Изобретение относится к индуцирующим воспринимаемость трансплантата клеткам моноцитарного происхождения и их получению, а также к их применению для создания воспринимаемости трансплантата.
Предпочтительно, изобретение относится к индуцирующим воспринимаемость трансплантата клеткам (в дальнейшем также называемым TAIC), происходящим из человеческих моноцитов.
Кроме того, изобретение относится к моноклональному антителу GM-7, специфически распознающему индуцирующие воспринимаемость трансплантата клетки по изобретению, получаемые от человека.
Кроме того, изобретение относится к применению антитела GM-7 для выявления и/или выделения индуцирующих воспринимаемость трансплантата клеток.
Термин "трансплантация" в области иммунологии применяют для обозначения пересадки клеток, ткани или органов от одного организма другому. Необходимость трансплантации является результатом открытия того, что многие заболевания можно лечить пересадкой (трансплантацией) здоровых органов, ткани или клеток от одного индивидуума, здоровому в данном отношении - донора, другому индивидууму, страдающему соответствующим заболеванием - реципиенту или хозяину.
В зависимости от родства между донором и реципиентом можно различать следующие типы трансплантатов:
1. Аутологичные трансплантаты. Они представляют собой ткани или клетки, трансплантированные у одного и того же индивидуума из одной области организма в другую.
2. Изогенные трансплантаты. Это включает в себя пересадку между генетически идентичными индивидуумами. Примерами данного типа являются инбредные линии крыс и мышей. У людей изогенными (сингенными) трансплантатами можно считать трансплантаты у генетически идентичных (монозиготных) близнецов.
3. Аллогенные трансплантаты. Данный термин обозначает трансплантаты у генетически различных представителей одного вида, такие как, например, от одного человеческого индивидуума другому или, в случае экспериментальных животных, от одной инбредной линии другой.
4. Ксеногенные трансплантаты. Они включают в себя пересадку между различными видами, такой как, например, пересадка сердца обезьяны человеку.
Как правило, аутологичные и изогенные трансплантаты не вызывают иммунологических проблем в смысле реакции отторжения. Однако это не верно в случае аллогенных и ксеногенных трансплантатов. Такие трансплантаты иммунной системой реципиента распознаются как чужеродные и отторгаются после сравнительно короткого промежутка времени. В случае реакции отторжения аллогенных/ксеногенных трансплантатов центральную роль играют T-лимфоциты (далее называемые T-клетками). Данные клетки распознают чужеродные клетки посредством их главного комплекса гистосовместимости (MHC). Чужеродный комплекс MHC сенсибилизирует реагирующие на антигены T-клетки реципиента, а затем активированные таким образом T-клетки приводят к разрушению донорных клеток или донорной ткани, соответственно. Клетки с другими комплексами MHC также называют "отличающимися по MHC".
Трансплантация аллогенных или ксеногенных клеток, тканей или органов неизбежно приводит к ситуации, когда клетки двух различных индивидуумов, т.е. отличающиеся по MHC клетки, представлены в крови реципиента одновременно. Данный феномен назвали "химеризм". Необходимо делать отличия между микрохимеризмом и макрохимеризмом. Микрохимеризм относится к состоянию, в котором полученные от донора клетки, например, после переливания крови или пересадки костного мозга или после трансплантации богатых лимфоцитами органов, таких как тонкий кишечник или печень, остаются персистирующими у отличающегося по MHC хозяина, но их можно обнаружить только в немногих отдельных случаях. Термин макрохимеризм применяют, когда более чем 5% клеток, обнаруживаемых у реципиента, происходят от донора, при этом в обоих случаях необходимо делать различия между химеризмом крови и органным химеризмом с соответствующим выявлением донорных клеток в крови или органах реципиента.
До некоторой степени химеризм может вести к толерантности по отношению к трансплантатам. Таким образом, в случае индивидуумов, подвергшихся, например, в результате лейкоза или лимфомы, трансплантации аллогенного костного мозга со значительным соответствием структуры MHC у донора и реципиента, после создания соответствующего миелоаблативного состояния (разрушение у индивидуума собственного костного мозга и происходящих из него клеток крови) и последующей трансплантации стволовых клеток наблюдали "донорный химеризм". В случае данных пациентов более чем 99% клеток, обнаруживаемых в крови, происходили от стволовых клеток донора, а данный химеризм формировал основу для толерантности по отношению ко всем органам, трансплантируемым от данного донора. Данное наблюдение подтверждено в ряде клинических примеров последующего донорства почек, печени и долей легкого [Dey B., et al., "Outcomes of Recipients of both bone marrow and solid organ transplants: A review". Medicine (Baltimore) 77, 355-369 (1989)]. При возникновения индукции толерантности существование полного донорного химеризма, определенного как присутствие более чем 90% донорных клеток в периферической крови аллогенного реципиента, не имеет решающего значения; напротив, обнаружено, что для индукции толерантности достаточно доли донорных клеток в размере 5%, называемой "смешанным" химеризмом [ср. обзор Acholonu I.N. и Ildstad S. T., "The role of bone marrow transplantation and tolerance: Organ-specific and cellular grafts", Curr. Opin. Organ. Transplant 4, 189-196 (1999)].
Хотя по существу, таким образом, предоставлено доказательство того, что донорный химеризм, сопровождающий трансплантацию костного мозга (BMTx), индуцирует толерантность, тем не менее, до настоящего времени не являлось возможным произвести успешную аллогенную BMTx в случае групп с полностью отличными по MHC донором и реципиентом. Это происходит вследствие того факта, что необходимая для этого предварительная подготовка, т.е. приведение реципиента в требуемое состояние (см. выше) содержит в себе три существенных вида риска для реципиента. Во-первых, токсичность для костного мозга применяемых химиотерапевтических средств, вводимых совместно с дополнительной радиационной обработкой или без нее, приводит к значительной смертности; во-вторых, существует риск отторжения трансплантированного костного мозга, несмотря на предшествующую подготовку; и, наконец, существует риск заболевания "трансплантат против хозяина" (GvHD) в том случае, когда пересаженные хозяину вместе с трансплантатом T-клетки действуют против организма хозяина [ср. Wolff S.N., "Hematopoietic cell transplant from volunteer unrelated or partially matched related donors: Recent developments." Curr. Opin. Organ. Transplant 5, 372-381 (2000)]. Данное заболевание имеет место каждый раз, когда иммунная система реципиента ослаблена предшествующей подготовкой до такой степени, что пересаженные вместе с костным мозгом T-клетки способны вызывать летальное отторжение у реципиента.
Во избежание указанных выше проблем в настоящее время предпринимаются попытки разработать настолько слабые режимы подготовки реципиента, насколько это возможно. Однако, чем больше различие между MHC донора и реципиента, тем более строгий режим подготовки необходимо выбрать для предотвращения отторжения трансплантированного костного мозга; это снова приводит к увеличенному риску заболевания "трансплантат против хозяина" (GvHD).
До настоящего времени не проводили подробного исследования в отношении времени, в течение которого в крови реципиента должен выявляться индуцированный химеризм для обеспечения состояния стабильной толерантности для донорского органа. Однако достоверные данные, полученные из модели на мышах, указывают на то, что выявляемого в течение двух недель макрохимеризма уже достаточно для возникновения толерантности [Weckerle, T. et al. "Allogeneic bone marrow transplantation with co-stimulatory blockade induces macro-chimerismand tolerance without cytoreductive host treatment. "Nat. Med. 6, 464-469 (2000)].
Необходимость разработки альтернатив доступным в настоящее время иммунодепрессантам для создания толерантности возникает с двух основных точек зрения. С одной стороны клинически доступные в настоящее время иммунодепрессанты не способны предотвращать хронического отторжения трансплантата, а с другой стороны, постоянный прием иммунодепрессантов сопровождается существенными побочными эффектами, приводящим к повышенной чувствительности пациента к вирусным, бактериальным и грибковым инфекциям и создающим значительные риски вследствие формирования малигномы и заболеваний сердечно-сосудистой системы, в частности, вследствие инфаркта миокарда и сердечной недостаточности [см. Wheeler, D. C. and Steiger, J. "Evolution and Etiology of Cardiovascular Diseases in Renal Transplant Recipients" Transplantation 70, 41-45 (2000)].
Munn et al. описали индуцированную макрофагами супрессию T-клеток, основанную на селективной элиминации триптофана и/или увеличении одного или нескольких метаболитов триптофана в окружающей клетку среде [см. Munn et al. "Inhibition of T Cell Proliferation by Macrophage Tryptophan Catabolism" J. Exp. Med. 189, 1363-1372 (1999)]. Начиная с данного исследования, авторы предлагают модифицировать опосредованные T-клетками иммунологические реакции изменением локальной внеклеточной концентрации триптофана, в частности ингибированием или увеличением опосредованного IDO метаболизма триптофана, см., например патенты США 6482416 и 6451840. Таким образом, данный подход предоставляет только дополнительную, независимую от антигена и неспецифическую иммуносупрессию, направленную против T-клеток.
Однако то, что экстренно необходимо, представляет собой клинически приемлемую концепцию для индукции зависящей от донора толерантности, т.е. концепцию, согласно которой иммунный ответ хозяина против тканевых антигенов на пересаженном от донора органе специфически подавлен, но при этом, в остальном, иммунокомпетентность хозяина полностью сохраняется.
Посредством такой концепции в средние сроки можно достичь значительного уменьшения несоответствия между необходимостью органов и предложением органов, так как в таком случае, в результате процессов острого или хронического отторжения, пропадет значительно меньшее количество органов. Недостаток подходящих органов можно продемонстрировать посредством статистических данных по США, датируемых февралем 2001 года:
Это объясняет те огромные усилия на государственном и международном уровне, прилагаемые для разработки концепций индукции зависящей от донора толерантности.
Следовательно, лежащая в основе изобретения проблема представляет собой предоставление способов воздействия на T-клетки организма реципиента таким образом, чтобы они оставались толерантными к соответствующим чужеродным клеткам, тканям или органам и/или воспринимали их в течение длительного времени без отторжения. Характер данных средств и их получение или применение не должны вызывать каких-либо этических и/или правовых проблем и должна существовать возможность их быстрого получения для планируемого терапевтического применения в необходимых для данной цели количествах и при приемлемых затратах на производство.
Для разрешения данной проблемы по настоящему изобретению предоставлены индуцирующие воспринимаемость трансплантата клетки моноцитарного происхождения позвоночных, конкретно млекопитающих, а более предпочтительно - людей. Неожиданно обнаружили, что модифицированные по изобретению моноциты донорской крови (донорского органа) в случае предоперационного и/или послеоперационного введения способны защищать организм реципиента от активированных чужеродным комплексом MHC T-клеток его собственного организма, таким образом предотвращая отторжение трансплантата. При применении в качестве "клеточных терапевтических средств" для индукции толерантности к трансплантату, модифицированные клетки у соответствующих пациентов не вызывают никаких или не вызывают заслуживающих внимания побочных эффектов в смысле клеточного отторжения, индукции опухолей, в частности злокачественных опухолей, и заболевания "трансплантат против хозяина".
Неожиданно обнаружили, что способ по изобретению приводит к модификации моноцитов in vitro таким образом, что получают клетки, способные после инъекции аллогенному реципиенту без иммуносупрессии предотвращать природный иммунный ответ против клеток или ткани донора и способные, таким образом, циркулировать в периферической крови, по меньшей мере, в течение трех недель. После введения приблизительно 105 клеток/кг массы тела (BW), образующийся в результате химеризм из связывающихся с GM-7 клеток (см. ниже) находится в области 5-20%. Данный временный химеризм индуцирует
a) долговременную воспринимаемость для последующих трансплантаций органов от того же донора, предпочтительно в пределах 10 дней после внутривенного введения клеток и
b) долговременную воспринимаемость в сочетании с кратковременным иммуносупрессирующим воздействием, когда применяют клетки после трансплантации органа.
Как показано в примере 10, иммуносупрессирующее действие клеток по изобретению не связано с индуцированной макрофагами супрессией T-клеток посредством экспрессии разрушающего триптофан фермента индоламин-2,3-диоксигеназы (IDO), как исследовано Munn et al., (см. выше). Предпочтительно, TAIC по изобретению индуцируют у реципиента специфическую толерантность к донору посредством инактивации аллореактивных T-клеток, с одной стороны, и посредством формирования у реципиента регуляторных T-клеток, с другой стороны, см. примеры 12 и 13.
Следовательно, полученные из моноцитов макрофаги, ингибирующие пролиферацию T-клеток посредством опосредованной IDO элиминации триптофана в окружающей клетки среде не включены в данное изобретение.
Описание фигур
Фиг.1: Определение посредством проточной цитометрии связывающей способности GM-7 в отношении исходных моноцитарных клеток перед модификацией по изобретению (диаграмма на левой стороне) и после нее (диаграмма на правой стороне). Ось x означает количество связавшихся клеток.
Фиг.2A: Кривые выживания Каплан-Мейер трансплантатов сердца после гетеротопической трансплантации сердца с предоперационным введением полученных от донора TAIC и без него в модели на крысах (n = 10 животных в группе).
Фиг.2B: Гистологический препарат аллогенного трансплантата LEW на POD (сутки после операции) 150 после гетеротопической трансплантации в брюшную полость реципиентной крысы DA, предварительно обработанной 106 TAIC, полученных из LEW на сутки -7. Кратность увеличения: 40.
Фиг.2C: Гистологический препарат из тимуса крысы DA, предварительно обработанной 106 TAIC, полученных из LEW, после мечения препарата моноклональными антителами I1.69, специфичными для MHC класса I LEW. Кратность увеличения: 40.
Фиг.2D: Определение посредством проточной цитометрии полученных от донора клеток у четырех реципиентных крыс DA, не подвергнутых иммуносупрессии с последующей инъекцией TAIC, полученных из LEW, (крысы 1-3: 106 клеток/кг × BW; крыса 4: 104 клеток/кг × BW).
Фиг.3: Кривые выживания Каплан-Мейер трансплантатов сердца после гетеротопической трансплантации сердца с предоперационным введением TAIC, полученных от донора, и без него в сочетании с начальным введением 4 циклов циклоспорина A (CSA; 5 мг/кг × BW) в модели на крысах (n=6-10 животных в группе).
Фиг.4A-C: Кривые выживания Каплан-Мейер трансплантатов сердца (A), печени (B) и почки (C) после гетеротопической (A и C) или ортотопической (B) трансплантации, соответственно, с предоперационным введением TAIC, полученных от донора, и без него (n=6 животных в группе).
Фиг.4D-F: Гистологические препараты трансплантатов почки (D), печени (E) и кожи (F) DA после гетеротопической трансплантации в реципиентную крысу LEW.
Фиг.5A: Кривые выживания Каплан-Мейер трансплантатов сердца после гетеротопических трансплантаций сердца при различных сочетаниях инбредных линий с предоперационным введением полученных от донора TAIC в сочетании с начальным введением 4 циклов CSA (5 мг/кг × BW) в модели на крысах (n=6 животных в группе).
Фиг.5B: Кривые выживания Каплан-Мейер трансплантатов сердца после гетеротопических трансплантаций сердца через 7 суток после введения различным образом обработанных моноцитов крови от донорных животных LEW (n=6 животных в группе).
Фиг.6A: Кривые выживания Каплан-Мейер трансплантатов легкого после ортотопических левосторонних трансплантаций легкого у свиней ("аутбредные карликовые свиньи") с применением тройной иммуносупрессии CSA, азатиоприном (AZA) и стероидами (STE) (n=4 животных в группе).
Фиг.6B: рентгеновский снимок грудной клетки (заднепередний способ) двух свиней с односторонней пересадкой левого легкого ("аутбредные карликовые свиньи") на POD (сутки после операции) 41 и 55. Тройную иммуносупрессию CSA, AZA и стероидами прерывали на 28 сутки после трансплантации.
Фиг.7: Смешанная культура лимфоцитов из CD14+-моноцитов индивидуума B
(GM-7-: серый столбец; GM-7+: черный столбец), отвечающих клеток донора A с несоответствующим MHC и облученных клеток донора B для сравнения супрессорной активности CD14+/GM-7+- и CD14+/GM-7--клеток.
Фиг.8: Смешанная культура лимфоцитов из стимулированных PHA лимфоцитов (PhaLy) и TAIC ("Mo+Ly" или "Mo"), предварительно инкубированная в двух экспериментах с ингибитором (1-MT) индоламин-2,3-диоксигеназы (IDO) для определения влияния 1-MT на супрессорную активность TAIC.
Фиг.9: Определение проточной цитометрией количества CD14+-моноцитов и CD2+-лимфоцитов в моноцитарной фракции, а также количества CD14+/CD3+-клеток, действующих в качестве TAIC, для определения влияния очистки клеток для обогащения моноцитами в начале культивирования на формирование иммуносупрессирующих CD14+/CD3+-клеток.
Фиг.10: Определение проточной цитометрией экспрессии GM-7 после операции в крови пациентов до (левая фигура) и после (правая фигура) инъекции TAIC, для определения влияния TAIC на экспрессию GM-7 в клетках крови in vivo.
Сущность изобретения
Основные стадии способа по изобретению для получения TAIC моноцитарного происхождения включают в себя:
(a) Выделение моноцитов из крови, предпочтительно из крови человека.
(b) Размножение моноцитов в подходящей культуральной среде, содержащей макрофагальный колониестимулирующий фактор (в дальнейшем обозначаемый как M-CSF) в качестве стимулирующего рост средства.
(c) Стимуляцию моноцитов интерфероном γ (в дальнейшем называемом γ-IFN); и
(d) Получение сформированных на стадии c) индуцирующих воспринимаемость трансплантата клеток посредством выделения клеток из культуральной среды.
По изобретению показано, что стимуляция γ-IFN представляет собой решающую стадию в получении TAIC (см. пример 7).
