ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к способу очистки рекомбинантных белков, накапливающихся в рекомбинантных подобных белковым тельцам скоплениях (RPBLA). Более конкретно это изобретение относится к выделению рекомбинантных слитых белков в рекомбинантных подобных белковым тельцам скоплениях, которые позволяют отделять их от других органелл клетки-хозяина вследствие различий в плотности, где требуемый рекомбинантный белок можно концентрировать, отделять от других компонентов клетки и легко выделять.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Белковые тельца (PB) представляют собой субклеточные органеллы (или крупные везикулы, окруженные мембраной, с диаметром приблизительно 1-3 микрон), которые специализируются на запасании белка. В природе они образуются в некоторых определенных тканях растений, таких как семена, и служат в качестве основного источника аминокислот для проращивания и роста проростка.
Запасные белки котрансляционно встраиваются в просвет эндоплазматической сети (ER) с помощью сигнального пептида для упаковки либо в ER, либо в вакуоли (Galili et al., 1993 Trends Cell Biol. 3:437-443) и собираются в мультимерные структуры внутри этих субклеточных компартментов, образуя определенные органеллы, называемые происходящими из (ER) белковыми тельцами (PB) или вакуолями запасных белков (PSV) (Okita and Rogers, 1996 Annu. Rev. Plant Physiol Mol. Biol. 47:327-350; Herman and Larkins, 1999 Plant Cell 11:601-613; Sanderfoot and Raikel, 1999 Plant Cell 11:629-642).
Запасные белки двудольных растений представляют собой, главным образом, растворимые белки, такие как белки глобулина или вицилина типа 7S, глобулина или легумина типа 11S, и они депонируются в PSV совместно с другими белками (т.е. ингибиторами протеаз, протеолитическими ферментами, лектинами и т.п.), сахарами и солями.
В противоположность PSV, PB (1-3 микрон) депонируют, главным образом, проламины, которые представляют собой высокогидрофобные запасные белки зерновых (такие как зеины маиса и глиадины пшеницы), и в них отсутствуют другие вспомогательные белки (Herman and Larkins, 1999 Plant Cell 11:601-613).
К настоящему времени в тканях, отличных от семян растений, PB не выявлены, за исключением телец ER. Тельца ER обладают небольшим размером (0,2-0,4 микрометров) и они образуются в листьях Arabidopsis только при повреждении и разжевывании насекомыми, однако они не образуются в нормальных условиях (Matsushima et al., 2003 Plant J. 33:493-502).
Подходы генетической инженерии применяли для исследования образования растительных PB, сборки и направления запасных белков. Было показано, что в случае, когда рекомбинантные белки, главным образом, растительные запасные белки, экспрессируются и упаковываются в Arabidopsis и табаке, ткани растений, которые не содержат PB (такие как вегетативные ткани), производят эти органеллы "de novo" (Bagga et al., 1997 Plant Cell 9:1683-1696 and Bagga et al., 1995 Plant Physiol. 107:13-23, и патенты США No. 5990384, No. 5215912 и No. 5589616; и Geli et al., 1994 Plant Cell 6:1911-1922).
Бета-зеин маиса при экспрессии в трансгенных растениях табака точно направлялся в новые образующиеся происходящие из ER PB в клетках листьев (Bagga et al., 1995 Plant Physiol. 107:13-23). кДНК гамма-зеина маиса и укороченная кДНК гамма-зеина, экспрессируемая в растениях Arabidopsis, также накапливаются в новых происходящих из ER PB в листьях (Geli et al., 1994 Plant Cell 6:1911-1922). Богатые лизином гамма-зеины, экспрессируемые в эндосперме маиса (Torrent et al. 1997 Plant Mol. Biol. 34(1):139-149), накапливаются в PB маиса и колокализуются с эндогенными зеинами. На трансгенных растениях табака, экспрессирующих ген альфа-зеина, было показано, что альфа-зеин не способен образовывать PB. Однако, когда альфа- и гамма-зеин коэкспрессировались, стабильность альфа-зеина повышалась, и оба белка колокализовались в происходящих из ER белковых тельцах (Coleman et al., 1996 Plant Cell 8:2335-2345). Также было описано образование новых PB в трансгенных бобах сои, трансформированных богатым метионином дельта-зеином массой 10 кДа (Bagga et al., 2000 Plant Sci. 150:21-28).
Рекомбинантные запасные белки также накапливаются в подобных PB органеллах в нерастительных системах-хозяевах, таких как ооциты Xenopus, и в дрожжах. Rosenberg et al., 1993 Plant Physiol 102:61-69 описали экспрессию гамма-глиадина пшеницы в дрожжах. Ген экспрессировался правильно, и белок накапливался в происходящих из ER PB. Torrent et al., 1994 Planta 192:512-518 показали, что в ооцитах Xenopus гамма-зеин также запасается в подобных PB органеллах после микроинъекции транскриптов, кодирующих белок, в ооциты. Hurkman et al., 1981 J. Cell Biol. 87:292-299, для альфа-зеинов и Altschuler et al., 1993 Plant Cell 5:443-450, для гамма-глиадинов в ооцитах Xenopus получили сходные результаты.
Одним из фундаментальных достижений в области биотехнологии (генетической инженерии) является возможность генетически воздействовать на организм для получения белка для терапевтического, нутрицевтического или промышленного применения. Получены способы продукции и выделения рекомбинантных белков из бульона для культивирования клеточных культур бактерий, дрожжей, из культур клеток сельскохозяйственных растений и млекопитающих. Описаны различные подходы для экспрессии белка в клетках-хозяевах. Основными целями этих подходов являлись: уровень экспрессии белка, стабильность белка и выделение белка (Menkhaus et al., 2004 Biotechnol. Prog. 20:1001-1014; Evangelista et al., 1998 Biotechnol. Prog. 14:607-614).
Одна стратегия, которая может решить проблему выделения белка, представляет собой секрецию. Однако секреция предполагает иногда низкие уровни экспрессии и нестабильность продукта. Другой стратегией является накопление рекомбинантного белка в наиболее подходящем отделе клетки. Эту стратегию широко использовали посредством направления рекомбинантных белков в ER с помощью конструирования C-концевого удлинения из тетрапептида (HDEL/KDEL) (Conrad and Fiedler, 1998 Plant Mol. Biol. 38:101-109).
Слитые белки, содержащие растительный запасной белок или домены запасного белка, слитые с гетерологичным белком, представляют собой альтернативный подход для направления рекомбинантных белков в ER (WO 2004003207). Одной интересной стратегией слияния является продукция рекомбинантных белков, слитых с олеозинами, составными белками масляных телец растений. Определенные свойства масляных телец дают преимущество в отношении легкого выделения белков с использованием двухфазной системы (van Rooijen and Moloney, 1995 Bio/Technology 13:72-77).
Гетерологичные белки были успешно экспрессированы в растительных клетках (обзоры Horn et al., 2004 Plant Cell Rep. 22:711-720; Twyman et al., 2003, Trends in Biotechnology 21:570-578; Ma et al., 1995, Science 268:716-719; Richter et al., 2000 Nat. Biotechnol. 18:1167-1171) и в некоторых из них, экспрессия рекомбинантного белка была направлена в происходящие из ER PB или PSV (PSV). Yang et al., 2003 Planta 216:597-603 экспрессировали лизоцим человека в семенах риса с использованием специфичных для семян промоторов запасных белков глутелина и глобулина. Результаты иммуноцитохимии показали, что рекомбинантный белок был локализован в ER-PB и что он запасался с эндогенными глобулинами и глутелинами риса. Экспрессию гликопротеина B цитомегаловируса человека (hCMV) в трансгенных растениях табака осуществляли с использованием промотора глутелина риса. Tackaberry et al., 1999 Vaccine 17:3020-3029. Недавно Arcalis et al., 2004 Plant Physiology 136:1-10 экспрессировали сывороточный альбумин человека (HSA) с C-концевым удлинением (KDEL) в семенах риса. Рекомбинантный HSA запасался в PSV с эндогенными запасными белками риса.
Одним препятствием для применения растений в качестве биофабрик является необходимость в проведении большего количества исследований, касающихся последующей переработки. Очистка белка из растений является трудной задачей вследствие сложности растительной системы. Растительные твердые вещества в экстракте являются крупными, плотными и их количество является относительно высоким (9-20 процентов по массе) (см. обзор Menkhaus et al., 2004 Biotechnol. Prog. 20:1001-1014). В настоящее время, способы очистки рекомбинантных белков включают в себя очистку экстрактов, обработку растворителями для удаления липидов и пигментов и очистку белков или пептидов с помощью нескольких колонок для ионообменной хроматографии и гель-фильтрации. Существующие протоколы основаны на применении определенных растворителей или водных растворов как для систем растений-хозяев, так и для рекомбинантного белка. В данной области существует необходимость в эффективных и универсальных способах выделения рекомбинантного белка из трансформированных хозяев. Эта необходимость особенно актуальна в случаях, когда необходимо выделить рекомбинантные белки, продуцируемые в хозяевах-растениях. Разнообразие хозяев и белков и их различные физико-химические характеристики требуют эффективного способа концентрирования и выделения рекомбинантных продуктов. Настоящее изобретение, описанное в настоящем документе, представляет собой один способ упрощения и повышения выделения рекомбинантно экспрессируемых белков из организмов, не относящихся к высшим растениям, таким как грибы и клетки млекопитающих.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к эффективному или универсальному процессу или способу выделения рекомбинантного белка из трансформированных хозяев. Представленная в настоящем описании разработка основана на открытии того факта, что отделение рекомбинантных подобных белковым тельцам скоплений (RPBLA) от белков других органелл клеток-хозяев можно осуществлять с неожиданно хорошим выходом с помощью способа на основе плотности, в частности, с помощью способов центрифугирования со слоем с определенной плотностью или с градиентом плотности.
Более конкретно, настоящее изобретение относится к способу очистки рекомбинантного слитого белка, который экспрессируется в качестве RPBLA клеток-хозяев. В соответствии с рассматриваемым способом получают водный гомогенат трансформированных клеток-хозяев, которые экспрессируют слитый белок в качестве RPBLA. Эти RPBLA обладают заранее определенной плотностью, которая может отличаться среди различных слитых белков, но является известной для конкретного слитого белка, подлежащего выделению. Заранее определенная плотность RPBLA, как правило, является более высокой, чем плотность по существу всех эндогенных белков клетки-хозяина, находящихся в гомогенате, и, как правило, составляет от приблизительно 1,1 до приблизительно 1,35 г/мл. В гомогенате образуют части различной плотности с получением части, которая содержит относительно повышенную концентрацию RPBLA, и части, которая содержит относительно сниженную концентрацию RPBLA. Часть со сниженной концентрацией RPBLA отделяют от части с относительно повышенной концентрацией RPBLA, таким образом, очищая указанный слитый белок. Часть с относительно повышенной концентрацией RPBLA можно собирать или обрабатывать одним или несколькими реагентами или подвергать одной или нескольким обработкам перед выделением RPBLA или слитого белка, находящегося в нем.
В предпочтительном варианте, слитый белок содержит две связанные друг с другом полипептидные последовательности, в которых одна последовательность представляет собой индуцирующую белковые тельца последовательность (PBIS), в то время как другая последовательность представляет собой последовательность представляющего интерес продукта, такого как молекула лекарственного средства и фермент или сходные с ними. Предпочтительные индуцирующие белковые тельца последовательности представляют собой последовательности соединений проламина, таких как гамма-зеин, альфа-зеин или проламин риса.
В настоящем описании клетки-хозяева представляют собой эукариотические клетки, такие как клетки высших растений, грибов и дрожжей, клетки животных, такие как клетки млекопитающих и клетки водорослей. Эти клетки могут быть свежими, которые получают непосредственно из свежей биомассы, или они могут быть высушенными, которые получают из высушенной биомассы; под биомассой понимают массу, получаемую из живых организмов или клеток, таких как культуральная среда или живой организм, например, такой как лист растения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На чертежах, являющихся частью этого описания, на фиг.1 представлено схематическое изображение бинарных векторов, используемых для временной (агроинфильтрация) и стабильной трансформации растений табака, представленны в верхней части фигуры. Два вектора, используемые для трансформации дрожжей, представлены в середине фигуры. Векторы, используемые для временной трансфекции культур клеток млекопитающих, представлены в нижней части. RX3, N-концевой домен гамма-зеина массой 27 кДа, 22aZ, альфа-зеин массой 22 кДа, 22aZt, N-концевой домен альфа-зеина массой 22 кДа, rP13, проламин риса массой 13 кДа и CS участок расщепления.
Фиг.2 представлена в четырех частях, фиг.2,A-D. На фиг.2,A представлено запасание слитых белков RX3-T20 и RX3-EGF в листьях трансгенных растений табака. Растворимые белки экстрагировали из листьев табака дикого типа (wt) и трансгенного табака (дорожки 2 и 4), анализировали в SDS-полиакриламидных гелях и переносили на нитроцеллюлозные мембраны, с последующим иммуноблот-анализом с использованием антисыворотки против гамма-зеина. Молекулярные массы указаны слева. Полученные из RX3 мономер (M) и димеры (D) слитых белков указаны стрелками.
