Область техники, к которой относится изобретение
Настоящее изобретение относится к области генной инженерии растений и биотехнологии. Оно может быть использовано для создания трансгенных растений с высоким уровнем экспрессии целевого гена. Это, в свою очередь, может применяться как для улучшения важных признаков ряда сельскохозяйственных культур, так и для биотехнологической наработки конкретных белков.
Уровень техники
Все большее применение в биотехнологической и сельскохозяйственной промышленности находят растения, свойства которых изменены посредством генетической трансформации. В настоящее время известны эффективные системы трансформации растений, с использованием которых можно создавать растения с необходимыми признаками. Из всех этих методов наибольшее практическое применение в настоящее время получили агробактериальный и биолистический методы доставки ДНК в геном растений. Агробактериальный метод трансформации растений - это использование в качестве векторов (переносчиков ДНК) бактерий A.tumefaciens. Он основан на способности патогенного организма Agrobacterium переносить из своей Ti-плазмиды в растительные клетки определенные фрагменты ДНК. Суть биолистического метода состоит в том, что металлическим частицам диаметром 1-4 мкм, покрытым ДНК, придается определенная скорость, благодаря чему они проникают через стенки интактных клеток и вносят в них молекулы ДНК. В обоих случаях целевой ген и связанные с ним последовательности вводятся в растительную клетку и стабильно интегрируются в растительный геном. Из этой клетки впоследствии образуется целое, полностью функционирующее растение с необходимыми свойствами.
Однако получение и последующее использование трансгенных растений сталкивается с проблемой достижения эффективной экспрессии белков в растениях из-за неустойчивой, низкой экспрессии гетерологичного гена при его интеграции в геном. Такая репрессия является результатом негативного влияния окружающего хроматина и регуляторных элементов, что можно считать следствием защитной реакции растительного генома на интеграцию чужеродной информации, экспрессию которой надо подавить. Основной причиной нестабильной экспрессии трансгена и ее затухания в течение поколений является репрессия на посттранскрипционном уровне (post-transcriptional gene silencing, PTGS). Механизм этого явления связан с тем, что если существуют две комплементарные РНК, одна из которых может находиться в крайне малой концентрации, то возникающая двуцепочечная РНК запускает как деградацию самой dsPHK, так и индукцию репрессии гена, ответственного за синтез этой РНК. Так же на вариабельность трансгенной экспрессии и на ее снижение влияет множество других факторов, таких как разное число встраиваемых в геном копий, сайты инсерции трансгена. Вариабельность трансгенной экспрессии сильно усложняет как анализ фенотипических признаков, так и создание коммерчески выгодных культур с предсказуемыми характеристиками. Таким образом, крайне актуальной задачей является получение растений с высокой и стабильной экспрессией трансгена. Однако к настоящему времени не известны универсальные сочетания регуляторных элементов и способ трансформации, которые бы эффективно работали в разнообразных растительных культурах, с разными типами промоторов и, кроме этого, не существует четких алгоритмов создания регуляторных элементов, способных сделать экспрессию фланкированного ими гена независимой от места интеграции в растительном геноме.
Широко известен тот факт, что получение трансгенных растений путем искусственного переноса генов (например, биолистическим методом) приводит к появлению в растении большого числа трансгенных копий, тогда как трансформация посредством Agrobacterium tumefaciens приводит к вставке значительно меньшего числа копий и появлению большого числа однокопийных трансгенных клонов. Таким образом, предпочтительно использовать агробактериальную трансформацию, так как мультикопийность часто бывает связана с низким уровнем экспрессии вследствие запуска РНК-сайленсинга. Однако, с другой стороны, низкая копийность трансгена не позволяет получать значительный уровень наработки белка.
Известно, что для снижения влияния эффекта РНК-сайленсинга при создании конструкций часто используются некоторые вирусные гены, кодирующие белки, которые подавляют умолкание РНК. Использование специфического вирусного супрессора сайленсинга, вспомогательного компонента протеиназы НС-Pro вируса гравировки табака, увеличивает уровень экспрессии белков в листьях взрослых растений табака (Mallory et al., 2002), а система транзиентной экспрессии, основанная на коэкспрессии трансгена и супрессора сайленсинга генов p19 вируса кустистой карликовости томатов, значительно увеличивает уровень экспрессии GFP за счет предотвращения умолкания этого гена в протрансформированных тканях (Voinnet et al., 2003). Для увеличения уровня экспрессии используется также 5' проксимальный регион потивируса, содержащий кодирующий регион Р1, вспомогательного компонента протеиназы, и небольшую часть Р3 (US Patent 5939541, 1999). Кроме этого, для стабилизации и увеличения уровня экспрессии проводили трансформацию растений A. thaliana, мутантных по генам sgs2 и sgs3, участвующим в механизме РНК-сайленсинга (Butaye et al. 2004). Однако все эти подходы сложны для практической реализации, так как во многих случаях нужно иметь либо мутантное растение-реципиент, либо специально экспрессировать в растении вирусные белки, что в свою очередь является нежелательным при последующем использовании полученного трансгенного растения в пищевой промышленности или медицине.
Выбор промотора, под контроль которого помещают гетерологичный ген, является еще одним важным фактором, определяющим уровень и стабильность экспрессии его в геноме реципиента. Особенно это важно при использовании тканеспецифичных растительных промоторов, которые намного слабее распространенных в настоящее время вирусных.
