Данное изобретение относится к оптимизации получения рекомбинантной сахарозосинтазы (SS) в растворимой активной форме с использованием подходящего штамма Escherichia coli, применению SS для приготовления наборов для определения сахарозы, конструированию оптимизированных форм SS для синтеза ADP-глюкозы (ADPG) и получению трансгенных растений, листья и запасающие ткани которых накапливают высокие уровни ADPG и обогащенного амилозой крахмала в результате сверхпродуцирования цитозольной ADPG в растениях, которые сверхэкспрессируют SS.
УРОВЕНЬ ТЕХНИКИ
Крахмал является основной запасной формой углеводов в растениях. Он накапливается в больших количествах в таких органах, как семена (пшеница, ячмень, кукуруза, горох и т.д.) и клубни (картофель и ямс, среди прочих), и является фундаментальным компонентом рациона человека. Кроме того, крахмал широко используется в бумажной, косметической, фармацевтической и пищевой промышленности, а также используется в качестве незаменимого компонента для изготовления биодеградируемых пластиков и благоприятных для окружающей среды красок. Поскольку он состоит из ковалентно связанных молекул глюкозы, исследования процессов, участвующих в синтезе этого полисахарида, является первоочередной задачей в различных областях промышленного производства.
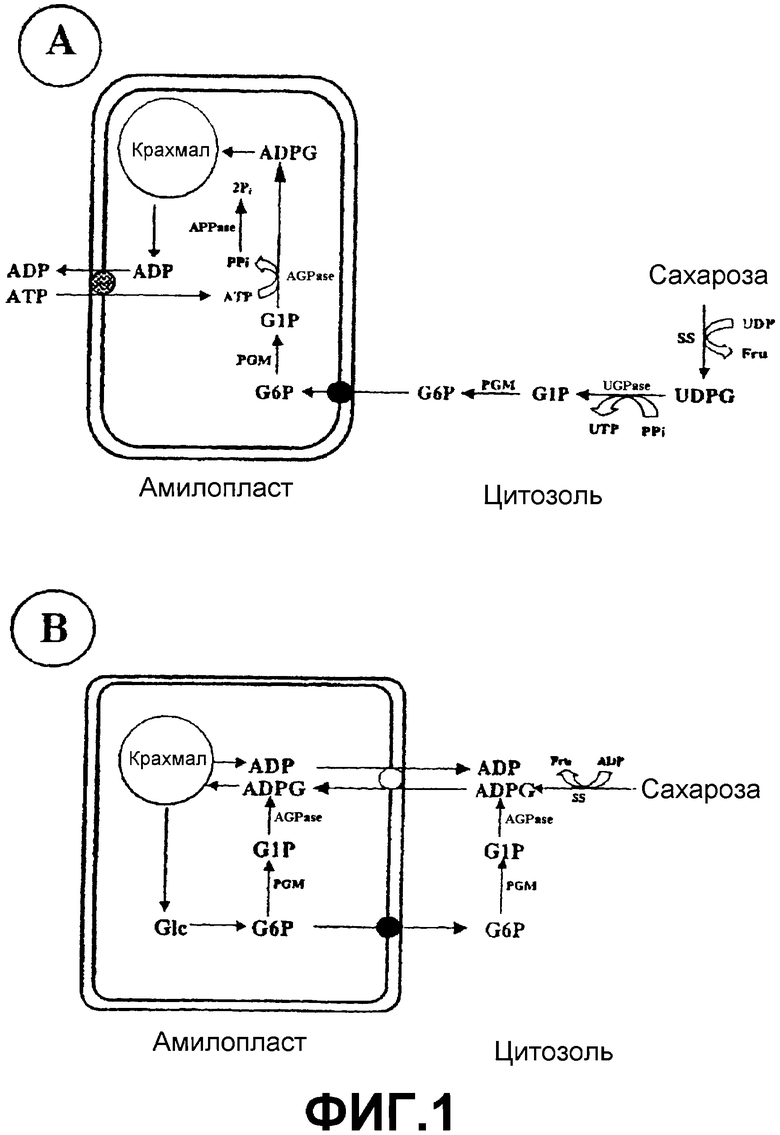
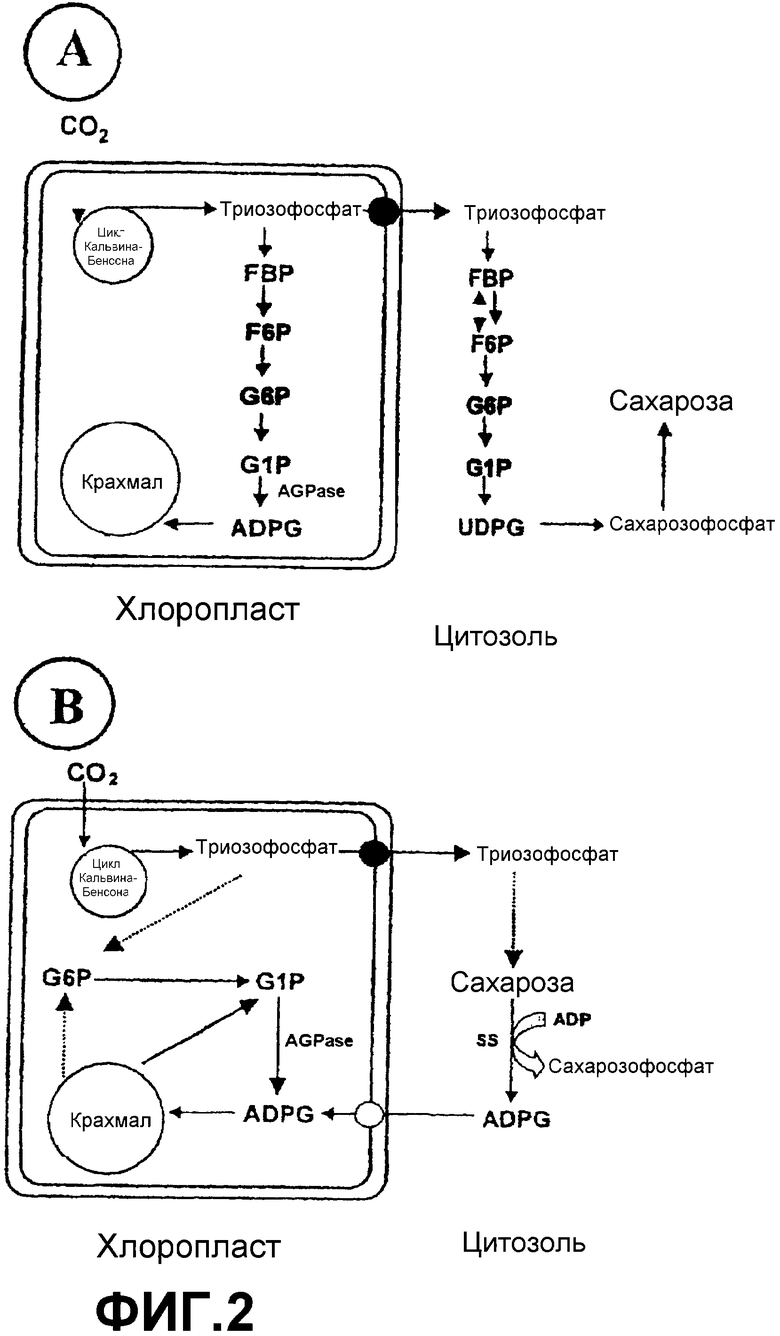
ADPG является универсальным предшественником биосинтеза крахмала в растениях, как в гетеротрофных органах (фиг.1А), так и в листьях (фиг.2А), и в большой степени предполагается, что ее продуцирование регулируется исключительно ферментом ADPG-пирофосфорилазой (AGPазой) или ADPG-синтазой (ЕС 2.7.7.27) (Okita, T.W. (1992) Is there an alternative pathway for starch synthesis? Plant Physiol. 100, 560-56; Müller-Röber, В., Sonnewald, U. Willmitzer, L. (1992) Inhibition of the ADPglucose pyrophosphorylase in transgenic potatoes leads to sugar-storing tubers and influences tuber formation and expression of tuber storage protein genes. EMBO J. 11, 1229-1238; Stark, D.M., Timmerman, K.P., Barry, G.F., Preiss, J., Kishore, G.M. (1992) Regulation of the amount of starch in plant tissues by ADPglucose pyrophosphorylase. Science 258, 287-282; Neuhaus, E.H., Häusler, R.E., Sonnewald, U. (2005) No time to shift the paradigm on the metabolic pathway to transitory starch in leaves. Trends Plant Sci. at press). Различные применения крахмала, продуцируемого в растении, основаны в основном на соотношении амилозы и амилопектина, которое определяет структуру крахмального зерна, а также его вязкость в водных суспензиях. Это соотношение амилозы и амилопектина зависит, среди прочего, от концентрации ADPG в клетке растения (Clarke, B.R., Denyer, K., Jenner, C.F., Smith, A.M. (1999) The relationship between the rate of starch synthesis, the adenosine 5'-diphosphoglucose concentration and the amylose content of starch in developing pea embryos. Planta 209, 324-329).
SS (ЕС° 2.4.1.13, °SS) (UDP-глюкозо:D-фруктозо-2-глюкозилтрансфераза) является обратимым ферментом, который катализирует продуцирование UDPG и фруктозы из сахарозы и UDP. Хотя, как показано на фиг.1А, SS классически рассматривалась как играющая роль в продуцировании UDPG, метаболический процессинг, который в конечном счете приводит к образованию крахмала в гетеротрофных тканях, таких как эндосперм и клубни (Zrenner, R., Salanoubat, M., Willmitzer, L., Sonnewald, U. (1995) Evidence for the crucial role of sucrose synthase for sink strength using transgenic potato plants. Plant J. 7, 97-107; Baroja-Fernández, E., Muсoz, F.J., Saikusa, Т., Rodríguez-López, M., Akazawa, Т., Pozueta-Romero, J. (2003) Sucrose synthase catalyzes the de novo production of ADPglucose linked to starch biosynthesis in heterotrophic tissues of plants. Plant Cell Physiol. 44, 500-509; Pozueta-Romero, J., Munoz, F.J., Rodríguez-López, M., Baroja-Fernández, E., Akazawa, T. (August 2003) New waves in the starch field. Lett. Plant Cell Physiol. 24-32), имеются ссылки на потенциальную способность этого фермента использовать другие нуклеотиддифосфаты in vitro для получения соответствующих сахаронуклеотидов (т.е. активированных нуклеотидами сахаров) (Murata, Т., Sugiyama, Т., Minamikawa, Т., Akazawa, T. (1966) Enzymic mechanism of starch synthesis in ripening rice grains. Mechanism of the sucrose-starch conversion. Arch. Biochem. Biophys. 113, 34-44; Delmer, D.P. (1972) The purification and properties of sucrose synthase from etiolated Phaseolus aureus seedlings. J. Biol. Chem. 247, 3822-3828). Хотя и существует сомнение в отношении физиологической релевантности (Okita, T.W. (1992) Is there an alternative pathway for starch synthesis? Plant Physiol. 100, 560-56; Müller-Röber, В., Sonnewald, U. Willmitzer, L. (1992) Inhibition of the ADPglucose pyrophosphorylase in transgenic potatoes leads to sugar-storing tubers and influences tuber formation and expression of tuber storage protein genes. EMBO J. 11, 1229-1238), было сделано предположение, что SS способна непосредственно продуцировать ADPG, которая может быть использована для продуцирования крахмала как в гетеротрофных тканях, так и в фотосинтетических тканях (фиг.1В и 2В) (Pozueta-Romero, J., Perata, P., Akazawa, T. (1999) Sucrose-starch conversion in heterotrophic tissues of plants. Crit. Rev. Plant Sci. 18, 489-525; Baroja-Fernández, E., Munoz, F.J., Akazawa, Т., Pozueta-Romero, J. (2001) Reappraisal of the currently prevailing model of starch biosynthesis in photosynthetic tissues: a proposal involving the cytosolic production of ADPglucose by sucrose synthase and occurrence of cyclic turnover of starch in the chloroplast. Plant Cell Physiol. 42, 1311-1320; Baroja-Fernández, E., Muсoz, F.J., Saikusa, Т., Rodríguez-López, M., Akazawa, Т., Pozueta-Romero, J. (2003) Sucrose synthase catalyzes the de novo production of ADPglucose linked to starch biosynthesis in heterotrophic tissues of plants. Plant Cell Physiol. 44, 500-509; Baroja-Fernández, E., Munoz, F.J., Zandueta-Criado, A., Morán-Zorzano, M.T., Viale, A.M., Alonso-Casajus, N., Pozueta-Romero, J. (2004) Most of ADPglucose linked to starch biosynthesis occurs outside the chloroplast in source leaves. Proc. Natl. Acad. Sci. USA 101, 13080-13085). Согласно этой гипотезе (основанной только и косвенно на доказательстве биохимического типа) SS является ответственной за синтез важного пула молекул ADPG, необходимых для биосинтеза крахмала. Однако эта гипотеза не была подтверждена экспериментально генной инженерией или традиционными способами улучшения сельскохозяйственных культур и не согласуется с бесчисленными тестами генетического и молекулярного типа, свидетельствующими о том, что AGPаза является единственным источником ADPG в растениях (Okita, T.W. (1992) Is there an alternative pathway for starch synthesis? Plant Physiol. 100, 560-56; Müller-Röber, В., Sonnewald, U. Willmitzer, L. (1992) Inhibition of the ADPglucose pyrophosphorylase in transgenic potatoes leads to sugar-storing tubers and influences tuber formation and expression of tuber storage protein genes. EMBO J. 11, 1229-1238; Neuhaus, E.H., Häusler, R.E., Sonnewald, U. (2005) No time to shift the paradigm on the metabolic pathway to transitory starch in leaves. Trends Plant Sci. at press).
