Данная заявка является частичным продолжением заявки №10/441089, зарегистрированной 19 мая 2003 г., которая является частичным продолжением заявки №10/299486, зарегистрированной 19 ноября 2002 г.
Железо является существенным микроэлементом, который требуется для роста и развития всех живых организмов; оно обязательно для синтеза ДНК и широкого спектра метаболических процессов. Однако нарушения метаболизма железа связаны с рядом существенных заболеваний млекопитающих, включая, но не ограничиваясь этим, железодефицитную анемию, гемосидероз или гемохроматоз, заболевание с перегрузкой железом (Pietrangelo, А., (2002) Am J Physiol. Gastrointest. Liver Physiol. 282, G403-414; Andrews, N.C. (2000) Annu. Rev. Genomics Hum. Genet. 1, 75-98; Philpott, C.C. (2002) Hepatology 35, 993-1001; Anderson and Powell, (2002) Int J Hematol 76, 203-203; Beutler et al., (2001) Drug-Metab. Dispos. 29, 495-499). В физиологических условиях содержание железа у человека регулируется контролируемой адсорбцией. У млекопитающих адсорбция железа осуществляется преимущественно в двенадцатиперстной кишке и верхних отделах тощей кишки, и это является единственным механизмом, с помощью которого железо запасается и физиологически контролируется (Philpott, С.С. (2002) Hepatology 35, 993-1001). После адсорбции железо связывается с циркулирующим трансферрином и доставляется к тканям организма. В печени, главном месте запасания железа, связанное с трансферрином железо захватывается клетками с помощью опосредованного рецепторами эндоцитоза через классический трансферриновый рецептор (TfR1) (Collawn et al. (1990) Cell 63, 1061-1072) и, вероятно, в больших количествах через недавно идентифицированный гомологичный трансферриновый рецептор 2 (TfR2) (Kawabata et al. (1999) J Biol Chem 274, 20826-20832). Внеклеточный домен данного белка на 45% идентичен соответствующей части TfR1 (там же). TfR2 может также связывать трансферрин в связи с двумя молекулами железа и способствовать захвату железа. Мутации TfR2 связаны с определенными формами гемохроматоза, демонстрируя важную роль TfR2 в гомеостазе железа (Philpott, С.С. (2002) Hepatology 35, 993-1001; Camasehella et al., (2000) Nat. Genet. 25, 14-15; Fleming et al., (2002) Proc. Natl. Acad. Sci. USA 99, 10653-10658). TfR2 экспрессируется преимущественно в печени (Fleming et al., (2000) Proc. Natl. Acad. Sci. USA 97, 2214-2219; Subramaniam et al., (2002) Cell Biochem. Biophys. 36, 235-239), однако его точная клеточная локализация до сих пор не известна.
Существует механизм обратной связи, который увеличивает абсорбцию железа у индивидуумов с дефицитом железа, в то время как абсорбция железа снижается у людей с избытком железа (Pietrangelo, А., (2002) Am J Physiol Gastrointest Liver Physiol 282, G403-414; Philpott, C.C. (2002) Hepatology 35, 993-1001; Anderson and Powell, (2002) Int J Hematol 76, 203-203). При наследственном гемохроматозе (НН), однако, данный регуляторный механизм, очевидно, нарушается; несмотря на избыток железа, повышенное количество железа абсорбируется из пищи, и это ведет к аккумуляции избытка железа во внутренних органах, приводя к дисфункции и недостаточности органов. Молекулярные механизмы, с помощью которых кишечник отвечает на изменения в потребностях организма в железе, плохо изучены. В данном контексте гепсидин, недавно идентифицированный пептид млекопитающих (Krause et al. (2000) FEBS Lett 489, 147-150: Park et al. (2001) J Biol Chem 276, 7806-7810), претендует на роль ключевого сигнального компонента, регулирующего гомеостаз железа (Philpott С.С. (2002) Hepatology 35, 993-1001; Nicolas et al. (2002) Proc Natl Acad Sci USA 99, 4596-4601).
Гепсидин представляет собой небольшой обогащенный цистеином пептид, продуцируемый преимущественно в печени. Данная молекула регулирует абсорбцию железа в кишечнике и ингибирует высвобождение железа из макрофагов. Гепсидин был первоначально выделен из плазмы крови и мочи человека как пептид из 25 аминокислот (а.к.), проявляющий антимикробную активность (Krause et al. (2000) FEBS Lett 489, 147-150; Park et al. (2001) J Biol Chem 276, 7806-7810). Впоследствии, при поиске специфичных для печени генов, которые регулируются железом, были идентифицированы кДНК гепсидина, кодирующие 83 а.к. предшественник у мышей и 84 а.к. предшественник у крысы и человека, включая предполагаемый 24 а.к. сигнальный пептид (Pigeon et al. (2001) J Biol Chem 276, 7811-7819). Структура кДНК гепсидина человека предполагает, что он транслируется как препропептид из 84 аминокислот, который в результате процессинга с аминоконца превращается в прогепсидиновый пептид из 60 аминокислотных остатков, который подвергается дополнительному процессингу до гепсидинового пептида из 25 аминокислот (Park et al. (2001)).
Экспрессия гепсидина исчезает у мышей с избытком железа из-за направленного разрушения гена вышележащего стимуляторного фактора 2 (Usf2), проявляющих тот же фенотип, что и найденный у мышей hfe-/- (Nicolas G., et al. (2001) Proc Natl Acad Sci USA 98, 8780-8785), приводя к заключению, что данный пептид играет центральную роль в метаболизме железа. Наоборот, гиперэкспрессия гепсидина, как показано, ведет к тяжелой железодефицитной анемии у трансгенных мышей (Nicolas et al. (2002) Proc Natl Acad Sci USA 99, 4596-4601), указывая на то, что гепсидин является центральным регулятором гомеостаза железа. Более того, недавние исследования показали, что экспрессия гепсидина в печени снижается у hfe нокаутной мыши (Ahmad et al. (2002) Blood Cells Mol Dis 29, 361-366), а мутации в пептиде гепсидине связаны с тяжелым ювенильным гемохроматозом (Roetto et al. (2003) Nat Genet 33, 21-22), что открывает новые перспективы для понимания молекулярного патогенеза избытка железа. Однако механизм, с помощью которого гепсидин поддерживает баланс запасов железа в организме или корректирует абсорбцию железа пищи при физиологических и патологических состояниях, все еще ожидает своего открытия.
В данном отношении клеточная локализация данного пептида и его регуляция при различном уровне железа имеют важное значение для исследования функции гепсидина. Хотя анализ методом Нозерн-блоттинга уровней мРНК гепсидина человека и мыши в различных органах позволил выявить, что гепсидин преимущественно экспрессируется в печени (Krause et al. (2000) FEBS Lett 489, 147-150; Park et al. (2001) J Biol Chem 276, 7806-7810; Nicolas et al. (2002) Proc Natl Acad Sci USA 99, 4596-4601), не существует данных о клеточной локализации данного пептида.
Краткое изложение существа изобретения
Настоящее изобретение касается регуляции гепсидином захвата железа клетками млекопитающих и применения гепсидина и/или специфических антител против гепсидина для диагностики заболеваний, связанных с нарушениями метаболизма железа. Наборы для диагностического определения согласно изобретению могут быть, в частности, использованы для скрининга всей популяции либо людей, либо животных и идентификации тех субъектов, которые имеют данные заболевания.
В одном аспекте изобретение представляет собой способ диагностики патологического состояния, характеризующегося нефизиологическими уровнями гепсидина, включающий получение образца ткани или жидкости от субъекта; контактирование образца с антителом или его фрагментом, который специфически связывает полипептид из средней части (аминокислоты от 20 до 50) или С-конца (аминокислоты от 65 до 84), и количественное определение уровня гепсидина с применением теста, основанного на связывании антитела и полипептида; где нефизиологический уровень гепсидина является показателем патологического состояния. В одном аспекте согласно изобретению чувствительные диагностические методы и наборы, как установлено, способны определять прогепсидин в плазме человека. Изобретение открывает широкий диапазон терапевтических перспектив, где антитело против гепсидина и диагностические методы и наборы могут быть использованы для определения гепсидина в качестве параметра, характеризующего прогрессирование заболеваний, указанных выше, и после терапии.
В одном осуществлении изобретение касается получения и очистки белка гепсидина, включая прогепсидин и его фрагменты. В другом осуществлении изобретение касается специфических антител против гепсидина, или их фрагментов, или вариантов, которые, в свою очередь, могут быть использованы в иммунных тестах для определения белка гепсидина, включая прогепсидин у предполагаемых людей или животных.
В другом аспекте изобретения диагностические способы и наборы для определения гепсидина могут быть использованы в генетических технологических подходах, таких как для определения гиперэкспрессии или снижения экспрессии гепсидина.
В еще одном аспекте изобретения гепсидин может быть использован для терапевтического лечения описанных здесь заболеваний путем лечения субъектов гепсидином и агонистами или антагонистами гепсидина. Захват железа клетками может модулироваться путем варьирования концентрации гепсидина, ингибирования связывания гепсидина с железом или рецептором TfR2. Соответственно, гепсидин и агонисты или антагонисты гепсидина могут быть использованы для лечения состояний, при которых существуют нарушения метаболизма железа. Например, такие вещества могут быть пригодны для лечения указанных выше заболеваний.
Эти и другие аспекты согласно изобретению будут лучше поняты при ссылке на последующие чертежи и подробное описание.
Краткое описание чертежей
На фиг.1 представлена аминокислотная последовательность белка-предшественника гепсидина человека, содержащая на N-конце типичный 24 а.к. сигнальный пептид (линия между а.к. 24 и 25 указывает на предполагаемый сайт отщепления сигнальной последовательности), 35 а.к. про-область и с-концевые 20-, 22- и 25- а.к. пептиды гепсидина, отличающиеся только по укорочению с N-конца, как отмечено стрелками. После отщепления сигнального пептида от предшественника гепсидина получается молекула прогепсидина, состоящая из 60 а.к. Предполагаемыми дисульфидными связями в гепсидине 25 являются 1-8, 2-7, 3-6 и 4-5, как показано прерывистыми линиями (из Hunter et al., 20). Антисыворотки EG (1 и 2)-HepN выработаны против предшественника гепсидина, а.к. 28-47, антисыворотка EG(1)-HepC, выработана против а.к. 70-84, как указано символами антител.
Фиг.2 иллюстрирует следующее: (А): РТ-ПЦР анализ печени человека (линии 2 и 3) и клеток HepG2 (линии 4 и 5), показывающий экспрессию гена гепсидина. Указан лэддер п.н. ДНК (линии 1 и 7). Линия 6 представляет собой негативный контроль. (B-D): анализ гепсидина иммуноблоттингом в экстрактах морской свинки (линии 1) и печени человека (линии 2), а также в клетках HepG2 (линии 3), сыворотке человека (линии 4) и скелетной мышце морской свинки (линии 5, контроль) с антителами EG(1)-HepN (В), EG(2)-HepN (С) и EG(1)-HepC (D). Отмечаются иммунореактивные полосы в области 10 и 20 кДа, полученные со всеми антителами, узнающими различные эпитопы в предшественнике гепсидина. (Используемые маркеры молекулярной массы: фосфорилаза В, 105 кДа; глутаматдегидрогеназа, 53 кДа; карбоангидраза, 34 кДа; миоглобин синий, 23 кДа; миоглобин красный, 17 кДа; лизоцим, 13 кДа; апротинин, 7 кДа; инсулин, 3 кДа).
Фиг.3 иллюстрирует определение гепсидина в клетках HepG2 с помощью флуоресцентной микроскопии с применением антител EG(1)-HepN (A), EG(2)-HepN (В) и EG(1)-HepC (С) (Масштаб отрезка 8 мкм).
Фиг.4 иллюстрирует клеточную локализацию гепсидина в печени морской свинки (A-F) и человека (G-I). Парафиновые срезы при иммуноокрашивании антителами, специфичными к разным областям EG(1)-HepN (A, D, G), EG(2)-HepN (В, Е, Н) и EG(1)-HepC (С, F, I), проявляют отчетливую иммунореактивность в базолатеральном домене мембраны гепатоцитов (стрелки). (Увеличение: А-С, X 180; D-I, X 540).
На фиг.5 иллюстрируются иммуногистохимически обработанные срезы печени морской свинки (А, антитело EG(1)-HepN; В, антитело EG(2)-HepN; С, антитело EG(1)-HepC, проявляющие четкую зональность гепсидина в печеночных дольках, со снижением иммунореактивности от перипортальных зон (звездочки) к центральным венам (головки стрелок). Отмечается, что не найдено никакой иммунореактивности в гепатоцитах вокруг центральных вен. (Стрелка в В указывает на портальную триаду.) (А-С, X 180).
На фиг.6 иллюстрируются результаты ИФА для циркулирующего прогепсидина человека. Представлена характерная стандартная кривая с концентрациями гепсидина-(28-47) в нг/мл и экстинкцией раствора ИФА при длине волны 450 нм. Отмечается высокая разрешающая способность в диапазоне от 4 до 400 нг/мл гепсидина - (28-47).
На фиг.7 иллюстрируются нанесенные рамки величин венозных сывороточных концентраций прогепсидина у 2 6 здоровых добровольцев (контроль), 40 больных хронической почечной недостаточностью, 19 больных хронической почечной недостаточностью и почечной анемией и 35 больных наследуемым гемохроматозом. Линия в рамке указывает медиану, кружок указывает среднее. Нижний и верхний конец рамки указывает 1-й и 3-й квартиль, усы - минимальные и максимальные величины. Прерывистая линия отмечает средний уровень циркулирующего иммунореактивного прогепсидина у контрольной группы (106,16 нг/мл).
На фиг.8 иллюстрируется корреляция между прогепсидином и железом (А), ферритином (В) и насыщением трансферрина (С) в пробах больных, леченных и не леченных НН. Авторы отмечают, что в их пробах заметной корреляции не найдено.
На фиг.9 иллюстрируется полная нуклеотидная (SEQ ID NO:1) и аминокислотная последовательности (SEQ ID NO:2) одной формы гепсидина, воспроизведенные из базы данных GenBank nos. поступления NM021175 и ААН20612 соответственно.
Фиг.10 иллюстрирует следующее: (А): РТ-ПЦР анализ почки человека (линия 2), мыши (линия 3) и крысы (линия 4), показывающий экспрессию гена гепсидина. Указан лэддер п.н. ДНК (линии 1 и 5). (В, С): анализ гепсидина иммуноблоттингом в экстрактах почки человека (линии 1), крысы (линии 2) и мыши (линии 3), а также в моче человека (линии 4) с антителами EG(2)-HepN (В) и EG(1)-HepC (С). Отмечаются иммунореактивные полосы в области 9,5 кДа, полученные с обоими антителами, узнающими различные эпитопы в предшественнике гепсидина. (Используемые маркеры молекулярной массы: фосфорилаза В, 105 кДа; глутаматдегидрогеназа, 53 кДа; карбоангидраза, 34 кДа; миоглобин синий, 23 кДа; миоглобин красный, 17 кДа; лизоцим, 13 кДа; апротинин, 7 кДа; инсулин, 3 кДа).
Фиг.11 иллюстрирует клеточную локализацию гепсидина в коре почки крысы. Парафиновые срезы при иммуноокрашивании антителами, специфичными к разным областям, EG(1)-HepC (A), EG(2)-HepC (В), EG(1)-HepN (С) и EG(2)-HepN (D), проявляют отчетливую иммунореактивность в дистальных отделах трубочек коры почки. В некоторых трубочках иммунореактивность распределяется в цитоплазме эпителиальных клеток (стрелки), но в других иммунореактивность локализуется у апикального полюса соответствующих клеток (D, головки стрелок). Отмечается, что в клубочках (звездочки) отсутствует какая-либо иммунореактивность гепсидина. (Увеличение: А, X 90; B-D, X 180).
Фиг.12 иллюстрирует тканевое распределение гепсидина в почке крысы (А и С) и мыши (В и D). Иммуногистохимия с антителом EG(2)-HepN выявляет внешнюю зону мозгового слоя (А и В), с заметным снижением иммунореактивности гепсидина между наружной полоской (os) и внутренней полоской (is), что показано черными пунктирными дугами. С и D показывают отсутствие иммунореактивности гепсидина во внутреннем мозговом слое (m). Сильная иммунореактивность наблюдается в коре (с). (Увеличение: А, В и D, X 90; С, X 180.)
Фиг.13 иллюстрирует субклеточную локализацию гепсидина в почке крысы при иммуноокрашивании антителами EG(1)-HepN (A), EG(2)-HepN (В и D) и EG(1)-HepC (С). В некоторых дистальных трубочках иммунореактивность гепсидина распределяется в цитоплазме эпителиальных клеток (стрелки), но в других иммунореактивность интенсивно концентрируется в направлении апикального полюса соответствующих клеток (черные головки стрелок). Отмечается, что в клубочках (звездочки) и проксимальных трубочках (полые головки стрелок) отсутствует какая-либо иммунореактивность гепсидина. (Увеличение: A-D, X 360.)
Фиг.14 иллюстрирует клеточную локализацию гепсидина в почке человека. Антитела EG(1)-HepN (A), EG(2)-HepN (В и D) и EG(1)-HepC (С) выявляют отчетливую иммунореактивность в дистальных трубочках коры почки (стрелки). В тех же самых трубочках существуют различия внутриклеточного распределения иммунореактивности гепсидина, с наличием интенсивно (черные головки стрелок) и слабо (полые головки стрелок) иммунореактивных эпителиальных клеток с цитоплазматическим окрашиванием. В клубочках не наблюдается какой-либо иммунореактивности (звездочки). (Увеличение: А-С, X 180; D, X 360.)
Фиг.15 иллюстрирует определение иммунореактивности гепсидина на апикальном полюсе дистальных клеток трубочек почки человека антителами EG(1)-HepN (A), EG(2)-HepN (В и D) и EG(2)-Hep С (С). Отмечается интенсивная иммунореактивность на апикальном полюсе секреторных эпителиальных клеток (черные головки стрелок, в некоторых клетках отсутствует иммунореактивность гепсидина (полые головки стрелок). Звездочка указывает клубочек. (Увеличение: А-С, X 180; D, X 360.)
На фиг.16 иллюстрируются нанесенные рамки величин венозных сывороточных концентраций и концентраций в моче прогепсидина у 22 здоровых добровольцев (контроль) и 22 больных хронической почечной недостаточностью. Линия в рамке указывает медиану, а кружок указывает среднее. Нижний и верхний конец рамки указывает 1-й и 3-й квартиль, усы - минимальные и максимальные величины. Прерывистая линия отмечает средний уровень циркулирующего иммунореактивного прогепсидина у контрольной группы (104,2 нг/мл).
Лучший вариант осуществления изобретения
В настоящем изобретении описывается, что гепсидин регулирует захват железа клетками млекопитающих и нефизиологическая экспрессия гепсидина ведет к заболеванию с вовлечением нарушения метаболизма железа. Применяемый здесь термин гепсидин означает прогепсидин, гепсидин или его фрагменты. Физиологическая концентрация гепсидина в крови составляет в диапазоне приблизительно от 50 приблизительно до 150 нг/мл. Нефизиологические концентрации находятся ниже или выше данного диапазона. Нефизиологические количества белка гепсидина или его фрагмента связаны с нарушениями метаболизма железа, ведущими к дефициту железа или его избытку, такими как железодефицитная анемия; заболевания с генетическим или не генетическим избытком железа, такие как гемосидероз и гемохроматоз или вторичный гемохроматоз, ацерулоплазминемия, гипотрансферринемия, атрансферринемия; заболевания с избытком железа неопределенного происхождения, например, в случае заболеваний билиарной системы, болезней печени, особенно алкогольных заболеваний печени, неалкогольного стеатогепатоза и хронических инфекций при гепатите В и С; заболевания с нарушением утилизации железа, такие как сидеробластная анемия, талассемия; гематологические заболевания, такие как лейкоз, полиглобулия, макроцитарная, микроцитарная или нормоцитарная анемия, анемия с ретикулоцитозом, гемолитическая анемия; нарушения ретикулоэндотелиальной системы, обусловленные инфекциями и заболеваниями; воспаления и инфекции, включая сепсис; иммунологические заболевания и опухоли, такие как карцинома, саркома, лимфома, которые ведут к нефизиологическим концентрациям гепсидина; нейродегенеративные заболевания, такие как болезнь Альцгеймера и болезнь Вильсона. Данное открытие позволяет разработать тесты для определения белка гепсидина и его фрагментов и его последующую очистку с сохранением его нативной конфигурации и физиологической активности. Изобретение частично основано на открытии, что у больных, страдающих от определенных нарушений, в ткани, крови и жидкостях организма, будь то человек или животное, присутствует белок гепсидин.
В данном изобретении впервые показано, что белок гепсидин, включая прогепсидин, у больных с данными нарушениями присутствует в ткани, крови и жидкостях организма, будь то человек или животное, в концентрациях, существенно превышающих найденные у нормальных людей или животных, которые не подвержены данным заболеваниям. Это достигнуто путем тестирования образца ткани, крови или жидкости организма у больного и определения присутствия и количества белка гепсидина и/или прогепсидина. Определение и количественное измерение любого белка гепсидина, включая прогепсидин или его фрагмент, в ткани, крови или жидкостях организма в соответствии с данным изобретением пригодно для подтверждения клинического диагноза описанных здесь заболеваний у заболевших пациентов и прослеживания течения заболевания. Изобретение пригодно также для мониторинга заболевания в течение и после периода лечения агентами, которые тестируются на их способность стабилизировать, снижать или предотвращать наступление таких заболеваний.
