Настоящее изобретение относится к синтетическому промотору, индуцируемому патогенами, который пригоден для регуляции транскрипции нуклеиновых кислот и включает минимальный промотор. Помимо этого настоящее изобретение относится к трансгенной клетке растения, а также к трансгенному растению. Также настоящее изобретение относится к способу получения растения, устойчивого к патогенам.
Известны различные способы получения растений, устойчивых к таким патогенам, как грибы, вирусы, бактерии и нематоды. В некоторых таких способах используют реакцию гиперчувствительности (РГ) растения, при которой в месте прямого контакта патогена и растения происходит развитие некроза. В процессе РГ в смежных клетках запускается широкий спектр действий по борьбе с патогенами, которые предотвращают дальнейшее проникновение патогенов в растительную ткань.
РГ может последовать за экспрессией эффекторных генов, таких как, например, гены авирулентности патогенов, и взаимодействием с продуктом соответствующего гена устойчивости (R-ген). При этом R-ген может присутствовать у растения или может быть внедрен в геном растения с помощью способов генной инженерии (Stuiver et al. 1998, Kelleretal., 1999, Belbahri et al., 2001). Кроме того, к запуску РГ может привести сверхэкспрессия или аутоактивирование R-генов (Тао et al., 2000, Tang et al., 1999, Bendahmane et al., 2002, Howles et al., 2005). Вследствие сверхэкспрессии R-генов происходит превышение пограничного значения, что приводит к запуску сигнального каскада, который запускается в норме только при проникновении патогенов или продуктов их генов авирулентности. Вследствие запуска этого каскада может быть достигнута устойчивость к патогенам широкого спектра (Oldroyd and Staskawicz, 1998, Tang et al., 1999, Тао et al., 2000, Howles et al., 2005). Под аутоактивными R-генами будут понимать R-гены, которые модифицированы таким образом, что для запуска сигнального каскада присутствие патогенов/авирулентных продуктов не является необходимым, и одновременно с этим для запуска сигнального каскада достаточна пониженная по сравнению с немодифицированными формами экспрессия.
Стюивер и коллеги (Stuiver et al., 1998) смогли показать, что трансформация растений томата, обладающих соответствующим Cf9-геном, avr9-геном из фитопатогенного гриба Cladosporium fulvum под контролем патоген-индуцируемого промотора Gst1 картофеля приводит к появлению устойчивости широкого спектра к грибам. Устойчивость к оомицетам Phytophthora parasitica var nicotianae может быть достигнута у Nicotiana tabacum после трансформации N. tabacum или элиситором Cryptogein из Р. cryptogea или бактериальным элиситором рорА из фитопатогенной бактерии Ralstonia solanacearum. Оба гена находятся под контролем патоген-индуцируемого промотора hsr203J из N. tabacum (Kelleret al., 1999, Belbahri et al., 2001).
Система запуска РГ требует обязательного контроля экспрессии эффекторных генов в месте инфекции. При неконтролируемой экспрессии экспрессия эффекторных генов оказывает негативный эффект на рост растения и, следовательно, на урожайность культурных растений (Stuiver and Güsters, 2001). Контролируемая экспрессия может быть, разумеется, достигнута с помощью выбора подходящих патоген-индуцируемых промоторов. В благоприятных условиях они не должны позволять совсем или должны позволять только слабую экспрессию, а при поражении в месте инфекции наоборот вызывать отчетливую экспрессию. После трансформации двух различных аутоактивных форм L6 генов устойчивости к ржавчине из льна (Linum usitatissimum) под контролем природного индуцируемого ржавчиной промотора Fis1 из льна можно получить два фенотипа: во первых, нормально выросшие растения, которые не демонстрируют улучшенной устойчивости к патогенам, и во вторых низкорослые растения с устойчивостью к патогенам широкого спектра (Howles et al., 2005). Эти результаты показывают, что в зависимости от применяемой формы аутоактивного R-гена имеет место активность промотора, которая может находиться выше пограничного значения для индукции сигнального каскада, в то время как у растений с непроявляющимся фенотипом индукция промоторов F/sf не достаточна, чтобы достичь пограничного значения. Специфичности природного промотора Fis1 не хватает для того, чтобы достичь высоко эффективной устойчивости к патогенам без негативных последствий для роста растения.
Природные патоген-индуцируемые промоторы демонстрируют зачастую неспецифическую активность и активируются с помощью многочисленных стимулов, таким образом, что их применение для экспрессии вышеуказанных эффекторных генов является бессмысленным, поскольку запуск РГ может происходить даже в отсутствие инфекции. Этот недостаток промоторов причиняет вред росту растений и, поэтому, приводит к снижению урожайности культурных растений. Поэтому были разработаны синтетические промоторы, содержащие мотив последовательности (цис-регуляторный элемент) из природных, патоген-индуцируемых промоторов, которые подходят для индукции патогеном. Мотивы последовательности для других стимулов были оттуда удалены. Цис-регуляторные элементы были клонированы в позицию upstream относительно минимального промотора (то есть предшествуют ему), в результате чего был создан функциональный промотор, проявляющий повышенную специфичность по сравнению с природными промоторами, из которых были выделены соответствующие цис-регуляторные элементы (Rushton et al., 2002). В качестве минимального промотора для двудольных растений применили область -46 до+8 гена 35S вируса мозаики цветной капусты. Кроме того, известно применение минимального промотора из природного промотора, из которого были клонированы соответствующие цис-регуляторные элементы (Perl-Treves et al., 2004). Для однодольных растений описано применение минимального промотора из гена Act1 риса (Lü et al., 2000).