Промоторы могут быть различных видов, конститутивные и специфические по ткани или времени экспрессии. Выбранные промоторы могут быть комбинированы дополнительными последовательностями для увеличения экспрессии целевой последовательности в определенной растительной ткани или органе. На сегодняшний день широко используются промоторы вирусный CaMV p35S, октопин- и нопалинсинтазные и растительные актиновые, убиквитиновые и RuBISCo промоторы. Кроме подбора оптимального для поставленной цели промотора, в экспрессирующих векторах, создаваемых для трансформации, необходимо использовать дополнительные регуляторные элементы, способные поддерживать эффективную работу гена, кодирующего целевой белок. Увеличение трансгенной экспрессии наблюдается при использовании экспрессионных кассет, регулируемых оптимизированными 5'- и 3'-регуляторными последовательностями. 5'-UTR последовательность из TMV (штамм U1), сейчас известная как Ω-последовательность, сильно увеличивает уровень экспрессии репортерного гена (Gallie et al., 1987). 5'-UTR последовательности высших растений также обладают энхансерным воздействием. Регуляторные последовательности белков, запасающихся в семенах фасоли обыкновенной (Phaseolus vulgaris), β-фазеолиновый промотор в комбинации с терминатором гена arcelin 5-1, повышают экспрессию гетерологичного белка в семенах двудольных до 36% от общего растворимого белка, вместо 1% при использовании 35S промотора (De Jaeger et al., 2002). При использовании rbcS1-кассеты уровень трансгенной экспрессии достиг в листьях табака 10% от общего растворимого белка (Outchkourov et al. 2003). При использовании репортерного гена GUS было показано, что применение 5'-UTR последовательности гена алкогольдегидрогеназы (NtADH) увеличивает уровень трансгенной экспрессии в 30-100 раз в случае культуры клеток BY-2 и в 30-60 раз для культуры клеток арабидопсиса Т87 (Satoh et al., 2004). 5'-некодирующая последовательность генов, кодирующих белки теплового шока, также увеличивает уровень экспрессии в растениях, при ее присутствии в химерных генах (US Patent 5362865, 1994). Кроме того, высокий уровень трансгенной экспрессии наблюдается при использовании интрон-содержащих генов в сравнении с конструкциями, не содержащими таковых. И положение интрона в гене, и его последовательность оказывают существенное влияние на уровень экспрессии. Использование интрона 5'-UTR последовательности гена rubi3 риса обеспечило 26-кратное увеличение уровня экспрессии репортерного гена GUS в культуре клеток риса Oruza sativa (Samadder et al., 2008). Компания Monsanto для увеличения продукции использовала интрон HSP70, кодирующего белок теплового шока кукурузы, в нетранслируемой лидерной области химерного гена (US Patent 5424412, 1995; US Patent 5593874, 1997; US Patent 5859347, 1999). Однако применение данных подходов ограничивается определенными видами растений и в настоящее время не существует универсальных элементов, которые бы эффективно увеличивали уровень экспрессии любого трансгена в разных растениях. Еще одной проблемой использования трансгенных растений является высокая вероятность последующей репрессии исходно эффективно экспрессирующегося гетерологичного гена в геноме растения-реципиента. С целью предотвращения такой репрессии и повышения уровня стабильности экспрессии трансгена часто используют последовательности ДНК, обычно A/T-богатые, которые, как было показано в экспериментах in vitro, способны взаимодействовать с фракцией ядерного матрикса (MAR, matrix attachment region). Существующая модель предполагает, что за счет взаимодействия с белками ядерного скелета снижается зависимость экспрессии генов, фланкированных MAR, от негативного эффекта окружающего хроматина. Для достижения более высоких уровней экспрессии используют MAR-элементы выделенные из различных источников, таких как человек, курица, дрожжи, соевые бобы, табак, арабидопсис, петуния, кукуруза и синтетические элементы. Была создана система идентификации таких элементов по участку последовательности, состоящей по меньшей мере из двадцати близко расположенных нуклеотидов, 90% которых являются А или Т остатками (US Patent 6245974, 2001). Наибольшее увеличения экспрессии, в 60 раз (Allen et al., 1996) и в 36 раз (Odell and Krebbers, 1998), получили биолистической трансформацией при использовании MAR из табака и курицы соответственно. Однако исследование проводилось на культуре клеток, а не на целом растении, а широко известным является факт недопустимости экстраполяции результатов полученных на культуре клеток на целый организм. Увеличение экспрессии в пять-десять раз в целом растении табака получили при использовании MAR-элементов арабидопсиса (Liu and Tabe, 1998). Для растений риса таких же значений добились комбинацией элементов MAR из табака и применением убиквитинового промотора (Xue et al., 2005). Известны патенты, включающие в себя использование RB7 MAR-элементов выделенных из растений табака (US Patent 5773689, 1998 и US Patent 5773695, 1998) и 5'-региона Bx7 глютенового гена эндосперм-специфичного запасающего белка Triticum aestivum (US Patent 6117612, 2001). Однако с использованием MAR-последовательностей в векторных конструкциях до сих пор не было получено стабильных результатов, которые позволили бы утверждать, что решена проблема константной экспрессии гетерологичного гена в растениях. Так, оказалось, что лишь некоторые MAR оказывают положительный эффект на экспрессию трансгена, который, однако, во многом зависит от конкретной системы выбранных промоторов, места интеграции гетерологичного гена в геном растений, используемого вида растений и способа встраивания конструкции в геном. Более того, способность эффективно связывать компоненты ядерного матрикса in vitro не имеет прямой корреляции с детектируемой активностью MAR в трансгенных линиях. В некоторых случаях было показано, что MAR является прямым стимулятором транскрипции гена в конструкции, а не нейтральной границей, как предполагалось ранее, кроме этого в двух работах (Breyne et al., 1992; Holmes-Davis and Comai, 2002) было показано негативное влияние MAR на экспрессию трансгена. Именно поэтому крайне актуальной является задача идентификации универсальных регуляторных элементов, способных эффективно защищать экспрессию трансгена в геноме растений от последующей репрессии. Таким образом, для создания трансгенного растения с искомым фенотипом и высоким уровнем продукции белка из обширного списка регуляторных последовательностей необходимо тщательно подбирать такой элемент, который в зависимости от рода и вида растения и метода трансформации наиболее полно отвечал поставленным целям, т.к. универсального элемента, дающего 100% гарантию достижения поставленных задач, пока не открыто.
