ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к области медицинских устройств и, более конкретно, к устройствам, направленным на отбор клеток, которые устойчивы к апоптотическим сигналам, способам применения устройств и областям их применения.
УРОВЕНЬ ТЕХНИКИ
Стволовые клетки представляют собой клетки, которые способны как делиться и дифференцироваться в разнообразные специализированные типы клеток, так и самообновляться с образованием большего количества стволовых клеток. У млекопитающих стволовые клетки встречаются либо в виде эмбриональных стволовых клеток, которые выделяют из внутренней клеточной массы бластоцист, либо в виде стволовых клеток взрослого организма, которые встречаются в различных тканях. У взрослых организмов стволовые клетки и клетки-предшественники выполняют функцию системы восстановления организма, восполняя ткани взрослого организма.
В отличие от всех современных методов лечения, основанных на хирургическом вмешательстве или лекарственных препаратах, которые модулируют физиологические процессы, стволовые клетки обеспечивают замещение дисфункциональной или дегенерирующей ткани. Таким образом, применение стволовых клеток в заместительной терапии может обеспечить кардинальное изменение прогноза многих неизлечимых в настоящее время болезней, восстановление функции поврежденных органов и коррекцию врожденных нарушений обмена веществ и дефектов. Важность технологий, ассоциированных с экспансией стволовых клеток, источником которых являются как ткани взрослого организма, так и/или эмбриональные ткани, иллюстрируется многочисленными доклиническими и клиническими применениями этих клеток в лечении широкого круга болезней.
Совсем недавние достижения показали, что несколько типов стволовых клеток в пределах гемопоэтического компартмента, включая гемопоэтические стволовые клетки и клетки-предшественники (HSPC) и мезенхимальные стромальные клетки (МСК), обладают способностью к дифференцировке в типы клеток, выходящие за пределы иммуно-гемопоэтической системы, создавая возможность применения этих типов клеток для восстановления и регенерации тканей при дегенеративных нарушениях широкого спектра, купирования органной недостаточности и дисфункции и, возможно, замещения органных трансплантатов методами клеточной терапии.
Одно из основных клинических применений стволовых клеток опосредовано трансплантациями гемопоэтических стволовых клеток (ТГСК). При этой методике некоторое количество клеток, полученных от донора, пересаживают реципиенту с целью восстановления иммунной и гемопоэтической систем реципиента. При осуществлении этих трансплантаций выяснилось, что восстановление иммунной системы фактически является наилучшей терапией для целого ряда раковых заболеваний, устойчивых к химиотерапии, таких как лейкемия, лимфома, множественная миелома и целого ряда солидных опухолей. Помимо применения в онкологии ТГСК обладает потенциалом для лечения доброкачественных нарушений, таких как аутоиммунные болезни (например, диабет Т1, СКВ и болезнь Крона), врожденных нарушений обмена веществ и энзимопатий, гемоглобинопатии или врожденного и приобретенного иммунодефицита. ТГСК применяли в случае неонкологических показаний, при которых отмечались значительные положительные результаты. Тем не менее, данную методику используют в настоящее время в случае состояний, угрожающих жизни, вследствие серьезных токсических последствий, из которых самой важной является болезнь «трансплантат против хозяина» (БТПХ).
При использовании данного подхода для лечения рака чаще всего его осуществляют после миелоаблятивного предварительного кондиционирования путем уничтожения клеток иммуно-гемопоэтической системы посредством агрессивной радио-химиотерапии, с тем, чтобы обеспечить предотвращение отторжения трансплантата. Современное понимание того, что клетки иммуно-гемопоэтической системы хозяина не обязательно необходимо уничтожать до трансплантации, было значительным достижением, соответственно произошла замена миелоаблативного режима кондиционирования немиелоаблативным режимом кондиционирования и кондиционированием сниженной интенсивности (RIC). Применение немиелоаблативного режима кондиционирования привело к значительному, пока еще недостаточному, улучшению, опасных для жизни ситуаций, вызванных дисфункцией жизненно важных органов, неприживлением трансплантата и некупируемыми инфекциями. Поскольку гемопоэтическая система хозяина сохраняется при RIC, она может восстановиться в том случае, если донорские клетки не приживились. С течением времени, в донорском трансплантате происходит процесс, который способствует возникновению реакций «трансплантат против опухоли» (ТПО) и «трансплантат против аутоиммунитета» (ТПА), при этом он еще дополнительно подвергает пациента критическому уровню заболеваемости и смертности от БТПХ.
Идентификация трансмембранного белка/адгезивной молекулы типа 1, сиаломуцина CD34, как маркера гемопоэтических стволовых клеток сделало возможным применение отбора клеток с помощью CD34+ как средства концентрирования гемопоэтических стволовых клеток с целью трансплантации. Маркеры CD34 отсутствуют у некоторых стволовых клеток и встречаются также на поверхности предшественников клеток крови, относящихся к различным субтипам. Применение такого способа положительного отбора приводит к потере некоторых из стволовых клеток с полезными свойствами. Более того, он обеспечивает получение смешанной клеточной популяции стволовых клеток и клеток-предшественников с некоторыми поздними клетками-предшественниками. Трансплантация такой смешанной клеточной популяции уменьшает успех процесса трансплантации [Askenasy N. et al., Current Stem Cell Research and Therapy 2006; 1:85-94]. Таким образом, возникла потребность в способе отбора, который бы обеспечивал сохранение всех клеток, необходимых для восстановления гемопоэза, при этом исключал возникновение неблагоприятных эффектов, оказывающих влияние на клетки.
Было подтверждено документально, что в отличие от соматических клеток гемопоэтические стволовые и клетки-предшественники (HSPC), мезенхимальные стромальные клетки и предшественники нейральных клеток (NP) являются нечувствительными к повреждающим факторам, как, например, факторам, которые наносят повреждение, как в результате радио-химиотерапии, так и в результате образования вторичных факторов, которые высвобождаются в костный мозг в результате массивной гибели резидентных гемопоэтических клеток. HSPC особенно устойчивы к апоптотическим сигналам, которые передаются через рецепторы семейства фактора некроза опухоли (ФНО), которые вместо этого используются для передачи ростовых сигналов в примитивные клетки-предшественники. В моделях на мышах было показано, что в гемопоэтических клетках-предшественниках в условиях повреждения и стресса сильно повышается экспрессия нескольких рецепторов семейства ФНО. В условиях трансплантации этот физиологический механизм способствует приживлению примитивных клеток-предшественников по сравнению с донорскими клетками, чувствительными к апоптозу. Таким образом, подвергание трансплантируемой популяции клеток воздействию апоптоз-индуцирующих лигандов семейства ФНО, таких как FasL, Trail, Tweak или ФНО-α, приводит к отрицательному отбору популяции стволовых клеток, поскольку клеточные популяции, чувствительные к апоптозу, индуцированному лигандом семейства ФНО, подвергаются апоптозу и удаляются из трансплантата. Применение такого способа в модели на мышах было раскрыто в заявке на патент WO 2007/138597.
Состав донорского трансплантата является важным параметром трансплантации стволовых клеток. Было показано, что для обеспечения приживления требуется пороговое количество клеток-предшественников. В дополнение к этому, присутствие некоторых субпопуляций нестволовых клеток существенно повышает вероятность приживления, так что трансплантация гетерогенных смесей клеток является более эффективной, чем трансплантация изолированных клеток-предшественников. Наиболее важными субпопуляциями в составе донорского трансплантата являются (CD4+ и CD8+) Т-клетки, поскольку было продемонстрировано, что они препятствуют отторжению и поддерживают приживление гемопоэтических клеток-предшественников внутри микроокружения костного мозга. Однако трансплантация аллогенных Т-клеток частично иммуносупрессированным реципиентам обеспечивает долговременное приживление, которое опосредует развитие потенциально летальной реакции «трансплантат против хозяина» (РТПХ) или болезни «трансплантат против хозяина» (БТПХ). Зрелые донорские Т-клетки опосредуют эту реакцию, тогда как донорские Т-клетки, которые образовались de novo после трансплантации, совместимы с хозяином. Прилагались большие усилия для разделения субпопуляций Т-клеток, которые опосредуют РТПХ и поддерживают приживление; однако до настоящего времени экспериментальные данные были неубедительными.
Болезнь «трансплантат против хозяина» (БТПХ) включает острофазовую форму реакции, которая обычно возникает в течение первых 100 суток после трансплантации, и хроническую форму реакции с более медленным прогрессированием, но с аналогичными пагубными последствиями. Важно отметить, что обе реакции запускаются первичной воспалительной реакцией, опосредованной зрелыми донорскими Т-клетками в пределах трех суток с момента трансплантации. Острую форму БТПХ обычно лечат с помощью иммуносупрессивной терапии, которая оказывает отрицательное влияние на восстановление гемопоэза, при этом в настоящее время не существует эффективной терапии хронической формы реакции. Традиционный подход к предотвращению БТПХ заключается в деплеции зрелых Т-клеток, получаемых из донорского инокулята, которое время от времени сопровождается деплециями В-лимфоцитов с применением поверхностных клеточных маркеров, таких как CD3, CD4, CD8. Огромные усилия по достижению более селективной деплеции Т-клеток (TCD) с применением различных поверхностных клеточных маркеров не принесли результата.
Более специфической деплеции достигли с приличными результатами путем удаления субпопуляций реактивных Т-клеток с применением апоптотических сигналов после сенсибилизации ex vivo против антигенов хозяина. Сенсибилизированные Т-клетки экспрессируют репертуары молекул, связанных с активацией, становятся чувствительными к гибели клеток, индуцированной активацией (AICD), и пролиферируют с высокими скоростями - это характеристики, которые используют в случае специфической деплеции. Однако в случае этого подхода существует несколько основных препятствий для селективного удаления. Во-первых, сенсибилизация Т-клеток представляет собой процесс повторного воздействия антигенов, который требует 3-7 суток инкубации ex vivo. Вследствие этого, активацию Т-клеток следует осуществлять по меньшей мере за 3 суток до трансплантации. Во-вторых, сенсибилизация и активация, которые делают быстроциркулирующие Т-клетки чувствительными к AICD, также индуцируют рост и экспансию эффекторных/Т-клеток памяти, чья персистирующая аллореактивность может инициировать и обеспечить распространение БТПХ. Эффекторные/Т-клетки памяти относительно устойчивы к образованию поперечных связей в Fas, отчасти из-за присущих низких уровней каспазы-3, что приводит к образованию фенотипа, характеризующегося устойчивостью к апоптозу, который вызывает у пациентов предрасположенность к острой и хронической форме БТПХ после инфузии Т-клеток, культивированных ex vivo. В-третьих, наиболее эффективная сенсибилизация ex vivo направлена против компонентов главного комплекса гистосовместимости (ГКГС), различие в которых приводит к основным аллоиммунным ответам в условиях трансплантации. Однако БТПХ стимулируется прежде всего антигенами минорного комплекса гистосовместимости (miHA) и нацелена главным образом на тканеспецифические антигены (ТСА). Вследствие повреждения, которое наносится в результате дотрансплантационного кондиционирования и воспаления, происходит экспонирование нормальных тканевых эпитопов и тканевых эпитопов с нарушенной структурой. Все же следует разработать эффективный способ селективной деплеции Т-клеток, которые индуцируют БТПХ, с сохранением субпопуляций клеток, которые поддерживают ТПО и приживление трансплантата.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение раскрывает устройство и набор, приспособленные для отбора клеток, которые устойчивы к опосредованному рецепторами апоптозу, и способ применения устройства и набора. Устройство обеспечивает отрицательный отбор зрелых иммунокомпетентных клеток, которые индуцируют реакцию «трансплантат против хозяина» (БТПХ) из гетерогенной клеточной популяции, которую вводят в устройство. Устройство обеспечивает эффективный отбор клеток с использованием упрощенного и более дешевого коммерчески доступного продукта - решение, которое на настоящий момент является новым. Настоящее изобретение дополнительно раскрывает области применения устройства.
В соответствии с одним аспектом настоящее изобретение раскрывает устройство, содержащее контейнер, который изготовлен из биосовместимого материала, и биологически активный апоптоз-индуцирующий лиганд, иммобилизованный на поверхности, отличающееся тем, что устройство приспособлено для отбора клеток.
В соответствии с одним вариантом реализации поверхность, на которой иммобилизируют биологически активный апоптоз-индуцирующий лиганд, является внутренней поверхностью контейнера.
В соответствии с другим вариантом реализации поверхность, на которой иммобилизируют биологически активный апоптоз-индуцирующий лиганд, является поверхностью микрогранул, присутствующих внутри контейнера.
В соответствии с другим вариантом реализации контейнер выбирают из группы, состоящей из пакета, колонки, трубки, бутылки, пузырька и флакона.
В соответствии с еще одним вариантом реализации биосовместимый материал, из которого устройство состоит, выбирают из группы, состоящей из: полипропилена, полистирола, силикона, поливинилхлорида или их комбинации.
В соответствии с еще одним вариантом реализации иммобилизованный апоптоз-индуцирующий лиганд выбирают из группы, состоящей из: фактора некроза опухоли (ФНО-α), Fas-лиганда (FasL), Trail, Tweak или любой их комбинации.
В соответствии с другим аспектом настоящее изобретение раскрывает способ отбора клетки, устойчивой к апоптотическим сигналам, из гетерогенной клеточной популяции; клеточная популяция состоит из клеток, устойчивых к апоптотическим сигналам, и клеток, чувствительных к апоптотическим сигналам. Способ заключается во введении образца, содержащего гетерогенную клеточную популяцию, в устройство настоящего изобретения и инкубации клеток внутри устройства, посредством этого проходит отбор Т-клеток, устойчивых к апоптотическим сигналам.
В соответствии с другим вариантом реализации отобранная клетка, устойчивая к апоптозу, представляет собой стволовую клетку, отобранную из группы, состоящей из: стволовой клетки пуповинной крови, мобилизованной стволовой клетки периферической крови, стволовой клетки костного мозга, раковой стволовой клетки и нейральной стволовой клетки.
В соответствии с другим вариантом реализации отобранная клетка, устойчивая к апоптозу, представляет собой иммунокомпетентную клетку, нечувствительную к гибели клеток, индуцированной активацией (AICD). Еще в другом варианте реализации иммунокомпетентная клетка, нечувствительная к гибели клеток, индуцированной активацией (AICD), представляет собой Т-клетку.
В соответствии с другим вариантом реализации отобранная клетка, устойчивая к апоптозу, представляет собой клетку-предшественник.
