Изобретение относится к молекуле ДНК, содержащей интронные последовательности и кодирующей человечий белок, который в зависимости от места действия называется стимулируемой солями желчных кислот липазой (ССЖЛ) или карбоксил-сложноэфирной липазой (КСЛ). ДНК молекула с успехом применима в получении рекомбинантных человечьих ССЖЛ/КСЛ, предпочтительно посредством продукции в трансгенных млекопитающих (но не человека). Рекомбинантные человечьи ССЖЛ/КСЛ могут быть использованы в качестве компонента детского питания в виде заменителя материнского молока, предназначенного для кормления младенцев, или для изготовления лекарственных средств против, например, малабсорбции жиров, муковисцидоза и хронического панкреатита.
Гидролиз пищевых липидов
Пищевые липиды являются важным источником энергии. Богатые энергией триацилглирецины составляют более 95% таких липидов. Некоторые липиды, например определенные жирные кислоты и растворимые в жирах витамины являются существенными компонентами пищи. Перед абсорбцией желудочно-кишечным трактом триацилглицерины, а также присутствующие в меньших количествах компоненты, например этерифицированные растворимые в жирах витамины и холестерин, и диацилфосфатидилглицерины, должны подвергнуться гидролизу по сложноэфирной связи с образованием менее гидрофобных абсорбируемых продуктов. Реакция гидролиза катализируется особой группой ферментов, называемых липазами.
В организме взрослого человека к важнейшим липазам относятся желудочная липаза, панкреатическая колипаза - зависимая липаза (гидролиз три- и диацилглицеринов), панкреатическая фосфолипаза A2 (гидролиз диацилфосфатидилглицеринов) и карбоксил-сложноэфирная липаза (КСЛ) (гидролиз сложных эфиров холестирина и растворимых в жирах витаминов). У вскармливаемых грудью новорожденных стимулируемая солями желчных кислот липаза (ССЖЛ) играет существенную роль в гидролизе некоторых из вышеупомянутых липидов. Совместно с солями желчных кислот продукты гидролиза липидов образуют смешанные мицеллы, из которых и происходит абсорбция.
Стимулируемая солями желчных кислот липаза
В лактирующей молочной железе человека происходит синтез и секреция с молоком стимулируемой солями желчных кислот липазы (ССЖЛ) (Blackberg и др., 1987), которая после особой активации первичными солями желчных кислот придает вскармливаемому грудью младенцу эндогенную способность усваивать в кишечнике жиры. Указанный фермент, составляющий примерно 1% всего молочного белка (Blackberg & Hernell, 1981), не разрушается при прохождении с молоком через желудок, а от содержащихся в двенадцатиперстной кишке панкреатических протеаз, например трипсина и химострипсина, липаза защищена от дезактивации солями желчных кислот. И тем не менее липаза дезактивируется при пастеризации молока, например при нагревании при 62,5oC, 30 мин (Bjorksten и др., 1980).
Опыты на моделях in vitro предполагают, что конечные продукты гидролиза триацилглицерина отличаются в присутствии ССЖЛ (Bernback и др., 1990; Hernell & Blackberg, 1982). Вследствие низкой внутрипросветной концентрации солей желчных кислот в течение неонатального периода, это может иметь важное значение для абсорбции продукта.
Карбоксил-сложноэфирная липаза
Карбоксил-сложноэфирная липаза (КСЛ) человеческого панкреатического сока (Lombardo и др.) функционально, видимо, идентична или по меньшей мере схожа с ССЖЛ (Blackberg и др., 1981). Кроме того, обе липазы характеризуются общими эпитопами, имеют идентичные N-концевые последовательности аминокислот (Abouakil и др., 1988) и ингибируются ингибиторами серин-эстераз, например эсерином и диизопропилфторфосфатом. В недавних исследованиях, проведенных в нескольких лабораториях, выявлено строение кДНК как молочной липазы, так и панкреатической липазы (Baba и др., 1991; Hui и др., 1991; Nilsson и др., 1990; Reue и др. , 1991), и исследователи пришли к выводу, что и молочный фермент, и панкреатический фермент являются продуктами одного и того же гена (в данной заявке называется CEL-геном, EC 3.1.1.1). Последовательность кДНК и определенная на ее основе аминокислотная последовательность описаны в WO 91/15234 (Оклахома Медикал Рисерч Фаундейш) и в WO 91/18923 (Актиэболагет Астра).
Таким образом, предполагается, что КСЛ идентичен ССЖЛ, и полипептид, кодируемый CEL-геном, в контексте настоящей заявки называется ССЖЛ/КСЛ.
Малабсорбция липидов
Общие причины малабсорбции липидов, а следовательно, нарушенного питания заключается в пониженном внутрипросветного содержания панкреатической колипазазависимой липазы и/или солей желчных кислот. Типичные примеры случаев такого дефицита липазы включают больных, страдающих муковисцидозом - обычным генетическим нарушением, сопровождающих 80% больных с таким дефицитом на протяжении всей их жизни, и хроническим панкреатитом, часто связанным с хроническим алкоголизмом.
Лечение больных, страдающих дефицитом панкреатической липазы, в настоящее время заключается во введении очень больших доз сырого препарата панкреатических ферментов свиньи. Однако колипазазависимая панкреатическая липаза дезактивируется при низких значениях pH, преобладающих в желудке. Этот феномен не может быть полностью преодолен применением больших доз фермента. Таким образом, вводимые в больших дозах препараты не приносят облегчения большинству больных, а сами препараты загрязнены и неприятны на вкус.
Приготовлены таблетки, способные пройти через кислотные области желудка с выделением фермента только в сравнительно щелочном окружении тощей кишки. Однако многие больные, страдающие панкреатическими нарушениями, отличаются необычайно высоким содержанием кислоты в тощей кишке, и в этих случаях фермент из таблеток может не выделиться.
Более того, поскольку выпускаемые в настоящее время промышленностью препараты животного происхождения, имеется риск иммунных реакций, могущих нанести вред больным или уменьшить эффективность лечения. Еще один недостаток существующих препаратов заключается в том, что для них не установлено проявлений других форм лополитической активности, отличной от активности колипазазависимой липазы. В самом деле, большинство таких препаратов в очень незначительной степени проявляют ССЖЛ/КСЛ активность. В этом может быть одна из причин того, что страдающие муковисцидозом больные, несмотря на поддерживающую терапию, испытывают недостаток в растворимых в жирах витаминах и в необходимых жирных кислотах.
Таким образом, существует большая необходимость в продуктах со свойствами и строением, аналогичными свойствами и строению человечьих липаз, и широкой специфичностью субстрата, при этом такие продукты могут быть введены перорально больным, страдающим дефицитом одного или нескольких панкреатических липолитических ферментов. Продукты, которые могут быть приготовлены использованием настоящего изобретения, отвечают такой необходимости как сами по себе, так и в комбинации с препаратами, содержащими другие липазы.
Детское питание
Хорошо известно, что вскармливание младенца материнским молоком считается более полезным, чем искусственное кормление. И не только потому, что молоко человека является хорошо сбалансированным источником питательных веществ, но еще и потому, что молоко матери легко усваивается младенцем. Так, некоторые биологически активные вещества, которые, как известно, выполняют физиологические функции в организме младенца, являются либо компонентами человечьего молока, либо образуются в ходе его усвоения, включая компоненты, участвующие в защите от инфекции, и компоненты, облегчающие усвоение питательных веществ человечьего молока.
Несмотря на серьезные попытки в направлении получения детского питания, так и не удалось создать состав, хоть в какой-то заметной степени отвечающего преимуществам материнского молока. Так, детское питание, часто приготовляемое на основе коровьего молока, как правило, не полностью усваивается младенцем, и в нем отсутствуют вещества, которые, как известно, оказывают влияние на физиологические функции организма младенца. С целью получения детского питания, аналогичного по своей питательной ценности материнскому, в состав питания предлагались разнообразные добавки, в том числе белковые фрагменты, витамины, минеральные вещества и т.д. с последующим риском повышенной нагрузки на важнейшие органы, например печень и почки, с их возможным длительным разрушением. Другой недостаток, связанный с применением составов на основе коровьего молока, заключается в повышенном риске возникновения у младенца аллергии к коровьим белкам.
В качестве альтернативы детскому питанию на основе коровьего молока используется человечье молоко, которое можно получить в так называемых молочных банках. Однако кормление новорожденных младенцев молоком из молочных банков в последние годы в значительной степени стараются избежать из боязни присутствия в молоке инфекционных агентов, например ВИЧ или ВОМ (вирус огуречной мозаики). Для разрушения инфекционных агентов в человечьем молоке молоко перед употреблением необходимо пастеризовать. Однако при пастеризации питательная ценность и биологическое действие компонентов молока уменьшается, например, как указано выше, происходит дезактивация ССЖЛ.
Введение в детское питание липаз
Функционирование поджелудочной железы и печени при рождении проявляется не в полной мере, что особенно заметно в случае недоношенного ребенка. По физиологическим причинам малабсорбция жиров является наиболее общим нарушением, которое, как полагают, возникает в результате низкой внутрипросветной концентрации панкреатической колипазазависимой липазы и солей желчных кислот Однако вследствие присутствия ССЖЛ такая малабсорбция отмечается гораздо реже в случаях вскармливания грудью младенцев по сравнению с младенцами, вскармливаемыми пастеризованным человечьим молоком или детским питанием (Bernback и др., 1990).
Чтобы обойти вышеуказанные недостатки, связанные с применением пастеризованного молока и детского питания на основе коровьего молока, желательно, таким образом, создать такое детское питание, состав которого был бы особенно близок составу материнского молока, то есть состав, содержащий белки материнского молока.
ССЖЛ/КСЛ обладают несколькими уникальными свойствами, делающими их идеально применимыми в качестве добавки к детскому питанию.
Самой природой эти липазы предназначены для перорального введения. Так липазы устойчивы к прохождению через желудок и активируются под действием содержимого тонкой кишки.
Вследствие своей широкой специфичности к субстрату липазы со своей стороны обладают потенциалом, способствующим полному усвоению большинства пищевых липидов, в том числе и растворимых в жирах сложных эфиров витаминов.
ССЖЛ/КСЛ могут превосходить панкреатическую колипазазависимую липазу по способности гидролизовать сложноэфирные связи в эфирах, содержащих длиноцепочечные полиненасыщенные жирные кислоты.
В присутствии желудочной липазы и в отсутствие или при низком содержании колипазазависимой липазы ССЖЛ/КСЛ способы обеспечить полный гидролиз триацилглицерина in vitro даже при низкой концентрации солей желчных кислот, как, например, в случае новорожденных младенцев. В присутствии ССЖЛ/КСЛ конечными продуктами гидролиза триацилглицерина являются свободные жирные кислоты и свободный глицерин, а не моноацилглицерин, образующийся в присутствии двух других липаз (Bernback и др., 1990). Это может благоприятствовать абсорбции продукта, особенно при низком внутрипросветном содержании солей желчных кислот.
Для применения ССЖЛ/КСЛ в качестве добавки к детскому питанию необходимо, однако, иметь доступ к большим количествам этого продукта. Хотя белки человечьего молока могут быть выделены непосредственно из молока, такой подход не является реалистичным и достаточно экономичным для получения больших количеств продукта, необходимых для широкомасштабного производства детского питания. В связи с этим необходимо создать другие методы для того, чтобы иметь возможность получать детское питание, содержащее белки человечьего молока. Настоящим изобретением даются такие способы получения в больших количествах ССЖЛ/КСЛ.
Продуцирование белков с молоком трансгенных животных
Выделение генов, кодирующих фармакологически активные белки, позволяет получать такие белки более дешевым путем в гетерологичных системах. Приемлемая экспрессионная система для молочных белков может быть представлена трансгенным животным (обзор см. Hennighausen и др., 1990). Пищевые составы, содержащие активированные солями желчных кислот липазы, полученные, например, технологией с использованием трансгенных животных, описаны в EP 317,355 (Оклахома Медикал Рисерч Фаундейшн).