Хотя описанные синтетические промоторы превосходят природные промоторы, они все же демонстрируют фоновую активность даже при неподходящих условиях. Эта фоновая активность колеблется у отдельных видов растений. Хотя у всех ранее изученных видов растений могла быть установлена способность к индукции патогенами, тем не менее, сила индукции и абсолютная активность промоторов ослабевают. При слишком сильной фоновой активности в неинфицированной ткани может быть установлена только слишком слабая способность к индукции патогенами как частное активности промотора в инфицированной ткани и активности промотора в неинфицированной ткани.
До настоящего времени отвечающими за величину базовой активности синтетического промотора считали только применяющиеся цис-регуляторные элементы. Они оказывают очень значительное влияние на силу промотора (Rushton et al., 2002). На настоящий момент влияние минимального промотора изучено мало. Из литературных источников известно, что минимальный промотор обладает очень слабым влиянием на регуляцию активности промотора (Singh, 1998). Бхуллар и коллеги (Bhullar et al., 2003) смогли установить только отчетливое снижение активности промотора 35S, когда минимальный промотор (от -46 до +1) заменяли на гетерологичные растительные минимальные промоторы. Эти различия объяснили различиями в последовательности ТАТА-боксов, в то время как, по их мнению, фланкирующие области ТАТА-бокса минимального промотора не были существенными для активности промотора.
Задачей настоящего изобретения является создать патоген-индуцируемый синтетический промотор со слабой базовой активностью.
Согласно настоящему изобретению, поставленная задача решается с помощью патоген-индуцируемого синтетического промотора с минимальным промотором, где минимальный промотор содержит мотив последовательности
a) dbrmwa или
b) twcccmt,
который находится downstream от (то есть после) ТАТА-области и перед сайтом начала транскрипции, расположенным в минимальном промоторе, в котором начинается транскрипция регулируемых нуклеиновых кислот. При этом мотив последовательности dbrmwa подходит для всех двудольных, а мотив последовательности twcccmt для всех однодольных растений.
Символы мотива последовательности имеют здесь следующее значение:
d = нуклеотид а или g или t/u
b = нуклеотид с или g или t/u
r = нуклеотид g или а
m = нуклеотид а или с
w = нуклеотид а или t/u
а = нуклеотид а
t = нуклеотид t
с = нуклеотид с
Согласно настоящему изобретению "минимальный промотор" является последовательностью ДНК промотора, которая необходима для функционирования промотора. С этой последовательностью ДНК могут связываться универсальные транскрипционные факторы, такие как, например, TFII-D, TFII-A, TFII-B, TFII-E и TFII-F, и образуется платформа для связывания комплексов РНК-полимеразы II/TFII-F. Поскольку в этой области начинается транскрипция мРНК на ДНК, сайт начала транскрипции (НТ) расположен внутри минимального промотора и он определяется как позиция +1. Минимальный промотор включает НТ и может, например, простираться от позиции -50 до позиции +15. В позиции -30 часто обнаруживают так называемый ТАТА-бокс, который, однако, встречается не во всех промоторах. ТАТА-бокс является участком с чередованием оснований тимина и аденина. ТАТА-бокс является сайтом связывания для ТАТА-бокс связывающих белков (ТББ).
Как "синтетические промоторы" будут обозначаться такие промоторы, которые не встречаются в природе, составленные из множества элементов, и содержащие минимальный промотор, а также по меньшей мере один цис-регуляторный элемент в направлении upstream от минимального промотора (то есть предшествуют ему), который служит в качестве сайта связывания для специальных транскрипционных факторов. Синтетические промоторы должны отвечать желательным требованиям, и быть индуцируемыми или блокироваться с помощью различных факторов.
"Производным" промотора является укороченные, или удлиненные, или идентичные на отдельных отрезках варианты этого промотора или гомологи с такими же модифицированными или особыми свойствами.
Термин "гомология" обозначает здесь гомологию матрицы ДНК по меньшей мере в 70%, которая может быть установлена широко известными способами, например, путем компьютерного сравнения последовательностей (Altschul, S.F. et al., 1990).