Только в целях описания изобретение будет описано в плане (а) получения белка гепсидина, включая прогепсидин или его фрагменты; (b) получения антител, которые специфически связывают белок гепсидин, включая прогепсидин или его фрагменты; (с) диагностических тестов и наборов для диагностики подтипов или мониторинга описанных здесь заболеваний; (d) способов гиперэкспрессии и отрицательной регуляции гепсидина или прогепсидина и (е) терапевтического лечения описанных здесь заболеваний.
В одном аспекте изобретения заявители предлагают способ определения роли гепсидина в физиологических условиях и при соответствующих заболеваниях. В другом аспекте изобретения заявители предлагают специфические антитела против средней части и С-конца молекулы предшественника гепсидина. В данном аспекте изобретения эти антитела применяли для определения клеточной локализации гепсидина в печени человека и морской свинки. Создан чувствительный ИФА, который определяет прогепсидин в человеческой сыворотке у больных с НН, хронической почечной недостаточностью (CRI) и почечной анемией (RA). Авторы показали, что прогепсидин выходит в кровь через базолатеральную мембрану гепатоцитов и экскретируется почками. Так как сывороточные уровни гепсидина существенно негативно регулируются при НН и при хронической RA, гепсидин должен играть роль в патофизиологии данных заболеваний.
Получение белка гепсидина
Выделение белка гепсидина из крови и жидкостей организма
В целях согласно изобретению термин белок гепсидин определяется как любой полипептид гепсидин млекопитающих, имеющий приблизительно 80-процентную идентичность аминокислотной последовательности с предсказанной аминокислотной последовательностью, опубликованной Pigeon и соавторами ((2001) J. Biol. Chem. 276, 7811-7819). Предлагаемые здесь белки гепсидины включают прогепсидин, гепсидин и его фрагменты. Предлагаемые здесь белки гепсидины включают также белки, характеризуемые аминокислотными последовательностями, сходными с таковыми очищенных белков гепсидинов, но в которых есть природные или преднамеренно созданные модификации. Например, модификации в пептиде гепсидине или последовательностях ДНК могут быть сделаны специалистами в данной области с применением известных способов. Интересующие модификации в последовательности белка гепсидина могут включать в себя изменение, замену, замещение, вставку или делецию выбранного аминокислотного остатка в кодирующей последовательности. Например, один или более остатков цистеина могут быть удалены или заменены другой аминокислотой для изменения конформации молекулы. Способы вызвать такое изменение, замещение, замену, вставку или делецию хорошо известны специалистам в данной области (см., например, патент США №4518584). Предпочтительно, чтобы такое изменение, замещение, замена, вставка или делеция сохраняли желаемую активность белка. Области белка гепсидина, которые важны для функции белка, могут быть определены различными способами, известными в данной области, включая метод сканирования аланина, который включает систематическую замену одной или ряда аминокислот на аланин, с последующим тестированием полученного содержащего аланин варианта на биологическую активность. Данный тип анализа дает возможность оценить важность замененной(ных) аминокислоты(от) для биологической активности.
Получение белка гепсидина может быть осуществлено с помощью выделения белка гепсидина из ткани, крови или жидкостей организма человека или животных, страдающих гемохроматозом, железодефицитной анемией, гемосидерозом, циррозом печени и другими такими описанными здесь заболеваниями, с применением стандартных способов, известных специалистам в данной области. Такие способы, включенные в изобретение, также относятся к способам получения белка гепсидина, включая выращивание клеток-хозяев в культуре в подходящей культуральной среде и очистку белка гепсидина из клеток или из культуры, в которой растут клетки.
Различные способы, известные в данной области, могут быть использованы для получения любого из выделенных белков гепсидинов согласно изобретению. Например, белок гепсидин может быть также получен путем химического синтеза аминокислотной последовательности белка гепсидина (Pigeon et al., (2001) J. Biol. Chem. 276, 7811-7819) по предсказанной при клонировании и секвенировании кДНК, кодирующей белок гепсидин. Данная информация о последовательности белка гепсидина может быть использована для предсказания подходящей аминокислотной последовательности фрагмента белка гепсидина для химического синтеза с применением стандартных способов пептидного синтеза, известных в данной области. Данные способы включают в себя твердофазный метод, разработанный R. Bruce Merrifield, Erickson and Merrifield, «Solid-Phase Peptide Synthesis», in The Proteins, Volume 2, H.Neurath & R.Hill (Eds.) Academic Press, Inc., New York pp.255-257; Merrifield, (1986) «Solid phase synthesis», Science, 242:341-347). В твердофазном методе аминокислоты постепенно добавляют к растущей пептидной цепи, которая соединена с нерастворимым матриксом, таким как полистироловые шарики. Главное преимущество данного способа состоит в том, что желаемый продукт на каждой стадии связывается с шариками, которые могут быть быстро отфильтрованы и промыты, и, таким образом, устраняется необходимость очищать промежуточные продукты. Все реакции выполняют в одном сосуде, что исключает потери, обусловленные повторяющимися переносами продуктов. Данный твердофазный способ химического пептидного синтеза может быть легко автоматизирован, что делает его подходящим для рутинного синтеза пептидов, содержащих приблизительно 50 остатков, с хорошими выходом и степенью чистоты (Stewart and Young, (1984) Solid Phase Peptide Synthesis, 2nd ed., Pierce Chemical Co.; Tam et al., (1983) J. Am. Chem. Soc, 105: 6442). Например, может быть синтезирован фрагмент белка гепсидина, соответствующий аминокислотным остаткам с 1 по 50 или с 34 по 84, как изображено на фиг.9. На самом простом уровне для получения небольших пептидов и фрагментов белка гепсидина особенно пригоден имеющийся в продаже аппарат для синтеза пептидов. Фрагменты применяют, например, для выработки антител против природного белка гепсидина.
Специалист в данной области может легко следовать известным способам выделения белков, с тем чтобы получить один из выделенных белков/пептидов гепсидина согласно изобретению. Они включают в себя, не ограничиваясь этим, иммунохроматографию, ВЭЖХ, исключающую по размеру хроматографию и иммуноаффинную хроматографию. См., например, Scopes, Protein Purification: Principles and Practice, Springer-Verlag (1994); Sambrook, et al., in Molecular Cloning: A Laboratory Manual; Ausubel et al., Current Protocols in Molecular Biology.
Наконец, для дополнительной очистки белка гепсидина может быть использована одна или более стадий высокоэффективной жидкостной хроматографии с обращенной фазой (ОФ-ВЭЖХ) с применением гидрофобных сред для ОФ-ВЭЖХ, например силикагеля, имеющего дополнительный метил или другие алифатические группы. Некоторые или все из указанных выше стадий очистки в различных сочетаниях могут быть также применены для обеспечения по существу гомогенного выделенного белка гепсидина. Очищенный таким образом белок гепсидин является по существу свободным от других белков млекопитающих и определяется в соответствии с настоящим изобретением как выделенный белок.
Последовательность белка гепсидина может быть идентифицирована с применением метода деградации белковой последовательности по Эдману. В данном методе последовательно за один цикл удаляется один аминокислотный остаток с аминоконца пептида для последующей идентификации последовательности с помощью хроматографических способов. См., например, способы, описанные в Konigsberg and Steinman, (1977) Strategy and Methods of Sequence Analysis, in Neurath and Hill (eds.), The Proteins (3rd ed.) Vol.3, pp.1-178, Academic Press. Кроме того, анализ последовательности белка гепсидина может быть ускорен при применении жидкофазного автоматического аминокислотного секвенатора в соответствии с описанным способом (Hewick et al., (1981) J. Biol. Chem., 256:7990-7997; Stein and Undefriend, (1984) Analy. Chem., 136:7-23), что позволяет тем самым анализировать пикомолярные количества белка гепсидина.
Очищенный белок гепсидин может быть применен в тестах связывания in vitro, которые хорошо известны в данной области, для идентификации молекул, которые связывают белок гепсидин. Данные молекулы включают в себя, не ограничиваясь этим, например, небольшие молекулы, молекулы из комбинаторных библиотек, антитела или другие белки. Молекулы, идентифицированные в тесте связывания, затем тестируют на предмет активности агониста или антагониста в тканевой культуре in vitro или на животных моделях, которые хорошо известны в данной области. Вкратце, молекулы титруют во множестве клеточных культур или на животных и затем тестируют, повлияет ли это на смерть клетки/животного или продлит выживаемость клеток/животного.
Кроме того, связывающие молекулы могут комплексироваться с токсинами, например рицином или токсинами холеры, или с другими соединениями, которые являются токсичными для клетки. Комплекс связывающей молекулы с токсином затем направленно доставляется к опухоли или другой клетке за счет специфичности связывания молекулы с белком гепсидином.
Клонирование и экспрессия рекомбинантного белка гепсидина
В других осуществлениях получение белка гепсидина может быть достигнуто с помощью технологии рекомбинантной ДНК. Например, подходящие нуклеотидные последовательности, кодирующие белок гепсидин, могут быть синтезированы, клонированы и экспрессированы в подходящих клетках-хозяевах. Так как последовательность ДНК, кодирующая белок гепсидин, известна (Pigeon et al., (2001) J. Biol. Chem. 276, 7811-7819), зонды ДНК могут быть синтезированы с помощью стандартных способов, известных в данной области, для скрининга библиотек кДНК, полученных из печеночной ткани человека или животных, страдающих гемохроматозом, железодефицитной анемией, гемосидерозом, циррозом печени и другими описанными здесь заболеваниями, на специфические кДНК белка гепсидина. Данные зонды кДНК могут быть дополнительно использованы для выделения полного семейства генов белка гепсидина из данных библиотек кДНК с применением способов, которые хорошо известны специалистам в данной области. См., например, способы, описанные в Maniatis et al., (1982) Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory, N.Y., Chapter 7.
Методы гибридизации пригодны для скрининга рекомбинантных клонов с применением меченых смешанных синтетических олигонуклеотидных зондов, где каждый зонд является потенциально полностью комплементарным специфической последовательности ДНК в гибридизуемом образце, который включает гетерогенную смесь денатурированной двуспиральной ДНК. Для такого скрининга гибридизацию предпочтительно выполняют либо на односпиральной ДНК, либо на денатурированной двуспиральной ДНК. При применении строгих условий гибридизации с целью избежания неспецифического связывания возможно, например, учитывать авторадиографическую визуализацию специфического клона ДНК путем гибридизации ДНК-мишени с таким единичным зондом в смеси, который является полностью ей комплементарным (Wallace, et al., (1981) Nucleic Acids Research, 9:879).
Альтернативно, экспрессионную библиотеку можно подвергнуть скринингу не прямо на белок гепсидин согласно изобретению, имеющий, по меньшей мере, один эпитоп, а с применением антител к белку. Такие антитела могут быть как поликлональными, так и моноклональными и использоваться для определения продукта экспрессии, указывая на присутствие белка гепсидина. Обычно конструируется библиотека лямбда gtll и проводится иммунологический скрининг в соответствии со способом Huynh, et al., (1985) (in DNA Cloning: A Practical Approach, D.M.Glover, ed., 1:49).
Создание специфических последовательностей ДНК, кодирующих белок гепсидин, может быть осуществлено также с помощью: (1) выделения последовательности двуспиральной ДНК из геномной ДНК и (2) химического получения последовательности ДНК для обеспечения необходимыми кодонами для интересующего белка.
Способ полимеразной цепной реакции (ПЦР) может быть применен для амплификации индивидуальных членов семейства гепсидина для последующего клонирования и экспрессии кДНК белков гепсидинов (см., например, патенты США №№4683202; 4683195; 4889818; Gyllensten et al., (1988) Proc. Nat′l Acad. Sci. USA, 85:7652-7656; Ochman et al., (1988) Genetics, 120:621-623; Triglia et al., (1988) Nucl. Acids. Res., 16:8156; Frohman et al., (1988) Proc. Nat′l Acad. Sci. USA, 85:8998-9002; Loh et al., (1989) Science, 243:217-220).
Для конструирования экспрессионных векторов, содержащих кодирующие последовательности белка гепсидина или его фрагментов и подходящие сигналы, контролирующие транскрипцию/трансляцию, могут быть применены методы, которые хорошо известны специалистам в данной области. Данные методы включают способы рекомбинантной ДНК in vitro, способы синтеза и способы рекомбинации in vivo/генетической рекомбинации. См., например, способы, описанные в Maniatis et al., 1982, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory, N.Y., Chapter 12.
Разнообразные системы хозяев экспрессионных векторов могут быть применены для экспрессии белка гепсидина или его фрагмента. Они включают, но не ограничиваются этим, микроорганизмы, такие как бактерии, трансформированные экспрессионными векторами рекомбинантной ДНК бактериофага, плазмидной ДНК или космидной ДНК, содержащими кодирующую последовательность белка гепсидина или его фрагментов; дрожжи, трансформированные рекомбинантными дрожжевыми экспрессионными векторами, содержащими кодирующую последовательность белка гепсидина или его фрагмента; клеточные системы насекомых, инфицированные рекомбинантными вирусными экспрессионными векторами (например, бакуловирусом), содержащими кодирующую последовательность белка гепсидина или его фрагмента; или системы животных клеток, инфицированные рекомбинантными вирусными экспрессионными векторами (например, аденовирусом, вирусом коровьей оспы), содержащими кодирующую последовательность белка гепсидина или его фрагмента.
Экспрессионные элементы данных векторов варьируются по их количеству и специфичности. В зависимости от применяемых хозяина/векторной системы любое количество подходящих транскрипционных и трансляционных элементов, включая конститутивные и индуцибельные промоторы, может быть применено в экспрессионном векторе. Например, при клонировании в бактериальных системах могут быть применены индуцибельные промоторы, такие как pL бактериофага лямбда, plac, ptrp, ptac (ptrp-lac гибридный промотор) и тому подобное; при клонировании в клеточных системах насекомых могут быть применены такие промоторы, как бакуловирусный полигедриновый промотор; при клонировании в клеточных системах млекопитающих могут быть применены такие промоторы, как поздний аденовирусный промотор или промотор вируса коровьей оспы 7,5К. Промоторы, получаемые с помощью рекомбинантной ДНК или синтетических способов, могут также быть применены для обеспечения транскрипции вставленной кодирующей последовательности белка гепсидина или его фрагмента.
В дрожжах может быть применен ряд векторов, содержащих конститутивные или индуцибельные промоторы. В качестве обзоров см. Current Protocols in Molecular Biology, Vol.2, (1988) Ed. Ausubel et al., Greene Publish. Assoc. & Wiley Interscience Ch. 13; Grant et al., (1987) Expression and Secretion Vectors for Yeast, in Methods in Enzymology, Eds. Wu & Grossman, (1987) Acad. Press, N.Y., Vol.153, hh.516-544; Glover, (1986) DNA Cloning, Vol.II, IRL Press, Wash., D.C. Ch.3; и Bitter, (1987) Heterologous Gene Expression in Yeast, Methods in Enzymology, Eds. Berger & Kimmel, Acad. Press, N.Y., Vol.152, pp.673-684 и The Molecular Biology of the Yeast Saccharomyces, (1982) Eds. Strathern et al., Cold Spring Harbor Press, Vols. I and II. Для тестов комплементарности в дрожжах кДНК белков гепсидинов или их фрагментов могут быть клонированы в эписомальные плазмиды дрожжей (Yep), которые автономно реплицируются в дрожжах из-за присутствия в дрожжах 2 мкм кольцевой плазмиды. Последовательность белка гепсидина или его фрагмента может быть клонирована ниже, либо конститутивного промотора дрожжей, такого как ADH или LEU2, либо индуцибельного промотора, такого как GAL (Cloning in Yeast, Ch. 3, R. Rothstein (1986) In DNA Cloning Vol.11, A Practical Approach, Ed. DM Glover, IRL Press, Wash., D.C). Конструкты могут содержать 5′ и 3′ нетранслируемые области родственной мРНК белка гепсидина или те, которые соответствуют гену дрожжей. Плазмиды YEp трансформируют с высокой эффективностью, и плазмиды являются крайне стабильными. Альтернативно, могут быть применены векторы, которые стимулируют интеграцию чужеродных последовательностей ДНК в хромосому дрожжей.
Особенно хорошей экспрессионной системой, которая может быть использована для экспрессии белка гепсидина или его фрагментов, является система насекомых. В одной такой системе Autographa californica nuclear polyhedrosis virus (AcNPV) применяют в качестве вектора для экспрессии чужеродных генов. Вирус растет в клетках Spodoptera frugiperda. Кодирующая последовательность белка гепсидина или его фрагмента может быть клонирована в несущественные области (например, в ген полигедрина) вируса и помещена под контроль промотора AcNPV (например, промотор полигедрина). Успешная вставка в ген полигедрина ведет к продукции незакрытого рекомбинантного вируса (т.е. вируса, не имеющего белковой оболочки, кодируемой геном полигедрина). Данные рекомбинантные вирусы затем используют для инфицирования клеток Spodoptera frugiperda, в которых вставленный ген экспрессируется. (См., например, Smith et al., (1983) J. Biol., 46:586; Smith, патент США №4215051.) Кроме того, материалы и методы для систем экспрессии бакуловируса/клеток насекомых имеются в продаже в виде наборов, например, от Invitrogen, San Diego, Calif., U.S.A. (the MaxBat™ kit), и такие методы хорошо известны в данной области, как описано в Summers and Smith, Texas Agricultural Experiment Station Bulletin No. 1555 (1987), включенном здесь в качестве ссылки. В применяемом здесь смысле клетка насекомых, способная экспрессировать полинуклеотид гепсидина согласно изобретению, является трансформированной.
В случаях, когда в качестве экспрессионного вектора применяют аденовирус, кодирующая последовательность белка гепсидина или его фрагмента может быть лигирована с аденовирусным комплексом, контролирующим транскрипцию/трансляцию, например с поздним промотором и тройной лидирующей последовательностью. Данный химерный ген может быть затем вставлен в аденовирусный геном путем рекомбинации in vivo или in vitro. Вставка в несущественную область вирусного генома (например, область Е1 или Е3) будет приводить к рекомбинантному вирусу, который является жизнеспособным и способным экспрессировать белок гепсидин или его фрагмент в инфицированных хозяевах. (См., например, Logan & Shenk, (1984) Proc. Natl. Acad. Sci., (USA) 81:3655-3659.) Альтернативно, может быть применен промотор 7,5К вируса коровьей оспы (см., например, Mackett et al., (1982) Proc. Natl. Acad. Sci., (USA) 79:7415-7419; Mackett et al., (1984) J. Virol., 49:857-864; Panicali et al. (1982) Proc. Natl. Acad. Sci., 79:4927-4931).
Для эффективной трансляции вставленных кодирующих последовательностей белка гепсидина или его фрагмента могут также потребоваться специфические сигналы инициации. Данные сигналы включают инициирующий кодон ATG и прилегающие последовательности. В тех случаях, когда полный геном белка гепсидина, включая его собственные кодон-инициации и прилегающие последовательности, вставляют в подходящие экспрессионные векторы, дополнительные сигналы контроля трансляции могут не понадобиться. Однако в тех случаях, когда вставляется только часть кодирующей последовательности белка гепсидина, должны быть предоставлены экзогенные сигналы контроля трансляции, включая инициирующий кодон ATG. Более того, инициирующий кодон должен быть в фазе с рамкой считывания кодирующей последовательности белка гепсидина или его фрагмента для уверенности в трансляции полной вставки. Данные экзогенные сигналы контроля трансляции и инициирующие кодоны могут быть различного происхождения, как природного, так и синтетического. Эффективность экспрессии может быть увеличена путем включения подходящих усиливающих транскрипцию элементов, терминаторов транскрипции и т.п. (см. Bitter et al., (1987) Methods in Enzymol., 153:516-544).
Кроме того, может быть выбран штамм клеток-хозяев, который модулирует экспрессию вставленных последовательностей или модифицирует и осуществляет процессинг продукта гена конкретным желаемым образом. Экспрессия, управляемая определенными промоторами, может быть увеличена в присутствии определенных индукторов (например, ионов цинка и кадмия для металлотионеиновых промоторов). Следовательно, экспрессия сконструированного генетически белка гепсидина или его фрагмента может контролироваться. Это важно, если белковый продукт клонированного чужеродного гена является летальным для клеток-хозяев. Более того, модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут быть важными для функции белка. Различные клетки-хозяева имеют характерные и специфические механизмы для посттрансляционного процессинга и модификации белков. Подходящие клеточные линии или системы хозяев могут быть выбраны для того, чтобы убедиться в правильности модификации и процессинга экспрессируемого чужеродного белка.
Клетки-хозяева, которые содержат кодирующую последовательность белка гепсидина или его фрагмента и которые экспрессируют биологически активный генный продукт белка гепсидина или его фрагмента, могут быть идентифицированы с помощью, по меньшей мере, четырех общих подходов: (а) гибридизацией ДНК-ДНК; (b) по присутствию или отсутствию «маркерных» генных функций; (с) при оценке уровня транскрипции, измеренной по экспрессии транскриптов мРНК белка гепсидина в клетках-хозяевах; и (d) при определении генных продуктов белка гепсидина при измерении с помощью иммунных анализов или по их биологической активности.
При первом подходе наличие вставленной в экспрессионный вектор кодирующей последовательности белка гепсидина или его фрагмента может быть выявлено с помощью ДНК-ДНК гибридизации с применением зондов, включающих нуклеотидные последовательности, которые гомологичны последовательности, кодирующей белок гепсидин или его отдельные части, по существу, как описано ранее (Pigeon et al., (2001) J. Biol. Chem. 276, 7811-7819).