В соответствии с еще одним вариантом реализации клеточную популяцию, которую используют в способе, получают из: костного мозга, периферической крови после мобилизации клеток-предшественников или пуповинной крови (ПК).
В соответствии с еще одним вариантом реализации время инкубации внутри устройства составляет от 2 часов до 72 часов.
В соответствии с еще одним аспектом настоящее изобретение раскрывает набор для отбора клеток, содержащий устройство настоящего изобретения и раствор для поддержания сохранности и активности апоптоз-индуцирующего лиганда внутри устройства. В некоторых вариантах реализации набор дополнительно содержит апоптоз-индуцирующий лиганд, отобранный из группы фактора некроза опухоли (ФНО-α), Fas-лиганда (FasL), Trail, Tweak или любой их комбинации.
В соответствии с другим вариантом реализации раствор для поддержания сохранности и активности апоптоз-индуцирующего лиганда представляет собой буфер или среду.
В соответствии с еще одним аспектом настоящее изобретение раскрывает способ применения вышеупомянутого устройства и способа. Способ представляет собой способ улучшения клинического исхода трансплантации гемопоэтических стволовых клеток и клеток-предшественников (HSPC). В данном способе получают образец, содержащий клеточную популяцию; клеточная популяция содержит стволовые и клетки-предшественники. Обеспечивают контакт популяции с апоптоз-индуцирующим лигандом и оставшиеся клетки выделяют и используют для трансплантации. В соответствии с дополнительным вариантом реализации контакт происходит внутри устройства настоящего изобретения.
В соответствии с другим вариантом реализации клеточную популяцию в настоящем способе получают из: костного мозга, периферической крови после мобилизации клеток-предшественников или пуповинной крови (ПК).
В соответствии с другим вариантом реализации стволовую клетку, которую выбирают с применением настоящего способа, выбирают из группы, состоящей из: стволовой клетки пуповинной крови, мобилизованной стволовой клетки периферической крови, стволовой клетки костного мозга, раковой стволовой клетки и нейральной стволовой клетки.
В соответствии с другим вариантом реализации популяцию клеток инкубируют с Fas-лигандом (FasL) в течение периода от 21 до 24 часов и с фактором некроза опухолей (ФНО-α) в течение периода от 24 до 48 часов.
В соответствии с другим вариантом реализации выделенные клетки в настоящем способе используют либо для аутологичных, аллогенных, либо гаплоидентичных трансплантаций.
В соответствии с другим вариантом реализации выделенные клетки в настоящем способе дополнительно содержат иммунокомпетентную клетку, нечувствительную к гибели клеток, индуцированной активацией (AICD). Еще в другом варианте реализации иммунокомпетентная клетка, нечувствительная к гибели клеток, индуцированной активацией (AICD), представляет собой Т-клетку.
В соответствии с еще одним аспектом настоящее изобретение раскрывает другой способ применения вышеупомянутого устройства и способа. Способ представляет собой способ удаления злокачественной клетки в композиции, содержащей трансплантат, состоящий из клеток-предшественников. В настоящем способе получают композицию, содержащую трансплантат, состоящий из клеток-предшественников, и затем обеспечивают контакт с апоптоз-индуцирующим лигандом. В соответствии с дополнительным вариантом реализации контакт происходит внутри устройства настоящего изобретения.
В соответствии с другим вариантом реализации обеспечивают контакт композиции с апоптоз-индуцирующим Fas-лигандом (FasL) в течение периода около 24 часов.
В соответствии с другим вариантом реализации трансплантат, состоящий из клеток-предшественников, используют в качестве аутологичного трансплантата.
В соответствии с другим аспектом настоящее изобретение раскрывает еще один способ применения вышеупомянутого устройства и способа. Способ представляет собой способ предотвращения болезни «трансплантат против хозяина» (БТПХ) с сохранением эффекта «трансплантат против опухоли» (ТПО). В данном способе получают образец, содержащий клеточную популяцию, причем клеточная популяция содержит HSPC и иммунокомпетентные клетки. Обеспечивают контакт популяции с апоптоз-индуцирующим лигандом, и оставшиеся клетки выделяют и используют для трансплантации. В соответствии с дополнительным вариантом реализации контакт происходит внутри устройства настоящего изобретения.
В соответствии с другим вариантом реализации обеспечивают контакт композиции с апоптоз-индуцирующим Fas-лигандом (FasL) в течение периода от 2-16 часов.
Дополнительные варианты реализации, отличительные признаки, преимущества и полный объем применимости настоящего изобретения станут очевидными на основе подробного описания и чертежей, приведенных в дальнейшем в этом документе. Однако следует отметить, что тогда как подробное описание, позволяет выявить предпочтительные варианты реализации настоящего изобретения, оно приводится лишь в иллюстративных целях, поскольку различные изменения и модификации внутри сущности и объема настоящего изобретения станут очевидными специалистам в данной области техники на основе данного подробного описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1. Ряд графиков, на которых показана чувствительность клеток ПК к опосредованному рецепторами апоптозу in vitro. (А) Свежие образцы клеток ПК инкубировали в среде без добавок хемокинов в течение различных периодов времени (n=35) и подвергали воздействию 50 нг/мл олигомеров FasL (n=21) и 20 нг/мл ФНО-альфа (n=18). Апоптоз определяли по поглощению аннексина V в мононуклеарных клетках (МНК), и гейтированных CD34+ клетках-предшественниках. (В) Экспрессия рецепторов Fas и ФНО в свежевыделенных мононуклеарных клетках ПК, гейтированных CD34+ и выделенных клетках-предшественниках негативной линии дифференцировки (lin-). (С) Апоптоз измеряли в зависимости от времени в гейтированных CD34+ клетках-предшественниках, экспрессирующих рецепторы Fas и ФНО под влиянием родственных лигандов (n=15-21). (D) Показатели пролиферации МНК и гейтированных CD34+ клеток-предшественников, экспрессирующих рецепторы Fas и ФНО, инкубированных с родственными лигандами (n=6). Пролиферацию измеряли по разведению CFSE с применением программного обеспечения ModFit. (Е) Экспрессия рецепторов Fas и ФНО в Т-клетках (CD3+), В-лимфоцитах (CD 19+) и миелоидных клетках (CD33+) в свежевыделенных образцах ПК (n=8-13). (F) Апоптоз, определенный по поглощению аннексина V в субпопуляциях клеток ПК позитивной линии дифференцировки, включая моноциты/макрофаги (CD 14+), которые подвергали воздействию в течение 48 часов FasL и ФНО- - (n=6-14).
Фигура 2. Фигура 2А представляет собой схему и фигуры 2B-2G представляют собой графики, на которых показано, что клетки-предшественники ПК устойчивы к опосредованному рецепторами апоптозу. (А) Мышей NOD.CID подвергали кондиционированию с помощью двух доз 25 мкг/г бусульфана и им трансплантировали через 2 суток равные количества клеток ПК, инкубированных в разных условиях. Уровень химеризма по линии человеческих клеток измеряли в костном мозге через 12 недель с применением селективных человеческих и мышиных антител к CD45. (В) Приживление свежевыделенных клеток ПК и через 24 часа после инкубации в среде с 50 нг/мл FasL и 20 нг/мл ФНО-α из одинаковой дозы ПК. Данные соответствуют 7 разным образцам ПК. (С) Приживление через 48 часов после инкубации в среде FasL и ФНО-α из одинаковой дозы ПК (соответствует 6 образам ПК). (D) Инкубация в течение 72 часов в среде с FasL и ФНО-α из одинаковой дозы ПК ухудшает приживление (соответствует 5 образцам ПК). (Е) Показатели пролиферации мононуклеарных клеток (МНК), гейтированных CD34+ и выделенных клеток-предшественников негативной линии дифференцировки (lin-) через 48 часов после инкубации в среде с FasL и ФНО-α (n=4-7). (F) Фракции митотически покоящихся субпопуляций ПК, находящихся в фазе клеточного цикла G0/G1, которые определили по включению в ядро пропидиум иодида (n=5-8). (G) Количество клеток, инициирующих долгосрочную культуру (LTC-IC), определяли после посева 103 клеток ПК на слоях мезенхимальных стромальных клеток в течение 5 недель и впоследствии переносили на полутвердую среду на основе метилцеллюлозы. FasL (50 нг/мл) и ФНО-α (20 нг/мл) присутствовали в течение всего периода культивирования, и их обновляли с недельными интервалами путем замены половины объема среды (n=8-13). Данные представляют сравнительные условия культивирования для одной дозы ПК.
Фигура 3. Фигура 3В представляет собой схему и фигуры 3А и 3С представляют собой графики, на которых показано, что подвергание клеток ПК воздействию лигандов рецепторов смерти повышает количество клеток-предшественников in vitro и степень миелоидной дифференцировки in vivo. (А) Экспрессия маркеров дифференцировки клеточных линий в свежевыделенных клетках ПК: клетки-предшественники (CD34), Т-клетки (CD3), В-лимфоциты (CD19) и миелоидные клетки (CD33). Изменение состава жизнеспособных клеток после инкубации в течение 24 и 48 часов под воздействием 50 нг/мл FasL и 20 нг/мл ФНО-α, что приводит к увеличению фракции клеток-предшественников (n=17-31). (В) После инкубации в течение различных периодов времени мертвые клетки удаляли путем центрифугирования в градиенте фиколла, а жизнеспособные клетки в равных количествах высевали на чашки с полужидкой средой на основе метилцеллюлозы, которые стимулировали с использованием фактора стволовых клеток (ФСК), интерлейкина-3 (IL-3) и гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ). Количество колониеобразующих клеток (КОЕ, в пересчете на 103 жизнеспособных клеток) определяли через 14 суток. (С) Относительное количество КОЕ в свежевыделенных образцах ПК (контроль) и после инкубации в среде с FasL и ФНО-α (n=12-27).
Фигура 4. Фигура 4А представляет собой схему и фигуры 4B-4D представляют собой графики, на которых показано комбинированное воздействие лигандов рецепторов смерти на клетки ПК. (А) Экспрессию Fas стимулировали путем инкубации с 20 нг/мл ФНО-α в течение 24 часов, и впоследствии клетки инкубировали в течение дополнительных 24 часов при наличии и отсутствии 50 нг/мл FasL. (В) Экспрессия Fas в мононуклеарных клетках (МНК) и гейтированных CD34+ клетках-предшественниках в различных условиях инкубации (n=6-9). (С) Апоптоз Fas-положительных МНК, гейтированных CD34+ и выделенных клеток-предшественников негативной линии дифференцировки (lin-) на протяжении 48 часов после инкубации с ФНО-α с добавлением и без добавления FasL (n=5-12). (D) После удаления мертвых клеток путем центрифугирования в фиколле повышается количество КОЕ в смешанных суспензиях клеток после различных инкубаций (n-7-15).
Фигура 5. Фигура 5С представляет собой схему и фигуры 5А, 5В и 5D представляют собой графики, на которых показано, что лиганды рецепторов смерти повышают количество клеток-предшественников на протяжении экспансии ex vivo клеток ПК. (A) CD34+ клетки-предшественники выделяли с помощью иммуномагнитной сепарации и подвергали экспансии в суспензионной культуре в течение 3 недель по протоколу, одобренному для использования в клинической практике. По сравнению с образцом свежей ПК количество клеток во фракции CD34+ клеток-предшественников увеличивалось на протяжении экспансии и дополнительно увеличивалось после добавления 50 нг/мл FasL на протяжении третьей и последней недели культивирования (n=4). (В) Абсолютные количества CD34+ клеток в культурах через 3 недели экспансии (n=4) нормализовали по общему количеству клеток 103. (С) Равные количества свежевыделенных клеток ПК и клеток после экспансии (из одного и того же образца) трансплантировали мышам NOD.SCID (H2Kg7), которых подвергали кондиционированию с помощью двух доз 25 мкг/г бусульфана. (D) Через 12 недель в костном мозге и селезенке определяли уровень химеризма по линии гемопоэтических клеток. Данные соответствуют 6 независимым дозам ПК и отражают сравнение свежевыделенных клеток, клеток-предшественников после экспансии при наличии и отсутствии FasL на протяжении последней третьей недели культивирования.
Фигура 6. Фигуры 6A-6F представляют собой графики, на которых показана экспрессия рецепторов смерти и чувствительность к апоптозу клеток мПК на протяжении краткого культивирования. Образцы мПК, криоконсервированные в течение 2-7 лет, отогревали, удаляли избыток ДМСО и клетки инкубировали в течение 4 и 16 часов при наличии и отсутствии 50 нг/мл FasL и 29 нг/мл ФНО-α. (А) Экспрессия рецепторов Fas и ФНО в мононуклеарных (МНК) клетках мПК и гейтированных CD34+ клетках-предшественниках (n=7-11). (В) Сравнительный анализ экспрессии рецепторов Fas и ФНО в Т-клетках позитивной линии дифференцировки (CD3), В-лимфоцитах (CD19) и миелоидных клетках (CD33) (n=7-11). (С) Экспрессия Fas существенно снижается на протяжении 16 часов культивирования во всех субпопуляциях (n=5-9). (D) Повышение экспрессии TNF-R2 на протяжении культивирования наиболее выражено в В-лимфоцитах (CD19) и миелоидных клетках (CD33) (n=7-11). (Е) Отогретые образцы мПК инкубировали в течение 4 и 16 часов при наличии и отсутствии FasL и ФНО-α (n=5-11). Апоптоз определяли по включению аннексина-V. (F) Сравнительные показатели апоптоза гейтированных CD34+ клеток-предшественников и CD3+ Т-клеток после инкубации отогретой мПК в течение 4 и 16 часов при наличии и отсутствии лигандов смерти.