Патоген-индуцируемый синтетический промотор по настоящему изобретению дает в результате транзитной биолистической трансформации ткани листа данного растения более низкую базовую активность по сравнению с обычно применяемыми промоторами с минимальным промотором, таким как минимальный промотор 35S двудольных растений, и минимальным промотором Mais-ubi1 однодольных растений. Более того, было обнаружено, что с помощью патоген-индуцируемых синтетических промоторов по настоящему изобретению также повышается уровень индукции.
Также патоген-индуцируемые синтетические промоторы по настоящему изобретению можно применять для изготовления трансгенных растений, которые обладают широкой устойчивостью к многочисленным патогенам, таким как грибы, оомицеты, бактерии, вирусы, насекомые и нематоды.
Мотивы последовательности dbrmwa и twcccmt располагаются на кодирующей цепочке между ТАТА-боксом и сайтом начала транскрипции, и могут повторяться два или более раза. Предпочтительными последовательностями минимального промотора являются последовательности, описанные в SEQ ID NOS: 1-9.
Цис-регуляторными элементами для изготовления патоген-индуцируемых синтетических промоторов являются такие элементы, которые встречаются в природных патоген-индуцируемых промоторах и отвечают в них за индукцию патогеном. Их идентификация описана Раштоном и коллегами (Rushton et al., 2002).
Предпочтительные цис-регуляторные элементы для изготовления синтетических промоторов с применением минимальных промоторов по настоящему изобретению также описаны в WO 00/29592. Из описанных там цис-регуляторных элементов прежде всего подходит D-бокс (SEQ ID NO: 10), в особенности в комбинации 2×S/2×D (SEQ ID NO: 11), а также Gst1-элемент, предпочтительно в комбинации 4×Gst1 (SEQ ID NO: 12).
К предпочтительным комбинациям цис-элементов причисляют обычно комбинации D-бокса (SEQ ID NO:10) с S-боксом, например, особенно предпочтительными являются Gst1-элемент рядом с вышеобозначенной комбинацией 2×S/2×D (SEQ ID NO: 11), комбинации 2×S/4×D (SEQ ID NO: 13); 4×S/2×D (SEQ ID NO: 14) и 2×Gst1/2×D (SEQ ID NO: 15). Комбинация 2×S/4×D элементов (SEQ ID NO: 13) с минимальным промотором согласно SEQ ID NO: 2 в трансгенном картофеле после инфицирования Phytophthora infestans демонстрирует среднее возрастание активности репортерного гена до величины 253.000 по сравнению с неинфицированным контролем. Если клонировать элемент 4×S/2×D (SEQ ID NO: 14) перед минимальным промотором (SEQ ID NO: 2), может быть установлено среднее повышение активности гена-репортера до величины 2.892. С помощью элемента 2×Gst1/2×D (SEQ ID NO: 15) было достигнуто среднее повышение до величины 2.967 по сравнению с контролем.
С помощью промоторов по настоящему изобретению можно получать, таким образом, трансгенные клетки растений, которые могут регенерировать в полные растения с улучшенными свойствами защиты от патогенов. В семенах таких трансгенных растений содержатся промоторы по настоящему изобретению. Настоящее изобретение не ограничено определенными сортами растений.
Настоящее изобретение относится далее также к способу получения растений, устойчивых к патогенам, с при котором в клетку растения вводят подходящий для возникновения защиты от патогенов ген, который находится под контролем патоген-индуцируемого синтетического промотора, после чего из этой клетки растения регенерируют растение, отличающийся тем, что патоген-индуцируемый синтетический промотор представляет собой патоген-индуцируемый синтетический промотор, который был описан выше.
Примеры
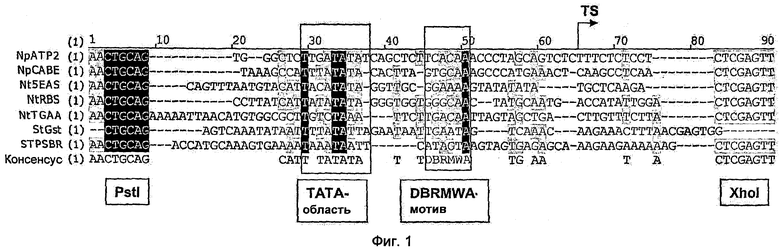
На Фиг.1 показано сравнение последовательностей предпочтительных минимальных промоторов двудольных растений (SEQ ID NOS: 1-7) с консервативными ТАТА-областями и мотивами dbrmwa, а также применяемыми для клонирования в плазмиде pMS23luc+сайтами рестрикции Pstl и Xhol.
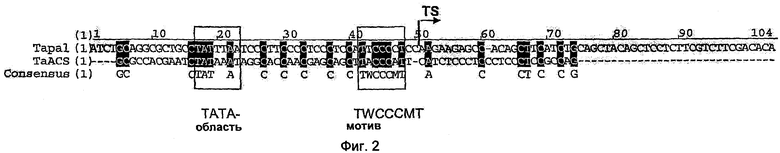
На Фиг.2 показано сравнение последовательностей предпочтительных минимальных промоторов однодольных растений (SEQ ID NOS: 8 и 9), которые применяли для транзитной трансформации листьев. Рядом с ТАТА-областью представлен мотив последовательности twcccmt как консервативная область.
