Перекрестная ссылка на родственные заявки
По заявке испрашивается приоритет предварительной заявки США, серийный номер 60/467188, поданной 1 мая 2003, которая приведена здесь в качестве ссылки.
Изложение интереса правительства
Изобретение сделано при поддержке правительства Соединенных Штатов, гранты № EY11254 и EY12598 Национальных Институтов Здоровья. Правительство Соединенных Штатов обладает определенными правами на настоящее изобретение.
Область техники, к которой относится изобретение
В целом, изобретение относится к антагонистам молекул клеточной адгезии млекопитающих. Более конкретно, изобретение относится к селективным пептиднам, которые являются антагонистами R-кадгерина (кадгерина-4) млекопитающих, и способам ингибирования клеточной адгезии и, следовательно, ангиогенеза сетчатки.
Уровень техники
Семейство молекул кадгерина состоит из трансмембранных гликопротеидов, которые функционируют в кальций-зависимых селективных межклеточных взаимодействиях. Эти молекулы играют важную роль в ходе эмбрионального развития и морфогенеза ткани, участвуя в клеточном распознавании и сортировке клеток. Подсемейства кадгеринов (классические кадгерины, протокадгерины, десмоколлины и другие белки, относящиеся к кадгеринам) характеризуются разнообразными внеклеточными доменами кадгерина, единичным трансмембранным сегментом и единичным цитоплазматическим доменом. Как сообщается, так называемые классические кадгерины (то есть E, P, N и R-кадгерин) содержат пять тандемно повторяющихся внеклеточных доменов кадгерина (EC1-EC5), которые участвуют, предпочтительно, в гомофильных взаимодействиях, и высококонсервативный цитоплазматический хвост, который опосредует специфические сигналы адгезии в клетке.
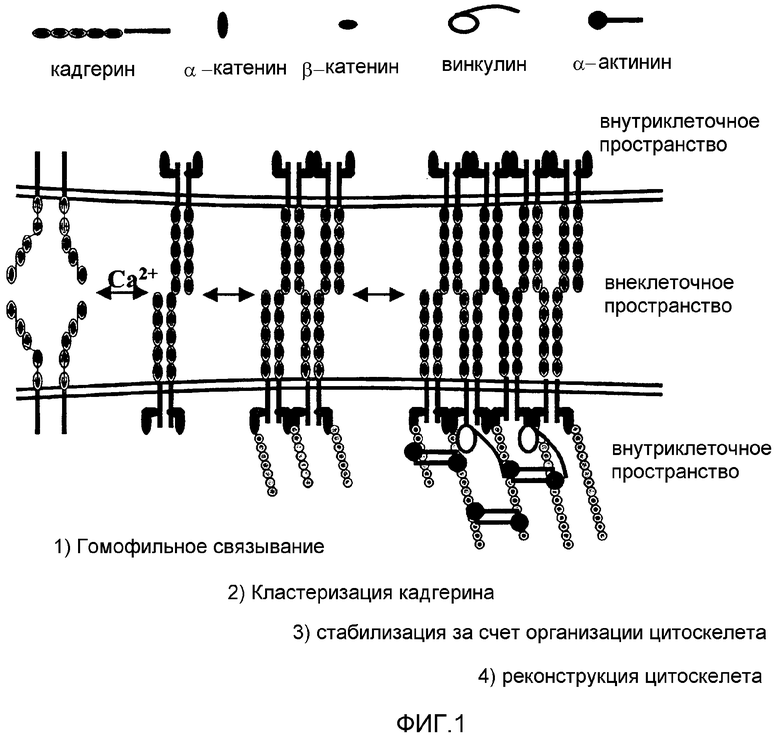
Опосредованная кадгерином межклеточная адгезия происходит как экспрессия множества молекул кадгерина при взаимодействии прилегающих клеток, что приводит к формированию адгезивных связей. Согласно модели зиппера для кадгерина, предложенной Shapiro et al. Nature 1995; 374 (6520): 327-37, молекулы кадгерина в мембране одной клетки образуют плотные димеры с параллельными цепочками (то есть так называемые цис-димеры). Как показано на фиг.1, эти цис-димеры затем связываются с димерами кадгерина, экспрессированными на прилегающих клетках (то есть происходит транс-димеризация). Когда поддерживается достаточное взаимодействие, может возникать кластеризация кадгерина, поскольку все большее количество молекул кадгерина вовлекается в сайт взаимодействия, что приводит к переплетению молекул поверхностей двух клеток. Таким образом, относительно слабые взаимодействия могут объединяться с образованием довольно прочной межклеточной адгезии.
Сначала, при адгезии кадгерина внутриклеточные сигналы, передаваемые через взаимодействия цитоплазматических хвостов кадгерина с молекулами α и β катенина, приводят к преобразованию цитоскелета. Несмотря на то, что объединение с актиновыми филаментами, как считается, не влияет на гомофильное связывание, их объединение помогает удерживать молекулы кадгерина в местах взаимодействия. В симбиотическом типе взаимоотношения кластеризация кадгерина вызывает преобразование цитоскелета и обеспечивает точки присоединения на мембране, которые являются важными для клеточных изменений, происходящих после формирования адгезивных контактов. В это время объединение с цитоскелетом удерживает кадгерины в местах взаимодействия и помогает вовлекать новые молекулы кадгерина, таким образом приводя к кластеризации кадгерина. В ходе кластеризации кадгерина кальций играет важную роль кофактора. В растворах с недостаточной концентрацией ионов кальция (то есть приблизительно ниже 2 мМ) кадгерин теряет свою функцию, и молекулы становятся более чувствительными к протеазной деградации. Кальций необходим для стабилизации структуры молекул кадгерина и обеспечивает правильную ориентацию прилегающих поверхностей кадгерина, поэтому в его отсутствие происходит потеря функции кадгерина и протеазная деградация.
Несмотря на то, что, как сообщается, каждый из пяти внеклеточных доменов классического кадгерина, от EC1 до EC5, играет важную роль в обеспечении димеризации кадгерина, на основе мутационного анализа было сделано предположение, что большинство остатков, которые образуют поверхность димеризации, было обнаружено на N-конце самого большего домена кадгерина (EC1) (Kitagawa, et al., Biochem. Biophys. Res. Commun., 2000; 271(2):358-63). Однако о механизмах специфичной гомодимеризации между молекулами кадгерина известно относительно немного.
Кадгерины играют значительную роль в контроле нейронов и развитии центральной нервной системы. Как сообщается, различные отделы мозга определяются экспрессией различных типов кадгерина при дифференциации. Кадгерины также играют важную роль в невральном развитии сетчатки за счет специфичной экспрессии в различных областях развивающейся сетчатки. Например, как сообщается, в ходе развития сетчатки эмбриона курицы, B-кадгерин обнаружен только в глии Мюллера, в то время как определенные популяции биполярных клеток экспрессируют R-кадгерин (также известный, как кадгерин-4). Амакринные клетки и субпопуляция ганглионарных клеток экспрессируют кадгерины 6B и 7. Во внутреннем плексиформном слое сетчатки эти же кадгерины экспрессируются только в подслоях, связанных с синапсин-I-позитивными нервными окончаниями, что дает основание для предположения, что различные профили экспрессии вносят свой вклад в формирование синапса между специфическими субпопуляциями нейронов в ходе развития сетчатки. В зародышевом зрительном нерве разрастание аксона ганглионарной клетки опосредовано адгезией N-кадгерина с глиальными клетками, экспрессирующими R-кадгерин.
Адгезия кадгеринов также играет роль в васкуляризации сетчатки при развитии (Dorrell, et al. Invest. Ophthalmol. Vis. Sci. 2002; 43 (11):3500-10). Нарушение адгезии R-кадгерина в ходе формирования поверхностного сплетения сосудов приводит к потере сложных соединений сосудов, наблюдаемых при нормальном формировании сосудов. При блокировании адгезии R-кадгерина в ходе последующего формирования глубоких сосудистых слоев ключевые направляющие сигналы утрачиваются, что приводит к миграции сосуда за пределы нормальной глубины сплетения сосудов в слой зрительных рецепторов.
Сетчатка состоит из четких слоев нейронных, глиальных и сосудистых элементов. Любое заболевание или состояние, которое даже незначительно изменяет слои сетчатки, может привести к дегенерации нейронов и значительной потере зрения. В течение более чем 70 лет в качестве модели для многих заболеваний, которые приводят к гибели клеток зрительных рецепторов, исследовали мышь с дегенерацией сетчатки (мышь rd/rd). У мыши rd/rd, дегенерация зрительных рецепторов начинается в первые три недели после рождения, поскольку палочковидные клетки подвергаются апоптозу, который возникает в результате мутации в β субъединице цГМФ-зависимой фосфодиэстеразы, что приводит к гибели колбочек, зрительных рецепторов. Сосудистая атрофия в сетчатке также связана во времени с гибелью клеток зрительных рецепторов у мышей rd/rd. По всей видимости, нормальное формирование сосудистой сети происходит при развитии трех функциональных слоев в течение первых трех недель. Однако когда сосуды в глубоком сосудистом слое начинают дегенерировать в течение второй недели, и к концу четвертой недели после рождения наблюдают существенное уменьшение числа сосудов, происходит почти полное исчезновение глубоких и промежуточных сосудистых сплетений.