В случае трансгенного животного кодирующая белок последовательность может быть введена в виде кДНК или в виде геномной последовательности. Поскольку для регулируемой экспрессии гена могут потребоваться интроны (Brinster и др., 1988; Whitelaw и др., 1991), во многих случаях рекомендуется применять геномную форму, а не кДНК форму структурного гена. В патенте WO 90/05188 (Фармасьютикал Протеинс Лимитид) описано применение в трансгенном животном кодирующей белок ДНК, включающей хотя бы один, но не все интроны, встречающиеся в природном кодирующем белок гене.
Целью изобретения является создание средств для получения с высоким выходом и по приемлемой цене человечьих рекомбинантных ССЖЛ/КСЛ, предназначенных для применения в детском питании с тем, чтобы избежать недостатков, характерных для пастеризованного молока и составов на основе коровьих белков.
Цель изобретения достигнута клонированием и секвенированием человечьего CEL-гена. Для повышения выхода ССЖЛ/КСЛ при продуцировании человечьих ССЖЛ/КСЛ в трансгенном млекопитающем (но не человеке) вместо известной к ДНК последовательности применена полученная ДНК молекула, содержащая интронные последовательности.
Соответственно, в одном из своих аспектов настоящее изобретение относится к молекуле ДНК, показанной в перечне последовательностей, как ПОСЛЕДОВАТЕЛЬНОСТЬ N 1, или аналог указанной молекулы ДНК, образованный гибридизацией с молекулой ДНК, показанной в перечне последовательностей, как ПОСЛЕДОВАТЕЛЬНОСТЬ N 1, или ее специфичной частью в жестких условиях гибридизации.
Методика, применяемая для выделения молекулы ДНК человечьих ССЖЛ/КСЛ, приведена в нижеследующих примерах.
Жесткие условия гибридизации, упомянутые выше, следует понимать в их обычном значении, т. е. как гибридизацию, проводимую согласно стандартному лабораторному руководству, например Sambrook и др., (1989).
Другим аспектом изобретения является экспрессирующая система млекопитающего, включающая ДНК последовательность, кодирующую человечьи ССЖЛ/КСЛ, встроенную в ген, кодирующий молочный белок отличного от человека млекопитающего, с образованием в результате гибридного гена, способного экспрессироваться в молочной железе взрослой самки млекопитающего, принявшей гибридный ген, с последующим продуцированием человечьих ССЖЛ/КСЛ при экспрессировании гибридного гена.
И в еще одном своем аспекте настоящее изобретение относится в способу создания трансгенного отличного от человека млекопитающего, способного экспрессировать человечьи ССЖЛ/КСЛ. Способ состоит с инъецировании вышеохарактеризованной экспрессионной системы млекопитающего в оплодотворенное яйцо или клетку эмбриона млекопитающего с введением в результате экспрессирующей системы в зародышевую линию млекопитающего с последующим развитием инъецированного оплодотворенного яйца или эмбриона во взрослую самку млекопитающего.
Молекула ДНК, показанная в перечне последовательностей, как ПОСЛЕДОВАТЕЛЬНОСТЬ N 1, общей длины в 11531 п.о. имеет следующие признаки (см. в конце текста).
В контексте настоящей заявки термин "ген" применяется для обозначения ДНК последовательности, участвующей в продуцировании полипептидной цепи и включающей области до и после области кодирования (5'-восходящая и 3'-нисходящая последовательности), а также промежуточные между отдельным кодирующими сегментами (так называемые экзоны) или в 5'-восходящей или 3'-нисходящей области. Восходящая 5'-область включает регуляторную последовательность, контролирующую экспрессию гена, как правило, промотор. Нисходящая 3'-область включает последовательности, участвующие в прекращении транскрипции гена, и возможно последовательности, ответственные за полиаденилирование транскрипта и нетранслируемой 3'-области.
Молекулы ДНК изобретения, о которых идет здесь речь, могут представлять собой естественные или синтетические ДНК последовательности, причем источником естественной последовательности обычно является непосредственно геномная ДНК, как правило, млекопитающего, например, описанная ниже. Синтетическая последовательность может быть получена обычными способами синтеза молекулы ДНК. Кроме того, ДНК последовательность может быть смешанного геномного и синтетического происхождения.
В еще одном своем аспекте изобретение относится к реплицируемому вектору экспрессии, при участии которого проводят и который способен поддержать экспрессию ДНК последовательности, кодирующей человечьи ССЖЛ/КСЛ.
В контексте настоящей заявки термин "реплицируемый" означает, что вектор способен реплицироваться в клетке-хозяине данного типа, в которую был введен. Непосредственно в восходящем направлении от ДНК последовательности человечьих ССЖЛ/КСЛ может быть предусмотрена последовательность, кодирующая сигнальный пептид, присутствие которого гарантирует секрецию человечьих ССЖЛ/КСЛ, экспрессируемых клетками-хозяевами, принявших вектор. Сигнальная последовательность может быть последовательностью, связанной в природе с ДНК последовательностью человечьих ССЖЛ/КСЛ, или иметь иной источник происхождения.
Вектор может быть представлен любым вектором, который может быть использован в методах биотехнологии, и выбор вектора часто зависит от клетки-хозяина, в которую вектор будет вводиться. Так вектор может быть представлен автономно реплицирующимся вектором, то есть вектором, существующим в виде внехромосомной частицы, репликация которой не зависит от хромосомной репликации. Примеры подобных векторов включают: плазмиды, фаги, космиды, минихромосомы или вирусы. Или же вектор может быть таким, который при введении в клетку-хозяина интегрируется в геном клетки-хозяина и реплицируется вместе с хромосомой-(ами), с которой был интегрирован. Примеры приемлемых векторов включают бактериальные вектора экспрессии и дрожжевые векторы экспрессии. Вектор изобретения может нести любую молекулу ДНК изобретения, охарактеризованную выше.
Изобретение, кроме того, относится к клетке, принявшей вышеохарактеризованный реплицируемый вектор экспрессии. В принципе, такая клетка может быть клеткой любого типа, то есть прокариотной клеткой, одноклеточным эукариотным микроорганизмом или клеткой, происходящей из многоклеточного организма, например, млекопитающего. Клетки млекопитающего особенно пригодны для целей изобретения и более подробно обсуждаются ниже.
В другом своем важном аспекте изобретение относится к способу продуцирования рекомбинантных человечьих ССЖЛ/КСЛ, в котором ДНК последовательность, кодирующую человечьи ССЖЛ/КСЛ, вводят в вектор, способный реплицироваться в специфичной клетке-хозяине, причем полученный рекомбинантный вектор вводят в клетку-хозяина, выращиваемую в соответствующей культурной среде в приемлемых для экспрессии человечьих ССЖЛ/КСЛ условиях с последующим выделением человечьих ССЖЛ/КСЛ.
Применяемой для выращивания клеток средой может служить любая пригодная для таких целей среда. Приемлемый вектор может быть представлен любым вышеописанным вектором и клетка-хозяин может относиться к любому из вышеперечисленных клеточных типов. Для конструирования вектора и осуществления его введения в клетку-хозяина могут быть любые известные способы, применяемые для этих целей в биотехнологии. Экспрессируемые клетками рекомбинантные человечьи ССЖЛ/КСЛ могут секретироваться, например, переноситься через клеточную мембрану по механизму, зависящему от типа клетки и состава вектора.
Если человечьи ССЖЛ/КСЛ продуцируются рекомбинантным хозяином внутриклеточно, то есть не секретируются клеткой, в этом случае их выделяют стандартным методом, включающими разрушение клетки механическими средствами, например обработкой ультразвуком или гомогенизацией или ферментативными или химическими средствами с последующей очисткой.
Для возможности секретирования ДНК последовательности, кодирующей человечьи ССЖЛ/КСД, ей должна предшествовать последовательность, кодирующая сигнальный пептид, присутствие которой гарантирует секретирование из клетки человечьих ССЖЛ/КСЛ таким образом, что по меньшей мере значительная часть экспрессируемых человечьих ССЖЛ/КСЛ секретируется в культурную среду с последующим их выделением из среды.
Рекомендуемый способ продуцирования рекомбинантных человечьих ССЖЛ/КСЛ настоящего изобретения состоит в использовании трансгенных, отличных от человека млекопитающих, способных выделять человека млекопитающих имеет то преимущество, что рекомбинантные человечьи ССЖЛ/КСЛ могут быть получены с высокими выходами и при разумных затратах, особенно, если отличным от человека млекопитающим является корова. В этом случае рекомбинантные человечьи ССЖЛ/КСЛ продуцируются с молоком, являющимся обычным компонентом, например, детского питания, в результате чего нет необходимости в интенсивной очистке при использовании рекомбинантных человечьих ССЖЛ/КСЛ в качестве питательной добавки к продуктам на основе молока. Более того, продуцирование в высшем организме, например отличном от человека млекопитающем, обычно ведет к точному процессингу белка млекопитающего, например пострансляционному процессингу, обсуждаемому выше, и надлежащей укладке белка. Кроме того, по существу чистые человечьи ССЖЛ/КСЛ могут быть получены в больших количествах.
Соответственно, еще одним своим важным аспектом настоящее изобретение относится к экспрессирующей системе млекопитающего, включающей кодирующую человечьи ССЖЛ/КСЛ ДНК последовательность, встроенную в ген, кодирующий молочный белок отличного от человека млекопитающего, с образованием в результате гибридного гена, экспрессируемого в молочной железе взрослой самки млекопитающего, принявшей гибридный ген.
В качестве ДНК последовательности, кодирующей человечьи ССЖЛ/КСЛ, рекомендуется ДНК последовательность, показанная в перечне последовательностей как последовательность N 1, или геномный человечий CEL-ген, или его аналог.
Молочная железа в качестве экспрессирующей ткани и гены, кодирующие молочные белки, как правило, считаются наиболее пригодными для применения с целью продуцирования гетерологичных белков в трансгенных, отличных от человека млекопитающих, поскольку молочные белки в естественных условиях продуцируются молочной железой на высоком уровне. Кроме того, молоко легко собирается и доступно в больших количествах. В этой связи, применение генов молочного белка в продуцировании рекомбинантных человечьих ССЖЛ/КСЛ имеет еще и то преимущество, что ССЖЛ/КСЛ продуцируются в условиях, аналогичных условиям их естественного продуцирования, с точки зрения регулирования экспрессии и места продуцирования (молочная железа).
В контексте настоящей заявки термин "гибридный ген" относится к ДНК последовательности, содержащей, с одной стороны, ДНК последовательность, кодирующую вышеохарактеризованные человечьи ССЖЛ/КСЛ, а с другой стороны, ДНК последовательность гена молочного белка, способного поддерживать экспрессию продукта гибридного гена. Термин "ген, кодирующий молочный белок", относится ко всему гену, а также к его последовательности, способных поддерживать и направлять экспрессию гибридного гена в представляющей интерес ткани, то есть в молочной железе. Обычно такая последовательность - это последовательность, включающая по меньшей мере одну или несколько областей промотора, участок начала транскрипции, некодирующие 3'- и 5'-области и структуральные последовательности. ДНК последовательность, кодирующая человечьи ССЖЛ/КСЛ, предпочтительно по существу не содержит прокариотных последовательностей, например векторных последовательностей, которые могут быть связаны с ДНК последовательностью после, например, ее клонирования.
Гибридный ген рекомендуют создавать вставкой in vitro известными специалистам методами в ген молочного белка ДНК последовательности, кодирующей человечьи ССЖЛ/КСЛ. Или же ДНК последовательность, кодирующая человечьи ССЖЛ/КСЛ, может быть вставлена in vivo гомологичной рекомбинацией.
Как правило, ДНК последовательность, кодирующая человечьи ССЖЛ/КСЛ, вставляется в один их первых экзонов выбранного гена молочного белка или в его эффективную последовательность, содержащую первые экзоны и предпочтительно значительную часть фланкирующей 5'-области, которые, как полагают, играют важную регулирующую роль.