Известен способ получения чужеродного белка в клетках E.coli с использованием экспрессионной кассеты, фланкированной терминаторами (RU 2201455), однако используемый в данном случае вектор, во-первых, предназначен для трансформации прокариот, у которых система транскрипции отличается от эукариотической. Во-вторых, при экспрессии белка в клетках E.coli не происходит линеаризации и интеграции плазмиды в геном клеток, поэтому негативное влияние геномного окружения в данном случае отсутствует. И наконец, терминирующие последовательности в RU 2201455 используют для того, чтобы в плазмиде с промоторных элементов до индукции не запускалась транскрипция через экспрессионную кассету, так как это может ухудшить эффективность индукции, при этом проблемы возможной репрессии транскрипции, запускаемой промоторной или РНК-интерференцией, в данном случае просто не существует.
Известна статья Grummt et al., 1986, в которой описан случай расположения терминирующей последовательности выше промотора рибосомального гена. В данном случае этот терминатор транскрипции усиливает инициацию транскрипции с нижележащего промотора. Однако стоит отметить, что при транскрипции рибосомальной РНК используется ферментная система РНК-полимеразы I, сильно отличающаяся по своему компонентному составу и механизму действия от РНК-полимеразы II, ответственной за синтез матричных РНК. Кроме этого в случае РНК-полимеразы I не наблюдается интерференция транскриптов, а терминатор работает не как защитный регуляторный элемент, обрывающий негативные транскрипты, а как участок, связывающий особый белковый фактор, усиливающий работу промотора гена.
Таким образом, задачей настоящего изобретения является создание векторов для трансформации растительных культур, в которых для поддержания высокого и стабильного уровня наработки целевого белка экспрессионная кассета изолируется эукариотическими терминаторами транскрипции.
Сущность изобретения
Нами предложен способ создания трансгенных линий растений, продуцирующих белок с высоким уровнем экспрессии, включающий трансформацию растений экспрессирующим вектором, включающим плазмиду, содержащую ген, кодирующий β-глюкуронидазу, 35S промотор, nos терминатор транскрипции, отличающийся тем, что выше промотора гена встраивают терминатор транскрипции, обладающий способностью эффективно обрывать геномные транскрипты в растительном геноме и эффективно защищать экспрессию трансгена в геноме растения от последующей репрессии.
Нами предложен способ использования регуляторных элементов, обладающих способностью обрывать геномные транскрипты в растительном геноме и влияющих на уровень экспрессии белка в сторону ее увеличения и стабилизации.
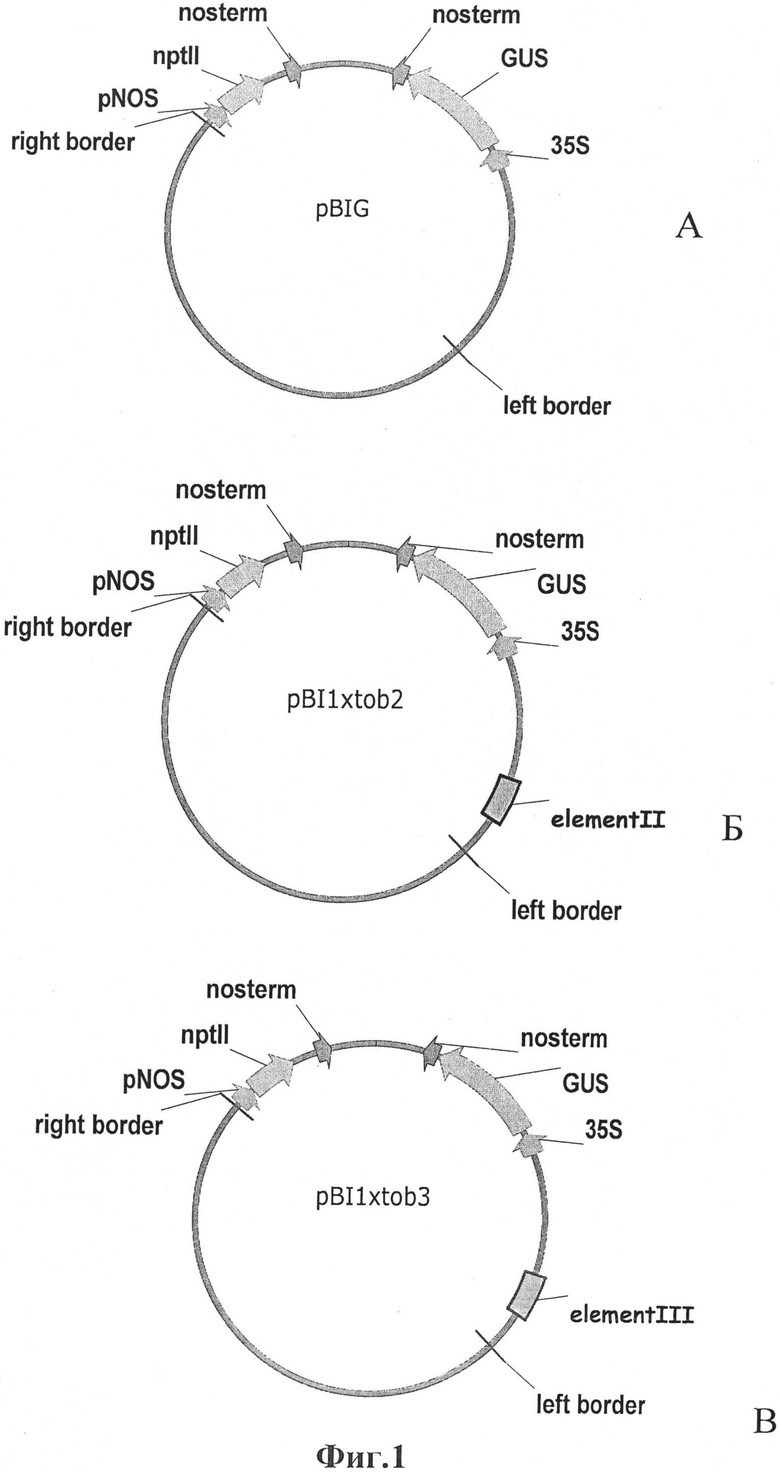
Также нами предложены векторные конструкции, которые содержат ген, кодирующий β-глюкуронидазу или другой целевой белок, 35S промотор, nos терминатор транскрипции и один из синтетических регуляторных элементов, встраиваемый выше последовательности промотора 35S (Фиг.1).
Для достижения результата выбирали оптимальный метод трансформации для создания трансгенных линий растений, продуцирующих белок со стабильным и высоким уровнем экспрессии.
Реализация предложенного способа поясняется чертежом, на котором представлены схемы конструкций для тестирования ДНК-элементов в тестовой культуре растений табака, где