На фиг.2,B представлен иммуноблот-анализ слитых белков RX3-T20 и RX3-EGF во фракциях градиента плотности. Очищенные гомогенаты влажных (свежих) листьев трансформированного табака помещали в ступенчатый градиент сахарозы (42%-49%-56%-65% масс./масс.). Накопление слитых белков RX3-EGF и RX3-T20 в гомогенате, супернатанте, интерфазе и фракции осадка анализировали с помощью иммуноблота с использованием антител против гамма-зеина. Каждая дорожка соответствует эквивалентным объемам всех фракций. H, гомогенат; S, супернатант; F42, интерфаза 42-49% масс./масс.; F49, интерфаза 49-56% масс./масс.; F56, интерфаза 56-65% масс./масс.; F65, осадок при 65% сахарозе. Молекулярные массы указаны слева. Полученные из RX3 мономер (M) и димеры (D) слитых белков указаны стрелками.
На фиг.2,C представлен анализ SDS-PAGE и анализ окрашиванием серебром слитого белка RX3-EGF, экспрессируемого pl9RX3EGF в трансформированных листьях табака, во фракциях градиента плотности. Очищенные гомогенаты влажных (свежих) листьев табака в буфере PBP помещали в ступенчатый градиент сахарозы (42%-49%-56%-65% масс./масс.). Накопление слитого белка RX3-EGF в фракциях гомогената, супернатанта, интерфазы и осадка анализировали посредством 15% SDS-PAGE и проявляли окрашиванием серебром. Каждая дорожка соответствует эквивалентным объемам всех фракций. Стрелкой указан белок RX3-EGF. H, гомогенат; S, супернатант; F42, интерфаза с 42-49% масс./масс. сахарозой; F49, интерфаза 49-56% масс./масс.; F56, интерфаза 56-65% масс./масс.; F65, осадок при 65% сахарозе. Молекулярные массы указаны слева.
На фиг.2,D представлены результаты SDS-PAGE и иммуноблотинга в отношении накопления RX3-T20 и RX3-EGF в влажных и сухих листьях табака. Листья табака высушивали при 37°C в течение одной недели и хранили в течение пяти месяцев в контейнере, не содержащем влаги. Растворимые белки, экстрагированные из эквивалентных количеств влажных (W) и сухих (D) трансформированных листьев табака, анализировали с помощью SDS-полиакриламидных гелей и иммуноблота с использованием антисыворотки против гамма-зеина (дорожки 1 и 2). Накопление RX3-EGF и RX3-T20 в плотных структурах в высушенных образцах анализировали фракционированием гомогенатов сухих листьев в градиентах сахарозы (20%-30%-42%-56% масс./масс.). Накопление слитых белков RX3-EGF и RX3-T20 в фракциях супернатанта, интерфазы и осадка анализировали с помощью иммуноблота с использованием антител против гамма-зеина. Наносили эквивалентные количества каждой фракции. S, супернатант; F20, интерфаза с 20%-30% масс./масс. сахарозой; F30, интерфаза с 30%-42% масс./масс. сахарозой; F42, интерфаза с 42%-56% масс./масс. сахарозой; F56, осадок при 56% сахарозе.
Фиг.3 представлена в двух частях, фиг.3,A и B. На фиг.3,A представлено накопление RX3-EGF и RX3-T20 в агроинфильтрированных проростках табака. Все растворимые белки анализировали с помощью SDS-PAGE и иммуноблота с использованием антисыворотки против гамма-зеина. Wt = контрольные проростки дикого типа; молекулярные массы указаны слева. Полученные из RX3 мономер (M), димеры (D) и тримеры (T) слитых белков указаны стрелками. На фиг.3,B представлено субклеточное фракционирование агроинфильтрированных проростков табака. Очищенные гомогенаты проростков помещали в ступенчатые градиенты сахарозы (20%-30%-42%-56% масс./масс.). Накопление слитых белков RX3-T20 и RX3-hGH во фракциях супернатанта, интерфазы и осадка анализировали с помощью иммуноблота с использованием антител против гамма-зеина. В каждую дорожку наносили эквивалентные количества каждой фракции. S, супернатант; F20, интерфаза с 20%-30% масс./масс. сахарозой; F30, интерфаза с 30%-42% масс./масс. сахарозой; F42, интерфаза с 42%-56% масс./масс. сахарозой; F56, осадок при 56% сахарозе. Молекулярные массы указаны слева. Полученные из RX3 мономер (M) и димеры (D) слитых белков указаны стрелками.
Фиг.4 представлена в трех частях в виде фиг.4,A, B и C. На фиг.4,A представлена концентрация белков RX3-T20 и RX3-EGF после центрифугирования через слой с сахарозой. Очищенные гомогенаты трансгенных листьев табака помещали в слой с сахарозой (42% масс./масс.). После центрифугирования накопление слитых белков RX3-EGF и RX3-T20 в супернатанте и осадке анализировали с помощью иммуноблота с использованием антител против гамма-зеина. Каждая дорожка соответствует эквивалентным количествам фракций. H, гомогенат; S, супернатант; P, слой осадка. Молекулярные массы указаны слева. Полученные из RX3 мономер (M) и димеры (D) слитых белков указаны стрелками.
На фиг.4,B представлена очистка белка RX3-EGF после центрифугирования через слой сахарозы. Очищенные гомогенаты трансгенных листьев табака помещали в слой сахарозы (42% масс./масс.). После центрифугирования образцы белков фракций гомогената, супернатанта и осадка анализировали с помощью 15% SDS-PAGE и окрашивания серебром. Каждая дорожка соответствует эквивалентным количествам фракций. Мономер (M) и димеры (D) RX3-EGF указаны стрелками. H, гомогенат; S, супернатант; P, слой осадка. Молекулярные массы указаны слева.
На фиг.4,C представлена концентрация белка RX3-EGF и очистка после центрифугирования при низкой скорости (LSC). Очищенные гомогенаты экспрессирующих RX3-EGF листьев табака центрифугировали при 1000 × g в течение 10 минут, и осадок (P1, дорожка 2) и супернатант (S, дорожка 1) анализировали с помощью гель-электрофореза и иммуноблота с использованием антител против гамма-зеина. Осадок P1 после центрифугирования на низкой скорости (LSC) промывали в буферной содержащей 5% TritonX-100 среде и, после второго центрифугирования, эквивалентные количества осадка P1 после LSC (дорожка 7), супернатант после промывания (W, дорожка 8) и конечный осадок P2 (дорожка 9) анализировали с помощью 15% SDS-PAGE и окрашивания серебром. Эти образцы сравнивали с эквивалентными образцами (дорожки 3-5) из осадка P1 (дорожка 3), полученного после одного центрифугирования в слое сахарозы и подвергнутого такому же процессу промывания, что и осадок LSC. Мономер (M) и димеры (D) RX3-EGF указаны стрелками.
Фиг.5 представлена в двух частях в виде фиг.5,A и B. На фиг.5,A представлено субклеточное распределение рекомбинантных слитых белков RX3-Ct, RX3-EGF и RX3-hGH, накопленных в трансфицированных клетках млекопитающих. Гомогенаты трансфицированных клеток помещали в ступенчатые градиенты сахарозы (20%-30%-42%-56% масс./масс.). После центрифугирования накопление слитых белков RX3-Ct, RX3-EGF и RX3-hGH во фракциях супернатанта, интерфазы и осадка анализировали с помощью иммуноблота с использованием антител против гамма-зеина. Клетки, трансфицированные плазмидой pECFP-N1 (Clontech), экспрессирующей ECGP, вариант GFP с голубой флуоресценцией, использовали в качестве контроля, и ECGP выявляли иммунологическим анализом с использованием антисыворотки против GFP. H, гомогенат; S, супернатант; F20, интерфаза с 20%-30% масс./масс. сахарозой; F30, интерфаза с 30%-42% масс./масс. сахарозой; F42, интерфаза с 42%-56% масс./масс. сахарозой; F56, осадок с 56% сахарозой.
На фиг.5,B представлена концентрация белка RX3-EGF, экспрессируемого CHO, после центрифугирования при низкой скорости. Гомогенаты из экспрессирующих RX3-EGF клеток CHO центрифугировали при 2500 × g в течение 10 минут, и осадок (P, дорожка 2) и супернатант (S, дорожка 1) анализировали с помощью гель-электрофореза и иммуноблота с использованием антител против гамма-зеина. Молекулярные массы указаны слева.
На фиг.6 представлено субклеточное распределение рекомбинантных слитых белков RX3-EGF и RX3-hGH, запасаемых в трансформированных клетках дрожжей. Лизированные сферопласты из трансформированных дрожжей помещали в ступенчатые градиенты сахарозы (20%-30%-42%-56% масс./масс.). После центрифугирования накопление слитых белков RX3-EGF и RX3-hGH во фракциях супернатанта, интерфазы и осадка анализировали с помощью иммуноблота с использованием антител против гамма-зеина. H, гомогенат лизированных сферопластов; S, супернатант; F20, интерфаза с 20%-30% масс./масс. сахарозой; F30, интерфаза с 30%-42% масс./масс. сахарозой; F42, интерфаза с 42%-56% масс./масс. сахарозой; F56, осадок в 56% сахарозе. Молекулярные массы указаны слева.
На фиг.7,A представлено субклеточное распределение рекомбинантных слитых белков, запасаемых в агроинфильтрированных растениях табака. Очищенные гомогенаты проростков помещали в ступенчатые градиенты сахарозы (20%-30%-42%-56% масс./масс.). Накопление слитых белков rP13-Ct, rP13-EGF и rP13-hGH во фракциях градиента супернатанта, интерфазы и осадка анализировали с помощью иммуноблота с использованием антител против кальцитонина, EGF и hGH. Эквивалентное исследование проводили с использованием двух вариантов гена альфа-зеина. Проводили слияние кальцитонина (Ct) и EGF с N-концевым доменом альфа-зеина (22aZt) и проводили слияние hGH с полным геном альфа-зеина (22aZ). В каждую дорожку помещали эквивалентные количества каждой фракции. S, супернатант; F20, интерфаза с 20%-30% масс./масс. сахарозой; F30, интерфаза с 30%-42% масс./масс. сахарозой; F42, интерфаза с 42%-56% масс./масс. сахарозой; F56, осадок в 56% сахарозе.
На фиг.7,B представлены результаты применения очищенных гомогенатов листьев из трансгенных линий табака, помещенных в ступенчатые градиенты сахарозы (10%-42%-56%-62% масс./масс.). Посредством иммуноблота с использованием антител против EGF было показано распределение rP13-EGF и 22aZt-EGF в эквивалентных количествах в каждой фракции. S, супернатант; F10, интерфаза с 10%-42% масс./масс. сахарозой; F42, интерфаза с 42%-56% масс./масс. сахарозой; F56, интерфаза с 56%-62% масс./масс. сахарозой; F62, осадок в 62% сахарозе.
На фиг.8 представлено выделение слитого белка RX3-T20 и RX3-EGF из RPBLA. Фракции RPBLA, полученные из слоя с определенной плотностью или ступенчатого градиента, ресуспендировали в присутствии восстанавливающих веществ. Растворенные (S) и нерастворенные белки (P) анализировали с помощью иммуноблота с использованием антисыворотки против гамма-зеина. Молекулярные массы указаны слева. Полученные из RX3 мономер (M), димеры (D) и тримеры (T) слитых белков указаны стрелками.
Настоящее изобретение обладает рядом преимуществ и положительных эффектов.
Одним преимуществом является то, что его применение обеспечивает возможность относительно простой быстрой очистки экспрессируемых белков на основе отличия в плотности экспрессируемого продукта от остальных растворимых веществ клетки.
Таким образом, преимущество этого изобретения состоит в том, что оно обеспечивает способ удаления эндогенных соединений (или нерекомбинантных продуктов) из организма хозяина и культур клеток.
Другим преимуществом этого изобретения является то, что оно обеспечивает надежный и воспроизводимый способ очистки рекомбинантных пептидов или белков из свежей или высушенной биомассы (организм-хозяин).
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Настоящее изобретение, главным образом, относится к последовательной обработке для выделения и очистки представляющих интерес рекомбинантных белков и пептидов из трансформированных организмов или культур клеток. Более конкретно, настоящее изобретение относится к способу очистки рекомбинантного слитого белка, который экспрессируется и запасается в клетках-хозяевах в качестве рекомбинантных подобных белковым тельцам скоплениях (RPBLA). RPBLA представляют собой скопления рекомбинантного слитого белка, индуцируемые доменами запасных белков, которые образуют внутри клеток отложения с высокой плотностью. Эти плотные отложения могут накапливаться в цитозоле, органеллах системы эндомембран, митохондриях, пластидах, или могут секретироваться. В соответствии с рассматриваемым способом, получают водный гомогенат трансформированных клеток-хозяев, которые экспрессируют слитый белок в качестве RPBLA. Для применения гомогенат предпочтительно является очищенным. В гомогенате образуются части с различной плотностью с получением части, которая содержит относительно повышенную концентрацию RPBLA, и части, которая содержит относительно сниженную концентрацию RPBLA. Часть со сниженной концентрацией RPBLA отделяют от части с относительно повышенной концентрацией RPBLA, таким образом, очищая слитый белок. Часть с относительно повышенной концентрацией RPBLA можно, таким образом, собирать или обрабатывать одним или несколькими реагентами или подвергать одному или нескольким процессам перед выделением RPBLA или слитого белка, находящегося в них.