Сахаронуклеотиды, такие как UDPG и ADPG, получают коммерчески из пирофосфорилазных реакций, катализируемых такими ферментами, как UDPG-пирофосфорилаза (UGP-аза) и AGP-аза, соответственно, основанных на использовании дорогостоящего вещества, называемого глюкозо-1-фосфатом (G1P). Альтернатива этой практике получения сахаронуклеотидов основывается на применении SS, разработка которого в значительной степени затруднялась ограничениями Escherichia coli для экспрессии и эффективного процессинга большого количества эукариотических белков. Это ограничение побудило некоторых исследователей к получению рекомбинантной SS с использованием биологических фабрик (кассет биореакторов для крупномасштабного производства клеток) эукариотического типа, таких как дрожжи (Zervosen, A., Römer, U., Elling, L. (1998) Application of recombinant sucrose synthase-large scale synthesis of ADP-glucose. J. Mol. Catalysis B: Enzymatic 5, 25-28; Römer, U., Schrader, H., Günther, N., Nettelstroth, N., Frommer, W.B., Elling, L. (2004) Expression, purification and characterization of recombinant sucrose synthase I from Solanum tuberosum L. for carbohydrate engineering. J. Biotechnology 107, 135-149). Альтернативно, SS, предназначенная для получения сахаронуклеотидов, должна была быть очищена дорогостоящими процессами очистки белков из экстрактов растений (Патент DE4221595 (1993), Purified sucrose synthase enzyme useful for production of nucleotide-activated sugars or oligosaccharides). Эта SS, полученная из экстрактов растений, имеет недостаток, заключающийся в том, что она имеет предпочтение в отношении UDP и очень низкую аффинность в отношении ADP (Pressey R (1969) Potato sucrose synthase: purification, properties, and changes in activity associated with maturation. Plant Physiol. 44, 759-764; Nguyen-Quock, В., Krivitzky, M., Huber, S.C., Lecharny, A. (1990) Sucrose synthase in developing maize leaves. Plant Physiol. 94, 516-523; Morell, M., Copeland, L. (1985) Sucrose synthase of soybean nodules. Plant Physiol. 78, 149-154). Production of recombinant SS from cultures of E.coli has recently been achieved (Nakai, Т., Tonouchi, N., Tsuchida, Т., Mori, H., Sakai, F., Hayashi, T. (1997) "Expression and characterization of sucrose synthase from mung bean seedlings in Escherichia coli" Biosci. Biotech. Biochem. 61, 1500-1503; Nakai, Т., Konishi, Т., Zhang, Z-Q., Chollet, R., Tonouchi, N., Tsuchida, Т., Yoshinaga, F., Mori, H., Sakai, F., Hayashi, T. (1997) "An increase in apparent affinity for sucrose of mung bean sucrose synthase is caused by in vitro phosphorylation or directed mutagenesis of Ser11" Plant Cell Physiol. 39, 1337-1341; Barratt, D.H.P., Barber, L., Kruger, N.J., Smith, A.M., Wang, T.L., Martin, C. (2001) Multiple, distinct isoforms of sucrose synthase in pea. Plant Physiol. 127, 655-664; Christopher, В., William, В., Robert, H. "Bacterial sucrose synthase compositions and methods of use" Patent WO9803637). Однако получение SS в этой прокариотической системе было связано с проблемами, заключающимися в том, что (1) количество полученной SS было очень низким (30 микрограммов/грамм бактерий (Nakai, Т., Tonouchi, N., Tsuchida, Т., Mori, H., Sakai, F., Hayashi, T. (1997) "Expression and characterization of sucrose synthase from mung bean seedlings in Escherichia coli" Biosci. Biotech. Biochem. 61, 1500-1503; Li, C.R., Zhang, X.B., Hew, C. S. (2003) "Cloning, characterization and expression analysis of a sucrose synthase gene from tropical epiphytic orchid Oncidium goldiana. Physiol. Plantarum 118, 352-360); (2) количество полученной активной SS было очень низким или равным нулю (0,05-1,5 единиц/мг) (Nakai, Т., Tonouchi, N., Tsuchida, Т., Mori, H., Sakai, F., Hayashi, T. (1997) "Expression and characterization of sucrose synthase from mung bean seedlings in Escherichia coli" Biosci. Biotech. Biochem. 61, 1500-1503; Li, C.R., Zhang, X.B., Hew, C.S. (2003) "Cloning, characterization and expression analysis of a sucrose synthase gene from tropical epiphytic orchid Oncidium goldiana. Physiol. Plantarum 118, 352-360); 5,6 U/mg (Rümer, U., Schrader, H., Gönther, N., Nettelstroth, N., Frommer, W.B., Elling, L. (2004) Expression, purification and characterization of recombinant sucrose synthase I from Solanum tuberosum L. for carbohydrate engineering. J. Biotechnology 107, 135-149), (3) рекомбинантная SS должна была быть очищена обычными способами очистки белков, такими как хроматография, электрофорез, изоэлектрическое фокусирование и т.д., которые, вместе, являются дорогостоящими и не гарантируют очистки этого белка до гомогенного состояния, и (4) большая часть этой SS направляется к тельцам включения или накапливается в форме неактивных агрегатов вследствие неспособности аппарата бактерий правильно укладывать этот белок (Miroux, B., Walker, J.E. (1996) “Over-production of proteins in Escherichia coli: mutant hosts that allow synthesis of some membrane proteins and globular proteins at high levels” J. Mol. Biol. 260, 289-298).
Данное изобретение описывает разработку системы на основе использования подходящего штамма E.coli и на основе использования подходящего экспрессирующего вектора, который делает возможными крупномасштабное получение и быструю и легкую очистку различных вариантов рекомбинантной SS в ее активной форме. Некоторые из этих вариантов имеют более высокую аффинность в отношении ADP, чем варианты, получаемые из экстрактов растений, и могут быть использованы как для получения UDPG, так и для получения ADPG из недорогих веществ, таких как сахароза, UDP и ADP.
Хроматографические способы составляют мощный инструмент для определения содержания сахарозы сложных проб, таких как экстракты растений, сыворотки, моча, фруктовый сок, вина, фрукты и пищевые продукты (D'Aoust, M-A., Yelle, S, Nguyen-Quock, B. (1999) Antisense inhibition of tomato fruit sucrose synthase decreases fruit setting and the sucrose unloading capacity of young fruit. Plant Cell 11, 2407-2418; Tang, G-Q., Sturm, A. (1999) Antisense repression of sucrose synthase in carrot affects growth rather than sucrose partitioning. Plant Mol. Biol. 41, 465-479; Frias, J., Price, K.R., Fenwich, G.R., Hedley, C.L., Sorensen, H., Vidal-Valverde, C. (1996) J. Chromatogr. A 719, 213-219). Такие способы требуют участия высококвалифицированного технического персонала и большого вложения в оборудование. К сожалению, альтернативные способы, основанные на гидролизе молекулы сахарозы под действием фермента инвертазы и последующем спектрофотометрическом или флуорометрическом определении молекул глюкозы и/или фруктозы (Sweetlove, L.J., Burrell, M.M., ap Rees, T. (1996) Starch metabolism in tubers of transgenic potato with increased ADPglucose pyrophosphorylase. Biochem. J. 320, 493-498; Stitt, M., Lilley, R.M., Gerhardt, R., Heldt, H.W. (1989) Metabolite levels in specific cells and subcellular compartments of plant leaves. Methods Enzymol. 174, 518-552; Holmes, E.W. (1997) Coupled enzymatic assay for the determination of sucrose. Anal. Biochem. 244, 103-109; Methods of Analysis (1996) Code of Practice for Evaluation of Fruit and Vegetable Juices. Association of the Industry of Juices and Nectars from Fruits and Vegetables of the European Economic Community), являются предметом ограничений технического характера, например, требующих вычитания измерений, соответствующих эндогенной глюкозе и/или фруктозе, присутствующим в пробе. Избыток глюкозы и/или фруктозы в пробе может увеличивать фоновый шум, который затрудняет достоверное и точное определение сахарозы. В огромном большинстве случаев необходимо проводить трудоемкие контрольные измерения перед принятием достоверного утверждения об истинном содержании сахарозы пробы (Worrell, A.C., Bruneau, J-M., Summerfelt, K., Boersig, M., Voelker, T.A. (1991) Expression of a maize sucrose phosphate synthase in tomato alters leaf carbohydrate partitioning. Plant, Cell 3, 1121-1130). Наборы для определения сахарозы, основанные на использовании инвертазы, доступны из таких компаний, как Sigma, Biopharm GmbH и Megazyme. Альтернативно, был разработан автоматизированный способ определения сахарозы, основанный на определении глюкозо-1-фосфата, высвобождаемого под действием сахарозофосфорилазы бактериального происхождения (Vinet, B., Panzini, B., Boucher, M., Massicotte, J. (1998) Automated enzymatic assay for the determination of sucrose in serum and urine and its use as a marker of gastric damage. Clin. Chem. 44, 2369-2371). Данное изобретение описывает развитие простого, достоверного и недорогого альтернативного способа для определения сахарозы в пробе, основанного на использовании SS и сопряженных ферментов, которые гидролизуют ADPG или UDPG.
Обсуждения, касающиеся факторов, управляющих внутриклеточными уровнями ADPG, вращались вокруг регуляции синтезирующего фермента, AGPазы (Preiss, (1988) Biosynthesis of starch and its regulation. The Biochemistry of Plants. Vol.14, Academic Press, New York, p.182-249; Pozueta-Romero, J., Perata, P., Akazawa, T. (1999) Sucrose-starch conversion in heterotrophic tissues. Crit. Rev. Plant. Sci. 18, 489-525). Действительно, высокая доля патентов и научных публикаций, касающихся получения ADPG и получения растений, продуцирующих крахмалы в количествах, представляющих интерес в промышленности, была сосредоточена на использовании AGPазы (Stark, D.M., Timmerman, K.P., Barry, G.F., Preiss, J., Kishore, G.M. (1992) Regulation of the amount of starch in plant tissues by ADPglucose pyrophosphorylase. Science 258, 287-282; Slattery, C.J., Kavakli, H., Okita, T.W. (2000) Engineering starch for increased quantity and quality. Trends Plant Sci. 5, 291-298). Однако, хотя они все еще должны быть подкреплены данными генетического/молекулярного типа, недавние научные исследования биохимического типа показывают, что, как показано на фигурах 1В и 2В, SS может участвовать в прямом синтезе ADPG, необходимого для биосинтеза крахмала (Baroja-Fernández, E., Muсoz, F.J., Saikusa, Т., Rodríguez-López, M., Akazawa, Т., Pozueta-Romero, J. (2003) Sucrose synthase catalyzes the de novo production of ADPglucose linked to starch biosynthesis in heterotrophic tissues of plants. Plant Cell Physiol. 44, 500-509). Эта гипотеза является особенно спорной, если иметь в виду, что (а) SS никогда не была связана с образованием крахмала в листьях, (b) необходимо присутствие переносчика (транслокатора) ADPG в мембраны пластид, соединяющего цитозольный пул ADPG, образуемой посредством SS, с крахмалсинтазой, присутствующей внутри пластиды, и (с) участие SS в качестве ADPG-продуцирующего источника находится в прямом противоречии со многими тестами биохимического/генетического/молекулярного типа, которые, по-видимому, показывают, что AGPаза является только источником ADPG (Okita, T.W. (1992) Is there an alternative pathway for starch synthesis? Plant Physiol. 100, 560-56; Müller-Röber, В., Sonnewald, U. Willmitzer, L. (1992) Inhibition of the ADPglucose pyrophosphorylase in transgenic potatoes leads to sugar-storing tubers and influences tuber formation and expression of tuber storage protein genes. EMBO J. 11, 1229-1238; Stark, D.M., Timmerman, K.P., Barry, G.F., Preiss, J., Kishore, G.M. (1992) Regulation of the amount of starch in plant tissues by ADPglucose pyrophosphorylase. Science 258, 287-282; Neuhaus, E.H., Häusler, R.E., Sonnewald, U. (2005) No time to shift the paradigm on the metabolic pathway to transitory starch in leaves. Trends Plant Sci. at press). Возможно, по всем этим причинам до настоящего времени никогда не конструировались растения, которые сверхэкспрессируют SS, для получения высоких уровней крахмала. Однако данное изобретение описывает, впервые, получение трансгенных растений, которые сверхэкспрессируют SS, для увеличения продуцирования этими растениями ADPG и крахмала. Напротив, авторы данного изобретения показывают, что растения, которые лишены крахмала вследствие отсутствия AGPазы, имеют нормальные уровни ADPG. Все это показывает, что, как показано на фигурах 1В и 2В, SS участвует в прямом синтезе ADPG, необходимой для биосинтеза крахмала, и является ответственной за синтез наибольшей части ADPG, накапливаемой в клетке растения.
Хотя на основе подхода, представленного на фиг.1А, в соответствии с которым SS участвует в синтезе UDPG (но не ADPG) в запасающих тканях, различные работы описали получение растений с уменьшенным содержанием крахмала в результате уменьшенной активности SS (Chourey, P.S., Nelson, O.E. (1976) The enzymatic deficiency conditioned by the shrunken-1 mutations in maize. Biochem. Genet. 14, 1041-1055; Zrenner, R., Salanoubat, M., Willmitzer, L., Sonnewald, U. (1995) Evidence for the crucial role of sucrose synthase for sink strength using transgenic potato plants. Plant J. 7, 97-107; Tang, G-Q., Sturm, A. (1999) Antisense repression of sucrose synthase in carrot (Daucus carota L.) affects growth rather than sucrose partitioning. Plant Mol. Biol. 41, 465-479). В этом смысле не существует экспериментального доказательства того, что сверхэкспрессия SS могла бы использоваться для получения растений с высоким содержанием крахмала вследствие увеличения уровней ADPG в соответствии с метаболическими схемами, показанными на фигурах 1В и 2В. Однако на основе способности SS продуцировать молекулу-предшественник биосинтеза полисахаридов клеточной стенки (UDPG), были опубликованы и запатентованы исследования, которые описывают получение растений хлопчатника с высоким содержанием волокна или зерновых с высоким содержанием целлюлоз вследствие сверхэкспрессии SS (Timothy, H.J., Xiamomu, N., Kanwarpal, S. "Manipulation of sucrose synthase genes to improve stalk and grain quality" Patent WО02067662; Robert, F., Danny, L., Yong-Ling, R. "Modification of sucrose synthase gene expression in plant tissue and uses therefor" Patent WО0245485; Christopher, В., William, В., Robert, H. "Bacterial sucrose synthase compositions and methods of use" Patent WО9803637).
