ОСНОВЫ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Белковые факторы и гормоны, участвующие в поддержании здоровья человека, в настоящее время производятся фармацевтической индустрией путем экстракции или - в последние десятилетия - с помощью рекомбинантной технологии. Экспрессии генетических конструкций, включенных в нужные гены, с успехом добивались в линиях клеток бактерий, дрожжей или млекопитающих. Однако применение культур клеток млекопитающих для получения комплексных белков, таких как белки, которые требуют точности в соблюдении характера гликозилирования, сопряжено с внедрением дорогостоящих процессов.
В последнее десятилетие все больше используется технология рекомбинантных ДНК для продуцирования коммерчески важных биологических материалов. С этой целью клонированы последовательности ДНК, кодирующие целый ряд белков человеческого организма, имеющих важное медицинское значение. К ним относятся инсулин, активатор плазминогена, альфа-1-антитрипсин и факторы VIII и IX свертывания крови. В настоящее время, несмотря на наличие новых технологий рекомбинантной ДНК, такие белки обычно очищают из крови и тканей - дорогостоящие и отнимающие много времени процессы, которые могут повлечь за собой риск переноса инфекционных агентов, таких как факторы, которые вызывают СПИД и гепатит.
Хотя экспрессия последовательностей ДНК в бактериальных клетках для продуцирования необходимых и важных с медицинской точки зрения белков выглядит привлекательно, на практике бактерия часто оказывается неподходящим объектом в качестве хозяина, потому что инородные белки в бактерии не стабильны, и процессинг протекает некорректно.
Осознавая существование данной проблемы, попытались осуществить экспрессию клонированных генов в культуре ткани млекопитающего, добиваясь в некоторых случаях жизнеспособной стратегии. Однако периодическое культивирование животных клеток является дорогостоящим и технически трудоемким процессом.
Следовательно, существует потребность в способе получения биологических веществ, таких как корректно модифицированные полипептиды эукариотов, отличающемся высоким выходом продукта и низкой себестоимостью. Отсутствие агентов, способных инфицировать человеческий организм, должно стать достоинством такого способа.
Возможность получения трансгенных животных с нужным геном, например рогатого скота, с целью получения значительных количеств белка человеческого организма в составе молока, представляет значительный интерес для индустрии. Некоторые группы исследователей опубликовали статьи о своих успехах в продуцировании человеческого сывороточного альбумина, альфа-антитрипсина и некоторых других белков у трансгенных коров или коз.
Предварительно было проведено множество экспериментов на мышах или крысах, и трансгенная экспрессия всегда ограничивалась молочными железами, так как использовали промоторы бета-казеина или лактальбумина, которые реагировали только с фактором транскрипции молочной железы женских особей в период лактации.
Экспрессия гетерологического белка исключительно в молоке связана с тем, чтобы избежать нежелательного влияния на здоровье животного-хозяина и обеспечить легкий способ очистки.
В настоящее время исследователи уделяют внимание нескольким системам для улучшения выхода продукции клеточной трансфекции или селекции и выбора источника гомологичной эмбриональной соматической клетки для улучшения жизнеспособности и иммунной системы клонированных животных.
С другой стороны, существует огромный интерес к явлению переноса ядра соматической клетки в основном в связи с тем, чтобы сделать возможным размножение элитных домашних животных и создание трансгенных животных для сельскохозяйственных и биомедико-биологических целей. Коротко, перенос ядра (ПЯ) включает в себя энуклеацию ооцита реципиента, с последующим переносом донорской клетки в перивителлиновое пространство в непосредственной близости от реципиентного цитопласта, и их слияние. Развитие индуцируется искусственно, посредством химической или физической активации. Путем переноса ядра соматической клетки успешно получено клонированное потомство у овцы (Campbell, K.H., et al., Nature 380: 64-66 (1996), 1996; Wells, D.N., et al., Biol Reprod 57: 385-393 (1997); Wilmut, I., et al, Nature 385:810-813 (1997)); козы (Baguisi, A., et al., Nat Biotechnol 17: 456-461 (1999)) и у коровы (Cibelli, J.B., et al., Science 280: 1256-1258 (1998); Kato, Y., et al., Science 282: 2095-2098 (1998); Wells, D.N., et al., Reprod Fertil Dev 10: 369-378 (1998)).
Существует несколько факторов, влияющих на результаты ПЯ, которые включают в себя способы энуклеации, слияния, активации и синхронизации цикла клетки донор-реципиент. Высокая эффективность энуклеации реципиентных ооцитов достигается путем применения ДНК-специфических жизнеспособных красителей для визуализации хроматина (Stice, S. L., and Keefer, C. L., Biol Reprod 48: 715-719 (1993); Westhusin, M.E., et al., J Reprod Fertil 95: 475-480 (1992)). Слияние донорской клетки с ооцитом реципиента зависит от безошибочности выравнивания клетки в пульсирующем поле, от контакта клетки донора с ооцитом реципиента и от размера донорских клеток (Collas, P., et al., Anal Biochem 208: 1-9 (1993)). Усовершенствована активация ПЯ воспроизведенного эмбриона, и темпы развития в бластоцист стали сравнимы с оплодотворенными in vitro ооцитами (Liu, L., et al., Mol Reprod Dev 49: 298-307 (1998)).
Продемонстрировано успешное развитие эмбрионов с ПЯ при использовании зрелых ооцитов (Willadsen, S. M., Nature 320: 63-65 (1986)), зигот (McGrath, J., and Solter, D., Dev Biol N Y 4:37-55 (1985)) и эмбрионов на стадии деления (Tsunoda, Y., et al., J Reprod Fertil 96: 275-281 (1992)) как реципиентов цитопластов, однако это зависит также и от источника донорских ядер. Совместимость клеточного цикла между реципиентом-цитопластом и донорскими клетками является одним из важных факторов, который влияет на развитие эмбрионов с ПЯ. Для сохранения плоидности воспроизводимого эмбриона необходима надлежащая синхронизация.
Митотический клеточный цикл имеет следующие последовательные фазы: предрепликационная гэп-фаза (G1), синтез ДНК (S-фаза), предмитотическая гэп-фаза (G2) и митоз (M-фаза). В течение одного клеточного цикла все геномные ДНК перед митозом единожды реплицируются. Донорское ядро в состоянии интерфазы переносили в энуклеированный зрелый ооцит (метафаза II), претерпевший несколько морфологических изменений. После слияния, но перед тем как произойдет разрыв оболочки донорского ядра (NEBD), хромосомы конденсируются (PCC). Эти процессы индуцируются посредством фактора-промотора созревания/митоза/мейоза (MPF) и митоген-активируемой протеинкиназы (MAPK) (Collas, P., and Robl, J. M., Biol Reprod 45: 455-465(1991)). Активность MPF и MAPK найдена во всех клетках в состоянии мейоза и митоза, и наивысшая активность обнаружена в метафазе, и в ооцитах млекопитающих высокие уровни активности могут также индуцировать угнетение в метафазе II. Уменьшение активности MPF и MAPK при оплодотворении или активации кальций-ионофором является сигналом для завершения мейоза, эмиссии второго полярного тельца, деконденсации ядра спермы и формирования пронуклеуса.
Непосредственное действие NEBD и PCC на донорский хроматин зависит от клеточного цикла ядра донора в момент переноса. Диплоидные G0/Gl-ядра конденсируются с образованием одинарных хроматид, но тетраплоидные G2-ядра конденсируются с образованием двойных хроматид. Однако ядра в S-фазе в момент переноса выглядят “пульверизованными”; PCC вызывает обширное повреждение ДНК. Следовательно, корректная плоидность может быть получена посредством перенесения G1- или G0-ядра в ооциты, находящиеся в метафазе II в момент активации или перед активацией. Второй способ состоит в переносе ядра в предварительно активированный ооцит в S-фазе, в данном случае возможно применение донорской клетки в G1, G0 или S-фазах. Поскольку активность MPF и MAPK низкая, хроматин деконденсируется и ДНК претерпевает репликацию без PCC и NEBD.
Третья схема синхронизации была продемонстрирована на мышах, когда развитие жизнеспособного потомства получали путем реконструкции эмбриона, используя донорскую клетку в G2-фазе или метафазе и энуклеированный ооцит в метафазе II (Cheong, H.T., et al., Biol Reprod 48: 958-963 (1993); Kwon, 0.Y., and Kono, T., Proc Natl Acad Sci USA 93: 13010-13013 (1996)). Обнаружено вытеснение полярного тельца из эмбриона, воспроизводимого посредством ПЯ, в результате чего получен одинарный диплоид эмбриона и диплоид полярного тельца (Kwon, O.Y., and Kono, T., Proc Natl Acad Sci USA 93: 13010-13013 (1996)). Однако о формировании полярного тельца после ПЯ в энуклеированный ооцит в MII-фазе у рогатого скота, овцы или свиней данных нет, что говорит о различии в специфике механизма, контролирующего формирование интактной веретенообразной структуры и вытеснение полярных телец.
Предполагается, что стадии клеточного цикла донорской клетки и реципиента также играют важную роль в перепрограммировании ядра донорской клетки. Увеличение промежутка времени между переносом донорского ядра и транскрипцией зиготы может улучшить перепрограммирование ядра. По этой причине некоторые авторы несколько часов активировали ооцит после слияния (Cibelli, J.B., et al., Science 280: 1256-1258 (1998); Wakayama, T., et al., Nature 394: 369-374 (1998); Wells, D.N., et al., Biol Reprod 60: 996-1005 (1999)). Другие авторы использовали последовательный перенос ядра (Stice, S.L., and Keefer, C. L., Biol Reprod 48: 715-719 (1993)).
Неизученный способ увеличения времени перепрограммирования донорского ядра состоит в переносе ядра перед метафазой II. После разрыва зародышевого пузырька (GVBD) все превращения ядра регулировались главным образом посредством увеличения MPF и MAPK в цитозоле ооцита, что предотвращает реорганизацию оболочки ядра и начало S-фазы до оплодотворения или активации. Следовательно, зрелый ооцит может быть универсальным реципиентом для донорской клетки, находящейся в метафазе или G2-фазе. Донорские клетки даже могут быть использованы в G1-фазе или G0-фазе, если индуцировать активацию в S-фазе перед делением клетки.
Перепрограммирование ядра возможно при использовании в качестве донорских клеток бластомеров в G2-фазе или M-фазе (Cheong, H.T., et al., Biol Reprod 48: 958-963 (1993); Kwon, O.Y., and Kono, T., Proc Natl Acad Sci USA 93: 13010-13013 (1996); Liu, L., et al., Mol Reprod Dev 47: 255-264 (1997)). Объяснением этому может быть тот факт, что в результате конденсации хромосомы некоторые факторы перемещаются из хроматина. Фактически, при переносе ядра NEBD и PCC рассматривались как морфологические признаки перепрограммирования ядра. Кроме того, во время оплодотворения хроматин спермы находится в крайне конденсированном состоянии, его объем значительно меньше, чем объем ядра соматических клеток, и ооцит имеет возможность удалить ядерный белок спермы. Хромосомы ооцита во время слияния спермы с ооцитом также конденсированы. Возможно, что конформация конденсированного хроматина может иметь некоторую биологическую значимость. Следовательно, посредством имитации этой ситуации через перенос ядра в метафазе, энуклеированный в метафазе реципиент может улучшить результат ПЯ. Тем не менее, несколько исследователей применили этот подход к домашним животным, и использовали при этом бластомеры в качестве донорских клеток (Liu, L., et al., Mol Reprod Dev 47: 255-264 (1997)).
Целью данного изобретения является характеристика и повышение качества известного переноса ядра соматической клетки, создание надежного и экономичного технического приема для получения генетически идентичных телят из зрелых донорских клеток.
КРАТКОЕ СОДЕРЖАНИЕ ИЗОБРЕТЕНИЯ
Изобретение относится к трансгенному млекопитающему, отличному от человека, характеризующемуся продуцированием неожиданно высоких уровней рекомбинантного гормона роста в своем молоке. Рекомбинантный гормон роста может представлять собой гормон роста человека, но не ограничиваться этим. Трансгенное млекопитающее, отличное от человека, может представлять собой животное рода быков, но не ограничиваться этим.