В предпочтительном варианте, слитый белок содержит две связанные друг с другом полипептидные последовательности, в которых одна последовательность представляет собой индуцирующую белковые тельца последовательность (PBIS), в то время как другая последовательность представляет собой последовательность представляющего интерес продукта, такого как молекула лекарственного средства, и фермент или сходные с ними. Предпочтительные PBIS представляют собой последовательности соединений проламина, таких как гамма-зеин, альфа-зеин или проламин риса.
Один аспект настоящего способа включает получение водного гомогената или другого пригодного экстракта (в собирательном значении обозначаемого в настоящем описании как гомогенат) из организма-хозяина или культуры клеток, которая экспрессирует и накапливает требуемый слитый белок в виде рекомбинантных подобных белковым тельцам скоплений (RPBLA). Гомогенат, как правило, предварительно очищают (очищают) перед применением для удаления клеточного дербиса, например, посредством фильтрации. Гомогенат, содержащий подобные белковым тельцам структуры, содержащие слитый белок, (RPBLA), липиды, растворимые белки, органеллы клеток, сахара, пигменты и алкалоиды непосредственно помещают в ступенчатый градиент плотности, и гомогенат разделяют на основе плотности его составляющих, например, посредством центрифугирования. В процессе центрифугирования в гомогенате образуются части с различной плотностью с образованием части, которая содержит относительно повышенную концентрацию RPBLA, и части, которая содержит относительно сниженную концентрацию RPBLA. Требуемые содержащие слитый белок RPBLA можно собирать в интерфазе с определенной плотностью. Этот способ дает возможность выделения более приблизительно 90 процентов экспрессируемого рекомбинантного слитого белка с более чем приблизительно 80 процентной чистотой.
Другой аспект этого изобретения относится к способу выделения RPBLA из предпочтительно очищенного гомогената с помощью слоя с однородной плотностью. Здесь, предпочтительно очищенный гомогенат помещают в слой с определенной плотностью так, чтобы эндогенные примеси не проходили через слой, и разделяют центрифугированием так, чтобы плотные RPBLA проходили через слой и их можно было собирать. В гомогенате образуются описанные ранее части с различной плотностью с образованием части, которая содержит относительно повышенную концентрацию RPBLA (часть под слоем), и части, которая содержит относительно сниженную концентрацию RPBLA (часть над слоем). Таким образом, плотность рекомбинантных подобных белковым тельцам скоплений выше, чем плотность слоя.
В другом варианте осуществления предпочтительно очищенный гомогенат разделяют непосредственно центрифугированием и в отсутствие сахарозы, или другого дополнительного обеспечивающего плотность раствора. Снова центрифугирование приводит к формированию частей различной плотности, образующихся в гомогенате, с образованием части, которая содержит относительно повышенную концентрацию RPBLA (осадок), и части, которая содержит относительно сниженную концентрацию RPBLA (супернатант). Затем для осуществления очистки RPBLA и, таким образом, слитого белка RPBLA можно разделять.
Настоящее изобретение относится к способу выделения рекомбинантных пептидов или белка, экспрессируемых в RPBLA, органеллах, образующихся в трансформированных клетках-хозяевах. В настоящем описании клетки-хозяева представляют собой эукариотические клетки, такие как клетки высших растений, дрожжей и грибов, клетки животных, таких как культивируемые клетки млекопитающих, клетки из трансгенных животных, яйцеклетки животных и сходные с ними, и клетки водорослей. Эти клетки могут быть свежими, которые получают непосредственно из культуральной среды или живого организма, такого как лист растений или животное, или могут быть высушенными.
Рекомбинантные подобные белковым тельцам скопления обладают заранее определенной плотностью, которая может отличаться для различных слитых белков, но которая известна для конкретного слитого белка, подлежащего выделению. Эта заранее определенная плотность RPBLA, как правило, выше, чем плотность по существу всех эндогенных белков клетки-хозяина, находящихся в гомогенате, и, как правило, составляет от приблизительно 1,1 до приблизительно 1,35 г/мл. Высокая плотность новых RPBLA является следствием универсальной способности рекомбинантных слитых белков собираться в мультимеры и запасаться.
Рассматриваемые RPBLA экспрессируются у эукариот и, как правило, характеризуются их плотностями, как указано выше. При экспрессии в клетках высших растений и животных RPBLA, как правило, обладают сферической формой, имеют диаметры приблизительно 1 микрон (мк) и окружены мембраной.
Слитые белки разделяют по их плотностям, которые обычно бывают выше, чем плотность любого другого белка, находящегося в трансфицированной клетке. Это разделение на основе плотности, как правило, проводят с использованием центрифуги, как широко распространено в биохимических лабораториях по всему миру. Иллюстративная коммерчески доступная центрифуга представляет собой Beckman Coulter AvantiTM, модель J-25, которую используют здесь для пропускания через один слой и прямого центрифугирования. Ультрацентрифугу Beckman Coulter OptimaTM XL-100K (ротор SW41Ti) использовали для исследований с градиентом. Центрифугирование часто проводят в присутствии дополнительных обеспечивающих неравномерную плотность растворенных веществ, таких как соль, такая как хлорид цезия, или сахар, такой как сахароза. Объединение гомогената и обеспечивающих неравномерную плотность растворенных веществ приводит к образованию смеси гомогенат-раствор.
В конкретном варианте осуществления рекомбинантные слитые белки содержат, или предпочтительно получены из них, индуцирующие белковые тельца последовательности (PBIS), связанные пептидной связью с представляющими интерес продуктами (мишенями) (например, пептиды или белки). PBIS представляют собой белок или аминокислотные последовательности, которые обеспечивает проникновение и/или накопление белка в RPBLA. Иллюстративные неограничивающие примеры PBIS включают запасные белки или модифицированные запасные белки, такие как, например, проламины, или модифицированные проламины, или домены проламина. Проламины рассмотрены в Shewry et al., 2002 J. Exp. Bot. 53(370):947-958.
Гамма-зеин, запасной белок маиса, последовательности ДНК и аминокислотных остатков которого указаны ниже, представляет собой один из четырех проламинов маиса и составляет 10-15 процентов от общего количества белка в эндосперме маиса. Как и другие проламины злаков, альфа- и гамма-зеины биологически синтезируются в мембраносвязанных полисомах с цитоплазматической стороны шероховатого ER, собираются в просвете, а затем депонируются в происходящих из ER PB (Herman et al., 1999 Plant Cell 11:601-613; Ludevid et al., 1984 Plant Mol. Biol. 3:277-234; Torrent et al., 1986 Plant Mol. Biol. 7:93-403).
Гамма-зеин состоит из четырех типичных доменов i) сигнального пептида из 19 аминокислот, ii) домена с повторами, содержащего восемь единиц гексапептида PPPVHL (SEQ ID NO: 1) (53 aa), iii) домена ProX, в котором остатки пролина чередуются с другими аминокислотами (29 aa) и iv) гидрофобного богатого цистеином C-концевого домена (111 aa).
Способность гамма-зеина собираться в происходящие из ER белковые тельца (PB) не ограничивается семенами. Действительно, когда ген гамма-зеина был конститутивно экспрессирован в трансгенных растениях Arabidopsis, запасной белок накапливался в происходящих из ER рекомбинантных PB в клетках мезофилла листьев (Geli et al., 1994 Plant Cell 6:1911-1922). При поиске сигнала, ответственного за накопление гамма-зеина в происходящих из ER PB (проламины не обладают сигналом KDEL), было показано, что богатый пролином N-концевой домен, включающий домен с тандемным повтором, был необходим для поддержания ER и что C-концевой домен был вовлечен в образование PB. Однако механизмы, посредством которых эти домены обеспечивают сборку PB, остаются неизвестными. Поскольку белковые тельца называют так соответственно только в семенах, аналогичные структуры, продуцируемые в других органах растений и в невысших растениях, называются, главным образом, рекомбинантными подобными белковым тельцам скоплениями (RPBLA).
Другие иллюстративные пригодные последовательности, подобные проламину, представлены ниже в таблице, совместно с их номером доступа в GenBank.
Кроме того, пригодные последовательности получают, проводя поиск посредством BLAST во всех неизбыточных базах данных CDS в GenBank translations+PDB+SwissProt+PIR+PRF (за исключением образцов из окружающей среды), как описано в Altschul et al., 1997 Nucleic acids Res. 25:3389-3402, с использованием запроса, такого как SEQ ID NO: 2 (белковая последовательность RX3), SEQ ID NO: 3 (белковая последовательность альфа-зеина), SEQ ID NO: 4 (белковая последовательность проламина риса).
Иллюстративный модифицированный проламин включает (a) последовательность сигнального пептида, (b) последовательность одной или нескольких копий гексапептида PPPVHL (SEQ ID NO: 1) домена с повторами белка гамма-зеина, полный домен, содержащий восемь элементов гексапептида; и (c) последовательность всего или фрагмента домена ProX гамма-зеина. Иллюстративные конкретные модифицированные проламины включают полипептиды, определенные ниже как R3, RX3 и P4, последовательности ДНК и аминокислотных остатков которых также представлены ниже.
Особенно предпочтительные проламины включают гамма-зеин и составляющие его компоненты, как описано в опубликованной заявке WO2004003207, белок rP13 риса и N-концевой фрагмент альфа-зеина маиса массой 22 кДа. Последовательности ДНК и аминокислотных остатков гамма-зеина (27 кДа), белков риса и альфа-зеина представлены в SEQ ID NO: 5 (последовательность ДНК) и SEQ ID NO: 6 (белковая последовательность); SEQ ID NO: 7 (последовательность ДНК RX3) и SEQ ID NO: 8 (белковая последовательность); SEQ ID NO: 9 (последовательность ДНК R3) и SEQ ID NO: 10 (белковая последовательность); SEQ ID NO: 11 (последовательность ДНК P4) и SEQ ID NO 12 (белковая последовательность); SEQ ID NO: 13 (последовательность ДНК X10) и SEQ ID NO: 14 (белковая последовательность).
rP13 - (белковая последовательность SEQ ID NO: 15 и последовательность ДНК SEQ ID NO: 16) проламин риса массой 13 кДа, гомологичный клону - (GenBank AB016504), Sha et al., 1996 Biosci. Biotechnol. Biochem. 60(2):335-337; Wen et al., 1993 Plant Physiol. 101(3):1115-1116; Kawagoe et al., 2005 Plant Cell 17(4):1141-1153; Mullins et al., 2004 J. Agric. Food Chem. 52 (8):2242-2246; Mitsukawa et al., 1999 Biosci. Biotechnol. Biochem. 63(11):1851-1858.
22aZt - (белковая последовательность SEQ ID NO: 17 и последовательность ДНК SEQ ID NO: 18) N-концевой фрагмент альфа-зеина маиса массой 22 кДа - (GenBank V01475), Kim et al., 2002 Plant Cell 14(3):655-672; Woo et al., 2001 Plant Cell 13(10):2297-2317; Matsushima et al., 1997 Biochim. Biophys. Acta 1339(1):14-22; Thompson et al., 1992 Plant Mol. Biol. 18(4):827-833.
Примеры представляющих интерес белков включают любой белок, обладающий терапевтическим, нутрицевтическим, биорегулирующим и промышленным применением, например, такой как моноклональные антитела (mAb, такие как IgG, IgM, IgA, и т.д.) и их фрагменты, антигены для вакцин (вирус иммунодефицита человека, HIV; антигены предповерхности, поверхности и сердцевины гепатита B, коронавирус гастроэнтерита, и т.д.), гормоны (кальцитонин, гормон роста, и т.д.), ингибиторы протеаз, антибиотики, коллаген, лактоферрин человека, цитокины, промышленные ферменты (гидролазы, гликозидазы, оксидоредуктазы и т.п.). Представлены иллюстративные последовательности ДНК и аминокислотных остатков представляющих интерес иллюстративных белков (белковая последовательность SEQ ID NO: 19 и последовательность ДНК SEQ ID NO: 20), кальцитонин лосося Genbank BAC57417.
hEGF - (белковая последовательность SEQ ID NO: 21 и последовательность ДНК SEQ ID NO: 22) конструкция, в основе которой лежит GenBank AAF85790 без сигнального пептида.
hGH - конструкция, в основе которой лежит P01241 без сигнального пептида (белковая последовательность SEQ ID NO: 23 и последовательность ДНК с использованием предпочтительных для растений кодонов SEQ ID NO: 24 и с использованием природных кодонов SEQ ID NO: 25).
Кроме того, в другом варианте осуществления рекомбинантный слитый белок в дополнение к последовательностям PBIS и представляющего интерес продукта содержит спейсерную аминокислотную последовательность. Спейсерная аминокислотная последовательность может представлять собой аминокислотную последовательность, расщепляемую ферментативными или химическими способами, или не расщепляемую. В конкретном варианте осуществления спейсерная аминокислотная последовательность помещена между указанным PBIS и представляющим интерес продуктом. Иллюстративная аминокислотная последовательность поддается расщеплению протеазой, такой как энтерокиназа, эндопротеаза Arg-C, эндопротеаза Glu-C, эндопротеаза Lys-C, фактор Xa и сходные с ними. Альтернативно, кодируется аминокислотная последовательность, которая поддается специфичному расщеплению химическим реагентом, например, таким как бромид цианогена, который расщепляет по остаткам метионина.