Рекомендуется, чтобы гибридный ген включал последовательность, кодирующую сигнальный пептид, необходимый для секретирования надлежащим образом продукта гибридного гена в молочную железу. Сигнальный пептид - это, как правило, пептид, находящийся обычно в рассматриваемом гене молочного белка, или пептид, связанный с ДНК последовательностью, кодирующей человечьи ССЖЛ/КСЛ. Однако также возможны и другие сигнальные последовательности, способные поддерживать секрецию продукта гибридного гена в молочную железу. Разумеется, различные элементы гибридного гена должны быть слиты путем, позволяющим правильную обработку и экспрессию генного продукта. Так, ДНК последовательность, кодирующая выбранный сигнальный пептид, как правило, должна быть точно слита с N-концевой частью ДНК последовательности, кодирующей человечьи ССЖЛ/КСЛ. В гибридном гене ДНК последовательность, кодирующая человечьи ССЖЛ/КСЛ, обычно включает их терминирующий кодон, но без их участка сигнала отщепления и полиаденилирования. В нисходящем направлении от ДНК последовательности, кодирующей человечьи ССЖЛ/КСЛ, обычно сохраняются процессинговые последовательности мРНК гена молочного белка.
Считается, что целый ряд факторов отвечает за реальный уровень экспрессии конкретного гибридного гена. Эффективность промотора, а также других вышеупомянутых регуляторных последовательностей, участок интеграции экспрессионной системы в геноме млекопитающего, участок интеграции ДНК последовательности, кодирующей человечьи ССЖЛ/КСЛ, в кодирующем молочный белок гене, элементы, создающие послетранскрипционную регуляцию, и другие аналогичные факторы могут иметь жизненно важное значение для достигаемого уровня экспрессии. Основываясь на знании разнообразных факторов, влияющих на уровень экспрессии гибридного гена, специалист способен создать экспрессионную систему, применимую для целей настоящего изобретения.
Молочной железой секретируется целый ряд различных молочных белков. Существует основная группа молочных белков, а именно казеины и сывороточные белки. Состав молока от животных различного вида меняется качественно, а также количественно по содержанию указанных белков. Большинство отличных от человека млекопитающих продуцируют казеин 3 различных типов, а именно α- казеин, β- казеин и κ- казеин. К наиболее обычным коровьим сывороточным белкам относятся α- лактальбумин и β- лактальбумин. Состав молока различного происхождения подробно раскрывается в работе Clark и др. (1987).
Применяемый ген молочного белка может происходить от того же вида, что и вид, в который вводится экспрессионная система, или может происходить от других видов животных. В этой связи показано, что регуляторные элементы, направляющие экспрессию гена на молочную железу, функциональны в границах вида, что может быть связано с возможным общим предшественником (Hennighausen и др., 1990).
Примеры приемлемых генов, кодирующих молочный белок, или его эффективных субпоследовательностей, предназначенных для конструирования экспрессирующей системы изобретения, как правило, могут быть обнаружены среди генов сывороточных белков разнообразных млекопитающих, например гена сывороточного кислотного белка (WAP-ген), предпочтительно мышиного происхождения, и гена β- лактоглобулина, предпочтительно овечьего происхождения. Кроме того, для трансгенного продуцирования человечьих ССЖЛ/КСЛ могут оказаться пригодны гены казеина различного происхождения, например коровий α S1-казеин и кроличий β- казеин. Рекомендуемым для применения в настоящем изобретении геном является мышиный WAP-ген, поскольку обнаружено, что этот ген способен обеспечить на высоком уровне ряд чужеродных человечьих белков в молоке различных трансгенных животных (Hennighausen и др., 1990).
Другой рекомендуемой последовательностью, связанной с экспрессирующей системой изобретения, является так называемая стабилизирующая экспрессирующая последовательность, способная поддерживать высокий уровень экспрессии. Существуют сильные доказательства того, что такая стабилизирующая последовательность находится вблизи и в восходящем направлении от генов молочных белков.
ДНК последовательность, кодирующая человечьи ССЖЛ/КСЛ и подлежащая вставке в экспрессирующую систему изобретения, может быть геномного или искусственного происхождения, или их комбинацией. Некоторые экспрессионные системы для достижения удовлетворительной экспрессии, как обнаружено, требуют присутствия интронов и иных регуляторных областей (Hennighausen и др., 1990). В некоторых случаях может оказаться желательным введение в векторные конструкции в качестве кодирующего полипептид элемента геномных структур, а не элементов кДНК (Brinster и др.). Применение интронных и экзонных структур может привести к более высокому уровню мРНК в устойчивом состоянии, чем в случае применения векторов на основе кДНК.
В еще одном своем аспекте изобретение относится к гибридному гену, включающему ДНК последовательность, кодирующую человечьи ССЖЛ/КСЛ и вставленную в ген, кодирующий молочный белок отличного от человека млекопитающего, при этом ДНК последовательность вставляется в ген молочного белка таким образом, что способна экспрессироваться в молочной железе млекопитающего, принявшего гибридный ген. Гибридный ген и его составляющие подробно обсуждены выше. Гибридный ген является важным промежуточным элементом в конструировании экспрессирующей системы изобретения, раскрытой выше.
В другом своем аспекте изобретение относится к клетке отличного от человека млекопитающего, принявшей вышеохарактеризованную экспрессирующую систему. Рекомендуемой клеткой млекопитающего является клетка эмбриона или пронуклеус. Экспрессирующую систему вводят соответствующим образом в клетку млекопитающего применением способа, разъясняемого далее и иллюстрируемого нижеследующими специальными примерами.
В еще одном своем важном аспекте изобретение относится к способу создания трансгенного, отличного от человека млекопитающего, способного экспрессировать человечьи ССЖЛ/КСЛ. Способ состоит в инъецировании вышеопределенной экспрессионной системы изобретения в оплодотворенную яйцеклетку или клетку эмбриона в зародышевую линию млекопитающего с последующим развитием полученного инъецированного оплодотворенного яйца или эмбриона во взрослую самку млекопитающего.
Введение экспрессионной системы в зародышевую линию млекопитающего может быть осуществлено применением любой приемлемой методики, приведенной, например, в издание "Манипуляции с эмбрионом мыши". Лабораторное руководство, Коулд Спринг Харбор Лаборетори Пресс, 1986. К примеру, несколько сот молекул экспрессионнной системы могут быть непосредственно инъецированы в оплодотворенную яйцеклетку, например в одну ее оплодотворенную клетку или ее пронуклеус, или в эмбрион выбранного млекопитающего, после чего микроинъецированные яйцеклетки могут быть перенесены в яйцеводы псевдобеременных приемных матерей с последующим развитием эмбрионов. Как правило, не все инъецированные яйцеклетки развиваются во взрослые самки, экспрессирующие человечьи ССЖЛ/КСЛ. Так, около половины животных со статистической точки зрения будут самцами, от которых, однако, в следующих поколениях могут быть получены и самки.
После интегрирования в зародышевую линию ДНК последовательность, кодирующая человечьи ССЖЛ/КСЛ, может быть экспрессирована на высоком уровне с продуцированием правильно обработанных и функциональных человечьих ССЖЛ/КСЛ в стабильных линиях рассматриваемого млекопитающего.
Особый интерес представляет способ создания трансгенных, отличных от человека млекопитающих, способных экспрессировать человечьи ССЖЛ/КСЛ и по существу не способных экспрессировать собственные ССЖЛ/КСЛ. Способ состоит в:
(a) уничтожении способности млекопитающим экспрессировать ССЖЛ/КСЛ с тем, чтобы по существу никакой экспрессии ССЖЛ/КСЛ не происходило, и введении вышеохарактеризованной экспрессионной системы изобретения или ДНК последовательности, кодирующей человечьи ССЖЛ/КСЛ, в зародышевую линию млекопитающего таким образом, что в млекопитающем экспрессируются человечьи ССЖЛ/КСЛ, и/или
(b) замене CEL-гена млекопитающего или части гена на вышеохарактеризованную экспрессионную систему изобретения или на ДНК последовательность, кодирующую человечьи ССЖЛ/КСЛ.
Способность млекопитающего экспрессировать ССЖЛ/КСЛ обычно уничтожают созданием мутаций в ДНК последовательности, отвечающей за экспрессию ССЖЛ/КСЛ. Такие мутации включают мутации, при которых ДНК последовательность выводится из рамки, вводится терминирующий кодон или происходит делеция одного или нескольких нуклеотидов ДНК последовательности.
Замена CEL-гена млекопитающего или его части на вышеохарактеризованную экспрессионную систему или на ДНК последовательность, кодирующую человечьи ССЖЛ/КСЛ, может быть осуществлена применением хорошо известных принципов гомологичной рекомбинации.
В еще одном своем аспекте изобретение относится к трансгенному, отличному от человека млекопитающему, созданному вышеприведенным способом.
Хотя трансгенное, отличное от человека млекопитающее изобретения в самом широком его аспекте не ограничено каким-либо конкретным типом млекопитающего, тем не менее млекопитающее обычно выбирают из группы, включающей мышь, крысу, кролика, овцу, свинью, козу и крупный рогатый скот. Для широкомасштабного продуцирования человечьих ССЖЛ/КСЛ обычно рекомендуются более крупные животные, например овцы, козы, свиньи и крупный рогатый скот, что связано с их способностью давать в больших количествах молоко. Однако интерес могут также представлять мыши, кролики и крысы вследствие большей простоты манипуляций с этими животными и более быстром получении результатов для трансгенных животных, чем в случае, например, крупного рогатого скота.
Объемом изобретения охватывается также потомство вышеохарактеризованных трансгенных животных, способное продуцировать человечьи ССЖЛ/КСЛ.
В еще одном своем аспекте настоящее изобретение включает молоко от млекопитающего (но не человека), содержащее рекомбинантные человечьи ССЖЛ/КСЛ.
И еще в одном своем аспекте настоящее изобретение относится к детскому питанию, содержащему рекомбинантные человечьи ССЖЛ/КСЛ, в частности вышеохарактеризованный полипептид изобретения. Детское питание может быть приготовлено добавлением к обычным компонентам детского питания рекомбинантных человечьих ССЖЛ/КСЛ или полипептида в очищенной или частично очищенной форме. Однако, как правило, рекомендуется изготовлять детское питание из вышеохарактеризованного молока изобретения, особенно, если это коровье молоко. Детское питание может быть изготовлено обычными методами и может содержать любые необходимые добавки, например минеральные вещества, витамины и т.д.
Пример 1. Геномная организация, анализ последовательности и локализация CEL-гена в хромосоме.
Если нет особых указаний, применялись стандартные методики молекулярной биологии (Maniatis и др., 1982; Ausubel и др., 1987; Sambrook и др., 1989).
Выделение геномных рекомбинантов
Гибридизацией в бляшках отобраны две различные геномные фаговые библиотеки человека: λ DASH (Клонтех Лабораториз Инк., Паоло Альто, Ка., США) и λ EMBL-3 SP6/T7 (Стратаген, Ла Джолла, Ка., США). В качестве зондов при гибридизации применялись различные субклонированные фрагменты рестрикции кДНК (Nilsson и др., 1990). Зонды метились методом введения метки в олигонулеотиды с применением [α-32P] дЦТФ (Feinberg и др., 1983).
Картирование, субклонирование и секвенирование геномных клонов
Положительные клоны гидролизуют в присутствии различных ферментов рестрикции, подвергают электрофорезу на 1%-ных агарозных гелях и затем переносят в вакууме (Фармация ЛКБ БТГ, Уппсала, Швеция) на найлоновую мембрану. Мембрану подвергают гибридизации с разнообразными кДНК зондами. Фрагменты рестрикции, гибридизованные с зондами, выделяют применением метода изтахофореза (Ofverstedt и др., 1984). Более мелкие фрагменты (<800 п.о.) непосредственно вставляют в вектора M13mp18, M13mp19, M13BM20 или M13BM21 и секвенируют применением в качестве бактерии-хозяина E. coli TG1, а более крупные фрагменты субклонируют в векторы pTZ18R или pTZ19R применением в качестве бактерии-хозяина E. coli DH5 α и дополнительно гидролизуют. (Плазмиды pS309, pS310 и pS451, применяемые в нижеследующем примере 2, получают соответственно). Некоторые из выделенных фрагментов применяются также в качестве зондов при гибридизации. Все последовательности нуклеотидов определены методом терминации дидезоксицепи (Sanger и др., 1977) применением фермента Кленова и M13 универсального секвенирующего праймера специфичных олигонуклеотидов. Информация о последовательности получена на основе авторадиограмм применением программного обеспечения MS-EdSeg по методике Sjoberg и др. (1989). Последовательности анализируют с помощью программ, входящих в пакет UWGCG программного обеспечения (Devereux и др., 1984).