При втором подходе рекомбинантный экспрессионный вектор/система хозяина могут быть идентифицированы и отобраны на основе наличия или отсутствия определенных "маркерных" функций гена (например, тимидинкиназной активности, устойчивости к антибиотикам, устойчивости к метотрексату, фенотипу трансформации, образования замкнутых телец в бакуловирусе и т.д.). Например, если кодирующая последовательность белка гепсидина или его фрагмента вставлена в последовательность маркерного гена вектора, рекомбинанты, содержащие кодирующую последовательность белка гепсидина или его фрагмента, могут быть идентифицированы по отсутствию функций маркерного гена. Альтернативно, маркерный ген может быть помещен в виде тандема с кодирующей последовательностью белка гепсидина или его фрагмента под контролем того же самого или другого промотора, применяемого для контроля экспрессии кодирующей гепсидин последовательности. Экспрессия маркера в ответ на индукцию или селекцию указывает на экспрессию кодирующей последовательности белка гепсидина.
При третьем подходе транскрипционная активность в отношении области, кодирующей белок гепсидин или его фрагмент, может быть оценена с помощью тестов гибридизации. Например, РНК может быть выделена и проанализирована с помощью нозерн-блоттинга с применением зонда, гомологичного кодирующей последовательности белка гепсидина или его фрагмента или ее отдельным частям, по существу, как описано Pigeon et al., ((2001) J. Biol. Chem. 276, 7811-7819). Альтернативно, суммарные нуклеиновые кислоты клетки-хозяина могут быть проэкстрагированы и проанализированы для гибридизации с такими зондами.
При четвертом подходе экспрессия белкового продукта гепсидина или его фрагмента может быть оценена иммунологически, например, с помощью иммуноблоттинга, иммунологических анализов, таких как радиоиммунопреципитация, иммуноферментные анализы и тому подобное.
При обнаружении того, что рекомбинант экспрессирует белок гепсидин или его фрагмент, следует проанализировать продукт гена. Это может быть осуществлено с помощью тестов, основанных на физических, иммунологических или функциональных свойствах продукта. Например, способы изобретения включают способ получения белка гепсидина, при котором клетка-хозяин, содержащая подходящий экспрессионный вектор, который включает гепсидиновый полинуклеотид согласно изобретению, культивируется при условиях, которые обеспечивают экспрессию кодируемого белка гепсидина. Белок гепсидин может быть выявлен в культуре, обычно в культуральной среде или в лизате, полученном из клеток-хозяев и дополнительно очищенном. Предпочтительные осуществления включают те, в которых белок, полученный с помощью таких способов, является полноразмерным или представляет собой зрелую форму белка.
В настоящем изобретении дополнительно предлагается выделенный белок гепсидин, кодируемый фрагментами нуклеиновой кислоты согласно изобретению или вырожденными вариантами фрагментов нуклеиновой кислоты согласно изобретению. Под "вырожденным вариантом" подразумеваются нуклеотидные фрагменты, которые отличаются от фрагмента нуклеиновой кислоты согласно изобретению (например, ORF) по нуклеотидной последовательности, но, благодаря вырожденности генетического кода, кодируют идентичную белковую последовательность. Предпочтительными фрагментами нуклеиновой кислоты согласно изобретению являются Orbs, которые кодируют белки.
Белок гепсидин согласно изобретению может быть альтернативно очищен из клеток, которые были изменены для экспрессии белка гепсидина. Как применяется здесь, клетка изменена для экспрессии желаемого полипептида или белка, когда посредством генетического манипулирования создается клетка для продукции белка гепсидина, которая обычно его не продуцирует или которая обычно продуцирует его на низком уровне. Специалист в данной области может легко адаптировать процедуры для введения и экспрессии либо рекомбинантных, либо синтетических последовательностей в эукариотные или прокариотные клетки для создания клетки, которая продуцирует белок гепсидин согласно изобретению.
Белок гепсидин согласно изобретению может быть также экспрессирован в виде продукта трансгенных животных, например в виде компонента молока трансгенных коров, коз, свиней или овец, которые отличаются соматическими или половыми клетками, содержащими нуклеотидную последовательность, кодирующую белок гепсидин.
Белок гепсидин может быть также получен известным традиционным химическим синтезом. Способы конструирования белка гепсидина согласно изобретению с помощью синтетических способов известны специалистам в данной области. Сконструированные синтетически последовательности белка гепсидина путем приведения первичных, вторичных или третичных структурных и/или конформационных свойств в соответствие с природным белком гепсидином могут обладать биологическими свойствами, общими для обоих, включая активность белка. Таким образом, они могут быть применены в качестве активных или иммунологических замен вместо природного, очищенного белка гепсидина при скрининге терапевтических соединений и при иммунологических способах выработки антител.
Белок гепсидин согласно изобретению может быть получен путем культивирования трансформированных клеток-хозяев при условиях культивирования, пригодных для экспрессии рекомбинантного белка. Полученный экспрессированный белок гепсидин может быть затем очищен из такой культуры (т.е. из культуральной среды или клеточных экстрактов) с помощью известных способов очистки, таких как гель-фильтрация и ионообменная хроматография. Очистка белка гепсидина может также включать аффинную колонку, содержащую агенты, которые должны связывать белок/одну или более дополнительных колоночных стадий, таких как аффинные смолы, такие как конкавалин А-агароза, гепарин-toyopearl™ или сефароза Cibacrom blue 3GA™; одну или более стадий, включающих хроматографию с гидрофобным взаимодействием с применением таких смол, как фениловый эфир, бутиловый эфир или пропиловый эфир; или аффинную хроматографию.
Альтернативно, белок гепсидин согласно изобретению может быть также экспрессирован в форме, которая должна облегчать очистку. Например, он может быть экспрессирован в виде гибридного белка, такого как гибриды с такими, как белок, связывающий мальтозу (МВР), глутатион-S-трансфераза (GST) или тиоредоксин (TRX), или в виде свободного конца с His. Имеются в продаже наборы для экспрессии и очистки таких гибридных белков от New England BioLab (Beverly, Mass.), Pharmacia (Piskataway, N.J.) и Invitrogen соответственно. Белок гепсидин может быть также снабжен эпитопом и затем очищен с применением специфичного антитела, направленного против такого эпитопа. Один такой эпитоп ("FLAG®") имеется в продаже от Kodak (New Haven, Conn.).
Другие фрагменты и производные последовательностей белков/пептидов гепсидина, которые, как ожидается, могли бы сохранять активность белка целиком или частично (например, связывание с рецептором TfR2, связывание с антителом, специфичным в отношении гепсидина и т.д.) и которые пригодны для скрининга или других иммунологических способов, также могут быть легко получены специалистами в данной области на основании представленных здесь раскрытий. Такие модификации охватываются настоящим изобретением.
Белок гепсидин или его фрагмент должен быть иммунореактивным независимо от того, получен ли он в результате экспрессии полной последовательности гена, части последовательности гена или двух или более последовательностей гена, которые лигированы для обеспечения продукции гибридных белков. Данная иммунореактивность может быть показана стандартными иммунологическими способами, такими как радиоиммунопреципитация, радиоиммунная конкуренция или иммуноблоттинг.
Получение антител, которые выявляют белок гепсидин или его фрагмент
В данной области известны различные способы, которые могут быть применены для получения антител против эпитопов средней части (аминокислоты с 20 по 50) или С-конца (аминокислоты с 65 по 84) белка гепсидина. Специфичные в отношении гепсидина антитела связывают данные эпитопы, но не другие известные последовательности. Такие антитела включают, но не ограничиваются этим, поликлональные, моноклональные, гибридные, одноцепочечные, Fab фрагменты и экспрессионную библиотеку Fab. Для получения антител различные животные-хозяева могут быть иммунизированы путем инъекции конкретного белка гепсидина или синтетического белка гепсидина, включая, но не ограничиваясь этим, кроликов, мышей, крыс и т.д. Для повышения иммунологического ответа в зависимости от вида хозяина могут быть применены различные адъюванты, включая, но не ограничиваясь этим, адъювант Фрейнда (полный или неполный), минеральные гели, такие как гидроксид алюминия, поверхностно активные вещества, такие как лизолецитин, плюроновые полиолы, полианионы, пептиды, масляные эмульсии, гемоцианин моллюска фиссуреллии, динитрофенол и потенциально пригодные для человека адъюванты, такие как BCG (bacille Calmette-Guerin) и corynebacterium parvum.
Поликлональные антитела могут быть легко получены специалистом в данной области от множества теплокровных животных, таких как лошади, коровы, различные домашние птицы, кролики, мыши или крысы. Вкратце, для иммунизации животного применяют гепсидин посредством внутрибрюшинных, внутримышечных, внутриглазных или подкожных инъекций и адъювант, такой как полный или неполный адъювант Фрейнда. После нескольких бустерных иммунизаций отбирают образцы сыворотки и тестируют на взаимодействие с гепсидином. Особенно предпочтительная поликлональная антисыворотка должна давать сигнал при одном из данных тестов, который, по меньшей мере, в три раза превышает фоновый сигнал. После достижения плато титра у животного в плане взаимодействия с гепсидином большие количества антисыворотки могут быть легко получены либо путем еженедельного отбора крови, либо путем обескровливания животного.
Моноклональные антитела против пептидов гепсидина могут быть получены с помощью любого способа, который обеспечивает получение молекул антитела постоянными линиями клеток в культуре. Это включает, но не ограничивается этим, способ гибридомы, исходно описанный Kohler and Milstein (Nature, (1975) 256:495-497), более поздний способ гибридомы В-клеток человека (Kosbor et al., (1983) Immunology Today, 4:72) и способ EBV-гибридомы (Cole et al., (1985) Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp.77-96). В дополнительном осуществлении изобретения моноклональные антитела, специфичные в отношении белков/пептидов гепсидина, могут быть получены у стерильных животных с помощью современного способа (патент PCT/US90/02545). Согласно изобретению могут быть применены антитела человека, и они могут быть получены с помощью гибридом человека (Cote et al., (1983) Proc. Natl. Acad. Sci., 80:2026-2030) или путем трансформации В-клеток человека вирусом EBV in vitro (Cole et al., (1985) in Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, pp.77-96). Действительно, согласно изобретению могут быть применены способы, разработанные для получения "гибридных антител" (Morrison et al., (1984) Proc. Natl. Acad. Sci., 81:6851-6855; Neuberger et al., (1984) Nature, 312:604-608; Takeda et al., (1985) Nature, 314:452-454), путем сплайсинга генов из молекулы антитела мыши с соответствующей антигенной специфичностью с генами из молекулы антитела человека с подходящей биологической активностью; такие антитела охватываются данным изобретением.
Согласно изобретению для получения специфичных в отношении белка гепсидина одноцепочечных антител могут быть адаптированы способы, описанные для получения одноцепочечных антител (патент США №4946778).
В дополнительном осуществлении изобретения применяют способы, описанные для конструирования экспрессионных библиотек Fab (Huse et al., (1989) Science, 246:1275-1281) для обеспечения быстрой и простой идентификации моноклональных Fab-фрагментов с желаемой специфичностью в отношении белков/пептидов гепсидина.
Фрагменты антител, которые содержат специфичные сайты связывания для белка гепсидина, могу быть получены с помощью известных способов. Например, такие фрагменты включают, но не ограничиваются этим: F(ab′)2 фрагменты, которые могут быть получены путем гидролиза пепсином молекулы антитела, и Fab фрагменты, которые могут быть получены восстановлением дисульфидных мостиков F(ab′)2 фрагментов.
Диагностические тесты и наборы
Еще одной целью согласно изобретению является предложение реактивов для применения в диагностических тестах для определения белка гепсидина у индивидуумов, страдающих гемохроматозом, железодефицитной анемией, гемосидерозом, циррозом печени и другими подобными им описанными здесь заболеваниями.
В одном способе данного осуществления белок гепсидин согласно изобретению может быть применен в качестве антигена в иммуноанализе для определения тех индивидуумов, которые страдают гемохроматозом, железодефицитной анемией, гемосидерозом, циррозом печени и другими подобными им описанными здесь заболеваниями. Белок, полипептид и/или пептид гепсидина согласно изобретению может быть применен в любой системе иммуноанализа, известной в данной области, включая, но не ограничиваясь этим, радиоиммуноанализы, иммуноферментный анализ, "сэндвич"-методы, реакции с преципитином, тесты иммунодиффузии в геле, тесты агглютинации, иммунофлуоресцентные тесты, иммунотесты с протеином А и иммуноэлектрофоретические тесты, перечисляя некоторые из них. Подходящие тесты описаны также в патенте США №4629783 и цитированных в нем патентах.
Согласно настоящему изобретению, моноклональные или поликлональные антитела, выработанные к различным формам белка гепсидина, могут быть применены для иммуноанализа образцов крови, спинномозговой жидкости или другой жидкости организма для выявления субъектов с гемохроматозом, железодефицитной анемией, гемосидерозом, циррозом печени и другими описанными здесь заболеваниями.
В одном осуществлении изобретения образец крови берут у больного из вены и приводят в контакт с антикоагулянтом, таким как ЭДТА, перемешивают, центрифугируют при 600g в течение 10 мин и плазму удаляют, как это обычно делают в данной области, или образец спинномозговой жидкости берут у больного с помощью люмбальной пункции.
Описанные здесь антитела могут быть применены в качестве основных реагентов в ряде различных иммуноанализов для определения присутствия белка гепсидина в образце ткани, крови или жидкости организма. Вообще говоря, антитела могут быть применены в любом типе иммуноанализа, будет ли он качественным или количественным. Они включают как сэндвич-метод с двумя сайтами, так и иммуноанализ с одним сайтом неконкурентного типа, а также традиционные конкурентные связывающие анализы.
Особенно предпочтительным из-за простоты определения и количественной природы является сэндвич-метод или метод двойных антител, для которого существует ряд вариаций, все из которых предназначены для охвата настоящим изобретением.
Например, при типичном прямом сэндвич-методе немеченое антитело иммобилизуется на твердом субстрате, например на лунках планшета для микротитрования, и тестируемый образец приводят в контакт со связанной молекулой. После подходящего периода инкубации в течение периода времени, достаточного для возможности образования двойного комплекса антитело-антиген, затем добавляют второе антитело, меченное репортерной молекулой, способной индуцировать определяемый сигнал, и инкубацию продолжают, давая достаточное время для связывания с антигеном через отличный сайт и для образования тройного комплекса антитело-антиген-меченое антитело. Любой непрореагировавший материал отмывается, и присутствие антигена определяется по наблюдению сигнала, который может быть измерен количественно с помощью сравнения с контрольным образцом, содержащим известные количества антигена. Вариации прямого сэндвич-метода включают одновременное определение, при котором как образец, так и антитело добавляют одновременно к связанному антителу, или обратный сэндвич-метод, при котором меченое антитело и тестируемый образец сначала объединяют, инкубируют и добавляют к немеченому, связанному с поверхностью антителу. Данные способы хорошо известны специалистам в данной области, и возможность небольших вариаций должна быть вполне очевидной. Применяемый здесь «сэндвич-метод» предназначен для охвата всех вариаций основного способа с двумя сайтами.
Для сэндвич-методов согласно изобретению лимитирующим фактором является только то, что оба антитела имеют разную специфичность связывания белка гепсидина. Таким образом, допустим ряд возможных вариаций.
В качестве более конкретного примера в типичном прямом сэндвич-методе первичное антитело либо ковалентно, либо пассивно связывается с твердым носителем. Твердая поверхность является обычно стеклянной или полимерной, причем наиболее часто применяемыми полимерами является целлюлоза, полиакриламид, нейлон, полистирол, поливинилхлорид или полипропилен. Твердые носители могут быть в форме пробирок, шариков, дисков или микропланшетов или любыми другими поверхностями, подходящими для проведения иммуноанализа. Процессы связывания хорошо известны в данной области. После связывания комплекс твердая фаза-антитело отмывают при подготовке для тестируемого образца. Аликвоту жидкости организма, содержащую тестируемый белок гепсидин, затем добавляют к твердофазному комплексу и инкубируют при 25°С в течение периода времени, достаточного для обеспечения связывания любого белка гепсидина, представленного антителу, специфичному в отношении белка гепсидина. Затем к комплексу с твердой фазой добавляют второе антитело и инкубируют при 25°С в течение дополнительного периода времени, достаточного для обеспечения связывания второго антитела с первым комплексом антиген-антитело на твердой фазе. Второе антитело связано с репортерной молекулой, видимый сигнал которой используют как показатель связывания второго антитела с любым антигеном в образце. Под «репортерной молекулой», как применяется в настоящем описании, подразумевается молекула, которая по своей химической структуре обеспечивает аналитически определяемый сигнал, который позволяет определить антиген, связанный с антителом. Определение должно быть, по меньшей мере, относительно количественным для возможности определения количества антигена в образце, это может быть рассчитано в абсолютных величинах или может быть сделано в сравнении со стандартом (или сериями стандартов), содержащим известный нормальный уровень антигена.
Наиболее обычно применяемыми репортерными молекулами в данном типе анализа являются либо ферменты, либо флуорофоры. В случае иммуноферментного анализа фермент конъюгируют со вторым антителом, часто посредством глутаральдегида или периодата. Однако, как легко понять, существует широкое разнообразие способов конъюгации, которые хорошо известны специалистам в данной области. Обычно применяемые ферменты включают среди прочих пероксидазу хрена, глюкозоксидазу, р-галактозидазу и щелочную фосфатазу. Субстраты, применяемые со специфическими ферментами, обычно выбираются для получения при гидролизе соответствующим ферментом определяемого изменения цвета. Например, п-нитрофенилфосфат подходит для применения с конъюгатами щелочной фосфатазы; для конъюгатов пероксидазы обычно применяют 1,2-фенилендиамин или толуидин. Возможно также применять флуорогенные субстраты, которые дают флуоресцирующий продукт в отличие от хромогенных субстратов, указанных выше. Во всех случаях меченное ферментом антитело добавляют к первому комплексу антитело-белок гепсидин и дают связаться с комплексом, а затем избыток реагента отмывают. Раствор, содержащий подходящий субстрат, затем добавляют к тройному комплексу антитело-антиген-меченое антитело. Субстрат взаимодействует с ферментом, связанным со вторым антителом, позволяя качественно оценить видимый сигнал, который может быть далее оценен количественно, обычно спектрофотометрически, что дает оценку количества антигена, который присутствует в образце сыворотки.
Альтернативно, флуоресцентные соединения, такие как флуоресцеин или родамин, могут быть химически присоединены к антителам без изменения их связывающей способности. При активации под действием освещения светом определенной длины волны антитело, меченное флуорохромом, поглощает энергию света, индуцируя в молекуле состояние возбудимости, с последующим излучением света при характерной большей длине волны. Излучение проявляется в виде характерного цвета, определяемого визуально с помощью светового микроскопа. Как и в иммуноферментном анализе (ИФА), флуоресцентно меченному антителу дают связаться с первым комплексом антитело-белок гепсидин. После отмывания не связавшегося реагента оставшийся тройной комплекс затем подвергают действию света с подходящей длиной волны, и наблюдаемая флуоресценция указывает на присутствие антигена. Способы ИФА и иммунофлуоресценции оба очень хорошо разработаны в данной области и особенно предпочтительны для настоящего способа. Однако могут быть также применены другие репортерные молекулы, такие как радиоизотопы, хемилюминисцентные или биолюминисцентные молекулы. Специалисту в данной области должно быть вполне понятно, как варьировать процедуру для соответствия требуемому применению.
Альтернативно, тестируемый образец либо крови человека, либо спинномозговой жидкости, содержащих белок гепсидин, можно использовать в иммуноанализе с одним сайтом, где он связывается с твердым субстратом либо ковалентно, либо не ковалентно. Немеченое антитело против белка гепсидина приводят в контакт с образцом, связанным с твердым субстратом. После подходящего периода инкубации в течение периода времени, достаточного для возможности образования двойного комплекса антитело-антиген, добавляют второе антитело, меченное репортерной молекулой, способной индуцировать определяемый сигнал, и инкубацию продолжают, давая пройти достаточному времени для образования тройного комплекса антиген-антитело-меченое антитело. В иммуноанализе с одним сайтом второе антитело может быть обычным антителом (т.е. ксеногенным антителом к иммуноглобулину, особенно анти-(IgM и IgG), соединенным с репортерной молекулой), которое способно связывать антитело, специфичное к интересующему белку гепсидину.
Ген гепсидина (мутированный или нормальный) может быть применен в тесте метаболизма железа. Ген экспрессируется с или без какой-либо сопровождающей молекулы в клеточных линиях или первичных клетках, происходящих от человека или животного, здоровых субъектов или клетках из других организмов (таких как грызуны, насекомые, бактерии, амфибии и т.п.). Измеряется захват железа данными клетками, например, с помощью применения радиоактивных изотопов. Далее, может быть также измерено связывание железа к продукту гена гепсидина. Такие эксперименты помогают в оценке роли гена гепсидина и генного продукта гепсидина в захвате железа, связывании и транспорте клетками и в клетках.
Терапевтическое применение
В одном аспекте изобретения диагностические способы и наборы могут быть применены для генетических технологических подходов, таких как для гиперэкспрессии или негативной регуляции гепсидина. При определенных терапевтических применениях желательно негативно регулировать экспрессию и/или функцию гена гепсидина, мутантного гена гепсидина, белка гепсидина или мутантного белка гепсидина. Например, негативная регуляция нормального гена гепсидина или нормального белка гепсидина желательна в таких ситуациях, когда железо недостаточно аккумулируется в организме, например при определенных анемиях (т.е. талассемиях, гемолитических анемиях, трансфузиях). С другой стороны, негативная регуляция мутантного гена гепсидина или белка гепсидина желательна в таких ситуациях, когда железо избыточно аккумулируется в организме.