В дополнительном варианте осуществления последовательность нуклеиновой кислоты, используемой для трансформации, представляет собой последовательность, как описано в соответствии с совместно поданной патентной заявкой WO 2004003207, которая включает поддающуюся расщеплению последовательность аминокислотных остатков между PBIS и представляющим интерес полипептидом. Кроме того, в другом варианте осуществления последовательность нуклеиновой кислоты представляет собой последовательность, как описано в соответствии с патентной заявкой WO 2004003207, однако последовательность нуклеиновой кислоты, кодирующая поддающуюся расщеплению аминокислотную последовательность, отсутствует.
В предпочтительном варианте осуществления слитые белки получают в соответствии со способом, который включает трансформацию системы клеток-хозяев, такой как животное, культура клеток животных, растение, культура клеток растений, грибы или водоросли, последовательностью нуклеиновой кислоты, содержащей (i) первую нуклеиновую кислоту, кодирующую PBIS, которая функционально связана в рамке считывания со (ii) второй последовательностью нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую представляющий интерес продукт; а именно последовательность нуклеиновой кислоты, которая кодирует PBIS, химически связана с последовательностью, которая кодирует представляющий интерес полипептид, так чтобы оба полипептида экспрессировались из соответствующих им рамок считывания. При экспрессии полученный слитый белок запасается в трансформированной системе-хозяине в виде рекомбинантных подобных белковым тельцам скоплений с высокой плотностью. В одном варианте осуществления 3'-конец первой последовательности нуклеиновой кислоты (i) соединен (связан) с 5'-концом второй последовательности нуклеиновой кислоты (ii). В другом варианте осуществления 5'-конец первой последовательности нуклеиновой кислоты (i) соединен (связан) с 3'-концом второй последовательности нуклеиновой кислоты (ii). В другом варианте осуществления PBIS содержит запасной белок или модифицированный запасной белок, его фрагмент или модифицированный фрагмент.
В другом конкретном варианте осуществления слитый белок получают в соответствии со способом, который включает трансформацию системы клеток-хозяев, такой как животное, культура клеток животных, растение, культура клеток растений, грибы или водоросли, с последовательностью нуклеиновой кислоты, содержащей, в дополнение к упомянутым ранее последовательностям нуклеиновых кислот (i) и (ii), последовательность нуклеиновой кислоты (iii), находящуюся в рамке считывания, которая кодирует спейсерную аминокислотную последовательность. Спейсерная аминокислотная последовательность может представлять собой аминокислотную последовательность, поддающуюся расщеплению ферментативными или химическими способами, или не поддающуюся расщеплению, как указано выше. В одном конкретном варианте осуществления последовательность нуклеиновой кислоты (iii) помещают между указанными последовательностями нуклеиновых кислот (i) и (ii), например, 3'-конец третьей последовательности нуклеиновой кислоты (iii) связан с 5'-концом второй последовательности нуклеиновой кислоты (ii). В другом варианте осуществления, 5'-конец третьей последовательности нуклеиновой кислоты (iii) связывают с 3'-концом второй последовательности нуклеиновой кислоты (ii).
Как используют в настоящем описании, термин "растительная клетка-хозяин" включает растения, включая как однодольные, так и двудольные, и, особенно, злаковые (например, маис, рис, овес и т.п.), бобовые (например, сою и т.п.), крестоцветные растения (например, Arabidopsis thaliana, кользу и т.п.) и пасленовые растения (например, картофель, томат, табак и т.п.).
Растительная система-хозяин также включает клетки растений. Клетки растений включают суспензионные культуры, зародыши, области меристемы, ткани каллюса, листья, корни, побеги, гаметофиты, спорофиты, пыльцу, семена и микроспоры. Растительная система клеток-хозяев может находиться на различных стадиях созревания и может быть выращена в жидкой или твердой культуре, или в почве, или в пригодной среде в горшках, теплицах или на полях. Экспрессия в растительных системах клеток-хозяев может быть временной или постоянной. Растительная система клеток-хозяев также относится к любому клону такого растения, семени, собственному или гибридному потомку, побегу, полученному либо половым, либо бесполым способом, и потомству любого из них, такому как побеги или семена.
Трансформацию клеток растений с использованием Agrobacterium tumefaciens, как правило, лучше всего проводить на двудольных растениях. Однодольные растения, как правило, легче трансформировать посредством так называемого генного переноса протопластов. Прямой генный перенос, как правило, проводят посредством электропорации, посредством опосредуемого полиэтиленгликолем переноса или бомбардировки клеток микроснарядами, несущими требуемую ДНК. Эти способы трансфекции хорошо известны в данной области и не требуют дополнительного обсуждения в рамках настоящего описания. Также показано, что по меньшей мере рис и маис можно трансформировать с помощью Agrobacterium. Способы регенерации целого растения из трансфицированных клеток и протопластов также хорошо известны, такими способами являются способы получения требуемого белка из тканей растений. См. также, патенты США No. 5618988 и No. 5679880 и ссылки в них.
Рассматриваемый способ также может включать выделение и растворение рекомбинантного слитого белка. Таким образом, например, RPBLA, выделенные из ступенчатого градиента плотности или слоя с однородной плотностью, суспендируют в буферном растворе, содержащем восстанавливающее вещество, и центрифугируют. Осадок удаляют, и рекомбинантный белок выделяют из супернатанта для дальнейшей очистки, если требуется, способами, такими как классические хроматографические способы.
Без дальнейшего уточнения, авторы полагают, что специалист в данной области может с использованием представленного выше описания и подробных примеров, представленных ниже, применять настоящее изобретение в его полном объеме. Следующие предпочтительные конкретные варианты осуществления, таким образом, следует толковать только как иллюстративные и не ограничивающие в любом случае остальной части описания.
Экспериментальный протокол
Пример 1: Конструирование плазмиды для трансформации растений
Кодирующие последовательности T20 и эпидермального фактора роста человека (hEGF) получали синтетически и модифицировали в целях оптимизации использования их кодонов для экспрессии в растениях.
Первую цепь последовательности кДНК, кодирующей 36 аминокислот T20, получали химическим синтезом олигонуклеотидов, и последовательность, соответствующую участку специфичного расщепления для фактора Xa и участку для фермента рестрикции, присоединяли к 5'-концу последовательности. Эту синтетическую конструкцию (SEQ ID NO: 26) очищали с помощью полиакриламидного денатурирующего геля.
Двухцепочечную кДНК получали посредством ПЦР с использованием специфичных для T20 праймеров, содержащих участки рестрикции для дальнейшего клонирования.
Праймеры:
V20 Прямой (SEQ ID NO: 27)
V20 Обратный (SEQ ID NO: 28)
Синтетический ген, кодирующий 53 аминокислоты активного hEGF получали с помощью способа ПЦР с перекрывающимся удлинением праймеров с использованием 4 олигонуклеотидов из приблизительно 60 оснований с 20 перекрывающимися основаниями. кДНК синтетического hEGF включала 5'-линкерную последовательность, соответствующую участку для специфичного расщепления фактором Xa. Олигонуклеотиды очищали с помощью полиакриламидного денатурирующего геля.
EGF1 (SEQ ID NO: 29)
EGF2 (SEQ ID NO: 30)
EGF3 (SEQ ID NO: 31)
EGF4 (SEQ ID NO: 32)
Синтетический ген, кодирующий 191 аминокислоту активного hGH получали с помощью способа ПЦР с перекрывающимся удлинением праймеров с использованием 15 олигонуклеотидов из приблизительно 60 оснований с 20 перекрывающимися основаниями. кДНК синтетического hGH включала 5'-линкерную последовательность, соответствующую специфичному для энтерокиназы участку расщепления. Олигонуклеотиды очищали с помощью полиакриламидного денатурирующего геля.
hGH1 (SEQ ID NO: 33)
hGH2 (SEQ ID NO: 34)
hGH3 (SEQ ID NO: 35)
hGH4 (SEQ ID NO: 36)
hGH5 (SEQ ID NO: 37)
hGH6 (SEQ ID NO: 38)
hGH7 (SEQ ID NO: 39)
hGH8 (SEQ ID NO: 40)
hGH9 (SEQ ID N0: 41)
hGH10 (SEQ ID NO: 42)
hGH11 (SEQ ID NO: 43)
hGH12 (SEQ ID NO: 44)
hGH13 (SEQ ID NO: 45)
hGH14 (SEQ ID NO: 46)
hGH15 (SEQ ID NO: 47)
кДНК синтетических T20 и hEGF очищали из агарозного геля (Amersham) и клонировали в вектор pGEM (Promega). Фрагмент кДНК RX3 (кодирующий N-концевой домен гамма-зеина), содержащий "липкие концы" BspHI и NcoI, встраивали в вектор pCKGFPS65C (Reichel et al., 1996 Proc. Natl. Acad. Sd. USA 93:5888-5893), предварительно расщепленный NcoI (как описано в патентной заявке WO2004003207). Проводили слияние последовательностей, кодирующих T20 и EGF, в рамке считывания с последовательностью RX3. Конструкции RX3-T20 и RX3-EGF получали посредством замены кодирующей GFP последовательности синтетическим геном T20 и EGF.
Полученные в результате конструкции, обозначенные pCRX3T20 и pCRX3EGF, содержали последовательность нуклеиновой кислоты, которая направляет транскрипцию белка, такую как усиленный промотор 35S (SED ID NO: 1), энхансер трансляции, такой как вирус гравировки табака (TEV), кодирующие T20 и EGF последовательности и 3'-последовательности для полиаденилирования из вируса мозаики цветной капусты (CaMV). В результате посредством встраивания экспрессирующих кассет HindIII/HindIII в бинарный вектор pBin19 получали эффективные векторы для трансформации растений pl9RX3T20 и pl9RX3EGF (Bevan, 1984 Nucleic acids Research 12:8711-8721).
Проводили слияние кДНК, кодирующей hGH с N-концевой кодирующей гамма-зеин последовательностью RX3 (патент WO2004003207), и ее встраивали в полученную из pUC18 плазмиду, содержащую усиленный промотор CaMV 35S и 3'-терминатор ocs. Экспрессирующую кассету из полученной из pUC18 плазмиды, названной pUC18RX3hGH, которая содержала последовательность соответствующего слитого белка RX3-hGH, встраивали в бинарный вектор pBin19 (Bevan, 1984 Nucleic acids Research 12:8711-8721).
кДНК, кодирующую альфа-зеин массой 22 кДа (22aZ) и проламин риса массой 13 кДа (rP13), амплифицировали посредством ОТ-ПЦР из библиотеки кДНК из маиса W64A и риса сорта Senia соответственно. Используемые в реакции ПЦР олигонуклеотиды представляли собой:
22aZ-5' (SEQ ID NO: 48)
22aZ-3' (SEQ ID NO: 49)
Rice13Prol-5' (SEQ ID NO: 50)
Rice13Prol-3' (SEQ ID NO: 51)
Соответствующие фрагменты ПЦР клонировали в вектор pCRII вектор (Invitrogen), секвенировали и клонировали в векторы pUC18, содержащие усиленный промотор CaMV 35S, последовательность TEV и 3'-терминатор ocs. pCRII-rP13 расщепляли посредством SalI и NcoI и клонировали в плазмиды pUC18RX3Ct, pUC18RX3hGH и pUC18RX3EGF, расщепленные указанными ферментами с получением соответственно: pUC18rP13Ct, pUC18rP13hGH и pUC18rPl3EGF. pCRII-22aZ расщепляли посредством SalI/NcoI и клонировали в плазмиды pUC18RX3Ct и pUC18RX3EGF, расщепленные указанными ферментами с получением pUC1822aZtCt и pUC1822aZtEGF соответственно. pCRII-22aZ также расщепляли посредством SalI/RcaI и клонировали в плазмиду pUC18RX3hGH, расщепленную посредством SalI/NcoI с получением клона pUC1822aZhGH. Наконец, все эти полученные из pUC18 векторы клонировали в pCambia 5300 посредством HindIII/EcoRI.
Пример 2: Конструирование плазмиды для трансформации клеток животных и дрожжей
Клетки животных
Проводили слияние синтетического гена, соответствующего последовательности зрелого кальцитонина (Ct, WO2004003207) и последовательностям EGF, а также кДНК, кодирующей hGH, с N-концевой кодирующей последовательностью гамма-зеина RX3 (патент WO2004003207) и его встраивали в вектор pUC18. Рестрикционные фрагменты SalI-BamHI из полученных из pUC18 плазмид pUC18RX3Ct, pUC18RX3EGF и pUC18RX3hGH, содержащих последовательности соответствующих слитых белков RX3-Ct, RX3-EGF и RX3-hGH, встраивали в вектор pсДНК3.1-(Invitrogen), расщепленный XhoI-BamHI. В полученных конструкциях, названных p3.1RX3CT, p3.1RX3EGF и p3.1RX3hGH, последовательности слитого белка находились под действием промотора CMV и терминатора pA BGH.
Клетки дрожжей
Рестрикционные фрагменты SalI (тупые концы)-BamHI из полученных из pUC18 плазмид, описанных выше, содержащие последовательности соответствующих слитых белков RX3-EGF и RX3-hGH, встраивали в вектор pYX243 (R&D Systems), расщепленный EcoRI (тупые концы)-BamHI. В полученных конструкциях, обозначенных c117 и c118 соответственно, последовательности слитых белков находились под действием индуцибельного промотора GAL.