В нормальном кровотоке и костном мозге взрослого организма присутствует популяция гематопоэтических стволовых клеток, от которой могут дифференцироваться различные субпопуляции клеток по положительной линии (Lin+HSC) или отрицательной линии (Lin-HSC). Кроме того, авторы настоящего изобретения обнаружили, что в субпопуляции Lin-HSC присутствуют эндотелиальные клетки-предшественники (EPC), способные формировать кровеносные сосуды in vitro и in vivo. EPC в популяции Lin-HSC могут воздействовать и стабилизировать дегенерирующую сосудистую сеть у мышей rd/rd при внутриглазном введении мышам. В поверхностный сосудистый слой вводили астроциты, мишени Lin-HSC и наблюдали за эндогенной развивающейся сосудистой сетью после инъекции на 2 день после рождения (P2). При достижении эндогенной сосудистой сетью периферии сетчатки, куда нацеливались Lin-HSC, клетки встраивались в новые кровеносные сосуды с образованием функциональных мозаичных сосудов со смешанными популяциями введенных Lin-HSC и эндогенными эндотелиальными клетками. Кроме того, Lin-HSC нацеливаются на области глубоких и промежуточных сосудистых слоев сетчатки до возникновения васкуляризации этих слоев за счет эндогенных эндотелиальных клеток. Включение Lin-HSC восстанавливает глубокую сосудистую сеть мышей rd/rd приблизительно в 2-3 раза сильнее по сравнению с нормальными мышами и контрольными мышами, которым вводили Lin+HSC. Кроме того, восстановление глубокой сосудистой сети предотвращает деградацию зрительных рецепторов во внешнем ядерном слое сетчатки. Однако, поскольку нет никакого основания для предложения, что эти стволовые клетки могут подвергаться дифференциации в нейроны сетчатки или глиальные клетки, механизм защиты нейронов остается неизвестным.
Нацеливание Lin-HSC на астроциты и глубокие сосудистые области перед естественной васкуляризацией предполагает, что Lin-HSC экспрессируют молекулы клеточной поверхности, которые действуют при нацеливании, сходном с нацеливанием эндогенных эндотелиальных клеток в ходе развития. Адгезия R-кадгерина играет важную роль в нацеливании эндотелиальных клеток на астроциты и сосудистые сплетения в ходе ангиогенеза сетчатки при развитии.
R-кадгерин был идентифицирован и секвенирован у большого числа млекопитающих. На фиг.2 показана аминокислотная последовательность (SEQ ID NO: 1) варианта препробелка R-кадгерина человека, описанного Kitagawa et al. в базе данных SWISS-PROT под номером NP 001785, версия NP 001785,2, GI: 14589893, соответствующее описание которого приведено здесь в качестве ссылки. SEQ ID NO: 1 включает аминокислотную последовательность IDSMSGR (SEQ ID NO: 2) в положениях 222-228.
На фиг.3 показана аминокислотная последовательность (SEQ ID NO: 3) еще одного варианта препробелка R-кадгерина человека, описанного Tanihara et al. в базе данных SWISS-PROT под номером NP P55283, версия P55283, GI: 1705542, соответствующее описание которого приведено здесь в качестве ссылки. SEQ ID NO: 3 включает аминокислотную последовательность INSMSGR (SEQ ID NO: 4) в положениях 222-228.
На фиг.4 показана аминокислотная последовательность (SEQ ID NO: 5) варианта препробелка R-кадгерина мыши (мышь домашняя), описанного Hutton et al. в базе данных SWISS-PROT под номером NP 033997, версия NP 033997,1, GI:6753376, соответствующее описание которого приведено здесь в качестве ссылки. SEQ ID NO: 5 включает аминокислотную последовательность IDSMSGR (SEQ ID NO: 2) в положениях 219-225.
Неселективные пептидные антагонисты кадгеринов, включая аминокислотную последовательность His-Ala-Val (HAV), описаны Blaschuket al. в патенте США № 6465427, № 63456512, № 6169071, и № 6031072. Blaschuket al. описали как линейные, так и циклические пептидные антагонисты кадгеринов, все из которых могут противодействовать множеству типов молекул кадгерина без различия.
Селективные пептидные антагонисты N-кадгерина, которые содержат аминокислотную последовательность Ile-Asn-Pro (INP), были описаны Williams et al., Mol. Cell Neurosci.,2000; 15(5): 456-64. В то время как пептиды HAV представляют собой неспецифичные антагонисты кадгеринов, пептидные антагонисты INP, описанные Williams et al., являются специфичными в отношении N-кадгерина и не проявляют существенного связывания с другими молекулами кадгерина, такими как R-кадгерин.
Вследствие различного распределения молекул клеточной адгезии в различных тканях организма, существует постоянная потребность в антагонистах, селективных к специфичным молекулам клеточной адгезии, в особенности в антагонистах, селективных к R-кадгерину. Селективные пептиды-антагонисты R-кадгерина настоящего изобретения удовлетворяют этой потребности.
Сущность изобретения
Выделенный пептид, применимый в качестве селективного антагониста R-кадгерина млекопитающих, содержит от 3 до 30 аминокислотных остатков, три соседних остатка пептида имеют аминокислотную последовательность Ile-Xaa-Ser (IXS); где Xaa представляет собой аминокислотный остаток, выбранный из группы, включающей Asp, Asn, Glu и Gln. Предпочтительно, Xaa представляет собой Asp или Asn. В одном из предпочтительных вариантов осуществления пептид содержит по меньшей мере семь аминокислотных остатков, и семь соседних аминокислотных остатков пептида имеют аминокислотную последовательность Ile-Xaa-Ser-Met-Ser-Gly-Arg (SEQ ID NO: 6), где Xaa представляет собой такие же остатки, как определено выше. Настоящее изобретение также относится к фармацевтическим композициям, содержащим пептиды-антагонисты R-кадгерина в фармацевтически приемлемом носителе.
В еще одном предпочтительном варианте осуществления пептид представляет собой циклический пептид, содержащий от 3 до 10 аминокислотных остатков, выстроенных в кольцо, три соседних остатка пептида имеют аминокислотную последовательность Ile-Xaa-Ser; где Xaa представляет собой аминокислотный остаток, выбранный из группы, включающей из Asp, Asn, Glu и Gln. Предпочтительно, Xaa представляет собой Asp или Asn.
Предпочтительный циклический пептид имеет формулу:

где X1 и X2 независимо представляют собой аминокислотный остаток или множество аминокислотных остатков до 10, связанных пептидными связями; Y1 и Y2 независимо представляют собой аминокислотные остатки, связанные друг с другом дисульфидной связью, и Xaa представляет собой аминокислотный остаток, выбранный из группы, включающей Asp, Asn, Glu и Gln.
Особенно предпочтительный циклический пептид имеет аминокислотную последовательность: циклическая Cys-Ile-Xaa-Ser-Cys (SEQ ID NO: 7); где Xaa представляет собой аминокислотный остаток, выбранный из группы, включающей Asp, Asn, Glu и Gln, и пептидное кольцо сформировано с помощью дисульфидной связи между двумя остатками цистеина.
Способ ингибирования опосредованной R-кадгерином клеточной адгезии предусматривает введение клеток, экспрессирующих R-кадгерин, в контакт с ингибирующим адгезию количеством селективного пептида-антагониста R-кадгерина настоящего изобретения. Например, ангиогенез сетчатки ингибируют путем введения пациенту, страдающему от патологического сосудистого ангиогенеза сетчатки, ингибирующего ангиогенез количества пептида-антагониста R-кадгерина настоящего изобретения. Таким же образом, нацеливание отрицательной линии гематопоэтических стволовых клеток на развивающуюся сосудистую сеть ингибируют путем введения стволовых клеток в контакт с ингибирующим нацеливание на сосудистую сеть количеством селективного пептида-антагониста R-кадгерина настоящего изобретения. Ингибированное нацеливания Lin-HSC, таких как эндотелиальные клетки-предшественники, на развивающуюся сосудистую сеть полезно для лечения заболеваний, связанных с патологическим развитием сосудов, таким как возрастная дегенерация желтого пятна и диабетическая ретинопатия.