Данное изобретение относится, во-первых, к развитию и оптимизации способа получения больших количеств рекомбинантной SS, которая является растворимой, может быть легко очищена и имеет высокую удельную активность, на основе использования подходящего штамма E.coli и экспрессирующего вектора, который делает возможным получение SS с гистидиновым хвостом. Далее, данное изобретение относится к процедуре, выполняемой для приготовления наборов для определения сахарозы, основанной на использовании ферментного продукта с активностью SS, сопряженного с ферментами, которые метаболизируют ADPG до UDPG. Кроме того, данное изобретение относится к оптимизации получения сахаронуклеотидов (активированных нуклеотидами сахаров), таких как ADPG или UDPG, которая исходит из вариантов SS, сконструированных специально для этой цели. Наконец, приведены подробности конструирования трансгенных растений с высоким содержанием сахарозы, ADPG и крахмала и высоким отношением амилоза/амилопектин после сверхэкспрессии SS.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Амплификация кДНК, которая кодирует SS
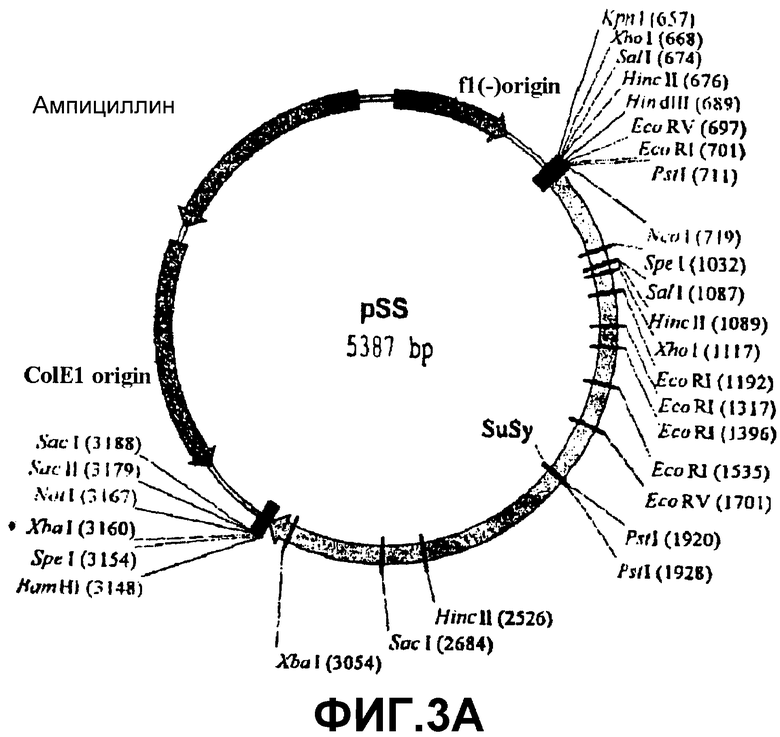
На основе знания нуклеотидной последовательности сахарозосинтазы дикого типа SS4 (Fu, H., Park, W.D. (1995) Sink- and vascular-associated sucrose synthase functions are encoded by different gene classes in potato. Plant Cell 7, 1369-1385), были созданы два специфических праймера, соответствующих 5′ - и 3′ - концам этого гена. С использованием этих праймеров ДНК-фрагмент 2418 п.н., обозначенный SSX, из кДНК-библиотеки листа картофеля был амплифицирован общепринятыми ПЦР-способами. Этот ПЦР-фрагмент встраивали в плазмиду pSK Bluescript (Stratagene) с получением pSS-конструкции (фиг.3А), которую амплифицировали в бактерии-хозяине XL1 Blue.
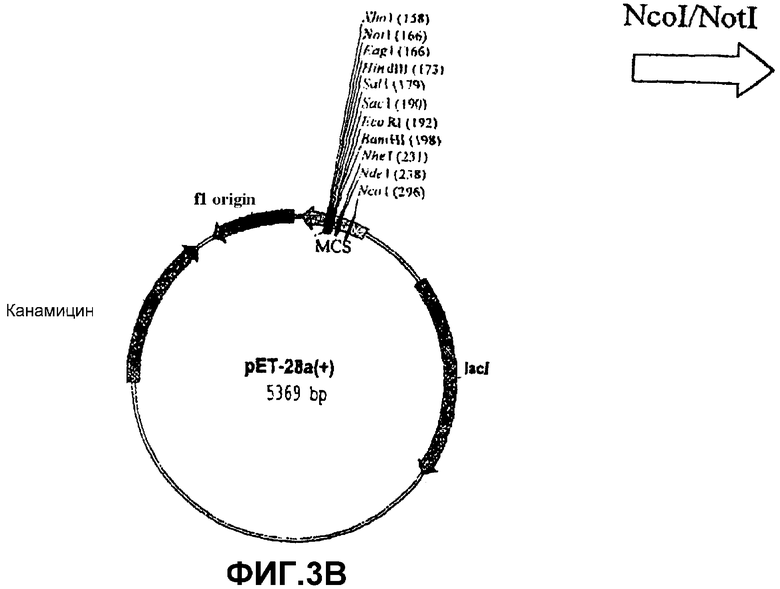
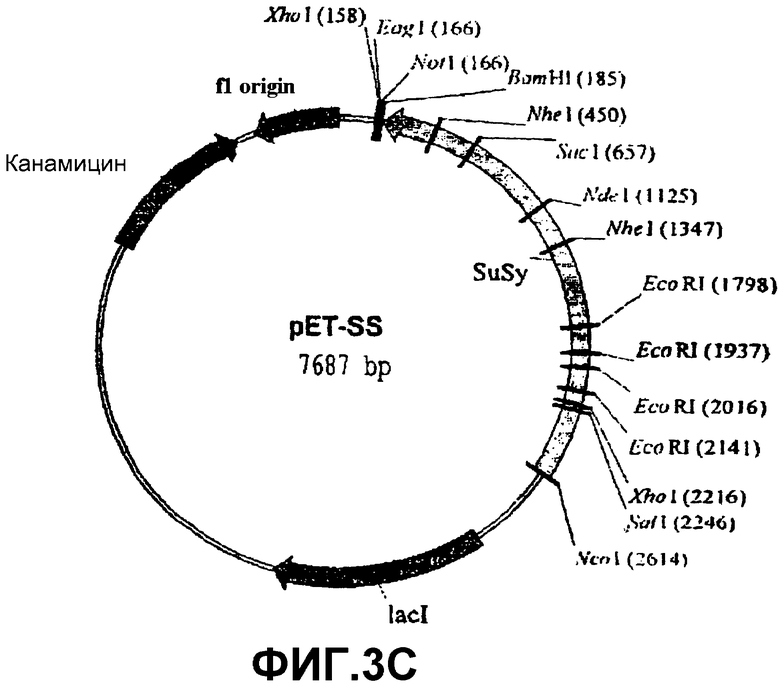
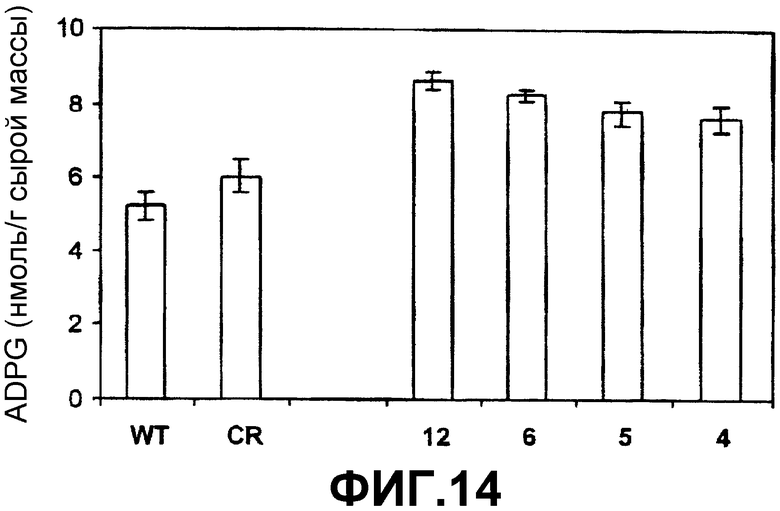
Получение активной рекомбинантной SS из специального штамма E.coli pSS расщепляли рестрикционными ферментами NcoI и NotI. Высвобождаемый фрагмент (который содержит кДНК, кодирующую SS, SSX) клонировали на тех же самых сайтах рестрикции экспрессионной плазмиды рЕТ-28а(+) (Novagen) (фиг.3В), которая имеет нуклеотидную последовательность в полилинкерном районе, которая кодирует богатую гистидином последовательность, которая становится слитой с этим рекомбинантным белком. Полученную плазмиду (обозначенную рЕТ-SS, фиг.3С) встраивали электропорацией в различные штаммы E.coli. Штамм E.coli BLR(DE3) (Novagen), трансформированный pET-SS, был депонирован 29 октября 2003 года в Испанской Коллекции Типовых Культур, расположенной в Research Building of Valencia University. Burjassot Campus, Burjassot 46100 (Valencia, Spain) с номером депозита СЕСТ:5850. Эти бактерии инкубировали при 20оС в среде LB (Луриа-Бертани). Сверхэкспрессию SSX выполняли добавлением 1 мМ изопропил-β-D-тиогалактопиранозида (IPTG) в 100 мл культуры клеток, выращиваемых при 20оС. После шести часов индуцированной культуры бактерии собирали и ресуспендировали в 4 мл связывающего буфера (Novagen, наборы для очистки His-bind), затем обрабатывали ультразвуком и центрифугировали при 40000 g в течение 20 минут.Супернатант, который содержит рекомбинантую SS с аминокислотной последовательностью, богатой остатками гистидина в N-концевой области, пропускали через аффинную колонку набора для очистки белков His-bind из Novagen. В соответствии с инструкциями, прилагаемыми к этому набору, SS элюировали 6 мл рекомендуемого буфера для элюции, который содержал 200 мМ имидазол вместо 1 молярного имидазола. После элюции белок быстро подвергали диализу для удаления любых следов имидазола, который необратимо инактивирует SS.
Получение изоформы SS, оптимизированной для получения ADPG
С использованием подходящих праймеров, с pSS в качестве матрицы, конструировали мутированный вариант SS5 с получением конструкции pSS5. Это выполняли с использованием набора для сайт-направленного мутагенеза QuickChange (Stratagene). pSS5 расщепляли NcoI и NotI. Высвобождаемый фрагмент (который содержит SS5) клонировали на тех же самых сайтах рестрикции экспрессионной плазмиды рЕТ-28а(+) с получением pET-SS5, которую встраивали электропорацией в E.coli BLR(DE3). Штамм E.coli XL1 Blue, трансформированный pSS5, был депонирован 29 октября 2003 года в Испанской Коллекции Типовых Культур, расположенной в Research Building of Valencia University, Burjassot Campus, Burjassot 46100 (Valencia, Spain) с номером депозита СЕСТ:5849.
Получение трансгенных растений, которые сверхэкспрессируют SS4
В данном изобретении SS сверхэкспрессировалась (а) конститутивно, (b) специфически в листьях и (с) специфически в запасающих органах, таких как клубни.
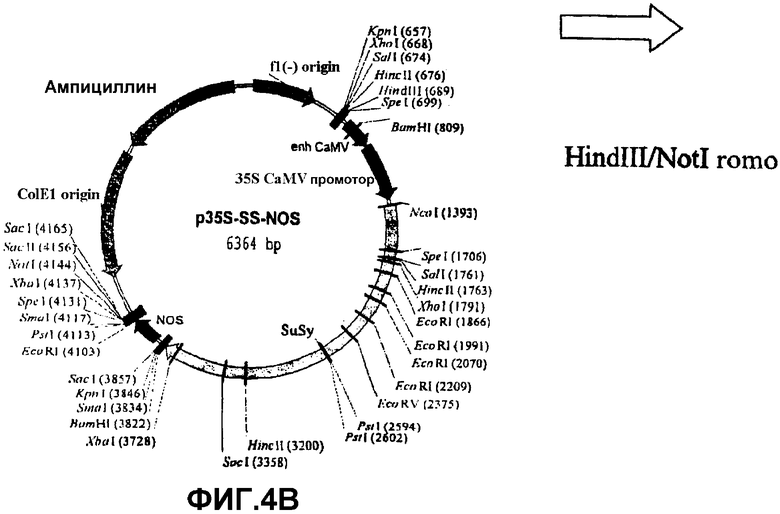
Для получения растений, которые сверхэкспрессировали конститутивно SS, были созданы конструкции, которые регулировались действием конститутивного промотора 35S вируса мозаичной болезни табака. Последовательное встраивание в pSS промотора 35S и терминатора NOS в 5′ - и 3′ - районах SSX приводило к получению плазмиды p35S-SS-NOS, рестрикционная карта которого показана на фиг.4В.
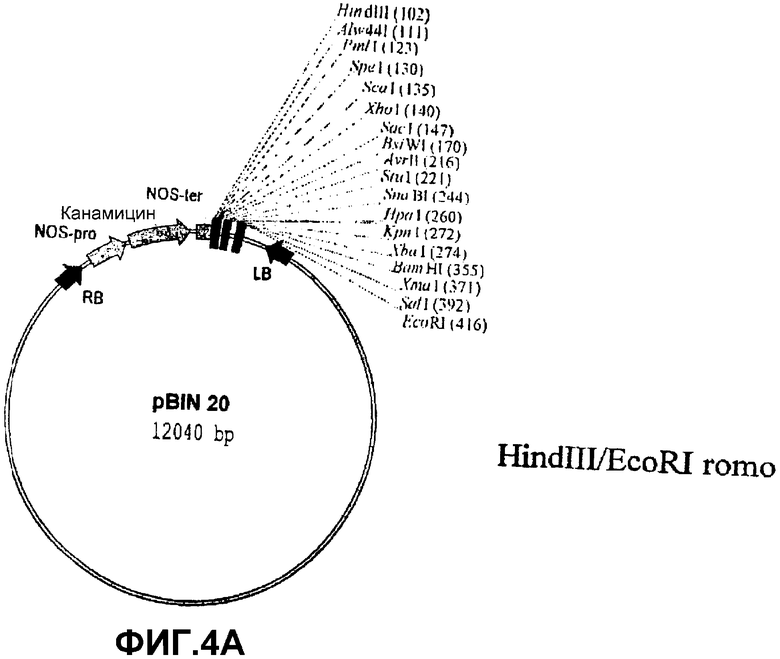
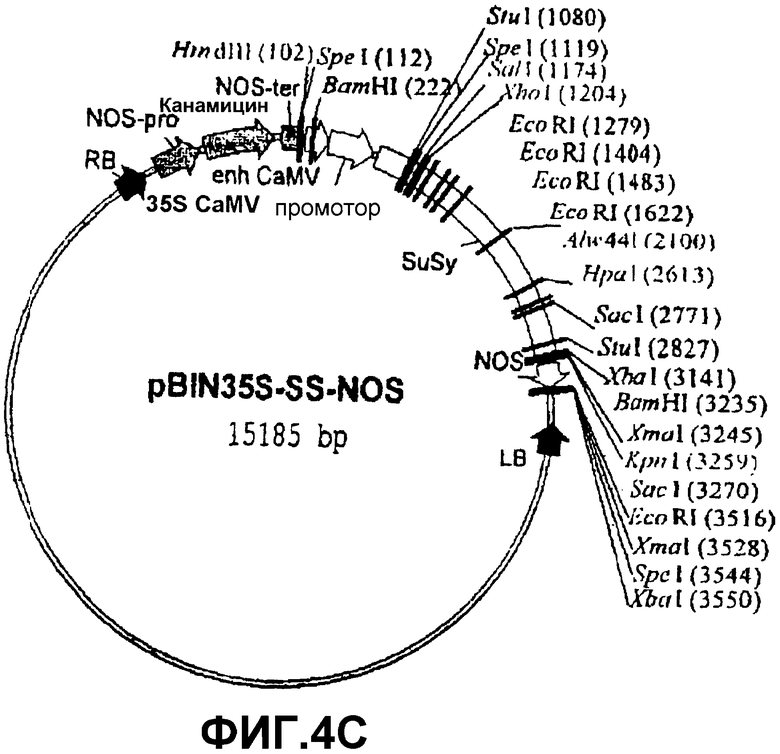
Для возможности переноса этой конструкции в геном растений посредством Agrobacterium tumefaciens ее нужно было сначала клонировать в бинарную плазмиду. Для этого, p35S-SS-NOS расщепляли последовательно ферментами NotI, ДНК-полимеразой Т4 и HindIII и клонировали в бинарной плазмиде pBIN20 (фиг.4А) (Hennegan, K.P., Danna, K.J. (1998) pBIN20: An improved binary vector for Agrobacterium-mediated transformation. Plant Mol. Biol. Rep.16, 129-131), которая была предварительно расщеплена последовательно ферментами EcoRI, ДНК-полимеразой Т4 и HindIII. Полученная таким образом плазмида была обозначена pBIN35S-SS-NOS (фиг.4С).
