с амилоидными бляшками in vitro в срезах головного мозга пациентов, страдающих связанными с амилоидом нарушениями типа болезни Альцгеймера. Предпочтительно, чтобы антитело/молекулы антител, предлагаемые в изобретении, могли препятствовать агрегации Аβ как in vivo, так и в опытах in vitro, что проиллюстрировано ниже в примерах. Аналогично этому, молекулы антител, предлагаемые в настоящем изобретении, предпочтительно обладают способностью деполимеризировать агрегат Aβ in vivo и/или in vitro, что описано ниже в примерах. Эту способность антител/молекул антител, предлагаемых в изобретении, можно применять в медицинских процедурах, прежде всего в описанных ниже фармацевтических композициях.
Изобретение относится также к молекуле нуклеиновой кислоты, кодирующей описанную молекулу антитела, предлагаемую в изобретении.
Такая молекула нуклеиновой кислоты может представлять собой встречающуюся в естественных условиях молекулу нуклеиновой кислоты, а также рекомбинантную молекулу нуклеиновой кислоты. Таким образом, молекула нуклеиновой кислоты, предлагаемая в изобретении, может иметь естественное происхождение или может представлять собой синтетическую или полусинтетическую молекулу. Она может представлять собой ДНК, РНК, а также ПНК (пентозонуклеиновую кислоту) и может представлять собой их гибрид.
Специалисту в данной области должно быть очевидно, что к молекуле нуклеиновой кислоты, предлагаемой в изобретении, можно добавлять регуляторные последовательности. Например, можно применять промоторы, энхансеры транскрипции и/или последовательности, которые могут индуцировать экспрессию полинуклеотида, предлагаемого в изобретении. Приемлемой индуцибельной системой является, например, регулируемая тетрациклином система экспрессии генов, описанная, например, у Gossen и Bujard (Proc. Natl. Acad. Sci. USA 89, (1992), сс.5547-5551) и у Gossen и др. (Trends Biotech. 12, 1994, сс.58-62), или индуцируемая дексаметазоном система экспрессии генов, описанная, например, у Crook, EMBO J. 8, 1989, сс.513-519.
Кроме того, для дополнительных целей молекула нуклеиновой кислоты может содержать, например, тиоэфирные связи и/или нуклеотидные аналоги. Такие модификации могут быть ценными для стабилизации молекулы нуклеиновой кислоты при воздействии эндо- и/или экзонуклеаз в клетке. Такие молекулы нуклеиновых кислот можно транскрибировать с помощью соответствующего вектора, содержащего химерный ген, который обеспечивает транскрипцию указанной молекулы нуклеиновой кислоты в клетке. В этой связи следует также понимать, что полинуклеотид, предлагаемый в изобретении, можно применять для «направленного переноса гена» или для методов «генной терапии». Согласно предпочтительному варианту осуществления изобретения указанные молекулы нуклеиновых кислот являются мечеными. Методы обнаружения нуклеиновых кислот хорошо известны в данной области и представляют собой Саузерн- или Нозерн-блоттинг, ПЦР или удлинение праймера. Этот вариант осуществления можно применять для осуществления скрининга с целью подтверждения успешной интродукции молекул нуклеиновых кислот, предлагаемых в изобретении, при генной терапии.
Молекулы(а) нуклеиновых(ой) кислот(ы), предлагаемые(ая) в изобретении, могут(жет) представлять собой полученную рекомбинантным путем химерную молекулу нуклеиновой кислоты, которая несет любую из вышеуказанных молекул нуклеиновых кислот либо индивидуально, либо в сочетании. Предпочтительно молекула нуклеиновой кислоты, предлагаемая в изобретении, является частью вектора.
Таким образом, настоящее изобретение относится к вектору, содержащему молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении.
Вектор, предлагаемый в настоящем изобретении, может представлять собой, например, плазмиду, космиду, вирус, бактериофаг или другой вектор, который, например, является общепринятым для генной инженерии и который может включать дополнительные гены, такие как маркерные гены, позволяющие осуществлять отбор этого вектора в пригодной клетке-хозяине и в приемлемых условиях.
Кроме того, вектор, предлагаемый в настоящем изобретении, может помимо нуклеотидных последовательностей, предлагаемых в изобретении, содержать элементы, контролирующие экспрессию, которые обеспечивают правильную экспрессию кодирующих областей в пригодных хозяевах. Такие контролирующие элементы являются известными специалисту и могут представлять собой промотор, кассету для сплайсинга, кодон инициации трансляции, трансляционный и инсерционный сайт для интродукции вставки в вектор. Предпочтительно молекула нуклеиновой кислоты, предлагаемая в изобретении, функционально связана с указанными контролирующими экспрессию последовательностями, которые обеспечивают экспрессию в эукариотических или прокариотических клетках.
Контролирующие элементы, которые обеспечивают экспрессию в эукариотических и прокариотических клетках, хорошо известны специалистам в данной области. Как отмечено выше, они, как правило, включают регуляторные последовательности, которые обеспечивают инициацию транскрипции, и необязательно поли-А-сигналы, обеспечивающие терминацию транскрипции и стабилизацию транскрипта. Дополнительные регуляторные элементы могут представлять собой энхансеры транскрипции, а также трансляции и/или встречающиеся в естественных условиях или гетерологичные промоторные области. Возможные регуляторные элементы, которые обеспечивают экспрессию, например, в клетках-хозяевах, полученных из организма млекопитающих, представляют собой промотор тимидинкиназы CMV-HSV (цитомегаловирус-вирус простого герпеса), промотор ОВ40, RSV (вирус саркомы Рауса), промотор человеческого фактора 1α удлинения, индуцируемый глюкокортикоидом промотор MMTV (вирус мышиного лейкоза Молони), индуцируемые металлотионеином или тетрациклином промоторы, или энхансеры типа CMV-энханмера или ОВ40-энхансера. Для экспрессии в нейронах можно применять промоторы нейрофиламента, PGDF (тромбоцитарный фактор роста), NSE (нейрон-специфическая энолаза), РгР (обогащенный пролином протеин слюнных желез) или thy-1-промоторы. Такие промоторы хорошо известны в данной области и описаны среди прочего у Charron, J. Biol. Chem. 270, 1995, сс.25739-25745. Для экспрессии в прокариотических клетках описана группа промоторов, включающая, например, tac-lac-промотор или trp-промотор. Помимо элементов, ответственных за инициацию транскрипции, регуляторные элементы могут также представлять собой сигналы терминации транскрипции, такие как ОВ40-поли-А-сайт или tk-поли-А-сайт, расположенные по ходу транскрипции полинуклеотида. В этом контексте приемлемыми экспрессионными векторами являются известные в данной области векторы, такие как вектор для экспрессии кДНК Окаяма-Берга (Okayama-Berg) pcDVl (фирма Pharmacia), pRc/CMV, рсДНК1, рсДНК3 (фирма In-vitrogene), pSPORTI (фирма GIBCO BRL), рХ (Pagano, Science 255, 1992, сс.1144-1147), дрожжевые двухгибридные векторы, такие как pEG202 и dpJG4-5 (Gyuris, Cell 75, 1995, сс.791-803), или прокариотические экспрессионные векторы, такие как лямбда gtll или pGEX (фирма Amersham-Pharmacia). Помимо молекул нуклеиновых кислот, предлагаемых в настоящем изобретении, вектор может содержать также нуклеотидные последовательности, кодирующие сигналы секреции. Такие последовательности хорошо известны специалисту в данной области. Кроме того, в зависимости от применяемой системы экспрессии к кодирующим последовательностям молекул нуклеиновых кислот, предлагаемых в изобретении, можно добавлять лидерные последовательности, которые обладают способностью обеспечивать направленный перенос пептидов, предлагаемых в изобретении, в клеточные компартменты, и они хорошо известны в данной области. Лидерную(ые) последовательность(и) объединяют во время соответствующей фазы с трансляционными, инициирующими и терминирующими последовательностями, и предпочтительно лидерная последовательность обладает способностью обеспечивать секрецию транслируемого протеина или образовавшегося из него протеина в периплазматическое пространство или во внеклеточную среду. Необязательно гетерологичная последовательность может кодировать слитый протеин, который включает С- или N-концевой служащий для идентификации пептид, придающий требуемые характеристики, например, стабилизацию или упрощение очистки экспрессируемого рекомбинантного продукта. После встраивания вектора в соответствующего хозяина, этого хозяина поддерживают в условиях, пригодных для достижения высокого уровня экспрессии нуклеотидных последовательностей, и затем при необходимости собирают и очищают молекулы антител или фрагменты, предлагаемые в изобретении. Таким образом, изобретение относится также к хозяевам/клеткам-хозяевам, которые содержат указанный вектор. Такие хозяева можно применять для получения антител/молекул антител, предлагаемых в изобретении, а также для медицинских/фармацевтических процедур. Указанные клетки-хозяева могут представлять собой трансдуцированные или трансфектированные нейроны, типа стволовых клеток нейронов, предпочтительно зрелых стволовых клеток нейронов. Такие клетки-хозяева можно применять для трансплантационной терапии.
Кроме того, вектор, предлагаемый в настоящем изобретении, может представлять собой также экспрессионный вектор, вектор для переноса гена или вектор для осуществления направленного переноса гена. Генная терапия, основой которой является интродукция терапевтических генов в клетку с помощью методов ex vivo или in vivo, является одним из наиболее важных путей применения переноса генов. Были созданы трансгенные мыши, экспрессирующие нейтрализующее антитело к фактору роста нерва, с помощью метода, основанного на применении «нейроантитела»; см. Capsoni, Proc. Natl. Acad. Sci. USA 97, 2000, сс.6826-6831 и Biocca, Embo J. 9, 1990, сс.101-108. Пригодные векторы, методы или системы для введения гена, применяемые в генной терапии in vitro или in vivo, описаны в литературе и известны специалисту в данной области; см., например, у Giordano, Nature Medicine 2, 1996, сс.534-539; Schaper, Circ. Res. 79, 1996, сс.911-919; Anderson, Science, 256, 1992, сс.808-813, Isner, Lancet, 348, 1996, сс.370-374; Muhlhauser, Circ. Res. 77, 1995, сс.1077-1086; Onodua, Blood, 91, 1998, сс.30-36; Verzeletti, Hum. Gene Ther., 9, 1998, сс.2243-2251; Verma, Nature, 389, 1997, сс.239-242; Anderson, Nature, 392 (Supp. 1998), сс.25-30; Wang, Gene Therapy, 4, 1997, сс.393-400; Wang, Nature Medicine, 2, 1996, сс.714-716; WO 94/29469; WO 97/00957; US 5580859; US 5589466; US 4394448 или у Schaper, Current Opinion in Biotechnology, 7, 1996, сс.635-640, и ссылки, процитированные в указанных публикациях. В частности, указанные векторы и/или системы для введения генов представлены также при описании методик, которые применяют для генной терапии пораженных неврологическим заболеванием тканей/клеток (см., среди прочего у Blomer, J. Virology, 71, 1997, сс.6641-6649) или гипоталамуса (см. среди прочего у Geddes, Front Neuroendocrinol, 20, 1999, сс.296-316 или Geddes, Nat. Med., 3, 1997, сс.1402-1404). Другие приемлемые конструкции, применяемые для генной терапии пораженных неврологическим заболеванием клеток /тканей, известны в данной области, например, описаны у Meier, J. Neuropathol. Exp. Neurol. 58, 1999, сс.1099-1110. Молекулы нуклеиновых кислот и векторы, предлагаемые в изобретении, можно создавать для непосредственной интродукции или для интродукции с помощью липосом, вирусных (например, аденовирусных, ретровирусных) векторов, электропорации, баллистических методов (например, генной пушки) или других систем введения в клетки. Кроме того, для молекул нуклеиновых кислот, предлагаемых в изобретении, можно использовать бакуловирусную систему в качестве эукариотической системы экспрессии. Интродукция и метод генной терапии должны предпочтительно приводить к экспрессии функционально активной молекулы антитела, предлагаемой в изобретении, при этом указанная экспрессируемая молекула антитела представляет собой молекулу, которая является наиболее предпочтительной для лечения, уменьшения интенсивности и/или предупреждения неврологических заболеваний, которые связаны с аномальным синтезом, сборкой и/или агрегацией амилоида, например, болезни Альцгеймера и т.п.
Таким образом, молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, и/или описанные выше векторы/хозяева, предлагаемые в настоящем изобретении, наиболее предпочтительно применять в виде фармацевтических композиций. Такие фармацевтические композиции можно применять в методах генной терапии. В этом контексте можно рассматривать вопрос о применении молекул нуклеиновых кислот и/или векторов, предлагаемых в настоящем изобретении, для модуляции, изменения и/или модификации (клеточной) экспрессии и/или концентрации молекул антител, предлагаемых в изобретении, или их фрагмента(ов).
Для применения в области генной терапии нуклеиновые кислоты, кодирующие пептид(ы), предлагаемый(ые) в изобретении, или их фрагменты можно клонировать в системе для введения генов, такой как вирус, и использовать вирус для заражения и уменьшения интенсивности симптомов заболевания или для лечения инфицированных клеток или организма.
Настоящее изобретение относится также к клетке-хозяину, трансфектированной или трансформированной вектором, предлагаемым в изобретении, или хозяину кроме человека, несущему вектор, предлагаемый в настоящем изобретении, т.е. к клетке-хозяину или хозяину, которые генетически модифицированы молекулой нуклеиновой кислоты, предлагаемой в изобретении, или вектором, который содержит такую молекулу нуклеиновой кислоты. Понятие «генетически модифицированый» означает, что клетка или хозяин несут помимо встречающегося в естественных условиях генома молекулу нуклеиновой кислоты или вектор, предлагаемые в изобретении, которые интродуцированы в клетку или хозяина или в одного из его предшественников/родителей. Молекула нуклеиновой кислоты или вектор могут присутствовать в генетически модифицированной клетке-хозяине или хозяине либо в виде независимой молекулы вне генома, предпочтительно в виде молекулы, которая обладает способностью к репликации, либо они стабильно интегрированы в геном клетки-хозяина или хозяина.
Клетка-хозяин, предлагаемая в настоящем изобретении, может представлять собой любую прокариотическую или эукариотическую клетку. Пригодными прокариотическими клетками являются клетки, которые обычно применяют для клонирования, типа Е. coli или Bacillus subtilis. A эукариотические клетки представляют собой, например, клетки грибов или животных. Примерами пригодных клеток грибов являются клетки дрожжей, предпочтительно дрожжей рода Saccharomyces и наиболее предпочтительно вида Saccharomyces cerevisiae. Приемлемыми клетками животных являются, например, клетки насекомых, клетки позвоночных, предпочтительно клетки млекопитающих, такие, например, как НЕК293, NSO, CHO, MDCK, U2-OSHela, NIH3T3, MOLT-4, Jurkat, PC-12, PC-3, IMR, NT2N, Sk-n-sh, CaSki, С33А. Эти клетки-хозяева, например, СНО-клетки, могут обеспечивать посттрансляционные модификации молекул антител, предлагаемых в изобретении, включая удаление лидерного пептида, укладку и сборку Н (тяжелой) и L (легкой) цепей, гликозилирование молекулы на правильных участках и секрецию функционально активной молекулы. Другие пригодные линии клеток, которые известны в данной области, получают из депозитариев клеточных линий, таких как Американская коллекция типовых культур (АТСС). Согласно настоящему изобретению также подразумевается, что в качестве клеток-хозяев могут функционировать первичные клетки/клеточные культуры. Такие клетки получают, в частности, из насекомых (например, насекомых р.р. Drosophila или Blatta) или млекопитающих (например, человека, свиней, мышей или крыс). Такие клетки-хозяева могут содержать клетки, полученные и/или выведенные из клеточных линий типа клеточной линии нейробластомы. Указанные выше первичные клетки хорошо известны в данной области и представляют собой среди прочего первичные астроциты, (смешанные) спинномозговые культуры или культуры клеток гиппокампа.
Согласно более предпочтительному варианту осуществления изобретения клетка-хозяин, трансформированная вектором, предлагаемым в изобретении, представляет собой нейрон, стволовую клетку нейрона (например, зрелую стволовую клетку нейрона), клетку головного мозга или клетку (линию), выведенную из вышеуказанных клеток. Кроме того, СНО-клетка, несущая молекулу нуклеиновой кислоты, предлагаемую в настоящем изобретении, может также представлять собой особый интерес в качестве хозяина. Такие клетки обеспечивают правильные вторичные модификации экспрессируемых молекул, т.е. молекул антител, предлагаемых в настоящем изобретении. Эти модификации включают среди прочего гликозилирование и фосфорилирование.
Хозяева могут представлять собой млекопитающих, кроме человека, наиболее предпочтительно мышей, крыс, овец, телят, собак, обезьян или человекообразных обезьян. Такие млекопитающие могут потребоваться для создания лекарственного средства, предпочтительно лекарственного средства для указанных неврологических и/или нейродегенеративных заболеваний. Кроме того, хозяева, предлагаемые в настоящем изобретении, наиболее предпочтительно можно применять для получения молекул антител (или их фрагментов), предлагаемых в изобретении. Предполагается, что указанные молекулы антител (или фрагментов) можно выделять из указанного хозяина. Также среди прочего указанные молекулы нуклеиновых кислот и/или векторы можно включать в последовательности для трансгенной экспрессии. Интродукцию молекул нуклеиновых кислот, предлагаемых в изобретении, в качестве трансгенов в хозяев кроме человека и их последующую экспрессию можно применять для получения антител, предлагаемых в изобретении. Например, экспрессия такого(их) трансгена(ов) в молоке трансгенного животного обеспечивает получение молекул антител, предлагаемых в изобретении, с количественным выходом; см. среди прочего US 5741957, US 5304489 или US 5849992. Ценные с этой точки зрения трансгены содержат молекулы нуклеиновых кислот, предлагаемые в изобретении, например, кодирующие последовательности легких и тяжелых цепей молекул антител, предлагаемых в изобретении, функционально связанные с промоторными и/или энхансерными структурами из гена, специфического для желез млекопитающих, типа казеина или бета-лактоглобулина.
Изобретение относится также к способу получения молекулы антитела, предлагаемой в изобретении, заключающемуся в том, что культивируют указанную выше клетку-хозяина в условиях, при которых происходит синтез указанной молекулы антитела, и выделяют молекулу антитела из культуры.
Изобретение относится также к композиции, содержащей молекулу антитела, предлагаемую в изобретении, или полученную с помощью описанного выше способа молекулу нуклеиновой кислоты, которая кодирует молекулу антитела, предлагаемую в изобретении, вектор, несущий указанную молекулу нуклеиновой кислоты или указанную выше клетку-хозяина и необязательно другие молекулы, либо индивидуально, либо в сочетании, например, молекулы, которые обладают способностью оказывать воздействие на формирование амилоидных бляшек, или которые обладают способностью осуществлять деполимеризацию уже сформировавшихся амилоидных бляшек. Понятие «композиция» в контексте настоящего описания относится к композиции, которая содержит по меньшей мере одно соединение, предлагаемое в изобретении. Предпочтительно такая композиция представляет собой фармацевтическую или диагностическую композицию.
Композиция может находиться в твердой или жидкой форме и среди прочего в форме порошка(ов), таблетки(ок), раствора(ов) или аэрозоля(ей). Такая композиция может содержать одно(у) или несколько антител/молекул антител, предлагаемых в изобретении, или молекулы нуклеиновых кислот, векторы или хозяева, предлагаемые в изобретении. Также предполагается, что указанная композиция содержит по меньшей мере две, предпочтительно три, более предпочтительно четыре, наиболее предпочтительно пять молекул антител, предлагаемых в изобретении, или молекулы(у) нуклеиновых(ой) кислот(ы), которые(ая) кодируют(ет) указанные(ую) молекулы(у) антител(а). Такая композиция может содержать также оптимизированные предлагаемые в изобретении антитела/молекулы антител, которые можно получать с помощью методов, описанных ниже и в прилагаемых примерах.
Предпочтительно, чтобы фармацевтическая композиция необязательно содержала фармацевтически приемлемый носитель и/или разбавитель. Указанную фармацевтическую композицию наиболее целесообразно применять для лечения неврологических и/или нейродегенеративных заболеваний. Такие заболевания включают (но, не ограничиваясь ими) болезнь Альцгеймера, амиотрофический боковой склероз (ALS), наследственное внутримозговое кровоизлияние с амилоидозом Дутча-типа, синдром Дауна, связанную с ВИЧ деменцию, болезнь Паркинсона и неврологические заболевания, связанные со старением. Фармацевтическая композиция, предлагаемая в изобретении, среди прочего может нести сильные ингибиторы образования амилоидных бляшек или сильный стимулятор деполимеризации амилоидных бляшек. Таким образом, настоящее изобретение относится к фармацевтическим композициям, которые содержат соединения, предлагаемые в изобретении, которые можно применять для лечения заболеваний/нарушений, связанных с патологическим протеолизом АРР и/или формированием амилоидных бляшек.
Примеры приемлемых фармацевтических носителей, эксципиентов и/или разбавителей хорошо известны в данной области и включают забуференные фосфатом физиологические растворы, воду, эмульсии, например, эмульсии типа масло/вода, различные типы смачивающих агентов, стерильные растворы и т.д. Композиции, содержащие такие носители, можно получать с помощью общепринятых хорошо известных методов. Такие фармацевтические композиции можно вводить пациенту в требуемой дозе. Введение приемлемых композиций можно осуществлять различными путями, например, посредством внутривенного, внутрибрюшинного, подкожного, внутримышечного, местного, внутрикожного, интраназального или внутрибронхиального введения. Наиболее предпочтительно обработку осуществляют путем инъекции и/или введения, например, в область артерии головного мозга или непосредственно в ткань головного мозга. Композиции, предлагаемые в изобретении, можно вводить также непосредственно в область-мишень, например, посредством биобаллистического введения снаружи или внутрь области-мишени, такой как головной мозг. Схему введения лекарственного средства может определять лечащий врач, и она зависит от клинических факторов. Как хорошо известно в медицинской практике, дозы для каждого пациента зависят от целого ряда факторов, включая размер пациента, площадь поверхности тела, возраст, конкретное применяемое соединение, пол, время и путь введения, общее состояние здоровья и одновременный прим других лекарственных средств. Однократная доза белковой фармацевтически активной субстанции может составлять от 1 нг до 10 мг/кг веса тела; однако можно применять также более низкие или более высокие дозы по сравнению с приведенными в качестве примера дозами, прежде всего с учетом вышеперечисленных факторов. Если схема представляет собой непрерывную инфузию, то дозы могут составлять также от 1 мкг до 10 мг стандартной дозы на кг веса тела в минуту.
Мониторинг процесса можно осуществлять путем периодической оценки. Композиции, предлагаемые в изобретении, можно наносить местно или вводить системно. Следует отметить, что периферически вводимые антитела могут проникать в центральную нервную систему, см. среди прочего у Bard, Nature Med. 6, 2000, сс.916-919. Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, в том числе физиологический раствор и забуференные среды. Носители для парентерального введения включают раствор хлорида натрия, декстрозу Рингена, декстрозу и хлорид натрия, раствор Рингера, дополненный лактатом, или масла с большим содержанием жира. Носители, предназначенные для внутривенного введения, включают жидкие и питательные добавки, электролитные добавки (например, основанные на декстрозе Рингера) и т.п. Могут присутствовать консерванты и другие добавки, такие, например, как противомикробные средства, антиоксиданты, хелатирующие агенты, инертные газы и т.п. Кроме того, фармацевтическая композиция, предлагаемая в изобретении, может содержать другие агенты, зависящие от предполагаемого применения фармацевтической композиции. Такие агенты могут представлять собой лекарственные средства, оказывающие воздействие на центральную нервную систему, например, нейрозащитные факторы, ингибиторы холинэстераз, агонисты мускаринового рецептора Ml, гормоны, антиоксиданты, ингибиторы воспаления и т.д. Особенно предпочтительно, когда фармацевтическая композиция содержит дополнительные агенты типа, например, нейромедиаторов и/или молекул, заменяющих нейромедиаторы, витамин Е или альфа-липоевую кислоту.