Фигура 7. Фигуры 7A-7D представляют собой графики, на которых показано, что лиганды рецепторов смерти предотвращают развитие БТПХ при трансплантациях криоконсервированных клеток после мобилизации периферической крови (мПК). (А) Выживаемость мышей NOD.SCID, прошедших кондиционирование с помощью двух доз 25 мкг/г, и которым трансплантировали через 2 суток 1,5×107 клеток мПК, инкубированных в среде с 50 нг/мл FasL и 20 нг/мл ФНОα в течение 4 часов (n=10 в каждой группе). (В) Уровень ксеногенного химеризма по линии человеческих клеток измеряли в костном мозге через 12 недель после трансплантации клеток, полученных из одного и того же образца мПК при наличии (n=10) и отсутствии (n=7) предварительной инкубации с FasL. (С) Масса тела через три недели после трансплантации у реципиентов отогретых клеток (n=18) и после инкубации в течение 4 часов в среде (n=12) при наличии и отсутствии FasL (n=9) и ФНОα (n=10). В случае экспериментальной точки 12 недель у мышей, которым трансплантировали клетки, инкубированные в среде и с FasL, оценивали: (D) клинические показатели в соответствии с нормой (0) и патологией (1): 1. болезнь кожи и выпадение волос, 2. слабость, 3. гиперкератоз подушечек лап и 4. диарея, и (Е) гистология печени, оцененная по показателям: 0 - отсутствие инфильтрации, 1 - редкая инфильтрация, 2 - очаговая инфильтрация, 3 - диффузная инфильтрация, 4 - разрушение структуры ткани.
Фигура 8. Фигуры 8В представляет собой схему и фигуры 8А, 8С, 8D и 8Е представляют собой графики, на которых показано, что воздействие лигандов рецепторов смерти не ухудшает антигенную стимуляцию и реактивность «трансплантат против опухоли». (А) Криоконсервированные клетки мПК отогревали и инкубировали в течение 4 или 16 часов в присутствии 50 нг/г FasL и 20 нг/г ФНО-α. Относительное распределение CD34+ клеток-предшественников, Т-клеток (CD3+), В-лимфоцитов (CD19+) и миелоидных клеток (CD33+) в пределах фракции жизнеспособных клеток показывает различные уровни чувствительности этих субпопуляций к апоптозу. (В) Образцы мПК инкубировали в течение 4 или 16 часов с лигандами смерти, мертвые клетки удаляли путем центрифугирования в фиколле, и равные количества жизнеспособных клеток высевали на чашки с полутвердой средой на основе метилцеллюлозы. (С) Клоногенную активность, которую выражали в виде количества колониеобразующих клеток (КОЕ) мПК после инкубации в среде и с 50 нг/мл FasL или 20 нг/мл ФНО-α (n=7-11 в каждой группе). (D) МПК, инкубированную в течение 4 часов с 50 нг/г FasL, совместно инкубировали с облученными аллогенными клетками-стимуляторами мПК в анализах с использованием реакции смешанной культуры лимфоцитов (MLR). Пролиферацию клеток-респондеров определяли по разведению CFSE и выражали в количественной форме с применением программного обеспечения ModFit. (Е) Мышам NOD.SCID инокулировали подкожно клетки человеческой карциномы толстой кишки НТ29 и проводили внутривенную инфузию 3×107 клеток мПК, предварительно инкубированных в среде и с FasL в течение 4 часов. Показатели опухолевого роста, которые измеряли с помощью штангенциркуля в соответствии с (мм3 = длина × ширина2 × 0,4), уменьшались за счет инфузии клеток мПК независимо от воздействия FasL.
Фигура 9. Фигуры 9А и 9Е представляют собой схемы и фигуры 9B-9D и 9F-9H представляют собой графики, на которых показано, что дотрансплантационная деплеция клеток, чувствительных к апоптозу, предотвращает БТПХ. (A-D) FasL-опосредованное удаление предварительно сенсибилизированных эффекторов БТПХ. (А) Мышей H2Kb иммунизировали дважды с использованием 107 спленоцитов H2Kd с интервалами в 3 суток. Через трое суток после второй иммунизации спленоциты (imH2Kb) культивировали в среде и с 50 нг/мл FasL в течение 24 часов. (В) Спленоциты, собранные у иммунизированных H2Kd мышей H2Kb, инкубировали в течение 24 часов в среде (n=6) и с 50 нг/мл FasL (n=5). Апоптоз и гибель измеряли по поглощению аннексина-V и 7-AAD, соответственно, в гейтированных CD4+ и CD8+ субпопуляциях Т-клеток. (С) У спленоцитов иммунизированных мышей отмечался повышенный ответ на облученные (3000 рад) аллогенные стимуляторы (H2Kd) по сравнению с посторонними стимуляторами (H2Kk). Аллоиммунные ответы в значительной степени подавлялись за счет инкубации с FasL в течение 24 часов, при этом сохранялась отвечаемость на посторонние стимулы.
Индекс пролиферации определяли по разведению CFSE (n=5 отдельных инкубаций). (D) Жизнеспособные спленоциты (1,5×106), полученные от иммунизированных доноров, подвергали адоптивному переносу сублетально облученным (650 рад) реципиентам F1 (H2Kb→H2Kb/d) после инкубации ex vivo в среде и с FasL. Различия по выживаемости поляризовали посредством введения 10 мкг липополисахарида (ЛПС) на +7 сутки (n=8 в каждой группе). (Е-Н) FasL-опосредованная деплеция нестимулированных донорских спленоцитов. (Е) Сублетально-облученным (650 рад) реципиентам F1 (H2Kb/d) проводили инфузию семиаллогенных спленоцитов (H2Kb), предварительно инкубированных в течение 24 часов в среде и с FasL. (F) Нестимулиованные спленоциты инкубировали в течение 24 часов в среде (n-7) и с 50 нг/мл FasL (n-9) для измерения апоптоза в гейтированных CD4+ и CD8+ субпопуляций Т-клеток. (G) Нестимулиованные спленоциты (H2Kb), инкубированные в течение 24 часов при наличии и отсутствии FasL, отвечают на облученные аллогенные клетки-стимуляторы (H2Kd). (Н) Жизнеспособные спленоциты (1,5×106), полученные от нестимулиованных доноров, подвергали адоптивному переносу сублетально облученным (650 рад) реципиентам F1 (H2Kb→H2Kb/d) после инкубации ex vivo в среде (n=10) и с FasL (n=10). Мышей подвергали стимуляции 10 мкг ЛПС (внутривенно) на +7 сутки.
Фигура 10. Фигуры 10А-10Е представляют собой графики, на которых показано, что Fas-опосредованная деплеция нестимулированных спленоцитов уменьшает эффект гаплоидентичных и аллогенных иммунокомпетентных клеток, приводящий к развитию БТПХ. Сублетально-облученным (650 рад) реципиентам F1 (H2Kh/d) проводили инфузию различных количеств (1,5-4,5×106) семиаллогенных спленоцитов, предварительно инкубированных в течение 24 часов в среде и с FasL. (А) У реципиентов спленоцитов, инкубированных в присутствии и в отсутствие FasL, определяли клинические показатели в соответствии с нормой (0) и патологией (1): 1. болезнь кожи и выпадение волос, 2. слабость, 3. Гиперкератоз подушечек лап и 4. диарея. (В) Потеря веса у реципиентов различных количеств донорских спленоцитов, инкубированных в присутствии и в отсутствие FasL (n=5-10 в каждой группе). (C-D) Гаплоидентичные трансплантации 3×106 спленоцитов сравнивали с аллогенными трансплантациями (H2Kb→H2Kd) после инкубации при наличии и отсутствии FasL в течение 24 часов (n=6-10 в каждой группе). Клинические показатели (С) и потерю веса (D) измеряли через одну неделю. (Е) Содержания Т- и В-лимфоцитов в селезенке постепенно уменьшается на протяжении первой недели после облучения (n=5 в каждой группе). Клетки, выжившие после стимуляции липополисахаридом (ЛПС), демонстрируют отчетливо выраженную стимуляцию этих субпопуляций (через 2 суток после инфузии ЛПС), которая снижалась за счет предварительной обработки нестимулированных спленоцитов FasL (n-7).
Фигура 11. Фигура 11С представляет собой схему и фигуры 11А, 11B и 11D-F представляют собой графики, на которых показано, что образование поперечных связей в Fas улучшает состояние мышей NOD.SCID при БТПХ. (А) Спленоциты, инкубированные в среде и с 50 нг/мл FasL, демонстрируют различные уровни спонтанного (светлые столбцы) и Fas-опосредованного апоптоза (темные столбцы). Апоптоз измеряли по поглощению аннексина V на гейтированных субпопуляциях CD4+ и CD8+ Т-клеток через 24 и 48 часов после инкубации (n=7-9). (В) Распределение по фракциям жизнеспособных CD4+ и CD8+ Т-клеток, CD4+CD25+ регуляторных Т-клеток и В-лимфоцитов (В220+) до и через 48 часов после инкубации в среде и с FasL. (С) Проводили инфузию спленоцитов, полученных от C57BI76 доноров, инкубированных в среде и с 50 нг/мл FasL, аллогенным мышам NOD.SCID (H2Kb→H2Kg7): 3×106 и 107 наивных спленоцитов, предварительно инкубированных в течение 24 и 48 часов, соответственно. (D) Выживаемость мышей NOD.SCID, которым проводили инфузию спленоцитов, предварительно инкубированных в контрольной среде (n=9) и с 50 нг/мл FasL (n=10). Летальную БТПХ вызывали путем внутривенной инъекции 10 мкг ЛПС. Через 3 недели у мышей оценивали клинические показатели с частотой вовлеченности желудочно-кишечного тракта (GI) (Е) и потерей веса (F).
Фигура 12. Фигура 12А представляет собой схему, фигуры 12В-С и 12E представляют собой графики, и фигура 12D представляет собой изображение и график, на которых показано удаление наивных иммунокомпетентных клеток, чувствительных к Fas при гаплоидентичных трансплантациях костного мозга (А) Реципиентам F1 (H2Kb/d) облученным при 850 рад, трансплантировали 5×106 ВМС (H2Kb, CD45.1) и различные количества донорских спленоцитов (H2Kb, CD45.2, GFP). У реципиентов спленоцитов, инкубированных в среде и с FasL в течение 48 часов (n=8 в каждой группе), оценивали клинические показатели (В) с частотой вовлеченности желудочно-кишечного тракта (GI) и потерей веса (С) через 3 недели после трансплантации. (D) Через 3 недели после трансплантации мышей выводили из эксперимента для гистологической оценки кожи и печени у реципиентов 3×106 спленоцитов, инкубированных в среде и с FasL (n=5): 0 - отсутствие инфильтрации, 1 - редкая инфильтрация, 2 - очаговая инфильтрация, 3 - диффузная инфильтрация, 4 - разрушение структуры ткани.
Фигура 13. Фигура 13А представляет собой график и FACS-сканограмму, фигуры 13В и 13D представляют собой FACS-сканограммы, и фигура 13С представляет собой график, на котором показано восстановление иммунной системы после селективной деплеции наивных иммунокомпетентных клеток, чувствительных к Fas, при гаплоидентичных трансплантациях костного мозга (А) Реципиентам F1 (H2Kb/d), облученным при 850 рад, трансплантировали 5×106 ВМС (
Фигура 14. Фигуры 14А, 14С и 14Е представляют собой схемы и фигуры 14В, 14D и 14F представляют собой графики, на которых показано, что FasL-опосредованная деплеция нестимулированных лимфоцитов поддерживает приживление гемопоэтических клеток. (А) Клетки цельного костного мозга (wBMC) инкубировали в течение 24 часов при наличии и отсутствии 50 нг/мл FasL, и 2×106 клеток трансплантировали сублетально облученным (800 рад) аллогенным хозяевам (H2Kd→H2Kb). (В) На химеризм периферической крови через 3 недели не влияла предварительная инкубация с FasL (n=6). (С) Трансплантация 106 ВМС негативной линии дифференцировки (n=7) облученным (800 рад) аллогенным хозяевам (H2Kd→H2Kb) была дополнена спленоцитами в количестве 106, полученными от доноров F1 (H2Kb/d) в отсутствие эффекта БТПХ после предварительной инкубации в среде (n=5) и с FasL (n=6) в течение 24 часов. (D) Добавление спленоцитов улучшало приживление гемопоэтических клеток независимо от воздействия FasL. (Е) Смешанный химеризм индуцировали с помощью трансплантации 5×105 lin-ВМС аллогенным реципиентам (H2Kd→H2Kb), облученным при 750 рад (контроль, n=6). Через 10 суток мышам проводили инфузию 106 аллогенных (H2Kd) лимфоцитов (DLI). (F) Уровни донорского химеризма повышались через 3 недели после DLI, предварительно инкубированных в течение 24 часов в среде и с FasL (n=6), по сравнению с мышами, которым не проводили инфузию лимфоцитов (контроль).
Фигура 15. Фигуры 15А, 15С, 15Е и 15G представляют собой схемы и фигуры 15В, 15D, 15F и 15Н представляют собой графики, на которых показано, что трансплантация нестимулиованных лимфоцитов после деплеции с помощью FasL обеспечивает сохранение эффекта «трансплантат против хозяина». (А) Мышам NOD.SCID (H2Kg7), имеющим подкожные аллогенные (H2Kd) опухоли, вызванные клетками карциномы толстой кишки СТ26, проводили инфузию (через 1 день после имплантации опухоли) 1,5×107 спленоцитов, полученных от совместимых с хозяином доноров NOD возрастом 5 недель (H2Kg7). (В) Инфузия лимфоцитов подавляла опухолевый рост у мышей NOD.SCID (n=10) независимо от предварительной инкубации в течение 24 часов в среде и с FasL (n-8). (С) Мышей H2Ka, имеющих совместимые по ГКГС подкожные нейробластомы (Neuro-2a, H2Ka), подвергали сублетальному облучению (750 рад) и трансплантации 2×106 клеток костного мозга негативной линии дифференцировки, полученных от аллогенных доноров (H2Kb). (D) Инфузия лимфоцитов, полученных от доноров F1, (H2Kb/d), в отсутствие действия РТПХ не оказывала значительного влияния на опухолевый рост (n=17). Инфузия аллогенных спленоцитов (H2Kb) инкубированных в среде в течение 24 часов, снижала показатели опухолевого роста (n=10), но 80% мышей погибло за 3 недели вследствие тяжелой формы БТПХ. Инфузия спленоцитов, предварительно инкубированных с FasL, в равной степени снижала опухолевый рост, но все мыши остались живыми (n=11). (Е) Сублетально облученным (750 рад) мышам BALB/c проводили инфузию смеси 5×106 изогенных (H2Kd) клеток костного мозга и 2×105 клеток лимфобластомы А20 (H2Kd). Смеси клеток предварительно инкубировали в течение 24 часов в среде и с FasL. (F) У реципиентов костного мозга и клеток А20, инкубированных в среде, развивались диссеминированные опухоли с летальным исходом (n=10), тогда как реципиенты клеток, предварительно инкубированных с FasL, выжили (n=10). 10 (G) Мышам NOD.SCID (H2Kg7) с ослабленным иммунитетом, облученным при 650 рад, проводили адоптивный перенос 5×107 клеток костного мозга, полученных от изогенных доноров NOD (H2Kg7), через 48 часов после инкубации в среде и с 50 нг/мл белка FasL. Гипергликемию принимали во внимание, когда после двух измерений уровни глюкозы в крови натощак превышали 200 мг/дл. (Н) Развитие гипергликемии после адоптивного переноса клеток цельного костного мозга, предварительно инкубированных в среде и с белком FasL (n=10).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В одном варианте реализации настоящее изобретение раскрывает устройство для отбора клетки, устойчивой к апоптотическим сигналам. В другом варианте реализации клетку, устойчивую к апоптотическим сигналам, выбирают из группы, состоящей из: стволовой клетки, клетки-предшественника и иммунокомпетентной клетки. В другом варианте реализации иммунокомпетентные клетки настоящего изобретения представляют собой субпопуляцию Т-клеток. В другом варианте реализации клетка настоящего изобретения представляет собой гемопоэтическую клетку. В другом варианте реализации клетку настоящего изобретения идентифицируют на основе поверхностного фенотипа, например, CD34+. В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, представляет собой клетку настоящего изобретения.