Удлинение праймера
Полную РНК выделяют из человечьей поджелудочной железы и жировой ткани по методике с применением гуанидинийизотиоцианата CsCl (Chirgwin и др., 1979). Удлинение праймера осуществляют по методике (Ausubel и др., 1987) применением полной РНК и десенсибилизированного 26-тимерного олигонуклеотида (5'-AGGTGAGGCCCAACACAACCAGTTGC-3'), положения 33-58. Гибридизацию праймера с 20 мкг полной РНК проводят при 30oC в течение ночи в 30 мкл 0,9 М NaCl, 0,15 М Хепеса (pH 7,5) и 0,3 М ЭДТК. После реакции удлинения в присутствии обратной транскриптазы продукты удлинения анализируют электрофорезом через 6%-ный денатурированный полиакриламидный гель.
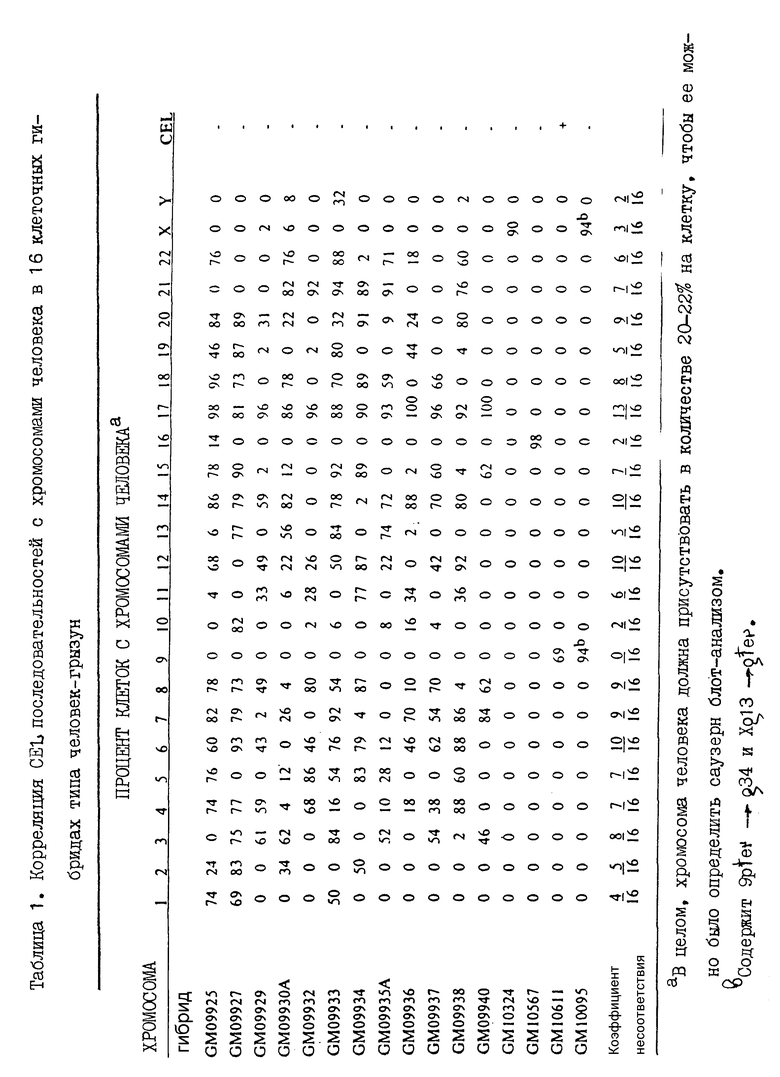
Соматические клеточные гибриды
Для хромосомного анализа CEL-гена использована ДНК из 16 соматических клеточных гибридных линий типа человек-грызун, полученных от NIGMS Хьюмен Генетик Мутант Селл Репозитори (Кориэлл Институт фор Медикал Рисерч, Камдэм, НДж). Человек-мышиные соматические клеточные гибриды GM09925-GM09950 происходят от слияния фибробластов мужского плода человека (IMR-91) с мышиной клеточной линией с дефицитом тимидин-киназы B-82 (Taggart и др., 1985; Mohandas и др., 1986). Гибриды GM10324 и GM102860 включают мышиную клеточную линию A9 с дефицитом HPRT и APRT (Callen и др., 1986), в то время как гибрид GM10611 получен микроклеточным слиянием клеточной линии GM07890 лимфобластов человека, инфицированных ретровирусным вектором SP-1, с клеточной линией UV-135 яичников китайского хомячка (Warburton и др., 1990). Гибрид GM10095 происходит от слияния лимфоцитов самки с балансированным 46, X, t (X; 9) (q13; 34) кариотипом с клеточной линией CHW1102 яичников китайского хомячка (Mohandas и др. , 1979). Содержание хромосом человека в гибридных линиях, определенное цитогенетическим анализом, а также саузерн-блот анализом и анализом гибридизации in vitro, приведено в табл. 1. Высокомолекулярные ДНК, выделенные из исходных клеточных линий мыши, китайского хомячка и человека и 16 гибридных клеточных линий, гидролизуют в присутствии EcoRI, фракционируют на 0,8 агарозных гелях и переносят на найлоновый фильтр. Методом мечения олигонуклеотидов (Feinberg и Vogelstein, 1983) получены [α-32P] дЦТФ- меченный CEL кДНК (кДНК полной длины), который гибридизован с содержимым фильтров. Фильтры промывают 60 мин каждый раз при 65oC в 6 x SSC /0,5% НДС и в 2 x SSC/0,5% НДС.
Полимеразная цепная реакция
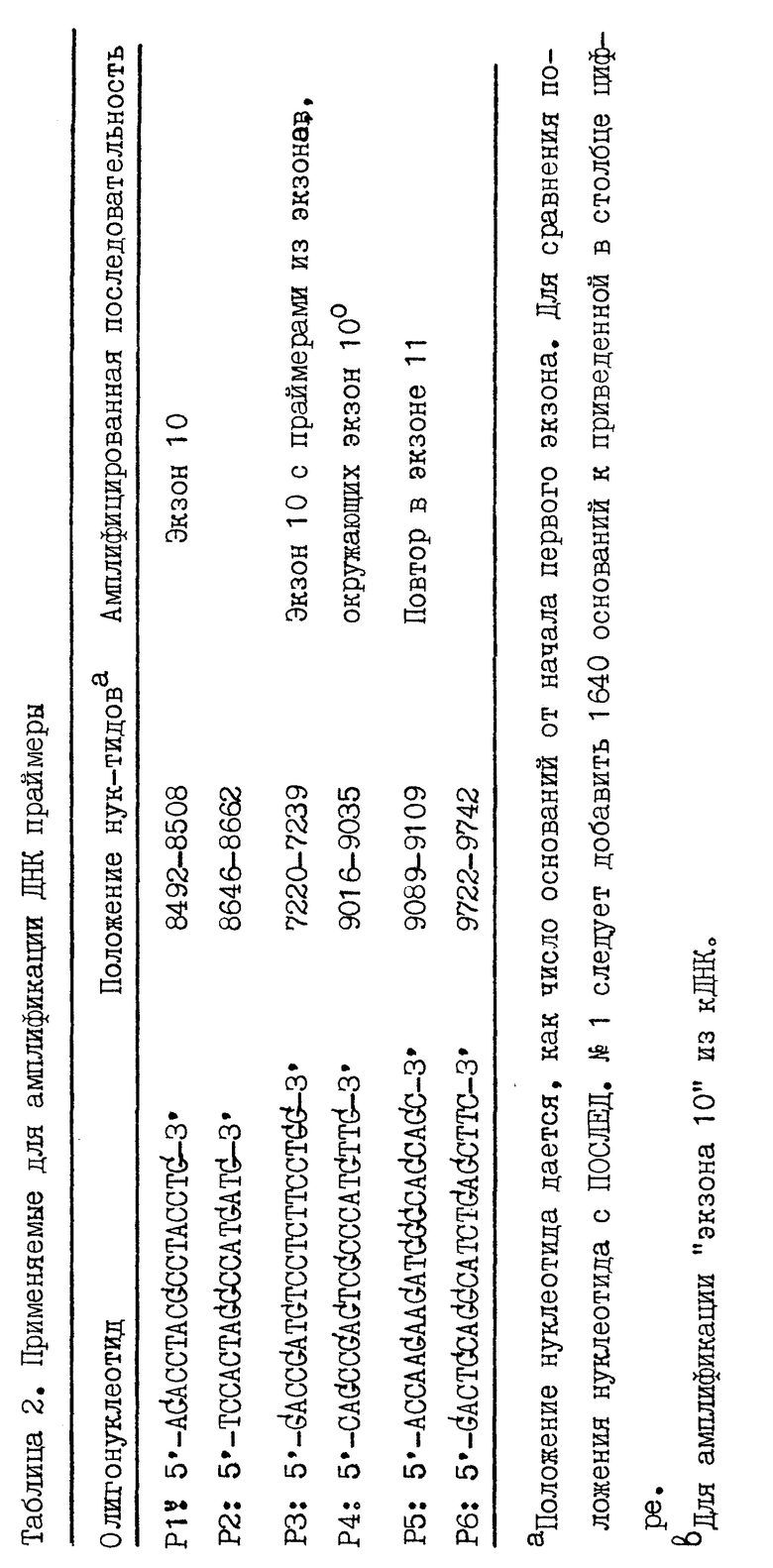
Полную человечью геномную ДНК, выделенную из лейкоцитов, ДНК из соматических клеточных гидридов и из некоторых положительных рекомбинантов, и полную РНК из лактирующей молочной железы человека и поджелудочной железы человека амплифицируют по экзону 10 и экзону 11. Применяют 2 мкг ДНК. Применяемые праймеры перечислены в табл. 2. Осуществляют тридцать циклов ПЦР в объеме 100 мкл [10 мМ Трис-HCl (pH 8,3), 50 мМ KCl, 1,5 мМ MgCl2, 200 мкМ каждого дНТФ, 100 мкг/мл желатины, 100 пмоль каждого праймера, 1,5 E Tag ДНК полимеразы (Перкин-Эльмер Цетус, Норуолк, СТ, США)] и температуре отжига 55oC для всех пар праймеров. РНК последовательность амплифицируют применением комбинированной комплементарной ДНК (кДНК) и методики ПЦР. кДНК синтезируют из 10 мкг полной РНК в 40 мл раствора, содержащего 50 мМ Трис-HCl (pH 8,3), 50 мМ KCl, 10 мМ MgCl2, 10 мкг/мл БСА, 1 мМ каждого дНТФ, 500 нг олиго(dt)12-18, 40 E ингибитора рибонуклеазы и 200 E обратной транскриптазы (MoMuLV) (БРЛ, Бетстеда Рисерч Лабораториз, Н-Й., США) выдерживанием 30 мин при 42oC. кДНК осаждают и вновь суспендируют в 25 мкл H2O, 2 мкл суспензии амплифицируют вышеприведенным способом. Амплифицированные фрагменты анализируют на 2%-ном агарозном геле. Некоторые фрагменты дополнительно субклонируют и секвенируют.