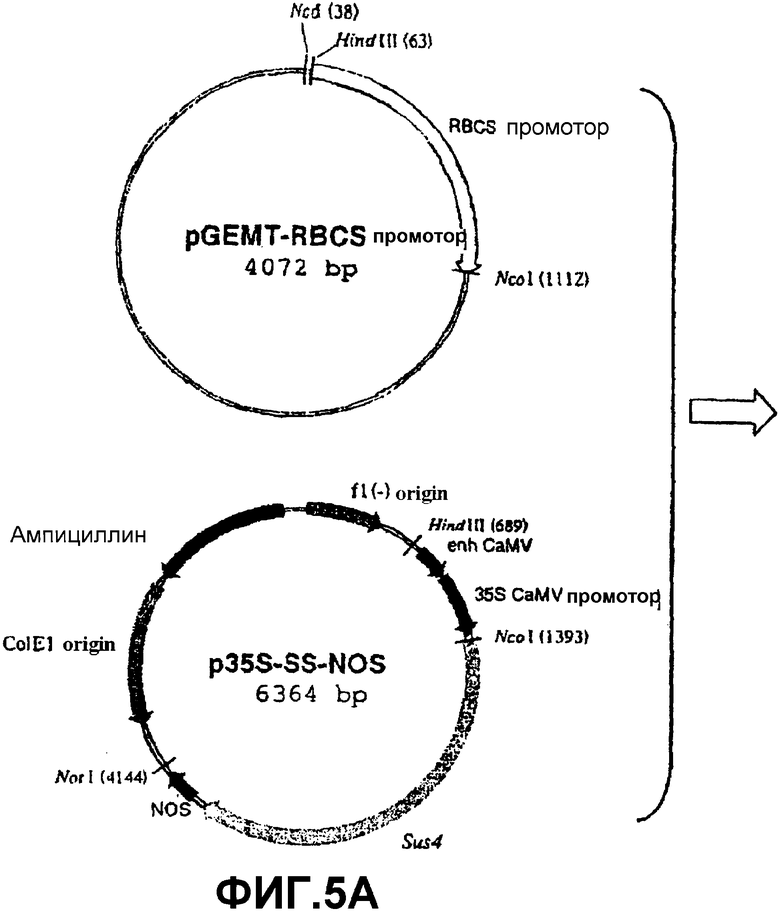
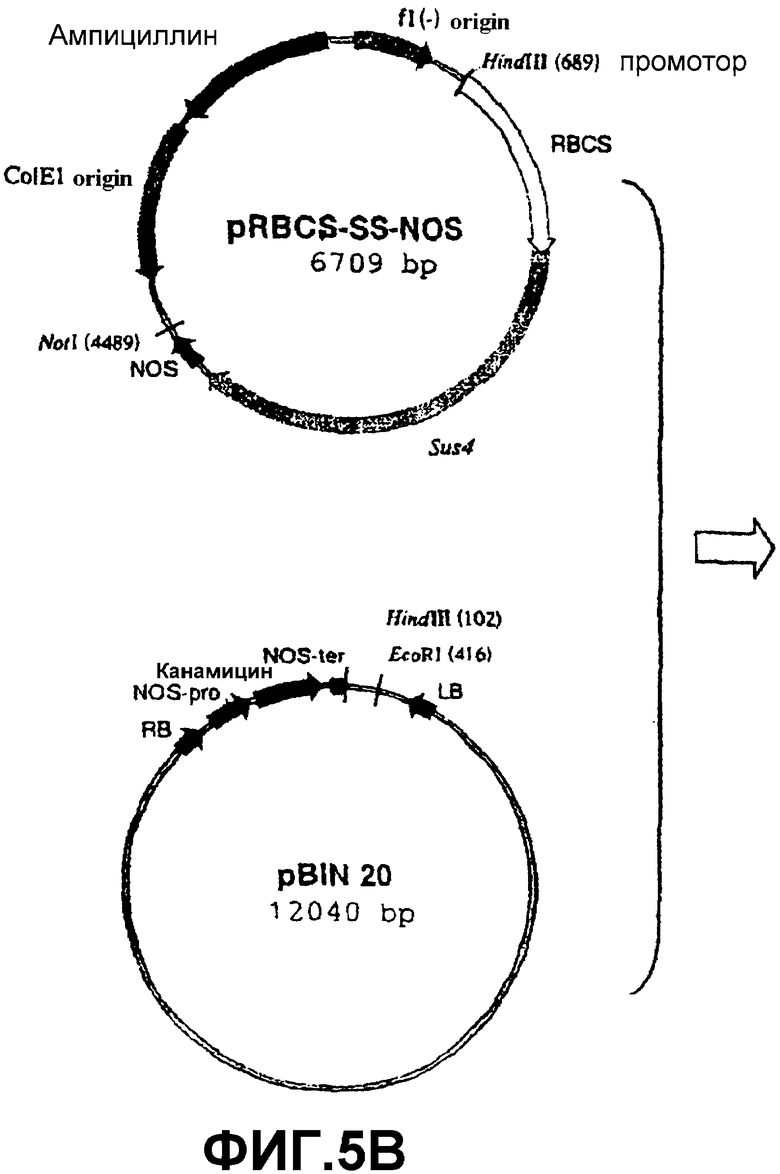
Для сверхэкспрессии SS специфически в освещенных листьях использовали ПЦР для амплификации промоторного района (обозначенного RBCS) гена, который кодирует малую субъединицу RUBISCO (рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы) табака (Barnes, S.A., Knight, J.S., Gray, J.C. (1994) Alteration of the amount of the chloropast phosphat translocator in transgenic tobacco affects the distribution of assimilate between starch and sugar. Plant Physiol. 106, 1123-1129). Эту нуклеотидную последовательность (которая сообщает специфическую экспрессию в фотосинтетически активных клетках) встраивали в вектор pGEMT-easy (Promega) с получением pGEMT-RBCSprom (фиг.5А). Эту конструкцию расщепляли HindIII и NcoI и высвобождаемый фрагмент клонировали в соответствующие сайты рестрикции p35S-SS-NOS с получением pRBCS-SS-NOS (фиг.5В). Эту конструкцию расщепляли последовательно HindIII, ДНК-полимеразой Т4 и NotI. Высвобождаемый фрагмент клонировали в плазмиду pBIN20, расщепленную последовательно HindIII, ДНК-полимеразой Т4 и EcoRI. Полученная конструкция была обозначена pBINRBCS-SS-NOS (фиг.5С).
После амплификации в E.coli (XL1 Blue) как pBIN35S-SS-NOS, так и pBINRBCS-SS-NOS встраивали в A. tumefaciens С58:GV2260 (Debleare, R., Rytebier, B., de Greve, H., Debroeck, F., Schell, J., van Montagu, M., Leemans, J. (1985) "Efficient octopine Ti plasmid-derived vectors of Agrobacterium mediated gene transfer, to plants" Nucl. Acids Res. 13, 4777-4788), которую использовали для трансформации таких видов, как томат (Lycopersicon esculentum), табак (Nicotiana tabacum), картофель (Solanum tuberosum) и рис, общепринятыми способами (Horsch, R.B., Fry, J.E., Hoffmann, N.L., Eichholtz, D., Rogers, S.G., Fraley, R.T. (1985) "A simple and general method for transferring genes into plants" Science 277, 1229-1231; Pozueta-Romero, J., Houlne, G., Schantz, R., Chamarro, J. (2001) "Enhanced regeneration of tomato and pepper seedling explants for Agrobacterium-mediated transformation" Plant Cell Tiss. Org. Cult. 67, 173-180; Hiei, Y., Ohta, S., Komari, Т., Kumashiro. T. (1994) "Efficient transformation of rice (Oryza saliva L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant (J. 6, 271-282). Штамм A. tumefaciens С58:GV2260, трансформированный pBIN35S-SS-NOS, был депонирован 29 октября 2003 года в Испанской Коллекции Типовых Культур, расположенной в Research Building of Valencia University, Burjassot Campus, Burjassot 46100 (Valencia, Spain) с номером депозита СЕСТ:5851.
Получение наборов для анализа для определения сахарозы
Один из наборов, предназначенных для определения сахарозы, показан в следующей Схеме I ферментативных реакций, проводимых с использованием этого набора, для спектрофотометрического/флуорометрического определения сахарозы, основанных на превращении сахарозы в сахаронуклеотид, и затем на превращении сахаронуклеотида в глюкозо-1-фосфат, глюкозо-6-фосфат и NAD(P)Н.
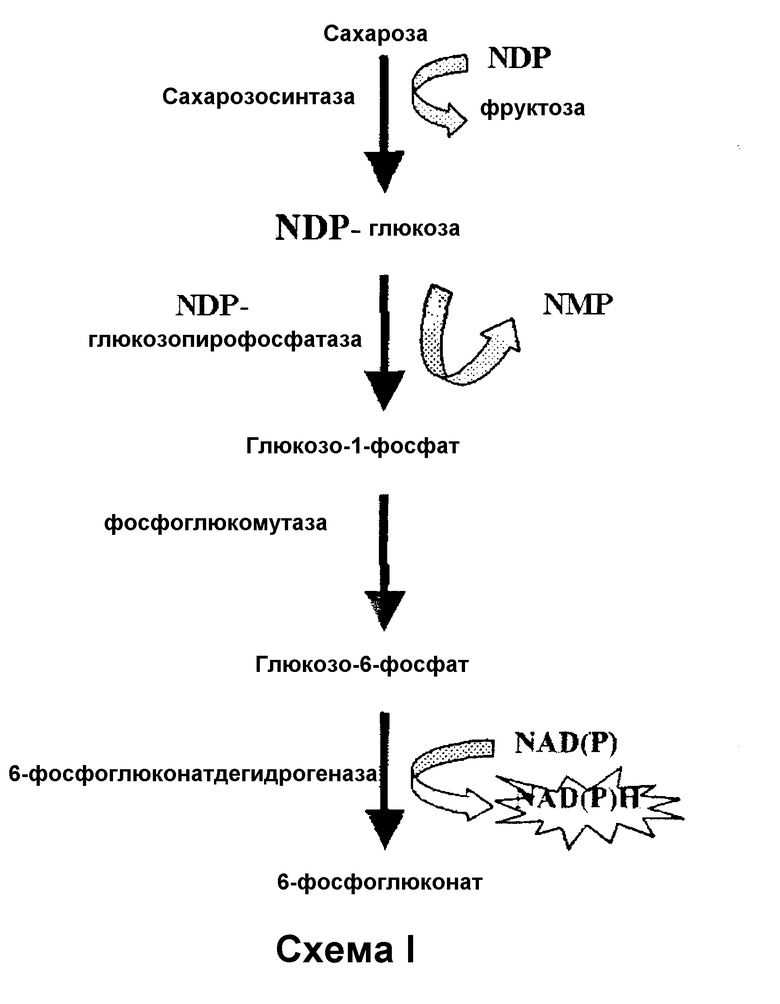
Этот набор основан на действии SS на молекулу сахарозы в присутствии нуклеотиддифосфата (например, UDP или ADP), с высвобождением эквимолярных количеств фруктозы и соответствующего сахаронуклеотида. Если сахаронуклеотидом, полученным из этой реакции, является UDPG, ее подвергают действию гидролитических ферментов UDPG, таких как UDPG-пирофосфатаза типа Nudix (EC 3.6.1.45) (Yagi, Т., Baroja-Fernández, E., Yamamoto, R., Munoz, F.J., Akazawa, Т., Pozueta-Romero, J. (2003) Cloning, expression and characterization of a mammalian Nudix hydrolase-like enzyme that cleaves the pyrophosphate bond of UDP-glucose. Biochem. J. 370, (409-415) or UDPG hydrolase (Burns, D.M., Beacham, I.R. (1986) Nucleotide sequence and transcriptional analysis of the E.coli ushA gene, encoding periplasmic UDP-sugar hydrolase (5′ - nucleotidase): regulation of the ushA gene, and the signal sequence of its encoded protein product. Nucl. Acids Res. 14, 4325-4342). G1P, высвобождаемый под действием этих гидролитических ферментов, превращается под действием фосфоглюкомутазы (PGM) с образованием глюкозо-6-фосфата (G6P), который, в свою очередь, может быть подвергнут реакции связывания с NAD(P)+под действием фермента G6P-дегидрогеназы (G6PDH) с образованием 6-фосфоглюконата и NAD(P)Н, который может быть легко определен флуориметрией и спектрофотометрией при 340 нм. В свою очередь, высвобождаемый NAD(P)Н может быть сопряжен с действием FMN-оксидоредуктазы/люциферазы с излучением света, который определяют количественно спектрофотометрически.
Альтернативно, как показано в Схеме II, полученная UDPG может быть сопряжена с UDPG-дегидрогеназой (ЕС 1.1.1.22), которая, в присутствии NAD, образует эквимолярные количества UDP-глюкуроната и NADН, который может быть определен флуорометрией или спектрофотометрически при 340 нм. В свою очередь, высвобождаемый NADН может быть сопряжен с действием FMN-оксидоредуктазы/люциферазы с обеспечением света, который определяют количественно спектрофотометрически.