Как обсуждалось выше, могут быть получены антитела, специфичные по отношению к нормальному или мутантному белку гепсидину. Такие антитела могут быть использованы терапевтически при описанных здесь заболеваниях. Например, для блокирования действия мутантного или нормального гена гепсидина, если функция, связанная с мутантным белком, заключается в положительной регуляции функции нормального белка гепсидина и ведет к избыточной аккумуляции железа в организме. Сходно, антитела могут быть использованы терапевтически для блокирования действия белка гепсидина, который вызывает понижение аккумуляции железа в организме.
Сходным образом ген гепсидина, либо нормальный, либо в мутантной форме, может отрицательно регулироваться путем применения антисмысловых олигонуклеотидов, направленных против гена или его транскриптов. Сходная стратегия может быть применена, как обсуждалось выше, в связи с антителами. В качестве особенно ценного обзора дизайна анализа и применения антисмысловых олигонуклеотидов см. Uhlmann et al., (1990) Chemical Reviews 90:543-584, раскрытие которых включены здесь в качестве ссылки. Антисмысловые олигонуклеотиды согласно изобретению могут быть синтезированы с помощью любых известных способов химического синтеза олигонуклеотидов. Такие способы обычно описаны, например, в Winnacker Chirurg (1992) 63:145. Антисмысловые олигонуклеотиды наиболее выгодно получать с помощью применения любого имеющегося в продаже автоматического синтезатора нуклеиновых кислот. В одном таком приборе, Applied Biosystems 380 В DNA Synthesizer, применяют бета-цианоэтилфосфорамидитную химию.
Так как известен полный нуклеотидный синтез ДНК, комплементарной гену гепсидина, также известен транскрипт мРНК последовательности кДНК. Как таковые антисмысловые олигонуклеотиды, гибридизуемые с любой частью таких транскриптов, могут быть получены с помощью методов олигонуклеотидного синтеза, известных специалистам в данной области. Несмотря на то что в практике изобретения может быть использован олигонуклеотид любой длины, последовательности короче 12 оснований могут быть менее специфичными в отношении гибридизации с мРНК-мишенями, могут легче разрушаться при ферментативном гидролизе и могут дестабилизироваться при ферментативном гидролизе. Следовательно, олигонуклеотиды, имеющие 12 или более нуклеотидов, являются предпочтительными. Длинные последовательности, особенно последовательности длиннее приблизительно 40 нуклеотидов, могут быть несколько менее эффективными в плане ингибирования трансляции из-за пониженного захвата клеткой-мишенью. Таким образом, предпочтительными являются олигомеры из 12-40 нуклеотидов, более предпочтительными из 15-30 нуклеотидов, наиболее предпочтительными из 18-26 нуклеотидов. Последовательности из 18-24 нуклеотидов являются особенно и наиболее предпочтительными.
В еще одном аспекте изобретения гепсидин может быть применен для терапии описанных здесь нарушений с помощью лечения субъектов гепсидином и агонистами или антагонистами гепсидина. Захват железа клетками может модулироваться путем варьирования концентрации гепсидина и/или ингибирования связывания гепсидина с железом или с рецептором трансферрина. Соответственно, гепсидин и агонисты или антагонисты гепсидина могут быть пригодны для лечения состояний, при которых существует нарушение метаболизма железа. Например, такие вещества могут быть пригодны для лечения таких состояний, как гемохроматоз, нейродегенеративные заболевания, ишемическое повреждение ткани, включая ишемический удар или травму, заболевание сердца и опухоли, в частности рак кожи, и другие такие заболевания, описанные выше.
Изобретение также охватывает способы модуляции метаболизма железа с применением гепсидина. В частности, настоящее изобретение относится к способу лечения состояний, связанных с нарушениями метаболизма железа, включающему введение, модулирующее уровень железа, количество гепсидина или стимулятора, агониста или антагониста гепсидина. Состояния, связанные с нарушениями метаболизма железа, которые можно лечить с применением способа изобретения, включают, в виде примера, гемохроматоз, нейродегенеративные заболевания, ишемическое повреждение ткани, включая ишемический удар или травму, заболевание сердца и опухоли, в частности рак кожи, и другие такие заболевания, описанные выше. Вещество, которое является агонистом или антагонистом гепсидина, может быть идентифицировано путем определения эффекта вещества на связывающую активность гепсидина и железа или гепсидина и трансферриновых рецепторов TfR1 или TfR2, или эффекта вещества на экспрессию гепсидина в клетках, способных экспрессировать гепсидин, включая клетки, сконструированные генетически для экспрессии гепсидина на их поверхности.
Изобретение, следовательно, в одном аспекте относится к способу идентификации агонистов или антагонистов гепсидина, включающему взаимодействие вещества, предположительно являющегося агонистом или антагонистом гепсидина, с гепсидином и железом в таких условиях, когда гепсидин способен связываться с железом; измерение количества гепсидина, связанного с железом; и определение эффекта вещества путем сравнения количества гепсидина, связанного с железом, с количеством, определенным в контроле. Изобретение также относится к способу идентификации агонистов или антагонистов гепсидина, включающему взаимодействие вещества, предположительно являющегося агонистом или антагонистом гепсидина, с гепсидином и рецептором трансферрина в таких условиях, когда гепсидин способен связываться с рецептором трансферрина; измерение количества гепсидина, связанного с рецептором трансферрина; и определение эффекта вещества путем сравнения количества гепсидина, связанного с рецептором трансферрина, с количеством, определенным в контроле.
Изобретение также относится к способу идентификации агонистов или антагонистов гепсидина, включающему взаимодействие вещества, предположительно являющегося агонистом или антагонистом гепсидина, с клеткой, которая продуцирует гепсидин; измерение количества гепсидина, экспрессируемого клеткой, и определение эффекта вещества путем сравнения уровня экспрессии гепсидина с уровнем, определенным в контроле. Изобретение дополнительно относится к способу идентификации агониста или антагониста опосредуемого гепсидином захвата железа, включающему: инкубацию клетки, экспрессирующей гепсидин на своей поверхности, и вещества, предположительно являющегося агонистом или антагонистом гепсидина, в присутствии железа и в отсутствие трансферрина, измерение уровня захвата железа клеткой и идентификацию агониста или антагониста опосредуемого гепсидином захвата железа путем сравнения уровня опосредуемого гепсидином захвата железа клеткой с уровнем захвата железа клеткой при контрольной инкубации в отсутствие вещества.
В некоторых осуществлениях изобретения пептиды гепсидина предлагаются для терапевтического применения у субъектов с симптомами заболевания с первичным избытком железа или с таким синдромом, как гемохроматоз, или другое состояние с избытком железа, вызываемое вторичными причинами, такими как повторяющиеся трансфузии. Пептид гепсидин может быть полноразмерным гепсидином или неким фрагментом гепсидина. Предпочтительно, чтобы пептид гепсидин включал аминокислотные остатки с 28 по 47 или с 70 по 80 гепсидина. Предсказанная аминокислотная последовательность и последовательности геномной и кДНК гепсидина предлагаются в Krause et al., (2000) FEBS Lett. 480, 147-150; Pigeon et al., (2001) J. Biol. Chem. 276, 7811-7819, включенные здесь полностью в качестве ссылки. Белок гепсидин или его фрагмент может быть введен с бета-2-микроглобулином, таким как в форме комплекса. В некоторых осуществлениях белок гепсидин, больший, чем приблизительно из 20 аминокислот, вводят в комплексе с бета-2-микроглобулином.
В некоторых осуществлениях изобретения предлагаются агонисты или антагонисты белка гепсидина или рецептора трансферрина. Агонисты полипептида гепсидина и/или антагонисты рецептора трансферрина применимы, например, для лечения заболеваний или синдромов с первичным или вторичным избытком железа, в то время как антагонисты полипептида гепсидина или агонисты рецептора трансферрина пригодны, например, для лечения состояний с дефицитом железа, таких как анемии. В других осуществлениях предлагаются мутантные белки/пептиды гепсидина, которые функционируют как антагонисты белка гепсидина дикого типа. Антагонисты или агонисты могут также быть антителами, направленными против рецептора трансферрина или средней части (аминокислоты с 20 по 50) или С-концевой области (аминокислоты с 65 по 84) белка гепсидина. В некоторых осуществлениях изобретения полипептиды гепсидина могут служить в качестве антагонистов рецептора трансферрина. В дополнительных осуществлениях изобретения с применением способов, хорошо известных в данной области, в качестве антагонистов или агонистов белка гепсидина и/или рецептора трансферрина могут быть разработаны пептидомиметики.
Скрининг лигандов рецептора трансферрина, являются ли они антагонистами или агонистами, может быть проведен с применением описанных здесь способов по способности связываться с рецептором трансферрина. Дополнительно, может быть определена конкуренция за связывание гепсидина с применением способов, хорошо известных в данной области. Скрининг лигандов или, в более общем виде, партнеров по связыванию с белком гепсидином может быть проведен, например, по способности ингибировать комплексирование полипептида гепсидина с бета-2-микроглобулином с применением описанных здесь способов.
В некоторых осуществлениях изобретения агонисты или антагонисты трансферрина сходно применяются для увеличения или снижения количества железа, транспортируемого в клетку, такую как гепатоциты или лимфоциты больного. Например, эффективность лекарства, терапевтического агента, агониста или антагониста может быть идентифицирована по программе скрининга, в которой модуляция прослеживается в клеточных системах in vitro. Клеточные системы-хозяева, которые экспрессируют различные мутантные белки/пептиды гепсидина, подходят для применения в качестве первичных систем скрининга. Кандидатные лекарства могут быть оценены путем инкубации с данными клетками и измерения клеточных функций, зависящих от гена гепсидина, или путем измерения присущих белку гепсидину укладки и процессинга. Такие тесты могут также повлечь за собой измерение рецептороподобной активности, транспорта и метаболизма железа, транскрипции гена или другой выше или ниже лежащей биологической функции, как это предписывается исследованиями функции гена гепсидина.
Альтернативно, могут быть применены бесклеточные системы. Очищенный белок гепсидин может быть воспроизведен в искусственных мембранах или везикулах и скрининг лекарств проведен в бесклеточной системе. Такие системы являются часто более удобными и по существу более поддающимися высокопроизводительным типам скрининга и автоматизации.
Критерии определения чистоты белка гепсидина включают те, которые являются стандартными в области белковой химии. Они включают определение N-концевой аминокислоты, одно- или двумерный электрофорез в полиакриламидном геле и окрашивание серебром. Очищенный белок пригоден для применения в исследованиях, относящихся к определению вторичной и третичной структуры, а также для помощи в дизайне лекарства и для исследования in vitro биологической функции молекулы.
В некоторых осуществлениях изобретения могут быть разработаны лекарства для модуляции гена гепсидина и активности белка гепсидина, исходя из знаний по структурным и функциональным корреляциям известного белка гепсидина. Для этого рациональный дизайн лекарства при применении рентгеноструктурной кристаллографии, компьютерного моделирования (САММ), качественных и количественных взаимоотношений между структурой и активностью (GSAR) и сходных способов может дополнительно сконцентрировать попытки разработки лекарства. Рациональный дизайн позволяет предсказать структуру белка или синтетического пептида, которые могут взаимодействовать с белком гепсидином и модифицировать его активность. Такие структуры могут быть синтезированы химически или экспрессированы в биологических системах. Данный подход рассмотрен в обзоре Capsey et al., Genetically Engineered Human Therapeutic Drugs, Stockton Press, New York (1988). Кроме того, комбинаторные библиотеки могут быть созданы, синтезированы и применены в программах скрининга.
Для того чтобы вводить терапевтические агенты, основанные на настоящем изобретении или происходящие из него, должно быть понятно, что для обеспечения улучшенного транспорта, доставки, толерантности и тому подобное в составы могут быть включены подходящие носители, наполнители и другие агенты.
Множество подходящих составов может быть найдено в своде составов, известных всем химикам фармакологам: Remington's Pharmaceutical Sciences, (15th Edition, Mack Publishing Company, Easton, Pa. (1975)), особенно глава 87, Blaug, Seymour. Данные составы включают, например, порошки, пасты, мази, желе, парафины, липиды, безводные поглощающие основания, эмульсии масла в воде или воды в масле, эмульсии твердых полиэтиленгликолей (полиэтиленгликоли различной молекулярной массы), полутвердые гели и полутвердые смеси, содержащие твердый полиэтиленгликоль.
Для лечения и терапии согласно настоящему изобретению может подходить любой из указанных выше составов, предлагаемый так, чтобы активный агент в составе не инактивировался под действием состава и состав был физиологически совместимым.
Изобретение не ограничено описанными здесь осуществлениями и может быть модифицировано или изменено без выхода за пределы объема изобретения.
Примеры
Пример 1: Экспрессия гепсидина в печени человека
Ткани и получение ткани.
Образцы печени человека (n=7), использованные в настоящем исследовании, были получены после гемигепатэктомии взрослых субъектов с метастазами печени. Здоровые ткани фиксировали в 4% параформальдегиде для иммуногистохимии или немедленно замораживали в жидком азоте для ОТ-ПЦР, иммуноблоттинга и иммунофлуоресцентного анализа.
Морских свинок (n=7) и мышей (n=S) анестезировали и затем забивали обезглавливанием. Образцы ткани печени, скелетной мышцы и сердца отделяли и немедленно замораживали в жидком азоте для анализа иммуноблоттингом или фиксировали в параформальдегиде.
Синтез пептидов, процедура иммунизации и антитела.
На основании опубликованной последовательности прогепсидина (Krause et al., (2000) FEBS Lett. 489, 147-150; Pigeon et al., (2001) J. Biol. Chem. 276, 7811-7819) были синтезированы пептиды гепсидин-(28-47) и гепсидин-(70-84) в виде С-концевых амидов с применением стандартного Fmoc протокола (Cetin et al., (1994), Proc. Natl. Acad. Sci. USA 91, 2935-2939). Пептиды конъюгировали с гемоцианином моллюска фиссуреллии с помощью м-малеимидбензоил-N-гидроксисукцинимидного эфира и двух кроликов SPF (Charles River Iffa Credo) иммунизировали каждым из пептидных конъюгатов (Eurogentec, Seraing, Бельгия). После титрования с помощью ИФА в настоящем исследовании были применены три антисыворотки [EG(1)-HepC, направленная против гепсидина-(70-84), и EG(1)-HepN и EG(2)-HepN, каждая из которых направлена против гепсидина-(28-47), (фиг.1), (гепсидин-28-47: PQQ TGQ LAE LQP QDR AGA RA SEQ. (SEQ ID NO:3), гепсидин-70-84: CGC CHR SKC GMC CKT (SEQ ID NO:4)). Примененные для получения антисыворотки пептидные эпитопы не обладали гомологией с каким-либо сообщавшимся до настоящего времени белком, как это было подтверждено с помощью поиска BLAST P2.
Антитело BT-TFR21 S против TfR2 мыши (BioTrend, Cologne, Германия) было получено против цитоплазматического N-конца TfR2-альфа (TfR2) мыши, который подвергается альтернативному сплайсингу с образованием альфа- и бета-изоформ, см. Fleming et al., (2000) Proc. Natl. Acad. Sci. USA 97, 2214-2219), с 68% гомологией последовательности, соответствующей области TfR2-альфа человека. Антитело получали у кроликов и очищали аффинным способом.
Экспрессионный анализ в печени человека.
Выделение РНК проводили с применением набора Qiagen RNA easy, включая гидролиз ДНК. Анализ обратная транскрипция (ОТ)-ПЦР проводили, как описано ранее (Kulaksiz et al., (2002) Proc. Natl. Acad. Sci. USA 99, 6796-6801; Kulaksiz et al., (2002) Am. J. Pathol. 161, 655-664), с применением следующих праймеров и описаний, приводимых в 5-3′ ориентации: гепсидин человека (база данных GenBank, регистрационный no. NM0211175), 5′-CTG САА ССС CAG GAC AGA G-3′ (SEQ ID NO 5) и 5′-GGA АТА ААТ AAG GAA GGG AGG TG-3′ (SEQ ID NO 6), соответствующие положениям нуклеотидов 147-165 и 338-316. TfR2 человека (#AF067864), 5′-GAT TCA GGG TCA GGG AGG GG-3′ (SEQ ID NO 7) и 5′-(GAA GGG GCT GTG ATT GAA GG-3′ (SEQ ID NO 8); соответствующие положениям нуклеотидов 2496-2515 и 2694-2675. После первоначальной денатурации при 94°С в течение 4 мин реакционные смеси подвергали 35 циклам следующей температурной программы: 94°С в течение 30 сек, 60°С в течение 30 сек и 72°С в течение 30 сек; данную программу завершали конечной стадией элонгации в течение 5 мин при 72°С. Продукты амплификации разгоняли на окрашиваемом этидия бромидом 1,8% агарозном геле в 89 мМ трис/8 9 мМ борная кислота/2 мМ ЭДТА (рН 8,3). Амплификацию значительного уровня геномной ДНК исключали с помощью соответствующих контролей.
Экспрессионный анализ в клетках HEPG2.
Клетки гепатомы человека HepG2 получали от Германской коллекции микроорганизмов и клеточных культур (Braunschweig, Германия) и выращивали при 37°С в 5% СО2 в среде RPMI 1640 (Gibco, Karlsruhe, Германия) с добавкой 10% (об./об.) инактивированной нагреванием FBS, пенициллина (100 единиц/мл) и стрептомицина (100 единиц/мл). Клетки анализировали с помощью ОТ-ПЦР с применением упомянутых выше описаний праймеров. Для иммунофлуоресцентной микроскопии клетки HepG2 выращивали на стеклянных пластинах, фиксировали при 4 мм в метаноле и пермеабилизировали с помощью 0,5% Тритона Х-100 в ЗФР. После инкубации с гепсидином (1:2000) и антителами TfR2 (1:1000) в течение 60 мин с последующей инкубацией с Cy-3-конъюгированным антителом против кролика (Dianova, Hamburg, Германия) производили исследования иммунологического окрашивания под микроскопом Olympus AX70 с применением соответствующих фильтров.
Экстракция гепсидина и TfR2 из сыворотки, тканей и клеток HepG2.
В виде более богатого источника гепсидина заявители использовали сыворотку, полученную от больных с хронической почечной недостаточностью. Для экстракции гепсидина 20-мл образцы сыворотки разбавляли 1:1 0,01 н. HCl и доводили рН до 3,0 концентрированной HCl. Замороженные ткани и клетки HepG2 перемешивали в 0,5 М уксусной кислоте и кипятили в течение 8 мин, как описано Cetin et al. (1994) Proc Natl Acad Sci USA 91, 2935-2939; Cetin et al. (1995) Proc Natl Acad Sci USA 92, 5925-5929. После гомогенизации с помощью гомогенизатора Ultra-Turrax (Janke & Kunkel, Staufen, Германия) образцы центрифугировали при 20000g в течение 20 мин при 4°С и супернатанты фильтровали через фильтр с размером пор 0,45-мкм. Для обогащения белка образцы сыворотки, экстракты клеток и цельно ткани наносили на картридж октадекасилила (С18) Sep-Pak (Waters, Massachusetts). Колонку промывали 0,01 М HCl и элюировали смесью 30% (об./об.) 2-пропанол/30% (об./об.) метанол/0,01 М HCl (Cetin et al. (1994) Proc Natl Acad Sci USA 91, 2935-2939). Белковые фракции лиофилизовали и хранили при -80°С до применения. Для анализа TfR2 ткани и клетки гомогенизировали в трис-HCl буфере, содержащем 100 мМ NaCl, 50 мМ трис-HCl, рН 7,4, 10% глицерина, 1% тритона Х-100, 2 мг/мл леупептина, 2 мг/мл пепстатина и 1 мМ фенилметилсульфонилфторида, и центрифугировали при 10000g в течение 30 мин при 4°С.
Анализ иммуноблоттингом.
Для анализа иммуноблоттингом белковые экстракты инкубировали в течение 7 мин при 94°С в буфере для образцов с 4% (мас./об.) ДДС-Na (Merck, Darmstadt, Германия), 50 мМ трис-HCl (рН 8,15), 1 мМ ЭДТА, 3,24 мМ дитиотреитом (Roth, Karlsruhe, Германия), 12,5% (мас./об.) глицерином (Merck) и 0,002% бромфеноловым синим (Merck). Для выявления гепсидина применяли 16,5% трицин-ДДС-Na-полиакриламидный гель согласно опубликованным протоколам (Cetin et al., (1994) Proc. Natl. Acad. Sci. USA 91, 2935-2939; Kulaksiz et al., (2002) Proc. Natl. Acad. Sci. USA 99, 6796-6801; Kulaksiz et al., (2002) Am. J. Pathol. 161, 655-664; Cetin et al., (1995) Proc. Natl. Acad. Sci. USA 92, 5925-5929). Иммуноблоттинг TfR2 проводили с применением 8% ДДС-Na-полиакриламидных гелей. После электрофореза белки переносили на гидрофобные мембраны на основе поливинилиденфторида (Pall, Portsmouth, Англия) полусухим блоттингом. Мембраны инкубировали в течение ночи с антителами против гепсидина или TfR2 в указанных выше разведениях. После промывки забуференным трис солевым раствором, содержащим 10 мМ трис-HCl (рН 8,0), 150 мМ NaCl и 0,05% твин 20, соответствующие иммунореактивные белки визуализировали после инкубации с конъюгированным с щелочной фосфатазой антителом козы против кролика (разведение 1:50000; Sigma) с применением нирросинего тетразолия и 5-бром-4-хлор-3-индолилфосфата в качестве хромогенов (Sigma). Иммунную реакцию при иммуноблоттинге специфично блокировали после предварительной инкубации антител с соответствующими пептидными иммуногенами. Перекрестное взаимодействие со вторым антителом козы против кролика исключали с помощью соответствующих контролей (Cetin et al. (1994) Proc Natl Acad Sci USA 91, 2935-2939; Kulaksiz et al. (2002) Proc Natl Acad Sci USA 99, 6796-6801; Kulaksiz et al. (2002) Am J Pathol 161, 655-664; Cetin et al. (1995) Proc Natl Acad Sci USA 92, 5925-5929).