В настоящем изобретении термин TAIC (индуцирующие воспринимаемость трансплантата клетки моноцитарного происхождения) обозначает клеточную популяцию, получаемую на стадии (d) описанного выше способа. Данная клеточная популяция вместе с происходящими из моноцитов эффективными по изобретению клетками также содержит лимфоциты, см. пример 11, а также, необязательно, дополнительные клетки, полученные из лейкоцитарной пленки, например гранулоциты. Количество полученных из моноцитов клеток в популяции TAIC предпочтительно составляет от 50 до 90%, более предпочтительно - от 60 до 70% относительно общего количества клеток.
В настоящем изобретении термин "общее количество клеток" относится к количеству живых клеток в рассматриваемой клеточной популяции. Данное количество можно определить "способом исключения по красителю трипановый синий", так как данный краситель позволяет отличать живые клетки от неживых оптическими средствами.
По изобретению, TAIC для индукции воспринимаемости трансплантата, как правило, можно применять в количестве 104-106 клеток на килограмм массы тела, предпочтительно - в количестве 105 клеток на килограмм массы тела. Введение TAIC в случае больших различий в MHC можно проводить повторно, предпочтительно три раза через интервалы длительностью приблизительно 10 суток, где введение клеток можно проводить до трансплантации или после нее (см. ниже). Общее количество клеток, необходимых для данной цели, можно получить в пределах от 6 до 8 суток после забора крови.
И в тестах на животных и в культуре доказано, что клетки по изобретению (TAIC) не создают риска формирования малигномы; данный результат являлось невозможным ожидать в любом другом случае вследствие природы исходных моноцитарных клеток, от которых происходят клетки по изобретению.
Как раскрыто ниже, из фракции клеток, существующей в TAIC, происходящих из моноцитов, можно выделить дополнительные субпопуляции клеток с оптимизированным действием по изобретению.
После культивации in vitro и стимуляции исходных клеток (моноцитов) интерфероном γ формируются TAIC, содержащие субпопуляцию клеток, см. пример 9, связывающуюся с моноклональным антителом GM-7, экспрессируемым гибридомной клеточной линией DSM ACC2542. Моноклональное антитело GM-7 представляет собой антитело иммуноглобулинового изотипа IgG2a, легкая цепь которого представляет собой изотип каппа. Характеристическим свойством данного антитела является его узконаправленная способность связываться с моноцитами, модифицированными посредством условий культивирования по изобретению, тогда как исходные моноцитарные клетки не распознаются, т.е. связывания антитела с исходными клетками не происходит (см. пример 9). Кроме того, на 20 добровольцах показано, что GM-7 не связывается с человеческими клетками в периферической крови, см. Фиг.10.
Антитело получали посредством иммунизации мышей TAIC, полученными из человеческих моноцитов, с применением способов, известных специалистам в данной области (Davis, W. C. "Methods in Molecular Biology: Monoclonal Antibody Protocols", New York: Humana Press Inc. Totowa, 1995). Затем посредством слияния образующей антитела B-клетки и миеломной клетки мыши получали гибридомную линию. Применяемые для получения таких клеточных линий способы в настоящее время известны в данной области (Davis, W. C. "Methods in Molecular Biology: Monoclonal Antibody Protocols", New York: Humana Press Inc. Totowa, 1995; Kohler, G., Milstein, C. "Continuous cultures of fused cells secreting antibody of predefined specificity", Nature 256, 495-497 (1975)). Продуцирующую антитела GM-7 гибридомную клеточную линию депонировали по правилам Будапештского соглашения в DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkultur GmbH, Braunschweig, Germany) под инвентарным № DSM ACC2542.
На Фиг.1 показана связывающая способность GM-7 по отношению к моноцитарным клеткам после модификации in vitro по изобретению, определенная посредством проточной цитометрии. Можно видеть, что полученные непосредственно из лейкоцитарной пленки несущие CD14 моноциты не связывают антитело GM-7 (заштрихованная серым область соответствует незаштрихованной с антителами контроля). Напротив, после культивирования в присутствие M-CSF и стимуляции γ-IFN, часть моноцитов экспрессирует антиген, распознаваемый моноклональным антителом GM-7. Моноклональное антитело GM-7 характеризуется как изотип κ-IgG2a. Следовательно, способ по изобретению приводит к изменению фенотипического профиля экспрессии антигенов на клеточной мембране модифицированных моноцитов (Фиг.1).
Моноклональное антитело GM-7 специфически связывается с той клеточной популяцией, которая среди образуемых способом по изобретению клеток индуцирует наиболее эффективную воспринимаемость трансплантата (см. Фиг.9).
Таким образом, предпочтительное осуществление изобретения относится к таким TAIC, которые способны к связыванию с антителом GM-7. Данные клетки впоследствии обозначали как TAICGM-7.
Следовательно, антитело по изобретению GM-7 представляет собой чрезвычайно эффективное и легкое в обращении средство для отбора и очистки индуцирующих воспринимаемость трансплантата клеток (TAIC). Посредством антитела по изобретению возможно получать гомогенную и высокоэффективную популяцию TAIC.
По предпочтительному осуществлению изобретения образуемые на стадии c) описанного выше способа по изобретению индуцирующие воспринимаемость трансплантата клетки, экспрессирующие связывающийся с антителом GM-7 антиген, можно или непосредственно выделить из клеточной среды после стадии c) или их можно выделить из клеточной популяции, полученной после выделения клеток из клеточной среды на стадии d) указанного выше способа по изобретению, посредством связывания с антителом GM-7, получаемого посредством гибридомной клеточной линии DSM ACC2542.
Для отбора TAIC по изобретению антитело приводят в контакт с образцом в условиях, позволяющих связывание антитела с индуцирующими воспринимаемость трансплантата клетками, присутствующими в образце. Затем образующиеся вследствие реакции связывания комплексы выделяют из образца. Для данной цели антитело можно иммобилизовать на носителе перед контактом с образцом; например, его можно связать с матриксом, пригодным для применения в хроматографии или с так называемыми "магнитными шариками". Данная процедура позволяет выделять и концентрировать индуцирующие воспринимаемость трансплантата клетки из образца большого объема.
Для получения индуцирующих воспринимаемость трансплантата клеток после выделения реакционного комплекса из образца связь между антителом и индуцирующими воспринимаемость трансплантата клетками разрушают. Это можно проводить известными в настоящее время в данной области способами, такими как, например, конкурентное замещение или отмывкой растворами солей. Соответствующие способы описаны, например, у Utz U. et al. ("Analysis of the T-cell Receptor repertoire of human T-cell leukemia virus type-1 (HTLV-1) Tax-specific CD8+ Cytotoxic T Lymphocytes from patients with HTLV-1 associated disease: Evidence for the oligoclonal expansion "J. of Virology Feb. 1996, 843-851).
Кроме того, моноклональное антитело GM-7 позволяет проводить качественное и количественное выявление индуцирующих воспринимаемость трансплантата клеток моноцитарного происхождения по изобретению в образцах крови и/или ткани пациента in vitro. Данный пациент, например, может представлять собой реципиента, которому необходимо трансплантировать или уже трансплантирован орган. Формирование реакционных комплексов в образце, указывающих на присутствие и, если применимо, количество индуцирующих воспринимаемость трансплантата клеток выявляют известными способами.
Для выявления реакционных комплексов, например, в данном случае, возможно непосредственно связывать ("метить") антитело GM-7 с детектируемой молекулой, например, ковалентно связанной с антителом. В области молекулярной диагностики описаны в большом количестве пригодные детектируемые молекулы и включают в себя, среди других, флуоресцентные красители, такие как флуоресцеинизотиоцианат или тетраметилродамин-5-изотиоцианат, люминесцентные красители, радиоактивно меченные молекулы и ферменты, такие как, пероксидазы (ср. Lottspeich, F., Zorbas, H. "Bioanalytik", Spektrum Akademischer Verlag GmbH, Heidelberg-Berlin, 1998).
Выявление антител проводят в зависимости от предварительно выбранной для мечения молекулы. В контексте настоящего изобретения антитело GM-7 связывали с флуоресцентной молекулой флуоресцеинизотиоцианата (FITC) так, чтобы выявление антитела являлось возможным проводить посредством проточной цитометрии и/или флуоресцентной микроскопии. Способы мечения антител FITC хорошо известны специалисту в данной области, работающему в данной сфере.
Альтернативно, реакционный комплекс можно также выявлять в двустадийном процессе с применением вторичных антител. В данном контексте немеченое антитело GM-7 в реакционном комплексе можно выявлять с применением дополнительного меченого антитела (ср. Lottspeich, F., Zorbas, H. "Bioanalytik", Spektrum Akademischer Verlag GmbH, Heidelberg-Berlin, 1998). Данный двустадийный способ выявления является значительно более чувствительным, чем прямое выявление связывания антитела по изобретению, так как с одним антителом GM-7 могут связываться несколько вторичных антител (усиление сигнала).
Следовательно, антитело GM-7 позволяет выявлять TAIC в периферической крови пациента, обработанного TAIC, например, в форме "мониторинга", в течение которого через конкретные временные интервалы определяют количество клеток в периферической крови. В настоящем изобретении в тестах на животных обнаружено, что присутствие полученных из моноцитов донора TAIC в периферической крови реципиента трансплантата коррелирует с толерантностью к трансплантируемому органу. Следовательно, данное открытие позволяет клиницистам постепенно прекратить или уменьшить дозировки необязательно вводимых иммунодепрессантов. Ранее с клинической точки зрения являлось невозможным предоставить точное доказательство того, проявляет ли иммунная система пациента толерантность в данный момент времени после трансплантации.
Как очевидно специалисту в данной области также возможно получение моноклональных антител к TAIC из моноцитов позвоночных, не являющихся людьми, в частности из моноцитов приматов и свиней, модифицированных по изобретению. В данном отношении иммунизацию соответствующего животного-хозяина и получение соответствующей гибридомной клеточной линии проводят, как описано для TAIC человеческого происхождения. Получение эффективных популяций TAIC других видов представляет собой важный вклад в область медицины ксеногенной трансплантации.
Особенно предпочтительное осуществление изобретения относится к субпопуляции TAIC по изобретению, одновременно экспрессирующей на своей клеточной поверхности антигены CD3 и CD14. Данные клетки далее обозначают как TAICCD3+/CD14+. О таких клетках в данной области до настоящего времени не сообщали. Моноциты и известные происходящие из моноцитов клетки несут поверхностный маркер CD14, однако они в то же время дополнительно не несут поверхностный маркер CD3.
Одновременно экспрессирующие поверхностные антигены CD3 и CD14 TAIC можно или непосредственно выделить из индуцирующих воспринимаемость трансплантата клеток, образуемых на стадии c) указанного выше способа по изобретению, или их можно выделить из клеточной популяции, полученной после выделения клеток из культуральной среды по стадии d) указанного выше способа по изобретению, или, альтернативно, их можно выделить из популяции TAICGM-7.
Кроме того, в настоящем изобретении показано, что TAICCD3+/CD14+ интенсивно экспрессируют гены Foxp3, CTLA4 и ген интегрина αEβ7 (см. пример 12). Напротив, исходные моноциты не экспрессируют данные гены совсем или экспрессируют их в малой степени. Следовательно, стимуляция экспрессии генов Foxp3, CTLA4 и гена интегрина αEβ7 представляет собой характеристику клеток TAICCD3+/CD14+.
Как обсуждается в примере 12, экспрессия маркеров Foxp3, CTLA4 и интегрина αEβ7 ранее описана только для регулирующих T-лимфоцитов. T-лимфоциты, одновременно экспрессирующие поверхностные антигены CD4 и CD25, представляют собой субпопуляцию регуляторных T-лимфоцитов, которые также обозначают как "супрессорные клетки". Их функцией является подавление иммунного ответа в организме. В частности, Foxp3 рассматривают как специфический транскрипционный фактор, служащий в качестве контрольного гена для развития регуляторных T-клеток, и специфически экспрессируемый данными клетками. По изобретению предпочтительно, чтобы клетки TAICCD3+/CD14+ экспрессировали, по меньшей мере, 1×10-9, более предпочтительно - по меньшей мере, 5×10-9, а в особенно предпочтительном способе - по меньшей мере, 1×10-8 мкг РНК Foxp3 на мкг общей РНК.
Подобным образом, CTLA4 рассматривают в качестве маркера для выявления регулирующего действия T-лимфоцитов, в частности, несущих CD4/CD25 T-лимфоцитов (см. литературу, цитируемую в примере 12). По изобретению клетки TAICCD3+/CD14+ предпочтительно должны экспрессировать, по меньшей мере, 5×10-7, более предпочтительно - по меньшей мере, 3×10-6, а в особенно предпочтительном способе - по меньшей мере, 5×10-6 мкг РНК CTLA4 на мкг общей РНК.
Интегрин αEβ7, распознающий эндотелиальный кадгерин, недавно описан Lehmann et al. в PNAS 99, страницы 13031-13036 (2002) в качестве нового маркера субпопуляции высокоэффективных регуляторных T-лимфоцитов, взаимодействующих с эпителиальным окружением. Экспрессия РНК интегрина αEβ7 по изобретению предпочтительно должна составлять в клетках TAICCD3+/CD14+, по меньшей мере, 1×10-12, более предпочтительно - по меньшей мере, 1×10-11, а в особенно предпочтительном способе - по меньшей мере, 1×10-10, а наиболее предпочтительно - по меньшей мере, 5×10-9 мкг РНК на 1 мкг общей РНК.
Как показано в таблице в примере 12, прямое совместное культивирование TAIC по изобретению с лимфоцитами приводит к значительному увеличению количества регуляторных T-лимфоцитов, в частности двойных позитивных по CD4/CD25 клеток в популяции лимфоцитов с сильной положительно регулируемой экспрессией генов Foxp3, CTLA4 и гена интегрина αEβ7. Кроме того, в примере показано, что данный эффект не наблюдают, если TAIC совместно культивируют с лимфоцитами непрямым способом.
Данные результаты означают, что стимуляция формирования и/или размножения регуляторных T-лимфоцитов посредством TAIC вовлечена в индукцию воспринимаемости трансплантата посредством TAIC по изобретению.
В примере 13 подтверждается данное предположение. В данном примере лимфоциты от реципиентных животных из примеров 3, 4, 5, 6 и 7 инкубировали in vitro с TAIC от соответствующих донорных животных. Для индукции толерантности, TAIC, предварительно инкубированные с лимфоцитами от реципиентов, инъецировали животным вместо TAIC. Таким образом, данным способом также можно индуцировать зависящую от донора толерантность, в то же время животные, которым вводили лимфоциты реципиентов, не культивированные совместно с полученными от доноров TAIC, не развивали толерантность.
TAIC по изобретению можно применять в качестве фармацевтического препарата. Клетки, полученные на стадии d) способа по изобретению, описанного выше, можно применять непосредственно. Приблизительно 10-50% общего количества полученных таким образом клеток в популяциях сформировано лимфоцитами и гранулоцитами, происходящими из исходного изолята моноцитов (лейкоцитарная пленка). Данные клетки поддерживают формирование происходящих из моноцитов TAIC на стадии культивирования по изобретению (см. пример 11); они не препятствуют индукции толерантности, если TAIC по изобретению применяют в качестве фармацевтического препарата.
Однако по дополнительным предпочтительным осуществлениям изобретения из общей популяции TAIC, полученной способом по изобретению (см. выше), можно выделить субпопуляции TAICGM-7 и/или TAICCD3+/CD14+ и их можно применять для индукции толерантности.
В культуральной среде (см. пример 2) без потери способности индуцировать толерантность TAIC или TAICGM-7 и/или TAICCD3+/CD14+ могут оставаться, по меньшей мере, 48 часов.
Для применения в качестве фармацевтического препарата TAIC или субпопуляции TAICGM-7 и/или TAICCD3+/CD14+, суспендированные, например, в человеческой сыворотке AB (универсально пригодной для применения), можно вводить внутривенно в качестве кратковременной трансфузии. Для индукции воспринимаемости трансплантата в случае аллогенной трансплантации, полученные из моноцитов донора TAIC или субпопуляции TAICGM-7 и/или TAICCD3+/CD14+ можно инъецировать отличающемуся по MHC реципиенту или до операции или после нее. В случае введения до операции TAIC необходимо инъецировать от одного до трех раз приблизительно за 1 неделю до операции. В случае введения после операции период между операцией и однократным введением клеток должен составлять не более чем 7 суток. Тогда TAIC по изобретению или субпопуляции TAICGM-7 и/или TAICCD3+/CD14+ способны оказывать сопротивление T-клеточному ответу иммунной системы реципиента против трансплантата и находиться в крови реципиента в течение достаточно длительного периода времени, чтобы гарантировать долговременную воспринимаемость трансплантата.
Предоперационную внутривенную инъекцию можно рассматривать в связи с донорством от живущих индивидуумов; однако, если рассматривать донорство от мертвых (кровь и органы трупов) предпочтительным может являться послеоперационное введение TAIC по изобретению или субпопуляций TAICGM-7 и/или TAICCD3+/CD14+. В случае донорства от трупов, тело донора с целью консервации промывают перфузионной средой посредством канализации основной артерии органа. В данном случае венозную кровь, как правило, отбирают через полую вену и выбрасывают. Для получения TAIC для применения по изобретению, венозную кровь можно собирать и обрабатывать как описано в примере 1. Альтернативно в случае донорства от мертвых, TAIC также можно получать из клеток (лимфоцитов и моноцитов) из селезенки донора.
В случае послеоперативного применения TAIC по изобретению применительно к донорству от мертвых промежуток между трансплантацией и применением клеток можно перекрыть сочетанием с иммунодепрессантами для предотвращения острого отторжения органа в промежутке между трансплантацией и приготовлением TAIC, полученных из крови донора. В данном контексте можно рассматривать терапевтические сочетания с традиционными иммунодепрессантами, например, такими как ингибиторы кальциневрина, такие как циклоспорин A (CSA) или такролимус или с азатиоприном (AZA), микофенолят мофетила, рапамицин, моноклональные антитела (ATG, ALG, но ни с диклизиумабом, ни с базиликсимабом) или стероиды (STE). Присутствие иммунодепрессантов (за исключением известных моноклональных антител к α-рецептору IL-2, таких как диклизиумаб и базиликсимаб) в крови реципиента не оказывает негативного воздействия на эффективность иммунодепрессантов с одной стороны или TAIC с другой стороны.