Изобретение, кроме того, относится к плазмиде, которая обеспечивает экспрессию нужного белка в клетках молочной железы млекопитающих, в которых экспрессия регулируется посредством промотора бета-казеина. Нужный белок может представлять собой гормон роста человека, но не ограничиваться этим.
Изобретение относится также к различным способам создания трансгенных млекопитающих, отличных от человека, которые продуцируют рекомбинантный гормон роста в своем молоке. Рекомбинантный гормон роста может, не ограничиваясь этим, представлять собой гормон роста человека. Трансгенное млекопитающее, отличное от человека, может, не ограничиваясь этим, представлять собой животное рода быков.
Изобретение относится также к способу продуцирования нужного белка, включающему в себя создание трансгенного млекопитающего, отличного от человека, которое продуцирует указанный белок в своем молоке, получение указанного молока от трансгенного млекопитающего, отличного от человека, и выделение указанного нужного белка из молока. Нужный белок может представлять собой, не ограничиваясь этим, гормон роста человека. Трансгенное млекопитающее, отличное от человека, может представлять собой, не ограничиваясь этим, животное рода быков.
Изобретение относится также к способу продуцирования и выделения рекомбинантного гормона роста из молока трансгенного млекопитающего. Рекомбинантный гормон роста может представлять собой, не ограничиваясь этим, гормон роста человека. Трансгенное млекопитающее может представлять собой, не ограничиваясь этим, животное рода быков.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фиг.1A-1B показан суточный объем молока, собираемого от трансгенной коровы, полученной путем слияния энуклеированного ооцита и фибробласта, предварительно трансфицированного плазмидой, содержащей ген, который кодирует гормон роста человека (hGH) и промотор, который направляет его экспрессию к клеткам молочной железы.
На фиг.2A-2C показана плотность бактерий, обнаруженных в молоке, собираемом от той же самой трансгенной коровы.
На фиг.3A-3B показана биологическая активность hGH, содержащегося в молоке той же самой трансгенной коровы.
На фиг.4A показана суточная масса hGH, получаемого из молока той же самой трансгенной коровы. Эта величина и суточный объем молока, собираемого от трансгенной коровы, вместе нанесены на график на фиг.4B.
На фиг.5A показана концентрация hGH и инсулиноподобного фактора роста 1 (IGF-1) в сыворотке той же самой трансгенной коровы, и суточная масса hGH, продуцируемого в молоке той же самой трансгенной коровы. Концентрация hGH в сыворотке трансгенной коровы и суточная масса hGH, продуцируемого в молоке трансгенной коровы, вместе нанесены на график на фиг.5B. На фиг.5C профили концентрации hGH и IGF-1 во времени в сыворотке трансгенной коровы вместе нанесены на график.
На фиг.6A и 6B показана концентрация hGH и инсулиноподобного фактора роста-1 (IGF-1) в сыворотке двух трансгенных телят, полученных посредством субклонирования коровы, которая является трансгенной в отношении продуцирования hGH в ее молоке.
На фиг.6C и 6D профили концентрации hGH и IGF-1 во времени в сыворотке каждого из трансгенных телят вместе нанесены на график.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Изобретение относится к отличному от человека трансгенному млекопитающему, характеризующемуся продуцированием неожиданно высокого уровня рекомбинантного гормона роста в своем молоке. Указанное млекопитающее может представлять собой, не ограничиваясь этим, животное рода быков. Другой род трансгенных млекопитающих может представлять собой, не ограничиваясь этим, род свиней, род овец, род коз или род грызунов.
Рекомбинантный гормон роста может представлять собой, не ограничиваясь этим, гормон роста человека. Данная молекула, известная также как соматотропин, является белком, состоящим из 191 аминокислоты с молекулярным весом приблизительно 22 кД. Она необходима для линейного роста, и применение ее хорошо изучено.
Изобретение также относится к трансгенному млекопитающему, характеризующемуся тем, что в его молоке продуцируется рекомбинантный гормон роста, стимулирующий молочные железы животного к продуцированию большего количества молока, содержащего указанный гормон.
Изобретение относится также к плазмиде, содержащей ген, кодирующий нужный белок, оперативно связанный с промотором бета-казеина и геном β-лактамазы. Указанный нужный белок может представлять собой, не ограничиваясь этим, гормон роста человека. Указанная плазмида может представлять собой плазмиду pRβhGH.
В следующем воплощении, плазмида дополнительно содержит устойчивый к неомицину ген для селекции генетически устойчивых клеток. Примером такой плазмиды является плазмида pRNeo.
В следующем воплощении плазмида содержит ген, кодирующий зеленый флуоресцирующий белок, такой как GFP, который контролируется промотором цитомегаловируса (CMV). Примером такой плазмиды является плазмида pRNeoGreen.
Кроме того, изобретение относится к плазмиде, такой как описана выше и которая линеаризована посредством рестрикционного расщепления. В частности, прибегая к использованию рестрикционного фермента ApaLI, вырезали ген β-лактамазы.
Ведется подготовка процедуры депонирования вышеназванных плазмид согласно Будапештскому Договору. Имя и адрес депонента: DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Mascheroder Weg 1b, 38124 Braunschweig, Germany. Номера соответствующих образцов будут получены в порядке очереди.
Далее, изобретение относится к плазмиде, сконструированной на основе Neo-резистентной генсодержащей плазмиды, такой как pVEβcashGH, в которую была встроена модифицированная укороченная область бета-казеинового промотора выше области, кодирующей hGH. Линейный фрагмент может быть получен из плазмиды pVEβcashGH посредством удаления гена бета-лактамазы.
Изобретение, кроме того, относится к способу трансфекции генетических конструкций с использованием комбинации катионных липидов для утилизации липосомами.
Описаны также способы селекции неомицин-резистентных клеток в подходящей среде, так же как и способы селекции зеленых флуоресцирующих трансгенных клеток. Данные клетки отбирали с осторожностью, чтобы избегать повреждения клетки.
Изобретение относится также к способу переноса ядер клеток, угнетенных в G0-фазе или в различные периоды времени клеточного цикла, в энуклеированные бычьи ооциты.
Изобретение относится к способу переноса трансгенного эмбриона в простимулированную гормоном матку коровы.
Согласно изобретению описан способ определения параметров здоровья животного. Для того чтобы определить такие параметры, анализы проводили как на основе сыворотки, так и на основе молока.
Далее, изобретение относится к способу создания трансгенного млекопитающего, отличного от человека, включающему в себя получение гена, который кодирует гормон роста, клонирование гена в плазмиду, где ген оперативно связывается с промотором, который будет направлять экспрессию гена в клетки молочной железы, что приводит к экспрессии плазмиды, трансфекцию соматических клеток с экспрессируемой плазмидой таким образом, что плазмида включается в геном клеток, что приводит к образованию трансгенных соматических клеток, энуклеацию зрелого ооцита, что приводит к образованию безъядерного ооцита, слияние одной трансгенной соматической клетки с энуклеированным ооцитом, что приводит к образованию моноклеточного эмбриона, имплантирование эмбриона в матку млекопитающего-реципиента и мониторинг беременности вплоть до рождения трансгенного млекопитающего.
Изобретение, кроме того, относится к способу создания трансгенного млекопитающего, отличного от человека, включающего в себя экстракцию соматических клеток из женской особи млекопитающего, которая является трансгенной по продуцированию рекомбинантного гормона роста в своем молоке, не обязательно фибробластов, энуклеацию зрелого ооцита, что приводит к образованию безъядерного ооцита, слияние одной трансгенной соматической клетки с энуклеированным ооцитом, что приводит к образованию моноклеточного эмбриона, имплантацию эмбриона в матку млекопитающего-реципиента и мониторинг беременности вплоть до рождения трансгенного млекопитающего.
Далее, изобретение относится к способу создания трансгенного млекопитающего, отличного от человека, включающего в себя суперовуляцию женской особи млекопитающего, отличного от человека, которая является трансгенной по продуцированию рекомбинантного гормона роста в своем молоке, искусственное оплодотворение млекопитающего спермой, полученной от мужской особи нетрансгенного млекопитающего, отличного от человека, с получением эмбриона, отбор эмбрионов, имплантирование эмбрионов в матку млекопитающего-реципиента и мониторинг беременности вплоть до рождения трансгенного млекопитающего.
Изобретение далее относится к способу создания трансгенного млекопитающего, отличного от человека, включающему в себя суперовуляцию женской особи млекопитающего, отличного от человека, которая трансгенна по продуцированию рекомбинантного гормона роста в своем молоке, искусственное оплодотворение млекопитающего спермой, полученной от мужской особи млекопитающего, отличного от человека, которая является трансгенной по продуцированию указанного рекомбинантного гормона роста, получение эмбрионов, отбор эмбрионов, имплантирование эмбрионов в матку млекопитающего-реципиента и моноторинг беременности вплоть до рождения трансгенного млекопитающего.
Изобретение далее относится к способу создания трансгенного млекопитающего, отличного от человека, включающему в себя суперовуляцию нетрансгенной женской особи, отличной от человека, искусственное оплодотворение млекопитающего спермой, полученной от мужской особи млекопитающего, отличного от человека, которая является трансгенной по продуцированию указанного рекомбинантного гормона роста, получение эмбрионов, отбор эмбрионов, имплантирование эмбрионов в матку млекопитающего-реципиента и моноторинг беременности вплоть до рождения трансгенного млекопитающего.
Рекомбинантный гормон роста может представлять собой гормон роста человека, но этим не ограничивается. Трансгенное млекопитающее, отличное от человека, может представлять собой животное рода быков, но этим не ограничивается.
Изобретение, кроме того, относится к способу продуцирования белка, включающему в себя создание трансгенного млекопитающего, отличного от человека, которое продуцирует нужный белок в своем молоке с неожиданно высокой продуктивностью, получение молока от трансгенного млекопитающего, отличного от человека, и выделение нужного белка из молока.
Изобретение также относится к способу продуцирования нужного белка у трансгенного млекопитающего, отличного от человека, выполненному посредством технологического приема, включающего в себя получение гена, который кодирует указанный нужный белок, клонирования гена в плазмиду, где ген оперативно связывается с промотором, который будет направлять экспрессию гена в клетки молочной железы, что приводит к экспрессии плазмиды, трансфекции соматических клеток, не обязательно фибробластов, с плазмидой, таким образом, что плазмида включается в геном указанных соматических клеток, что приводит к образованию трансгенных соматических клеток, энуклеацию зрелого ооцита, что приводит к образованию безъядерного ооцита, слияние одной трансгенной соматической клетки с энуклеированным ооцитом, что приводит к образованию моноклеточного эмбриона, имплантирование эмбриона в матку млекопитающего-реципиента и мониторинг беременности вплоть до рождения трансгенного млекопитающего.
Изобретение относится также к способу продуцирования нужного белка трансгенным млекопитающим, отличным от человека, выполненному посредством технологического приема, включающего в себя экстракцию соматических клеток из женской особи млекопитающего, трансгенной по продуцированию указанного нужного белка в своем молоке, не обязательно фибробластов, энуклеацию зрелого ооцита, что приводит к образованию безъядерного ооцита, слияние одной трансгенной соматической клетки с энуклеированным ооцитом, что приводит к образованию моноклеточного эмбриона, имплантацию эмбриона в матку млекопитающего-реципиента и мониторинг беременности вплоть до рождения трансгенного млекопитающего.
Изобретение относится также к способу продуцирования нужного белка у трансгенного млекопитающего, отличного от человека, выполненному посредством технологического приема, включающего в себя суперовуляцию женской особи млекопитающего, отличного от человека, трансгенного по продуцированию указанного нужного белка в своем молоке, искусственное оплодотворение млекопитающего спермой, полученной от мужской особи нетрансгенного млекопитающего, отличного от человека, продуцирование эмбрионов, отбор эмбрионов, имплантацию эмбрионов в матку млекопитающего-реципиента и мониторинга беременности вплоть до рождения трансгенного млекопитающего.