Иммуногистохимия и иммунофлуоресценция.
Ткани фиксировали в 4% параформальдегиде в течение 18 час при 4°С. После обезвоживания в последовательных сменах этанола с разной концентрацией образцы заключали в парафин. Парафиновые срезы (5 мкм) подвергали иммунологическому окрашиванию на гепсидин (антитела EG(1)-HepN, EG(2)-HepN и EG(1)-HepC, каждое из которых разводили 1:2000) или TfR2 (антитело BT-TFR21-S, разведенное 1:1000) с помощью метода авидин-биотин-пероксидазного комплекса (ABC) и последовательности инкубации, описанных ранее (Kulaksiz et al., (2002) Proc. Natl. Acad. Sci. USA 99, 6796-6801; Kulaksiz et al., (2002) Am. J. Pathol. 161, 655-664). Срезы инкубировали с соответствующими антителами в течение 24 час при 4°С с последующей инкубацией с биотинилированным IgG против кролика (Jackson Iimunoresearch, West Grove, PA, США) в течение 30 мин, разведенным 1:200. Срезы затем инкубировали в течение 30 мин с предварительно образованным комплексом биотин-пероксидаза/стрептавидин (Jackson Immunoresearch), разведенным в ЗФР (конечные концентрации: биотин-пероксидаза, 0,7 мкг/мл; стрептавидин, 5 мкг/мл). Сайты связывания антиген-антитело визуализировали инкубацией срезов в смеси 07 мМ диаминобензидина гидрохлорида/0,002% Н2О2 в 0,05 М трис-HCl (рН 7,6).
Для иммунофлуоресцентной микроскопии получали срезы ткани печени человека (2-4 мкм) с помощью криотома (FrigoCut 2800E; Leica, Nussloch, Германия), сушили на воздухе в течение 2 часов и фиксировали в течение 10 мин в холодном ацетоне (-20°С). Двойное иммунофлуоресцентное промечивание производили, как описано ранее (Rost et al., (1999) Hepatology 29, 814-821), с применением специфичных антител против гепсидина (разведенных 1:1000) и моноклонального антитела С219 (то же), направленного против канальцевых Р-гликопротеинов (Gentocor, Malvern, PA), разведенного 1:30. После инкубации с соответствующей антисывороткой производили окрашивание путем инкубации с Cy2-(1:200) и Cy3-(1:600) мечеными антителами против IgG мыши и кролика (Dianova, Hamburg, Германия). Микроизображения получали с помощью микроскопа Olympus AX7O, снабженного цифровой камерой (набор цветов 12, программа системы изображения SIS, Munster, Германия) и программой анализа (SIS, Munster, Германия).
Контроли специфичности.
Зависимые от способа неспецифичные величины исключали с помощью контролей, как было описано (Cetin et al., (1994) Proc. Natl. Acad. Sci. USA 91, 2935-2939; Cetin et al., (1995) Proc. Natl. Acad. Sci. USA 92, 5925-5929). Специфичность антител тестировали путем предварительной адсорбции антител гомологичными или гетерологичными антигенными пептидами (6,25-100 мкг/мл антисыворотки) (Kulaksiz et al., (2002) Proc. Natl. Acad. Sci. USA 99, 6796-6801; Kulaksiz et al., (2002) Am. J. Pathol. 161, 655-664). Предварительная адсорбция антител гомологичными антигенами в такой низкой концентрации, как 6,25 мкг/мл, полностью блокировала иммунологическое окрашивание в тканях и клетках печени, тогда как предварительная адсорбция антител гетерологичными антигенами в концентрации до 100 мкг/мл не оказывала влияния на иммунологическое окрашивание.
ИФА анализ гепсидина с помощью конкурентного связывания.
Образцы сыворотки получали от 26 здоровых индивидуумов (13 женщин, 13 мужчин в возрасте 26-64, средний возраст 43 года), от 35 больных с НН, гомозиготных в отношении мутации C282Y в HFE (14 женщин, 21 мужчина в возрасте 23-82 года, средний возраст 54 года), с (15 больных) и без (20 больных) терапией кровопусканием и от 59 больных с почечной недостаточностью, подвергавшихся хроническому гемодиализу (33 женщины, 26 мужчин в возрасте 26-96 лет, средний возраст 57 лет). При взятии крови были предприняты меры против инфицирования больных. У 19 больных из группы с почечной недостаточностью была почечная анемия, отличавшаяся максимальным уровнем гемоглобина 11 г/дл. Все больные с хронической почечной недостаточностью подвергались лечению 2-3 раза в неделю 3000 IE рекомбинантного эритропоэтина человека (ЕРО). Отбирали 10 мл образцы крови в охлажденные на льду пробирки для сыворотки и центрифугировали при 2500g в течение 10 мин при 4°С. Измерения проводили в двух параллельных определениях с применением 96-луночных планшетов для микротитрования, покрытых 200 мкл/лунку антителом кролика против гепсидина EG(2)-HepN, разбавленным 1:4000 в забуференном трис солевом растворе (TBS), содержащем 40 мМ трис-HCl (рН 7,3), 100 мМ NaCl, 50 мкл стандартов, содержащих различное количество синтетических пептидов (0, 20, 100, 500 и 1000 нг/мл), или образцы сыворотки человека, и в каждую лунку добавляли 150 мкл биотинилированного по N-концу гепсидина-(28-47) (Peptide Specialty Laboratories GmbH, Heidelberg, Германия) (2 нг/лунку) и инкубировали в течение 1 часа при комн. темп. После промывки TBST (TBS с 0,05% твином 20) комплексы биотинилированный антиген-антитело выявляли с помощью стрептавидин-пероксидазного фермента (Dako, Hamburg, Германия) с субстратом тетраметилбензидина (DRG Instruments GmbH, Marburg, Германия); цветную реакцию останавливали с помощью 1 М H2SO4 и считывали поглощение раствора при длине волны 450/630 нм.
Измеренные величины гепсидина для четырех интересующих групп были введены в таблицу EXCEL и оценены с помощью SAS WIN, версия 8.2. Измеренные величины были суммированы посредством следующей суммарной статистики путем группового диагноза: количество наблюдений, арифметическое среднее, стандартное отклонение, минимум, медиана и максимум. Возможные различия между группами анализировали с помощью парных U-тестов Вилькоксона. Уровень значимости α был установлен равным 5% (0,05). Корреляцию между прогепсидином и железом, ферритином и трансферрином анализировали с помощью ранговой корреляции Спирмана.
Экспрессия гепсидина и TfR2 в печени и клетках HEPG2.
Анализ с помощью ОТ-ПЦР показал, что гепсидин экспрессируется в печени человека (Gehrke et al. (2003) Blood MS#2002-11-3610.R2). Сходным образом, ожидаемый продукт ПЦР 192 п.н. был выявлен в клетках HepG2 (контроль), которые, как уже было показано, экспрессируют гепсидин (Pigeon С et al. (2001) J Biol Chem 276, 7811-7819; Gehrke et al. (2003) (фиг.2, A). Кроме того, анализ ОТ-ПЦР четко показал, что TfR2 экспрессируется в печени человека и клетках HepG2 (данные не показаны).
При иммуноблоттинге все антитела против гепсидина [EG(1)-HepN, EG(2)-HepN и EG(1)-HepC] одинаковым образом идентифицировали иммунореактивную полосу ~10 кДа в экстрактах печени человека и морской свинки. Данный пептид печени двигался совместно с иммунореактивной полосой, узнаваемой антителами против гепсидина, в гомогенатах клеток HepG2 (фиг.2, B-D). Все антитела выявляли также иммунореактивный белок ~20 кДа на всех дорожках, содержащих экстракты печени человека и морской свинки или экстракты клеток HepG2. Анализ иммуноблоттингом экстрактов скелетной мышцы (контроль) не выявил ни 10 кДа, ни 20 кДа иммунореактивных полос (фиг.2, B-D). Анализ с помощью иммуноблоттинга с применением антитела против TfR2 BT-TFR21-S привел к окрашиванию ожидаемого (Fleming et al., (2000) Proc. Natl. Acad. Sci. USA 97, 2214-2219) белка ~105 кДа в экстрактах печени мыши. В экстрактах печени человека и клеток HepG2 тем же антителом узнавались ~95 кДа иммунореактивный TfR2 и в меньшей степени ~105 кДа иммунореактивный белок (данные не показаны). Иммунореактивность не обнаружена в сердце (контрольная ткань).
Иммунофлуоресценция в клетках HepG2.
Экспрессию пептида гепсидина в клетках HepG2 исследовали с помощью иммунофлуоресцентного анализа с применением специфичных в отношении эпитопа антител против гепсидина. Все антитела сходным образом идентифицировали гепсидин в клетках HepG2, что отражалось в сильной иммунореактивности (фиг.3). В соответствии с клеточной локализацией гепсидина антитело против TfR2 выявляло TfR2 в тех же клетках (данные не показаны).
Клеточная и внутриклеточная локализация гепсидина и TfR2.
Иммуногистохимические исследования с помощью различных специфичных в отношении области антител постоянно устанавливали локализацию гепсидина в гепатоцитах в печени человека (фиг.4). Клетки Купфера, клетки эндотелия, желчные протоки и сосудистая система были полностью лишены иммунореактивности гепсидина. Те же антитела выявляли сильную иммунореактивность гепсидина также в печени морской свинки (фиг.4). Печеночные дольки были неоднородны в отношении иммунореактивности гепсидина: в пределах печеночной дольки иммунореактивные в отношении гепсидина клетки преимущественно располагались в перипортальной зоне, и частота позитивных в отношении гепсидина клеток постепенно снижалась от портальной триады в направлении к центральной вене (фиг.5). Примечательно, что между позитивными в отношении гепсидина клетками имелись различия: в то время как большая часть гепатоцитов была сильно позитивной в отношении гепсидина, другие проявляли лишь слабое окрашивание или были полностью неспособны к реакции на гепсидин (фиг.5). На внутриклеточном уровне иммунореактивность в отношении гепсидина была ограничена доменом базолатеральной (=синусоидальной) мембраны гепатоцитов согласно иммуногистохимии; иммунореактивность не обнаруживалась на домене апикальной мембраны соответствующих клеток (фиг.2). Сходным образом иммунофлуоресцентный анализ показал сильную иммунореактивность в отношении гепсидина на домене базолатеральной мембраны; иммунореактивность отсутствовала на домене апикальной мембраны, как это было обнаружено двойным окрашиванием с помощью антитела С219, направленного против канальцевых Р-гликопротеинов (Rost et al. (1999) Hepatology 29, 814-821 (данные не показаны).
В соответствии с локализацией гепсидина специфичное в отношении белка антитело BT-TFR21-S выявляло TfR2 в печени человека и мыши. На клеточном уровне TfR2 обнаруживался на базолатеральной мембране гепатоцитов при наличии различий между клетками, касающихся интенсивности иммунореактивности (данные не показаны). Неоднородность также наблюдалась в пределах печеночной дольки, с повышением иммунореактивности от центральных вен к портальным триадам.
Определение пропептида гепсидина в плазме человека.
Был разработан стабильный ИФА анализ прогепсидина (DRG Instruments GmbH, Marburg, Германия) с высокой воспроизводимостью и чувствительностью с применением специфичного в отношении N-конца гепсидина антитела EG(2)-HepN. Как видно на фиг.6, ИФА проявлял наибольшую разрешающую способность в диапазоне между 4 и 400 нг/мл, в котором проводили определение концентрации прогепсидина в сыворотке человека. В качестве контроля специфичности производили инкубацию ИФА с гетерологичными пептидами. При применении гетерологичных пептидов не наблюдалось перекрестной реактивности.
Наличие прогепсидина в крови было подтверждено с помощью анализа иммуноблоттингом. Все антитела против гепсидина идентифицировали в экстрактах сыворотки человека единственной иммунореактивной в отношении гепсидина полосы с молекулярной массой ~10 кДа, которая двигалась в точности так же, как и иммунореактивный гепсидин в тканях печени и экстрактах клеток HepG2 (фиг.2, B-D).
Свойства ИФА.
Чувствительность анализа составляла 3,95 нг/мл. Не было перекрытия с наименьшим стандартом (20 нг/мл). Последовательные разведения прогепсидина человека, разведенные в нулевом стандарте, шли параллельно со стандартной кривой ИФА прогепсидина с диапазоном открываемости между 90,6-111,6%. Открываемость, выраженная в процентах от ожидаемой концентрации, находилась между 91,8 и 105,7%. Была показана высокая точность (суммарный CV<10%) при трех концентрациях прогепсидина в пределах диапазона анализа.
Уровень прогепсидина при наследуемом гемохроматозе, хронической почечной недостаточности и почечной анемии.
С помощью чувствительного ИФА гепсидина в сыворотке здоровой контрольной группы из 26 добровольцев был определен прогепсидин в диапазоне 51,6-153,4 нг/мл (среднее±SE; 106,2±32,1 нг/мл) (фиг.7, таблица 1). У больных с НН концентрация прогепсидина составляла от 12,1 до 153,9 нг/мл сыворотки (среднее+SE; 70,2±38,1 нг/мл). Данная концентрация была значительно ниже по сравнению с контрольными субъектами (Р<0,05) (фиг.7, таблица 1). В сыворотке больных, страдающих CRI, концентрация прогепсидина варьировалась от 31,1 до 471,3 нг/мл (среднее±SE; 1481±88,0 нг/мл) и была значительно повышена по сравнению с контрольными субъектами (Р<0,01) и НН (Р<0,001). Напротив, уровень прогепсидина у больных RA с гемодиализом был значительно снижен (115,0±53,1 нг/мл; диапазон 20,5-252,4 нг/мл) (Р=0,05) по сравнению с больными CRI (фиг.7, табл.1).
Не было обнаружено значимой корреляции в наших образцах (сыворотке от НН, CRI и RA) между прогепсидином и насыщением железом, ферритином и трансферрином (фиг.8). Тест отличия от нуля показал отсутствие отличия.
Обсуждение
Анализ с помощью ОТ-ПЦР с применением специфичных праймеров показал, что гепсидин интенсивно экспрессируется в клетках HepG2 (контроль), хорошо дифференцированной гепатоцеллюлярной карциномной линии клеток (Aden et al. (1979) Nature 282, 615-616), проявляющих многие физиологические свойства нормальных гепатоцитов. При применении подходящих описаний праймеров и их сочетаний, успешно использованных на клетках HepG2, исследования ОТ-ПЦР подтвердили экспрессию гепсидина в печени человека. Три разных антитела, узнающие разные эпитопы в молекуле предшественника гепсидина, одновременно идентифицировали иммунореактивный пептид ~10 кДа при иммуноблоттинге не только в клетках HepG2, но и в экстрактах печени двух видов, человека и морской свинки. Кажущаяся молекулярная масса данного иммунореактивного пептида находится в соответствии с молекулярной массой, рассчитанной для прогормона гепсидина на основании последовательности кДНК (Pigeon С et al. (2001) J Biol Chem 276, 7811-7819) (фиг.1). Интересно, что с помощью всех антител против гепсидина в экстрактах клеток HepG2 и печени человека и морской свинки выявлялась вторая иммунореактивная полоса ~20 кДа, отсутствующая в контрольной ткани. Данный иммунореактивный белок может представлять собой димерный тип гепсидина. Действительно, в предшествующем исследовании для гепсидина-25, но не для гепсидина-20 была описана способность к агрегации и возможному образованию мультимеров (Hunter et al. (2002) J Biol Chem 277, 37597-37603).
Иммуноцитохимические исследования с применением антител, специфичных в отношении областей и молекулярных доменов гепсидина, выявили сильную иммунореактивность в клетках HepG2, что продемонстрировало экспрессию гепсидина в данных клетках, как это уже было показано с помощью молекулярно-биологических способов (Gehrke et al. (2003) Blood MS#2002-11-3610.R2). Иммуногистохимические и иммунофлуоресцентные исследования с данными различными антителами против гепсидина показали, что в печени человека и морской свинки гепсидин специфично локализован в гепатоцитах, расположенных преимущественно вокруг портальной триады. Одновременное окрашивание различными антителами, специфичными в отношении областей не только печени человека и морской свинки, но и клеток HepG2, указывает на то, что источником гепсидина являются гепатоциты. Иммунореактивность гепсидина снижается от перипортальных зон в направлении к центральным венам. Данное зональное распределение в пределах портальных долек может иметь функциональное значение, поскольку перипортальные гепатоциты обладают доступом к первичному прохождению через портальные вены обогащенной железом крови из кишечника. Примечательно, что между гепсидин-положительными клетками в отношении плотности иммунореактивности в отношении гепсидина имелись определенные межклеточные различия, что может отражать различия между клетками в экспрессии или секреции гепсидина.
На внутриклеточном уровне гепсидин концентрируется на домене базолатеральной мембраны гепатоцитов. Иммунореактивность не была обнаружена в домене апикальной мембраны. Паттерн дискретного распределения гепсидина на внутриклеточном уровне может указывать на направленную секрецию гепсидина в синусоиды печени. Данный направленный путь секреции дополнительно подтверждается выявлением прогормона гепсидина (фиг.1) в сыворотке человека (см. ниже); следовательно, эти данные служат дополнительным подтверждением того, что гепатоциты могут регулировать обмен железа эндокринным способом посредством секреции прогепсидина.
Для анализа экспрессии и клеточного распределения TfR2, a также соответствующих доменов-мишеней мембраны были проведены исследования с помощью иммуноблоттинга и иммуногистохимии на клеточном уровне. Как было показано в предшествующих исследованиях, ОТ-ПЦР показал, что TfR2 интенсивно экспрессируется в печени человека (Fleming et al., (2000) Proc. Natl. Acad. Sci. USA 97, 2214-2219). Наличие данного белка было подтверждено с помощью иммуноблоттинга с применением антитела BT-TFR21-S, специфичного в отношении TfR2 человека и мыши. В экстрактах печени мыши был выявлен иммунореактивный белок ~105 кДа; данная молекулярная масса иммунореактивного TfR2 несколько превышает ожидаемую массу 95 кДа (Fleming et al., (2000) Proc. Natl. Acad. Sci. USA 97, 2214-2219) и может отражать некоторые посттрансляционные модификации, как описано ранее (Kawabata et al., (2000) J. Biol. Chem. 275, 16618-16625). При идентичных условиях, однако, антитело против TfR2 идентифицировало в экстрактах печени человека белок с ожидаемой молекулярной массой 95 кДа и белок 105 кДа с меньшим сродством. Расхождение между иммуноблоттингом печени человека и мыши может быть обусловлено межвидовыми различиями.
Иммуногистохимические исследования показали, что TfR2 локализован в гепатоцитах печени человека и мыши; в соответствии с клеточной локализацией гепсидина специфичное в отношении белка антитело показало локализацию TfR2 исключительно на базолатеральной мембране. Данный тип специфичной для мембраны связи TfR2 свидетельствует, в частности, о базолатеральной активации TfR2, который участвует в метаболизме железа путем связывания содержащего два атома железа трансферрина и опосредования захвата связанного трансферрином железа гепатоцитами из крови (Philpott, С.С.(2002) Hepatology 35, 993-1001; Subramaniam et al., (2002) Cell Biochem. Biophys. 36, 235-239). Примечательно, что сходное с описанным для гепсидина зонирование в дольке наблюдалось для TfR2 со снижением иммунореактивности от перипортальных зон в направлении к центральным венам.
Поскольку взаимодействие между гепсидином и TfR2 на клеточном уровне было обсуждено в предшествующих исследованиях (Nicolas et al., (2001) Proc. Natl. Acad. Sci. USA 98, 8780-8785; Frazer et al., (2002) Gastroenterology 123, 835-844), был проведен анализ совместного присутствия гепсидина и TfR2 в клетках HepG2 - хорошо дифференцированной клеточной линии (Aden et al., (1979) Nature 282, 615-616), проявляющей многие особенности физиологии нормальных гепатоцитов. Исследования с помощью RT-PCR с применением подходящих описаний и сочетаний праймеров, примененные в отношении печени человека, выявили экспрессию гепсидина и TfR2 в клетках HepG2. На уровне трансляции наличие гепсидина и TfR2 в клетках HepG2 было подтверждено исследованиями с помощью иммуноблоттинга, который выявил полосы иммунореактивного белка соответствующей молекулярной массы, двигающиеся сходно с соответствующими иммунореактивными полосами из тканей печени. Совместная локализация соответствующих белков в клетках HepG2 была в значительной мере подтверждена иммуногистохимией с применением соответствующих специфичных в отношении области и молекулярного домена антител. Все антитела были способны промечивать гепсидин в клетках HepG2 с выявлением гранулярного паттерна иммунореактивности в данных клетках, что свидетельствует о локализации пептида в малых секреторных везикулах, как это уже было показано в гепатоцитах с помощью электронной микроскопии (Schwartz et al., (1985) EMBO J. 4, 899-904). TfR2 был локализован иммуногистохимически с особым паттерном распределения в клетках HepG2.
На основе настоящих данных на транскрипционном и трансляционном уровне гепсидин и TfR2 совместно экспрессируются в печени и совместно локализуются на домене базолатеральной мембраны гепатоцитов. В дополнение к совместной локализации TfR2 и гепсидина на клеточном уровне было выявлено также сходное распределение данных молекул в пределах печеночных долек с концентрированием иммунореактивности в перипортальных зонах и снижением окрашивания в направлении центральных вен. Координированная экспрессия данных белков в общем (базолатеральном) мембранном домене и их сходное распределение в зонах дольки свидетельствует о морфофункциональном сопряжении регуляторного пептида гормона гепсидина и захвата связанного трансферрином железа посредством TfR2. Действительно, различные данные подтверждают взаимодействие между гепсидином и TfR2. Во-первых, изменения в насыщенности трансферрина, возможно, воспринимаемые TfR2, модулируют экспрессию гепсидина печени (Philpott, С.С. (2002) Hepatology 35, 993-1001). Во-вторых, как показано с помощью количественного анализа ОТ-ПЦР на печени человека, экспрессия TfR2 в печени в значительной мере коррелирует с экспрессией гепсидина, регулируемой насыщенностью трансферрина (S.G.Gehrke, H.Kulaksiz et al., неопубликованные данные). В-третьих, гепсидин и TfR2 совместно локализуются в общем домене клеточной мембраны и проявляют сходное распределение в дольке с сильной иммунореактивностью в перипортальных зонах, месте, где в случае мутаций, которые блокируют экспрессию TfR2 (Fleming et al., (2002) Proc. Natl. Acad. Sci. USA 99, 10653-10658) и гепсидина (Nicolas et al., (2001) Proc. Natl. Acad. Sci. USA 98, 8780-8785), но где также имеет место перегрузка гепсидина (Zhou et al., (1998) Proc. Natl. Acad. Sci. USA 95, 2492-2497; Levy et al., (1999) Blood 94, 9-11) и B2m (Santos et al., (1996) J. Exp. Med. 184, 1975-1985) железом печени. В-четвертых, сообщалось, что мутации гена TfR2 ведут к гемохроматозу (Camasehella et al., (2000) Nat. Genet. 25, 14-15); это может быть результатом снижения экспрессии гепсидина, которая, в свою очередь, приводит к снижению абсорбции железа (Nicolas et al., (2001) Proc. Natl. Acad. Sci. USA 98, 8780-8785).
Одновременное наличие гепсидина и TfR2 в клетках HepG2 и их общая поляризованная локализация и распределение в дольке печени могут указывать на то, что гепсидин является внутренне присущим печени пептидом, морфофункционально сопряженным с TfR2, который регулируется насыщенностью трансферрина и, в свою очередь, модулирует экспрессию гепсидина. Таким образом, можно ожидать соответствующих данных от исследований пути сигнализации гепсидина.
Поскольку образующие кровь ткани и места хранения железа, такие как печень, передают клеткам кишечника сигналы, которые указывают на потребности организма в железе пищи (Philpott С.С. (2002) Hepatology 35, 993-1001), гепсидин служит кандидатом на роль сигнализирующего фактора, секретируемого гепатоцитами и регулирующего всасывание железа кишечником. Однако до настоящего исследования имелось противоречие относительно присутствия определенных молекулярных форм гепсидина в крови (Krause et al. (2000) FEBS Lett 489, 147-150; Park et al. (2001) J Biol Chem 276, 7806-7810; Hunter et al. (2002) J Biol Chem 277, 37597-37603).
Для анализа того, секретируется ли прогормон гепсидина в кровь, и оценки диапазона уровня прогепсидина в сыворотке человека у здоровых добровольцев и больных с различными заболеваниями был разработан ИФА с применением антитела EG(2)-HepN против N-конца прогормона гепсидина. Хотя антитело EG(1)-НерС против С-конца обеспечивало специфичные результаты в экспериментах с точечным блоттингом (данные не показаны), иммуноблоттингом, иммуногистохимией и иммунофлуоресценцией (фиг.1-4), при ИФА иммунореактивность не удавалось получить. Неспособность антитела EG(1)-HepC идентифицировать гепсидин в кровяном русле, возможно, является следствием компактного характера укладки гепсидина и его третичной структуры в крови.
ИФА с антителом EG(2)-HepN отличался высокой воспроизводимостью, стабильностью и чувствительностью с границей определения 3,95 нг/лунку и хорошим разрешением в диапазоне 4-400 нг/мл, диапазоне, в котором производится определение концентрации гепсидина. В сыворотке человека от здоровых индивидуумов (n=26) уровень прогепсидина находился в диапазоне от 51,6 до 153,4 нг/мл (среднее ± ст.ош.; 106,2±32,1 нг/мл), что сопоставимо с концентрацией известных регуляторных пептидных гормонов и приблизительно в 11 раз выше концентрации гепсидина в моче человека (Park et al. (2001) J Biol Chem 276, 7806-7810). Интересно, что измеренная концентрация находилась в широком диапазоне прогепсидина, что может указывать на то, что пептид может быть объектом эффективной регуляции.
Структура кДНК позволяет предполагать, что гепсидин транслируется в виде 84 а.к. препропептида, который подвергается N-концевому процессингу до 20-25 аминокислотных пептидов (Park et al. (2001) (фиг.1 и 9). Хотя между Gly24 и Ser25 находится сильная консенсусная последовательность для сайта отщепления сигнальной последовательности, что должно было бы давать пропептид из 60 остатков, в предшествующих исследованиях не удалось выделить более крупный пропептид из природных источников, таких как ткань печени и кровь (Park et al. (2001)). Дополнительным техническим затруднением служит то, что в печени имеется множество конвертаз пропептида, которые могут подавлять выделение определенных пропептидов. В данном контексте недавние исследования показали, что циркулирующая форма гепсидина у человека, как описано двумя исследовательскими группами для крови (Krause et al. (2000) FEBS Lett 489, 147-150) и мочи (Park et al. (2001) J Biol Chem 276, 7806-7810), состоит из С-концевых 2 0-25 аминокислот белка. Однако измерения с помощью ИФА согласно изобретению производились с применением специфичного антитела, направленного против N-конца предшественника гепсидина в предположении, что, наряду с 20-25-аминокислотными подвергшимися процессингу формами, в кровь человека секретируется и находится в кровотоке прогормон гепсидина. Действительно, предполагаемое выделение прогепсидина в кровь было подтверждено с помощью иммуноблоттинга. Все антитела против гепсидина выявляли в экстрактах сыворотки человека единственную полосу гепсидина ~10 кДа, которая двигалась точно так же, как и иммунореактивный гепсидин в тканевых экстрактах печени и клеток HepG2 (положительный контроль; фиг.1). Фрагменты гепсидина, меньшие 10 кДа, не выявлялись. Наличие прогепсидина в сыворотке человека указывает на то, что гепатоциты секретируют прогормон гепсидина, который может снижать всасывание железа пищи эндокринным путем.
Для анализа значимости гепсидина для больных с избытком железа в настоящем изобретении представлена концентрация гепсидина в сыворотке 35 НН больных, гомозиготных в отношении мутации C282Y в НРЕ с типичными признаками избыточности железа, выявленными у всех исследованных больных НН. Концентрация гепсидина у данных индивидуумов не увеличивалась для снижения всасывания железа в кишечнике, как это предполагалось ранее (Fleming and Sly (2001) Proc Natl Acad Sci USA 98, 8160-8162). Уровень прогепсидина в сыворотке больных НН был неожиданно снижен не только у больных, не подвергавшихся лечению, но также и у индивидуумов, подвергавшихся еженедельной терапии кровопусканием. По сравнению со здоровыми добровольцами концентрация прогепсидина была заметно снижена со 106,2 до 70,2 нг/мл сыворотки. Не было обнаружено различий между больными НН, подвергавшихся и не подвергавшихся лечению. Данные наблюдения находятся в соответствии с предшествующими исследованиями НН, показавшими, что экспрессия гепсидина в печени значительно снижена у мышей с нокаутом hfe (Ahmad et al. (2002) Blood Cells Mol Dis 29, 361-366; Muckenthaler et al. (2003) Nat Genet 34, 102-107) и больных с ассоциированным с HFE гемохроматозом. Эти данные находятся также в соответствии с исследованиями in vitro, показавшими, что нагрузка железом первичных гепатоцитов человека и клеток HepG2 снижает мРНК гепсидина (Gehrke et al. (2003) Blood MS#2002-11-3610.R2; Nemeth et al. (2003) Blood 101, 2461-2463). Поскольку всасывание железа повышено у больных НН несмотря на перегрузку железом (Pietrangelo А. (2002) Am J Physiol Gastrointest Liver Physiol 282, G403-414; Philpott C.C. (2002) Hepatology 35, 993-1001; Anderson and Powell (2002) Int J Hematol 76, 203-207), a конститутивная экспрессия гепсидина предотвращала перегрузку железом на мышиной модели гемохроматоза (Nicolas et al. (2003) Nat Genet 34, 97-101), полагают, что у больных НН нарушена регуляция гепсидина. Пониженная концентрация гепсидина, очевидно, не способна в достаточной мере подавлять повышенное всасывание железа в кишечнике. Более того, на основании того, что экспрессия гепсидина значительно снижена в печени мышей с нокаутом hfe (Ahmad et al. (2002) Blood Cells Mol Dis 29, 361-366; Muckenthaler et al. (2003) Nat Genet 34, 102-107) и у больных НН (Bridle et al. (2003) Lancet 361, 669-673), отсутствие повышения прогепсидина у НН, несмотря на перегрузку железом, указывает на то, что HFE может принимать участие в регуляции уровня гепсидина сыворотки.
Хотя предшествующие исследования показали, что экскреция гепсидина с мочой хорошо коррелирует с ферритином сыворотки (Nemeth et al. (2003) Blood 101, 2461-2463), в настоящем исследовании не было обнаружено корреляции между прогепсидином крови и уровнем железа или ферритина в сыворотке больных НН или находящихся на диализе. Сходным образом, не было обнаружено корреляции между прогепсидином и насыщением трансферрина, которое, как предполагается, регулирует экспрессию гепсидина в печени (Gehrke et al. (2003)), хотя исследованные больные НН не страдали анемией, гипоксией или воспалением, являющимися влияющими на гепсидин параметрами. Данные позволяют предполагать, что регуляция уровня прогепсидина в сыворотке запасами железа включает сложные непрямые эффекты (Nemeth et al. (2003) Blood 101, 2461-2463).
Поскольку гепсидин был также выделен из мочи, в настоящем изобретении предлагается оценка регуляции гепсидина у больных с почечной недостаточностью. В отличие от больных НН и здоровых субъектов, концентрация иммунореактивного прогепсидина в сыворотке больных CRI была значительно повышена с 106,2 нг/мл у здоровых субъектов до 148,1 нг/мл. Повышенный уровень прогепсидина у больных, подвергающихся диализу, позволяет предполагать, что в метаболизм и/или удаление находящегося в кровяном русле пептида могут быть вовлечены почки. Однако в настоящее время не ясно, просто ли гепсидин мочи фильтруется из крови или он имеет почечное происхождение. На основании согласно изобретению нельзя исключить, что гепсидин секретируется, по меньшей мере, частично почками, поскольку он был также обнаружен в клетках канальцев почек (Kulaksiz et al. (2003), неопубликованные данные).
В настоящем изобретении предлагается определение уровня прогепсидина в сыворотке у больных, подвергающихся диализу, с RA, хорошо выявляемым осложнением прогрессирующей почечной недостаточности, которое отличается нормохромными, нормоцитными эритроцитами. По сравнению со здоровыми субъектами концентрация иммунореактивного прогепсидина у больных с RA не была существенно увеличена (среднее 115,0 нг/мл). Несмотря на терминальную почечную недостаточность у данных больных, ведущую к накоплению пептидного гормона, уровень прогепсидина был существенно ниже, чем у больных, подвергающихся диализу, без анемии (среднее 148,1 нг/мл). На основании согласно изобретению можно заключить, что регуляция гепсидина при RA отличается от таковой при анемии воспаления или аденомах печени. Снижение прогепсидина при RA отражает активную физиологическую модуляцию пептида, направленную на повышение всасывания железа в кишечнике и высвобождение железа из ретикулоэндотелиальных макрофагов. Настоящее изобретение показывает, что прогепсидин увеличивается в группе больных CRI без анемии, несмотря на терапию ЕРО. Таким образом, можно заключить, что гепсидин снижается при RA из-за потери крови, которая может быть причиной подавления гепсидина (Nicolas et al. (2002) J Clin Invest 110, 1037-1044).
В настоящем изобретении предлагается ИФА для измерения уровня прогепсидина в сыворотке человека. Данный анализ является неинвазивным и прост в применении, и таким образом соответствует обычной работе. Анализ прогепсидина отличается своей точностью, чувствительностью, воспроизводимостью и четким определением гепсидина-(28-47) в образцах сыворотки человека. Применение настоящего ИФА впервые обеспечивает выявление и определение прогепсидина у больных, страдающих несколькими нарушениями обмена железа. Для выяснения точного молекулярного механизма действия прогепсидина при различных состояниях обмена железа необходимы дальнейшие исследования. Настоящим изобретением также предусматривается, что агонисты и антагонисты гепсидина могли бы быть потенциальными лекарствами для профилактики и лечения заболеваний, связанных с железом.
Для понимания роли гепсидина необходимы знания относительно клеточного происхождения и сигнального пути пептида. В данном отношении в настоящем изобретении описана иммунореактивность гепсидина в печени человека и морской свинки, где он локализуется на домене базолатеральной мембраны гепатоцитов. В предшествующих исследованиях предполагалась возможная связь между данными клетками и всасывающими энтероцитами (Hunter et al., (2002) J. Biol. Chem., M205305200; Anderson et al., (2002) Biochem. Soc. Trans. 30, 724-726). В настоящем изобретении описано выявление прогепсидина в плазме человека, что тем самым указывает на то, что гепатоциты секретируют прогормон гепсидина, который может снижать всасывание железа пищи эндокринным путем. Более того, гепсидин был обнаружен в клетках HepG2, где был также выявлен недавно открытый рецептор типа 2 трансферрина (данные не показаны).
Иммуноферментный анализ для количественного измерения гепсидина в сыворотке человека или животного и других жидкостях организма.
В одном осуществлении изобретения применяют иммуноферментный анализ («ИФА») гепсидина. ИФА представляет собой твердофазный ИФА, основанный на конкурентном принципе. Лунки 96-луночного планшета для микротитрования покрывают поликлональным антителом кролика против гепсидина, направленным против гепсидина-(28-47). За сайты связывания антител против гепсидина, иммобилизованных в лунках, конкурируют неизвестное количество имеющегося в образце прогепсидина и фиксированное количество гепсидина-(28-47), конъюгированного с молекулой биотина. Через один час инкубации планшет для микротитрования промывают для прекращения реакции конкуренции. При последующей инкубации связанные молекулы биотина выявляют с помощью стрептавидина-пероксидазы хрена. После получасовой инкубации планшет промывают второй раз. После добавления раствора субстрата концентрация гепсидина обратно пропорциональна измеренной оптической плотности.
Материалы/
Лунки для микротитрования: лунки, покрытые антителом против гепсидина (96 лунок); реагент: конъюгат биотина (гепсидин, конъюгированный с биотином) 7 мл; набор стандартов сравнения, 1,0 мл каждого; 0, 20, 100, 500, 1000, 2000 нг/мл; контроли прогепсидина, низкий и высокий, 2 флакона (лиофилизат); реагент: ферментный комплекс (стрептавидин, конъюгированный с пероксидазой хрена ("HRP")) 14 мл; реагент: раствор субстрата, -HS-ТМВ, 14 мл; останавливающий раствор, 0,5 М H2SO4, 14 мл; раствор для промывки, 40x, 30 мл; микропланшетный ридер (450±10 нм) (например, DRG Instruments Microtiterplate Reader); прецизионные микропипетки с одноразовыми наконечниками на 50 и 100 мкл; обычный холодильник; фильтровальная бумага; деионизированная вода.
Хотя данное осуществление было описано в плане предпочтительных материалов, специалист в области изобретения должен понимать, что в изобретении могут быть применены другие материалы. Например, специалист в данной области должен понимать, что в изобретении могут быть применены комплементарные связывающие части, отличные от биотина/стрептавидина, а также сочетания фермент/субстрат, отличные от пероксидазы хрена/пероксида.
Условия хранения.
При хранении при температуре от 2°С до 8°С нераспечатанные реагенты должны сохранять реакционноспособность до срока годности. Лунки для микротитрования должны храниться при температуре от 2°С до 8°С. После раскрытия пакета из фольги следует предпринять предосторожности для его повторного плотного закрытия. Иммунореактивность покрытых лунок для микротитрования стабильна в течение приблиз. 6 недель после вскрытия, но при тщательном закрытии пакета с молнией, содержащего влагопоглотитель.
Сбор и подготовка образцов.
В тесте следует применять сыворотку или ЭДТА-плазму человека или животного. Нет необходимости в специальной предварительной обработке биологического образца. Биологический образец может храниться при 2-8°С в течение до 24 часов и должен быть заморожен при -20°С или ниже на более долгие периоды. Не следует применять сильно гемолизованные образцы или образцы с высоким содержанием липидов. Для другого материала образца может потребоваться специальный протокол экстракции.
Проведение анализа: общие замечания.
Всем реагентам и образцам дают возможность достичь комнатной температуры перед применением.
После начала теста все стадии следует завершать без перерыва.
Следует применять новые пластиковые наконечники для каждого реагента, стандарта или образца во избежание загрязнения. При внесении раствора субстрата и останавливающего раствора следует исключить пипетки с металлическими частями.
Внесение пипеткой стандартов и образцов на дно лунки. Для внесения конъюгата фермента и останавливающего раствора рекомендуется держать пипетку в вертикальном положении над лункой и вносить соответствующий раствор в центр лунки так, чтобы достигалось полное перемешивание конъюгата фермента с образцом или стандартом или останавливающего раствора с раствором субстрата.
Рекомендуется, чтобы перед началом анализа были готовы все реагенты, крышки сняты, все необходимые лунки были помещены в держатель и т.д. Это должно обеспечивать равную продолжительность времени, затраченного на каждую стадию пипетирования без перерыва.
Как общее правило, ферментативная реакция линейно пропорциональна времени и температуре. Это делает возможным интерполяцию для фиксированных физико-химических условий. Если при пробном тестировании поглощение нулевым стандартом составляет менее 1,0 или превышает верхний предел Вашего спектрофотометра для микротитрования, Вы можете увеличить или снизить время инкубации при конечном ферментативном образовании окраски до 30 или 10 минут соответственно. Поскольку при каждом тестировании применяют калибраторы, колебания поглощения не влияют на результат.
Раствор субстрата должен быть бесцветным или слабо-синим или зеленым. Если раствор является темно-синим, реагент является непригодным, и он должен быть отвергнут.
При инкубации с раствором субстрата следует избегать попадания солнечного света на планшет для микротитрования.
Подготовка реагентов.
Стандарты сравнения и контроли: восстановить лиофилизованное содержимое флаконов стандарта/контроля с помощью 1,0 мл бидистиллированной воды. Примечание: восстановленные стандарты/контроли стабильны в течение 6 дней при 2-8°С. Для более длительного хранения заморозить при 20°С. Раствор для промывки: добавить деионизированную воду к концентрированному 40х раствору для промывки (содержимое: 30 мл) до конечного объема 1200 мл. Разбавленный раствор для промывки стабилен в течение 2 недель при комнатной температуре.
Процедура анализа.
Вставить желаемое количество покрытых стрипов в держатель.
Внести 50 мкл стандартов гепсидина в соответствующие лунки.
Внести 50 мкл образца в отобранные лунки.
Внести в каждую лунку 50 мкл конъюгата биотина.
Тщательно перемешать планшет в течение 10 секунд. Важно получить полное перемешивание на данной стадии.
Инкубировать в течение 60 минут при комнатной температуре.
Резко вытряхнуть содержимое лунок.
Промыть лунки 3 раза разбавленным раствором для промывки (400 мкл на лунку). Резко вытряхнуть лунки на фильтровальную бумагу для удаления оставшихся капелек.
Добавить в каждую лунку 100 мкл комплекса стрептавидин HRP.
Инкубировать в течение 30 минут при комнатной температуре.
Резко вытряхнуть содержимое лунок.
Промыть лунки 3 раза разбавленным раствором для промывки (400 мкл на лунку). Резко вытряхнуть лунки на фильтровальную бумагу для удаления оставшихся капелек.
Добавить в каждую лунку 100 мкл раствора субстрата с отмеренными интервалами.
Инкубировать в течение 15 минут при комнатной температуре.
Остановить ферментативную реакцию добавлением в каждую лунку останавливающего раствора с теми же временными интервалами, как и на стадии 10, и измерить поглощение каждой лунки при 450±10 нм.
Стабильность конечной реакции.
Рекомендуется, чтобы лунки считывались в пределах 30 минут после стадии 15.
Расчет результатов.