Можно видеть, что минимальные промоторы StGst (SEQ ID NO: 6), NtTGAA (SEQ ID NO: 5), StPSBR (SEQ ID NO: 7), NpCABE (SEQ ID NO: 2), NtRBS (SEQ ID NO: 3), NpATP2 (SEQ ID NO: 1) и Nt5EAS (SEQ ID NO: 4) проявляют отчетливо сниженную по сравнению с минимальным промотором 35S активность (<70%).
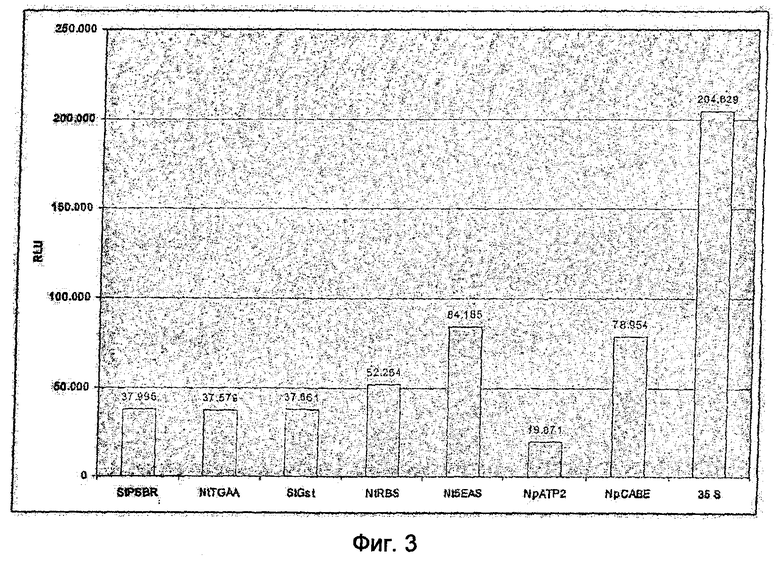
На Фиг.3 показан обзор средней активности репортерных генов для неинфицированных трансгенных растений картофеля с синтетическим промотором, состоящим из 4xGst1 элемента (SEQ ID NO: 12) и представленных минимальных промоторов, клонированных с геном люциферазы из Photinus pyralis в качестве репортерного гена (RLU (Relative Light Unit) = сравнительные световые единицы). Стабильные трансгенные линии с минимальными промоторами, несущими мотив последовательности dbrmwa, в контрольных условиях демонстрируют экспрессию гена-репортера, отчетливо сниженную по сравнению с минимальным промотором 35S. Наименьшая средняя активность была достигнута при применении минимального промотора гена NpATP2 (SEQ ID NO: 1). У этих растений измерения показали только 9,7% средней активности минимального промотора 35S. При применении минимальных промоторов StPSBR (SEQ ID NO: 7), NtTGAA (SEQ ID NO: 5) или StGst (SEQ ID NO: 6) измерения показали 18% от активности минимального промотора 35S, при применении минимального промотора NtRBS (SEQ ID NO: 3) - 26%, при применении минимального промотора NpCABE (SEQ ID NO: 2) - 39% и при применении минимального промотора Nt5EAS (SEQ ID NO: 4) - 41%.
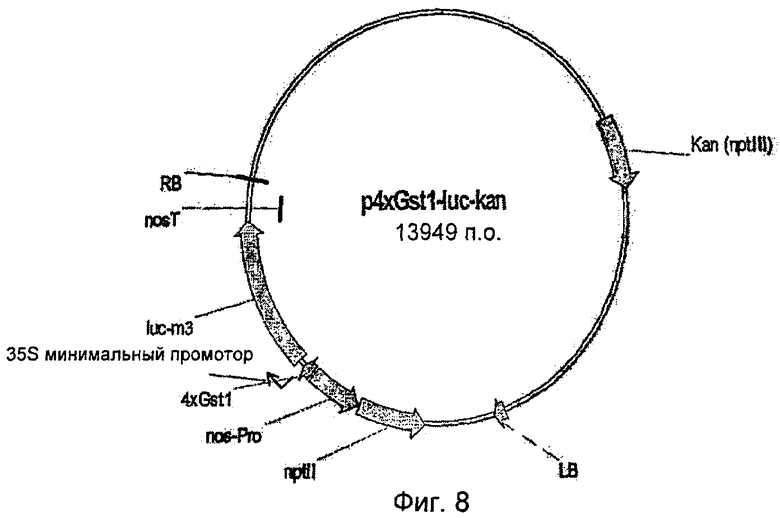
Для специалиста изготовление походящей конструкции для трансформации растений промоторами по настоящему изобретению не является проблемой. Например, можно изготовить бинарный вектор p4xGst1-luc-kan (Фиг.8), который будет применяться для стабильной трансформации растений картофеля сорта "Baltica". Этот вектор является производным бинарного вектора pGPTV (Becker et al., 1992). Бинарный вектор p4xGst1 luc-kan несет ген люциферазы из Photinus pyralis под контролем минимального промотора синтетического промотора 4xGst1:35S (Rushton et al., 2002). В качестве концевой последовательности плазмида содержит терминатор гена нопалинсинтазы из Agrobacterium tumefaciens. Описанный модуль экспрессии локализован на Т-ДНК вместе с функциональным модулем экспрессии гена неомицинфосфотрасферазы (nptll) в качестве селективного маркера.