Изобретение относится также к способу продуцирования нужного белка трансгенным млекопитающим, отличным от человека, выполненному посредством технологического приема, включающего в себя суперовуляцию женской особи млекопитающего, отличного от человека, которая трансгенна по продуцированию указанного нужного белка в своем молоке, искусственное оплодотворение млекопитающего спермой, полученной от мужской особи млекопитающего, отличного от человека, которая трансгенна по продуцированию указанного нужного белка, продуцирование эмбрионов, отбор эмбрионов, имплантацию эмбрионов в матку млекопитающего-реципиента и моноторинг беременности вплоть до рождения трансгенного млекопитающего.
Изобретение относится также к способу продуцирования нужного белка трансгенным млекопитающим, отличным от человека, выполненному посредством технологического приема, включающего в себя суперовуляцию женской особи нетрансгенного млекопитающего, отличного от человека; искусственное оплодотворение млекопитающего спермой, полученной от мужской особи млекопитающего, отличного от человека, являющейся трансгенной по продуцированию указанного нужного белка, продуцирование эмбрионов, отбор эмбрионов, имплантацию эмбрионов в матку млекопитающего-реципиента и моноторинг беременности вплоть до рождения трансгенного млекопитающего.
Трансгенное млекопитающее, отличающееся тем, что продуцирует неожиданно высокие уровни нужного белка в своем молоке, может представлять собой, не ограничиваясь этим, животное рода быков. Трансгенные млекопитающие могут представлять собой и другой род млекопитающих, например, род свиней, род овец, род коз или род грызунов, но не ограничиваться этим. Нужный белок может представлять собой, но не ограничиваясь этим, гормон роста человека.
Изобретение далее относится к трансгенному млекопитающему, отличному от человека, относящемуся к роду быков, которое продуцирует рекомбинантный гормон роста человека в своем молоке, и геном которого содержит интегрированную плазмиду, и указанная плазмида содержит ген гормона роста человека и промотор бета-казеина, который направляет экспрессию указанного гена в клетки молочной железы млекопитающего.
Изобретение далее относится к трансгенному млекопитающему, которое продуцирует неожиданно высокие уровни hGH, хотя при этом до сих пор не показан физический рост, ожидаемый при таком высоком уровне продуцирования hGH. Так как трансгенные быки находятся под воздействием присутствующих гормонов роста человека, ожидалось, что расти животные должны быстрее, чем в норме растут нетрансгенные животные, и страдать от таких заболеваний, как сахарный диабет, гипертензия, увеличение риска кардиососудистых заболеваний и гипертрофия органов, включая печень, селезенку, почки и сердце. Такие высокие уровни hGH теоретически должны приводить к нежизнеспособному состоянию. Однако дело не в этом. Млекопитающее, такое как корова, с вызывающими тревогу повышенными уровнями чужеродного белка в крови, представляет собой совершенно здоровое животное, и исключительная продуктивность по количеству производимого рекомбинантного белка представляется неожиданным и технологически прогрессивным благоприятствующим фактором.
Рекомбинантный гормон роста человека согласно изобретению продуцируется в неожиданно высоких количествах. Уровень продуцируемого гормона роста человека приблизительно составляет более чем 1,0 г/л молока. Уровень продуцируемого hGH может составлять более чем приблизительно 2,0 г/л молока. Уровень продуцируемого hGH также может быть больше, чем приблизительно 3,0 г/л молока. В другом воплощении, уровень продуцируемого hGH может составлять больше, чем приблизительно 4,0 г/л молока. Однако в другом воплощении, уровень продуцируемого hGH может составлять более чем приблизительно 5,0 г/л молока. В следующем воплощении, уровень продуцируемого hGH может составлять более чем приблизительно 6,0 г/л молока. Однако в следующем воплощении, уровень продуцируемого hGH составляет приблизительно от 1,0 г/л молока приблизительно до 7,0 г/л молока. В следующем воплощении, уровень продуцируемого hGH составляет приблизительно от 2,0 г/л молока приблизительно до 6,0 г/л молока. Однако в другом воплощении, уровень продуцируемого hGH составляет приблизительно от 2,0 и приблизительно до 5,0 г/л молока.
В дополнение к этому, настоящее изобретение относится к способу выделения рекомбинантного гормона роста из молока трансгенного млекопитающего, а также к исследованию указанного гормона. Способы выделения могут включать в себя хроматографию и приемы по накоплению гормона. Могут применяться различные виды хроматографии, которые включают в себя ионообменную хроматографию, хроматографию с обращенной фазой, заместительную молекулярную хроматографию или аффинную хроматографию. Ионообменная хроматография может представлять собой анионообменную хроматографию. Аффинная хроматография может представлять собой иммуноаффинную хроматографию. Кроме того, может проводиться многократная хроматография.
Изобретение далее относится к способу выделения рекомбинантного гормона роста из молока трансгенного млекопитающего, отличного от человека, которое продуцирует рекомбинантный гормон роста, включая осветление молока трансгенного млекопитающего, отличного от человека, что приводит к тому, что молоко становится прозрачным, и проведение хроматографии прозрачного молока, что приводит к получению очищенного рекомбинантного гормона роста.
Изобретение далее относится к способу выделения рекомбинантного гормона роста из молока трансгенного млекопитающего, отличного от человека, которое продуцирует рекомбинантный гормон роста, и включающему в себя осветление молока трансгенного млекопитающего, отличного от человека, что приводит к тому, что молоко становится прозрачным, проведение анионообменной хроматографии осветленного молока на расширенном слое, что приводит к получению вещества после анионообменной хроматографии, проведение хроматографии с обращенной фазой после анионообменной хроматографии, что приводит к получению вещества после хроматографии с обращенной фазой, проведение после хроматографии с обращенной фазой анионообменной хроматографии, что приводит к получению вещества после анионообменной хроматографии, проведение после анионообменной хроматографии заместительной молекулярной хроматографии, что приводит к получению вещества после заместительной молекулярной хроматографии, концентрирование вещества после заместительной молекулярной хроматографии, что приводит к получению концентрированного вещества, и проведение заместительной молекулярной хроматографии концентрированного вещества, что приводит к получению очищенного рекомбинантного гормона роста.
Изобретение также относится к способу выделения рекомбинантного гормона роста из молока трансгенного млекопитающего, отличного от человека, которое продуцирует рекомбинантный гормон роста, и включающему в себя осветление молока, полученного от трансгенного млекопитающего, что приводит к тому, что молоко становится прозрачным, проведение иммуноаффинной хроматографии прозрачного молока, что приводит к получению вещества после иммуноаффинной хроматографии, проведение хроматографии с обращенной фазой после иммуноаффинной хроматографии, что приводит к получению вещества после хроматографии с обращенной фазой, проведение после хроматографии с обращенной фазой анионообменной хроматографии, что приводит к получению вещества после анионообменной хроматографии, проведение после анионообменной хроматографии молекулярной вытеснительной хроматографии, что приводит к получению вещества после молекулярной вытеснительной хроматографии, проведение после молекулярной вытеснительной хроматографии стадии концентрирования, что приводит к получению концентрированного материала, проведение молекулярной вытеснительной хроматографии концентрированного вещества, что приводит к очищенному рекомбинантному гормону роста.
Рекомбинантный гормон роста, описанный выше при описании способов очищения, может представлять собой, не ограничиваясь этим, гормон роста человека. Трансгенное млекопитающее может представлять собой, не ограничиваться этим, животное вида быков.
Последующие примеры иллюстрируют, без ограничения, способ и композиции согласно изобретению. Что совершенно понятно специалистам в данной области, применимы и другие подходящие модификации и адаптации различных условий и параметров, обычно встречающихся в ферментативном производстве химикалий и процедурах очистки белка, если они не выходят за рамки изобретения.
ПРИМЕР 1
Конструкция экспрессирующих плазмид
Авторы создали конструкцию, несущую большой сегмент бычьего гена промотора бета-казеина, включающий в себя 5ґ-концевой короткий фрагмент некодирующей области гена бета-казеина, слитый с кодирующей последовательностью гена гормона роста человека. Используемая в различных конструкциях область бета-казеина убывала приблизительно от 3,8 т.п.н. и приблизительно до 1,3 т.п.н. Ген hGH охватывает приблизительно от 2 до 2,2 т.п.н., в зависимости от того, включена ли в него сигнальная поли-A-последовательность.
Экспрессирующую кассету помещали в полилинкер обычного вектора клонирования типа pUC или pBS.
Этот промотор, в данном случае, такой как бета-казеин и гетерологичный hGH, обеспечивает специфику ткани и экспериментальную регуляцию экспрессии генов под его контролем.
Типичным представителем плазмиды является pRβhGH, которая несет в себе полную длину промотора бета-казеина быка, слитого с кодирующей последовательностью гена гормона роста человека.
Другие созданные конструкции получены главным образом из исходной, для улучшения селекции трансфекции клеток или улучшения эффективности интеграции ДНК в геном бычьей клетки.
На первом этапе, для того, чтобы облегчить селекцию, выполнялась ко-трансфекция с генетицин-резистентным геном, содержащим плазмиду, но на следующем этапе применяли другую конструкцию в сходном векторе, содержащем hGH-экспрессирующую кассету, несущую ген NPT для резистентности к неомицину. Примером такой плазмиды является pRNeo.
Для конститутивной экспрессии зеленого флуоресцирующего белка, была получена другая плазмида, которая включала в себя CMV-промотор, энхансер растительной природы (alfalfa) и ген зеленого флуоресцирующего белка из медузы A. victoria. Примером такой плазмиды является pRNeoGreen.
Более того, была создана другая плазмида. Она была сконструирована на основе Neo резистентной ген-содержащей плазмиды, в которую выше кодирующей области hGH была включена модифицированная короткая область промотора бета-казеина. Данной плазмидой является pVEβcashGH.
Были созданы другие конструкции, в которых область β-лактамазы была вырезана посредством рестрикции ферментом ApaLI, и после электрофореза в агарозном геле и гель-экстракции был выделен линейный фрагмент, содержащий полную экспрессирующую кассету.
Конструкции анализировали при помощи рестрикционных ферментов и определения последовательности ДНК, и их способность сопровождать экспрессию hGH была предварительно протестирована на линии клеток молочной железы посредством распознавания флуоресцентным антителом.
В качестве примера, в деталях будет описано создание плазмиды pVEβcashGH, включая генетические конструкции.
Создание плазмиды pVEβcashGH
Цель данной конструкции состоит в обеспечении минимального удлинения области промотора бета-казеина, в направлении специфического регулирования транскрипции гена hGH, присоединенного непосредственно книзу вместе с его сигнальным поли-А в организме хозяина.
Использование pVEX в качестве исходного вектора обеспечивает использование неомицин-резистентного гена, регулируемого посредством промотора tk, уже присутствующего в данной плазмиде. Ранее промотор SV40, содержащий 554 п.н., был удален посредством рестрикции ферментами StuI и NdeI, и сайт последнего заполнен ДНК-полимеразой Кленова, и самолигатурой полученного вектора.
Короткий промотор бета-казеина был получен после PCR-амплификации фрагмента 1,3 т.п.н. из области в 3,8 т.п.н. исходного гена промотора, посредством применения следующих олигомеров в качестве праймеров:
PB1 5ґ TCTACTCGAGGATCATCTATCTGTCCCAAAG(SEQ ID NO:1) и
PB2 5ґCTAGGATCCAATGATCTGATTTTGTGG(SEQ ID NO:2)
Данный фрагмент охватывает 1230 п.н. классического промотора плюс 49 п.н. первого, некодирующего экзона гена бета-казеина.
Фрагмент гена hGH получали посредством технических приемов PCR на исходном бычьем геномном клоне hGH, используя следующие олигомеры в качестве праймеров:
PB4 5ґCTAGGATCCATGGCTACAGGTAAGCGCC (SEQ ID NO:3) и
GHTE 5ґATGCTGTGTCTGGACGTCCT(SEQ ID NO:4)
У фрагмента промотора бета-казеина “затупляли концы” ферментом Кленова и включали в замещенный BamHI сайт pVEX. После отбора рекомбинантных клонов авторы выбирали определенное направление, предназначенное для использования уникального сайта HindIII, локализованного ниже в pVEX для включения кодирующей области hGH.