В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, устойчива к ФНО-альфа. В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, устойчива к Fas-лиганду. В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, устойчива к TRAIL. В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, устойчива к Tweak. В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, устойчива к ФНО-альфа, Fas-лиганду, TRAIL, Tweak или любой их комбинации.
В другом варианте реализации в настоящем изобретении предложены способы и устройства, которые позволяют преодолеть трудности и неточности, ассоциированные со способами окрашивания клеток для идентификации стволовой клетки или "стволовитости". В известном уровне техники обычно предлагаются способы идентификации стволовой клетки, тогда как в другом варианте реализации в настоящем изобретении предложены способы отбора стволовой клетки из культуры клеток, причем культура клеток содержит различные типы клеток, включая нестволовые клетки. В другом варианте реализации отбор осуществляют в один этап.
До подробного толкования по меньшей мере одного варианта реализации настоящего изобретения, следует понять, что применение настоящего изобретения не ограничивается деталями, которые изложены в следующем описании или приведены в примерах. Настоящее изобретение допускает либо практическое воплощение, либо выполнение других вариантов реализации различными способами. Также, следует понять, что формулировка и терминология, использованные в данном документе, используются в целях описания и не носят ограничительного характера.
В другом варианте реализации устройство настоящего изобретения содержит контейнер с апоптоз-индуцирующим лигандом, который иммобилизован на внутренней поверхности контейнера. В другом варианте реализации устройство настоящего изобретения содержит контейнер с апоптоз-индуцирующим лигандом, который иммобилизован на поверхности микрогранул, которые в него ввели. В другом варианте реализации устройство настоящего изобретения содержит контейнер с апоптоз-индуцирующим лигандом, который иммобилизован на поверхности микрогранул, которые содержатся внутри устройства. В другом варианте реализации устройство настоящего изобретения используют для отбора стволовых клеток с применением одного этапа. В другом варианте реализации один этап включает инкубацию гетерогенной клеточной популяции внутри контейнера. В другом варианте реализации посредством инкубации клеточная популяция подвергается воздействию апоптических лигандов в соответствии с данным документом. В другом варианте реализации инкубация приводит к выживанию клеток, устойчивых к апоптозу, таких как стволовые клетки, и апоптотической гибели типов клеток, чувствительных к апоптозу.
В другом варианте реализации отобранную популяцию клеток, устойчивых к апоптозу (таких как стволовые клетки), используют для трансплантации в ходе лечения болезни, как, например, но без ограничений: рак, иммунопатологические болезни, рак, устойчивый к химиотерапии, или врожденный или приобретенный иммунодефицит. В другом варианте реализации специалист в данной области техники может без труда найти применение клеткам настоящего изобретения, устойчивым к апоптозу.
В другом варианте реализации контейнер сконструирован из по меньшей мере одного биосовместимого материала. В другом варианте реализации биосовместимый материал включает, но без ограничений: полипропилен, полистирол, силикон, поливинилхлорид или их комбинацию. В другом варианте реализации биосовместимый материал выбирают в соответствии параметрами, как, например, но без ограничений: прочность или способность обеспечивать иммобилизацию апоптоз-индуцирующих лигандов на внутренней поверхности или прозрачность. В другом варианте реализации биосовместимый материал является инертным в отношении оказания любого нежелательного местного или системного действия на реципиента или человека, получающего лечение, но вызывает наиболее подходящий благоприятный клеточный или тканевой ответ в этой конкретной ситуации и оптимизирует клинически значимый эффект такого лечения (биосовместимые материалы приведены в качестве примера в патенте США 5998024, патенте США 6526984 и патенте США 4979959). В другом варианте реализации биосовместимый материал включает гидрофобный полимер, покрытый по меньшей мере одним слоем, который обеспечивает выживаемость клеток настоящего изобретения, устойчивых к апоптозу.
В другом варианте реализации контейнер представляет собой пакет. В другом варианте реализации контейнер представляет собой колонку. В другом варианте реализации контейнер представляет собой трубку. В другом варианте реализации контейнер представляет собой бутылку. В другом варианте реализации контейнер представляет собой пузырек. В другом варианте реализации контейнер представляет собой флакон. В другом варианте реализации контейнер представляет собой любую другую емкость, которая, известна в данной области техники и подходит для конкретного определенного использования устройства.
В соответствии с одним вариантом реализации устройство состоит из одного контейнера, в который вводят клеточную популяцию. В другом варианте реализации контейнер содержит две взаимосвязанные камеры, разделенные с помощью фильтровального прибора, приспособленного для отделения цельных клеток от клеточных остатков и белков. В другом варианте реализации клеточную популяция вводят в одну камеру, инкубируют внутри камеры, а после инкубации можно использовать фильтр, чтобы дополнительно отделить отобранные клетки от клеточных остатков и белков в растворе.
В соответствии с другим вариантом реализации представлен контейнер в форме колонки, в которой инкубируют клеточную популяцию, что обеспечивает прикрепление цельных клеток, или конкретно стволовых клеток, к поверхности колонки. В другом варианте реализации после инкубации удаляют оставшиеся клеточные остатки и белки, а отобранные клетки извлекают из колонки. Применение такой колонки в качестве примера приведено патенте США 5098842 и в заявке на патент США 2011/0256581.
В другом варианте реализации для отбора клетки, устойчивой к апоптозу, в качестве апоптоз-индуцирующих средств используют представителей семейства фактора некроза опухоли (ФНО). В другом варианте реализации апоптоз-индуцирующий лиганд, принадлежащий к семейству ФНО, который используют в соответствии с настоящим изобретением, представляет собой: ФНО-α, FasL, Trail (лиганд Аро2) или Tweak (лиганд Аро3). В другом варианте реализации проапоптотическое средство представляет собой рекомбинантный белок.
В другом варианте реализации биологически активный апоптоз-индуцирующий лиганд представляет собой лиганд, который обладает апоптоз-индуцирующей активностью при иммобилизации на внутренней поверхности контейнера. В другом варианте реализации биологически активный апоптоз-индуцирующий лиганд является любым из: ФНО-α, FasL, Trail и Tweak, которые содержат по меньшей мере активный участок, способный связываться с соответствующими рецепторами, индуцирующими апоптоз, в иммобилизованной форме.
В другом варианте реализации источником апоптоз-индуцирующий лиганда является млекопитающее. В другом варианте реализации апоптоз-индуцирующий лиганд представляет собой человеческий апоптоз-индуцирующий лиганд. В другом варианте реализации апоптоз-индуцирующий лиганд представляет собой апоптоз-индуцирующий лиганд грызуна.
В соответствии с другим вариантом реализации биологически активные апоптоз-индуцирующие лиганды иммобилизованы на внутренней поверхности контейнера. В другом варианте реализации биологически активные апоптоз-индуцирующие лиганды иммобилизованы на микрогранулах внутри контейнера. В другом варианте реализации биологически активные апоптоз-индуцирующие лиганды иммобилизованы таким способом, который обеспечивает свободное взаимодействие лигандов с клетками, которые содержатся внутри контейнера.
Существует много способов иммобилизации белка на поверхности, известных в данной области техники. Большинство способов иммобилизации включают модификацию или покрытие поверхности соответствующими веществами для изменения свойств поверхности или обеспечения наличия функциональных групп для связывания белка. С другой стороны, иммобилизация белков на непокрытую поверхность в отсутствие модификации требует применения аффинного пептида, который обладает специфичностью к конкретной поверхности. В другом варианте реализации иммобилизация достигается с использованием хелатора, совместимого с апоптоз-индуцирующим лигандом настоящего изобретения. В другом варианте реализации специалист в данной области техники может без труда идентифицировать надлежащий хелатор. В другом варианте реализации хелатор и является совместимым с апоптоз-индуцирующим лигандом настоящего изобретения, и поддерживает его биологическую активность. В другом варианте реализации хелатор и является совместимым с апоптоз-индуцирующим лигандом настоящего изобретения, и обеспечивает проявление его биологической активности.
В другом варианте реализации иммобилизация апоптоз-индуцирующего лиганда достигается за счет: физической адсорбции, взаимодействия между His-б и Na+ ионами, ассоциации белков с лейциновой молнией в гетеродимер по типу coiled coil, хемисорбции SH-групп, образования оснований Шиффа между альдегидными и аминогруппами, ацилтрансферазной реакции TGase, сродства между стрептавидин-биотином, сродства между FLAG и антителом к FLAG, сродства между глутатионом и GST или связывание белка, слитого с PS-аффинным пептидом с гидрофильным полистиролом (различные способы иммобилизации приведены в качестве примера в патенте США 6040182, патенте США 4885234, заявке на патент США 2010/0209945 и заявке на патент США 2006/0009623).
В другом варианте реализации апоптоз-индуцирующий лиганд представляет собой производное или аналог полноразмерного белка. В другом варианте реализации апоптоз-индуцирующий лиганд представляет собой небольшую органическую молекулу. В другом варианте реализации апоптоз-индуцирующий лиганд представляет собой вариант, производное, модифицированный вариант или процессированный вариант полноразмерного белка. В другом варианте реализации FasL представляет собой человеческий FasL, как, например, изложенный в SEQ ID NO: 1. В другом варианте реализации человеческий ФНО-α настоящего изобретения содержит SEQ ID NO: 2. В другом варианте реализации человеческий Trail настоящего изобретения содержит SEQ ID NO: 3. В другом варианте реализации человеческий Tweak настоящего изобретения содержит SEQ ID NO: 4.
Белки, раскрытые в данном документе можно получить с помощью технологии рекомбинантной ДНК или способов химического синтеза.
В другом варианте реализации в данном документе предложен способ отбора клеток, устойчивых к апоптозу, из гетерогенной клеточной популяции, отличающийся тем, что гетерогенная популяция состоит из клеток, устойчивых к апоптотическим сигналам, и клеток, чувствительных к апоптотическим сигналам. В другом варианте реализации способ заключается во введении гетерогенной клеточной популяции в устройство и инкубации в нем. В другом варианте реализации способ включает введение гетерогенной клеточной популяции в устройство и инкубацию в нем.
В другом варианте реализации термин "отбор", как используется в данном документе, относится к способу, при котором сохраняется только клетка, отобранная из гетерогенной клеточной популяции. В другом варианте реализации сохранившаяся клетка представляет собой клетку, устойчивую к апоптотическим сигналам.
В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, представляет собой стволовую клетку. В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, представляет собой иммунокомпетентную клетку, нечувствительную к гибели клеток, индуцированной активацией (AICD). В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, представляет собой клетку-предшественник. В другом варианте реализации клетка, устойчивая к апоптотическим сигналам, представляет собой стволовую клетку, иммунокомпетентную клетку, нечувствительную к гибели клеток, индуцированной активацией (AICD), клетку-предшественник или любую их комбинацию.
В другом варианте реализации стволовая клетка представляет собой стволовую клетку пуповинной крови. В другом варианте реализации стволовая клетка представляет собой мобилизованную стволовую клетку периферической крови. В другом варианте реализации стволовая клетка представляет собой стволовую клетку костного мозга. В другом варианте реализации стволовая клетка представляет собой раковую стволовую клетку. В другом варианте реализации стволовая клетка представляет собой нейральную стволовую клетку. В другом варианте реализации стволовая клетка представляет собой стволовую клетку пуповинной крови, стволовую клетку мобилизованной периферической крови, стволовую клетку костного мозга, раковую стволовую клетку, нейральную стволовую клетку или их комбинации.
В другом варианте реализации "иммунокомпетентная клетка, нечувствительная к гибели клеток, индуцированной активацией (AICD)", как используется в данном документе, относится к клетке иммунной системы, которая не подвергается апоптозу после активации. В другом варианте реализации иммунокомпетентная клетка, нечувствительная к гибели клеток, индуцированной активацией (AICD), представляет собой неактивированную Т-клетку.
Сбор стволовых клеток для терапевтических целей требует извлечения ткани, которая содержит стволовые клетки, либо аутологичные, либо от донора, и затем отделения стволовых клеток от других клеточных популяций, которые могут оказывать отрицательное воздействие при котрансплантации наряду со стволовыми клетками.
В другом варианте реализации словосочетание "устойчивая к апоптозу" означает "устойчивая к опосредованному рецепторами апоптозу". В другом варианте реализации способ отбора клеток настоящего изобретения представляет собой одноэтапный способ отрицательного отбора типов клеток, которые устойчивы к опосредованному рецепторами апоптозу. В другом варианте реализации настоящий способ обеспечивает параллельное выделение, как стволовых клеток, так и иммунокомпетентных клеток, которые поддерживают клинический исход трансплантации, посредством однократного применения устройства, что приведено ниже в качестве примера.
В соответствии с другим вариантом реализации клеточные популяции, которые можно использовать в способе настоящего изобретения, получают из, но без ограничений, костного мозга, пуповинной крови (ПК) и периферической крови после мобилизации клеток-предшественников (мПК). Однако специалисту в данной области техники понятно, что клеточную популяцию можно также получить из других тканей взрослого организма, которые содержат клетки, устойчивые к апоптозу. В другом варианте реализации клеточную популяцию получают из эмбриональной ткани.