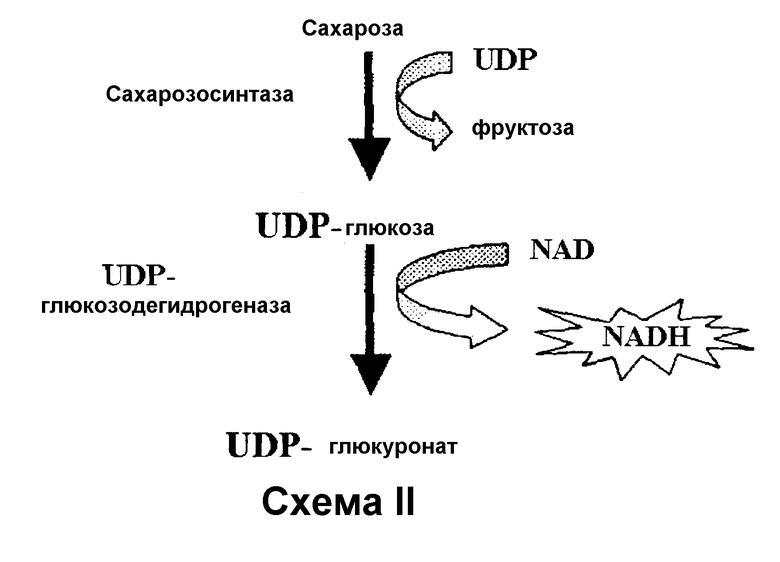
Если продуктом реакции, катализируемой SS, является ADPG, ее подвергают действию гидролитических ферментов ADPG, таких как бактериальная ADPG-пирофосфатаза (ЕС 3.6.1.21) (Moreno-Bruna, В., Baroja-Fernández, E., Muсoz, F.J., Bastarrica-Berasategui, A., Zandueta-Criado, A., Rodríguez-López, M., Lasa, I., Akazawa, Т., Pozueta-Romero, J. (2001) Adenosine diphosphate sugar pyrophosphatase prevents glycogen biosynthesis in Escherichia coli. Proc. Natl. Acad. Sci. USA 98, 8128-8132). Высвобождаемый G1P превращают под действием фосфоглюкомутазы с образованием глюкозо-6-фосфата (G6P), который, в свою очередь, может быть подвергнут реакции связывания с NAD(P)+под действием фермента G6P-дегидрогеназы с образованием 6-фосфоглюконата и NAD(P)Н, который может быть легко определен флуорометрией или спектрофотометрией при 340 нм.
В любом случае, схемы ферментативных реакций, связанных с образованием сахаронуклеотида (т.е. активированного нуклеотидом сахара), опосредованные SS, являются превосходным образом пригодными для применения к амперометрическому детектированию.
ПРИМЕРЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже описаны примеры, которые показывают подробно процедуру для клонирования кДНК, которая кодирует изоформу SS картофеля, в подходящем экспрессирующем векторе и в штамме E.coli, оптимизированном для получения и накопления этого фермента в его активной форме. Другие примеры описывают применение рекомбинантной SS для приготовления наборов для анализов для определения сахарозы в пробах растений, сыворотке, моче, фруктовых соках, подслащенных фруктовых напитках, прохладительных напитках и т.д. Другой пример описывает применение вариантов SS, оптимизированных для крупномасштабного получения сахаронуклеотидов, таких как UDPG и ADPG. Наконец, другой пример описывает получение растений с высоким содержанием сахарозы, ADPG и крахмала и высоким отношением амилоза/амилопектин вследствие высокой ADPG-продуцирующей активности в растениях, которые сверхэкспрессируют SS.
Пример 1: Экспрессия в Escherichia coli BLR (DE3) рекомбинантной SS с гистидиновым хвостом, которая может быть легко очищена и имеет высокую удельную активность.
На основе знания нуклеотидной последовательности гена SS4, который кодирует изоформу SS картофеля, можно создать два специфических праймера, последовательностями которых являются, в направлении 5′-3′, SEQ ID NO:1 и SEQ ID NO:2. С использованием этих праймеров ДНК-фрагмент, обозначенный SSX, амплифицировали обычными способами ПЦР из кДНК-библиотеки клубней картофеля и встраивали в плазмиду pSK Bluescript (Stratagene), которую амплифицировали в бактерии-хозяине XL1 Blue. Нуклеотидной последовательностью SSX является SEQ ID NO:3, которая незначительно отличается от SS4 (номер доступа GenBank U24087). Аминокислотная последовательность, расшифрованная из SEQ ID NO:3, незначительно отличается от SS4 и, следовательно, обозначена как SSX. Аминокислотной последовательностью, расшифрованной после экспрессии SEQ ID NO:3 в плазмиде pET-28a(+), является SEQ ID NO:4, которая включает в себя богатую гистидином последовательность из 38 аминокислот, слитую с аминоконцевой областью аминокислотной последовательности, расшифрованной из SEQ ID NO:3.
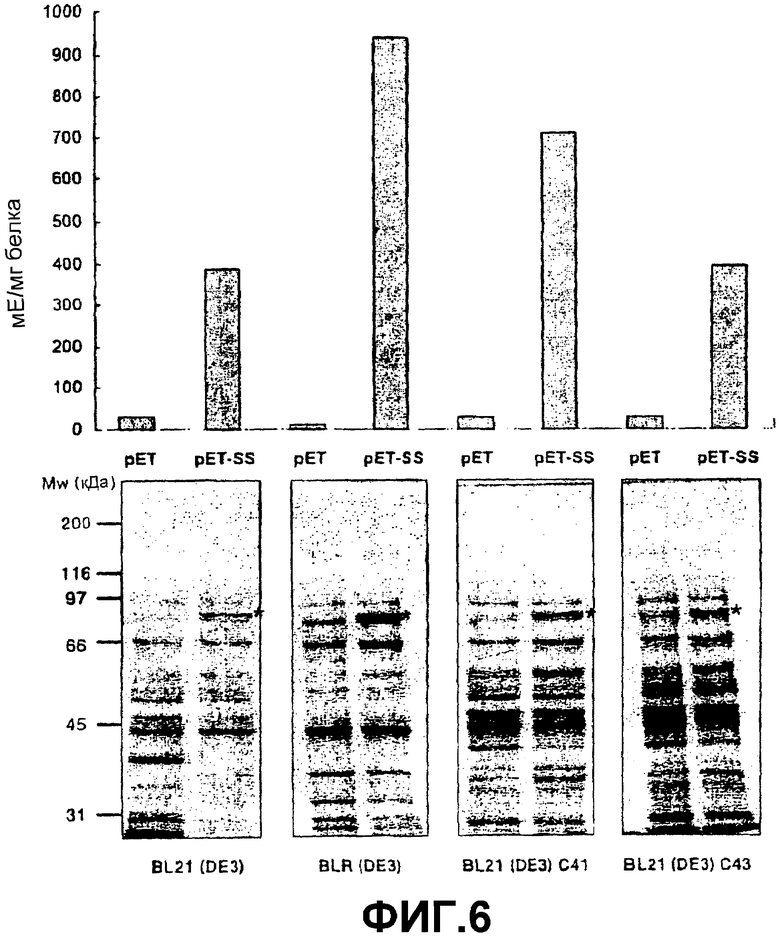
Получение SSX в бактериях BL21(DE3), трансформированных pET-SS, индуцировали добавлением 1 мМ IPTG. После шести дополнительных часов культивирования при 37оС наблюдали, что бактерии, трансформированные pET-SS, накапливали белок в агрегированной форме, размер которой соответствует SS. Однако эти бактерии не имели активности SS. Эта неспособность экспрессии активной формы SS может быть приписана трудностям, которые имеет E.coli в отношении правильной укладки некоторых эукариотических белков высокой молекулярной массы (Miroux, B., Walker, J.E. (1996) “Over-production of proteins in Escherichia coli: mutant hosts that allow synthesis of some membrane proteins and globular proteins at high levels” J. Mol. Biol. 260, 289-298). С целью преодоления этой проблемы исследовали способность получения активной SS в других бактериальных штаммах и при температуре 20оС. Во всех из них продуцирование SSX индуцировали добавлением 1 мМ IPTG. После шести часов дополнительной инкубации бактерии обрабатывали ультразвуком и центрифугировали. Полученный супернатант анализировали на активность SS. В этих условиях, как показано на фиг.6, штамм BLR(DE3) оказался наиболее эффективным с точки зрения продуцирования растворимой, активной SS. Штамм E.coli BLR(DE3) (Nivagen), трансформированный pET-SS, был депонирован 29 октября 2003 года в Испанской Коллекции Типовых Культур с номером депозита СЕСТ:5850. Вклад рекомбинантного SSX в общий пул белка СЕСТ:5850 составляет приблизительно 20%, в сравнении с очень низкой продуктивностью рекомбинантной SS (30 микрограммов на грамм бактерий), описанной в литературе (Nakai, Т., Tonouchi, N., Tsuchida, Т., Mori, H., Sakai, F., Hayashi, T. (1997) "Expression and characterization of sucrose synthase from mung bean seedlings in Escherichia coli" Biosci. Biotech. Biochem. 61, 1500-1503; Li, C.R., Zhang, X.B., Hew, C.S. (2003) "Cloning, characterization and expression analysis of a sucrose synthase gene from tropical epiphytic orchid Oncidium goldiana. Physiol. Plantarum 118, 352-360). Супернатант пропускали через аффинную колонку His-Bind (Novagen), в которой этот рекомбинантный белок, имеющий гистидиновый хвост, специфически удерживается. После элюции и диализа очищенной SS ее инкубируют с 50 мМ HEPES, рН 7,0 / 1 мМ ЭДТА / 20% полиэтиленгликоль / 1 мМ MgCl2 / 15 мМ KCl /2° мМ° UDP. Удельная активность, определенная в виде продуцирования UDPG, была 80 единиц/мг белка, что гораздо выше, чем активность 0,05-5 единиц/мг рекомбинантной SS, описанная в литературе (Nakai, Т., Tonouchi, N., Tsuchida, Т., Mori, H., Sakai, F., Hayashi, T. (1997) "Expression and characterization of sucrose synthase from mung bean seedlings in Escherichia coli" Biosci. Biotech. Biochem. 61, 1500-1503; Li, C.R., Zhang, X.B., Hew, C.S. (2003) "Cloning, characterization and expression analysis of a sucrose synthase gene from tropical epiphytic orchid Oncidium goldiana. Physiol. Plantarum 118, 352-360); Römer, U., Schrader, H., Günther, N., Nettelstroth, N., Frommer, W.B., Elling, L. (2004) Expression, purification and characterization of recombinant sucrose synthase I from Solanum tuberosum L. for carbohydrate engineering. J. Biotechnology 107, 135-149) и больше, чем 3 единицы/мг, что соответствует удельной активности SS, очищенной из экстрактов растений (Pressey R (1969) Potato sucrose synthase: purification, properties and changes in activity associated with maturation. Plant Physiol. 44, 759-764. Эта единица определяется как количество фермента, которое катализирует продуцирование одного микромоля UDPG в минуту. Аффинность в отношении UDP в присутствии 500 мМ сахарозы была Km (UDP)=0,25 мМ, в то время как Km в отношении сахарозы была 30 мМ в присутствии 1 мМ UDP. Эта аффинность в отношении сахарозы в присутствии UDP является незначительно более высокой, чем аффинность, которая обнаруживалась рекомбинантной SS, полученной в дрожжах (Km=95 мМ, Romer, U., Schrader, H., Gunther, N., Nettelstroth, N., Frommer, W.B., Elling, L. (2004) Expression, purification and characterization of recombinant sucrose synthase I from Solanum tuberosum L. for carbohydrate engineering. J. Biotechnology 107, 135-149).
Пример 2: Крупномасштабное получение UDPG и ADPG на основе использования рекомбинантной SS из E.coli
Три грамма UDPG высокой чистоты получали эффективно и экономично после инкубации в течение 12 часов при 37оС 100 миллилитров раствора, содержащего 1 М сахарозу, 50 мМ HEPES, рН 7,0 / 1 мМ ЭДТА / 20% полиэтиленгликоль / 1 мМ MgCl2 / 15 мМ KCl / 100 мМ UDP и 30 единиц рекомбинантной SS из картофеля, полученной после экспрессии pET-SS в BLR(DE3) и последующей очистки. Реакция завершалась после нагревания этого раствора при 100оС в течение 90 секунд и затем центрифугирования при 10000 g в течение 10 минут. Супернатант наносили на ВЭЖХ-хроматограф препаративного масштаба (Waters Associates) и UDPG очищали, как описано в литературе (Rodríguez-López, M., Baroja-Fernàndez, E., Zandueta-Criado, A., Pozueta-Romero, J. (2000) Adenosine diphosphate glucose pyrophosphatase: a plastidial phosphodiesterase that prevents starch biosynthesis. Proc. Natl. Acad. Sci. USA 97, 8705-8710).
Получение ADPG требовало генерирования мутированной формы SS с аффинностью в отношении ADP, гораздо большей, чем аффинность, описанная для SS, экстрагированной из тканей растений (Pressey R (1969) Potato sucrose synthase: purification, properties, and changes in activity associated with maturation. Plant Physiol. 44, 759-764; Nguyen-Quock, В., Krivitzky, M., Huber, S.C., Lecharny, A. (1990) Sucrose synthase in developing maize leaves. Plant Physiol. 94, 516-523; Morell, M., Copeland, L. (1985) Sucrose synthase of soybean nodules. Plant Physiol. 78, 149-154).
Эту изоформу, обозначенную SS5, получали точечным мутагенезом SSX с использованием набора для сайт-направленного мутагенеза QuickChange (Stratagene) и последовательного использования следующих пар праймеров, последовательностями которых являются [SEQ ID NO:5, SEQ ID NO:6], [SEQ ID NO:7, SEQ ID NO:8] и [SEQ ID NO:9, SEQ ID NO:10]. Полученной нуклеотидной последовательностью, обозначенной SS5, является SEQ ID NO:11. Изменения в аминокислотной последовательности SS5 (Susy 5) относительно SS4 - Susy 4 - (присутствующей в базах данных) показаны в виде затененных последовательностей в таблице I. Аминокислотной последовательностью, расшифрованной после экспрессии SEQ ID NO:11 в плазмиде pET-28a(+), является SEQ ID NO:12, которая включает в себя богатую гистидином последовательность из 38 аминокислот, слитую с аминоконцевой областью аминокислотной последовательности, расшифрованной из SEQ ID NO:11.
Таблица I включает указанную богатую гистидином последовательность из 38 аминокислот, слитую с амино-концевой частью SS5.