А - контрольная pBI121, в которой тестируемые ДНК-элементы отсутствуют;
Б - конструкция pBI1tod2 с новым синтетическим элементом II, встроенным выше 35S промотора.
В - конструкция pBI1tod3 с новым синтетическим элементом III, встроенным выше 35S промотора.
Предлагаемое изобретение может быть использовано для конструирования трансгенных конструкций, содержащих эффективно работающие регуляторные элементы, в биотехнологии, молекулярной биологии, представляет интерес для крупномасштабного получения рекомбинантных белков, которые могут быть использованы в медицинских и исследовательских целях и для использования растений с данной конструкцией для сельскохозяйственной промышленности.
Таким образом, нами предложен способ создания трансгенных линий растений, продуцирующих белок с высоким уровнем экспрессии, включающий трансформацию растений экспрессирующим вектором, включающим плазмиду, содержащую ген, кодирующий β-глюкуронидазу, 35S промотор, nos терминатор транскрипции, отличающийся тем, что выше промотора гена встраивают терминатор транскрипции, обладающий способностью эффективно обрывать геномные транскрипты в растительном геноме и эффективно защищать экспрессию трансгена в геноме растения от последующей репрессии.
Пример 1
Материалы и способы, используемые при получении трансгенного растения.
Анализ экспрессии репортерного гена проводился на растениях табака Nicotiana tabacum, как наиболее широко применяемой в растительной биотехнологической практике для тестирования подобных регуляторных элементов.
Конструирование векторных плазмид.
В качестве репортерного гена использовался ген β-глюкуронидазы (GUS), помещенный в экспрессионной кассете под 35S промотор. Известно, что данный промотор обеспечивает высокий уровень транскрипции целевого гена в большинстве известных растительных культур. Перед промотором гена помещались синтетические ДНК-элементы, обладающие терминирующей активностью в клетках растений и способствующие получению трансгенной экспрессии высокого уровня.
Данные ДНК-элементы, обладающие способностью обрывать геномные транскрипты, собраны путем комбинирования классического сигнала полиаденилирования (AATAAA), окруженного T-богатой зоной, с последовательностью, обладающей активностью рибозима и способной саморазрезаться. Рибозим - это последовательность РНК с собственной ферментативной активностью. Данный подход позволяет усилить классическую систему терминации/полиаденилирования за счет внесения в молекулу новосинтезиованной РНК дополнительного разрыва.
Для оценки эффективности применения этих элементов при поддержании высокого уровня наработки целевого белка использовались следующие плазмиды:
В качестве отрицательного контроля использовался коммерческий вектор pBI121 (Clontech), содержащий ген ген β-глюкуронидазы (GUS), находящийся под контролем 35S промотора. Данная плазмида содержит ген неомицинфосфотрансферазы II устойчивости к канамицину, который использовался в качестве маркерного для последующего отбора рекомбинантных клонов.
Векторные плазмиды для переноса в растительные клетки репортерного гена с элементами II и III конструировались с использованием хорошо известных методов молекулярного клонирования (Sambrook et al., 1989).