В данном смысле клетки моноцитарного происхождения, модифицированные по изобретению, можно применять в качестве "носителей для переноса толерантности" для любого клеточного трансплантата (такого как островковые клетки, гепатоциты, стволовые клетки взрослого и для любого другого запрограммированного типа клеток или типа тканей) и органа (такого как, например, почка, печень, сердце) поскольку они являются генетически идентичными трансплантируемым клеткам (органам), т.е. они должны происходить от самого донора или от его идентичного близнеца. TAIC выполняют свою защитную функцию, позволяя трансплантированным клеткам/органам адаптироваться в новом окружении, таким образом, предохраняя реципиента от побочных эффектов долговременного лечения иммунодепрессантами.
Подробное описание изобретения
Исходные клетки для способа по изобретению представляют собой моноциты крови. Предпочтительно они представляют собой моноциты из человеческой крови. С целью индукции воспринимаемости трансплантата клетки должны происходить от донора трансплантата (или его идентичного близнеца). В случае ксеногенных трансплантаций, например, органов обезьяны или свиньи человеку, TAIC по изобретению должны, следовательно, происходить из моноцитов соответствующего донорного животного.
Для получения моноцитов кровь после обычной обработки антикоагулянтом с применением известных в данной области способов, предпочтительно центрифугирования, вначале разделяют на плазму и белые и красные кровяные клетки. После центрифугирования плазма находится в супернатанте; ниже его находится слой, содержащий целые белые кровяные клетки. Данный слой также обозначают как лейкоцитарная пленка. Ниже его находится фаза, содержащая красные кровяные клетки (гематокрит).
Применительно к способу по изобретению слой лейкоцитарной пленки вначале выделяют и разделяют для получения моноцитов, например, посредством центрифугирования известными способами. По предпочтительному осуществлению способа слой лейкоцитарной пленки помещают на разделяющую лимфоциты среду (Ficoll-Hypaque) и центрифугируют (см. пример 1). В примере 1 описано предпочтительное осуществление изобретения, где эритроциты и мертвые клетки, еще содержащиеся в лейкоцитарной пленке, отделяют центрифугированием, а белые клетки крови, включающие в себя моноциты, присутствуют в виде отдельной фракции на среде для разделения. Далее светлую фракцию моноцитов можно осторожно отобрать пипеткой и, для обогащения выделенной фракции моноцитами, повторно центрифугировать и отмыть. В ходе процесса моноциты собираются на дне центрифужного сосуда вместе с частью лимфоцитов.
По особенно предпочтительному осуществлению способа по изобретению условия для получения содержащей моноциты фракции контролируют так, чтобы фракция вместе с моноцитами относительно общего количества клеток содержала 10-50% лимфоцитов. Предпочтительно фракция содержит приблизительно 50-90%, а в особенно предпочтительном способе - 60-70% моноцитов и приблизительно 10-50 %, а в особенно предпочтительном способе - 20-50 % лимфоцитов, каждые по отношению к общему количеству клеток, где различия необязательно возникают из-за гранулоцитов.
Как показано в примере 11, присутствие лимфоцитов, в пределах от 20 до 30% относительно общего количества клеток, во время культивирования исходных моноцитов с M-CSF и интерфероном γ, приводит к образованию значительно большего количества двойных положительных по CD3/CD14 TAIC, чем в том случае, когда присутствует только малое количество лимфоцитов (приблизительно 5%).
Для получения достаточного качества TAIC вначале необходимо обеспечить размножение моноцитов. Для данной цели можно применять известную пригодную для моноцитов ростовую среду; однако, среда должна содержать ростовой фактор M-CSF (макрофагальный колониестимулирующий фактор). M-CSF (также называемый CSF-1) продуцируют моноциты, фибробласты, лимфоциты и эндотелиальные клетки. Концентрация M-CSF в культуральной среде, может предпочтительно составлять от 2 до 20 мкг/л среды, более предпочтительно - от 4 до 6 мкг/л и в особенно предпочтительном способе - 5 мкг/л.
Потом или одновременно клетки необходимо стимулировать γ-IFN, т.е. культивировать в присутствие γ-IFN. Стимуляцию моноцитов γ-IFN проводят после начальной фазы размножения продолжительностью от 3 до 6 суток в среде культивирования с ростовым фактором. Предпочтительно на 4 сутки после начала культивирования проводят стимуляцию γ-IFN, и данную стимуляцию продолжают в течение периода предпочтительно от 24 до 72 часов, более предпочтительно - 48 часов в условиях инкубатора, т.е. при 37°C и в атмосфере с 5% CO2.
Концентрация γ-IFN в среде может составлять от 0,1 до 20 нг/мл, предпочтительно - от 1 до 10 нг/мл и особенно предпочтительно - 5 нг/мл.
Стимуляцию γ-IFN можно начинать одновременно с размножением моноцитов в содержащей ростовой фактор среде. Однако как указано выше предпочтительна стимуляция после периода от 3 до 6 суток начальной стадии размножения. Предпочтительно, чтобы размножение клеток и стимуляция γ-IFN, в сумме, не продолжались более 8 суток. В любом случае обработку γ-IFN необходимо проводить так, чтобы после стадии размножения она продолжалась, по меньшей мере, в течение 24 часов и не более 72 часов, предпочтительно 48 часов. Следовательно, период для размножения и стимуляции клеток в сумме должен длиться предпочтительно от 4 до 8 суток.
По предпочтительному осуществлению изобретения размножение и стимуляцию интерфероном γ проводят так, как указано в примере 2, таким образом, что моноциты сначала размножаются в культуральной среде с ростовым фактором, а γ-IFN добавляют к культуральной среде после периода от 3 до 6 суток в таком количестве, чтобы получить в среде концентрацию от 0,1 до 20 нг/мл, предпочтительно - от 1 до 10 нг/мл, а особенно предпочтительно - 5 нг/мл.
Предпочтительно способ по изобретению проводят в сосуде, поверхность которого предварительно покрыта фетальной телячьей сывороткой (FCS) или альтернативно человеческой сывороткой AB (см. пример 2). Нанесение FCS можно проводить посредством покрывания поверхности сосудов для культивирования FCS перед применением, и после периода взаимодействия в течение нескольких часов, в частности от 4 до 72 часов, предпочтительно 12-48 часов и в особенности 24 часов, и удалять FCS, не прикрепившейся к поверхности подходящим образом.
В течение стадии культивирования клетки оседают на дне сосуда для культивирования приблизительно после 24 часов. Вследствие их адгезивных свойств моноциты и полученные из моноцитов в течение процесса TAIC прикрепляются к дну соответствующего культурального сосуда. Если культуральную среду, как описано в примере 2, меняют в течение культивации, вначале осторожно удаляют супернатант, например, посредством отбора пипеткой или декантацией и потом заливают свежую культуральную среду. Однако, предпочтительно, клетки, прикрепляющиеся ко дну, не промывают или промывают очень осторожно, так что любые присутствующие лимфоциты не удаляются.
Удаление прикрепившихся клеток проводят механически, например посредством тонкого клеточного скребка или шпателя.
Однако по предпочтительному осуществлению способа по изобретению полное удаление клеток проводят обработкой пригодным ферментом, например, трипсином (см. пример 2). Раствору трипсина (от 0,1 до 0,025 г/л, предпочтительно 0,05 г/л) можно дать возможность действовать на клетки от 2 до 10 минут при температуре от 35°C до 39°C, предпочтительно - при 37°C, в присутствии 5% CO2.
Активность фермента затем блокируют обычным способом, а освободившиеся после данного воздействия плавающие TAIC можно получить обычным способом посредством центрифугирования. Далее они пригодны для немедленного применения, необязательно в суспензии в пригодной среде, например, в PBS. Однако их также можно в течение нескольких суток, в частности приблизительно в течение от 2 до 3 суток, сохранять в питательной среде (см. пример 2); где данная среда для сохранения не должна содержать ни ростового фактора, ни γ-IFN. В такой питательной среде клетки можно сохранять в виде TAIC, по меньшей мере, в течение 48 часов.
Для хранения в течение более длительных периодов клетки можно подвергнуть глубокой заморозке. Протоколы для глубокой заморозки живых клеток в настоящее время известны в данной области, ср. Griffith M. et al., ("Epithelial Cell Culture, Cornea", в Methods of tissue engineering, Atala A. и Lanza R. P., Academic Press 2002, chupter 4, pages 131-140). Предпочтительная суспензионная среда для глубокой заморозки клеток по изобретению представляет собой FCS, содержащую DMSO.
По одному из осуществлений изобретения клеточную суспензию, содержащую индуцирующие воспринимаемость трансплантата клетки, полученные на стадиях c) или d) можно дополнительно очистить относительно клеток, связывающихся с антителом GM-7, так чтобы получить субпопуляцию TAICGM-7. Способы для такой очистки подробно описаны выше.
По дополнительному предпочтительному осуществлению изобретения из популяции TAIC, выделяют такие клетки, которые одновременно экспрессируют на своих клеточных поверхностях антигены CD3 и CD14.
Способы для выделения таких клеток известны в данной области. Примеры таких способов представляют собой "сортировку клеток с активированной флуоресценцией" (FACS), "иммунологическую сортировку на магнитных шариках" и "магнитную сортировку активированных клеток" (MACS) или так называемый способ "розеткообразования" [см Gmelig-Meyling F. etal. "Simplified procedure for the separation of human T- and non-T-cells", Vox Sang. 33, 5-8 (1977)].
Выделение субпопуляции TAIC TAICCD3+/CD14+ можно проводить непосредственно из популяции TAIC, полученной на стадиях c) или d) описанного выше способа по изобретению, или из субпопуляции TAICGM-7. Последний способ обработки означает, что произойдет постепенное обогащение TAICCD3+/CD14+.
В предпочтительном осуществлении изобретения для получения фармацевтической композиции для подавления отторжения трансплантата in vivo непосредственно применяют TAIC по изобретению.
Такой фармацевтический препарат может содержать полученные на стадии d) способа по изобретению живые TAIC по изобретению, суспендированные в фармацевтически приемлемом носителе, предпочтительно в количестве приблизительно от 1×105 до 1×107 клеток/мл, а более предпочтительно - приблизительно 1Ч106 клеток/мл препарата.
В дополнительном предпочтительном осуществлении изобретения для получения фармацевтической композиции для подавления отторжения трансплантата in vivo непосредственно применяют клетки субпопуляции TAICGM-7 по изобретению.
Такой фармацевтический препарат может содержать связывающиеся с антителом GM-7 живые клетки TAICGM-7 по изобретению, суспендированные в фармацевтически приемлемом жидком носителе, предпочтительно в количестве приблизительно от 1×106 до 1×108 клеток/мл, а более предпочтительно - приблизительно 1×106 клеток/мл препарата.
В особенно предпочтительном осуществлении изобретения для получения фармацевтической композиции для подавления отторжения трансплантата in vivo непосредственно применяют клетки субпопуляции TAICCD3+/CD14+ по изобретению.
Такой фармацевтический препарат может содержать одновременно экспрессирующие антигены CD3 и CD14 живые клетки TAICCD3+/CD14+ по изобретению, предпочтительно в количестве приблизительно от 5×105 до 5×107 клеток/мл, а более предпочтительно - приблизительно 5×106 клеток/мл препарата.
Описанные выше фармацевтические препараты могут содержать клетки по изобретению, суспендированные в хорошо переносимой физиологической среде. Подходящими средами являются, например, раствор Рингера, физиологический раствор или раствор человеческого альбумина в концентрации от 5 до 20% и т.п.
Клеточные препараты по изобретению могут содержать живые TAIC, полученные на стадии d) способа по изобретению. Альтернативно препараты могут содержать клетки, относящиеся к субпопуляциям связывающихся с антителом GM-7 клеток
TAICGM-7 или одновременно экспрессирующих на своей клеточной поверхности антигены CD3 и CD14 клеток TAICCD3+/CD14+. Препараты могут содержать соответствующие клетки в количестве предпочтительно, по меньшей мере, 1×105, более предпочтительно - по меньшей мере, 5×105, а наиболее предпочтительно - по меньшей мере, 1×106 клеток на мл, суспендированные в жидкой среде-носителе. Среда может представлять собой среду клеточной культуры или среду доставки, хорошо переносимую клетками, такую как раствор человеческого альбумина в концентрации от 5 до 20%. Альтернативно клетки в препарате можно подвергнуть глубокой заморозке и содержать в пригодной для хранения среде, такой как, например, RPMI с 50% раствором человеческого альбумина и 10% DMSO.
Наконец, изобретение также относится к способу, в котором индуцирующие воспринимаемость трансплантата клетки по изобретению (TAIC, TAICGM-7 или
TAICCD3+/CD14+) применяют для получения или роста регуляторных T-лимфоцитов in vitro. Как показано в примере 12, прямое совместное культивирование TAIC с лимфоцитами in vitro приводит к значительной пролиферации регуляторных T-лимфоцитов, особенно CD4+/CD25+-лимфоцитов, см. Wood и Sakaguchi: "Regulatory Cells in Transplantation Tolerance", Nature Review Immunology 3, 199-210 (2003). Следовательно, прямым совместным культивированием TAIC с лимфоцитами, как описано в примере 12, возможно получать и/или выращивать регуляторные T-лимфоциты, особенно CD4+/CD25+-лимфоциты.
Прямое культивирование in vitro по настоящему изобретению означает, что TAIC и лимфоциты культивируют совместно в непосредственном физическом контакте в одной среде, как среди прочего, показано в примере 12.
В данном способе среда предпочтительно содержит соответствующие клетки, т.е. TAIC и лимфоциты приблизительно в равных количествах клеток и каждые в количестве предпочтительно, по меньшей мере, 1×105, более предпочтительно - по меньшей мере, 5×105, а наиболее предпочтительно - по меньшей мере, 1×106 клеток на мл, растворенные в жидкой среде-носителе; среда может представлять собой среду клеточной культуры или среду доставки, хорошо переносимую клетками, такую как раствор человеческого альбумина в концентрации от 5 до 20%. Совместное культивирование предпочтительно необходимо проводить в физиологических условиях приблизительно при 37°C, например, в инкубаторе, в течение предпочтительно от 3 до 5, а более предпочтительно - 4 суток.
Как показано в примере 13 индуцирующие воспринимаемость трансплантата клетки можно индуцировать не только посредством введения реципиенту TAIC, полученных из моноцитов донора, но также посредством обратного введения реципиенту лимфоцитов реципиента, ранее непосредственно совместно культивируемых in vitro с полученными от донора, как описано выше TAIC.
Следовательно, по дополнительному осуществлению изобретения регуляторные T-лимфоциты можно получить in vitro из лимфоцитов, происходящих от реципиента трансплантата, посредством прямого совместного культивирования лимфоцитов реципиента с TAIC, происходящими от донора данного трансплантата. Обратное введение совместно культивируемых лимфоцитов реципиенту приведет к воспринимаемости трансплантата реципиентом, как показано в примере 13.
Полученные таким образом происходящие от реципиента регуляторные T-лимфоциты можно выделить посредством FACS, как описано здесь (см. выше), и применять в фармацевтическом препарате, в котором клетки суспендированы в фармацевтически приемлемом носителе, как описано выше, для предотвращения отторжения трансплантата у реципиента.
Изобретение более подробно иллюстрируется посредством примеров.
Если в примере не определено, состав среды и применяемые вещества представляют собой следующее:
1. Раствор пенициллина/стрептомицина:
10000 единиц пенициллина в виде натриевой соли пенициллина G и 1000 мкг стрептомицина в виде сульфата стрептомицина на мл физиологического раствора хлорида натрия (NaCl 0,85%) (Каталог Gibco № 15140122).
2. Трипсин-ЭДТА
0,5 г трипсина и 0,2 г ЭДТА (4 Na)/л
3. Среда RPMI 1640 (1×, жидкая (11875)), содержащая L-глутамин
Среды RPMI (Roswell Park Memorial Institute) 1640 обогащены композициями, которые можно широко использовать для клеток млекопитающих.
Ссылка: Moore G. E., et al., J. A. M. A. 199: 519 (1967)
4. PBS (фосфатно-солевой буфер по Дульбекко) ср. J. Exp. Med. 98: 167 (1954):
5. Ficoll-Hypaque (Фиколл-гипак):
Среда для разделения лимфоцитов (сополимеризат сахарозы/эпихлоргидрина Mg 400000; Плотность 1,077, доведенный диатризоатом натрия).
6. L-глутамин
Жидкий: 29,2 мг/мл.
7. Макрофагальный колониестимулирующий фактор (M-CSF)
Рекомбинантный человеческий M-CSF из E. coli; содержит в качестве мономера (18,5 кДа) 135 аминокислотных остатков, включающих в себя N-концевой метионин; находится в виде гомодимера с молярной массой 37 кДа; (Каталог SIGMA № M 6518).
8. Интерферон γ (γ-IFN)
Рекомбинантный человеческий γ-IFN из E. coli; белок 16,7 кДа, содержащий 143 аминокислотных остатка (Каталог CHEMICON № IF002).
Пример 1
Выделение моноцитов из цельной крови
Во избежание коагуляции крови и для питания клеток 450 мл цельной человеческой крови смешивали в наборе из трехкамерных пакетов с 63 мл стабилизирующего раствора, содержащего на литр H2O, 3,27 г лимонной кислоты, 26,3 г трехзамещенного цитрата натрия, 25,5 г декстрозы и 22,22 г дигидроксифосфата натрия. Величина pH раствора составляла 5,6-5,8.