Пример 3: Трансформация хозяина
Дрожжи
Штамм leu2 Saccharomyces cerevisiae трансформировали плазмидными конструкциями c117 и c118 способом LiAc (Ito et al. 1983, J. Bacteriol. 153:163-168) и трансформанты отбирали на чашках Leu-. Анализ экспрессии проводили посредством выращивания трансформантов в содержащей галактозу среде.
Растительный материал
Растения табака (Nicotiana tabacum var. Wisconsin) выращивали в камере для выращивания in vitro при 24-26°C с 16-часовым световым периодом. Взрослые растения выращивали в теплице между 18-28°C, влажность поддерживали между 55 и 65% со средним световым периодом 16 часов.
Проростки для способа агроинфильтрации (Vaquero et al., 1999 Proc. Natl. Acad. Sci., USA 96 (20): 11128-11133; Kapila et al., 1997 Plant Sci. 122:101-108) выращивали из семян в течение 4-6 недель в условиях in vitro, описанных выше.
Стабильная трансформация табака
Бинарные векторы переносили в штамм LBA4404 A. tumefaciens. Пластины листьев табака (Nicotiana tobaccum, W38) трансформировали, как описано Draper and Hamil 1988. В: Plant Genetic Transformation and Gene Expression. A Laboratory Manual (Eds. Draper, J., Scott, R., Armitage, P. and Walden, R.), Blackwell Scientific Publications. Регенерированные растения отбирали на среде, содержащей 200 мг/л канамицина, и переносили в теплицу. Трансгенные растения табака, обладающие наивысшими уровнями трансгенного продукта, культивировали с получением поколений T1 и T2.
Уровень рекомбинантного белка определяли с помощью иммуноблота. Общие экстракты белков из листьев табака количественно определяли посредством анализа Брэдфорда, разделяли на 15% SDS-PAGE и переносили на нитроцеллюлозные мембраны с использованием Mini Trans-Blot Electrophoretic Transfer Cell (Bio Rad). Мембраны инкубировали с антисывороткой против гамма-зеина (разведение 1/7000) (Ludevid et al. 1985, Plant Science 41:41-48), а затем инкубировали с конъюгированными с пероксидазой хрена антителами (разведение 1/10000, Amersham Pharmacia). Иммунореактивные полосы определяли по повышенной хемилюминесценции (система для вестерн-блоттинга ECL, Amersham Pharmacia).
Агроинфильтрация табака
Проростки для способа агроинфильтрации выращивали из семян в течение 4-6 недель в камере для выращивания in vitro при 24-26°C со световым периодом 16 часов.
Штамм LB4404 A. tumefaciens, содержащий требуемую конструкцию, выращивали на среде LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl), дополненной канамицином (50 мг/л) и рифампицином (100 мг/л) при 28°C с помощью устройства для встряхивания (250 об/мин) в течение ночи (приблизительно 18 часов). Затем агробактерии инокулировали в 30 мл LB, также дополненную канамицином (50 мг/л) и рифампицином (100 мг/л). После культивирования в течение ночи при 28°C (приблизительно 18 часов), клетки агробактерий собирали посредством центрифугирования в течение 10 минут при 3000 × g и ресуспендировали в 10 мл жидкой среды MS с 4,9 г/л MES (Sigma Chemical) и 30 г/л сахарозы при pH 5,8. Для агроинфильтрации бактериальную культуру доводили до конечной OD600, составляющей 0,1. Затем культуру клеток дополняли ацетосирингоном до конечной концентрации, составляющей 0,2 мМ, и инкубировали в течение 90 минут при 28°C.
Для агроинфильтрации проростки полностью покрывали суспензией и применяли вакуум (100 кПа) в течение 5-6 секунд. Суспензию удаляли и поддерживали в камере для выращивания при 24-26°C со световым периодом 16 часов в течение четырех суток. Материал проростков выделяли, и общий экстракт белков анализировали с помощью иммуноблота с использованием антител против гамма-зеина.
Трансформация клеток животных
Конструкции p3.1RX3.EGF и p3.1RX3.hGH встраивали в культивируемые клетки млекопитающих 293T, Cos1 или CHO посредством способа трансфекции на основе липофектамина (Invitrogen). Клетки, трансфицированные плазмидой pECFP-N1 (Clontech), содержащей последовательность гена усиленного модифицированного GFP с голубой флуоресценцией, использовали в качестве контроля.
Пример 4: Экстракция белка из листьев табака
Свежий растительный материал
Растительный материал (влажные или сухие листья табака или проростки) погружали в жидкий азот и гомогенизировали с помощью буфера для экстракции T, содержащего 50 мМ Трис-HCl, pH 8, 200 мМ дитиотреитол (DTT) и ингибиторы протеаз [10 мкМ апротинин, 1 мкМ пепстатин, 100 мкМ леупептин, 100 мкМ фенилметилсульфонилфторид (PMSF) и 100 мкМ E64 (Sigma Chemical)]. Гомогенаты центрифугировали при 10000 × g в течение 30 минут при 4°C для удаления нерастворимых веществ. Все растворимые белки (TSP) количественно определяли с использованием анализа белков Бредфорда (BioRad).
Сухие листья табака и экстракция белка
Взрослые трансгенные листья табака и взрослые листья табака дикого типа высушивали в помещении при температуре 37°C в течение двух недель на фильтровальной бумаге. Через две недели листья разрезали и хранили при комнатной температуре в течение 5 месяцев. Все растворимые белки сухого материала экстрагировали, как описано для свежего материала, и анализировали с помощью вестерн-блота.
Пример 5: Получение RPBLA
Гомогенизация
Свежие и высушенные трансгенные листья табака и агроинфильтрированные проростки табака (временная трансформация) помещали в ступку и растирали при 0°C в буфере для экстракции PBP, содержащем 100 мМ Трис, pH 8, 50 мМ KCl, 6 мМ MgCl2, 10 мМ ЭДТА, дополненном 10% сахарозой и ингибиторами протеаз (PMSF, леупептин, апротинин, E-64). Гомогенат дополнительно помещали в лед с использованием политрона (IKA T25 Basic, 24,000 об/мин) с небольшим ротором (диаметр 7,5 мМ), приблизительно десять раз в течение 3-4 секунд. Твердое вещество удаляли фильтрованием через четыре слоя Miracloth (22-24 микрометра) (Calbiochem) для удаления неразрушенной ткани и клеток.
Пример 6: Белки из клеток животных
Трансфицированные клетки выделяли из культуральных планшетов соскабливанием и их суспендировали в среде для гомогенизации B (10 мМ Трис-HCl, pH 8,0, 0,9% NaCl, 5 мМ ЭДТА с ингибиторами протеаз). Суспензию отбирали в 5 мл шприц, с установленной иглой калибра 23, и ее выталкивали приблизительно 30 раз. Разрушение клеток контролировали с помощью фазово-контрастного микроскопа. Гомогенат помещали в ступенчатый градиент сахарозы и центрифугировали, как описано для гомогенатов листьев табака.
Накопление слитых белков во временно трансфицированных клетках анализировали с помощью вестерн-блота с использованием антител против гамма-зеина, индуцированных против гамма-зеина. Через 48 часов трансфекции все растворимые белки клеток экстрагировали с помощью буфера A (100 мМ Трис-HCl, pH 8,0, 150 мМ NaCl, 5 мМ EDTA, 0,5% SDS, 0,5% Triton X-100, 2% 2-меркаптоэтанол и ингибиторы протеаз). Аликвоты среды для инкубации осаждали и хранили при -20°C. Белки, экстрагированные из эквивалентных количеств трансфицированных клеток и среды, разделяли посредством SDS-гель-электрофореза в полиакриламидном геле и переносили на нитроцеллюлозные пластинки для иммунодетекции.
Пример 7: Разделение с помощью прерывистого градиента
Гомогенат центрифугировали при 50 × g в течение 5 минут при 4°C с получением очищенного экстракта. В случае прерывистого градиента сахарозы, этот очищенный гомогенат (супернатант) наслаивали на ступенчатый градиент, состоящий из 2,5 мл 20%-30%-42%-56% (масс./масс.) или 42%-49%-56%-65% (масс./масс.) сахарозы в буфере PBP и его центрифугировали (ультрацентрифуга Beckman Coulter OptimaTM XL-100K) при 80000 × g в роторе с качающейся емкостью (SW41Ti) при 4°C в течение 120 минут без торможения.
Отбирали фракции супернатанта, интерфаз и осадка. Эквивалентные аликвоты фракций супернатанта, интерфазы и осадка осаждали посредством 15% TCA и анализировали с помощью SDS-PAGE и иммуноблота с использованием специфичных антител против экспрессируемых слитых белков. Белки RX3-EGF, RX3-T20, RX3-Ct и RX3-INF определяли с помощью вестерн-блота с использованием антител против гамма-зеина. Электрофоретические гели анализировали посредством окрашивания серебром в соответствии с Morrissey et al., 1981 Anal. Biochem. 117:307-310, для оценки повышения содержания рекомбинантного белка относительно нежелательных белков внутри PB.
Пример 8: Разделение посредством однородного слоя
Гомогенат, полученный как описано выше, центрифугировали в 42% (масс./масс.) слое сахарозы объемом 8 мл (1,18 г/см3) в течение 120 минут при 24000 g при 4°C. Выделяли фракции супернатанта, интерфазы и осадка. RPBLA осаждаются на в нижней части слоя. Для анализа белка эквивалентные аликвоты этих фракций осаждали в 15% TCA, и образцы разделяли в 15% SDS-PAGE и анализировали окрашиванием серебром. Рекомбинантные белки, находящиеся во фракции PB, определяли с помощью иммуноблота с использованием антител против гамма-зеина.
Пример 9: Выделение рекомбинантного белка из выделенных RPBLA
RPBLA, выделенные из 42%-56% (масс./масс.) интерфазы ступенчатых градиентов сахарозы или выделенные посредством слоя с определенной плотностью с 42% (масс./масс.) сахарозой, промывали в буфере PBP и выделяли с помощью кратковременного центрифугирования в течение 5 минут при 16000 × g. Рекомбинантные белки, накопившиеся внутри RPBLA, растворяли в одном объеме буфера SB, содержащего 12,5 мМ борат натрия, pH 8, 0,1% SDS и 2% 2-меркаптоэтанол. Раствор инкубировали в течение ночи (приблизительно 18 часов) при 37°C. Одну аликвоту центрифугировали 10 минут при 16000 × g при комнатной температуре, и супернатант и осадок анализировали посредством SDS-PAGE и вестерн-блота для оценки полного растворения белка.
Пример 10: Белки из трансфицированных клеток дрожжей
Осаждали S. cerevisiae, экспрессирующие рекомбинантные слитые белки. Аликвоты соответствующей среды для инкубации осаждали и хранили при -20°C для анализа. Осадки клеток также замораживали, и после размораживания клетки разбивали стандартными способами с использованием стеклянных гранул и среды Y (50 мМ HCl-Tris pH 8,0, 150 мМ NaCl, 5 мМ ЭДТА, 200 мМ DTT и ингибиторы протеаз). Эквивалентные количества и клеток, и среды анализировали с помощью SDS-PAGE и иммуноблота с использованием специфичных антител против рекомбинантных экспрессируемых белков.
Способ разрушения, применяемый для выделения органелл из трансформированных клеток дрожжей, основан на мягком лизисе сферопластов, описанном в Zinser et al., 1995 Yeast, 11:493-536. Тридцать мл культивируемых трансформированных клеток дрожжей (DO600 приблизительно 0,5) осаждали, промывали 1 M сорбитом и суспендировали в 1 мл буфере для сферопластов (1 M сорбит, 50 мМ фосфат калия, pH 7,5, 14 мМ 2-меркаптоэтанол), содержащем 100 единиц/мл зимолазы. В течение 20-30 минут при 30°C при периодическом осторожном перемешивании давали возможность образоваться сферопластам. После осаждения при 1000 g в течение 6 минут сферопласты промывали буфером для сферопластов без 2-меркаптоэтанола и ресуспендировали в 0,5 мл ледяном буфере для лизиса (0,3 M сорбит, 10 мМ триэтаноламин, 1 мМ ЭДТА и ингибиторы протеаз). После 20 минут во льду при периодическом осторожном перемешивании, лизаты доводили до конечной концентрации 1,0 M сорбита. Лизаты помещали на ступенчатый градиент сахарозы и центрифугировали, как описано для гомогенатов листьев табака. Фракции анализировали с помощью SDS-PAGE и иммуноблота.
Результаты
Пример A: Выделение (очистка) RPBLA с помощью градиента плотности из вегетативных тканей трансгенных растений
Гены, кодирующие полученные на основе гамма-зеина слитые белки RX3-EGF и RX3-T20, встраивали в растения табака посредством Agrobacterium tumefaciens. Для определения тех растений, у которых экспрессия рекомбинантного белка была более высокой, трансформированные растения анализировали с помощью иммуноблота. На фиг.2,A представлен образец обоих белков RX3EGF и RX3T20. Следует отметить, что оба рекомбинантных белка представлены в виде правильно запасаемых белков во всех линиях трансгенных растений. Преобладающие нижние полосы соответствуют мономерным формам слитых белков, и более высокие полосы соответствуют димерам. Слитые белки, как правило, запасаются в виде мультимеров, и количество мономеров и олигомеров, определяемых в иммуноблотах, зависит от степени восстановления дисульфидных связей.