Плазмида pBI1tob2, содержащая синтетическую последовательность II перед геном GUS выше 35S промотора, была получена путем встраивания данного фрагмента ДНК длиной 714 п.н. по сайту рестрикции KpnI.
Плазмида pBI1tod3, содержащая синтетическую последовательность III перед геном GUS выше 35S промотора, была получена путем встраивания данного фрагмента ДНК длиной 585 п.н. по сайту рестрикции KpnI.
Подготовка бактериальной суспензии.
Для генетической трансформации бинарными векторами pBI121, pBI1tob2 и pBI1tob3 использовали супервирулентный штамм Agrobacterium tumefaciens CBE21 (Ревенкова и др. 1994), сконструированный на основе дикого штамма A. tumefaciens с Ti-плазмидой pTiBo542.
Бактериальные суспензии штаммов CBE21/pB1, CBE21/pBI1tob2 и CBE21/pBI1tob3 для инокуляции эксплантов наращивали в течение ночи при 28°C в 50 мл среды LB (см. табл. 1), содержащей 100 мг/л антибиотика канамицин. Перед инокуляцией суспензию клеток центрифугировали при 5 тыс.об/мин в течение пяти минут. Осадок промывали дважды 50 мл жидкой среды MS (Murashige and Skoog, 1962), содержание сахарозы в которой было 30 г/л, для удаления остатков среды LB. Промытый осадок ресуспендировали в 50 мл жидкой среды MS, затем плотность суспензии доводили до ОП600, равной 1.
Подготовка маточных растений
В качестве маточных использовались растения Nicotiana tabacum сорта Petite Havana SRI культуры in vitro, находящиеся на среде 1/2 MS с добавлением витаминов MS, в соответствующем количестве, с содержанием сахарозы 10% и агара 0.8%. Растения культивировались при 23°C и световом режиме дня 16 ч/8 ч (день/ночь) и подвергались пересадке один раз в два месяца. Для эксперимента использовались растения на вторую-третью неделю после очередного пассажа.
Сбор и подготовка листьев
С маточных растений табака двух- трехнедельного возраста, содержащихся в культуре in vitro, непосредственно перед постановкой генетической трансформации срезали молодые, полностью развернувшиеся листья. Чтобы уменьшить усыхание листьев в процессе последующих операций, их содержали в закрытых чашках Петри с небольшим количеством (20 мл) дистиллированной воды. На одну чашку приходится по 20-25 нарезанных фрагментов листа, примерным размером 0.5×2 см. Трансформация
Подготовка, инокуляция и кокультивация эксплантов проводят известным способом, основанным на работах Horsch et al. (1985) и McCormick (1991). Регенерация листовых эксплантов происходила на среде MS, содержащей витамины, сахарозу 30 г/л и гормоны БАП (бензиламинопурин) 1 мг/л, ИУК (индолилуксусную кислоту) 0.1 мг/л.
Селекция трансгенной ткани, отбор трансформантов и элиминация агробактерий.
Экспланты после инокуляции и кокультивации с агробактериями раскладывают в чашки Петри на поверхность среды MS для регенерации. Чашки заматывают парафильмом и инкубируют в термостате в темноте при температуре 23-25°C. Через три дня экспланты для регенерации переносят на среду, дополненную антибиотиками цефотоксимом 500 мг/л и канамицином 100 мг/л. Цефотоксим используют для элиминации остатков агробактерий на эксплантах. Канамицин выступает в роли селективного агента, поскольку в векторных конструкциях в качестве селективного маркера используется ген неомицинфосфотрансферазы nptII.
Мультипликация и укоренение трансформантов.
В процессе прересадок постепенно снижали содержание гормонов, уже через два месяца концентрацию БАП уменьшали до 0.3 мг/л, а ИУК убирали полностью, а потом убирали полностью. Концентрацию цефотоксима изменяют в течение пассажей: с каждым месячным пассажем уровень снижают на 100 мг/л от исходных 500 мг/л до конечных 0 мг/л. Побеги размером один-два сантиметра помещали уже на полностью безгормональную среду. Для укоренения экспланты переносились на среду MS со сниженным содержанием сахарозы, постепенно доводя ее концентрацию до 10 мг/л. При последнем пассаже использовалась среда MS, свободная от антибиотиков. Укорененные растения подвергались предтепличной адаптации и высаживались в тепличный грунт.
Пример 2
Выделение тотальной растительной ДНК из табака
Для экстракции геномной ДНК использовался растительный материал in vivo. С тепличных растений срезался третий лист побега. Выделение проводилось по модифицированному нами протоколу. За основу бралась методика Rogers et al. (1994) с применением 2xCTAB-буфера.
В отличие от протокола Rogers et al. (1994) гомогенезированная ткань ресуспендируется в 1 мл промывочного буфера: 100 мМ ацетата калия, pH 4.5, 20 мМ ЭДТА, 1% PVP, 1% 2-МЭ. Полученная суспензия центрифугируется 5 мин при 4,5 т.о., надосадочная жидкость удаляется. Осадок снова ресуспендируется в 600 мкл экстракционного CTAB-буфера следующего состава: 100 мМ трис-HCl, pH 8, 2.5. М NaCl, 20 мМ ЭДТА, 2% СТАВ, 40 мМ 2-МЭ.