Для выделения компонентов крови затем в течение 7 минут проводили "сильное центрифугирование" данной смеси при 4000 об/мин при 20°C. Это приводило к расслоению корпускулярного и некорпускулярного компонентов на три слоя. С применением набора пакетов в предоставленном для данной цели устройстве для сдавливания эритроциты продавливали в нижний пакет, плазму продавливали в верхний пакет, а так называемая лейкоцитарная пленка оставалась в среднем пакете и составляла объем приблизительно 50 мл.
Количество 50 мл вновь полученной лейкоцитарной пленки затем разделяли на 2 части по 25 мл каждая и наслаивали на 25 мл предварительно помещенной в две 50 мл пробирки Falcon со средой для разделения Ficoll-Hypaque, соответственно.
Данный препарат 30 минут центрифугировали при 2500 об/мин без торможения. После этого эритроциты и мертвые клетки, все еще присутствующие в лейкоцитарной пленке находились ниже фазы фиколла, тогда как белые клетки крови, включающие в себя моноциты, отделялись на Ficoll в виде интерфазы белого цвета.
Затем интерфазу белого цвета, включающую в себя моноциты, осторожно забирали пипеткой и смешивали с 10 мл фосфатно-солевого буфера (PBS).
Затем данный препарат три раза центрифугировали в течение 10 минут при 1800 об/мин. с торможением, супернатант пипеткой удаляли после каждой стадии центрифугирования и вносили свежий PBS.
Клеточные осадки, образовывающиеся на дне сосуда для центрифугирования (пробирки Falcon), содержали фракцию мононуклеарных клеток, т.е. моноцитов.
Для получения моноцитов, необходимых для экспериментов на крысах, 25 мл цельной крови забирали у четырех генетически идентичных крыс-доноров, соответственно, и каждые из них наслаивали на 25 мл среды для разделения Ficoll-Hypaque. Затем процедуру продолжали таким же способом, как описано выше. Альтернативно, аналогичные стадии разделения на Ficoll можно также применять для суспензии клеток селезенки.
Пример 2
Размножение и модификация моноцитов
Культивирование и размножение моноцитов проводили в питательной среде со следующими компонентами:
Питательная среда содержала 2,5 мкг/500 мл M-CSF.
Выделенные в примере 1 моноциты суспендировали в общем количестве 106 клеток в 10 мл питательной среды и переносили на чашку Петри (диаметром 100 мм). Чашку Петри предварительно заполняли чистой инактивированной FCS, а через 24 часа FCS сливали для получения, таким образом, покрытой FCS чашки.
Чашку Петри покрывали соответствующей крышкой и оставляли в течение 3 суток в инкубаторе при 37°C. Клетки оседали на дно чашки Петри после 24 часов. На вторые сутки супернатант удаляли пипеткой, а чашку Петри снова заполняли 10 мл свежей питательной среды.
На 4 сутки добавляли 50 нг интерферона γ в 10 мл питательной среды, а чашки снова закрывали и оставляли в течение дополнительных 48 часов в инкубаторе при 37°C.
Затем в чашку Петри пипеткой добавляли 10 мл раствора трипсина, разведенного в соотношении 1:10 PBS. Закрытую чашку Петри оставляли на 10 минут в инкубаторе при 37°C.
Затем клеточным скребком проводили отделение клеток, прикрепленных к дну чашки Петри таким образом, что большая часть (>90%) клеток плавала в супернатанте.
Весь супернатант (10 мл раствора трипсина + 10 мл среды) забирали пипеткой, помещали в 50 мл пробирку Falcon и центрифугировали 10 минут при 1800 об/мин. Затем супернатант удаляли и добавляли свежую питательную среду для осаждения (оставшегося клеточного осадка), на 106 клеток добавляли 1 мл питательной среды. Определение количества клеток для определения точной дозы проводили известными способами, ср. Hay R.J., "Cell Quantification and Characterisation", в Methods of Tissue Engineering, Academic Press 2002, Chapter 4, S. 55-84.
Данную клеточную суспензию центрифугировали (1800 об/мин, 10 минут, см. выше), а клеточный осадок помещали или в PBS или, для применения у людей, в NaCl (физиол.). Затем проводили внутривенное введение сразу или в пределах 48 часов.
Альтернативно, после центрифугирования и удаления супернатанта, содержащего трипсин, к клеткам в качестве среды для замораживания добавляли FCS/DMSO и клетки в объеме 10 мл подвергали глубокой заморозке.
Среда для замораживания содержала 95% FCS и 5% DMSO. В каждом случае добавляли приблизительно 106 клеток в 1 мл среды и замораживали при следующих стадиях:
30 минут на льду;
2 часа при -20°C в предварительно охлажденной камере Styropor;
24 часа при -80°C в Styropor;
Хранение в малых пробирках в жидком азоте (N2) при -180°C.
На Фиг.1 показаны фенотипические изменения в экспрессии антигенов на моноцитах, применяемых после культивирования и стимуляции γ-IFN исходных моноцитарных клеток, определенные посредством проточной цитометрии.
Пример 3
Применение индуцирующих воспринимаемость трансплантата клеток (TAIC) для предварительной подготовки иммунной системы реципиента трансплантата
Применение TAIC для обработки потенциального реципиента перед трансплантацией предлагают, например, в клиническом случае донорства от живого индивида, где перед трансплантацией органа ясно, кто будет донором, а кто получит орган донора.
Для данного случая проводили гетеротопическую трансплантацию сердца в крысиной модели на взрослых инбредных крысах сочетания линий LEW [RT1.l] (применительно к настоящему изобретению обозначаемой как "LEW") → DA [RTl.avl] (применительно к настоящему изобретению обозначаемой как "DA") с применением технологии, описанной Ono и Lindsey [Ссылка Ono, K. and Lindsey, E.S. "Improved technique of heart transplantation in rats". J. Thorac. Cardiovasc. Surg. 57, 225-229 (1969)]. За 7 суток перед трансплантацией реципиентные крысы DA внутривенно в 1 мл PBS получали 106 TAIC, происходящих из моноцитов LEW. Затем через семь суток проводили трансплантацию взятого у линии крыс LEW сердца, которое гетеротопически имплантировали в брюшко (брюшную полость) реципиентного животного DA. Так как применяемые линии мышей являлись инбредными линиями, TAIC по изобретению экспрессировали идентичные трансплантированному сердцу LEW тканевые антигены.
С целями контроля проводили так называемую трансплантацию от третьей линии. С данной целью TAIC (106), происходящие из того же донорного животного LEW, снова внутривенно инъецировали животным DA на сутки -7 (т.е. за 7 суток перед хирургическим вмешательством). Через 7 суток проводили трансплантацию сердца, полученного от инбредной линии крыс CAP [RT1.c] (применительно к настоящему изобретению обозначаемой как "CAP"). Крысы CAP экспрессируют гаплотип RT1.c, следовательно, они полностью не согласованы по MHC (различаются по MHC) с донорными животными LEW. Ниже подробно представлены отдельные тестовые группы, в которых проводили гетеротопическую трансплантацию сердца (n=10):
1. LEW → DA; без обработки;
2. LEW → DA; 106 происходящих от LEW TAIC в/в, сутки -7 перед трансплантацией сердца LEW;
3. CAP → DA; 106 происходящих от LEW TAIC в/в, сутки -7 перед трансплантацией сердца САР;
4. CAP → DA; без обработки;
5. LEW → LEW, без обработки;
6. LEW → LEW, 106 происходящих от LEW TAIC в/в, сутки -7 перед трансплантацией сердца LEW.
Как можно видеть из кривых выживания Каплан-Мейер для сердца (Фиг.2A), 9 из 10 сердец LEW длительно время (>150 суток) воспринимались реципиентными животными DA, тогда как сердца третьей линии CAP остро отторгались после 6,7±0,8 суток. Когда не проводили предварительного введения TAIC, сердца LEW также остро отторгались после 7,2±1,0 суток. Отторжение сердец CAP посредством иммунного ответа, вызванного у животного DA после 6,7±0,8 суток, происходил в пределах того же временного интервала, что и в необработанной контрольной группе, в которой сердца CAP крыс DA отторгались после 6,9±1,0 суток, при сравнении группы 3 и группы 4, соответственно. Все сердца, трансплантированные сингенно по схеме LEW → LEW, выживали длительное время (>150 суток) вне зависимости от того проводили ли предварительную обработку TAIC или нет. Из данного можно сделать заключение, что TAIC не подвергали организм реципиента риску в смысле заболевания "трансплантат против хозяина". Кроме того, из данных результатов можно сделать заключение, что в/в. введение 106 TAIC за 1 неделю перед трансплантацией делает возможным индуцировать долговременную воспринимаемость органов. Индуцированная толерантность зависит от донора, так как соответствующая предварительная обработка крыс DA происходящими от LEW TAIC не вызывала толерантности для трансплантатов сердца третьей линии CAP. При сингенной схеме LEW → LEW дополнительное введение TAIC не влияло ни на выживание реципиентных животных, ни на выживание трансплантата.
На Фиг.2B показана гистологическая оценка аллогенных трансплантатов LEW на 150 сутки после операции (POD), после гетеротопической трансплантации в брюшную полость реципиентной крысы DA. Реципиентную крысу за 7 суток перед трансплантацией предварительно обрабатывали 106 происходящих из LEW TAIC. Трансплантат удовлетворительно функционировал (сильный пульс) до времени его удаления из реципиентной крысы (POD 150). На гистологическом препарате (×40) выявляли морфологию здоровой сердечной мышцы, нормальную сосудистую эндотелиальную ткань и только минимальную инфильтрацию мононуклеарными клетками без признаков острого или хронического процессов отторжения.
Примечательным в связи с этим является также тот факт, что происходящие от донора клетки (выявляемые иммуногистохимической детекцией с применением специфичных к MHC класса I LEW антител I1.69) можно длительное время в течение года выявлять в тимусе реципиентных крыс DA. На Фиг.2C представлен гистологический препарат тимуса крысы DA, предварительно обработанной 106 происходящих из LEW TAIC, полученный на POD 150. Препарат метили моноклональным антителом I1.69, специфичным к MHC класса I LEW. Как показано на фигуре, введение TAIC приводит к колонизации вилочковой железы, где происходит представление происходящих от донора антигенов MHC зрелым T-клеткам; можно легко распознать небольшие гнезда клеток, которые можно метить антителом.
Пример 4
Выявление происходящих от донора модифицированных моноцитов у реципиента
Индукцию смешанного химеризма внутривенной инъекцией происходящими от LEW TAIC у соответствующего животного DA можно рассматривать как возможную причину толерантности, индуцируемой по изобретению. Соответственно, для проверки того, можно ли с применением проточной цитометрии после инъекции происходящими от LEW TAIC длительное время (>45 суток) выявлять в крови реципиентного животного более чем 5% клеток провели тест. Проточную цитометрию проводили, как описано в существующем состоянии технологии [Ссылка Preffer F.I., "Flow cytometry" In: Diagnostic Immunopathology, Colvin RB, Bhan AK, McCluskey, RT (eds.), Raven Press New York, pp. 725-449 (1994)]. Выявление донорных клеток LEW в крови реципиента DA проводили с применением моноклонального антитела I1.69, специфически выявляющего антигены MHC класса I LEW. [см. Fandrich F., et al. Nature Med. 8, 171-178, (2002)]. Данные о химеризме, определенные для 4 из описанных в примере 3 трансплантаций LEW → DA после введения в периферическую кровь реципиентных животных 106 или 104 TAIC/кг × BW, соответственно, представлены на Фиг.2D.
После инъекции происходящих от LEW TAIC четырем реципиентным крысам DA не подвергнутым иммуносупрессии (106 клеток/кг × BW крысам 1-3 и 104 клеток/кг × BW крысе 4), у крыс 1-3 в крови реципиента DA выявили долговременный (60-80 суток) химеризм, который можно приписать происходящим от донора клеткам. Выявление происходящих от донора клеток проводили с применением определения посредством проточной цитометрии с применением специфичного к MHC класса I LEW антитела I1.69 (моноклональное антитело, связывающееся только с несущими MHC класса I клетками крыс линии LEW и не обнаруживающее перекрестной реактивности с клетками крыс линий DA). Так как почти все клетки периферической крови на клеточной поверхности экспрессируют антиген MHC класса I, с применением I1.69 можно очень точно определить долю донорных клеток в периферической крови крысы DA. Как показано на диаграммах на Фиг.20, у 3 и 4 животных более чем 10% клеток экспрессировали метящийся I1.69 антиген донора в течение первых 6 недель после внутривенной инъекции TAIC. Через 60 дней после инъекции, данная доля химеризма в периферической крови данных крыс DA значительно уменьшается и полностью пропадает на сутки 100. Однако данный преходящий химеризм тесно коррелирован с выживанием трансплантатов сердец LEW, трансплантированных после двойного времени (т.е. через 7 суток после инъекции TAIC), который больше не отторгается, несмотря на отсутствие донорского химеризма (после 100 суток) (крысы 1-3 в примере). С другой стороны индукция кратковременного химеризма с последующим введением 104 клеток/кг × BW (крыса 4; <20 суток) не гарантировала долговременной воспринимаемости трансплантата в виде пересаженного сердца.
Пример 5
Введение происходящих от донора клеток после трансплантации
Затем проводили оценку для клинического случая донорства от мертвого (трупа) для определения того, до какой степени послеоперативное введение TAIC способно индуцировать соответствующую долговременную воспринимаемость трансплантированного органа у реципиента. Для данной цели проводили гетеротопические трансплантации сердца в модели инбредных линий LEW → DA. Реципиентные крысы DA получали 4 цикла циклоспорина A (CSA) внутривенно в дозах в размере 5 мг CSA/кг × BW на сутки 0, 1, 2 и 3 после трансплантации. На сутки 7 производили введение 106 происходящих от LEW TAIC внутривенно в хвостовую вену реципиентных крыс DA. В качестве контроля служили следующие экспериментальные группы (n=6-10).
1. LEW → DA; без обработки
2. LEW → DA; CSA, 5 мг/кг × BW, в/в, сутки 0, 1, 2 и 3
3. LEW → DA; CSA, 5 мг/кг × BW, в/в, сутки 0, 1, 2 и 3 и 106 происходящих от LEW TAIC, в/в, сутки 7
4. LEW → DA; 106 происходящих от LEW TAIC, в/в, сутки 7
5. CAP → DA; CSA, 5 мг/кг × BW, в/в, сутки 0, 1, 2 и 3 и 106 происходящих от LEW TAIC, в/в, сутки 7.
Послеоперативная внутривенная обработка реципиентных крыс DA 106 происходящих от LEW TAIC на POD (сутки после операции) 7 приводила к долговременной толерантности, если в течение 0-3 суток проводили начальное внутривенное введение 4 циклов CSA (5 мг/кг × BW, в/в). Начальное введение CSA является необходимым, так как самостоятельная обработка единичным введением 106 TAIC на POD 7 (сутки после операции) предотвращала острое отторжение только в 1 случае из 6. Выбранная в данном случае доза CSA оказывает только субтерапевтический эффект, так как одним введением CSA долговременную воспринимаемость индуцировать невозможно, даже если выживание трансплантата увеличено. Индуцированная воспринимаемость трансплантата является зависимой от донора, так как соответствующая обработка крыс DA происходящими от LEW TAIC не вызывала толерантности для трансплантатов сердца третьей линии CAP.
Как показано на Фиг.3, сердца экспериментальной группы 1 остро отторгались через 7,0±0,8 суток. Сердца группы 2 отторгались с задержкой через 14,4±7,0 суток. Сердца группы 3 воспринимались в течение более чем 150 суток в 5 случаях из 6. В крысиной системе воспринимаемость органов в течение более чем 100 суток рассматривают как долговременную воспринимаемость, так как средняя продолжительность жизни инбредной крысы составляет приблизительно только 2 года. Самостоятельное послеоперативное введение TAIC (106 клеток) являлось способным предотвратить острое отторжение только в 1 случае из 6, так как сердца переставали сокращаться через 22,6±31,6 суток (группа 4). Контроль третьей линии с сердцами CAP показал специфичность индуцированной воспринимаемости трансплантата, так как сердца третьей линии снова остро отторгались через 7,4±2,2 суток. Данные результаты также доказывают то, что дополнительное применение TAIC не может вызывать какой-либо генерализованной иммуносупрессии, которая не предотвращала острого отторжения сердец CAP.
Пример 6
Исследование в отношении органной специфичности индуцированной TAIC долговременной воспринимаемости
Для исключения возможности того, что TAIC способны обеспечивать защиту от направленного реципиентом против антигена донора аллогенетически специфического иммунного ответа только для трансплантатов сердца, в указанных ниже экспериментальных группах (n=6) в сильно отторгающей крысиной инбредной системе DA → LEW трансплантировали следующие органы:
1. DA (почка) → LEW; без обработки
2. DA (почка) → LEW; 106 происходящих от DA TAIC, в/в, сутки -7 и -1 перед трансплантацией
3. LEW (почка) → LEW; без обработки
4. DA (печень) → LEW; без обработки
5. DA (печень) → LEW; 106 происходящих от DA TAIC, в/в, сутки -7 и -1 перед трансплантацией
6. LEW (печень) → LEW; без обработки
7. DA (кожа) → LEW; без обработки
8. DA (кожа) → LEW; 106 происходящих от DA TAIC, в/в, сутки -7 и -1 перед трансплантацией
9. LEW (кожа) → LEW; без обработки.
Как показано с помощью графиков Каплан-Мейер, приведенных на Фиг.4A-C, 6 из 6 трансплантатов почек, 5 из 6 трансплантатов печени и 5 из 6 кожных трансплантатов реципиентными крысами LEW воспринимались долговременно (>150 суток). Все сингенные трансплантации органов служили в качестве контроля технического исполнения без осложнений и воспринимались донорным животным без ограничений (не показано на кривых выживания). Следовательно, данные тестовые схемы также демонстрируют то, что внутривенная предварительная инъекция 2×106 TAIC индуцирует долговременную воспринимаемость органа для более чем 80% трансплантируемых органов.