Может быть применен любой планшетный ридер, пригодный для определения поглощения при 450±10 нм. Количество тестостерона в каждом образце получают следующим образом.
a. С помощью линейной или полулогарифмической миллиметровой бумаги строят стандартную кривую путем отложения среднего поглощения (Y) каждого стандарта сравнения против соответствующей его концентрации (X) в нг/мл. Для построения стандартной кривой заявители рекомендуют логистическую функцию с четырьмя параметрами.
b. Используя среднее поглощение каждого образца, определяют соответствующую величину тестостерона простой интерполяцией данной стандартной кривой, умножая при необходимости на разведение исходного образца.
ИФА DRG МАТ 3000 и программа регрессии DRG обеспечивают считывание и обеспечиваемую компьютером интерпретацию с применением логистической функции с четырьмя параметрами.
Пример стандартной кривой.
Следующие данные предназначены лишь для иллюстрации и не могут быть применены вместо получения данных во время тестирования.
Воспроизведение показателей чувствительности.
На фиг.6 показан типичный ИФА для стандартной кривой для прогепсидина крови человека с концентрацией гепсидина-(28-47) в нг/ил с поглощением раствором ИФА при длине волны 450 нм.
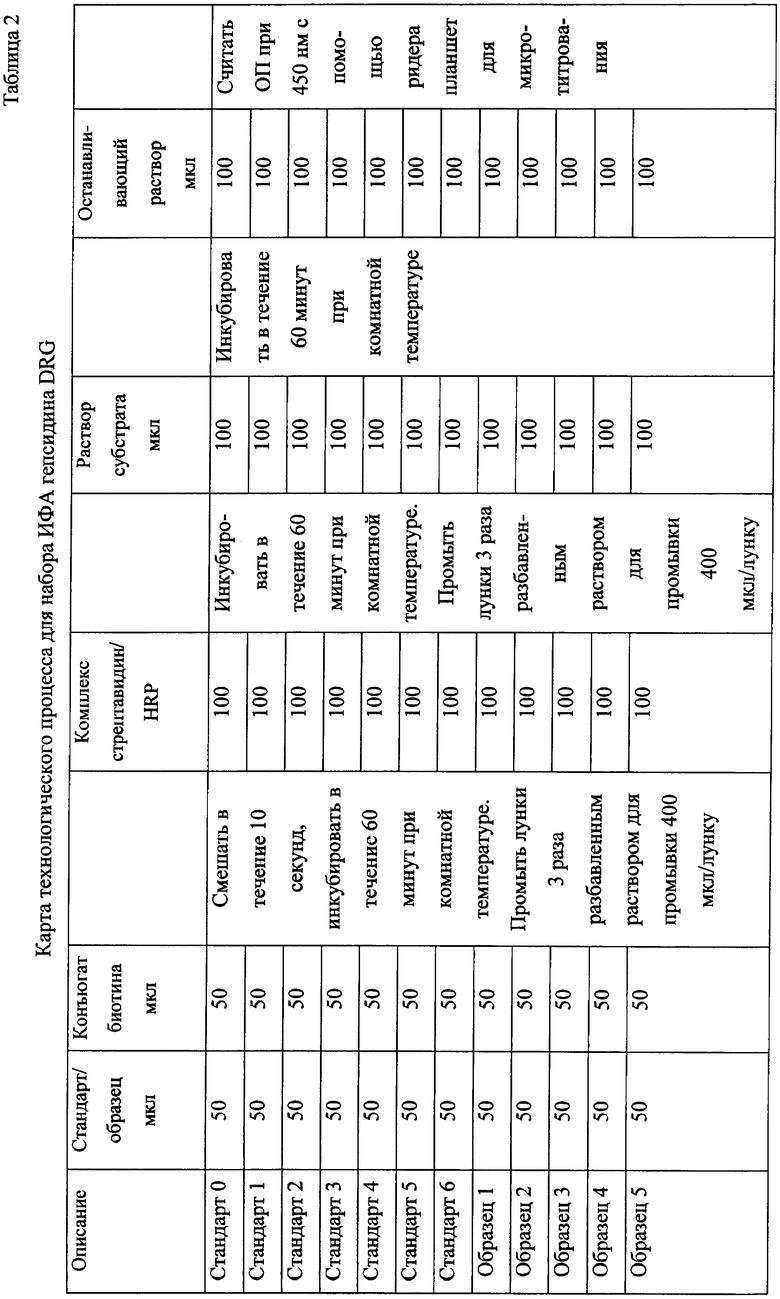
Аналитическую чувствительность рассчитывали как среднее минус 2 ст.откл. (ст.откл.=0,055) для 21 воспроизведения (n=21) анализа для нулевого стандарта.
Чувствительность анализа равна 3,95 нг/мл. Линейность анализа оценивали разведением образцов (сыворотки), содержащих различный уровень гепсидина, нулевым стандартом. Содержание гепсидина в разбавленных образцах оценивали с помощью ИФА. Для каждого образца было получено три разведения, и был рассчитан показатель процентного открытия.
Аналитическую открываемость гепсидина устанавливали при 3 различных концентрациях образцов сыворотки. К образцам с различными исходными концентрациями гепсидина добавляли возрастающее количество немеченого гепсидина (50 нг/мл, 250 нг/мл, 500 нг/мл). Анализировали каждый образец (без добавки и с добавкой). Измеряли концентрацию и рассчитывали показатель процентного выхода гепсидина.
Внутреннюю точность (сходимость результатов ряда измерений) как отклонение определяли с помощью повторных измерений (n=12) 3 контрольных образцов с различным содержанием гепсидина.
Образец 1: Среднее=426,7; Ст.откл.=20, 2; Коэфф.вар.(%)=4,69
Образец 2: Среднее=210,7; Ст.откл.=8,58; Коэфф.вар.(%)=4,07
Образец 3: Среднее=110,7; Ст.откл.=4,74; Коэфф.вар.(%)=4,28
Точность в разных тестах (сходимость результатов разных измерений) как отклонение определяли с помощью повторных измерений (n=23) (3×) 3 различных контрольных образцов с применением наборов из трех различных партий.
Образец 1: Среднее=431,96; Ст.откл.=20,8; Коэфф.вар.(%)=4,82
Образец 2: Среднее=216,17; Ст.откл.=14,44; Коэфф.вар.(%)=6,68
Образец 3: Среднее=109,8; Ст.откл.=10,72; Коэфф.вар.(%)=9,76
Пример 2: Экспрессия гепсидина в почках человека
Гепсидин экспрессируется в дневальных канальцах почек и выделяется в мочу.
Принято считать, что гомеостаз железа контролируется главным образом всасыванием железа в желудочно-кишечном тракте. Однако недавние исследования показывают, что в обмене железа участвуют также почки. Поскольку регулирующий железо и противомикробный пептид гепсидин был первоначально выделен из мочи человека, заявители исследовали клеточную и внутриклеточную локализацию гепсидина в почках млекопитающих и разработали ИФА тест для анализа концентрации прогепсидина в сыворотке и моче.
Экспрессия и клеточная локализация гепсидина была показана с помощью ОТ-ПЦР, иммуноблоттинга и иммуногистохимии в почках человека, мыши и крысы с применением специфичных в отношении гепсидина поликлональных антисывороток. Его концентрация в сыворотке и моче была определена с помощью чувствительного ИФА.
Гепсидин экспрессируется в почках человека, мыши и крысы. Анализ с помощью иммуноблоттинга с применением специфичных в отношении области антисывороток выявил пептид ~9,5 кДа, соответствующий кажущейся молекулярной массе прогепсидина. Исследования по локализации показали, что гепсидин экспрессируется в дистальных канальцах коркового и наружной части мозгового слоя почек. На внутриклеточном уровне гепсидин локализуется на домене апикальной мембраны секреторных клеток канальцев, который на основании его дополнительного присутствия в моче, очевидно, секретируется апикально в мочу. Повышенный уровень прогепсидина (156,8 нг/мл, здоровые добровольцы 104,2 нг/мл) был выявлен у больных с CRI, что указывает на то, что почки могут метаболизировать или элиминировать гормон крови.
На основании данных об экспрессии гепсидина в почках млекопитающих заявители полагают, что регулирующий железо гормон гепсидин является присущим почкам пептидом, который не только метаболизируется/элиминируется почками, но и синтезируется в канальцевой системе почек и секретируется люминально в мочу. Локализация гепсидина в почках указывает на регуляторную роль данного пептида в канальцевой системе почек.
Введение
Недавние исследования показали отличную от нормальной экспрессию гепсидина (Muckenthaler et al., (2003) Nat Genet, 34:102-107) и нарушенную регуляцию гепсидина (Bridle et al., (2003) Lancet, 361:669-673; и Kulaksiz et al., (2003) Gut, в печати) при ассоциированном с HFE гемохромацитозе и связь мутаций гепсидина с тяжелым юношеским гемохромацитозом (Roetto et al., (2003) Nat Genet, 33:21-22). На основании данных наблюдений было предположено, что гепсидин является ключевым компонентом гомеостаза железа, который действует в качестве негативного регулятора всасывания железа в тонком кишечнике и высвобождения железа из макрофагов (Nicolas et al. (2002) Proc Natl Acad Sci USA 99, 4596-4601).
Хотя большинство исследований сконцентрировано на регуляции и функции гепсидина в печени, основном месте образования гепсидина (Park et al.; Kulaksiz et al., (2003) Gut, в печати), накапливаются указания на то, что данный пептид может также играть роль в почках и мочевом тракте (Id., Wareing et al., (2003) Am J Physiol Renal Physiol, публикация до печати; и Ferguson et al., (2003) Kidney Int, 64:1755-1764). Принято считать, что гомеостаз железа контролируется главным образом в желудочно-кишечном тракте на уровне захвата из пищи. Существующая догма состоит в том, что не существует пути экскреции железа из организма. Однако недавние исследования показали, что почки играют важную роль в гомеостазе железа (Wareing et al., (2003) Am J Physiol Renal Physiol, публикация до печати; Ferguson et al., (2003) Kidney Int, 64:1755-1764 и Gunshin et al., (1997) Nature, 388:482-488); значительная часть железа сыворотки подвергается ультрафильтрации клубочками, и основная часть фильтрованного клубочками железа реабсорбируется (Wareing et al., (2000) J Physiol, 524.2:581-586).
Поэтому имеются основания для анализа того, находится ли также гепсидин в почках в качестве местного пептида. В связи с этим заявители разработали антисыворотки против различных эпитопов молекулы предшественника гепсидина и исследовали три вида млекопитающих на транскрипционном и трансляционном уровне. Результаты заявителей указывают на то, что помимо элиминации гепсидина сыворотки в почках пептид также продуцируется в виде внутреннего гормона в клетках дистальных канальцев почек млекопитающих и секретируется люминально в мочу, что указывает на регуляторную роль гепсидина в почках и/или мочевом тракте.
Материалы и способы
Ткани и препараты тканей. Образцы почек человека (n=5), использованные в настоящем изобретении, были получены после резекции почки у взрослых больных с гипернефромой. Образцы печени человека (n=7), использованные в настоящем изобретении, были получены после гемигепатэктомии взрослых больных с метастазами печени (Kulaksiz et al., (2003) Gut, в печати). Здоровые ткани фиксировали в 4% параформальдегиде или в фиксаторе Буэна для иммуногистохимии или немедленно замораживали в жидком азоте для ОТ-ПЦР и иммуноблоттинга. Крыс (n=5) и мышей (n=5) анестезировали и затем забивали обезглавливанием. Образцы ткани почек и печени вырезали и немедленно замораживали в жидком азоте для ОТ-ПЦР или анализа иммуноблоттингом или фиксировали в параформальдегиде.
Синтез пептидов, процедура иммунизации и антитела. На основании опубликованной последовательности прогепсидина (Krause et al., (2000) FEBS Lett. 480:147-150; Pigeon et al., (2001) J Biol Chem, 276:7811-7819) были синтезированы пептиды гепсидин-(28-47) и гепсидин-(70-84) в виде С-концевых амидов с применением стандартного Fmoc протокола (Kulaksiz et al., (2002) Proc Natl Acad Sci USA 99, 6796-6801; и Kulaksiz et al., (2002) Am J Pathol, 161:655-664). Пептиды конъюгировали с гемоцианином моллюска фиссуреллии с помощью м-малеимидбензоил-N-гидроксисукцинимидного эфира, и двух кроликов SPF (Charles River Iffa Credo) иммунизировали каждым из пептидных конъюгатов (Eurogentec, Seraing, Бельгия). Были получены, охарактеризованы и применены (Kulaksiz et al., (2003) Gut, в печати) антитела EG(1)HepC, EG(2)HepC [каждый из которых направлен против прогепсидина-(70-84)] и EG(1)HepN и EG(2)HepN [каждый из которых направлен против прогепсидина-(28-47)].
Анализ экспрессии в почках. На основании последовательностей кДНК GenBank были сконструированы и применены следующие праймеры: гепсидин человека (номер регистрации данных NM021175), представлены в ориентации 5′-3′, 5′-CTG САА ССС CAG GAC AGA G-3′ и 5′-GGA ATA AAT AAG GAA GGG AGG GG-3′; гепсидин крысы (# NM 053469), 5′-АСА GAA GGC AAG ATG GCA СТ-3′ и 5′-GAA GTT GGT GTC TCG СТТ СС-3′, гепсидин-1 мыши (# NM 032541), 5′-CGA ТАС САА TGC AGA AGA GAA GG-3′ и 5′-ТТС AAG GTC ATT GGT GGG GA-3′. Праймеры не проявляли гомологии с любыми ранее сообщавшимися последовательностями.
Выделение РНК производили с применением набора Qiagen RNAeasy, включающего гидролиз ДНК. Анализ с помощью обратной транскрипции (ОТ)-ПЦР проводили, как описано ранее (Kulaksiz et al., (2002) Proc Natl Acad Sci USA 99:6796-6801; и Kulaksiz et al., (2002) Am J Pathol, 161:655-664). После первоначальной денатурации при 94°С в течение 4 мин реакцию производили на протяжении до 30 циклов при следующей температурной программе: 94°С в течение 30 сек, 60°С в течение 30 сек и 72°С в течение 30 сек; данную программу завершали конечной 5-мин стадией элонгации при 72°С. Продукты амплификации подвергали разделению на окрашиваемом этидия бромидом 1,8% агарозном геле со смесью 89 мМ трис/89 мМ борная кислота/2 мМ ЭДТА (рН 8,3). В качестве контроля специфичности амплифицированные продукты ПЦР секвенировали с помощью MWG-Biotech.
Анализ иммуноблоттингом. Эксперименты с иммуноблоттингом проводили на 16,5% трицин-ДДС-Na-полиакриламидных гелях. Белки из почек и печени человека, мыши и крысы, а также их мочи человека (50 мл для каждого эксперимента) экстрагировали согласно опубликованным протоколам (Kulaksiz et al., (2003) Gut, в печати; Kulaksiz et al., (2002) Proc Natl Acad Sci USA 99:6796-6801; и Kulaksiz et al., (2002) Am J Pathol, 161:655-664). После электрофореза белки переносили на гидрофобные мембраны на основе поливинилиденфторида (Pall, Portsmouth, Англия) с помощью полусухого блоттинга. Мембраны инкубировали в течение ночи с антителами против гепсидина, разбавленными 1:1000. После промывки забуференным трис солевым раствором, содержащим 10 мМ трис-HCl (рН 8,0), 150 мМ NaCl и 0,05% твин 20, иммунореактивные белки визуализировали после инкубации с конъюгированным с щелочной фосфатазой антителом козы против кролика (разбавленным 1:50000, Sigma) с применением в качестве хромогенов нитросинего тетразолия и 5-бром-4-хлор-3-индолилфосфата (Sigma). Иммунологическую реакцию при иммуноблоттинге специфично блокировали иммунопреципитацией антител с помощью соответствующих пептидных иммуногенов. Перекрестное взаимодействие со вторым антителом козы против кролика исключали с помощью соответствующих контролей (Kulaksiz et al., (2002) Proc Natl Acad Sci USA 99:6796-6801; и Kulaksiz et al., (2002) Am J Pathol, 161:655-664).
Иммуноцитохимический протокол. Ткани фиксировали в 4% параформальдегиде или фиксаторе Боуэна в течение 18 час при 4°С и заключали в парафин. Парафиновые срезы (4-5 мкм) иммунологически окрашивали на гепсидин (антитела EG(1)HepN, EG(2)HepN, EG(1)HepC и EG(2)HepC, каждое из которых разбавляли 1:2000) с помощью способа авидин-биотин-пероксидазного комплекса (ABC); последовательности инкубации и визуализацию сайтов связывания антиген-антитело производили, как подробно описано (Kulaksiz et al., (2003) Gut, в печати; Kulaksiz et al., (2002) Proc Natl Acad Sci USA 99:6796-6801; и Kulaksiz et al., (2002) Am J Pathol, 161:655-664). Вкратце, срезы инкубировали с соответствующими антителами в течение 24 час при 4°С с последующей инкубацией с биотинилированным IgG против кролика (Jackson Immunoresearch, West Grove, PA., США) в течение 30 мин при разбавлении 1:200. Срезы затем инкубировали в течение 30 мин с предварительно образованным комплексом биотин-пероксидаза/стрептавидин (Jackson Immunoresearch), разбавленным ЗФР (конечные концентрации: биотин-пероксидаза, 0,7 мкг/мл; стрептавидин, 5 мкг/мл). Сайты связывания антиген-антитело определяли инкубацией срезов в 0,7 мМ диаминобензидина гидрохлориде/0,002% Н2О2 в 0,05 М трис-HCl (рН 7,6).
Контроли специфичности. В зависимости от способа неспецифичность исключали с помощью опубликованных контролей (Kulaksiz et al., (2003) Gut, в печати). Специфичность антител тестировали путем предварительной адсорбции антител с помощью гомологичных или гетерологичных антигенных пептидов (6,25-100 мкг/мл антисыворотки) (Kulaksiz et al., (2002) Proc Natl Acad Sci USA 99, 6796-6801; и Kulaksiz et al., (2002) Am J Pathol, 161, 655-664). Предварительная адсорбция антител с помощью гомологичных антигенов в таких низких концентрациях, как 6,25 мкг/мл, полностью блокировала иммунологическую окраску в почках, тогда как предварительная адсорбция антител с помощью гетерологичных антигенов в концентрации до 100 мкг/мл не оказывала влияния на иммунологическое окрашивание.
Конкурентный ИФА гепсидина. Образцы сыворотки и мочи были получены от 22 индивидуумов (11 женщин, 11 мужчин в возрасте 23-59, средний возраст 39 лет) и образцы сыворотки были получены от 22 больных хронической почечной недостаточностью, подвергавшихся хроническому гемодиализу (11 женщин, 11 мужчин в возрасте 25-77 лет, средний возраст 48 лет). Всех больных с хронической почечной недостаточностью лечили 2-3 раза в неделю 3000 ME рекомбинантного эритропоэтина человека (ЕРО). При отборе образцов были предприняты меры, чтобы у здоровых добровольцев и больных не было инфекций и кровотечения. Образцы 10 мл крови отбирали в пробирки для сыворотки и образцы 10 мл мочи отбирали в пробирки для мочи, центрифугировали при 2500×g в течение 10 мин при 4°С. Измерения производили в двух параллелях с применением 96-луночных планшетов для микротитрования, как описано (8). Вкратце, планшеты для микротитрования покрывали 200 мкл/лунку антителом кролика против EG(2)-HepN, разбавленным 1:4000. В каждую лунку добавляли 50 мкл стандартов, содержащих различное количество синтетических пептидов (0, 20, 100, 500 и 1000 нг/мл), или образцов сыворотки и мочи человека, и 150 мкл биотинилированного по N-концу гепсидина-(28-47) (Peptide Specialty Laboratories GmbH, Heidelberg, Германия) (2 нг/лунку), и инкубировали в течение 1 часа при комн. темп. После промывки с помощью TBST (TBS с 0,05% твина 20) биотинилированные комплексы антиген-антитело выявляли с помощью стрептавидина/пероксидазного фермента (Dako, Hamburg, Германия) с применением субстрата тетраметилбензидина (DRG Instruments GmbH, Marburg, Германия); цветную реакцию останавливали с помощью 1 М H2SO4 и поглощение раствором считывали при длине волны 450/630 нм.
Статистический анализ. Данные представлены в виде среднего ± ст.ош.ср. Статистический анализ производили по t-критерию Стьюдента. Различия считали значимыми при Р<0,05.
Результаты
Экспрессия гепсидина в почках млекопитающих. Анализ ОТ-ПЦР четко показал экспрессию гепсидина не только в печени (положительный контроль, см. Kulaksiz et al., (2003) Gut, в печати), но и в почках человека, крысы и мыши (фиг.10). В печени (данные не показаны) и почках указанных видов были обнаружены ожидаемый 192-п.н. продукт ПЦР для человека, 193-п.н. продукт ПЦР для мыши, 201-п.н. продукт ПЦР для крысы. Анализ последовательности показал, что образованные ПЦР продукты имеют полную гомологию с кДНК соответствующих пептидов.
На трансляционном уровне наличие гепсидина было подтверждено с помощью исследований иммуноблоттингом с применением региоспецифичных антител (фиг.10). Антисыворотки, направленные против С- и N-конца молекулы предшественника гепсидина, одинаково обнаруживали иммунореактивную полосу ~9,5 кДа в экстрактах почек человека, крысы и мыши.