Неомицинфосфотрансфераза придает трансгенным растениям устойчивость против канамицина или паромицина. Чтобы поменять минимальный промотор 35S на описанные выше минимальные промоторы, бинарный вектор p4xGst1 luc-kan лизировали с помощью Xhol/Sall, в результате чего минимальный промотор 35S удаляли, а тетрамер элемента Gst1 оставался на месте. Сайт рестрикции для Sail достраивали с помощью ферментов полимеразы Кленова и dNTP's для получения «тупого конца». Минимальный промотор, представленный в плазмиде pMS23luc+, вырезали лизисом Pdil/Xhol, вставляли в бинарный вектор, и, вслед за тем, трансформировали им Е. coll. Штамм агробактерии GV3101:pMP90 (Koncz and Schell, 1986) трансформировали бинарными векторами с новой последовательностью (An, 1987), и проводили селекцию с помощью антибиотика канамицина (50 мг/л). Трансгенные агробактерии использовали для трансформации картофеля сорта "Baltica" (Dietze et al., 1995).
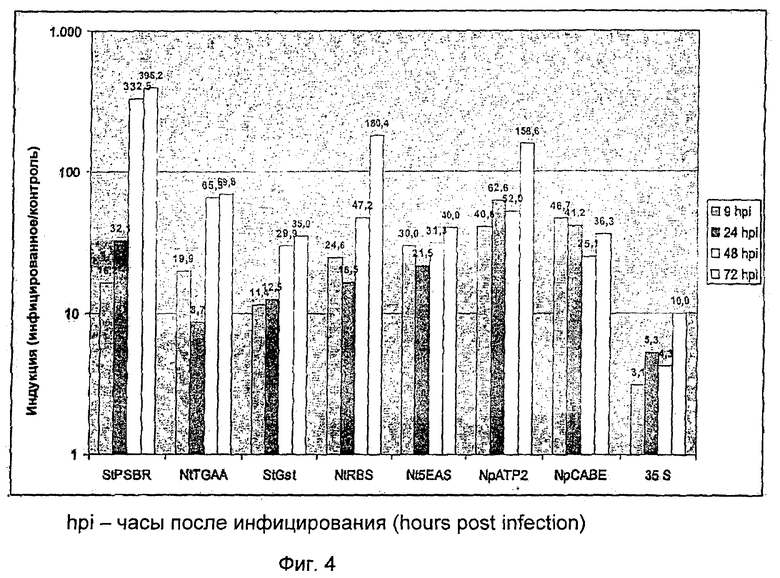
На Фиг.4 показан обзор достигнутых индукций после инфекции in vitro для стабильных, трансгенных растений картофеля с синтетическим промотором, состоящим из элемента 4xGst1 (SEQ ID NO: 12) и описанного минимального промотора. Инфицирование растений осуществляли in vitro суспензией зооспор Phytophthora infestans.
Спустя различные временные промежутки после прививки отбирали in vitro пробы листьев растений, определяли вес проб, и добавляли к ним 10 объемов 1xCCLR буфера (Promega, Mannheim). Материал гомогенизировали с помощью гомогенизатора RW20 Homogenisators (IKA Labortechnik, Staufen) в буфере на льду. В ходе центрифугирования при >10.000×g в течение 10 минут гомогенат становился прозрачным, и 10 мл супернатанта смешивали с 50 мкл субстрата LAR (Promega, Mannheim) в пробирках для люминометра, и определяли световое излучение как показатель активности люциферазы в люминометре (Sirius, Berthold Detection System GmbH, Pforzheim). Для сравнения in vitro использовали растения, помещенные в такие же условия и обработанные водой вместо зооспор. Средняя величина коэффициентов пяти независимых линий активности люциферазы в инфицированных и контрольных вариантах отражает индукцию синтетического промотора при инфицировании. Как видно на Фиг.7, при применении минимального промотора 35S максимальная индукция активности люциферазы с коэффициентом десять достигалась только спустя 72 часа после инфицирования. Все новые минимальные промоторы продемонстрировали здесь отчетливо улучшенную индукцию. Самая сильная индукция с коэффициентом 395 была достигнута спустя 72 часа после инфицирования с минимальным промотором StPSBR (SEQ ID NO: 7). В общем индукция может быть улучшена при применении новых минимальных промоторов через 72 часа после инфицирования, от коэффициента 3,5 при применении минимального промотора StGst (SEQ ID NO: 6) до коэффициента 39,5 при применении минимального промотора StPSBR (SEQ ID NO: 7) по сравнению с минимальным промотором 35S. Примечательно, что у минимальных промоторов наблюдается отчетливое различие в кинетике индукции после инфицирования патогеном. При применении минимального промотора 35S через 72 после инфицирования индукция является отчетливо измеряемой, тоже происходит при применении минимальных промоторов StPSBR, NtTGAA, StGst, NtRBS, а также NpATP2. При применении промоторов NpCABE и NtoEAS сильная активация проявляется уже через 9 часов после инфицирования, и индукция на протяжении оставшегося времени исследования оставалась примерно на достигнутом уровне.