Для этой цели у фрагмента hGH также были “затуплены концы” и сайт плазмиды HindIII также был замещен. Отбирали клоны, которые содержали промотор бета-казеина и hGH, соответствующим образом слитые для экспрессирования hGH только под контролем данного промотора.
Размер указанной плазмиды составляет приблизительно 8,5 т.п.н.
Трансфекция соматических клеток
Плазмиды pRβhGH (вместе с другой плазмидой, содержащей генетицин-резистентный ген), pRNeo, pRNeoGreen или pVEβcashGH, использовали затем для трансфекции исходной культуры соматических клеток, применяя фосфат кальция или липосомный способ. Обычно для трансфекции использовали эмбриональные фибробласты теленка.
Трансфицированные клетки отбирали, добавляя в культуру генетицин. По прошествии периода от 2 до 8 недель клетки, резистентные к генетицину, считали подходящими для использования в качестве донорских клеток для получения трансгенных клонов. Клетки, отобранные для трансфекции, анализировали посредством PCR на содержание экспрессирующей кассеты, для уверенности в надлежащем переносе ядра для создания трансгенных эмбрионов.
ПРИМЕР 2
Энуклеация ооцитов и метафазный перенос ядра в зрелые энуклеированные ооциты
Сбор и созревание бычьих ооцитов In vitro
Бычьи ооциты отбирали (отсасывали) из яичников на скотобойне и доводили до стадии зрелости в TCM-199 + 5% FCS при 39°C в течение 24 часов. Среда созревания уравновешивалась CO2 по меньшей мере в течение 2 часов до начала ее использования. Зрелые ооциты отбирали смывом посредством интенсивного встряхивания в течение 2 минут в теплом TL-HEPES с 1 мг/мл бычьей гиалуронидазы семенников.
Перенос ядра с использованием клеток cumulus
Энуклеация (удаление ядер)
Ооциты энуклеировали механическим способом, применяя гидротехнические микроманипуляции Narishige и Nikon Diaphot микроскопию. Энуклеацию проводили в 20 микролитровых срезанных на конус и заостренных пипетках. Предварительно ооциты окрашивали в 5 мкг/мл красителя бисбензимидина (Hoechst 33342) (Sigma Chemical Co.,St. Louis, MO, USA) в течение 20 минут. Клетки в метафазе энуклеировали при визуализации окрашенных хромосом в ультрафиолетовом свете. Хромосомы в метафазе анализировали после аспирации внутри пипетки. Трансгенную соматическую клетку переносили в перивителлиновое пространство и располагали вплотную к энуклеированному ооциту.
Слияние
Трансгенную соматическую клетку и энуклеированный ооцит вручную выравнивали в камере слияния таким образом, чтобы мембраны, подготовленные для слияния, были параллельны электродам. Это было сделано при использовании стеклянной пипетки для проведения манипуляций с эмбрионами.
Слияние электрическим способом
Слияние проводили, используя электрический импульс 180 В/см в течение 15 мксек (манипулятор клеток BTX Electro 200) (BTX Inc., San Diego, Ca, USA), и проводили мониторинг при помощи оптимизатора-анализатора графических импульсов BTX. Импульсная камера для воздействия на эмбрионы состояла из двух проводов из нержавеющей стали по 0,5 мм в качестве электродов, установленных на расстоянии 0,5 мм на предметном стекле скользящей части микроскопа. Проводили мониторинг предполагаемых зигот в отношении слияния, лизиса и фрагментации.
Оценка развития компетенции
Зиготы оценивали через 48 часов после оплодотворения в отношении дробления, и через 7-9 дней - в отношении развития морулы или бластоцист.
ПРИМЕР 3
Культура клеток и эмбрионов
С целью упрощения процедур и увеличения степени выживаемости эмбриона в программе клонирования быка, в эксперименте исследовали различные донорские клетки, комплексы культур и варианты обработки реципиентного ооцита. В случае использования зрелых фибробластов в качестве донорских клеток, применяли три комплекса культуры для реконструкции эмбрионов: TCM-199 + 5% FCS, Menezo + 5% FCS (оба с клетками VERO в качестве сопутствующей культуры) и SOF без сопутствующей культуры, но с низкой концентрацией О2. Когда донорскими клетками были генетически и негенетически модифицированные эмбриональные фибробласты, в качестве среды для культуры реконструкции эмбрионов применяли также среду SOF. Наконец, когда в качестве донорских клеток использовали генетически модифицированные эмбриональные фибробласты, реципиентные ооциты предварительно обрабатывали росковитином (R) для задержки мейоза и оптимизации доступности реципиента. Ооциты доставали аспирацией из яичников на скотобойне и доводили до зрелого состояния в TCM-199 + 5% FCS при 39°C в течение 24 часов. Для группы ооцитов, обрабатываемых посредством R, их до созревания инкубировали с 25 мкМ R в TCM 199 + 5% FCS в течение 24 часов при 39°С. Зрелые ооциты лишали оболочки путем интенсивного встряхивания в течение 3 минут в TL HEPES с 1 мг/мл бычьей семенниковой гиалуронидазой. Выявляли метафазы, и ооциты энуклеировали при визуализации с Hoechst 33342 (5 мкг/мл) под УФ-светом (<6 секунд). В качестве донорских клеток использовали зрелые фибробласты, взятые от самца Ангусской породы скота и эмбриональные фибробласты от самки 45-дневного зародыша Джерсейской породы. Трансфекцию проводили с помощью конструкции, содержащей неомицин-резистентный ген и используя липосомы. После отбора с генетицином в течение 10-15 дней донорские клетки в фазах G0/G1 соединяли с энуклеированными ооцитами посредством электрического импульса. Через 3 часа активацию индуцировали путем инкубации в TL-HEPES с 5 мкМ иономицина в течение 4 минут и 2 мМ 6-DMAP в течение 3 часов. Ооциты затем промывали TL-HEPES и со-культурой в каждом из TCM-199 + 5% FCS + 10 г/л альбумин или Menezo + 2% FCS, оба с VERO клетками, или в среде SOF и в атмосфере 5% CO2 + 5% О2 + 90% N2. Обычно две бластоцисты переносили в организм реципиентной коровы нехирургическим путем и беременность определяли ультразвуковым исследованием на 30-35 день. Дробление (48 часов), развитие до бластоцист (дни от 7 до 9) регистрировали и анализировали посредством критерия Хи-квадрат. Темпы деления и развития до бластоцист были более высокими, когда эмбрионы культивировали в SOF. Однако различий в темпах развития беременности, в зависимости от разных условий культивирования или источника донорских клеток, не наблюдали. В суспензии мейотических 24-часовых клеток сохранялась компетентность реципиентных ооцитов. Следовательно, обработка росковитином может использоваться для увеличения доступности ооцитов для процедур ядерного переноса. См. таблицу 1 ниже.
Процентное содержание в колонках с различными показателями различно (P<0,05)
Коровы, подвергшиеся имплантации, нормально переносили беременность вплоть до естественного родоразрешения. При соответствующих условиях для родоразрешения возможно хирургическое вмешательство (кесарево сечение). Новорожденных кормили молозивом, обогащенным иммуноглобулином, в течение первых 48 часов, далее использовали сначала синтетическую, а позже естественную пищу (вся пища не содержала компонентов животного происхождения).
ПРИМЕР 4
Тесты, выполняемые на трансгенных телятах и продуцируемом рекомбинантном белке
В настоящем примере авторы изобретения приводят полное описание тестов, выполняемых на конкретном трансгенном теленке, который был получен как результат процедуры, описанной в примерах 1-3, и тестов продуцируемого им рекомбинантного белка. С другой стороны, должно быть ясно, что такие же анализы проводились на животных, которые родились как естественный результат других способов получения трансгенных телят, таких как те, что будут описаны в примерах 5 и 6 ниже.
Было подтверждено способами, основанными на реакциях PCR, выполняемыми на ДНК, выделенной из клеток белой крови теленка, с использованием ДНК из нетрансгенных телят Джерсейской породы в качестве отрицательного контроля, что промотор бета-казеина и кодирующий ген hGH включались в геном клеток трансгенного теленка. Они могут быть обнаружены вместе как уникальный ДНК-фрагмент, отличный от гомолога гена бета-казеина животного.
Применяя фармацевтический автоматический секвенатор, получили подтверждение того, что последовательность включенного гена на 100% соответствует кодирующему hGH гену. Она включает в себя интроны, секреторный сигнал и терминатор. Также была определена последовательность бычьего промотора бета-казеина, который контролирует экспрессию гена hGH у теленка. Все эти элементы точно совпадают с теоретически ожидаемой последовательностью из генетической конструкции, применяемой для трансформации клеток, из которых будут созданы клоны.
При известной точности генетического аспекта эксперимента авторы доказали, что продуцирование рекомбинантного белка есть явление ожидаемое, и каждая его физическая или химическая характеристика совпадает с соответствующей характеристикой природного hGH.
Для этого авторы должны были получить молоко от телки, так как бета-казеиновый промотор обеспечивает экспрессию рекомбинантного белка только в молочных железах, в то время, когда животное продуцирует молоко.
Поэтому трансгенную телку стимулировали введением гормона для начала продуцирования ею молока по истечении десяти месяцев после рождения. В это время телка имела вес 240 кг (приблизительно 530 фунтов).
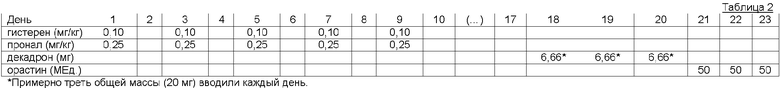
Первая фаза указанного стимулирования включает в себя подкожное комбинированное введение эстрогенов (эстрадиол бензоат, Histeren®, Instituto Rosenbusch) и прогестагены (медроксипрогестерон ацетат, Pronal®, Aton), включающее 5 следующих один за другим введения каждого лекарства в дозе 0,1 мг/кг и 0,25 мг/кг, соответственно, каждые 48 часов (то есть в дни 1, 3, 5, 7 и 9, с учетом того, что стимуляция начинается в 1 день).
Вторая фаза включала в себя подкожное введение дексаметазона (Decadron, Sidus) и окситоцина (Orasthin®, Hoechst Marion Roussel), полная сумма из 20 мг первого лекарства, будучи введенной в течение периода от 18 до 20 дня (третья часть от указанной полной массы каждый день), и 3 введения по 50 МЕ. последнего из двух лекарств в дни от 21 до 23.
Вышесказанная информация суммирована в таблице 2:
Как и ожидали, корова начинала продуцировать молозиво на следующий день после окончания стимулирования, а затем, постепенно, по качеству продуцируемая жидкость превращалась в молоко. Собранная жидкость должным образом сохранялась и основательно анализировалась.
Несколько тестов, результаты которых показаны на фиг.1-4, проводили на молозиве и молоке (для упрощения, и молозиво, и молоко в дальнейшем в этом документе будут именоваться как “молоко", исключая те случаи, когда различие должно быть подчеркнуто). Во-первых, измеряли объем собираемого молока. Первоначальная продуктивность молока (первые пять дней лактации) составила приблизительно 1650 мл/день. В течение первого месяца продуцирования молока проводили два разовых надоя вручную в день, один - утром и один - после обеда (вклад каждого вымени в итоговый объем мог быть учтен), хотя со второго месяца и в дальнейшем, когда продуцирование молока стало увеличиваться, проводили три разовых надоя в день (утром, в обед и после обеда). Дневные объемы увеличивались более или менее постоянно, до достижения почти 10000 мл через три месяца после первого разового надоя. Детальная информация, касающаяся этой темы (фиг.1A), представлена на кривой суточных объемов молока по датам (фиг.1B).
Параллельно с измерениями объема молока проводили микробиологические анализы молока, результаты которых (фиг.2A) показаны на соответствующих графиках КОЕ на мл (Колониеобразующие единицы), в зависимости от даты (фиг. 2B-2C).
