Настоящее изобретение претендует на приоритет по следующим предварительным патентным заявкам: №60/541911, поданной 6 февраля 2004 года, №60/547584, поданной 26 февраля 2004 года, №60/553948, поданной 18 марта 2004 года, и №60/599014, поданной 6 августа 2004 года, которые включены в настоящее описание в виде ссылок во всей их полноте.
Предшествующий уровень техники
CD38 представляет собой мембранный гликопротеид второго типа, относящийся к семейству эктоэнзимов благодаря своей ферментативной активности в качестве АДФ-рибозил-циклазы и цАДФ-гидролазы. В ходе онтогенеза CD38 появляется на CD34+ коммитированных стволовых клетках и линиеспецифических коммитированных клетках-предшественниках лимфоидных, эритроидных и миелоидных клеток. Считается, что экспрессия CD38 сохраняется только в лимфоидных клетках-предшественниках в течение ранних стадий развития T- и В-лимфоцитов.
Повышенная регуляция CD38 служит маркером активации лимфоцитов, в частности дифференцировки В-лимфоцитов в плазмоциты. (Ко)рецепторные функции рецептора CD38, ведущего к внутриклеточной передаче сигналов или межклеточной коммуникации через его лиганд СD31, так же как и его роль в качестве внутриклеточного регулятора второго мессенджера, циклической АДФ-рибозы (АДФ-р), постулируются в разнообразии каскадов передачи сигналов. Однако необходимо пояснить его физиологическое значение, начиная с нокаута его мышиного аналога или человеческих анти-CD38-аутоантител, не проявляющих вредных свойств.
Кроме наблюдения его экспрессии в гематопоэтической системе, исследователи отметили повышающую сверхрегуляцию CD38 в различных клеточных линиях, полученных из B-, T- и миелоидных/моноцитарных опухолей, включая B- или Т-клеточный острый лимфобластный лейкоз (ОЛЛ), острый миелоидный лейкоз (ОМЛ), неходжкинскую лимфому (НХЛ) и множественную миелому (ММ). Например, выраженная экспрессия CD38 доказана у большинства из всех наблюдавшихся пациентов с ММ.
Следовательно, сверхэкспрессия CD38 в злокачественных клетках обеспечивает выгодную терапевтическую мишень для иммунотерапии. Особый интерес вызывает тот факт, что наиболее примитивные полипотентные стволовые клетки гематопоэтической системы являются CD38-негативными, и что величина цитотоксических эффектов антителозависимой клеточно-опосредованной цитотоксичности ADCC или комплементзависимой цитотоксичности CDC хорошо коррелирует с уровнями экспрессии соответствующей мишени.
Современные подходы к анти-CD38-терапии можно разделить на две группы, представляющие собой подходы in vivo и ex vivo. При подходе in vivo анти-CD38-антитела вводят нуждающемуся в терапии субъекту, с тем чтобы вызвать антитело-опосредованное истощение злокачественных клеток, сверхэкспрессирующих CD38. Истощение может достигаться путем или ADCC, и/или CDC с помощью клеток-эффекторов или путем использования анти-CD38-антител в качестве направленных функциональных групп для транспорта цитотоксических веществ, например сапорина, к клеткам-мишеням, и последующей интернализации. При подходе ех vivo у нуждающегося в терапии субъекта удаляют клеточную популяцию, например клетки костного мозга, содержащие CD38-свехэкспрессирующие злокачественные клетки, и обеспечивают их контакт с анти-CD38-антителами. Клетки-мишени или разрушают цитотоксическими веществами, например сапорином, как описано в подходе in vivo, или удаляют путем приведения в контакт с клеточной популяцией с иммобилизованными анти-CD38-антителами, таким образом, удаляя свехэкспрессирующие CD38-клетки-мишени из смеси. После этого истощенную клеточную популяцию обратно вводят пациенту.
В зависимости от различных свойств антитела, специфические в отношении CD38, можно разделить на разные группы. Связывание некоторых антител с молекулой CD38 (преимущественно с аминокислотами 220-300) может запускать в клетке-мишени такие активности, как высвобождение Ca2+, высвобождение цитокина, явления фосфорилирования и стимуляцию роста, основанную на соответствующей специфичности антитела (Konopleva et al., 1998; Ausiello et al., 2000), но не выявлено четкой корреляции между сайтом связывания различных известных антител и их (не-)агонистическими свойствами (Funaro et al., 1990).
Об эффективности опубликованных анти-CD38-антител известно относительно немного. Известно, что все известные антитела распознают эпитопы (аминокислотные остатки от 220 до 300), локализованные только на C-концевом участке CD38. До настоящего времени не обнаружены антитела, специфичные в отношении эпитопов на N-концевом участке CD38, располагающихся в первичной последовательности белка вдали от активного сайта. Однако авторы настоящего изобретения обнаружили, что OKT10, в отношении которого проводили клиническое тестирование, имеет относительно низкую аффинность и эффективность при анализе химерной конструкции, содержащей участки человеческого Fc. Кроме того, OKT10 представляет собой мышиное антитело, что делает его неподходящим для введения человеку. Недавно был описан фрагмент человеческого анти-CD38 scFv-антитела (WO 02/06347). Однако это антитело является специфичным для селективно экспрессируемого эпитопа CD38.
Соответственно, в свете высокого потенциала для терапии анти-CD38-антителами существует большая потребность в человеческих анти-CD38-антителах с высокой аффинностью и с высокой эффективностью ADCC и/или CDC-опосредованного лизиса злокачественных клеток, сверхэкспрессирующих CD38.
Настоящее изобретение удовлетворяет этим и другим потребностям путем обеспечения полностью человеческих и высокоэффективных анти-CD38-антител, описание которых приводится ниже.
Сущность изобретения
Целью настоящего изобретения является обеспечение человеческих и гуманизированных антител, способных эффективно опосредовать лизис клеток, сверхэкспрессирующих CD38.
Другой целью изобретения является обеспечение антител, безопасных для введения человеку.
Целью настоящего изобретения является также обеспечение способов лечения заболевания или и/или состояний, ассоциированных с повышенной регуляцией CD38, с использованием одного или более антител согласно изобретению. Эти и другие цели настоящего изобретения ниже описаны более полно.
В одном аспекте настоящее изобретение обеспечивает выделенное антитело, или функциональный фрагмент антитела, которые содержат антигенсвязывающую область, являющуюся специфичной в отношении эпитопа CD38, при этом антитело или его функциональный фрагмент способны опосредовать лизис клеток-мишеней CD38+ (LP-1 (DSMZ: ACC41) и RPMI-8226 (ATCC: CCL-155)) путем антителозависимой клеточно-опосредованной цитотоксичности ("ADCC") с эффективностью, по меньшей мере в два-пять раз превышающей эффективность химерного антитела OKT10, имеющего последовательность SEQ ID NO:23 и 24 (в одинаковых или по существу одинаковых условиях), при использовании человеческих мононуклеарных клеток периферической крови PBMC в качестве клеток-эффекторов, и при соотношении количества клеток-эффекторов к количеству клеток-мишеней в диапазоне от 30:1 до 50:1. Такое антитело или его функциональный фрагмент может содержать антигенсвязывающую область, которая содержит область H-CDR3, приведенную в SEQ ID NO:5, 6, 7 или 8; антигенсвязывающая область может дополнительно включать в себя область H-CDR2, приведенную в SEQ ID NO:5, 6, 7 или 8; и антигенсвязывающая область также может содержать область H-CDR1, приведенную в SEQ ID NO:5, 6, 7 или 8. Такое CD38-специфическое антитело согласно изобретению может содержать антигенсвязывающую область, которая содержит область L-CDR3, приведенную в SEQ ID NO:13, 14, 15 или 16; антигенсвязывающая область может дополнительно включать в себя область L-CDR1, приведенную в SEQ ID NO:13, 14, 15 или 16; и антигенсвязывающая область также может содержать область L-CDR2, приведенную в SEQ ID NO:13, 14, 15 или 16.
В другом аспекте настоящее изобретение связано c выделенным антителом или функциональным фрагментом антитела, содержащим антигенсвязывающую область, которая является специфичной в отношении эпитопа CD38, при этом антитело или его функциональный фрагмент способны опосредовать лизис CD38-трансфицированных клеток CHO путем CDC с эффективностью, по меньшей мере вдвое превышающей эффективность химерного OKT10 (SEQ ID NO:23 и 24) в одинаковых или по существу одинаковых условиях, как указано в предыдущем параграфе. Антитело, удовлетворяющее этим критериям, может содержать антигенсвязывающую область, которая содержит область H-CDR3, приведенную в SEQ ID NO:5, 6 или 7; антигенсвязывающая область может дополнительно включать в себя область H-CDR2, приведенную в SEQ ID NO:5, 6 или 7; и антигенсвязывающая область также может содержать область H-CDR1, приведенную в SEQ ID NO:5, 6 или 7. Такое CD38-специфическое антитело согласно изобретению может содержать антигенсвязывающую область, которая содержит область L-CDR3, приведенную в SEQ ID NO:13, 14 или 15; антигенсвязывающая область может дополнительно включать в себя область L-CDR1, приведенную в SEQ ID NO:13, 14 или 15; и антигенсвязывающая область также может содержать область L-CDR2, приведенную в SEQ ID NO:13, 14 или 15.
Антитела (и их функциональные фрагменты) согласно изобретению могут содержать антигенсвязывающую область, которая специфична в отношении эпитопа CD38, и указанный эпитоп содержит один или более аминокислотных остатков CD38 43-215, как приведено в SEQ ID NO:22. Более конкретно, эпитоп, с которым связывается антигенсвязывающая область, может содержать один или более аминокислотных остатков, обнаруживаемых в одном или более аминокислотных фрагментах секвенирования, взятых из каталога аминокислотных фрагментов 44-66, 82-94, 142-154, 148-164, 158-170 и 192-206. Для некоторых антител эпитоп может быть линейным, в то время как для других он может быть конформационным (то есть прерывистым). Антитело или его функциональный фрагмент, обладающие одним или более из этих свойств, могут содержать антигенсвязывающую область, которая содержит область H-CDR3, приведенную в SEQ ID NO:5, 6, 7 или 8; антигенсвязывающая область может дополнительно включать в себя область H-CDR2, приведенную в SEQ ID NO:5, 6, 7 или 8; и антигенсвязывающая область также может содержать область H-CDR1, приведенную в SEQ ID NO:5, 6, 7 или 8. Такое CD38-специфическое антитело согласно изобретению может содержать антиген-связывающую область, которая содержит область L-CDR3, приведенную в SEQ ID NO:13, 14, 15 или 16; антигенсвязывающая область может дополнительно включать в себя область L-CDR1, приведенную в SEQ ID NO:13, 14, 15 или 16; и антигенсвязывающая область также может содержать область L-CDR2, приведенную в SEQ ID NO:13, 14, 15 или 16.
Пептидные варианты раскрываемых последовательностей также входят в объем настоящего изобретения. Соответственно, настоящее изобретение включает в себя анти-CD38-антитела, имеющие тяжелую цепь аминокислотной последовательности, обладающей последовательностью, по меньшей мере на 60 процентов идентичной в гипервариабельных CDR-областях областям CDR, приведенным в SEQ ID NO:5, 6, 7 или 8; и/или последовательностью, по меньшей мере на 80 процентов идентичной в CDR-областях CDR-областям, приведенным в SEQ ID NO:5, 6, 7 или 8. Дополнительно включены в объем настоящего изобретения анти-CD38-антитела, имеющие легкую цепь аминокислотной последовательности, обладающей последовательностью, по меньшей мере на 60 процентов идентичной в CDR-областях CDR-областям, приведенным в SEQ ID NO:13, 14, 15 или 16; и/или последовательностью, по меньшей мере на 80 процентов идентичной в CDR-областях, гомологичных CDR-областям, приведенным в SEQ ID NO:13, 14, 15 или 16.
Антитело согласно изобретению может представлять собой IgG (например, IgGI), в то время как фрагментом антитела может быть, например, Fab или scFv. Фрагмент антитела согласно изобретению может, соответственно, представлять собой или может содержать антигенсвязывающую область, которая может проявляться одним или несколькими описанными в настоящем описании путями.
Настоящее изобретение относится также к выделенным последовательностям нуклеиновых кислот, каждая из которых может кодировать область связывания с человеческим антителом или его функциональным фрагментом, являющимся специфичным в отношении эпитопа CD38. Такая последовательность нуклеиновых кислот может кодировать вариабельную тяжелую цепь антитела и включает в себя последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 2, 3 или 4, или последовательность нуклеиновых кислот, которая гибридизуется в жестких условиях с комплементарной нитью SEQ ID NO:1, 2, 3 или 4. Нуклеиновая кислота может кодировать вариабельную легкую цепь выделенного антитела или его функционального фрагмента и может содержать последовательность, выбранную из группы, состоящей из SEQ ID NO:9, 10, 11 или 12, или последовательности нуклеиновых кислот, которая гибридизуется в жестких условиях с комплементарной нитью SEQ ID NO:9, 10, 11 или 12.
Нуклеиновые кислоты согласно изобретению подходят для рекомбинантной продукции. Таким образом, изобретение также относится к векторам и клеткам-хозяевам, содержащим последовательность нуклеиновых кислот согласно изобретению.
Композиции согласно изобретению можно использовать для терапевтического или профилактического применения. В этой связи настоящее изобретение включает в себя фармацевтическую композицию, содержащую антитело согласно изобретению (или функциональный фрагмент антитела) и его фармацевтически приемлемый носитель или эксципиент. В родственном аспекте настоящее изобретение связано со способом лечения нарушения или состояния, связанного с нежелательным наличием CD38 или клеток, экспрессирующих CD38. Как указано или предполагается в настоящем изобретении, такой способ содержит стадии введения нуждающемуся в этом субъекту эффективного количества фармацевтической композиции, которая содержит антитело согласно изобретению.
Настоящее изобретение также относится к выделенным эпитопам CD38, как в линейной, так и в конформационной форме, и их применению для выделения антитела или его функционального фрагмента; и это антитело или фрагмент антитела включают в себя антигенсвязывающую область, которая специфична в отношении указанного эпитопа. В этом отношении линейный эпитоп может содержать аминокислотные остатки 192-206, в то время как конформационный эпитоп может содержать один или более аминокислотных остатков, выбранных из группы, состоящей из аминокислот 44-66, 82-94, 142-154, 148-164, 158-170 и 202-224 CD38. Эпитоп CD38 можно использовать, например, для выделения антител или их функциональных фрагментов (и каждое из этих антител или фрагментов антител содержит антигенсвязывающую область, которая специфична в отношении такого эпитопа), содержащей стадии приведения в контакт указанного эпитопа CD38 с библиотекой антител и выделение антитела (антител) или его функционального фрагмента (фрагментов).
В другом варианте осуществления настоящее изобретение связано с выделенным эпитопом CD38, который по существу состоит из аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 44-66, 82-94, 142-154, 148-164, 158-170, 192-206 и 202-224 CD38. Для целей настоящего изобретения такой эпитоп "по существу состоит из" одной из непосредственно предшествующих аминокислотных последовательностей плюс имеет дополнительные признаки, при условии, что дополнительные признаки существенно не затрагивают исходных и новых характеристик эпитопа.
В еще одном варианте осуществления настоящее изобретение связано с выделенным эпитопом из CD38, который состоит из аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 44-66, 82-94, 142-154, 148-164, 158-170, 192-206 и 202-224 из CD38.
Настоящее изобретение связано также с набором, содержащим (i) выделенный эпитоп из CD38, содержащий один или более отрезков аминокислотной последовательности, взятых из списка аминокислот 44-66, 82-94, 142-154, 148-164, 158-170, 192-206 и 202-224; (ii) библиотеку антител и (iii) инструкции для использования библиотеки антител для выделения одного или более элементов такой библиотеки, который специфически связывается с таким эпитопом.
Краткое описание чертежей
На фиг.1а представлены последовательности нуклеиновых кислот различных новых вариабельных тяжелых областей антител.
На фиг.1b показаны аминокислотные последовательности различных новых вариабельных тяжелых областей антител. CDR-области HCDR1, HCDR2 и HCDR3 обозначены от N- до C-конца жирным шрифтом.
На фиг.2a показаны последовательности нуклеиновых кислот различных новых вариабельных легких областей антител.
На фиг.2b показаны аминокислотные последовательности различных новых вариабельных легких областей антител. CDR-области LGDR1, LCDR2 и LCDR3 обозначены от N- до C-конца жирным шрифтом.
На фиг.3 показаны аминокислотные последовательности вариабельных тяжелых областей различных консенсусных последовательностей основного гена антитела HuCAL. CDR-области HCDR1, HCDR2 и HCDR3 обозначены от N- до C-конца жирным шрифтом.
На фиг.4 показаны аминокислотные последовательности вариабельных легких областей различных консенсусных последовательностей основного гена антитела HuCAL. CDR-области LCDR1, LCDR2 и LCDR3 обозначены от N- до C-конца жирным шрифтом.
На фиг.5 показана аминокислотная последовательность CD38 (первичное присоединение SWISS-PROT № P28907).