Экстракты листьев табака помещали в ступенчатые градиенты плотности, и накопление рекомбинантных белков в различных фракциях анализировали с помощью иммуноблота (фиг.2,B). Результаты, представленные на фиг.2,B, показывают, что RX3-EGF был представлен во фракциях, соответствующих плотным RPBLA. Большинство из этих органелл обладали плотностями выше 1,2632 (F56, дорожка 6), и для значительной части из них показана плотность выше 1,3163 г/см3; (F65, дорожка 7). Слитый белок RX3-T20 находился в интерфазе 49%-56% сахарозы (дорожка 12), указывая на то, что RPBLA, содержащие RX3-T20, обладают плотностью выше 1,2241 г/см3, где значительная часть из них обладает плотностью выше 1,2632, и значительная часть обладает плотностями выше 1,2632 (дорожка 13).
Эти новые RPBLA, сформированные в листьях табака, обладают плотностями, находящимися в диапазоне плотностей природных белковых телец маиса (Ludevid et al., 1984 Plant Mol. Biol. 3:227-234; Lending et al., 1989 Plant Cell 1:1011-1023), или являются еще более плотными. Следует отметить, что PB RX3-T20 являются в некоторой степени (немного) менее плотными, чем PB RX3-EGF, это объясняется некоторыми конкретными свойствами представляющего интерес белка. Таким образом, несмотря на то что RPBLA, накапливающие рекомбинантные слитые белки, обладают высокими плотностями относительно обычно представленных растворимых компонентов клеток, свойства белка, слитого с доменом RX3, могут определять изменения такой плотности.
Было установлено, что более 90 процентов обоих рекомбинантных белков были выделены из фракций плотных RPBLA и осадка (см. фиг.2,B). Таким образом, выделение RPBLA на основе плотности, по-видимому, является пригодной системой для очистки (концентрации) слитых белков.
Для оценки очистки рекомбинантного белка RX3-EGF посредством выделения RPBLA, фракции с различной плотностью анализировали посредством окрашивания серебром (фиг.2,C). Как можно видеть на окрашенном геле, более 90 процентов эндогенных белков табака были локализованы в растворимой (S) фракции и фракции интерфазы (F422 и F49) градиента, во фракциях, в которых белок RX3-EGF отсутствовал или слабо определялся (см. фиг.2,B). Таким образом, растворимые белки и основной объем белков, представленных в менее плотных органеллах, можно удалить, отбирая одну или две фракции (F56 и F65) градиента.
В отношении степени очистки слитых белков во фракциях RPBLA (F56 и F65) было установлено, что белок RX3-EGF составляет приблизительно 80 процентов белков, определяемых в содержащих PBLS фракциях. Этот результат показывает, что с использованием процесса выделения RPBLA можно добиться существенного увеличения содержания слитых белков всего за одну стадию очистки.
Пример B: Выделение рекомбинантных белков в RPBLA, выделенных из сухих тканей растений
Важным моментом молекулярного культивирования является наличие простых способов хранения растительной биомассы. В связи с этим, высушивание может представлять собой удобный способ снижения объема для хранения и сохранения продукта. Тем не менее, высушивание часто приводит к деградации представляющих интерес белков. Применение высушенных растений для выделения RPBLA, содержащих рекомбинантные белки, представляло бы большой интерес для промышленных целей.
Трансформированные листья табака, запасающие слитые белки RX3-EGF и RX3-T20, как описано выше, высушивали, как также описано выше. Через 5 месяцев хранения в сухом виде, анализировали стабильность рекомбинантных белков. Экстракты белков из эквивалентных количеств влажной (свежей) (W, дорожка 1) и сухой (D, дорожка 2) ткани листьев анализировали с помощью иммуноблота (фиг.2,D). Как можно видеть на фигуре, белок RX3-EGF был стабильным в высушенных трансформированных растениях, где количества, выделенные из влажных и сухих растений, оказались сходными (для сравнения дорожки W и D).
Распределение ступенчатых градиентов плотности слитых белков (RX3EGF и RX3T20) из гомогенатов высушенных листьев анализировали с помощью иммуноблота (фиг.2,D, дорожки 3-7). Интересно, что оба слитых белка, главным образом, выделяли из плотных структур, обладающих плотностями выше 1,1868 г/см3 (фракция F42) и 1,2632 г/см3 (фракция F56).
Таким образом, рекомбинантные белки можно очищать из высушенных тканей посредством выделения RPBLA, что говорит о том, что сбор трансгенных растений и экстракция и очистка рекомбинантного белка, таким образом, могут быть независимы по времени. В соответствии с этими результатами слитые белки гамма-зеина также накапливались в RPBLA в семенах риса.
Пример C: Выделение рекомбинантного белка посредством выделения RPBLA из временно трансформированных проростков табака
Системы с временной экспрессией могут представлять собой удобный инструмент для исследования характера накопления рекомбинантных белков в короткий период времени. Так рекомбинантные белки RX3-EGF и RX3-T20 также экспрессировались и накапливались во временно трансформированных посредством агроинфильтрации проростках табака. Для экстрактов белков из трансформированных проростков, проанализированных с помощью иммуноблота (фиг.3,A), показана характерная сложная электрофоретическая картина, которую наблюдали для стабильно трансформированных растений (для сравнения фиг.3,A, дорожка 4 и фиг.2,A, дорожка 4), указывая на то, что слитые белки запасаются правильно при использовании этого способа трансформации.
Экспрессию слитого белка RX3-hGH с более высокой молекулярной массой также анализировали во временно трансформированном табаке (фиг.3,A, дорожка 4). После субклеточного фракционирования в градиентах плотности оба слитых белка RX3-T20 и RX3-hGH были выделены в плотных фракциях, соответствующих фракциям RPBLA (фиг.3,B, дорожки 4, 5 и 9, 10), демонстрируя плотности выше 1,1868 г/см3 (F42) и 1,2632 г/см3 (F56). Временную экспрессию, таким образом, можно использовать для анализа в течение короткого периода времени конкретных свойств, касающихся плотности, PB, содержащих требуемый рекомбинантный белок.
Пример D: Выделение рекомбинантных белков посредством центрифугирования при низкой и средней скорости
Для упрощения процесса, используемого для очистки рекомбинантных белков посредством плотных рекомбинантных подобных белковым тельцам скоплений, проводили два дополнительных альтернативных способа: i) очищенные гомогенаты центрифугировали только через один плотный слой сахарозы (фиг.4,A, B), и ii) очищенные гомогенаты просто центрифугировали посредством центрифугирования при низкой скорости (т.е. 1000-2500 × g в течение 10 минут).
В соответствии с ранее описанными результатами, оба белка RX3-EGF и RX3-T20 были выделены с высоким выходом (более 90%) в осадках, полученных после центрифугирования через слой сахарозы с плотностью 1,1868 г/см3 (фиг.4,A, дорожки 4 и 6). Кроме того, степень очистки белка RX3-EGF была очень высокой, как можно видеть на окрашенном серебром геле на фиг.4,B, где нежелательные эндогенные белки табака слабо определялись в соответствующем осадке (дорожка 4).
Принципиальное преимущество этого способа по сравнению со ступенчатыми градиентами плотности основано на его простой расширяемости для промышленного производства рекомбинантных белков. Следует отметить, что слой с определенной плотностью, а также другие свойства, такие как вязкость и осмолярность, можно подбирать в каждом случае в целях оптимизации выделения и очистки рекомбинантных белков.
Кроме того, для концентрирования и очистки содержащих слитый белок подобных белковым тельцам структур также анализировали центрифугирование при низкой скорости (LSC) (фиг.4,C, LSC). Результаты показали, что после 1000 × g в течение 10 минут практически весь слитый белок RX3-EGF был выделен из осадка (фиг.4,C дорожка 2). Однако окрашивание белков, находящихся в этом осадке, выявило, что слитый белок не был высоко чистым по сравнению с белком, полученным после центрифугирования через слой сахарозы с плотностью 1,1868 г/см3 (фиг.4,C, для сравнения дорожки 3 и 7).
Соответственно, первый осадок, полученный посредством центрифугирования при низкой скорости, промывали с использованием буфера, содержащего 5% Triton X-100. После промывания образец центрифугировали при 12000 × g в течение 5 минут и, что интересно, большое количество нежелательных белков, находящихся в осадке P1, удалялось после промывания и центрифугирования, и новый осадок (P2, фиг.4,C, дорожка 9) содержал значительно увеличенное содержание белка RX3-EGF. Следует отметить, что количество, а также картина для белков на дорожке 9, являются сходными с количеством и картиной для белков, полученных после промывания осадка, полученного после центрифугирования через слой сахарозы в содержащем Triton X-100 буфере (фиг.4,C, дорожка 5). В основе варианта с центрифугированием при низкой скорости лежит высокая плотность структур, содержащих слитые белки, и условия центрифугирования можно оптимизировать для каждой мишени перед масштабированием.
Пример E: Выделение рекомбинантного белка посредством выделения RPBLA из трансфицированных клеток животных
Были проведены исследования для определения того, индуцируют ли происходящие из запасных белков слитые белки образование плотных рекомбинантных подобных PB скоплений также в трансфицированных клетках животных. Субклеточное распределение органелл из гомогенизированных трансфицированных клеток млекопитающих анализировали посредством применения ступенчатых градиентов плотности. Три различных типа культур клеток, 293T (из человека), Cos1 (из обезьяны) и CHO (из хомяка), трансфицировали с использованием кДНК, кодирующей три различных слитых белка, RX3-Ct, RX3-EGF и RX3-hGH. Клетки Cos1, трансфицированные pECFP-N1 (Clontech), использовали в качестве контроля. Фракции градиента собирали, как описано ранее, и анализировали с помощью иммуноблота (фиг.5,A).
Рекомбинантные полученные из RX3 белки, экспрессированные в трансфицированных клетках, определяли с использованием антисыворотки против гамма-зеина. Определения контрольного ECGP в различных собранных фракциях проводили с использованием антисыворотки против GFP, индуцированной у кроликов.
Как ожидалось, растворимый белок ECGP выделяли из фракции супернатанта (S, фиг.5,A, дорожка 2), и никаких следов этого белка не было выявлено во фракциях интерфазы и осадка, где осаждались дисперсные фракции клеток. Напротив, RX3CT, RX3EGF и RX3hGH, главным образом, находились в плотных фракциях F30, F42 и F56 (фиг.5,A), что говорит о том, что полученные из гамма-зеина слитые белки можно выделять из этих плотных фракций (плотности от 1,1270 до 1,2632 г/см3). Эти результаты согласуются с результатами, полученными посредством иммуноцитохимии, где слитые белки были локализованы в ER и в рекомбинантных подобных белковым тельцам скоплениях с диаметром от приблизительно 1 до приблизительно 1,4 микрон.
Значительное количество рекомбинантного белка было выделено из растворимой фракции градиентов из трансфицированных RX3-Ct и RX3-hGH клеток. Это, возможно, является следствием избыточного разрушения клеток в процессе гомогенизации, которое приводит к растворению еще не собранных слитых белков, находящихся в ER.
Центрифугирование при низкой скорости (LSC, фиг.5,B) также оценивали с использованием гомогенатов из экспрессирующих RX3-EGF клеток CHO. Как можно видеть на фиг.5,B, большое количество слитого белка было выделено из осадка после 2500 × g (дорожка 2), что подтверждает то, что слитые белки накапливаются в плотных подобных белковым тельцам структурах в клетках животных, которые можно выделять способами на основе плотности.
Пример F: Выделение рекомбинантных белков из трансформированных дрожжей с помощью градиентов плотности
Образование плотных структур, содержащих слитые белки, в трансформированных дрожжах также анализировали посредством ступенчатых градиентов плотности. кДНК, кодирующие слитые белки RX3-EGF и RX3-hGH, встраивали посредством вектора для трансформации дрожжей в Sacharomyces cerevisiae с использованием стандартных способов. Способ разрушения, примененный для выделения органелл, был основан на мягком лизисе сферопластов, как описано в способах. Лизаты помещали на ступенчатый градиент сахарозы и центрифугировали, как описано для клеток млекопитающих или гомогенатов листьев табака. Фракции анализировали с помощью SDS-PAGE и иммуноблота.
Результаты фракционирования представлены на фиг.6, где можно видеть, что большая часть обоих белков RX3-EGF и RX3-hGH была локализована в интерфазе F30 (фиг.6, дорожка 4) градиентов. Эта фракция содержит субклеточные структуры с плотностями между 1,1270 и 1,1868 г/см3. Значительных количеств слитых белков в супернатанте и фракции F20 выявлено не было, что указывает на то, что слитые белки накапливаются в дрожжевых клетках. Возможно, что только небольшой размер клеток дрожжей (вплоть до 3 микрон) обеспечивает образование небольших RPBLA, которые обладают меньшей плотностью, чем RPBLA, выявляемые в растениях и клетках. В любом случае, их можно выделять и очищать от других клеточных белков посредством центрифугирования.
Пример G: Отделение слитых белков от различных доменов запасных белков посредством градиентов плотности
Способ очистки полученных из гамма-зеина слитых белков на основе их свойств, касающихся плотности, можно применять для слитых белков, полученных из других запасных белков. Здесь авторы настоящего изобретения показывают, каким образом слитые белки, полученные из проламина риса массой 13 кДа (rP13) и из альфа-зеина массой 22 кДа (22aZt), накапливаются в плотных фракциях, соответствующих RPBLA на ступенчатых градиентах сахарозы (фиг.7). Выбор проламина риса массой 13 кДа (rP13) и альфа-зеина массой 22 кДа [в полноразмерном варианте (22aZ) или N-концевого домена (22aZt)] был основан на отсутствии гомологии между ними и относительно домена RX3. Неожиданно, оба запасных белка образовывали RPBLA, которые были выделены в более плотных интерфазах, после того, как их подвергали ступенчатому градиенту плотности (фиг.7,A).