Генная структура человечьего CEL-гена
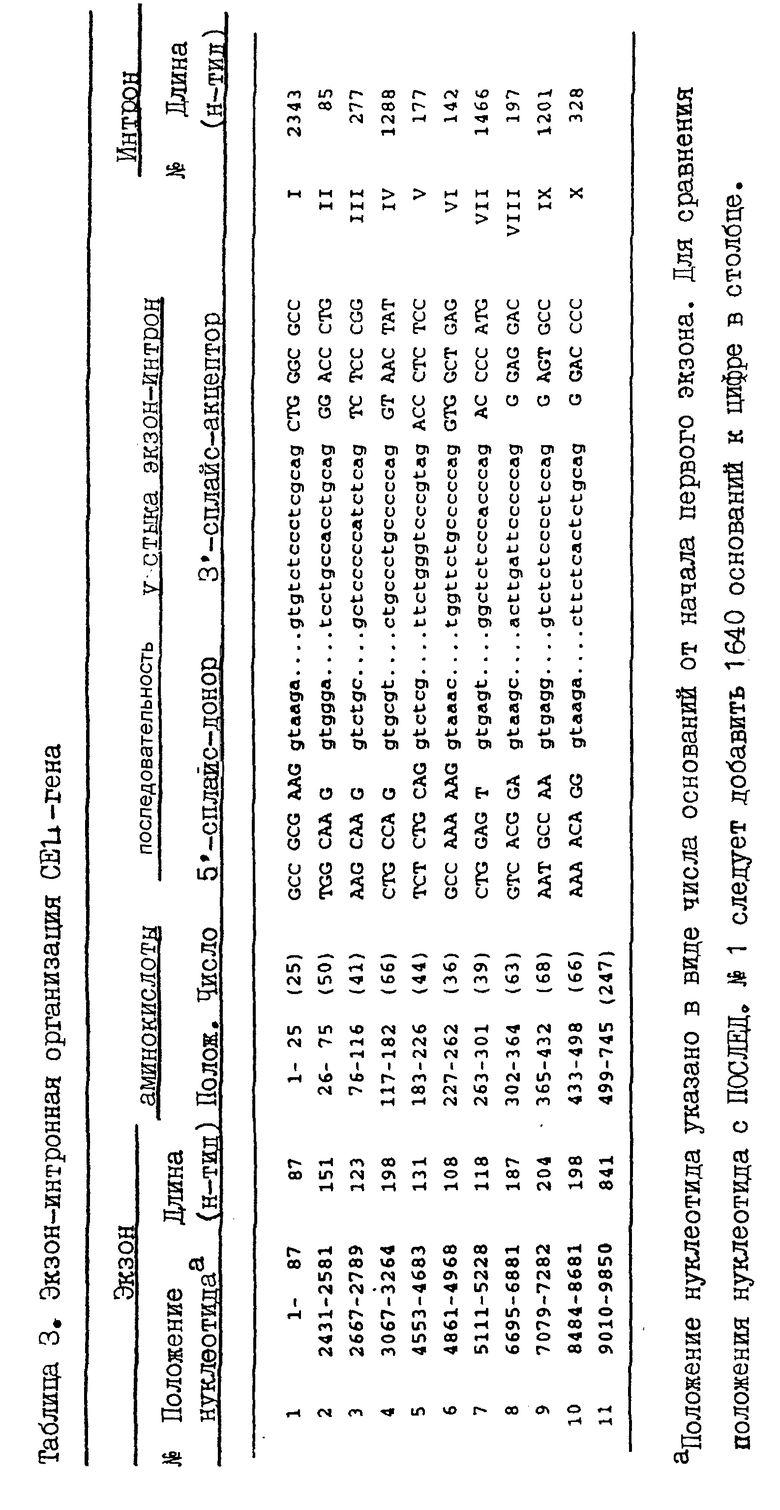
В каждой геномной библиотеке было скринировано 106 рекомбинантов и скрининги давали несколько положительных клонов, которые были изолированы и картированы. Два клона, обозначенные λ BSSL1 и λ BSSL5A, подвергнуты дополнительному анализу. Гидролизом в присутствии нескольких ферментов рестрикции, саузерн-блоттингом с последующей гибридизацией с кДНК зондами показано, что λ BSSL 5A клон охватывает CEL-ген целиком и что λ BSSL1 клон охватывает 5'-половину и примерно 10 т.п.о. 5'-фланкирующей области (фиг. 1). Совместно эти два клона охватывают 25 т.п.о. генома человека.
После субклонирования и гидролиза ферментами рестрикции получены приемлемые для секвенирования фрагменты, позволяющие определить последовательность CEL-гена полностью, включая 1640 п.о. 5'-фланкирующей области и 41 п. о. 3'-фланкирующей области. На основании этих данных установлено, что CEL-ген человека (Последовательность N1) охватывает область в 9850 п.о., содержащую 11 экзонов, разделяемых 10 интронами (фиг. 1). Это означает, что экзоны и особенно интроны сравнительно малы. В самом деле, размер экзонов 1 - 10 колеблется в интервале 97 - 204 п.о., а длина экзона 11 - 841 п.с. Размер интронов колеблется в пределах 85 - 2343 п.о. Как видно из табл. 3, все границы экзон/интрон подчиняются AG/GT правилу и хорошо согласуются с консенсусной последовательностью, предложенной Mount и др. (1982). При сравнении кодирующей части CEL-гена с кДНК (Nilsson и др., 1990) обнаружено единственное отличие в нуклеотидной последовательности: вторым нуклеотидом в экзоне 1 является C, которому в кДНК последовательности соответствует T. Поскольку это положение расположено на расстоянии в 10 нуклеотидов в восходящем направлении от кодона АТС начала трансляции, указанное отличие не влияет на аминокислотную последовательность.
В секвенированной области присутствуют семь членов Alu класса повторяющихся ДНК элементов, обозначенные Alu1-Alu7 (5'-3') (фиг. 1), один из которых находится в 5'-фланкирующей области, а шесть остальных - в пределах CEL-гена.
Сайты инициирования транскрипции и 5'-фланкирующая область
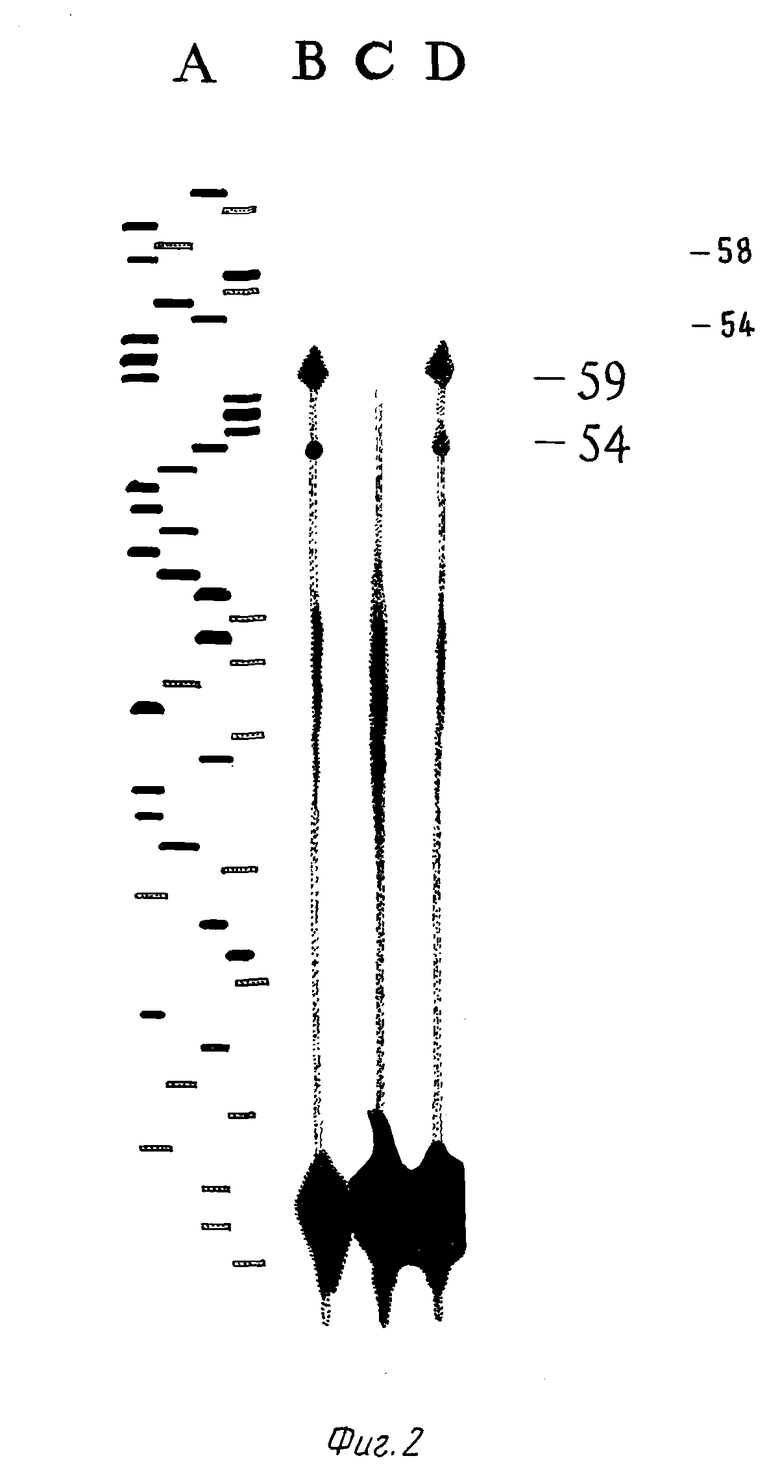
Для картирования сайта (ов) инициирования транскрипции CEL-гена человека осуществлен анализ с удлинением праймера, в котором использована полная РНК из поджелудочной железы, лактирующей молочной железы и жировой ткани человека. Результаты анализа указывают на основной сайт начала транскрипции, расположенный в 12 п.о., и меньший сайт начала, расположенный на расстоянии в 8 п.о. в восходящем направлении от инициатора метионина. Сайты начала транскрипции одинаковы как для поджелудочной железы, так и для лактирующей молочной железы, в то время как жировой ткани никакого сигнала не было обнаружено (фиг. 2). Секвенированная область включает 1640 нуклеотидов 5'-фланкирующей ДНК. На основе схожести последовательностей на расстоянии в 30 нуклеотидов в восходящем направлении от сайта инициации транскрипции обнаружена подобная рамке Хогнесса последовательность CATAAAT (фиг. 4). Никаких свидетельств присутствия в этой области структур типа рамки Хогнесса или CC блоков не обнаружено.
5'-фланкирующая последовательность подвергнута в обоих нитях компьютерному скринингу на нуклеотидные последовательности, известные как последовательности транскрипционного фактора связывания в других специфичных для молочной железы и поджелудочной железы генах. Обнаружено несколько предполагаемых последовательностей (фиг. 4).
Хромосомная локализация CEL-гена
В контрольной ДНК человека применением CEL кДНК зонда обнаружены четыре EcoRI фрагмента примерно в 13 т.п.о., 10 т.п.о., 2,2 т.п.о. и 2 т.п.о., в то время как в контрольных ДНК мыши и хомячка обнаружены единственные фрагменты соответственно в 25 т.п.о. и 8,6 т.п.о. Присутствие в гибридных клонах последовательностей CEL-гена человека коррелирует только с присутствием хромосомы 9 человека (табл. 1). Всего лишь один из 16 проанализированных гибридов оказался положительным на присутствие CEL-гена, и данный гибрид содержит хромосому 9 в качестве единственной хромосомы человека. Никакого несоответствия в локализации этой хромосомы не обнаружено, в то время как в локализации любой другой хромосомы выявлено по меньшей мере два несоответствия (табл. 1). Для дополнительной сублокализации CEL-гена нами использован гибрид человека - китайский хомячок (GM 10095), сохранивший der (9) транслокационную хромосому (9pter ---> 9q34 : Xq13 ---> Xqter) в качестве единственной ДНК человека. Саузерн-блоттингом нам не удалось обнаружить в этом гибриде какие-либо последовательности CEL-гена, что указывает на присутствие остатков CEL-гена в пределах 9q34-qter области.
Пример 2. Конструирование векторов экспрессии.
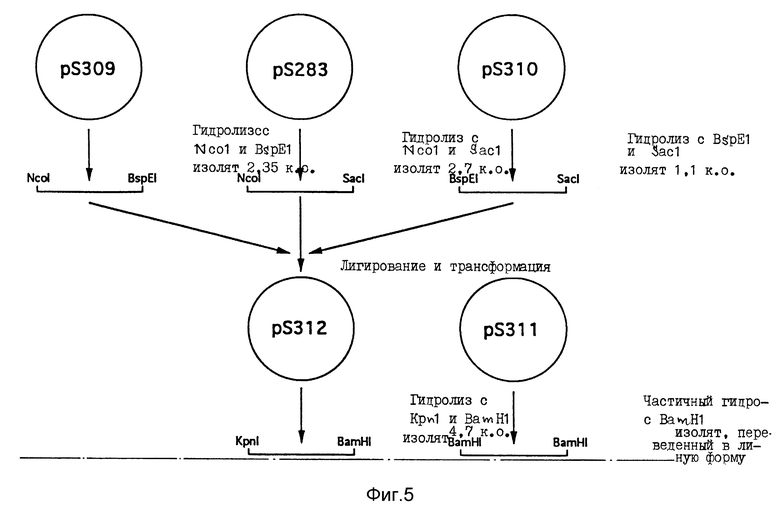
Для конструирования вектора экспрессии для продуцирования рекомбинантной человечьей КСЛ в молоке от трансгенных животных применена следующая стратегия (фиг. 5).