Краткое описание чертежей
В разделе чертежей на фиг.1 представлена схема кластеризации кадгерина и кадгерин-регулируемой клеточной адгезии;
на фиг.2 показана аминокислотная последовательность варианта препробелка R-кадгерина человека (SEQ ID NO: 1), содержащего последовательность IDSMSGR (SEQ ID NO: 2) при остатках 222-228;
на фиг.3 показана аминокислотная последовательность варианта препробелка R-кадгерина человека (SEQ ID NO: 3), содержащего последовательность INSMSGR (SEQ ID NO: 4) при остатках 222-228;
на фиг.4 показана аминокислотная последовательность варианта препробелка R-кадгерина мыши (SEQ ID NO: 5), содержащего последовательность IDSMSGR (SEQ ID NO: 2) при остатках 229-225;
на фиг.5 (A) отражена гомология последовательностей при остатках 24-92 N-кадгерина мыши и различных R-кадгеринов (консервативные остатки (синие) и неконсервативные (красные) остатки); отмечается гомология между R-кадгеринами млекопитающих от человека, мыши и крысы, каждый из которых содержит последовательность IDS или INS при остатках 53-55, в отличие от R-кадгерина курицы и N-кадгерина мыши, которые содержат последовательность IDP и INP, соответственно, при остатках 53-55; (B) циклические и линейные пептиды, в этой области, соответствующие остаткам R-кадгерина мыши и человека и N-кадгерина мыши, синтезировали наряду с соответствующими контрольными пептидами: циклический CIDSC (SEQ ID NO: 8), циклический CINPC (SEQ ID NO: 9), IDSMSGR (SEQ ID NO: 2), IDSASGR (SEQ ID NO: 10), INPASGQ (SEQ ID NO: 11), циклический CSDIC (SEQ ID NO: 12) и циклический CRADC (SEQ ID NO: 13); частичные последовательности кадгерина, приведенные на фиг.5 (A), представляют собой сверху вниз: N-кадгерин мыши (SEQ ID NO: 14), R-кадгерин мыши (SEQ ID NO: 15), R-кадгерин крысы (SEQ ID NO: 16), R-кадгерин человека (SEQ ID NO: 17), другой R-кадгерин человека (SEQ ID NO: 18) и R-кадгерин курицы (gallus gallus) (SEQ ID NO: 19);
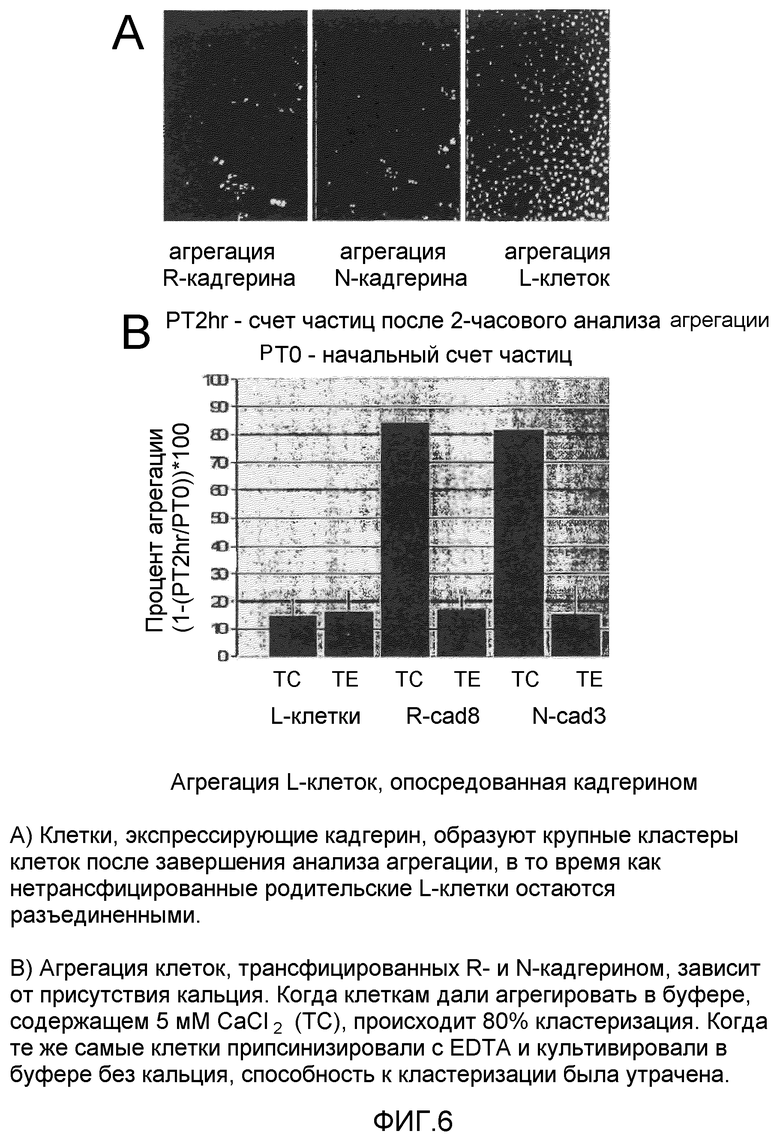
на фиг.6 (A) изображены микрофотографии, демонстрирующие агрегацию L-клеток, экспрессирующих R-кадгерин и N-кадгерин; (B) представляет собой столбчатую диаграмму процента агрегации L-клеток, опосредованной R и N-кадгеринами в присутствии и в отсутствие кальция;
на фиг.7 продемонстрирована устойчивая трансфекция L-клеток R-кадгерином и N-кадгерином; (A) иммуноблоттинг R-кадгерина; (B) иммуноблоттинг N-кадгерина; (C-E) микрофотографии окрашенных L-клеток, демонстрирующих экспрессию R-кадгерина (C) и N-кадгерина (D), по сравнению с клетками, не экспрессирующими ни R-кадгерин, ни N-кадгерин (E);
на фиг.8 графически представлено селективное ингибирование агрегации L-клеток, экспрессирующих кадгерин, с помощью пептидов, содержащих IDS, которые связываются с клетками, экспрессирующими R-кадгерин, по сравнению с пептидами, содержащими INP, которые связываются с клетками, экспрессирующими N-кадгерин;
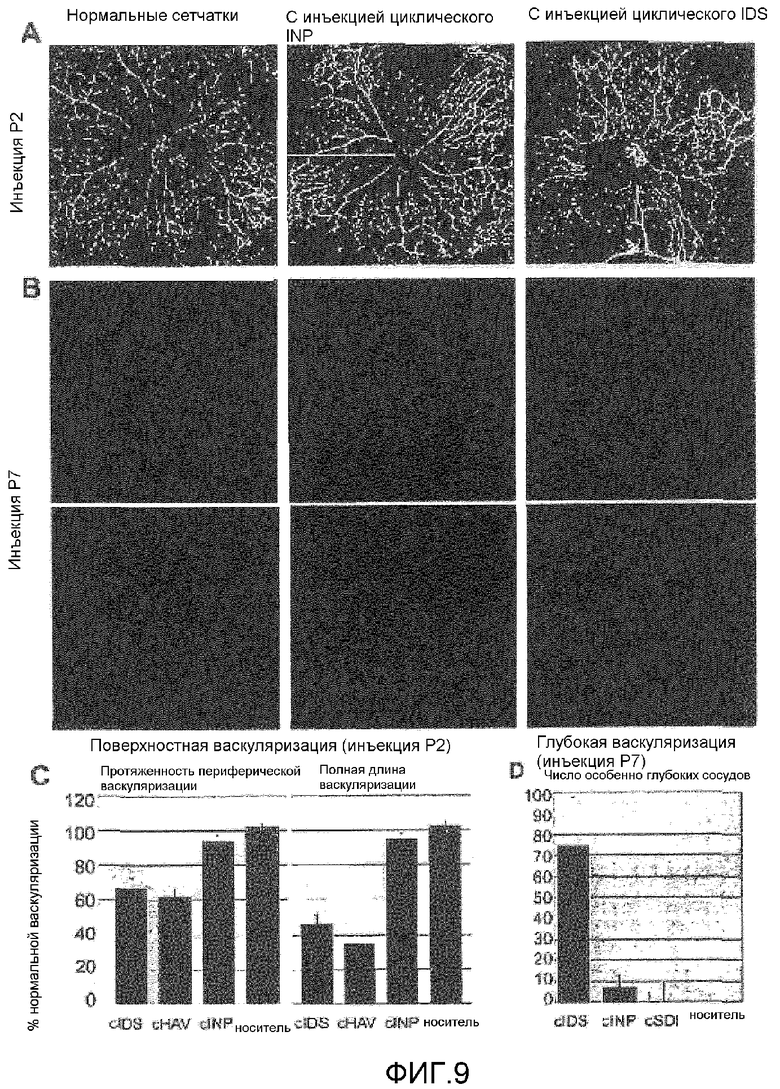
на фиг.9 отражено селективное ингибирование васкуляризации сетчатки мыши после интравитреальной инъекции циклического CIDSC (SEQ ID NO: 8) по сравнению с циклическим CINPC (SEQ ID NO: 9); на (A) отображены микрофотографии сетчаток мышей rd/rd на стадии развития P2; на (B) отображены микрофотографии сетчаток мышей rd/rd на стадии развития P7; (C) представляет собой столбчатую диаграмму поверхностной васкуляризации; (D) представляет собой столбчатую диаграмму глубокой васкуляризации;
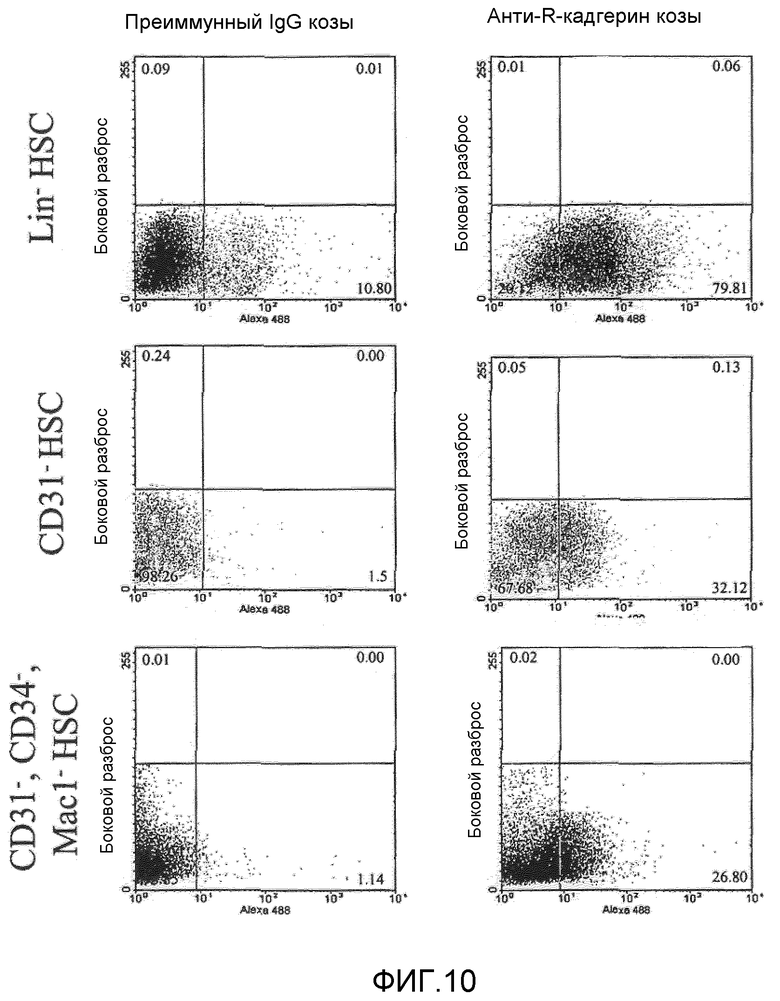
на фиг.10 отражены результаты анализа проточной цитометрии экспрессии R-кадгерина в гематопоэтических стволовых клетках (HSC);
на фиг.11 отображены поперечные микрофотографии сетчаток мышей rd/rd, обработанных различными пептидами изобретения и контрольными пептидами; и
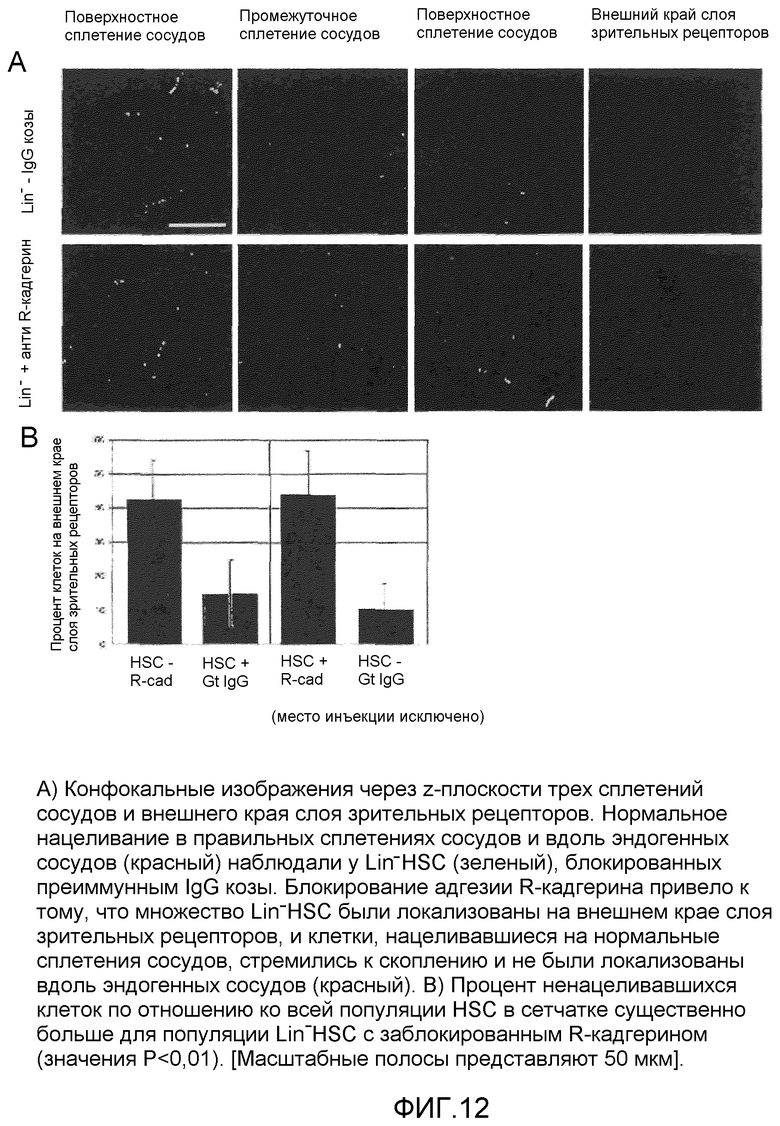
на фиг.12 отражено блокирование нацеливания Lin-HSC на развивающуюся сосудистую сеть сетчатки пептидами-антагонистами R-кадгерина изобретения.
Подробное описание предпочтительных вариантов осуществления