Затем перечисленные выше трансплантаты почек, печени и кожи, трансплантированные с предварительным введением TAIC и без него, проверяли гистологически. На Фиг.4D показано, что как результат внутривенной инъекции 106 TAIC на кг массы тела, вводимой дважды на сутки -7 и -1 до трансплантации можно показать, что морфология трансплантированной почки осталась неповрежденной (в сравнении с контрольной почкой и сингенным трансплантатом LEW → LEW), тогда как в необработанной аллогенной контрольной группе DA → LEW можно увидеть явную инфильтрацию трансплантата в интерстициальном пространстве (на сутки 14 после операции). Аллогенный трансплантат печени (Фиг.4E), полностью отторгнутый на POD 12 без предварительной обработки TAIC (DA → LEW без TAIC), вел себя аналогичным образом, тогда как предварительное введение TAIC позволяло паренхиме трансплантатов печени выглядеть как полностью гомогенной и интактной по сравнению с сингенными трансплантатами. Аллогенный кожный трансплантат (DA) также приживлялся без осложнений (при сравнении с сингенным трансплантатом LEW), тогда как трансплантированный совместно трансплантат третьей линии CAP остро отторгался уже на 12 сутки после операции. Данный эксперимент доказывает специфичность индуцированной TAIC толерантности, так как предварительная обработка реципиентного животного LEW происходящими от DA TAIC на сутки -7 и -1 приводит к воспринимаемости трансплантатов DA, тогда как трансплантированный одновременно на сутки 0 кожный трансплантат CAP остро отторгается (Фиг.4F).
Пример 7A
Исследования в отношении специфичности к гаплотипу индуцированной TAIC долговременной воспринимаемости органов
Для исключения возможности того, что TAIC индуцируют долговременную воспринимаемость органов только в выбранном сочетании инбредных линий крыс LEW → DA, в модели гетеротопической трансплантации сердца тестировали указанные ниже дополнительные сочетания инбредных линий (n=6). Для данной цели наряду с другими линиями также применяли инбредную линию BN[RT1.n] (применительно к данному изобретению обозначаемую как "BN").
1. DA → LEW; без обработки
2. DA → LEW; CSA, 5 мг/кг × BW, в/в, сутки 0, 1, 2, 3 и 106 происходящих от DA TAIC/кг × BW, в/в, сутки 7 и 10 после операции
3. CAP → LEW; без обработки
4. CAP → LEW; CSA, 5 мг/кг × BW, в/в, сутки 0, 1, 2, 3 и 106 происходящих от CAP TAIC/кг × BW, в/в, сутки 7 и 10
5. BN → LEW; без обработки
6. BN → LEW; CSA, 5 мг/кг × BW, в/в, сутки 0, 1, 2, 3 и 106 происходящих от BN TAIC/кг ×Ч BW, в/в, сутки 7 и 10.
Из предварительных экспериментов уже являлось известным, что с применением единственного послеоперационного в/в. введения 106 TAIC в так называемом "высокоотвечающем" сочетании (DA → LEW), ассоциированном с сильнейшим известным в крысиной инбредной модели ответом отторжением, нельзя достичь долговременной воспринимаемости. Поэтому предприняли попытку увеличить присутствие TAIC у реципиентного животного посредством введения их дважды. Как показано посредством соответствующих кривых выживания, приведенных на Фиг.5A, двукратное введение TAIC на сутки 7 и 10 после операции обеспечивало долговременную воспринимаемость в 4 из 6 случаях в группе 2 и в 5 из 6 случаях в экспериментальной группе 4. В сочетании линий BN → LEW все трансплантированные органы выживали длительное время без какого-либо признака острого отторжения. Во всех тех случаях, в которых трансплантированные сердца долговременно воспринимались (>150 суток) в течение первых 20-45 суток после введения TAIC, у реципиентных животных являлось возможным выявить смешанный химеризм. Однако ранняя органная недостаточность всегда сопровождалась отсутствием клеток донора в периферической крови реципиента. Из этих данных можно сделать заключение, что в/в введение TAIC приводит к долговременной воспринимаемости органов, только если клетки индуцируют смешанный донорный химеризм, который можно выявить, по меньшей мере, в течение 21 суток. Это также подтверждается экспериментальными результатами, показанными на Фиг.2D.
Следовательно, посредством послеоперационного введения 106 TAIC на кг массы тела, внутривенно вводимых реципиентным животным на сутки 7 и 10, являлось возможным индуцировать долговременную толерантность в 3 сочетаниях инбредных линий в более чем 80% случаев, если, дополнительно, вначале проводили 4 цикла CSA.
Пример 7B
Исследования в отношении значения стимуляции γ-IFN обработанных M-CSF моноцитов для индукции толерантности in vivo
Для того чтобы появилась возможность охарактеризовать действенность дополнительной стимуляции интерфероном γ обработанных M-CSF моноцитов крови на развитие толерантности in vivo, более подробно исследовали следующие тестовые группы в модели инбредных крыс LEW → DA для гетеротопической трансплантации сердца (n=6 животных на группу):
1. LEW → DA; 106/кг × BW происходящих от LEW моноцитов крови (не культивированных моноцитов), в/в, сутки -7 перед трансплантацией сердца
2. LEW → DA; 106/кг × BW происходящих от LEW моноцитов крови (обработанных M-CSF), в/в, сутки -7 перед трансплантацией сердца
3. LEW → DA; 106/кг × BW происходящих от LEW моноцитов крови (стимулированных γ-IFN), в/в, сутки -7 перед трансплантацией сердца
4. LEW → DA; 106/кг × BW происходящих от LEW моноцитов крови (обработанных M-CSF и стимулированных γ-IFN), в/в, сутки -7 перед трансплантацией сердца.
Как показано посредством кривых выживания Каплан-Мейер на Фиг.5B, внутривенное введение свежих некультивированных происходящих от LEW моноцитов крови, за 7 суток перед трансплантацией крови не может предотвратить острого отторжения аллогенного трансплантата сердец LEW у донорного животного DA. Трансплантаты данной экспериментальной группы отторгались в среднем (±SD) через 7,8±0,8 суток. Однако предварительное введение культивированных в течение 6 суток с M-CSF моноцитов приводило к значительному увеличению времени выживания органа, так как в данной экспериментальной группе отторжение наблюдали в среднем только через 35,0±51,5 суток. Стимуляция свежих моноцитов крови только γ-IFN в течение 48 часов в среде RPMI, не содержащей M-CSF, не могла предотвратить острого отторжения трансплантатов сердца, прекращавших сокращаться в среднем через 7,8±1,2 суток. Только обработка свежих моноцитов крови M-CSF в течение 6 суток с дополнительной стимуляцией данных клеток γ-IFN в течение 48 часов (на 5 и 6 сутки) приводила к долговременной толерантности (>150 суток) 4 из 6 трансплантатов. Результаты показывают решающее значение дополнительной стимуляции обработанных M-CSF моноцитов γ-IFN для образования толерантности in vivo.
Пример 8
Исследования в отношении видовой специфичности индуцируемой TAIC долговременной воспринимаемости органов
Для исключения возможности того, что TAIC эффективно индуцируют долговременную воспринимаемость только у выбранных видов животных, т.е. крыс, клетки моноцитарного происхождения выделяли от донорной свиньи сходным с подробно описанным в примере 2 способом, где стимуляцию γ-IFN проводили посредством добавления к питательной среде 1000 Ед/10 мл на 24 часа. Далее проводили исследование эффективности TAIC в отношении подавления острого отторжения в модели левосторонней трансплантации легкого на "карликовых свиньях с несоответствующим SLA", обладающих потенциальной возможностью к отторжению их органов по сравнению с человеком. В данном случае левое легкое донорной свиньи удаляли и ортотопически трансплантировали генетически отличной реципиентной свинье (т.е. в левую часть грудной полости после удаления левого легкого у реципиентной свиньи). В течение удаления легкого одновременно забирали 500 мл донорской крови, служившей для получения TAIC из моноцитов данной свиньи.
Все донорные животные обладали массой 30-35 кг. Среди свиней образовывали следующие экспериментальные группы (n=4):
1. CSA, 5 г/сутки, 50 мг азатиоприна/сутки, 25 мг метилпреднизолона/сутки, в течение 28 суток
2. Как в группе 1, и 108 клеток костного мозга (BM) донорной свиньи/кг × BW реципиентной свиньи при 4,5 Гр тотального облучения тела (WBI) реципиентной свиньи
3. Как в группе 1, и 106 происходящих от донорной свиньи TAIC/кг × BW на 7 и 10 сутки после операции после трансплантации легкого.
Как показано на Фиг.6A, отторжение трансплантатов легкого в группе 1 происходило в среднем через 50,3±9,9 суток после прекращения иммуносупрессии. С другой стороны животные в группе 2 отторгали их органы через 102,3±81,0 суток. Тройная иммуносупрессия в сочетании с обработкой TAIC, повторенной дважды, продлевала выживание органов до 292±135,5 суток, 3 из 4 животных продемонстрировали долговременную воспринимаемость органов (>350 суток). Во всех случаях группы 3 в крови реципиента являлось возможным наблюдать смешанный донорный химеризм в течение более чем 21 суток, тогда как у животных из групп 1 и 2 выявляли только кратковременный смешанный химеризм в течение 6-8 суток. На Фиг.6B представлены легочные трансплантаты свиньи из экспериментальной группы 1 (POD 41) и обработанной TAIC свиньи, из экспериментальной группы 3 (55 суток после трансплантации; B). Хотя в проекции левой грудной стенки, вокруг силуэта сердца, можно наблюдать уменьшение прозрачности (затемнение), как показатель отторгнутого трансплантата легкого у свиньи из экспериментальной группы 1, на рентгенограмме обработанного TAIC животного представлено нерентгеноконтрастные легкое и место расположения трансплантата с хорошим контурированием по отношению к силуэту сердца.
Пример 9
Связывание антитела GM-7 с TAIC
Моноклональное антитело GM-7 получали иммунизацией мыши человеческими TAIC, полученными как описано в примере 2. Клетки гибридомы, продуцирующие антитела депонировали в "Deutsche Sammlung fur Mikroorganismen" под инвентарным № DSM ACC2542. Представленные ниже результаты демонстрируют, что антитело специфически связывается с антигеном, экспрессируемым только на CD14+-клетках, подвергнутых по изобретению модификации M-CSF в течение 6 суток и стимуляции γ-IFN в течение 2 суток ex situ.
На Фиг.1 представлена определенная проточной цитометрией связывающая способность GM-7 в отношении моноцитарных клеток после модификации in vitro, т.е. после трансформации в TAIC. Можно видеть, что несущие CD14 моноциты, полученные непосредственно из лейкоцитарной пленки, не связываются антителом GM-7 (заштрихованная серым область соответствует антителу контроля). Напротив, часть моноцитов после культивирования с M-CSF и стимуляции γ-IFN, экспрессируют антиген, распознающийся моноклональным антителом GM-7. После культивирования как описано в примере 2, приблизительно 80% трансформированных моноцитов являются способными связывать моноклональное антитело GM-7.
В дополнительном эксперименте в смешанной культуре лимфоцитов (MLC) сравнивали супрессорную активность CD14+/GM-7+-клеток с супрессорной активностью CD14+/GM-7--клеток. MLC получали как описано у Kurnick, J.T. "Cellular Assays" in: Diagnostic Immunopathology, [Colvin R.B., Bhan A.K., McCluskey, R.U. (ed.), Raven Press, New York, pages 751-771 (1994)].
В данном примере TAIC происходили от индивида B. Как показано на Фиг.7, между супрессорной активностью положительных по GM-7 и отрицательных по GM-7 TAIC существует значимое отличие. Только положительная по GM-7 часть популяции TAIC, полученных через 6 суток обработки посредством M-CSF и 2 суток стимуляции посредством γ-IFN, оказывает значимое ингибирующее действие на T-клеточную пролиферативную активность отвечающих клеток индивида A после стимуляции клетками индивида B.
Эти данные означают, что для клинических целей, из популяции TAIC, полученной после культивирования исходных моноцитов с M-CSF и стимуляции γ-IFN, вследствие доступности моноклонального антитела GM-7 можно выделить клеточную популяцию с оптимизированной супрессорной функцией.
Пример 10
Определение влияния ингибитора IDO 1-метилтриптофана на иммуносупрессорную активность TAIC
Для выяснения того препятствует ли ингибитор фермента индоламин-2,3-диоксигеназы (IDO) 1-метилтриптофана (1-MT) супрессорной функции TAIC, образуемых в системах "Mo" и "Mo+Ly" (см. пример 11), исследовали ряд смешанных культур лимфоцитов со стимулированными PHA (фитогемагглютинином) T-клетками в присутствии и отсутствии 1-MT.
В данных смешанных культурах лимфоцитов 50000 лимфоцитов с 2 мкг PHA переносили в лунки 96-луночного планшета с последующим периодом пролиферации в течение 144 часов (обозначенном как "PhaLy"). Лимфоциты, только культивированные в среде без добавления PHA, служили в качестве дополнительного контроля (обозначено как "Ly"). Для определения пролиферации стимулированных PHA лимфоцитов в различных совместных культурах создавали и анализировали следующие 4 системы:
[105 клеток из системы "Mo+Ly" по примеру 11]
[105 клеток из системы "Mo" по примеру 11]
После 144 часов культивирования все контроли и совместные культуры дополнительно культивировали 24 часа в присутствии 3[H]-тимидина ("импульсно") а потом определяли количество радиоактивно меченного включившегося тимидина, как количество импульсов в минуту (имп./мин), см. Фиг.8. На данной схеме количество определенной радиоактивности представляет собой меру количества меченного тимидина включившегося в ДНК и, следовательно, меру скорости пролиферации лимфоцитов. Представленные на Фиг.8 значения соответствуют средним значениям для тройных определений в 3 экспериментах, с указанием стандартного отклонения для каждого.
Результаты показывали, что лимфоциты, не стимулированные PHA ("Ly") значительно не пролиферировали, средняя наблюдаемая радиоактивность составляла 367 имп./мин. (см. Фиг.8). Напротив, стимуляция 2 мкг PHA приводила к значительному увеличению скорости пролиферации лимфоцитов ("PhaLy"), наивысшее измеренное включение при среднем значении 18459 имп./мин. в данных образцах.
Добавление TAIC к культурам лимфоцитов отчетливо уменьшало скорость пролиферации: сильно, если добавляли клетки из системы примера 11 "Mo+Ly" (PhaLy +"Mo+Ly", определенное значение составляло 1498 имп./мин), и менее сильно, когда добавляли клетки из системы "Mo" примера 11 (PhaLy + "Mo", измеренное значение составляло 3369 имп./мин).
Полученные после добавления к системам, содержащим "Mo+Ly" и "Mo" со стимулированными лимфоцитами, по 2 мкмоль 1-метилтриптофана (1-MT) результаты продемонстрировали, что 1-метилтриптофан (1-MT) синергично увеличивает супрессорную функцию TAIC, в соответствии с чем уменьшение являлось более сильным с TAIC из системы "Mo+Ly" (измеренное значение составляло 267 имп./мин), чем с TAIC из системы "Mo" (измеренное значение составляло 390 имп./мин).
Пример 11
Влияние лимфоцитов на формирование CD3+/CD14+-клеток в течение культивирования моноцитов
Влияние лимфоцитов, присутствующих в моноцитарной фракции на образование CD14+/CD3+-клеток, действующих как TAIC, определяли посредством сравнения двух различных систем.
Для первой системы (далее обозначаемой как "Mo") сначала, как описано в примере 1, из интерфазы лейкоцитарной пленки забирали моноцитарную фракцию. Затем, как описано в примере 2, клетки переносили на стадию культивирования с M-CSF. В пределах одного часа после начала культивирования, моноциты, прикрепившиеся ко дну флакона с тканевой культурой, пять раз промывали по 10 мл PBS каждый раз, и, таким образом, количество присутствующих в культуре лимфоцитов уменьшалось до <5% (4,8±2,4%), тогда как количество полученных таким образом обогащенных моноцитов (CD14+) составляло приблизительно 90% (92±5,6%). Дополнительные клеточные компоненты в системе представляли собой B-лимфоциты и гранулоциты
Клетки во второй системе (далее обозначаемой как "Mo+Ly") также забирали из интерфазы "лейкоцитарной пленки" как моноцитарную фракцию как описано в примере 1. Однако в отличие от системы "Mo" клетки, прикрепившиеся ко дну флакона с тканевой культурой, отмывали однократно после 24 часов от начала фазы культивирования, описанной в примере 2. Как результат получали клеточную популяцию, состоявшую из 45±5,3% CD14+-моноцитов и 23,5±8,9% CD2+-лимфоцитов. Как и в системе "Mo" лимфоциты и гранулоциты также присутствовали в данной системе.
Определение количеств соответствующих клеточных типов в целой клеточной популяции в системе проводили посредством проточной цитометрии в трех экспериментальных системах для каждой (см. Фиг.9). Результаты представлены как средние значения, включающие в себя стандартное отклонение.
В начале культивирования CD14+/CD3+-клетки не выявляли ни в системе "Mo" ни в системе "Mo+Ly"(d 0 = сутки 0). После периода культивирования в течение пяти суток, эксперимент завершали и клетки охарактеризовывали посредством анализа FACS после отделения клеток от дна флакона с тканевой культурой, как описано в примере 2. Обнаружено, что относительное количество CD14+-клеток уменьшалось в данных системах от 92% до 42% в системе "Mo" и от 45 % до 28% в системе "Mo+Ly". По-видимому, лимфоциты пролиферируют быстрее, чем моноциты; относительное количество лимфоцитов увеличивалось в системе "Mo" от 4,8% до 69,8%, а в системе "Mo+Ly" от 23,5% до 50,6%. В течение культивирования CD14+/CD3+-клетки, действующие как TAIC, формируются в обеих культурах. В данном контексте важно, что в системе "Mo+Ly" наблюдали значительно большее увеличение содержания CD14+/CD3+-клеток с количеством 32,0±5,3%, чем в системе "Mo" только с 7,2±3,2%.