В другом варианте реализации гетерогенную клеточную популяцию получают из органа или ткани. В другом варианте реализации гетерогенную клеточную популяцию получают из эмбриона, где эмбрион не является эмбрионом человека. В другом варианте реализации гетерогенную клеточную популяцию получают из ткани, содержащей эмбриональные клетки, стволовые клетки, иммунокомпетентные клетки или любую их комбинацию. В другом варианте реализации гетерогенную клеточную популяцию получают из костного мозга. В другом варианте реализации гетерогенную клеточную популяцию получают путем механического отделения от стромы костного мозга путем аспирации или путем афереза после мобилизации в периферическую кровь (мПК) посредством активации и дезинтеграции молекулярных анкеров. В другом варианте реализации гетерогенную клеточную популяцию получают из периферической крови.
В другом варианте реализации стволовую клетку выбирают из гетерогенной популяции клеток. В другом варианте реализации словосочетание "гетерогенная популяция клеток", как используется в данном документе, относится к смеси клеток различных типов, содержащей стволовую клетку согласно вышеприведенному определению и по меньшей мере одну клетку, чувствительную к апоптозу. В другом варианте реализации гетерогенную популяцию клеток получают из любого организма. В другом варианте реализации источником гетерогенной популяции клеток является млекопитающее. В другом варианте реализации источником гетерогенной популяции клеток является человек.
В другом варианте реализации гетерогенная популяция клеток содержит смесь из клетки позитивной линии дифференцировки и стволовой клетки. В другом варианте реализации "клетка позитивной линии дифференцировки", как используется в данном документе, относится к клетке, экспрессирующей маркеры зрелых клеток клеточной линии. В другом варианте реализации маркер зрелых клеток клеточной линии представляет собой белок кластера дифференцировки (CD).
В другом варианте реализации способ настоящего изобретения используют для осуществления деплеции клеточной линии.
В другом варианте реализации гетерогенная популяция клеток представляет собой ткань или ее часть. В другом варианте реализации гетерогенная популяция клеток представляет собой клеточный агрегат. В другом варианте реализации гетерогенная популяция клеток представляет собой суспензию отдельных клеток. В другом варианте реализации гетерогенная популяция клеток представляет собой первичную культуру. В другом варианте реализации гетерогенная популяция клеток представляет собой клеточный образец. В другом варианте реализации гетерогенная популяция клеток содержит популяцию, которая чувствительна к проапоптотическим средствам настоящего изобретения.
В соответствии с другим вариантом реализации инкубация внутри устройства длится от 2 часов до 72 часов. В другом варианте реализации инкубация внутри устройства длится от 2 часов до 4 часов. В другом варианте реализации инкубация внутри устройства длится от 4 часов до 10 часов. В другом варианте реализации инкубация внутри устройства длится от 10 часов до 24 часов. В другом варианте реализации инкубация внутри устройства длится от 12 часов до 36 часов. В другом варианте реализации инкубация внутри устройства длится от 24 часов до 48 часов. В другом варианте реализации инкубация внутри устройства длится от 36 часов до 72 часов. В другом варианте реализации инкубация внутри устройства длится от 48 часов до 72 часов.
Неожиданно авторы настоящего изобретения обнаружили, что разные времена инкубации с разными комбинациями апоптоз-индуцирующих лигандов могут привести к функционально различимым клиническим исходам. Таким образом, время инкубации зависит от конкретного применения, которое в данном документе приведено ниже в качестве примера.
В соответствии с другим вариантом реализации настоящее изобретение раскрывает набор для отбора клеток. В другом варианте реализации набор содержит устройство настоящего изобретения, которое дополнительно содержит раствор для поддержания сохранности и активности апоптоз-индуцирующего лиганда. В другом варианте реализации набор дополнительно содержит вкладыш с инструкциями по осуществлению отбора клеток в соответствии со способами настоящего изобретения. В другом варианте реализации раствор для поддержания сохранности и активности апоптоз-индуцирующего лиганда представляет собой раствор. В другом варианте реализации указанный раствор представляет собой буфер или среду, которая обеспечивает активность апоптоз-индуцирующих лигандов с поддержанием их сохранности и структуры. В другом варианте реализации компоненты раствора варьируют в зависимости от используемых апоптоз-индуцирующих лигандов. В другом варианте реализации раствор содержит ингибитор протеазы. В другом варианте реализации ингибитор протеазы выбирают из, но без ограничений: фенилметилсульфонилфторида, бензамидина, пепстатина А, леупептина, апротинина, антипаина, ЭДТА, ЭГТА или любой их комбинации. В другом варианте реализации раствор содержит буферную систему, выбранную из, но без ограничений, Трис-буфера или глицин-NaOH (разные условия, которые влияют на выбор буфера для поддержания стабильности и активности белков, приведены в качестве примера в Uguw S.O. and Apte S.P., Pharmaceutical Technology: 2004; March: 86-113).
В другом варианте реализации раствор содержит элементы, которые способствуют и/или обеспечивают выживаемость клеток, клеточный рост, пролиферацию клеток или любую их комбинацию. В другом варианте реализации элементы, которые способствуют и/или обеспечивают выживаемость клеток, клеточный рост, пролиферацию клеток или любую их комбинацию, включают: ростовые среды, сыворотку или противобактериальные средства. В соответствии с данным вариантом реализации раствор способствует поддержанию жизнеспособности клеток внутри устройства набора.
В другом варианте реализации раствор содержит факторы, которые способствуют пролиферации стволовых клеток. В другом варианте реализации факторы, которые способствуют пролиферации стволовых клеток, выбирают из, но без ограничений, ростовых факторов, гормонов, ферментов или химических веществ.
В другом варианте реализации набор дополнительно содержит дополнительный апоптоз-индуцирующий лиганд, который добавляют в устройство или в раствор, соответственно обеспечивая селективность раствора по отношению к устойчивым к апоптозу клеткам настоящего изобретения.
В другом варианте реализации набор настоящего изобретения обеспечивает свободный выбор вида или концентрации апоптоз-индуцирующих лигандов, которые добавляют к популяции клеток. В соответствии с данным вариантом реализации набор содержит дополнительные контейнеры, причем каждый содержит разный апоптоз-индуцирующий лиганд. Пользователь набора выбирает предпочтительную комбинацию апоптоз-индуцирующих лигандов в соответствии с необходимостью, как приводится ниже в данном документе в качестве примера, вводит лиганды в устройство и обеспечивает инкубацию с лигандами.
В другом варианте реализации набор настоящего изобретения предполагает временную гибкость в плане добавления апоптоз-индуцирующих лигандов. Как приводится ниже в данном документе в качестве примера, разные комбинации, последовательность введения или времена инкубации апоптоз-индуцирующих лигандов приводят к разным клиническим исходам. Пользователь набора может выбрать необходимый порядок добавления апоптоз-индуцирующих лигандов и период инкубации с каждым лигандом внутри устройства, вследствие чего достигается предпочтительный клинический исход.
В соответствии с другим вариантом реализации настоящее изобретение дополнительно раскрывает способ улучшения клинического исхода трансплантации гемопоэтических стволовых и клеток-предшественников (HSPC). В другом варианте реализации получают клеточную популяцию, содержащую HSPC. В другом варианте реализации обеспечивают контакт клеточной популяции с апоптоз-индуцирующим лигандом внутри устройства настоящего изобретения и инкубируют внутри устройства. В другом варианте реализации выделяют выжившие клетки. В другом варианте реализации выделенные клетки трансплантируют субъекту.
В другом варианте реализации в устройство набора настоящего изобретения вводят аллогенную пуповинную кровь. В соответствии с данным вариантом реализации FasL иммобилизован на внутренней поверхности контейнера. Кровь инкубируют в устройстве в течение менее чем 24 часов. По окончанию инкубации в устройство вводят ФНО-α и продолжают инкубацию в течение 24-48 часов. После второй инкубации клетки извлекают из устройства и трансплантируют пациенту. В другом варианте реализации между окончанием инкубации и трансплантацией используют этап отделения, для того чтобы отделить живые клетки от клеточных остатков и белков в растворе.
В другом варианте реализации клеточную популяцию согласно настоящему способу получают из аутологичного трансплантата. В другом варианте реализации клеточную популяцию согласно настоящему способу получают из аллогенного трансплантата.
В соответствии с другим вариантом реализации настоящее изобретение дополнительно раскрывает способ удаления злокачественной клетки в композиции. В данном способе обеспечивают контакт композиции и инкубируют с апоптоз-индуцирующим лигандом. В другом варианте реализации композиция содержит трансплантат, состоящий из клеток-предшественников.
В другом варианте реализации апоптоз-индуцирующий лиганд представляет собой FasL. В другом варианте реализации инкубацию проводят в течение около 24 часов. Авторы настоящего изобретения показали in-vivo, что добавление FasL к трансплантату, содержащему злокачественную клетку, на время около 24 часов, неожиданно приводит к удалению злокачественного компонента и повышению коэффициента выживаемости (Фигура 15E-F).
В соответствии с другим вариантом реализации этот метод удаления потенциально обеспечивает удаление злокачественных клеток в трансплантатах, полученных от клинически здоровых доноров, которые страдают скрытой формой злокачественной болезни.
В соответствии с другим вариантом реализации трансплантат извлекают из костного мозга пациента, страдающего раком, и вводят в устройство настоящего изобретения. В соответствии с данным вариантом реализации трансплантат инкубируют с иммобилизованным FasL в течение около 24 часов. По завершении инкубации отобранные клетки трансплантируют обратно пациенту, страдающему раком. В другом варианте реализации трансплантации предшествует этап отделения, с тем, чтобы отделить отобранные клетки от клеточных остатков и белков.
В соответствии с другим вариантом реализации настоящее изобретение дополнительно раскрывает способ предотвращения болезни «трансплантат против хозяина» (БТПХ) с сохранением эффекта «трансплантат против опухоли» (ТПО). В соответствии с данным способом получают образец. В одном варианте реализации образец содержит клеточную популяцию. В другом варианте реализации клеточная популяция содержит HSPC. В другом варианте реализации клеточная популяция содержит иммунокомпетентную клетку. В другом варианте реализации клеточная популяция содержит HSPC, иммунокомпетентную клетку или их комбинацию. В соответствии с одним вариантом реализации обеспечивают контакт клеточной популяции с апоптоз-индуцирующим лигандом. В другом варианте реализации выжившую клетку извлекают из устройства. В другом варианте реализации извлеченную клетку трансплантируют субъекту.
В соответствии с другим вариантом реализации способ настоящего изобретения обеспечивает отбор только субпопуляций Т-клеток, которые не индуцируют БТПХ. Авторы настоящего изобретения неожиданно обнаружили, что краткая инкубация с FasL в течение 2-16 часов без сопутствующей сенсибилизации Т-клеток, приводит к эффективному удалению эффекторов БТПХ (Фигура 9). В соответствии с данным вариантом реализации достигается выживаемость субпопуляций Т-клеток, которые поддерживают приживление гемопоэтических клеток-предшественников, поскольку обработка трансплантируемых клеток с помощью FasL не нарушает приживление (Фигура 14).
В другом варианте реализации селективное удаление Т-клеток, чувствительных к апоптозу, из донорского инокулята не нарушает возникновение сильной реакции «трансплантат против опухоли» (ТПО).
В соответствии с другим вариантом реализации способ настоящего изобретения можно применять на трансплантате, состоящем из аллогенных мобилизованных клеток периферической крови, недостатком которого, как известно в данной области техники, является развитие тяжелой формы БТПХ. В данном варианте реализации трансплантат, состоящий аллогенных мобилизованных клеток периферической крови, вводят в устройство настоящего изобретения. Трансплантат затем инкубируют с иммобилизованным FasL в течение краткого периода из 2-16 часов. После инкубации оставшиеся клетки выделяют и трансплантируют пациенту. В соответствии с другим вариантом реализации трансплантации может предшествовать этап отделения, с тем, чтобы отделить отобранные клетки от клеточных остатков и белков.
В соответствии с другим вариантом реализации способ настоящего изобретения можно использовать для селективной деплеции нестимулированных Т-клеток до инфузии донорских лимфоцитов (DLI).
Рекомбинантная экспрессия в соответствии с некоторыми вариантами реализации
В другом варианте реализации белок настоящего изобретения можно синтезировать путем экспрессии полинуклеотидной молекулы, кодирующей белок, в клетке хозяина, например, клетке микроорганизма, трансформированной молекулой нуклеиновой кислоты.
Последовательности ДНК, кодирующие белки, можно выделить из любых клеток, в которых они образуются, с применением различных способов, широко известных в данной области техники (см., например, Sambrook, et al., Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor, N.Y., (2001)). Например, ДНК, кодирующую белок дикого типа, можно амплифицировать исходя из геномной ДНК, плазмиды или космиды соответствующего микроорганизма с помощью полимеразной цепной реакции (ПЦР) с применением специфических праймеров, сконструированных на основе нуклеотидной последовательности с известной последовательностью. Подходящие методы широко известны в данной области техники, описаны, например, в патентах США No. 4683195; 4683202; 25 4800159 и 4965188.
ДНК можно экстрагировать из клетки до амплификации с применением различных способов, известных в данной области техники, см., например, Marek P.М et al., "Cloning and expression in Escherichia coli of Clostridium thermocellum DNA encoding p-glucosidase activity", Enzyme and Microbial Technology Volume 9, Issue 8, August 1987, Pages 474-478.
Выделенный полинуклеотид, кодирующий белок, можно клонировать в вектор, такой как плазмида рЕТ28а.
После выделения и клонирования полинуклеотида, кодирующего белок, можно ввести мутацию(и) путем модификации одной или нескольких пар оснований, с применением способов, известных в данной области техники, таких как, например, сайт-специфический мутагенез (см., например, Kunkel Proc. Natl. Acad. Sci. USA 1985, 82: 488-492; Weiner et al., Gene 1994, 151: 119-123; Ishii et al., Methods Enzymol. 1998, 293: 53-71); кассетный мутагенез (см., например, Kegler-Ebo et al., Nucleic Acids Res. 1994 May 11; 22(9): 1593-1599); рекурсивный множественный мутагенез (см., например, Delagrave et al., Protein Engineering 1993, 6(3): 327-331), и сайт-насыщающий мутагенез (см., например, заявку на патент США No. 2009/0130718).
Также широко известны способы введения многочисленных мутаций в полинуклеотид (см., например, Michaelian et al., Nucleic Acids Res. 1992, 20:376; Dwivedi et al., Anal. Biochem. 1994, 221:425-428; Bhat Methods Mol. Biol. 1996, 57:269-277; Meetei et al., Anal. Biochem. 1998, 264:288-291; Kim et al., Biotechniques 2000, 28: 196-198; и международные публикации заявок на патенты No. WO 03/002761 A1 и WO 99/25871).