Биологическую активность (hGH) также анализировали посредством анализа биологической активности in vitro в культуре клеток NB2 (фиг.3A-3B). Доказано, что биологическая активность рекомбинантного hGH, продуцируемого в молоке первотелки, соответствует, в пределах уровня ошибки способа, нормальным величинам активности природного hGH человека. Более того, у полученного молока, анализируемого методом Вестерн-блоттинга, обнаружили в основной зоне hGH, соответствующий интактному. Дополнительные минорные зоны соответствовали расщепленным вариантам и агрегатам, как и ожидалось в таких системах продуцирования.
Вместе с информацией, относящейся к биологической активности и дневным объемам, с применением уравнения 1, возможен подсчет дневного продуцирования hGH, где mhGH - это масса продуцируемого за день hGH в миллиграмах, BA - биологическая активность в МЕд., SA - специфическая активность hGH (3 МЕд./мг), и V - суточный объем собираемого молока. Данные показаны на фиг.4A. График суточной массы hGH показан на фиг.4B. Для того чтобы обосновать визуальную корреляцию между суточной массой hGH и суточным объемом молока, последний также был нанесен на график на фиг.4B.
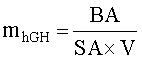
Из этой информации можно понять, что суточная масса hGH в молоке и суточный объем молока, по мере развития коровы, имеют возрастающую тенденцию (причем первый увеличивается в большей пропорции), которая представляет собой направленную вверх тенденцию продуктивности рекомбинантного гормона (грамм hGH на литр молока). Нужно отметить, что продуктивность увеличивается от приблизительно 2 г hGH на литр (первый разовый надой), когда продуцировалось молозиво, до 5 г hGH в среднем на литр молока, от второго месяца лактации до четвертого. Таким образом, получена неожиданно высокая полезная продуктивность гормона роста человека, так же как и продуктивность, более или менее постоянно увеличивалось качество жидкости, и молозиво превращалось в молоко.
Хотя в данное время масса hGH, продуцируемого за день, является безусловно заметной величиной, нужно отметить, что пока корова не полностью выросла, тенденция увеличения массы рекомбинантного белка, продуцируемого за день, будет продолжаться и в будующем, пока не достигнет своего максимума.
Параллельно с тестами, проводимыми на молоке, различные анализы проводили на сыворотке коровы, результаты этих анализов представлены на фиг.5А-5С. Сначала проводили измерения концентрации hGH в сыворотке коровы. Результаты (фиг.5А) нанесены на график, вместе с суточной массой hGH в молоке, что позволяет сравнить обе величины (фиг.5В).
Ссылочная область изменения GH в сыворотке (у человека) составляет 0,06-5 нг/мл. Предполагая наличие сходного диапазона изменения у быков, нужно отметить, что, исключая начальную область, полная кривая для hGH в сыворотке трансгенных коров, при сравнении данных, располагается значительно выше верхнего предела указанной области изменения. Данный факт, тем не менее, нужно принимать во внимание, и, хотя hGH и соответствующий бычий гормон роста (bGH) очень схожи в отношении их аминокислотной последовательности и 3D-структуры, первый, хотя и полностью функциональный, не в полной мере активен у крупного рогатого скота.
Следовательно, чтобы оценить потенциальный риск состоянию здоровья коровы, связанный с высоким уровнем hGH в ее сыворотке, параллельно проводили измерения концентрации в сыворотке вещества IGF-1 (инсулиноподобный фактор роста 1, известный также как соматомедин C) (фиг.5A). Гормон роста осуществляет свои функции прежде всего через IGF-1, который синтезируется в печени. Поскольку IGF-1 значительно содействует делению клетки in vivo и метаболическому эффекту гормона роста, оценка IGF-1 является важным диагностическим инструментом для косвенного определения предполагаемых нарушений функции гормона роста. Таким образом, IGF-1 представляет собой зависимый индикатор биологического усвоения гормона роста. Результаты этих анализов представлены на фиг.5C, вместе с одновременным соотнесением к hGH, что позволяет одновременно визуализировать обе величины.
Более того, величину IGF-1 и hGH измеряли в сыворотке группы нетрансгенных коров, чтобы иметь контрольную группу и, таким образом, выяснить сопоставимость с данными, соответствующими трансгенной корове. Средние величины измерений обоих белков для нетрансгенной группы и трансгенной коровы приведены в таблице 3.
Важно заметить, что средняя концентрация hGH в сыворотке у нетрансгенной группы заключена в пределах предполагаемой ссылочной области, с учетом того факта, что среднее для трансгенной коровы значительно выше верхнего предела указанной области. С другой стороны, бесспорно, что средний уровень IGF-1 в сыворотке трансгенных животных существенно выше, чем соответственно у нетрансгенных коров, что составляет фундаментальное различие.
Следовательно, несмотря на то, что высокая концентрация hGH (который, как известно, в человеческом организме ведет к нарушениям с тяжелыми последствиями, такими как сахарный диабет, гипертензия, увеличение риска кардиососудистых заболеваний, гипертрофия органов, включая печень, селезенку, почки и сердце) в сыворотке трансгенных коров теоретически должна приводить животного в нежизнеспособное состояние, очевидно, это не должно представлять помеху здоровью и хорошему самочувствию. Корова с тревожно высокими уровнями чужеродного гормона в крови, но при этом абсолютно здоровая и исключительно продуктивная по количеству производимого продукта рекомбинантного белка, представляется неожиданным и технологически прогрессивным благоприятствующим явлением.
Другим новаторским аспектом согласно изобретению является то, что рекомбинантный hGH, который входит в кровоток коровы, стимулирует молочную железу продуцировать больше молока. Этот эффект достижим в результате действия IGF-1. Эта молекула увеличивает кровоток через молочную железу, обеспечивая крайне необходимый предшественник для синтеза молочного жира, белка и лактозы. Таким образом, IGF-1 направляет питание через кровь к клеткам вымени, где он помогает продуцированию молока. Таким образом достигается самостимуляция животного, так как рекомбинантный hGH, продуцируемый в молоке трансгенной коровы, способствует поддержанию увеличения объема молока, продуцируемого животным через стимуляцию молочной железы с соответствующей большей секрецией hGH в молоке.
ПРИМЕР 5
Получение трансгенных телят посредством субклонирования
Пять образцов ткани были взяты из уха трансгенного теленка с применением иглы диаметром 1,5 мм, конец которой для этой цели предварительно был срезан на конус. Образцы транспортировали с охлаждением в лабораторию в среде с PBS-основой, содержащей антибиотики и фунгициды.
После этого ткань образцов инкубировали в течение 72 часов в среде MEM с 10% бычьей эмбриональной сывороткой и антибиотиками при 39°C и в атмосфере 5% CO2. В конечном итоге ткань образцов удаляли и фибробластам у периферийной границы чашки позволяли расти до степени смыкания монослоя. Как только это было достигнуто, фибробласты инкубировали по меньшей мере 5 дней без замены культуральной среды для того, чтобы достигнуть их синхронизации в стадии G0, что анализировали способом визуализации под микроскопом.
После обработки трипсином единичные фибробласты были слиты с энуклеированными бычьими ооцитами, согласно примеру 2, и полученные таким образом эмбрионы культивировали в среде SOF и в атмосфере, содержащей 5% CO2 + 5% O2 + 90% N2 до стадии бластоцисты. После этого обычно две бластоцисты переносили нехирургическим путем в реципиентную корову и беременность определяли на 30-35 день ультразвуковым исследованием.
Коровы, подвергшиеся имплантации, нормально переносили беременность вплоть до естественного разрешения. При соответствующих условиях, для разрешения, возможно хирургическое вмешательство (кесарево сечение). Новорожденных кормили молозивом, обогащенным иммуноглобулином, в течение первых 48 часов, далее использовали сначала синтетическую, а позже естественную пищу (вся пища не содержала компонентов животного происхождения).
На фиг.6A и 6B показана концентрация hGH и IGF-1, измеренная в сыворотке и проводимая параллельно на двух трансгенных животных, полученных субклонированием трансгенных коров, и результаты этих анализов изображены на фиг.6C и 6D соответственно для получения одновременной визуализации обеих величин для каждого животного. Так как в это время телята еще молоды, значения обеих величин, и hGH и IGF-1, все еще располагаются в соответствующей области ссылки, но вполне ожидаемо, что они будут расти таким же образом, как у трансгенной коровы, от которой эти два клона получены.
ПРИМЕР 6
Получение трансгенных телят искусственным оплодотворением трансгенной коровы, подвергшейся суперовуляции
В данном примере будет представлен альтернативный подход для получения трансгенных быков. Этот способ включает в себя суперовуляцию трансгенной коровы посредством гормональной обработки, искусственное оплодотворение указанной коровы, сбор полученных таким образом эмбрионов, имплантация указанных эмбрионов в суррогатную корову и развитие беременности вплоть до рождения животных. Описание данной процедуры представлено ниже.
Суперовуляция
Утром первого дня (то есть дня, когда процедура стартовала) 150 мкг простагландина (D(+)-clorprostenol, Arsaprost®, Arsa) было введено трансгенной корове посредством внутримышечной инъекции. Животное содержали на диете, состоящей из 4 кг дневного рациона, включающего в себя приблизительно 15% белков (перед началом процедуры животному давали по 2 кг пищи с тем же содержанием белков). Утром на восьмой день внутри влагалища размещали прибор контроля высвобождения лекарственного средства CIDR, в данном случае прогестерона. Вместе с этим, внутримышечно вводили 50 мг прогестерона и эстрадиола. Количество пищи, употребляемое животным, возрастало до 6 кг/день с тем же содержанием белков. Затем в дни 12, 13 и 14 были сделаны две внутримышечные инъекции FSH и LH (PLUSET®, Calier) (одна утром, а другая после обеда). На следующий день (15-й день) обработка продолжалась внутримышечным введением двух инъекций PLUSET® (одна утром и другая после обеда) и двух инъекций по 150 мкг простагландина каждая (в том же режиме введения). На утро 16-го дня CIDR удаляли. Общее количество PLUSET®, введенного в течение фазы суперовуляции, равнялось 350 МЕд.
Оплодотворение
Данная фаза включает в себя три поочередных инъекции (первая и вторая - утром и после обеда соответственно на 17 день, и последняя - утром на 18 день) семенной жидкости от донора, быка Джерсейской породы, которую получали предварительно и хранили в заморозке, чтобы сохранить жизнеспособность сперматозоидов.
Сбор эмбрионов и их имплантация в суррогатных коров
Эмбрионы собирали посредством вымывания струей из маточных рогов коровы с 1 л DMPBS (Nutricell®) утром на 26 день. Сразу после этого два эмбриона переносили нехирургическим путем в реципиентную корову и беременность определяли на 30-35 день посредством ультразвукового обследования.
Коровы, подвергшиеся имплантации, нормально переносили беременность вплоть до естественного разрешения. При соответствующих условиях для разрешения возможно хирургическое вмешательство (кесарево сечение). Новорожденных кормили молозивом, обогащенным иммуноглобулином, в течение первых 48 часов, далее использовали сначала синтетическую, а позже естественную пищу (вся пища не содержала компонентов животного происхождения).
Так как рождение биологического приплода подчиняется закону Менделя, половина животных, рожденных вследствие процедуры, описанной выше, должны быть трансгенными, и половина из них должны быть мужскими особями, а другая половина - женскими. Следовательно, существует высокая вероятность получения трансгенной мужской особи (животного-основателя), семенная жидкость которого может быть использована для размещения в Master Bank трансгенной семенной жидкости Джерсейской породы, для использования ее с целью оплодотворения трансгенных/нетрансгенных коров, подвергнутых суперовуляции, с тем, чтобы увеличить трансгенное стадо.