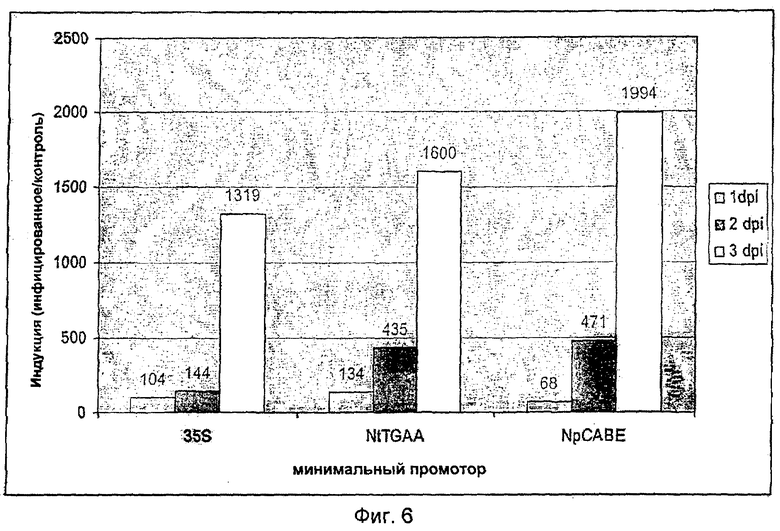
Преимущество новых минимальных промоторов также было показано после слияния с комбинацией цис-элементов 2×S/2×D. Для этого растения картофеля были стабильно трансформированы бинарными векторами p2×S/2×Dluc-kan, p2×S/2×DNpCABEluc-kan и p2×S/2×DNtTGAAluc-kan. Были изготовлены бинарные векторы, в которых 4×Gst1-элемент из вышеописанных бинарных векторов с новыми минимальными промоторами и 4×Gst1-элементом был удален с помощью лизиса BcuI/Eco147I, а элемент 2×S/2×D (SEQ ID NO: 11) вставлен в виде фрагмента BcuI/Eco32I. Бинарными векторами с новой последовательностью трансформировали штамм агробактерий GV3101::pMP90 (Koncz and Schell, 1986) (An, 1987) и проводили селекцию с помощью антибиотика канамицина (50 мг/л). Трансгенные агробактерий внедряли в картофель сорта "Baltica" (Dietze et al., 1995). Трансгенные побеги размножали и в условиях in vitro прививали суспензией зооспор (50.000 спор/мл) Phytophthora infestans. Можно видеть, что при применении цис-регуляторных элементов 2×S/2×D (SEQ ID NO: 11) с минимальными промоторами по настоящему изобретению может быть достигнута фоновая активность, сниженная по сравнению с минимальным промотором 35S (Фиг.5). Одновременно с этим может наблюдаться более сильная индукция синтетических промоторов после прививания трансгенного картофеля Р. infestans (Фиг.6). Усиление индукции в более позднее время (3 дня после инфицирования = 3 dpi (days post infection)) было не столь отчетливо выраженным как то, которое наблюдали при применении 4xGst1-элемента. Через 2 дня после инфицирования при применении новых минимальных промоторов могла быть установлена отчетливо более сильная индукция после проникновения патогенов. Итак, использование этих минимальных промоторов приводит к улучшению кинетики синтетических промоторов, так, что реакция на внедрение патогена развивается раньше по сравнению с применением синтетических промоторов с минимальным промотором 35S.
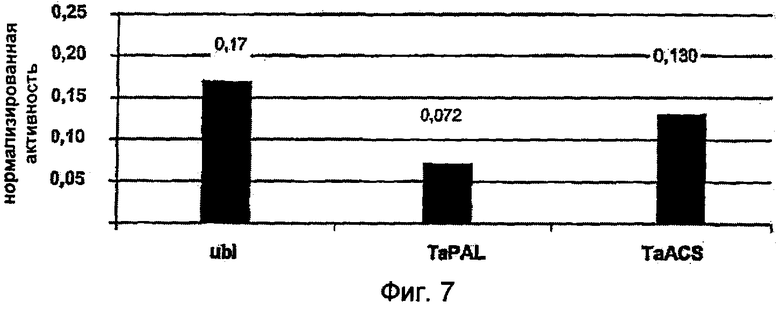
На Фиг.7 показано сравнение нормализированной активности патоген-индуцируемых синтетических промоторов, включающих элемент 2×S/2×D (SEQ ID NO: 11) и минимальные промоторы ubi1 (контрольный промотор), TaPAL (SEQ ID NO: 9) и TaACS (SEQ ID NO: 8), после биолитической трансформации зачаточных листьев пшеницы сорта "Taifun". Как известно, новые минимальные промоторы TaPAL и TaACS в пшенице обладают базовой активностью, сниженной по сравнению с минимальным промотором ubi1. В то время как при использовании минимального промотора ubi1 нормализированная активность равна 0,17, при применении минимального промотора TaPAL она может снижаться до 0,072, а при применении минимального промотора TaACS до 0,13.