Альтернативным способом получения полинуклеотида с необходимой последовательностью является применение синтетического гена. Полинуклеотид, кодирующий белок настоящего изобретения, можно получить синтетическим методом, например, с применением фосфорамидитного способа (см. Beaucage et al., Curr Protoc Nucleic Acid Chem. 2001 May; Chapter 3: Unit 3.3; Caruthers et al., Methods Enzymol. 1987, 20 154: 287-313).
Применение синтетических генов позволяет получить искусственный ген, который содержит оптимизированную последовательность нуклеотидов, которую необходимо экспрессировать у необходимого вида (например, Е. coli). Переконструирование гена является средством улучшения экспрессии генов во многих случаях. Представляется возможным переделать открытую рамку считывания вследствие избыточности генетического кода. Таким образом, можно изменить приблизительно до трети нуклеотидов в открытой рамке считывания и все еще получить тот же самый белок. Например, для типичной белковой последовательности из 300 аминокислот существует более 10150 комбинаций кодонов, которые будут кодировать идентичный белок. Применение способов оптимизации, например, замена редко используемых кодонов чаще используемыми кодонами может привести к сильному влиянию на уровни экспрессии белка, кодируемого геном-мишенью. Также можно включить дополнительные оптимизации, такие как удаление вторичных структур РНК. Для осуществления этих и других одновременных оптимизаций доступны компьютерные программы. Вследствие большого количества изменений нуклеотидов, сделанных в исходной последовательности ДНК, единственный практический путь создания по-новому сконструированных генов заключается в применении генного синтеза.
Полинуклеотид, полученный таким образом, затем можно подвергать дополнительным манипуляциям, включая одну или несколько из очистки, отжига, лигирования, амплификации, расщепления рестрикционными эндонуклеазами и клонирования в подходящие векторы. Полинуклеотид первоначально можно лигировать либо в клонирующий вектор, либо непосредственно в экспрессирующий вектор, который подходит для экспрессии в клетке хозяина конкретного типа.
Специалисты в данной области техники без труда поймут, что кодон, используемый в полинуклеотиде для кодирования конкретной аминокислоты, которая должна заменить аминокислоту, которая первоначально присутствует в последовательности, кодирующей фермент дикого типа, необходимо выбрать в соответствии с известной и предпочтительной частотой использования кодона клеткой хозяина, которую выбрали для экспрессии полинуклеотида.
Специалисту в данной области техники понятно взаимоотношение между последовательностью нуклеиновой кислоты и белковой последовательностью, в частности, генетическим кодом и вырожденностью этого кода, и он сможет без труда сконструировать нуклеиновые кислоты, кодирующие белки настоящего изобретения. Например, специалисту в данной области техники понятно, что для каждой аминокислотной замены в белковой последовательности, имеется один или несколько кодонов, которые кодируют заменяемую аминокислоту. Соответственно, очевидно, что, в зависимости от вырожденности генетического кода в отношении конкретного аминокислотного остатка, можно составить одну или несколько последовательностей нуклеиновых кислот, соответствующих определенному варианту белковой последовательности.
Полинуклеотиды настоящего изобретения могут включать некодирующие последовательности, включая например, некодирующие 5'- и 3'-последовательности, такие как транскрибируемые, нетранслируемые последовательности, стоп-кодоны, сайты связывания рибосом, последовательности, которые стабилизируют мРНК, интроны и сигналы полиаденилирования. Дополнительно включены полинуклеотиды, которые содержат кодирующие последовательности для дополнительных аминокислот, гетерологичных для вариантного белка, в частности маркерную последовательность, такую как поли-His метка, которая способствует очистке белка в форме слитого белка.
Белки настоящего изобретения можно получить в виде меченых белков, например, для упрощения выделения и очистки. Неограничивающим примером конструкта, обеспечивающего мечение, является His-Tag (шесть идущих подряд гистидиновых остатков), который можно выделить и очистить с помощью традиционных способов. Также для удобства можно включить сайт протеолитического расщепления между участком метки и представляющей интерес белковой последовательностью для обеспечения удаления меток, как, например, сайт расщепления тромбином.
Полинуклеотид, кодирующий белок настоящего изобретения, можно встроить во множество экспрессирующих векторов, которые можно трансформировать во множество клеток хозяина. Клетка хозяина может быть прокариотической или эукариотической.
Введение полинуклеотида в клетку хозяина можно осуществить с помощью широко известных способов, таких как химическая трансформация (например, обработка хлоридом кальция), электропорация, конъюгирование, трансдукция, трансфекция с помощью фосфата кальция, трансфекция, опосредованная ДЭАЭ-декстраном, трансвекция, микроинъекция, трансфекция, опосредованная катионными липидами, введение при соскабливании, баллистическое введение и инфицирование.
В некоторых вариантах реализации клетка представляет собой прокариотическую клетку. Репрезентативные, неограничивающие примеры подходящих прокариотических хозяев включают бактериальные клетки, такие как клетки Escherictahia coli и Bacillus subtilis. В других вариантах реализации клетка представляет собой эукариотическую клетку. В некоторых иллюстративных вариантах реализации клетка представляет собой грибную клетку, как, например, дрожжи. Репрезентативные, неограничивающие примеры подходящих дрожжевых клеток включают Saccharomyces cerevisiae и Pichia pastoiis. В дополнительных иллюстративных вариантах реализации клетка представляет собой растительную клетку.
Белки можно экспрессировать в любых векторах, подходящих для экспрессии. Подходящий вектор определяют в зависимости от выбранной клетки хозяина. Векторы для экспрессии белков в Е. coli, например, включают, но без ограничений, рЕТ, pK233, рТ7 и lambda pSKF. Другие экспрессирующие векторные системы основаны на бета-галактозидазе (рЕХ); мальтоза-связывающем белке (pMAL); и глутатион-S-трансферазе (pGST).
Отбор клетки хозяина, трансформированной необходимым вектором, можно провести с применением стандартных протоколов для отбора, включающих выращивание на селективной среде, которая является токсичной для нетрансформированной клетки. Например, Е. coli можно выращивать на среде, содержащей селективный антибиотик; клетки, трансформированные экспрессирующим вектором, который дополнительно несет ген устойчивости к антибиотикам, выращивают на селективной среде.
После трансформации подходящей клетки хозяина и культивирования в условиях, подходящих для экспрессии белка, можно идентифицировать необходимый белок в клеточных экстрактах трансформированных клеток. Трансформированных хозяев, экспрессирующих представляющий интерес белок, можно идентифицировать путем анализа белков, экспрессированных хозяином, с применением SDS-PAGE и путем сравнения полученного геля с гелем, полученным после SDS-PAGE в случае хозяина, которого трансформировали тем же самым вектором, но в отсутствие последовательности нуклеиновой кислоты, кодирующей представляющий интерес белок.
Представляющий интерес белок также можно идентифицировать с помощью других известных способов, таких как иммуноблот-анализ с применением подходящих антител, дот-блоттинг общего количества клеточных экстрактов, ограниченный протеолиз, масс-спектрометрический анализ и их комбинации.
Представляющий интерес белок можно выделить и очистить с помощью традиционных способов, включая фракционирование сульфатом аммония и этиловым спиртом, экстрагирование кислотой, фракционирование солями, ионообменную хроматографию, гидрофобную хроматографию, гельпроникающую хроматографию, аффинную хроматографию и их комбинации.
С применением способов, известных в данной области техники, некоторые из которых описаны ниже в данном документе, можно проанализировать различные свойства выделенного представляющего интерес белка, например, специфическую активность и термостабильность.
Специалисты в данной области техники могут без труда определить условия для проведения вышеупомянутых процедур, а также другие пригодные способы (см., например, Current Protocols in Protein Science, 1995 John Wiley & Sons).
В отдельных вариантах реализации белки настоящего изобретения можно получить и/или использовать в отсутствие стартового кодона (метионин или валин) и/или в отсутствие лидерного (сигнального) пептида, чтобы облегчить получение и очистку рекомбинантных белков. Известно, что клонирование генов в отсутствие последовательностей, кодирующих лидерные пептиды, будет приводить к локализации белков в цитоплазме клетки хозяина и будет облегчать их выделение (см., например, Glick, В.R. и Pasternak, J.J. (1998) в "Molecular biotechnology: Principles and applications of recombinant DNA", 2nd edition, ASM Press, Washington D.C., стр. 109-143).
Получение белков с помощью синтеза в соответствии с некоторыми вариантами реализации
Белки настоящего изобретения можно синтезировать с помощью любых методов, которые известны специалистам в области белкового синтеза. В случае твердофазного белкового синтеза краткое описание многих методов можно найти в: Stewart, J.М. и Young, J.D. (1963), "Solid Phase Peptide Synthesis," W.H. Freeman Co. (San Francisco); и Meienhofer, J (1973). "Hormonal Proteins and Peptides," vol. 2, p. 46, Academic Press (New York). Для ознакомления с классическим синтезом в растворе см. Schroder, G. и Lupke, K. (1965). The Peptides, vol. 1, Academic Press (New York).
В целом, способы пептидного синтеза включают последовательное присоединение одной или нескольких аминокислот или аминокислот, защищенных подходящим образом, к растущей пептидной цепи. Обычно обеспечивают защиту либо аминной, либо карбоксильной группы первой аминокислоты с помощью подходящей защитной группы. Защищенная или дериватизированная аминокислота затем может либо прикрепляться к инертной твердой подложке, либо использоваться в растворе при присоединении следующей аминокислоты к последовательности, имеющей дополнительную (аминную или карбоксильную) группу, защищенную подходящим образом, в условиях подходящих для образования амидной связи. Защитную группу затем удаляют из этого вновь присоединенного аминокислотного остатка и затем присоединяют следующую аминокислоту (защищенную подходящим образом) и так далее; также традиционно этот процесс сопровождается этапами отмывки. После соединения всех необходимых аминокислот с образованием соответствующей последовательности, последовательно или одновременно удаляют любые оставшиеся защитные группы (и любую твердую подложку) для получения конечной молекулы пептида. За счет простой модификации этой общей методики представляется возможным за один раз присоединить несколько аминокислот к растущей цепи, например, путем связывания (в условиях, которые не способствуют рацемизации хиральных центров) защищенного трипептида с дипептидом, защищенным подходящим образом, с образованием пентапептида после снятия защиты и так далее.
Дополнительное описание пептидного синтеза раскрыто в патенте США No. 6472505.
Дополнительные цели, преимущества и новые признаки настоящего изобретения станут очевидными специалисту в данной области техники после изучения следующих примеров, которые не носят ограничительного характера. Дополнительно каждый из различных вариантов реализации и аспектов настоящего изобретения, которые описаны выше в данном документе и которые заявляются ниже в разделе формулы изобретения, находят экспериментальное подтверждение в следующих примерах.


Пример 1
Подвергание клеток пуповинной крови воздействию лигандов рецепторов смерти ex vivo
Апоптотическое действие активации рецепторов смерти in vitro
Основная функция суперсемейства ФНО заключается в транедукции апоптотических сигналов. Свежую пуповинную кровь (ПК), полученную при своевременных родах после информированного согласия доноров, подвергали воздействию FasL и ФНО-α в течение различных периодов времени в суспензионной культуре без добавления ростовых факторов. В случае гейтированных CD34+ клеток-предшественников в составе смешанных культур ПК отмечалось снижение показателей апоптоза с возрастающей чувствительностью в зависимости от времени (Фигура 1А), однако апоптотическая гибель клеток не усиливалась за счет подвергания воздействию лигандов рецепторов смерти семейства ФНО. Ранее предполагали, что нечувствительность к апоптотическим сигналам вызвана низким уровнем экспрессии рецепторов в гемопоэтических клетках-предшественниках. С помощью данного анализа выявили более низкий уровень экспрессии рецепторов Fas и ФНО в гейтированных CD34+ и выделенных lin- клетках-предшественниках, чем в общей популяции клеток ПК (Фигура 1В). Более точная оценка субпопуляций клеток, экспрессирующих рецепторы, показала повышенную чувствительность CD34+ клеток-предшественников к апоптозу, однако индукция апоптоза за счет родственных рецепторов не происходила за одним исключением (Фигура 1С). Кратковременное подвергание воздействию ФНО-α (<24 часов) было единственным случаем обнаруживаемого опосредованного рецепторами апоптоза CD34+ клеток, экспрессирующих как TNF-R1, так и TNF-R2. Эти данные предполагают, что экспрессия рецепторов смерти является характерной для клеток, которые особенно чувствительны к спонтанному апоптозу в суспензионной культуре. Различия в апоптозе могут возникать из-за разных скоростей пролиферации, однако повышенная чувствительность к апоптозу рецептороположительных CD34+ клеток-предшественников ассоциировалась со сниженными скоростями пролиферации, которые также не изменялись из-за присутствия родственных лигандов (Фигура 1D).
Чтобы определить, какие субпопуляции клеток ПК чувствительны к апоптозу, дополнительно оценивали клетки миелоидной и лимфоидной линий. Тогда как CD3+ Т-клетки экспрессируют высокие уровни Fas, в CD33+ миелоидных клетках преобладают рецепторы ФНО (Фигура IE). В целом TNF-R1 экспрессирован на более уровнях, чем TNF-R2 во всех субпопуляциях позитивной линии дифференцировки. С помощью апоптотической стимуляции лигандами смерти в течение 48 часов выявили повышение показателей спонтанного апоптоза всех субпопуляций позитивной линии дифференцировки по сравнению с lin- клетками-предшественниками, которые в значительной степени не изменялись из-за присутствия лигандов (Фигура 1F). Наблюдали два примечательных исключения: а) Fas-опосредованный апоптоз у CD14-CD33+ миелоидных клеток, соответствующих нейтрофилам ПК, которые экспрессируют высокие уровни рецепторов ФНО; и b) ФНО-индуцированный апоптоз у CD3+ Т-клеток. Устойчивость Т-клеток, полученных из ПК, к апоптозу в суспензионной культуре объясняется наивной природой этой не контактировавшей с антигеном субпопуляции, которая требует стимуляции для повышения чувствительности к отрицательной регуляции AICD-типа. Таким образом, апоптоз нестимулированных клеток ПК в культуре определяется гибелью CD19+ В-лимфоцитов и миелоидных клеток (CD14+, CD33+), опосредованной функцией рецепторов семейства ФНО в разных субпопуляциях, которая обеспечивает отбор с помощью апоптоза.