Таблица I

Рекомбинантная SS5, полученная после экспрессии pET-SS5, имела Vmax 80 единиц/мг белка и 65 единиц/мг белка в присутствии NDP и ADP, соответственно. Аффинности в отношении UDP и ADP в присутствии 500 мМ сахарозы были очень сходными (Km=0,2 мМ как для ADP, так и для UDP), в то время как Km в отношении сахарозы была 30 мМ и 100 мМ в присутствии насыщенных концентраций UDP и ADP, соответственно. Эти кинетические параметры являются очень отличающимися от кинетических параметров, описанных для SS, экстрагированной из клубня картофеля и других органов других видов, согласно которым Vmax этого фермента является в 10 раз более высокой в присутствии UDP, чем в присутствии ADP (Pressey R (1969) Potato sucrose synthase: purification, properties, and changes in activity associated with maturation. Plant Physiol. 44, 759-764; Morell, M., Copeland, L. (1985) Sucrose synthase of soybean nodules. Plant Physiol. 78, 149-154; Nguyen-Quock, В., Krivitzky, M., Huber, S.C., Lecharny, A. (1990) Sucrose synthase in developing maize leaves. Plant Physiol. 94, 516-523). Штамм E.coli XL1 Blue, трансформированный pSS5, был депонирован в Испанской Коллекции Типовых Культур с номером депозита СЕСТ:5849.
Три грамма ADPG высокой чистоты получали эффективно и экономично после инкубации в течение 12 часов при 37оС 100 миллилитров раствора, содержащего 1 М сахарозу, 50 мМ HEPES, рН 7,0 / 1 мМ ЭДТА / 20% полиэтиленгликоль / 1 мМ MgCl2 / 15 мМ KCl / 100 мМ ADP и 30 единиц рекомбинантной SS из картофеля, полученной после экспрессии pET-SS в BLR(DE3) и последующей очистки в колонке His-bind. Реакция завершалась после нагревания этого раствора при 100оС в течение 90 секунд и затем центрифугирования при 10000 g в течение 10 минут. Супернатант наносили на ВЭЖХ-хроматограф препаративного масштаба (Waters Associates) для очистки ADPG.
Пример 3: Получение ферментных наборов для определения сахарозы
Для определения сахарозы готовили следующие реакционные смеси (коктейли) со следующими компонентами и конечными количествами/концентрациями:
1. НАБОРЫ, ОСНОВАННЫЕ НА ИСПОЛЬЗОВАНИИ ГИДРОЛИТИЧЕСКИХ ФЕРМЕНТОВ САХАРОНУКЛЕОТИДОВ:
а. 2 единицы SS (рекомбинантной или нерекомбинантной)
b. 2 мМ ADP или UDP (в зависимости от того, получают ли ADPG или UDPG, соответственно)
с. 2 единицы ADPG-пирофосфатазы или 2 единицы UDPG-пирофосфатазы (в зависимости от того, подлежат включению в реакционную смесь ADP или UDP, соответственно)
d. 2 единицы PGM
e. 2 единицы G6PDH
f. 0,5 мМ NAD(P)
g. реакционный буфер: 50 мМ HEPES, рН 7,0 / 1 мМ ЭДТА / 20% полиэтиленгликоль / 1 мМ MgCl2 / 15 мМ KCl
h. предварительно отфильтрованная тест-проба.
2. НАБОР, ОСНОВАННЫЙ НА ИСПОЛЬЗОВАНИИ UDPG-ДЕГИДРОГЕНАЗЫ
а. 2 единицы SS (рекомбинантной или нерекомбинантной)
b. 2 мМ UDP
с. 2 единицы UDPG-дегидрогеназы
d. 0,5 мМ NAD
е. реакционный буфер: 50 мМ HEPES, рН 7,0 / 1 мМ ЭДТА / 20% полиэтиленгликоль / 1 мМ MgCl2 / 15 мМ KCl
h. предварительно отфильтрованная тест-проба.
Определение количества сахарозы, присутствующей в тест-пробе, основано на флуорометрическом определении или спектрофотометрическом определении (при 340 нм) NAD(P)Н, полученного в соответствии с сопряженными реакциями, показанными в схемах I и II.
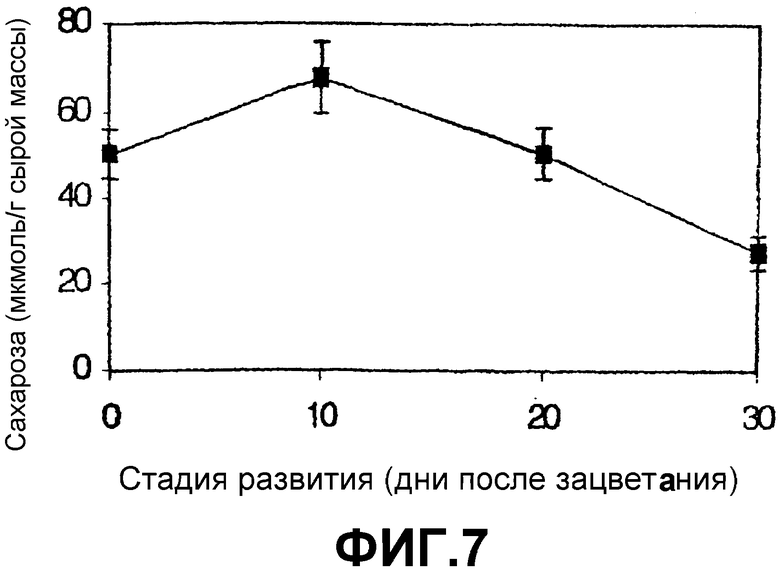
Для определения содержания сахарозы семян ячменя на различных стадиях развития (фиг.7) эти реакции выполняли в лунках 300 микролитров планшета ELISA в течение 3 минут при 37оС. Объем тест-пробы составлял 20 микролитров, а объем реакционной смеси (коктейля), полученной из комбинации реагентов a-g (набор №1) и а-е (набор №2), составлял 280 микролитров. Контроли содержали все эти компоненты смесей, за исключением SS. Измерение проводили с использованием спектрофотометра MultiScan. Было обнаружено, что величины, полученные как с набором типа «1», так и с набором типа «2», были сравнимы с величинами, определенными с использованием хроматографических способов, описанных во введении (Baroja-Fernández, E., Muсoz, F.J., Saikusa, Т., Rodríguez-López, M., Akazawa, Т., Pozueta-Romero, J. (2003) Sucrose synthase catalyzes the de novo production of ADPglucose linked to starch biosynthesis in heterotrophic tissues of plants. Plant Cell Physiol. 44, 500-509).
Пример 4: Получение трансгенных растений, которые сверхэкспрессирует SS.
Фигуры 8-10 представляют результаты, полученные для листьев растений картофеля, которые сверхэкспрессируют SS как конститутивно (35S-SS-NOS), так и специфически (RBCS-SS-NOS).
Как показано на фиг.8, SS-активность в листьях любого из этих растений является в 2-10 раз более высокой, чем в том же самом органе растения дикого типа (WT). Эти листья имели следующие свойства:
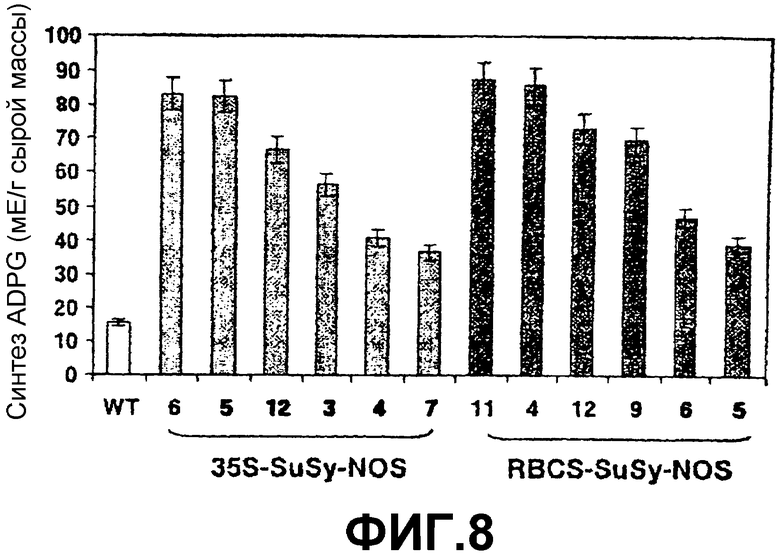
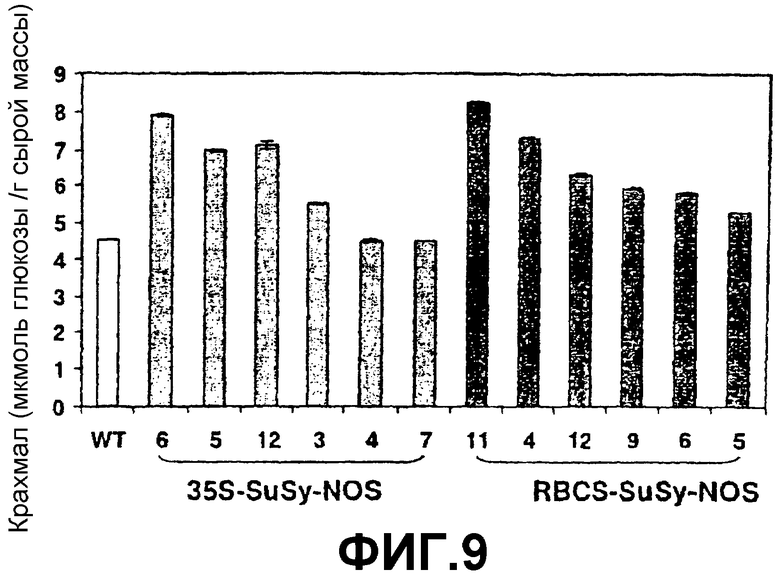
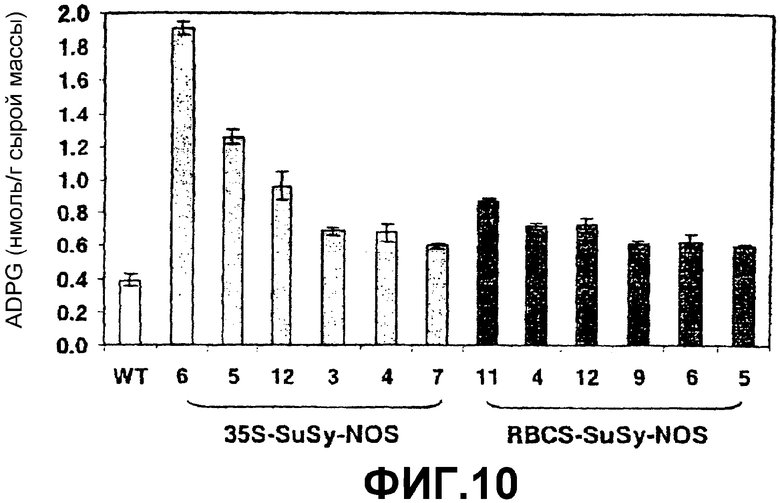
1. Явная корреляция между ADPG-продуцирующей активностью SS (фиг.8) и уровнями крахмала (фиг.9) и ADPG (фиг.10). Это свойство наблюдали не только в листьях, но также в запасающих тканях, таких как клубни и семена (см. ниже).
2. Высокое содержание крахмала (фиг.9) относительно листьев растений дикого типа. Например, содержание крахмала листа растения картофеля «дикого типа», выращиваемого при фотопериоде 8 часов света/16 часов темноты и при 20оС, равно 5 микромоль/грамм сырой массы, в то время как лист трансгенного растения, которое сверхэкспрессирует SS, имел содержание крахмала 8 микромоль/грамм сырой массы. Различия между растениями дикого типа и трансгенными растениями становятся более явными, когда фотопериод является продолжительным, так что листья растения, которое сверхэкспрессирует SS, содержат в 4 раза больше крахмала, чем листья растения дикого типа.
3. Высокое содержание ADPG относительно той же самой ткани или того же самого органа нетрансформированного растения (фиг.10). Среднее содержание в листе растения картофеля дикого типа, выращиваемого при фотопериоде 8 часов света/16 часов темноты и при 20оС, равно 0,35 наномоль/грамм сырой массы, в то время как листья растений, которые сверхэкспрессируют SS, могут иметь содержание 2,5 наномоль/грамм сырой массы.
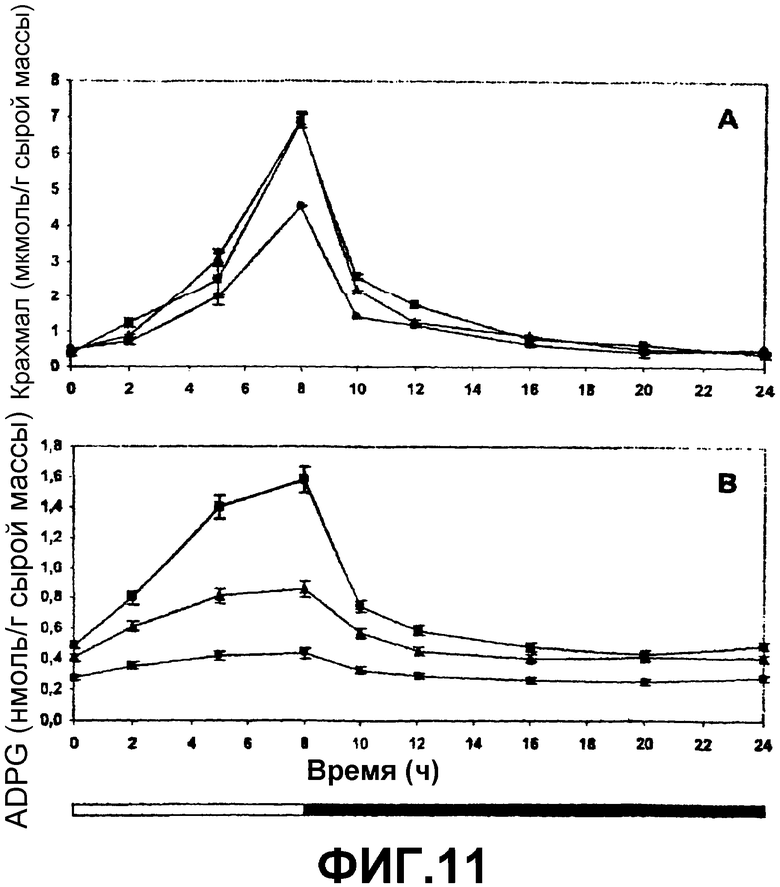
4. Как ADPG, так и крахмал обнаруживают транзиторное (временное) накопление на протяжении этого фотопериода (фиг.11). Скорость накопления обоих веществ сохраняет положительную корреляцию с активностью SS, что свидетельствует о том, что в противоположность тому, что предполагается «классической» моделью биосинтеза крахмала (фиг.2А), и в подтверждение гипотезы «альтернативной» модели, показанной на фиг.2В, SS играет фундаментальную роль в образовании ADPG и в связи между метаболизмом сахарозы и метаболизмом крахмала.
5. Нормальные уровни растворимых сахаров, таких как глюкоза и фруктоза. Однако уровни глюкозо-6-фосфата и сахарозы в трансгенных листьях являются более высокими, чем уровни этих соединений в листьях картофеля дикого типа (таблица 2).