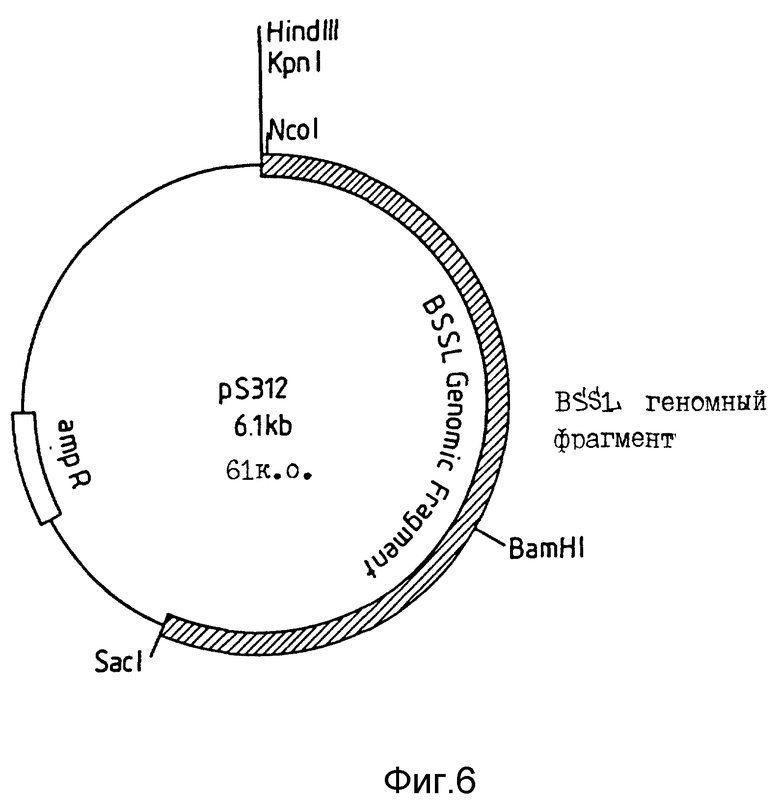
По вышеприведенным методикам получены три основанные на pTZ плазмиды (Фармация, Уппсала, Швеция), содержащие различные части CEL-гена человека, а именно: pS309, pS310 и pS311. Плазмида pS 309 содержит SphI фрагмент, охватывающий CEL-ген от 5'-нетранскрибированной области до части четвертого интрона. Плазмида pS310 содержит SacI фрагмент, охватывающий последовательность CEL-гена от части первого интрона до части шестого интрона. Третья плазмида pS311 содержит BamHI фрагмент, охватывающий вариант CEL-гена от основной части пятого интрона до остальной части структуры интрон/экзон. В этой плазмиде повторяющаяся последовательность экзона 11, обычно кодирующая 16 повторов, мутирована с кодированием усеченного варианта, имеющего 9 повторов.
Для слияния геномных последовательностей использована другая плазмида (pS283), содержащая часть CEL кДНК человека, клонированную в плазмиду pUC19 по HindIII и SacI сайтам. Плазмида pS283 использована также для доступа к удобному участку рестрикции ферментом (Kpn1), расположенному в 5'-нетранслируемой лидерной последовательности CEL-гена. Затем плазмиду pS283 гидролизуют в присутствии Ncol и SacI с последующим выделением фрагмента в 2,7 т.п. о. Гидролизом плазмиды pS309 в присутствии Ncol и BspEI выделен фрагмент в 2,3 т.п.о., содержащий 5'-часть CEL-гена. Гидролизом плазмиды pS310 в присутствии BspEI и SacI выделен фрагмент в 2,7 т.п.о., содержащий часть средней области CEL-гена. Эти три фрагмента лигируют и трансформируют в компетентном ТС2 штамме E. coli и трансформанты выделяют отбором по ампициллину. Из ряда трансформантов получают плазмиды и одну из плазмид, названную pS312 и содержащую целевую конструкцию (фиг. 6), применяют в дальнейших экспериментах.
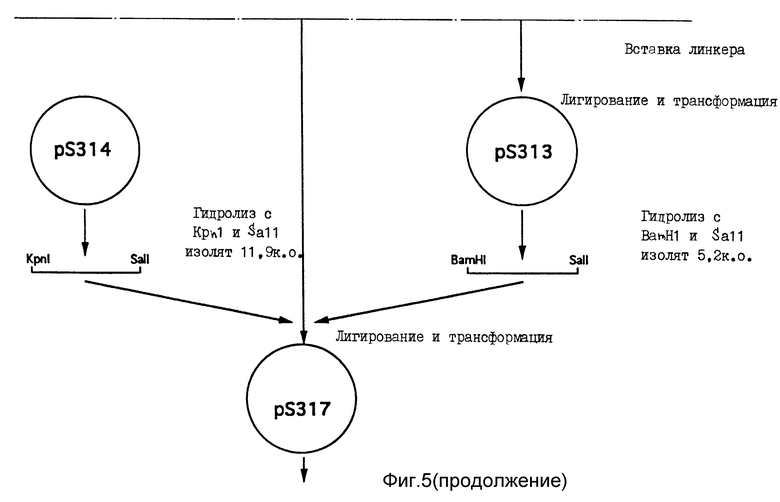
Для получения модификации pS311, в которой для облегчения последующего клонирования расположенный в нисходящем направлении от терминирующего кодона BamHI сайт превращен в Sall сайт, используют следующий способ. Частичным гидролизом в присутствии BamHI pS311 переводят в линейную форму. Выделяют фрагмент в линейной форме и в него вставляют синтетический ДНК линкер (5'-GATCGTCGAC-3'), превращающий BamHI сайт в SalI сайт с разрушением в результате BamHI сайта. Поскольку существуют два потенциальных положения для интеграции синтетического линкера, полученные плазмиды анализируют расщеплением в присутствии фермента рестрикции. Выделенную плазмиду со вставкой линкера в целевом положении, расположенном в нисходящем направлении от экзона 11, обозначают как pS313.
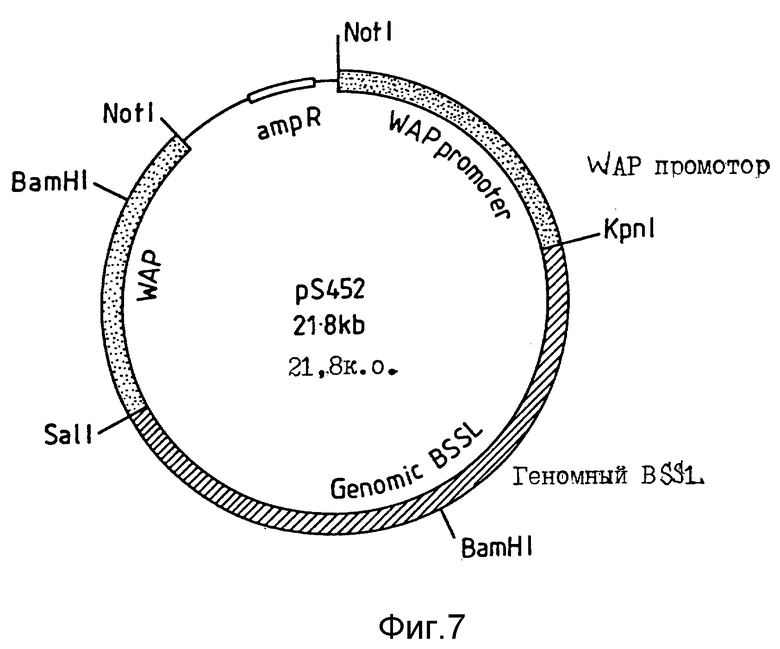
Для получения вектора экспрессии, принявшего CEL геномные последовательности и кодирующего усеченный CEL вариант, применяют плазмиду pS314, предназначенную для поддержания постадийно- и тканево-специфичной экспрессии в клетках молочной железы в период лактации. Плазмида pS314 содержит геномный фрагмент из мышиного гена сывороточного молочного белка (WAP-гена) (Campbell и др. , 1984), клонированного в виде NotI фрагмента. Геномный фрагмент содержит примерно 4,5 т.п.о. восходящих регуляторных последовательностей (ВРП), полную транскрибированную экзон-интронную область и 3 т.п. о. последовательности последнего экзона в нисходящем направлении. Единственный KpnI сайт расположен в первом экзоне на расстоянии в 24 п.о. в восходящем направлении от природного WAP кодона инициирования трансляции. Другим уникальным сайтом рестрикции ферментом является SalI сайт, расположенный в экзоне 3. В плазмиде pS314 этот SalI сайт разрушают гидролизом, наполняют применением фрагмента Кленова и повторным лигированием. Взамен вводят новый SalI сайт непосредственно в нисходящем направлении от KpnI сайта в экзоне 1. Вставку осуществляют гидролизом в присутствии KpnI и введением в указанное положение ренатурированных синтетических олигомеров SYM 2401 (5'-CGTCGACGTAC-3') и SYM 2402 (5'-GTCGACGGTAC-3') (фиг. 8). CEL геномную последовательность человека вставляют между указанными сайтами (KpnI и SalI) применением следующей стратегии. Во-первых, pS314 гидролизуют в присутствии KpnI и SalI и фрагмент, представляющий отщепленную плазмиду, выделяют электрофорезом. Во-вторых, плазмиду pS312 гидрозируют в присутствии KpnI и BamHI и выделяют фрагмент в примерно 4,7 т.п.о., представляющий 5'-часть CEL-гена человека. В-третьих, плазмиду pS313 гидролизуют в присутствии BamHI и SalI и выделяют 3'-часть CEL-гена человека. Три полученных фрагмента лигируют, трансформируют в компетентном штамме E. coli и трансформанты выделяют после отбора по ампициллину. Из нескольких трансформантов получают плазмиды, которые тщательно анализируют картированием ферментами рестрикции и секвенс-анализом. Выявлена одна плазмида, представляющая целевой вектор экспрессии и названная pS317.
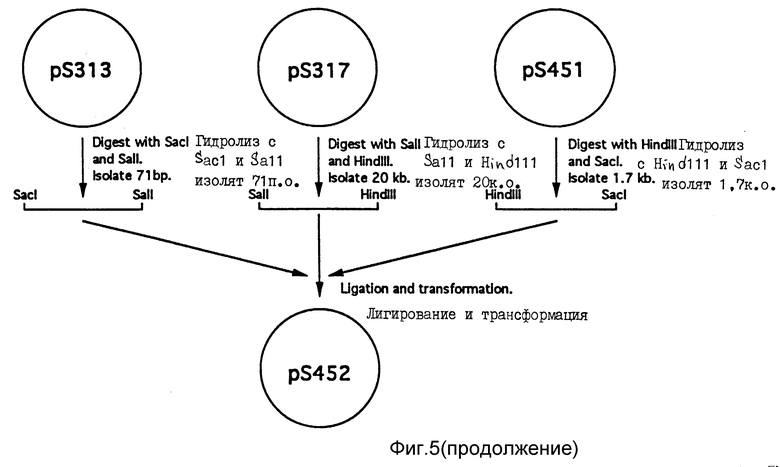
Для конструирования вектора экспрессии геномного CEL, кодирующего CEL полной длины, плазмиду pS317 модифицируют следующим образом (фиг. 5). Во-вторых, плазмиду pTZ18R (Фармация), содержащую BamHI фрагмент в 5,2 т.п.о. CEL-гена человека, простирающийся от пятого интрона в нисходящем направлении одиннадцатого экзона (pS451), гидролизуют в присутствии HindIII и SacI. В результате гидролиза получают фрагмент в 1,7 т.п.о., простирающийся от HindIII сайта, расположенного в интроне 9, SacI сайта, расположенного в экзоне 11. Во-вторых, плазмиду pS313 гидролизуют в присутствии SacI и SaiI и выделяют фрагмент в 71 п.о., содержащий 3'-часть экзона 11 и созданный SalI сайт. В-третьих, из плазмиды pS317 выделяют остальную часть WAP/CEL рекомбинантного гена и плазмидные последовательности в виде SalI/HindIII фрагмента в 20 т.п.о. Три полученных фрагмента лигируют и трансформируют в бактерию. Из нескольких трансформантов получают плазмиды. Плазмиды гидролизуют в присутствии различных ферментов рестрикции и подвергают секвенс-анализу. Идентифицирована одна плазмида, содержащая целевой рекомбинантный ген. Этот конечный вектор экспрессии получил название pS452 (фиг. 7).