Как используется здесь и в прилагаемой формуле изобретения, термин "циклический пептид" относится к молекулам, содержащим множество аминокислот, соединенных в цепь пептидными связями, концы данной цепи соединены с образованием кольца аминокислотных остатков. Циклический пептид может быть соединен пептидной связью, дисульфидной связью между двумя аминокислотными остатками, такими как остатки цистеина, или посредством любой другой подходящей связующей группы. Непептидные связующие группы могут представлять собой любое химическое соединение, способное реагировать с функциональными группами на каждом конце пептидной цепи для образования связи между ними. Например, два конца пептидной цепи можно связать с помощью небелковой аминокислоты, такой как, например, 3-аминомасляная кислота, или дисульфида, образованного непептидными тиоловыми группами, такими как амид тиогликолевой кислоты на амино-конце и амид, образованный из 2-аминоэтантиола на карбокси-конце.
Как используется здесь и в прилагаемой формуле изобретения, термин "фармацевтически приемлемый" и его грамматические варианты, при указании на носители и другие инертные наполнители, означает, что такие материалы могут использоваться для введения пациенту человеку без образования нежелательных физиологических побочных эффектов, таких как раздражение сетчатки или глаз, тошнота, головокружение, неясное или ослабленное зрение, цитотоксичность и тому подобное.
Термин "аминокислота", как используется здесь и в прилагаемой формуле изобретения, в целом, относится к любой альфа-аминокислоте. Предпочтительно, пептиды настоящего изобретения содержат 21 аминокислоту, кодируемую генетическим кодом, хотя также могут быть включены измененные аминокислотные остатки. Аминокислоты могут быть в L, D или D, L форме. Предпочтительно, пептиды по настоящему изобретению содержат аминокислоты L-формы. Чтобы минимизировать вероятность протеиназнойдеградации in vivo, вводимые пептиды настоящего изобретения могут включать один или более аминокислотных остатков в D-форме.
Выделенный пептид, являющийся селективным антагонистом R-кадгерина млекопитающего, содержит от 3 до 30 аминокислотных остатков, три соседних остатка пептида имеют аминокислотную последовательность Ile-Xaa-Ser. Xaa представляет собой аминокислотный остаток, выбранный из группы, состоящей из Asp, Asn, Glu и Gln. Предпочтительно, Xaa представляет собой Asp или Asn. Пептид-антагонист R-кадгерина настоящего изобретения может быть линейным или циклическим.
Селективные пептиды-антагонисты R-кадгерина по настоящему изобретению воспроизводят последовательности Ile-Asp-Ser и Ile-Asn-Ser, найденные в домене EC1 R-кадгерина млекопитающих, но не в других молекулах кадгерина. Пептиды, содержащие последовательность Ile-Xaa-Ser, могут связываться с молекулами R-кадгерина млекопитающих и противодействовать им. Xaa, предпочтительно, представляет собой остаток аспарагиновой кислоты (Asp) или остаток аспарагина (Asn), для соответствия природной последовательности в молекулах R-кадгерина млекопитающих. Остатки глутаминовой кислоты (Glu) и глутамина (Gln) также подходят в качестве Xaa из-за химического сходства с Asp и Asn, соответственно.
В одном предпочтительном варианте осуществления пептид содержит, по меньшей мере, семь аминокислотных остатков, и семь соседних аминокислотных остатков пептида имеют аминокислотную последовательность Ile-Xaa-Ser-Met-Ser-Gly-Arg (SEQ ID NO: 6). Xaa представляет собой аминокислотный остаток, выбранный из группы, состоящей из Asp, Asn, Glu и Gln. Предпочтительно, Xaa представляет собой Asp или Asn.
В другом предпочтительном варианте осуществления пептид представляет собой циклический пептид, содержащий от 3 до 10 аминокислотных остатков, выстроенных в кольцо, три соседних остатка пептида имеют аминокислотную последовательность Ile-Xaa-Ser; где Xaa представляет собой аминокислотный остаток, выбранный из группы, включающей Asp, Asn, Glu и Gln. Предпочтительно, Xaa представляет собой Asp или Asn.