Данные результаты демонстрируют, что очистка клеток в начале культивации для обогащения моноцитами до относительного количества более чем 90% оказывает отрицательное воздействие на образование иммуносупрессирующих CD14+/CD3+-клеток в популяции TAIC, тогда как описанный в примерах 1 и 2 способ приводит к значительно большему выходу CD14+/CD3+-клеток.
Пример 12
Совместное культивирование человеческих TAIC с аллогенными лимфоцитами in vitro для образования регуляторных T-клеток
Для определения того, индуцируют ли TAIC формирование у реципиента регуляторных T-клеток, т.е. CD4+/CD25+-лимфоцитов, создавали несколько различных культур TAIC с лимфоцитами и анализировали формирование получающихся из них регуляторных T-клеток.
С данной целью TAIC донора A прямо или опосредовано совместно культивировали in vitro вместе с лимфоцитами донора B. В прямой совместной культуре делали возможным прямой контакт клетка-клетка между TAIC (донора A) и лимфоцитами (донора B), тогда как в опосредованной совместной культуре мембрана ("вставка в клеточную культуру", с размером проб 0,4 мкм, Falcon, № заказа 353090) позволяла обмениваться средой, но препятствовала физическому контакту двух клеточных популяций. Прямую и опосредованную культуру обрабатывали предпочтительно от 3 до 5 суток, более предпочтительно 4 суток в условиях инкубатора, т.е. при 37°C и в атмосфере 5% CO2.
После культивирования в обеих системах, а также в контролях, где TAIC или лимфоциты, соответственно, культивировали отдельно, определяли соответствующие количества регуляторных T-клеток (CD4+/CD25+). Дополнительно в контрольной системе, где культивировали только TAIC, определяли количество CD3+/CD14+-клеток, представлявших собой компонент популяции клеток TAIC с наиболее значительным супрессорным действием в смешанной культуре лимфоцито.
Во всех системах посредством анализа FACS определяли поверхностные антигены клеток и анализировали количество соответствующих клеточных популяций в общем количестве клеток.
Кроме того, в соответствующей клеточной популяции посредством ПЦР определяли относительную экспрессию трех новых контрольных генов, специфически экспрессирующихся в регуляторных T-клетках (Foxp3, CTLA4 и интегрин αEβ7) (см. таблицу). Foxp3 представляет собой специфический транскрипционный фактор, рассматриваемый как ген для контроля развития регуляторных T-клеток и специфически экспрессируемый данными клетками [см Hori, S. et al.,"Control of Regulatory T-cell Development by the Transcription Factor Foxp3", Science 299, 1057-1061 (2003)].
CTLA4 представляет собой дополнительный фактор, применяемый в качестве маркера для определения регуляторной функции CD4+/CD25+-T-клеток (см. Khattri, R. et al., "An essential role for Scurfin in CD4+/CD25+ T regulatory cells", Nature Immunology, публикация онлайн, doi:10.1038/ni909 (2003); Shimizu, J. et al. "Stimulation of CD25+/CD4+ regulatory T cells through GITR breaks immunological self-tolerance", Nature Immunology, публикация онлайн, doi:10.1038/ni759 (2003); Cobbold, S. P. et al. "Regulatory T cells in the induction and maintenance of peripheral transplantation tolerance", Transpl. Int. 16 (2), 66-75 (2003)].
Интегрин αEβ7 представляет собой интегрин, который связывается с эпителиальным кадгерином и который можно применять в качестве маркера для большинства эффективных субпопуляций регуляторных CD25+-T-клеток [см. Lehmann, J. et al. "Expression of the integrin αEβ7 identifies unique subsets of CD25+ as well as CD25-regulatory T cells" PNAS 99 (20), 13031-13036 (2002)].
Определение экспрессии Foxp3, CTLA4 и интегрина αEβ7 проводили посредством количественной ПЦР с применением GAPDH и β-актина, двух "домашних генов", в качестве контроля; определенные значения, с одной стороны, находились в связи друг с другом, значения, полученные для CD14+/CD3--клеток обозначены как 1; с другой стороны, абсолютные количества РНК указывали в мкг на мкг общей РНК, для установления соотношения измеренной супрессии, осуществляемой TAIC in vitro и формированием двойных позитивных CD4+/CD25+-клеток со скоростью экспрессии соответствующих генов. Для количественной ПЦР применяли стандартные способы, известные специалистам (см. Lottspeich, F., Zorbas, H. "Bioanalytik", Spektrum Akademischer Verlag GmbH, Heidelberg Berlin, 1998).
Таблица демонстрирует, что в популяции лимфоцитов, полученных из прямой совместной культуры, доля двойных позитивных CD4+/CD25+-клеток многократно увеличивается до значения 8,7% в сравнении с количеством CD4+/CD25+-лимфоцитов, полученных из непрямой совместной культуры или контроля с 2,38% и 2,65%, соответственно.
Субпопуляция CD4+/CD25+-клеток экспрессирует наивысшие относительные количества мРНК всех тестируемых контрольных генов в сравнении с экспрессией у CD4+/CD25--клеток (см. таблицу). Хотя экспрессия Foxp3 увеличена приблизительно в 10 раз (37 против 3,75), экспрессия CTLA4 достигает даже большей максимальной экспрессии (4699 против 0,376). Увеличение экспрессии в CD4+/CD25+-клетках в течение периода совместного культивирования третьего контрольного гена - интегрина αEβ7 - является приблизительно такой же высокой, как и для CTLA4, так как абсолютное количество мРНК интегрина αEβ7 достигает в CD4+/CD25+-клетках
1,4×10-9 в сравнении с 3,4×10-12 мкг мРНК/мкг общей РНК в CD4+/CD25--клетках.
После опосредованной совместной культуры относительное количество мРНК в субпопуляции CD4+/CD25+ составляет только 10 и является таким же низким как относительное количество, экспрессируемое лимфоцитами в контроле, экспрессирующем относительное количество мРНК Foxp3 в размере 15.
Относительное количество мРНК CTLA4, экспрессируемое лимфоцитами в контроле, является таким низким как 0,375 в популяции CD4+/CD25+ и 0,1 в популяции
CD4+/CD25-.
CD3+
CD3-
CD25+
CD25-
CD25+
CD25-
CD25+
CD25-
10-10
10-9
10-8
10-9
10-9
10-10
10-10
10-10
10-10
10-6
10-10
10-9
10-12