Фармацевтические композиции, а также способы, предлагаемые в изобретении, или описанное изобретение можно применять для лечения всех типов болезней, которые к настоящему времени еще неизвестны или которые связаны или зависят от патологической агрегации АРР или патологического процессинга АРР. Они могут оказаться особенно ценными для лечения болезни Альцгеймера и других заболеваний, в проявлении которых, по-видимому, играют роль внеклеточные отложения амилоида-β. Их предпочтительно применять для лечения человека, хотя под объем изобретения подпадает также лечение животных с использованием способов, путей применения и композиций, которые представлены в настоящем описании.
Предпочтительным вариантом осуществления изобретения является описанная выше композиция, предлагаемая в настоящем изобретении, которая представляет собой диагностическую композицию, которая необязательно дополнительно содержит приемлемые средства для обнаружения. Диагностическая композиция содержит по меньшей мере одно из вышеуказанных соединений, предлагаемых в изобретении.
Диагностическая композиция может содержать соединения, предлагаемые в изобретении, в частности и предпочтительно молекулы антител, предлагаемые в настоящем изобретении, в растворенной форме/жидкой фазе, но также возможно, чтобы эти соединения были связаны/присоединены и/или сшиты с твердой подложкой.
Твердые подложки можно применять в сочетании с указанной диагностической композицией или соединения, предлагаемые в настоящем изобретении, можно непосредственно связывать с твердыми подложками. Такие подложки хорошо известны в данной области и включают среди прочего поступающие в продажу материалы для заполнения колонок, гранулы из полистирола, гранулы из латекса, магнитные гранулы, коллоидные металлические частицы, стеклянные и/или силиконовые чипы и поверхности, нитроцеллюлозные полоски, мембраны, пластины, дурациты, лунки и лунки реакционных планшетов, пластиковые пробирки и т.д. Соединение(я), предлагаемое(ые) в изобретении, в частности, антитела, предлагаемые в настоящем изобретении, можно связывать с многочисленными различными носителями. Примеры хорошо известных носителей включают стекло, полистирол, поливинилхлорид, полипропилен, полиэтилен, поликарбонат, декстран, найлон, амилозы, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. В зависимости от целей изобретения носитель по своей природе может быть либо растворимым, либо нерастворимым. Соответствующие метки и методы их введения указаны выше и будут более подробно описаны ниже. Приемлемые методы фиксации/иммобилизации указанного(ых) соединения(й), предлагаемого(ых) в изобретении, хорошо известны и включают (но, не ограничиваясь ими) ионные, гидрофобные, ковалентные взаимодействия и т.п.
Наиболее предпочтительно диагностическую композицию, предлагаемую в изобретении, применять для обнаружения и/или количественной оценки АРР и/или продуктов процессинга АРР, типа амилоида-β, или для обнаружения и/или количественной оценки патологических и/или (генетически) модифицированных областей расщепления АРР.
Как проиллюстрировано в приведенных ниже примерах, соединения, предлагаемые в настоящем изобретении, в частности молекулы антител, предлагаемые в изобретении, являются особенно ценными в качестве диагностических реагентов для обнаружения истинных человеческих амилоидных бляшек в срезах головного мозга пациентов, страдающих болезнью Альцгеймера, путем непрямой иммунофлуоресценции.
Предпочтительно с целью обнаружения метить указанные соединения, предлагаемые в настоящем изобретении, которые входят в диагностическую композицию. Различные методы, которые можно использовать для мечения биологических молекул, известны специалисту в данной области и подпадают под объем настоящего изобретения. Такие методы описаны, например, у Tijssen в: «Practice и theory of enzyme immuno assays». Burden, RH и von Knippenburg (ред-ры), том 15 (1985), «Basic methods in molecular biology»; Davis LG, Dibmer MD; Battey Elsevier (1990), Mayer и др., (ред-ры) «Immunochemical methods in cell и molecular biology» Academic Press, London (1987) или в сериях «Methods in Enzymology», Academic Press, Inc.
Специалисту в данной области известны многочисленные разнообразные метки и методы введения меток. Примеры типов меток, которые можно применять согласно настоящему изобретению, включают ферменты, радиоизотопы, коллоидные металлы, флуоресцентные соединения, хемилюминесцентные соединения и биолюминесцентные соединения.
Обычно применяемые метки включают среди прочего флуорохромы (типа флуоресцеина, родамина, Texas Red и т.д.), ферменты (типа пероксидазы из хрена, β-галактозидазы, щелочной фосфатазы), радиоактивные изотопы (типа 32Р или 125I), биотин, дигоксигенин, коллоидные металлы, хеми- или биолюминесцентные соединения (типа диоксетанов, люминола или акридиниума). Технология введения метки, например, ковалентное связывание ферментов или биотинильных групп, иодирование, фосфорилирование, биотинилирование и т.д., хорошо известны в данной области.
Методы обнаружения включают (но, не ограничиваясь ими) авторадиографию, флуоресцентную микроскопию, прямые или косвенные ферментативные реакции и т.д. Общепринятые методы обнаружения включают радиоизотопные и нерадиоизотопные методы. Они представляют собой среди прочего Вестерн-блоттинг, анализы в поверхностном слое, РИА (радиоиммунный анализ) и ИРМА (иммунорадиометрический анализ), ФИА (иммуноферментный анализ), ELISA (твердофазный иммуноферментный анализ), ИФА (иммунофлуоресцентный анализ) и ХЛИА (иммунохемилюминесцентный анализ).
Кроме того, настоящее изобретение относится к применению молекулы антитела, предлагаемой в изобретении, или молекулы антитела, полученной с помощью способа, предлагаемого в изобретении, молекулы нуклеиновой кислоты, вектора или хозяина, предлагаемых в изобретении, для приготовления фармацевтической или диагностической композиции, которая предназначена для предупреждения и/или диагностики, и/или лечения заболевания, связанного с амилоидогенезом и/или образованием амилоидных бляшек. Кроме того, предпочтительным является применение указанных соединений, в частности, молекул антител, предлагаемых в изобретении, для предупреждения и/или лечения нейропатологических состояний, которые связаны с модифицированным или аномальным процессингом АРР и/или амилоидогенезом. Молекулы антител, например, в форме (сконструированных) иммуноглобулинов, типа антител, несущих каркасный участок IgG, в частности IgG-1, или в форме химерных антител, биспецифических антител, одноцепочечных Fv (scFv) или биспецифических scFv и т.п., можно применять для приготовления представленных в описании фармацевтических композиций. Кроме того, молекулы антител можно применять в диагностических наборах, описанных ниже в примерах, поскольку молекулы антител, предлагаемые в изобретении, специфически взаимодействуют/выявляют Аβ4 и/или амилоидные отложения/бляшки.
Таким образом, изобретение относится к применению соединений, предлагаемых в настоящем изобретении, для приготовления фармацевтической композиции, которая предназначена для облегчения состояния, например, путем разрушения β-амилоидных бляшек или путем пассивной иммунизации, направленной против образования β-амилоидных бляшек. Как проиллюстрировано ниже в примерах, молекулы антител, предлагаемые в изобретении, особенно предпочтительно применять для предупреждения Аβ-агрегации и для деполимеризации уже образовавшихся амилоидных агрегатов. Таким образом, антитела, предлагаемые в изобретении, можно применять уменьшения патологических амилоидных отложений/бляшек, для клиренса амилоидных бляшек/предшественников бляшек, а также для защиты нервной ткани. В частности молекулы антител, предлагаемые в изобретении, можно применять для предупреждения образования амилоидных бляшек in vivo, а также для клиренса in vivo уже существующих амилоидных отложений/бляшек. Кроме того, молекулы антител, предлагаемые в изобретении, можно применять для пассивной иммунизации против Аβ4. Клиренс Аβ4/отложений Аβ4 можно среди прочего осуществлять путем медицинского применения антител, предлагаемых в изобретении, которые несут Fc-фрагмент. Этот Fc-фрагмент антитела может оказаться особенно полезным при иммунных ответах, опосредуемых Fc-рецептором, таких как притяжение макрофагов (фагоцитарных клеток и/или микроглии) и/или клеток-хелперов. Для опосредования связанных с Fc-фрагментом иммунных ответов молекула антитела, предлагаемая в изобретении, предпочтительно должна нести каркасный участок (человеческого) IgG-1. Как будет обсуждаться ниже, предпочтительно пациент, которого следует лечить с помощью молекул антител, предлагаемых в изобретении, молекул нуклеиновых кислот, кодирующих указанные антитела или их фрагменты, векторов, предлагаемых в изобретении, или клеток-хозяев, предлагаемых в изобретении, представляет собой человека. Под объем изобретения подпадают также другие каркасные участки, типа IgG2a- или IgG2b-каркасные участки молекул антител, предлагаемых в изобретении. Каркасные участки иммуноглобулинов в формате IgG2a и IgG2b особенно предпочтительны для применения в опытах с использованием мышей, например, для применения молекул антител, предлагаемых в изобретении, в научных целях, например, для опытов на трансгенных мышах, которые экспрессируют (человеческие) АРР или АРР-фрагменты дикого типа и/или Аβ4.
Перечисленные выше заболевания, связанные с амилоидогенезом и/или образованием амилоидных бляшек, включают (но, не ограничиваясь ими) деменцию, болезнь Альцгеймера, моторную невропатию, болезнь Паркинсона, ALS (амиотрофический боковой склероз), скрепи (почесуха), связанную с ВИЧ деменцию, а также болезнь Крейтцфельда-Якоба, наследственное внутримозговое кровоизлияние с амилоидозом Дутча-типа, синдром Дауна и неврологические заболевания, связанные со старением. Молекулы антител, предлагаемые в изобретении, и композиции, представленные в описании, можно применять также для облегчения и/или предупреждения воспалительных процессов, связанных с амилоидогенезом и/или образованием амилоидных бляшек.
Таким образом, настоящее изобретение относится также к способу лечения, предупреждения и/или замедления развития неврологических и/или нейродегенеративных заболеваний, заключающемуся в том, что пациенту, который страдает указанным неврологическим и/или нейродегенеративным заболеванием, и/или пациенту, предрасположенному к указанному неврологическому и/или нейродегенеративному заболеванию, вводят эффективное количество молекулы антитела, предлагаемой в изобретении, молекулы нуклеиновой кислоты, предлагаемой в изобретении, и/или описанной выше композиции.
Еще одним вариантом осуществления настоящего изобретения является набор, содержащий по меньшей мере одну молекулу антитела, по меньшей мере одну молекулу нуклеиновой кислоты, по меньшей мере один вектор или по меньшей мере одну клетку-хозяина, предлагаемую в изобретении. Предпочтительно набор, предлагаемый в настоящем изобретении, необязательно содержит также буфер(ы), заранее приготовленные растворы и/или остальные реагенты или материалы, необходимые для осуществления медицинских, научных или диагностических анализов и других целей. Кроме того, компоненты набора, предлагаемого в изобретении, можно упаковывать индивидуально в пузырьки или флаконы, или они могут находиться в виде комбинации в контейнерах или наборах, состоящих из нескольких контейнеров.
Набор, предлагаемый в настоящем изобретении, целесообразно использовать, среди прочего, для осуществления способа, предлагаемого в изобретении, и его можно применять для различных указанных в описании целей, например, в качестве диагностического набора, средств для исследования или медицинских средств. Кроме того, набор, предлагаемый в изобретении, может содержать средства для обнаружения, пригодные для научных, медицинских и/или диагностических целей. Производство наборов осуществляют предпочтительно с помощью стандартных процедур, известных специалисту в данной области.
Изобретение относится также к способу оптимизации молекулы антитела, указанной в описании, заключающемуся в том, что
(а) конструируют библиотеку разнообразных Fab-фрагментов антитела, полученных из антитела, которое несет по меньшей мере один CDR3 VH-области, кодируемый молекулой нуклеиновой кислоты, которая представлена в SEQ ID NO:21, 23 или 25, или меньшей мере одну аминокислотную последовательность CDR3 VH-области, представленную в SEQ ID NO:22, 24 или 26;
(б) тестируют полученную оптимизированную Fab-библиотеку с помощью пэннинга через Аβ/Аβ4;
(в) идентифицируют оптимизированные клоны и
(г) экспрессируют отобранные оптимизированные клоны.
Оптимизация антител/молекул антител, предлагаемых в изобретении, описана также ниже в примерах и может включать отбор, например, по признаку более высокой аффинности к одной или обеим указанным областям/эпитопам β-А4, или отбор по признаку повышенной экспрессии и т.п. Согласно одному из вариантов осуществления изобретения отбор по признаку более высокой аффинности к одной или обеим областям/эпитопам β-А4 предусматривает отбор по признаку высокой аффинности к (а) аминокислотной последовательности β-А4, содержащей аминокислоты 2-10 (или ее фрагменту(ам)), и/или (б) аминокислотной последовательности (3-А4, содержащей аминокислоты 12-25 (или ее фрагменту(ам)) (SEQ ID NO:27).
Специалист в данной области легко может осуществлять способ, предлагаемый в изобретении, основываясь на данных, представленных в настоящем изобретении. Протоколы оптимизации антител известны в данной области. Эти протоколы оптимизации предусматривают среди прочего мутагенез посредством «прогулки» по CDR, который представлен в настоящем описании и описан у Yang, J. Mol. Biol. 25, 1995, сс.392-403; Schier, J. Mol. Biol. 263, 1996, сс.551-567; Barbas, Trends. Biotech 14, 1996, сс.230-34 или у Wu PNAS 95, 1998, сс.6037-6042; Schier, Human Antibody Hybridomas 7, 1996, с. 97; Moore, J. Mol. Biol. 272, 1997, с. 336. Методики «пэннинга» также известны в данной области, см., например, у Кау, Gene 128, 1993 сс.59-65. Кроме того, в таких публикациях как Borrebaeck (1995), «Antibody Engineering», изд-во Oxford University, cc.229-266; McCafferty (1996), «Antibody Engineering», изд-во Oxford University Press;
Kay (1996), A Laboratory Manual, изд-во Academic Press, описаны протоколы оптимизации, которые можно модифицировать в зависимости от целей настоящего изобретения.
Способ оптимизации может дополнительно включать стадию (ва), заключающуюся в том, что оптимизированные клоны дополнительно оптимизируют с помощью кассеты для мутагенеза, как это описано ниже в примерах.
Способ оптимизации молекулы антитела, представленной в описании, дополнительно проиллюстрирован ниже в примерах в виде способа созревания аффинности родительских антител /молекул антител, которые обладают способностью специфически распознавать две области бета-А4 пептида/Абета4/Аβ/Аβ4/βА4.
Предпочтительно указанный на стадии (б) описанного выше способа Аβ/Аβ4 (в контексте настоящего изобретения также обозначенный как βА4) представляет собой агрегированный Аβ/Аβ4. Пэннинг можно осуществлять (как описано ниже в примерах) с использованием повышенной силы связывания. Силу можно повышать, например, путем снижения концентрации Аβ/Аβ4 или путем повышения температуры (анализа). Тестирование оптимизированной библиотеки с помощью пэннинга известно специалисту в данной области и описано у Кау (1993), loc. cit. Предпочтительно идентификацию на стадии (в) осуществляют путем упорядочивания по самым низким значениям KD.
Наиболее предпочтительно идентификацию на стадии (в) осуществляют с помощью koff упорядочивания. Метод koff упорядочивания известен специалистам в данной области и описан у Schier (1996), loc. cit.; Schier, J. Mol. Biol. 255, 1996, сс.28-43 или у Duenas, Mol. Immunol. 33, 1996, сс.279-286. Метод koff упорядочивания дополнительно проиллюстрирован ниже в примерах. Константу off-скорости (koff) можно измерять согласно методу, описанному ниже в примерах.
Как указывалось ранее, выявленные клоны можно экспрессировать для дополнительной оценки. Экспрессию можно осуществлять известными методами, которые среди прочего проиллюстрированы ниже в примерах. С помощью экспрессии можно получать среди прочего экспрессированные Fab-фрагменты, scFv, биспецифические иммуноглобулины, биспецифические молекулы антител. Fab- и/или Fv-слитые протеины или полноразмерные антитела, типа IgG, в частности IgGl.
Оптимизированные антитела, в частности, оптимизированные Fab-фрагменты или оптимизированные IgG, предпочтительно IgGl, можно оценивать с помощью методов, проиллюстрированных ниже в примерах. Такие методы включают (но не ограничиваясь ими) оценку аффинности связывания, определение значений КD, pepspot-анализ, ELISA-анализы, РИА-анализы, ХЛИА-анализы, (иммуно-)гистологические исследования (например, окрашивание амилоидных бляшек), анализы деполимеризации или антителозависимого фагоцитоза β-А4.
Следующим вариантом осуществления настоящего изобретения является способ, заключающийся в том, что оптимизированные антитела получают с помощью перекрестного клонирования. Этот метод проиллюстрирован также ниже в примерах и включает стадию, заключающуюся в том, что объединяют независимо оптимизированные CDR-участки, например, путем объединения независимо оптимизированных H-CDR2 и L-CDR2 из зрелых клонов с H-CDR3, предпочтительно с этим же H-CDR3.
Одним из предпочтительных вариантов осуществления изобретения является способ приготовления фармацевтической композиции, заключающийся в том, что
(а) осуществляют оптимизацию антитела с помощью метода, который представлен в описании и проиллюстрирован ниже в примерах; и (в) объединяют оптимизированное(ую) антитело/молекулу антитела с физиологически приемлемым носителем согласно описанному выше методу.