фосфат
фосфат
6. Внешняя морфология растений, которые сверхэкспрессируют SS, не отклоняется от нормы, в сравнении с внешней морфологией нетрансформированного растения.
Фиг.12-14 показывают результаты, полученные в клубнях картофеля, которые сверхэкспрессируют SS конститутивно (35S-SS-NOS). Эти результаты являются по существу идентичными результатам, наблюдаемым в клубнях, которые сверхэкспрессируют SS под контролем специфического для клубней промотора (промотора гена patatina).
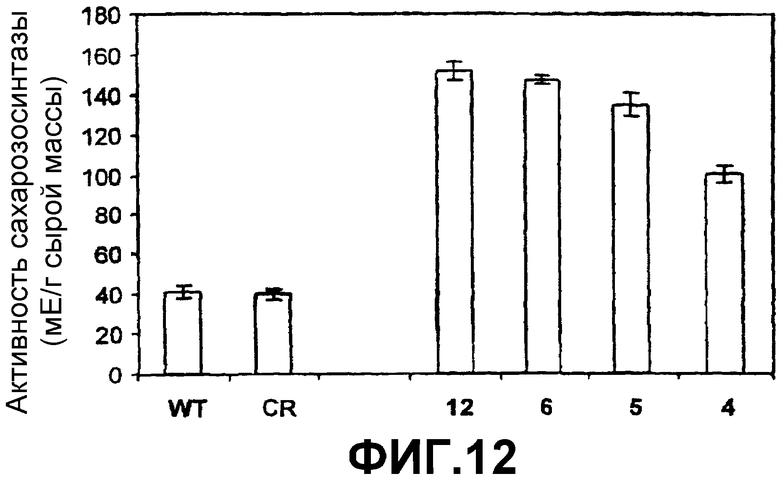
Как показано на фиг.12, активность SS в клубнях любого из этих растений является более высокой, чем в том же самом органе растения дикого типа. Эти клубни имели следующие свойства:
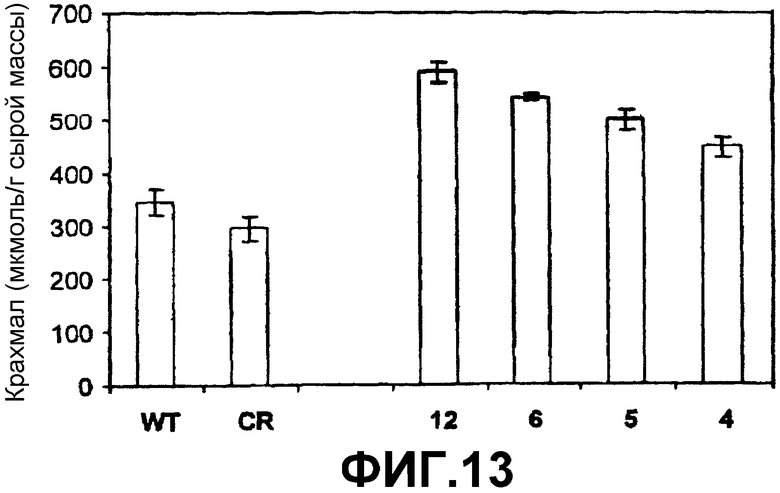
1. Явная корреляция между ADPG-продуцирующей активностью SS (фиг.12) и уровнями крахмала (фиг.13) и ADPG (фиг.14).
2. Высокое содержание крахмала (фиг.13) относительно клубней нетрансформированных растений. Например, содержание крахмала в клубне растения «дикого типа» равно приблизительно 300 микромоль/грамм сырой массы (эквивалентно 54 мг крахмала на грамм сырой массы), в то время как клубень, который сверхэкспрессирует SS, имеет 450-600 микромоль/грамм сырой массы.
3. Высокое содержание ADPG относительно клубней растений дикого типа (фиг.14). Среднее содержание в клубне дикого типа равно 5 наномоль/грамм сырой массы, в то время как клубни, которые сверхэкспрессируют SS, могут иметь содержание 7-9 наномоль/грамм сырой массы.
Результаты, полученные в семенах риса, листьях томата и табака, а также в плодах томата, являются качественно сходными с результатами, показанными на фиг.8-14. Во всех случаях было увеличение содержания крахмала и увеличение отношения амилоза/амилопектин.
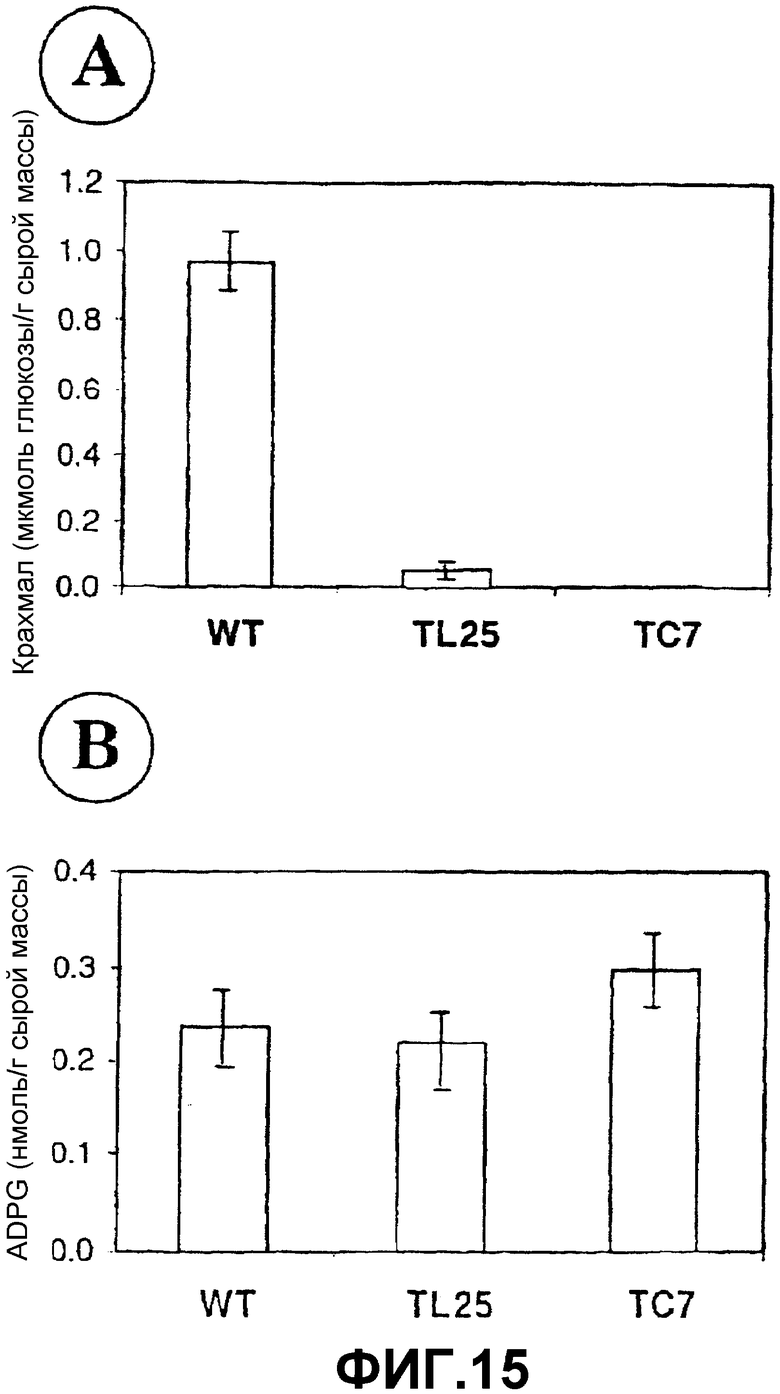
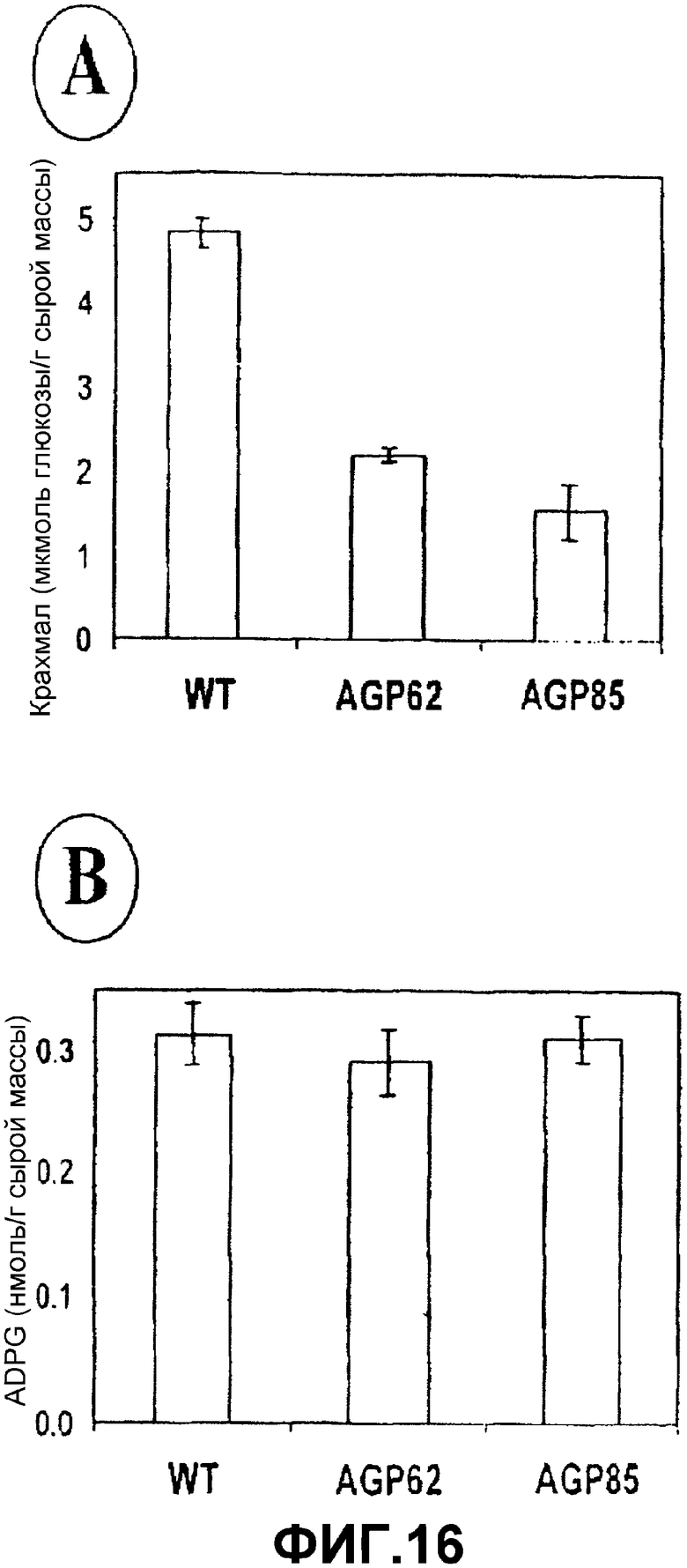
Получение растений с высоким содержанием ADPG и крахмала после сверхэкспрессии SS является результатом, который является полностью неожиданным в соответствии с существующими идеями в отношении биосинтеза крахмала (иллюстрированного на фиг.1А и 2А) и, возможно, объясняет, почему конструирование растений, которые сверхэкспрессируют SS, не принималось ранее в качестве стратегии для увеличения образования крахмала. Результаты, полученные на основании этого исследования, предполагают, что SS, а не AGPаза является фундаментальным источником ADPG, которая накапливается в растениях. Согласно моделям, которые все еще существуют, AGPаза является единственным источником ADPG. Однако удивительным образом, уровни ADPG никогда не исследовались в недостаточных в отношении AGPазы растениях. Для исследования значимости данного изобретения авторы изобретения впервые анализировали уровни ADPG и крахмала в растениях Arabidopsis и картофеля с пониженной активностью AGPазы. Как показано на фиг.15А, уровни крахмала в AGPаза-недостаточных растениях TL25 Arabidopsis (Lin, T.P., Caspar, T., Somerville, C.R., Preiss, J. (1988) Isolation and characterization of a starchless mutant of Arabidopsis thaliana lacking ADPglucose pyrophosphorylase activity. Plant Physiol. 88, 1131-1135) являются более низкими, чем уровни, наблюдаемые в растениях WT. Однако уровни ADPG являются нормальными (фиг.15 В). В противоположность этому, уровни крахмала в растениях картофеля AGP62 и AGP85 (Müller-Röber, B., Sonnewald, U., Willmitzer, L. (1992) Inhibition of the ADPglucose pyrophosphorylase in transgenic potatoes leads to sugar-storing tubers and influences tuber formation and expression of tuber storage protein genes, EMBO J. 11, 1229-1239) являются уменьшенными относительно уровней, наблюдаемых в листьях растений дикого типа (фиг.16А). Однако уровни ADPG являются полностью нормальными (фиг.16В). Взятые вместе, эти наблюдения (а) показывают, что SS, а не AGPаза, является основным источником ADPG в растениях, и (b) выдвигают на первый план значимость изобретения авторов после демонстрации, что сверхэкспрессия SS приводит к растениям с высоким содержанием крахмала.
ОПИСАНИЕ ДИАГРАММ
Фиг.1: Механизмы биосинтеза крахмала в гетеротрофных органах. (А) «Классический» механизм, в соответствии с которым SS участвует в образовании UDPG, которая в конечном счете превращается в крахмал после объединенного действия UDPG-пирофосфорилазы (UGP-азы), цитозольной фосфоглюкомутазы (PGM), пластидной фосфоглюкомутазы, ADPG-пирофосфорилазы (AGPазы) и крахмалсинтазы. (В) «Альтернативный» механизм, в соответствии с которым SS участвует в прямом образовании ADPG в цитозоле. Затем эта ADPG транспортируется в амилопласт при помощи транслокатора (переносчика). После попадания ADPG в амилопласт крахмалсинтаза использует ADPG для образования крахмала.
Фиг.2: Механизмы биосинтеза крахмала в листьях. (А) «Классический» механизм, в соответствии с которым весь процесс биосинтеза крахмала происходит внутри хлоропласта. Согласно этой точке зрения метаболизм крахмала и сахарозы не являются связанными. Кроме того, SS не принимает участия в процессе глюконеогенеза. (В) «Альтернативный» механизм биосинтеза крахмала, в соответствии с которым SS участвует в прямом образовании ADPG в цитозоле. Затем эта ADPG транспортируется во внутреннее пространство пластиды, где крахмалсинтаза использует ее в качестве субстрата для реакции синтеза крахмала.