Экспрессия CTLA4 в субпопуляции CD14+/CD3+ клеток TAIC являлась сходной с экспрессией Foxp3 и также являлась значительно большей, чем во всех других клеточных популяциях. В этом смысле, примечательно, что экспрессия CTLA4 (относительное значение 12500) в субпопуляции CD14+/CD3+ во много раз более сильная, чем экспрессия Foxp3, демонстрирующая относительное значение 50.
В субпопуляции TAIC CD14+/CD3- выявить экспрессию интегрина αEβ7 являлось невозможным, так как подобно поведению экспрессии других анализируемых генов, экспрессия интегрина в субпопуляции TAIC CD14+/CD3-, измеряемая как абсолютное значение в мкг РНК на мкг общей РНК, достигала максимального значения (3,4×10-9 мкг/мкг общей РНК).
Значительно увеличенная экспрессия трех маркеров лимфоцитов Foxp3, CTLA4 и интегрина αEβ7 у происходящих из моноцитов TAIC в сравнении с нормальными лимфоцитами является совершенно неожиданной, и до настоящего времени ее никогда не наблюдали у моноцитов или происходящих из моноцитов клеток.
В заключение, приведенные выше результаты данного примера демонстрируют следующее: Если отталкиваться от предположения, что уровень экспрессии Foxp3, CTLA4 и интегрина αEβ7 коррелирует с иммунорегуляторными свойствами соответствующих клеток, здесь показано, что супрессорная активность субпопуляции TAIC CD3+/CD14+ связана с высокой экспрессией данных трех генов. Это является даже более неожиданным, если принять во внимание, что данные маркеры до настоящего времени описаны только для лимфоцитов, но никогда для клеток моноцитарного происхождения.
Дополнительно показано, что прямое совместное культивирование TAIC с лимфоцитами приводит к значительно большим количествам CD4+/CD25+-лимфоцитов по сравнению с опосредованным совместным культивированием и культивированием только лимфоцитов. Это поддерживает предположение о том, что in vivo (т.е. у пациента) после введения TAIC также формируются регуляторные T-клетки. Данные результаты находятся в согласии с известной иммунорегулирующей функцией CD4+/CD25+-лимфоцитов и с тем фактом, что содержание в данных клетках мРНК Foxp3, CTLA4 и интегрина αEβ7 значительно выше, чем в лимфоцитах при опосредованном культивировании или чем в контрольных лимфоцитах.
Пример 13
Индукция воспринимаемости трансплантата in vivo культивируемыми in vitro с TAIC лимфоцитами
Представленные в примере 12 результаты и заключения подтверждены в экспериментах на животных in vivo. В отличие от описанной в примерах 3, 4, 5, 6 и 7 процедуры, в данном примере животным в выбранных сочетаниях инбредных линий LEW → DA не инъецировали полученные из LEW TAIC для индукции толерантности, а животным инъецировали лимфоциты DA от реципиента, которые в течение периода 5 суток предварительно прямым способом совместно культивировали с полученными от LEW TAIC от донора или которые культивировали в среде отдельно в качестве контроля.
У животных, получавших до аллогенной трансплантации сердца аутологичные совместно культивируемые лимфоциты DA, развилась специфичная толерантность, тогда как у контрольных животных, получавших не культивируемые совместно контрольные лимфоциты DA, во временном интервале от 10 до 14 суток развилось острое отторжение трансплантированного сердца.
Данные результаты указывают на то, что физическое взаимодействие клетка-клетка между TAIC донора и лимфоцитами реципиента индуцирует образование регуляторных T-клеток, которые сами по себе способны к модификации сингенной иммунной системы реципиентов таким образом, чтобы супрессировать потенциально аллореактивные T-клетки и, таким образом, предотвратить отторжение органа.
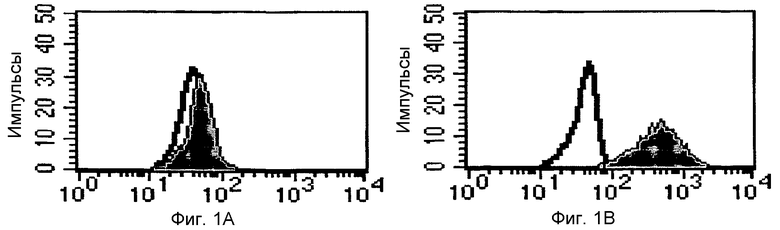
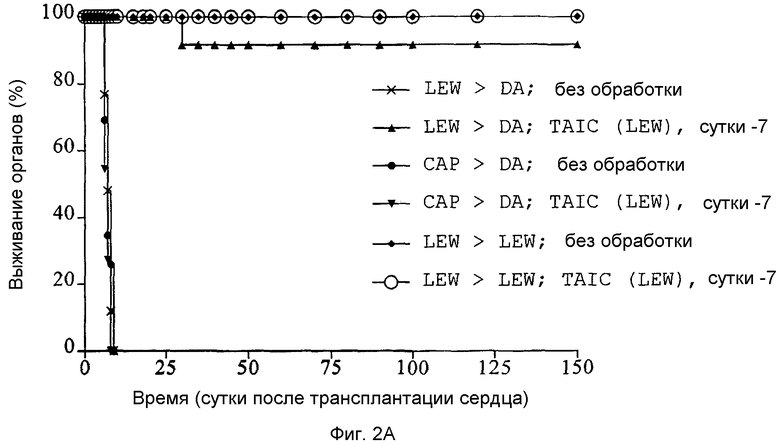
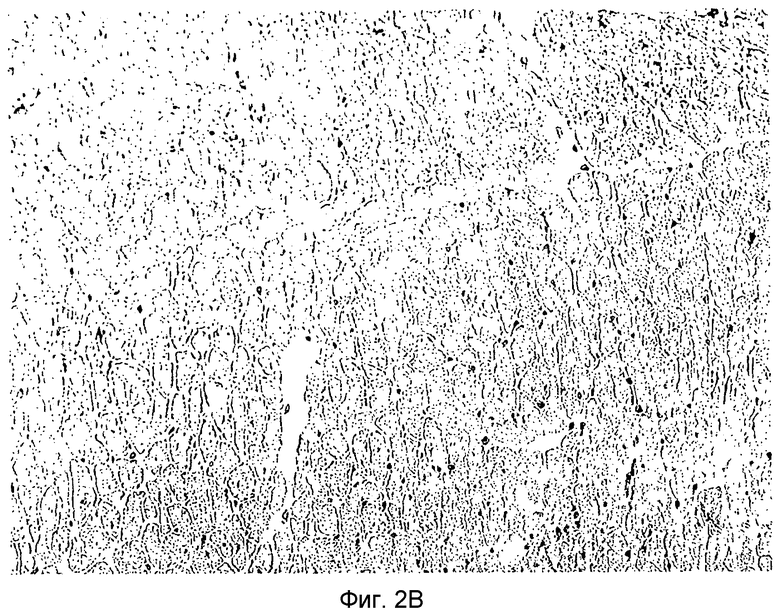
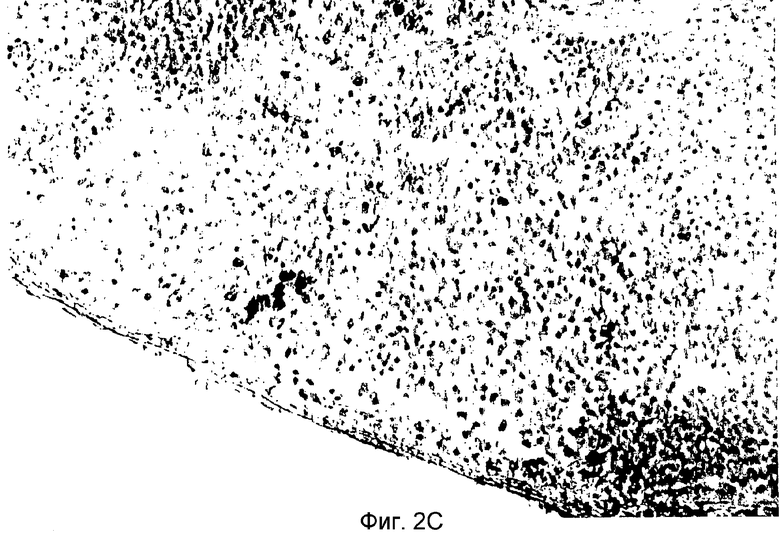
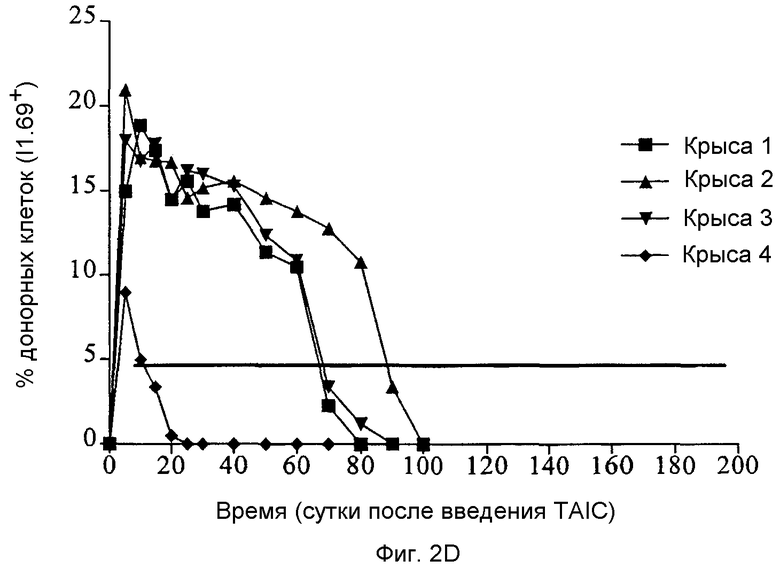
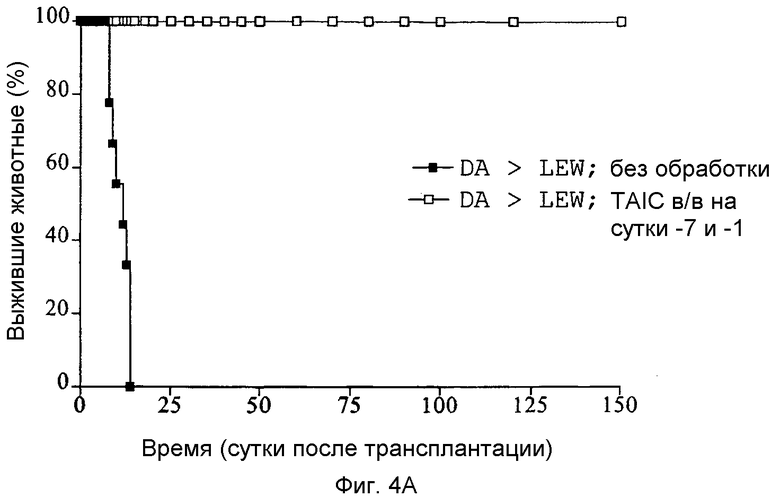
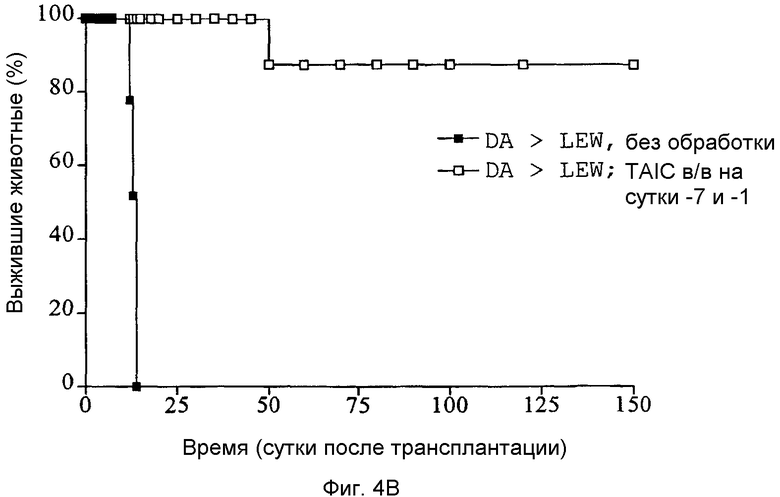
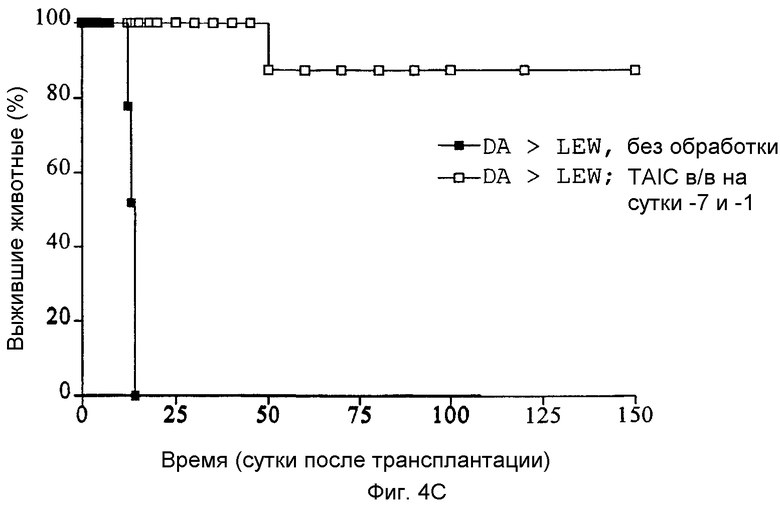
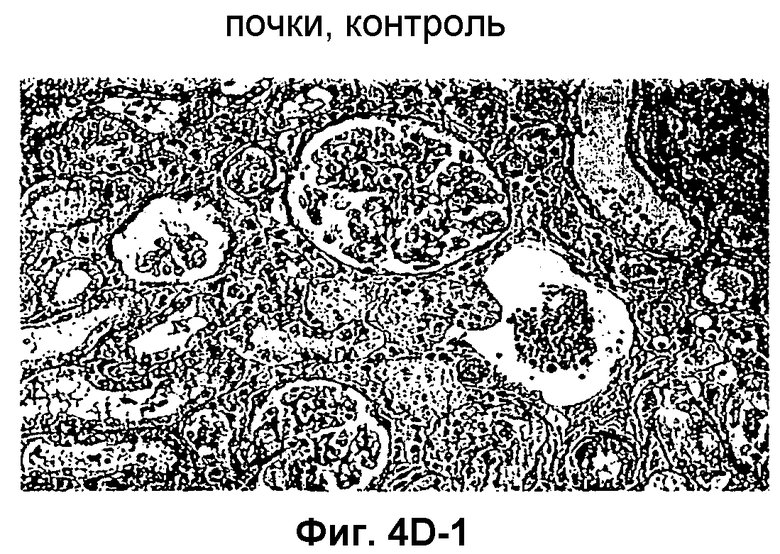
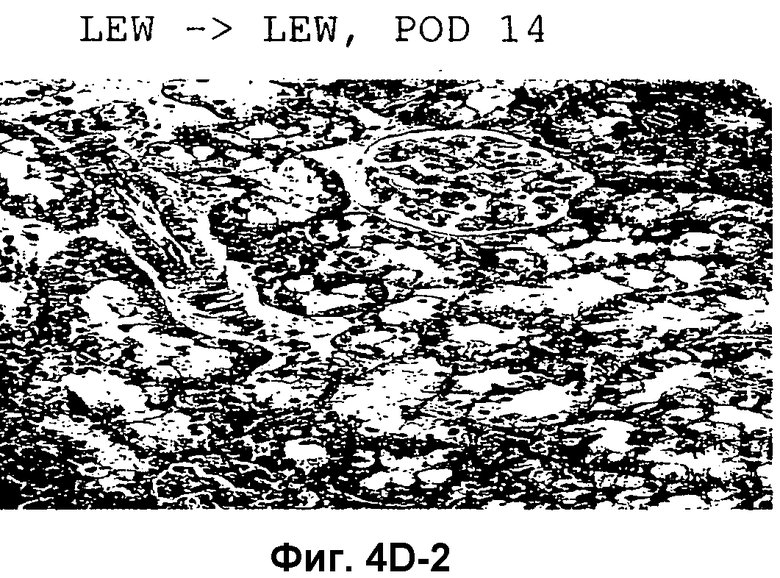
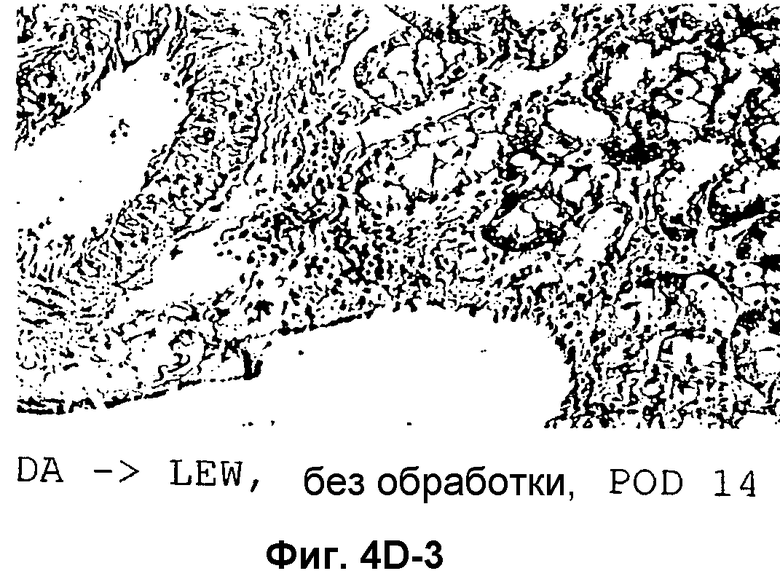
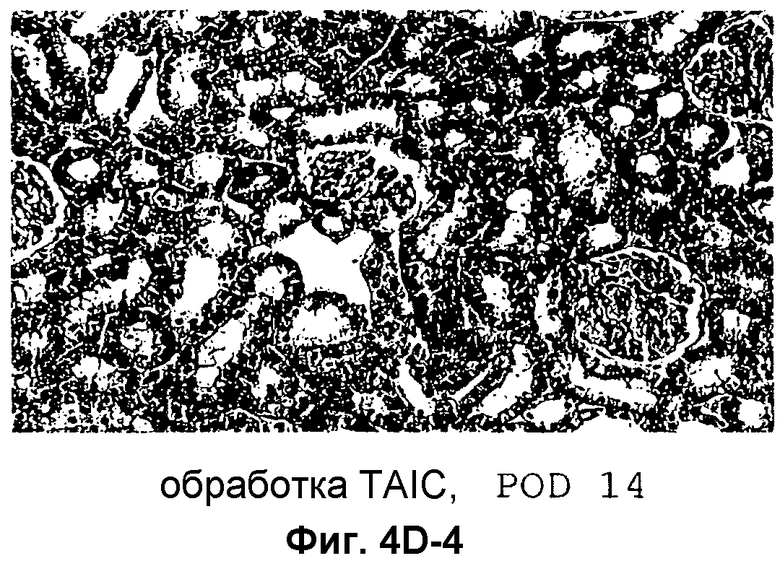

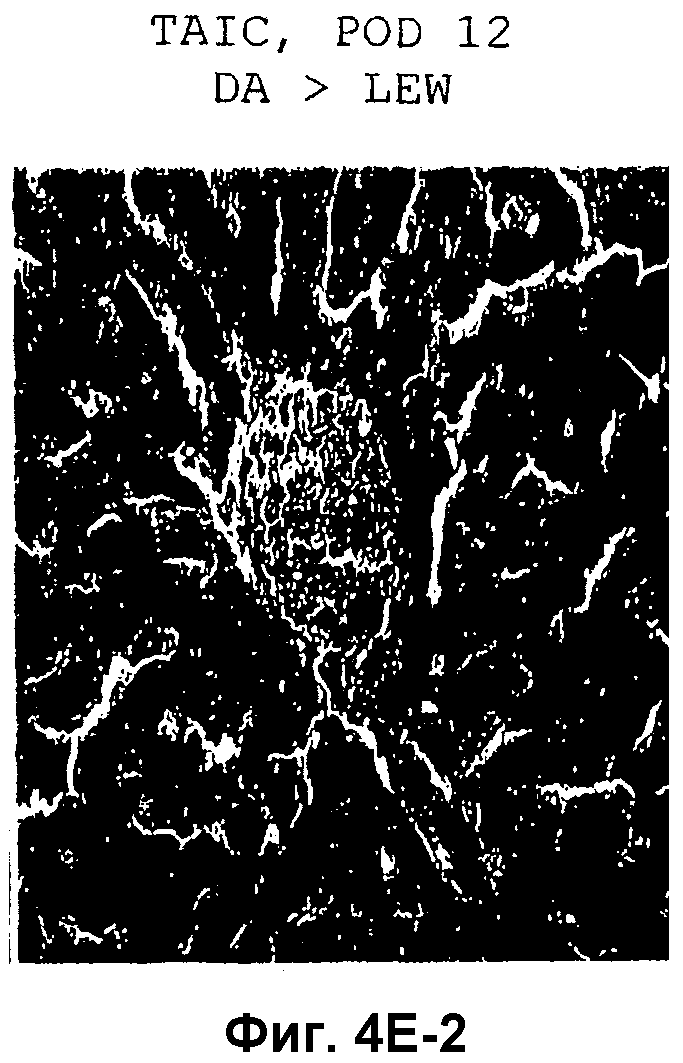

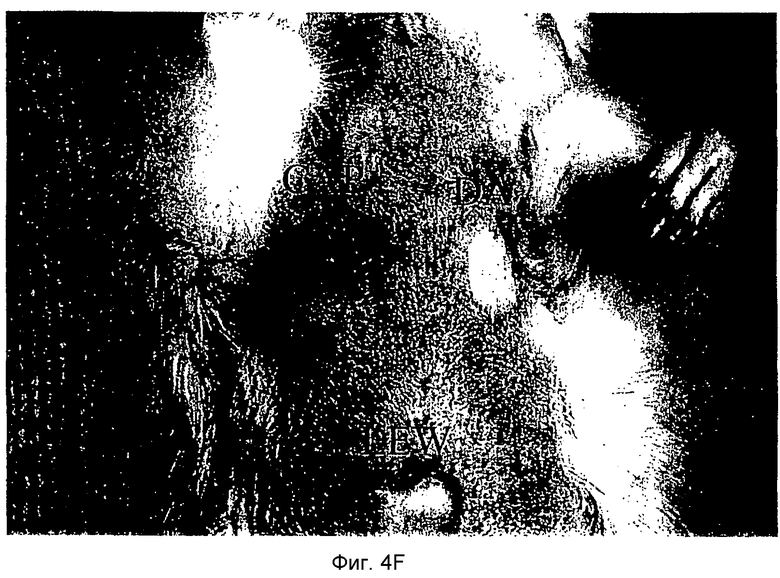
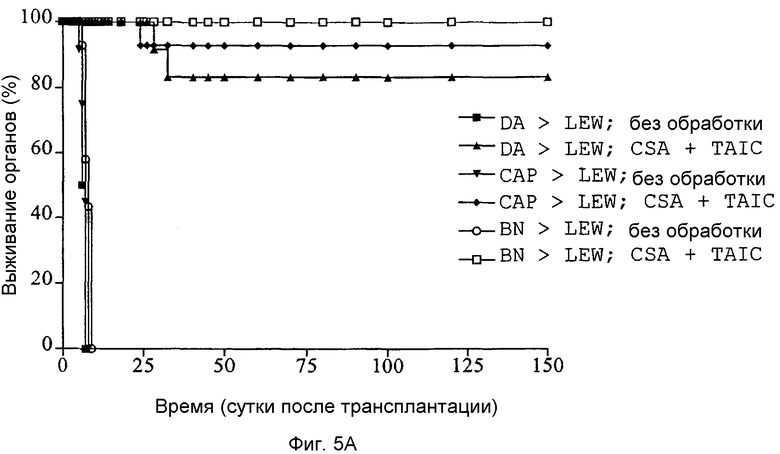
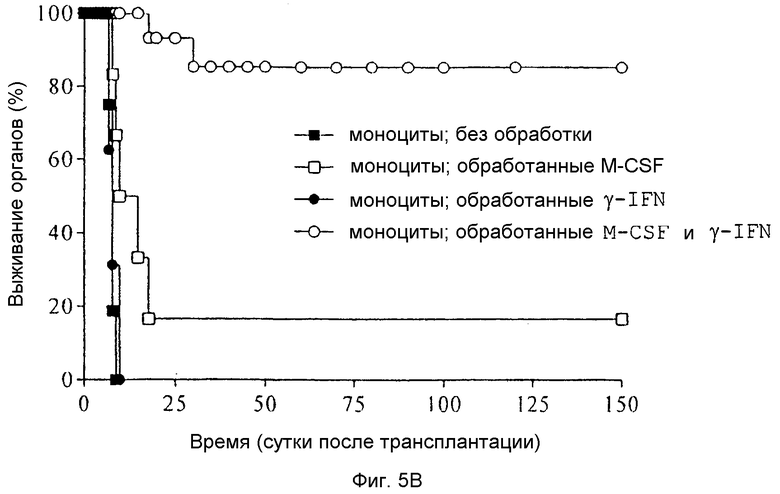
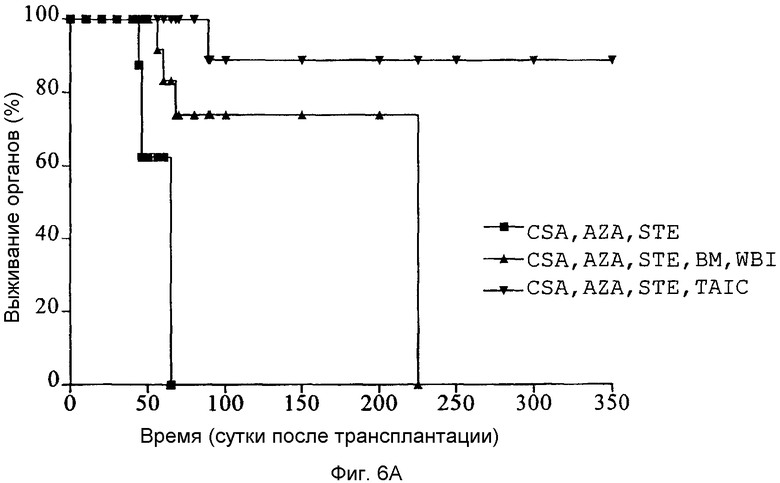
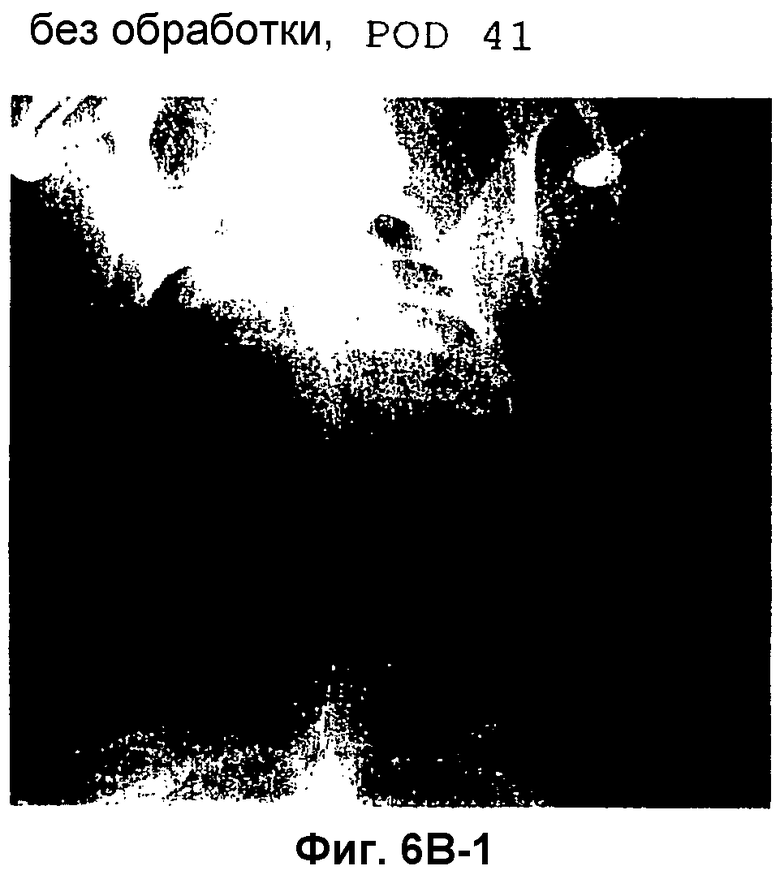

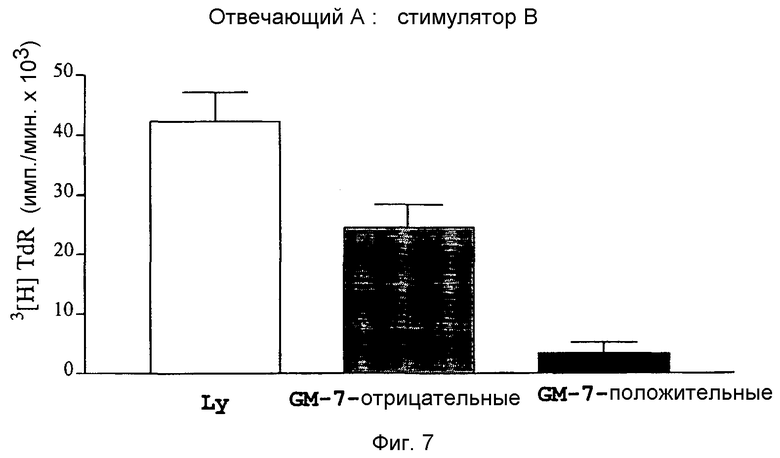
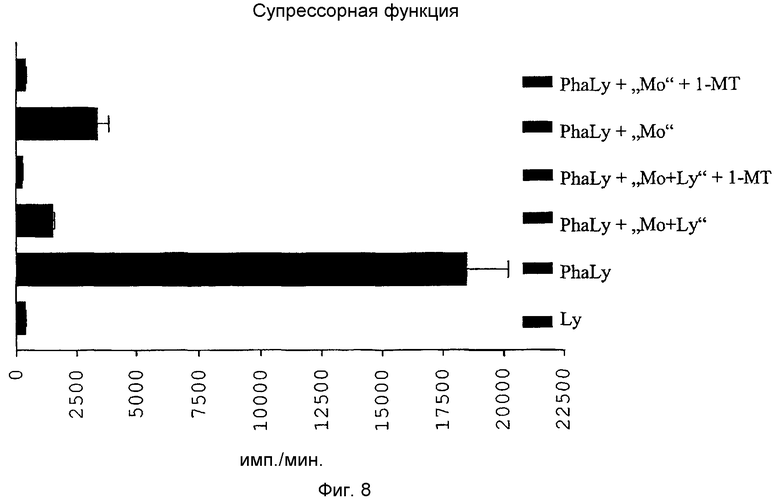
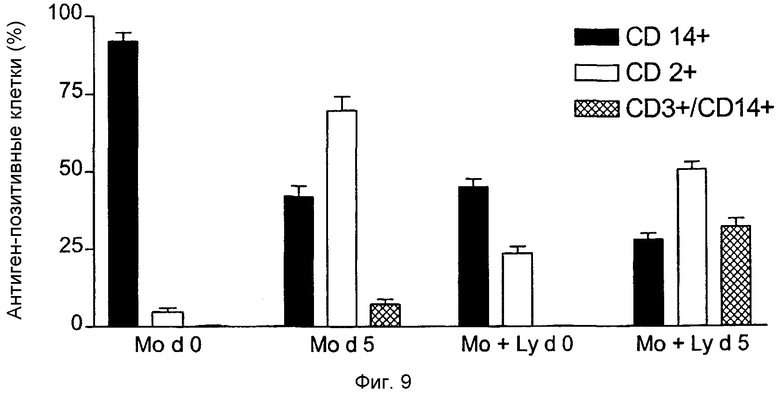
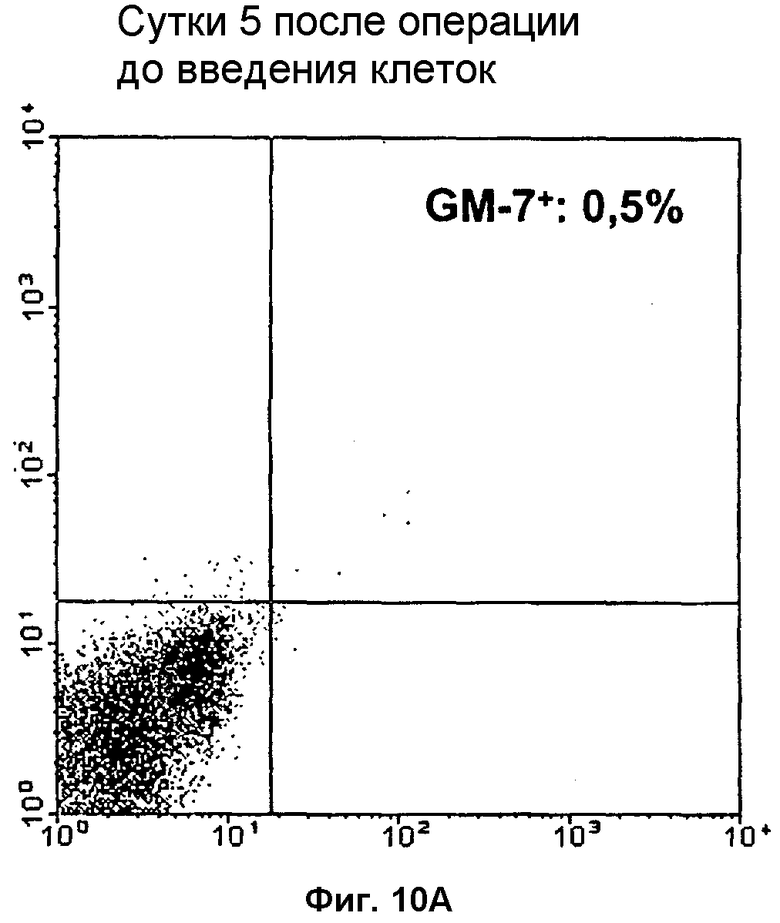
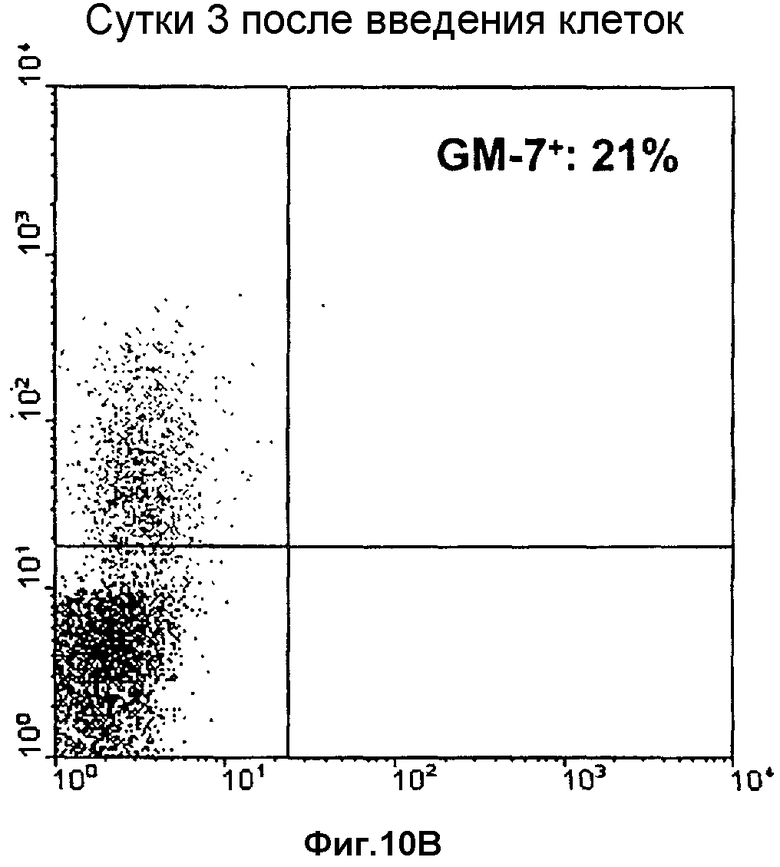
Изобретение относится к области биотехнологии, конкретно к получению индуцирующих воспринимаемость трансплантата клеток моноцитарного происхождения, экспрессирующих антигены CD3 и CD14, и может быть использовано в трансплантологии. Из крови выделяют моноциты, размножают их в культуральной среде с 1-20 мкг/л фактора роста клеток M-CSF и культивируют в течение 24-72 часов в культуральной среде с 0,1-20 нг/мл интерферона γ-IFN. После этого выделяют клетки из культуральной среды. Для выделения CD3 и CD14 клеток используют антитела, продуцируемые гибридомой DSM АСС2542. Полученные клетки при необходимости суспендируют и подвергают заморозке. Изобретение позволяет получить индуцирующие воспринимаемость трансплантата клетки моноцитарного происхождения, которые обладают эффектом подавления реакций отторжения трансплантата. 11 н. и 15 з.п. ф-лы, 29 ил., 1 табл.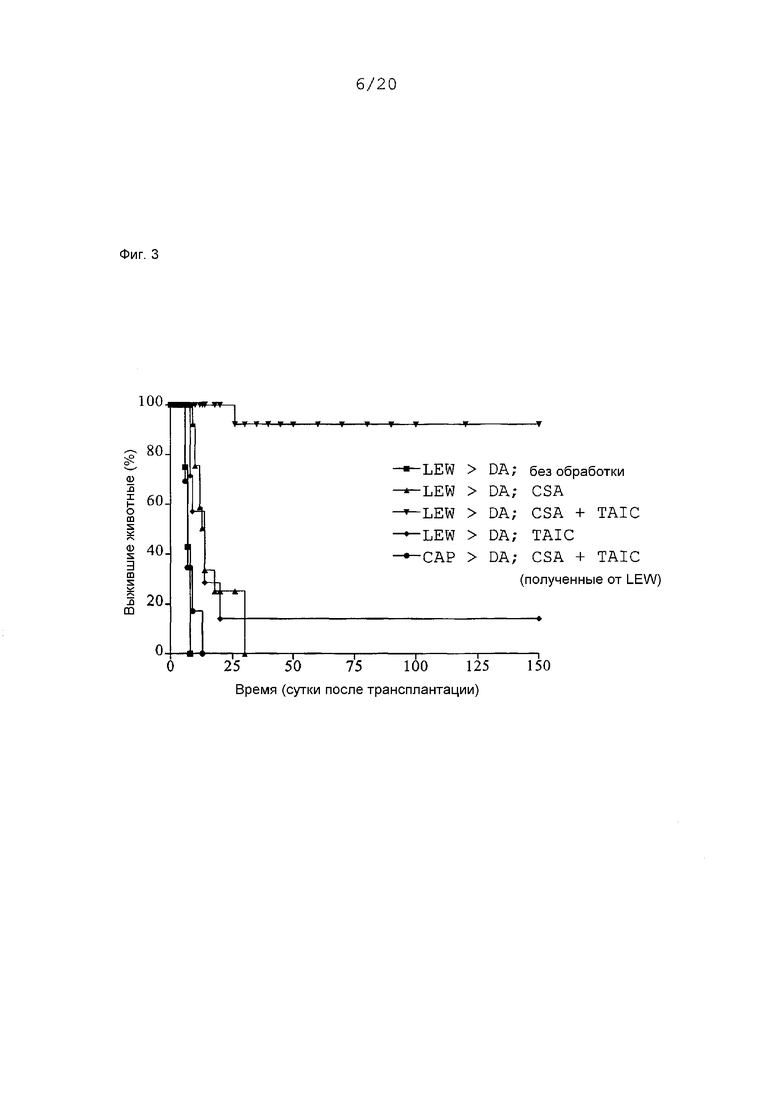
1. Способ получения индуцирующих воспринимаемость трансплантата клеток моноцитарного происхождения, экспрессирующих антигены CD3 и CD14, отличающийся тем, что:
a) из крови выделяют моноциты;
b) моноциты размножают в подходящей культуральной среде, которая содержит 1-20 мкг/л фактора роста клеток M-CSF;
c) моноциты одновременно со стадией b) или после нее культивируют в течение 24-72 ч в культуральной среде, содержащей 0,1-20 нг/мл интерферона γ-IFN; и
d) получают сформированные на стадии с) индуцирующие воспринимаемость трансплантата клетки посредством выделения клеток из культуральной среды,