Таким образом, изобретение относится также к фармацевтической композиции, приготовленной с помощью указанного метода и содержащей дополнительно оптимизированные молекулы антител, которые обладают способностью специфически распознавать две области пептида бета-А4/Абета4Аβ/Аβ4/βА4, как они описаны выше. Примеры указанных последовательностей:
SEQ ID NO:1
AEFRHDSGY
Первая область пептида β-А4, «N-концевая(ой) область/эпитоп»
SEQ ID NO:2
VHHQKLVFFAEDVG
Вторая область пептида β-А4, «центральная(ый)/срединная(ый) область/эпитоп»
SEQ ID NO:3
VH-область MS-Roche №3 (нуклеотидная последовательность)
CAGGTGCAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGC
CTGCGTCTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGA
GCTGGGTGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGCGATTA
GCGGTAGCGGCGGCAGCACCTATTATGCGGATAGCGTGAAAGGCCGTTTTAC
CATTTCACGTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTG
CGTGCGGAAGATACGGCCGTGTATTATTGCGCGCGTCTTACTCATTATGCTC
GTTATTATCGTTATTTTGATGTTTGGGGCCAAGGCACCCTGGTGACGGTTAGC
TCAGC (SEQ ID NO:3)
SEQ ID NO:4
VH-область MS-Roche №3 (аминокислотная последовательность)
QVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISG
SGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLTHYARYY
RYFDVWGQGTLVTVSS (SEQ ID NO:4)
SEQ ID NO:5
VH-область MS-Roche №7 (нуклеотидная последовательность)
CAGGTGCAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGC
CTGCGTCTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGA
GCTGGGTGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGCGATTA
GCGGTAGCGGCGGCAGCACCTATTATGCGGATAGCGTGAAAGGCCGTTTACC
ATTTCACGTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTGC
GTGCGGAAGATACGGCCGTGTATTATTGCGCGCGTGGTAAGGGTAATACTCA
TAAGCCTTATGGTTATGTTCGTTATTTTGATGTTTGGGGCCAAGGCACCCTGG
TGACGGTTAGCTCAGC (SEQ ID NO:5)
SEQ ID NO:6
VH-область MS-Roche №7 (аминокислотная последовательность)
QVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISG
SGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGKGNTHKP
YGYVRYFDVWGQGTLVTVSS (SEQ ID NO:6)
SEQ ID NO:7
VH-область MS-Roche №8 (нуклеотидная последовательность)
CAGGTGCAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGC
CTGCGTCTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGA
GCTGGGTGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGCGATTA
GCGGTAGCGGCGGCAGCACCTATTATGCGGATAGCGTGAAAGGCCGTTTTAC
CATTTCACGTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTG
CGTGCGGAAGATACGGCCGTGTATTATTGCGCGCGTCTTCTTTCTCGTGGTTA TAATGGTTATTATCATAAGTTTGATGTTTGGGGCCAAGGCACCCTGGTGACG
GTTAGCTCAGC (SEQ ID NO:7)
SEQ ID NO:8
VH-область MS-Roche №8 (аминокислотная последовательность)
QVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISG
SGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLLSRGYNG
YYHKFDVWGQGTLVTVSS (SEQ ID NO:8)
SEQ ID NO:9
VL-область MS-Roche №3 (нуклеотидная последовательность)
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGAGCGTGAGCAGCAGCTATCTGG
CGTGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTATGGCGC
GAGCAGCCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGC
ACGGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGGTTT
ATTATTGCCAGCAGGTTTATAATCCTCCTGTTACCTTTGGCCAGGGTACGAAA
GTTGAAATTAAACGTACG (SEQ ID NO:9)
SEQ ID NO:10
VL-область MS-Roche №3 (аминокислотная последовательность)
DIVLTQSPATLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSR
ATGVPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQVYNPPVTFGQGTKVEIKRT
(SEQ ID NO:10)
SEQ ID NO:11
VL-область MS-Roche №7 (нуклеотидная последовательность)
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGAGCGTGAGCAGCAGCTATCTGG
CGTGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTATGGCGC
GAGCAGCCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGC
ACGGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGACTT
ATTATTGCTTTCAGCTTTATTCTGATCCTTTTACCTTTGGCCAGGGTACGAAA
GTTGAAATTAAACGTACG (SEQ ID NO.11)
SEQ ID NO:12
VL-область MS-Roche №7 (аминокислотная последовательность)
DIVLTQSPATLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSR
ATGVPARFSGSGSGTDFTLTISSLEPEDFATYYCFQLYSDPFTFGQGTKVEIKRT
(SEQ ID NO:12)
SEQ ID NO:13
VL-область MS-Roche №8 (нуклеотидная последовательность)
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGAGCGTGAGCAGCAGCTATCTGG
CGTGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTATGGCGC
GAGCAGCCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGC
ACGGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGACTT
ATTATTGCCAGCAGCTTTCTTCTTTTCCTCCTACCTTTGGCCAGGGTACGAAA
GTTGAAATTAAACGTACG (SEQ ID NO:13)
SEQ ID NO:14
VL-область MS-Roche №8 (аминокислотная последовательность)
DIVLTQSPATLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSR
ATGVPARFSGSGSGTDFTLTISSLEPEDFATYYCQQLSSFPPTFGQGTKVEIKRT
(SEQ ID NO:14)
SEQIDNO:15
CDR3 VL-области MSR-3 (нуклеотидная последовательность)
CAGCAGGTTTATAATCCTCCTGTT
(SEQ ID NO:15)
SEQIDNO:16
CDR3 VL-области MSR-3 (аминокислотная последовательность)
QQVYNPPV (SEQ ID NO:16)
SEQIDNO:17
CDR3 VL-области MSR-7 (нуклеотидная последовательность)
TTTCAGCTTTATTCTGATCCTTTT
(SEQ ID NO:17)
SEQ ID NO:18
CDR3 VL-области MSR-7 (аминокислотная последовательность)
FQLYSDPF (SEQ ID NO.18)
SEQ ID NO:19
CDR3 VL-области of MSR-8 (нуклеотидная последовательность)
CAGCAGCTTTCTTCTTTT CCT CCT
(SEQ ID NO.19)
SEQ ID NO:20
CDR3 VL-области MSR-8 (аминокислотная последовательность)
QQLSSFPP (SEQ ID NO:20)
SEQ ID NO:21
CDR VH-области MSR-3 (нуклеотидная последовательность)
CTTACTCATTATGCTGCTTATTATCGTTATTTTGATGTT
(SEQ ID NO:21)
SEQ ID NO:22
CDR VH-области MSR-3 (аминокислотная последовательность)
LTHYARYYRYFDV (SEQ ID NO:22)
SEQ ID NO:23
CDR VH-области MSR-7 (нуклеотидная последовательность)
GGTAAGGGTAATACTCATAAGCCTTATGGTTATGTTCGTTATTTTGATGTT
(SEQ ID NO:23)
SEQ ID NO:24
CDR VH-области MSR-7 (аминокислотная последовательность)
GKGNTHKPYGYVRYFDV (SEQ ID NO:24)
SEQ ID NO:25
CDR VH-области MSR-8 (нуклеотидная последовательность)
CTTCTTTCTCGTGGTTATAATGGTTATTATCATAAGTTTGATGTT
(SEQ ID NO.25)
SEQ ID NO:26
CDR VH-области MSR-8 (аминокислотная последовательность)
LLSRGYNGYYHKFDV (SEQ ID NO:26)
SEQ ID NO:27
Aβ4 (аминокислоты 1-42)
DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVVIA (SEQ ID NO:27)
SEQ ID NO:28 праймер
5'-GTGGTGGTTCCGATATC-3' (SEQ ID NO:28)
SEQ ID NO:29 праймер
5'-AGCGTCACACTCGGTGCGGCTTTCGGCTGGCCAAGAACGGTTA-3' (SEQ ID NO:29)
SEQ ID NO:30 праймер
5'-CAGGAAACAGCTATGAC-3' (SEQ ID NO:30)
SEQ ID NO:31 праймер
5'-TACCGTTGCTCTTCACCCC-3' (SEQ ID NO:31)
SEQ ID NO:32
VH MS-Roche №3.6Н5×3.6L2; ДНК; искусственная последовательность
CAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGCCTGCGT
CTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGAGCTGGG
TGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGCTATTTCTGAGTC
TGGTAAGACTAAGTATTATGCTGATTCTGTTAAGGGTCGTTTTACCATTTCAC
GTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTGCGTGCGGA
AGATACGGCCGTGTATTATTGCGCGCGTCTTACTCATTATGCTCGTTATTATC
GTTATTTTGATGTTTGGGGCCAAGGCACCCTGGTGACGGTTAGCTCA (SEQ ID NO:32)
SEQ ID NO.33
протеин VH-области MS-Roche №3.6Н5×3.6L2; протеин/1; искусственная последовательность
QLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISESG
KTKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLTHYARYYRY
FDVWGQGTLVTVSS (SEQ ID NO:33)
SEQ ID NO:34
VH-область MS-Roche №3.6Н8×3.6L2; ДНК; искусственная последовательность
CAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGCCTGCGT
CTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGAGCTGGG
TGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGCTATTTCTGAGTA
TTCTAAGTTTAAGTATTATGCTGATTCTGTTAAGGGTCGTTTTACCATTTCAC
GTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTGCGTGCGGA
AGATACGGCCGTGTATTATTGCGCGCGTCTTACTCATTATGCTCGTTATTATC
GTTATTTTGATGTTTGGGGCCAAGGCACCCTGGTGACGGTTAGCTCA (SEQ ID NO:34)
SEQ ID NO:35
протеин VH-области MS-Roche №3.6Н8×3.6L2; протеин/1; искусственная последовательность
QLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISEYS
KFKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARLTHYARYYRY
FDVWGQGTLVTVSS (SEQ ID NO:35)
SEQ ID NO:36
VH-область MS-Roche №7.4Н2×7.2L1; ДНК; искусственная последовательность
CAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGCCTGCGT
CTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGAGCTGGG
TGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGCTATTAATTATAA
TGGTGCTCGTATTTATTATGCTGATTCTGTTAAGGGTCGTTTTACCATTTCAC
GTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTGCGTGCGGA
AGATACGGCCGTGTATTATTGCGCGCGTGGTAAGGGTAATACTCATAAGCCT
TATGGTTATGTTCGTTATTTTGATGTTTGGGGCCAAGGCACCCTGGTGACGGT
TAGCTCA (SEQ ID NO:36)
SEQ ID NO:37
протеин VH-области MS-Roche №7.4Н2×7.2L1; протеин/1; искусственная последовательность
QLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAINYNG
ARIYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGKGNTHKPYG
YVRYFDVWGQGTLVTVSS (SEQ ID NO:37)
SEQ ID NO:38
VH-область MS-Roche №7.9Н2×7.12 L2; ДНК; искусственная последовательность
CAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGCCTGCGT
CTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGAGCTGGG
TGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGCTATTAATGCTGA
TGGTAATCGTAAGTATTATGCTGATTCTGTTAAGGGTCGTTTTACCATTTCAC
GTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTGCGTGCGGA
AGATACGGCCGTGTATTATTGCGCGCGTGGTAAGGGTAATACTCATAAGCCT
TATGGTTATGTTCGTTATTTTGATGTTTGGGGCCAAGGCACCCTGGTGACGGT
TAGCTCA (SEQ ID NO:38)
SEQ ID NO:39
протеин VH-области MS-Roche №7.9Н2×7.12 L2; протеин/1; искусственная последовательность
QLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAINADG
NRKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGKGNTHKPY
GYVRYFDVWGQGTLVTVSS (SEQ ID NO:39)
SEQ ID NO:40
VH-область MS-Roche №7.9Н4×7.12L2; ДНК; искусственная последовательность
CAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGCCTGCGT
CTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGAGCTGGG
TGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGA
GCGCTATTAATGCTGT
TGGTATGAAGAAGTTTTATGCTGATTCTGTTAAGGGTCGTTTTACCATTTCAC
GTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTGCGTGCGGA
AGATACGGCCGTGTATTATTGCGCGCGTGGTAAGGGTAATACTCATAAGCCT
TATGGTTATGTTCGTTATTTTGATGTTTGGGGCCAAGGCACCCTGGTGACGGT
TAGCTCA (SEQ ID NO:40)
SEQ ID NO:41
протеин VH-области MS-Roche №7.9Н4×7.12L2; протеин/1; искусственная последовательность
QLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAINAVG
MKKFYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGKGNTHKPY
GYVRYFDVWGQGTLVTVSS (SEQ ID NO:41)
SEQ ID NO:42
VH-область MS-Roche №7.11H1×7.11L1; ДНК; искусственная последовательность
CAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGCCTGCGT
CTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGAGCTGGG
TGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGGTATTAATGCTGC
TGGTTTTCGTACTTATTATGCTGATTCTGTTAAGGGTCGTTTTACCATTTCAC
GTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTGCGTGCGGA
AGATACGGCCGTGTATTATTGCGCGCGTGGTAAGGGTAATACTCATAAGCCT
TATGGTTATGTTCGTTATTTTGATGTTTGGGGCCAAGGCACCCTGGTGACGGT
TAGCTCA (SEQ ID NO:42)
SEQ ID NO.43
протеин VH-области MS-Roche №7.11Н1×7.11L1; протеин/1; искусственная последовательность
QLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSGINAAG
FRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGKGNTHKPYG
YVRYFDVWGQGTLVTVSS (SEQ ID NO:43)
SEQ ID NO:44
VH-область MS-Roche №7.11H1×7.2L1; ДНК; искусственная последовательность
CAATTGGTGGAAAGCGGCGGCGGCCTGGTGCAACCGGGCGGCAGCCTGCGT
CTGAGCTGCGCGGCCTCCGGATTTACCTTTAGCAGCTATGCGATGAGCTGGG
TGCGCCAAGCCCCTGGGAAGGGTCTCGAGTGGGTGAGCGGTATTAATGCTGC
TGGTTTTCGTACTTATTATGCTGATTCTGTTAAGGGTCGTTTTACCATTTCAC
GTGATAATTCGAAAAACACCCTGTATCTGCAAATGAACAGCCTGCGTGCGGA
AGATACGGCCGTGTATTATTGCGCGCGTGGTAAGGGTAATACTCATAAGCCT
TATGGTTATGTTCGTTATTTTGATGTTTGGGGCCAAGGCACCCTGGTGACGGT
TAGCTCA (SEQ ID NO:44)
SEQ ID NO:45
протеин VH-области MS-Roche №7.11H1×7.2L1; протеин/1; искусственная последовательность
QLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSGINAAG
FRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGKGNTHKPYG
YVRYFDVWGQGTLVTVSS (SEQ ID NO:45)
SEQ ID NO:46 VL-область MS-Roche №3.6Н5×3.6L2; ДНК; искусственная последовательность
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGTTTCTTTCTCGTTATTATCTGGCG
TGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTATGGCGCGA
GCAGCCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGCAC
GGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGGTTTATT
ATTGCCAGCAGACTTATAATTATCCTCCTACCTTTGGCCAGGGTACGAAAGT
TGAAATTAAACGTACG (SEQ ID NO:46)
SEQ ID NO:47
протеин VL-области MS-Roche №3.6Н5×3.6L2; протеин/1; искусственная последовательность
DIVLTQSPATLSLSPGERATLSCRASQFLSRYYLAWYQQKPGQAPRLLIYGASSR
ATGVPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQTYNYPPTFGQGTKVEIKRT
(SEQ ID NO:47)
SEQ ID NO:48
VL-область MS-Roche №3.6Н8×3.6L2; ДНК; искусственная последовательность
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGTTTCTTTCTCGTTATTATCTGGCG
TGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTATGGCGCGA
GCAGCCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGCAC
GGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGGTTTATT
ATTGCCAGCAGACTTATAATTATCCTCCTACCTTTGGCCAGGGTACGAAAGT
TGAAATTAAACGTACG (SEQ ID NO:48)
SEQ ID NO:49
протеин VL-области MS-Roche №3.6Н8×3.6L2; протеин/1; искусственная последовательность
DIVLTQSPATLSLSPGERATLSCRASQFLSRYYLAWYQQKPGQAPRLLIYGASSR
ATGVPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQTYNYPPTFGQGTKVEIKRT
(SEQ ID NO:49)
SEQ ID NO:50
VL-область MS-Roche №7.4Н2×7.2L1; ДНК; искусственная последовательность
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGTATGTTGATCGTACTTATCTGGC
GTGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTATGGCGCG
AGCAGCCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGCA
CGGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGACTTA
TTATTGCCAGCAGATTTATTCTTTTCCTCATACCTTTGGCCAGGGTACGAAAG
TTGAAATTAAACGTACG (SEQ ID NO:50)
SEQ ID NO:51
протеин VL-области MS-Roche №7.4Н2×7.2L1; протеин/1; искусственная последовательность
DIVLTQSPATLSLSPGERATLSCRASQYVDRTYLAWYQQKPGQAPRLLIYGASSR
ATGVPARFSGSGSGTDFTLTISSLEPEDFATYYCQQIYSFPHTFGQGTKVEIKRT
(SEQ ID NO:51)
SEQ ID NO:52
VL-область MS-Roche №7.9Н2×7.12 L2; ДНК; искусственная последовательность
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGCGTTTTTTTTATAAGTATCTGGC
GTGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTCTGGTTCT
TCTAACCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGCA
CGGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGGTTTA
TTATTGCCTTCAGCTTTATAATATTCCTAATACCTTTGGCCAGGGTACGAAAG
TTGAAATTAAACGTACG (SEQ ID NO:52)
SEQ ID NO:53
протеин VL-области MS-Roche №7.9Н2×7.12 L2; протеин/1; искусственная последовательность
DIVLTQSPATLSLSPGERATLSCRASQRFFYKYLAWYQQKPGQAPRLLISGSSNR
ATGVPARFSGSGSGTDFTLTISSLEPEDFAVYYCLQLYNIPNTFGQGTKVEIKRT
(SEQ ID NO:53)
SEQ ID NO:54
VL-область MS-Roche №7.9Н4×7.12L2; ДНК; искусственная последовательность
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGCGTTTTTTTTATAAGTATCTGGC
GTGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTCTGGTTCT
TCTAACCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGCA
CGGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGGTTTA
TTATTGCCTTCAGCTTTATAATATTCCTAATACCTTTGGCCAGGGTACGAAAG
TTGAAATTAAACGTACG (SEQ ID NO:54)
SEQ ID NO:55
протеин VL-области MS-Roche №7.9Н4×7.12L2; протеин/1; искусственная последовательность
DIVLTQSPATLSLSPGERATLSCRASQRFFYKYLAWYQQKPGQAPRLLISGSSNR
ATGVPARFSGSGSGTDFTLTISSLEPEDFAVYYCLQLYNIPNTFGQGTKVEIKRT
(SEQ ID NO:55)
SEQ ID NO:56
VL-область MS-Roche №7.11H1×7.11L1; ДНК; искусственная
последовательность
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGCGTATTCTTCGTATTTATCTGGC
GTGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTATGGCGCG
AGCAGCCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGCA
CGGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGACTTA
TTATTGCCAGCAGGTTTATTCTCCTCCTCATACCTTTGGCCAGGGTACGAAAG
TTGAAATTAAACGTACG (SEQ ID NO:56)
SEQ ID NO:57
протеин VL-области MS-Roche №7.11H1×7.11L1; протеин/1; искусственная последовательность
DIVLTQSPATLSLSPGERATLSCRASQRILRIYLAWYQQKPGQAPRLLIYGASSRA
TGVPARFSGSGSGTDFTLTISSLEPEDFATYYCQQVYSPPHTFGQGTKVEIKRT (SEQ ID NO:57)
SEQ ID NO:58
VL-область MS-Roche №7.11H1×7.2L1; ДНК; искусственная последовательность
GATATCGTGCTGACCCAGAGCCCGGCGACCCTGAGCCTGTCTCCGGGCGAAC
GTGCGACCCTGAGCTGCAGAGCGAGCCAGTATGTTGATCGTACTTATCTGGC
GTGGTACCAGCAGAAACCAGGTCAAGCACCGCGTCTATTAATTTATGGCGCG
AGCAGCCGTGCAACTGGGGTCCCGGCGCGTTTTAGCGGCTCTGGATCCGGCA
CGGATTTTACCCTGACCATTAGCAGCCTGGAACCTGAAGACTTTGCGACTTA
TTATTGCCAGCAGATTTATTCTTTTCCTCATACCTTTGGCCAGGGTACGAAAG
TTGAAATTAAACGTACG (SEQ ID NO:58)
SEQ ID NO:59
протеин VL-области MS-Roche №7.11H1×7.2L1; протеин/1; искусственная последовательность
DIVLTQSPATLSLSPGERATLSCRASQYVDRTYLAWYQQKPGQAPRLLIYGASSR
ATGVPARFSGSGSGTDFTLTISSLEPEDFATYYCQQIYSFPHTFGQGTKVEIKRT
(SEQ ID NO:59)
SEQ ID NO:60
HCDR3-область MS-Roche №3.6Н5×3.6L2; ДНК; искусственная последовательность
CTTACTCATTATGCTCGTTATTATCGTTATTTTGATGTT (SEQ ID NO:60)
SEQ ID NO:61
протеин HCDR3-oбласти MS-Roche №3.6Н5×3.6L2; протеин/1; искусственная последовательность
LTHYARYYRYFDV (SEQ ID NO:61)
SEQ ID NO:62
НСDR3-область MS-Roche №3.6Н8×3.6L2; ДНК; искусственная последовательность
CTTACTCATTATGCTCGTTATTATCGTTATTTTGATGTT (SEQ ID NO:62)
SEQ ID NO:63
протеин HCDR3-oбласти MS-Roche №3.6Н8×3.6L2; протеин/1; искусственная последовательность
LTHYARYYRYFDV (SEQ ID NO:63)
SEQ ID NO:64
HCDR3-область MS-Roche №7.4Н2×7.2L1; ДНК; искусственная последовательность
GGTAAGGGTAATACTCATAAGCCTTATGGTTATGTTCGTTATTTTGATGTT
(SEQ ID NO:64)
SEQ ID NO:65
протеин HCDR3-oбласти MS-Roche №7.4Н2×7.2L1; протеин/1; искусственная последовательность
GKGNTHKPYGYVRYFDV (SEQ ID NO:65)
SEQ ID NO:66
HCDR3-область MS-Roche №7.9Н2×7.12L2; ДНК; искусственная последовательность
GGTAAGGGTAATACTCATAAGCCTTATGGTTATGTTCGTTATTTTGATGTT
(SEQ ID NO:66)
SEQ ID NO:67
протеин HCDR3-oбласти MS-Roche №7.9Н2×7.12L2; протеин/1; искусственная последовательность
GKGNTHKPYGYVRYFDV (SEQ ID NO:67)
SEQ ID NO:68
НСDR3-область MS-Roche №7.9Н4×7.12L2; ДНК; искусственная последовательность
GGTAAGGGTAATACTCATAAGCCTTATGGTTATGTTCGTTATTTTGATGTT (SEQ ID NO:68)
SEQ ID NO:69
протеин HCDR3-области MS-Roche №7.9Н4×7.12L2; протеин/1; искусственная последовательность
GKGNTHKPYGYVRYFDV (SEQ ID NO:69)
SEQ ID NO:70
НCDR3-область MS-Roche №7.11Н1×7.11L1; ДНК; искусственная последовательность
GGTAAGGGTAATACTCATAAGCCTTATGGTTATGTTCGTTATTTTGATGTT
(SEQ ID NO:70)
SEQ ID NO:71
протеин HCDR3-области MS-Roche №7.11Н1×7.11L1; протеин/1; искусственная последовательность
GKGNTHKPYGYVRYFDV (SEQ ID NO:71)
SEQ ID NO:72
HCDR3-область MS-Roche №7.11H1×7.2L1; ДНК; искусственная последовательность
GGTAAGGGTAATACTCATAAGCCTTATGGTTATGTTCGTTATTTTGATGTT
(SEQ ID NO:72)
SEQ ID NO:73
протеин HCDR3-oбласти MS-Roche №7.11H1×7.2L1; протеин/1; искусственная последовательность
GKGNTHKPYGYVRYFDV (SEQ ID NO:73)
SEQ ID NO:74
LСDR3-область MS-Roche №3.6Н5×3.6L2; ДНК; искусственная последовательность
CAGCAGACTTATAATTATCCTCCT (SEQ ID NO:74)
SEQ ID NO:75
протеин LCDR3-области MS-Roche №3.6Н5×3.6L2; протеин/1; искусственная последовательность
QQTYNYPP (SEQ ID NO:75)
SEQ ID NO:76
LCDR3-область MS-Roche №3.6Н8×3.6L2; ДНК; искусственная последовательность
CAGCAGACTTATAATTATCCTCCT (SEQ ID NO:76)
SEQ ID NO:77
протеин LCDR3-область MS-Roche №3.6Н8×3.6L2; протеин/1; искусственная последовательность
QQTYNYPP (SEQ ID NO:77)
SEQ ID NO:78
LCDR3-область MS-Roche №7.4Н2×7.2L1; ДНК; искусственная последовательность
CAGCAGATTTATTCTTTTCCTCAT (SEQ ID NO:78)
SEQ ID NO:79
протеин LCDR3-области MS-Roche №7.4Н2×7.2L1; протеин/1; искусственная последовательность
QQIYSFPH (SEQ ID NO:79)
SEQ ID NO:80
LCDR3-область MS-Roche №7.9Н2×7.12 L2; ДНК; искусственная последовательность
CTTCAGCTTTATAATATTCCTAAT (SEQ ID NO:80)
SEQ ID NO:81
протеин LСDR3-области MS-Roche №7.9Н2×7.12 L2; протеин/1; искусственная последовательность
LQLYNIPN (SEQ ID NO:81)
SEQ ID NO:82
LCDR3-область MS-Roche №7.9Н4×7.12L2; ДНК; искусственная последовательность
CTTCAGCTTTATAATATTCCTAAT (SEQ ID NO:82)
SEQ ID NO:83
протеин LCDR3-область MS-Roche №7.9Н4×7.12L2; протеин/1; искусственная последовательность
LQLYNIPN (SEQ ID NO:83)
SEQ ID NO:84
LCDR3-область MS-Roche №7.11H1×7.11L1; ДНК; искусственная последовательность
CAGCAGGTTTATTCTCCTCCTCAT (SEQ ID NO:84)
SEQ ID NO:85
протеин LCDR3-области MS-Roche №7.11H1×7.11L1; протеин/1; искусственная последовательность
QQVYSPPH (SEQ ID NO:85)
SEQ ID NO:86
LCDR3-область MS-Roche №7.11H1×7.2L1; ДНК; искусственная последовательность
CAGCAGATTTATTCTTTTCCTCAT (SEQ ID NO:86)
SEQ ID NO:87
протеин LCDR3-области MS-Roche №7.11Н1×7.2L1; протеин/1; искусственная последовательность
QQIYSFPH (SEQ ID NO:87)
SEQ ID NO:88
VH-область MS-Roche №7.9Н7; ДНК; искусственная последовательность
Caggtgcaattggtggaaagcggcggcggcctggtgcaaccgggcggcagcctgcgtctgagctgcgcggcctccgga
tttacctttagcagctatgcgatgagctgggtgcgccaagcccctgggaagggtctcgagtgggtgagcgctattaatgcttc
tggtactcgtacttattatgctgattctgttaagggtcgttttaccatttcacgtgataattcgaaaaacaccctgtatctgcaaat
gaacagcctgcgtgcggaagatacggccgtgtattattgcgcgcgtggtaagggtaatactcataagccttatggttatgttc
gttattttgatgtttggggccaaggcaccctggtgacggttagctca (SEQ ID NO:88)
SEQ ID NO:89
протеин VH-области MS-Roche №7.9Н7; протеин/1; искусственная последовательность
QVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAINA
SGTRTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARGKGNTHKP
YGYVRYFDVWGQGTLVTVSS (SEQ ID NO:89)
SEQ ID NO:90
VL-область MS-Roche №7.9Н7; ДНК; искусственная последовательность
Gatatcgtgctgacccagagcccggcgaccctgagcctgtctccgggcgaacgtgcgaccctgagctgcagagcgagc
cagagcgtgagcagcagctatctggcgtggtaccagcagaaaccaggtcaagcaccgcgtctattaatttatggcgcgagc
agccgtgcaactggggtcccggcgcgttttagcggctctggatccggcacggattttaccctgaccattagcagcctggaa
cctgaagactttgcgacttattattgccttcagatttataatatgcctattacctttggccagggtacgaaagttgaaattaaacgt
acg (SEQ ID NO:90)
SEQ ID NO:91
протеин VL-области MS-Roche №7.9Н7; протеин/1; искусственная последовательность
DIVLTQSPATLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSR
ATGVPARFSGSGSGTDFTLTISSLEPEDFATYYCLQIYNMPITFGQGTKVEIKRT
(SEQ ID NO:91)
SEQ ID NO:92
HCDR3-область MS-Roche №7.9Н7; ДНК; искусственная последовательность
Ggtaagggtaatactcataagccttatggttatgttcgttattttgatgtt (SEQ ID NO:92)
SEQ ID NO:93
протеин HCDR3-oбласти MS-Roche №7.9H7; протеин/1; искусственная последовательность
GKGNTHKPYGYVRYFDV (SEQ ID NO:93)
SEQ ID NO:94
LCDR3-область MS-Roche №7.9Н7; ДНК; искусственная последовательность
Cttcagatttataatatgcctatt (SEQ ID NO:94)
SEQ ID NO:95
протеин LCDR3-oбласти MS-Roche №7.9H7; протеин/1; искусственная последовательность
LQIYNMPI (SEQ ID NO:95)
Дополнительные приведенные с целью иллюстрации последовательности представлены ниже в Перечне последовательностей, а также ниже в таблицах, в частности в таблицах 1, 8 и 10.