Предпочтительный циклический пептид с пятью аминокислотами, выстроенными в кольцо, имеет формулу:

где X1 и X2 независимо представляют аминокислотный остаток или множество аминокислотных остатков до 10, связанных пептидными связями; Y1 и Y2 независимо представляют аминокислотные остатки, связанные друг с другом дисульфидной связью, и Xaa представляет собой аминокислотный остаток, выбранный из группы, включающей Asp, Asn, Glu и Gln. Предпочтительно, Y1 и Y2 представляют собой остатки цистеина, соединенные дисульфидной связью (то есть остаток цистина).
Особенно предпочтительный циклический пептид имеет аминокислотную последовательность: цикло Cys-Ile-Xaa-Ser-Cys (SEQ ID NO: 7); где Xaa представляет собой аминокислотный остаток, выбранный из группы, включающей Asp, Asn, Glu и Gln, и данное кольцо сформировано с помощью дисульфидной связи между двумя остатками цистеина. Предпочтительно, Xaa представляет собой Asp или Asn.
Способ ингибирования опосредованной R-кадгерином клеточной адгезии предусматривает введение клеток, экспрессирующих R-кадгерин, в контакт с ингибирующим адгезию количеством селективного пептида-антагониста R-кадгерина настоящего изобретения. Клетки можно ввести в контактс пептидомантагонистом in vivo путем введения ингибирующего клеточную адгезию количества антагониста млекопитающему, страдающему заболеванием или состоянием, лечение которого можно осуществлять путем ингибирования опосредованной R-кадгерином клеточной адгезии (например, заболевания сетчатки, характеризующиеся патологической пролиферацией сосудов). Например, человека, страдающего возрастной дегенерацией желтого пятна или диабетической ретинопатией, можно лечить с помощью селективного пептида-антагониста R-кадгерина по настоящему изобретению. Предпочтительно, антагонист вводят в виде фармацевтической композиции, содержащей антагонист и фармацевтически приемлемый носитель.
Для селективного нацеливания или антагонизма R-кадгерина пептиды и композиции по настоящему изобретению можно вводить в терапевтически эффективном количестве парентерально, перорально, с помощью ингаляции или местно в стандартной лекарственной форме вместе с фармацевтически приемлемыми носителями, средствами доставки и адъювантами. Термин "парентеральный", как используется здесь, включает внутривенное, подкожное, внутримышечное, надчревное, внутриглазничное (например, в стекловидное тело) и интраперитонеальное введение, а также введение путем инфузии.
Можно использовать любой подходящий путь введения, и фармацевтическую композицию, включая селективный пептид-антагонист R-кадгерина по настоящему изобретению, вводят в дозе, эффективной для предполагаемого лечения. Специалист в данной области легко определит терапевтически эффективные количества, необходимые для лечения конкретного медицинского состояния или ингибирования его развития, применяя предклинические и клинические исследования, известные в области медицины.
Термин "терапевтически эффективное количество", как используется здесь, относится к количеству активного ингредиента, которое оказывает биологический или лечебный эффект на медицинскую ткань, систему, животное или человека, которого добивается врач или исследователь.
Термин "ингибировать", как используется здесь, относится к замедлению, задержке или прекращению медицинского состояния или биохимического взаимодействия, но не обязательно указывает на полное прекращение состояния или полное устранение взаимодействия. Длительное сохранение эффекта у пациента или длительное снижение тяжести симптомов указывает на то, что медицинское состояние положительно контролируется (то есть ингибировано).
Режимы дозировки для пептидов-антагонистов R-кадгерина по настоящему изобретению и содержащих его композиций основаны на нескольких факторах, таких как возраст, вес, пол и тип заболевания у пациента, тяжесть состояния, путь введения и антагонистическая активность конкретного используемого пептида-антагониста. Режим дозировки можно изменять в зависимости от вышеупомянутых факторов. Например, для ингибирования ангиогенеза сетчатки применяют уровни дозировки с порядком от приблизительно 0,01 мг до приблизительно 1000 мг на килограмм веса тела. Предпочтительные уровни дозировки находятся в диапазоне от приблизительно 0,01 мг до приблизительно 100 мг на килограмм веса тела.
Для введения путем инъекции композицию, содержащую пептид по настоящему изобретению, составляют вместе с фармацевтически приемлемым носителем, таким как вода, солевой раствор или водный раствор декстрозы. Для инъекции типичная суточная доза составляет от приблизительно 0,01 мг до приблизительно 100 мг на килограмм веса тела, которую вводят ежедневно в качестве однократной дозы или в качестве многократной дозы в зависимости от указанных выше факторов.
Фармацевтические композиции по настоящему изобретению, содержащие селективный пептид-антагонист R-кадгерина по изобретению и фармацевтически приемлемый носитель, могут также содержать фармацевтически приемлемые инертные наполнители. Фармацевтически приемлемые инертные наполнители, которые могут входить в состав фармацевтических композиций по настоящему изобретению, включают, например, физиологически переносимые поверхностно-активные вещества, растворители, забуференные агенты, консерванты и тому подобное, хорошо известные из уровня техники.
Например, для ингибирования ангиогенеза сетчатки пациенту, страдающему от патологической пролиферации сосудов сетчатки, вводят терапевтически эффективное количество пептида антагониста R-кадгерина по настоящему изобретению. Введенный пептид селективно связывается с R-кадгерином в сетчатке, таким образом, прерывая и ингибируя ангиогенез в ней. Предпочтительно, пептидный антагонист вводят путем инъекции в стекловидное тело.
Нацеливание Lin-HSC на развивающуюся сосудистую сеть ингибируют путем введения стволовых клеток в контакт с ингибирующим нацеливание на сосудистую сеть количеством селективного пептида-антагониста R-кадгерина по настоящему изобретению. Ингибированное нацеливание Lin-HSC, таких как эндотелиальные клетки-предшественники, на развивающуюся сосудистую сеть можно использовать при лечении заболеваний, связанных с патологическим развитием сосудов, таким как возрастная дегенерация желтого пятна и диабетическая ретинопатия. Предпочтительно, Lin-HSC приводят в контакт in vivo путем введения пептидов-антагонистов R-кадгерина по настоящему изобретению млекопитающему, такому как человек, страдающему сосудистым пролиферативным заболеванием или состоянием.
Следующие неограничивающие примеры представлены для дополнительной иллюстрации различных аспектов изобретения. Для специалиста в данной области техники очевидно, что модификации примеров и проиллюстрированных вариантов осуществления, раскрытых здесь, можно осуществить без отклонения от сущности и объема изобретения.
Пример 1. Синтез пептида
Пептиды по настоящему изобретению и различные контрольные пептиды синтезировали в центральном учреждении белков и нуклеиновых кислот The Scripps Research Institute с применением твердофазного синтеза и очищали до максимально возможной степени чистоты (чистота более 95%) в соответствии с данными анализа ВЭЖХ. Последовательности пептидов анализировали с помощью масс-спектрометрии для подтверждения синтеза правильных пептидов. Все пептиды блокировали на амино-конце амидной группой и ацетилировали на карбокси-конце. Циклические пептиды готовили с цистеиновыми остатками на амино- и карбокси-концах, для создания дисульфидной связи и формирования кольца, содержащего пять аминокислотных остатков. На фиг.5 (B) показаны полученные пептиды: циклический CIDSC (SEQ ID NO: 9), циклический CINPC (SEQ ID NO: 9), IDSMSGR (SEQ ID NO: 2), IDSASGR (SEQ ID NO: 10), INPASGQ (SEQ ID NO: 11), циклический CSDIC (SEQ ID NO: 12) и циклический CRADC (SEQ ID NO: 13).
Пример 2. Трансфекции L-клетки
R-кадгерин мыши и плазмиды N-кадгерина были любезно предоставлены доктором Masatoshi Takeichi (Kyoto University, Japan). Плазмиды субклонировали в векторы pDsRed2 N1 (Clontech) для кодирования составных белков с красным флуоресцентным белком (RFP), присоединенным к C-концу молекул кадгерина. L-клетки (клетки фибробластов L929 мыши, ATCC #CRL-2148) устойчиво трансфицировали либо R, либо N-кадгерином pDsRed2 N1 с применением системы трансфекции фосфата кальция (Life Technologies) в соответствии с протоколом изготовителя. После селекции путем выращивания в питательных средах с добавкой генетицина (700 нг/мл генетицина G418, Gibco BRL) отобрали положительные клоны. Клетки исследовали на экспрессию RFP и проверили на экспрессию кадгерина с помощью иммунофлюоресцентного окрашивания и иммуноблоттинга. На фиг.6 проиллюстрирована агрегация L-клеток, экспрессирующих R и N-кадгерины. На фиг.6 (A) показаны микрофотографии L-клеток, экспрессирующих R-кадгерин (слева) и N-кадгерин (посередине), агрегирующих в средах, содержащих кальций, по сравнению с нетрансфицированными L-клетками, которые не агрегировали. Фиг.6 (B) представляет собой столбчатую диаграмму, иллюстрирующую процент агрегации клеток, показанных на фиг.6 (A). Клетки, трансфицированные N-кадгерином и R-кадгерином, трипсинизировали в буфере, содержащем приблизительно 5 мМ хлорида кальция (обозначенные как TC), образовывали крупные кластеры клеток, тогда как эндогенные L-клетки демонстрировали незначительную агрегацию в буфере, содержащем кальций. Клетки, трипсинизированные с EDTA в буфере без кальция (обозначенные как ТЕ), демонстрировали незначительную агрегацию, независимо от того, трансфицировали клетки кадгеринами или нет.
Пример 3. Клеточная культура и иммунофлюоресценция
Трансфицированные L-клетки или L-клетки дикого типа выращивали в модифицированной среде Eagles (MEM) с добавкой основного раствора соли Earl's, 2 мМ Glutamax, 1 мМ пирувата натрия, 0,1 мМ заменимых аминокислот и 10% эмбриональной бычьей сыворотки. Трансфицированные линии клеток выращивали в средах с добавкой приблизительно 700 мкг/мл G418 (все реактивы питательных сред были от Gibco BRL). Для иммунофлюоресценции клетки выращивали до приблизительно 75% слияния на покровных стеклянных полосках. Клетки фиксировали в 4% параформальдегиде в течение приблизительно получаса с последующей блокировкой 5% нормальной козьей сывороткой и 5% эмбриональной бычей сывороткой в 1x фосфатном буферном солевом растворе (PBS). Поликлональные антитела козы против R-кадгерина или N-кадгерина (Santa Cruz) использовали с разбавлением 1:200, и флюоресценцию обеспечили за счет инкубации с маркированными вторичными IgG анти-коза Alexa488 (Molecular Probes). Изображения сделали с применением флуоресцентного конфокального микроскопа Radiance 200 (BioRad). Для иммуноблот-анализа клетки растворили в буфере, содержащем 1% Triton X-100. Приблизительно 50 мкг полного лизата клеток добавили на каждую дорожку 8% полиакриламидного геля и белки разделили с помощью электрофореза. Для визуализации соответствующих полос использовали моноклональные антитела (1:1000, BD Biosciences), специфичные для N-кадгерина или R-кадгерина.
На фиг.7 (A) показан иммуноблоттинг нативных L-клеток и L-клеток, трансфицированных R-кадгерином и N-кадгерином и окрашенных антителом R-кадгерина. Только клетки, трансфицированные R-кадгерином, показали значительные уровни экспрессии R-кадгерина. На фиг.7 (В) показан иммуноблоттинг нативных L-клеток и L-клеток, трансфицированных R-кадгерином и N-кадгерином и окрашенных антителом N-кадгерина. Только клетки, трансфицированные N-кадгерином, показали значительные уровни экспрессии N-кадгерина. Фиг.7 (C) представляет собой флуоресцентную микрофотографию клеток, экспрессирующих R-кадгерин, маркированных флуоресцентными антителами кадгерина, демонстрирующую экспрессию на поверхности клетки только молекул R-кадгерина. Фиг.7 (D) представляет собой флуоресцентную микрофотографию клеток, экспрессирующих N-кадгерин, маркированных флуоресцентными антителами кадгерина, демонстрирующую экспрессию на поверхности клетки только молекул N-кадгерина. Фиг.7 (E) представляет собой флуоресцентную микрофотографию нативных L-клеток, подвергнутых воздействию флуоресцентных антител кадгерина, но не показывающую экспрессии на поверхности клетки молекул кадгерина любого типа.
Пример 4. Проба на агрегацию
L-клетки выращивали до высокого уровня слияния с последующей трипсинизацией с помощью 0,01% трипсина + 5 мМCaCl2 и без EDTA (TC) или с помощью 0,01% трипсина с 0,1 мМ EDTA и без кальция (ТЕ). Клетки собрали и промыли с последующим ресуспендированием в буферном растворе Hanks (HBSS) + 1% BSA с (TC) или без (ТЕ) 5 мМ CaCl2. Клетки культивировали при 37°C в 0,5 мл раствора при 2 × 105 клеток на каждую лунку 24-луночной плашки со встряхиванием при приблизительно 60-70 об/мин с переменными концентрациями пептида. Все пробы выполняли трижды. Степень агрегации клеток представляли отношением полного количества частиц после 2 часов инкубации (N2hr) к начальному количеству частиц (N0). Частицы подсчитывали на гемоцитометре с применением суммы 8 отдельных 20 мкл счетов на лунку до (N0), и после (Nt) инкубации. Результаты проиллюстрированы на фиг.8.
Пример 5. Обработка мышей посредством инъекции пептидов в стекловидное тело
Пептиды растворили в PBS + 10% диметилсульфоксид до концентрации 10 мМ. Приблизительно 0,5 мкл или 1,0 мкл 10 мМ раствора пептида вводили в полость стекловидного тела 2-дневных или 7-дневных мышей соответственно. В P5 или Р11 сетчатки препарировали в соответствии с описанием, и сосуды, и астроциты визуализировали с помощью иммуногистохимии. Количественный анализ периферической васкуляризации, длину сосудов и площадь сосудов в ходе поверхностного формирования сосудов получили с помощью отображения сетчаток с инъекцией при тех же настройках микроскопии. Затем получили количества с применением программного обеспечения LASERPIX® (BioRad) с помощью контрольных мышей того же помета, используемых для начальной нормализации степени васкуляризации сетчатки. Количественный анализ действия на формирование глубоких сосудов получили путем фокусировки на передней камере глаза до сплетения сосудов нормальной глубины с применением конфокальной микроскопии и подсчета количества сосудов, мигрировавших в слой зрительных рецепторов. Результаты представлены на фиг.9.
Пример 6. Выделение стволовой клетки и обогащение
Клетки костного мозга выделили из взрослых трансгенных мышей, в которых размноженный GFP был объединен с промотором β-актина (ACTbEGFP, the Jackson Laboratory, Bar Harbor, Maine). Затем собрали моноциты путем разделения с помощью градиента плотности с применением Histopaque (Sigma) и пометили панельными антителами сопряженной с биотином линии (CD45, CD3, Ly-6G, CD11, TER-119, Pharmingen, San Diego, CA) для отбора Lin-. Клетки Lin+ и Lin- разделяли с применением колонки магнитного разделения (MACS, Miltenyi Biotech, Auborn, California). Так как было определено, что клетки CD31- представляют лучший контроль нефункционирующей субпопуляции HSC, что определили с помощью нацеливания на сосуды, клетки CD31- отделили от моноцитов с помощью разделения MACS с применением антител CD31 и использовали в качестве отрицательного контроля для функционирующей Lin-HSC. HSC от мышей дикого типа проанализировали на экспрессию R-кадгерина с помощью маркирования клеток антителами анти-R-кадгерина (sc-6456, Santa Cruz Biotech) и меченными Alexa-488 вторичными антителами анти-коза осла (молекулярные зонды) и с применением проточного цитометра FACS calibur (Beckton Dickinson, Franklin Lakes, NJ). Результаты представлены на фиг.10.
Пример 7. Культивирование клетки HSC, инъекции и количественный анализ
До инъекции Lin-HSC культивировали с 100 нМ антитела, блокирующего R-кадгерин (SC-6456, Santa Cruz Biotech) или преиммунным IgG козы в солевом растворе с фосфатным буфером в течение приблизительно 1 часа при приблизительно 37°C. Интравитреальные инъекции в глаза P6 выполняли с применением 0,5 мкл раствора HSC. Затем сетчатки исследовали в P12 в виде тотального препарата или срезов. Нацеливание Lin-HSC определили количественно путем подсчета общего количества стволовых клеток в сетчатке с применением восьми различных областей обзора сетчатки: левый, правый, верхний и нижний квадранты (3/4 расстояния до периферии сетчатки), два промежуточных квадранта (1/4-1/2 расстояния до периферии), место инъекции и область диска зрительного нерва. Эти клетки характеризовали локализацией в поверхностных, промежуточных или глубоких слоях или отсутствием нацеливания (клетки, которые лежат в задней части слоя зрительных рецепторов). Количество ненацеливающихся клеток в слое зрительных рецепторов представляют как процент от общего количества наблюдаемых стволовых клеток. Результаты представлены на фиг.11 и фиг.12.
ОБСУЖДЕНИЕ
Селективные пептиды-антагонисты R-кадгерина по настоящему изобретению действуют как пептиды, имитирующие ключевые мотивы узнавания R-кадгерина (то есть последовательности IDS и INS, найденные в R-кадгеринах млекопитающих). Без привязки к теории, пептиды-антагонисты по настоящему изобретению вероятно блокируют функцию адгезии молекул R-кадгерина за счет конкурентного взаимодействия с доменами EC1 молекул R-кадгерина на поверхностях клеток. Пептиды-антагонисты по настоящему изобретению могут использоваться для изучения молекулярных функций и при лечении заболеваний, связанных с клеточной адгезией, и, в целом, они легче диффундируют в ткани послеинъекции in vivo, чем антагонисты на основе антител.