SCID-репопулирующие клетки и клетки, инициирующие долгосрочную культуру, устойчивы к опосредованному рецепторами апоптозу
Для оценки функции клеток-предшественников, которые постепенно подвергаются коммитированию, включая SCID-репопулирующие клетки (SRC), клетки, инициирующие долгосрочную культуру (LTC-IC), использовали несколько суррогатных анализов и кратковременные анализы клоногенных свойств. Действие SRC частично согласуется с восстановлением человеческих клеток и служит в качестве суррогатного функционального анализа для определения наиболее примитивных клеток-предшественников в составе образца. Изначально использовали равные количества клеток, инкубированных до трансплантации в присутствии и в отсутствие FasL и ФНО-α, потому что сильное колебание уровней ксеногенного химеризма делает невозможным точную оценку улучшения приживления (Фигура 2А). Функция SRC сохранялась после воздействие FasL и ФНО-α в течение 24-48 часов до трансплантации(Фигура 2B-D), что свидетельствовало о том, что наиболее примитивные гемопоэтические клетки-предшественники были нечувствительными к спонтанному и опосредованному рецепторами апоптозу в суспензионной культуре. Продленная инкубация ex vivo ассоциирована с потерей способности к приживлению человеческих гемопоэтических клеток-предшественников, независимо от присутствия FasL и ФНО-α (Фигура 2Е). Продолжительная инкубация клеток ПК ex vivo в жидкой среде, дополненной хемокинами и ростовыми факторами, ассоциирована с резкими изменениями фенотипа и, как известно, ухудшает хоминг клеток и приживление у мышей с ослабленным иммунитетом. Однако в отличие от прежних исследований, в которых недостаточное приживление SRC объяснялось рецептор-опосредованным апоптозом, вызванным лигандами смерти, пагубное влияние продленного культивирования в значительной степени не зависит от активации мембранных рецепторов смерти.
Недостаточное приживление человеческих клеток после длительных периодов культивирования ex vivo ассоциировано с пролиферацией и выходом из G0/G1 фазы клеточного цикла. Как показатели пролиферации (Фигура 2Е), так и фаза клеточного цикла (Фигура 2F) клеток ПК, клеток-предшественников CD34+ и lin- не изменялись из-за присутствия FasL и ФНО-а через 48 часов после инкубации. Сохранение фракций с высокой плотностью митотически покоящихся клеток-предшественников в присутствии лигандов смерти объясняет равные уровни приживления SRC, которое ограничено клетками, находящимися в G0/G1.
Суррогатные анализы ex vivo на активность гемопоэтических клеток-предшественников включают долгосрочное культивирование на мезенхимальных стромальных клетках (LTC-IC), которые представляют собой более примитивную субпопуляцию, чем колонии, формируемые коммитированными клетками-предшественниками на полутвердой среде на основе метилцеллюлозы. Подвергание воздействию FasL и ФНО-α в концентрациях, которые являются токсичными для соматических клеток, на протяжении всего 5-недельного периода долгосрочного культивирования не влияло на активность LTC-IC (Фигура 2G), что согласовалось с общей устойчивостью CD34+ и lin- клеток-предшественников к апоптотическим сигналам, опосредованным рецепторами Fas и ФНО. Таким образом, несмотря на заметное изменение состава жизнеспособных субпопуляций на протяжении подвергания воздействию лигандам смерти, вызванное разной присущей чувствительностью различных субпопуляций ПК к апоптозу, сохраняются активности SRC и LTC-IC наиболее примитивных клеток-предшественников, что отражает устойчивость к опосредованному рецепторами апоптозу.
Обогащение миелоидными клетками-предшественниками за счет удаления мертвых клеток
Присущая чувствительность различных субпопуляций клеток ПК к спонтанному и рецептор-индуцированному апоптозу приводит к отчетливо выраженным различиям по составу жизнеспособных клеток после инкубации ex vivo. Например, через 48 часов после инкубации с токсичными дозами FasL происходит отчетливо выраженное обогащение жизнеспособными клетками негативной линии дифференцировки, тогда как инкубация с ФНО-α приводит к относительному обогащению жизнеспособными миелоидными клетками (Фигура 3А). Предполагалось, что обогащение клетками-предшественниками после подвергания воздействию лигандов рецепторов смерти будет увеличивать клоногенную активность в полужидких культурах. Различия по количеству колониеобразующих единиц (КОЕ) можно оценить после удаления мертвых клеток путем осаждения в фиколле и посева на чашках равных количеств жизнеспособных клеток ПК на полутвердой среде на основе метилцеллюлозы (Фигура 3В), которые стимулировали с использованием фактора стволовых клеток (ФСК), интерлейкина-3 (IL-3) и гранулоцитарно-макрофагального колониестимулирующего фактора (ГМКСФ). Согласно преобладающему апоптозу зрелых клеток ПК, количество КОЕ-ГМ увеличивалось от 1:143 в контрольных культурах до 1:37 и 1:31 через 48 часов после воздействия FasL и ФНО-α, соответственно (Фигура 3С). Тогда как максимальное обогащение наблюдали через 48 часов после инкубации, продленное культивирования ex vivo дольше этого периода оказывало отрицательное влияние на функцию коммитированных миелоидных клеток-предшественников. Этот анализ раскрывает селективное рецептор-опосредованное удаление клеток, отличных от клеток-предшественников, в образцах ПК, которое несмотря на похожие общие показатели апоптоза приводит к 4-5-кратному увеличению количества КОЕ-ГМ.
Взаимное влияние рецепторов семейства ФНО не обеспечивает повышение чувствительности к апоптозу
Взаимодействия рецепторов семейства ФНО/лигандов отличаются частичной гомологией, но разной спецификой активации рецепторов. Большинство лигандов связывается с несколькими родственными рецепторами, однако ничего не известно о перекрестной активации рецепторов несколькими лигандами. Взаимное влияние в этом семействе, таким образом, опосредовано индуцированным повышением экспрессии рецепторов в ответ на активацию другого рецептора семейства ФНО. Наиболее известным примером взаимного влияния является индуцированная экспрессия Fas после активации рецептора ФНО в CD34+ клетках-предшественниках. Для определения того, повышает ли экспрессия Fas, индуцированная ФНО, чувствительность клеток ПК к апоптозу, клетки сначала подвергали воздействию ФНО-α и затем подвергали воздействию FasL 15 (Фигура 4А), что приводило к отчетливо выраженному повышению экспрессии рецептора Fas (Фигура 4В). Тогда как в клетках Fas+ происходила индукция апоптоза за счет воздействия FasL на протяжении вторых суток культивирования, клетки ПК, гейтированные CD34+ и выделенные lin- клетки-предшественники были относительно защищенными за счет присутствия ФНО-α (Фигура 4С), что свидетельствовало о том, что повышение экспрессии Fas не зависело от чувствительности к апоптозу. Функциональные анализы обогащения клетками-предшественниками путем лиганд-опосредованного удаления апоптоз-чувствительных клеток показали, что совместное подвергание воздействию FasL и ФНО-α повышает количество КОЕ-ГМ (Фигура 4D) и сообразно с этим, совместное подвергание клеток ПК воздействию обоих лигандов не ухудшает активность SRC in vivo (не показано). Эти данные подтверждают устойчивость гемопоэтических клеток-предшественников к апоптотической сигнальной трансдукции, опосредованную двумя независимыми путями, их совместной активацией и взаимным влиянием рецепторов семейства ФНО.
Деплеция дифференцирующихся клеток на протяжении экспансии клеток ПК ех vivo
Пуповинная кровь представляет собой хороший источник гемопоэтических клеток-предшественников для восстановления иммуно-гемопоэтической системы после агрессивной химиолучевой терапии, однако имеет два главных недостатка: небольшое число клеток и медленное приживление. Для преодоления данных ограничений был разработан целый ряд подходов к экспансии клеток ПК ex vivo до трансплантации. Происходит численная экспансия клеток и активация клеток-предшественников, что улучшает качество восстановления гемопоэза. Предполагалось, что подвергание клеточных культур воздействию лигандов семейства ФНО предоставит относительное преимущество по отношению к более ранним клеткам-предшественникам за счет ограничения клональной экспансии дифференцированных клеток, которые незначительно влияют на эффективность приживления. С применением стандартного протокола экспансии, который обеспечивает увеличение количества фракций (Фигура 5А) и абсолютных количеств CD34+ клеток-предшественников (Фигура 5В), добавление FasL на протяжении третьей и последней недели культивирования ex vivo обепечивало дополнительную значительную экспансию субпопуляции CD34+. Для определения количества и функции SRC с помощью наиболее примитивного анализа человеческих гемопоэтических клеток-предшественников, мышам NOD.SCED с ослабленным иммунитетом, которых обработали бусульфаном, трансплантировали равные количества клеток ПК из одинаковой дозы ПК (Фигура 5С). Воздействие FasL на протяжении третьей недели культивирования приводило к равным и/или более высоким уровням ксеногенного химеризма по человеческим гемопоэтическим клеткам (Фигура 5D), что указывало на то, что произошло улучшение функции клеток-предшественников. Сильная вариабельность приживления у отдельных мышей делает невозможным более точное определение главных функциональных свойств возросшего числа CD34+ клеток-предшественников после экспансии. Таким образом, селективная деплеция клеток, чувствительных к апоптозу, представляет собой эффективный путь увеличения количества фракций и числа клеток-предшественников ПК после экспансии ex vivo для трансплантации.
Пример 2
Подвергание мобилизованных клеток периферической крови воздействию лигандов рецепторов смерти ex vivo
Апоптотическое действие активации рецепторов смерти in vitro
Преобладающим источником гемопоэтических клеток-предшественников для трансплантации является периферическая кровь после мобилизации клеток с помощью гранулоцитарного колониестимулирующего фактора (Г-КСФ) или антагонистов c-kit и CXCR4. Впоследствии путем афереза из периферической крови собирают мобилизованные мононуклеарные клетки, содержащие существенное количество CD34+ клеток-предшественников. Клетки, собранные из периферической крови, обычно активированы, таким образом, для селективной деплеции используются значительно более короткие периоды воздействия лигандов рецепторов смерти. Дополнительное отличие от данных, представленных для клеток ПК, заключается в применении криоконсервированных образцов мПК, отогрев которых ассоциирован с апоптотической гибелью 15-25% клеток. Fas экспрессирован у ~25% CD34+ клеток-предшественников (Фигура 6А) и значительной доли В-лимфоцитов и миелоидных клеток (50-65%, Фигура 6В). Рецепторы ФНО экспрессированы главным образом в клетках позитивной линии дифференцировки мПК с преобладающей экспрессией TNF-R1. После краткого культивирования экспрессия Fas снижается (Фигура 6С), тогда как у всех субпопуляциях отмечается отчетливо выраженное повышение экспрессии TNF-R2 (Фигура 6D). Эти данные раскрывают динамические различия в экспрессии рецепторов смерти в отогретых клетках мПК, на которую, с одной стороны, влияет гибель клеток Fas+ и с другой стороны повышенная экспрессия TNF-R2.
Тогда как 25-45% клеток позитивной линии дифференцировки подвергаются апоптозу в пределах 4 часов инкубации ex vivo, в 60-90% этих клеток происходит индукция апоптоза в пределах 16 часов после инкубации (Фигура 6Е). Присутствие FasL и ФНО-α не уменьшает значимо высокие показатели апоптоза клеток позитивной линии дифференцировки, наблюдаемого в среде. Наиболее выраженное отличие от клеток ПК наблюдается в случае субпопуляции активированных CD3+ Т-клеток, находящихся в мобилизованной периферической крови (мПК), в которой происходит индукция апоптоза лигандами в пределах короткого периода времени. Эти относительные уровни чувствительности к опосредованному рецепторами апоптозу приводят к значительным изменениям состава жизнеспособных клеток с заметным снижением количества Т-клеток через 4 часа после инкубации и В-лимфоцитов и миелоидных клеток через 16 часов после инкубации. В случае обеих временных точек CD3+ Т-клетки подвергаются селективной деплеции, которая приводит к обогащению фракции жизнеспособных клеток CD34+ клетками-предшественниками (Фигура 6F). Таким образом, краткая инкубация мобилизованной периферической крови с лигандами смерти обеспечивает деплецию субпопуляций зрелых клеток и обогащение клетками-предшественниками криоконсервированных образцов мПК.
Деплеция Т-клеток, полученных из мПК, чувствительных к образованию поперечных связей в Fas, улучшает состояние при болезни «трансплантат против хозяина»
Наиболее значимой причиной заболеваемости при трансплантациях клеток мПК является БТПХ, как последствие характерного состояния активированных Т-клеток в периферической крови. Для оценки влияния FasL на БТПХ при ксеногенной трансплантации, мышам NOD.SCID трансплантировали 1,5×107 жизнеспособных клеток мПК, полученных из одинаковой дозы при наличии и отсутствии воздействия лиганда. Предварительная инкубация с FasL в течение 4 часов приводила к выживанию всех мышей, тогда как инкубация в среде вызывала гибель одной трети реципиентов (Фигура 7А). Выжившие клетки проявили похожие уровни химеризма по человеческим гемопоэтическим клеткам (Фигура 7В), что снова продемонстрировало, что SRC, находящиеся в мПК, были устойчивыми к Fas-опосредованному апоптозу. Тяжелая форма БТПХ была причиной смертности мышей, инокулированных мПК, инкубированной в среде, что определили по сильной потери веса (Фигура 7С), клиническим показателям (Фигура 7D) и гистологии печени (Фигура 7Е). Выживаемость и все эти признаки БТПХ при ксеногенной трансплантации улучшались за счет кратковременного воздействия FasL на клетки мПК. В дополнение к этому, исходя из функционального анализа, становится очевидным, что, несмотря на явно похожие показатели общего апоптоза, присутствие лигандов влияет на различные субпопуляции клеток позитивной линии дифференцировки мПК.