На чертежах показано:
на фиг.1 - полная последовательность библиотеки HuCAL®-Fab1.
Нумерация соответствует VBASE за исключением бреши в положении 9 VLλ. В VBASE брешь находится в положении 10 (Chothia и др., 1992). В полной последовательности показаны все консервативные остатки CDR3. Соответствующие последовательности, применяемые для библиотеки HuCAL-Fab1, описаны в прилагаемом перечне последовательностей
А: аминокислотная последовательность
Б: последовательность ДНК;
на фиг.2- вектор pMORPH®18_Fab, предназначенный для проявления Fab-фрагментов.
Векторная карта и последовательность ДНК, включающая сайты рестрикции;
на фиг.3 - Экспрессирующий Fab вектор pMORPH®×9_Fab.
Векторная карта и последовательность ДНК, включающая сайты рестрикции;
на фиг.4 - последовательности Fab-фрагментов родительских антител MS-Roche-3, MS-Roche-7 и MS-Roche 8.
А: аминокислотная последовательность
Б: последовательность ДНК
на фиг.5 - полученное методом непрямой иммунофлуоресценции изображение амилоидных бляшек на криостатном срезе человеческой височной коры головного мозга. Бляшки метили с помощью MS-R №3.2 Fab-фрагмента (верхние панели) и MS-R №7.4 Fab-фрагмента (нижние панели) в концентрации 20 мкг/мл (левые панели) и 5 мкг/мл (правые панели) в условиях строгой блокады. Связывание MS-R Fab-фрагмента подтверждали с помощью козьего античеловеческого антитела, конъюгированного с Су3;
на фиг.6 - полученное методом непрямой иммунофлуоресценции изображение амилоидных бляшек на криостатном срезе человеческой височной коры головного мозга. Бляшки метили с помощью MS-R №3.3 IgG1 (верхние панели) и MS-R №7.12 IgG1 (нижние панели) в концентрации 0,05 мкг/мл (левые панели) и 0,01 мкг/мл (правые панели) в условиях строгой блокады. Связывание антитела MS-R IgG1 подтверждали с помощью козьего антитела к человеческому Ig (H+L), конъюгированного с Су3;
на фиг.7 - полученное методом непрямой иммунофлуоресценции изображение амилоидных бляшек на криостатном срезе человеческой височной коры, полученное с использованием антител после конечного созревания аффинности. Бляшки метили с помощью MS-R №7.9.Н7 IgG1 (МАт 31, верхняя панель), MS-R №7.11.H1×7.2.L1 IgG1 (МАт 11, средняя панель) и MS-R №3.4.Н7 (нижняя панель). Антитела применяли в концентрации 0,05 мкг/мл (левые панели) и 0,01 мкг/мл (правые панели) в условиях строгой блокады. Связывание антитела MS-R IgG1 подтверждали с помощью козьего антитела к человеческому Ig (H+L), конъюгированного с Су3;
масштаб: 8,5 мм =150 мкм;
на фиг.8 - анализ полимеризации.
Антитела к Аβ предупреждают включение биотинилированного Аβ в ранее образовавшиеся агрегаты Аβ;
на фиг.9 - анализ деполимеризации.
Антитела кАβ индуцируют высвобождение биотинилированного Аβ из агрегатов Аβ.
На фиг.10 - выявление с помощью окрашивания in vivo амилоидных бляшек в мышах, несущих двойной трансген APP/PS2, после внутривенной инъекции 1 мг MS-Roche IgG №7.9.Н2×7.12.L2. Через 3 дня мышей подвергали перфузии забуференным фосфатом физиологическим раствором и умерщвляли. Присутствие человеческого IgG, связанного с амилоидными бляшками, подтверждали с помощью конфокальной микроскопии после мечения криостатных срезов лобной коры головного мозга конъюгатом козье антитело к человеческому IgG - Су3 (панель Б). Этот же срез подвергали контрастному окрашиванию с использованием мышиного моноклонального антитела к Абета (конъюгат ВАР-2-Аlеха488, панель А) для визуализации положения амилоидных бляшек. Показаны индивидуальные красные и зеленые каналы, слитое изображение и сигналы совместной локализации;
масштаб: 1 см = 50 мкм.
На фиг.11 - выявление путем окрашивания in vivo амилоидных бляшек в мышах, несущих двойной трансген APP/PS2, после внутривенной инъекции 1 мг MS-Roche IgG №7.9.Н4×7.12.L2. Экспериментальные условия и процедура окрашивания идентичны приведенным в комментарии к фиг.10;
масштаб: 1,6 см = 50 мкм.
На фиг.12 - выявление путем окрашивания in vivo амилоидных бляшек в мышах, несущих двойной трансген APP/PS2, после внутривенной инъекции 1 мг MS-Roche IgG №7.11. HI×7.2.L1 (МАт 11). Экспериментальные условия и процедура окрашивания идентичны приведенным в комментарии к фиг.10;
масштаб: 1,4 см = 70 мкм.
На фиг.13 - выявление путем окрашивания in vivo амилоидных бляшек в мышах, несущих двойной трансген APP/PS2, после внутривенной инъекции 2 мг MS-Roche IgG №7.9.Н7 (МАт 31) в день 0, 3 и 6. Через 9 дней мышей подвергали перфузии забуференным фосфатом физиологическим раствором и умерщвляли. Присутствие человеческого IgG, связанного с амилоидными бляшками, подтверждали с помощью конфокальной микроскопии после мечения криостатных срезов лобной коры головного мозга конъюгатом козье антитело к человеческому IgG - Су3 (панель Б). Этот же срез подвергали контрастному окрашиванию с использованием мышиного моноклонального антитела к Абета (конъюгат ВАР-2-Аlеха488, панель А) для визуализации положения амилоидных бляшек. Показаны индивидуальные красные (панель Б) и зеленые (панель А) каналы, слитое изображение (панель Г) и сигналы совместной локализации (панель В);
масштаб: 1,6 см = 80 мкм (панель А. Б, В); 1,0 см= 50 мкм панель (Г).
На фиг.14 - выявление путем окрашивания in vivo амилоидных бляшек в мышах, несущих двойной трансген APP/PS2, после внутривенной инъекции 2 мг MS-Roche IgG №7.11.H1×7.2.L1 (МАт 11) в день 0, 3 и 6. Экспериментальные условия и процедура окрашивания идентичны приведенным в комментарии к фиг.13;
масштаб: 1,6 см = 80 мкм.
На фиг.15 - результаты анализа связывания антител к Аβ с находящимся на клеточной поверхности АРР. Связывание антитела с трансфектированными человеческим АРР клетками линии НЕК293 и нетрансфектированными контрольными клетками оценивали с помощью проточной цитометрии. Приведенные ниже примеры служат для иллюстрации изобретения.
Пример 1: Конструирование и скрининг комбинаторной библиотеки человеческих химерных антител (HuCAL®-Fab1)
Клонирование HuCAL®-Fab1
HuCA®-Fab1 представляет собой библиотеку полностью синтетических модулированных человеческих антител в формате Fab-фрагментов антитела. HuCAL®-Fab1 конструировали, используя в качестве исходной библиотеку антител в формате одноцепочечного антитела (HuCAL®-scFv; Knappik, J. Mol. Biol. 296, 2000, сс.57-86).
Положения 1 и 2 Vλ. Исходные основные гены HuCAL® конструировали с аутентичными для них N-концами: VLλ1: QS (CAGAGC), VLλ2: QS (CAGAGC) и VLλ3: SY (AGCTAT). Последовательности, содержащие эти аминокислоты, описаны в WO 97/08320. При конструировании библиотеки HuCAL® первые две аминокислоты заменяли на DI для облегчения клонирования библиотеки (сайт EcoRI). Все библиотеки HuCAL® содержали гены VLλ с сайтом EcoRV GATATC (DI) на 5'-конце. Все гены каппа-цепи HuCAL® (основные гены и все гены,входящие в библиотеку) содержали DI на 5'-конце (фиг.1 А и Б).
Положение 1 VH. Исходные основные гены HuCAL® конструировали с аутентичными для них N-концами: Н1А, VH1B, VH2, VH4 и VH6, в которых первая аминокислота представляла собой Q(=CAG), и VH3 и VH5, в которых первая аминокислота представляла собой Е(=GAA). Последовательности, содержащие эти аминокислоты, описаны в WO 97/08320. В процессе клонирования библиотеки HuCAL®-Fab1 аминокислоту в положении 1 VH заменяли на Q (CAG) во всех VH-генах (фиг.1А и Б).
Создание библиотек CDR
Положение 85 Vκ1/Vκ3. Поскольку для интродукции библиотеки CDR3 использовали процедуру кассетного мутагенеза (Knappik, (2000), loc. cit), то в положении 85 Vκ1 и Vκ3 может находиться либо Т, либо V. Таким образом, в процессе конструирования библиотеки HuCAL®-scFv1 положение 85 в Vκ1 и Vκ3 может варьироваться следующим образом: Vκ1, исходная последовательность, 85Т (кодон АСС); Vκ1, библиотека, 85Т или 85V (TRIM кодоны ACT или GTT); Vκ3, исходная последовательность, 85V (кодон GTG); Vκ3, библиотека, 85Т или 85V (TRIM кодоны ACT или GTT); аналогично применяемым в HuCAL®-Fab1.
Конструирование CDR3. Все консервативные остатки CDR3 представлены на фиг.1А и Б.
Длины CDR3. Ниже представлено полученное распределение длин CDR3. Остатки, которые можно заменять, представлены в скобках (x) на фиг.1. В CDR3 V-каппа цепи, состоящей из 8 аминокислотных остатков (положение 89-96) (иногда 7-10 остатков), Q89, S90 и D92 являются фиксированными; а в CDR3 VH-цепи, состоящей из 5-28 аминокислотных остатков (положение 95-102) (иногда 4-28), D101 является фиксированным.
HuCAL®-Fab1 клонировали в фагмидном экспрессионном векторе pMORPH®18_Fab1 (фиг.2). Этот вектор содержит Fd-фрагмент с сигнальной последовательностью phoA, слитой на С-конце с укороченным геном III протеина нитчатого фага, и содержит также легкую цепь VL-CL с сигнальной последовательностью отрА. Обе цепи находятся под контролем оперона lac. Константные области Сλ, Сκ и СН1 представляют собой синтетические гены, полностью совместимые с модуляторной системой HuCAL®(Knappik, (2000), loc. cit).
Полную VH-цепь (MunI/StyI-фрагмент) заменяли на «пустой» фрагмент длиной 1205 пар оснований, содержащий транскрипционную единицу β-лактамазы (bla), облегчая тем самым последующие стадии получения векторного фрагмента и возможность отбора в отношении полного удаления VH. После замены VH-области VLλ удаляли с помощью EcoRI/DraIII, a VLκ с помощью EcoRI/BslWIs заменяли на фрагмент длиной 1420 пар оснований гена бактериальной щелочной фосфатазы (bap). Поскольку изменчивость легких цепей ниже, чем тяжелых цепей, клонирование начинали с библиотек легких цепей. Библиотеки VLλ- и VLκ-легких цепей, различающиеся по L-CDR3, которые создавали для библиотеки HuCAL®-scFv (Knappik, (2000), loc. cit.), использовали также для клонирования HuCAL®-Fabi. В случае λ-цепи они состояли из λ1-, λ2- и λ3-каркасного участка HuCAL®, и их общая изменчивость составляла 5,7×106. VLλ-фрагменты амплифицировали с помощью 15 циклов ПЦР (с использованием Pwo-полимеразы) с праймерами 5'-GTGGTGGTTCCGATATC-3' (SEQ ID NO:28) и 5'-AGCGTCACACTCGGTGCGGCTTTCGGCTGGCCAAGAACGGTTA-3' (SEQ ID NO:29). ПЦР-продукты расщепляли с помощью EcoRV/DraIII и очищали на геле. В случае библиотеки VLλ несущий bap «пустой» фрагмент удаляли из библиотеки вектора с помощью EcoRV/DraIII. 2 мкг очищенного с помощью геля вектора лигировали с 3-кратным молярным избытком VLλ-цепей в течение 16 ч при 16°С, и полученные в результате лигирования смеси встраивали путем электропорации в 800 мкл клеток Е. coli TOP I OF (фирма Invitrogen), получая всего 4,1×108 независимых колоний. Трансформанты амплифицировали примерно в 2000 раз в среде: 2 × YT/1% глюкозы/34 мкг/мл хлорамфеникола/100 мкг/мл ампициллина, собирали и хранили в 20% (мас./об.) глицерине при -80°С.
Библиотеки κ-цепей содержали основные гены κ1-, κ2-, κ3- и κ4-HuCAL® с общей изменчивостью 5,7×106. VLλ-цепи получали рестрикцией с использованием EcoRy/BsiWI и очищали на геле. В случае библиотеки VLκ несущий bap «пустой» фрагмент удаляли из библиотеки вектора с помощью EcoRV/BslWI. 1 мкг очищенного с помощью геля вектора смешивали с 5-кратным молярным избытком VLκ-цепей. Лигирование и трансформацию клеток Е. coli TOPI OF (фирма Invitrogen) осуществляли с помощью процесса, описанного для VLλ-цепей, получая всего 1,6×108 независимых колоний.
Получали ДНК двух библиотек легких цепей и несущий bla «пустой» фрагмент удаляли с помощью MunI/StyI, создавая тем самым два вектора для вставки подбиблиотек VH.
VH-библиотеки HuCAL®-scFv применяли для создания HuCAL®-Fabl. VH-библиотеки HuCAL®-scFv состояли из основных генов VH1A/B-6, которые различались двумя кассетами тринуклеотидных VH-CDR3, каждая из которых отличалась по длине CDR3, и каждую VH-библиотеку объединяли с VLκ- и VLλ-библиотекой. Для создания HuCAL®-Fabl получали ДНК из этих VH-библиотек, сохраняя исходную изменчивость. ДНК расщепляли с помощью MunI/StyI и очищали на геле. 5-кратный молярный избыток VH-цепей лигировали с 3 мкг вектора, содержащего библиотеку VLλ, и с 3 мкг вектора, содержащего библиотеку VLκ, в течение 4 ч при 22°С. Для каждого вектора полученные в результате лигирования смеси встраивали путем электропорации в 1200 мкл клеток Е. coli TOP I OF (фирма Invitrogen), получая всего 2,1×1010 независимых колоний. Трансформанты амплифицировали примерно в 4000 раз в среде: 2 × YT/1% глюкозы/34 мкг/мл хлорамфеникола/100 мкг/мл тетрациклина, собирали и хранили в 20% (мас./об.) глицерине при -80°С.
Для качественного контроля легкой цепи и тяжелой цепи индивидуальные клоны секвенировали с использованием 5'-CAGGAAACAGCTATGAC-3' (SEQ ID NO:30) и 5'-TACCGTTGCTCTTCACCCC-3' (SEQ ID NO:31) соответственно.
«Спасение» фагмиды, амплификация и очистка фага. HuCAL®-Fab 1 амплифицировали в 2 × TY-среде, содержащей 34 мкг/мл хлорамфеникола, 10 мкг/мл тетрациклина и 1% глюкозы (2 × TY-CG). После заражения фагом-хелпером (VCSM13) при 37°С при значении ОП600 примерно 0,5, центрифугирования и ресуспендирования в среде: 2 × TY/34 мкг/мл хлорамфеникола/50 мкг/мл канамицина клетки выращивали в течение ночи при 30°С. Фаги осаждали из супернатанта с помощью ПЭГ (Ausubel, (1998), Current protocols in molecular biology. John Wiley & Sons, Inc., New York, USA), ресуспендировали в ЗФР/20% глицерина и хранили при -80°С. Амплификацию фага между двумя циклами пэннинга осуществляли следующим образом: находящиеся на средней логарифмической фазе роста TG1-клетки заражали элюированным фагом и высевали на LB-агар, дополненный 1% глюкозы и 34 мкг/мл хлорамфеникола. После инкубации при 30°С колонии соскабливали, доводили значение ОП600 до 0,5 и добавляли фаг-хелпер согласно описанному выше процессу.
Пример 2: Твердофазный пэннинг
Лунки титрационных микропланшетов MaxiSorpTM типа F96 (фирма Nunc) сенсибилизировали 100 мкл 2,5 мкМ человеческого пептида Аβ (1-40) (фирма Bachem), растворенного в TBS, который содержал NaN3 (0,05 об. %), и запечатанный планшет инкубировали в течение 3 дней при 37°С, за это время пептид мог агрегироваться на планшете. После блокирования с помощью 5% обезжиренного сухого молока в TBS добавляли 1-5×1012 фага HuCAL®-Fab, очищенного согласно описанному выше методу, и выдерживали в течение 1 ч при 20°С. После нескольких стадий отмывки связанные фаги подвергали элюированию с использованием 500 мМ NaCl, 100 мМ глицина, рН 2,2 и затем нейтрализовали 1М Трис-Cl-буфером, рН 7. Осуществляли 3 цикла пэннинга, при этом между каждым циклом амплифицировали фаг согласно описанному выше методу, строгость отмывки повышали от цикла к циклу.
Пример 3: Субклонирование отобранных для экспрессии Fab-фрагментов
Кодирующие Fab вставки отобранных HuCAL®-Fab-фрагментов субклонировали в экспрессионном векторе pMORPH®×7_FS для облегчения быстрой экспрессии растворимого Fab. Препараты ДНК отобранных клонов HuCAL -Fab расщепляли с помощью XbaI/EcoRI, вырезая тем самым кодирующую Fab вставку (ompA-VL и phoA-Fd). После субклонирования очищенных вставок в расщепленном с помощью XbaI/EcoRI векторе pMORPH®×7, который ранее нес вставку scFv, получали вектор, предназначенный для экспрессии Fab, обозначенный pMORPH®×9_Fabl (фиг.3). Fab-фрагменты, экспрессируемые в этом векторе, несли две С-концевых метки (FLAG и Strep), предназначенные для обнаружения и очистки.
Пример 4: Идентификация связывающихся с Аβ Fab-фрагментов с помощью ELISA
Лунки титрационных микропланшетов Maxisorp типа F384 (фирма Nunc) сенсибилизировали с помощью 20 мкл 2,5 мкМ человеческого пептида Ар (1-40) (фирма Bachem), растворенного в TBS, который содержал NaN3 (0,05 об. %), и запечатанный планшет инкубировали в течение 3 дней при 37°С, за это время пептид мог агрегироваться на планшете. Экспрессию индивидуальных Fab-фрагментов индуцировали с помощью 1 мМ ИПТГ в течение 16 ч при 22°С. Растворимый Fab-фрагмент экстрагировали из Е. coli посредством лизиса с использованием BEL-буфера (буфера, содержащего борную кислоту, NaCl, ЭДТК и лизозим, рН 8) и применяли для анализа методом ELISA. Для обнаружения Fab-фрагмента применяли конъюгированное с щелочной фосфатазой козье антитело к Fab (фирма Dianova/Jackson Immuno Research). После возбуждения при длине волны 340 нм испускание при длине волны 535 нм регистрировали после добавления флуоресцентного субстрата AttoPhos (фирма Roche Diagnostics).
Пример 5: Оптимизация фрагментов антитела
Для оптимизации аффинности к связыванию отобранных связывающихся с Аβ фрагментов антитела некоторые из Fab-фрагментов, т.е. MS-Roche-3 (MSR-3), MS-Roche-7 (MSR-7) и MS-Roche-8 (MSR-8) (фиг.4) применяли для конструирования библиотеки Fab-фрагментов антитела, заменяя родительскую κ3-цепь VL пулом всех каппа-цепей κ1-3, которые различались по CDR3, из библиотеки HuCAL® (Knappik и др., 2000).
Fab-фрагменты MS-Roche-3, -7 и -8 клонировали с использованием сайтов рестрикции XbaI/EcoRI из pMORPH®×9_FS в pMORPH®18, фагмидном векторе, предназначенном для фагового проявления Fab-фрагментов, получая pMORPH®18_Fabl (фиг.2). Пул каппа-цепей клонировали в pMORPH®18_Fabl с использованием сайтов рестрикции XbaI/SphI.
Полученную библиотеку оптимизированных Fab-фрагментов подвергали скринингу путем пэннинга через агрегированный человеческий пептид Аβ (1-40), нанесенный на твердую подложку, согласно методу, описанному в примере 2. Оптимизированные клоны идентифицировали путем koff-упорядочивания с использованием Biacore-анализа, описанного в примере 8. Оптимизированные клоны MS-Roche-3.2, 3.3, 3.4, 3.6, 7.2, 7.3, 7.4, 7.9, 7.11, 7.12, 8.1, 8.2 дополнительно характеризовали и установлено, что они обладали повышенной аффинностью и биологической активностью по сравнению с исходным фрагментом MS-Roche-3, MS-Roche-7 и MS-Roche-8 (фиг.4). Перечисленные CDR-участки относятся к библиотеке HuCAL®, основанной на консенсусном гене антитела VН3каппа3. Fab-фрагмент MS-Roche-7.12 получали путем клонирования HCDR3 родительского клона MS-R 7 в библиотеке HuCAL®-Fab, несущей различия во всех шести CDR-участках, используя процедуру, идентичную применяемой для СDR3-кассет, которая описана у Knappik и др., 2000. Библиотеку кассет создавали, строго основываясь на известном естественном распределении аминокислот и следуя концепции канонических конформаций CDR, разработанной Allazikani (Allazikani и др., 1997). Однако в противоположность основным генам HuCAL® клон MS-Roche 7.12 содержал аминокислоту S в положении 49 VL-цепи (см. приложение к таблице 1).
Оптимизированные Fab-фрагменты после первого цикла созревания аффинности обладали улучшенными характеристиками по сравнению с исходными клонами MS-Roche-3, MS-Roche-7 и MS-Roche-8 (фиг.4). Аффинности связывания созревших Fab-фрагментов с Aβ1-40 и Aβ1-42 существенно возрастали и характеризовались значениями KD 22-240 нМ по сравнению с 850-1714 нМ родительских клонов (таблица 3). Иммуногистохимические анализы амилоидных бляшек в срезах головного мозга пациентов, страдающих болезнью Альцгеймера (БА), также выявили заметно возросший уровень окрашивания созревших клонов, т.е. были получены более высокие соотношения сигнала и основного уровня, а позитивное окрашивание бляшек было обнаружено при относительно низких концентрациях созревших Fab-фрагментов (фиг.5).
Для дополнительной оптимизации СDR2-участки VH и CDR1-участки VL наборов фрагментов антител, полученных из клонов MS-Roche-3, -7 и -8 с оптимизированным L-CDR3 (таблица 1; фиг.4), оптимизировали с помощью кассетного мутагенеза, применяя для этой цели сайтнаправленный мутагенез с использованием трех нуклеотидов (Virnekas и др., 1994). Для этого конструировали основанную на трех нуклеотидах НСОК2-кассету и основанную на трех нуклеотидах LCDR1-кассету, применяя метод, идентичный методу создания CDR3-кассет, который описан у Knappik и др., 2000. Библиотеку кассет создавали, строго основываясь на известном естественном распределении аминокислот и следуя концепции канонических конформаций CDR, разработанной Allazikani (Allazikani и др., 1997). Протокол, применяемый для оптимизации первоначально отобранных фрагментов антител, должен имитировать процесс созревания аффинности с помощью соматической сверхмутации, происходящей при естественном иммунном ответе.
Каждую из полученных библиотек по отдельности подвергали скринингу, согласно описанному выше методу, получая клоны с оптимизированным либо H-CDR2-, либо L-CDR1-участком. Все клоны идентифицировали согласно описанному выше методу по улучшенному kof в отношении Aβ1-40-фибрилл после koff-упорядочивания с использованием Biacore-анализа и было установлено, что они обладают улучшенной аффинностью либо к Аβ1-40 или Аβ-42, либо к обоим пептидам при сравнении с соответствующим родительским клоном (таблица 3). В таблице 1 представлены характеристики последовательностей родительских, а также последовательности оптимизированных клонов. Перечисленные CDR участки относятся к библиотеке HuCAL®, основанной на консенсусном гене антитела VН3каппа3.
Например, аффинность родительского Fab MS-Roche-7 к Аβ1-40 улучшилась в 35 раз (от 1 100 нМ до 31 нМ) после оптимизации L-CDR3 (MS-Roche-7.9) и еще более улучшилась (до 5 нМ) после оптимизации H-CDR2 (MS-Roche-7.9H2), как проиллюстрировано в таблице 3.
Процедура оптимизации H-CDR2 и L-CDR1 не только приводит к улучшению аффинности, но также способствует значительному улучшению окрашивания некоторыми клонами амилоидных бляшек в срезах головного мозга страдающих БА пациентов, что доказано для клонов MS-Roche 7.9H2 и 7.9Н3.
Последовательности, принадлежащие к консенсусным последовательностям VH3 и Vκ3 HuCAL, см. на фиг.1A.
Пример 6
Конструирование экспрессионных векторов HuCAL® иммуноглобулина
Клонирование тяжелой цепи. Множественный сайт клонирования рсДНК3.1+ (фирма Invitrogen) изымали (NheI/ApaI) и «лишний» фрагмент, совместимый с рестрикционными сайтами, которые использовали для создания HuCAL®, встраивали для лигирования лидерных последовательностей (NheI/EcoRI), VH-областей (MunI/) и константных областей иммуноглобулина (BlpI/АраГ). В лидерную последовательность (EMBL 83133) встраивали последовательность Козака (Kozak, 1987). Константные области человеческого IgG (PIR A02146), IgG4 (EMBL K01316) и сывороточного IgAl (EMBL J00220) расщепляли на перекрывающиеся олигонуклеотиды длиной примерно 70 оснований. Интродуцировали молчащие мутации для изымания сайтов рестрикции, несовместимых с конструкцией HuCAL®. Сплайсинг олигонуклеотидов осуществляли с помощью удлиняющей перекрытия ПЦР.
В процессе субклонирования из Fab в IgG, последовательность ДНК VH Fab-фрагмента вырезали с помощью Mfe I/Blp I и встраивали путем лигирования в несущий IgG вектор, расщепленный («открытый») с помощью EcoR I/Blp I. EcoR I (g/aattc) и Mfe I (c/aattg) разделяли совместимые «липкие» концы (aatt) и последовательность ДНК исходного сайта Mfe I в Fab-фрагменте заменяли с: c/aattg на: g/aattg после встраивания путем лигирования в экспрессионный вектор, несущий IgG, тем самым разрушая оба сайта Mfe I и EcoR I и получая при этом аминокислотную замену Q (кодон: саа) на Е (кодон: gaa).
Клонирование легкой цепи. Множественный сайт клонирования рсДНК3.1/Zeo+ (фирма Invitrogen) заменяли двумя различными «лишними» фрагментами. «Лишний» κ-фрагмент обеспечивал создание сайтов рестрикции, предназначенных для встраивания κ-лидера (NheI/EcoRV), Vκ-областей HuCAL®-scFv(EcoRV/BsiWI) и константной области κ-цепи (BsiWI/ApaI). Соответствующие сайты рестрикции в «лишнем» λ-фрагменте представляли собой NheI/EcoRV (λ-лидер), EcoRV/Hpal (Vλ-области) и HpaI/ApaI (константная область λ-цепи). Как в κ-лидерную последовательность (EMBL Z00022), так и в λ-лидерную последовательность (EMBL J00241) встраивали последовательности Козака. Константные области человеческой κ-цепи (EMBL L00241) и λ-цепи (EMBL M18645) объединяли с помощью описанной выше удлиняющей перекрытия ПЦР.
Создание экспрессирующих IgG СНО-клеток. СНО-клетки линии К1 совместно трансфектировали эквимолярной смесью экспрессионных векторов тяжелой и легкой цепей IgG. Трансфектанты, обладающие двойственной устойчивостью, отбирали с помощью 600 мкг/мл G418 и 300 мкг/мл зеоцина (фирма Invitrogen) с последующим предельным разбавлением. В супернатанте индивидуальных клонов оценивали экспрессию IgG с помощью основанного на применении иммобилизованных антител метода ELISA. Позитивные клоны размножали в среде RPMI-1640, дополненной 10% «ультра-низким» IgG-ФТС (фирма Life Technologies). После доведения значения рН супернатанта до 8,0 и стерилизации фильтрацией раствор подвергали стандартной хроматографии на колонках с протеином А (размер пор 20 Å, фирма РЕ Biosystems).
Пример 7: Pepspot-анализ с использованием декапептидов
Приведенную ниже аминокислотную последовательность, содержащую Аβ (1-42), разделяли на 43 перекрывающихся декапептида со сдвигом рамки считывания на одну аминокислоту.
ISEVKM1DAEF RHDSGYEVHH QKLVFFAEDV GSNKGAIIGL MVGGVVI42ATV IV (SEQ ID NO:414). Таким образом, включенная в нее последовательность DAEF RHDSGYEVHH QKLVFFAEDV GSNKGAIIGL MVGGVVIA (SEQ ID NO: 27) соответствовала аминокислотам 1-42 пептида Аβ4/β-А4.
Синтезировали 43 декапептида с ацетилированными N-концами и С-концами, которые ковалентно связывали с целлюлозной пластиной («pepspot») с использованием метода поставщика (Jerini BioTools, Берлин). Целлюлозную пластину инкубировали в течение 2 ч на качающейся платформе с моноклональным антителом (2 мкг/мл) в блокирующем буфере (50 мМ Трис×HCl, 140 мМ NaCl, 5 мМ NaЭДТК, 0,05% NP40 (фирма Fluka), 0,25% желатина (Sigma), 1% фракции V бычьего сывороточного альбумина (фирма Sigma), рН 7,4). Пластину промывали трижды по 3 мин на качающейся платформе TBS (10 мМ Трис×HCl, 150 мМ NaCl, рН 7,5). Затем смачивали катодным буфером (25 мМ Трис-основание, 40 мМ 6-аминогексановая кислота, 0,01% ДСН, 20% метанола) и переносили на полусухой блок для блоттинга с пептидом, нанесенным на мембрану из поливинилиденфторида (ПВДФ-мембрану) (фирма Biorad) такого же размера.
Полусухой блок для блоттинга состоял из свежесмоченных листов фильтровальной бумаги (Whatman No.3), по размеру слегка превышающих пластину с пептидом:
3 листа, смоченные катодным буфером,
пластина с пептидом,
пластина ПВДФ-мембраны, смоченная метанолом,
3 листа бумаги, смоченные анодным буфером 1 (30 мМ Трис-основание, 20% метанола),
3 листа бумаги, смоченные анодным буфером 2 (0,3 мМ Трис-основание, 20% метанола).
Перенос осуществляли при плотности электрического тока между катодом 5 и анодом 0,8 мА/см2 в течение 40 мин, что достаточно для элюирования большей части антител из целлюлозной пластины и отложения их на ПВДФ-мембране. Затем ПВДФ-мембрану заменяли второй ПВДФ-мембраной и осуществляли перенос в течение еще 40 мин для гарантии полного элюирования из целлюлозной пластины.
ПВДФ-мембрану погружали на 10 мин в блокирующий буфер. Затем добавляли меченное пероксидазой из хрена (HRP) антитело к человеческому Ig H+L (фирма Pierce), используя разбавление 1:1000, и мембрану инкубировали на качающейся платформе в течение 1 ч. Затем ее промывали 3×10 мин TBST (TBS с 0,005% Твин 20). Для появления окраски мембрану погружали в раствор, который содержал 3 мг 4-хлорнафтола, растворенного в 9 мл метанола, с добавлением 41 мл ЗФР (20 мМ Na-фосфат, 150 мМ NaCl, pH 7,2), 10 мкл 30% перекиси водорода (фирма Merck). После появления сине-черных пятен мембрану интенсивно промывали водой и сушили.
Оценку дающих реакцию с антителом пятен (pepspot) осуществляли путем визуального мониторинга через прозрачную матрицу для блоттинга (spot). В качестве эпитопов представляющего интерес антитела принимали минимальную аминокислотную последовательность реакционноспособных пептидов. Для сравнения аналогичным образом анализировали мышиные моноклональные антитела (ВАР-2, ВАР-1, ВАР-17 ВАР-21, ВАР-24 и 4G8), используя при этом меченное с помощью HRP антитело к мышиному Ig вместо антитела к человеческому Ig.
Следует отметить, что созревание аффинности и превращение моновалентных Fab-фрагментов в полноразмерные антитела в виде IgGI, как правило, приводит к некоторому расширению распознающей эпитоп последовательности, что подтверждено с помощью pepspot-анализа и ELISA. Этот может быть связано с рекрутментом контактных точек в области взаимодействия антитело-антиген в результате созревания аффинности или с более сильным связыванием с минимальным эпитопом, что, как обнаружено, также может ослаблять взаимодействия с соседней аминокислотой. Последний факт может быть доказан путем зондирования полученных из Аβ пептидов полноразмерными антителами в виде IgG. Как проиллюстрировано в таблице 2, в которой представлены данные pepspot-анализа, последовательности, распознающие N-концевые и срединные эпитопы, удлиняются на несколько (вплоть до трех) аминокислот, что установлено при сравнении родительских Fab и соответствующих полностью созревших антител в виде IgG. Однако следует учитывать также, что декапептиды модифицировали для ковалентного связывания с С-концевой аминокислотой, и эта аминокислота может представлять собой аминокислоту, которая менее доступна для полноразмерного антитела из-за связанных со стерическим строением помех. Если имеет место этот случай, то последняя С-концевая аминокислота не участвует должным образом в функции, распознающей эпитоп последовательность, и это возможное уменьшение длины минимальной распознающей последовательности на одну аминокислоту на С-конце следует принимать во внимание при pepspot-анализе, применяемом в настоящем изобретении.
Таблица 2: Pepspot-анализ связывания Fab-фрагментов и полноразмерных антител в виде IgG с декапептидами на целлюлозной пластине. Номера соответствуют незаменимым аминокислотам из последовательности Аβ1-40, которые должны присутствовать в декапептиде для оптимального связывания с антителом. Скобками обозначены пептиды со слабой реакционной способностью и, как следствие, с низкой активностью в эпитопе.
Пример 8: Определение значений КD антител MS-R Fab и MS-R IgG1, характеризующих связывание фибрилл Аβ1-40 и Аβ1-42, in vitro с помощью поверхностного плазменного резонанса (SPR)
Связывание антител к Аβ (Fab-фрагментов и IgGI) с фибриллярным Аβ оценивали в оперативном режиме «online» с помощью поверхностного плазменного резонанса (SPR) и аффинность молекулярных взаимодействий определяли согласно методам, описанным у Johnson, Anal. Biochem., 198, 1991, сс.268 -277 и у Richalet-Secordel, Anal.Biochem., 249, 1997, сс.165-173. Для этих измерений использовали устройства типа Biacore2000 и Biacore3000. Фибриллы Аβ1-40 и Аβ1-42 получали in vitro путем инкубации синтетических пептидов в концентрации 200 мкг/мл в 10 мМ Na-ацетатном буфере (рН 4,0) в течение 3 дней при 37°С. С помощью электронной микроскопии подтверждали фибриллярную структуру обоих пептидов, причем фибриллы Aβ1-40 в основном были более короткими (<1 мкм), а фибриллы Аβ1-42 в основном были более длинными (>1 мкм). Эти фибриллы, вероятно, в большей степени соответствуют агрегированным пептидам Аβ, которые присутствуют в головном мозге пациентов, страдающих БА, чем не имеющие определенной формы смеси аморфных агрегатов и неструктурированных осадков. Фибриллы разбавляли в соотношении 1:10 и непосредственно связывали с сенсорным чипом «Pioneer Sensor Chip F1» согласно инструкции производителя (BIAapplication Handbook, версия АВ, фирма Biacore AB, Уппсала, 1998). В предварительных экспериментах установлено, что отобранные MS-Roche Fab-фрагменты значительно различаются по своим реакционным кинетическим характеристикам и в зависимости от этого следует подбирать систему анализа данных. Для связывающихся агентов, характеризующихся медленной кинетикой, значения КD рассчитывали путем аппроксимации кривой зависимости сенсорных ответов от времени, т.е. из соотношения koff/kon. Связывающиеся агенты с более быстрой кинетикой анализировали путем аппроксимации кривой зависимости сенсорных ответов от концентрации при равновесии (адсорбция-изотермы). Значения KD рассчитывали с помощью сенсограмм фирмы Biacore на основе общей концентрации Fab-фрагментов, определенной путем анализа протеинов. Для клонов, полученных после первого и второго цикла созревания аффинности, содержание активных Fab-фрагментов в каждом препарате определяли с помощью Biacore согласно методу, описанному у Christensen, Analytical Biochemistry, 249, 1997, сс.153-164. В целом метод состоял в следующем: зависящее от времени связывание протеинов с фибриллами Аβ1-40, иммобилизованными на чипе Biacore, оценивали во время фазы ассоциации в условиях ограничения массы при различных скоростях потока раствора аналита. Условия ограничения массы реализовали путем иммобилизации больших количеств Аβ-фибрилл (2300 реактивных единиц) на поверхности исследуемого канала чипа и путем работы при относительно низких концентрациях аналита, т.е. 160 нМ (в пересчете на общую концентрацию Fab-протеина).
Обобщенные данные о значениях KD отобранных клонов MS-Roche, которые были идентифицированы при первичном скрининге библиотеки HuCAL, и их соответствующих зрелых производных, полученных после первого и второго цикла созревания аффинности, представлены в таблице 3. В первом цикле CDR3 тяжелой цепи (VH-CDR3) сохраняли без изменений, и оптимизация затрагивала изменения CDR3 легкой цепи (VL-CDR3). Во втором цикле созревания аффинности осуществляли изменение VL-CDR1 и VH-CDR2.
Некоторые из связывающихся агентов, полученных в первом цикле созревания, превращали в полноразмерные человеческие антитела в виде IgG1 с помощью технологии MorphoSys, описанной в примере 6, и значения KD определяли с помощью Biacore согласно описанному выше методу. Значения KD полноразмерных IgGI, которые связывались с фибриллами Аβ1-40 и Аβ1-42, представлены в таблице 4.
После второго цикла идентифицировали зрелые производные как L-CDR1-, так и Н-СDR2-библиотеки и давали объединяться легким и тяжелым цепям. Стратегия перекрестного клонирования описана в примере 13. Обменивали либо полные легкие цепи, либо LCDR1, либо L-CDR1+2. Значения KD отобранных полученных перекрестным клонированием Fab-фрагментов представлены в таблице 8.
Некоторые из Fab-фрагментов, полученных после первого и второго циклов созревания и после перекрестного клонирования связывающихся агентов, превращали в полноразмерные человеческие антитела в виде IgGI согласно технологии, разработанной MorphoSys, которая описана в примере 6. Значения KD связывания IgG с фибриллами Аβ1-40 и Аβ1-42 определяли с помощью Biacore. В целом, метод состоит в следующем: использовали кинетическую модель ступенчатого получения бивалентного комплекса и значения KD рассчитывали с помощью анализа равновесия связывания методом Скатчарда (Scatchard). Из-за очень медленного процесса ассоциации при низкой концентрации антитела (несколько часов до достижения равновесия) данные о равновесии связывания получали экстраполяцией кривых ассоциации для длительных интервалов времени. «On-» и «off-» скорости образования моновалентного и бивалентного комплекса определяли с помощью процедуры аппроксимации кривой и использовали для экстраполяции. На основе этих значений Req осуществляли Скатчард-анализ и определяли значения KD для образования моновалентного и бивалентного комплекса. Данные обобщены в таблице 3. Из нелинейных графиков зависимости Скатчарда определяли более высокую (бивалентный комплекс) и более низкую (моновалентный комплекс) аффинность к взаимодействию для MS-R IgG, полученных после второго цикла созревания аффинности и перекрестного клонирования. Эти две величины аффинности соответствуют более низким и более высоким значениям KD из диапазона, представленного в таблице 3.
№
Aβ1-40,
нМ
Aβ1-42,
нМ
№
Aβ1-40,
нМ
Aβ1-42,
нМ
№
Aβ1-40,
нМ
Aβ1-42,
нМ
Таблица 3: Значения KD, характеризующие связывание MS-R Fab с фибриллами Аβ1-40 и Aβ1-42, определяли с помощью Biacore. Для клонов, полученных после первого и второго цикла созревания аффиности, значения скорректированы с учетом содержания активного Fab-фрагмента в каждом образце, как это описано выше.
а - значения рассчитывали из зависимости сенсорных ответов от концентрации при равновесии; н.о. - не определяли.
Таблица 4: Значения КD, характеризующие связывание MS-R IgG1 с фибриллами Аβ1-40 и Аβ1-42, определяли с помощью Biacore. IgG получали из MS-R Fab-фрагментов, отобранных после первого цикла созревания аффинности. Значения скорректированы с учетом содержания активных MS-R IgG в каждом образце, как это описано выше.
Таблица 5: Значения КD, характеризующие связывание MS-R IgG1 с фибриллами Аβ1-40 и Аβ1-42, определяли с помощью Biacore. IgG получали из MS-R Fab-фрагментов, отобранных после первого и второго циклов созревания аффинности. Значения скорректированы с учетом содержания активных MS-R IgG в каждом образце, как это описано выше. Два значения KD, представленные для MS-R IgG, которые получены после второй стадии созревания аффинности и перекрестного клонирования связывающихся агентов, характеризуют более высокую и более низкую аффинность к взаимодействию, рассчитанную из нелинейных графиков зависимости Скатчарда. Для ряда других MS-R IgG (например, MS-R IgG 7-9.H2×7.12.L2 и MS-R IgG 7.9.H4×7.12.L2) комплекс линейных графиков Скатчарда имел погрешности (оказался размытым) и поэтому нельзя было рассчитать значения КD.
Пример 9: Окрашивание истинных человеческих амилоидных бляшек в срезах головного мозга страдающих болезнью Альцгеймера пациентов с помощью непрямой иммунофлуоресценции
Отобранные Fab-фрагменты и полноразмерные IgG1 MS-Roche тестировали в отношении связывания с β-амилоидными бляшками путем иммуногистохимического анализа. Криостатные срезы нефиксированной ткани человеческой височной коры головного мозга (полученной после смерти пациента, имеющего положительный диагноз болезни Альцгеймера) метили для оценки методом непрямой иммунофлуоресценции с помощью различных концентраций Fab-фрагментов или полноразмерных антител в виде IgGI MS-Roche. Наличие Fab-фрагментов и антител в виде IgG1 подтверждали с помощью козьего антитела к очищенному с помощью аффинной хроматографии человеческому F(ab')2-фрагменту, конъюгированного с Су3, и козьего антитела к человеческому Ig (H+L), конъюгированного с Су3, соответственно. Оба вторичных реагента получали от фирмы Jackson Immuno Research. Для всех контрольных образцов, которые включали неродственный Fab-фрагмент и только вторичные антитела, получены отрицательные результаты. Репрезентативные примеры окрашивания бляшек отобранными MS-Roche Fab-фрагментами и MS-Roche антителами в виде IgG1 представлены на фиг.5-7.
Пример 10: Анализ полимеризации: предупреждение агрегации Аβ
Синтетический Аβ при инкубации в водном буфере в течение нескольких дней спонтанно агрегируется и образует фибриллярные структуры, которые аналогичны обнаруженным в амилоидных отложениях в головном мозге пациентов, страдающих болезнью Альцгеймера. Авторами изобретения был разработан анализ in vitro для оценки включения биотинилированного Аβ в ранее образовавшиеся Аβ-агрегаты с целью выявления способности нейтрализовать Аβ с помощью антител к Аβ и других связывающихся с Аβ протеинов, таких как альбумин (Bohrmann и др., J. Biol. Chem. 274, 1999, сс.15990-15995). С помощью этого анализа можно оценивать также воздействие небольших молекул на агрегацию Аβ.
Экспериментальная процедура:
Титрационные микропланшеты фирмы NUNC типа Maxisorb (МТП) сенсибилизировали смесью 1:1 Аβ1-40 и Aβ1-42 (по 2 мкМ каждого, 100 мкл на лунку) при 37°С в течение 3 дней. В этих условиях высоко агрегированный фибриллярный Аβ адсорбировался и иммобилизовывался на поверхности лунки. Затем раствор для сенсибилизации удаляли, и планшеты сушили при комнатной температуре в течение 2-4 ч. (Высушенные планшеты можно хранить при -20°С). Сохранившиеся сайты связывания блокировали, добавляя 300 мкл/лунку забуференного фосфатом физиологического раствора, содержащего 0,05% Твин 20 (Т-ЗФР) и 1% бычьего сывороточного альбумина (БСА). После инкубации в течение 1-2 ч при комнатной температуре планшеты промывали 1×300 мкл Т-ЗФР. Добавляли раствор 20 нМ биотинилированного Аβ1-40 в 20 мМ Tris-HCl, 150 мМ NaCl, рН 7,2 (TBS), дополненного 0,05% NaN3, и серийное разбавление антитела (100 мкл/лунку) и планшет инкубировали при 37°С в течение ночи. После трехкратной отмывки 300 мкл Т-ЗФР добавляли (100 мкл/лунку) конъюгат стрептавидин-POD (фирма Roche Molecular Biochemicals), разбавленный 1:1000 в Т-ЗФР, содержащем 1% БСА, и инкубировали при комнатной температуре в течение 2 ч. Лунки промывали трижды Т-ЗФР и добавляли 100 мкл/лунку свежеприготовленного раствора триметилбензидина (ТМБ). [Приготовление раствора ТМБ: 10 мл 30 мМ лимонной кислоты, рН 4,1 (регулирование рН с помощью КОН) + 0,5 мл ТМБ (12 мг ТМБ в 1 мл ацетона + 9 мл метанола) + 0,01 мл 35% Н2O2]. Реакцию прекращали, добавляя 100 мкл/лунку 1н. H2SO4, и абсорбцию оценивали при 450 нм с помощью планшет-ридера для микропланшетов.