Проводили слияние последовательности кальцитонина в рамке считывания с последовательностями rP13 и 22aZt под действием промотора CaMV35S и осуществляли встраивание в Agrobacterium tumefaciens. Проростки табака временно трансформировали, и гомогенаты листьев подвергали ступенчатым градиентам плотности. Собранные фракции анализировали с помощью SDS-PAGE и иммуноблота с использованием антисыворотки против кальцитонина, индуцированной у кроликов.
Как можно видеть на фиг.7,A (дорожки 4-5, и 9-10), большие количества обоих слитых белков rP13Ct и 22aZtCt были локализованы во фракциях F42 и F56, указывая на наличие рекомбинантных подобных PB скоплений с плотностями, превышающими 1,1868 г/см3, которые можно выделять для очистки слитого белка. Результаты также указывают на то, что плотность рекомбинантных подобных PB скоплений, содержащих слитые белки, может варьировать в зависимости от запасного белка, участвующего в слиянии.
Исследования по агроинфильтрации проводили с hGH, слитым с проламином риса (rP13-hGH) и полноразмерным альфа-зеином (22aZ-hGH). Снова, большая часть RPBLA наблюдалась во фракциях F42 и F56, 7,A (дорожка 4-5 и дорожки 9-10), однако интересно, что в то же время не образовавший скоплений слитый белок также был выявлен в супернатанте и интерфазах с низкой плотностью (дорожки 1-2 и 6-7). Этот эффект можно объяснить частичным эффектом растворения hGH на rP13 и 22aZ.
Трансгенные растения табака, экспрессирующие слитые белки rP13-EGF и 22aZ-EGF, получали посредством трансформации с помощью Agrobacterium tumefasciens. Растения с наивысшей экспрессией определяли с помощью иммуноблота с использованием антитела против EGF, и эти клеточные линии использовали в сравнительном анализе с проростками табака, агроинфильтрированными теми же конструкциями. Как можно видеть на фиг.7 (A, дорожки 4 и 9; B, дорожка 3) во всех случаях, RPBLA были выделены в определенной интерфазе (F12), что подразумевает, что RPBLA являются высоко плотными и гомогенными.
Все эти результаты, взятые вместе, позволяют понять, что проламины способны индуцировать RPBLA с высокой плотностью, даже когда они являются слитыми с другими белками. Это представляет собой неожиданный результат, главным образом, в связи с тем, что почти никакой гомологии между ними не выявлено. Более того, существуют некоторые данные, позволяющие предположить, что проламины взаимодействуют для стабилизации белковых телец и что некоторые из них не являются стабильными при экспрессии в вегетативной ткани по отдельности, например, такие как альфа-зеин (Coleman et al., 1996 Plant Cell 8:2335-2345).
Пример H: Экстракция рекомбинантных белков из выделенных PBLS
Было показано, что выделение плотных рекомбинантных подобных PB скоплений является преимущественным способом для выделения рекомбинантных белков с высоким выходом и высоким уровнем очистки из трансгенных организмов. Здесь было показано, что эти рекомбинантные белки можно экстрагировать из запасающих органелл. Несмотря на то что рекомбинантные PBLA можно непосредственно использовать в некоторых способах применения (т.е. получение пероральной вакцины), в некоторых других случаях может быть необходимым использование очищенных рекомбинантных белков.
После инкубации в течение ночи (приблизительно 18 часов) фракций PB при 37°C в буфере, содержащем восстановители (обработка буфером SB, который содержит 12,5 мМ борат натрия, pH 8, 0,1% SDS и 2% 2-меркаптоэтанол), белки RX3-EGF и RX3-T20 растворялись. Как можно видеть на фиг.8, дорожки 1-4 для RX3-EGF и дорожки 6-7 для RX3-T20, оба экстрагированных слитых белка были выделены в их растворимых формах (S). Впоследствии, в зависимости от способа их применения, экстрагированные белки можно подвергать дальнейшей очистке или использовать в качестве частично очищенных экстрактов.
Все патенты и статьи, цитированные в настоящем описании, включены в настоящее описание в качестве ссылок. Использование неопределенного артикля подразумевает, что он включает один или несколько.
Приведенное выше описание и примеры представлены в качестве иллюстративных и их не следует толковать как ограничивающие. Возможны другие варианты в пределах сущности и объема этого изобретения, и они будут легко доступны специалистам в данной области.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ БИОЛОГИЧЕСКИ АКТИВНЫХ БЕЛКОВ | 2007 |
|
RU2441911C2 |
| ПРОИЗВОДСТВО БЕЛКОВ | 2005 |
|
RU2429243C2 |
| ПОЛУЧЕНИЕ ПЕПТИДОВ И БЕЛКОВ ПУТЕМ АККУМУЛИРОВАНИЯ В ЭНДОПЛАЗМАТИЧЕСКОМ РЕТИКУЛУМЕ РАСТЕНИЙ | 2002 |
|
RU2381275C2 |
| РАСТИТЕЛЬНЫЙ ПЕПТИД ГАММА-ЗЕИН ДЛЯ ДОСТАВКИ БИОМОЛЕКУЛ В РАСТИТЕЛЬНЫЕ КЛЕТКИ | 2011 |
|
RU2563805C2 |
| КОМПОЗИЦИЯ (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ НЕ ПРИРОДНОЙ УПОРЯДОЧЕННОЙ И СОДЕРЖАЩЕЙ ПОВТОРЫ АНТИГЕННОЙ МАТРИЦЫ, СПОСОБ ТЕРАПЕВТИЧЕСКОГО ЛЕЧЕНИЯ И СПОСОБ ИММУНИЗАЦИИ | 1999 |
|
RU2245163C2 |
| ИММУНИЗАЦИЯ ПРОТИВ CHLAMYDIA TRACHOMATIS | 2002 |
|
RU2331435C2 |
| ПОЛУЧЕНИЕ ВИРУСОПОДОБНЫХ ЧАСТИЦ В РАСТЕНИЯХ | 2011 |
|
RU2655431C2 |
| ИЗОПРОПИЛМАЛАТ СИНТАЗА ИЗ NICOTIANA TABACUM И СПОСОБЫ И ЕЕ ПРИМЕНЕНИЕ | 2012 |
|
RU2644238C2 |
| УСТОЙЧИВЫЕ К БАКТЕРИЯМ ТРАНСГЕННЫЕ РАСТЕНИЯ | 2012 |
|
RU2640246C2 |
| ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ, СОДЕРЖАЩИЕ БЕЛКИ-МИШЕНИ, СЛИТЫЕ С БЕЛКАМИ ОБОЛОЧКИ РАСТИТЕЛЬНЫХ ВИРУСОВ | 2010 |