Для удаления прокариотных плазмидных последовательностей pS452 гидролизуют в присутствии NotI. Последующим электрофорезом на агарозном геле выделяют элемент рекомбинантного вектора, состоящий из мышиной WAP последовательности, фланкирующей человечий CEL геномный фрагмент. Перед инъецированием в эмбрионы мыши выделенный фрагмент подвергают дополнительной очистке применением электроэлюирования.
Рекомбинантный WAP/CEL-ген для экспрессии в молочной железе трансгенных животных показан на фиг. 8.
Депозиты
Следующие плазмиды депонированы согласно Будапештскому Соглашению в (Deutsche Sammlung von Mikroorganismen und Zellkulturen) (см. в конце текста).
Пример 3. Создание трансгенных животных.
По методике примера 2 из плазмиды pS452 выделяют NotI фрагмент. Полученный ДНК фрагмент содержит мышиный WAP промотор, связанный с геномной последовательностью, кодирующей человечьи ССЖЛ/КСЛ. Выделенный фрагмент в концентрации 3 нг/мкл инъецируют в пронуклеус 350 C57B1/6JxCBA/2J-f2 эмбрионов, полученных от донорной мыши, праймированной для сверховуляции 5 МЕ сывороточного гонадотропина жеребой кобылы. Мыши линии C57B1/6JxCBA/2J-f1 получены от Бомхольтгард Бридинг энд Рисерч Сентр ЛТД, Рай, Дания. После сбора эмбрионов из яйцевода их обработкой гиалуронидазой в среде М2 отделяют от клеток кумулуса (Hogan и др., 1986). После промывания эмбрионы переносят в среду М 16 (Hogan и др., 1986) и выдерживают в инкубаторе в атмосфере 5% CO2. Инъецирование осуществляют в микрокапле М2 под легким парафиновым маслом с помощью гидравлического микроманипулятора фирмы Наришиги и инверсионного микроскопа Никон, снабженного оптикой фирмы Номарски. После инъецирования выглядевшие здоровыми эмбрионы имплантируют внутрибрюшинно псевдобеременным C57B1/6JxCBA/2J-f1 реципиентам, получавшим 0,37 мл 2,5% авертина. Мыши с интегрированным трансгеном выявлены ПЦР анализом ДНК из образцов хвоста, отобранных биопсией через три недели после рождения животных. Положительные результаты подтверждены саузерн-блот анализом.
Пример 4. Экспрессия ССЖЛ/КСЛ в трансгенной мыши.
Трансгенную мышь идентифицируют анализом ДНК, полученной из образца отрезанного хвоста. Образцы ткани инкубируют с протеиназой K и экстрагируют смесью фенола с хоороформом. Выделенную ДНК используют в полимеразной цепной реакции с праймерами, амплифицирующими специфичные фрагменты при наличии введенной гетерологичной ЛНК, представляющей фрагмент вектора экспрессии. Животных также оценивают в тестах гибридизации ДНК с целью подтверждения данных ПЦР и выявления возможных перегруппировок, строения элементов интегрированного вектора и для получения информации о числе копий элементов интегрированного вектора.
В одной серии опытов анализу подвергнуто 18 мышей. Анализ осуществлен двумя способами и полученные результаты показывают, что одна из мышей несет гетерологичный ДНК векторный элемент, происходящий из pS452. Результаты ПЦР анализа и опытов с гибридизацией были идентичны (фиг. 9).
Мышь, для которой выявлено присутствие элемента векторной ДНК (животное-основатель), спаривают и F1 помет подвергают анализу на трансген теми же методами.
Самкам лактирующих животных инъецируют внутрибрюшинно 2 МЕ окситоцина и 10 мин спустя анестезируют введением внутрибрюшинно 0,4 мл 2,5 авертина. Устройство для сбор молока присоединяют к соску силиконовой трубочкой и молоко собирают в пробирки Эппендорфа на 1,5 мл острожным массажем молочной железы. Количество молока менялось в зависимости от дня сбора в пределах 0,1 - 0,5 мл на мышь и на сбор.
Анализ на присутствие рекомбинантных человеческих ССЖЛ/КСЛ проводят проведением НДС-ПАГЭ, переносом на нитроцеллюлозные мембраны и инкубированием с поликлональными антителами, созданными против нативных человечьих ССЖЛ/КСЛ. Полученные результаты указывают на экспрессию рекомбинантных человечьих ССЖЛ/КСЛ в молоке трансгенных мышей. Присутствие рекомбинантных человечьих ССЖЛ/КСЛ в молоке от трансгенных мышей показало на фиг. 10 в виде полосы при 116,5.
Созданы стабильные линии трансгенных животных.
Аналогичным путем могут быть созданы и другие трансгенные животные, например коровы или овцы, способные экспрессировать человечьи ССЖЛ/КСЛ.
REFERENCES ЛИТЕРАТУРА
Abouakil, N., Rogalska, E., Bonicel, J. & Lombardo, D. (1988): Biochim. Biophys. Acta 961, 299 - 308.
Ausubel, F.M., Brent, R.E., Moore, D.D., Smiyh, J.A., Seidman, J.G. and Struhl, K.: Current Protocols in Molecular Biology. (Wiley Interscience, New York 1987)
Baba, Т., Downs, D., Jackson, K.W., Tang, J. and Wang, C.S. (1991): Biochemistry 30, 500 - 510.
Beato, M. (1989): Cell 56, 335 - 344.
Bernback, S., Blackberg, L. & Hernell, O. (1990): J. Clin. Invest. 221 - 226.
Bjorksten,B., Burman, L.G., deChateau, P., Fredrikzon, B., Gothefors, L. & Hernell, O. (1980): Br. Med. J. 201, 267 - 272.
Blackberg, L., Angquist, K.A, & Hernell, O. (1987): FEBS Lett. 217, 37 - 41.
Biackberg, L. & Hernell, O. (1981): Eur. J. Biochem 116, 221 - 225.
Blackberg, L. Lombardo, D. , Hernell, O. , Guy, O. & Olivecrona, T. (1981): FEBS Lett. 136, 284 - 288.
Boulet, A. M. , Erwin, C.R. and Rutter, W.J. (1986): Proc. Natl. Acad. Sci. U.S.A. 83, 3599 - 3603.
Brinster, R.L., Alien, J.M., Behringer, R.R., Gelinas, R.E. & Palmiter, R.D. (1988): Proc. Natl. Acad. Sci. U.S.A. 85, 836 - 840.
Callen, D.F. (1986): Ann. Genet. 29, 235 - 239.
Campbell, S. M. , Rosen, J.M., Hennighausen, L.G., Strech-Jurk, U. and Sippel, A.E. (1984): Nucleic Acid Res. 12, 8685 - 8697.
Chirgwin, J.M., Przybyla, A.E., MacDonald, R.J. and Rutter, W.J. (1979): Biochemistry 18, 5294 - 5299.
Clark, A.J., Simons, P., Wilmut, 1. and Lahte, R. (1987): TIBTECH 5, 20 - 24.
Devereux, J., Haeberli, P. and Smithies. (1984): Nucleic Acids Res. 12, 387 - 395.
Feinberg, A. and Vogelstein, B. (1983): Anal. Biochem. 132, 6-13.
Hennighausen, L., Ruiz, L. & Wall, R. (1990): Current Opinion in Biotechnology 1, 74 - 78.
Hernell, O. & Biackberg, L. (1982): Pediatr. Res. 16, 882 - 885.
Hogan, В. , Constantini, F. and Lacy, E. (1986): Manipulating the mouse embryo. A Laboratory Manual. Cold Spring Harbor Laboratory Press.
Hui, D. and Kissel, J.A. (1990): Febs Lett. 276, 131 - 134.
Lombardo, D. , Guy, O. & Figarella, С. (1978): Biochim. Biophys. Acta 527, 142-149.
Maniatis, Т. , Fritsch, E.F. & Sambrook, J.: Molecular Cloning. A Laboratory Manual. (Cold Spring Harbor, NY, 1982)
Mohandas, Т., Sparkes, R.S., Sparkes, M.C., Shulkin, J.D., Toomey, K.E. and Funderburk, S.J. (1979): Am. J. Hum. Genet. 31, 586 - 600.
Mohandas, Т., Heinzmann, С., Sparkes, R.S. Wasmuth, J., Edwards, P. and Lusis, A.J. (1986): Somatic Cell. Mol. Genet. 12, 89 - 94.
Mount, S.M. (1982): Nucleic Acids Res. 10, 459 - 472.
Nilsson, J., Blackberg, L., Carlsson, P., Enerback, S., Hernell, O. and Bjursell, G. (1990): Eur. J. Biochem. 192, 543 - 550.
Qasba, M., and Safaya, S.K. (1984): Nature 308, 377 - 380.
Reue, К. , Zambaux, J., Wong, H., Lee, G., Leete, Т.Н., Ronk, M., Shively, J. E., Sternby, B., Borgstrom, B., Ameis, D. and Schotz, M.C. (1991): J. Lipid. Res. 32, 267 - 276.
Sambrook, J. , Fritsch, E.F. and Maniatis, Т.Е.: Molecular Cloning. A Laboratory Manual. (Cold Spring Harbor, NY. 1989)
Sanger, F., Nicklen, S. and Coulson, A.R. (1977): Proc. Natl. Acad. Sci. U.S.A. 74, 5463 - 5467.
Sjoberg, S., Carlsson, P., Enerback, S. and Bjursell, G. (1989): Comput. Appl. Biol. Sci. 5, 41 - 46.
Taggart. R. T. , Mohandas, Т., Shows, T.B. and Bell, G.I. (1985): Proc. Nati. Acad. Sci. U.S.A. 82, 6240 - 6244.
Warburton, D., Gersen, S., Yu, M.T., Jackson, C., Handelin, B. and Housman, D. (1990): Genomics 6, 358 - 366.
Whitelaw et al. (1991): Transgenic Research 1, 3 - 13.
Yu-Lee, L., Richter-Mann, L., Couch, C., Stewart, F., Mackinlay, G. and Rosen, J. (1986): Nucleic. Acid. Res. 14, 1883 - 1902.
Ofverstedt, L. G., Harnmarstrom, K., Balgobin, N., Hjerten, S., Petterson, U. and Chattopadhyaya, J. (1984): Biochim. Biophys. Acta 782, 120 - 126.
Пояснения к чертежам
Фиг. 1. Локус CEL-гена. Показаны локализация и карта рестрикции ферментами двух частично перекрывающихся клонов: λ BSSL1 и λ BSSL5A. Экзон-интронная организация и используемые сайты рестрикции ферментами показаны ниже. Экзоны представлены блоками, пронумерованными цифрами 1 - 11. Asp = Asp700, B = BamH1, E = EcoR1, S = Sac1, Sa = Sal1, Sp = Sph1 и X = Xba1. Положение и ориентация A1 повторяющихся элементов показаны жирными стрелками. Буквами a - h обозначены различные субклонированные фрагменты.
Фиг. 2. Анализ РНК с удлинением праймера. РНК выделена из лактирующей молочной железы, поджелудочной железы и жировой ткани человека. Для праймирования РНК обратной транскрипцией использован радиомеченный с конца 26-тимерный олигонуклеотид, комплементарный нуклеотидам в положениях 33 - 58 CEL-гена. Вертикальная полоса A - маркер молекулярного размера (лэддер последовательности нуклеотидов), вертикальная полоса B - панкреатическая РНК, вертикальная полоса C - РНК жировой ткани и вертикальная полоса С - РНК лактирующей молочной железы.