ПРИМЕР 7
Выделение рекомбинантного hGH из молока
Чтобы проверить правильность приблизительного молекулярного веса, результатов Вестерн-блоттинга и биологической активности рекомбинантного hGH, продуцируемого в молоке коров, проведен полный процесс выделения, для чего необходимо, чтобы при производстве биофармацевтического продукта нужный белок был очищен до состояния гомогенности, чтобы избежать присутствия возможных примесей в указанном продукте. Этот процесс включает в себя следующие этапы: получение обезжиренного молока способом центрифугирования и разведения полученного супернатанта для достижения лучшей растворимости рекомбинантного hGH в результате удерживания в мицеллах казеина (осветление), пропускание этого раствора через колонку анионообменной хроматографии в расширенном слое (альтернативой этому этапу является применение иммуноаффинной хроматографии, см. пример 8, ниже), полученный раствор уменьшали этапом с обращенной фазой HPLC (C4), после этого фракции с большим содержанием рекомбинантного hGH подвергали анионообменной хроматографии.
Очищенный материал обессоливали, концентрировали и подвергали молекулярной заместительной хроматографии. Для получения чистого рекомбинантного hGH, на этом этапе вещество разделяется по молекулярному весу.
Процедура выделения гормона роста человека (hGH) из молока включает в себя следующие этапы: (a) осветление, (b) анионообменную хроматографию в расширенном слое, (c) хроматографию с обращенной фазой, (d) анионообменную хроматографию, (e) заместительную молекулярную хроматографию (обессоливание), (f) концентрирование и (g) заместительную молекулярную хроматографию.
Осветление
Свежее молоко смешивали с достаточным количеством Tween 80 для того чтобы получить 0,5% раствор. После добавления Tween 80, прибавляли 2M Tris-HCl до pH 7,3±0,1. После этого продукт гомогенизировали в течение 30 минут, а затем центрифугировали при 14000 g, для того чтобы отделить слой жира. Затем полученный раствор разводили с 0,5% Tween 80 до проводимости менее чем или равной 1500 мкСм/см. pH доводили до 7,3±0,1 добавлением 2M Tris-HCl, а затем продукт фильтровали через пористую мембрану с размером пор 0,8 мкм и хранили в удобном виде.
Анионообменная хроматография в расширенном слое
Материал, полученный на предыдущем этапе, хроматографировали, используя анионообменный матрикс, согласно следующим параметрам:
1. Оборудование:
A. Колонка:
1) Диаметр: 5 см.
2) Высота слоя: 30 см (компановка слоя).
3) Матрикс:
a. Проточный Q XL (Amersham).
b. Объем: 600 мл.
2. Растворы и буферы:
А. 0,5н. NaOH.
В. 20% этанол.
C. буфер A: 20 мм Tris.HCl, pH 7,3.
D. буфер B: 500 мм Tris.HCl, pH 7,3.
E. буфер C: 20 мМ Tris.HCl, 150 мМ NaCl, pH 7,3.
F. буфер D: 20 мМ Tris.HCl, 500 мМ NaCl, pH 7,3.
3. Материал для хроматографии:
A. Осветленное молоко.
B. Свойства образца:
1) Объем: 25±5 л.
2) Проводимость: <1500 мкСм/см.
3) pH: 7,3±0,1.
Для уравновешивания колонки через нее пропускали 1,5 объема колонки ("vc") (900 мл) дистиллированной воды, со скоростью элюирования 115±5 см/час (нисходящее течение). После этого следующие растворы или буферы в количествах, приведенных далее, последовательно пропускали через колонку со скоростью течения 230±30 см/час (восходящее течение): 3,0 vc (1800 мл) 0,5н NaOH, 3,0 vc (l800 мл) дистиллированной воды, 1,0 vc (600 мл) буфера B, и, в конце, 3,0 vc (1800 мл) буфера A.
Как только колонка уравновешивалась, загружали материал для хроматографии. Указанную загрузку производили при температуре 12±3°C и при скорости элюции 230±30 см/час. После этого проводили элюирование со скоростью 115±15 см/час и при той же температуре. Во-первых, через колонку пропускали значительный объем буфера А (восходящим течением), во-вторых, растворы и буферы, далее приведенные, пропускали в следующем порядке (нисходящим течением): 1,5 vc (900 мл) буфера А, 2,0 vc (1200 мл) буфера С, и, в конце, 2,0 vc (1200 мл) буфера D.
Как только этот этап заканчивался, для того чтобы очистить колонку, через нее последовательно пропускали следующие растворы или буферы в количествах, приведенных ниже: 1,5 vc (900 мл) дистиллированной воды, 1,5 vc (900 мл) 0,5н. NaOH, 2,0 vc (1200 мл) дистиллированной воды, 1,0 vc (600 мл) буфера B, 1,5 vc (900 мл) дистиллированной воды и, в конце, 1,5 vc (900 мл) 20% этанола.
Отобранные фракции, содержащие hGH, анализировали на совокупный белок (способом Bradford) и на нужный белок (посредством RIA) и хранили при температуре 2-8°C.
Хроматография с обращенной фазой
Материал, полученный на предыдущем этапе, хроматографировали согласно следующим параметрам:
1. Оборудование:
A. Колонка:
1) Диаметр: 4 см.
2) Высота слоя: 48 см.
3) Матрикс:
a. BakerBond-бутил широкопористый с размером пор в 15 мкм, подготовленный для ЖХ-колонки (Baker).
b. Объем: 600 мл.
2. Растворы и буферы:
A. Мобильная фаза 1 (MP1): 30 мМ NaHCO3, pH 7,2: дистиллированная вода: ацетонитрил (35:55:10).
B. Мобильная фаза 2 (MP2): 30 мМ NaHCO3, pH 7,2: дистиллированная вода:ацетонитрил(20:10:70).
C. 50% метанол.
3. Материал для хроматографии:
A. Пул отобранных фракций, полученных на предыдущем этапе.
B. Свойства образца:
1) Объем: 30±15 л.
2) pH: 7,3±0,3.
Для уравновешивания колонки последовательно пропускали через нее следующие растворы или буферы в количествах, приведенных ниже, со скоростью, меньшей чем или равной 478 см/час: 0,3 vc (180 мл) 50% метанол, после этого применяли градиент 50% метанол-MP2, начиная от соотношения 100:0 указанных растворов, до достижения соотношения 0:100 указанных растворов в суммарном объеме 1,0 vc (600 мл), как только градиент заканчивался, через колонку пропускали 1,0 vc (600 мл) MP2, далее применяли градиент MP2-MP1, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 1,0 vc (600 мл), и, наконец, через колонку пропускали 2,0 vc (1200 мл) MP1.
Как только колонка уравновешивалась, материал для хроматографии фильтровали через пористую мембрану с размером пор 0,45 мкм и после этого немедленно загружали в колонку. Указанную загрузку проводили при температуре 20±5°C и при скорости течения, меньшей или равной 238 см/час. После этого проводили элюирование при скорости 478±78 см/час и при той же температуре и растворы и буферы, приведенные ниже, пропускали в следующем порядке: 1,0 vc (600 мл) MP1, градиент MP1-MP2, начиная от соотношения указанных растворов 65:35 до достижения соотношения указанных растворов 45:55 в суммарном объеме 18,0 vc (9000 мл), 1,0 vc (600 мл) MP1-MP2 в соотношении 45:55, и, в конце, 2,0 vc (1200 мл) MP2.
Как только этот этап заканчивался, для того чтобы очистить колонку, применяли градиент MP2-50% метанол, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 1,0 vc (600 мл), и, в заключение, через колонку пропускали 2,0 vc (1200 мл) 50% метанол.
Фракции, полученные после этой хроматографии, по анализу методом ДСН-ПААГ оказались гомогенными на 20% для окисленного hGH, и проводили отбор, в зависимости от результатов. После этого отобранные фракции, содержащие hGH, анализировали на содержание суммарного белка (способом Bradford) и хранили при температуре 2-8°C.
Анионообменная хроматография
Материал, полученный после предыдущего этапа, хроматографировали, используя анионообменный матрикс, такой как следующий:
1. Оборудование:
A. Колонка:
1) Диаметр: 5 см.
2) Высота слоя: 25 см.
3) Матрикс:
а. Source 30Q (Pharmacia).
b. объем: 500 мл.
2. Растворы и буферы:
A. 20% этанол.
B. Раствор K: 0,5н. NaOH, 3M NaCl.
C. Раствор L: 50 мМ Tris, pH 7,50.
D. Раствор M: 0,1н. HCl, 3 M NaCl.
E. Мобильная фаза 3 (MP3): Раствор L: ацетонитрил (70:30).
F. Мобильная фаза 4 (MP4): 50 мМ Tris, 0,1M NaCl, pH 7,50: ацетонитрил (70:30).
3. Материал для хроматографии:
A. Отобранные фракции, полученные на предыдущем этапе.
B. Свойства образца:
1) Объем: 4,5±1 л.
2) pH: 7,2±0,2.
Чтобы уравновесить и санировать колонку, через нее последовательно пропускали следующие растворы или буферы в количествах, приведенных далее, со скоростью течения, меньшей или равной 183±20 см/час: 1,0 vc (500 мл) дистиллированной воды, 1,0 vc (500 мл) раствора K, 1,0 vc (500 мл) раствора L и, в заключение, 1,0 vc (500 мл) MP3.
Как только колонка уравновешивалась, загружали материал для хроматографии. Указанную загрузку проводили при температуре 20±5°C и скорости течения, меньшей или равной 183 см/час. После этого проводили элюирование при скорости течения 183±20 см/час и при той же температуре, затем пропускали растворы и буферы, представленные далее, в следующем порядке: 1,0 vc (500 мл) MP3, градиент MP3-MP4, начиная от соотношения указанных растворов 15:85 до достижения соотношения указанных растворов 25:75 в суммарном объеме 5,0 vc (2500 мл), и, в заключение, через колонку пропускали 2,0 vc (1000 мл) MP4.
Как только этот этап заканчивался, для того чтобы очистить колонку, через нее последовательно пропускали следующие растворы или буферы, в количествах, представленных далее в этом документе: градиент MP4-дистиллированная вода, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 0,5 vc (250 мл), 0,5 vc (250 мл) дистиллированной воды, градиент дистиллированная вода-раствор K, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 0,5 vc (250 мл), 1,0 vc (500 мл) раствора K, градиент раствор К-дистиллированная вода, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 0,5 vc (250 мл), 0,5 vc (250 мл) дистиллированной воды, градиент дистиллированная вода-раствор M, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 0,5 vc (250 мл), 1,0 vc (500 мл) раствора M, градиент раствор M-дистиллированная вода, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 0,5 vc (250 мл), 1,0 vc (500 мл) дистиллированной воды, градиент дистиллированная вода-раствор L, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 0,5 vc (250 мл), 0,5 vc (250 мл), 0,5 vc (250 мл) раствора L, градиент раствор L-дистиллированная вода, начиная от соотношения указанных растворов 100:0 до достижения соотношения указанных растворов 0:100 в суммарном объеме 0,5 vc (250 мл), и, в заключение, 1,5 vc (750 мл) дистиллированной воды.
Отобранные фракции, содержащие hGH, анализировали на суммарный белок (способом Bradford) и хранили при температуре 2-8°C.
Молекулярная вытеснительная хроматография
Материал, полученный на предыдущем этапе, хроматографировали, применяя следующий молекулярный заместительный матрикс:
1.Оборудование:
A. Колонка:
1) Диаметр: 5 см.
2) Высота слоя: 25 см.
3) Матрикс:
а. Cellufine GH25 (Millipore).
b. Объем: 500 мл.
2. Растворы и буферы:
А. 0,5н. NaOH.
B. 20% этанол.
C. Буфер C: 150 мМ NaH2PO4, pH 7,2.
D. Буфер G: 320 мМ глицин, 10 мМ NaH2PO4, 0,1% Tween 80, pH 6,9.
3. Материал для хроматографии:
А. Отобранные фракции, полученные на предыдущем этапе.
B. Свойства образца:
1) Объем: 0,5±0,2 л.
2) pH: 7,5±0,5.
Чтобы уравновесить и санировать колонку, через нее последовательно пропускали следующие растворы или буферы в количествах, приведенных далее, со скоростью течения, меньшей или равной 180 см/час: 1,0 vc (500 мл) дистиллированной воды, 1,0 vc (500 мл) 0,5н. NaOH, 0,5 vc (250 мл) дистиллированной воды, 0,5 vc (250 мл) буфера C и, в заключение, 2,0 vc (1000 мл) буфера G.