e) необязательно выделяют индуцирующие воспринимаемость трансплантата CD3 и CD14 клетки посредством связывания с антителами, продуцируемыми гибридомой DSM АСС2542,
f) при необходимости полученные клетки суспендируют и подвергают заморозке.
2. Способ по п.1, отличающийся тем, что моноциты представляют собой моноциты человеческого происхождения.
3. Способ по п.2, отличающийся тем, что моноциты выделяют из крови таким образом, что вместе с моноцитами также присутствуют и лимфоциты, по меньшей мере, в количестве 10% по отношению к общему количеству клеток в изоляте.
4. Способ по п.3, отличающийся тем, что образующиеся на стадии с) или полученные на стадии d) индуцирующие воспринимаемость трансплантата клетки выделяют посредством связывания с антителами, полученными посредством гибридомной клеточной линии DSM АСС2542.
5. Способ по п.1 или 4, отличающийся тем, что среди образующихся на стадии с) или полученных на стадии d) по п.1 или полученных на стадии выделения по п.4 индуцирующих воспринимаемость трансплантата клеток выделяют такие клетки, которые на своей клеточной поверхности одновременно экспрессируют антигены CD3 и CD14.
6. Способ по п.1, отличающийся тем, что после стадии b) моноциты в течение от 24 до 72 ч культивируют в содержащей γ-IFN культуральной среде и культивирование в присутствии γ-IFN начинается с 3 по 6 сутки от начала культивирования на стадии b).
7. Способ по п.1, отличающийся тем, что общий период культивирования на стадиях b) и с) составляет от 4 до 8 суток.
8. Способ по п.1 или 4, отличающийся тем, что после стадии d) по п.1 или после стадий выделения по п.4 клетки суспендируют в пригодной клеточной культуральной среде, или в PBS, или растворе NaCl.
9. Способ по п.5, отличающийся тем, что после стадий выделения по п.5 клетки суспендируют в пригодной клеточной культуральной среде, или в PBS, или растворе NaCl.
10. Способ по п.8, отличающийся тем, что клетки суспендируют в среде для замораживания и далее подвергают глубокой заморозке.
11. Способ по п.9, отличающийся тем, что клетки суспендируют в среде для замораживания и далее подвергают глубокой заморозке.
12. Способ по п.10 или 11, отличающийся тем, что среда для замораживания содержит фетальную телячью сыворотку (FCS) или человеческую сыворотку АВ и DMSO.
13. Способ получения фармацевтической композиции для подавления реакций отторжения трансплантата, отличающийся тем, что клетки, полученные на стадии d) по пп.1-3 или 6-9, или клетки, полученные на стадии выделения по п.4 или 5, включают в состав композиции с фармацевтически приемлемым носителем.
14. Индуцирующие воспринимаемость трансплантата клетки моноцитарного происхождения, получаемые по любому из способов по пп.1-12, где указанные клетки одновременно экспрессируют антигены CD3 и CD14 на своей клеточной поверхности.
15. Индуцирующие воспринимаемость трансплантата клетки по п.14, отличающиеся тем, что они представляют собой клетки человеческого происхождения.
16. Клеточный препарат для индукции воспринимаемости транплантата, содержащий индуцирующие воспринимаемость трансплантата клетки по пп.14 и 15 или клетки, полученные на стадии d) по п.1, или клетки, полученные на стадии выделения по п.4, в среде, хорошо переносимой клетками.
17. Фармацевтическая композиция для подавления реакций отторжения трансплантата, содержащая индуцирующие воспринимаемость трансплантата клетки по пп.14 и 15, или клеточный препарат по п.16, или клетки, полученные на стадии d) по п.1, или клетки, полученные на стадии выделения по п.4, в фармацевтически приемлемом носителе.
18. Применение индуцирующих воспринимаемость трансплантата клеток по пп.14 и 15, или клеточного препарата по п.16, или клеток, полученных на стадии d) по п.1, или клеток, полученных на стадии выделения по п.4, для получения фармацевтической композиции для подавления реакций отторжения трансплантата.
19. Применение индуцирующих воспринимаемость трансплантата клеток по пп.14 и 15, или клеточного препарата по п.16, или клеток, полученных на стадии d) по п.1, или клеток, полученных на стадии выделения по п.4, для получения и/или размножения in vitro регуляторных Т-лимфоцитов.
20. Применение по п.19, где регуляторные Т-лимфоциты одновременно экспрессируют антигены CD4 и CD25 на своей клеточной поверхности.
21. Способ получения и/или размножения регуляторных Т-лимфоцитов, характеризующийся тем, что:
а) одновременно культивируют индуцирующие воспринимаемость трансплантата клетки по пп.14 и 15, или клеточный препарат по п.16, или клетки, полученные на стадии d) по п.1, или клетки, полученные на стадии выделения по п.4, с препаратом Т-лимфоцитов и b) необязательно выделяют регуляторные Т-лимфоциты из клеточной среды.
22. Способ по п.21, отличающийся тем, что регуляторные Т-лимфоциты одновременно экспрессируют антигены CD4 и CD25 на своей клеточной поверхности.
23. Способ по п.21 или 22, отличающийся тем, что регуляторные Т-лимфоциты выделяют из клеточной среды сортировкой FACS.
24. Гибридомная клеточная линия DSM АСС2542, продуцирующая антитела, которые специфически распознают индуцирующие воспринимаемость трансплантата клетки, получаемые способом по п.1.
25. Антитело, продуцируемое гибридомной клеточной линией DSM АСС2542, пригодное для выделения клеток в способе по п.1.
26. Применение антитела по п.25 для выявления и/или выделения индуцирующих воспринимаемость трансплантата клеток.
| BUCKLE A.M | |||
| et al., Colony-stimulating factors and interferon-gamma differentially affect cell surface molecules shared by monocytes and neutrophils, Clin | |||
| Exp.Immunol., 1990, v.81, n.2, p.339-345 | |||
| TANAKA J | |||
| et al., The role of accessory cells in allogeneic peripheral blood stem cell transplantation, Int | |||
| J | |||
| Hematol., 1999, v.69, n.2, p.70-74 | |||
| СПОСОБЫ ИНДУЦИРОВАНИЯ Т-КЛЕТОЧНОЙ ТОЛЕРАНТНОСТИ К ТКАНЕВОМУ ИЛИ ОРГАННОМУ ТРАНСПЛАНТАТУ | 1995 |
|
RU2169009C2 |