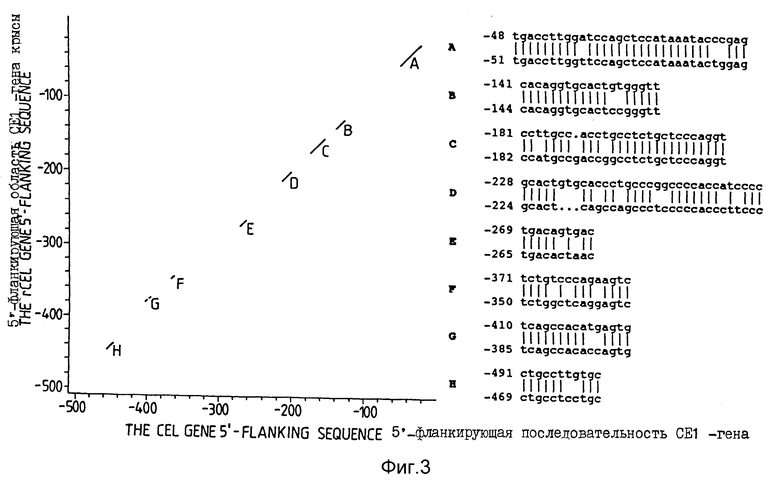
Фиг. 3. Дот-плот анализ 5'-фланкирующих областей CEL-генов человека и крысы. Области гомологии отмечены буквами A - H и прописными буквами написаны представляющие эти области последовательности: сверху - для человека и снизу - для крысы.
Фиг. 4. Анализ 5'-фланкирующей последовательности CEL-гена человека. Предполагаемые распознающие последовательности либо выделены подчеркиванием, либо подчеркиванием представлена комплементарная нить. Жирными буквами показано местоположение гомологичности к CEL-гену крысы (области A - H). Рамка Хогнесса подчеркнута пунктиром.
Существуют две последовательности, каждая из которых показывает 80%-ое сходство с консенсусной последовательностью участка связывания глюкокортикоидного рецептора (GGTACANNNTGTTCT) (Beato M. 1989), первая из них находится у нуклеотида в положении - 231 (1A), а вторая - у нуклеотида в положении - 811 (1B). Более того, у нуклеотида в положении - 861 находится последовательность, показывающая 87%-ное сходство с консенсусной последовательностью участка связывания рецептора эстрогена (AGGTCANNNTGACCT) (Beato M. 1989).
В работе Lubon и Henninghausen (1987) осуществлен анализ промотора и 5'-фланкирующих последовательностей гена сывороточного кислотного белка (WAP-гена) и выявлены участки связывания белков ядра клеток лактирующей молочной железы. Один из них, а именно участок консервативной последовательности в 11 п.о. (AAGAAGGAAGT) присутствует в ряде изученных генов молочных белков, например гене крысиного α- лактальбумина (Qasba и др., 1984) и гене крысиного α- казеина (Yu-Lee и др., 1986). В 5'-фланкирующей области CEL-гена на комплементарной нити у нуклеотида в положении - 1299 находится последовательность, показывающая 82%-ное сходство с указанной консервативной последовательностью.
При исследовании регуляции гена β- казеина в ядерных экстрактах беременных и лактирующих мышей обнаружен тканевоспецифичный фактор молочной железы (ФМЖ) и была идентифицирована его распознающая последовательность (ANTTCTTGGNA). В 5'-фланкирующей области CEL-гена человека имеются две последовательности, одна из которых находится на комплементарной нити у нуклеотида в положении - 368 (4A), а другая - у нуклеотида в положении - 1095 (4B), каждая из которых показывает 82%-ное сходство с консенсусной последовательностью участка связывания ФМЖ. Кроме этих двух предполагаемых участков связывания ФМЖ в 5'-фланкирующей области на комплементарной нити у нуклеотида 275 в интроне 1 находится последовательность: AGTTCTTGGCA, показывающая 100%-ную идентичность консенсусной последовательности участка связывания ФМЖ.
Кроме того, существует четыре последовательности, каждая из которых показывает 65%-ное сходство с консенсусной последовательностью крысиного панкреоспецифичного усиливающего элемента (GTCACCTGTGCTTTTCCCTG) (Boulet и др. , 1986), одна из которых находится у нуклеотида в положении - 359 (5A), вторая - у нуклеотида в положении - 718 (5B), третья - у нуклеотида в положении - 1140 и последняя - у нуклеотида в положении - 1277 (5D).
Фиг. 5. Способ получения плазмиды pS452. Дополнительные подробности см. пример 2.
Фиг. 6. Схематическое строение плазмиды pS312.
Фиг. 7. Схематичное строение плазмиды pS452.
Фиг. 8. Физическая карта, представляющая физическое введение CEL-геномной структуры человека в первый экзон WAP-гена по методике примера 2.
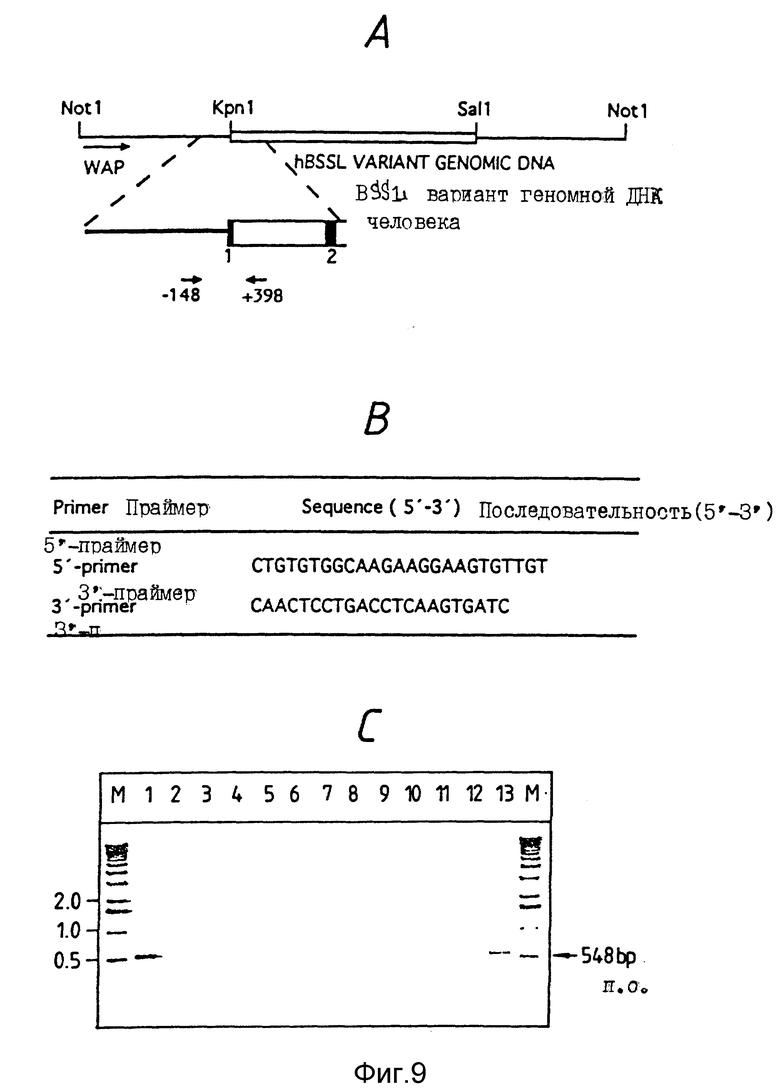
Фиг. 9.
A. Схематический показ локализации ПЦР-праймеров, применяемых для выявления трансгенных животных. 5'-Праймер расположен в пределах WAP последовательности, начинаясь в положении - 148 п.о. в восходящем направлении от места слияния WAP и CEL. 3'-Праймер расположен в первом CEL интроне, заканчиваясь на расстоянии 398 п.о. в нисходящем направлении от места слияния.
B. Последовательности применяемых ПЦР-праймеров.
C. Агарозный гель, показывающий типичный анализ методом ПЦР потенциальных животных-основателей. М - маркеры молекулярной массы, вертикальная полоса 1 - контрольный ПЦР-продукт, полученный из плазмиды pS452, вертикальные полосы 2 - 13 - ПЦР, проведенные с ДНК препаратами от потенциальных животных-основателей.
Фиг. 10. Иммуноблот-анализ молока от мышиной линии, трансгенный по рекомбинантному мышиному WAP/человечьему CEL-гену плазмиды pS452. Белки разделяют методом НДС-ПАГЭ, переносят на Иммобилоновые мембраны (Миллипор) и визуально изучают продукты реакции с поликлональными кроличьими антителами, созданными с применением высокоочищенной нативной человечьей СЖЛ, с последующей реакцией со свиным антикроличьим IgC, меченным щелочной фосфатазой (Дакопаттс). Вертикальная полоса 1: низкомолекулярные маркеры 106, 80, 49,5, 32,5, 27,5 и 18,5 кДа соответственно. Вертикальная полоса 2: высокомолярные маркеры 205, 116,5, 80 и 49,5 кДа соответственно. Вертикальная полоса 3: 25 нг очищенной нерекомбинантной СЖЛ из человечьего молока. Вертикальная полоса 4: 2 мкл образца молока от CEL трансгеной мыши в разбавлении 1:10. Вертикальные полосы 5 и 6: образцы молока по 2 мкл от двух различных не-CEL трансгенных мышей в разбавлении 1:10 в качестве контрольных образцов.
Перечень последовательностей
(2) Информация для Последовательности N 1:
(i) Характеристики последовательности:
(A) Длина: 11531 пара оснований
(B) Тип: нуклеиновая кислота
(C) Число нитей: две
(D) Топология: линейная
(ii) Молекулярный тип: ДНК (геномная)
(vi) Первоначальный источник:
(A) Организм: Homo Sapiens
(B) Тип ткани: молочная железа
(ix) Признак:
(A) Название /ключ: CDS
(B) Местоположение: стык (1653...1727, 4071...4221, 4307...4429, 4707... 4904, 6193. . . 6323, 6501. ..6608, 6751...6868, 8335...8521, 8719...8922, 10124...10321, 10650...11394)
(ix) Признак:
(A) Название/ключ: зрелый пептид
(B) Местоположение: стык (1722...1727, 4071...4221, 4307...4429, 4707... 4904, 6193. . . 6323, 6501. ..6608, 6751...6868, 8335...8521, 8719...8922, 10124...10321, 10650...11394)
(D) Прочая информация: /EC. номер 3.1.1.1 /продукт = "Стимулируемая солями желчных кислот липаза"
(ix) Признак:
(A) Название/ключ: 5' UTR
(B) Местоположение: 1...1640
(ix) Признак:
(A) Название/ключ: рамка Хогнесса
(B) Местоположение: 1611...1617
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 1641...1727
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 4071...4221
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 4307...4429
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 4707...4904
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 6193...6323
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 6501...6608
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 6751...6868
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 8335...8521
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение:8719...8922
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 10124....10321
(ix) Признак:
(A) Название/ключ: экзон
(B) Местоположение: 10650...11490
(ix) Признак:
(A) Название/ключ: 3' UTR
(B) Местоположение: 11491...11531
(XI) Описание последовательности: Последовательность N 1:







Изобретение относится к генетической инженерии и касается ДНК, кодирующей BSSL/CEL и векторы экспрессии. ДНК имеет нуклеотидную последовательность SEQID NO: 1. Вектор содержит 2,7 т.п.о. NotI - NatI фрагмент рИС 19. Фрагмент включает ген устойчивости к ампициллину amp R, 5'-фланкирующую и 5'-нетранслируемую области WAP мыши на фрагменте NotI - KpnI размером в 4,1 т. п. о., структурный ген BSSL человека на 9,9 т.п.о. KpnI - SalI фрагменте, 3'-фланкирующую область и структурный ген WAP мыши на SalI - Not фрагменте размером 4,3 т. п. о. ДНК позволяет получать рекомбинантные человечьи ССЖЛ/КСЛ, которые можно использовать как компонент детского питания. 2 с.п. ф-лы, 5 табл., 10 ил.
| WO 9118923 A1, 1991 | |||
| WO 9115234 A1, 1991 | |||
| Рекомбинантная плазмидная ДНК pBG М3, определяющая синтез эндонуклеазы рестрикции С @ BI | 1988 |
|
SU1573026A1 |