Результаты:
Из фиг.8 видно, что антитела MS-Roche IgG1 предупреждали включение биотинилированного Аβ1-40 в сформировавшиеся ранее агрегаты Aβ1-40/Aβ1-42. Способность этих полноразмерных человеческих IgG нейтрализовать Аβ такая же, как у мышиного моноклонального антитела ВАР-1, которое было получено с использованием стандартной процедуры иммунизации и которое специфически распознает аминокислотные остатки 4-6 пептида Аβ при pepspot-анализе, описанном в примере 7. Мышиное моноклональное антитело ВАР-2, которое также дает реакцию только с аминокислотами 4-6 (Brockhaus, неопубликованные данные), обладало существенно более низкой активностью при анализе указанным методом. Еще более низкая активность была обнаружена при использовании специфического в отношении С-конца Аβ1-40 антитела ВАР-17 (Brockhaus, Neuroreport 9, 1998, сс.1481-1486) и моноклонального антитела 4G8, которое распознает эпитоп, расположенный между положениями 17 и 24 в последовательности Аβ (Kim, Neuroscience Research Communication, т. 2, 1988, сс.121-130). БСА в концентрации вплоть до 10 мкг/мл не оказывал влияния на включение биотинилированного Аβ и служил отрицательным контролем. Однако установлено, что БСА в более высоких концентрациях, т.е. >100 мкг/мл ингибировал связывание биотинилированного Аβ со сформировавшимися ранее Аβ-фибриллами (Bohrmann, J Biol Chem, 274 (23), 1999, сс.15990-5), что свидетельствует о том, что взаимодействие БСА с Аβ не является высокоаффинным.
Пример 11: Анализ деполимеризации: высвобождение биотинилированного Аβ из агрегированного Аβ
С помощью аналогичного набора экспериментов оценивали потенциальную способность антитела MS-Roche IgG индуцировать деполимеризацию агрегированного Аβ. Биотинилированный Aβ1-40 первоначально до обработки различными антитела к Аβ включался в сформировавшиеся ранее фибриллы Аβ1-40/Аβ1-42. Высвобождение биотинилированного Аβ оценивали с помощью такого же анализа, который описан для анализа полимеризации.
Экспериментальная процедура:
Титрационные микропланшеты фирмы NUNC типа Maxisorb (МТП) сенсибилизировали смесью 1:1 Aβ1-40 и Аβ1-42 согласно процедуре, описанной для анализа полимеризации. Для включения биотинилированного Аβ сенсибилизированные планшеты инкубировали с 200 мкл/лунку 20 нМ биотинилированного Aβ1-40 в TBS, содержащем 0,05% NaN3, при 37°С в течение ночи. После отмывки планшета с помощью 3×300 мкл/лунку Т-ЗФР, добавляли серийные разведения антител в TBS, содержащем 0,05% NaN3, и инкубировали при 37°С в течение 3 ч. Планшет промывали и анализировали присутствие биотинилированного Аβ1-40 с помощью описанного выше метода.
Результаты:
На фиг.9 А-Г показано, что антитела, предлагаемые в изобретении, индуцировали деполимеризацию агрегированного Аβ, что установлено по высвобождению включенного биотинилированного Aβ1-40. Антитела MS-R и мышиное моноклональное антитело ВАР-1 обладали сопоставимой активностью, в то время как антитела ВАР-2, ВАР-17 и 4G8 обладали существенно более низкой эффективностью в отношении высвобождения биотинилированного Аβ из массы иммобилизованных агрегатов Аβ. ВАР-1 легко можно отдифференцировать от антитела MS-R по его реакционной способности с расположенным на клеточной поверхности полноразмерным АРР (см. фиг.15), и антитела типа ВАР-1, обладающие такими свойствами, не могут найти терапевтическое применение из-за возможных аутоиммунологических реакций, которые они могут индуцировать. Интересно отметить, что антитело ВАР-2, несмотря на его специфичность в отношении аминокислотных остатков 4-6, которые являются доступными в агрегированном Аβ, обладало существенно более низкой активностью в этом опыте, что свидетельствует о том, что не все специфические в отношении N-концов антитела априори обладают одинаковой эффективностью в отношении высвобождения Аβ из сформировавшихся ранее агрегатов. MS-Roche IgG явно превышали ВАР-2 по деполимеризующей активности. Относительно низкая эффективность ВАР-17 (антитело, специфическое в отношении С-конца) и 4G8 (антитело, специфическое в отношении аминокислотных остатков 16-24), установленная в этом опыте, является результатом криптической природы этих двух эпитопов в агрегированном Аβ. Как уже отмечалось ранее при анализе полимеризации, БСА в применяемых концентрациях не оказывал влияния на агрегированный Аβ.
Антитела MS-R, полученные после второго цикла созревания аффинности и после перекрестного клонирования связывающихся агентов, обладали в целом более высокой эффективностью в опыте по оценке деполимеризации (ср. фиг.9А с фиг.9Б и В), что согласуется с повышенной аффинностью к связыванию этих антител (см. таблицы 3-5). Было установлено, что моноклональные антитела AMY-33 и 6F/3D могут препятствовать агрегации Аβ in vitro в определенных экспериментальных условиях (Solomon, Proc. Natl. Acad. Sci. USA, 93, 1996, сс.452-455; антитела AMY-33 и 6F/3D получали от фирмы Zymed Laboratories Inc., Сан-Франциско (заказ №13-0100) и фирмы Dako Diagnostics AG, Зуг, Швейцария (заказ № М087201)). Как видно из фиг.9Г, оба эти антитела совсем не обладали активностью в опыте по оценке деполимеризации.
Пример 12: Анализ эпитопов с помощью ELISA с использованием конъюгатов пептидов
Приведенные ниже гептапептиды (обозначены с помощью однобуквенного кода) получали с помощью твердофазного синтеза и очищали жидкостной хроматографией с использованием известных в данной области методов.
AEFRHDC
EFRHDSC
FRHDSGC
RHDSGYC
HDSGYEC
DSGYEVC
SGYEVHC
YEVHHQC
EVHHQKC
VHHQKLC
HHQKLVC
HQKLVFC
QKLVFFC
KLVFFAC
LVFFAEC
VFFAEDC
FFAEDVC
FAEDVGC
AEDVGSC
EDVGSNC
DVGSNKC
VGSNKGC
GSNKGAC
CSNKGAI
CNKGAII
CKGAIIG
CGLMVGG
CMVGGVV
CGGVVIA
Пептиды растворяли в ДМСО до достижения концентрации 10 мМ. Бычий альбумин (практически свободный от жирных кислот БСА, фирма Sigma, партия 112F-9390) растворяли до достижения концентрации 10 мг/мл в 0,1М бикарбонате натрия и активировали, добавляя на каждый 1 мл по 50 мкл раствора с концентрацией 26 мг/мл N-сукцинимидилмалеинимидопропионата (NSMP, фирма Pierce) в ДМСО. После выдерживания реакционной смеси в течение 15 мин при комнатной температуре активированный БСА очищали гель-фильтрацией (NAP-10, фирма Pharmacia) в ЗФР с 0,1% азида натрия в качестве растворителя. 50 мкл активированного с помощью NSMP БСА (6,7 мг/мл) разбавляли 50 мкл ЗФР, содержащего 0,1% азида натрия, и добавляли 10 мкл раствора пептида (1 мМ в ДМСО). Для получения отрицательного контроля активированную БСА обрабатывали имитатором без добавления пептида. После выдерживания в течение 4 ч при комнатной температуре реакцию прекращали, добавляя 10 мкл 10 мМ цистеина. Аликвотные количества содержащей конъюгат смеси разбавляли в соотношении 1:100 0,1М натрий-бикарбонатным буфером и сразу же вносили в лунки (100 мкл) ELISA-планшетов (тип Nunc Immuno-Plate). После выстаивания в течение 16 ч при 4°С в каждую лунку добавляли 100 мкл блокирующего буфера (описанного выше) и инкубировали в течение еще 30 мин. Планшеты промывали, используя 2×300 мкл/лунку TBST (описанного выше), и вносили 100 мкл антитела в концентрации 10 мкг/мл или 2 мкг/мл в блокирующем буфере. Планшеты выдерживали в течение 16 ч при 4°С и промывали, используя 2×300 мкл TBST. Добавляли 100 мкл/лунку конъюгированного с HRP антитела к человеческому Ig H+L (фирма Pierce, разбавление блокирующим буфером 1:1000) и инкубировали в течение 1 ч при температуре окружающей среды. Планшеты промывали, используя 3×300 мкл/лунку TBST. Появление окраски начиналось после добавления 100 мкл реагента, представляющего собой тетраметилбензидин /перекись водорода. Реакцию прекращали через 5 мин, добавляя 100 мкл/лунку 1М серной кислоты, и оценивали оптическую плотность с помощью оптического ридера (Microplate Reader 3550, фирма BioRad) при 450 нм. Для сравнения аналогичным образом анализировали мышиные моноклональные антитела, за исключением того, что в качестве проявляющего агента применяли меченое с помощью HRP антитело к мышиному Ig вместо антитела к человеческому Ig.
На основе специфичности описанных выше гептапептидов, полученных из AJ3, осуществляли описанные выше анализы ELISA. Предпочтительно антитела, предлагаемые в изобретении, представляли собой антитела, для которых при оценке по оптической плотности соотношение их сигнала и фонового сигнала превышало «10», когда их реактивность в отношении полученного из А-бета пептида (AEFRHD; аминокислоты 2-7 пептида А-бета) сравнивали с неродственным протеином/пептидом типа БСА. Наиболее предпочтительно соотношение оптических плотностей превышало «5» для соответствующей реакции с использованием по меньшей мере одного из перечисленных ниже трех полученных из Аβ пептидов: (VFFAED; аминокислоты 18-23 Аβ) или (FFAEDV; аминокислоты 19-24 Аβ) или (LVFFAE; аминокислоты 17-22 Aβ). Соответствующие результаты для предлагаемых в изобретении родительских и/или созревших антител, представлены далее в двух таблицах:
Таблица 6: Реакционная способность MS-R Fab-фрагментов в отношении конъюгированных с БСА А-бета гептапептидов 2-7 (AEFRHD), 17-22 (LVFFAE), 18-23 (VFFAED) и 19-24 (FFAEDV).
Представлены соотношения при анализе с помощью ELISA (оптическая плотность), полученные с использованием конъюгированных с БСА пептидов и неконъюгированных пептидов. Также представлены данные об интенсивности сигналов при использовании пептидов 17-22, 18-23 и 19-24 по сравнению с пептидом 2-7.
№
2-7/БСА
17-22/БСА
18-23/БСА
19-
24/БСА
17-22/2-7
18-23/2-7
19-24/2-7
8.2.H2×8.2.Ll
13
20
25
25
1,54
1,92
1,92
Мат
Таблица 7: Реакционная способность MS-R IgG и мышиных моноклональных антител ВАР-1, ВАР-2, 4G8, 6Е10 Amy-33 и 6F/3D в отношении конъюгированных с БСА Аβ-гептапептидов 2-7 (AEFRHD), 17-22 (LVFFAE), 18-23 (VFFAED) и 19-24 (FFAEDV).
Представлены соотношения при анализе с помощью ELISA (оптическая плотность), полученные с использованием конъюгированных с БСА пептидов и неконъюгированных пептидов. Также представлены данные об интенсивности сигналов при использовании пептидов 17-22, 18-23 и 19-24 по сравнению с пептидом 2-7.
*Это антитело является специфическим для последовательности 8-17 и не распознает последовательности N-концевых или срединных эпитопов.
Пример 13: Объединение оптимизированных H-CDR2 и L-CDR1 посредством перекрестного клонирования
Модульное конструирование библиотеки HuCAL позволяет осуществлять замену гипервариабельных участков (CDR) двух различных кодирующих Fab генов с помощью одной стадии клонирования. Для дополнительного улучшения аффинности объединяли независимо оптимизированные клоны H-CDR2 и L-CDR1, полученные из зрелых клонов, с этими же H-CDR3, поскольку существовала высокая вероятность того, что это объединение может приводить к дополнительному повышению аффинности (Yang и др., J.Mol.Biol. 254, 1995, сс.392-403; Schier и др., J.Mol.Biol. 263, 19966, сс.551-567; Chen и др., J.Mol.Biol. 293, 1999, сс.865-881). Полные легкие цепи или их фрагменты переносили из клона-донора с оптимизированным L-CDR1 в клон-реципиент с оптимизированным H-CDR2. Клон-донор и клон-реципиент объединяли только тогда, когда они оба несли идентичные H-CDR3-последовательности. Все клоны-доноры и клоны-реципиенты несли каркасный участок VН3-Vκ3.
Процедуру осуществляли путем переноса полных легких цепей из клона-донора с оптимизированным L-CDR1 в клон-реципиент с оптимизированным Н-CDR2. Специфичность в отношении эпитопа сохранялась только при объединении клонов с одинаковыми H-CDR3. При замене оптимизированного Н-CDR2-клона легкой цепи получали только клон с оптимизированным L-CDR1, если этот обмен осуществляли между клонами с одинаковыми L-CDR3. Если L-CDR3 клонов, подлежащих объединению, были различными, то клон с оптимизированным H-CDR2 приобретал помимо оптимизированного L-CDR1 другой L-CDR3 (L-CDR2 оставался консенсусной последовательностью в HuCAL (Knappik и др., 2000)), а при использовании в качестве доноров легкой цепи производных MS-Roche №7.12 L-CDR1, 2 и 3 происходила замена в клоне-акцепторе с оптимизированным H-CDR2. Применяли три различные стратегии клонирования:
1) с использованием рестриктаз XbaI и SphI фрагмент легкой цепи антитела вырезали из плазмиды 1 (например, pM×9_Fab_MS-Roche №7.11.H1_FS) и в полученный таким образом каркас вектора затем встраивали путем лигирования фрагмент легкой цепи плазмиды 2 (например, pMx9_Fab_MS-Roche №7.2.L1_FS), полученной с помощью расщепления с использованием Xbal и SphI. Тем самым создавали новую плазмиду (обозначена как: pM×9_Fab_MS-Roche №7.11.H1×7.2.L1_FS), кодирующую L-CDR1,2,3 родительского клона №7.2.L1 и H-CDR1,2,3 родительского клона №7.11.H1;
2) с использованием рестриктаз XbaI и Асс65I фрагмент, кодирующий L-CDR1, вырезали из плазмиды 1 (например, pM×9_Fab_MS-Roche №7.11.H2_FS) и в полученный тем самым каркас вектора встраивали путем лигирования L-CDR1 -фрагмент плазмиды 2 (например, pM×9_Fab_MS-Roche №7.12.Ll_FS), полученной с помощью расщепления с использованием Xbal и Асс65I. Тем самым создавали новую плазмиду (обозначена как: pM×9_Fab_MS-Roche №7.11.H2×7.12.L1(L-CDR1)_FS), кодирующую L-CDR1 родительского клона №7.12.L1, в то время как L-CDR2,3 и H-CDR1,2,3 получали из родительского клона №7.11.Н2;
3) с использованием рестриктаз XbaI и BamHI фрагмент, кодирующий L-CDR1 и L-CDR2, вырезали из плазмиды 1 (например, pMx9_Fab_MS-Roche №7.11.H2_FS) и в полученный тем самым каркас вектора встраивали путем лигирования L-CDR1- и L-СDR2-фрагмент плазмиды 2 (например, pM×9_Fab_MS-Roche №7.12.L1_FS), полученной с помощью расщепления с использованием XbaI и BamHI. Тем самым получали новую плазмиду (обозначена как: pM×9_Fab_MS-Roche №7.11.H2×7.12.L1(L-CDR1+2)_FS), кодирующую L-CDR1 и L-CDR2 родительского клона №7.12.L1, в то время как L-CDR3 и H-CDR1,2,3 получали из родительского клона №7.11.Н2.
Приведенные с целью иллюстрации примеры различных стратегий клонирования, а также последовательности клонов-доноров и клонов-реципиентов представлены в таблице 8.
После крупномасштабной экспрессии и очистки определяли их аффинность к фибриллам Аβ (1-40). Кроме того, ниже в таблице 9 представлены значения КD для отобранных полученных перекрестным клонированием антител MS-R Fab.
Таблица 9: Значения KD, характеризующие связывание полученных перекрестным клонированием MS-R Fab с фибриллами Аβ1-40 и Аβ1-42, которые оценивали с помощью Biacore. Получение перекрестным клонированием Fab-фрагментов описано в примере 13. Значения KD определяли путем аппроксимации кинетических кривых и коррекции с учетом содержания активного Fab в каждом образце согласно методу, представленному в описании. Некоторые Fab-фрагменты дополнительно очищали с помощью гель-хроматографии или препаративного ультрацентрифугирования для удаления агрегированного материала. (L1) обозначает несущий зрелый H-CDR2 клон-акцептор, в который встроен L-CDR1 только из клона-донора с оптимизированным L-CDR1; (L1+2) обозначает несущий зрелый H-CDR2 клон-акцептор, в который встроен L-CDR1+2 из клона-донора с оптимизированным L-CDR1.
Пример 14: Выявление in vivo амилоидных бляшек на мышиной модели болезни Альцгеймера с помощью конфокальной лазерной сканирующей микроскопии и анализа совместной локализации
Отобранные антитела MS-R IgG1 тестировали на мышах, несущих двойной трансген APP/PS2 (ссылка: Richards и др., Soc. Newosci. Abstr., том. 27, программа №5467, 2001) в отношении способности выявлять амилоидные бляшки in vivo. Антитела (1 мг/мышь) вводили i.v. и через 3 дня осуществляли перфузию головного мозга физиологическим раствором и подготавливали для изготовления криостатных срезов. В другом исследовании мышей обрабатывали более высокими концентрациями антител, т.е. инъецировали i.v. 2 мг в дни 0, 3 и 6, и животных умерщвляли на девятый день. Присутствие связанного с амилоидными бляшкам антитела оценивали на нефиксированных криостатных срезах с помощью непрямой иммунофлуоресценции с двойной меткой, используя козье антитело к человеческому IgG (H+L), конъюгированное с любым Су3 (№109-165-003, фирма Jackson Immuno Research), а затем иммуноконъюгат ВАР-2-А1еха488. Для визуализации применяли конфокальную лазерную микроскопию и изображение обрабатывали для количественной оценки совместной локализации с помощью пакета программ IMARIS и CO - LOCALIZATION (фирма Bitplane, Швейцария). Репрезентативные примеры представлены на фиг.10-14. Было обнаружено, что все изученные антитела MS-R являлись позитивными с точки зрения иммунного окрашивания амилоидных бляшек in vivo, хотя при этом была обнаружена определенная вариабельность.
Пример 15: Изучение связывания различных моноклональных антител с амилоидным протеином-предшественником (АРР) на поверхности НЕК293-клеток
АРР экспрессируется в широком разнообразии структур центральной нервной системы. Связывание антитела с расположенным на клеточной поверхности АРР может приводить к активации комплемента и разрушению клеток в здоровых областях головного мозга. Поэтому для терапевтического применения антител к А-бета обязательным является отсутствие реактивности в отношении АРР. Обладающие высокой аффинностью антитела к N-концевому домену А-бета (например, ВАР-1, ВАР-2) распознают также соответствующий эпитоп в каркасном участке АРР. В противоположность этому неожиданно было установлено, что антитела к срединному эпитопу (например, 4G8) и антитела, предлагаемые в изобретении, не обладают способностью распознавать находящийся на клеточной поверхности АРР. Таким образом, антитела, предлагаемые в изобретении, которые окрашивают А-бета, но не окрашивают APP in vivo, являются более предпочтительными по сравнению с неизбирательными антителами.
Метод проточной цитометрии хорошо известен в данной области. Относительные единицы флуоресценции (FL1-H), оцененные с помощью проточной цитометрии, характеризуют связывание с клеточной поверхностью соответствующего антитела. Сдвиг флуоресценции на поверхности клеток трансфектированной APP линии НЕК293 по сравнению с нетрансфектированными НЕК293-клетками свидетельствует о нежелательной реакции с расположенным на клеточной поверхности APP. Например, при оценке антител к N-концевому домену ВАР-1 и ВАР-2 был выявлен значительный сдвиг FL-1-сигнала в линии НЕК293/АРР (жирная линия) по сравнению с нетрансфектированным НЕК293-клетками (пунктирная линия). При оценке антитела 4G8 (специфического для срединного эпитопа А-бета) и всех антител, предлагаемых в изобретении (специфических в отношении N-концевого и срединного эпитопов А-бета), не выявлено никакого заметного сдвига флуоресценции. Различия в основном уровне флуоресценции между клетками НЕ293/АРР и НЕК293 связаны с различным размером клеток. Применяли устройство FACScan в сочетании с пакетом программ Cellquest Pro Software package (оба фирмы Becton Dickinson).
Пример 16: Перечень идентифицированных SEQ ID NO, связанных с молекулами антител, предлагаемыми в изобретении
Ниже в таблице 10 представлены последовательности, которые выявлены для некоторых специфических молекул антител, предлагаемых в изобретении.
Таблица 10: Идентификация SEQ ID NO родительских антител, а также оптимизированных зрелых и/или полученных перекрестным клонированием молекул антител
прот.






































































































































































| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К БЕТА-АМИЛОИДУ | 2013 |
|
RU2651486C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ АМИЛОИДА-БЕТА ПРИ ГЛАЗНЫХ ЗАБОЛЕВАНИЯХ | 2008 |
|
RU2571859C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К АМИЛОИДУ БЕТА | 2007 |
|
RU2498999C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К АМИЛОИДУ БЕТА | 2013 |
|
RU2668161C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К АМИЛОИДУ БЕТА | 2008 |
|
RU2567151C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ АМИЛОИДА БЕТА ПРИ ГЛАЗНЫХ ЗАБОЛЕВАНИЯХ | 2008 |
|
RU2542967C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО | 2008 |
|
RU2538709C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ПРОТИВ АМИЛОИДА БЕТА ПРИ ГЛАЗНЫХ ЗАБОЛЕВАНИЯХ | 2008 |
|
RU2604181C2 |
| БЕЗОПАСНЫЕ И ФУНКЦИОНАЛЬНЫЕ ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА | 2011 |
|
RU2607368C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИ-CD38-АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2402568C2 |
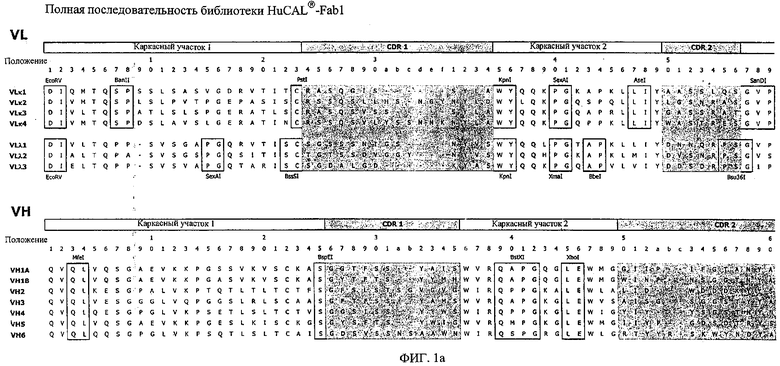
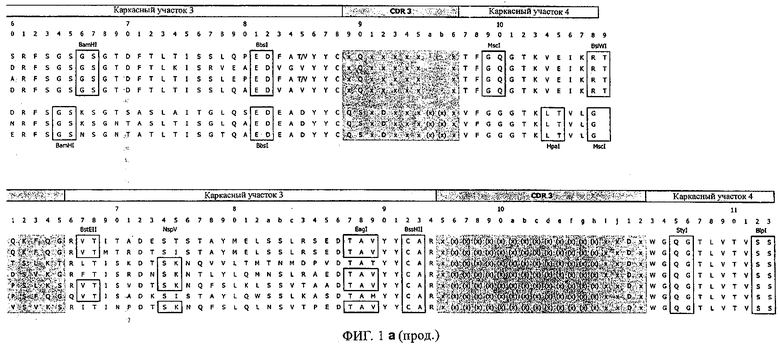

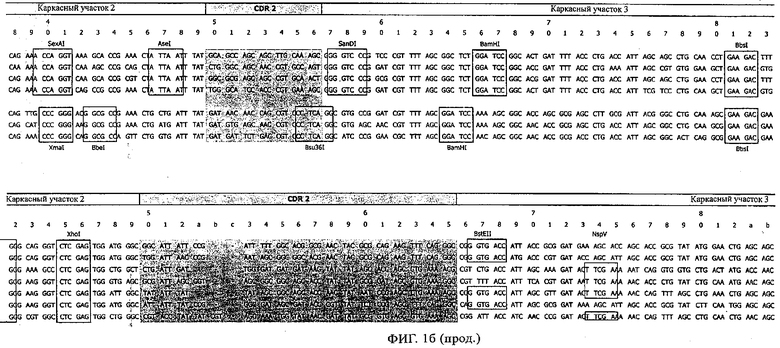
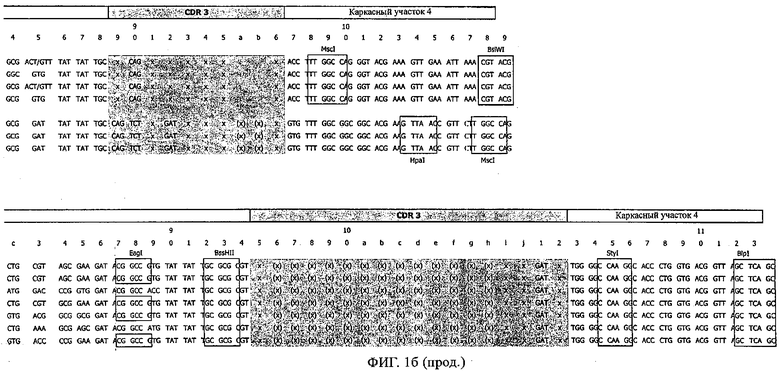
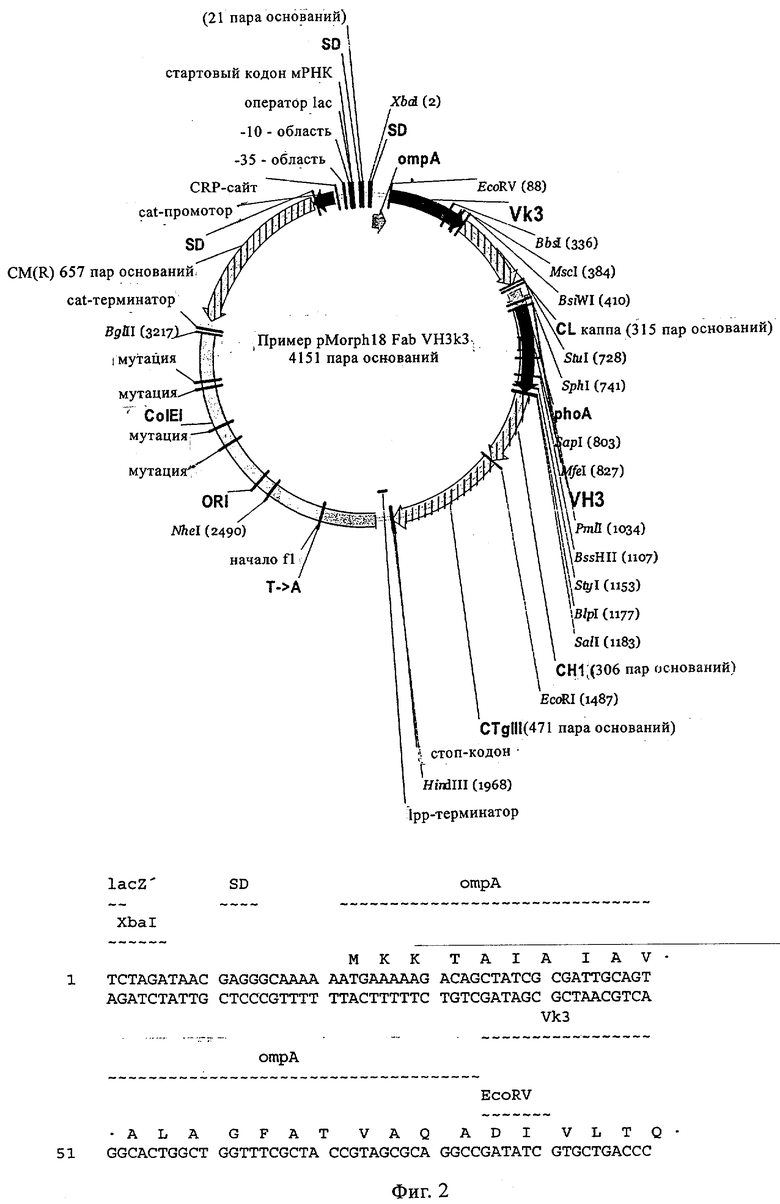


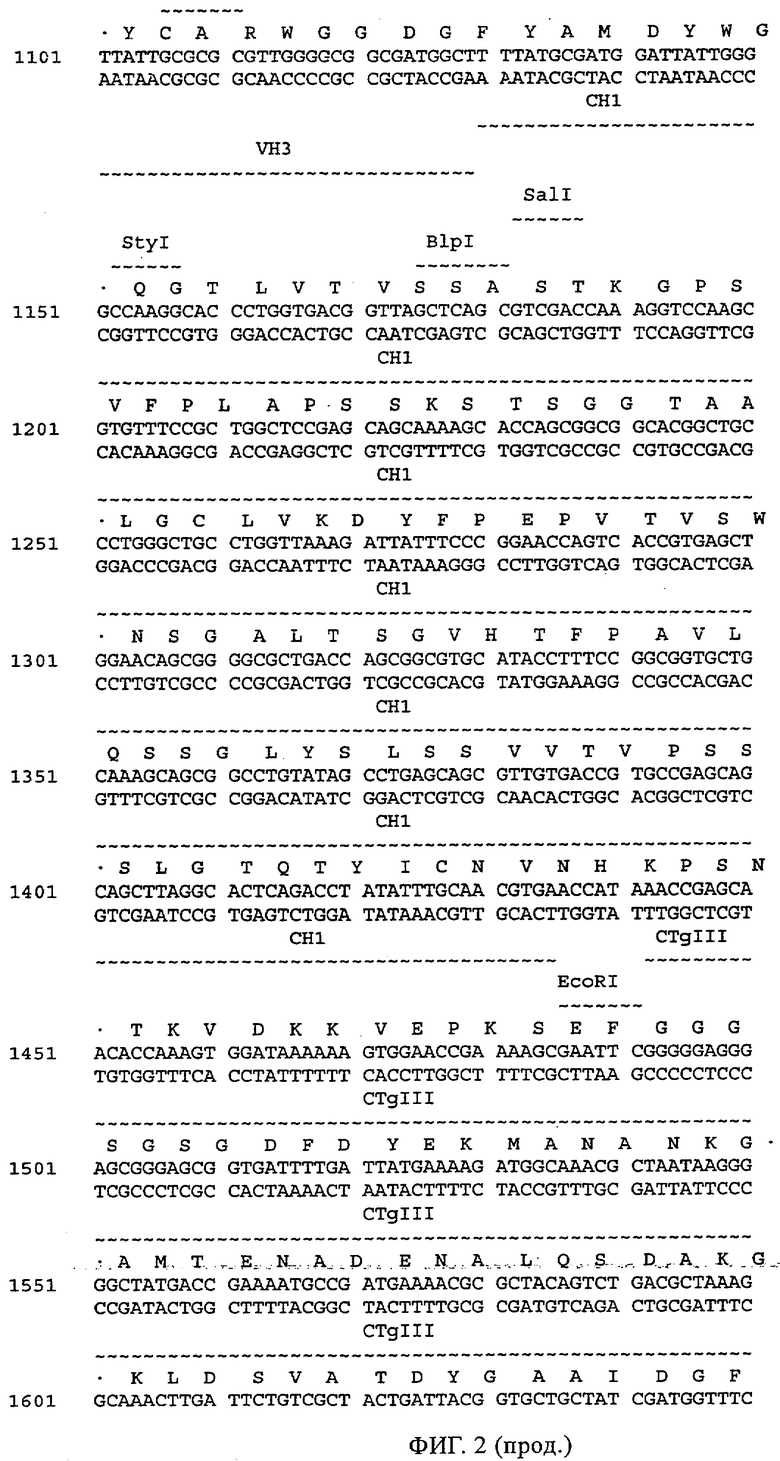





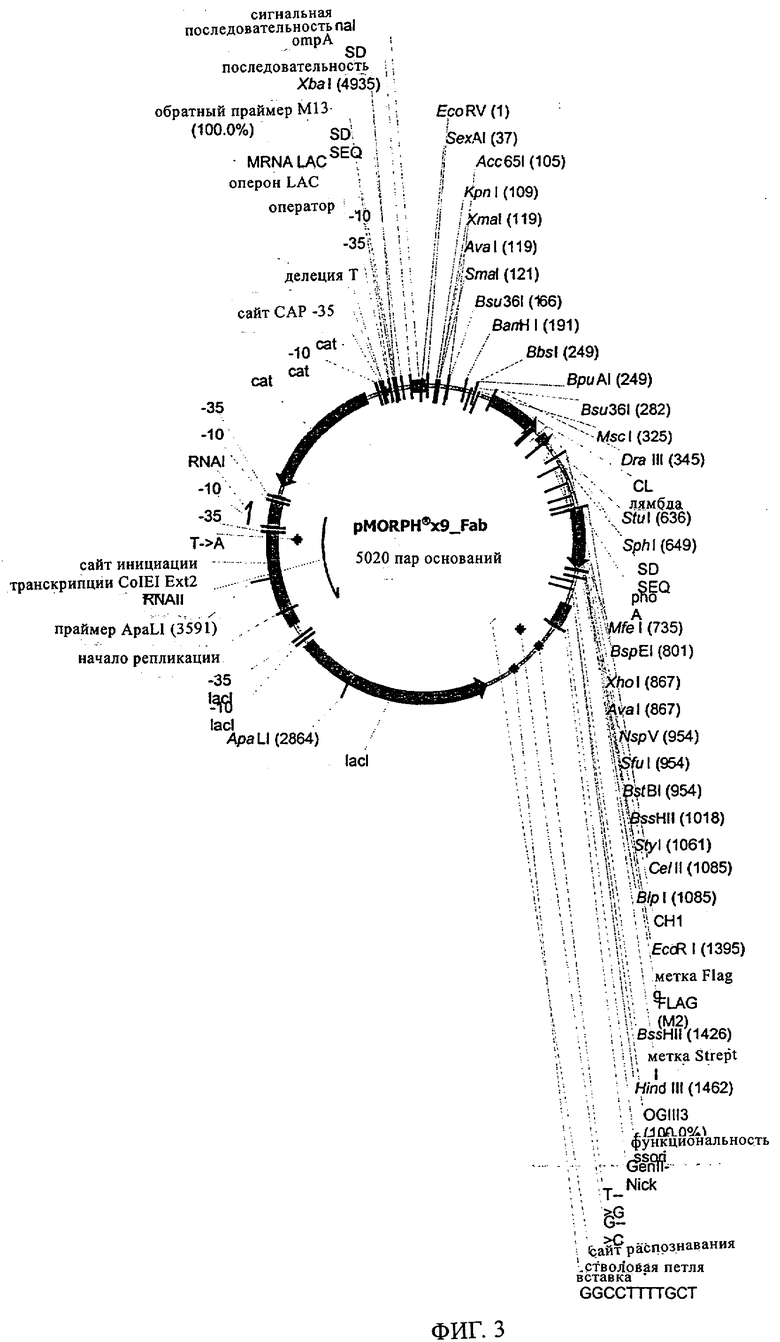




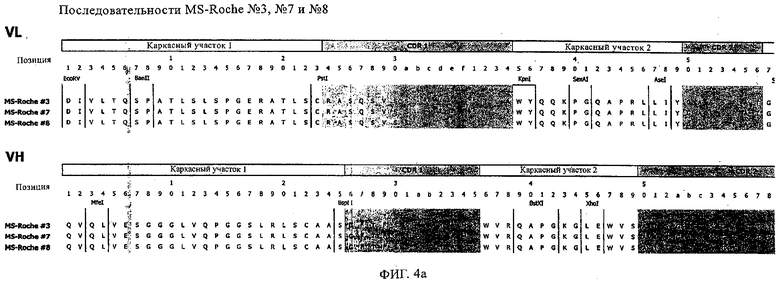





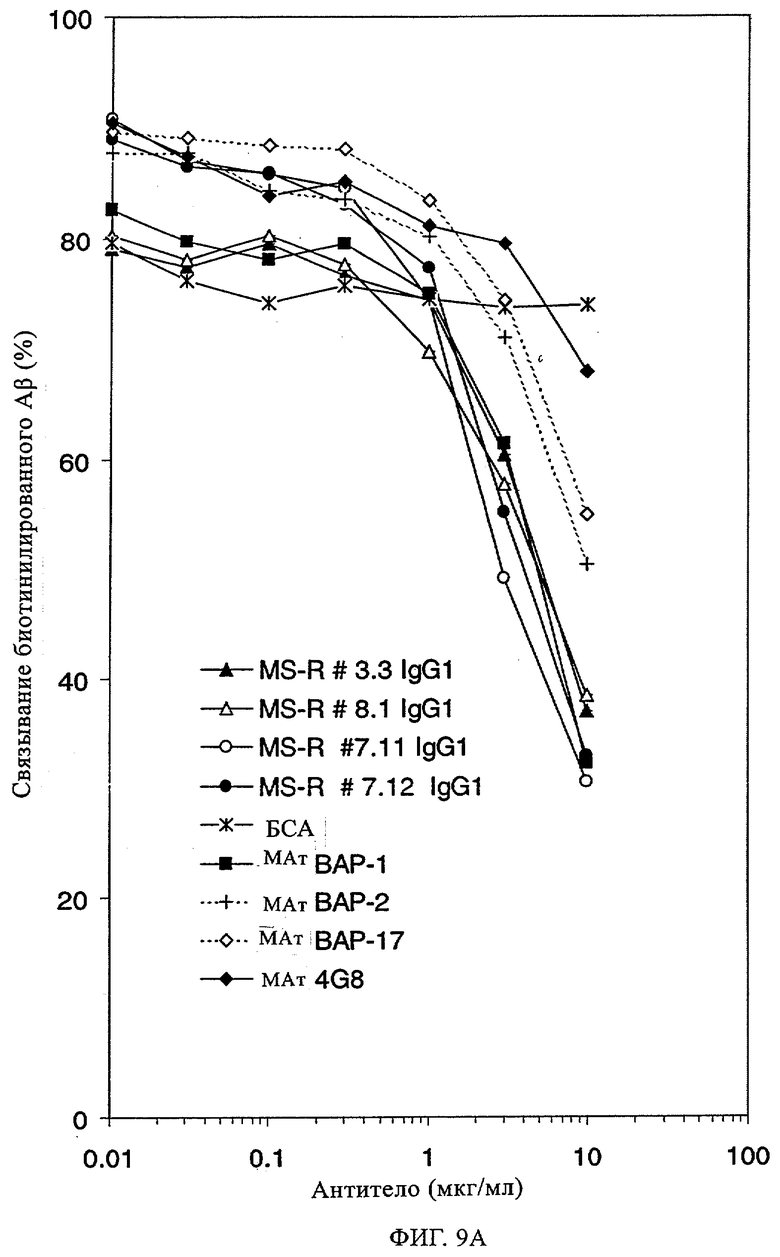
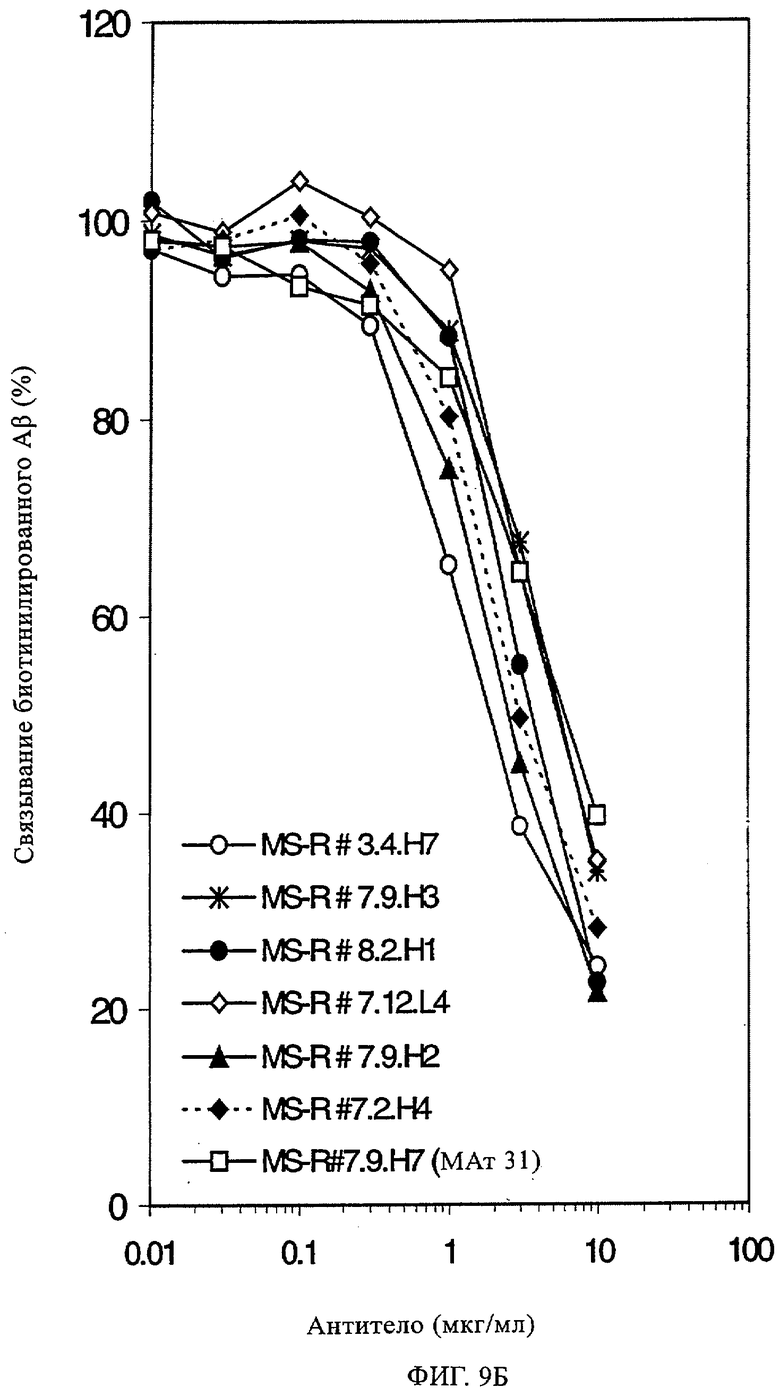
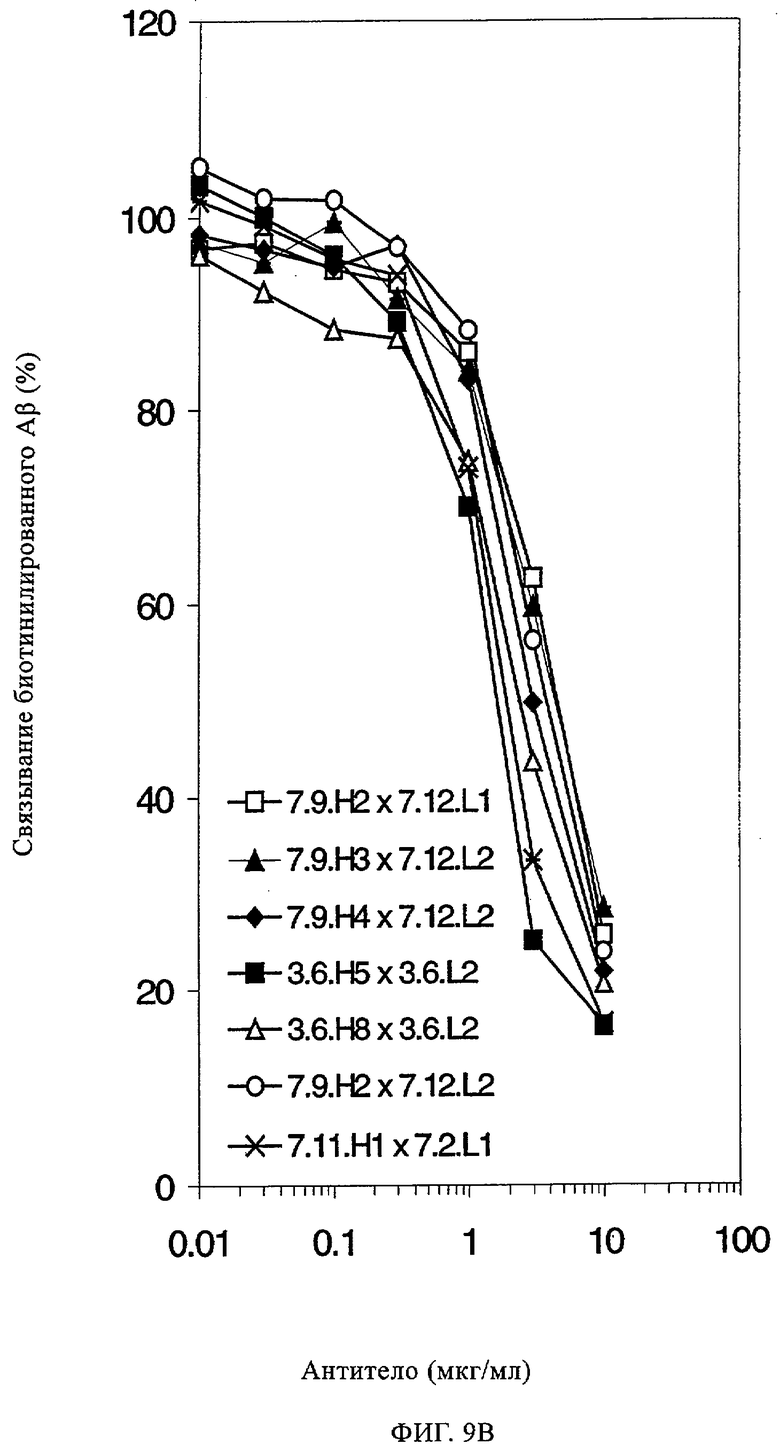
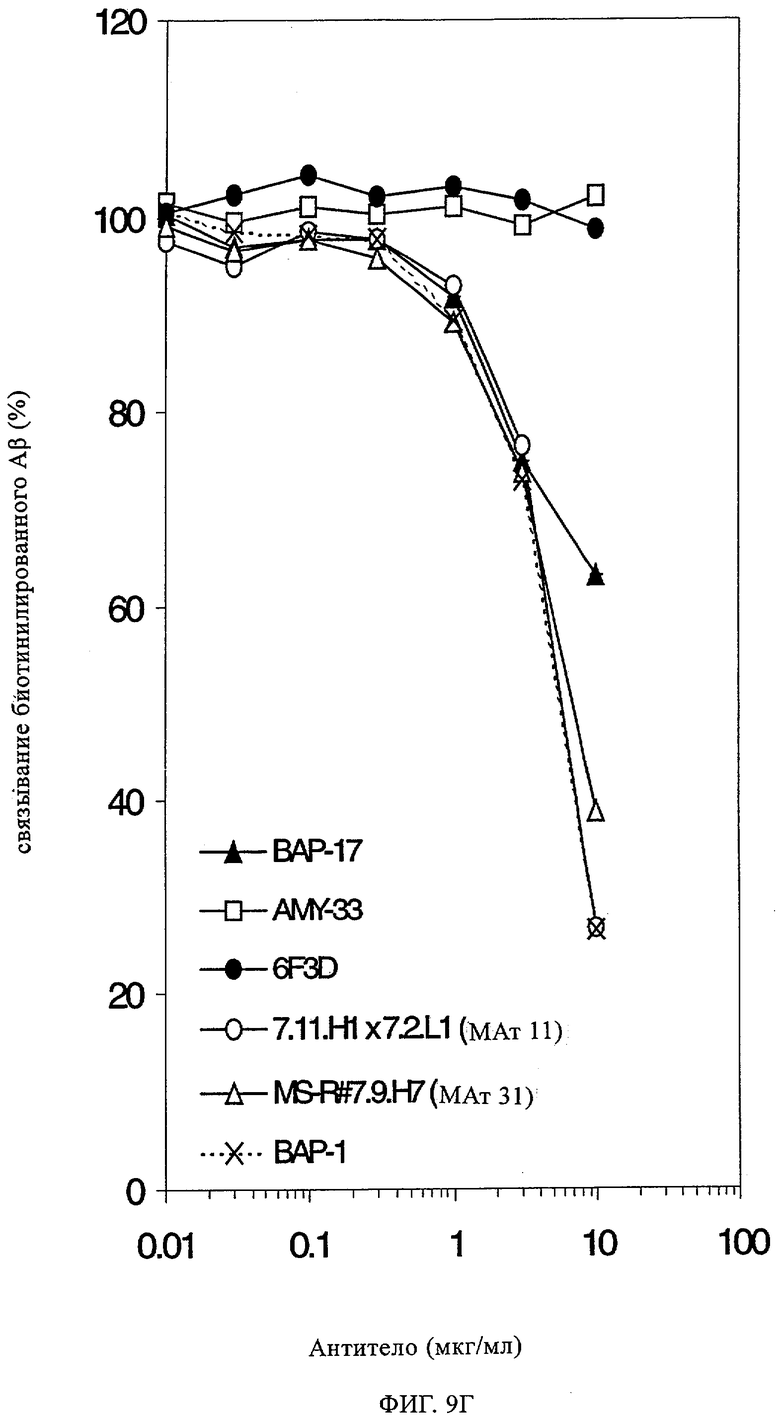
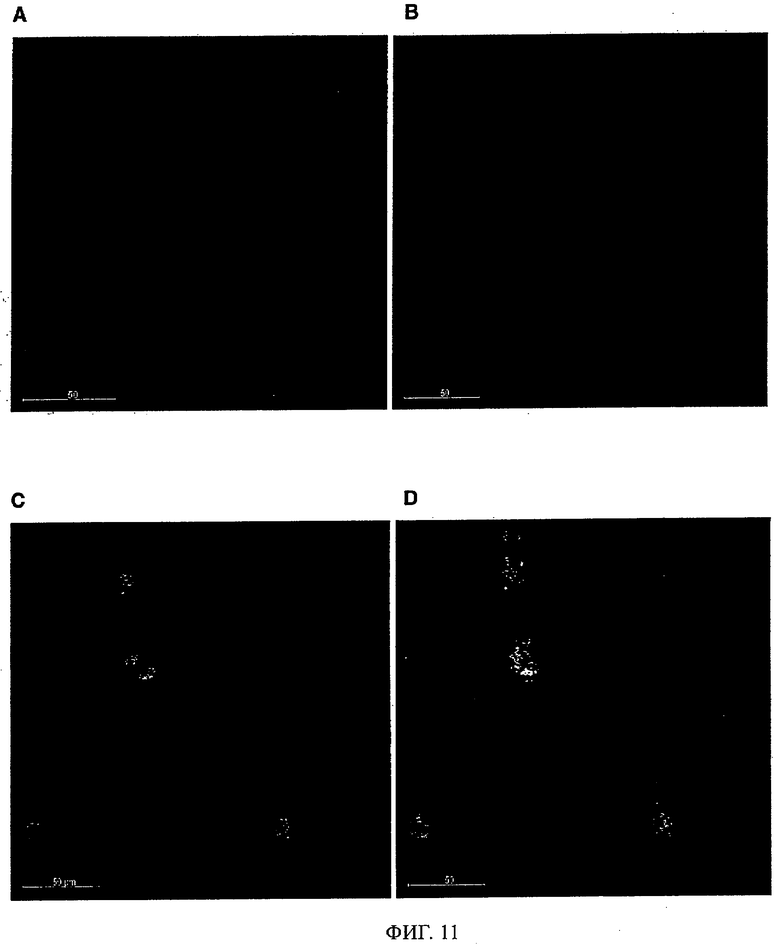
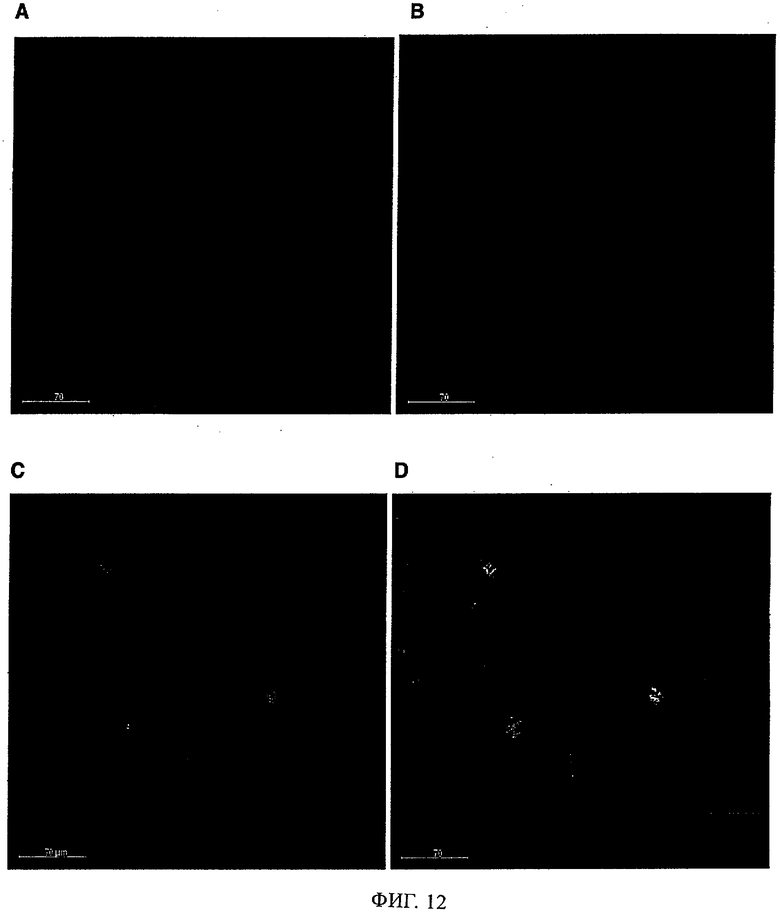
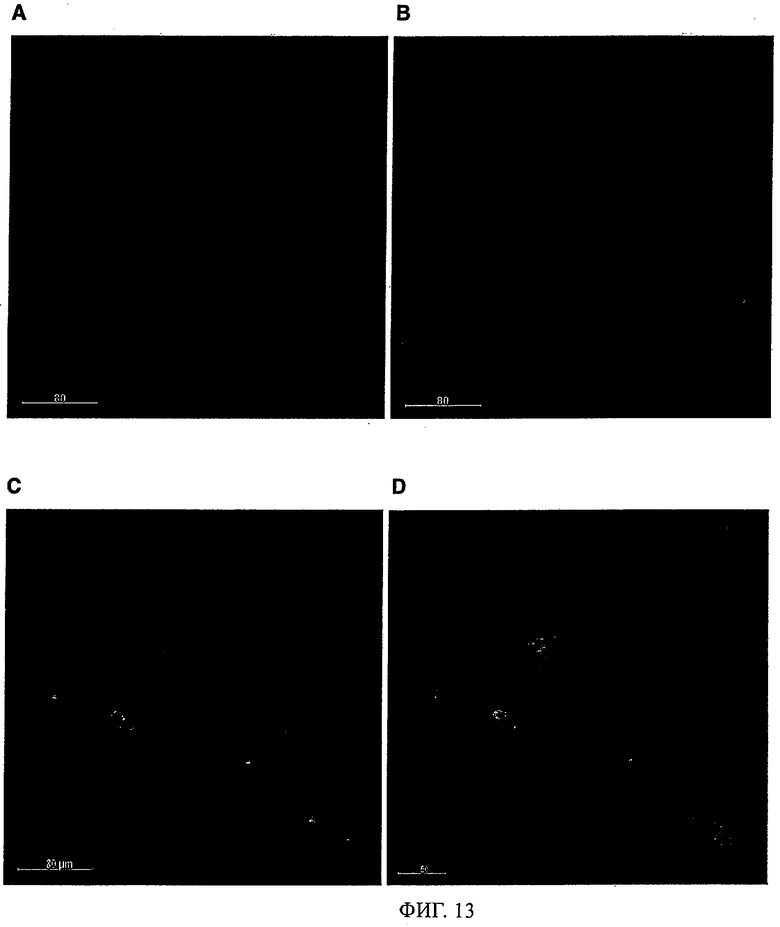

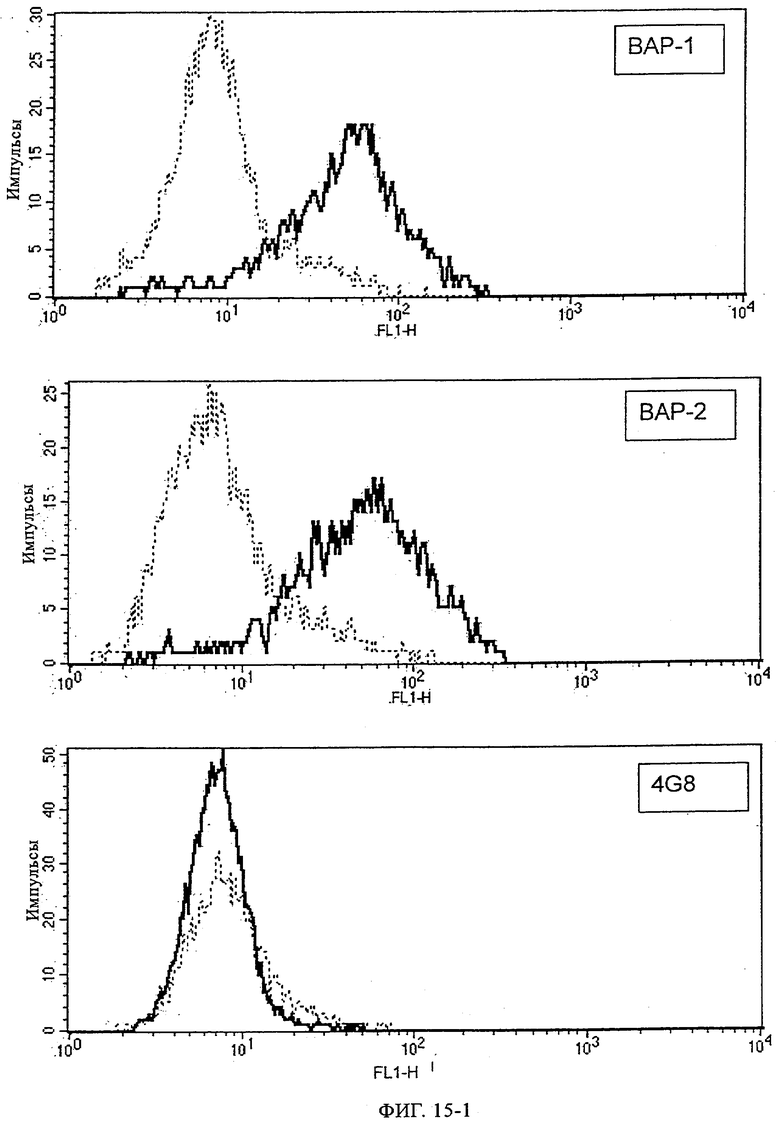
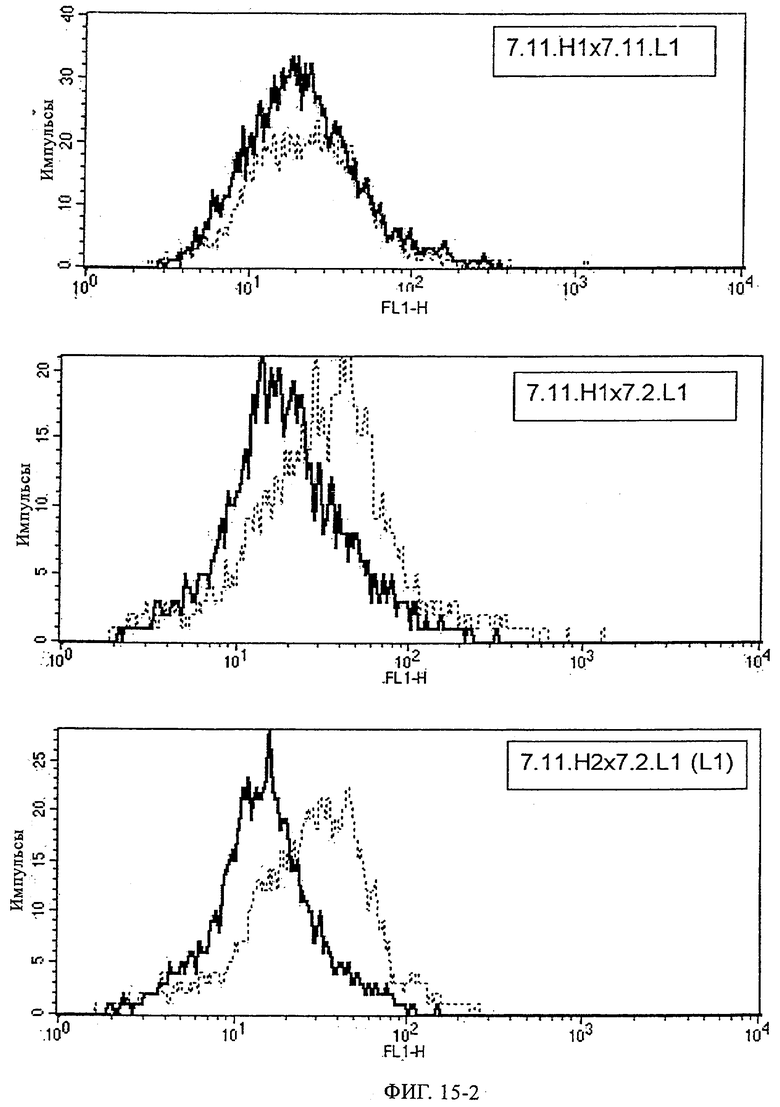
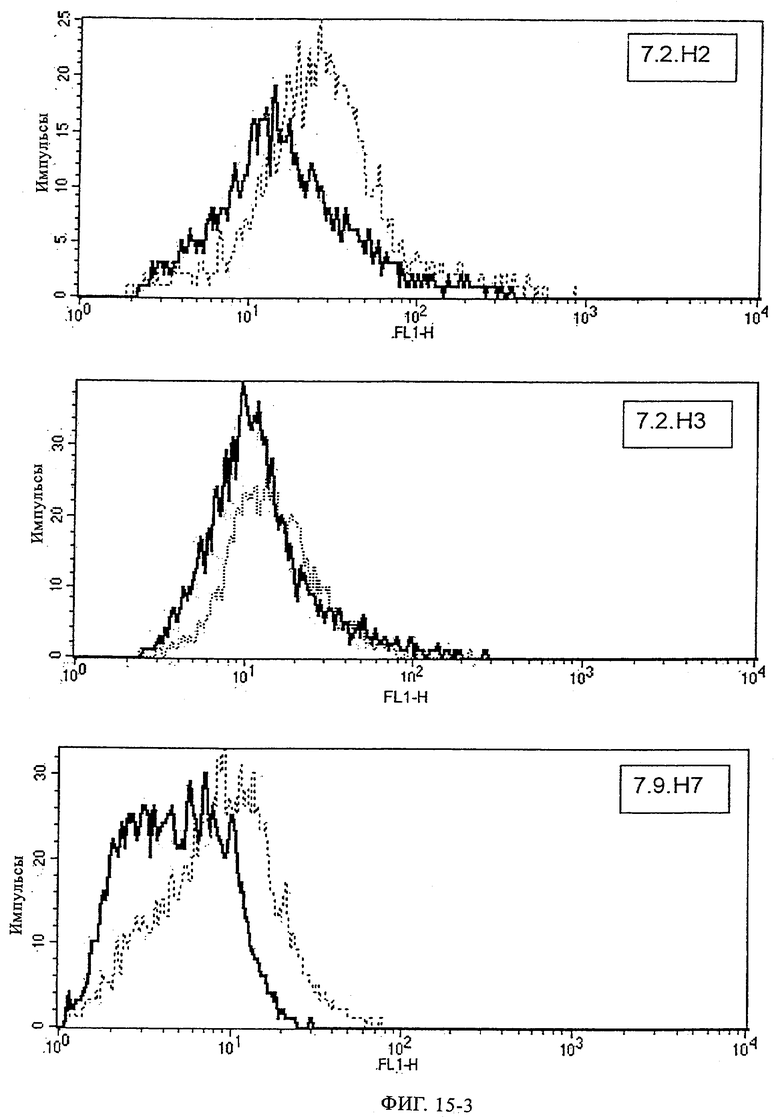
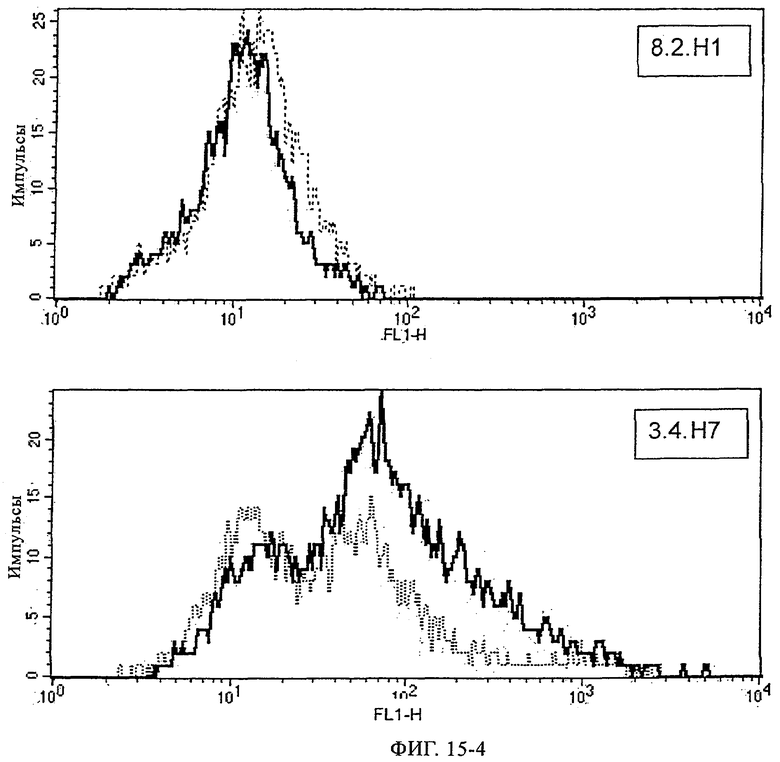
Настоящее изобретение относится к области биотехнологии. Предложены варианты антител, специфически распознающие две области пептида β-A4, характеризующиеся перечнем аминокислотных остатков. Первая область распознаваемого пептида содержит аминокислотную последовательность AEFRHDSGY или ее фрагмент, а вторая область содержит аминокислотную последовательность VHHQKLVFFAEDVG или ее фрагмент. Описаны