Клеточная локализация гепсидина. Иммуногистохимические исследования с помощью региоспецифичных антисывороток против гепсидина постоянно выявляли гепсидин в системе дистальных канальцев почек человека, мыши и крысы (фиг.11-15). Проксимальные канальцы почек, собирательные трубочки и клубочки были полностью лишены иммунореактивности гепсидина. Иммунореактивные дистальные канальцы были ограничены корой и наружным мозговым слоем почек, внутренний мозговой слой почек не проявлял иммунореактивности на гепсидин (фиг.11, 12). Следует отметить наличие четких межклеточных различий между гепсидин-положительными тубулярными клетками: в то время как большинство тубулярных клеток было сильно положительным в отношении гепсидина, некоторые из них проявляли лишь слабую иммунореактивность или были полностью лишены иммунореактивности в отношении гепсидина (фиг.14). Показательно, что на всех исследованных срезах антисыворотки против гепсидина выявляли гранулярный паттерн иммунореактивности в цитоплазме эпителиальных клеток, выстилающих дистальные канальцы (фиг.11, 12). В некоторых тканях позитивные в отношении гепсидина клетки проявляли сильную иммунореактивность, концентрированную на апикальном полюсе секреторных клеток (фиг.13, 15), и иммунореактивность не была обнаружена на домене базолатеральной мембраны соответствующих клеток.
Определение пропептида гепсидина в сыворотке и моче: Был разработан стабильный тест ИФА гепсидина с высокой воспроизводимостью и чувствительностью с применением специфичного в отношении N-конца гепсидина антитела EG(2)-HepN (Kulaksiz et al., (2003) Gut, в печати). Как показано на фиг.16, ИФА обнаружил, что прогепсидин имеется в сыворотке человека. В сыворотке здоровых субъектов измеренный уровень прогепсидина находился в диапазоне от 68,5 до 139,2 нг/мл (среднее±ст.ош; 104,2±19,5 нг/мл). Концентрация прогепсидина в сыворотке больных, страдающих хронической почечной недостаточностью, варьировалась от 63,9 до 327,3 нг/мл (среднее ± ст.ош; 156,8±61,9 нг/мл) и была значительно увеличена по сравнению с таковой в контрольной группе.
С помощью чувствительного ИФА гепсидина был измерен уровень прогепсидина в моче человека, находящийся в диапазоне от 13,9 до 456,0 нг/мл (среднее±ст.ош.; 180,1±94,8 нг/мл) в контрольной группе. Наличие прогепсидина в моче человека было также подтверждено с помощью анализа иммуноблоттингом. В экстрактах мочи человека сыворотки против гепсидина выявили единственную иммунореактивную в отношении гепсидина полосу с молекулярной массой ~9,5 кДа, которая двигалась точно так же, как и иммунореактивный гепсидин в тканях почек (фиг.10).
Обсуждение
Новый гормон гепсидин представляет собой противомикробный пептид и центральный регулятор гомеостаза железа (Park et al., (2001) J Biol Chem 276:7806-7810; Krause et al., (2000) FEBS Lett 480:147-150; Pigeon et al., (2001) J Biol Chem 276:7811-7819; Nicolas et al., (2001) Proc Natl Acad Sci USA, 98:8780-8785; и Nicolas et al., (2001) Proc Natl Acad Sci USA, 99:4596-4601). В предшествующих исследованиях было показано, что печень служит главным источником гепсидина (Park et al., CH (2001) J Biol Chem 276:7806-7810; и Kulaksiz et al., (2003) Gut, в печати). Хотя гепсидин был первоначально выделен из мочи человека (Park et al., (2001) и гемофильтрата (Krause et al., (2000)), экспрессия данного регуляторного пептида не была обнаружена в почках (Pigeon et al., (2001).
С помощью подходящих описаний и сочетаний праймеров, успешно примененных для печени (Kulaksiz et al., (2003) Gut, в печати; и Gehrke et al., (2003) Blood, 102:371-376), настоящий ОТ-ПЦР анализ ясно показал, что гепсидин экспрессируется не только в печени, но и в почках трех видов млекопитающих человека, крысы и мыши. Анализ секвенированием показал специфичность полученных ПЦР продуктов.
Для подтверждения наличия транслированного пептида гепсидина в почках заявители создали набор региоспецифичных антисывороток против гепсидина и применили их в анализе с помощью иммуноблоттинга и иммуногистохимии. Анализ с помощью иммуноблоттинга подтвердил экспрессию гепсидина в почках. Четыре различные антисыворотки, узнающие разные эпитопы в молекуле предшественника гепсидина, одновременно идентифицировали иммунореактивный пептид ~9,5 кДа в почках трех разных видов, который соответствует молекулярной массе прогормона гепсидина, рассчитанной на основании соответствующей последовательности кДНК (Pigeon et al., (2001)). Кажущаяся молекулярная масса данного иммунореактивного пептида находится также в соответствии с молекулярной массой прогормона гепсидина, определенной в печени (Kulaksiz et al., (2003) Gut, в печати). Результаты заявителей однозначно показали, что гепсидин не является специфичным для печени, поскольку он находится также в почках.
Иммуноцитохимические исследования с четырьмя региоспецифичными антисыворотками против гепсидина показали, что в почках человека, мыши и крысы гепсидин специфично локализуется в системе канальцев коры и наружного мозгового слоя почек. Данные иммунореактивные канальцы были идентифицированы как дистальные канальцы почек по их характерным морфологическим особенностям, выявленным при световой микроскопии. Одновременное окрашивание различными региоспецифичными антителами не только в почках человека, но и в почках мыши и крысы, указывает на то, что источником гепсидина почек служат дистальные канальцы. Не было обнаружено иммунореактивности гепсидина в проксимальных почечных канальцах, собирательных трубочках и клубочках или внутренней части мозгового слоя почек.
Иммунореактивность гепсидина в коре и наружной части мозгового слоя почек была ограничена эпителиальными секреторными клетками дистальных канальцев. Примечательно, что все антисыворотки против гепсидина давали гранулярный характер иммунореактивности, что позволяет предполагать локализацию пептида в малых секреторных пузырьках или лизосомах соответствующих клеток, которые уже были выявлены в данных клетках с помощью электронной микроскопии (van Katachalan MA, Kritz W: Pathology of the kidney. Edited by JC Jennette, JL Oldson, MM Schwarz, SG Silver: Philadelphia, Heptinstall′s, 1998, pp 3-66). Примечательно, что имеются четкие межклеточные различия между эпителиальными клетками даже одних и тех же канальцев в отношении плотности иммунореактивности гепсидина, что может отражать различия между клетками в экспрессии или секреции гепсидина. Бросается в глаза то, что в некоторых канальцах иммунореактивность гепсидина локализовалась во всей цитоплазме эпителиальных клеток, тогда как в других канальцах сильная иммунореактивность гепсидина концентрировалась на апикальном полюсе секреторных клеток. Данный особый характер распределения гепсидина на клеточном уровне позволяет предполагать люминально направленную секрецию гепсидина. Заявители не обнаружили экспрессию гепсидина на домене базолатеральной мембраны клеток канальцев почек. Это позволяет предполагать, что почечный гепсидин не выделяется в кровь секретирующими клетками, выстилающими канальцы.
Принято считать, что контроль гомеостаза железа в организме зависит главным образом от жесткой регуляции захвата железа пищи в проксимальном тонком кишечнике. Однако недавние исследования показали, что в гомеостазе железа важную роль играют почки (Wareing et al., (2003) Am J Physiol Renal Physiol, публикация до печати; Ferguson et al., (2003) Kidney Int, 64:1755-1764; и Gunshin et al., (1997) Nature, 388:482-488). Wareing с соавторами удалось убедительно показать, что метаболически значительное количество железа фильтруется в клубочке, и лишь 0,8-1,5% фильтрованного железа действительно экскретируется с мочой (Wareing et al., (2000) J Physiol, 524.2:581-586). Таким образом, должен существовать очень эффективный путь и строгая регуляция реабсорбции железа вдоль почечных канальцев. Действительно, Ferguson с соавторами смогли локализовать транспортер 1 двухвалентных металлов (DMT-1) в канальцевой системе почек (Ferguson et al., (2001) Am J Physiol Renal Physiol, 280:F803-F814). Предполагается, что данный белок служит главным путем захвата железа пищи желудочно-кишечным трактом (Gunshin et al., (1997)). Примечательно, что было показано, что экспрессия DMT-1 является наибольшей на домене апикальной мембраны клеток канальцев коры и наружной части мозгового слоя почек, где заявители обнаружили также гепсидин. Более того, недавние исследования показали, что нарушение поглощения железа пищи сильно влияет на экспрессию DMT-1 в почках (Wareing et al., (2003)). На основании этих результатов и данных, показывающих, что экспрессия гепсидина обратно коррелирует с экспрессией DMT-1 в двенадцатиперстной кишке (Frazer et al., (2002) Gastroenterology 123:835-844), заявители предполагают регуляторную роль гепсидина в транспорте железа в почках.
Предполагаемое выделение гепсидина в мочу было подтверждено исследованиями с применением иммуноблоттинга. Региоспецифичные антисыворотки против гепсидина одинаковым образом идентифицировали сильно промечиваемую полосу правильной молекулярной массы (Kulaksiz et al., (2003) Gut, в печати), которая двигалась точно так же, как и иммунореактивный прогепсидин в экстрактах ткани почек. Эти данные четко показывают, что прогепсидин секретируется секреторными клетками дистальных канальцев и выделяется люминально в мочу, где он, по-видимому, избегает канальцевого протеолиза и повторного использования. Для измерения концентрации прогепсидина в моче человека был разработан чувствительный ИФА с чувствительностью выявления 3,95 нг/лунку. ИФА анализ с антисывороткой против гепсидина EG(2)-HepN, которая уже успешно применялась в экспериментах с ИФА (Kulaksiz et al., (2003) Gut, в печати), показал высокую концентрацию прогепсидина в диапазоне от 13,9 до 456,0 нг/мл (среднее±ст.ош.; 180,1±94,8 нг/мл) в моче здоровых субъектов. Данная концентрация значительно превышает концентрацию прогепсидина в кровяном русле тех же самых лиц (от 68,5 до 139,2 нг/мл; среднее±ст.ош., 104,2±19,5 нг/мл). Примечательно, что не было обнаружено корреляции между прогепсидином в кровяном русле и уровнями железа или ферритина в сыворотке (Kulaksiz et al., (2003) Gut, в печати). Сходным образом, не было обнаружено корреляции между прогепсидином мочи и уровнями железа или ферритина в сыворотке (данные не показаны), которые, как предполагается, регулируют экспрессию гепсидина печени (Pigeon et al., (2001) J Biol Chem 276:7811-7819; Nemeth et al. (2002) Blood, 101:2461-2463; и Ganz T, (2003) Blood, 102:783-788). Поэтому заявители предполагают, что на регуляцию прогепсидина почек/мочи железо или ферритин сыворотки не оказывают прямого влияния.
Оценка регуляции прогепсидина в почках больных, подвергающихся хроническому гемодиализу, показала, что концентрация прогепсидина в сыворотке данных больных была значительно увеличена с 104,2 нг/мл у здоровых субъектов до 156,8 нг/мл. Повышенный уровень прогепсидина у больных с диализом позволяет предполагать, что почки не только участвуют в синтезе гепсидина, но и участвуют в метаболизме и/или элиминации циркулирующего пептида. Интересно, что в современном исследовании было показано, что гормон почек эритропоэтин снижает экспрессию гена гепсидина в печени (Nicolas (2002) Blood Cells, Molecules, and Diseases, 29:327-335). Таким образом, другое объяснение повышенной концентрации прогепсидина у больных с диализом могло бы быть отнесено к относительной недостаточности эритропоэтина, которая обычно встречается на конечной стадии почечной недостаточности (Eckardt KU, (2000) Clin. Nephrol, 53:S2-8; и Santoro A: (2002) Rev Clin Exp Hematol, Suppl 1:12-20). Однако заявители сообщают о повышенном уровне прогепсидина, измеренном у больных с хронической почечной недостаточностью, хотя их лечили ингибирующим гепсидин гормоном эритропоэтином, поддерживающим фильтрацию гепсидина в почках. В одном осуществлении согласно изобретению предусматривается, что гепсидин мочи имеет частично почечное и частично печеночное происхождение; следовательно, следует отметить, что измеренный прогепсидин мочи представляет собой сумму высвобожденного пептида почек и удаленного из кровяного русла пептида.
В заключение недавние исследования показывают, что почки играют важную роль в гомеостазе железа (Wareing et al., (2003) Am J Physiol Renal Physiol, опубл. до печати; Ferguson et al., (2003) Kidney Int, 64:1755-1764; Gunshin et al., (1997) Nature, 388:482-488; Wareing et al., (2000) J Physiol, 524.2:581-586 и Ferguson et al., (2001) Am J Physiol Renal Physiol, 280:F803-F814), однако нет данных относительно регуляторного механизма транспорта железа в почках. В связи с этим заявители впервые локализовали гепсидин в почках трех видов млекопитающих. Данные заявителей указывают на то, что гепсидин не является специфичным для печени. Помимо удаления гепсидина сыворотки в почках пептид может также образовываться в качестве внутреннего гормона в секреторных клетках дистальных канальцев почек и выделяться люминально в мочу, что предполагает регуляторную роль гепсидина в почках и/или мочевом пути. В последующих исследованиях следует проанализировать регуляторный механизм гепсидина в канальцевой системе почек.
Пример 3: Экспрессия гепсидина в поджелудочной железе человека
Ткани поджелудочной железы, примененные в настоящем исследовании, были получены после операции Whipple у больных, страдающих раком поджелудочной железы. С помощью подходящих описаний и сочетаний праймером, успешно примененных в отношении печени и почек, настоящий анализ ОТ-ПЦР показал, что гепсидин экспрессируется не только в печени и почках, но и в поджелудочной железе человека. Секвенирующий анализ показал специфичность полученных продуктов ПЦР.
Анализ с помощью иммуноблоттинга со специфичными антителами подтвердил экспрессию гепсидина в поджелудочной железе на трансляционном уровне. С помощью тех же антител гепсидин был локализован в поджелудочной железе иммуногистохимически. На парафиновых срезах было показано, что иммунореактивность гепсидина локализуется в эндокринной части поджелудочной железы; не было обнаружено иммунореактивности в экзокринной части поджелудочной железы.
Промышленная применимость
В изобретении имеются применения в связи с диагностикой патологического состояния, характеризуемого нефизиологическими уровнями белка гепсидина, включая прогепсидин и его фрагменты.
Хотя изобретение было здесь описано со ссылкой на конкретные осуществления, следует понимать, что данные осуществления являются лишь иллюстрацией принципов и применений согласно изобретению. Следовательно, следует понимать, что могут быть сделаны разнообразные модификации иллюстративных осуществлений и что могут быть изобретены другие применения, не выходя за пределы духа и объема согласно изобретению, как определено прилагаемой формулой изобретения.
Все патентные и не патентные публикации, цитированные в данном описании, являются указанием на достижения специалистов в данной области, к которой относится данной изобретение. Все данные публикации и заявки на изобретение включены здесь в качестве ссылок в той же мере, как если бы отдельная публикация или патентная заявка была конкретно и отдельно указана как включаемая здесь в качестве ссылки.



| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К ГЕПСИДИНУ И ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2668391C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ С ПРИМЕНЕНИЕМ ИНГИБИТОРОВ КОСТНОГО МОРФОГЕНЕТИЧЕСКОГО БЕЛКА 6 (ВМР6) | 2017 |
|
RU2790023C2 |
| ИСПОЛЬЗОВАНИЕ БЕЛКОВ ТЕПЛОВОГО ШОКА ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ ТЕРАПИЙ АНТИТЕЛАМИ | 2003 |
|
RU2375077C2 |
| СПОСОБ ЛЕЧЕНИЯ ОПУХОЛИ У СУБЪЕКТА | 2006 |
|
RU2404193C2 |
| ИНГИБИРОВАНИЕ АНГИОГЕНЕЗА | 2008 |
|
RU2471498C2 |
| ИНГИБИРОВАНИЕ МЕТАСТАЗИРОВАНИЯ ОПУХОЛИ ПРИ ПОМОЩИ АНТАГОНИСТОВ Bv8 ИЛИ G-CSF | 2010 |
|
RU2567803C2 |
| ПОЛИПЕПТИД С АНТИВИРУСНОЙ АКТИВНОСТЬЮ, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2004 |
|
RU2372356C2 |
| БЕЛКИ СПЕЦИФИЧЕСКОГО СВЯЗЫВАНИЯ И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2549678C2 |
| ГОМОГЕННЫЕ ПРЕПАРАТЫ IL-28 И IL-29 | 2004 |
|
RU2518324C2 |
| ПРИМЕНЕНИЕ ИНГИБИТОРА РАК ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ СУСТАВОВ | 2004 |
|
RU2360696C2 |
В настоящем изобретении описываются способы и наборы для диагностики заболеваний, которые характеризуются нефизиологическим уровнем содержания белка гепсидина в организме млекопитающих. Сущность изобретения основывается на связывании антитела и полипептида, где антитело или его фрагмент специфически связываются с одним или более эпитопами средней части или эпитопами карбоксильного конца последовательности SEQ ID NO:2. Описаны также соответствующие антитела. Изобретение может быть использовано для скрининга заболеваний, характеризующихся нефизиологическим уровнем содержания белка гепсидина в организме. 3 н. и 17 з.п. ф-лы, 16 ил., 2 табл.
1. Способ диагностики патологического состояния, характеризующегося нефизиологическими уровнями гепсидина, подразумевающий получение образца ткани или жидкости от субъекта; приведение образца в контакт с антителом или его фрагментом, которые специфически связываются с одним или более эпитопами средней части или эпитопами карбоксильного конца из SEQ ID NO:2, и количественное определение уровня комплекса указанного антитела с фрагментом из SEQ ID NO:2 в образце; где нефизиологический уровень указанного комплекса является показателем патологического состояния и где указанное антитело способно обнаружить пептид, включающий аминокислоты 25-84 из SEQ ID NO:2 в анализе методом Вестерн-блоттинга.
2. Способ по п.1, в котором антитело специфически связывает эпитоп средней части, находящийся между аминокислотами с 28 по 47 из SEQ ID NO:2.
3. Способ по п.1, в котором антитело специфически связывает эпитоп карбоксильного конца, находящийся между аминокислотами с 70 по 84 из SEQ ID NO:2.
4. Способ по п.1, в котором количественное определение подразумевает проведение анализа, выбранного из группы, состоящей из радиоиммуноанализа, иммуноферментного анализа, сэндвич-метода, реакции с преципитином, теста иммунодиффузии в геле, теста агглютинации, иммунофлуоресцентного теста, иммунотеста с протеином А и иммуноэлектрофоретического теста.
5. Набор для выявления патологического состояния, характеризующегося нефизиологическими уровнями гепсидина, включающий в себя антитело или его фрагмент, которые специфически связываются с одним или более эпитопами средней части или эпитопами карбоксильного конца из SEQ ID NO:2, и реагент, который напрямую или не напрямую связывается с антителом или его фрагментом, где указанное антитело способно обнаружить пептид, включающий аминокислоты 25-84 из SEQ ID NO:2 в анализе методом Вестерн-блоттинга.
6. Набор по п.5, где антитело или его фрагмент иммобилизованы на подложке.
7. Набор по п.5, где реагент включает в себя один или более эпитопов средней части или эпитопов карбоксильного конца из SEQ ID NO:2 в комплексе с первой связывающей молекулой.
8. Набор по п.7, где первая связывающая молекула представляет собой биотин.
9. Набор по п.8, который дополнительно включает в себя фермент в комплексе со второй связывающей молекулой и субстрат фермента.
10. Набор по п.9, где вторая связывающая молекула представляет собой стрептавидин.
11. Набор по п.9, где фермент представляет собой пероксидазу хрена, а субстрат содержит пероксид.
12. Антитело или его фрагмент, которые специфически связываются с одним или более эпитопами средней части или эпитопами карбоксильного конца из SEQ ID NO:2 и где указанное антитело способно обнаружить пептид, включающий аминокислоты 25-84 из SEQ ID NO:2 в анализе методом Вестерн-блоттинга.
13. Антитело по п.12, где эпитоп средней части находится между аминокислотами с 28 по 47 из SEQ ID NO:2.
14. Антитело по п.12, где эпитоп карбоксильного конца находится между аминокислотами с 70 по 84 из SEQ ID NO:2.
15. Способ по п.1, где указанный фрагмент SEQ ID NO:2 включает в себя аминокислоты 25-59 из SEQ ID NO:2 или аминокислоты 60-84 из SEQ ID NO:2.
16. Способ по п.1, где указанный фрагмент SEQ ID NO:2 включает в себя аминокислоты 25-59 из SEQ ID NO:2.
17. Набор по п.5, где указанное антитело связывается с аминокислотами 25-59 из SEQ ID NO:2 или аминокислотами 60-84 из SEQ ID NO:2.
18. Набор по п.5, где указанное антитело связывается с аминокислотами 25-59 из SEQ ID NO:2.
19. Антитело по п.12, способное обнаружить менее чем приблизительно 50 нг SEQ ID NO:3.
20. Антитело по п.19, способное обнаружить приблизительно 1 нг SEQ ID NO:3.
| Электромешалка для приготовления мыльно-пенистых ванн для размешивания в ваннах медикаментов | 1948 |
|
SU73454A1 |
| US 5420016 А, 30.05.1995 | |||
| PIGEON С | |||
| et al., A new mouse liver-specific gene, encoding a protein homologous to human antimicrobial peptide hepcidin, is overexpressed during iron overload, 2001, J | |||
| Biol | |||
| Chem | |||
| СПОСОБ СОСТАВЛЕНИЯ ЗВУКОВОЙ ЗАПИСИ | 1921 |
|
SU276A1 |
| NICOLAS G | |||
| et al., Lack of hepcidin gene expression and severe tissue iron overload | |||