На фиг.6 показаны нуклеотидные последовательности тяжелых и легких цепей химерного OKT10.
На фиг.7 схематически показан перечень эпитопов представителей антител согласно изобретению.
На фиг.8 показана последовательность ДНК pMORPH®_h_IgGl_1 (601-2100 пар нуклеотидов (п.н.)) (SEQ ID NO:32): Вектор основан на плазмидной пкДНК3.1+векторы (Invitrogen). Аминокислотная последовательность «лишней» VH-последовательности выделена жирным шрифтом, тогда как концевые рамки считывания лидерной VH-последовательности и ген константной области напечатаны обычным шрифтом. Сайты рестрикции обозначены над последовательностью. Примирующие сайты секвенируемых праймеров подчеркнуты.
На фиг.9 показана последовательность ДНК легкой цепи экспрессирующего вектора Ig каппа pMORPH®_h_IgK_1 (601-1400 п.н.) (SEQ ID NO:33): Вектор основан на пкДНК3.1+векторы (Invitrogen). Аминокислотная последовательность «лишней» Vk-последовательности выделена жирным шрифтом, тогда как концевые рамки считывания лидерной Vk-последовательности и ген константной области напечатаны обычным шрифтом. Сайты рестрикции обозначены над последовательностью. Примирующие сайты секвенируемых праймеров подчеркнуты.
На фиг.10 показана последовательность ДНК легкой цепи вектора HuCAL Ig лямбда pMORPH®_h_Igλ_1 (601-1400 п.н.) (SEQ ID NO:34): Аминокислотная последовательность «лишней» Vλ-последовательности выделена жирным шрифтом, тогда как концевые рамки считывания лидерной Vλ-последовательности и ген константной области напечатаны обычным шрифтом. Сайты рестрикции обозначены над последовательностью. Примирующие сайты секвенируемых праймеров подчеркнуты.
На фиг.11 показаны результаты исследования пролиферации: клетки PBMC от 6 разных здоровых доноров (показанных отдельными точками) культивировали в течение 3 дней в присутствии антител HuCAL® Мон.АТ#1 (=MOR03077), Мон.АТ#2 (=MOR03079) и Мон.АТ#3 (=MOR03080), ссылочного антитела chOKT10, контрольного агонистического (ag) IB4, неродственного HuCAL® отрицательного контроля IgGI (NC) и мышиного IgQ2a (Iso) в качестве родственного по изотипу контроля для IB4. Использовали стандартные метки BrdU для измерения активности пролиферации и их включение (в RLU, относительных световых единицах) анализировали путем твердофазного иммуноферментного анализа ELISА на основе хемилюминесценции.
На фиг.12 показаны результаты анализа высвобождения IL-6: Клетки PBMC от 4-8 различных здоровых доноров (показанных отдельными точками) культивировали в течение 24 часов в присутствии только антител HuCAL® Мон.АТ#1 (=MOR03077), Мон.АТ#2 (=MOR03079) и Мон.АТ#3 (=MOR03080), ссылочного антитела chOKT10, контрольного агонистического (ag) IB4, неродственного HuCAL® отрицательного контроля IgGI (NC) и питательной среды (среда). Содержание IL-6 в относительных световых единицах (RLU) анализировали в супернатантной культуре путем хемилюминесцентного способа ELISA.
На фиг.13 приведены данные о цитотоксичности по отношению к клеткам-предшественникам CD34+/CD38+: Клетки PBMC от здоровых доноров, среди которых содержатся аутогенные клетки-предшественники CD34+/CD38+, инкубировали в присутствии HuCAL® Мон.АТ#1 (=MOR03077), Мон.АТ#2 (=MOR03079) и Мон.АТ#3 (=MOR03080), положительного контроля (РС=chOKT10) и неродственного отрицательного контроля HuCAL®, соответственно, в течение 4 часов. После этого клеточную суспензию смешивали с кондиционированной метилцеллюлозной средой и инкубировали в течение 2 недель. Колониеобразующие единицы (CFU), полученные из эритроидной колониеобразующей "взрывообразующей" единицы (BFU-E; столбец B) и гранулоцитарных/эритроидных/макрофагных/мегакариоцитарных стволовых клеток (CFU-GEMM; столбец B) и гранулоцитарных/макрофагных стволовых клеток (CFU-GM; столбец C) подсчитывали и стандартизировали против контрольной среды ("отсутствует”=среда). В столбце А показано общее количество CFU (общее количество колоний СFU) для всех предшественников. Приводятся средние значения по меньшей мере от 10 различных доноров PBMC. Планки погрешностей представляют стандартную ошибку средних значений.
На фиг.14 приведены данные, относящиеся к ADCC у различных клеточных линий:
a: Единственные измерения (за исключением RPMI8226: среднее значение 4 отдельных анализов); соотношение E:T имеет значение 30:1.
b: Namba et al., 1989.
c: 5 мкг/мл, используемые для последовательности антитела (за исключением Raji с 0,1 мкг/мл).
d: Дополнительный ретиноевый анализ для стимуляции специфического лизиса CD38-экспрессии [%]=[(экспериментальный лизис - средний лизис)/(1-средний лизис)]×100.
PC: положительный контроль (=chOKT10).
ММ: Множественная миелома.
ХЛЛ: Хронический В-клеточный лейкоз.
ОЛЛ: Острый лимфобластный лейкоз.
ОМЛ: Острый миелолейкоз.
DSMZ: Немецкая коллекция микроорганизмов и клеточных культур GmbH.
ATCC: Американская коллекция типовых культур.
ECACC: Европейская коллекция клеточных культур.
MFI: Средняя интенсивность флуоресценции.
На фиг.15 приведены данные относительно ADCC к образцам с ММ:
a: 2-4 отдельных исследования.
На фиг.16 приведены экспериментальные результаты по средним объемам опухоли ксенотрансплантата человеческой миеломы после лечения с помощью MOR03080: группа 1: носитель; группа 2: MOR03080 в качестве hIgG1 в дозе 1 мг/кг в течение 32-68 дней через день; группа 3: MOR03080 в качестве hIgG1 в дозе 5 мг/кг в течение 32-68 дней через день; группа 4: MOR03080 в качестве chIgG2a в дозе 5 мг/кг в течение 32-68 дней через день; группа 5: MOR03080 в качестве hIgG1 в дозе 1 мг/кг, в течение 14-36 дней через день, группа 6: без лечения.
Подробное описание изобретения
Настоящее изобретение основано на открытии новых антител, являющихся специфичными в отношении CD38 или имеющих высокую аффинность к CD38 и способных приносить субъекту терапевтическую пользу. Антитела согласно изобретению, которые могут быть человеческими или гуманизированными, можно использовать во многих контекстах, которые более полно описаны ниже.
"Человеческое" антитело или функциональный фрагмент человеческого антитела в настоящем изобретении определены как не являющиеся химерными (например, не "гуманизированными") и не полученные (как полностью, так и частично) из видов, отличных от нечеловека. Человеческое антитело или функциональный фрагмент антитела можно получать от человека, или они могут быть синтетическими человеческими антителами. "Синтетическое человеческое антитело" в настоящем изобретении определено как антитело, имеющее последовательность, полностью или частично полученную in silico из синтетических последовательностей на основе анализа известных последовательностей человеческих антител. Конструирования in silico последовательности человеческого антитела или его фрагмента можно достичь путем анализа базы данных последовательностей человеческого антитела или фрагмента антитела и путем разработки полипептидной последовательности с использованием полученных из нее данных. Другим примером человеческого антитела или функционального фрагмента антитела является антитело, кодируемое нуклеиновой кислотой, выделенной из библиотеки последовательностей антител человеческого происхождения (то есть такой библиотеки, которая основана на антителах, взятых из человеческого естественного источника).
"Гуманизированное антитело" или функциональный гуманизированный фрагмент антитела определены в настоящем изобретении как (i) полученные из источника нечеловеческого происхождения (например, у трансгенной мыши, несущей ксеногенную иммунную систему), и такое антитело основано на последовательности зародышевой линии человеческих клеток; или (ii) являющиеся химерными, в которых вариабельный домен имеет нечеловеческое происхождение, и константный домен имеет человеческое происхождение, или (iii) являются CDR-трансплантированными, в которых области CDR вариабельных доменов имеют нечеловеческое происхождение, в то время как один или более каркасов вариабельных доменов имеют человеческое происхождение, и константный домен (при его наличии) имеет человеческое происхождение.
В настоящем описании подразумевается, что антитело "связывается специфично с", является "специфичным к/для" или "специфично распознает" антиген (здесь CD38), если такое антитело способно различать такой антиген от одного или более ссылочных антигенов, поскольку специфичность связывания не является абсолютной, а является относительным свойством. В наиболее общем виде (и при отсутствии упоминания о конкретных ссылках) "специфическое связывание" относится к способности антитела отличать представляющий интерес антиген от неродственных антигенов, как определяется, например, один из следующих способов. К таким способам относятся без ограничения Вестерн-блоттинг, ELISA-, RIA-, ECL-, IRMA-анализы и пептидное сканирование. Например, можно проводить стандартный анализ ELISA. Можно проводить оценку по стандартному развитию окраски (например, вторичное антитело с пероксидазой хрена и тетраметилбензидин с перекисью водорода). Реакцию в некоторых лунках оценивают по оптической плотности, например, при 450 нм. Обычный исходный уровень (отрицательная реакция) может составлять 0,1 OD; обычная положительная реакция может составлять 1 OD. Это означает, что положительная и отрицательная реакции могут отличаться более чем в 10 раз. Как правило, определение специфичности связывания проводят, используя не один ссылочный антиген, а набор примерно из трех-пяти неродственных антигенов, таких как молочный порошок, BSA, трансферрин и пр.
Вместе с тем "специфическое связывание" может относиться также к способности антитела определять различие между антигеном-мишенью и одним или более близкородственными антигенами, которые используются в качестве ссылочных пунктов, например, между CD38 и CD157. Дополнительно "специфическое связывание" может относиться к способности антитела отличать различные части его антигена-мишени, например, различные домены или области CD38, такие как эпитопы в N-концевой или C-концевой области CD38, или один или более остатков ключевой аминокислоты или отрезки [последовательностей] аминокислотных остатков CD38.
Также используемый в настоящем изобретении термин "иммуноглобулин" (Ig) определяет белок, принадлежащий к классам IgG, IgM, IgE, IgA или IgD (или их любым подклассам), и включает в себя все общеизвестные антитела и их функциональные фрагменты. Термин "функциональный фрагмент" антитела/иммуноглобулина определяет фрагмент антитела/иммуноглобулина (например, вариабельную область IgG), который сохраняет антиген-связывающую область. "Антигенсвязывающая область" антитела обычно выявляется в одной или более гипервариабельной области (областях) антитела, например в области CDR-1, -2, и/или -3; однако вариабельные "каркасные" области могут также играть важную роль в связывании антигена, например, путем обеспечения скелета для областей CDR. Предпочтительно, "антигенсвязывающая область" содержит по меньшей мере 4-103 аминокислотных остатков из вариабельных легких (VL) цепей, и 5-109 из вариабельных тяжелых (VH) цепей, более предпочтительно 3-107 аминокислотных остатков из VL и 4-111 из VH, и особенно предпочтительно содержит полные VL и VH цепи (положения аминокислот 1-109 в VL и 1-113 в VH; нумерация согласно WO 97/08320). Предпочтительным классом иммуноглобулинов для использования в настоящем изобретении является IgG. "Функциональные фрагменты" согласно изобретению включают в себя домен F(ab')2-фрагмента, Fab-фрагмента и scFv. F(ab')2 или Fab можно конструировать так, чтобы минимизировать или полностью исключить межмолекулярные дисульфидные взаимодействия, которые возникают между доменами CHI и CL.
Антитело согласно изобретению может быть получено из библиотеки рекомбинантных антител на основе аминокислотных последовательностей, которые были разработаны in silico и кодируются искусственно созданными нуклеиновыми кислотами. В дизайне silico последовательность антитела получают, например, путем анализа базы данных по человеческим последовательностям и разработки полипептидной последовательности с использованием полученных в анализе данных. Способы дизайна и получения in silico-созданных последовательностей описаны, например, у Knappik et al., J. Mol. Biol. (2000) 296:57; Rrebs et al., J. Immunol. Methods. (2001) 254:67; и в патенте США № 6300064, выданном на имя Knappik et al., ссылки на которые во всей их полноте включены в настоящее описание.
Антитела согласно изобретению
Во всем тексте настоящего описания имеется ссылка на следующие антитела, согласно изобретению: "антитело №" или "LACS" или "MOR" 3077, 3079, 3080 и 3100. LAC 3077 представляет собой антитело, имеющее вариабельную тяжелую область, соответствующую последовательности SEQ ID NO:1 (ДНК)/SEQ ID NO:5 (белок), и вариабельную легкую область, соответствующую SEQ ID NO:9 (ДНК)/SEQ ID NO:13 (белок). LAC 3079 представляет собой антитело, имеющее вариабельную тяжелую область, соответствующую SEQ ID NO:2 (ДНК)/SEQ ID NO:6 (белок), и вариабельную легкую область, соответствующую SEQ ID NO:10 (ДНК)/SEQ ID NO:14 (белок). LAC 3080 представляет собой антитело, имеющее вариабельную тяжелую область, относящуюся к SEQ ID NO:3 (ДНК)/SEQ ID NO:7 (белок), и вариабельную легкую область, соответствующую SEQ ID NO:11 (ДНК)/SEQ ID NO:15 (белок). LAC 3100 представляет собой антитело, имеющее вариабельную тяжелую область, соответствующую SEQ ID NO:4 (ДНК)/SEQ ID NO:8 (белок), и вариабельную легкую область, соответствующую SEQ ID NO:12 (ДНК)/SEQ ID NO:16 (белок).
В одном аспекте настоящее изобретение связано с антителами, имеющими антиген-связывающую область, способными специфически связываться или имеющими высокую аффинность к одной или более областям CD38, аминокислотная последовательность которых приведена в SEQ ID NO:22. Считается, что антитело имеет "высокую аффинность" к антигену, если значение аффинности составляет по меньшей мере 100 нм (моновалентная аффинность фрагмента Fab). Антитело или антигенсвязывающая область согласно изобретению предпочтительно могут связываться с CD38 с аффинностью, составляющей менее 100 нм, более предпочтительно меньше чем около 60 нм и еще более предпочтительно меньше чем около 30 нм. Дополнительно предпочтительными являются антитела, которые связываются с CD38 с аффинностью, составляющей меньше чем около 10 нм, и более предпочтительно меньше чем около 3 нм. Например, аффинность антитела согласно изобретению в отношении CD38 может составлять около 10,0 нм или 2,4 нм (моновалентная аффинность фрагмента Fab).
В таблице 1 приводятся общие показатели аффинности антител согласно изобретению, определяемые путем анализа поверхностного плазмонного резонанса (Biacore) и FACS- Scatchard-анализа.
Показатели аффинности антител
b: клеточная линия ММ RPMI8226, используемая в FACS-Scatchards-анализе
Согласно таблице 1 аффинность антител LAC 3077, 3079, 3080 и 3100 измеряли путем поверхностного плазмонного резонанса (Biacore) на иммобилизованном рекомбинантном CD38 и способом проточной цитометрии, используя CD38-экспрессирующую человеческую клеточную линию RPMI8226. Исследования Biacore проводили на непосредственно иммобилизованном антигене (слитый белок CD38-Fc). Формат Fab антител LAC 3077, 3079, 3080 и 3100 показывает диапазон моновалентной аффинности, составляющей между около 2,4 и 56 нм на иммобилизованном слитом белке CD38-FC, с наиболее высокой аффинностью, демонстрируемой LAC 3079, и затем Fab 3100, 3080 и 3077.
Для определения аффинности на клеточной основе (FACS Scatchard) использовали формат IgGI. Правый столбец таблицы 1 означает прочность связывания антител LAC в этом формате. LAC 3080 проявлял наиболее сильное связывание, которое является слегка более сильным, чем у LAC 3079 и LAC 3077.
Другим предпочтительным признаком предпочтительных антител согласно изобретению является их специфичность в отношении зоны в пределах N-концевой области CD38. Например, антитела согласно изобретению LAC 3077, 3079, 3080, и 3100 могут специфически связываться с N-концевой областью CD38.
Тип эпитопа, с которым связывается антитело согласно изобретению, может представлять собой линейный тип (то есть один последовательный отрезок аминокислот) или конформационный тип (то есть множественные отрезки аминокислот). Чтобы определить, является ли эпитоп конкретного антитела линейным или конформационным, специалист в данной области может провести анализ связывания антител с перекрывающимися пептидами (например, 13-мерные пептиды с перекрыванием из 11 аминокислот), покрывающими различные домены CD38. Используя этот анализ, авторы настоящего изобретения обнаружили, что антитела LAC 3077, 3080 и 3100 распознают прерывистые эпитопы в N-концевой области CD38, тогда как эпитоп LAC 3079 можно описать как линейный (см. Фиг.7). В сочетании со сведениями, представленными в настоящем изобретении, специалист поймет, как использовать один или более выделенных эпитопов CD38 для создания антител, имеющих антиген-связывающую область, которая специфична в отношении указанных эпитопов (например, использование синтетических пептидов из эпитопов CD38 или клеток, экспрессирующих эпитопы CD38).
Антитело согласно изобретению предпочтительно является видовым антителом, перекрестно реагирующим с человеческим и по меньшей мере с одним другим видом, который может представлять собой вид грызунов или вид не принадлежащих к человеческому роду приматов. Приматом нечеловеческого рода может являться резус, бабуин и/или циномолгус. Видом грызунов может быть мышь, крыса и/или хомяк. Антитело, которое является перекрестно реагирующим, например, по меньшей мере с одним видом грызунов, по сравнению с известными анти-CD38-антителами, может обеспечивать большую гибкость и выгоду для проведения исследований in vivo с одним и тем же антителом у множества видов.
Предпочтительно, антитело согласно изобретению способно не только связываться с CD38, но также и способно опосредовать лизис клетки, экспрессирующей CD38. Более конкретно, антитело согласно изобретению может опосредовать свой терапевтический эффект путем истощения CD38-позитивных (например, злокачественных) клеток посредством антитело-эффекторных функций. Эти функции включают в себя антителозависимую клеточно-опосредованную цитотоксичность (ADCC) и комплементзависимую цитотоксичность (CDC).
В таблице 2 приведены суммарные показатели определения EC50 (средней дозы, вызывающей эффект в 50% случаев) антител, представленных в настоящем изобретении, как по ADCC, так и по CDC.
Значения EC50 антител
b: единственное определение
с: среднее значение из 2 определений ЕС50
d: среднее значение из 3 определений ЕС50
е: среднее значение из 4 определений ЕС50
Вместе с тем экспрессия CD38 выявлена не только в иммунных клетках, имеющих миелоидное (например, моноциты, гранулоциты) и лимфоидное происхождение (например, активированные B- и Т-клетки; плазматические клетки), но также и в соответствующих клетках-предшественниках. Поскольку важно, что на эти клетки не действует антитело-опосредованный лизис злокачественных клеток, антитела согласно изобретению предпочтительно не являются цитотоксическими в отношении клеток-предшественников.
Наряду со своей каталитической активностью в качестве циклической АДФ-рибозил-циклазы и -гидролазы, CD38 проявляет способность к трансдукции сигналов биологической релевантности (Hoshino et al., 1997; Ausiello et al., 2000). Эти функции in vivo можно индуцировать, например, путем рецептор-лигандного взаимодействия или перекрестного связывания с агонистическими анти-CD38-антителами, что приводит, например, к мобилизации кальция, пролиферации лимфоцитов и высвобождению цитокинов. Предпочтительно антитела согласно изобретению представляют собой неагонистические антитела.
Варианты пептидов
Антитела согласно изобретению не ограничены специфически пептидными последовательностями, обеспечиваемыми в настоящем изобретении. В настоящем изобретении осуществляются также варианты этих полипептидов. Со ссылкой на настоящее описание и общедоступные технологии специалист в данной области сможет изготавливать, тестировать и использовать функциональные варианты раскрываемых в настоящем изобретении антител, принимая во внимание, что варианты, обладающие способностью опосредовать лизис CD38+ клеток-мишеней, входят в объем настоящего изобретения. Используемый в этом контексте термин "способность опосредовать лизис CD38+ клеток-мишеней" означает функциональную характеристику, приписываемую анти-CD38-антителу согласно изобретению. Способность опосредовать лизис CD38+ клеток-мишеней, таким образом, включает в себя способность опосредовать лизис CD38+ клеток-мишеней, например, путем ADCC и/или CDC, или с использованием конструкции токсина, конъюгированного с антителом согласно изобретению.
Вариант может включать в себя антитело, имеющее по сравнению с раскрываемой в настоящем изобретении пептидной последовательностью, например, по меньшей мере одну измененную область, определяющую комплементарность (CDR) (гипервариабельную область), и/или каркасный (FR) (вариабельный) домен/положение. Для лучшей иллюстрации этой концепции ниже приводится краткое описание структуры антитела.
Антитело составлено из двух пептидных цепей, каждая из которых содержит один (легкая цепь) или три (тяжелая цепь) константных домена и вариабельную область (VL, VH), которая в каждом случае составлена из четырех FR-областей и трех промежуточных областей CDR. Сайт связывания с антигеном образован одной или более областями CDR, тогда как FR-области обеспечивают структурный каркас для областей CDR и, следовательно, играют важную роль в связывании антигена. Изменяя один или более аминокислотных остатков в CDR- или FR-области, специалист обычно может производить мутантные или разнообразные последовательности антитела, которые можно отбирать по антигену, например, для получения новых или улучшенных свойств.
В таблицах 3a (VH) и 3b (VL) очерчены области CDR и FR для некоторых антител согласно изобретению, и в них сравниваются аминокислоты в заданном положении друг с другом и с соответствующей последовательностью или с последовательностью "основного гена" (как описано в патенте США № 6300064):

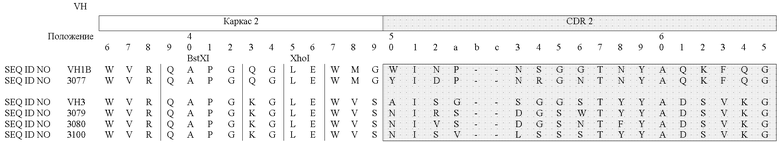

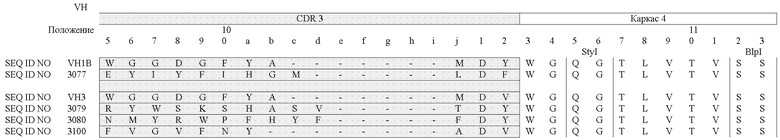

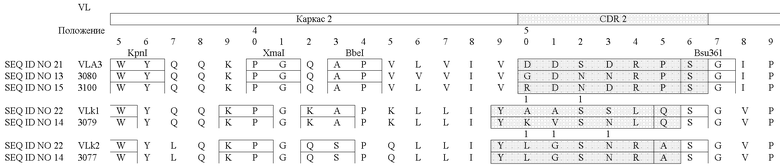


Специалист может использовать данные из таблиц 3a и 3b для дизайна пептидных вариантов, которые входят в объем настоящего изобретения. Предпочтительно, когда варианты сконструированы путем замены аминокислот в пределах одной или более областей CDR; вариант также может иметь одну или более измененные каркасных областей. Что касается сравнения новых антител друг с другом, вероятные остатки, возможные для замены, включают в себя, например, остатки вариабельных легких цепей 4 или 37 и, например, остатки вариабельных тяжелых цепей 13 или 43 из областей LAC 3080 и 3077, поскольку эти вариабельные положения находятся напротив друг друга. Замены также можно производить в каркасных областях. Например, пептид FR-домена может быть изменен при наличии отклонения в остатке по сравнению с последовательностью клетки-предшественника.
Что касается сравнения новых антител с соответствующей последовательностью или с последовательностью "основного гена", вероятные остатки, возможные для замены, включают в себя, например, остатки вариабельных легких цепей 27, 50 или 90 из LAC 3080 по сравнению с VLλ3 и, например, остатки вариабельных тяжелых цепей 33, 52 и 97 из LAC 3080 по сравнению с VH3. С другой стороны, специалист может провести аналогичный анализ путем сравнения аминокислотных последовательностей, раскрываемых в настоящем изобретении, с известными последовательностями этого же класса таких антител, используя, например, способ, описанный Knappik et al., 2000, и описанный в патенте № 6300064, выданном на имя Knappik et al.
Кроме того, варианты можно получать, используя одну область LAC в качестве исходной точки для оптимизации, разнообразя один или более аминокислотных остатков в LAC, предпочтительно аминокислотных остатков в одной или более областях CDR и путем скрининга полученной в результате коллекции вариантов антител для определения вариантов с улучшенными свойствами. В особенности предпочтительно введение разнообразия в один или более аминокислотных остатков в областях CDR-3 VL, CDR-3 VH, CDR-1 VL и/или CDR-2 VH. Разнообразие можно осуществлять путем синтеза коллекции молекул ДНК с использованием технологии тринуклеотидного мутагенеза (TRIM) (Vimeks, B., Ge, L., Plckthun, A., Schneider, K.C., Wellnhofer, G., and Moroney S.E. (1994) Trinucleotide phosphoramidites: ideal reagents for the synthesis of mixed oligonucleotides for random mutagenesis. Nucl. Acids Res. 22, 5600).
Варианты консервативных аминокислот
Можно изготовлять полипептидные варианты, которые сохраняют общую молекулярную структуру описываемой в настоящем изобретении пептидной последовательности антитела. Учитывая свойства отдельных аминокислот, специалист распознает некоторые рациональные замены. Замены аминокислот, а именно "консервативные замены", можно проводить, например, на основе сходства по полярности, заряду, растворимости, гидрофобности, гидрофильности и/или амфипатическому характеру вовлеченных остатков.
Например, (a) неполярные (гидрофобные) аминокислоты включают в себя аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин; (b) полярные нейтральные аминокислоты включают в себя глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; (c) положительно заряженные (основные) аминокислоты, включают в себя аргинин, лизин и гистидин; и (d) отрицательно заряженные (кислые) аминокислоты включают в себя аспарагиновую кислоту и глутаминовую кислоту. Обычно замены можно делать в пределах групп (a)-(d). Кроме того, глицин и пролин могут быть заменены друг на друга, что основано на их способности нарушать [структуру] α-спирали. Подобно этому некоторые аминокислоты, такие как аланин, цистеин, лейцин, метионин, глутаминовая кислота, глутамин, гистидин и лизин, обычно чаще выявляются в α-спиралях, в то время как валин, изолейцин, фенилаланин, тирозин, триптофан и треонин обычно чаще выявляются в β-складчатых структурах. Глицин, серин, аспарагиновая кислота, аспарагин и пролин обычно обнаруживаются в местах поворотов [в структуре]. Некоторые предпочтительные замены можно сделать среди следующих групп: (i) S и T; (ii) P и G; и (iii) A, V, L и I. Учитывая известный генетический код и рекомбинантные и синтетические технологии ДНК, специалист с легкостью может сконструировать ДНК путем кодирования консервативных вариантов аминокислот. В одном конкретном примере аминокислота в положении 3 в SEQ ID NO:5, 6, 7 и/или 8 может быть заменена с Q на E.
Используемый в настоящем изобретении термин "идентичность последовательности" между двумя полипептидными последовательностями указывает на процент аминокислот, идентичных у двух последовательностей. Термин "подобие последовательности" указывает на процент аминокислот, являющихся либо идентичными, либо являющихся результатом консервативных аминокислотных замен. Предпочтительные полипептидные последовательности согласно изобретению имеют степень идентичности последовательности в CDR-областях по меньшей мере 60% более предпочтительно по меньшей мере 70% или 80%, еще более предпочтительно по меньшей мере 90% и, наиболее предпочтительно по меньшей мере 95%. Предпочтительные антитела также имеют подобие последовательности в CDR-областях, составляющее по меньшей мере 80%, более предпочтительно 90% и наиболее предпочтительно 95%.
Молекулы ДНК согласно изобретению
Настоящее изобретение относится также к молекулам ДНК, кодирующим антитело согласно изобретению. Эти последовательности включают в себя молекулы ДНК, обозначенные на фиг.1а и 2a, но не ограниченные вышеперечисленным.
Молекулы ДНК согласно изобретению не ограничены последовательностями, раскрываемыми в настоящем описании, но включают в себя также и их варианты. Варианты ДНК в рамках настоящего изобретения можно описать в отношении их физических свойств в гибридизации. Специалист без труда определит, что можно использовать ДНК для идентификации ее комплемента и ее эквивалента или гомолога, используя технологию гибридизации нуклеиновой кислоты, поскольку ДНК имеет двойную спираль. Также можно определить, что гибридизация может происходить с комплементарностью менее 100%. Однако при соответствующем выборе условий для различения последовательностей ДНК можно использовать технологию гибридизации на основе структурного сродства их последовательностей с конкретным зондом. Методическими руководствами по таким условиям являются следующие публикации: Sambrook et al., 1989 (Sambrook, J., Fritsch, E.F. and Matriatis, T. (1989) Molecular Cloning: A laboratory manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, USA) and Ausubel et al., 1995 (Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Sedman, J.G., Smith, J.A., & Struhl, K. eds. (1995). Current Protocols in Molecular Biology. New York: John Wiley and Sons).
Структурное подобие между двумя полинуклеотидными последовательностями можно выразить как функцию от «жесткости» условий, при которых эти две последовательности будут гибридизоваться друг с другом. Используемый в настоящем изобретении термин «жесткость» существует в том смысле, что условия невыгодны для гибридизации. Жесткие условия очень невыгодны для гибридизации, и при таких условиях только наиболее структурно родственные молекулы будут гибридизоваться друг с другом. Напротив, нежесткие условия являются благоприятными для гибридизации молекул, проявляющих меньшую степень структурного родства. Поэтому жесткость гибридизации непосредственно коррелирует со структурными взаимоотношениями двух последовательностей нуклеиновых кислот. Следующие соотношения полезны для [установления] корреляции между гибридизацией и структурным родством (где Tm является температурой плавления дуплекса нуклеиновой кислоты):
a. Tm=69,3+0,41(G+C)%;
b. Tm ДНК-дуплекса уменьшается на 1°C с увеличением на каждый 1% количества несовпадающих пар оснований;
c. (Tm)μ2-(Tm)μ1=18,5 log10μ2/μ1, где μ2 и μ1 представляют собой ионную силу двух растворов.
Жесткость гибридизации представляет собой функции многих факторов, включая общую концентрацию ДНК, ионную силу, температуру, размер зонда и наличие веществ, которые разрушают водородные связи. Факторы, облегчающие гибридизацию, включают в себя высокие концентрации ДНК, высокие значения ионной силы, низкие температуры, увеличенный размер зонда и отсутствие веществ, разрушающих водородные связи. Обычно гибридизацию осуществляют в две стадии: стадию "связывания" и стадию "промывки".
Во-первых, на стадии связывания зонд связан с мишенью в условиях, благоприятных для гибридизации. Жесткость на этой стадии обычно регулируют путем изменения температуры. При высокой жесткости температура обычно соответствует 65-70°C в случае, если не используются короткие (<20 п.н.) олигонуклеотидные зонды. Представляемый раствор гибридизации содержит 6× раствора хлорида и цитрата натрия SSC, 0,5% додецилсульфата натрия SDS, 5× раствора Денхардта и 100 мкг неспецифического носителя ДНК. См. Ausubel et al., раздел 2,9, приложение 27 (1994). Безусловно, известно множество различных, но при этом функционально эквивалентных буферных условий. Там, где степень родства ниже, можно выбирать более низкую температуру. Значение низкой температуры жесткого связывания составляет примерно от 25 до 40°C. Средняя жесткость предусматривает температуру в диапазоне по меньшей мере от 40°C до менее чем примерно 65°C. Высокая жесткость предусматривает температуру по меньшей мере примерно 65°C.
Во-вторых, излишки зонда удаляют промывкой. Именно в этой фазе обычно применяют более жесткие условия. Следовательно, именно эта стадия "промывки" является наиболее важным в определении родства посредством гибридизации. Промывающие растворы обычно содержат низкие концентрации солей. В одном примере раствор средней жесткости содержит 2X SSC и 0,1% SDS. Промывающий раствор высокой жесткости содержит эквивалентное количество (по ионной силе) менее чем около 0,2X SSC, с раствором предпочтительной жесткости, содержащим около 0,1X SSC. Температуры, связанные с различной жесткостью, сходны с вышеуказанными температурами для "связывания". Промывающий раствор также обычно неоднократно меняют в течение промывки. Например, обычные условия промывки высокой жесткости включают в себя двукратную промывку в течение 30 минут при 55°C и трехкратную промывку в течение 15 минут при 60°C.
Соответственно, настоящее изобретение включает в себя молекулы нуклеиновой кислоты, которые гибридизуются с молекулами, представленными в фиг.1 и 2a в условиях связывания и промывки высокой жесткости, в которых такие нуклеиновые молекулы кодируют антитело или его функциональный фрагмент, имеющие свойства, описанные в настоящем изобретении. Предпочтительными молекулами (с точки зрения мРНК) являются молекулы, имеющие последовательности, по меньшей мере на 75% или 80% (предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% и наиболее предпочтительно по меньшей мере на 95%) гомологичные или идентичные одной из молекул ДНК, описываемой в настоящем изобретении. В одном конкретном примере варианта согласно изобретению в нуклеиновой кислоте в положении 7 в SEQ ID NO:1, 2, 3 и/или 4 можно заменить C на G, таким образом изменив кодон CAA на GAA.
Функционально эквивалентные варианты
Можно описать еще один класс вариантов ДНК, входящих в объем настоящего изобретения, по отношению к продукту, который они кодируют (см. белки, приведенные на фиг.1b и 2b). Эти функционально эквивалентные гены отличаются тем, что они кодируют сходные пептидные последовательности, приведенные на фиг.1b и 2b, благодаря вырожденности генетического кода. Последовательности SEQ ID NO:1 и 31 представляют собой пример функционально эквивалентных вариантов, поскольку эти последовательности нуклеиновых кислот различны, тем не менее они кодируют сходный полипептид, то есть SEQ ID NO:5.
Общепризнанно, что варианты обеспечиваемых настоящим изобретением молекул ДНК могут быть сконструированы несколькими различными способами. Например, они могут быть сконструированы как полностью синтетические ДНК. Способы эффективного синтеза олигонуклеотидов в диапазоне от 20 примерно до 150 нуклеотидов общедоступны. См. Ausubel et al., раздел 2,11, приложение 21 (1993). Перекрывающиеся олигонуклеотиды можно синтезировать и проводить сборку способом, впервые изложенным Khorana et al., в J. Mol Biol. 72:209-217 (1971); см. также Ausubel et al., выше, раздел 8,2. Конструирование синтетических ДНК предпочтительно проводят с подходящими сайтами рестрикции, созданными на 5'- и 3'-концах гена для облегчения клонирования в подходящий вектор.
Как указано, способ создания вариантов состоит в том, чтобы начать с одной из ДНК, раскрываемых в настоящем изобретении, и затем проводить сайт-направленный мутагенез. См. Ausubel et al., выше, глава 8, приложение 37 (1997). При обычном способе ДНК-мишень клонируют в фаг, являющийся носителем однонитевой ДНК. Однонитевую ДНК выделяют и гибридизуют с олигонуклеотидом, содержащим желаемую замену нуклеотида. Синтезируют комплементарную нить, и двунитевой фаг вводят в клетку-хозяина. Часть получившегося потомства будет нести желаемую мутацию, которую можно подтвердить, используя секвенирование ДНК. Кроме того, доступны различные способы, повышающие вероятность того, что фаговое потомство будет желательным мутантом. Эти способы известны специалистам в данной области, и наборы для создания таких мутантов коммерчески доступны.
Конструкция и экспрессия рекомбинантной ДНК
Настоящее изобретение дополнительно обеспечивает конструкции рекомбинантной ДНК, содержащие одну или более нуклеотидных последовательностей согласно изобретению. Рекомбинантные конструкции согласно изобретению используют в связи с вектором, таким как плазмида или вирусный вектор, в которые вставлена молекула ДНК, кодирующая антитело согласно изобретению.
Кодируемый ген может быть произведен с помощью технологий, описанных Sambrook et al., 1989 и Ausubel et al., 1989. С другой стороны, последовательности ДНК могут быть синтезированы химическим путем, например, с использованием синтезаторов. Например, можно обратиться к технологии, описанной в Oligonucleotide synthesis (1984, Gait, ed., IRL Press, Oxford), ссылка на которое включена в настоящее описание во всей ее полноте. Рекомбинантные конструкции согласно изобретению содержат экспрессирующие векторы, которые способны к экспрессии РНК и/или белковых продуктов кодируемой ДНК или нескольких ДНК. Вектор может дополнительно содержать регуляторные последовательности, включающие в себя промотор, функционально связанный с открытой рамкой считывания (ORF). Вектор может дополнительно содержать выбранную маркерную последовательность. Специфическая инициация и сигналы бактериальной секреции также могут быть необходимы для эффективной трансляции встроенного гена-мишени, кодирующего последовательности.
В настоящем изобретении дополнительно обеспечиваются клетки-хозяева, содержащие по меньшей мере одну из ДНК согласно изобретению. Клеткой-хозяином может быть фактически любая клетка, для которой приемлемы экспрессирующие векторы. Например, это может быть клетка-хозяин высших эукариот, такая как клетка млекопитающего, клетка-хозяин низших эукариот, такая как дрожжевая клетка, но предпочтительной является прокариотическая клетка, такая как бактериальная клетка. Введение рекомбинантных конструкций в клетку-хозяина можно осуществлять путем трансфекции фосфатом кальция, диэтиламиноэтилом ДЭМЭ, декстран-опосредованной трансфекции, электропорации или фаговой инфекции.
Бактериальная экспрессия
Полезные экспрессирующие векторы для использования у бактерий конструируют вставкой структурной последовательности ДНК, кодирующей желаемый белок вместе с подходящим сигналами инициации, трансляции и терминации в функциональной фазе считывания с функциональным промотором. Вектор будет содержать один или более выбираемые по фенотипу маркеры и начало репликации для гарантии сохранения вектора и, если это желательно, для обеспечения амплификации в клетке-хозяине. Подходящие для трансформации прокариотические клетки-хозяева включают в себя E. coli, Bacillus subtilis, Salmonella typhimurium и различные виды в пределах рода Pseudomonas, Streptomyces и Stafylococcus.
Бактериальными векторами могут являться векторы, например, на основе фага, плазмиды или фагемиды. Эти векторы могут содержать выбранный маркер и бактериальное начало репликации, выведенные из коммерчески доступных плазмид, обычно содержащих элементы хорошо известного вектора клонирования pBR322 (ATCC 37017). В последующем происходит трансформация подходящей нити хозяина и рост нити хозяина до адекватной клеточной плотности, выбранный промотор дерепрессируют/индуцируют адекватными способами (например, изменением температуры или химической индукцией) и в течение дополнительного времени клетки культивируют. Клетки обычно собирают центрифугированием, разрушают физическими или химическими способами, и полученный неочищенный экстракт сохраняют для дальнейшей очистки.
Некоторые экспрессирующие векторы в бактериальных системах можно предпочтительно выбирать в зависимости от предназначения использования экспрессируемого белка. Например, когда необходимо произвести большое количество такого белка, например, для образования антител или для скрининга библиотек белков, желательными могут являться векторы, которые направляют экспрессию высоких уровней слитых белковых продуктов, которые легко очищаются.
Терапевтические способы
Терапевтические способы охватывают введение нуждающемуся в лечении субъекту терапевтически эффективного количества антитела согласно изобретению. "Терапевтически эффективное" количество в настоящем изобретении определяют, как количество антитела, которое является достаточным количеством для истощения CD38-положительных клеток в зоне лечения у субъекта, как в виде единственной дозы, так и в соответствии с режимом многократного дозирования, и единственное или в комбинации с другими средствами, которое приводит к облегчению неблагоприятного состояния, и вместе с тем это количество является токсикологически толерантным. Субъектом может являться человек или млекопитающее, не принадлежащее к человеческому виду (например, кролик, крыса, мышь, обезьяна или другой примат низшего рода).
Антитело согласно изобретению можно совместно вводить с известными лекарственными средствами, и в некоторых случаях само антитело может быть модифицировано. Например, антитело можно конъюгировать с иммунотоксином или радиоизотопом для потенциально дополнительного повышения эффективности.
Антитела согласно изобретению можно использовать в качестве терапевтического или диагностического инструмента в различных ситуациях, когда выявлен CD38 или его нежелательная экспрессия. Особенно подходящими нарушениями и состояниями для лечения антителом согласно изобретению являются множественная миелома (ММ) и другие гематологические заболевания, такие как хронический лимфоцитарный лейкоз (ХЛЛ), хронический миелоидный лейкоз (ХМЛ), острый миелоидный лейкоз (ОМЛ) и острый лимфоцитарный лейкоз (ОЛЛ). Антитело согласно изобретению также можно использовать для лечения воспалительных заболеваний, таких как ревматоидный артрит (РА) или системная красная волчанка (СКВ).
Для лечения любого из вышеперечисленных нарушений фармацевтические композиции для применения согласно настоящему изобретению можно образовывать общепринятыми способами, с использованием одного или более физиологически приемлемых носителей или эксципиентов. Антитело согласно изобретению можно вводить любым подходящим способом, который можно изменять в зависимости от типа заболевания, предназначенного для лечения. Возможные пути введения включают в себя парентеральный (например, внутримышечный, внутривенный, внутриартериальный, внутрибрюшинный или подкожный), внутрилегочный и интраназальный, и, если желательно, путь введения внутрь пораженной ткани для местной иммуносупрессивной терапии. Дополнительно, антитело согласно изобретению можно вводить импульсным вливанием, например, с уменьшением дозы антитела. Предпочтительно, дозу вводят путем инъекции, с наиболее предпочтительным внутривенным или подкожным введением, отчасти зависящим от того, является ли введение краткосрочным или постоянным. Количество, необходимое для введения, будет зависеть от различных факторов, таких как клинические симптомы, масса индивидуума, введение других препаратов. Специалист в данной области определит, что путь введения будет варьировать в зависимости от нарушения или состояния, предназначенного для лечения.
Определение терапевтически эффективного количества нового полипептида, согласно настоящему изобретению, в значительной степени будет зависеть от конкретных особенностей пациента, пути введения и характера заболевания, предназначенного для лечения. Общее практическое руководство можно найти, например, в публикациях Международной конференции по гармонизации International Conference on Harmonisation и в Remington's pharmaceutical sciences, главы 27 и 28, стр.484-528 (18th ed., Alfonso R. Gennaro, Ed., Easton, Pa.: Mack Pub. Co., 1990). Более конкретно, определение терапевтически эффективного количества будет зависеть от таких факторов, как токсичность и эффективность лекарственного средства. Токсичность можно определить, используя способы, известные в данной области и изложенные в вышеприведенных ссылках. Эффективность можно определить, используя указанное практическое руководство в соединении со способами, описанными ниже в примерах.
Способы диагностики
CD38 является высокоэкспрессируемым в гематологических клетках в некоторых злокачественных образованиях; таким образом, анти-CD38-антитело согласно изобретению можно использовать для отображения или визуализации участка возможного скопления злокачественных клеток у пациента. При этом антитело может быть меченым для выявления с помощью радиоизотопов, аффинных меток (таких как биотин, авидин и т.д.), флуоресцентных меток, парамагнитных атомов и т.д. Порядок выполнения таких способов мечения известны в данной области. Обзор клинического применения антител для диагностического изображения приведены Grossman, H.B., Urol. Clin. North Amer. 13:465-474 (1986)), Unger, Е.C. et al., Invest. Radiol. 20:693-700 (1985)) и Khaw, B.A. et al., Science 209:295-297(1980).
Обнаружение фокусов таких выявляемых с помощью меток антител может быть, например, показателем локализации развития опухоли. В одном варианте осуществления это исследование проводят путем отбора образцов ткани или крови, и инкубирования таких образцов в присутствии выявляемых с помощью меток антител. В предпочтительном варианте осуществления эту технологию осуществляют неинвазивным образом с помощью магнитного изображения, флюорографии и т.д. Такое диагностическое исследование можно использовать для контроля успешности лечения заболеваний, при которых наличие или отсутствие CD38-положительных клеток является существенным индикатором. Настоящее изобретение рассматривает также, как описано ниже, использование анти-CD38-антител для диагностики в установке ex vivo.
Терапевтические и диагностические композиции
Антитела согласно изобретению можно образовывать согласно известным способам изготовления фармацевтически полезных композиций, в которых антитело согласно изобретению (включая любой его функциональный фрагмент) объединено в смеси с фармацевтически приемлемым носителем. Подходящие носители и их рецептура описаны, например, в Remington's pharmaceutical sciences (18th ed., Alfonso R. Gennaro, Ed., Easton, Pa,: Mack Pub. Co., 1990). Для образования фармацевтически приемлемой композиции, подходящей для эффективного введения, такие композиции будут содержать эффективное количество одного или более антител согласно изобретению, совместно с подходящим количеством носителя.
Рецептура композиций может быть образована таким образом, чтобы задать регулируемое высвобождение активного соединения. Препараты с регулируемым высвобождением можно получить при использовании полимеров для образования комплекса или абсорбции анити-CD38-антитела. Регулируемую доставку можно осуществлять выбором подходящих макромолекул (например, полиэфиров, полиаминокислот, поливинила, пирролидона, этиленвинилацетата, метилцеллюлозы, карбоксиметилцеллюлозы или протамина сульфата) и концентрации макромолекул, а также способом включения для достижения регулируемого высвобождения. Другой возможный способ регуляции продолжительности действия с помощью препаратов, регулирующих высвобождение, состоит в том, чтобы включить анти-CD38-антитело в частицы полимерного материала, такого как полиэфиры, полиаминокислоты, гидрогели, поли(молочная кислота) или сополимеры этиленвинилацетата. Альтернативно, вместо включения этих средств в полимерные частицы можно захватывать эти материалы в приготовленные микрокапсулы, например, способами коацервации или поверхностной полимеризации, например, в гидроксиметилцеллюлозные или желатиновые микрокапсулы и, соответственно, в поли(метилметакрилатные) микрокапсулы, или в коллоидные системы доставки лекарственных средств, например, в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы или в макроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences (1980).
Соединения могут быть составлены в композицию для парентерального введения путем инъекции, например болюсной инъекции, или непрерывным вливанием. Композиция для введения может представлять собой единичную дозированную форму, например быть в ампулах, или в виде мультидозных контейнеров с добавлением консерванта. Композиции могут находиться в таких формах, как суспензии, растворы или эмульсии в масляных или водных носителях, и могут содержать средства составления препаратов, такие как суспендирующие, стабилизирующие и/или диспергирующие средства. Альтернативно, активный компонент может находиться в виде порошка для объединения с подходящим носителем, например стерильной апирогенной водой, перед применением.
Если желательно, композиции могут представлять собой упаковку или раздаточное устройство, которое может содержать одну или более единичных дозированных форм, содержащих активный компонент. Упаковка, такая как блистерная упаковка, может содержать, например, металлическую или пластиковую пленку. Упаковка или раздаточное устройство может дополняться инструкциями для введения.
Настоящее изобретение будет дополнительно понято по нижеприведенным рабочим примерам, предназначенным для иллюстрации настоящего изобретения, и таким образом, не ограничивают его объем.
ПРИМЕРЫ
Клеточные линии
Следующие клеточные линии были получены из Европейской коллекции клеточных культур (ECACC), Немецкой коллекции микроорганизмов (DSMZ) или Американской коллекции типовых культур (ATCC): клеточная линия гибридомы, продуцирующая CD38 мышиное IgGI моноклональное антитело OKT10 (ECACC, #87021903), клетки Jurkat (DSMZ, ACC282), LP-1 (DSMZ, ACC41), RPMI8226 (ATCC, CCL-155), HEK293 (ATCC, CRL-1573), CHO-K1 (ATCC, CRL-61) и Raji (ATCC, CGL-86).
Клетки и культуральные условия
Все клетки культивировали в стандартных условиях при 37°C и 5% CO2 в увлажненном инкубаторе. Клеточные линии LP-1, RPMI8226, Jurkat и Raji культивировали в среде RPMI 1640 (Pan biotech GmbH, #P04-16500) с добавлением 10% фетальной телячьей сыворотки FCS (Pan biotech GmbH, #P30-3302), 50 ед/мл пенициллина, 50 мкг/мл стрептомицина (Gibco, #15140-122) и 2 мМ глутамина (Gibco, #25030-024) и, в случае клеток Jurkat и Raji, c дополнительным добавлением 10 мМ Hepes (Pan biotech GmbH, #P05-01100) и 1 мМ пирувата натрия (Pan biotech GmbH, #P04-43100).
CHO-K1 и HEK293 выращивали в модифицированной по способу Дульбекко среде Игла DMEM (Gibco, #10938-025) с добавлением 2 мМ глутамина мм и 10% FCS. Стабильные CD38 CHO-K1 трансфектанты сохраняли в присутствии G418 (PAA GmbH, Pll-012), тогда как для HEK293 добавление 1 мМ пирувата натрия было обязательным. После транзитной трансфекции HEK293 10% FCS заменяли ультранизким IgG FCS (Invitrogen, #16250-078). Клеточную линию OKT10 культивировали в среде IDMEM (Gibco, #1980-022) с добавлением 2 мМ глутамина и 20% FCS.
Приготовление суспензионной культуры клеток периферической крови
Все образцы крови отбирали после информированного согласия. Мононуклеарные клетки периферической крови (PBMC) от здоровых доноров выделяли с помощью прибора Histopaque®-1077 (Sigma) согласно инструкциям изготовителя. Эти клеточные суспензии очищали от эритроцитов инкубированием в лизирующем буфере ACK (ACK Lysis Buffer) (0,15М NH4Cl, 10 мМ KHCO3, 0,1М EDTA) в течение 5 минут при комнатной температуре (КТ) или в коммерческом варианте (Bioscience, #00-4333). Клетки дважды промывали PBS (забуференный фосфатом физиологический раствор) и затем дополнительно обрабатывали для проточной цитометрии или ADCC (см. ниже).
Проточная цитометрия ("FACS")
Все окрашивание осуществляли в 96-луночных культуральных планшетах с круглым дном (Nalge Nunc) с 2×105 клеток на лунку. Клетки инкубировали с антителами Fab или IgG при указанных концентрациях в 50 мкл FACS-буфера (PBS, 3% FCS, 0,02% NaN3) в течение 40 минут при 4°C. Клетки дважды промывали и затем инкубировали с R-фикоэритрином (PE), конъюгированным с козьим-анти-человеческим или козьим-анти-мышиным IgG (H+L) F(ab')2 (Jackson Immuno Research), разводили 1:200 FACS буфером в течение 30 минут при 4°C. Клетки снова промывали, ресуспендировали в 0,3 мл FACS буфера и затем анализировали методом проточной цитометрии в приборе FACSCalibur (Becton Dickinson, San Diego, CA).
Для анализа Scatchard на основе FACS клетки RPMI8226 окрашивали в 12 разных разведениях (1:2n), начиная с конечной концентрации, составляющей 12,5 мкг/мл (IgG). Использовали по меньшей мере два независимых измерения для каждой концентрации и значения KD экстраполировали по средней интенсивности флуоресценции согласно Chamow et al. (1994).
Поверхностный плазмонный резонанс
Кинетические константы kon и koff определяли путем последовательных разведений соответствующего Fab, связанного с ковалентно иммобилизованным слитым белком CD38-Fс, используя инструментарий BIAcore 3000 (Biacore, Uppsala, Sweden). Для ковалентной антигенной иммобилизации использовали стандартный способ химического присоединения аминов EDC-NHS. Для прямого присоединения CD38 Fc-слитого белка сенсорные чипы CM5 (Biacore) покрывали ~600-700 RU в 10 мМ ацетатного буфера, pH 4,5. Для эталонного клеточного потока использовали соответствующее количество HSA (человеческого сывороточного альбумина). Кинетические измерения проводили в PBS (136 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,76 мМ KH2PO4, pH 7,4) при скорости потока 20 мкл/мин с использованием концентрации Fab в диапазоне от 1,5 до 500 нМ. Время введения для каждой концентрации составляло 1 минуту, с последующими 2-минутными фазами диссоциации. Для регенерации использовали 5 мкл 10 мМ HCl. Все сенсограммы подбирали по локализации, используя программное обеспечение для оценки BIA 3,1 (Biacore).
ПРИМЕР 1
Создание антител из библиотек HuCAL
Для создания терапевтических антител против CD38 проводили отбор на основе библиотеки фаговых дисплеев MorphoSys HuCAL GOLD. HuCAL GOLD® представляет собой Fab-библиотеку, основанную на концепции HuCAL® (Knappik et al., 2000; Krebs et al., 2001), где все шесть областей CDR различны, и где используется технология CysDisplayTМ для соединения Fab-фрагментов с поверхностью фага (Löhning, 2001).
A. Получение фагмид, амплификация фага и очистка
Фагмидную библиотеку HuCAL GOLD® амплифицировали в среде 2×TY, содержащей 34 мкг/мл хлорамфеникола и 1% глюкозы (2×TY-CG). После инфицирования фага-хелпера (VCSM13) в 0,5 OD600 (30 минут при 37°C без шейкера; 30 минут при 37°C с шейкером при 250 об/мин) клетки осаждали (4120×g; 5 минут; при 4°C), ресуспендировали в 2×TY/34 мкг/мл хлорамфеникола/50 мкг/мл канамицина и выращивали в течение ночи при 22°C. Фаги преципитировали из супернатанта на полиэтиленгликоле (ПЭГ), ресуспендировали в PBS/20% глицерине и сохраняли при -80°C. Фаговую амплификацию между двумя стадиями пэннинга проводили следующим образом: фазу середины логарифмического роста mid-log клеток ТG1 инфицировали элюированными фагами и помещали в планшеты на агар Лурия-Бертани LB, с добавлением 1% глюкозы и 34 мкг/мл хлорамфеникола (LB-CG). После инкубации в течение ночи при 30°C колонии снимали механически, доводили OD600 до значения 0,5 и добавляли фаг-хелпер, как описано выше.
B. Пэннинги с HuCAL GOLD®
Для селекции антител из фаговых библиотек HuCAL GOLD® их разделяли на три пула, соответствующих различным контрольным VH-генам (пул 1: VH1/5λκ, пул 2: VH3/λκ, пул 3: VH2/4/6λκ). Эти пулы были индивидуально подвергнуты 3 стадиям пэннинга цельных клеток на CD38-экспрессирующие клетки CHO-K1, с последующим pH-разведением и постадсорбционной стадией на CD38-отрицательные клетки CHО-K1 для истощения неродственных фаговых антител. Наконец, оставшиеся фаговые антитела использовали для инфицирования клетки E. coli TG1. После центрифугирования бактериальный осадок ресуспендировали в среде 2×TY, помещали в чашки на агар и инкубировали в течение ночи при 30°C. Выбранные клоны механически снимали с планшетов, фаги высвобождали и амплифицировали. Вторую и третью стадию отбора осуществляли аналогично начальной стадии.
Кодирующие Fab включения выбранного HuCAL GOLD® фага субклонировали в экспрессирующий вектор pMORPH®x9_Fab_FS (Rauchenberger et al., 2003), для облегчения быстрой экспрессии растворимого Fab. ДНК выбранных клонов перерабатывали с помощью Xbal и EcoRI, таким образом отсекая кодирующую Fab вставку (ompA-VLCL и phoA-Fd), и клонировали в секущий вектор Xbal/EcoRI pMORPH®×9_Fab_FS. Fab, экспрессируемый в этом векторе, несет две С-концевых метки (FLAGT и Strep-tag® II) для обнаружения и очистки.
ПРИМЕР 2
Биологические пробы
Антителозависимую клеточную цитотоксичность (ADCC) и комплементзависимую цитотоксичность измеряли согласно опубликованному протоколу на основе анализа проточной цитометрии (Naundorf et al., 2002) следующим образом:
ADCC:
Для измерения ADCC клетки-мишени (T) доводили до 2,0E+05 клеток/мл и метили кальцеином АМ (Molecular Probes, C-3099), 100 нг/мл, в среде RPMI 1640 (Pan biotech GmbH) в течение 2 минут при комнатной температуре. Остаточный кальцеин удаляли в 3 стадии промывки в среде RPMI 1640. Одновременно клетки PBMC приготавливали в качестве источника клеток-эффекторов (E) (природных киллеров), их доводили до 1,0E+07 и смешивали с мечеными клетками-мишенями, для получения соотношения Е:Т, составляющего 50:1 или менее, в зависимости от условий анализа. Клетки один раз промывали и смесь клеток ресуспендировали в 200 мкл среды RPMI 1640, содержащей соответствующее антитело в различных разведениях. Планшет инкубировали в течение 4 часов в стандартных условиях при 37°C и 5% CO2 в увлажненном инкубаторе. До анализа FACS клетки метили йодидом пропидия (PI) и анализировали методом проточной цитометрии (Becton-Dickinson). Подсчет проводили между точками 50,000 и 150,000 в каждой пробе.
Следующее уравнение позволяет получить степень повышения активности лизиса [в %]:
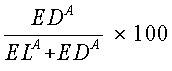
где EDА=точки мертвых клеток (клетки, окрашенные кальцеином + PI), и
ELA=точки живых клеток (клетки, окрашенные кальцеином).
CDC:
Для измерения CDC трансфектанты CD38 CHO-K1 в количестве 5,0E+04 добавляли в планшет с микролитровыми лунками (Nunc), вместе с человеческой сывороткой (Sigma, #S-1764) в разведении 1:4 и соответствующим антителом. Все реактивы и клетки разводили средой RPMI 1640 (Pan biotech GmbH), с добавлением 10% FCS. Реакционную смесь инкубировали в течение 2 часов в стандартных условиях при 37°C и 5% CO2 в увлажненном инкубаторе. В качестве отрицательной контрольной группы служили как инактивированный высокой температурой комплемент, так и CD38-трансфектанты без антитела. Клетки метили PI и подвергали FACS-анализу.
Всего подсчитали 5000 точек и количество мертвых клеток при различных концентрациях антитела, используемых для определения значений EC50. Следующее уравнение дает степень повышения активности лизиса [в %]:
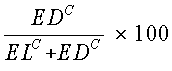
где EDС=точки мертвых клеток (клетки, окрашенные PI), и
ELС=точки живых клеток (неокрашенные клетки).
Значения цитотоксичности всего из 12 различных разведений антитела (1:2n) в трех повторах использовали в ADCC и в двух повторах - в CDC для каждого антитела для получения значений ЕС50, используя стандартное программное обеспечение (PRISM®, Graph Pad Software).
ПРИМЕР 3
Создание устойчивого CD38-трансфектанта и Fc-слитых белков CD38
Для создания белка CD38 для пэннинга и скрининга необходимо установление двух различных экспрессирующих систем. Одна стратегия включает в себя создание слитого белка СD38-Fс, который очищали от супернатантов после транзитной трансфекции клеток HEK293. Вторая стратегия охватывает создание устойчивой клеточной линии CHO-KL для высокой степени поверхностной экспрессии CD38 для использования в селекции фагового антитела посредством пэннинга цельной клетки.
В качестве начальной стадии использовали клетки Jurkat (DSMZ ACC282) для создания кДНК (Invitrogen), с последующей амплификацией полноразмерных CD38-кодирующих последовательностей, используя праймеры, комплементарные, соответственно, первым 7 и последним 9 кодонам CD38, (праймер MTE001 и MTE002rev; таблица 4). Анализ последовательности вставки CD38 подтверждал опубликованную Jackson et al. (1990) аминокислотную последовательность, за исключением положения 49, в котором выявляли глутамин вместо тирозина, как описано Nata et al. (1997). Для введения сайтов рестрикции эндонуклеаз и клонирования в различные производные экспрессирующего вектора пкДНК3.1 (Stratagene) очищенный продукт ПЦР служил матрицей для реамплификации полного гена (праймеры MTE006 и MTE007rev, таблица 4) или части гена (праймеры MTE004 и MTE009rev, таблица 4). В последнем случае фрагмент, кодирующий внеклеточный домен (аминокислоты 45-300), амплифицировали и клонировали в рамке считывания между человеческой лидирующей последовательностью Vkарра и матричной последовательностью Fc-гамма 1. Этот вектор служит вектором экспрессии для создания растворимого слитого белка CD38-Fс. Другое пкДНК3.1-производное без лидирующей последовательности использовали для вставки непроцессированного гена CD38. В этом случае стоп-кодон перед Fc-кодирующей областью и отсутствующая лидирующая последовательности приводили к поверхностной экспрессии CD38. Клетки HEK293 подвергали транзитной трансфекции вектором слитого с Fc белка для создания растворимого слитого белка Fc-CD38, и в случае непроцессированного производного, клетки CHO-K1 трансфицировали для создания устойчивой клеточной линии, экспрессирующей CD38.

ПРИМЕР 4
Клонирование, экспрессия и очистка IgG1 HuCAL®
Для экспрессии непроцессированного IgG, вариабельные фрагменты доменов тяжелых (VH) и легких цепей (VL) субклонировали из экспрессирующих векторов Fab в подходящие векторы pMORPH®_hIg (см. Фиг.8-10). Пары рестрикционных эндонуклеаз BlpI/MfeI (для получения вставки) и BlpI/EcoRI (для получения вектора) использовали для субклонирования фрагмента VH-домена в pMORPH®_hIgG1. Пары ферментов EcoRV/HpaI (лямбда-вставка) и EcoRV/BsiWI (kappa-вставка) использовали для субклонирования фрагментов VL-домена в соответствующие векторы pMORPH®_hIgκ_l или pMORPH®_h_Igλ_1. Полученные конструкции IgG экспрессировали в клетках HEK293 (ATCC CRL-1573) путем транзитной трансфекции с использованием стандартной технологии ДНК-кальций-фосфатного соосаждения. IgG очищали от супернатантов клеточной культуры путем аффинной хроматографии на сефарозной колонке с протеином А. Дальнейший процесс включал в себя буферный обмен путем гель-фильтрации и стерильной фильтрации очищенного IgG. Контроль качества давал чистоту >90% при редуцирующем электрофорезе в полиакриламидном геле с додецилсульфатом натрия SDS-PAGE и >90% мономерных IgG, что определяли с помощью аналитической эксклюзионной хроматографии. Содержание эндотоксина в материале определяли в результате кинетической реакции с лизатом амебоцитов LAL (Cambrex European Endotoxin Testing Service, Belgium).
ПРИМЕР 5
Создание и производство химерного OKT10 (chOKT10; SEQ ID NO:23 и 24)
Для конструирования chOKT10 мышиные области VH и VL амплифицировали ПЦР с использованием кДНК, приготовленной из клеточной линии мышиной гибридомы OKT10 (ECACC #87021903). Использовали набор праймеров согласно публикациям (Dattamajumdar et al., 1996; Zhou et al., 1994). Продукты ПЦР использовали для топо-клонирования (Invitrogen; вектор рСRII), и единственные колонии подвергали анализу последовательности (обратный праймер M13), который выявил две различных последовательности легкой цепи каппа и одну последовательность тяжелой цепи. Согласно выравниванию последовательностей (EMBL-база данных нуклеотидных последовательностей) и литературным данным (Krebber et al., 1997) одна из каппа-последовательностей принадлежит собственному спектру слитого партнера опухолевой клетки Х63Аg8.653 и, следовательно, не является принадлежащим антителу OKT10. Таким образом, только новая каппа-последовательность и единственный VH-фрагмент использовали для дальнейшего клонирования. Оба фрагмента реамплифицировали для дополнения сайтов рестрикции эндонуклеаз, с последующим клонированием в соответствующие экспрессирующие векторы-IgG1 pMORPH®. Последовательности тяжелой цепи (SEQ ID NO:23) и легкой цепи (SEQ ID NO:24) приведены на фиг.6. Клетки HEK293 подвергали транзитной трансфекции и супернатант анализировали путем FACS на связывание химерного антитела OKT10 с клеточной линией Raji, сверхэкспрессирующей CD38 (ATCC).
ПРИМЕР 6
Картирование эпитопа
Материалы и способы:
Антитела:
Для картирования эпитопа использовали следующие анти-CD38-IgG:
Последовательность CD38:
Аминокислотная (aa) последовательность (положения 44-300) основана на человеческом CD38, взятом из опубликованной последовательности из первичного дополнения SWISS-PROT под номером P28907. При конструировании пептида в положении 49 использовали аминокислоту Q (вместо T).
Анализ белковых пятен PepSpot:
Пептиды антигена синтезировали на целлюлозной мембране постадийно, с получением определенного расположения (пептидная матрица), и ковалентно связывали с целлюлозной мембраной. Анализ связывания проводили непосредственно на пептидной матрице. Обычно пептидную матрицу антигена инкубировали с блокирующим буфером в течение нескольких часов для уменьшения неспецифического связывания антител. После инкубирования с первичным (антиген-пептидное связывание) антителом в блокирующем буфере проводили инкубирование со вторичным антителом, меченным пероксидазой (POD), которое селективно связывает первичное антитело. Осуществляли кратковременную промывку в (Tween)-Tris-буферном растворе (TBS) непосредственно после инкубации пептидной матрицы антигена со вторичным антителом, с последующим первым хемилюминесцентным экспериментом, проводимым с целью получения предварительных результатов, определяющих какие именно пептиды антигена связываются с первичным антителом. Последующие несколько стадий промывки буфером (T-TBS и ТBS-буфер) были предназначены для уменьшения ложноположительного связывания (неспецифическое связывание антитела непосредственно с целлюлозной мембраной). После указанных стадий промывки проводили заключительный анализ хемилюминесценции. Эти данные анализировали с помощью систем изображения, показывающей интенсивность сигнала (Boehringer Light units, BLU) в виде отдельных измерений для каждого пептида. Для оценки неспецифического связывания вторичных антител (против человеческого IgG), эти антитела в качестве первой стадии инкубировали с пептидной матрицей в отсутствие первичных антител. Если первичное антитело совсем не обнаруживало связывания с пептидами, его можно было непосредственно метить POD, что повышало чувствительность системы (как осуществляли для MOR3077). В этом случае осуществляли общепринятые способы химического связывания посредством свободных аминогрупп. Антиген сканировали 13-мерными пептидами (11 перекрывающихся аминокислот). Результат представлял собой матрицу из 123 пептидов. Анализы связывания проводили непосредственно на матрице. Связанные с пептидом антитела MOR03077, MOR03079, MOR03080, MOR03100 и химерный OKT10 выявляли, используя вторичное антитело, меченное пероксидазой (конъюгированного с пероксидазой козьего антитела против человеческого IgG, специфичного в отношении гамма-цепи выделенного на основе аффинности антитела; Sigma-Aldrich, A6029). Картирование проводили с хемилюминисцентным субстратом в комбинации с системой изображения. Дополнительно осуществляли прямое POD-мечение MOR03077 для повышения чувствительности системы.
2. Краткое изложение и выводы
Все пять антител проявляли различные профили в анализе белковых пятен. Схематично краткий обзор представлен на фиг.7, который иллюстрирует различные аминокислотные последовательности распознаваемого CD38. Эпитоп для MOR03079 и химерного OKT10 вполне можно рассматривать как линейный. Эпитоп для MOR03079 можно постулировать на отрезке аминокислот 192-206 (VSRRFAEAACDVVHV) CD38, тогда как последовательность между aминокислотами 284 и 298 (FLQCVKNPEDSSCTS) для химерного OKT10 признана преобладающей. Последние результаты подтверждают опубликованные данные для парентерального мышиного OKT10 (Hoshino et al., 1997), согласно которым этот эпитоп постулирован между аминокислотами 280-298. Вместе с тем, для более точного определения эпитопа и определения ключевых аминокислот (основные сайты взаимодействия антиген-антитело) должно быть предусмотрено укорочение пептидов VSRRFAEAACDVVHV и FLQCVKNPEDSSCTS и аланиновое сканирование этих двух пептидов.
Эпитопы для MOR03080 и MOR03100 вполне можно рассматривать как прерывистые, так как было распознано несколько пептидов, охватывающих различные сайты белковых сайтов. Эти пептиды содержат аминокислоты 82-94 и 158-170 MOR03080 и 82-94, 142-154, 158-170, 188-200 и 280-296 MOR03100. Однако можно постулировать некоторые перекрывания между обоими эпитопами, так как два различных сайта, находящихся в пределах аминокислот 82-94 (CQSVWDAFKGAFI; пептид #20) и 158-170 (TWCGEFNTSKINY; пептид #58), распознаются обоими антителами.
Эпитоп для MOR03077 можно рассматривать, как четко отличающийся от двух последних эпитопов, и он может быть описан как разделенный на множество сегментов прерывистый эпитоп. Эпитоп включает в себя аминокислоты 44-66, 110-122, 148-164, 186-200 и 202-224.
ПРИМЕР 7
Анализ высвобождения/пролиферации IL-6
1. Материалы и способы
Анализ пролиферации и высвобождения IL-6 осуществляли согласно публикации Ausiello et al., (2000) со следующими модификациями: клетки PBMC от различных здоровых доноров (с их согласия) очищали путем центрифугирования в градиенте плотности с использованием системы разделения клеток Histopaque, согласно инструкциям поставщика (Sigma), и культивировали в стандартных условиях (5% CO2, 37°C) в среде RPMI 1640 с добавлением с 10% FCS и глутамина ("полная среда RPMI 1640"). Для обоих анализов использовали следующие антитела: моноклональные анти-CD38-IgGl MOR03077, MOR03079 и MOR03080 HuCAL®, агонистическое мышиное моноклональное антитело IgG2a (IB4; Malavasi et al., 1984), неродственное антитело IgG1 HuCAL®, контрольные парные изотипы (мышиный IgG2a: анти-тринитрофенол, гаптен-специфическое антитело; в каталоге: #555571, клон G155-178; Becton Dickinson) или контрольную среду. Для анализа высвобождения IL-6 1,0E+06 клеток PBMC в 0,5 мл полной среды RPMI 1640 инкубировали в течение 24 часов в культуральной 15 мл-пробирке (Falcon) в присутствии 20 мкг/мл антитела. Супернатанты клеточной культуры собирали и анализировали на предмет высвобождения IL-6, используя набор Quantikine согласно протоколу изготовителя (R&D systems). Для анализа пролиферации 2,0E+05 клеток PBMC инкубировали в течение 3 дней в 96-луночном плоскодонном планшете (Nunc) в присутствии 20 мкг/мл антитела. Каждый анализ проводили в двух повторах. Спустя 4 дня в каждую лунку добавляли BrdU, и перед фиксацией клеток и денатурацией ДНК клетки дополнительно инкубировали в течение 24 часов при 37°C согласно протоколу поставщика (Roche). Включение BrdU измеряли посредством анти-BrdU пероксидазо-связанного антитела на установке с хемилюминесцентной основой.
2. Краткое изложение и выводы
Анализ пролиферации
В дополнение к своей каталитической активности в качестве циклазы и гидролазы циклической АДФ-рибозы, CD38 проявляет способность передавать сигналы биологической релевантности (Hoshino et al., 1997; Ausiello et al., 2000). Эти функции могут быть индуцированы in vivo, например, рецептор-лигандными взаимодействиями или перекрестным связыванием с анти-CD38-антителами. Такие сигналы приводят, например, к мобилизации кальция, пролиферации лимфоцитов и высвобождению цитокинов. Однако эта передача сигналов зависит не только от антигенного эпитопа, но также может варьировать у различных доноров (Ausiello et al., 2000). В свете иммунотерапии неагонистические антитела являются предпочтительными по сравнению с агонистическими антителами. Поэтому анти-CD38-антитела HuCAL® (моноклональные антитела MOR03077; MOR03079, MOR03080) имели дополнительные отличия в анализе пролиферации и анализе высвобождения IL-6 (важного фактора роста ММ) по сравнению со ссылочным антителом chOKT10 и агонистическим анти-CD38 моноклональным антителом IB4.
Как показано на фиг.11 и фиг.12, анти-CD38-антитела Мон.АТ #1, 2 и 3 HuCAL, а также ссылочное антитело chOKT10 и соответствующий отрицательный контроль не вызывали или вызывали лишь слабую индукцию пролиферации и не вызывали высвобождения IL-6 по сравнению с агонистическим антителом IB4.
ПРИМЕР 8
Анализ клоногенности
1. Материалы и способы
Клетки PBMC, содержащие аутогенные CD34+/CD38+ клетки-предшественники, выделяли у здоровых индивидуумов (с их согласия) путем центрифугирования в градиенте плотности, с использованием системы разделения клеток Histopaque, согласно инструкциям поставщика (Sigma), и инкубировали с различным антителами анти-CD38-IgGI HuCAL® (Мон.АТ MOR03077, MOR03079 и MOR03080) и положительным контролем (PC) с chOKT10 в количестве 10 мкг/мл. Питательная среда и неродственный IgGl HuCAL® служил в качестве исходного контроля. В каждом анализе ADCC использовали 4,0E+05 клеток РBMC, которые инкубировали в течение 4 часов при 37°C в среде RPMI 1640 с добавлением 10% FCS. Для анализа клоногенности 2,50 мл "полной” метилцеллюлозы (CellSystems) вводили в 2,5E+05 клеток из анализа ADCC и инкубировали для развития колоний в течение по меньшей мере 14 дней в регулируемой среде (37°C; 5% CO2). Колонии анализировались двумя независимыми операторами и группировались в BFU-E+CFU-GEMM (эритроидные колониеобразующие "взрывообразующие” единицы и гранулоцитарные/эритроидные/макрофагные/мегакариоцитарные стволовые клетки) и CFU-GM (гранулоцитарные/макрофагные стволовые клетки).
2. Краткое изложение и выводы
Поскольку экспрессия CD38 выявлена не только в иммунных клетках миелодного (например, в моноцитах, гранулоцитах) и лимфоидного (например, в активированных B- и Т-клетках; плазматических клетках) происхождения, но также и в соответствующих клетках-предшественниках (CD34+/CD38+), представляется важным, что эти клетки не повреждаются антитело-опосредованным лизисом. В этой связи применяли анализ клоногенности для изучения этих эффектов в отношении клеток предшественников CD34+/CD38+.
Клетки PBMC от здоровых доноров инкубировали с анти-CD38-антителами HuCAL® (Мон.АТ #1, Мон.АТ #2 и Мон.АТ #3) или несколькими контрольными группами (неродственное антитело HuCAL®, питательная среда и ссылочное антитело chOKT10 в качестве положительного контроля) согласно стандартному протоколу ADCC, с последующим дополнительным инкубированием в кондиционированной метилцеллюлозе для развития колонии. Как показано на фиг.13, не выявлено никакого значительного уменьшения колониеобразующих единиц для всех анти-CD38-антител HuCAL® по сравнению с неродственным антителом или ссылочным антителом.
ПРИМЕР 9
Анализы ADCC с различными клеточными линиями и клетками первичной множественной миеломы
1. Материалы и способы
Выделение и образцов ADCC у больных с ММ: пунктаты костного мозга получали от больных с множественной миеломой (с их согласия). Злокачественные клетки очищали по стандартному протоколу, используя анти-CD138-магнитные шарики (Milteny Biotec) после центрифугирования в градиенте плотности (Sigma). Анализ ADCC осуществляли, как описано выше.
2. Краткое изложение и выводы
Для анализа ADCC использовали несколько клеточных линий, полученных из различных злокачественных новообразований, для проявления цитотоксического эффекта анти-CD38-антител HuCAL® в более широком спектре клеточных линий, включающих в себя различное происхождение и уровни экспрессии CD38. Как показано на фиг.14, все клетки претерпели лизис в ADCC при постоянной концентрации антитела (5 мкг/мл) и соотношении Е:Т, составляющем 30:1. Цитотоксичность посредством ADCC была также выявлена в нескольких образцах у больных со множественной миеломой. Все анти-CD38-антитела HuCAL® были способны осуществлять дозозависимой лизис клеток ММ, и значения EC50 варьировали от 0,006 до 0,249 нм (фиг.15).
ПРИМЕР 10
Анализ перекрестной реактивности посредством FACS и иммуногистохимии (ИГХ)
1. Материалы и способы
Иммуногистохимия миндалин: Для ИГХ-анализа моноклональные антитела против CD38 HuCAL® и неродственное антитело отрицательного контроля преобразовывали в бивалентный формат dHLX (Plückthun & Pack, 1997). Криосрезы в 5 мкм из лимфатических узлов, взятых у макаки циномолгус, макаки резус и человека (получены из архива Института патологии Университета Граца, Австрия) выполняли на криостате Leica CM3050. Срезы сушили на воздухе в течение от 30 минут до 1 часа, фиксировали в ледяном метаноле в течение 10 минут и промывали PBS. Для обнаружения dHLX-формата использовали мышиное анти-His-антитело (Dianova) в комбинации с набором Envison (DAKO). Для обнаружения мышиных анти-CD38-антител (например, ссылочного моноклонального мышиного OKT10) использовали только набор Envison.
FACS-анализ лимфоцитов: обработанные EDTA образцы крови получали от здоровых людей (с их согласия), от макак-резус и циномолгус и подвергали центрифугированию в градиенте плотности, используя систему сепарации клеток Histopaque согласно инструкциям поставщика (Sigma). Для анализа FACS инкубировали интерфазные клетки с первичными антителами (анти-CD38 HuCAL® и моноклональные антитела (Мон.АТ) в качестве отрицательного контроля, такие как мышиный IgG2a или Fab-формат, с положительным контролем в виде мышиного антитела ОKT10 и контролем по родственным изотипам) с последующим инкубированием с анти-M2-Flag (Sigma; только для Fab-формата) и меченным фикоэритрином (PE) антимышиным конъюгатом (Jackson Research). FACS-анализ проводили на популяции лимфоцитов, дающих сигнал выше порогового значения.
2. Краткое изложение и выводы
Анти-CD38 HuCAL® анализировали на предмет межвидовой перекрестной реактивности. Хотя все моноклональные анти-CD38-антитела и обладали способностью обнаруживать человеческий CD38 на лимфоцитах в анализах FACS и ИГХ, только MOR03080 вместе с положительным контролем OKT10 проявлял дополнительную реактивность в отношении CD38 макак циномолгус и резус (см. Таблицу 5: анализ перекрестной реактивности).
ПРИМЕР 11
Лечение с помощью MOR03080 ксенотрансплантатов человеческой миеломы у мышей (с использованием клеточной линии RPMI8226)
1. Постановка подкожной модели мыши
Подкожная мышиная модель опухолевой клеточной линии RPMI8226 миеломы человека у самок мышей C.B.-17-SCID была выполнкена согласно Aurigon Life Science GmbH (Tutzing, Германия) следующим образом: в день -1, 0 и 1 внутривенно вводили поликлональные анти-асиало-GM1-антитела (ASGM) (WAKO-Chemicals), истощающие ксенореактивные NK-клетки у мышей SCID, для дезактивирования любой остаточной специфической иммунной реактивности у мышей C.B.-17-SCID. В день 0 подкожно инокулировали опухолевые клетки RPMI8226 в количестве либо 5×106, либо 1×107 в 50 мкл PBS в правый бок мышей как подвергнутых терапии ASGM (как описано выше), так и не подвергавшихся (каждая группа состояла из пяти мышей). Во всех 4 инокулированных группах опухоль развивалась сходным образом без значительных отличий, что было выявлено при введении или без введения анти-асиало GM1-антител или путем инокулирования различного количества клеток. Опухоли обнаруживали медленный рост, с тенденцией замедления или изменения размеров в течение нескольких дней. Две опухоли обнаружили изменение в размерах в течение всего периода исследования, и одну опухоль после пикового объема в 321 мм3 даже расценивали как полностью исчезнувшую. Для изучения лечения на этой модели опухоли необходимо включение большого количества животных с привитой опухолью на группу.
2. Лечение с помощью MOR03080
2.1 Цель исследования
Это исследование проводили согласно Aurigon Life Science GmbH (Tutzing, Германия) для сравнения противоопухолевой эффективности применяемых внутрибрюшинно антител (анти-CD38 HuCAL®) с эффективностью носителя (РBS). Человеческое антитело hMOR03080 (изотип IgGl) тестировали при различных количествах и схемах лечения. Дополнительно тестировали химерное антитело chMOR03080 (изотип IgG2a: химерное антитело, содержащее вариабельные области MOR03080 и константные мышиные области, конструировали способом, аналогично описанному в примере 5 для химерного OKT10 (мышиные VH/VL и человеческие константные области). Линия злокачественных клеток RPMI8226 была выбрана в качестве модели и инокулирована подкожно самкам мышей SCID, как описано выше. Конечными показателями исследования являлись масса тела (м.т.), объем опухоли и клинические симптомы.
2.2 Антитела и носитель
Готовые к использованию антитела фармы Aurigon в концентрациях 2,13 мг/мл (MOR03080 hIgG1) и 1,73 мг/мл (MOR03080 chIgG2a до применения хранили при -80°C. Антитела подвергали оттаиванию и разводили PBS до соответствующей конечной концентрации. Готовый к использованию носитель (PBS) фирмы Aurigon до применения хранили при в 4°C.
2.3 Характеристика животных
Вид: мышь
Нагрузка: C.B.-17-SCID (C.B-Igh-lb/IcrTac) от Fox chase
Количество и пол: 75 самок
Поставщик: Taconic M&B, Bomholtvej 10, DK-8680 Ry
Состояние здоровья: SPF
Предписанная масса: примерно 18 граммов
Адаптация: 9 дней
2.4 Опухолевая клеточная линия
Клетки опухоли (клеточная линия RPMI8226) выращивали и переносили в Aurigon Life Science GmbH, где клетки разделяли и выращивали для другого цикла. В Aurigon приготавливали клетки для инъекций в день инокулирования. Культуральной средой, используемой для репродукции клеток, являлась среда RPMI 1640 с добавлением 5% FCS, 2 мМ L-глутамина и пенстрепа. У клеток не было отмечено никаких неожиданных проявлений в скорости роста или поведении.
Для инокулирования клетки опухоли суспендировали в PBS и доводили до конечной концентрации 1×107 клеток/в 50 мкл PBS. Суспензию опухолевых клеток тщательно перемешивали перед инъекцией.
2.5 Порядок эксперимента
В день 0 опухолевые клетки RPMI8226 в количестве 1×107 подкожно инокулировали в правую дорсальную область 75 мышам SCID. Первую группу образовали из 15 рандомизированных животных (группа 5) непосредственно после инокулирования. Этой группе вводили 1 мг/кг массы тела hIgG1-MOR03080 через день в период между 14 и 36 днем. Из остальных 60 животных на 31 день образовали 4 группы по десять животных в каждой группе (объем опухоли составлял около 92 мм3). Группы 1-4 образовывали согласно сопоставимым размерам опухоли и стандартным отклонениям. Дополнительно выбрали группу из 5 животных (группа 6), у которых выявили относительно малые объемы опухоли (объем опухоли около 50 мм3) для сравнения с группой, подвергшейся предварительному лечению (у всех, кроме трех мышей, объем опухоли составлял менее 10 мм3, у одной около 22 мм3, у одной около 44 мм3 и у одной около 119 мм3). Группам 1-4 через день, начиная с 32 дня до 68 дня, вводили или PBS (носитель; группа 1), 1 мг/кг веса тела hIgG1-MOR03080 (группа 2) или 5 мг/кг м.т. hIgG1-MOR03080 (группа 3), или 5 мг/кг м.т. chIgG2a-MOR03080 (группа 4). Группа 6 не получала никакого лечения (см. Таблицу 6). Объемы опухоли, массу тела и клинические симптомы измеряли два раза в неделю до конца исследования.
2.6 Результаты
Клинические наблюдения и летальность
Не выявлено никаких специфических клинических проявлений или летальности, связанных с опухолью или вводимыми веществами. В группе 3 (5 мг/кг hIgG1) четверо животных умерли во время забора крови (одна мышь на 3 день, одна на 34 день; две на 52 день). В группе 4 (1 мг/кг muIgG2a) единственное животное умерло во время забора крови (на 34 день). Всех остальных животных, умерших в ходе исследования, специально умерщвляли в связи с размером опухоли.
Изменение массы тела
Не выявлено никакого влияния на изменение массы, связанного с лекарственным средством, по сравнению с группой 1 (носитель). На массу тела значительно влиял забор крови в группе 3 (5 мг/кг hIgG1) и в группе 4 (5 мг/кг muIgG2a). Несмотря на такие разрывы, среднее значение увеличения массы во всех группах было постоянным.
Развитие опухоли (см. Фиг.16)
В группе 1 (носитель) рост опухоли происходил с ожидаемой скоростью при медленном прогрессировании. В силу того, что эта клеточная линия имеет сильно выраженные стандартные отклонения, из дальнейшего статистического анализа были исключены значения для самой большой и самой маленькой опухоли. Рост опухоли у животных в группе 1 был сопоставим с ростом опухоли в группе 6 (без лечения), притом, что в этой группе начальный средний объем опухоли на 31 день был меньше. Таким образом, лечение могло оказывать небольшое влияние на скорость роста опухоли. В группе 1 пришлось подвергнуть умерщвлению двух мышей перед 83 днем в связи с размерами опухоли, и дополнительно одну мышь перед 87 днем, поэтому среднее значение объема опухоли после 80 дня было нерепрезентативным. В группе 6 пришлось подвергнуть умерщвлению одну мышь перед 80 днем в связи с размерами опухоли, двух мышей перед 83 днем, и дополнительно перед 87 днем, поэтому среднее значение объема опухоли после 76 дня было нерепрезентативным.
В группе 2, которой водили 1 мг/кг массы тела hIgG1, одно животное исключили из дальнейшего анализа в силу того, что опухоль переросла в мышечную ткань, и обычно это является причиной увеличения скорости роста опухоли. По сравнению с контрольной группой 1 (носитель) средние размеры опухоли начинали значительно различаться, начиная с 45 дня до конца исследования. После окончания лечения не было выявлено никакого увеличения роста опухоли (день 68).
У животных в группе 3 (5 мг/кг м.т. hIgG1) отмечалось заметное уменьшение роста опухоли по сравнению с группой 1 (носитель), которое становилось статистически значимым с 38 дня до 83 дня. Средний объем опухоли вновь начинал значительно расти примерно через две недели после окончания лечения. Одна из десяти опухолей исчезла на 45 день и не начинала повторно расти вплоть до 19 дня после окончания лечения
Лучшие показатели у всех леченых групп, с начальным объемом опухоли в 92 мм3, выявили в группе 4 (5 мг/кг м.т. muIgG2a), где проявлялся явный регресс среднего объема опухоли, и у 4 животных даже выявили исчезновение опухоли до окончания периода наблюдения. Различия в среднем объеме опухоли группы 1 (носитель) были очень значительными, начиная с 38 дня и до конца исследования.
Начальное введение 1 мг/кг м.т. hIgG1 между 14 и 36 днями (группа 5) оказывало ранний и продолжительный эффект на развитие опухоли, одно животное было исключено из дальнейшего анализа, поскольку опухоль переросла в мышечную ткань. На 31 день только пять животных имели поддающуюся измерению опухоль в месте инокулирования, по сравнению с остальной частью инокулированных животных, среди которых только 2 из 60 не отвечали на инокулирование опухоли. Прогрессирование опухоли было отложено примерно на 31 день (при сравнении 52 дня из контрольной группы 1 с 83 днем из группы 5). Приблизительно у 50% животных в месте инокулирования в конце исследования опухоль не выявлялась.
2.7 Выводы
Не выявлено никаких специфичных опухолевых или связанных с веществами клинических данных или данных по летальности по сравнению с группой 1 (контроль).
Не выявлено никакого влияния на изменение массы, связанного с лекарственным средством.
Рост опухоли из опухолевых клеток RPMI8226 после лечения уменьшался следующим образом, в порядке эффективности: 1 мг/кг hIgG1, с 14 до 36 дня через день (группа 5)>5 мг/кг muIgG2a, с 32 до 68 дня через день (группа 4)>5 мг/кг hIgG1, с 32 до 68 дня через день (группа 3)>1 мг/кг hIgG1, с 32 до 68 дня через день (группа 2). В группах 2-4 средние объемы опухоли снова увеличивались после окончания лечения в варьируемых пределах.
Литература
Ausiello C.M., Urbani F., Lande R., la Sala A., Di Carlo В., Baj G., Surico N., Hilgers J., Deaglio S., Funaro A., Malavasi F. (2000) Functional topography of discrete domains of human CD38. Tissue Antigens. 2000 Dec; 56(6):539-47.
Chamow. S.M., Zhang, D.Z., Tan, X.Y, Mathre, S.M., Marsters, SA., Peers, D.H., Byrn, R.A., Ashknazi, A., Junghans, R.P (1994). Humanized, bispecific immunoadhesin-antibody that retargets CD3+ effectors to kill HIV-1-infected cells. J Immunol. 1994 Nov 1; 153(9):4268-80.
Dattamajumdar, A.K., Jacobsen, D.P., Hood, L.E., Osman, G.B. (1996). Rapid cloning of rearranged mouse immunoglobulin variable genes. Immunogentetics 43, 141-151.
Funaro, A., Spagnoli, G.C., Ausiello, C.M., Alessio, M., Roggero, S., Delia, D., Zaccolo, M., and Malavasi, F. (1990) Involvement of the multilineage CD38 molecule in a unique pathway of cell activation and proliferation. J. Immunol. 145, 2390-2396.
Hoshino S., Kukimoto I., Kontani K., Inoue S., Kanda Y., Malavasi F., Katada T. (1997). Mapping of the catalytic and epitopic sites of human CD38/NAD+ glycohydrolase to a functional domain in the caiboxyl terminus. J Immunol. 158(2):741-7.
Jackson D.G., Bell J.I. (1990) Isolation of a cDNA encoding the human CD38 (T10) molecule, a cell surface glycoprotein with an unusual discontinuous pattern of expression during lymphocyte differentiation. J Immunol. 144(7):2811-5.
Knappik, A., Ge, L., Honegger, A., Pack, Р., Fischer, M., Wellnhofer. G., Hoess, A., Wolle, J., Pluckthun A., and Virnekas, B. (2000). Fully synthetic human combinatorial antibody libraries (HuCAL) based on modular consensus frameworks and CDRs randomized with trinucleotides. J Mol Biol 296, 57-86.
Konopleva M., Estrov Z., Zhao S., Andreeff M., Mehta K. (1998) Ligation of cell surface CD38 protein with agonistic monoclonal antibody induces a cell growth signal in myeloid leukemia cells. J Immunol. 161(9):4702-8.
Krebber, A., Bornhauser, S., Burmester, J., Honegger, A., Willuda, J., Bossard, H.R., Plückthun, A. (1997). Reliable cloning of functional antibody variable domains from hybridomas and spleen cell repertoires employing a reengineered phage display system. J. Imm. Meth. 201, 35-55.
Krebs, B., Rauchenberger, R., Reiffert, S., Rothe, C., Tesar, M., Thomassen, E., Cao, M., Dreier, T., Fischer, D., Hoss, A., Inge, L., Knappik, A., Marget, M., Pack, P., Meng, X.Q., Schier, R., Sohlemann, P., Winter, J., Wolle, J., and Kretzschmar, T. (2001). High-throughput generation and engineering of recombinant human antibodies. J Immunol Methods 254, 67-84.
Löhning, С. (2001). Novel methods for displaying (poly)peptides/proteins on bacteriophage particles via disulfide bonds. WO 01/05950.
Malavasi, F., Caligaris-Cappio, F., Milanese, C., Dellabona, P., Richiardi, P., Carbonara, A.O. (1984). Characterization of a murine monoclonal antibody specific for human early lymphohemopoietic cells. Нum. Immunol 9:9-20.
Namba, M., Otsuki, Т., Mori, Togawa, A., Wada, H., Sugihara, Т., Yawata, Y., Kimoto, T. (1989). Establishment of five human myeloma cell lines. In Vitro Cell Dev. Biol. 25:723.
Nata K., Takamura Т., Karasawa Т., Kumagai Т., Hashioka W., Tohgo A., Yonekura H., Takasawa S., Nakamura S., Okamoto H. (1997). Human gene encoding CD38 (ADP-ribosyi cyclase/cyclic ADP-ribose hydrolase): organization, nucleotide sequence and alternative splicing. Gene 186(2):285-92.
Naundorf, S., Preithner, S., Mayer, P., Lippold, S., Wolf, A., Hanakam, F., Fichtner, I., Kufer, P., Raum, Т., Riethmller, G., Baeuerle, P.A., Dreier, T. (2002). Int J. Cancer 100, 101-110.
Pluckthun A, and Pack P. (1997) New protein engineering approaches to multivalent and bispecific antibody fragments. Ummunotechnology 3(2):83-105.
Rauchenberger R., Borges E., Thomassen-Wolf E., Rom E., Adar R., Yaniv Y., Malka M., Chumakov L, Kotzer S., Resnitzky D., Knappik A., Reiffert S., Prassler J., Jury K., Waldherr D., Bauer S., Kretzschmar Т., Yayon A., Rothe C. (2003). Human combinatorial Fab library yielding specific and functional antibodies against the human fibroblast growth factor receptor 3. J Biol Chem. 278(40):38194-205.
Zhou, H., Fisher, R.J., Papas, T.S. (1994). Optimization of primer sequences for mouse scFv repertoire display library construction. Nucleic Acids Res. 22:888-889.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ MST1R И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2534890C2 |
| ANTI-CXCR4 АНТИТЕЛА И КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВО | 2014 |
|
RU2675516C2 |
| АНТИ-GM-CSF АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2447085C2 |
| Молекулы, связывающиеся с 4-1ВВ | 2011 |
|
RU2710717C2 |
| ВЫДЕЛЕННОЕ АНТИ-CD30 АНТИТЕЛО (ВАРИАНТЫ), ХОЗЯЙСКАЯ КЛЕТКА, СПОСОБ ПОЛУЧЕНИЯ ХИМЕРНОГО ИЛИ ГУМАНИЗИРОВАННОГО ВАРИАНТА АНТИ-CD30 АНТИТЕЛ (ВАРИАНТЫ), СПОСОБ ИНГИБИРОВАНИЯ РОСТА КЛЕТОК CD30+ И СПОСОБ ИНГИБИРОВАНИЯ РОСТА ОПУХОЛЕВЫХ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ CD30 | 2006 |
|
RU2492186C2 |
| АНТИТЕЛА РЕЦЕПТОРА 1 ИНТЕРФЕРОНА АЛЬФА И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2600884C2 |
| АНТИТЕЛА, НАЦЕЛЕННЫЕ НА РЕЦЕПТОР, СВЯЗАННЫЙ С G-БЕЛКАМИ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2725819C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ РАКА | 2013 |
|
RU2631804C2 |
| НОВЫЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1(PD-1) | 2016 |
|
RU2757316C2 |
| АНТИ-С4.4А АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2577977C2 |




























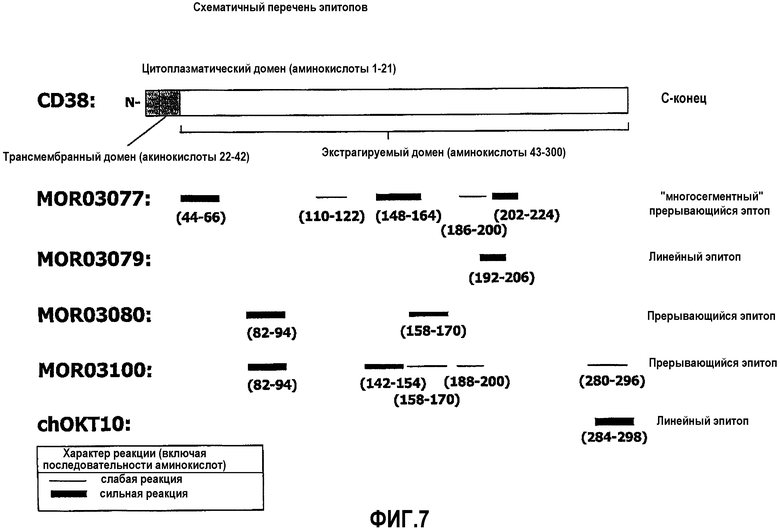

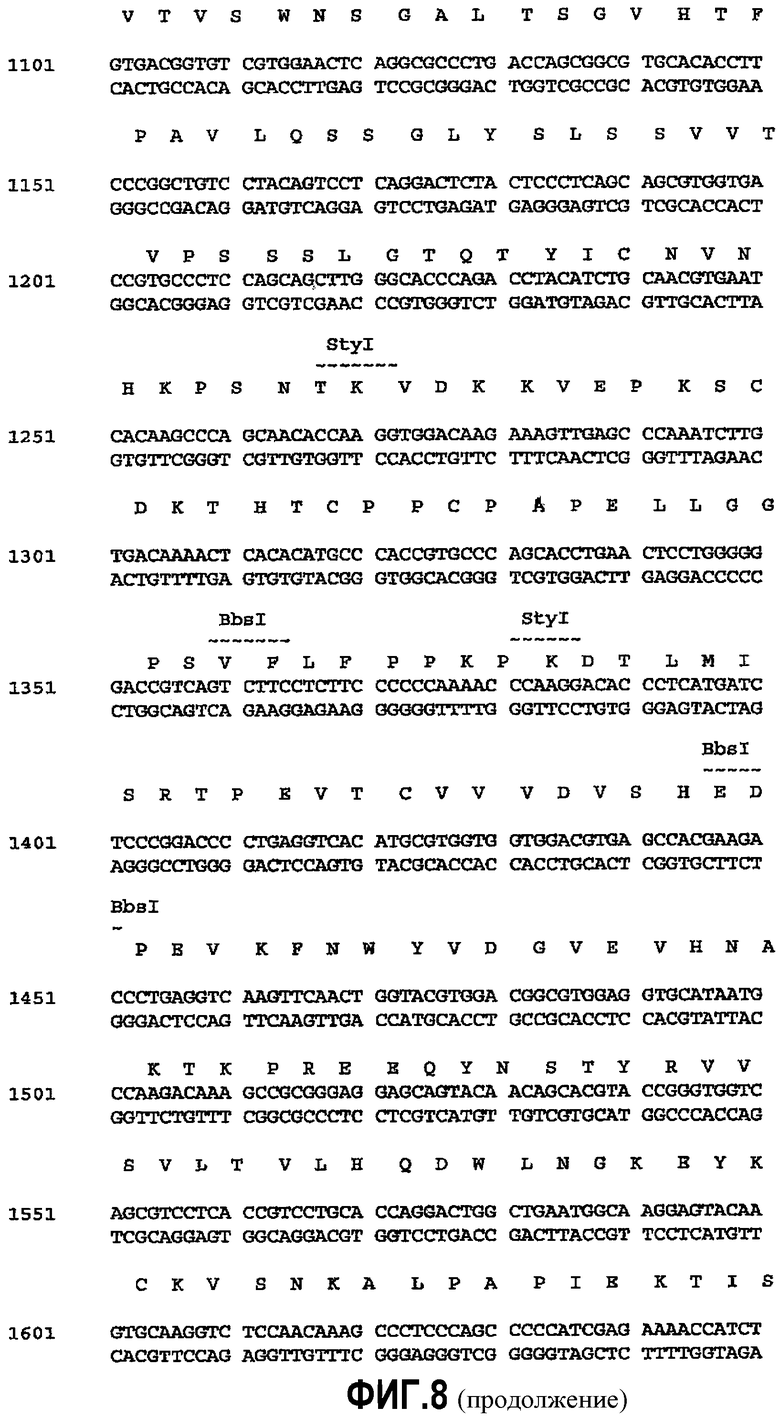





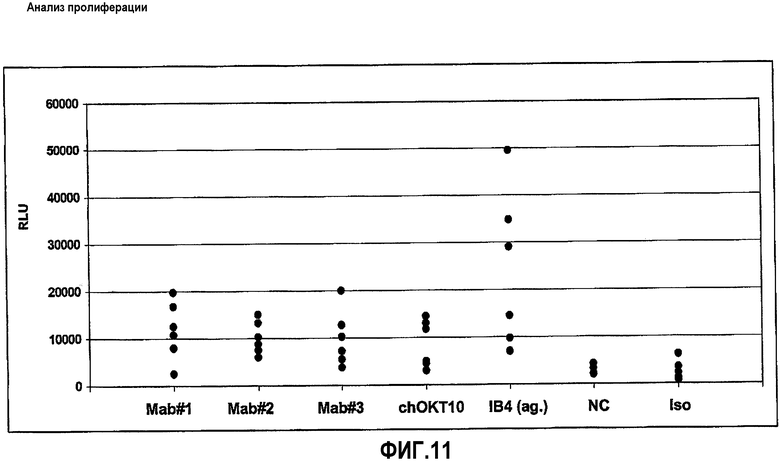
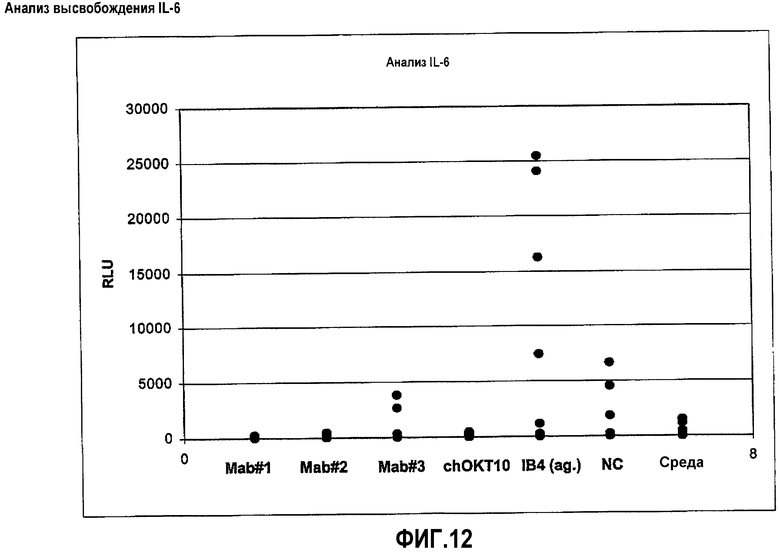
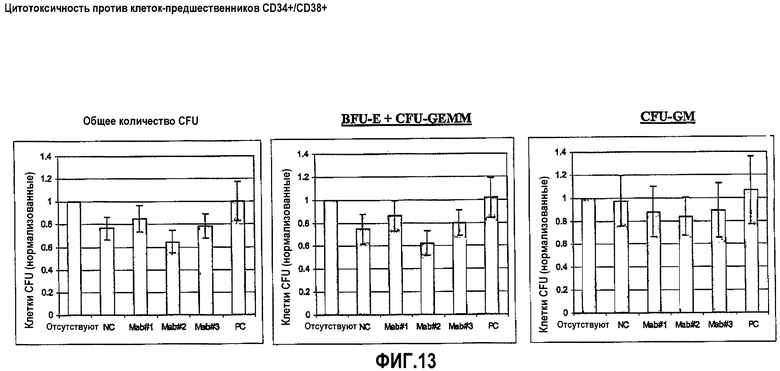
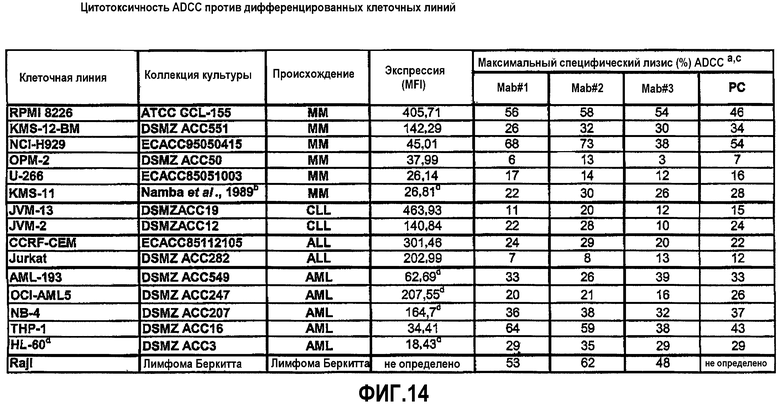

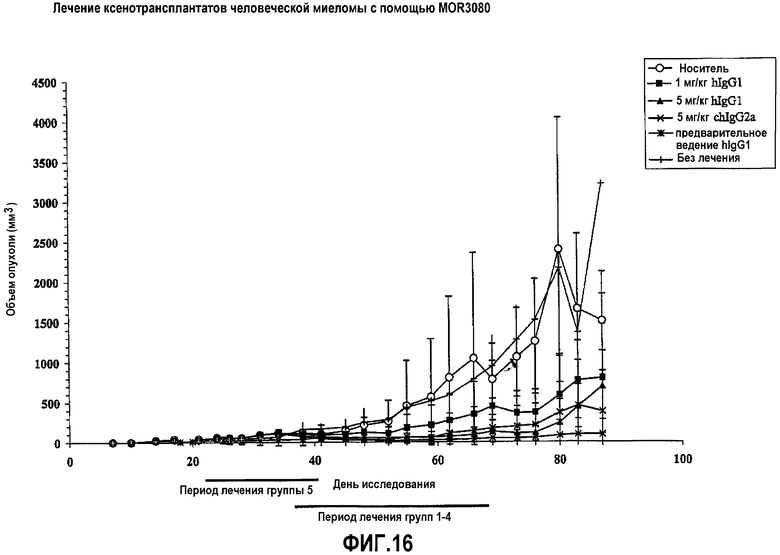
Изобретение относится к биотехнологии. Описаны выделенные антитела человека или гуманизированные антитела, или их функциональные фрагменты, специфичные в отношении эпитопа CD38. Представлены вариабельные области тяжелой и легкой цепи описанных антител. Предложена фармацевтическая композиция, содержащая описанное антитело, или его функциональный фрагмент. Предложен способ лечения нарушения или состояния, связанного с нежелательным присутствием клеток CD38+, включающий введение индивиду эффективного количества предложенной фармацевтической композиции. Описан выделенный эпитоп CD38. Изобретение позволяет эффективно опосредовать лизис клеток, сверхэкспрессирующих CD38. 8 н. и 45 з.п. ф-лы, 18 ил., 8 табл.
1. Выделенное антитело человека, или гуманизированное антитело, или его функциональный фрагмент, специфичное в отношении эпитопа CD38 (SEQ ID NO:22), при этом указанное антитело, или его функциональный фрагмент, способно опосредовать лизис клеток-мишеней CD38+ путем ADCC (антитело-зависимой клеточной цитотоксичности), с эффективностью, по меньшей мере, в пять раз превышающей эффективность химерного ОКТ 10 (SEQ ID NO:23 и 24) в одинаковых или, по существу, одинаковых условиях при использовании клеток РВМС человека в качестве эффекторных клеток, где указанные клетки-мишени CD38+ выбраны из группы, состоящей из LP-1 (DSMZ: АСС41) и RPMI-8226 (АТСС: CCL-155), и где отношение эффекторных клеток к клеткам-мишеням составляет приблизительно от 30:1 примерно до 50:1, и где выделенное антитело человека, его функциональный фрагмент, специфично в отношении эпитопа CD38, при этом указанный эпитоп содержит один или более аминокислотных остатков из аминокислотных остатков 1-215 CD38 (SEQ ID NO:22); предпочтительно эпитоп содержит один или более аминокислотных остатков, содержащихся в одном или более из аминокислотных отрезков, взятых из списка аминокислотных отрезков 44-66, 82-94, 142-154, 148-164, 158-170 и 192-206 CD38.
2. Выделенное антитело или его функциональный фрагмент по п.1, которое является антителом человека или его функциональным фрагментом.
3. Выделенное антитело или его функциональный фрагмент по п.1 или 2, содержащее область H-CDR3, область H-CDR2 и область H-CDR1, которые, по существу, соответствуют SEQ ID NO:5, 6, 7 или 8; и содержащее область L-CDR3, область L-CDR2 и область L-CDR1, которые, по существу, соответствуют SEQ ID NO:13, 14, 15 или 16.
4. Выделенное антитело или его функциональный фрагмент по п.3, где указанное антитело или его функциональный фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:5; и где указанное антитело или его функциональный фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:13.
5. Выделенное антитело или его функциональный фрагмент по п.4, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:5, и вариабельную область легкой цепи, представленную как SEQ ID NO:13.
6. Выделенное антитело или его функциональный фрагмент по п.3, где указанное антитело или его функциональный фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:6; и где указанное антитело или его функциональный фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:14.
7. Выделенное антитело или его функциональный фрагмент по п.6, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:6, и вариабельную область легкой цепи, представленную как SEQ ID NO:14.
8. Выделенное антитело или его функциональный фрагмент по п.3, где указанное антитело или его функциональный фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:7; и где указанное антитело или его функциональный фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:15.
9. Выделенное антитело или его функциональный фрагмент по п.8, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:7, и вариабельную область легкой цепи, представленную как SEQ ID NO:15.
10. Выделенное антитело или его функциональный фрагмент по п.3, где указанное антитело или его функциональный фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:8; и где указанное антитело или его функциональный фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:16.
11. Выделенное антитело или его функциональный фрагмент по п.10, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:8, и вариабельную область легкой цепи, представленную как SEQ ID NO:16.
12. Выделенное антитело или его функциональный фрагмент по п.1 или 2, содержащее вариабельную область тяжелой цепи, представленную как SEQ ID NO:5, 6, 7 или 8.
13. Выделенное антитело или его функциональный фрагмент по п.1 или 2, содержащее вариабельную область легкой цепи, представленную как SEQ ID NO:13, 14, 15 или 16.
14. Выделенное антитело или его функциональный фрагмент по п.1 или 2, которое представляет собой Fab или scFv.
15. Выделенное антитело по п.1 или 2, которое представляет собой IgG.
16. Выделенное антитело по п.15, где указанный IgG представляет собой IgGl.
17. Выделенное антитело человека, или гуманизированное антитело или его функциональный фрагмент специфичное в отношении эпитопа CD38 (SEQ ID NO:22), при этом указанное антитело или его функциональный фрагмент способно опосредовать гибель CD38-трансфицированной клетки СНО путем CDC (комплемент-зависимой цитотоксичности), с эффективностью, по меньшей мере в два раза превышающей эффективность химерного ОКТ 10 (SEQ ID NO:23 и 24) в одинаковых или, по существу, одинаковых условиях, где указанное антитело или его функциональный фрагмент специфично в отношении эпитопа CD38, при этом указанный эпитоп содержит один или более аминокислотных остатков из аминокислотных остатков 1-215 CD38 (SEQ ID NO:22); предпочтительно указанный эпитоп содержит один или более аминокислотных остатков, содержащихся в одном или более из аминокислотных отрезков, взятых из списка аминокислотных отрезков 44-66, 82-94, 142-154, 148-164, 158-170 и 192-206 CD38.
18. Выделенное антитело или его функциональный фрагмент по п.17, которое является антителом человека или его функциональным фрагментом.
19. Выделенное антитело или его функциональный фрагмент по п.17 или 18, содержащее область H-CDR3, область H-CDR2 и область H-CDR1, которые, по существу, соответствуют SEQ ID NO:5, 6, 7 или 8; и содержащее область L-CDR3, область L-CDR2 и область L-CDR1, которые, по существу, соответствуют SEQ ID NO:13, 14, 15 или 16.
20. Выделенное антитело или его функциональный фрагмент по п.19, где указанное антитело или его функциональный фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:5; и где указанное антитело или его функциональный фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:13.
21. Выделенное антитело или его функциональный фрагмент по п.20, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:5, и вариабельную область легкой цепи, представленную как SEQ ID NO:13.
22. Выделенное антитело или его функциональный фрагмент по п.19, где указанное антитело или его функциональный фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:6; и где указанное антитело или его функциональный фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:14.
23. Выделенное антитело или его функциональный фрагмент по п.22, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:6, и вариабельную область легкой цепи, представленную как SEQ ID NO:14.
24. Выделенное антитело или его функциональный фрагмент по п.19, где указанное антитело или его функциональный фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:7; и где указанное антитело или его функциональный фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:15.
25. Выделенное антитело или его функциональный фрагмент по п.24, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:7, и вариабельную область легкой цепи, представленную как SEQ ID NO:15.
26. Выделенное антитело или его функциональный фрагмент по п.19, где указанное антитело или его функциональный фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:8; и где указанное антитело или его функциональный фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:16.
27. Выделенное антитело или его функциональный фрагмент по п.26, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:8, и вариабельную область легкой цепи, представленную как SEQ ID NO:16.
28. Выделенное антитело или его функциональный фрагмент по п.17 или 18, содержащее вариабельную тяжелую цепь, представленную как SEQ ID NO:5, 6, 7 или 8.
29. Выделенное антитело или его функциональный фрагмент по п.17 или 18, содержащее вариабельную легкую цепь, представленную как SEQ ID NO:13, 14, 15 или 16.
30. Выделенное антитело или его фрагмент по п.17 или 18, которое представляет собой Fab или scFv.
31. Выделенное антитело или его фрагмент по п.17 или 18, которое представляет собой IgG.
32. Выделенное антитело по п.31, где указанный IgG представляет собой IgGl.
33. Выделенное антитело или его функциональный фрагмент, специфичное в отношении эпитопа CD38 (SEQ ID NO:22), содержащее область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:5, 6, 7 или 8; и где указанное антитело или его фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:13, 14, 15 или 16.
34. Выделенное антитело или его фрагмент по п.33, где указанное антитело или его фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:5; и где указанное антитело или его фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:13.
35. Выделенное антитело или его функциональный фрагмент по п.34, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:5, и вариабельную область легкой цепи, представленную как SEQ ID NO:13.
36. Выделенное антитело или его фрагмент по п.33, где указанное антитело или его фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:6; и где указанное антитело или его фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:14.
37. Выделенное антитело или его функциональный фрагмент по п.36, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:6, и вариабельную область легкой цепи, представленную как SEQ ID NO:14.
38. Выделенное антитело или его фрагмент по п.33, где указанное антитело или его фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:7; и где указанное антитело или его фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:15.
39. Выделенное антитело или его функциональный фрагмент по п.38, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:7, и вариабельную область легкой цепи, представленную как SEQ ID NO:15.
40. Выделенное антитело или его фрагмент по п.33, где указанное антитело или его фрагмент содержит область H-CDR1, область H-CDR2 и область H-CDR3, которые представлены как SEQ ID NO:8; и где указанное антитело или его фрагмент содержит область L-CDR1, область L-CDR2 и область L-CDR3, которые представлены как SEQ ID NO:16.
41. Выделенное антитело или его функциональный фрагмент по п.40, где указанное антитело или его функциональный фрагмент содержит вариабельную область тяжелой цепи, представленную как SEQ ID NO:8, и вариабельную область легкой цепи, представленную как SEQ ID NO:16.
42. Выделенное антитело или его функциональный фрагмент по п.33, которое представляет собой Fab или scFv.
43. Выделенное антитело по п.33, которое представляет собой IgG.
44. Выделенное антитело по п.43, где указанный IgG представляет собой IgGl.
45. Вариабельная область тяжелой цепи выделенного антитела или его функционального фрагмента по любому из пп.1-44, кодируемая (i) последовательностью нуклеиновой кислоты, содержащей SEQ ID NO:1, 2, 3 или 4; или (ii) последовательностью нуклеиновой кислоты, которая гибридизируется в жестких условиях с комплементарной цепью SEQ ID NO:1, 2, 3 или 4, где указанное антитело или его функциональный фрагмент специфично в отношении эпитопа CD38, или (iii) последовательностью нуклеиновой кислоты, которая на 85% идентична последовательности SEQ ID NO:1, 2, 3 или 4, где указанное антитело или его функциональный фрагмент специфично в отношении эпитопа CD38.
46. Вариабельная область тяжелой цепи по п.45, которая кодируется последовательностью нуклеиновой кислоты, содержащей SEQ ID NO:1, 2, 3 или 4.
47. Вариабельная область легкой цепи выделенного антитела или его функционального фрагмента по любому из пп.1-44, кодируемая (i) последовательностью нуклеиновой кислоты, которая по меньшей мере на 85% идентична SEQ ID NO:9, 10, 11 или 12; или (ii) последовательностями нуклеиновых кислот, которые гибридизируются в жестких условиях с комплементарной цепью SEQ ID NO:9, 10, 11 или 12, где указанное антитело или его функциональный фрагмент специфично в отношении эпитопа CD38; или (iii) последовательностью нуклеиновой кислоты, которая на 85% идентична последовательности SEQ ID NO:9, 10, 11 или 12, где указанное антитело или его функциональный фрагмент специфично в отношении эпитопа CD38.
48. Вариабельная область легкой цепи по п.47, которая кодируется последовательностью нуклеиновой кислоты, содержащей SEQ ID NO:9, 10, 11 или 12.
49. Фармацевтическая композиция для анти-СD38 терапии, содержащая эффективное количество антитела или его функционального фрагмента по любому из пп.1-44 и фармацевтически приемлемый носитель или эксципиент.
50. Способ лечения нарушения или состояния, связанного с нежелательным присутствием клеток CD38+, включающий введение индивиду эффективного количества фармацевтической композиции по п.49.
51. Способ по п.50, где указанное нарушение или состояние представляет собой гематологическое заболевание, множественную миелому, хронический лимфоцитарный лейкоз, хронический миелолейкоз, острый миелолейкоз и острый лимфоцитарный лейкоз.
52. Способ по п.50, где указанное нарушение или состояние представляет собой воспалительное заболевание, выбранное из ревматоидного артрита и системной красной волчанки.
53. Выделенный эпитоп CD38, по существу, состоящий из аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 44-66, 82-94, 142-154, 148-164, 158-170, 192-206 и 202-224 CD38; или выделенный эпитоп CD38, состоящий из аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 44-66, 82-94, 142-154, 148-164, 158-170, 192-206 и 202-224 CD38; или набор, содержащий выделенный эпитоп CD38, содержащий одну или несколько аминокислотных фрагментов из перечня 44-66, 82-94, 142-154, 148-164, 158-170, 192-206 и 202-224, и библиотека антител и инструкция для применения.
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| RU 2002100905 А, 23.06.2000. | |||