Как только колонка уравновешивалась, загружали материал для хроматогафии. Указанную загрузку проводили при температуре 20±5°C и скорости течения равной 183 см/час. После этого проводили элюирование при той же скорости течения и при той же температуре, через колонку пропускали 1,0 vc (500 мл) Буфера G и делали столько прогонов, сколько необходимо.
Как только этот этап заканчивался, для того чтобы очистить колонку, через нее последовательно пропускали следующие растворы или буферы, в количествах, представленных далее в этом документе: 0,5 vc (250 мл) дистиллированной воды, 1,0 vc (500 мл) 0,5н. NaOH, 0,5 vc (250 мл) дистиллированной воды, 0,5 vc (250 мл) буфера C, 0,5 vc (250 мл) дистиллированной воды и, в заключение, 1,5 vc (750 мл) 20% этанола.
Отобранные фракции, содержащие hGH, анализировали на суммарный белок и хранили при температуре 2-8°C.
Концентрирование
Фракции, полученные на предыдущем этапе, концентрировали, согласно условиям, описанным ниже:
1. Оборудование:
A. Перистальтический насос: Watson Marlow - Cat. No.302S.
B. Насосные трубки: Marlow - Cat. No.902.0080.016.
C. Концентратор: Prep Scale Millipore - Cat. No.CDU F006LC.
2. Растворы и буферы:
A. 0,28% додецилсульфат натрия (SDS).
B. 0,06% Тритон.
C. 0,125н. NaOH.
D. Буфер G: 320 мМ Глицин, 10 мМ NaH2PO4, 0,1% Tween 80, pH 6,9.
3. Материал для обработки:
A. Отобранные фракции, полученные на предыдущем этапе.
B. Свойства образца:
1) Объем: 1,0±0,5 л.
2) Проводимость: 1200±100 мкСм/см.
3) pH: 6,9±0,1.
Во-первых, оборудование очищали, санировали и уравновешивали, растворы и буферы в следующей последовательности протекали через оборудование: 2 л 0,125н NaOH, 10 л дистиллированной воды и, в заключение, 2 л буфера G. После этого оборудование готово к использованию для концентрирования отобранных фракций, следуя принятой технологии. Процедуру концентрирования проводили до достижения концентрации белка 15 мг/мл (анализировали способом Bradford).
Отобранные фракции фильтровали через мембрану с порами размером 0,22 мкм, анализировали на суммарный белок (способом Bradford) и хранили при температуре 4°C.
Проводимость и pH отобранных фракций равнялись соответственно 1100-1300 мкСм/см и 6,9±0,1.
Молекулярная заместительная хроматография
Материал, полученный на предыдущем этапе, хроматографировали, используя следующий матрикс молекулярного замещения:
1. Оборудование:
A. Колонка:
1) Диаметр: 5 см.
2) Высота слоя: 92 см.
3) Матрикс:
а. Сефакрил S-200 высокого разрешения (Amersham Pharmacia).
b. Объем: 1800 мл.
2. Растворы и буферы:
A. 0,5н NaOH.
B. 20% этанол.
C. Буфер H: 320 мм Глицин, 2,2 мМ NaH2PO4, 1,8 мМ Na2HPO4, pH 7,30.
3. Материал для хроматографии:
A. Отобранные на предыдущем этапе и концентрированные фракции.
B. Свойства образца:
1) Объем: 40±20 мл.
2) Проводимость: 1200±100 мкСм/см.
3) pH: 7,3±0,1.
Чтобы уравновесить и санировать колонку, через нее последовательно пропускали следующие растворы или буферы в количествах, приведенных далее, со скоростью течения менее чем 46 см/час: 1,0 vc (1800 мл) дистиллированной воды, 1,0 vc (1800 мл) 0,5н. NaOH и, в заключение, - 2,0 vc (3600 мл) буфера H.
Как только колонка уравновешивалась, загружали материал для хроматографии. Указанную загрузку проводили при температуре 20±5°C и скорости течения 46±15 см/час. После этого проводили элюирование при той же скорости течения и температуре, через колонку пропускали 1,0 vc (1800 мл) буфера H и делали столько прогонов, сколько необходимо.
Как только этап заканчивался, чтобы очистить колонку, через нее последовательно пропускали следующие растворы или буферы в количествах, приведенных далее: 1,0 vc (1800 мл) дистиллированной воды и 1,5 vc (2700 мл) 20% этанола.
Фракции, содержащие чистый hGH, асептически фильтровали через мембрану с порами размером 0,22 мкм в стерильные апирогенные пластиковые бутылки, анализировали на суммарный белок и хранили при температуре -20°C.
ПРИМЕР 8
Альтернативная процедура выделения hGH из молока
Для того чтобы выделить рекомбинантный hGH, содержащийся в молоке, вместо ранее описанного способа выделения может быть применена альтернативная схема. Основное различие между процедурой, описанной в примере 7, и альтернативной процедурой, представленной в данном примере, заключается в том, что второй этап первого из способов включает в себя анионообменную хроматографию в расширенном слое, в то время как соответствующий этап альтернативного способа содержит в себе иммуноаффинную хроматографию. Этапы осветления в обоих способах также незначительно отличаются. Так как остальные этапы у обеих схем выделения идентичны, ниже будут описаны только два первых этапа альтернативной процедуры.
Осветление
Свежее молоко смешивалось со значительным количеством Tween 80, чтобы получить 0,5% раствор. После добавления Tween 80, был добавлен 1M Tris, чтобы довести pH до 7,3±0,3. После этого продукт гомогенизировали в течение 30 минут и центрифугировали при 14000 g, чтобы отделить слой жира. Затем полученный раствор разбавляли 20-кратно буфером S (50 мМ Tris HCl, 500 мМ NaCl, 0,5% Tween 80, pH 7,3), потом фильтровали через мембрану с порами размером 0,45 мкм и хранили в удобном виде.
Иммуноаффинная хроматография
Материал, полученный на предыдущем этапе, хроматографировали, используя матрикс иммуноаффинного взаимодействия (Аффигель 10 Сложный эфир Агарозы, изготовленный BioRad, ковалентно соединенный с анти-GH моноклональными антителами, изготовленными Bio Sidus) согласно следующим параметрам:
1. Оборудование:
A. Колонка:
1) Диаметр: 30 см.
2) Высота слоя: 15 см.
3) Матрикс:
a. Аффигель 10 Сложный эфир Агарозы (BioRad), с ковалентно присоединенными анти-GH моноклональными антителами (Bio Sidus).
b. Объем: 10 л.
2. Растворы и буферы:
A. Буфер A: 50 мМ Tris.HCl, 500 мМ NaCl, pH 7,2.
B. Буфер B: 100 мМ лимонная кислота, pH 3,0.
C. Буфер C: 150 мМ NaH2PO4, pH 7,2.
D. Буфер D: 50 мМ Tris.HCl, 500 мМ NaCl, 500 мМ гуанидин.HCl, pH 7,2.
E. Буфер E: 50 мМ Tris.HCl, 500 мМ NaCl, pH 7,2, 0,2% азид натрия, гентамицин 0,1 г/л.
3. Материал для хроматографии.
A. Осветленное молоко.
B. Свойства образца:
1) Объем: 30-50 л.
2) Проводимость: 45±15 мСм/см.
3) pH:7,3±0,3.
Для уравновешивания и санирования колонки, если она не была использована в течение последних семи дней, через нее последовательно пропускали следующие растворы или буферы в количествах, приведенных далее, со скоростью течения менее чем 51 см/час: 1,0 объем, равный объему колонки ("vc") (10 л) буфера A, 2,0 vc (20 л) буфера D, 2,0 vc (20 л) буфера A, 1,0 vc (10 л) буфера B, 2,0 vc (20 л) буфера C и, в заключение, 1,0 vc (10 л) буфера A.
С другой стороны, если колонку использовали в последние семь дней, через нее последовательно пропускали следующие растворы или буферы в количествах, приведенных далее, со скоростью течения менее чем 51 см/час: 1,0 vc (10 л) буфера C и, в заключение, 2,0 vc (20 л) буфера A.
Как только колонка уравновешивалась, в нее загружали материал для хроматографии. Указанную загрузку проводили при температуре 5±3°C и при скорости течения менее чем 51 см/час. После этого проводили элюирование при скорости течения 42±9 см/час и при той же температуре и пропускали растворы и буферы в порядке и количестве, приведенном далее: 2,0 vc (20 л) буфера A и 1,5 vc (15 л) буфера B.
Как только этот этап заканчивался, для того чтобы очистить колонку, через нее последовательно пропускали следующие растворы или буферы в количествах, приведенных далее: 2,0 vc (20 л) буфера D, 2,0 vc (20 л) буфера A и, в заключение, 2,0 vc (20 л) буфера E.
Отобранные фракции, содержащие hGH, анализировали на суммарный белок (способом Bradford) и на нужный белок (посредством RIA) и хранили при температуре 4°C.
Пример 9
Качественный контроль чистого рекомбинантного hGH
Две партии чистого рекомбинантного hGH подвергали серии анализов для выяснения, чем продукт отличается от природного hGH. Данные процедуры включали в себя следующие анализы, но не ограничивались ими: ДСН/ПААГ, Вестерн-блоттинг, анализ биологической активности in vitro (клетки nb2) и in vivo (крысы с гипофизэктомией), пептидное картирование, определение полной аминокислотной последовательности, изоэлектрическое фокусирование (IEF), хроматографию с обращенной фазой и гель-хроматографию HPLC. Результаты, полученные во всех этих анализах, в точности совпадали для рекомбинантного и природного hGH, что доказывает тот факт, что чистый рекомбинантный hGH в точности соответствует природному hGH и, следовательно, подходит для производства биофармацевтического продукта.
Теперь, имея полное описание изобретения, специалисту в данной области должно быть понятно, что подобное может быть проведено в пределах широкой и эквивалентной области условий, рецептур и других параметров без изменения рамок изобретения или любых его воплощений. Все патенты и публикации, процитированные в данном документе, полностью включены в данное описание посредством ссылки во всей полноте.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЛЕКОПИТАЮЩИХ, КОТОРЫЕ ПРОДУЦИРУЮТ ЭКЗОГЕННЫЕ БЕЛКИ В МОЛОКЕ, И ТРАНСГЕННЫЕ МЛЕКОПИТАЮЩИЕ, ПОЛУЧЕННЫЕ ТАКИМ СПОСОБОМ | 2004 |
|
RU2390562C2 |
| ТРАНСГЕН ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА В МОЛОКЕ ТРАНСГЕННЫХ КОРОВ, СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОЙ КОРОВЫ (ВАРИАНТЫ), МОЛОКО ОТ ТРАНСГЕННОЙ КОРОВЫ, ПИЩЕВОЙ СОСТАВ | 1990 |
|
RU2095414C1 |
| ПРОИЗВОДСТВО БЕЛКОВ | 2005 |
|
RU2429243C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ В МОЛОЧНОЙ ЖЕЛЕЗЕ НЕТРАНСГЕННЫХ МЛЕКОПИТАЮЩИХ | 2003 |
|
RU2345088C2 |
| ПОЛУЧЕНИЕ БИОЛОГИЧЕСКИ АКТИВНЫХ БЕЛКОВ | 2007 |
|
RU2441911C2 |
| ТРАНСГЕННЫЕ КОПЫТНЫЕ ЖИВОТНЫЕ, ИМЕЮЩИЕ ПОНИЖЕННУЮ АКТИВНОСТЬ ПРИОННОГО БЕЛКА, И ИХ ПРИМЕНЕНИЯ | 2003 |
|
RU2384059C2 |
| ВАРИАНТЫ СТИМУЛИРУЕМОЙ СОЛЯМИ ЖЕЛЧИ ЛИПАЗЫ, КОДИРУЮЩИЕ ИХ МОЛЕКУЛЫ ДНК И ТРАНСГЕННЫЕ МЛЕКОПИТАЮЩИЕ, НЕ ПРИНАДЛЕЖАЩИЕ К ЧЕЛОВЕКУ | 1994 |
|
RU2219239C2 |
| ВЫДЕЛЕНИЕ И ОЧИСТКА БЕЛКА | 2005 |
|
RU2420585C2 |
| МОЛЕКУЛА ДНК, ВЕКТОР ЭКСПРЕССИИ | 1993 |
|
RU2128708C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭМБРИОНОВ ЖИВОТНЫХ И СПОСОБ ВЫРАЩИВАНИЯ ЖИВОТНОГО ИЗ ЭМБРИОНОВ | 1998 |
|
RU2216592C2 |
Изобретение относится к области биотехнологии и генной инженерии. Способ включает в себя клонирование генетической конструкции, кодирующей ген hGH, и, соответственно, промотор бета-казеина в экспрессирующий вектор. Далее проводят трансфекцию в соматические клетки женской особи, особенно фибробласты, и переносят ядра в энуклеированные бычьи ооциты, в результате чего образуются трансгенные эмбрионы. Предложены также способы очистки полученного рекомбинантного гормона роста из молока трансгенных животных. Таким образом, трансгенные эмбрионы дают начало трансгенному рогатому скоту, который продуцирует в своем молоке гормон роста человека в огромных количествах и из которого гормон полностью выделялся и анализировался для выполнения всех необходимых условий для производства чистого биофармацевтического продукта. Изобретение может быть использовано в медицине и ветеринарии. 4 н. и 44 з.п. ф-лы, 6 ил., 3 табл.