|
RU2571223C2 |



















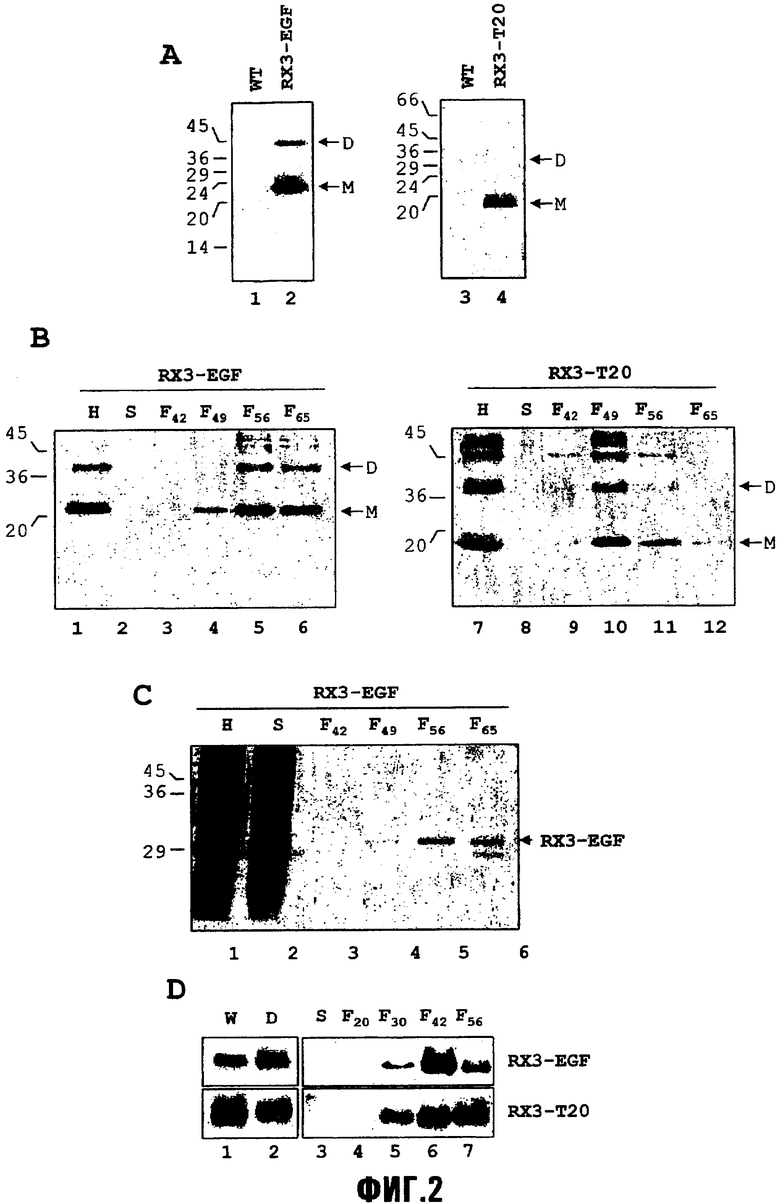

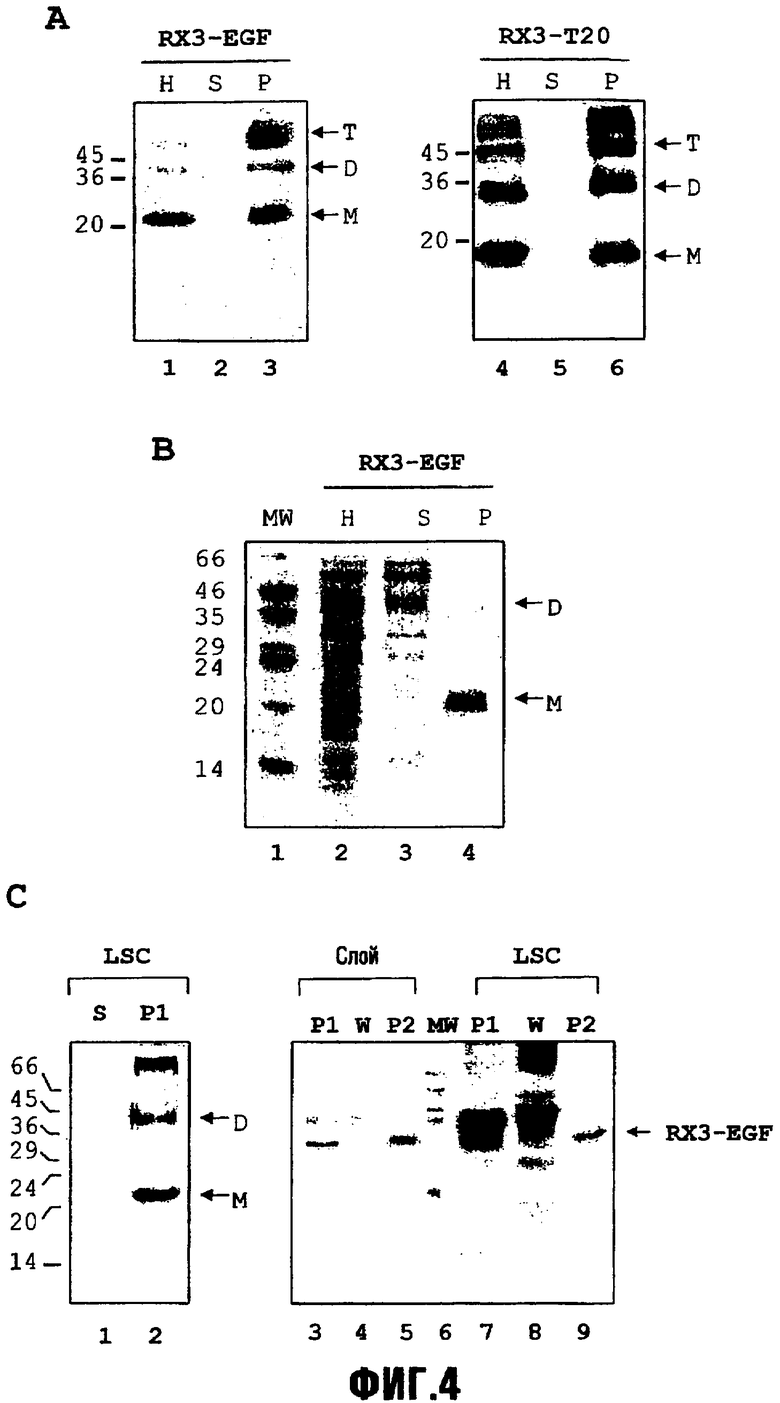

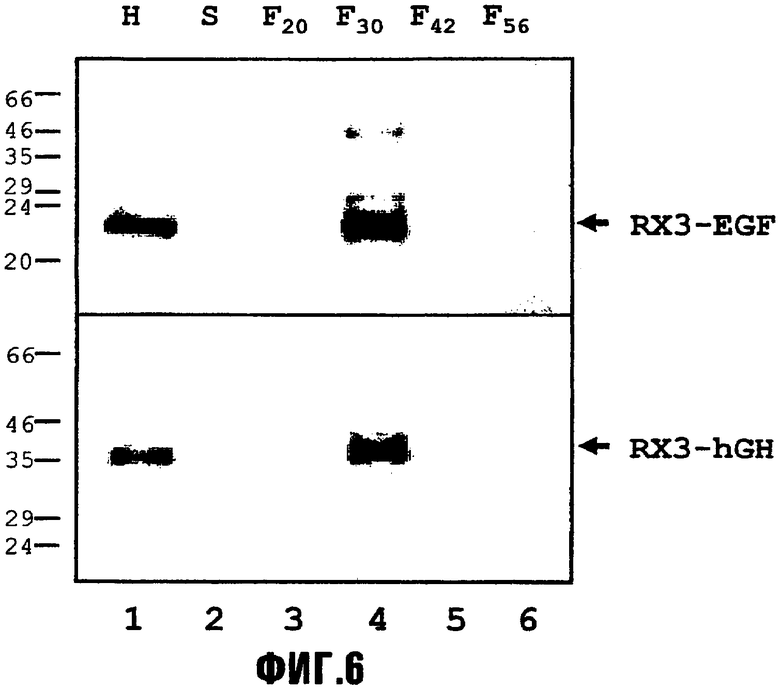
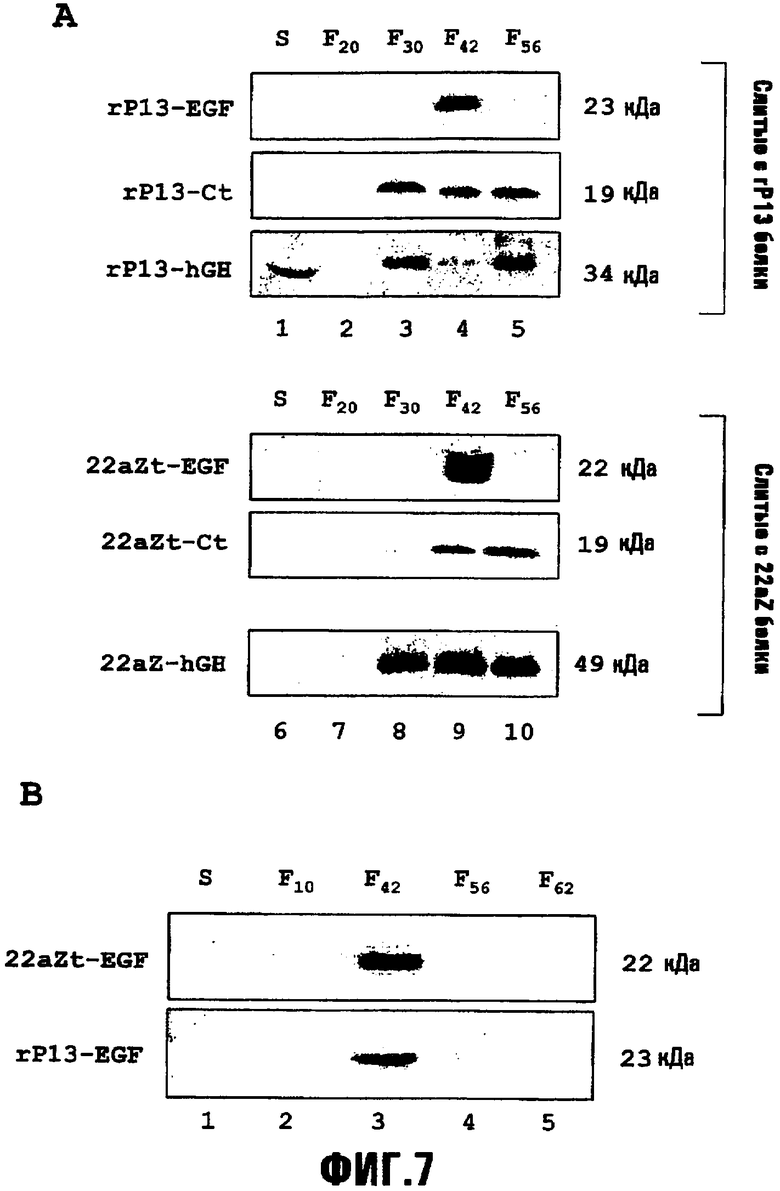
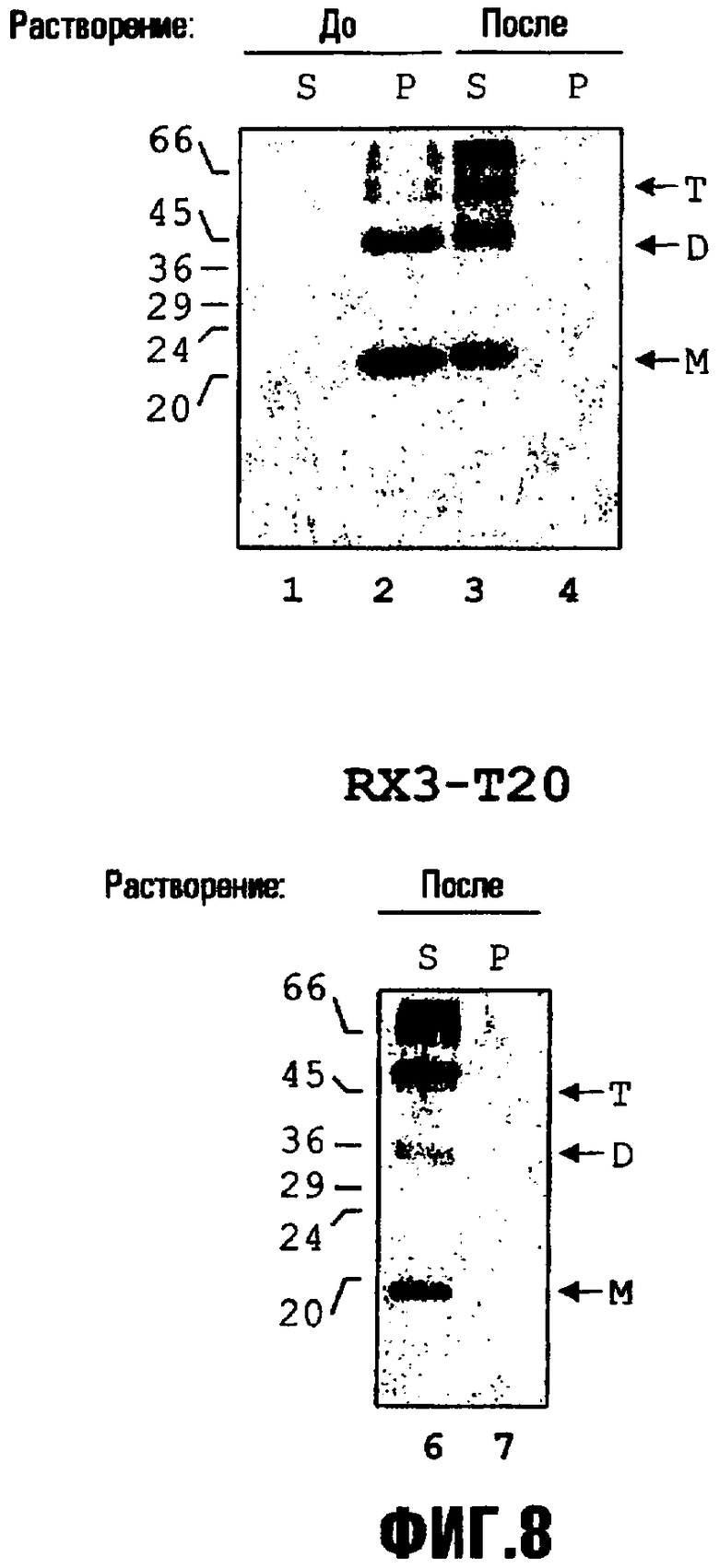
Изобретение относится к способу выделения рекомбинантных белков из трансформированных хозяев. Данные рекомбинантные белки содержат индуцирующую белковые тельца последовательность, слитую с последовательностью представляющего интерес продукта. При осуществлении способа получают водный гомогенат трансформированных клеток-хозяев, которые экспрессируют слитый белок в виде рекомбинантных подобных белковым тельцам скоплений (RPBLA), обладающих плотностью приблизительно от 1,1 до 1,35 г/мл. Гомогенат центрифугируют с образованием фракций с различной плотностью, либо в отсутствие дополнительного обеспечивающего плотность раствора, либо с помощью слоя с определенной плотностью, которая меньше чем плотность RPBLA. В результате образуются фракции, которые содержат относительно сниженную концентрацию RPBLA, и фракция, которая содержит относительно повышенную концентрацию RPBLA. Фракцию со сниженной концентрацией RPBLA отделяют от фракций с относительно повышенной концентрацией RPBLA. Затем очищают указанный слитый белок. Изобретение обеспечивает простоту осуществления, быстроту и эффективную очистку рекомбинантных белков. 14 з.п. ф-лы, 8 ил., 1 табл.
1. Способ очистки рекомбинантного слитого белка, экспрессированного в качестве рекомбинантных подобных белковым тельцам скоплений (RPBLA) в клетках-хозяевах, который включает стадии:
(a) получения водного гомогената, полученного путем гомогенизации в буфере трансформированных клеток-хозяев, которые продуцируют слитый белок в виде рекомбинантных подобных белковым тельцам скоплений (RPBLA), где указанные RPBLA обладают заранее определенной плотностью, где указанные RPBLA обладают плотностью, составляющей от приблизительно 1,1 до приблизительно 1,35 г/мл, и где указанный слитый белок содержит две последовательности, связанные друг с другом, в которых одна последовательность представляет собой индуцирующую белковые тельца последовательность, и другая представляет собой последовательность представляющего интерес продукта, где индуцирующая белковые тельца последовательность содержит проламин или модифицированный проламин;
(b) образования фракций с различной плотностью в гомогенате путем центрифугирования гомогената в отсутствие дополнительного обеспечивающего плотность раствора или с помощью слоя с определенной плотностью, обладающего плотностью меньшей, чем плотность указанных RPBLA, с получением фракции, которая содержит относительно повышенную концентрацию RPBLA, и фракции, которая содержит относительно сниженную концентрацию RPBLA;
(c) отделения указанной фракции, содержащей относительно сниженную концентрацию RPBLA, от указанной фракции с относительно повышенной концентрацией RPBLA и отбора фракции с относительно повышенной концентрацией RPBLA; и
(d) очищения указанного слитого белка.
2. Способ по п.1, в котором указанная заранее определенная плотность RPBLA является большей, чем плотность, по существу, всех эндогенных белков клетки-хозяина, находящихся в гомогенате.
3. Способ по п.1 или 2, в котором указанные фракции с различной плотностью получают с помощью слоя с определенной плотностью, обладающего плотностью меньшей, чем плотность указанных RPBLA.
4. Способ по п.1 или 2, в котором указанные фракции с различной плотностью получают с помощью супернатанта и осадка, полученных после центрифугирования гомогената при низкой скорости.
5. Способ по п.1, в котором указанные клетки-хозяева представляют собой клетки высших растений.
6. Способ по п.1, в котором указанные клетки-хозяева представляют собой клетки грибов.
7. Способ по п.6, в котором указанные клетки-хозяева грибов представляют собой клетки дрожжей.
8. Способ по п.1, в котором указанные клетки-хозяева представляют собой клетки водорослей.
9. Способ по п.1, в котором указанные клетки-хозяева представляют собой клетки животных.
10. Способ по п.9, в котором указанные клетки-хозяева животных представляют собой клетки млекопитающих.
11. Способ по п.1 или 2, в котором указанный слитый белок дополнительно включает линкерную последовательность между индуцирующей белковые тельца последовательностью и последовательностью представляющего интерес продукта.
12. Способ по п.1 или 2, в котором последовательность проламина представляет собой гамма-зеин, альфа-зеин или проламин риса.
13. Способ по п.1 или 2, в котором гомогенат получают из свежей биомассы.
14. Способ по п.1 или 2, в котором гомогенат получают из высушенной биомассы.
15. Способ по п.1 или 2, включающий дополнительную стадию выделения RPBLA.
| MAINIERI D | |||
| ЕТ AL.: "Zeolin | |||
| A new recombinant storage protein constructed using maize g-zein and bean phaseolin", Plant Physiology, 2004, v | |||
| Регулятор для ветряного двигателя в ветроэлектрических установках | 1921 |
|
SU136A1 |
| TULLY R.E | |||
| ЕТ AL.: "Protein bodies of castor bean endosperm", Plant | |||
| Physiol., 1976, v.58, p.710-716 | |||
| BAGGA S | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |

