Перекрестная ссылка на родственные заявки
По данной заявке испрашивается приоритет предварительной заявки на патент США №60/656037, поданной 24 февраля 2005, включенной в данное описание в виде ссылки.
Заявление о государственном интересе
Часть работы, описанной в данном описании, поддержана грантами за номерами EY11254 и EY12598 от National Eye Institute of the National Institutes of Health. Правительство Соединенных Штатов имеет некоторые права на данное изобретение.
Область техники, к которой относится изобретение
Данное изобретение относится к изолированным клеткам костного мозга млекопитающих. Конкретнее, изобретение относится к изолированным популяциям клеток костного мозга, которые имеют характеристики миелоидных клеток и способны к внедрению в сосудистую сеть сетчатки при интравитреальной инъекции в глаз. Изобретение также относится к способам лечения глазных дегенеративных болезней путем введения изолированных клеток костного мозга в глаз млекопитающего.
Предпосылки создания изобретения
Связанная с возрастом дегенерация желтого пятна (ARMD) и диабетическая ретинопатия (DR) являются ведущими причинами потери зрения в индустриальных странах и являются результатом аномального образования новых сосудов в сетчатке. Так как сетчатка состоит из хорошо установленных слоев нейронных, глиальных и васкулярных элементов, относительно небольшие нарушения, такие как нарушения, заметные при разрастании сосудов или отеке, могут привести к существенной потере зрительной функции. Наследственные дегенерации, такие как пигментный ретинит (RP), также ассоциируются с сосудистыми аномалиями, такими как артериолярное сужение и васкулярная атрофия. Большинство наследственных дегенераций сетчатки специфически влияет на фоторецепторы палочки, но также имеется сопутствующая потеря колбочек, принципиального компонента пятна, участка сетчатки у людей, который ответственен за остроту центрального тонкого зрения. Специфичные факторы выживания колбочек недавно описаны (Mohand-Said et al., 1998, Proc. Natl. Acad. Sci. USA, 95: 8357-8362) и могут облегчить выживание колбочек на мышиных моделях дегенерации сетчатки.
Наследственные дегенерации сетчатки поражают 1 из 3500 индивидуумов и характеризуются прогрессирующей куриной слепотой, скотомой, атрофией зрительного нерва, артериолярной аттенуацией, измененной проницаемостью сосудов и дефектом центрального зрения, часто прогрессирующим до полной слепоты (Heckenlively J.R., editor, 1988; Retinitis Pigmentosa, Philadelphia: JB Lippincott Co.). Молекулярный генетический анализ таких заболеваний идентифицирует мутации в более чем 110 различных генах для только относительно небольшого процента известных пораженных индивидуумов (Humphries et al., 1992, Science, 256: 804-808; Farrar et al., 2002, EMBO J., 21: 857-864). Многие такие мутации ассоциируются с ферментативными и структурными элементами механизма фототрансдукции, включая родопсин, cGMP-фосфодиэстеразу, rds-периферин и RPE65. Несмотря на такие сведения, пока не имеется эффективных способов лечения для замедления или реверсии развития таких дегенеративных болезней сетчатки. Последние достижения в области генной терапии привели к успешной реверсии фенотипов rds (Ali et al., 2000, Nat. Genet., 25: 306-310) и rd (Takahashi et al., 1999, J. Virol., 73: 7812-7816) у мышей и фенотипа RPE65 у собак (Acland et al., 2001, Nat. Genet., 28: 92-95), когда к фоторецепторам или пигментированному эпителию сетчатки (RPE) животных со специфической мутацией доставляется трансген дикого типа.
На протяжении многих лет известно, что популяция стволовых клеток существует в нормальных зрелых кровотоке и костном мозге. Различные субпопуляции таких клеток могут дифферицировать дальше в направлении гемопоэтической позитивной линии (Lin+) или негативной линии (Lin-). Более того, недавно показано, что популяция негатиной гемопоэтической клеточной линии стволовых клеток (HSC) содержит эндотелиальные клетки-предшественники (ЕРС), способные образовывать кровеносные сосуды in vitro и in vivo (см. Asahara et al., 1997, Science, 275: 964-7). Такие клетки могут принимать участие в нормальном и патологическом постнатальном ангиогенезе (см. Lyden et al., 2001, Nat. Med., 7, 1194-201; Kalka et al., 2000, Proc. Natl. Acad. Sci. U.S.A., 97: 3422-7; и Kocher et al., 2001, Nat. Med., 7: 430-6), а также дифференцировать в различные неэндотелиальные типы клеток, включая гепатоциты (см. Lagasse et al., 2000, Nat. Med., 6: 1229-34), микроглии (см. Priller et al., 2002, Nat. Med., 7: 1356-61), кардиомиоциты (см. Orlic et al., 2001, Proc. Natl. Acad. Sci. U.S.A., 98: 10344-9), и эпителий (см. Lyden et al., 2001, Nat. Med., 7, 1194-1201). Хотя такие клетки используют в некоторых экспериментальных моделях ангиогенеза, механизм нацеливания ЕРС на новую сосудистую сеть не известен и не определена стратегия, которая будет эффективно повышать число клеток, которые вносят вклад в определенную сосудистую сеть.
Гемопоэтические стволовые клетки из костного мозга в настоящее время являются единственным типом стволовых клеток, обычно используемых для лечебных применений. HSC костного мозга используются в трансплантатах свыше 40 лет. В настоящее время исследуются передовые методы сбора очищенных стволовых клеток с целью разработки способов лечения лейкоза, лимфомы и наследственных изменений крови. Исследуются клинические применения стволовых клеток для лечения диабета и запущенного рака почек у ограниченного числа больных людей.
Сущность изобретения
Настоящее изобретение относится к изолированной популяции миелоидоподобных клеток костного мозга (MLBM), продуцируемых клетками позитивного отбора, которые экспрессируют CD44, CD11b или оба антигена, из костного мозга млекопитающего. Такие клетки обнаруживают благоприятную васкулотрофическую и нейротрофическую активность при интраокулярном введении в глаз млекопитающего, страдающего от дегенеративной болезни глаз. Популяцию клеток MLBM по настоящему изобретению можно изолировать обработкой костного мозга антителами против CD44 (рецептор гиалуроновой кислоты), антителами против CD11b или антителами против обоих антигенов, и случаем могут быть клетки позитивного отбора, которые вступают в иммунную реакцию с антителами одного или обоих видов. Большинство клеток клеточной популяции MLBM по изобретению являются негативной линией и экспрессируют как антиген CD44, так и антиген CD11b.
Настоящее изобретение также относится к способу лечения васкулотрофических и нейротрофических болезней сетчатки у млекопитающего. Способ включает введение изолированных клеток из популяции клеток MLBM в больной глаз млекопитающего, предпочтительно, внутриглазной инъекцией. Предпочтительно, популяция клеток MLBM является аутологичной млекопитающему, которого лечат (т.е. популяцию клеток MLBM выделяют из костного мозга индивидуума млекопитающего, которого лечат). Способ лечения по настоящему изобретению уменьшает интенсивность дегенерации сосудов и дегенерации нейронов фоторецепторов в сетчатке млекопитающего, которое страдает от болезни глаз. Клетки вводят в количестве, достаточном для задержки сосудистой и нейронной дегенерации в сетчатке. Благоприятно клетки из популяции MLBM проникают в сосудистую сеть сетчатки и дифференцируют в эндотелиальные клетки, причем в то же время проникают в нейронную сеть и уменьшают интенсивность дегенерации клеток колбочек в сетчатке. Изолированная популяция клеток MLBM млекопитающего включает клетки, которые селективно нацеливаются на активированные ретинальные астроциты, когда инъецированы интравитреально в глаз, и устойчиво остаются в новой сосудистой сети и нейронной сети глаза. Предпочтительно, млекопитающим является человек.
В предпочтительном воплощении, по меньшей мере, примерно 75% клеток в изолированной популяции миелоподобных клеток костного мозга экспрессируют CD44, предпочтительнее, по меньшей мере, примерно 90%.
В одном предпочтительном воплощении клетки из популяции клеток MLBM трансфицируют терапевтически применимым геном. Например, клетки можно трансфицировать полинуклеотидами, которые операбельно кодируют нейротрофические средства или средства против ангиогенеза, которые селективно нацеливаются на новую сосудистую сеть и ингибируют образование новых сосудов, не поражая уже установившиеся сосуды, через форму генной терапии на основе клеток. В одном предпочтительном воплощении изолированная популяция клеток MLBM по изобретению включает ген, кодирующий пептид, ингибирующий ангиогенез. Ингибирующие ангиогенез клетки из популяции клеток MLBM применимы для модуляции аномального роста кровеносных сосудов при заболеваниях, таких как ARMD, DR и некоторые дегенерации сетчатки, связанные с аномальной сосудистой сетью. В другом предпочтительном воплощении изолированные клетки из популяции клеток MLBM по настоящему изобретению трансфицируют для включения гена, кодирующего нейротрофический пептид. Нейротрофические трансфицированные клетки MLBM применимы для промотирования спасения нейронов при болезни глаз, в том числе нейронной дегенерации сетчатки, такой как глаукома, пигментный ретинит и т.п.
Особым преимуществом лечения глаз изолированной популяцией клеток MLBM по настоящему изобретению является эффект васкулотрофического и нейроторофического спасения, наблюдаемый в глазах, обработанных интравитреально клетками из популяции клеток MLBM. Нейроны и фоторецепторы сетчатки, в частности колбочки, сохраняются, и можно поддерживать в некотором объеме зрительную функцию глаз, обработанных клетками из популяции клеток MLBM по изобретению.
Настоящее изобретение также относится к способу изоляции популяции миелоподобных клеток костного мозга негативным отбором клеток с маркером. Способ включает контактирование множества клеток костного мозга с антителами, специфичными для Ter119, CD45RB220 и CD3e, удаление клеток из множества клеток костного мозга, вступающих в иммунную реакцию с антителами к Ter119, CD45RB220 и CD3e, и извлечение миелоподобных клеток костного мозга, делетированных в клетках, экспрессирующих Ter119, CD45RB220 и CD3e. С использованием такого способа можно извлечь клеточную популяцию, в которой более 90% клеток экспрессируют CD44.
Предпочтительно, пораженная сетчатка, которую лечат популяцией клеток MLBM и способами по изобретению, содержит активированные астроциты. Этого можно достичь ранним лечением глаза, когда имеется ассоциированный глиоз, или использованием лазера для стимуляции локальной пролиферации активированных астроцитов.
Кроме лечебных применений, изолированная популяция миелоподобных клеток костного мозга по изобретению применима в качестве исследовательского инструмента для исследования физиологии развития сосудов в глазу и для доставки специфических генов к определенным местам размещения (например, в астроциты) в глазу. Такие применения дают ценный инструмент для исследования функции генов и возможных лечебных механизмов.
Краткое описание чертежей
На прилагаемых чертежах приводится описанное далее.
Фиг.1 отображает принципиальные схемы развития сетчатки у мыши. (А) Развитие первичного сплетения. (В) Вторая фаза образования сосудов сетчатки. GCL - слой ганглиозных клеток; IPL - внутренний слой сплетения; INL - внутренний ядерный слой; OPL - наружный слой сплетения; ONL - наружный ядерный слой; RPE - ретинальный пигментный эпителий; ON - зрительный нерв; Р - периферия. Часть (С) отображает характеризацию методом проточной цитометрии выделенных полученных из костного мозга клеток Lin+ HSC и Lin- HSC. Верхний ряд: точечный график распределения клеток, не меченных антителами, где R1 определяет определяемую количественно площадь дискриминационного окна РЕ-позитивного окрашивания; R2 показывает GFP-позитивное окрашивание; средний ряд - клетки Lin- HSC (С57В/6), и нижний ряд - клетки Lin+ HSC (С57В/6), каждую клеточную линию метят РЕ-конъюгированными антителами к Sca-1, c-kit, Flk-1/KDR, CD31. Данные по Tie-2 получают на мышах Tie-2-GFP. Проценты показывают процент позитивно меченых клеток из всей популяции Lin- HSC или Lin+ HSC.
Фиг.2 отображает внедрение HSC Lin- в развивающуюся сетчатку мыши. (а) На четвертые сутки после инъекции (Р6) интравитреально инъецированные клетки eGFP+ Lin- HSC присоединяются и дифференцируют на сетчатке. (b) Lin- HSC (мыши В6.129S7-Gtrosa26, окрашивание антителами β-gal) сами определяют будущее сосудистой сети, окрашенной антителами к коллагену IV (звездочка показывает верхнюю границу сосудистой сети). (с) Большинство клеток Lin+ HSC (eGFP+) на четвертый день после инъекции (Р6) неспособны к дифференцировке. (d) Мезентериальные мышиные ЕС eGFP+, четыре дня после инъекции (Р6). (е) Lin- HSC (eGFP+), инъецированные в зрелые глаза мышей. (f) Lin- HSC eGFP+ при малом увеличении, возвращающиеся и дифференцирующие вдоль предсуществующей астроцитарной матрицы в трансгенной мыши GFAP-GFP, (g) Ассоциация клеток Lin- (eGFP) и нижележащих астроцитов (стрелки) при большем увеличении. (h) Контроль - трансгенные неинъецированные GFAP-GFP. (i) На четвертые сутки после инъекции (Р6) Lin- HSC eGFP+ мигрируют и претерпевают дифференцировку на месте будущего глубокого сплетения. На чертеже слева - активность Lin- HSC во всей препарированной сетчатке; на чертеже справа показано местоположение клеток Lin- (стрелки) в сетчатке (верхняя часть со стороны стекловидного тела, нижняя часть - со стороны склеры). (j) Двойное мечение антителами к α-CD31-PE и α-GFP-alexa 488. Семь дней после инъекции, инъецированные Lin- HSC (eGFP, красный цвет) включаются в сосудистую сеть (CD31). Стрелки-указатели показывают области включения. (k) Клетки Lin- HSC eGFP+ образуют сосуды на четырнадцатый день после инъекции (Р17). (l и m) Интракардиальная инъекция родамина-декстрана показывает, что сосуды являются интактными и функционируют как в первичном (l), так и в глубоком сплетении (m).
Фиг.3 показывает, что клетки Lin- HSC eGFP+ возвращаются к глиозу (показано астроцитами, экспрессирующими GFAP, крайнее левое изображение), индуцированному повреждением, вызванным как лазером (а), так и механически (b), в зрелой сетчатке (звездочка показывает место повреждения). Изображения, крайние справа, получены при большем увеличении и показывают тесную связь Lin- HSC и астроцитов. Масштабная полоса = 20 мкМ.
Фиг.4 показывает, что клетки Lin- HSC спасают сосудистую сеть мыши с дегенерацией сетчатки. (a-d) Сетчатки на 27 сутки после инъекции (Р33) антигенами к коллагену IV; (a) и (b), сетчатки, инъецированные клетками Lin+ HSC (Balb/c), не показывают отличия в сосудистой сети от здоровой мыши FVB; (с) и (d), сетчатки, инъецированные клетками Lin- HSC (Balb/c), обнаруживают сильно развитую сосудистую сеть, аналогичную сети мыши дикого типа; (а) и (с), замороженные срезы целой сетчатки (верхняя часть со стороны стекловидного тела, нижняя часть со стороны склеры), окрашенные DAPI; (b) и (d) - глубокое сплетение во всем объеме сетчатки; (е) гистограмма, иллюстрирующая разрастание сосудистой сети глубокого сосудистого сплетения, образовавшегося в сетчатках, инъецированных клетками Lin- HSC (n=6). Степень глубокой ретинальной васкуляризации определяют количественно, вычисляя общую длину сосудов на каждом изображении. Сравнивают отношения (средняя длина сосудов)/область высокой степени (в микронах) для сетчаток с Lin- HSC, Lin+ HSC или контрольных. (f) Сравнение длины глубокого сосудистого сплетения после инъекции клеток Lin- HSC (R, правый глаз) или Lin+ HSC (L, левый глаз) у мышей rd/rd. Показаны результаты, полученные с шестью независимыми мышами (каждый цвет представляет отдельную мышь). (g) и (h) - клетки Lin- HSC также спасают сосудистую сеть rd/rd, когда инъецированы в глаза Р15. Показано промежуточное и глубокое сосудистое сплетение сетчаток, инъецированных клетками Lin- HSC (G) или Lin+ HSC (Н) (один месяц после инъекции).
Фиг.5 отображает микрофотографии ткани сетчатки мыши: (а) глубокий слой препарата всей сетчатки (rd/rd мыши) виден через 5 дней после инъекции (Р11) Lin- HSC eGFP+ (серый цвет); (b) и (с) - сосудистая сеть сетчатки Р60 мышей Tie-2-GFP (rd/rd), которые получили инъекцию клеток Lin- (b) или Lin+ HSC Balb/c, в Р6. В левых частях (b) и (с) видны только эндотелиальные клетки (окрашенные GFP). Средние части (b) и (с) показывают окрашивание антителами CD31; стрелки показывают сосуды, окрашенные CD31, но не GFP, правые части (b) и (с) показывают окрашивание как GFP, так и CD31. (d) Окрашивание α-SMA сетчатки, инъецированной Lin- HSC (левая часть), и контрольной сетчатки (правая часть).
Фиг.6 показывает, что трансфицированные Т2-TrpRS Lin- HSC ингибируют развитие сосудистой сети мыши. (а) Схематическое представление TrpRS, Т2-TrpRS и Т2-TrpRS с сигнальной последовательностью Igk человека по аминоконцам. (b) Сетчатки, инъецированные трансфицированными Т2-TrpRS Lin- HSC, экспрессируют белок Т2-TrpRS in vivo. (1) Рекомбинантный Т2-TrpRS, продуцированный в E. coli; (2) рекомбинантный Т2-TrpRS, продуцированный в E. coli; (3) рекомбинантный Т2-TrpRS, продуцированный в E. coli; (4) контрольная сетчатка; (5) сетчатка, инъецированная Lin- HSC + pSecTag2A (только вектор); (6) сетчатка, инъецированная Lin- HSC + pKLe135 (Igk-Т2-TrpRS в pSecTag). (а) Эндогенный TrpRS. (b) Рекомбинантный TrpRS. (с) Т2-TrpRS сетчатки, инъецированной Lin- HSC. (е-f) Характерные первичные (поверхностные) и вторичные (глубокие) сплетения инъецированных сетчаток, семь дней после инъекции; (с) и (d) - глаза, инъецированные Lin- HSC, трансфицированными пустой плазмидой, развиваются нормально; (е) и (f) - в большинстве глаз, инъецированных Lin- HSC, трансфицированными Т2-TrpRS, обнаруживают подавление глубокого сплетения; (с) и (е) - первичное (поверхностное) сплетение; (d) и (f) - вторичное (глубокое) сплетение. Отмеченные на (f) слабо выраженные сосуды являются "кровоточащими" изображениями первичных сосудов сети, показанных на (е).
Фиг.7 показывает последовательность ДНК, кодирующую Т2-TrpRS, меченный His6, SEQ ID NO: 1.
Фиг.8 показывает аминокислотную последовательность Т2-TrpRS, меченного His6, SEQ ID NO: 2.
Фиг.9 представляет микрофотографии и электроретинограммы (ERG) сетчаток мышей, получивших в глаза инъекцию Lin- HSC и Lin+ HSC (контроль).
Фиг.10 представляет статистические графики, показывающие корреляцию между спасением нейронов (ось Y) и спасением сосудов (ось X) как для промежуточных (Int.), так и для глубоких слоев сосудов глаз мыши rd/rd, обработанных Lin- HSC.
Фиг.11 представляет статистические графики, показывающие отсутствие корреляции между спасением нейронов (ось Y) и спасением сосудов (ось X) для глаз мыши rd/rd, обработанных Lin+ HSC.
Фиг.12 представляет собой диаграмму длины сосудов (ось Y) в произвольных относительных единицах в случае обработанных Lin- HSC (темные столбики) и необработанных (светлые столбики) глаз мыши rd/rd в моменты времени 1 месяц (1 М), 2 месяца (2 М) и 6 месяцев (6 М) после инъекции.
Фиг.13 включает три гистограммы, показывающие число ядер в наружном нейронном слое (ONR) мышей rd/rd через 1 месяц (1 М), 2 месяца (2 М) и 6 месяцев (6 М) после инъекции, и показывает существенное увеличение числа ядер в случае глаз, обработанных Lin- HSC (темные столбики), относительно контрольных глаз, обработанных Lin+ HSC (светлые столбики).
Фиг.14 представляет графики числа ядер в наружном нейронном слое отдельных мышей rd/rd, причем правый глаз (R, обработан Lin- HSC) сравнивают с левым глазом (L, контрольный глаз, обработан Lin+ HSC), в моменты времени (после инъекции) 1 месяц (1 М), 2 месяца (2 М) и 6 месяцев (6 М); каждая линия на данном графике соответствует сравнению глаз отдельной мыши.
Фиг.15 отображает изменения сосудистой сети и нейронов сетчатки у мышей rd1/rd1 (C3H/HeJ, левый ряд изображений) или мышей дикого типа (C57BL/6, правый ряд). Показана ретинальная сосудистая сеть промежуточных (верхний ряд) или глубоких (средний ряд) сосудистых сплетений в препаратах цельных сетчаток (красный цвет - коллаген IV, зеленый - CD31) и срезах (красный цвет - DAPI, зеленый - CD31, нижний ряд) одних и тех же сетчаток (Р - постнатальный день). (GCL - слой ганглиозных клеток, INL - внутренний ядерный слой, ONL - наружный ядерный слой).
Фиг.16 показывает, что инъекция Lin- HSC спасает от дегенерации нейроны у мышей rd1/rd1. (А, В и С) - Ретинальная сосудистая сеть промежуточного (Int.) или глубокого сплетения и срезы глаза, инъецированного Lin- HSC (правый ряд), и контралатерального контрольного глаза, инъецированного клетками (CD31-) (левый ряд), в Р30 (А), Р60 (В) и Р180 (С). (D) - средняя общая длина сосудистой сети (среднее + или - среднеквадратическая ошибка) в сетчатках, инъецированных Lin- HSC или контрольными клетками (CD31-), в Р30 (слева, n=10), P60 (посередине, n=10) и Р180 (справа, n=6). Данные для промежуточного (Int.) и глубокого сплетения показаны отдельно (ось Y - относительная длина сосудистой сети). (Е) - Средние числа ядер клеток в ONL в Р30 (слева, n=10), P60 (посередине, n=10) или Р180 (справа, n=6) сетчаток, инъецированных контрольными клетками (CD31-) или Lin- HSC (ось Y - относительное число ядер клеток в ONL). (F) - Линейная корреляция между длиной сосудистой сети (ось Х) и числом ядер клеток в ONL (ось Y) в Р30 (слева), P60 (посередине) и Р180 (справа) сетчаток, инъецированных Lin- HSC или контрольными клетками.
Фиг.17 показывает, что функция сетчатки спасается инъекцией Lin- HSC. Используют электроретинографическую (ERG) запись для измерения функции сетчаток, инъецированных Lin- HSC или контрольными клетками (CD31-). (А и В) - Характерные случаи спасенных и неспасенных сетчаток через 2 месяца после инъекции. Показаны срезы сетчатки правого глаза, инъецированного Lin- HSC (А), и левого глаза, инъецированного контрольными клетками CD31- (В), одного и того же животного (зеленый цвет - сосудистая сеть, окрашенная CD31, красный - ядра, окрашенные DAPI). (С) - Результаты ERG для того же животного, к которому относятся (А) и (В).
Фиг.18 показывает, что популяция клеток костного мозга человека может спасать дегенерирующие сетчатки у мыши rd1 (А-С). Спасение также наблюдают на другой модели дегенерации сетчатки rd10 (D-K). А - Lin- HSC человека (hLin- HSC), меченные зеленым красителем, могут дифференцировать в клетки сосудистой сети сетчатки после интравитреальной инъекции мышам C3SnSmn. CB17-Prkdc SCID. (В и С) - сосудистая сеть сетчатки (левые изображения; верх - промежуточное сплетение, низ - глубокое сплетение) и нейроны (правые изображения) в глазу, инъецированном hLin- HSC (В), или контралатеральном контрольном глазу (С), через 1,5 месяца после инъекции. (D-K) - Спасение мышей rd10 Lin- HSC (инъецированных в Р6). Показаны характерные сетчатки в Р21 (D - Lin- HSC, Н - контрольные клетки), Р30 (Е - Lin- HSC, I - контрольные клетки), Р60 (F - Lin- HSC, J - контрольные клетки) и Р105 (G - Lin- HSC, K - контрольные клетки) (обработанные и необработанные глаза одного и того же животного в каждый момент времени). Сосудистую сеть сетчатки (верхние изображения на каждой части представляют собой промежуточное сплетение; средние изображения на каждой части представляют собой глубокое сплетение) окрашивают CD31 (зеленый цвет) и коллагеном IV (красный). Нижние изображения на каждой части показывают поперечный срез той же сетчатки (красный цвет - DAPI, зеленый - CD31).
Фиг.19 показывает, что кристаллин αA позитивно регулируется в спасенных клетках наружного ядерного слоя после обработки Lin- HSC, но не в контралатеральных глазах, обработанных контрольными клетками. Левое изображение - контроль IgG в спасенной сетчатке, среднее изображение - кристаллин αA в спасенной сетчатке, правое изображение - кристаллин αA в неспасенной сетчатке.
Фиг.20 включает таблицы генов, которые позитивно регулируются в сетчатках мышей, обработанных Lin- HSC по настоящему изобретению. (А) - Гены, экспрессия которых возрастает в 3 раза в сетчатках мышей, обработанных мышиными Lin- HSC. (В) Гены кристаллина, которые позитивно регулируются в сетчатках мышей, обработанных мышиными Lin- HSC. (С) Гены, экспрессия которых возрастает в 2 раза в сетчатках мышей, обработанных Lin- HSC человека. (D) - Гены для нейротрофических факторов или факторов роста, экспрессия которых позитивно регулируются в сетчатках мышей, обработанных Lin- HSC человека.
Фиг.21 иллюстрирует распределение CD31 и поверхностных антигенов интегрина α6 в популяциях CD133-позитивных (CD133+) и CD133-негативных (CD133-) Lin- HSC человека. Левые изображения показывают графики рассеяния по результатам проточной цитометрии. Центральные и правые части представляют собой гистограммы, показывающие уровень экспрессии специфических антител в клеточной популяции. Ось Y представляет число событий, и ось Х показывает интенсивность сигнала. Заштрихованная гистограмма, смещенная вправо от контурной (контрольной) гистограммы, представляет возросший сигнал флуоресценции и экспрессию антител выше фонового уровня.
Фиг.22 иллюстрирует постнатальное развитие сетчатки у мышей дикого типа С57/В16, происходящее при нормальных уровнях кислорода (нормоксия), в постнатальные дни Р0-Р30.
Фиг.23 иллюстрирует модель вызванной кислородом ретинопатии у мышей С57/В16, возникшей при высоких уровнях кислорода (гипероксия; 75% кислорода) в дни Р7-Р12 с последующей нормоксией в Р12-Р17.
Фиг.24 показывает спасение сосудов путем обработки популяциями Lin- HSC на модели ретинопатии, вызванной кислородом (OIR).
Фиг.25 показывает, что спасенные фоторецепторы в наружном ядерном слое (ONL) мыши rd1 после интравитреальной инъекции Lin- HSC представляют собой, преимущественно, колбочки. Небольшой процент фоторецепторов сетчатки мышей дикого типа (верхние изображения) составляют колбочки, как показывает экспрессия красного/зеленого опсина колбочек (А), в то время как большинство клеток ONL позитивны в отношении красного специфического родопсина (В). Сосудистая сеть сетчатки показывает аутофлуоресценцию с неиммунной сывороткой (С), но ядерные слои полностью негативны для окрашивания специфическими опсинами палочек или колбочек. Сетчатки мышей rd/rd (нижние изображения) имеют уменьшенный внутренний ядерный слой и почти полностью атрофированный ONL, которые оба негативны в отношении опсина колбочек (D) или палочек (часть G). Контроль - глаза, обработанные CD31-HSC, идентичен сетчаткам rd/rd, не получавшим инъекцию, без окрашивания для опсина кобочек (Е) или палочек (Н). Контралатеральные глаза, обработанные Lin-HSC, показывают заметно уменьшенный, но явно присутствующий слой ONL, который состоит преимущественно из колбочек, что показывает позитивная иммунореактивность в отношении красного/зеленого опсина колбочек (F). Также отмечают небольшое число палочек (I).
Фиг.26 показывает графики рассеяния по определению проточной цитометрией свойств популяций стволовых клеток негативной линии и позитивной линии (верхние левые и нижние левые графики, соответственно), показывающие проценты клеток, экспрессирующих антиген CD44 (красные точки); а также графики популяций CD31-негативных и CD31-позитивных клеток (верхние правые и нижние правые графики, соответственно), показывающие проценты клеток, экспрессирующих антиген CD44 (красные точки).
Фиг.27 показывает графики рассеяния по определению проточной цитометрией свойств популяций клеток негативной линии, экспрессирующих антиген CD44 (левый ряд графиков), и субпопуляций клеток костного мозга, которые не экспрессируют антиген CD44 на значительном уровне (правый ряд графиков), иллюстрирующие относительные проценты клеток, экспрессирующих другие антигены клеточной поверхности.
Фиг.28 показывает микрофотографические изображения сетчатки мыши, интравитреально инъецированной клетками популяции клеток MLBM по изобретению (слева), в сравнении с сетчаткой мыши, интравитреально инъецированной клетками CD44lo.
Фиг.29 показывает микрофотографические изображения сетчатки глаз, инъецированных клетками популяции клеток MLBM (CD44hi) и клетками CD44lo.
Фиг.30 показывает графики, демонстрирующие благоприятное действие популяции клеток MLBM для уменьшения интесивности патогенного ангиогенеза и промотирования благоприятной физиологической реваскуляризации сетчаток мышей на модели вызванной кислородом ретинопатии - ретролетальной фиброплазии. На верхнем графике сравниваются площадь пучков преретинальных новых образовавшихся сосудов в случае контрольной сетчатки (первый столбик), сетчатки, обработанной клетками CD44lo (средний столбик), и сетчаток, обработанных клетками из популяции клеток MLBM (правый столбик). На нижнем графике сравнивается площадь сосудистой облитерации контрольной сетчатки (первый столбик), сетчатки, обработанной клетками CD44lo (средний столбик), и сетчаток, обработанных клетками из популяции клеток MLBM (правый столбик).
Фиг.31 представляет собой микрофотографическое изображение, показывающее, что только клетки из популяции клеток MLBM включаются в сосудистую сеть сетчатки, клетки экспрессируют васкулярный эндотелиальный фактор роста (VEGF), как показывает окрашивание в зеленый цвет клеток в нижней части изображения.
Фиг.32 представляет микрофотографические изображения, показывающие, что клетки из популяции клеток MLBM CD11b+ по изобретению селективно выбирают мишенью сосудистую сеть сетчатки.
Фиг.33 представляет микрофотографические изображения, показывающие, что клетки костного мозга CD44-CD11b- сосудистую сеть сетчатки селективно мишенью не выбирают.
Фиг.34 показывает последовательность аминокислотных остатков фрагмента Т2 TrpRS (SEQ ID NO: 3) и его вариации Т-2-TrpRS-GD (SEQ ID NO: 4).
Фиг.35 показывает последовательность аминокислотных остатков мини-TrpRS (SEQ ID NO: 5).
Фиг.36 показывает последовательность аминокислотных остатков Т1-TrpRS (SEQ ID NO: 6).
Фиг.37 показывает нормальное развитие сосудов сетчатки у мыши-модели, вызванной кислородом ретинопатии (OIR), и эффект спасения после интравитреальной трансплантации клеток, полученных из линии клеток костного мозга. Мышь рождается с бессосудистой сетчаткой, как видно в постнатальный день 2 (Р2) (изображение а, тотальный препарат сетчатки), когда сосуды обнаруживают на поверхности сетчатки, занимающей одну плоскость, как видно на b. Изображения b, d и f представляют собой изображения, взятые из наборов данных трехмерного (3D) исполнения анфас конфокальной z-серии, повернутых на 90%. В течение первой недели после рождения поверхностная сосудистая сеть сетчатки растет радиально от диска зрительного нерва, почти достигая периферии в Р10 (с). Затем в течение второй недели создается глубокая ретинальная сосудистая сеть из ответвления поверхностного слоя (d). Наконец, между первыми двумя сплетениями образуется третье сплетение сосудов, и зрелая сосудистая сеть сетчатки устанавливается в примерно Р30 (е, f). Изображение g показывает, что воздействие гипероксии на модель OIR вызывает центральную вазооблитерацию, как в данном случае видно в Р10. Изображение h показывает, что после возвращения к нормоксии в Р12 в центральной сетчатке начинается реваскуляризация, и характерные пучки преретинальных новых сосудов образуются на поверхности между васкуляризованной (периферической) и бессосудистой (центральной) сетчаткой. Такие пучки интенсивно окрашиваются изолектином. Части i-n показывают, что гемопоэтические клетки-предшественники Lin- промотируют восстановление сосудов на модели OIR. Клетки Lin-, инъецированные интравитреально перед интенсивным воздействием кислорода, существенно ускоряют реваскуляризацию центральной сетчатки при сравнении с другим глазом, обработанным носителем, в Р17. В то время как сетчатка, обработанная носителем, показывает частичное отсутствие поверхностной сосудистой сети (i) и полное отсутствие глубокой ретинальной сосудистой сети (k, m), обработанный клетками Lin- другой глаз показывает относительно нормальную сосудистую сеть (j) с наличием всех трех сплетений (k, m). Часть о показывает, что в Р17 глаза OIR, обработанные клетками Lin-, полностью реваскуляризуются значительно чаще, чем глаза, не подвергавшиеся инъекции, или глаза с инъекцией носителя. Сосуды визуализируют кардиальной перфузией флуоресцеина-декстрана, как показано на изображениях a-f, i j, и лектином GS, изображения g, h, k-n. Ядра на изображениях k-n мечены DAPI.
Фиг.38 показывает, что клетки Lin- ускоряют ретинальную реваскуляризацию и уменьшают образование пучков преретинальных новых сосудов при OIR. Изображения a-d показывают применение метода компьютерного анализа изображений для вычисления площади облитерации сосудов сетчатки, а также образования пучков преретинальных новых сосудов (красный цвет) на тотальных препаратах сетчатки из глаз OIR в постнатальный день 17. Часть е показывает, что сетчатки, обработанные клетками Lin- перед гипероксией, показывают почти 6-кратное снижение площади облитерации относительно контрольных не подвергавшихся инъекции образцов и приблизительно 5-кратное снижение по сравнению с сетчатками глаз, обработанных одним носителем. Часть f показывает, что обработка клетками Lin- существенно уменьшает двухмерную площадь пучков новых сосудов по сравнению с сетчатками необработанных глаз и обработанных одним носителем. Часть g показывает, что трансплантация клеток Lin- эффективна для снижения площади облитерации не только когда клетки вводят до гипероксии, но также в Р9-Р12 во время гипероксии и также после возвращения в условия нормоксии (графики представляют среднее ± SEM, *p<0,001).
Фиг.39 показывает, что обработка клетками костного мозга оказывает незначительное или непродолжительное токсическое действие. Сетчатки, оцененные через 5 или 6 месяцев после обработки клетками Lin-, имеют нормальную на вид сосудистую сеть, и нейронная сетчатка на поперечных срезах гистологически выглядит сохранившейся (a-f, сетчатка, не подвергавшаяся инъекции, против инъецированной клетками Lin-, 6 месяцев после трансплантации). Опухолей не отмечают, и единственной аномалией является случайная "розетка" в нейронной сетчатке, которую также можно видеть в случае контрольных глаз, не подвергавшихся инъекции (g, h).
Фиг.40 показывает, что клетки CD44HI превалируют в популяции Lin- и эффективно промотируют восстановление сосудов на модели OIR. Часть а показывает, что костный мозг содержит фракции CD44HI и CD44LO, и популяция обогащена клетками CD44HI по сравнению с контрольными клетками CD. Вставки показывают свойства слабого рассеяния клеток CD44HI, которые являются типично моноцитами и гранулоцитами, в то время как свойства слабого рассеяния клеток CD44LO являются свойствами типичных лимфоцитов. Часть b показывает в Р17 характерные сетчатки глаз, обработанных клетками костного мозга CD44LO и CD44HI перед воздействием кислорода. Нижние части представляют примеры количественного определения площадей облитерации и образования новых сосудов в Р17, используемых для представления данных, показанных на фрагменте с. Часть с показывает, что облитерация сосудов и преретинальное образование новых сосудов уменьшаются в случае глаз, обработанных клетками CD44HI, с эффективностью, схожей со случаем, когда глаза обрабатывают клетками Lin-. Площади облитерации сосудов (*) и преретинального образования новых сосудов (**) существенно меньше при инъекции в глаз CD44HI и Lin- по сравнению с инъекцией носителя или без инъекции (p<10-5 во всех случаях). Площадь облитерации для глаз, обработанных Lin-, также уменьшается по сравнению с CD44HI (р=0,03), но в значительно меньшей степени. Площади преретинального образования новых сосудов для глаз, обработанных Lin- и CD44HI, существенно не отличаются (р=0,25).
Фиг.41 показывает, что субпопуляция CD44HI экспрессирует маркеры миелоидных линий. На части а отражается применение проточной цитометрии с использованием двухцветной маркировки для дополнительной характеризации популяций CD44. Все клетки метят антителами против CD44 и метят совместно указанными антителами. Популяция клеток CD44HI показывает сильное мечение в случае CD11a, CD11b и Ly6GC. Фракции клеток CD44hi позитивны в отношении CD14, F4/80, cKit и CD115. Большинство указанных антигенов присутствует в миелоидных клеточных линиях. Клетки CD44lo сильно метятся Ter119 и CD45R B220, которые являются маркерами для эритробластов и В-клеток соответственно.
Фиг.42 показывает, что клетки CD44HI закрепляются в периваскулярном пространстве сетчатки. Конфокальное изображение используют для создания ряда изображений в z-измерении, которые приводят к 3D. На части а показана проекция этого и показаны меченый CD31 сосудистый эндотелий и экспрессия GFP из введенных клеток костного мозга. Предполагается, что клетка костного мозга появляется в периваскулярной позиции. Данные 3D показывают, что визуализируются просвет сосуда и относительное положение клетки костного мозга GFP+. Числа, указанные на (b), соответствуют позициям поперечного сечения, указанным на (а). Сигнал GFP детектируют вне просвета во всех случаях, за исключением №3, часть b, который представляет срез клеточного тела с интенсивной флуоресценцией, где очевиден сигнал просачивания крови.
Фиг.43 показывает анализ in situ инъецированных клеток костного мозга CD44HI на модели OIR. Мечение контрольной сетчатки, которая не получает обработки клетками, показывает присутствие эндогенных периваскулярных клеток F4/80+ (a-c). Инъецированные клетки CD44HI выбирают мишенью сосудистую сеть сетчатки и имеют локализацию, морфологию и картину экспрессии F4/80, схожие с эндогенными клетками (d-i). Трансплантированные периваскулярные клетки костного мозга утрачивают экспрессию CD44, в то время как клетки, не ассоциированные с сосудистой сетью сетчатки, сохраняют экспрессию CD44 (j-o).
Фиг.44 показывает матричный анализ экспрессии, который показывает высокую экспрессию миелоидоассоциированных генов в популяции CD44HI, в то время как клетки CD44LO экспрессируют гены, ассоциированные с лимфоидными клетками. Используют матрицы AFFYMETRIX® для сравнения профилей экспрессии генов между указанными двумя популяциями клеток костного мозга. Показанные гены имеют минимум 5-кратное различие в экспрессии. Наблюдают существенно более высокие уровни экспрессии CD44 в популяции CD44HI по сравнению с клетками CD44LO.
Фиг.45 показывает, что клетки CD44HI могут дифференцировать в клетки с микроглиальными характеристиками. Части А и В показывают, что инъецированные клетки CD44HI экспрессируют CD11b и F4/80 и имеют морфологию и периваскулярную локализацию, схожие с эндогенными микроглиями. Часть С представляет трехмерное изображение периваскулярной локализации инъецированной клетки CD44HI. Часть D показывает вид морфологии инъецированных клеток CD44HI при большом увеличении.
Фиг.46 показывает, что клетки CD44HI можно изолировать негативным отбором. Часть А показывает, что истощение костного мозга мыши MACS с использованием антител, селективных для CD45R/B220, TER119 и CD3e, дает популяцию клеток, которые более чем на 90% являются клетками CD44HI. Часть В показывает, что негативная фракция (популяция CD44HI), по существу, не содержит клетки CD45R/B220, TER119 и CD3e. Часть С показывает, что негативно отобранные клетки CD44HI сохраняют способности к выбору мишени и дифференцировке.
Подробное описание предпочтительных воплощений
Клетки костного мозга включают субпопуляцию клеток, которые экспрессируют антиген CD44 (т.е. рецептор гиалуроновой кислоты) и CD11b (т.е. интегрин αM). Миелоидоподобную популяцию клеток костного мозга, обогащенную клетками, экспрессирующими CD44 и CD11b, можно выделить из костного мозга обработкой клеток костного мозга антителами к антигену CD44 (анти-CD44) и/или антителами к антигену CD11b (анти-CD11b) и последующим отбором клеток, которые иммунореактивны в отношении антител. Затем антитела удаляют из клеток способами, хорошо известными в технике. Клетки можно отобрать, например, с использованием проточной цитометрии, с использованием антител, связанных с или нанесенных на гранулы, с последующей фильтрацией, или другими способами разделения, хорошо известными в технике. Большинство отобранных клеток являются негативными линиями и экспрессируют как антиген CD44, так и антиген CD11b, в зависимости от того, какие антитела используют при изоляции.
Клетки костного мозга включают стволовые клетки. Стволовые клетки типично идентифицируются распределением антигенов на поверхности клеток (подробное обсуждение см. в Stem Cells: Scientific Progress and Future Directions, сообщение подготовлено National Institute of Health, Office of Science Policy, June 2001, Appendix E: Stem Cell Markers, сообщение включено в настоящее описание в качестве ссылки). Приблизительно 75% гемопоэтических стволовых клеток негативной линии, изолированных из костного мозга, также являются CD44-позитивными. В предпочтительном воплощении большинство клеток из популяции клеток MLBM являются гемопоэтическими стволовыми клетками негативной линии (т.е. CD44+Lin-HSC).
Настоящее изобретение относится к способу уменьшения интенсивности сосудистой и нейронной дегенерации в сетчатке млекопитающего, страдающего от болезни глаз. Изолированную популяцию клеток MLBM по изобретению вводят в сетчатку млекопитающего, предпочтительно, интравитреальной инъекцией. Клетки вводят в количестве, достаточном для уменьшения интенсивности сосудистой и/или нейронной дегенерации в сетчатке. Предпочтительно, изолированная популяция клеток MLBM аутологична млекопитающему, которого лечат. Предпочтительно, клетки из популяции клеток MLBM вводят в физиологически переносимой среде, такой как забуференный фосфатом физиологический раствор (PBS).
Предпочтительный способ включает выделение популяции клеток MLBM из костного мозга млекопитающего, которого лечат, и последующее введение клеток млекопитающему в количестве, достаточном для уменьшения интенсивности сосудистой и/или нейронной дегенерации в сетчатке. Клетки можно выделить из млекопитающего, страдающего от дегенеративной болезни глаз, предпочтительно, на ранней стадии болезни глаз, или из здорового млекопитающего, о котором известно, что оно предрасположено к дегенеративной болезни глаз (т.е. по генетической предрасположенности). В последнем случае изолированную популяцию клеток MLBM можно сохранить после выделения и затем можно инъецировать для профилактики на ранних стадиях развивающейся позднее болезни глаз. Предпочтительно, больная сетчатка включает активированные астроциты, которые становятся мишенью для клеток из популяции клеток MLBM. Соответственно, раннее лечение глаз, когда имеется ассоциированный глиоз, является благоприятным. С другой стороны, сетчатку можно обработать лазером для стимуляции локальной пролиферации активированных астроцитов в сетчатке перед введением аутологичной популяции клеток MLBM.
Гемопоэтические стволовые клетки представляют собой стволовые клетки, способные развиваться в различные типы клеток крови, например, В-клетки, Т-клетки, гранулоциты, тромбоциты и эритроциты. Линиеспецифические поверхностные антигены представляют собой группу белков клеточной поверхности, которые являются маркерами зрелых линий клеток крови, включая CD2, CD3, CD11, CD11a, Mac-1 (CD11b:CD18), CD14, CD16, CD19, CD24, CD33, CD36, CD38, CD45, CD45RA, мышиный Ly-6G, мышиный TER-119, CD56, CD64, CD68, CD86 (B7.2), CD66b, лейкоцитарный антиген человека DR (HLA-DR) и CD235a (гликофорин А). Гемопоэтические клетки, которые не экспрессируют значительные уровни таких антигенов, обычно относят к негативной линии (Lin-). Гемопоэтические стволовые клетки человека обычно экспрессируют другие поверхностные антигены, такие как CD31, CD34, CD117 (c-kit) и/или CD133. Мышиные гемопоэтические стволовые клетки обычно экспрессируют другие поверхностные антигены, такие как CD34, CD117 (c-kit), Thy-1 и/или Sca-1.
Изолированные гемопоэтические стволовые клетки, которые не экспрессируют "значительные уровни поверхностного антигена" (Lin) на своей клеточной поверхности, относят в данном описании к гемопоэтическим стволовым клеткам "негативной линии" или "Lin-", т.е. Lin- HSC. Большинство клеток популяций клеток MLBM по настоящему изобретению являются Lin- и экспрессируют как относительно большое количество антигена CD44 (CD44hi), так и антигена CD11b. Такие CD44+CD11b+ Lin- HSC способны внедряться в развивающуюся сосудистую сеть и затем дифференцировать и становиться эндотелиальными клетками сосудистой сети.
Используемое в данном описании и прилагаемой формуле изобретения определение "зрелый" в отношении костного мозга и клеток костного мозга включает костный мозг, выделенный постнатально, т.е. из молодых и взрослых особей, в отличие от эмбрионов. Соответственно, термин "взрослое млекопитающее" относится как к молодым (постнатальным), так и созревшим млекопитающим, в отличие от эмбриона или пренатального индивидуума.
Изолированные популяции клеток MLBM по настоящему изобретению селективно нацеливаются на астроциты и включаются в новую сосудистую сеть сетчатки, когда инъецированы интравитреально в глаз млекопитающего такого вида, как мышь или человек, из организма которого выделены клетки.
Изолированные популяции клеток MLBM по настоящему изобретению включают клетки, которые дифференцируют в эндотелиальные клетки и образуют сосудистые структуры в сетчатке. В частности, популяция клеток MLBM по настоящему изобретению применима для лечения ретинальных неоваскулярных и ретинальных сосудистых дегенеративных болезней и для восстановления повреждения сосудов сетчатки. Популяция клеток MLBM по настоящему изобретению также промотирует спасение нейронов в сетчатке и промотирует позитивную регуляцию антиапоптозных генов. Кроме того, популяцию клеток MLBM по настоящему изобретению можно использовать для лечения дефектов сетчатки глаз новорожденных млекопитающих, таких как млекопитающие, страдающие от ретинопатии, вызванной кислородом, или ретролетальной фиброплазии.
Обнаружено, что клетки костного мозга, которые не экспрессируют CD44 (клетки CD44LO), как правило, экспрессируют один или несколько следующих маркеров: Ter119, CD45RB220 и CD3e. Используя такой факт, клетки MLBM CD44HI по настоящему изобретению можно изолировать способом, включающим негативный отбор с маркером. Способ включает контактирование множества клеток костного мозга с антителами, специфическими к Ter119, CD45RB220 и CD3e, удаление клеток из множества клеток костного мозга, которые вступают в иммунные реакции с антителами к Ter119, CD45RB220 и CD3e, и извлечение миелоподобных клеток костного мозга, из которых удалены клетки, экспрессирующие Ter119, CD45RB220 и CD3e. С использованием такого способа можно извлечь клеточную популяцию, в которой более 90% клеток экспрессируют CD44.
Настоящее изобретение также относится к способу лечения болезней глаз у млекопитающего, включающему выделение из костного мозга млекопитающего популяции клеток MLBM и интравитреальную инъекцию клеток из популяции клеток MLBM в глаз млекопитающего в количестве, достаточном для приостановки заболевания. Способ по настоящему изобретению можно использовать для лечения болезней глаз, таких как дегенеративные болезни сетчатки, дегенеративные болезни сосудов сетчатки, ишемические ретинопатии, кровоизлияния, просачиваемость сосудов и хориоретинопатии у новорожденных, молодых или полностью созревших млекопитающих. Примерами таких болезней являются связанная дегенерация желтого пятна (ARMD), диабетическая ретинопатия (DR), (предположительно) глазной гистоплазмоз (POHS), ретролетальная фиброплазия (ROP), серповидно-клеточная анемия и пигментный ретинит, а также повреждения сетчатки.
Число клеток из популяции клеток MLBM, инъецируемых в глаз, является достаточным для угнетения болезненного состояния глаза. Например, количество инъецируемых клеток может быть эффективным для восстановления повреждения сетчатки глаза, стабилизации новой сосудистой сети сетчатки и предупреждения или заживления просачивания сосудов и кровоизлияния.
Клетки из популяции клеток MLBM по настоящему изобретению можно трансфицировать терапевтически применимыми генами, такими как гены, кодирующие антиангиогенные белки, для применения при лечении глаз и в генной терапии на основе клеток, и генами, кодирующими нейротрофические средства, для усиления эффектов спасения нейронов.
Трансфицированные клетки могут включать любой ген, который терапевтически полезен для лечения ретинальных расстройств. В одном предпочтительном воплощении трансфицированные клетки из популяции клеток MLBM по настоящему изобретению включают ген, операбельно кодирующий антиангиогенный пептид, в том числе, белки или фрагменты белков, такие как TrpRS или его антиангиогенные (т.е. ангиостатические) фрагменты, например фрагменты TrpRS, названные Т2-TrpRS (SEQ ID NO: 3 на фиг.34), Т2-TrpRS-GD (SEQ ID NO: 4 на фиг.34), которые оба являются предпочтительными ангиостатическими пептидами, а также мини-TrpRS (SEQ ID NO: 5 на фиг.35) и Т1-TrpRS (SEQ ID NO: 6 на фиг.36). Трансфицированные клетки из популяции клеток MLBM, кодирующие антиангиогенный пептид по настоящему изобретению, применимы для лечения болезней сетчатки, включая аномальное развитие сосудов, такое как диабетическая ретинопатия, и подобные заболевания. Предпочтительно, клетки из популяции клеток MLBM являются клетками человека.
В другом предпочтительном воплощении трансфицированные клетки из популяции клеток MLBM по настоящему изобретению включают ген, операбельно кодирующий нейротрофическое средство, такое как фактор роста нервов, нейротрофин-3, нейротрофин-4, нейротрофин-5, цилиарный нейротрофический фактор, образованный ретинальным пигментированным эпителием нейротрофический фактор, инсулиноподобный фактор роста, образованный глиальной клеточной линией нейротрофический фактор, образованный головным мозгом нейротрофический фактор, и подобные средства. Такие нейротрофические клетки из популяции клеток MLBM применимы для промотирования спасения нейронов при ретинальных нейронных дегенеративных заболеваниях, таких как глаукома и пигментный ретинит, при лечении повреждений нервов сетчатки и т.п. Имеются сообщения, что имплантаты цилиарного нейротрофического фактора применимы для лечения пигментного ретинита (см. Kirby et al., 2001, Mol. Ther., 3(2): 241-8; Farrar et al., 2002, EMBO Journal, 21: 857-864). Сообщается, что нейротрофический фактор, образованный головным мозгом, модулирует ассоциированные с ростом гены в поврежденных ганглиях сетчатки (см. Fournier et al., 1997, J. Neurosci. Res., 47: 561-572). Линия глиальных клеток, образованная нейротрофическим фактором, как сообщается, замедляет дегенерацию фоторецепторов при пигментном ретините (см. McGee et al., 2001, Mol. Ther., 4(6): 622-9).
Настоящее изобретение также относится к способам лечения глазных ангиогенных болезней введением трансфицированных клеток из популяции клеток MLBM по настоящему изобретению интравитреальной инъекцией клеток в глаз. Такие трансфицированные клетки из популяции клеток MLBM содержат клетки из популяции клеток MLBM, трансфицированных терапевтически применимым геном, таким как ген, кодирующий антиангиогенный или нейротрофический генный продукт. Предпочтительно, трансфицированные клетки из популяции клеток MLBM являются клетками человека.
Предпочтительно, по меньшей мере, примерно 1×105 клеток из популяции клеток MLBM или трансфицированных клеток из популяции клеток MLBM вводят интравитреальной инъекцией в глаз млекопитающего, страдающего от дегенеративной болезни сетчатки. Число клеток, которые следует инъецировать, может зависеть от тяжести дегенерации клетчатки, возраста млекопитающего и других факторов, которые будут вполне очевидны для специалиста в области лечения болезней сетчатки. Клетки из популяции клеток MLBM можно вводить в однократной дозе или вводить несколькими дозами в течение некоторого времени, как определит клиницист при назначении лечения.
Популяции клеток MLBM по настоящему изобретению применимы для лечения повреждений сетчатки и дефектов сетчатки, включая внедрение в или деградацию сосудистой сети сетчатки или деградацию нейронов сетчатки. Популяции клеток MLBM человека также можно использовать для разрушения линии генетически идентичных клеток, т.е. клонов, для применения при регенеративном или восстановительном лечении сосудистой сети сетчатки, а также для лечения или уменьшения интенсивности деградации нейронов сетчатки. Более того, популяции клеток MLBM по настоящему изобретению применимы в качестве исследовательских инструментов для исследования развития сосудов сетчатки и для доставки генов в избранные клетки-мишени, такие как астроциты.
Развитие сосудов сетчатки у мыши
Модель глазного ангиогенеза. Мышиный глаз обеспечивает адекватную модель для исследования разития сосудов сетчатки млекопитающего. Во время развития сосудов сетчатки мыши, подгоняемые ишемией кровеносные сосуды развиваются в тесной связи с астроцитами. Такие глиальные элементы мигрируют на третий триместр сетчатки плода человека, или у новорожденного грызуна, от диска зрительного нерва вдоль слоя ганглиозных клеток и быстро распространяются. Так как сосудистая сеть сетчатки мыши развивается, эндотелиальные клетки используют такую уже установившуюся астроцитарную матрицу для определения рисунка сосудов сетчатки (см. фиг.1 (а и b)). Фиг.1 (а и b) отображает принципиальные схемы развития сетчатки у мыши. Часть (а) отображает развитие первичного сплетения (темные полосы на схеме вверху слева), расположенные над астроцитарной матрицей (светлые линии), в то время как (b) отображает вторую фазу образования сосудов сетчатки. На фиг.1 GCL обозначает слой ганглиозных клеток; IPL обозначает внутренний слой сплетения; INL обозначает внутренний ядерный слой; OPL обозначает наружный слой сплетения; ONL обозначает наружный ядерный слой; RPE обозначает ретинальный пигментный эпителий; ON обозначает зрительный нерв; Р обозначает периферию.
При рождении сосудистая сеть сетчатки фактически отсутствует. В постнатальный день 14 (Р14) в сетчатке развит комплекс первичного (поверхностного) и вторичного (глубокого) слоев сосудов сетчатки, что совпадает с появлением зрения. Сначала напоминающие спицы сосуды растут радиально над уже существующей сетью астроцитов к периферии, причем становятся прогрессивно взаимосвязанными путем образования капиллярного сплетения. Указанные сосуды растут в виде монослоя в нервном волокне до Р10 (фиг.1 (а)). В промежутке Р7-Р8 коллатеральные ответвления начинают вырастать из такого первичного сплетения и проникать в сетчатку до наружного плексиформного слоя, где они образуют вторичное или глубокое ретинальное сплетение. В Р21 вся сеть претерпевает экстенсивное исправление, и третичное, или промежуточное, сплетение образуется на внутренней поверхности внутреннего ядерного слоя (фиг.1 (b)).
Модель ретинального ангиогенеза у новорожденной мыши применима для исследования роли HSC во время глазного ангиогенеза по нескольким причинам. У данной физиологически релевантной модели большая астроцитарная матрица существует до появления эндогенных кровеносных сосудов, позволяя оценить роль клетки в нацеливании на клетку во время процесса образования новых сосудов. Кроме того, известно, что такой последовательный и воспроизводимый процесс образования сосудов сетчатки у новорожденных управляется гипоксией, причем в этом отношении существует сходство для многих болезней сетчатки, в которых, как известно, играет роль ишемия.
Накопление эндотелиальных клеток-предшественников (ЕРС) из костного мозга
Хотя экспрессия маркеров на поверхности широко оценена на популяции ЕРС, обнаруженной в препаратах HSC, маркеры, которые уникально идентифицируют ЕРС, пока недостаточно определены. Для того чтобы накопить ЕРС, из одноядерных клеток костного мозга мыши извлекают гемопоэтические клетки маркерпозитивной линии (Lin+), т.е. В-лимфоциты (CD45), Т-лимфоциты (CD3), гранулоциты (Ly-6G), моноциты (CD11) и эритроциты (TER-119). Для дальнейшего накопления ЕРС используют антиген Sca-1. Сравнение результатов, полученных после интравитреальной инъекции идентичных количеств или клеток Lin- Sca-1+, или клеток Lin-, не обнаруживает различия между указанными двумя группами. Действительно, когда инъецируют только клетки Lin- Sca-1-, наблюдают гораздо большее включение в развивающиеся кровеносные сосуды.
Популяции Lin- HSC обогащаются ЕРС, как показывают функциональные анализы. Более того, популяции Lin+ HSC функционально ведут себя вполне отлично от популяций Lin- HSC. Эпитопы, обычно используемые для идентификации ЕРС для каждой фракции (на основании предварительных сообщений об исследованиях свойств in vitro), также оцениваются. Хотя ни один из указанных маркеров не ассоциируется исключительно с фракцией Lin-, во всех случаях имеется возрастание от 70 до примерно 1800% в Lin- HSC по сравнению с Lin+ HSC (фиг.1 (с)). Фиг.1, часть (с), иллюстрирует характеризацию проточной цитометрией Lin+ HSC и Lin- HSC, выделенных из костного мозга. Верхний ряд части (с) представляет точечный график распределения немеченных антителами клеток. R1 показывает определяемую количественно площадь дискриминационного окна РЕ-позитивного окрашивания; R2 показывает GFP-позитивное окрашивание. Точечные графики Lin- HSC приводятся в среднем ряду, и точечные графики Lin+ HSC приводятся в нижнем ряду. Клетки С57В/6 метят РЕ-конъюгированными антителами к Sca-1, c-kit, Flk-1/KDR, CD31. Данные по Tie-2 получают на мышах Tie-2-GFP. Проценты, указанные в углах точечных графиков, показывают процент позитивно меченых клеток из всей популяции Lin- HSC или Lin+ HSC. Представляет интерес то, что принятые для ЕРС маркеры, подобные Flk-1/KDR, Tie-2 и Sca-1, экспрессируются плохо и поэтому не используются для дальнейшего фракционирования.
Lin- HSC можно выделить (а) экстрагированием костного мозга взрослого млекопитающего; (b) разделением множества моноцитов из костного мозга; (с) мечением моноцитов биотинилированными линиеспецифическими панелированными антителами к одному или нескольким линиеспецифическим поверхностным антигенам, предпочтительно, линиеспецифическим поверхностным антигенам, выбранным из группы, состоящей из CD2, CD3, CD4, CD11, CD11a, Mac-1, CD14, CD16, CD19, CD24, CD33, CD36, CD38, CD45, Ly-6G (мышиный), TER-119 (мышиный), CD45RA, CD56, CD64, CD68, CD86 (B7.2), CD66b, DR главного комплекса гистосовместимости человека (HLA-DR) и CD235a (гликофорин А); (d) удалением из множества моноцитов моноцитов, позитивных для указанных одного или нескольких линиеспецифических поверхностных антигенов; и (е) извлечением из них популяции линии негативных гемопоэтических стволовых клеток.
Когда Lin- HSC выделяют из костного мозга взрослого человека, предпочтительно, моноциты метят биотинилированными линиеспецифическими панелированными антителами к линиеспецифическим поверхностным антигенам CD2, CD3, CD4, CD11a, Mac-1, CD14, CD16, CD19, CD33, CD38, CD45RA, CD64, CD68, CD86 (B7.2) и CD235a. Когда Lin- HSC выделяют из костного мозга взрослой мыши, предпочтительно, моноциты метят биотинилированными линиеспецифическими панелированными антителами к линиеспецифическим поверхностным антигенам CD3, CD11, CD45, Ly-6G и TER-119.
Интравитреально инъецированные клетки Lin - HSC содержат ЕРС, которые выбирают мишенью астроциты и внедряются в развивающуюся ретинальную сосудистую сеть.
Для того чтобы определить, могут ли интравитреально инъецированные Lin- HSC выбрать мишенью специфические типы клеток сетчатки, использовать астроцитарную матрицу и принимать участие в ретинальном ангиогенезе, приблизительно 105 клеток из композиции Lin- HSC по настоящему изобретению или клеток Lin+ HSC (контроль, примерно 105 клеток), выделенных из костного мозга взрослых мышей (GFP или трансгенных LacZ), инъецируют в глаза мышей на 2 день после рождения (Р2). Через четыре дня после инъекции (Р6) многие клетки из композиции Lin- HSC по настоящему изобретению, полученные от GFP или трансгенных LacZ мышей, прилипают к сетчатке и имеют характерный удлиненный вид эндотелиальных клеток (фиг.2 (а)). Фиг.2 отображает внедрение клеток Lin- в разивающуюся сетчатку мыши. Как видно на фиг.2, изображение (а), на четвертые сутки после инъекции (Р6) интравитреально инъецированные eGFP+ Lin- HSC прикрепляются и дифферецируют на сетчатке.
Во многих участках сетчатки GFP-экспрессирующие клетки располагаются в виде, соответствующем нижележащим астроцитам и имеющем сходство с кровеносными сосудами. Такие флуоресцирующие клетки наблюдают в будущей эндогенной развивающейся сосудистой сети (фиг.2 (b)). Напротив, только небольшое число Lin+ HSC (фиг.2 (с)) или зрелых мезентериальных эндотелиальных клеток мыши (фиг.2 (d)) прикрепляются к поверхности сетчатки. Для того чтобы определить, могут ли клетки из инъецированной популяции Lin- HSC также присоединяться к сетчаткам с уже установившимися сосудами, Lin- HSC инъецируют в зрелые глаза. Интересно, что не наблюдают прикрепления клеток к сетчатке или включения в установившиеся здоровые кровеносные сосуды сетчатки (фиг.2 (е)). Это показывает, что композиция Lin- HSC по настоящему изобретению не разрушает нормально развитую сосудистую сеть и не будет инициировать аномальную васкуляризацию в нормально развитой сетчатке.
Для того чтобы определить соотношение между инъецированными композициями Lin- HSC по настоящему изобретению и ретинальными астроцитами, используют трансгенную мышь, которая экспрессирует глиальный фибриллярный кислый белок (GFAP - маркер астроцитов) и управляемый промотором зеленый белок флуоресценции (GFP). Проверка сетчаток таких GFAP-GFP трансгенных мышей с Lin- HSC от eGFP трансгенной мыши показывает совместную локализацию инъецированных eGFP ЕРС и существующих астроцитов (фиг.2 (f-h), стрелки). Наблюдают, что картины GFP + Lin- HSC соответствуют нижележащей сети астроцитов (фиг.2 (g)). Обследование таких глаз показало, что инъецированные меченые клетки присоединяются только к астроцитам; в Р6 в сетчатках мышей, в которых на периферии сетчаток пока не имеется эндогенных сосудов, инъецированные клетки наблюдают как прилипшие к астроцитам в пока не васкуляризованных участках. Неожиданно оказалось, что инъецированные меченые клетки наблюдают в более глубоких слоях сетчатки точно в тех местах, где потом будут развиваться нормальные сосуды (фиг.2 (i), стрелки).
Для того чтобы определить, устойчиво ли инъецированные Lin- HSC включаются в развивающуюся ретинальную сосудистую сеть, сосуды сетчатки обследуют позднее в несколько моментов времени. Как и ранее в Р9 (семь дней после инъекции), Lin- HSC включаются в структуры CD31+ (фиг.2 (j)). В Р16 (14 дней после инъекции) клетки уже в большой мере включены в структуры, подобные сосудам сетчатки (фиг.2 (k)). Когда родамин-декстран инъецируют интраваскулярно (для идентификации функциональных кровеносных сосудов сетчатки) перед умерщвлением животных, большинство Lin- HSC объединяются с открытыми сосудами (фиг.2 (l)). Наблюдают две картины распределения меченых клеток: (1) на одном изображении клетки размещены вдоль сосудов в клетках между немечеными эндотелиальными клетками; и (2) другое изображение показывает, что сосуды полностью состоят из меченых клеток. Инъецированные клетки также включаются в сосуды глубокого сосудистого сплетения (фиг.2 (m)). Хотя о спорадическом включении ЕРС, полученных из Lin- HSC, в новую сосудистую сеть сообщалось ранее, это первое сообщение о том, что сосудистые сетки целиком состоят из таких клеток. Это показывает, что клетки из популяции Lin- HSC, полученные из костного мозга, инъецированные интрвитреально, могут эффективно включаться в любой слой формирующегося сосудистого сплетения сетчатки.
Гистологическое обследование тканей, не относящихся к сетчатке (например, головного мозга, печени, сердца, легких, костного мозга), не показывает наличия каких-либо GFP-позитивных клеток при обследовании в пределах 5 или 10 после интравитрельной инъекции. Это показывает, что субпопуляция клеток в фракции Lin- HSC селективно нацеливается на ретинальные асторциты и устойчиво включается в развивающуюся ретинальную сосудистую сеть. Так как такие клетки имеют многие свойства эндотелиальных клеток (ассоциация с ретинальными астроцитами, вытянутая морфология, устойчивое включение в открытые сосуды и отсутствие вне сосудов), такие клетки представляют ЕРС, представленные в популяции Lin- HSC. Астроциты-мишени имеют тот же тип, который наблюдают при многих гипоксических ретинопатиях. Хорошо известно, что глиальные клетки являются заметным компонентом ветвей новых сосудов пучков, наблюдаемых при DR и других формах повреждения сетчатки. В условиях реактивного глиоза и вызванного ишемией образования новых сосудов активированные астроциты пролиферируют, продуцируют цитокины и позитивно регулируют GFAP, подобно тому, что наблюдают во время образования сосудов сетчатки при рождении у многих видов млекопитающих, включая людей.
Популяции Lin- HSC будут нацеливаться на активированные астроциты в глазах взрослой мыши так, как это происходит в отношении глаз новрожденных. Клетки Lin- HSC инъецируют в зрелые глаза с сетчатками, поврежденными фотокоагуляцией (фиг.3 (а)) или кончиком иглы (фиг.3 (b)). На обеих моделях популяцию клеток с заметным окрашиванием GFAP наблюдают только вокруг места повреждения (фиг.3 (а и b)). Клетки из инъецированных композиций Lin- HSC локализуются в месте повреждения и остаются специфически ассоциированными с GFAP-позитивными астроцитами (фиг.3 (а и b)). В таких местах также наблюдают, что клетки Lin- HSC мигрируют в более глубокий слой сетчатки на уровне, подобном уровню, наблюдаемому во время образования глубокой сосудистой сети сетчатки у новорожденных. Неповрежденные части сетчатки не содержат клетки Lin- HSC, идентично тому, что наблюдают, когда Lin- HSC инъецируют в здоровые неповрежденные зрелые сетчатки (фиг.2 (е)). Такие результаты показывают, что композиции Lin- HSC могут селективно нацеливаться на активированные глиальные клетки в поврежденных зрелых сетчатках с глиозом, а также на неонатальные сетчатки, претерпевающие образование кровеносных сосудов.
Интравитреально инъецированные Lin - HSC могут спасать и стабилизировать дегенерирующую сосудистую сеть.
Так как интравитреально инъецированные композиции Lin- HSC нацеливаются на астроциты и включаются в нормальную ретинальную сосудистую сеть, такие клетки также стабилизируют дегенерацию сосудистой сети при ишемических или дегенеративных болезнях сетчатки, ассоциированных с глиозом и дегенерацией сосудов. Мышь rd/rd является моделью в случае дегенерации сетчатки, которая показывает полную дегенерацию фоторецепторных и сосудистых слоев сетчатки через одни месяц после рождения. Сосудистая сеть сетчатки у таких мышей развивается нормально до Р16, когда происходит регрессия глубокого сосудистого сплетения; у большинства мышей глубокое и промежуточное сплетения почти полностью дегенерируют на Р30.
Для того чтобы определить, могут ли HSC спасти регрессирующие сосуды, Lin+ или Lin- HSC (от мышей Balb/c) инъецируют мышам rd/rd интравитреально в Р6. В Р33 после инъекции клеток Lin+ сосуды самого глубокого слоя сетчатки почти полностью отсутствуют (фиг.4 (а и b)). Напротив, большинство сетчаток, инъецированных Lin- HSC, в Р33 имеют почти нормальную ретинальную сосудистую сеть с тремя параллельными хорошо сформированными слоями (фиг.4 (а и d)). Количественное определение такого эффекта показывает, что средняя длина сосудов в глубоком сосудистом сплетении глаз rd/rd, инъецированных Lin-, почти в три раза больше, чем в необработанных или обработанных Lin+ глазах (фиг.4 (е)). Неожиданно оказалось, что инъекция композиции Lin- HSC, полученных из костного мозга взрослой мыши rd/rd (FVB/N), также уменьшает дегенерацию сосудистой сети сетчатки новорожденной мыши rd/rd (фиг.4 (f)). Дегенерацию сосудистой сети глаз мыши rd/rd наблюдают уже через 2-3 недели после рождения. Инъекция Lin- HSC достаточно поздно - в Р15 также приводит к частичной стабилизации дегенерирующей сосудистой сети у мышей rd/rd в течение, по меньшей мере, одного месяца (фиг.4 (g и h)).
Композиция Lin- HSC, инъецированная молодым мышам rd/rd (например, Р2), также включается в развивающуюся поверхностную сосудистую сеть. В Р11 наблюдают, что такие клетки мигрируют до уровня глубокого сосудистого сплетения и образуют рисунок, идентичный рисунку, наблюдаемому в наружном слое сосудов сетчатки у дикого типа (фиг.5 (а)). Для того чтобы более четко описать, каким образом клетки из инъецированных композиций Lin- HSC включаются в дегенерирующую сосудистую сеть сетчатки мышей rd/rd и стабилизируют ее, в глаза мышей Tie-2-GFP FVB инъецируют композицию Lin- HSC, полученных от мышей Balb/c. Мыши FVB имеют генотип rd/rd, и поскольку они экспрессируют слитый белок Tie-2-GFP, все эндогенные кровеносные сосуды флуоресцируют.
Когда немеченые клетки из композиции Lin- HSC инъецируют в глаза новорожденных Tie-2-GFP FVB, и затем они включаются в развивающуюся сосудистую сеть, должны существовать немеченые промежутки в эндогенных меченых Tie-2-GFP сосудах, соответствующие включенным немеченым инъецированным Lin- HSC. Затем последующее окрашивание другим маркером сосудов очерчивает весь сосуд, позволяя определить, являются ли неэндогенные эндотелиальные клетки частью сосудистой сети. Через два месяца после инъекции в сетчатках глаз, инъецированных композицией Lin- HSC, наблюдают CD31-позитивные, Tie-2-GFP-негативные сосуды (фиг.5 (b)). Интересно, что большинство спасенных сосудов сетчатки содержат Tie-2-GFP-позитивные клетки (фиг.5 (с)). Распределение перицитов при определении окрашиванием актина гладких мышц не изменяется инъекцией Lin- HSC, независимо от того, имеется ли спасение сосудов (фиг.5 (d)). Такие данные четко демонстрируют, что инъецированные интравитреально клетки Lin- HSC мигрируют в сетчатку, принимают участие в формировании нормальных кровеносных сосудов сетчатки и стабилизируют эндогенную дегенерирующую сосудистую сеть генетически дефективных мышей.
Ингибирование ретинального ангиогенеза трансфицированными клетками из Lin - HSC
Большинство болезней сосудов сетчатки включает аномальную пролиферацию сосудов, а не дегенерацию. Трансгенные клетки, нацеленные на астроциты, можно использовать для доставки антиангиогенного белка и ингибирования ангиогенеза. Клетки из композиций Lin- HSC трансфицируют Т2-триптофанил-тРНК-синтетазой (Т2-TrpRS). Т2-TrpRS представляет собой фрагмент в 43 кД TrpRS, который сильно ингибирует ретинальный ангиогенез (фиг.6 (а)). В Р12 сетчатки глаз, инъецированных контрольной композицией Lin- HSC, трансфицированной плазмидой (без гена Т2-TrpRS) в Р2, имеют нормальные первичные (фиг.6 (с)) и вторичные (фиг.6 (d)) ретинальные сосудистые сплетения. Когда в глаза в Р2 инъецируют трансфицированную Т2-TrpRS композицию Lin- HSC и оценивают через 10 дней, первичная сетка имеет много аномалий (фиг.6 (е)), и образование глубокой сосудистой сети сетчатки почти полностью подавляется (фиг.6 (f)). Наблюдают, что несколько сосудов в таких глазах заметно разрежены большими промежутками между сосудами. Степень ингибирования Lin- HSC, секретирующими Т2-TrpRS, указана в таблице 1.
Т2-TrpRS продуцируется и секретируется клетками в композиции Lin- HSC in vitro, и после инъекции таких трансфицированных клеток в стекловидное тело в сетчатке обнаруживают фрагмент Т2-TrpRS в 30 кД (фиг.6 9b)). Такой фрагмент в 30 кД специфически обнаруживают только в сетчатках, инъецированных трансфицированными Lin- HSC, и такое снижение кажущейся молекулярной массы по сравнению с рекомбинантным или синтезированным in vitro белком может происходить из-за процессинга или распада Т2-TrpRS in vivo. Такие результаты показывают, что композиции Lin- HSC можно использовать для доставки функционально активных генов, таких как гены, экспрессирующие ангиостатические молекулы, в сосудистую сеть сетчатки за счет нацеливания на активированные астроциты. Хотя вероятно, что наблюдаемое ангиостатическое действие имеет место из-за клеточно-опосредованной активности, это невозможно, так как глаза, обработанные идентичными, но Т2-нетрансфицированными композициями Lin- HSC, имеют нормальную ретинальную сосудистую сеть.
Инъецированные интравитреально популяции Lin- HSC локализуются у астроцитов сетчатки, включаются в сосуды и могут применяться при лечении многих болезней сетчатки. Хотя большинство клеток из инъецированных композиций Lin- HSC прилипает к астроцитарной матрице, небольшое число мигрирует глубоко в сетчатку, располагаясь в участках, где впоследствие будет развиваться глубокая сосудистая сеть. Несмотря на то, что GFAP-позитивные астроциты в данной области не обнаруживают до 42 дня после рождения, это не исключает возможности того, что GFAP-негативные клетки уже присутствуют, обеспечивая сигнал локализации Lin- HSC. Предыдущие исследования показали, что многие болезни ассоциируются с реактивным глиозом. При DR, в частности, глиальные клетки и их внеклеточная матрица ассоциируются с патологическим ангиогенезом.
Так как клетки из инъецированных композиций Lin- HSC специфически прикрепляются к GFAP-экспрессирующим глиальным клеткам, независимо от типа повреждения, композиции Lin- HSC по настоящему изобретению можно использовать для нацеливания на предангиогенные повреждения в сетчатке. Например, при ишемических ретинопатиях, таких как диабет, образование новых сосудов является реакцией на гипоксию. Нацеливая композиции Lin- HSC в места паталогического образования новых сосудов, можно стабилизировать новую сосудистую сеть, предотвратить аномалии новой сосудистой сети, такие как кровоизлияния или отек (случаи потери зрения, связанные с DR), и потенциально можно смягчить гипоксию, которая первоначально стимулировала образование новых сосудов. Аномальные кровеносные сосуды можно восстановить до нормального состояния. Кроме того, можно доставлять ангиостатические белки, такие как Т2-TrpRS, в области паталогического ангиогенеза, используя трансфицированные композиции Lin- HSC и вызванную лазером активацию астроцитов. Так как лазерную фотокоагуляцию обычно используют в клинической офтальмологии, такой подход имеет применение в случае многих болезней сетчатки. Хотя такие подходы на оонове клеток используются в терапии рака, их применение при болезнях глаз более выгодно, так как внутриглазная инъекция делает возможной доставку большого числа клеток в место болезни.
Нейротрофическое и васкулотрофическое спасение с помощью Lin - HSC
Используют MACS для извлечения Lin- HSC из костного мозга усиленного белка зеленой флуоресценции (eGFP), С3Н (rd/rd), мышей FVB (rd/rd), так, как описано выше. Lin- HSC, содержащие ЕРС от таких мышей, инъецируют интравитреально в глаза Р6 мышей С3Н или FVB. Сетчатки собирают в различные моменты времени (1 месяц, 2 месяца и 6 месяцев) после инъекции. Сосудистую сеть анализируют с помощью сканирующего лазерного конфокального микроскопа после окрашивания антителами к CD31 и гистологии сетчатки после окрашивания ядер DAPI. Также используют микроматричный анализ экспрессии генов мРНК из сетчаток в различные моменты времени для идентификации генов, потенциально вовлеченных в действие.
Глаза мышей rd/rd в Р21 имеют глубокую дегенерацию как нейросенсорной сетчатки, так и сосудистой сети сетчатки. Глаза мышей rd/rd, обработанные Lin- HSC в Р6, сохраняют нормальную сосудистую сеть сетчатки на протяжении 6 месяцев; как глубокие, так и промежуточные слои существенно лучше при сравнении с контролем во все моменты времени (1М, 2М и 6М) (см. фиг.12). Кроме того, авторы наблюдали, что сетчатки, обработанные Lin- HSC, также более толстые (1М - в 1,2 раза, 2М - в 1,3 раза, 6М - в 1,4 раза) и имеют большее число клеток в наружном ядерном слое (1М - в 2,2 раза, 2М - в 3,7 раза, 6М - в 5,7 раза) относительно глаз, обработанных Lin+ HSC в качестве контроля. Крупномасштабный геномный анализ "спасенных" (например, Lin- HSC) сетчаток rd/rd при сравнении с контролем (необработанными или обработанными не Lin-) показывает значительную позитивную регуляцию генов, кодирующих sHPS (малые белки теплового шока) и специфические факторы роста, что коррелирует со спасением сосудов и нейронов, включая гены, кодирующие белки, перечисленные на фиг.20, части А и В.
Популяции Lin- HSC, полученные из костного мозга, существенно и воспроизводимо вызывают сохранение нормальной сосудистой сети и существенно усиливают фоторецепторные и другие нейронные слои мыши rd/rd. Такой эффект нейронного спасения коррелирует с существенной позитивной регуляцией малых белков теплового шока и факторов роста и обеспечивает понимание терапевтических подходов к дегенеративным расстройствам сетчатки, не поддающимся лечению в настоящее время.
Сетчатки мышей rd/rd обнаруживают глубокую сосудистую и нейронную дегенерацию.
Нормальное постнатальное развитие сосудов и нейронов сетчатки у мышей хорошо описано и аналогично изменениям, наблюдаемым в третьем триместре развития плода человека (Dorrell et al., 2002, Invest. Ophtalmol. Vis. Sci., 43: 3500-3510). Мыши, гомозиготные к гену rd1, разделяют многие характеристики с дегенерацией сетчатки у человека (Frasson et al., 1999, Nat. Med., 5: 1183-1187) и обнаруживают быструю потерю фоторецептора (PR), сопровождаемую тяжелой атрофией сосудов в результате мутации гена, кодирующего PR-cGMP-фосфодиэстеразу (Bowes et al., 1990, Nature, 347: 677-680). Для того чтобы обследовать сосудистую сеть во время развития сетчатки и ее последующую дегенерацию, используют антитела против коллагена IV (CIV), белок внеклеточного матрикса (ЕСМ) зрелой сосудистой сети и CD31 (PECAM-1) - маркер эндотелиальных клеток (фиг.15). Сетчатки rd1/rd1 (C3H/HeJ) развиваются нормально до приблизительно 8 дня после рождения (Р), когда начинается дегенерация содержащего фоторецепторы наружного ядерного слоя (ONL). ONL быстро дегенерирует, и клетки гибнут путем апоптоза, так что на Р20 сохраняется только один слой ядер. Двойное окрашивание тотальных препаратов сетчаток антителами как к CIV, так и к CD31 показывает детали дегенерации сосудов у мышей rd1/rd1, подобные описанным другими авторами (Blahks et al., 1986, J. Comp. Neurol., 254: 543-553). Оказывается, что первичные и глубокие сосудистые слои развиваются нормально до Р12, после чего происходит быстрая потеря эндотелиальных клеток, о чем свидетельствует отсутствие окрашивания CD31. CD31-Позитивные эндотелиальные клетки присутствуют при нормальном распределении до Р12, но после этого быстро исчезают. Интересно, что CIV-позитивное окрашивание присутствует во все моменты времени, когда проводят проверку, причем предполагается, что сосуды и ассоциированные ЕСМ формируются нормально, но после Р13 остается только матрикс, после чего CD31-позитивные клетки не обнаруживают (фиг.15, средний ряд). Промежуточное сосудистое сплетение также дегенерирует после Р21, но процесс происходит медленнее, чем наблюдаемый в глубоком сплетении (фиг.15, верхний ряд). Ретинальные сосудистые и нейронные слои здоровой мыши показаны, для сравнения, для мыши rd1/rd1 (правые ряды, фиг.15).
Нейрозащитное действие Lin - HSC, полученных из костного мозга, на мышей rd1/rd1
Инъецированные интравитреально Lin- HSC включаются в эндогенную сосудистую сеть сетчатки во все три сосудистые сплетения и предохраняют сосуды от дегенерации. Представляет интерес то, что инъецированные клетки фактически никогда не обнаруживают в наружном ядерном слое. Такие клетки или включаются в формирующиеся сосуды сетчатки или обнаруживаются в непосредственной близости к таким сосудам. Мышиные Lin- HSC (от C3H/HeJ) инъецируют интравитреально в глаза мышей C3H/HeJ (rd1/rd1) в Р6 непосредственно перед началом дегенерации. В Р30 глаза, получившие инъекцию контрольных клеток (CD31-), обнаруживают типичный фенотип rd1/rd1, т.е. почти полную дегенерацию глубокого сосудистого сплетения, и ONL наблюдают в каждой проверенной сетчатке. Глаза, получившие инъекцию Lin- HSC, сохраняют кажущиеся нормальными промежуточные и глубокие сплетения. Неожиданно значительно больше клеток наблюдают в межъядерном слое (INL) и ONL глаз, инъецированных Lin- HSC, чем глаз, инъецированных контрольными клетками (фиг.16 (А)). Такое спасающее действие Lin- HSC можно наблюдать через 2 месяца (фиг.16 (В)) и на протяжении 6 месяцев после инъекции (фиг.16 (С)). Различия в сосудистой сети промежуточного и глубокого сплетений глаз, инъецированных Lin- HSC, а также содержащих нейроны INL и ONL, существенны во все моменты времени измерения, когда сравнивают спасаемые и неспасаемые глаза (фиг.16 (В и С)). Такой эффект определяют количественно, измеряя общую длину сосудистой сети (фиг.16 (D)) и подсчитывая число ядер DAPI-позитивных клеток, обнаруживаемых в ONL (фиг.16 (Е)). Подобный линейно-регрессионный анализ применяют к результатам во все моменты времени.
Для глаз, инъецированных Lin- HSC, наблюдают статистически значимую корреляцию между спасением сосудов и спасением нейронов (например, толщиной ONL) в Р30 (p<0,024) и Р60 (p<0,034) (фиг.16 (F)). Корреляция остается высокой, хотя статистически незначимой (p<0,14) в Р180, когда сравнивают сетчатки глаз, инъецированных Lin- HSC и контрольными клетками (фиг.16 (F)). Напротив, сетчатки глаз, инъецированных контрольными клетками, не показывают значимой корреляции между сохранением сосудистой сети и ONL в любой момент времени (фиг.16 (F)). Такие результаты показывают, что интравитреальная инъекция Lin- HSC приводит к сопутствующему спасению сосудов и нейронов в сетчатках мышей rd1/rd1. Инъецированные клетки не обнаруживают в ONL или каком-либо другом месте, кроме как в пределах или непосредственной близости к кровеносным сосудам сетчатки.
Функциональное спасение сетчаток rd/rd, инъецированных Lin - HSC
Получают элетроретинограммы (ERG) мышей через 2 месяца после инъекции контрольных клеток или мышиных Lin- HSC (фиг.17). Осуществляют иммуногистохимический и микроскопический анализ каждого глаза после записи ERG для того, чтобы подтвердить, что имеет место спасение сосудов и нейронов. Характерные записи ERG для обработанных спасенных и контрольных неспасенных глаз показывают, что в спасенных глазах субтрактивный сигнал в цифровой форме (для обработанных глаз минус для необработанных) дает четко обнаруживаемый сигнал с амплитудой порядка 8-10 микровольт (фиг.17). Ясно, что сигналы от обоих глаз весьма аномальные. Однако, в случае глаз, обработанных Lin- HSC, можно записать соответствующие и детектируемые ERG. Во всех случаях ERG контрольных глаз не являются детектируемыми. Хотя амплитуды сигналов в спасенных глазах существенно ниже нормальных, сигналы наблюдают соответственно всякий раз, когда имеется гистологическое спасение, и их величина порядка величин, сообщенных в других исследованиях спасения на основе генов. В целом такие результаты показывают некоторую степень функционального спасения в случае глаз, обработанных Lin- HSC.
Спасенные типы клеток сетчатки rd/rd представляют собой, преимущественно, колбочки
Спасенные и неспасенные сетчатки анализируют иммуногистохимически с антителами, специфическими для опсина палочек и колбочек. Те же глаза, которые использовали для записей ERG, представленных на фиг.17, анализируют на опсин палочек и колбочек. В сетчатках мышей дикого типа менее примерно 5% имеющихся фоторецепторов представляют собой колбочки (Soucy et al., 1998, Neuron, 21: 481-493), и картины иммуногистохимического окрашивания, наблюдаемые с красным/зеленым опсином колбочек, как видно на фиг.25 (А), или красным родопсином, как видно на фиг.25 (В), соответствуют такому проценту колбочковидных зрительных клеток. Когда сетчатки дикого типа окрашивают преиммунным IgG, не наблюдают окрашивания где-либо в нейросенсорных сетчатках иного, чем аутофлуоресценция кровеносных сосудов (фиг.25 (С)). Через два месяца после рождения сетчатки неинъецированных мышей rd/rd имеют по существу атрофический наружный ядерный слой, который не показывает какого-либо окрашивания с антителами к красному-зеленому опсину колбочек (фиг.25 (D)) или родопсину (фиг.25 (G)). Глаза, инъецированные контролем CD31- HSC, также не окрашиваются позитивно на присутствие опсина ни колбочек (фиг.25 (Е)), ни палочек (фиг.25 (Н)). Напротив, контралатеральные глаза, инъецированные Lin- HSC, имеют от примерно 3 до примерно 8 рядов ядер в сохранившемся наружном ядерном слое; большинство таких клеток являются позитивными на опсин колбочек (фиг.25 (F)) с приблизительно 1-3% клеток, позитивных на опсин палочек (фиг.25 (I)). Примечательно, что такая картина, почти обратная той, которую обычно наблюдают в нормальной сетчатке мыши, где преобладают палочки. Такие результаты показывают, что инъекция Lin- HSC предохраняет колбочки на длительное время во время, когда они обычно дегенерируют.
Lin - HSC, полученные из костного мозга человека (hBM), также спасают дегенерирующие сетчатки.
Lin- HSC, выделенные из костного мозга человека, ведут себя подобно мышиным Lin- HSC. Костный мозг берут у людей-доноров и извлекают Lin- HSC, получая популяцию человеческих Lin- HSC (hLin- HSC). Такие клетки метят ex-vivo флуоресцентным красителем и инъецируют в глаза мышей C3SnSmn.CB17-Prkdc SCID. Инъецированные hLin- HSC мигрируют к и нацеливаются на участки ретинального ангиогенеза идентично тому, что наблюдают, когда инъецируют мышиные Lin- HSC (фиг.18 (А)). Кроме нацеливания на сосудистую сеть, человеческие Lin- HSC также обеспечивают сильный эффект спасения как сосудистых, так и нейронных слоев у мышей rd1/rd1 (фиг.18 (В и С)). Такое наблюдение подтверждает присутствие клеток костного мозга, которые избрали мишенью сосудистую сеть сетчатки и могут предотвращать дегенерацию сетчатки.
Lin - HSC оказывают васкуло- и нейротрофическое действие на мышей rd10/rd10
Хотя мыши rd1/rd1 являются наиболее широко используемой и наилучшим образом охарактеризованной моделью дегенерации сетчатки (Chang et al., 2002, Vision Res., 42: 517-525), дегенерация происходит очень быстро, и с этой точки зрения отличается от обычного более медленного течения, которое наблюдают при болезни человека. При такой особенности дегенерация фоторецепторных клеток начинается примерно в Р8 - время, когда сосудистая сеть сетчатки еще быстро разрастается (фиг.15). Последующая дегенерация глубокой сосудистой сети сетчатки происходит даже в то время, когда промежуточное сплетение еще формируется, и таким образом сетчатки мышей rd1/rd1 никогда полностью не развиваются, в отличие от того, что наблюдают у большинства людей с таким заболеванием. Мышиную модель rd10, которая имеет более медленное течение дегенерации и имеет более близкое сходство с дегенеративным состоянием сетчатки человека, используют для исследования спасения сосудов, опосредуемого Lin- HSC. У мыши rd10 дегенерация фоторецепторных клеток начинается примерно в Р21 и несколько позже начинается дегенерация сосудов.
Так как нормальное нейросенсорное развитие сетчатки в основном завершается в Р21, наблюдают, что дегенерация начинается после того, как завершилась дифференциация сетчатки, и такой путь более аналогичен дегенерациям сетчатки человека, чем у мышиной модели rd1/rd1. Lin- HSC или контрольные клетки от мышей rd10 инъецируют в глаза в Р6 и оценивают сетчатки в различные моменты времени. В Р21 сетчатки глаз, инъецированных как Lin- HSC, так и контрольными клетками, оказываются нормальными с завершенным развитием всех слоев сосудов и нормальным развитием INL и ONL (фиг.10 (D и Н)). Приблизительно в Р21 начинается дегенерация сетчатки и развивается с возрастом. В Р30 сетчатки, инъецированные контрольными клетками, показывают тяжелую сосудистую и нейронную дегенерацию (фиг.18 (I)), в то время как сетчатки, инъецированные Lin- HSC, сохраняют почти нормальные слои сосудов и фоторецепторные клетки (фиг.18 (Е)). Различие между спасенными и неспасенными глазами более выражено в более поздние моменты времени (ср. фиг.18 (F и G) - фиг.18 (J и K)). В глазах, обработанных контролем, развитие дегенерации сосудов очень четко наблюдают с помощью иммуногистохимического окрашивания CD31 и коллагеном IV (фиг.18 (I-K)). Глаза, обработанные контролем, почти полностью негативны в отношении CD31, в то время как сохраняются явными "следы" сосудов, позитивных к коллагену IV, что указывает, что происходит регрессия сосудов, а не незавершенное образование сосудов. Напротив, глаза, обработанные Lin- HSC, имеют как CD31-, так и коллаген IV-позитивные сосуды, которые кажутся весьма схожими с нормальными глазами дикого типа (сравните фиг.18 (F и I)).
Анализ экспрессии генов сетчаток мыши rd/rd после обработки Lin - HSC
Крупномасштабные геномные (микроматричные) анализы используют для анализа спасенных и неспасенных сетчаток для идентификации предполагаемых медиаторов нейротрофического спасения. Экспрессию генов в сетчатках мышей rd1/rd1, обработанных Lin- HSC, сравнивают с экспрессией в неинъецированных сетчатках, а также в сетчатках, инъецированных контрольными клетками (CD31-). Такое сравнение осуществляют каждый раз при трехкратном повторе. Для того чтобы такое сравнение рассматривать, требуется, чтобы уровни экспрессии генов были, по меньшей мере, в 2 раза выше, чем фоновые уровни при всех трех повторах. Гены, которые позитивно регулируются в сетчатках мышей rd/rd, защищенных Lin- HSC, в 3 раза чаще в сравнении с сетчатками, инъецированными контрольными клетками и неинъецированными, показаны на фиг.20, части А и В. Вычисляют коэффициент уровней дисперсии (COV) для экспрессированных генов путем деления стандартного отклонения на средний уровень экспрессии каждой реплики кРНК. Кроме того, вычисляют корреляцию между уровнями экспрессии и дисперсией шума, коррелируя среднее и стандартное отклонение (SD). Получают корреляцию между уровнем экспрессии гена и стандартным отклонением для каждого гена, создавая возможность определения фоновых уровней и границ достоверных уровней экспрессии. В целом результаты хорошо вписываются в приемлемые пределы (Tu et al., 2002, Proc. Natl. Acad. Sci. USA, 99:14031-14036). Гены, которые отдельно обсуждаются ниже, показывают уровни экспрессии выше таких критических уровней экспрессии. Также определяют парные величины "t-критерия" для обсуждаемых генов. В каждом случае величины р являются приемлемыми (около или ниже 0,05), что показывает, что существует подобие между повторами и вероятные значимые различия между различными испытываемыми группами. Многие из генов, нерегулируемых позитивно в значимой степени, включая MAD и Ying Yang-1 (YY-1) (Austen et al., 1997, Curr. Top. Microbiol. Immunol., 224: 123-120), кодируют белки с функциями, включающими защиту клеток от апоптоза. Ряд генов кристаллина, имеющих гомологию последовательностей и схожие функции с известными белками теплового шока, включая защиту клеток от стресса, также позитивно регулируются обработкой Lin- HSC. Экспрессия α-кристаллина согласно иммуногистохимическому анализу локализуется на ONL (фиг.19). Фиг.19 показывает, что кристаллин αA позитивно регулируется в спасенных клетках наружного ядерного слоя после обработки Lin- HSC, но не в контралатеральных глазах, обработанных контрольными клетками. Левое изображение показывает окрашивание IgG (контроль) в спасенных сетчатках. Среднее изображение показывает кристаллин αA в спасенной сетчатке. Правое изображение показывает кристаллин αA в неспасенной сетчатке.
Матричную РНК из сетчаток мышей rd1/rd1, спасенных человеческими Lin- HSC, гибридизируют со специфическими для человека микроматричными чипами Affymetrix U133F. После анализа в строгих условиях обнаруживают ряд генов, экспрессия мРНК которых специфична для человека, выше фона и существенно выше в сетчатках, спасенных человеческими Lin- HSC, по сравнению с сетчатками, спасенными мышиными Lin- HSC, и неспасенными сетчатками, инъецированными человеческими контрольными клетками (фиг.20, часть С). Фактор клеточной адгезии CD6, экспрессированный на поверхности сперматогоний и вновь дифференцированных CD34+ гемопоэтических стволовых клеток, и интерферон-альфа 13 - другой ген, экспрессируемый стволовыми клетками, обнаружены оба методом микроматричной биоинформатики (microarray bioinformatics technique), что подтверждает протокол оценок. Кроме того, в образцах мышиных сетчаток, спасенных человеческими Lin- HSC, некоторые факторы роста и нейротрофические факторы экспрессируются выше уровня фона (фиг.20, часть D).
Маркеры для линиекоммитированных гемопоэтических клеток используют для негативного отбора популяции полученных из костного мозга Lin- HSC, содержащих ЕРС. Хотя субпопуляция полученных из костного мозга Lin- HSC, которые могут служить в качестве ЕРС, не характеризуется обычно используемыми маркерами клеточной поверхности, поведение таких клеток при развитии или повреждении сосудистой сети клетчатки полностью отличается от поведения, наблюдаемого для Lin+ или популяций зрелых эндотелиальных клеток. Такие клетки селективно нацеливаются на участки ретинального ангиогенеза и принимают участие в формировании открытых кровеносных сосудов.
Наследственные дегенеративные болезни клетчатки часто сопровождаются потерей ретинальной сосудистой сети. Эффективное лечение таких заболеваний требует восстановления функции, а также сохранения сложной формы ткани. Хотя в некоторых последних работах исследуют применение основанной на клетках доставки трофических факторов или самих клеток, может потребоваться некоторое сочетание того и другого. Например, применение терапии с факторами роста для лечения дегенеративной болезни клетчатки приводит к позитивно регулируемому избыточному росту кровеносных сосудов, приводящему к серьезному нарушению нормальной формы ткани сетчатки. Применение нейронных или ретинальных стволовых клеток для лечения дегенеративной болезни клетчатки может восстановить нейронную функцию, но также будет необходима функциональная сосудистая сеть для сохранения функциональной целостности сетчатки. Включение клеток из популяции Lin- HSC в сосуды сетчатки мышей rd/rd стабилизирует дегенеративную сосудистую сеть без разрушения структуры сетчатки. Когда клетки инъецируют мышам rd/rd в Р15, также наблюдают эффект спасения. Так как сосудистая дегенерация начинается у мышей rd/rd в Р16, такое наблюдение расширяет терапевтическое окно для эффективного лечения Lin- HSC. Нейроны и фоторецепторы сетчатки сохраняются и сохраняется зрительная функция глаз, инъецированных клетками Lin- HSC.
Полученные из костного мозга зрелые Lin- HSC вызывают основательное васкуло- и нейротрофическое действие, когда инъецируются интравитреально мышам с дегенеративной болезнью сетчатки. Такой эффект спасения сохраняется до 6 месяцев после лечения и наибольший эффект имеет место, когда Lin- HSC инъецируют до наступления полной дегенерации сетчатки (до 16 дней после рождения у мышей, у которых полная дегенерация сетчатки обычно обнаруживается в пределах 30 дней после рождения). Такое спасение наблюдают на двух мышиных моделях дегенерации сетчатки и, что примечательно, его осуществляют полученными из костного мозга человека зрелыми Lin- HSC, когда реципиентом является грызун с иммунодефицитом с дегенерацией сетчатки (например, мышь SCID), или когда донором является мышь с дегенерацией сетчатки. Хотя в некоторых последних сообщениях описывается частичное фенотипическое спасение у мышей или собак с дегенерацией сетчатки после спасения генов на основе вирусов геном дикого типа (Ali et al., 2000. Nat. Genet., 25: 306-310; Takahashi et al., 1999, J. Virol., 73: 7812-7816; Acland et al., 2001, Nat. Genet., 28: 92-95), настоящее изобретение является первым общим лечебным действием, имеющим в основе клетки, достигнутым путем спасения сосудов. Таким образом, возможное применение такого подхода при лечении группы заболеваний (например, пигментного ретинита) с более 100 известными ассоциированными мутациями является более практичным, чем создание способов лечения отдельными генами для лечения каждой известной мутации.
Точная молекулярная основа нейротрофического эффекта спасения остается неизвестной, но такой эффект наблюдается только тогда, когда имеются сопутствующие стабилизация/спасение сосудов. Присутствия инъецированных стволовых клеток самих по себе недостаточно для генерации нейротрофического спасения, и отчетливое отсутствие происходящих из стволовых клеток нейронов в наружном ядерном слое исключает возможность того, что инъецированные клетки трансформируются в фоторецепторы. Результаты, полученные микроматричным анализом экспрессии генов, показывают существенную позитивную регуляцию генов, антиапоптозное действие которых известно. Так как наибольшая гибель нейронов, наблюдаемая при дегенерациях сетчатки, является апоптозом, такая защита может представлять собой большое лечебное благоприятное воздействие для продления жизни фоторецепторов и других нейронов, критичных для зрительной функции при таких заболеваниях. С-myc является фактором транскрипции, который принимает участие в апоптозе путем позитивной регуляции различных конечных вызывающих апоптоз факторов. Экспрессия c-myc повышается в 4,5 раза у мышей rd/rd по сравнению с диким типом, что указывает на возможное участие в дегенерации фоторецепторов, наблюдаемой у мышей rd1/rd1. Известно, что оба гена Mad1 и YY-1, которые в значительной мере позитивно регулируются в защищенных Lin- HSC сетчатках (фиг.20, часть А), подавляют активность c-myc, ингибируя таким образом вызванный c-myc апоптоз. Также показано, что сверхэкспрессия Mad1 подавляет вызванную Fas активацию каспазы-8 - другого критичного компонента пути апоптоза. Позитивная регуляция таких двух молекул может играть некую роль в защите сетчатки от сосудистой и нейронной дегенерации путем предотвращения инициации апоптоза, который обычно ведет к дегенерации у мышей rd/rd.
Другой набор генов, которые в значительной мере позитивно регулируются в защищенных Lin- HSC сетчатках, включает членов семейства кристаллинов (фиг.20, часть В). Подобно белкам теплового шока и другим белкам, вызванным стрессом, кристаллины могут активироваться ретинальным стрессом и обеспечивать защитное действие против апоптоза. Аномально низкая экспрессия кристаллина αA коррелирует с потерей фоторецепторов на крысиной модели дистрофии сетчатки, и современный анализ белков (proteomic analysis) сетчатки у мышей rd/rd показывает индукцию позитивной регуляции кристаллина в ответ на дегенерацию сетчатки. На основании полученных с микроматрицами результатов авторов по сетчаткам мышей rd/rd, спасенным ЕРС, оказывается, что позитивная регуляция кристаллинов играет ключевую роль в опосредуемой ЕРС защите нейронов сетчатки.
Гены, такие как c-myc, Mad1, YX-1 и кристаллины, вероятно, являются даун-медиаторами спасения нейронов. Нейротрофические агенты могут регулировать экспрессию апоптозных генов, хотя осуществленный авторами микроматричный анализ сетчаток, спасенных мышиными стволовыми клетками, не показывает индукции возросших уровней известных нейротрофических факторов. С другой стороны, анализ спасения, опосредованного стволовыми клетками, полученными из костного мозга человека, со специфическими для человека чипами показывает небольшое, но заметное усиление экспрессии генов нескольких факторов роста.
Позитивно регулируемые гены включают несколько членов семейства фактора роста фибробластов и отоферлин. Мутации в гене отоферлина ассоциируются с генетическими расстройствами, ведущими к глухоте вследствие невропатии органов слуха. Возможно, что продуцирование отоферлина инъецированными Lin- HSC также вносит вклад в предупреждение ретинальной невропатии. Исторически долгое время считалось, что изменения сосудов, наблюдаемые у пациентов и животных с дегенерацией сетчатки, являлись вторичными к пониженной потребности метаболизма, так как фоторецепторы погибают. Результаты данной заявки показывают, что, по меньшей мере, для мышей с наследственной дегенерацией сетчатки сохранение нормальной сосудистой сети также может способствовать сохранению компонентов наружного ядерного слоя. Последние имеющиеся в литературе сообщения могут поддержать концепцию, что тканеспецифическая сосудистая сеть обладает трофическим действием, которое выходит за пределы, ожидаемые от простого предоставления "питания" сосудам. Например, эндотелиальные клетки печени можно индуцировать к продуцированию, после ативации VEGFR1, факторов роста, критичных для регенерации и сохранения гепатоцитов вопреки повреждению печени (LeCouter et al., 2003, Science, 299: 890-893).
Подобные указывающие взаимодействия между сосудистыми эндотелиальными клетками и соседними паренхимными клетками печени, как сообщается, вовлекаются в органогенез печени также до формирования функциональных кровеносных сосудов. Эндогенная сосудистая сеть сетчатки у индивидуумов с дегенерацией сетчатки не может настолько способствовать спасению, но если такую сосудистую сеть поддерживают эндотелиальные предшественники, полученные из популяций гемопоэтических стволовых клеток костного мозга, они могут сделать сосудистую сеть более устойчивой к дегенерации и в то же время облегчить выживание нейронов сетчатки так же, как и сосудов. У людей с дегенерацией сетчатки отсрочка начала полной дегенерации сетчатки может предоставить дополнительные годы с сохранением зрения. Животные, обработанные Lin- HSC, в существенной степени сохраняют ERG, что может быть достаточным для поддержания зрительной функции.
Объективно широко признано, что может происходить существенная потеря фоторецепторов и других нейронов, в то время как зрительная функция все еще сохраняется. В некоторый момент граница пересекается, и зрение теряется. Так как почти все наследственные дегенерации сетчатки у людей начинаются рано, но постепенно, индивидуума с дегенерацией сетчатки можно идентифицировать и лечить интравитреально пересадкой аутологичных стволовых клеток костного мозга по изобретению для задержки дегенерации сетчатки и сопутствующей потери зрения. Для того чтобы усилить нацеливание и включение стволовых клеток по изобретению, желательно наличие активированных астроцитов (Otani et al., 2002, Nat. Med., 8: 1004-1010); это можно осуществить ранним лечением, когда имеется ассоциированный глиоз, или применением лазера для стимулированной локальной пролиферации активированных астроцитов. Необязательно, для усиления эффекта спасения можно использовать трансфекцию ex vivo стволовых клеток одним или несколькими нейротрофическими веществами перед внутриглазной инъекцией. Такой подход можно применить для лечения других зрительных нейронных дегенеративных расстройств, таких как глаукома, при которых имеется дегенерация ганглиозных клеток сетчатки.
Популяции Lin- HSC из зрелого костного мозга содержат популяцию ЕРС, которые могут промотировать ангиогенез путем нацеливания на реактивные астроциты и включаются в установившуюся матрицу без разрушения структуры сетчатки. Lin- HSC также обеспечивают длительный нейротрофический эффект спасения для глаз, страдающих от дегенерации сетчатки. Кроме того, композиции генетически модифицированных аутологичных Lin- HSC, содержащие ЕРС, можно трансплантировать в глаза с ишемическими или аномальными сосудами и можно устойчиво включить в новые сосуды и нейронные слои и непрерывно локально доставлять лечебные молекулы в течение длительных периодов времени. Такая локальная доставка генов, которые экспрессируют фармакологические средства в физиологически значительных дозах, представляет новую разновидность лечения болезней глаз, в настоящее время не поддающихся лечению.
Фоторецепторы в здоровой сетчатке мыши, например, представляют собой преимущественно палочки, но наружный ядерный слой, наблюдаемый после спасения Lin- HSC по изобретению, содержит преимущественно колбочки. Наибольшие наследственные дегенерации сетчатки у людей имеют место в результате первичных специфических дефектов палочек, и полагают, что потеря колбочек является вторичной к дисфункции палочек, которая, вероятно, соотносится с потерей какого-то трофического фактора, экспрессируемого палочками.
ПРИМЕРЫ
Пример 1. Изоляция и накапливание клеток; получение популяций А и В мышиных Lin - HSC
Общая процедура. Все оценки in vivo осуществляют согласно руководству NIH по содержанию и использованию лабораторных животных, и все процедуры оценок одобрены The Scripps Research Institute (TSRI, La Jolla, CA) Animal Care and Use Committee. Клетки костного мозга экстрагируют из костного мозга В6.129S7-Gtrosa26, Tie-2GFP, ACTbEGFP, FVB/NJ (мыши rd/rd) или взрослых мышей Balb/cBYJ (The Jackson Laboratory, ME).
Затем отделяют моноциты разделением в градиенте плотности с использованием градиента полисахарозы гистопак® (HISTOPAQUE®) (Sigma; St. Louis, MO) и метят биотинилированными линиеспецифическими панельными антителами (CD45, CD3, Ly-6G, CD11, TER-119, Pharmingen, San Diego, CA) для отбора Lin- в мышах. Клетки позитивной линии (Lin+) отделяют и удаляют из Lin- HSC с использованием магнитного сепаратора (сортер AUTOMACS™, Miltenyl Biotech, Auburn, CA). Далее, полученную популяцию Lin- HSC, содержащую эндотелиальные клетки-предшественники, характеризуют с использованием проточного цитометра FACS™ Calibur (Becton Dickinson, Franklin Lakes, NJ) с использованием следующих антител: РЕ-конъюгированных Sca-1, c-kit, KDR и CD31 (Pharmingen, San Diego, CA). Клетки костного мозга Tie-2GFP используют для характеризации Tie-2.
Для того чтобы собрать зрелые эндотелиальные мышиные клетки, из мыши ACTbEGFP хирургически извлекают мезентериальную ткань и помещают в коллагеназу (Worthington, Lakewood, NJ) для расщепления ткани с последующей фильтрацией с использованием фильтра 45 мкм. Фильтрат собирают и инкубируют со средами для выращивания эндотелия (Clonetics, San Diego, CA). Характеристики эндотелия подтверждают наблюдением морфологической "булыжной мостовой", окрашиванием mAb CD31 (Pharmingen) и проверкой культур на образование трубкообразных структур в матриксе матригеля™ (Beckton Dickinson, Franklin Lakes, NJ).
Популяция А мышиных Lin - HSC. Клетки костного мозга извлекают из мышей ACTbEGFP общей процедурой, описанной выше. Клетки Lin- HSC характеризуют проточной цитометрией FACS для маркеров поверхностных антигенов CD-31, c-kit, Sca-1, Flk-1 и Tie-2. Результаты приводятся на фиг.1 (с). Примерно 81% Lin- HSC показывают маркер CD-31, примерно 70,5% Lin- HSC показывают маркер c-kit, примерно 4% Lin- HSC показывают маркер Sca-1, примерно 2,2% Lin- HSC показывают маркер Flk-1, и примерно 0,91% Lin- HSC показывают маркер Tie-2. Напротив, Lin+ HSC, извлеченные из таких клеток костного мозга, имеют существенно отличающийся маркерный профиль (т.е. CD 31-37,4%; c-kit - 20%; Sca-1-2,8%; Flk-1-0,05%).
Популяция В мышиных Lin - HSC. Клетки костного мозга извлекают из мышей Balb/C, ACTbEGFP и С3Н общей процедурой, описанной выше. Клетки Lin- HSC анализируют на наличие маркеров клеточной поверхности (Sca-1, Flk-1/KDR, c-kit (CD117), CD34, CD31 и различные интегрины: α1, α2, α3, α4, α5, α6, αL, αM, αV, αX, αIIb, β1, β2, β3, β4, β5 и β7). Результаты приводятся в таблице 2.
Пример 2. Интравитреальное введение клеток мышиной модели
На веке мыши создают бороздку тонким лезвием для воздействия на глазное яблоко в Р2-Р6. Затем инъецируют интравитреально популяцию А HSC негативной линии по настоящему изобретению (приблизительно 105 клеток в примерно 0,5 мкл среды для культивирования клеток) с использованием шприца с иглой 33 (Hamilton, Reno, NV).
Пример 3. Трансфекция ЕРС
Мышиные Lin- HSC (популяция А) трансфицируют ДНК, кодирующей фрагмент Т2 TrpRS, также заключающий метку His6 (SEQ ID NO: 1, фиг.7), с использованием реагента для трансфекции FuGENE™6 (Roche, Indianapolis, IN) согласно протоколу изготовителей. Клетки Lin- HSC (примерно 106 клеток на мл) суспендируют в среде OPTI-MEM® (Invitrogen, Carlsbad, CA), содержащей фактор стволовых клеток (PeproTech, Rocky Hill, NJ). Затем добавляют смесь ДНК (примерно 1 мкг) и реагента FuGENE (примерно 3 мкл) и смеси инкубируют при примерно 37°С в течение примерно 18 часов. После инкубации клетки промывают и собирают. Степень трансфекции такой системы составляет приблизительно 17%, что подтверждает анализ FACS. Продуцирование Т2-TrpRS подтверждают вестерн-блоттингом. Аминокислотная последовательность Т2-TrpRS, меченного His6, приводится как SEQ ID NO: 2, фиг.8.
Пример 4. Иммуногистохимия и конфокальный анализ
Мышиные сетчатки собирают в различные моменты времени и препарируют как тотальные препараты или замороженные срезы. Для тотальных препаратов сетчатки фиксируют 4% параформальдегидом и блокируют в 50% фетальной сыворотке теленка (FBS) и 20% сыворотке здоровой козы в течение одного часа при комнатной температуре. Сетчатки обрабатывают первичными антителами и детектируют со вторичными антителами. Используемыми первичными антителами являются анти-коллаген IV (Chemicon, Temecula, CA), анти-β-gal (Promega, Madison, WI), анти-GFAP (Dako Cytomation, Carpenteria, CA), против α-актина гладких мышц (α-SMA, Dako Cytomation). Вторичными используемыми антителами являются антитела, конъюгированные с маркерами флуоресценции или Alexa 488 или 594 (Molecular Probes, Eugene, OR). Изображения получают с использованием конфокального микроскопа MRC 1024 (Bio-Rad, Hercules, CA). Трехмерные изображения создают с использованием программного обеспечения LASERSHARP® (Bio-Rad) для того, чтобы обследовать три различных слоя развития сосудов в сетчатке в тотальном препарате. Различия в интенсивности элементов изображения GFP между усиленными GFP (eGFP) мышами и мышами GFAP/wtGFP, установленные с помощью конфокального микроскопа, используют для создания трехмерного изображения.
Пример 5. Анализ для количественной оценки ретинального ангиогенеза in vivo на мышах
Для анализа Т2-TrpRS из трехмерных изображений мышиных сетчаток восстанавливают изображения первичного и глубокого сплетения. Первичное сплетение делят на две категории: нормальное развитие или прекратившееся развитие сосудов. Категории ингибирования развития глубоких сосудов интерпретируют на основе процента ингибирования сосудов, включая следующие критерии: полное ингибирование образования глубокого сплетения обозначают "полное", нормальное развитие сосудов (включая ингибирование менее чем на 25%) обозначают "нормальное", а остальное обозначают "частичное". В случае результатов спасения мышей rd/rd с использованием объектива 10х, внимание привлекают четыре отдельные области более глубокого сплетения в каждой сетчатке в тотальном препарате. Вычисляют общую длину сосудистой сети для каждого изображения, суммируют и сравнивают между группами. Для того чтобы получить точную информацию, Lin- HSC инъецируют в один глаз, а Lin+ HSC инъецируют в другой глаз одной и той же мыши. Неинъецированные контрольные сетчатки берут от того же помета.
Пример 6. Мышиные модели повреждения зрелой сетчатки
Создают модели с повреждением лазером или шрамом или с использованием диодного лазера (150 мВт, 1 секунда, 50 мм) или механически, прокалывая сетчатку иглой 27 размера. Через пять дней после повреждения инъецируют клетки с использованием интравитреального способа. Пятью днями позже собирают глаза от подопытных мышей.
Пример 7. Нейротрофическое спасение при дегенерации сетчатки
Гемопэтические стволовые клетки негативной линии (Lin- HSC) из костного мозга взрослых мышей обладают действием васкулотрофического и нейротрофического спасения на мышиной модели дегенерации сетчатки. Правые глаза 10-дневных мышей инъецируют интравитреально примерно 0,5 мкл, содержащими примерно 105 Lin- HSC по настоящему изобретению, и оценивают через 2 месяца на наличие ретинальной сосудистой сети и число ядер в нейронном слое. Левые глаза тех же мышей инъецируют примерно таким же числом Lin+ HSC как контроля и оценивают подобным образом. Как видно на фиг.9, в глазах, обработанных Lin- HSC, сосудистая сеть сетчатки выглядит почти нормально, внутренний ядерный слой почти нормальный, и наружный ядерный слой (ONL) имеет примерно 3 - примерно 4 слоя ядер. Напротив, контралатеральный глаз, обработанный Lin+ HSC, имеет заметно атрофированный средний слой сосудов сетчатки и полностью атрофированный наружный слой сосудов сетчатки; внутренний ядерный слой заметно атрофирован, и наружный ядерный слой полностью исчез. Это наглядно иллюстрируют мышь 3 и мышь 5. У мыши 1 не имеется эффекта спасения, и это справедливо для приблизительно 15% инъецированных мышей.
Когда зрительную функцию оценивают по электроретинограммам (ERG), восстановление позитивной ERG наблюдают, когда наблюдают спасение как сосудов, так и нейронов (мыши 3 и 5). Позитивную ERG не наблюдают, когда не происходит спасения ни сосудов, ни нейронов (мышь 1). Такая корреляция между спасением сосудов и нейронов глаз мышей rd/rd Lin- HSC по настоящему изобретению иллюстрируется графиком регрессионного анализа, приведенным на фиг.10. Корреляцию между восстановлением нейронов (ось Y) и сосудов (ось X) наблюдают для сосудистой сети промежуточного типа (r=0,45) и для глубокой сосудистой сети (r=0,67).
Фиг.11 показывает отсутствие какой-либо статистически значимой корреляции между спасением сосудов и нейронов Lin+ HSC. Спасение сосудов определяют количественно, и результаты представлены на фиг.12. Результаты для мышей через 1 месяц (1М), 2 месяца (2М) и 6 месяцев (6М) после инъекции, приведенные на фиг.12, показывают, что длина сосудистой сети существенно возрастает в случае глаз, обработанных Lin- HSC по настоящему изобретению (темные столбики), относительно длины сосудистой сети в случае необработанных глаз для одной и той же мыши (светлые столбики), в частности, через 1 месяц и 2 месяца после инъекции. Эффект нейротрофического спасения определяют количественно, подсчитывая ядра во внутреннем и наружном слоях примерно через два месяца после инъекции Lin- HSC или Lin+ HSC. Результаты представлены на фиг.13 и 14.
Пример 8. Популяция человеческих Lin - HSC
Клетки костного мозга берут у здоровых взрослых добровольцев общей процедурой, описанной выше. Затем отделяют моноциты разделением в градиенте плотности с использованием градиента полисахарозы гистопак® (Sigma; St. Louis, MO). Для того чтобы изолировать популяцию Lin- HSC от одноядерных клеток костного мозга человека, используют следующие биотинилированные линиеспецифические панельные антитела с магнитным сепаратором (сортер AUTOMACS™, Miltenyl Biotech, Auburn, CA): CD2, CD3, CD4, CD11a, Мас-1, CD14, CD16, CD19, CD33, CD38, CD45R, CD64, CD68, CD86, CD235a (Pharmingen).
Затем популяцию человеческих Lin- HSC разделяют на две субпопуляции на основании экспрессии CD133. Клетки метят биотинилированными антителами CD133 и разделяют на CD133-позитивную и CD133-негативную популяции.
Пример 9. Интравитреальное введение человеческих и мышиных клеток в мышиные модели дегенерации сетчатки
Линии мышей С3Н/HeJ, C3SnSmn.CB17-Prkdc SCID и rd10 используют в качестве моделей дегенерации сетчатки. Мыши С3Н/HeJ и C3SnSmn.CB17-Prkdc SCID являются гомозиготными для мутации 1 (rdl) дегенерации сетчатки - мутации, которая вызывает только начало тяжелой дегенерации сетчатки. Мутация локализуется в экзоне 7 гена Pde6b, кодирующего β-субзвено cGMP-фосфодиэстеразы палочковых фоторецепторов. Мутация в данном гене обнаружена у людей, больных аутосомным рециссивным пигментным ретинитом (RP). Мыши C3SnSmn.CB17-Prkdc SCID также являются гомозиготными для спонтанной мутации тяжелого комбинированного иммунодефицита (Prkdc SCID) и используются для экспериментов с переносом человеческих клеток. Дегенерация сетчатки у мышей rd10 вызывается мутацией в экзоне 13 гена Pde6b. Это также клинически релевантно модели RP с более поздним началом и более легкой дегенерацией сетчатки, чем у rd1/rd1. Все оценки осуществляют согласно руководству NIH по содержанию и использованию лабораторных животных, и все процедуры оценок одобрены The Scripps Research Institute Animal Care and Use Committee.
На веке мыши создают бороздку тонким лезвием для воздействия на глазное яблоко в Р2-Р6. Затем в мышиный глаз инъецируют интравитреально клетки HSC негативной линии для мышиной популяции А или человеческой популяции С (приблизительно 105 клеток в от примерно 0,5 мкл до примерно 1 мкл среды для культивирования клеток) с использованием шприца с иглой 33 (Hamilton, Reno, NV). Для того чтобы визуализировать человеческие инъецированные клетки, перед инъекцией клетки метят красителем (зеленый краситель для отслеживания клеток CMFDA, Molecular Probes).
Сетчатки собирают в различные моменты времени и фиксируют 4% параформальдегидом (PFA) и метанолом с последующей блокировкой в смеси 50% FBS/20% NGS в течение одного часа при комнатной температуре. Для того чтобы окрасить сосудистую сеть, сетчатки инкубируют с антителами анти-CD31 (Pharmingen) и антиколлаген IV (Chemicon), а затем вторичными антителами, конъюгированными с Alexa 488 или 594 (Molecular Probes, Eugene, OR). Сетчатки делают плоскими четырьмя релаксирующими разрезами и получают тотальный препарат. Изображения сосудистой сети в промежуточных или глубоких ретинальных сосудистых сплетениях (см. Dorrell et al., 2002, Invest. Ophthalmol. Vis. Sci., 43: 3500-3510) получают с использованием конфокального микроскопа Radiance MP2100 и программного обеспечения LASERSHARP® (Biorad, Hercules, California). Для количественной оценки сосудистой сети выбирают произвольно четыре независимых поля (900 мкм×900 мкм) из средней части промежуточного или глубокого сосудистого слоя и измеряют общую длину сосудистой сети с использованием анализирующего программного обеспечения LASERPRIX® (Biorad). Общие длины указанных четырех полей используют для дальнейшего анализа.
Сетчатки в плоских препаратах повторно заделывают для криостатических срезов. Сетчатки помещают в 4% PFA на ночь с последующей инкубацией с 20% сахарозы. Сетчатки заделывают в соединении с оптимальной температурой резания (ОСТ: Tissue-Tek; Sakura FineTech, Torrance, CA). Срезы из криостата (10 мкм) регидратируют в PBS, содержащем краситель для ядер DAPI (Sigma-Aldrich, St. Louis, Missouri). Изображения помеченных DAPI ядер трех различных участков (ширина 280 мкм, беспристрастный отбор образцов) в одном срезе, содержащие диск зрительного нерва и полную периферическую сетчатку, получают на конфокальном микроскопе. Число ядер, локализованных в ONL трех независимых полей в срезе, подсчитывают и суммируют для анализа. Осуществляют простой линейно-регриссионный анализ для проверки соотношения между длиной сосудистой сети в глубоком сплетении и числом клеток в ONL.
После темновой адаптации в течение ночи мышам дают наркоз интраперитонеальной инъекцией 15 мкг/г кетамина и 7 мкг/г ксилазина. Регистрируют электроретинограммы (ERG) с поверхности роговицы каждого глаза после расширения зрачка (1% сульфата атропина) с использованием роговичного электрода с золотой петлей вместе с электродом сравнения во рту и заземляющим электродом на хвосте. Стимуляцию проводят фотостимулятором Grass (PS33 Plus, Grass Instruments, Quincy, MA), прикрепленным к наружной стороне высокоотражающего купола Ганцфилда (Ganzfield). Регистрируют реакцию палочек на коротковолновые (Wratten 47A; λmax=470 нм) вспышки света в интервале интенсивностей до максимальной, которую можно получить с фотостимулятором (0,668 кд.с/м2). Ответные сигналы усиливают (усилитель СР511 АС, Grass Instruments), преобразуют в цифровую форму (PCI-1200, National Instruments, Austin, TX) и анализируют с помощью ЭВМ. Каждая мышь служит в качестве собственного внутреннего контроля с ERG, регистрируемыми как от обработанного, так и от необработанного глаза. Усредняют до 100 колебаний для самых слабых сигналов. Усредненные ответы от необработанного глаза вычитают в цифровой форме из ответов от обработанного глаза и эту разницу в сигнале используют для индексации функционального спасения.
Используют микроматричный анализ для оценки экспрессии ретинальных генов - мишеней для Lin- HSC. Мышей rd/rd, Р6, инъецируют или Lin- или CD31- HSC. Сетчатки таких мышей иссекают через 40 дней после инъекции в свободной от РНКазы среде (спасение сосудистой сети сетчатки и фоторецепторного слоя наблюдают в данный момент времени после инъекции). Один квадрант каждой сетчатки анализируют с помощью тотального препарата для того, чтобы убедиться, что достигается нормальное нацеливание HSC, а также защита сосудистой сети и нейронов. РНК из сетчаток после успешных инъекций очищают с использованием TRIzol (Life Technologies, Rockville, MD), протокола выделения РНК с фенолом/хлороформом. РНК гибридизируют с чипами Affymetrix Mu74Av2 и анализируют экспрессию генов с использованием программного обеспечения GENESPRING® (SiliconGenetics, Redwood City, CA). Очищенные человеческие или мышиные HSC инъецируют интравитреально мышам Р6. В Р45 иссекают сетчатки и собирают во фракции: 1) инъецированные человеческими HSC спасенные мышиные сетчатки, 2) инъецированные человеческими HSC неспасенные мышиные сетчатки и 3) инъецированные мышиными HSC спасенные мышиные сетчатки для очистки РНК и гибиридизации со специфическими для человека чипами Affymetrix. Используют программное обеспечение GENESPRING® для идентификации генов, которые экспрессированы выше уровня фона и с большей экспрессией в сетчатках, спасенных человеческими HSC. Затем анализируют по отдельности профили экспрессии с парой зондов для каждого из таких генов и сравнивают с моделью экспериментов с микроматрицами с нормальными человеческими U133A с использованием dChip для того, чтобы определить специфическую для человека гибридизацию и устранить ложные позитивы из-за перекрестновидовой гибридизации.
Фиг.21 иллюстрирует данные проточной цитометрии для сравнения экспрессии поверхностных антигенов CD31 и интегрина альфа-6 на популяциях CD133-позитивных (CD133+) и CD133-негативных (CD133-) человеческих Lin- HSC по настоящему изобретению. Левые части показывают графики рассеяния по проточной цитометрии. Центральная и правая части представляют собой гистограммы, показывающие уровень специфической экспрессии антител на клеточной популяции. Ось Y представляет число событий, и ось Х показывает интенсивность сигнала. Контурные гистограммы относятся к контрольным антителам изотипа IgG, показывающим уровень неспецифического фонового окрашивания. Заштрихованные гистограммы показывают уровень специфической экспрессии антител на клеточной популяции. Заштрихованные гистограммы, сдвинутые вправо от контурной (контроль) гистограммы, представляют повышенный сигнал флуоресценции и экспрессию антител выше фонового уровня. Сравнение положения пиков на заштрихованных гистограммах между двумя клеточными популяциями представляет различие в экспрессии белка на клетках. Например, CD31 экспрессируется выше фонового уровня как на CD133+, так и на CD133- клетках по изобретению; однако клеток, экспрессирующих более низкие уровни CD31, имеется больше в CD133+ популяции клеток, чем в популяции CD133-. Из таких данных очевидно, что экспрессия CD31 в двух популяциях различная и что экспрессия интегрина альфа-6 больше ограничивается для клеток в популяции Lin- и таким образом может служить в качестве маркера клеток с функцией васкуло- и нейротрофического спасения.
Когда CD133-позитивную и CD133-негативную субпопуляцию Lin- HSC инъецируют интравитреально в глаза новорожденных мышей SCID, наибольшую степень включения в развивающуюся сосудистую сеть наблюдают для CD133-негативной субпопуляции, которая экспрессирует поверхностные антигены как CD31, так и интегрин α6 (см. фиг.21, нижняя часть). Оказывается, что CD133-позитивная субпопуляция, которая не экспрессирует ни CD31, ни интегрин α6 (фиг.21, верхняя часть), нацеливается на места периферического управляемого ишемией образования новых сосудов, но не тогда, когда инъецирована в глаза, претерпевающие ангиогенез.
Спасенные и неспасенные сетчатки анализируют иммуногистохимически с антителами, специфичными для опсина палочек или колбочек. Те же глаза, которые использовали для записи ERG, представленных на фиг.17, анализируют на опсин палочек или колбочек. В сетчатках мышей дикого типа менее 5% имеющихся фоторецепторов представляют собой колбочки (Soucy et al., 1998, Neuron, 21: 481-493) и картины иммуногистохимического окрашивания, наблюдаемые с красным/зеленым опсином колбочек, как показано на фиг.25 (А), или родопсином палочек, как показано на фиг.21 (В), содержат такой процент клеток колбочек. Антитела, специфичные для родопсина палочек (rho4D2), поставлены Dr. Robert Molday из University of British Columbia, и используются так, как описано ранее (Hicks et al., 1986, Exp. Eye Res., 42: 55-71). Кроличьи антитела, специфичные для красного/зеленого опсина, закупают у Chemicon (AB5405) и используют согласно рекомендациям изготовителя.
Пример 10. Интавитреальное введение мышиных клеток мышиным моделям дегенерации сетчатки, вызванной кислородом
Новорожденных мышей дикого типа С57В16 подвергают гипероксии (75% кислорода) между постнатальными днями Р7-Р12 как модели вызванной кислородом дегенерации сетчатки (OIR). Фиг.22 иллюстрирует нормальное постнатальное развитие сосудистой сети у мышей С57В16 от Р0 до Р30. В Р10 можно наблюдать только прорастающие поверхностные сосуды вокруг диска зрительного нерва. В течение нескольких дней первичная поверхностная сеть расширяется до периферии, достигая дальней периферии в Р10. Между Р7 и Р12 развивается вторичное (глубокое) сплетение. В Р17 имеются расширенная поверхностная и глубокая сеть сосудов (фиг.22, вставки). В последующие дни происходит ремоделирование вместе с развитием третичного (промежуточного) слоя сосудов до тех пор, пока не достигается зрелая структура приблизительно в Р21.
Напротив, на модели OIR, описанной в данном описании, после воздействия 75% кислорода в Р7-Р12 нормальная последовательность событий серьезно нарушается (фиг.23). Популяции зрелых мышиных Lin- HSC по изобретению инъецируют интравитреально в Р3 в глаз мыши, который затем подвергают OIR, в другой глаз инъецируют PBS или CD31-негативные клетки в качестве контроля. Фиг.24 поясняет, что популяции Lin- HSC могут устранить дегенеративное действие высоких уровней кислорода на развивающуюся мышиную сетчатку. Полностью развитую поверхностную и глубокую ретинальную сосудистую сеть наблюдают в Р17 в обработанных глазах, в то время как контрольные глаза показывают большие области, лишенные сосудов, при отсутствии глубоких сосудов (фиг.24). Обследовали приблизительно 100 глаз мышей на модели OIR. Нормальная сосудистая сеть развивается в 58% глаз, обработанных популяциями Lin- HSC, в сравнении с 12% контрольных глаз, обработанных CD31- клетками, и 3% контрольных глаз, обработанных PBS.
Пример 11. Выделение миелоидоподобных клеток костного мозга из мышиного костного мозга отбором с CD44
Клетки костного мозга извлекают из взрослой мыши (The Jackson Laboratory, ME). Цельный костный мозг обрабатывают мышиными антителами к CD44 и используют проточную цитометрию для выделения из костного мозга клеток, экспрессирующих CD44. Клетки отделяют от антител и хранят в буферном растворе для дальнейшего применения. Также выделяют популяцию клеток, которые существенно не экспрессируют CD44 (CD44loBM).
Пример 12. Выделение миелоидоподобных клеток костного мозга из мышиного костного мозга отбором с CD44
Клетки костного мозга также позитивно выделяют с использованием антител к CD11b вместо CD44, так, как описано в примере 11. Изолируют популяцию миелоидоподобных клеток костного мозга, которые являются CD44hi и CD11b+, которые имеют характеристики активности, схожие с популяцией CD44hi, выделенной в примере 11 с использованием CD44. Также выделяют популяцию CD44hi CD11b-, которая, как находят, является неактивной.
Пример 13. Характеризация популяций клеток MLBM
Хотя роль CD44 в данном контексте не является ясной, возможно, что данный рецептор опосредует выживание клеток, миграцию клеток и/или дифференцировку клеток в богатом гиалуроновой кислотой стекловидном теле после инъекции клеток в глаз. В нефракционированном мышином костном мозгу имеются различные популяции клеток CD44hi (т.е. MLBM) и CD44lo. Популяция клеток MLBM представляет 76% популяции Lin-, используемой в предыдущих примерах, в то время как только примерно 37% и 4%, соответственно, клеточные популяции Lin+ и CD31-/CD34-/CD11b- из CD44, экспрессированного костным мозгом (фиг.26). Соответственно существует превосходная корреляция между экспрессией CD44 и васкулотрофической и нейротрофической активностями, наблюдаемыми в указанных трех популяциях, т.е. Lin- клетки являются наиболее эффективными, в то время как CD31-/CD34-/CD11b- клетки являются, сообразно, наименее эффективными. С использованием панели линиеспецифических антител определяют, что большинство CD44hi клеток имеют строго миелоидные характеристики (фиг.27). Подобным образом почти все CD44hi клетки костного мозга также являются CD11b+ (фиг.27).
MLBM, позитивно отобранные в примере 12 с использованием антител к CD11b (CD44hi CD11b+), дают результаты по активности, схожие с результатами, полученными с MLBM, изолированными с использованием отбора с антителами к CD44, в экспериментах с нацеливанием на сосуды.
Характеристики антигенов клеточной поверхности популяции клеток MLBM из примера 12 и клеток CD44hi CD11b+, выделенных в примере 12, приводятся ниже в таблице 3. В таблице 3 большее число знаков (+) показывает относительно более высокую экспрессию антигена. Знак минус (-) показывает, что экспрессия не обнаруживается.
Пример 14. Васкулотрофическое и нейротрофическое действие популяции клеток MLBM
Популяция клеток MLBM из примера 11 сохраняет свойства Lin- клеток в отношении нацеливания на сосуды и васкуло- и нейротрофического действия, в то время как клетки CD44loBM показывают незначительную активность или отсутствие активности. Активность нацеливания на сосуды демонструется инъекцией клеток из популяции клеток GFP+ MLBM интравитреально мышам в постнатальный день 7 (Р7) и анализом сетчаток в Р14. После мечения кровеносных сосудов изолектином GS наблюдают, что клетки GFP+ направляются к сосудистой сети сетчатки и предпринимают периваскулярную локализацию без видимого включения. Такие события являются обычными при использовании MLBM, но нечастыми или отсутствуют в глазах, обработанных CD44loBM (фиг.28).
Васкуло- и нейротрофическую активность популяции клеток MLBM из примера 11 оценивают с использованием мышиной модели дегенерации сетчатки, как описано выше для Lin- HSC. Мыши rd1/rd1 показывают характерные признаки дегенеративной болезни сетчатки, включая гибель фоторецепторов и атрофию глубокой ретинальной сосудистой сети. Как описано выше, клетки костного мозга Lin- HSC сохраняют глубокую ретинальную сосудистую сеть и частично спасенные фоторецепторы. Популяция клеток MLBM по настоящему изобретению выполняет ту же функцию (фиг.29).
Модель вызванной кислородом ретинопатии разделяет признаки ретинопатии преждевременности. Патология, ассоциированная с такой моделью, существенно уменьшается, когда глаза обрабатывают клетками из популяции клеток MLBM. Действие клеток из популяции клеток MLBM на данной модели схоже с действием, наблюдаемым с использованием Lin- HSC, описанным выше. Глаза, обработанные клетками из популяции клеток MLBM, показывают существенное уменьшение по двум параметрам, используемым для количественной оценки степени патологии на данной модели: площади облитерации сосудов и площади пучков вновь образованных сосудов. Напротив, глаза, обработанные клетками CD44loBM, не показывают улучшения по сравнению с глазами, обработанными контролями-носителями (фиг.30).
Кроме нацеливания на сосудистую сеть клетки из популяции клеток MLBM дифференцируют в макрофагоподобные клетки (F4/80+), проникают в сетчатку и занимают положение непосредственно против ретинального пигментного эпителия (RPE). Такая локализация облегчает наблюдаемое действие клеток из популяции клеток MLBM по спасению сосудов и фоторецепторов. Более того, только в месте вблизи RPE клетки из популяции клеток MLBM продуцируют васкулярный эндотелиальный фактор роста (VEGF), что демонстрируется инъекцией клеток из популяции клеток MLBM, полученных от мышей VEGF-GFP, при которой белок зеленой флуоресценции (GFP) экспрессируется после активации гена VEGF (фиг.31). Таким образом, оказывается, что клетки из популяции клеток MLBM находятся в состоянии, "активированном" VEGF. Оказывается, что введенные клетки из популяции клеток MLBM восстанавливают эндогенные клетки того же типа, так как в области RPE наблюдают как GFP+ (введенные), так и GFP- (эндогенные) клетки. Такую локализацию наблюдают у мышей дикого типа во время нормального развития сосудистой сети сетчатки, в спасенных сетчатках у мыши rd1/rd1 и на модели ретинопатии, вызванной кислородом.
Подобные результаты по нацеливанию на сосуды получают для популяции клеток MLBM из примера 12. Фиг.32 показывает, что в Р20 клетки CD44hi CD11b+ из примера 12 (зеленый цвет) специфически нацеливаются на сосудистую сеть (красный цвет), когда инъецированы в Р2, подобно CD44-высокой популяции из примера 11. Фиг.33 показывает, что клетки CD44lo CD11b- из примера 12 не нацеливаются специфически на сосудистую сеть.
Популяция клеток MLBM по настоящему изобретению обеспечивает эффективное и разностороннее лечение глазных болезней. Клетки легко извлекаются из аутологичного костного мозга, причем таким образом минимизируется возможная иммуногенность, часто наблюдаемая при терапиях на основе клеток. Кроме того, популяцию клеток MLBM по настоящему изобретению можно трансфицировать полезными генами для доставки функциональных генов в сетчатку.
Пример 15. Дополнительная характеризация субпопуляций клеток костного мозга
Как описано в предыдущих примерах, все эксперименты осуществляют согласно руководству NIH по содержанию и использованию лабораторных животных, и все экспериментальные процедуры одобрены the TSRI Animal Care and Use Committee. OIR вызывают у мышей С57В16 согласно протоколу, описанному выше. Мышат на 7 день после рождения и их матерей переносят из комнатной воздушной атмосферы в окружающую среду, содержащую 75% кислорода, на 5 суток, и затем возвращают в комнатную атмосферу. Уровни кислорода контролируют с использованием анализатора кислорода, одобренного FDA (AX-300, Teledyne Analytical Instruments, CA, USA). В таких условиях во время гипероксии в центральной сетчатке образуются большие гиповаскулярные участки, и после возвращения к нормоксии происходит аномальное образование новых сосудов, достигающее максимума в примерно Р17, и в конце концов, расщепление (фиг.37, части g-i; фиг.2, части а, с).
Получение клеток. Осуществляют экстракцию клеток мышиного костного мозга, по существу, следующим образом. Клетки костного мозга собирают из берцовых костей и большеберцовой кости мышей actGFP и обрабатывают с использованием двух различных способов. По первому способу одноядерные клетки отделяют в градиенте плотности с использованием FICO/LITE LM® (Atlanta Biologicals, Norcross, Georgia) и метят биотинилированными линеспецифическими антителами (CD45R/B220, CD3e, Ly-6G/C, CD11b, TER119, Pharmingen, San Diego, CA). За этим следует инкубация со стрептавидином или антибиотиновыми магнитными гранулами и сортировка с использованием системы сортировки клеток MACS (Miltenyl Biotech, Auburn, CA) для получения популяций Lin- HSC. По второму способу цельный костный мозг инкубируют с антителами, направленными против CD44, которые конъюгированы с флуоресцентной меткой. Затем используют клеточный сортер с возбуждением флуоресценции (FACS) для изоляции клеток CD44HI (т.е. популяции клеток MLBM, в которой большинство клеток экспрессирует CD44) и клеток CD44LO (т.е. популяции клеток, в которой меньшинство клеток экспрессирует CD44).
Характеризация клеток костного мозга. Дальнейший анализ субпопуляций клеток, полученных вышеуказанными способами, выполняют с использованием двух процедур: (1) проточной цитометрии с использованием двухцветной маркировки в сочетании с антителами против различных линий и поверхностными маркерами клеток-предшественников, включая CD11a, CD11b, Ly6G/C, CD43, F4/80, CD14, cKit, CD34, α6-интегрина и CD115 (все от Pharmingen, San Diego, CA); и (2) анализа экспрессии генов с использованием чипов AFFYMETRIX® Mu430 (Affymetrix, Santa Clara, CA) с использованием стандартных способов, известных в технике. Экспрессию генов анализируют с использованием программного обеспечения GENESPRING® (Agilent Technologies, Palo Alto, CA).
Интравитреальная инъекция. Делают бороздку на веке аккуратным надрезанием для воздействия на глазное яблоко мышей в Р2-Р7 (предгипероксия). В один глаз каждого животного инъецируют в стекловидное тело примерно 150000-250000 клеток, полученных из костного мозга, в 0,5 мкл носителя (PBS, содержащий 0,5% BSA и 2 мМ ЭДТК) c использованием шприца Гамильтона с иглой 33 (Hamilton, Reno, Nevada). В противоположный контрольный глаз инъецируют приблизительно равное число контрольных клеток или один носитель, и в некоторых случаях инъекцию не делают вовсе для того, чтобы наблюдать за естественным ходом болезни. В подмножестве экспериментов трансплантацию клеток осуществляют в более поздние сроки между Р9 и Р12.
Окрашивание сосудистой сети сетчатки. Сетчатки собирают в Р17 для получения изображения сосудистой сети и локализации и характеризации инъецированных клеток. В некоторых случаях животным дают наркоз и перед иссечением сетчаток делают интракардиальную инъекцию меченного флуоресцеином высокомолекулярного декстрана (FITC Dextran, Sigma) для визуализации открытых сосудов. В других случаях используют иммуногистологические методы для окрашивания кровеносных сосудов и экспрессирующих GFP клеток. Сетчатки фиксируют в 4% перфторуксусной кислоте (ПФК) и метаноле с последующей блокировкой в 20% FBS/20% NGS в течение одного часа при комнатной температуре. За этим для идентификации сосудов следует инкубация в течение ночи с изолектином GS-IB4, конъюгированным с ALEXA® 594 (Molecular Probes, Eugene, Oregon). Сетчатки делают плоскими четырьмя радиальными релаксирующими разрезами и получают тотальные препараты или заделывают в ОСТ и делают криосечения и получают криосрезы и перед заливкой осуществляют контрастирующее окрашивание DAPI.
Для того чтобы охарактеризовать трансплантированные клетки, используют иммуногистологические методы для идентификации следующих клеточных маркеров в подмножествах глаз: F4/80 (Caltag, Burlingame, CA), CD44, CD31 (Pharmingen, San Diego, CA) и NG2 (Chemicon, Temecula, CA). Все сетчатки при трехкратном повторе окрашивают лектином, анти-GFP и одним из вышеперечисленных маркеров.
Получение изображений и анализ изображений. Изображения сосудистой сети сетчатки получают с использованием лазерного сканирующего конфокального микроскопа RADIANCE® 2100MP (Biorad, Hercules, California). Количественное определение вазооблитерации и образования новых сосудов осуществляют следующим образом. Площадь сосудистой облитерации измеряют, тщательно очерчивая всю площадь, с использованием программного обеспечения PHOTOSHOP® (Abode) или VOLOCITY® (Improvision, Lexington, MA). Подобным образом вычисляют площадь преретинального образования новых сосудов ("пучков"), используя конфокальные изображения, фокусированные в преретинальной плоскости и выбирая пучки, основываясь на интенсивности элементов изображения (пучки мечены ярче, чем нормальная сосудистая сеть). Затем выбранные участки суммируют и получают общую площадь образования новых сосудов. Используют Т-критерий для статистического сравнения различных экспериментальных групп.
Трехмерные изображения сосудистой сети сетчатки и периваскулярных клеток костного мозга получают, собирая z-серии конфокальных изображений и преобразуя их в объемы с использованием программного обеспечения VOLOCITY®. Затем можно рассмотреть ретинальные сосуды в поперечном срезе и определить положение трансплантированных клеток костного мозга относительно просвета сосуда.
Развитие сосудистой сети сетчатки и мышиная модель ретинопатии, вызванной кислородом. Нормальное развитие сосудистой сети сетчатки у мышей после рождения, растущих в условиях нормоксии, показано на фиг.37, части а-f. В постнатальный день 2 (Р2) наблюдают только прорастающие поверхностные сосуды, занимающие одну плоскость вокруг диска зрительного нерва (фиг.37, части а, b). В течение следующей недели первичная поверхностная сеть расширяется до периферии, достигая дальней периферии приблизительно в Р12 (фиг.37, часть с). В промежутке Р7-Р12 развивается вторичное (глубокое) сплетение (фиг.37, часть d). В конце первого месяца происходит ремоделирование в полностью васкуляризованной сетчатке (фиг.37, часть е) вместе с развитием третичного (промежуточного) слоя сосудов и достигается зрелая структура (фиг.37, часть f).
Напротив, на модели OIR, воздействие 75% кислорода в Р7-Р12 серьезно нарушает нормальную последовательность событий: происходит заметная регрессия поверхностной сети сосудов, которые уже сформировались в центральной сетчатке, в особенности вдоль артерий (фиг.37, часть g (Р10) и части h, i (Р17)), и развитие глубокого сплетения существенно замедляется (фиг.37, части k, m, поперечные срезы сетчатки в Р17). Аномальный рост сосудов снова начинается только после возвращения в условия нормоксии в Р12. По существу, теперь имеются условия относительной гипоксии для сетчатки со значительной нехваткой сосудов. В Р17 некоторые глубокие сосуды можно идентифицировать на периферии, но аномальные преретинальные пучки новых сосудов, ассоциированные с прониканием интраваскулярного красителя, можно видеть в средней периферии на границе между гиповаскулярной центральной сетчаткой и периферией с более развитыми сосудами (фиг.37, часть h). В последующие дни поверхностные и глубокие сосуды постепенно развиваются в участках, лишенных сосудов, но пучки новых сосудов, выступающие над внутренней ограничивающей мембраной (ILM) сетчатки и входящие в стекловидное тело, часто существуют до Р21 или даже долее. В Р25-Р30 сосудистая сеть сетчатки ремоделируется и похожа на нормальную сосудистую сеть в такое время.
Инъекция гемопоэтических клеток-предшественников перед гипероксией промотирует восстановление сетчатки после вызванной кислородом вазооблитерации. Инъекция клеток Lin- HSC по изобретению в Р2-Р7 существенно изменяет способность ретинальной сосудистой сети восстанавливаться после воздействия гипероксии (фиг.37, части j, l, n, o, и фиг.38, части b, d, e, f, g). Инъекция одного носителя не вызывает таких изменений. В Р17 в более 50% случаев полностью развитая поверхностная и глубокая ретинальная сосудистая сеть видна в глазах, инъецированных Lin- HSC, в то время как контралатеральные контрольные глаза показывают большие участки, лишенные сосудов, и практически отсутствие глубоких сосудов (фиг.37, части l, n, ср. с частями h, i, k, m и частью о). В некоторых случаях, в особенности когда повреждение в контралатеральном глазу очень серьезное, к Р17 восстановление в глазу, инъецированном Lin- HSC, не завершается, но происходит существенно лучше в огромном большинстве случаев. Сравнение разных глаз одного и того же животного также подтверждает эффективность Lin- HSC, эффективно уравнивая большинство других генетических факторов и факторов окружающей среды.
Сосудистая облитерация была свойством, которому недостаточно уделяли внимания на этой модели, так как в большинстве исследований анализируется только образование преретинальных пучков новых сосудов в серийных срезах сетчатки. Сосудистую облитерацию и образование пучков можно оценивать в одной и той же сетчатке с использованием конфокальной микроскопии и цифрового анализа изображений (см., например, фиг.38, части а-d). Р17 выбирают главным моментом времени для анализа, поскольку образование пучков часто является максимальным в это время, в то время как существенная сосудистая облитерация еще имеется в контрольных глазах. С использованием нового способа комбинированного анализа идентифицируют существенные различия между обработанными и контрольными глазами. Сосудистая облитерация, измеренная в Р17, существенно уменьшается (более 75% восстановление в области облитерации) в обработанных Lin- сетчатках по сравнению с сетчатками глаз, получающих один носитель, или не подвергавшихся инъекции (фиг.38, часть е). Не имеется существенных различий между инъекцией носителя и отсутствием инъекции в данном отношении. Подобным образом, глаза, обработанные Lin-, имеют приблизительно 70% восстановление в области пучков новых сосудов по сравнению с глазами, инъецированными носителем, и более чем 80% восстановление по сравнению с неинъецированным контролем (фиг.38, часть f). Таким образом, обработка глаз Lin- HSC оказывает существенное влияние на два основных повреждения сосудов и параметры восстановления на мышиной модели OIR, т.е. одновременно снижается образование пучков новых сосудов, причем в то же время ускоряется "физиологическая" реваскуляризация в сетчатке.
Ускоренное восстановление также наблюдают, когда обработку осуществляют в период гипероксии и после возвращения к нормоксии, но эффект уменьшается. Эксперименты, описанные ранее, включали инъекции, осуществленные в дни Р2-Р7, до воздействия гипероксии. Для того чтобы определить, влияют ли клетки Lin- также на восстановление сосудов, если инъецированы позднее, во время фазы гипероксии цикла и после возвращения к нормоксии, осуществляют инъекции в Р9, Р11 или Р12, и сетчатки оценивают в различные более поздние моменты времени. Результаты приводятся на фиг.38, часть g, и показывают, что инъекция Lin- HSC эффективна для ускорения восстановления сосудов и уменьшения площади облитерации даже тогда, когда клетки введены во время гипероксии и в Р12. Однако оказывается, что эффект иногда ослабленный, что указывает, что максимальная эффективность достигается, когда обработку осуществляют до воздействия высоких концентраций кислорода.
После обработки гемопоэтическими Lin- клетками-предшественниками структура и функция сетчатки хорошо сохраняются в течение длительного времени. Длительные эффекты и возможное побочное действие обработки Lin- HSC также исследуется. С этой целью берут сетчатки мышей в возрасте 3-6 месяцев, которые подвергались инъекции Lin- HSC и воздействию гипероксии согласно установленной модели (фиг.39). Опухолей не находят, и нейронная сетчатка выглядит сохранившейся гистологически во всех случаях. Единственной заметной аномалией является случайное образование "розетки" в сетчатке, что также имеется в контрольных глазах (фиг.39, части g, h). Сосудистая сеть сетчатки глаз, инъецированных Lin- HSC, имеет нормальный вид, и явные отличия от контрольных глаз, не подвергавшихся инъекции, не обнаружены (фиг.39, части а-f).
Также исследуется продолжительность существования трансплантированных клеток. Клетки GFP+ наблюдают только в небольшом проценте глаз (10%), что указывает, что большинство инъецированных клеток не выживает долее нескольких месяцев. Когда выжившие клетки присутствуют, они часто локализуются в непосредственной близости к сосудистой сети сетчатки. Функция сетчатки, измеренная методом электроретинографической записи, выполненной в период от 17 дней до 6 месяцев после трансплантации, не показывает различия между глазами, в которые трансплантировали Lin- HSC, и здоровыми соответствующими по возрасту глазами без OIR. Для того чтобы проверить возможность того, что трансплантированные клетки могут покинуть глаз и распространиться системно, анализируют селезенки и/или печени 15 мышей на присутствие клеток GFP+ примерно через 7-10 дней после инъекции. Клетки вне глаз не наблюдают.
Проверка активного типа клеток: популяция Lin - обогащена клетками CD44 HI. В усилиях для лучшего понимания механизмов, которые могут действовать во время таких процессов, и упрощения процедуры отбора клеток, предпринята попытка идентифицировать один маркер, который можно использовать для выделения активных HSC из костного мозга. На основании таких характеристик, как вовлечение в миграцию клеток и дифференцировку, собирают большую панель кандидатов в маркеры клеток-предшественников костного мозга. С использованием проточной цитометрии такие маркеры сканируют, сравнивая их экспрессию в активных клетках Lin- против экспрессии в контрольных ВМ клетках, которые, как показано ранее, неактивны в ряде экспериментальных систем. Доказано, что CD44 дифференциально экспрессируются в таких двух популяциях: клетки CD44HI присутствуют в существенно большей пропорции в Lin- клетках (76%), чем в популяции контрольных ВМ клеток (4%) (фиг.40, часть а). Как указано выше, CD44 является клеточным поверхностным рецептором для гиалуроновой кислоты, и, как показано, принимает участие в регуляции некоторых клеточных функций, которые, как полагают, являются важными в опосредовании эффекта спасения, включая выживание, миграцию и дифференцировку. Распределение клеток CD44HI, которые весьма преобладают в популяции активных клеток и достаточно редко встречаются в контрольных клетках, показывает, что CD44 действительно являются эффективным индикатором активности.
Например, клетки CD44HI промотируют восстановление сосудов на модели OIR, в то время как клетки CD44LO не промотируют восстановление. Эффективность клеток CD44HI подтверждают на модели OIR по их способности облегчать восстановление сосудов. С использованием такого же экспериментального плана, какой описан в случае инъекций клеток Lin-, показывают, что клетки CD44HI промотируют восстановление сосудов на такой модели с эффективностью, схожей с эффективностью, которую наблюдают с клетками Lin- (фиг.40, части b, c). Напротив, клетки CD44lo не оказывают позитивного действия на восстановление. Важно, что показано, что в сетчатках животных, обработанных CD44LO, часто наблюдают небольшое число клеток, или их не наблюдают, что предполагает, что такие клетки имеют пониженную способность выживать в стекловидном теле и/или мигрировать в сетчатку. Неизвестно, являются ли клетки CD44HI единственной субпопуляцией активных клеток костного мозга или одной из нескольких субпопуляций, обладающих такой активностью.
Клетки CD44 HI экспрессируют гены и маркеры, напоминающие о миелоидном источнике. Также осуществляют характеризацию популяции CD44HI крупномасштабным анализом экспрессии и мечением антителами Lin- и маркеров, специфических для предшественников, с последующей проточной цитометрией (фиг.41 и фиг.44). Оба способа показывают, что клетки CD44HI имеют профиль экспрессии, напоминающий миелоидный источник. Сильную экспрессию CD11a, CD11b и Ly6G/C наблюдают на таких клетках на уровне белков, в то время как проточной цитометрией обнаруживают менее интенсивное позитивное мечение для F4/80, CD14, cKit и CD115. Некоторые миелоидспецифические гены, включая CD204, CD114, CD33 и CD115, при анализе экспрессии экспрессируются в большой степени по сравнению с клетками CD44lo (фиг.44). Напротив, на уровне белков популяция CD44LO имеет существенную экспрессию на Ter119 и CD45R B220, которые являются маркерами эритробластов/эритроцитов и В-клеток соответственно. На экспрессионной матрице ряд генов, ассоциированных с лимфоцитами, в большой степени экспрессируется в CD44LO по сравнению с клетками CD44HI, включая CD19, CD79a и CD22 (фиг.44). Таким образом, анализ на транскрипционном и белковом уровнях идентифицирует активную популяцию CD44HI как первичную миелоидную по происхождению, в то время как неактивные клетки CD44LO являются, большей частью, лимфоидными.
Анализ трансплантированных клеток in situ - свидетельство дифференцировки. Имея более четко определенную популяцию активных клеток из костного мозга, исследуют судьбу клеток после введения в глаз. С этой целью сетчатки модели OIR, инъецированные CD44HI, анализируют методами иммуногистохимии с различными маркерами. Значительное большинство включенных клеток селективно нацеливаются на сосудистую сеть сетчатки и выбирают преваскулярную локализацию, часто образуя удлиненные структуры, тесно ассоциированные с сосудами хозяина (фиг.42, часть а). С использованием антител против CD31 и NG2 такие маркеры не обнаруживают на экспрессирующих GFP периваскулярных клетках костного мозга, что наводит на мысль, что такие клетки не дифференцируют в эндотелиальные клетки или перициты соответственно. Кроме того, оказывается, что трансплантированные клетки не образуют какую-либо часть просвета сосуда (фиг.42, часть b), демонстрируя, таким образом, что невозможно, чтобы указанные клетки дифференцировали в эндотелиальные клетки. Напротив, микрофаговый/микроглийный маркер F4/80 метит многие, но не все, периваскулярные GFP+ клетки в глазах, обработанных CD44HI (фиг.43, части d-i). Такие включенные F4/80+ клетки имеют вид, весьма схожий с эндогенными периваскулярными клетками, которые также мечены F4/80 (фиг.43, части а-с), что наводит на мысль, что трансплантированные клетки предполагают идентичность, схожую с нативными клетками в модели OIR.
Одним из возможных преимуществ клеточной терапии, в частности, по сравнению с обычным лечением фармацевтическими препаратами, является возможность клеток реагировать на локальные раздражители и претерпевать модификацию при изменениях окружающей среды. Наблюдают, что трансплантированные клетки в Р10 (10 дней после инъекции), которые нацеливаются на сосудистую сеть сетчатки и предпринимают периваскулярную локализацию, имеют CD44, даун-регулированные до уровней, неподдающихся обнаружению (фиг.43, части j-o). Однако клетки, которые не ассоциируются с сосудистой сетью, сохраняют экспрессию CD44. Таким образом, субпопуляции имплантированных клеток, которые изначально отбирают при помощи FACS на основании высокой экспрессии CD44, даун-регулируют этот рецептор in vivo, что коррелирует с местоположением клеток в сетчатке. Это наводит на мысль, что введенные клетки действительно претерпевают селективные изменения (дифференцировку) в окружающей среде глаза.
Результаты, подробно описанные выше, показывают, что терапию на основе клеток можно применять для лечения ROP и других ишемических ретинопатий. Результаты, полученные на мышиной модели, показывают, что такой подход эффективен для уменьшения сосудистой патологии, ассоциированной с воздействием высоких концентраций кислорода, и показывает незначительную токсичность или ее отсутствие. Преимуществом применения клеточной терапии в сравнении с однофакторной терапией может заключаться в возможности клетки адаптироваться и реагировать на изменения окружающей среды. Эволюция от однофакторных средств к сочетаниям лекарственных средств и интервенций, к отбору и доставке усложненных адаптируемых клеток, которые могут распределять и проводить сложную последовательность реакций в процессе взаимодействия с тканью хозяина, является новой интересной концепцией. В этом отношении настоящее изобретение относится к "смещению парадигмы" в подходе к ишемическим ретинопатиям/васкулопатиям, т.е. приданию особого значения лечению и стабилизации вместо ингибирования и облитерации.
Изолированные популяции клеток MLBM по изобретению, нацеленные на сосудистую сеть, могут быть использованы для доставки ангиостатических средств и обладают васкуло- и нейротрофическим действием на модели дегенерации сетчатки. В настоящем исследовании специфические субпопуляции Lin- изолированных популяций клеток MLBM являются высокоэффективными при ускорении восстановления OIR. Представляет интерес то, что активные клетки экспрессируют маркеры, которые, как предполагается, имеют миелоидное происхождение и, вероятно, претерпевают дифференцировку и модификацию после трансплантации.
Применение клеточной терапии для промотирования васкуляризации инициировано областью кардиологии с целью коллатерализации инфаркта артерий. Значительное количество доказательств показывает, что некоторые клетки костного мозга эффективны при улучшении перфузии и функции сердца. Однко пока неясно, какой(ие) тип(ы) клеток ответственен(ны) за наблюдаемые эффекты. Многочисленные исследования, изучающие потенциальную роль эндотелиальных клеток-предшественников, полученных из костного мозга (ЕРС), приходят к заключению, что такие клетки присутствуют в новых или коллатеральных сосудах, но небольшое число включенных клеток, о которых сообщается в таких исследованиях, ставит вопросы рассмотрения их важности. Кроме того, гетерогенные популяции клеток костного мозга, такие как одноядерные клетки, или нефракционированные клетки, которые содержат очень небольшое число стволовых клеток и/или ЕРС, также могут существенно усилить коллатеральное развитие, что приводит к мысли о работе других механизмов, кроме непосредственного включения в сосуды. Хотя не имеется предположения о связи с какой-либо теорией, возможно, что такие клетки играют вспомогательную паракринную роль, за счет которой факторы, секретированные из них, действуют, оптимизируя состояние сосудистой сети хозяина. Показано, что многие субпопуляции клеток костного мозга являются источником ангиогенных факторов, и известно, что моноциты секретируют ряд таких факторов. Таким образом, у клеток костного мозга имеется возможность служить паракринным элементом, дополняя роль ЕРС в образовании коллатеральных сосудов и взаимодействии с иммунной системой хозяина.
Хотя точные механизмы работы в такой системе пока неясны, имеется существенный прогресс в смысле понимания природы функциональных клеток костного мозга. На основании идентификации активной миелоидной популяции в костном мозгу, что обеспечивается клетками по настоящему изобретению, можно сделать некоторые предположения, касающиеся механизма. Миелоидные клетки, особенно моноциты и макрофаги, имеют установленную способность влиять на рост кровеносных сосудов через секрецию ангиогенных факторов роста. Кроме того, показано, что макрофаги более толерантны к гипоксии, чем другие типы клеток, и отвечают на условия низкого содержания кислорода секрецией ангиогенных факторов. Таким образом, включение миелоидных предшественников в ишемические сетчатки может дать клетку, которая может противостоять условиям гипоксии и может промотировать восстановление сосудов как паракринный элемент. Присутствие полученных от хозяина F4/80+-периваскулярных клеток в сетчатке при OIR наводит на мысль, что такой тип клеток играет некую роль в процессе, и, вероятно, доставка большого пула подобных клеток (или их предшественников) прямой трансплантацией в глаз усиливает такое действие. Такой ход событий выдвигает на первый план парадоксальное наблюдение, что, как наблюдалось в настоящем исследовании, инъекция популяций клеток по настоящему изобретению промотирует реваскуляризацию сетчатки, причем в то же время подавляется преретинальное образование новых сосудов. Хотя основа этого пока полностью неизвестна, возможно, что ускоренная "физиологическая" реваскуляризация может уменьшить гипоксию, испытываемую сетчаткой, так что стимулируемые ишемией пучки новых сосудов не образуются в той же степени.
Мысль о том, что миелоидоподобные клетки помогают росту сосудов, может иметь отношение к некоторой ранней работе, относящейся к rd1 и rd10 мышиным моделям дегенерации сетчатки. Инъецированные миелоидные предшественники могут действовать, поддерживая глубокую ретинальную сосудистую сеть через секретированные факторы и предупреждая дегенерацию сосудов, что наблюдают на указанных моделях. Некоторые секретированные макрофагами ангиогенные факторы, такие как bFGF, также показывают нейротрофическую активность. Таким образом, наблюдаемая уменьшенная гибель фоторецепторов после инъекции популяций клеток по настоящему изобретению мышам rd может опосредоваться через паракринный механизм, при котором нейротрофические факторы продуцируются трансплантированными миелоидными клетками, полученными из костного мозга. В поддержку такого механизма настоящее исследование показывает, что ТНЕ изолированные популяции клеток MLBM по изобретению способны к спасению сосудов и нейронов на модели rd с эффективностью, схожей с той, какую наблюдают после инъекции изолированных клеток MLBM.
При клиническом лечении ROP клетки крови плодного тяжа собирают во время родов недоношенного ребенка с высокой опасностью, затем клетки сортируют для обогащения специфической субпопуляцией, которая опосредует эффект спасения, и затем такие аутологичные клетки-предшественники инъецируют в глаз ребенка.
Одним из основных существующих ограничений применения клеточной терапии является тот факт, что во многих случаях точные молекулярные механизмы действия еще неясны, и в действительности такие механизмы могут различаться между моделями. Однако это может в действительности являться самым большим преимуществом терапий на основе клеток, т.е. способность реагировать различными путями и широким разнообразием на изменяющиеся условия и раздражители. Это справедливо не только для различных экспериментальных систем и испытаний, но также всегда в одной системе. Иными словами, такие клетки могут секретировать некоторые факторы в один момент времени и другие факторы в другой, и в итоге, если потребность в них падает, могут прекратить действие в целом. Важно, что существующие способы лечения лекарственными средствами на основе химических веществ этого не могут, и основой является тот факт, что клетки фундаментально применяют и получают ответную реакцию. Модификация клеточных маркеров в трансплантированных клетках in vivo, наблюдаемая в настоящем исследовании, поддерживает такую концепцию.
Пример 16. Клетки MLBM дифференцируют в клетки с микроглиальными характеристиками
Анализ сетчаток после инъекции клеток CD44HI показывает, что популяция CD44HI клеток костного мозга после инъекции в глаз дифференцирует в микроглии. Микроглии являются постоянной миелоидной популяцией в сетчатке и экспрессируют характерные маркеры, включая CD11b. Такие клетки также различаются своей разветвленной (ветвистой) морфологией и предполагают периваскулярную локализацию. Анализируют локализацию, морфологию и экспрессию маркеров клеток CD44hi в различные моменты после инъекции в глаза. Наблюдают, что инъецированные CD44HI GFP+ клетки отображают все описанные характеристики эндогеныых ретинальных микроглий (фиг.45). Части А и В на фиг.45 показывают, что инъецированные клетки CD44HI экспрессируют CD11b и F4/80 и имеют морфологию и периваскулярную локализацию, схожие с эндогенными микроглиями. Часть С показывает 3D-анализ изображений, который показывает, что инъецированные клетки CD44HI локализуются в периваскулярной области. Часть D показывает морфологию инъецированных клеток CD44HI при большом увеличении.
Пример 17. Изоляция клеток MLBM негативным отбором
Желательно для целей экспериментов и клинических применений инъецировать клетки, которые не содержат агенты отбора, связанные с поверхностью, такие как антитела и/или магнитные гранулы. Одним из способов достижения такой цели является использование стратегии негативного отбора для изоляции клеток CD44HI. Через характеризацию профилей экспрессии поверхностных маркеров популяций клеток CD44HI и CD44LO, описанных в данном описании, обнаруживают, что клетки CD44LO отображают высокую экспрессию Ter119 и CD45RB220 - маркеров эритроидных клеток и В-клеток, соответственно. Антитела против указанных маркеров с добавлением маркера Т-клеток CD3e эффективно метят популяцию CD44LO и позволяют удалять их магнитным или FACS разделением, оставляя "незатронутыми" клетки CD44HI как продукт. Клетки, разделенные FACS с использованием такой стратегии, показывают типичные функциональные характеристики популяций клеток MLBM по настоящему изобретению (фиг.46).
Фиг.46, часть А, показывает, что обработка мышиного костного мозга MACS с использованием антител, селективных для CD45R/B220, Ter119 и CD3e, дает популяцию клеток, которые более чем на 90% являются клетками CD44HI. Часть В показывает, что негативная фракция (популяция CD44HI), по существу, свободна от клеток CD45R/B220, Ter119 и CD3e. Часть С показывает, что негативно отобранные клетки CD44HI сохраняют способности к нацеливанию на сетчатку и дифференцировке.
Многочисленные варианты и модификации воплощений, описанных выше, можно осуществить без отхода от сущности и объема новых признаков изобретения. Не предполагаются или не следует предполагать ограничений в отношении конкретных воплощений, поясненных в данном описании.







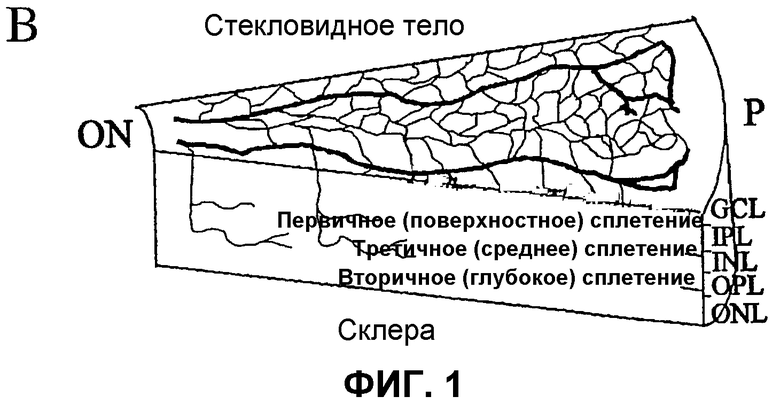

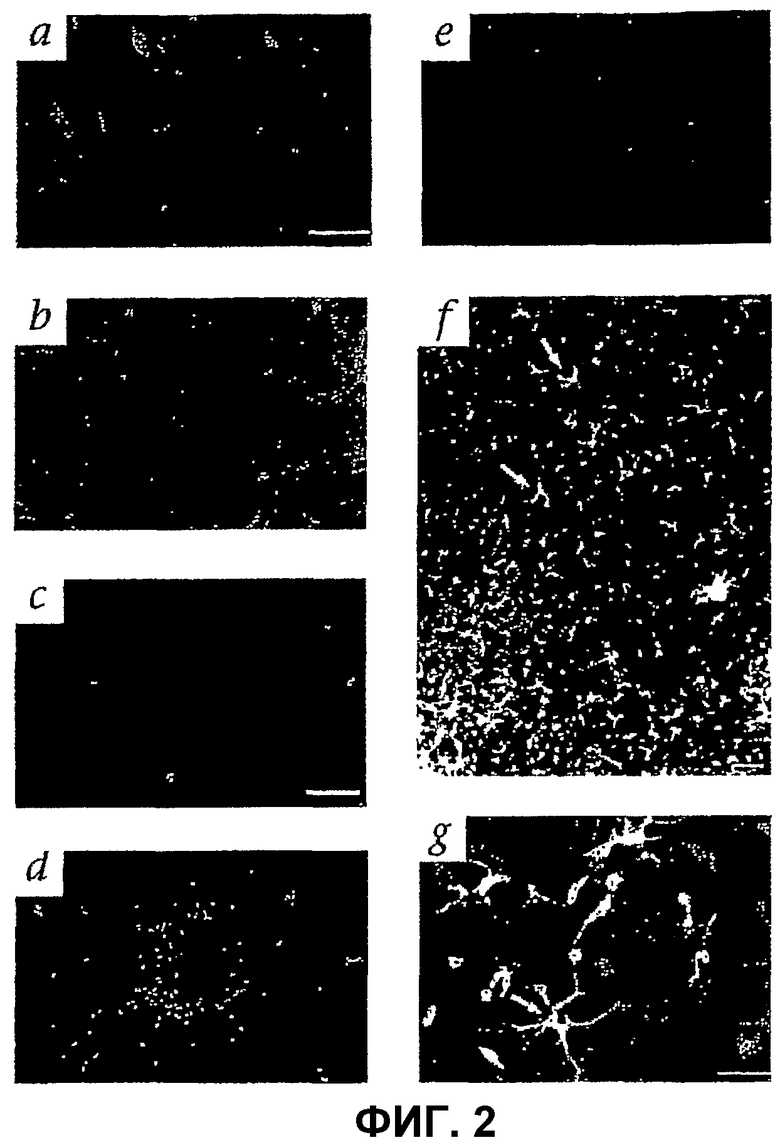

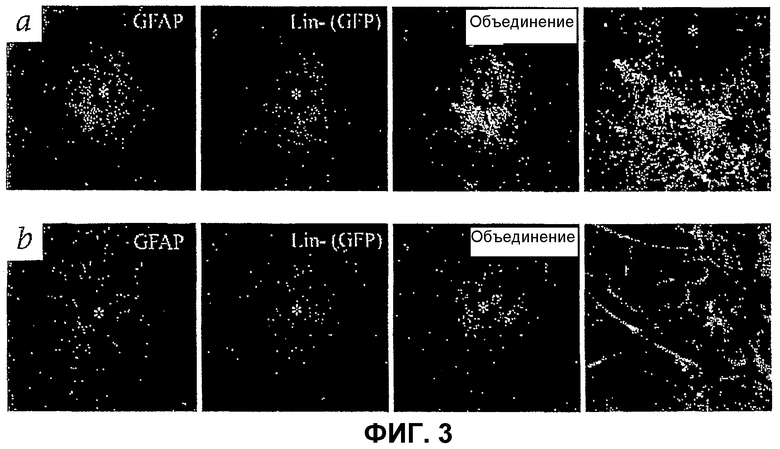
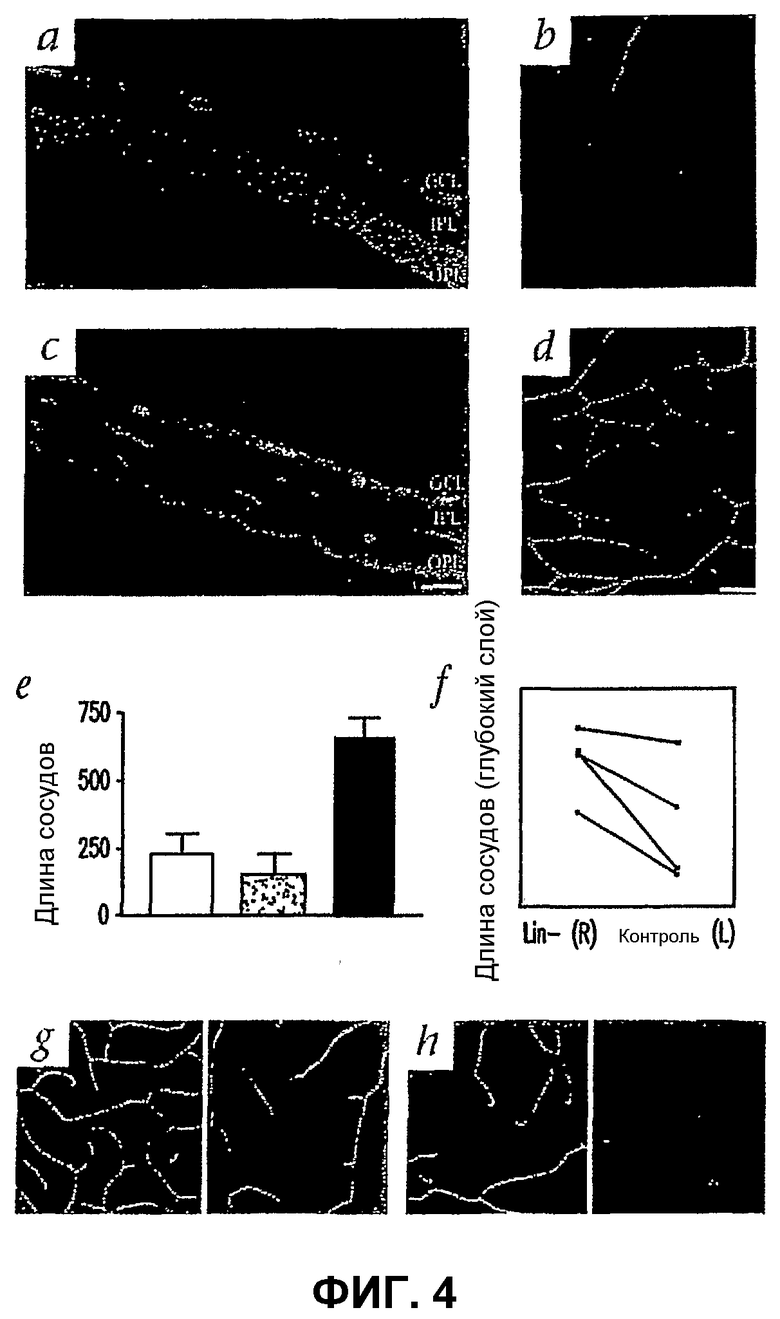
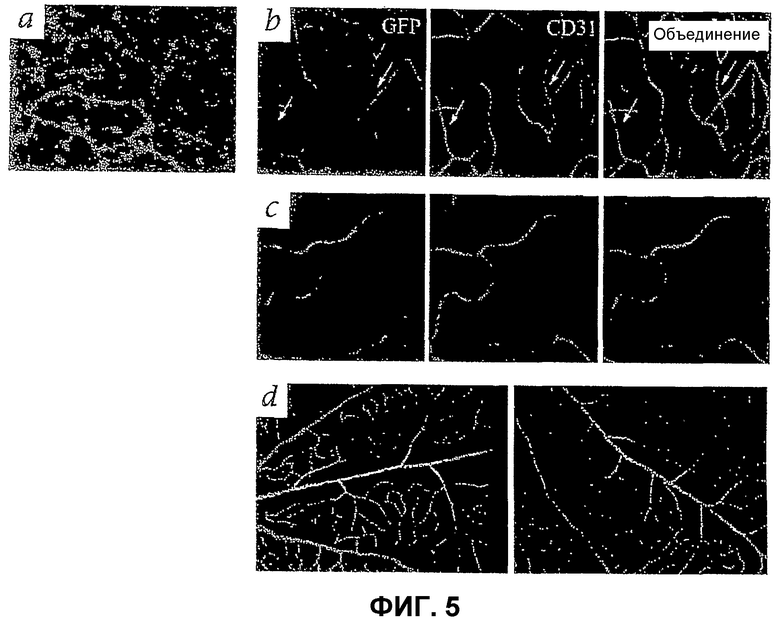
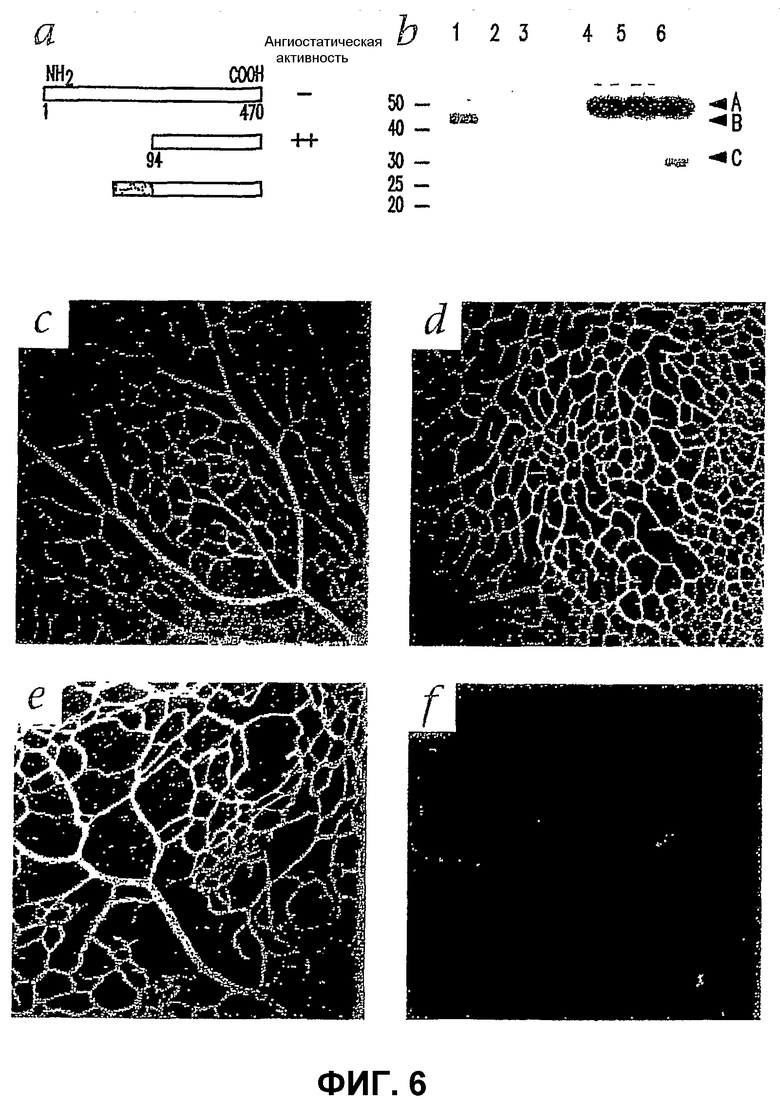



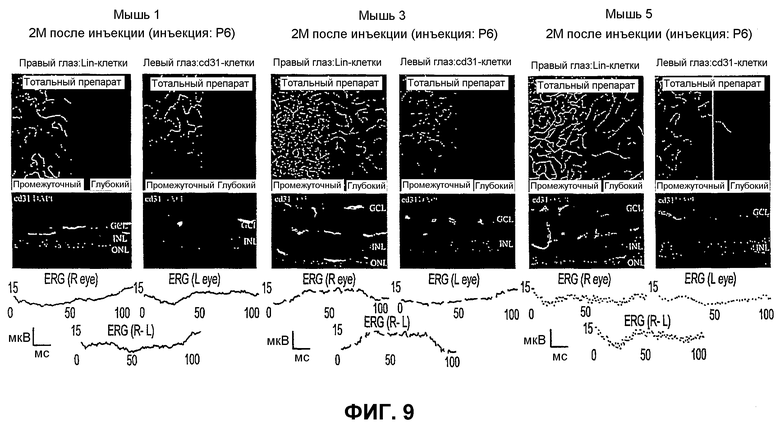
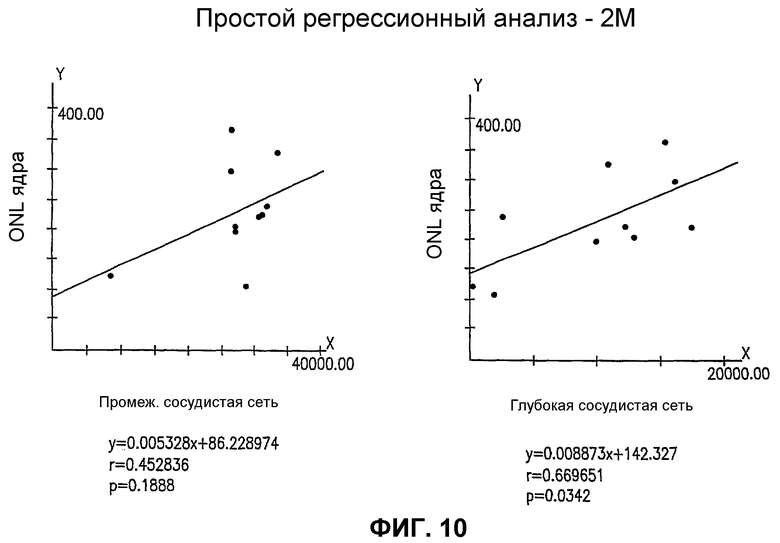
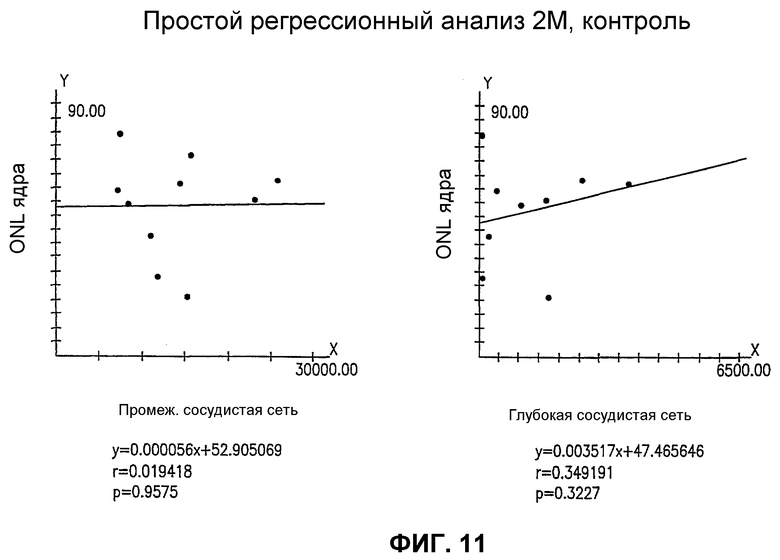
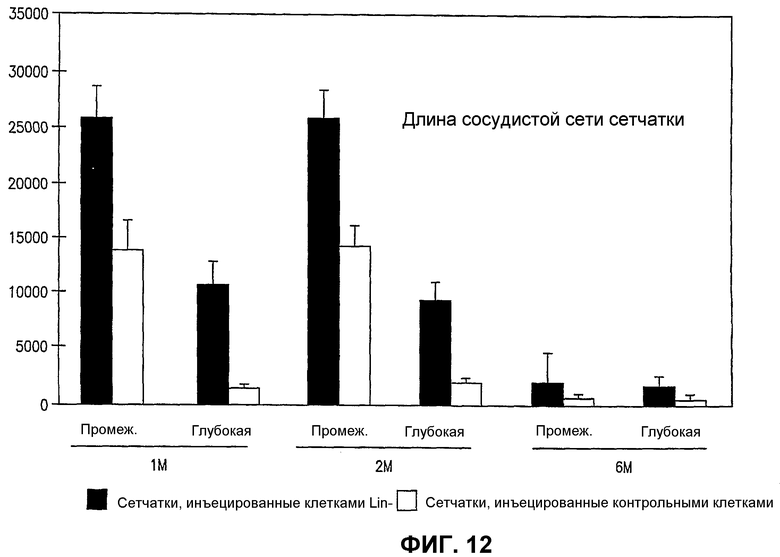
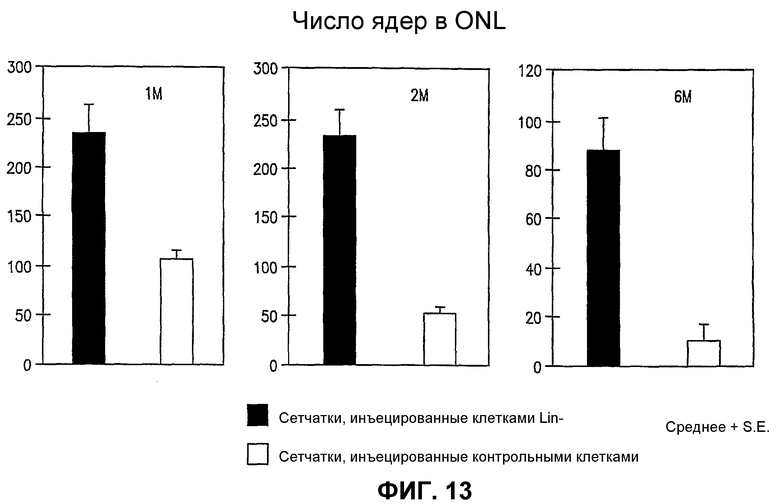
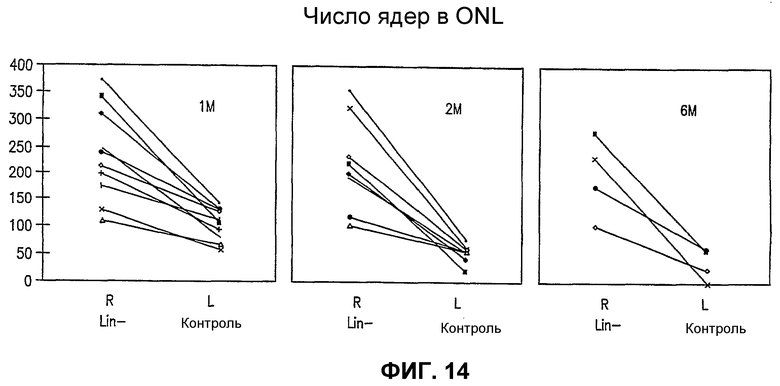
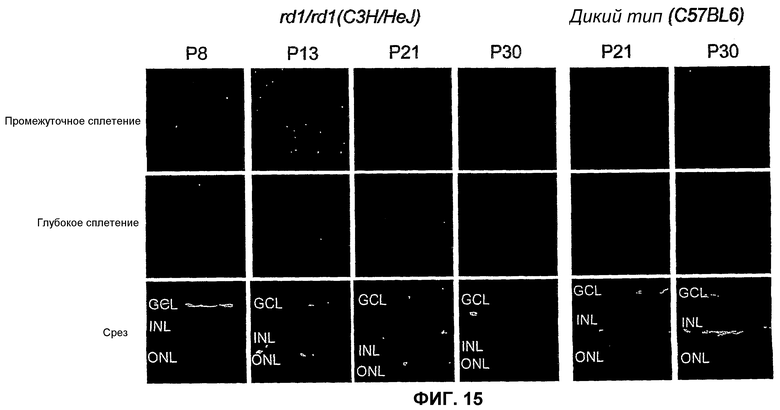
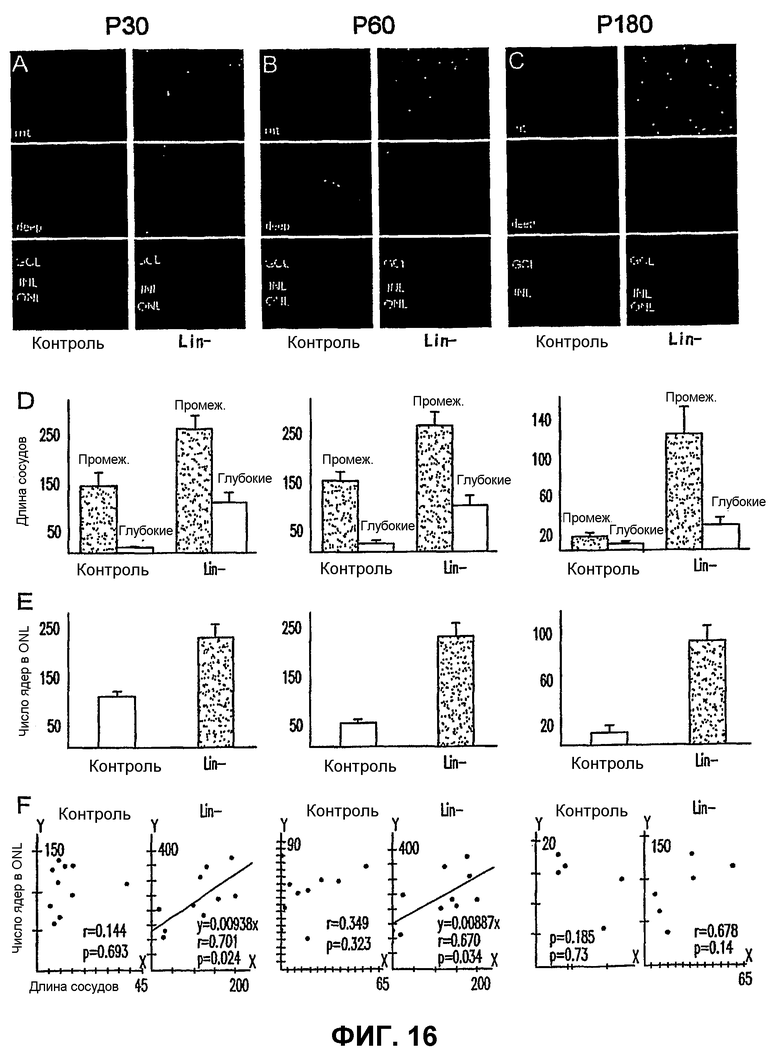
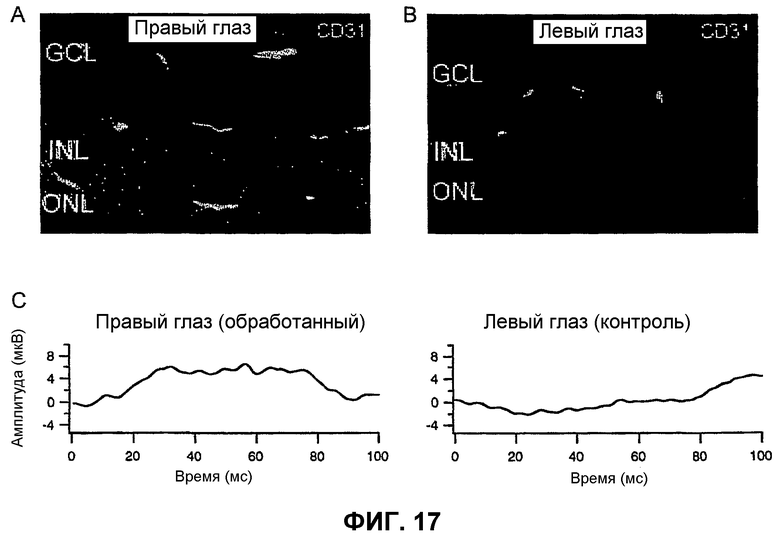
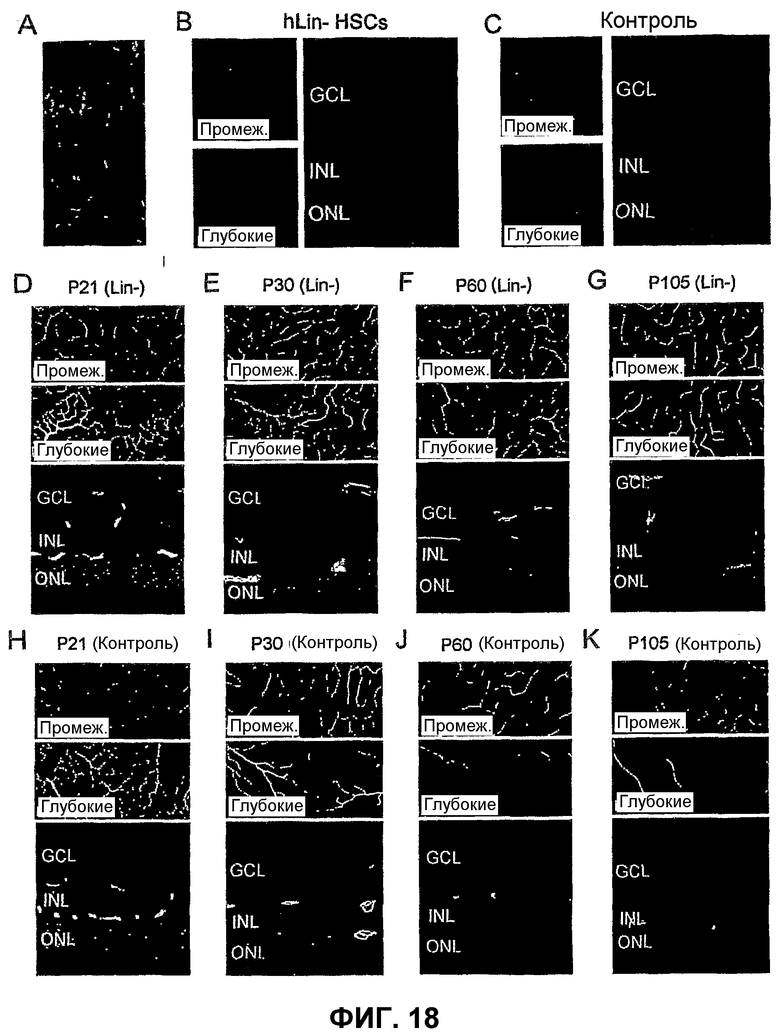
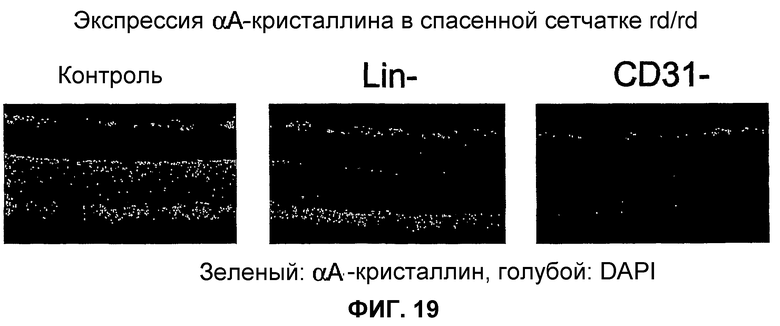
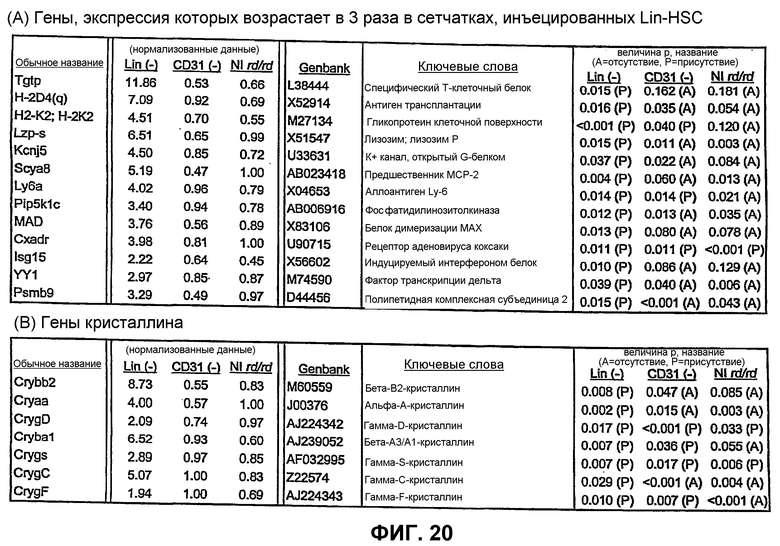
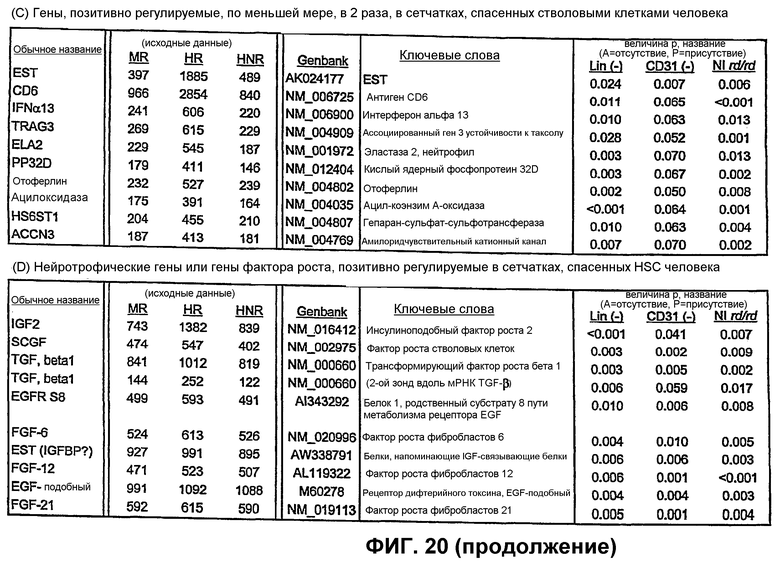
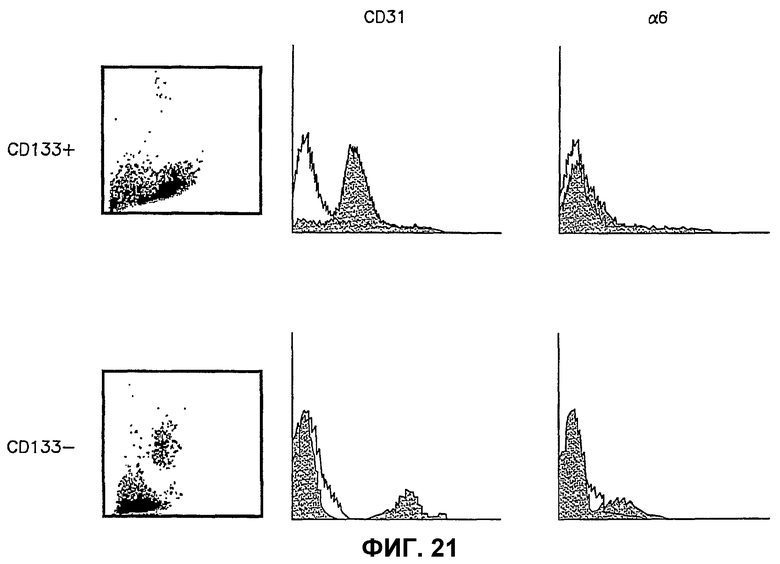
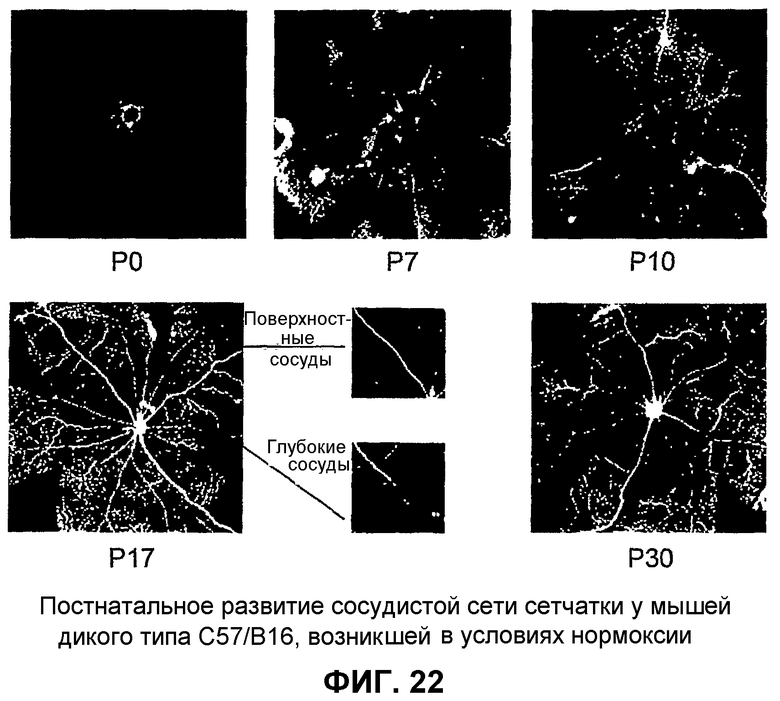
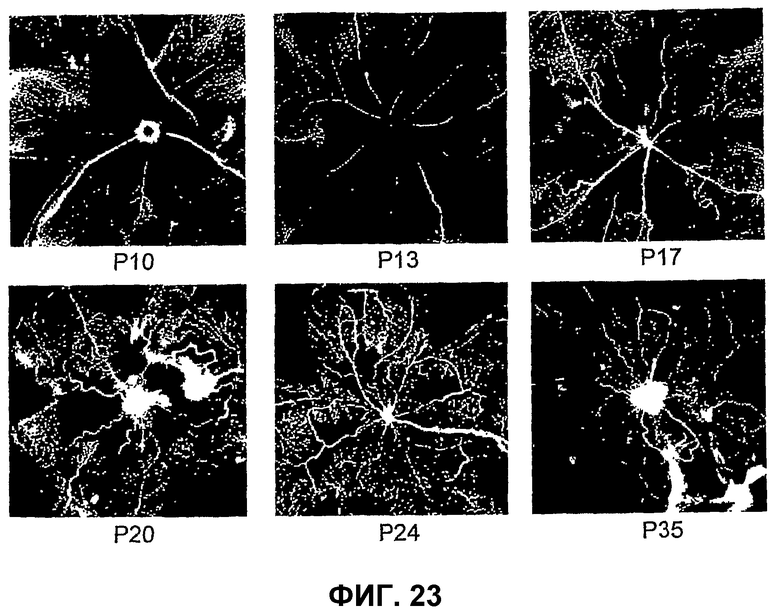
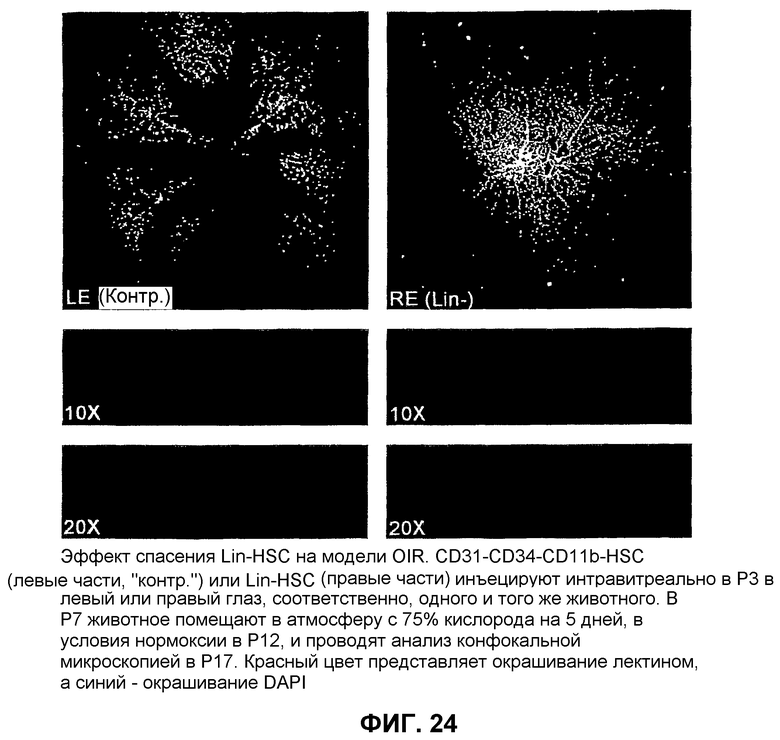
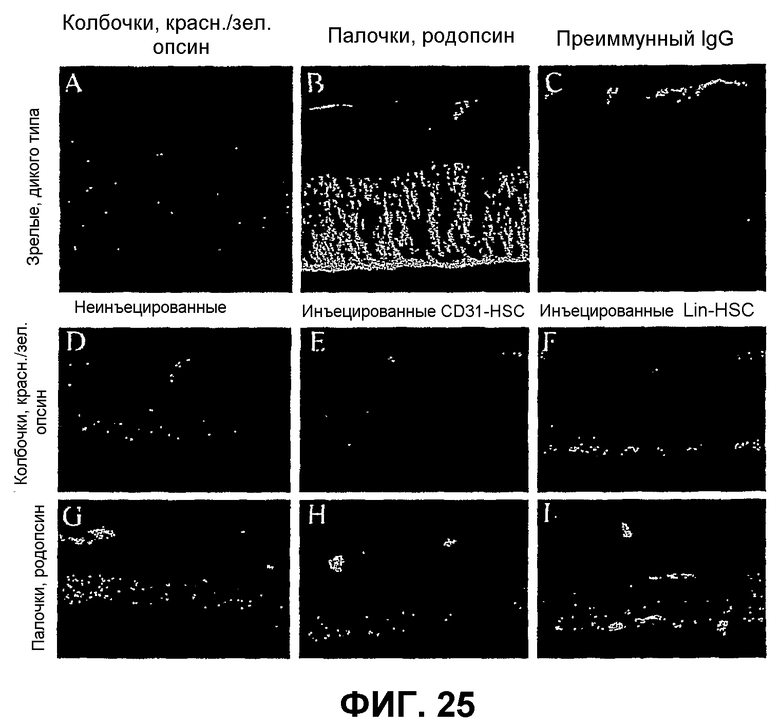
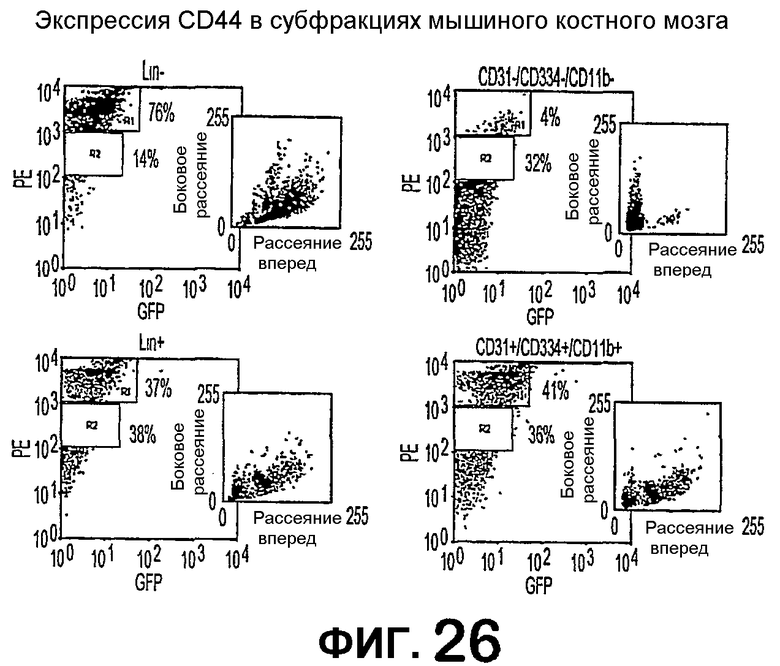
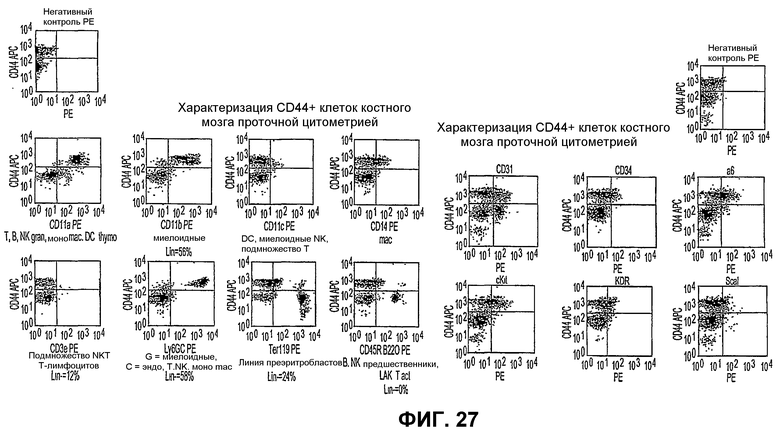
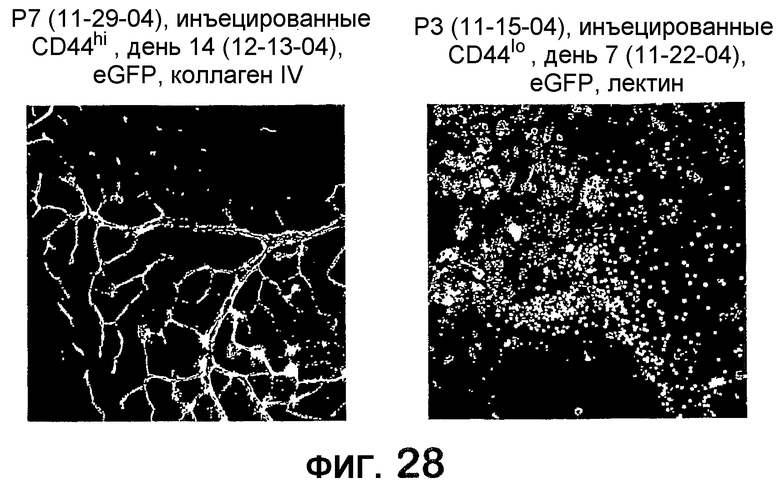
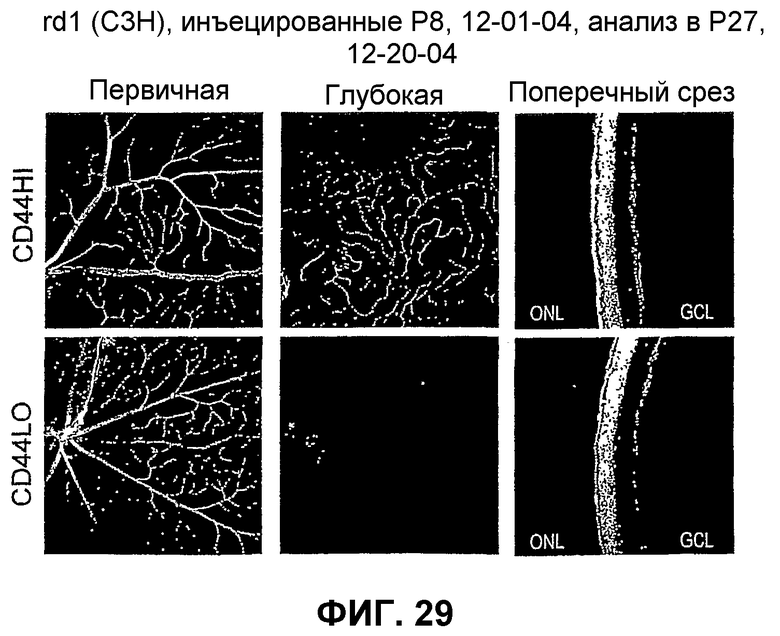
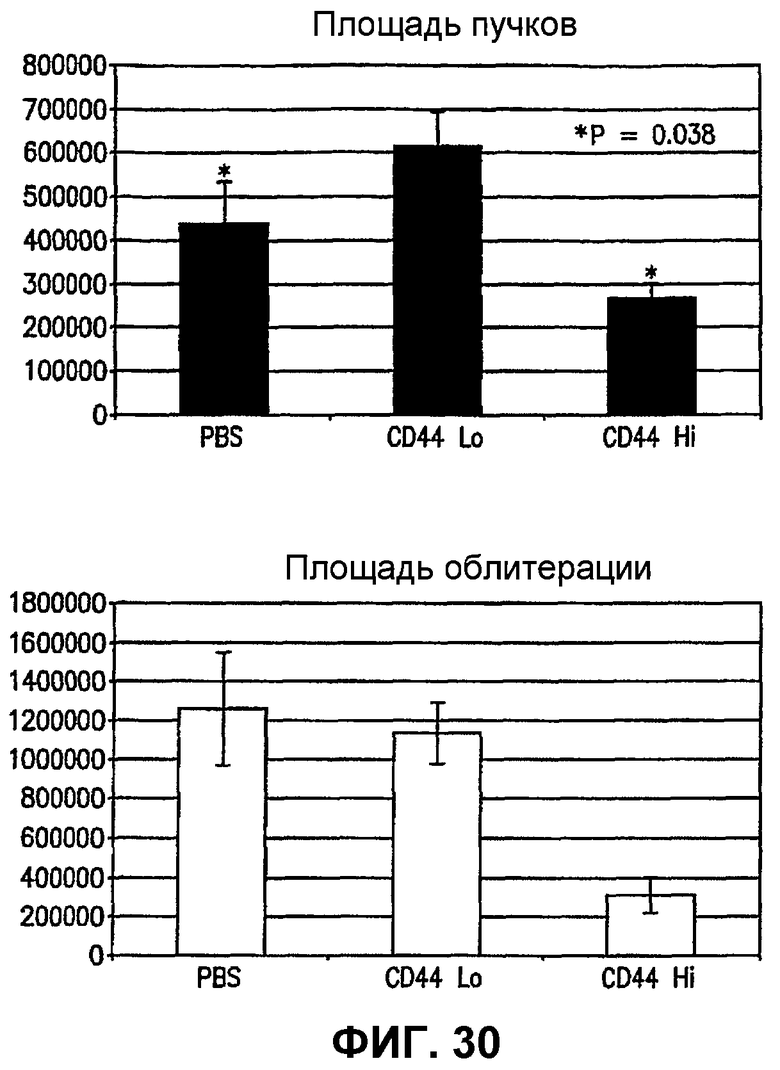

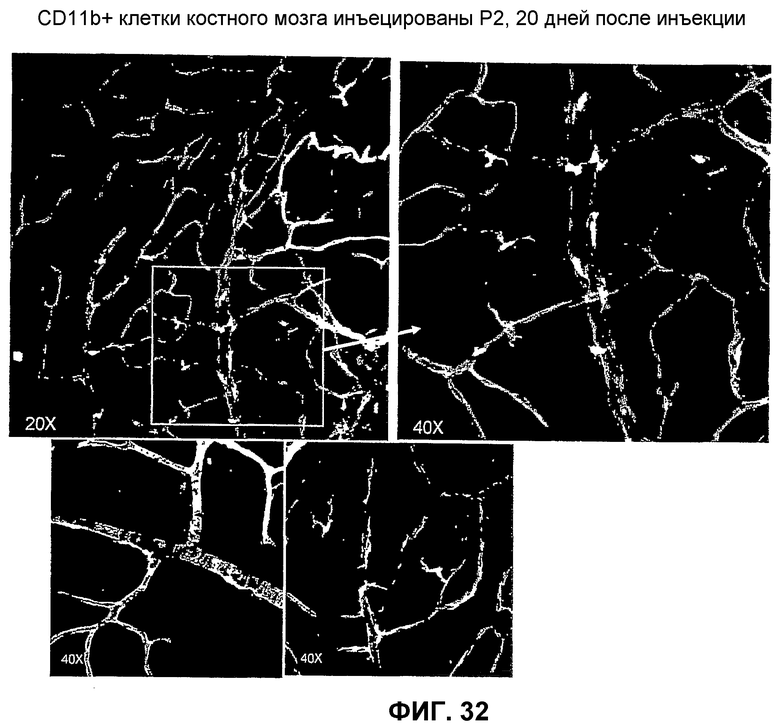
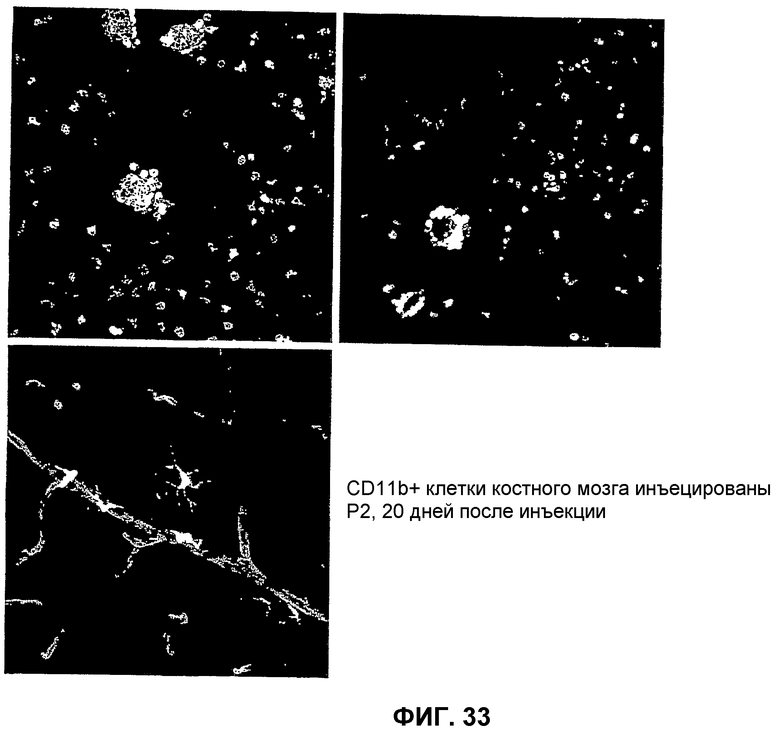



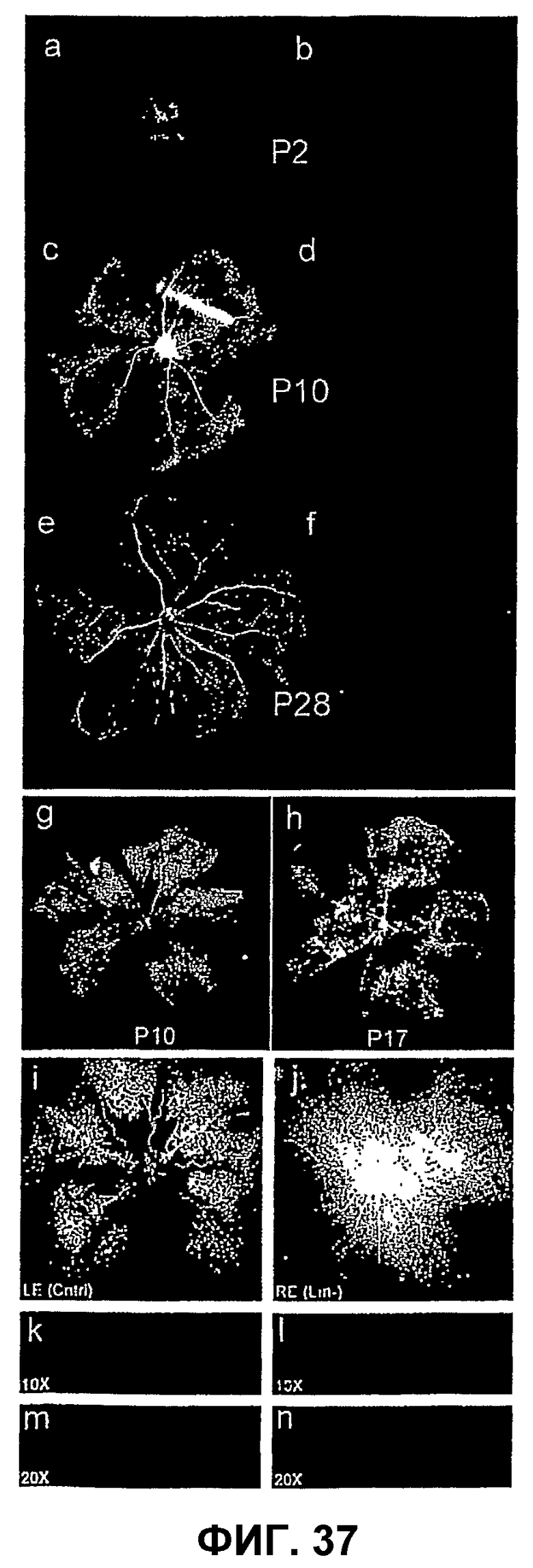
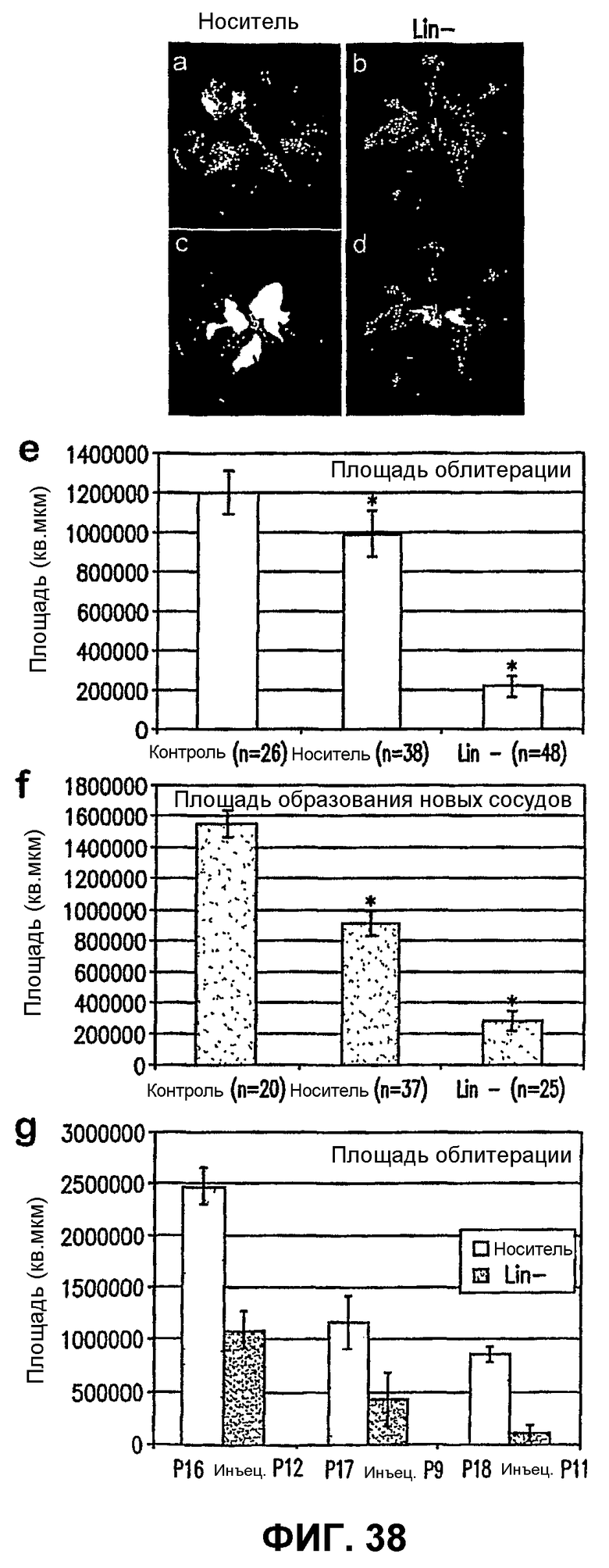
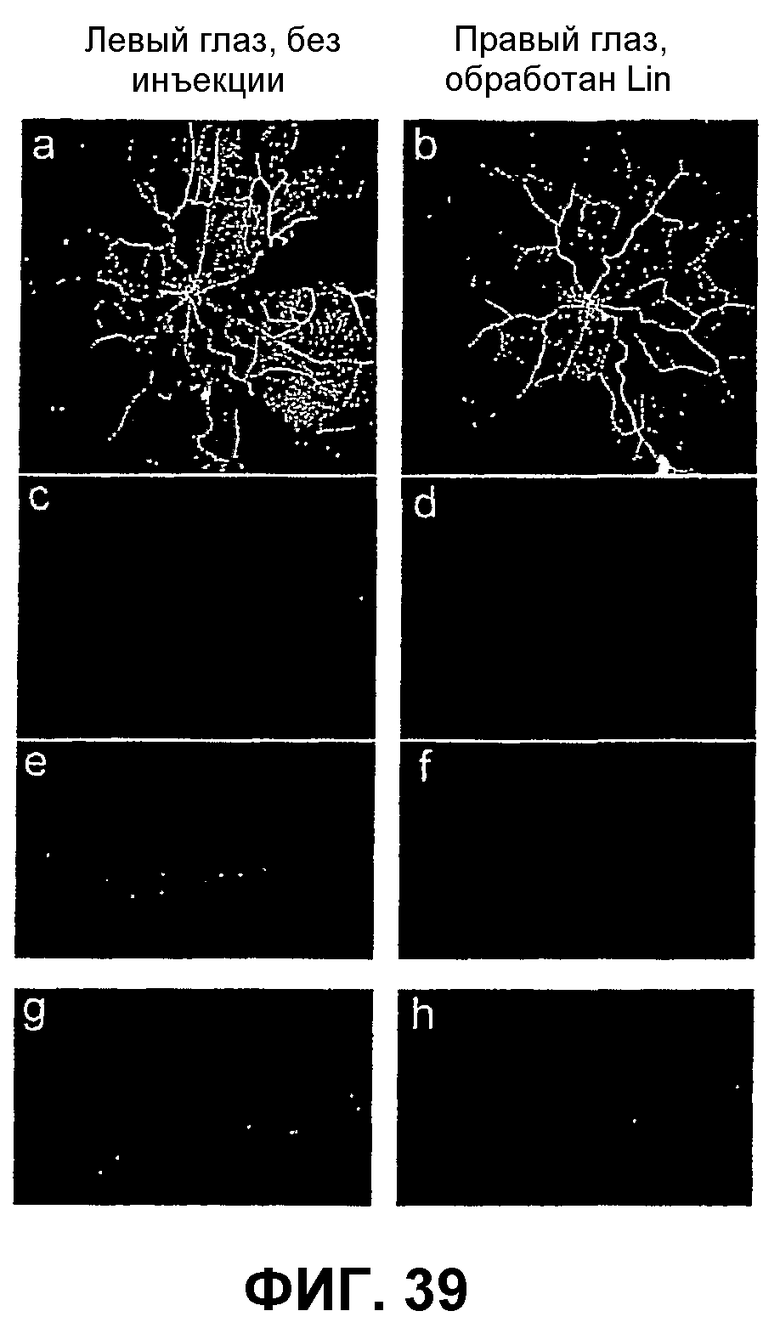
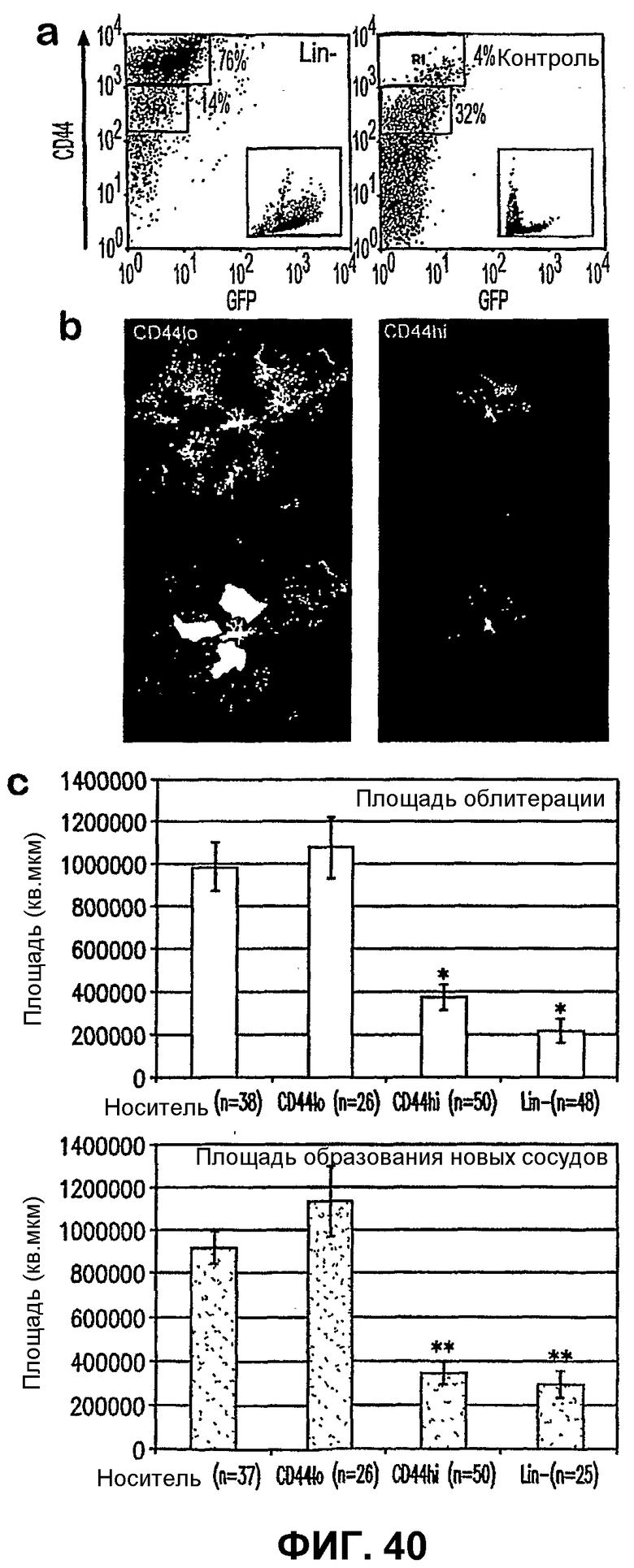
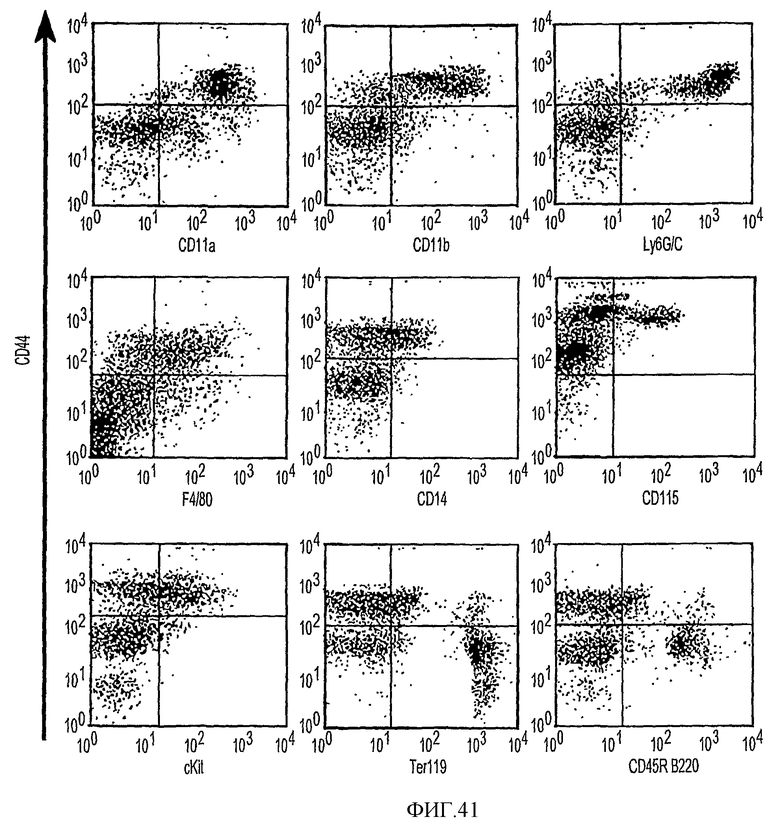

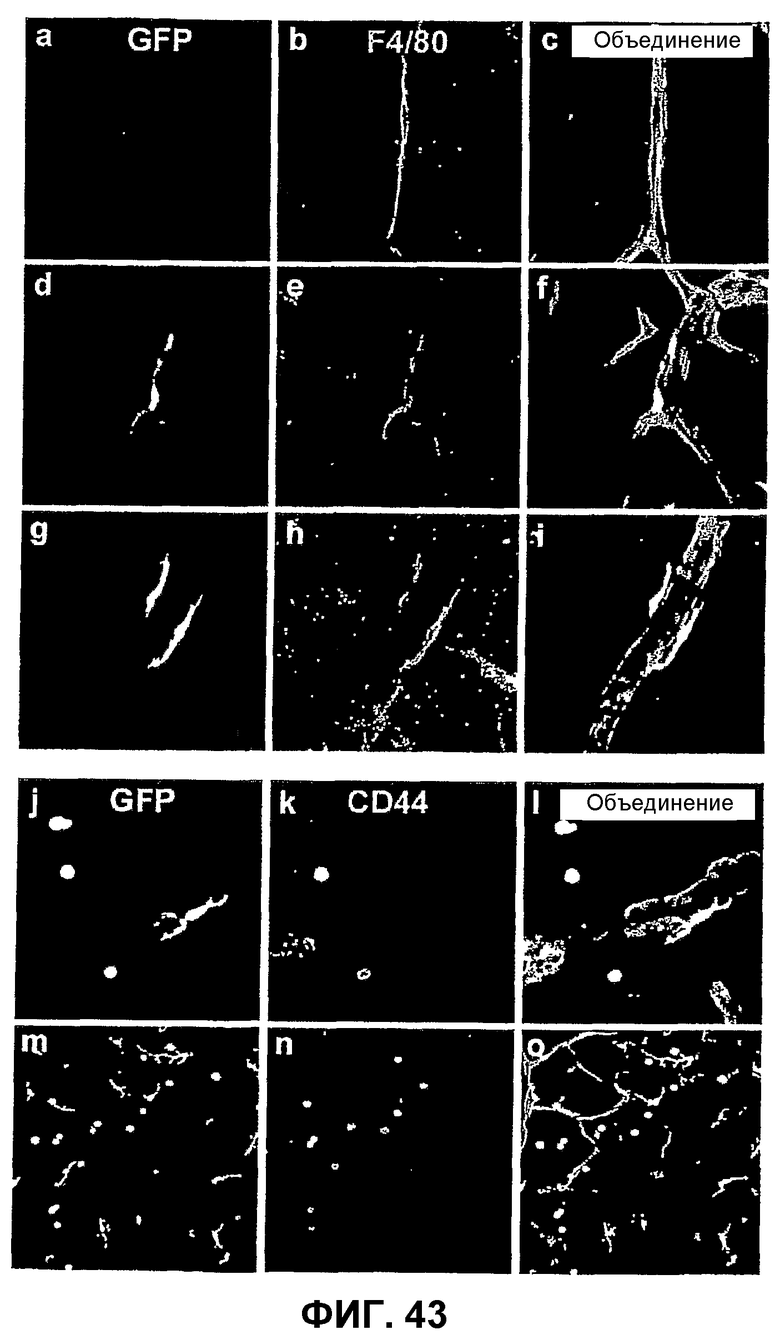
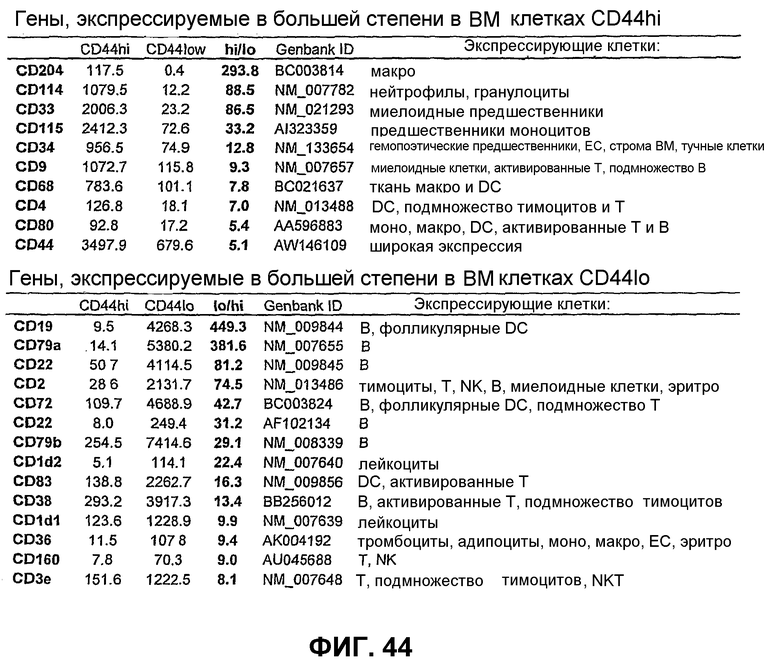
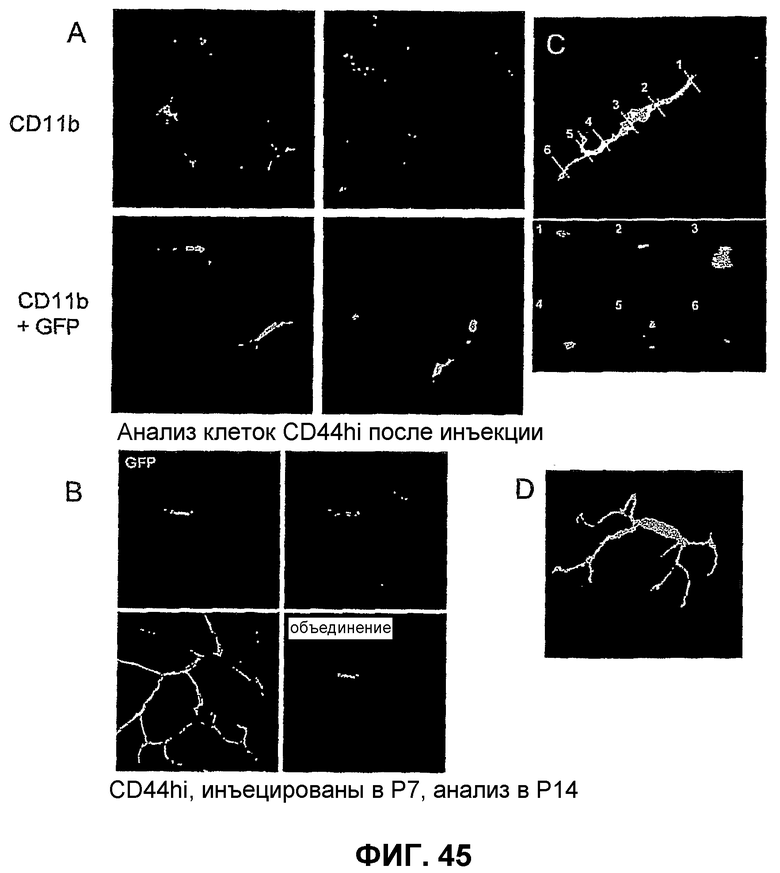
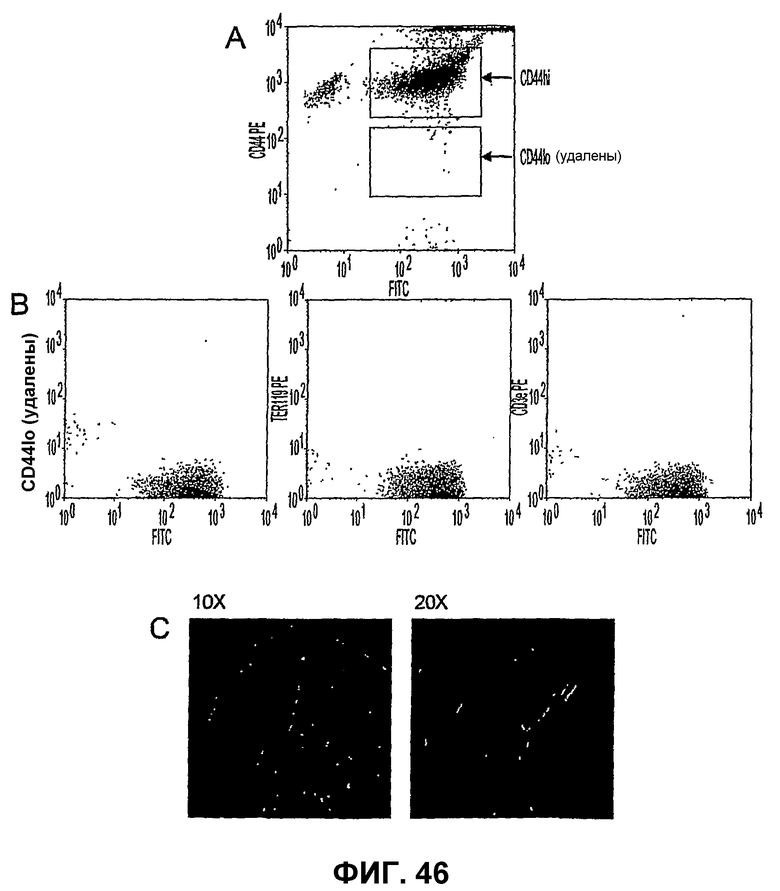
Изобретение относится к области биотехнологии, клеточной инженерии и медицине. Способ выделения миелоидоподобных клеток костного мозга осуществляют путем обработки клеток костного мозга антителами против CD44 (рецептора гиалуроновой кислоты), против CD11b или против того и другого и с использованием проточной цитометрии. Далее проводят позитивный отбор из них клеток, экспрессирующих CD44 и/или CD11b. Изолированные миелоидоподобные клетки костного мозга можно трансфицировать геном, кодирующим терапевтически применимый белок, для доставки гена в сетчатку. Полученные клетки обладают благоприятной васкулотрофической и нейротрофической активностью, когда введены интраокулярно в глаз млекопитающего, в частности млекопитающего, страдающего от дегенеративной болезни глаз. 2 н. и 19 з.п. ф-лы, 46 ил., 3 табл.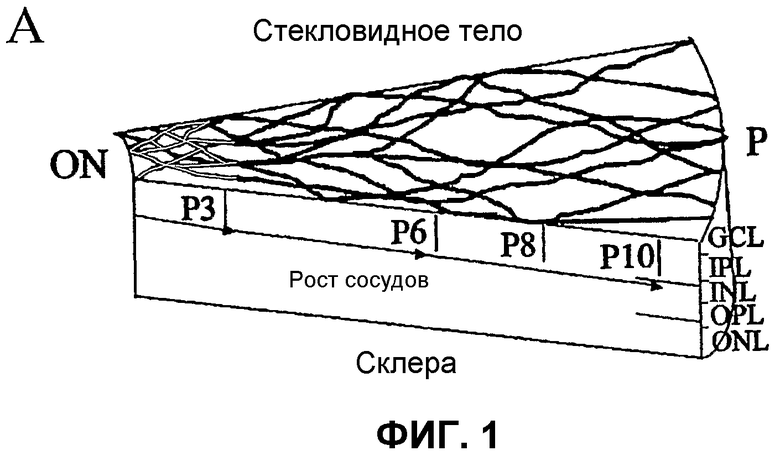
1. Способ получения изолированной популяции миелоидоподобных клеток костного мозга, которые экспрессируют как антиген CD44, так и антиген CD11b, и которые способны к интеграции в сосудистую сеть сетчатки глаза, куда вводят указанную популяцию клеток, включающий выделение клеток костного мозга из млекопитающего и позитивный отбор клеток из костного мозга, которые иммунореактивны в отношении антител, выбранных из группы, состоящей из анти-СП44, анти-CD11b и их сочетания, и последующее разделение клеток, которые экспрессируют CD44 и CD11b антигены, от антитела или антител.
2. Способ уменьшения интенсивности дегенеративной болезни глаз у млекопитающего, включающий введение в глаз млекопитающего, страдающего от болезни глаз, терапевтически эффективного количества клеток из изолированной популяции миелоидоподобных клеток костного мозга, которые экспрессируют как антиген CD44, так и антиген CD11b, где изолированная популяция миелоидоподобных клеток костного мозга получена способом по п.1, и количество клеток является достаточным для замедления дегенерации сосудов, дегенерации нейронов или того и другого в сетчатке глаза, в который вводят клетки.
3. Способ по п.2, где клетки являются аутологичными млекопитающему, которому вводят клетки.
4. Способ по п.2, где млекопитающим является человек.
5. Способ по п.2, где клетки вводят внутриглазной инъекцией.
6. Способ по п.2, где болезнь представляет собой дегенеративную болезнь сетчатки.
7. Способ по п.2, где болезнь представляет собой ишемическую ретинопатию.
8. Способ по п.2, где болезнь представляет собой кровоточивость сосудов.
9. Способ по п.2, где болезнь представляет собой просачивание сосудов.
10. Способ по п.2, где болезнь представляет собой хориоретинопатию.
11. Способ по п.2, где болезнь представляет собой связанную с возрастом дегенерацию желтого пятна.
12. Способ по п.2, где болезнь представляет собой диабетическую ретинопатию.
13. Способ по п.2, где болезнь представляет собой (предположительно) глазной гистоплазмоз.
14. Способ по п.2, где млекопитающим является новорожденное млекопитающее.
15. Способ по п.2, где болезнь представляет собой ретролетальную фиброплазию.
16. Способ по п.2, где болезнь представляет собой серповидно-клеточную анемию.
17. Способ по п.2, где болезнь представляет собой пигментный ретинит.
18. Способ по п.2, где клетки перед введением в глаз млекопитающего трансфицируют геном, который операбельно кодирует терапевтически применимый пептид.
19. Способ по п.18, где терапевтически применимый пептид представляет собой антиангиогенный пептид.
20. Способ по п.18, где терапевтически применимый пептид представляет собой нейротрофическое средство.
21. Способ по п.20, где нейротрофическое средство выбирают из группы, состоящей из фактора роста нерва, нейротрофина-3, нейротрофина-4, нейротрофина-5, цилиарного нейротрофического фактора, нейротрофического фактора, полученного из пигментированного эпителия сетчатки, инсулиноподобного фактора роста, нейротрофического фактора, полученного из глиальной клеточной линии, и нейротрофического фактора, полученного из головного мозга.
| WILLHEIM M | |||
| et al | |||
| Immunophenotyping of human bone marrow-derived macrophages | |||
| Scand | |||
| J | |||
| Immunol | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| KANSAS G.S | |||
| et al | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Blood | |||
| Способ приготовления консистентных мазей | 1919 |
|
SU1990A1 |
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| KRIZANAC-BENGEZ L. | |||