Фиг.3: Стадии в конструировании экспрессионной плазмиды pET-SS из pET-28a(+) и pSS.
Фиг.4: Стадии в конструировании экспрессионной плазмиды pBIN35S-SS-NOS из pBIN20 и p35S-SS-NOS.
Фиг.5: Стадии в конструировании экспрессионной плазмиды pRBCS-SS-NOS из pGEMT-RBCSprom, p35S-SS-NOS и pBIN20.
Фиг.6: Экспрессия pET-SS в различных штаммах Escherichia coli. (А) Активность SS (в миллиединицах (мЕ) на миллиграмм бактериального белка) в бактериальных экстрактах, трансформированных pET или pET-SS. Реакция происходила в направлении деградации сахарозы и образования ADPG. Реакционная смесь содержала 50 мМ HEPES (рН 7,0), 1 мМ ЭДТА, 20% полиэтиленгликоль, 1 мМ MgCl2, 15 мМ KCl и 2 мМ ADP. Реакция происходила в течение 10 минут при 37оС. (В) Электрофорез в ДСН-ПААГ белковых экстрактов из различных E.coli, трансформированных pET и pET-SS. Положение рекомбинантной SSX указано звездочкой.
Фиг.7: Определение сахарозы на различных стадиях развития эндосперма ячменя с использованием набора на основе сопряженных реакций SS, ADPG- (UDPG-) пирофосфатазы, PGM и G6PDH. Эти результаты были идентичными результатам, полученным параллельно (а) с использованием набора на основе сопряженных реакций SS и UDPG-дегидрогеназы и (b) с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ) с амперометрическим детектированием в системе DX-500 Dionex, соединенной с колонкой Carbo-Pac PA1.
Абсцисса: Дни после зацветания
Ордината: Содержание сахарозы (мкмоль/г сырой массы)
Фиг.8: Активность SS в листьях растений картофеля дикого типа (WT) и растениях картофеля, которые сверхэкспрессируют SSX после интеграции конструкций 35S-SS-NOS (при помощи штамма Agrobacterium tumefaciens СЕСЕ:5851) или RBCS-SS-NOS в их геном. Активность выражена в миллиединицах (мЕ) на грамм сырой массы. Эта единица определяется как количество SS, требуемое для образования одного микромоля ADPG в минуту.
Фиг.9: Содержание крахмала в листьях растений картофеля дикого типа (WT) и растениях картофеля, которые сверхэкспрессируют SSX после интеграции конструкций 35S-SS-NOS (при помощи штамма Agrobacterium tumefaciens СЕСЕ:5851) или RBCS-SS-NOS в их геном.
Фиг.10. Содержание ADPG в листьях растений картофеля дикого типа (WT) и растениях картофеля, которые сверхэкспрессируют SSX после интеграции конструкций 35S-SS-NOS (при помощи штамма Agrobacterium tumefaciens СЕСЕ:5851) или RBCS-SS-NOS в их геном.
Фиг.11: Транзиторное накопление (А) крахмала и (В) ADPG во время фотопериода 8 часов света и 16 часов темноты в листьях растений WT (•), 35S-SS-NOS (■) и RBCS-SS-NOS (▲).
Фиг.12: Активность SS (отнесенная к сырой массе, FW) в клубнях растений картофеля дикого типа (WT), регенерационных контролях (RG) и растениях картофеля, которые сверхэкспрессируют SSX (линии 4, 5, 6 и 12) после интеграции конструкции 35S-SS-NOS в их геном (при помощи штамма Agrobacterium tumefaciens СЕСЕ:5851). Активность выражена в миллиединицах (мЕ) на грамм сырой массы. Эта единица определяется как количество SS, требуемое для образования одного микромоля ADPG в минуту.
Фиг.13: Содержание крахмала (отнесенное к сырой массе, FW) в клубнях растений картофеля дикого типа (WT), регенерационных контролях (RG) и растениях картофеля, которые сверхэкспрессируют SSX (линии 4, 5, 6 и 12) после интеграции конструкции 35S-SS-NOS в их геном (при помощи штамма Agrobacterium tumefaciens СЕСТ:5851).
Фиг.14: Содержание ADPG (отнесенное к сырой массе, FW) в клубнях растений картофеля дикого типа (WT) и растениях картофеля, которые сверхэкспрессируют SSX после интеграции конструкции 35S-SS-NOS в их геном (при помощи штамма Agrobacterium tumefaciens СЕСТ:5851).
Фиг.15: Содержание (А) крахмала и (В) ADPG в листьях AGPаза-недостаточного Arabidopsis thaliana TL25.
Фиг.16: Содержание (А) крахмала и (В) ADPG в листьях AGPаза-недостаточного картофеля AGP62 и AGP85.










| название | год | авторы | номер документа |
|---|---|---|---|
| КАРТОФЕЛЬ С ПОВЫШЕННЫМ ВЫХОДОМ КРАХМАЛА В ПЕРЕСЧЕТЕ НА ОТДЕЛЬНОЕ РАСТЕНИЕ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2004 |
|
RU2289914C2 |
| КАРТОФЕЛЬ С ПОВЫШЕННЫМ ВЫХОДОМ КРАХМАЛА В ПЕРЕСЧЕТЕ НА ОТДЕЛЬНОЕ РАСТЕНИЕ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2006 |
|
RU2326527C2 |
| СПОСОБЫ МОДУЛЯЦИИ ПРОВОДИМОСТИ УСТЬИЦА И РАСТИТЕЛЬНЫЕ ЭКСПРЕССИОННЫЕ КОНСТРУКЦИИ ДЛЯ ИХ ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2664461C2 |
| СПОСОБ УВЕЛИЧЕНИЯ УРОЖАЙНОСТИ РАСТЕНИЯ, СПОСОБ КОНТРОЛИРОВАНИЯ РЕЖИМА ЦВЕТЕНИЯ РАСТЕНИЯ И КОНСТРУКЦИЯ ДНК ДЛЯ ИХ ОСУЩЕСТВЛЕНИЯ | 1998 |
|
RU2241753C2 |
| ИНКАПСУЛИРОВАНИЕ ПОЛИПЕПТИДОВ В МАТРИЦУ КРАХМАЛА | 1997 |
|
RU2268301C9 |
| ТЕРМОСТАБИЛЬНЫЕ МУТАНТЫ ФЕРМЕНТОВ БИОСИНТЕЗА КРАХМАЛА | 2002 |
|
RU2303633C2 |
| СПОСОБ ВЫЯВЛЕНИЯ СОБЫТИЙ РЕДАКТИРОВАНИЯ ГЕНА SSIIA У ЗЕРНОВЫХ КУЛЬТУР С ПОМОЩЬЮ НАБОРА ОЛИГОНУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2023 |
|
RU2839687C1 |
| СПОСОБ СОЗДАНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ С ВЫСОКИМ УРОВНЕМ ЭКСПРЕССИИ ТРАНСГЕННОГО БЕЛКА | 2011 |
|
RU2507736C2 |
| ПАТОГЕН-ИНДУЦИРУЕМЫЙ СИНТЕТИЧЕСКИЙ ПРОМОТОР | 2007 |
|
RU2406761C2 |
| СПОСОБ РЕГУЛИРОВАНИЯ КЛУБНЕОБРАЗОВАНИЯ И ПРОДУКТИВНОСТИ РАСТЕНИЙ КАРТОФЕЛЯ В УСЛОВИЯХ ГИДРОПОНИКИ | 2017 |
|
RU2660918C1 |
Изобретение относится к биотехнологии. Описано получение трансгенных растений, которые сверхэкспрессируют ген SS, либо конститутивно, либо в листьях, либо в запасающих органах и которые имеют высокое содержание (как в листьях, так и в запасающих тканях) сахарозы, ADPG, G6P и крахмала в результате высокой ADPG-синтезирующей активности SS. Раскрыто трансгенное растение, полученное описанным способом. Изобретение позволяет получать трансгенные растения, имеющие высокое содержание (как в листьях, так и в запасающих тканях) сахарозы, ADPG, G6P и крахмала. 2 н. и 9 з.п. ф-лы, 16 ил., 2 табл.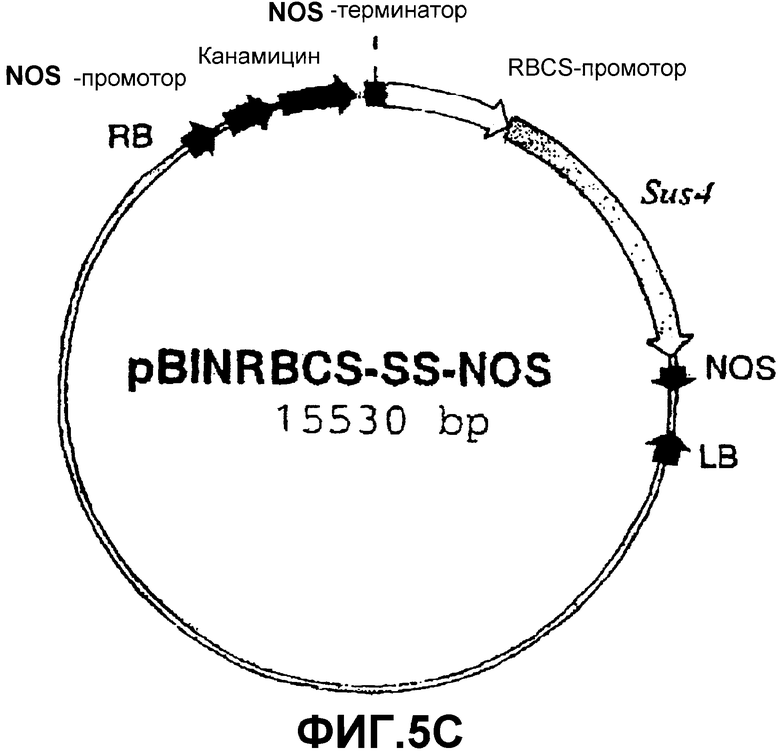
1. Способ получения трансгенных растений, который включает трансформирование или трансфицирование соответствующих растений дикого типа генетической конструкцией, содержащей нуклеотидную последовательность, выбранную из SEQ ID NO:3 или SEQ ID NO:11, которая сверхэкспрессирует фермент сахарозосинтазы, вовлеченный в непосредственную продукцию ADPG.
2. Способ по п.1, в котором экспрессия фермента сахарозосинтазы является в 2-10 раз более высокой, чем экспрессия, наблюдаемая в том же органе растения дикого типа.
3. Способ по п.2, отличающийся тем, что он предусматривает следующие стадии:
a) последовательное встраивание в плазмиду pSS промотора 35S и терминатора NOS в 5′- и 3′-районы соответственно, гена SSX или любой другой версии, которая кодирует SS, для получения плазмиды p35S-SS-NOS, рестрикционная карта которой показана на фиг.4В,
b) последовательное расщепление p35S-SS-NOS ферментами NotI, ДНК-полимеразой Т4 и HindIII,
c) клонирование полученного фрагмента в бинарной плазмиде pBIN20, предварительно расщепленной последовательно EcoRI, ДНК-полимеразой Т4 и HindIII, с получением плазмиды pBIN35S-SS-NOS, показанной на фиг.4С,
d) амплификацию pBIN35S-SS-NOS в E.coli (XL1 Blue),
e) встраивание генетической конструкции, амплифицированной на предыдущей стадии, в Agrobacterium tumefaciens C58:GV2260 с получением трансформированного штамма СЕСТ 5851,
f) трансфекцию растений трансформированным штаммом СЕСТ 5851.
4. Способ по п.2, отличающийся тем, что он предусматривает следующие стадии:
a) последовательное встраивание в плазмиду pGEMT промотора гена, который кодирует малую субъединицу RUB I SCO, для получения плазмиды pGEMT-RBCSprom, рестрикционная карта которой показана на фиг.5А,
b) расщепление pGEMT-RBCS ферментами HindIII и Ncol для встраивания фрагмента, высвобожденного в соответствующих сайтах рестрикции p35S-SS-NOS, с получением pRBCS-SS-NOS, рестрикционная карта которой показана на фиг.5В,
c) последовательное расщепление pRBCS-SS-NOS ферментами HindIII, ДНК-полимеразой Т4 и NotI и клонирование фрагмента, высвобожденного в pBIN20, расщепленной последовательно ферментами HindIII, ДНК-полимеразой Т4 и EcoRI, с получением pBINRBCS-SS-NOS, рестрикционная карта которой показана на фиг.5С,
d) амплификацию pBINRBCS-SS-NOS в E.coli (XL1 Blue),
e) встраивание генетической конструкции, амплифицированной на предыдущей стадии, в Agrobacterium tumefaciens C58:GV2260 и трансфекцию растений этим трансформированным штаммом.
5. Трансгенные растения, трансформированные или трансфицированные генетической конструкцией, содержащей нуклеотидную последовательность, выбранную из SEQ ID NO:3 или SEQ ID NO:11, которая сверхэкспрессирует фермент сахарозосинтазы, вовлеченный в непосредственную продукцию ADPG, листья и запасающие ткани которых имеют более высокие уровни ADPG и, соответственно, обогащенного амилозой крахмала, чем уровни, наблюдаемые в тех же самых листьях и запасающих тканях соответствующих растений дикого типа, выращенных в идентичных условиях.
6. Трансгенные растения по п.5, отличающиеся тем, что их листья содержат по меньшей мере в 1,6 раза больше крахмала, чем листья растения дикого типа.
7. Трансгенные растения по п.5, отличающиеся тем, что их листья содержат по меньшей мере в 7,1 раза больше ADPG, чем листья растения дикого типа.
8. Трансгенные растения по п.5, отличающиеся тем, что их клубни содержат по меньшей мере в 1,5 раза больше крахмала, чем клубни растения дикого типа.
9. Трансгенные растения по п.5, отличающиеся тем, что их клубни содержат по меньшей мере в 1,4 раза больше ADPG, чем клубни растения дикого типа.
10. Трансгенные растения по п.5, отличающиеся тем, что экспрессия фермента сахарозосинтазы является в 2-10 раз более высокой, чем экспрессия, наблюдаемая в том же органе растения дикого типа.
11. Трансгенные растения по п.5, отличающиеся тем, что они предпочтительно выбраны из растений табака, картофеля, томата или риса.
| СПОСОБ ИЗВЛЕЧЕНИЯ НЕФТИ ИЗ НЕФТЯНОГО РЕЗЕРВУАРА (ВАРИАНТЫ) | 1989 |
|
RU2067662C1 |
| WO 9803637 A1, 29.01.1998 | |||
| WO 0245485 А1, 13.06.2002 | |||
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОГО РАСТЕНИЯ КУКУРУЗЫ | 1991 |
|
RU2114911C1 |