Морфогенез ткани в ходе развития большинства тканей, включая нервную ткань сетчатки, подразумевает селективное связывание молекул адгезии между клетками. Такая селективность связывания позволяет группироваться сходно дифференцированным клеткам и не позволяет типам клетки вторгаться в несоответствующие структуры ткани. Однако, несмотря на обширные изучения свойств и функции кадгеринов, в особенности N- и E-кадгеринов, общий механизм, отвечающий за специфичность кадгерина, еще не открыт. Пептиды-антагонисты R-кадгерина по настоящему изобретению селективно взаимодействуют с молекулами R-кадгерина млекопитающих без значительного связывания с другими классами кадгерина. Эти пептиды содержат последовательность IDS (или ее гомологи INS, IES и IQS), которая соответствует области в домене EC1 кадгерина, остатки 53-55 из SEQ ID NO: 17 и 18, в которых, как сообщается, расположены важные взаимодействия на поверхности адгезии на основе структурного, мутационного анализа и анализа гомологии последовательностей.
Без привязки к теории, считается, что мотив IDS устанавливает прямые контакты с последовательностью VDI прилегающей молекулы кадгерина на поверхности адгезии. Поскольку остатки 53-55 кадгеринов, по всей видимости, необходимы для трансдимеризации, и поскольку в отличие от других областей, важных для адгезии, эта короткая аминокислотная последовательность не является консервативной среди классических членов семейства кадгеринов, эта область действует как определяющий фактор для специфичности кадгерина. В действительности, циклическая IDS (CIDSC, SEQ ID NO: 8) селективно ингибирует клеточную агрегацию, опосредованную R-кадгерином, в то время как соответствующий аналог N-кадгерина, циклический INP (CINPC, SEQ ID NO: 9), селективно ингибирует клеточную агрегацию, опосредованную N-кадгерином. N-кадгерин и R-кадгерин являются наиболее гомологичными из членов семейства кадгеринов. Фактически, хотя все кадгерины, включая R- и N-кадгерин, преимущественно взаимодействуют гомофильным образом, R- и N-кадгерин являются единственными двумя классическими членами семейства кадгеринов, среди которых наблюдали функциональные гетеродимеры. Таким образом, очень маловероятно, что циклический IDS и циклический INP обладают различными функциональными свойствами в отношении N и R-кадгерина, но совпадающими свойствами в отношении любого другого члена семейства кадгеринов. Данные исследования демонстрируют, что мотив IXS (где X представляет собой D, N, E, или Q, то есть соответствует остаткам 53-55 R-кадгерина и их гомологам) играет важную роль в обеспечении гомологичного связывания молекул R-кадгерина. По всей видимости, на основании специфичности IXS к R-кадгерину и INP к N-кадгерину, соответственно размещенные остатки в других членах семейства кадгеринов (например, мотив IER E-кадгерина и мотив IEK P-кадгерина) также обеспечивают специфичность к другим классическим кадгеринам.
Предыдущие исследования показали, что антитела против R-кадгерина прерывают васкуляризациюсетчатки in vivo. По всей видимости, васкуляризация поверхностного сплетения прервалась по причине нарушения опосредованных R-кадгерином направляющих сигналов, передаваемых астроцитами, расположенными перед эндотелиальными клетками. Экспрессию R-кадгерина также наблюдали в областях, в которых впоследствии обнаруживают глубокие сплетения сосудов, непосредственно перед инвазией сосудов. Считается, что молекулы R-кадгерина в этих областях направляют эндотелиальные клетки на правильное сплетение сосудов, так как инъекция блокирующих R-кадгерин антител заставляет сосуд обходить нормальные васкуляризованные слои.
В настоящее время, схожие фенотипы сосудов произвели с помощью инъекции циклической IDS (CIDSC, SEQ ID NO: 8) как в ходе поверхностной, так и в ходе глубокой васкуляризации сетчатки. Так как циклическая IDS селективно в значительной степени прерывала агрегацию, опосредованную R-кадгерином, in vitro (то есть без значительного прерывания агрегации, опосредованной N-кадгерином), вероятно, что in vivo фенотип сосудов также производился за счет взаимодействий с высоким сродством циклического IDS с R-кадгерином. Кроме того, инъекция циклического INP (CINPC, SEQ ID NO: 9), который представлял собой эффективный ингибитор агрегации, опосредованной N-кадгерином, но не агрегации R-кадгерина in vitro, не приводила к значимому фенотипу сосудов сетчатки. В совокупности, эти результаты подтверждают специфическую роль R-кадгерина в контроле над сосудами.
Разработка селективных пептидов-антагонистов R-кадгерина по настоящему изобретению была основана на структурном, биохимическом и мутационном анализе различных членов семейства классических кадгеринов. Как известно, триптофан-2 является важным для функции кадгеринов наряду с последовательностью HAV при аминокислотных остатках 79-81 N-кадгерина и R-кадгерина. Фактически, как сообщается, линейные и циклические пептиды, содержащие последовательность HAV, блокируют опосредованное N-кадгериномразрастание неврита in vitro. Однако эти последовательности абсолютно консервативны среди всех молекул кадгерина и поэтому не могут придавать специфичность связывания. Другие остатки также должны устанавливать важные контакты на поверхности димеризации, при этом некоторые неконсервативные остатки являются важными для узнавания кадгеринов. Внимание сосредоточили на остатках в амино-концевом повторе кадгерина (EC1). Большинство важных для контакта остатков было локализовано в трех областях, аминокислоты 35-45, аминокислоты 53-59 и аминокислоты 79-86. Остатки 53-59, содержащиеся в большинстве данных кадгерин-специфичных остатках, являются потенциально важными в формировании поверхности димеризации. Из них остатки 53-55 имели особое значение. Таким образом, пептиды-аналоги разрабатывали из этой области для увеличения вероятности получения R-кадгерин-специфичных пептидов. Разработали схожие пептиды против последовательности от N-кадгерина мыши, наиболее близкого члена семейства кадгеринов, и другие контрольные пептиды и использовали их для сравнительного анализа.
Агрегация, опосредованная кадгерином
Выбрали клетки фибробластов мыши (линия L929), обычно называемые L-клетками, поскольку они, как известно, не содержат эндогенной экспрессии кадгерина. Создали трансфектанты, устойчивые к R-кадгерину, и использовали их для проверки действия разработанных пептидов на агрегацию, опосредованную R-кадгерином. Также создали трансфектанты, устойчивые к N-кадгерину, и использовали их для оценивания пептидов с точки зрения селективности к кадгерину, на основании их действия на агрегацию, опосредованную N-кадгерином. Иммуноблот-анализ выявил высокие уровни R-кадгерина и отсутствие экспрессии N-кадгерина в клоне 8 R-кадгерина (R-cad8), в то время как в клоне N-кадгерина 3 (N-cad3) обнаружили высокие уровни экспрессии N-кадгерина и отсутствие экспрессии R-кадгерина, как показано на фиг.7. Иммунофлюоресценция подтверждала экспрессию в данных выбранных клонах соответствующего кадгерина. При тестировании в анализе агрегации выявили изменение морфологии трансфицируемых линий клеток в результате трансфекции кадгерина. В то время как родительские (то есть нетрансфицированные) L-клетки оставались разделенными в виде отдельных клеточных частиц, мыши, трансфицированные R-кадгерином и N-кадгерином, образовывали большие, кальций-зависимые (буфер TC) кластеры клеток из-за плотной межклеточной связи, как показано на фиг.6 (A). Кластеры опосредованной R-кадгерином агрегации устраняли с помощью начальной трипсинизации клеток с раствором EDTA и агрегацией в буфере без кальция (буфер ТЕ), как показано на фиг.6 (B).
Действие пептида на агрегацию клеток
Пептиды добавляли с переменными концентрациями в лунки агрегации для проверки их эффективности в блокировке опосредованной кадгерином агрегации. Циклический IDS ингибировал опосредованную R-кадгерином агрегацию с IC50 приблизительно 300 мкМ. Линейный пептид EDSMSGR (SEQ ID NO: 2) также блокировал опосредованную R-кадгерином агрегацию. Однако его эффективность (IC50 ˜ 900 мкМ) была приблизительно в 3 раза ниже, чем для циклического IDS. Поскольку циклические пептиды также оказались намного лучше растворимыми и более простыми в обращении, чем линейные пептиды, дальнейший акцент сделали исключительно на анализе циклических пептидов (фиг.8 (A)). Действие циклической IDS было специфичным к R-кадгерину, поскольку на агрегацию N-кадгерина наблюдали несильное действие. В отличие от этого, соответствующая последовательность, специфичная к N-кадгерину, циклический INP, ингибировала опосредованную N-кадгерином агрегацию с IC50 несколько ниже 300 мкМ (фиг.8 (B)) подобно действию циклического IDS на агрегацию R-кадгерина. Циклический INP оказывал несильное действие на опосредованную R-кадгерином агрегацию.