1. Способ продуцирования рекомбинантного гормона роста, включающий в себя
a) получение отличного от человека трансгенного млекопитающего из рода быков, которое в своем молоке продуцирует гормон роста, причем в сыворотке указанного млекопитающего продуцируется в среднем на месяц по меньшей мере 92 нг/мл гормона роста;
b) получение указанного молока от указанного трансгенного млекопитающего, отличного от человека; и
c) выделение указанного гормона роста из указанного молока.
2. Способ по п.1, где указанное отличное от человека трансгенное млекопитающее получено способом, включающим в себя
a) клонирование гена, который кодирует гормон роста в плазмиду, посредством чего указанный ген оперативно связывается с промотором, который будет направлять экспрессию указанного гена в клетках молочной железы, приводя к экспрессии плазмиды;
b) трансфекцию бычьих соматических клеток указанной экспрессирующей плазмидой так, чтобы указанная плазмида включалась в геном указанных соматических клеток, что приводит к получению трансгенных соматических клеток;
c) энуклеацию зрелого бычьего ооцита, что приводит к получению энуклеированного ооцита;
d) слияние одной из указанных трансгенных соматических клеток с указанным энуклеированным ооцитом, что приводит к получению моноклеточного эмбриона;
e) имплантацию указанного эмбриона в матку реципиентного млекопитающего, что приводит к беременности млекопитающего; и
f) мониторинг беременности вплоть до рождения трансгенного млекопитающего.
3. Способ по п.2, где указанный промотор является промотором бета-казеина, и где указанный гормон роста является гормоном роста человека.
4. Способ по п.3, где экспрессирующей плазмидой является плазмида pRβhGH.
5. Способ по п.4, где указанная экспрессирующая плазмида дополнительно содержит ген резистентности к неомицину.
6. Способ по п.5, где указанной экспрессирующей плазмидой, которая дополнительно содержит ген резистентности к неомицину, является плазмида pRNeo.
7. Способ по п.2, где указанное млекопитающее, геном которого содержит интегрированную плазмиду, относится к роду быков и продуцирует в своем молоке рекомбинантный гормон роста, причем указанная плазмида содержит ген гормона роста человека и промотор бета-казеина, который направляет экспрессию указанного гена в клетках молочной железы указанного млекопитающего.
8. Способ по п.7, где указанной плазмидой является плазмида pRβhGH.
9. Способ по п.8, где указанная плазмида дополнительно содержит ген резистентности к неомицину.
10. Способ по п.9, где указанной плазмидой, которая дополнительно содержит ген резистентности к неомицину, является плазмида pRNeo.
11. Способ по п.2, где указанные соматические клетки являются фибробластами.
12. Способ по п.2, где указанные трансгенные соматические клетки получены путем их выделения из женской особи трансгенного млекопитающего из рода быков для продуцирования в молоке указанного млекопитающего указанного гормона роста.
13. Способ по п.12, где указанные трансгенные соматические клетки являются фибробластами.
14. Способ по п.1, где указанное отличное от человека трансгенное млекопитающее получено способом, включающим в себя
a) суперовуляцию женской особи млекопитающего, отличного от человека, которое является трансгенным по продуцированию в своем молоке указанного представляющего интерес белка;
b) искусственное оплодотворение указанного млекопитающего спермой, полученной от мужской особи нетрансгенного млекопитающего, отличного от человека, с получением эмбрионов;
c) сбор указанных эмбрионов;
d) имплантацию указанных эмбрионов в матку млекопитающего-реципиента; и
e) мониторинг беременности вплоть до рождения трансгенного млекопитающего.
15. Способ по п.1, где указанное трансгенное млекопитающее, отличное от человека, получено способом, включающим в себя
a) суперовуляцию женской особи млекопитающего, отличного от человека, которая является трансгенной по продуцированию в своем молоке указанного представляющего интерес белка;
b) искусственное оплодотворение указанного млекопитающего спермой, полученной от мужской особи млекопитающего, отличного от человека, являющегося трансгенным по продуцированию указанного представляющего интерес белка, с получением эмбрионов;
c) сбор указанных эмбрионов;
d) имплантацию указанных эмбрионов в матку млекопитающего-реципиента; и
e) мониторинг беременности вплоть до рождения трансгенного млекопитающего.
16. Способ по п.1, где указанное трансгенное млекопитающее, отличное от человека, получено способом, включающим в себя
a) суперовуляцию женской особи нетрансгенного млекопитающего, отличного от человека;
b) искусственное оплодотворение указанного млекопитающего спермой, полученной от мужской особи млекопитающего, отличного от человека, являющегося трансгенным по продуцированию указанного представляющего интерес белка, с получением эмбрионов;
c) сбор указанных эмбрионов;
d) имплантацию указанных эмбрионов в матку млекопитающего-реципиента; и
e) мониторинг беременности вплоть до рождения трансгенного млекопитающего.
17. Способ по любому из пп.2, 12, 14, 15 или 16, где указанное млекопитающее представляет собой млекопитающее из рода быков, рода свиней, рода овец, рода коз или рода грызунов.
18. Способ по п.17, где указанное млекопитающее относится к роду быков.
19. Способ по любому из пп.2, 12, 14, 15 или 16, где указанный представляющий интерес белок является гормоном роста млекопитающих.
20. Способ по п.19, где указанный гормон роста млекопитающих является гормоном роста человека, бычьим гормоном роста, свиным гормоном роста, овечьим гормоном роста, козьим гормоном роста или гормоном роста грызуна.
21. Способ по п.20, где указанный гормон роста млекопитающих является гормоном роста человека.
22. Способ по п.21, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 1,0 г hGH/л молока.
23. Способ по п.22, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 2,0 г hGH/л молока.
24. Способ по п.23, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 3,0 г hGH/л молока.
25. Способ по п.24, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 4,0 г hGH/л молока.
26. Способ по п.25, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 5,0 г hGH/л молока.
27. Способ по п.26, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 6,0 г hGH/л молока.
28. Способ по п.19, где продуцирование указанного гормона роста млекопитающих указанным млекопитающим стимулирует указанное млекопитающее продуцировать больше молока, содержащего указанный гормон роста млекопитающего.
29. Способ по любому из пп.2, 12, 14, 15 или 16, где ген, кодирующий указанный гормон роста, связанный с промотором, который направляет экспрессию указанного гена в клетках молочной железы, находится в соматических клетках и клетках эмбрионов указанного млекопитающего.
30. Способ очистки рекомбинантного гормона роста из молока отличного от человека трансгенного млекопитающего из рода быков, причем в сыворотке указанного млекопитающего продуцируется в среднем на месяц по меньшей мере 92 нг/мл гормона роста, включающий в себя
a) осветление молока трансгенного млекопитающего, отличного от человека, что дает осветленное молоко; и
b) хроматографирование указанного осветленного молока, что дает очищенный рекомбинантный гормон роста.
31. Способ по п.30, где указанная хроматография представляет собой ионообменную хроматографию, хроматографию с обращенной фазой, молекулярную заместительную хроматографию или аффинную хроматографию.
32. Способ по п.31, где проводят многократные стадии хроматографирования.
33. Способ по п.31, где указанная ионообменная хроматография представляет собой анионообменную хроматографию.
34. Способ по п.31, где аффинная хроматография представляет собой иммунохроматографию.
35. Способ по п.31, дополнительно включающий в себя стадию концентрирования.
36. Способ очистки рекомбинантного гормона роста из молока отличного от человека трансгенного млекопитающего из рода быков, причем в сыворотке указанного млекопитающего продуцируется в среднем на месяц по меньшей мере 92 нг/мл гормона роста, включающий в себя
a) осветление молока трансгенного млекопитающего, отличного от человека, что дает осветленное молоко;
b) хроматографирование указанного осветленного молока, с получением результирующего материала после анионообменной хроматографии;
c) хроматографирование указанного материала после анионообменной хроматографии, с получением результирующего материала после хроматографии с обращенной фазой;
d) анионообменную хроматографию указанного материала после хроматографии с обращенной фазой, с получением результирующего материала после анионообменной хроматографии;
e) молекулярную вытеснительную хроматографию указанного материала после анионообменной хроматографии, с получением результирующего материала после молекулярной вытеснительной хроматографии;
f) концентрирование указанного материала после молекулярной вытеснительной хроматографии, с получением концентрированного материала; и
g) молекулярную вытеснительную хроматографию указанного концентрированного материала, с получением чистого рекомбинантного гормона роста.
37. Способ очистки рекомбинантного гормона роста из молока отличного от человека трансгенного млекопитающего из рода быков, причем в сыворотке указанного млекопитающего продуцируется в среднем на месяц по меньшей мере 92 нг/мл гормона роста, включающий в себя
a) осветление молока трансгенного млекопитающего, что дает осветленное молоко;
b) хроматографирование указанного осветленного молока, с получением результирующего материала после иммуноаффинной хроматографии;
c) хроматографию с обращенной фазой указанного материала после иммуноаффинной хроматографии, с получением результирующего материала после хроматографии о обращенной фазой;
d) анионообменную хроматографию указанного материала после хроматографии с обращенной фазой, с получением результирующего материала после анионообменной хроматографии;
e) молекулярную вытеснительную хроматографию указанного материала после анионообменной хроматографии, с получением результирующего материала после молекулярной вытеснительной хроматографии;
i) концентрирование указанного материала после молекулярной вытеснительной хроматографии, с получением концентрированного материала; и
g) молекулярную вытеснительную хроматографию указанного концентрированного материала, с получением чистого рекомбинантного гормона роста.
38. Способ по п.36 или 37, где указанный гормон роста представляет собой гормон роста млекопитающего.
39. Способ по п.38, где указанный гормон роста млекопитающего представляет собой гормон роста человека, бычий гормон роста, свиной гормон роста, овечий гормон роста, козий гормон роста или гормон роста грызуна.
40. Способ по п.39, где указанный гормон роста млекопитающего представляет собой гормон роста человека.
41. Способ по п.40, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 1,0 г hGH/л молока.
42. Способ по п.41, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 2,0 г hGH/л молока.
43. Способ по п.42, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 3,0 г hGH/л молока.
44. Способ по п.43, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 4,0 г hGH/л молока.
45. Способ по п.44, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 5,0 г hGH/л молока.
46. Способ по п.45, где указанное млекопитающее продуцирует гормон роста человека приблизительно в количестве, составляющем более чем 6,0 г hGH/л молока.
47. Способ по п.36 или 37, где указанное отличное от человека трансгенное млекопитающее относится к роду быков.
48. Способ по п.36 или 37, где указанное отличное от человека трансгенное млекопитающее представляет собой свинью, овцу, козу или грызуна.
| CERDAN M.G | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| US 6451527, 17.09.2002 | |||
| US 5741957, 21.04.1998 | |||
| US 6395958, 28.05.2002. | |||