На фиг.9 показана плазмида pubiTATARucll, содержащая кДНК с геном люциферазы Renilla reniformis, такая же, как содержащаяся в имеющейся в продаже плазмиде pRL-Null. кДНК находится под контролем минимального промотора ubi1. Минимальный промотор ubi1 содержит область последовательности от -45 до +76 по отношению к сайту начала транскрипции. Для повышения силы экспрессии первый интрон гена ubi1 в его натуральном контексте в плазмиде вставлен перед репортерным геном. Плазмида служит для клонирования цис-регуляторных элементов 2×S/2×D (SEQ ID NO: 11), чтобы изготовить из них синтетический промотор, индуцируемый патогенами. Минимальный промотор ubi1 заменяли на новые минимальные промоторы для улучшения свойств синтетических промоторов.
Литература
An, G. (1987). Binary Ti vectors for plant transformation and promoter analysis. Methods Enzymol. 153: 292-305
Altschul, S. F. et al. (1990), Basic Local Alignment search tool, J. Mol. Biol. 215: 403-410
Becker, D. et al. (1992). New plant binary vectors with selectable markers located proximal to the left T-DNA border. Plant Mol Biol. 29: 1195-1197
Belbahri, L. et al. (2001). A local accumulation of the Ralstonia solanacearum PopA protein in transgenic tobacco renders a compatible plant-pathogen interaction incompatible. Plant J. 28: 419-430
Bendahmane, A. et al. (2002). Constitutive gain-of-function mutants in a nucleotide binding site-leucine rieh repeat protein encoded at the Rx locus of potato. Plant J. 32:195-204
Bhullar, S. (2003). Strategies for the development of functionally equivalent Promoters with minimum sequence homology for transgene expression in plants: cis-elements in a novel DNA context versus domain swapping. Plant Physiol. 132: 988-998
Dietze, J. et al. (1995). Agrobacterium-mediated transformation of potato (Solanumtuberosum). In: Gene Transfer to Plants XXII (Potrykus, I. and Spangenberg, G., eds.). Berlin: Springer Verlag, pp 24-29
Howles, P. et al. (2005). Autoactive alleles of the flax L6 rust resistance gene induce non-race-speeifie rust resistance associated with the hypersensitive response. Mol. Plant-Microbe Interact. 18: 570-582
Keller, H. et al. (1999). Pathogen-induced elicitin production in transgenic tobacco generates a hypersensitive response and non-specific disease resistance. Plant Celi 11:223-235
Koncz, C. and Schell, J. (1986). The promoter of TL-DNA gene 5 controls the tissue specific expression of chimeric genes carried by a novel type of Agrobacterium vector. Mol. Gen. Genet., 204: 383-396
Lü, H. et al. (2000). Construction of chimeric inducible Promoters by elicitors of rice fungal blast pathogen and their expression in transgenic rice. Chinese Science Bulletin 45: 242-246
Oldroyd, G.E.D. and Staskawicz, B.J. (1998). Genetically engineered broad spectrur disease resistance in tomato. Proc. Natl. Acad. Sei. U. S. A. 95: 10300 -10350
Maas, С.et al. (1991). The combination of a novel stimulatory element in the first exon of the maize Shrunken-1 gene with the following introni enhances reporter gene expression up to 1000-fold. Plant Mol Biol. 16: 199-207
Perl-Treves, R. et al. (2004). Early induetion of the Arabidopsis GSTF8 Promoter by specific strains of the fungal pathogen Rhizactonia solani. Mol. Plant-Microbe Interact. 17: 70-80
Rushton, P.J. et al. (2002). Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen- and wound-induced signalling. Plant Cell 14, 749-762
Singh, K.B. (1998). Transcriptional regulation in plants: the importance of combinatorial control. Plant Physiol. 118:1111-1120
Stuiver, M.H. and Gusters, J.H.H.V. (2001). Engineering disease resistance in plants. Nature 411: 865-868
Stuiver, M.H. et al. (1998). Infection-induced expression of the avirulence gene avr9 in transgenic Cf9 tomato plants confers resistance to fungal pathogen attack. 7th International congress of plant pathology, 9-16 August, Edinburgh, Scotland Tang, X. et al. (1999). Overexpression of Pto activates defense responses and confers broad resistance. Plant Cell 11:15-29