Другие контрольные пептиды, циклический RAD (CRADC, SEQ ID NO: 13) и циклический SDI (CSDIC, SEQ ID NO: 12) оказывали незначительное действие на агрегацию, опосредованную либо R-кадгерином, либо N-кадгерином. В качестве сравнения протестировали циклический пептид HAV (CHAVC, SEQ ID NO: 20), уже известный, как эффективный при блокировке адгезии, опосредованной любыми классическими молекулами кадгерина. В анализе циклический HAV блокировал агрегацию, опосредованную R-кадгерином и N-кадгерином с IC50 в диапазоне между 150 и 200 мкМ. Таким образом, циклический IDS и циклический INP селективно блокировали адгезию R или N кадгерина соответственно, только с несколько более низкими сродствами, чем у неспецифичного пептида, блокирующего все кадгерины. Было показано, что в предыдущих исследованиях с применением антител против R-кадгерина, нормальная васкуляризация развития сетчатки прерывается. Эти антитела также были эффективны при прерывании агрегации, опосредованной кадгерином в системе анализа с IC50 приблизительно 10 нМ, как показано на фиг.8 (C).
Действие пептидов на васкуляризацию сетчатки
Пептиды вводили в полость стекловидного тела глаз новорожденных мышей. Когда пептиды циклический IDS или циклический HAV вводили в глаза двухдневных мышей, и формирующуюся в результате этого сосудистую сеть исследовали три дня спустя, на 5 день после рождения (P5), формирование сосудов прерывалось, при этом результаты были схожи с наблюдаемыми при инъекциях антител. Эти сетчатки характеризовали как содержащие менее обширную периферическую васкуляризацию и меньшее количество взаимосвязанных сосудов в областях васкуляризации по сравнению с нормальными контрольными мышами того же помета без инъекций. В целом, васкуляризация поверхностного слоя была снижена на половину за счет пептидов, блокирующих R-кадгерин, в то время как сетчатки с инъекциями N-кадгерин-специфичного циклического INP были относительно нормальными (см. фиг.9(A-C)).
Селективные пептиды-антагонисты R-кадгерина настоящего изобретения также прерывали нормальную васкуляризацию глубоких слоев сетчатки. При введении циклического пептида IDS в P7 непосредственно перед проникновением в глубину сосудов поверхностной сосудистой сети и началом формирования глубокого сплетения сосудов формирующаяся сосудистая сеть Р11 характеризовалась многочисленными отростками сосудов, которые мигрировали за пределы нормальной глубины сплетения сосудов в аваскулярный слой зрительных рецепторов. Кроме того, это сходно с действием, наблюдаемым прежде при введении антител R-кадгерина. Напротив, глубокое сплетение сосудов глаз с инъекциями циклического пептида INP формировалось нормально, как показано на фиг.9 (B и D).
R-кадгерин экспрессируется Lin-HSC
Экспрессию R-кадгерина гематопоэтической стволовой клеткой (HSC) проанализировали для определения, экспрессировались ли молекулы клеточной адгезии R-кадгерина на клеточной поверхности клеток, функционально являющихся нацеливающимися. С применением анализа проточной цитометрии, R-кадгерин экспрессировался на клеточной поверхности почти 80% субпопуляции Lin- HSC, в то время как только 30% клеток Lin+ экспрессируют R-кадгерин (фиг.10). На основании относительных интенсивностей флуоресценции между двумя популяциями клеток, вероятно, что клетки Lin- также экспрессируют более высокие концентрации R-кадгерина на клеточной поверхности, чем малая часть R-кадгерин-положительных клеток Lin+. Таким образом, большинство клеток в субпопуляции, которые функционально нацеливаются на сосудистую сеть сетчатки, экспрессируют R-кадгерин, в отличие от большинства клеток ненацеливающейся субпопуляции. Интересно отметить, что HSC других субпопуляций, которые представлены CD31, CD34 и Mac1-отрицательной и вообще не обладают функцией нацеливания, содержали даже меньше клеток, экспрессирующих R-кадгерин.
Антитела и пептиды, блокирующие R-кадгерин, прерывающие нацеливание HSC
Для исследования степени, до которой межклеточная адгезия R-кадгерина функционирует в нацеливании HSC на различные слои сосудов сетчатки, Lin-HSC заблокировали с помощью R-кадгерин-специфичных, блокирующих антител перед инъекцией. Спустя шесть дней после инъекции нормальные Lin-HSC обнаружили локализованными только в трех слоях сосудов: 1) поверхностное сплетение сосудов, локализованное в слое ганглионарных клеток, 2) глубокое сплетение сосудов, локализованное вблизи внешнего плексиформного слоя и 3) промежуточный слой, локализованный в переднем крае внутреннего ядерного слоя. На фиг.11 (A) показаны поперечные сечения сетчаток после инъекции нормальных Lin-HSC (слева), Lin-HSC, культивированных с антителами R-кадгерина, блокирующими адгезию (посередине), и Lin-HSC, культивированных с преиммунным IgG козы (справа).
При предварительном культивировании Lin-HSC с антителами анти-R-кадгерина перед инъекцией многие из этих клеток теряли способность к правильному нацеливанию, в то время как клетки, предварительно культивированные с преиммунным IgG, функционируют так же, как и незаблокированные HSC. По всей видимости, на нацеливание на глубокие и промежуточные слои сосудов влияет, в особенности, блокировка адгезии R-кадгерина, поскольку в этих областях обнаружили локализованными относительно немного HSC с заблокированным R-кадгерином. Клетки, локализованные в поверхностном сплетении сосудов, также казались менее организованными и не были локализованными совместно с эндогенной сетью сосудов в такой же степени, как нормальные Lin-HSC или предварительно культивированные с преиммунным IgG.
Многие из Lin-HSC, предварительно культивированных с антителом R-кадгерина, мигрировали через сетчатку, через все три слоя сосудов и присоединились к внешнему краю зрительных рецепторов вблизи слоя RPE. Почти половину HSC с заблокированным R-кадгерином обнаружили на внешнем крае слоя зрительных рецепторов (фиг.11(B)). Для сравнения, в контрольных сетчатках с инъекцией HSC, предварительно культивированных с преиммунным IgG, только 15% HSC не нацеливались на эту область. Большую часть ненацеливавшихся клеток из контрольных сетчаток обнаружили вблизи места инъекции, и их вероятно можно отнести к клеткам, которые были выпущены под сетчатку при удалении иглы. При исключении места инъекции из вычисления количество ненацеливавшихся HSC, культивированных с преиммунным IgG, уменьшилось до 10%. Поскольку обычно Lin-HSC почти не наблюдали в данном "особенно глубоком" слое, этот незначительный процент ненацеливавшихся HSC, культивированных с преиммунным IgG, вероятно, можно отнести к тому факту, что преиммунный IgG мог связаться приблизительно с 10% клеток Lin- (фиг.10). Эти связанные молекулы IgG могут неспецифично предотвратить нормальную адгезию просто за счет стерических затруднений. Однако разница между количеством клеток, ненацеливавшихся по причине специфического блокирования R-кадгерина и неспецифического блокирования IgG, является значительной.
На фиг.12 (A) показаны конфокальные изображения через z-плоскости трех сплетений сосудов и внешнего края слоя зрительных рецепторов. Нормальное нацеливание в соответствующих сплетениях сосудов и вдоль эндогенных сосудов (красный) наблюдали у Lin-HSC (зеленый), блокированных преиммунным IgG козы. Блокирование адгезии R-кадгерина привело к локализации множества Lin-HSC на внешнем краю слоя зрительных рецепторов, и клетки, нацеливавшиеся на нормальные сплетения сосудов, стремились к скоплению и не были локализованы вдоль эндогенных сосудов (красный). Фиг.12 (B) представляет собой столбчатую диаграмму, демонстрирующую, что процент ненацеливавшихся клеток по отношению ко всей популяции HSC в сетчатке был существенно больше для популяции Lin-HSC с заблокированным R-кадгерином (значения P<0,01).
Предшествующее описание следует рассматривать в качестве иллюстрации, но не в качестве ограничения. Тем не менее, другие варианты в пределах сущности и объема настоящего изобретения могут быть легко представлены специалистами в данной области техники.







Предложен выделенный циклический пептид, имеющий аминокислотную последовательность Cys-Ile-Xaa-Ser-Cys (SEQ ID NO:7); где Хаа представляет собой аминокислотный остаток, выбранный из группы, состоящей из Asp, Asn, Glu и Gin, и содержащий дисульфидную связь между двумя остатками Cys, который можно использовать в качестве селективного антагониста R-кадгерина млекопитающих. Селективные пептиды-антагонисты R-кадгерина по изобретению могут использоваться для ингибирования нацеливания гематопоэтической стволовой клетки (HSC) на развивающуюся сосудистую сеть, для ингибирования опосредованной R-кадгерином клеточной адгезии и ингибирования ангиогенеза сетчатки. 5 н. и 3 з.п. ф-лы, 12 ил.
где Хаа представляет собой аминокислотный остаток, выбранный из группы, состоящей из Asp, Asn, Glu и Gln, и содержащий дисульфидную связь между двумя остатками Cys.
| ПОЛИНУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЮЩАЯ ПРОТОКАДГЕРИН PC3, PC4, PC5 (ВАРИАНТЫ), ПОЛИПЕПТИД ПРОТОКАДГЕРИНА PC3, PC4, PC5 (ВАРИАНТЫ) | 1995 |
|
RU2197525C2 |
| US 6472368 В1, 29.10.2002 | |||
| WO 00/02917 А2, 20.01.2000 | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Peri F | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