молекулы нуклеиновых кислот, кодирующие молекулы антител, предлагаемые в изобретении, векторы и хозяева, которые содержат указанные молекулы нуклеиновых кислот. Раскрыты способы получения и оптимизации антител, фармацевтические композиции на основе антител и способ их получения, а также набор на основе антител и различные применения антител. Использование изобретения обеспечивает высокоспецифичные антитела к пептиду β-А4, что может найти применение для диагностики различных заболеваний, опосредованных пептидом β-А4. 14 н. и 15 з.п. ф-лы, 15 ил., 10 табл.
(а) аминокислотную последовательность, содержащую AEFRHD, EF, EFR, FR, EFRHDSG, EFRHD или HDSG в указанной первой области β-А4; и
(б) аминокислотную последовательность, содержащую HHQKL, LV, LVFFAE, VFFAED или VFFA, FFAEDV в указанной второй области.
MS-R 3.6H5×3.6L2;
MS-R 3.6H8×3.6L2;
MS-R 7.4H2×7.2L1;
MS-R 7.9H2×7.12L2;
MS-R 7.9H4×7.12L2;
MS-R 7.11H1×7.11L1;
MS-R 7.11H1×7.2L1;
MS-R 3.3H1×3.4L1;
MS-R 3.4H1×3.4L9;
MS-R 3.4H3×3.4L7;
MS-R 3.4H3×3.4L9;
MS-R 3.4H7×3.4L9;
MS-R 3.4H7×3.4L7;
MS-R 3.6H5×3.6L1;
MS-R 7.2H2×7.2L1;
MS-R 7.4H2×7.12L2;
MS-R 7.9H2×7.2L1;
MS-R 7.9H2×7.12L1;
MS-R 7.11H2×7.2L1;
MS-R 7.11H2×7.9L1;
MS-R 7.11H2×7.12L1 или
MS-R 8.1H1×8.2L1,
как указано в табл.10.
(а) конструируют библиотеку разнообразных Fab-фрагментов антитела, полученных из антитела, которое несет по меньшей мере один CDR3 VH-области, кодируемый молекулой нуклеиновой кислоты, представленной в SEQ ID NO:21, 23 или 25, или меньшей мере одну аминокислотную последовательность CDR3 VH-области, представленную в SEQ ID NO:22, 24 или 26;
(б) тестируют полученную оптимизированную Fab-библиотеку с помощью пэннинга через Aβ/Aβ4;
(в) идентифицируют оптимизированные клоны и
(г) экспрессируют отобранные оптимизированные клоны.
(а) оптимизируют антитело с помощью способа по одному из пп.24-27 и
(б) приготавливают композицию, включающую оптимизированное(ую) антитело/молекулу антитела и физиологически приемлемый носитель.
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Способ осветления мыла из контакта Петрова | 1947 |
|
SU72880A1 |
| FRENKEL et al | |||
| "Immunization against Alzheimer′s beta -amyloid plaques via EFRH phage administration", Proc Natl | |||
| Acad | |||
| Sci | |||
| USA, 2000; 97(21), реферат | |||
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ С ФАКТОРОМ РОСТА НЕРВА | 1994 |
|
RU2126265C1 |