ПЦР-анализ трансгенных линий
Линии, полученные с помощью любой из использованных векторных конструкций, анализируются двумя парами праймеров: на вставку селективного маркера (nptII, Nos/nptII, 742), вставку репортерного гена (uidA, GUSA1/GUSA2,1812) и на отсутствие вирусной контаминации, проверкой наличия генов вирулентности ряда vir (virA/virB, 670). Линии, содержащие новые синтетические элементы, проверялись на наличие элемента II или элемента III (2-1/2-2, 715 и 3-1/3-2, 597, соответственно).
ПЦР-анализ интеграции различных гетерологичных элементов в геном табака проводят в реакционной смеси, которая содержит 67 мМ Tris-HCl, pH 9,0, 16 мМ (NH4)2SO4, 2 мМ MgCl2, 0,01% БСА, по 200 µМ каждого dNTP. Концентрации праймеров и полимеразы, а также температурный режим подбирают для каждого конкретного случая. Для амплификации фрагмента nptII в реакционную смесь вносят праймеры до 0,6 µМ конечной концентрации и 0.05 U/µl Taq полимеразы. Режим амплификации: 5 мин денатурация при 94°C (горячий старт), 30 с денатурация при 93°C, отжиг - 45 с 62°C, элонгация 45 с 72°C, циклов амплификации 30. Ожидаемый размер амплифицируемого фрагмента 742 п.н.
Вставка репортерного гена uidA определялась при условиях, по большей части схожих с таковыми для nptII, только концентрация праймеров составила 0,15 µМ каждого, 0.1 U/µl Taq, а температурный режим был изменен следующим образом: горячий старт 2 мин при 94°C, денатурация 30 с 93°C, отжиг - 40 с 60°C, элонгация 55 с 72°C, 30 циклов амплификации. Ожидаемый размер амплифицируемого фрагмента 1812 п.н. Вставка элемента II определялась при условиях, по большей части схожих с таковыми для uidA, только концентрация праймеров составила 0,25 µМ каждого, а температурный режим был изменен следующим образом: горячий старт 2 мин при 94°C, денатурация 30 с 93°C, отжиг - 40 с 60°C, элонгация 35 с 72°C, циклов амплификации 30. Ожидаемый размер амплифицируемого фрагмента 715 п.н.
Вставка элемента III определялась при условиях, по большей части схожих с таковыми для uidA и элемента II, только концентрация праймеров составила 0,25 µМ каждого, а температурный режим был изменен следующим образом: горячий старт 2 мин при 94°C, денатурация 30 с 93°C, отжиг - 40 с 60°C, элонгация 35 с 72°C, циклов амплификации 30. Ожидаемый размер амплифицируемого фрагмента 597 п.н.
Гистохимический анализ GUS активности.
Гистохимическое определение активности GUS проводили по способу Jefferson (1987). Гистохимическое определение GUS активности проводили с использованием 5-бром-4-хлор-3-индолилглюкоронида (X-GLUC, Duchefa). Для этого растительную ткань помещали в буфер 50 мМ NaPO4, рН 7.0, 10 мМ Na2EDTA, 0.1% тритон Х-100, содержащий 1 мг/л X-GLUC, инкубировали в течение шести часов при 37°C. После чего несколько раз промывали 50% этанолом и хранили окрашенные ткани при 4°C в 70% этаноле.
Флюориметрический анализ GUS активности.
Измерение проводилось при использовании реактива 4-MUG (4-метилумбеллиферилглюкоронида), который при расщеплении β-глюкуронидазой до 4-MU имеет оптимумы возбуждения при 365 нм и излучения при 455 нм. Для этого белки экстрагировались из растительной ткани в растворе для экстракции состава: 50 мМ NaPO4, pH 7.0, 10 мМ Na2EDTA, 0.1% тритон Х-100 0.1% саркозил, 10 мМ β-меркаптоэтанол, следующим образом: 200 мг ткани растиралось в ступке с жидким азотом и порошок опускался в буфер для экстракции. В дальнейшем проводилось центрифугирование при 10.000 g, и надосадок, содержащий белки, пререносился в отдельный эппендорф. Для каждого образца предварительно инкубировалось по 100 µл буфера для экстракции GUS с добавлением 1 мМ реактива 4-MUG. Добавляли в реакционную смесь 10 µл белкового экстракта и через 10 с 900 µл 0.2 М Na2CO3, для остановки реакции. В дальнейшем измерение проводилось на флюориметре, откалиброванном по свежеприготовленным стандартным растворам 4-MU в буфере для экстракции с добавлением 0.2 М Na2CO3 в соотношении 1:9. Количество репортерного белка оценивалось относительно общего белка экстракта, определение концентрации которого проводилось по реакции Бредфорда. Окончательные концентрации были получены в размерности нмоль 4-MU/ч на 1 µг общего белка.
Пример 3
Данные ПЦР анализа трансгенных растений табака, полученных с помощью трансформации экспрессионными векторами.
Основываясь на результатах ПЦР анализа, из полученних трансгенных линий табака для дальнейшего анализа было выбрано четыре клона, содержащих вставку элемента II и восемь клонов со вставкой элемента III (см. табл. 2). Также было выбрано 14 контрольных линий.
Праймеры
VirB1 GGCTACATCGAAGATCGTATGAATG
VirB2 GACTATAGCGATGGTTACGATGTTGAC
NOS CGCGGGTTTCTGGAGTTTAATGAGCTAAG
nptII-2 GCATGCGCGCCTTGAGCCTGG
CaMV35S-R TGAATCTTTTGACTGCATCTTTAACCTTCTT
nptII-F ACGACGGGCGTTCCTTGCG
3-1 TCATAGTGTTACCATCAACCACCTTAACTtc
3-2 TTCCGCAAAAATCACCAGTCTCTCTCTAC
2-1 СATAGTGTTACCATCAACCACCTTAACTTC
2-2 TCCTGTCACCGGATGTGTTTTCC
Пример 4
Данные гистохимического анализа GUS активности
Для оценки влияния новых регуляторных элементов на уровень экспрессии репотерного белка еще на стадии размножения трансгенных линий in vitro проводился гистохимический анализ GUS-активности тканей табака. Такой анализ позволил быстро оценить наличие экспрессии белка в тканях растения (см. табл 3).
Пример 5
Влияние регуляторных элементов на стабильную GUS экспрессию в тканях растений табака.
Для полноценного анализа влияния синтетических элементов II и III был проведен флюориметрический анализ активности GUS в растениях, протрансформированных экспрессионными векторами, содержащими, в том числе, и эти регуляторные элементы. Для анализа было взято по 200 мг молодой ткани растений, находящихся в теплице (как правило это был 3-4 лист). Пробы были взяты с пяти индивидуальных растений каждой линии. Измерение проводилось на флюориметре, откалиброванном по свежеприготовленным стандартным ратсворам 4-MU. Количество репортерного белка оценивалось относительно общего белка экстракта, определение концентрации которого проводилось по реакции Бредфорда. Окончательные концентрации были получены в размерности нмоль 4-MU/ч на µг общего белка. Была проведена статистическая обработка полученных данных в программе Microsoft Excel и результаты сведены к следующей таблице (см. табл 4). Так как полученные нами значения уровня экспрессии отклоняются от нормального распределения (согласно тестам Лилифора и Шапиро-Уилка), к ним невозможно применить дальнейший статистический анализ, рассчитанный на нормальное распределение.
bнмоль 4-MU/ч на µг общего белка;
cСтандартное отклонение;
dкоэффициент вариации;
Исходя из этих данных видно, что при расположении терминирующих элементов выше промотора репортерного гена общий уровень экспрессии трансгена увеличивается в три-четыре раза, что позволяет отметить данные элементы как возможные компоненты для создания экспрессионных кассет, направленных на получение высоких и стабильных уровней экспрессии целевого гена в различных растительных объектах. Данные элементы могут быть использованы в создании трансгенных растений практически для всех отраслей их применения, таких как биотехнология и сельскохозяйственная промышленность.
Литература
1. Kwaks Т.H.J., Barnett P., Hemrika W. et al. Identification of anti-repressor elements that confer high and stable protein production in mammalian cells Nature Biotechnology, 2003, 21: 553-558.
2. Allen G.C., Hall G., Michalowski S., Newmann W., Spiker S., Weissinger A.K., Thompson W.F. (1996). High-Level Transgene Expression in Plant Cells: Effects of a Strong Scaffold Attachment Region from Tobacco. Plant Cell. 8: 899-913.
3. Breyne P, van Montagu M, Depicker N, Gheysen G. (1992). Characterization of a plant scaffold attachment region in a DNA fragment that normalizes transgene expression in tobacco. Plant Cell. 4: 463-471.
4. Butaye K.M., Goderis I.J., Wouters P.F., Pues J.M., Delaure S.L., Broekaert W.F., Depicker A., Cammue B.P., De Bolle M.F. (2004). Stable high-level transgene expression in Arabidopsis thaliana using gene silencing mutants and matrix attachment regions. Plant J. 39: 440-449.
5. Gailie D.R., David E. Sleat, John W. Watts, Philip C. Turner, T. Michael A. Wilson (1987). The 5'-Leader Sequence of Tobacco Mosaic Virus RNA Enhances the Expression of Foreign Ene Transcripts in Vitro and in Vivo. Nucleic Acids Res. 15: 3257-3273.
6. Geert De Jaeger, Stanley Scheffer, Anni Jacobs, Mukund Zambre, Oliver Zobell, Alain Goossens, Ann Depickerl & Geert Angenon. (2002). Boosting heterologous protein production in transgenic dicotyledonous seeds using Phaseolus vulgaris regulatory sequences. Nature Biotechnol. 20: 1265-1268.
7. Grummt Ingrid, Kuhn Anne, Bartsch Ingrid, Rosenbauer Horst. (1986). A transcription terminator located upstream of the mouse rDNA initiation site affects rRNA synthesis. Cell. 47: 901-911.
8. Holmes-Davis R, Comai L. (2002). The matrix attachment regions (MARs) associated with the Heat Shock Cognate 80 gene (HSC80) of tomato represent specific regulatory elements. Mol Genet Genomics. 266: 891-898.
9. Horsch, R.В., Fry, J.E., Hoffmann, N.L., Eichholtz, D., Rogers, S.G., Fraley, R.T. (1985). A simple and general method for transferring genes into plants. Science. 227: 1229-1231.
10. Jefferson R.A., Kavanagh T.A., Bevan M.W.. (1987). GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 6: 3901-3907.
11. Liu, J.W. and L.M. Tabe (1998). The influences of two plant nuclear Matrix Attachment Regions (MARs) on gene expression in transgenic plants. Plant Cell Physiol. 39: 115-123.
12. Mallory, A.C, Reinhart, B.J., Battel, D., Vance, V.B. & Bowman, L.H. (2002). A viral suppressor of RNA silencing differentially regulates the accumulation of short interfering RNAs and micro-RNAs in tobacco. Proc Natl Acad Sci USA. 99:15228-15233.
13. McCormick, S. (1991) Transformation of tomato with Agrobacterium tumefaciens. Plant Tissue Culture Manual B6: 1-9. Kluwer Academic Publishers.
14. Murashige T. and Skoog F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant. 15: 473-497.
15. Odell, J.T. and Krebbers, E. (1998). Enhanced transgene expression in a population of monocot cells employing scaffold attachment regions. World Patent Office.
16. Outchkourov, N.S., Peters, J., de Jong, J., Rademakers, W. and Jongsma, M.A. (2003). The promoter-terminator of chrysanthemum rbcSl directs very high foreign gene expression levels in plants. Planta. 216: 1003-1112.
17. Rogers S., Bendich A. (1994). Extraction of DNA from plant, fungal and algal tissues. In: Gelvin SB, Schilperoort RA (eds) Plant Molecular Biology Manual. Boston, MA: Kluwer Academic Publishers, D 1: 1-8.
18. Samadder P, Sivamani E, Lu J, Li X, Qu R. (2008). Transcriptional and post-transcriptional enhancement of gene expression by the 5' UTR intron of rice rubi3 gene in transgenic rice cells. Mol Genet Genomics. 279: 429-39.
19. Sambrook, J., E.F.Fritsch, and T.Maniatis. (1989). Molecular cloning: a laboratory manual. Second edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA.
20. Satoh J., Kato K., Shinmyo A.J. (2004). The 5'-untranslated region of the tobacco alcohol dehydrogenase gene functions as an effective translational enhancer in plant. Biosci Bioeng. 98: 1-8.
21. Voinnet O., Rivas S., Mestre P., Baulcombe D.C. (2003). An enhanced transient expression system in plants based on suppression of gene silencing by the pl9 protein of tomato bushy stunt virus. Plant J. 33: 949-956
22. Xue, H., Yang, Y.T., Wu, C.A., Yang, G.D., Zhang, M.M., Zheng, C.C. (2005). TM2, a novel strong matrix attachment region, isolated from tobacco, increases transgene expression in transgenic rice calli and plants. Theor. Appl. Genet. 110: 620-627
23. Ревенкова E.B., Багян И.Л., Позмогова Г.Е., Краев А.С. (1994) Разработка новой векторной системы на базе штамма Agrobacterium tumefaciens А281. Молекул, генет., микробиол. и вирусол. - Т 5. - С.36.
Изобретение относится к области биохимии, в частности к способу создания трансгенных линий растений, продуцирующих белок с высоким уровнем экспрессии, включающему трансформацию растений экспрессирующим вектором, включающим плазмиду, содержащую ген, кодирующий β-глюкуронидазу, 35S промотор, nos терминатор транскрипции. При этом выше промотора гена встраивают терминатор транскрипции, обладающий способностью эффективно обрывать геномные транскрипты в растительном геноме и эффективно защищать экспрессию трансгена в геноме растения от последующих репрессий. Изобретение позволяет создавать трансгенные линии растений, продуцирующие белок с высоким уровнем экспрессии. 1 ил., 3 табл., 5 пр.
Способ создания трансгенных линий растений, продуцирующих белок с высоким уровнем экспрессии, включающий трансформацию растений экспрессирующим вектором, включающим плазмиду, содержащую ген, кодирующий β-глюкуронидазу, 35S промотор, nos терминатор транскрипции, отличающийся тем, что выше промотора гена встраивают терминатор транскрипции, обладающий способностью эффективно обрывать геномные транcкрипты в растительном геноме и эффективно защищать экспрессию трансгена в геноме растения от последующей репрессии.
| СПОСОБ ПОЛУЧЕНИЯ ЧУЖЕРОДНОГО ПРОТЕИНА В КЛЕТКАХ E.COLI, ПЛАЗМИДНЫЙ ВЕКТОР И ТРАНСФОРМИРОВАННЫЙ ШТАММ Е.COLI ДЛЯ ЭКСПРЕССИИ ЧУЖЕРОДНОГО ПРОТЕИНА | 1996 |
|
RU2201455C2 |
| СПОСОБ МОДИФИКАЦИИ ФРАГМЕНТА ДНК, КОДИРУЮЩЕГО ИНСЕКТИЦИДНЫЙ БЕЛОК BACILLUS THURINGIENSIS, ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ИНСЕКТИЦИДНЫЙ БЕЛОК BACILLUS THURINGIENSIS (ВАРИАНТЫ), ФРАГМЕНТ ДНК, СОДЕРЖАЩИЙ СТРУКТУРНЫЙ ГЕН, КОДИРУЮЩИЙ ИНСЕКТИЦИДНЫЙ БЕЛОК (ВАРИАНТЫ), И ФРАГМЕНТ ДНК, СОДЕРЖАЩИЙ СТРУКТУРНЫЙ ГЕН, КОДИРУЮЩИЙ СЛИТЫЙ БЕЛОК | 1990 |
|
RU2107725C1 |
| СКРИПКИН Е.А | |||
| и др | |||
| Векторы для клонирования и определения активности элементов транскрипционной системы бактерий | |||
| Клонирование терминатора транскрипции оперона рРНК из архебактерий Halobacterium halobium | |||
| Биополимеры и клетка, 1987, т.3, №5, с.263-268 | |||
| GRUMMT I | |||
| Et al., A transcription | |||