Tao, Y. et al. (2000). Mutational analysis of the Arabidopsis nucleotide binding site-leucine-rich repeat resistance gene RPS2. Plant Cell 12: 2541-2554
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ РАСТИТЕЛЬНЫЕ ЦИС-РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ ДЛЯ РАЗРАБОТКИ ПАТОГЕН-ЧУВСТВИТЕЛЬНЫХ ХИМЕРНЫХ ПРОМОТОРОВ | 2012 |
|
RU2660569C2 |
| АУТОАКТИВИРУЮЩИЙСЯ БЕЛОК УСТОЙЧИВОСТИ | 2006 |
|
RU2375453C2 |
| СПОСОБ ПОВЫШЕНИЯ РЕЗИСТЕНТНОСТИ ТРАНСГЕННЫХ РАСТЕНИЙ К ПАТОГЕННЫМ ВОЗДЕЙСТВИЯМ | 2007 |
|
RU2428480C9 |
| БОЛЕЗНЕУСТОЙЧИВЫЕ РАСТЕНИЯ | 2006 |
|
RU2393227C2 |
| СПОСОБ И КОНСТРУКТ ДЛЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА SCBV | 2012 |
|
RU2627595C2 |
| КОНСТРУКЦИЯ И СПОСОБ КОНСТРУИРОВАНИЯ СИНТЕТИЧЕСКОГО ДВУНАПРАВЛЕННОГО РАСТИТЕЛЬНОГО ПРОМОТОРА UBI1 | 2012 |
|
RU2639538C2 |
| ИСКУССТВЕННЫЙ ПРОМОТОР ДЛЯ ЭКСПРЕССИИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДНК В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2003 |
|
RU2326167C2 |
| ПОНИЖАЮЩАЯ РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНА С ПОМОЩЬЮ ИСКУССТВЕННЫХ МИКРОРНК | 2008 |
|
RU2492239C2 |
| КОНСТИТУТИВНЫЕ ПРОМОТОРЫ СОИ | 2014 |
|
RU2714724C2 |
| Промотор pro-SmAMP-D1 из растения звездчатка средняя (Stellaria media L.) для биотехнологии растений | 2022 |
|
RU2799014C1 |











Синтетический патоген-индуцируемый промотор включает минимальный промотор предшествующий ему по меньшей мере один цис-регуляторный элелмент. Минимальный промотор включает мотив последовательности a) dbrmwa или b) twcccmt, который находится после ТАТА-области и перед сайтом начала транскрипции, находящимся в минимальном промоторе. Введение к клетку растения нуклеиновой кислоты, подходящей для возникновения защиты от патогенов, под контролем синтетического промотора с последующей регенерацией из упомянутой клетки целого растения обеспечивает этому растению устойчивость к патогенам. 6 н. и 3 з.п. ф-лы, 9 ил.
1. Патоген-индуцируемый синтетический промотор, подходящий для регуляции транскрипции нуклеиновой кислоты и содержащий минимальный промотор и предшествующий минимальному промотору по меньшей мере один цис-регуляторный элемент, отличающийся тем, что минимальный промотор содержит мотив последовательности
a) dbrmwa или
b) twcccmt
и находится после ТАТА-области и перед сайтом начала транскрипции, находящимся в минимальном промоторе, в котором начинается транскрипция регулируемой нуклеиновой кислоты.
2. Патоген-индуцируемый синтетический промотор по п.1, отличающийся тем, что мотив последовательности в минимальном промоторе повторяется два или более раз.
3. Патоген-индуцируемый синтетический промотор по п.1 или 2, минимальный промотор которого содержит нуклеотидную последовательность согласно любой из SEQ ID NO: 1-9.
4. Патоген-индуцируемый синтетический промотор по п.1, который содержит рядом с минимальным промотором по меньшей мере один цис-регуляторный элемент с нуклеотидной последовательностью согласно любой из SEQ ID NOs: 10-15.
5. Рекомбинантный ген с патоген-индуцируемым синтетическим промотором по любому из пп.1-4.
6. Клетка растения, в ДНК которой интегрирован патоген-индуцируемый синтетический промотор по любому из пп.1-4.
7. Трансгенное растение с клеткой растения по п.6.
8. Семя трансгенного растения по п.7, содержащее патоген- индуцируемый синтетический промотор по п.1.
9. Способ получения растения, устойчивого к патогенам, при котором в клетку растения вводят нуклеиновую кислоту, подходящую для возникновения защиты от патогенов, находящуюся под контролем патоген-индуцируемого синтетического промотора, после чего из этой клетки растения регенерируют растение, отличающийся тем, что патоген-индуцируемый синтетический промотор представляет собой патоген-индуцируемый синтетический промотор по пп.1-4.
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| DATABASE EMBL, Accession no | |||
| Фильтр для улавливания паров ртути из воздуха | 1938 |
|
SU60077A1 |