Воздействие лигандов рецепторов смерти повышает количество миелоидных клеток-предшественников
Состав субпопуляций жизнеспособных клеток в пределах культур клеток мПК после краткой инкубации отражает различные уровни чувствительности иммунокомпетентных клеток, полученных из мПК, к апоптозу, опосредованному рецепторами Fas и ФНО. Относительное распределение клеток согласуется с высокой чувствительностью CD3+ Т-клеток к апоптозу на протяжении 4 часов инкубации и последующей гибелью CD19+ В-лимфоцитов и CD33+ миелоидных клеток на протяжении более продолжительных периодов инкубации (Фигура 8А). Эти различия навели на мысль, что в образцах мПК количество клеток-предшественников можно повысить за счет краткой инкубации с лигандами смерти и удаления мертвых клеток (Фигура 8В). Воздействие FasL и ФНО-α приводит к 4-кратному обогащению миелоидными клетками-предшественниками, что обеспечивает двойное обогащение по сравнению с наблюдаемым после инкубации в среде (Фигура 8С). Как и в случае с клетками ПК, воздействие лигандов рецепторов смерти приводит к увеличению количества КОЕ, однако в отличие от клеток ПК используется более краткий период подвергания мПК воздействию лигандов смерти. Эти данные подтверждают эффективность и безопасность предложенного функционального удаления эффекторов БТПХ и показывают, что происходит заметное обогащение клетками-предшественниками, нечувствительными к апоптозу.
Дотрансплантационное воздействие FasL не ухудшает иммунную реактивность и реактивность «трансплантат против опухоли»
Решающий вопрос в экспериментах с применением селективной деплеции Т-клеток для предотвращения и уменьшения тяжести БТПХ заключается в том, сохраняет ли трансплантат способность вызывать сильные реакции «трансплантат против опухоли». Несмотря на эти изменения состава, мПК, инкубированная в среде и с FasL, сохраняет реактивность против человеческих облученных аллогенных клеток-стимуляторов мПК (Фигура 8D). Сообразно с этим иммунизация мПК при наличии и отсутствии воздействия FasL оказывает похожее влияние на подавление опухолевого роста у мышей с ослабленным иммунитетом, имеющих подкожные опухоли, вызванные клетками человеческой карциномы толстой кишки НТ29 (Фигура 8Е). Таким образом, нестимулированные криоконсервированные клетки мПК после воздействия FasL подавляют развитие летальной БТПХ, обеспечивая приживление и сохранение ТПО-реактивности.
Пример 3
Дотрансплантационная деплеция Т-клеток предотвращает летальную БТПХ
Предотвращение летальной БТПХ за счет селективной деплеции ex vivo Т-клеток, сенсибилизированных к хозяину
Как физическая деплеция Т-клеток из донорского инокулята, так и FasL-опосредованное удаление несовместимых с хозяином Т-клеток обеспечивают надежное предотвращение БТПХ. Трансплантации гаплоидентичным мышам (от родителей к потомству) представляют огромный риск развития БТПХ, отличающейся высокими уровнями смертности. На первой стадии эксперименты повторяли с применением деплеция Т-клеток с помощью апоптотических сигналов после сенсибилизации к антигенам хозяина. Антиген-специфическая сенсибилизация in vitro в течение 2-3 суток вызывает опосредованную Т-клеточным рецептором (TCR) стимуляцию чувствительных Т-клеток, в которых одновременно повышается экспрессия Fas и родственного лиганда, что приводит к запуску апоптотического каскада параллельно с деактивацией защитных антиапоптотических механизмов. Для сравнения данного метода с исследуемым подходом, который характеризуется применением краткого подвергания воздействию апоптических лигандов ex vivo в отсутствие продолжительной инкубации, доноров подвергали предварительной иммунизации против антигенов хозяина in vivo (Фигура 9А). Как и следовало ожидать, подвергание сенсибилизированных спленоцитов воздействию FasL индуцировало значительный апоптоз как CD4+, так и CD8+ Т-клеток (Фигура 9В). Эффективность сенсибилизации стала очевидной из более сильных пролиферативных ответов лимфоцитов, собранных у мышей В6 (H2Kb), иммунизированных лимфоцитами BALB/c (H2Kd) против стимулирующих аллоантигенов H2Kd, по сравнению с посторонними антигенами (H2Kk) (Фигура 9С). Эти ответы продолжались после краткой (24 часов) инкубации клеток-респондеров в контрольной среде, однако добавление FasL снижало отвечаемость спленоцитов, предварительно сенсибилизированных к стимуляторам H2Kd. Сохранялись ответы на посторонние антигены H2Kk.
Для оценки влияния FasL на способность сенсибилизированных лимфоцитов вызывать БТПХ, жизнеспособные клетки, полученные от иммунизированных H2Kd родительских доноров H2Kb, подвергали адоптивному переносу сублетально облученным гаплоидентичным F1-реципиентам (H2Kb→H2Kb/d). Инфузия предварительно сенсибилизированных спленоцитов, инкубированных в контрольной среде, вызывала развитие тяжелой формы БТПХ, которая в одинаковой степени приводила к летальному исходу после стимуляции липополисахаридом (ЛПС), что привело к гибели всех мышей (Фигура 9D). Стимуляция ЛПС вызывает сильную активацию иммунного ответа и высвобождение провоспалительных цитокинов, которая вызывает максимальное обострение реакций, происходящих вследствие БТПХ, и вызывает гибель клеток. В отличие от этого, инкубация ex vivo спленоцитов, полученных от доноров, предварительно сенсибилизированных с помощью FasL до инфузии, привела к выживанию 50% мышей даже после введения ЛПС. Эта in vivo модель подтверждает предыдущие исследования, в которых показано, что запускаемая аллоантигенами стимуляция лимфоцитов приводит к образованию популяции эффекторных клеток, вызывающих развитие БТПХ, которые можно специфически удалить с применением проапоптотических средств.
Предотвращение летальной БТПХ за счет селективной деплеции ex vivo нестимулированных Т-клеток
Мы предположили, что дотрансплантационное подвергание донорских лимфоцитов воздействию антигенов хозяина может усилить стимуляцию Т-клеток и что эти аллоиммунные ответы могут продолжаться из-за ограниченной чувствительности некоторых субпопуляций эффекторньгх/лимфоцитов памяти, образовавшихся в культуре, к Fas-опосредованному апоптозу или из-за невозможности образования поперечных связей в Fas для удаления совершенно всех аллореактивных Т-клеток. В дополнение к этому, адаптация этого подхода на основе предварительной сенсибилизации к использованию человеческих пар донор:хозяин для использования в клинике может быть достаточно сложной. В поисках более простой и более надежной модели отбора аллореактивных Т-клеток с помощью FasL, модифицировали метод инкубации для применения наивных донорских клеток, которые предварительно не подвергали воздействию аллоантигенов реципиента (Фигура 9Е). В случае нового подхода наивные спленоциты подвергали деплеции ex vivo за счет воздействия среды, содержащей FasL, в отсутствие предварительного или одновременного подвергания воздействию антигенов хозяина и в отсутствие пролиферативных стимулов. FasL усиливал апоптоз наивных CD4+ и CD8+ Т-клеток (Фигура 9F), однако сохранялись аллоиммунные ответы жизнеспособных клеток, которые впоследствии стимулировали in vitro (Фигура 9G). Таким образом, подвергание нестимулированных спленоцитов воздействию FasL сохраняет способность клеток, нечувствительных к апоптозу, отвечать на аллогенные антигены.
Для оценки влияния удаления нестимулированных лимфоцитов ex vivo, проводили инфузию жизнеспособных спленоцитов сублетально облученным гаплоидентичным F1-реципиентам (H2Kb-GFP→H2Kb/d). Несенсибилизированные лимфоциты, инкубированные в среде, в меньшей степени опосредовали развитие летальной БТПХ, поэтому для индукции цитокиновой бури и поляризации донорских лимфоцитов использовали ЛПС. Выживаемость 70% реципиентов нестимулиованных спленоцитов, предварительно обработанных FasL, после стимуляции ЛПС превосходила выживаемость реципиентов спленоцитов после стимуляции в организме хозяина и деплеции ex vivo, тогда как у всех реципиентов спленоцитов, инкубированных в контрольной среде, ускорялось развитие летальной БТПХ (Фигура 9Н). Таким образом, обработка донорских лимфоцитов FasL предотвращала развитие летальной БТПХ (включая стимуляцию ЛПС) в модели с высоким уровнем риска развития острой БТПХ у родителей по отношению к F1-гибридам в отсутствие предварительного подвергания воздействию антигенов хозяина.
Количественные и качественные аспекты предотвращения БТПХ
Адоптивный перенос жизнеспособных спленоцитов после инкубации в контрольной среде вызывал сильную БТПХ у сублетально облученных гаплоидентичных F1-реципиентов, развитие которой замедлялось вследствие предварительной инкубации донорских клеток с FasL (Фигура 10А). Дозозависимое снижение массы тела является свидетельством тяжести прогрессирующей БТПХ у реципиентов гаплоидентичных спленоцитов, инкубированных в среде (Фигура 10В). В отличие от этого, потеря веса значительно снижалась у F1-гибридов мышей, которым проводили инфузию наивных родительских спленоцитов, которые предварительно инкубировали с FasL, что согласовалось с уменьшением тяжести БТПХ. Клинические показатели БТПХ (Фигура 10С) и потеря веса (Фигура 10D) при гаплоидентичных трансплантациях (H2Kb→H2Kb/d) превышали показатели, наблюдаемые в случае мышей с комбинаций аллогенных трансплантаций (H2Kb→H2Kd), и сообразно с этим защитное воздействие предварительной инкубации с FasL было более эффективным в отношении предотвращения БТПХ при аллогенных трансплантациях. Следует отметить, что адоптивный перенос 1,5-5×106 Т-клеток мышам равен дозам 60-200×106 Т-клеток/кг в случае людей, тогда как пороговая безопасная концентрация для предотвращения БТПХ посредством деплеции Т-клеток по фенотипу составляет 0,02×106 Т-клеток/кг при гаплоидентичных трансплантациях.
Для оценки различий в составе иммуннокомпетентных выживших клеток в разных экспериментальных группах, оценивали состав селезенок до и через 2 суток после стимуляции ЛПС (Фигура 10Е). Независимо от адоптивного переноса спленоцитов на протяжении первой недели после сублетального облучения уменьшалось количество всех субпопуляций Т- и В-клеток. У реципиентов лимфоцитов, инкубированных в контрольной среде, формировался ответ на ЛПС за счет резкого увеличения количества селезеночных CD4+ и CD8+ Т-клеток и В-лимфоцитов (p<0,001), количество которых значительно уменьшалось у реципиентов спленоцитов, предварительно обработанных FasL (p<0,001). Повторение этих экспериментов с применением очищенных Т-клеток в противоположность нефракционированным спленоцитам показала похожие результаты (данные не показаны).
Моделирование БТПХ у мышей NOD.SCID для определения времени воздействия лигандов рецепторов смерти ex vivo
Предотвращение БТПХ за счет селективной деплеции нестимулированных донорских Т-клеток ставит вопрос о вовлечении двух возможных механизмов. Во-первых, частичная иммуно- и миелодеплеция, индуцированная с помощью сублетального облучения, наводит на мысль, что остаточные элементы иммунной системы хозяина могут эффективно противодействовать донорским Т-клеткам, которые подвергаются деплеции с помощью FasL, что приводит к подавлению БТПХ. Во-вторых, кондиционирование с помощью тотального облучения всего организма индуцирует повреждение ткани, которое играет важную роль в афферентном звене БТПХ. Для оценки обоих этих механизмов у мышей NOD.SCID с отсутствием лимфоцитов индуцировали болезнь в отсутствие дотрансплантационного кондиционирования. Подвергание нестимулированных спленоцитов воздействию FasL в течение 24 и 48 часов вызывало значительный уровень апоптоза (Фигура 11А) с соответствующим уменьшением количества фракций жизнеспособных CD4+ и CD8+ Т-клеток (Фигура 1IB).
После серии калибровочных экспериментов мышам NOD.SCID проводили инфузию 3×106 или 107 наивных аллогенных спленоцитов (H2Kb→H2Kg7) через 24 или 48 часов после инкубации ex vivo в среде, содержащей FasL, соответственно (Фигура 11С). Инфузия спленоцитов, инкубированных в среде, вызывала ~20% смертность при использовании более высокой дозы, и после проведения стимуляции ЛПС регистрировалась сильная смертность (Фигура 11D). Защитное воздействие инкубации с FasL на развитие летальной БТПХ, опосредованное донорскими лимфоцитами, снова отличалось преобладающей выживаемостью мышей после инъекции ЛПС (Фигура 11D). Это защитное воздействие сопровождалось заметным улучшением клинических показателей (Фигура 11Е) и уменьшением потери веса (Фигура 11F). Предотвращение развития БТПХ у мышей NOD.SCID в отсутствие компетентного остаточного иммунитета хозяина и в отсутствие предрасполагающих факторов хозяина, таких как индуцированное кондиционированием повреждение ткани, указывает на то, что БТПХ главным образом является последствием модуляции, опосредованной инокулятом донорских Т-клеток за счет подвергания нестимулированных лимфоцитов воздействию FasL.
Пример 4
Влияние селективной деплеции нестимулированных лимфоцитов ex vivo на приживление гемопоэтических клеток
Деплеция наивных спленоцитов, чувствительных к Fas, ослабляет развитие БТПХ при трансплантациях костного мозга
Чтобы подтвердить результаты, полученные с использованием модели, имитирующей РТПХ, в условиях трансплантации, оценивали влияние Fas-опосредованной деплеции наивных клеток хозяина у мышей, которых подвергали гаплоидентичной ТКМ наряду с инфузией донорских лимфоцитов (Фигура 12А). У облученных реципиентов F1, которым вводили 5×106 гаплоидентичных ВМС наряду с 1,5, 3 и 4,5 миллионами спленоцитов, предварительно инкубированных в контрольной среде (H2Kb→H2Kb/d), проявилась тяжелая форма БТПХ (Фигура 12В) и значительная потеря веса на 10-15% (Фигура 12С). Клинические показатели БТПХ подтвердились гистологическим анализом препаратов кожи уха и печени (Фигура 12D). Предварительная инкубация вводимых посредством инфузии наивных спленоцитов с FasL в течение 24 часов приводила к снижению клинических показателей, характерных для БТПХ (р<0,005), уменьшению потери веса (р<0,005) и снижению уровня лимфоцитарной инфильтрации пораженных органов-мишеней (р<0,005). Однако после инфузии 3-4,5×106 гаплоидентичных спленоцитов, предварительно обработанных FasL, все же имел место остаточный слабовыраженный эффект БТПХ. Следует отметить, что эта доза Т-клеток соответствует дозам 4-6×107 Т-клеток/кг, количествам Т-клеток, которые часто вводят при трансплантациях неманипулированных аллогенных клеток цельного костного мозга, полученного от совместимых неродственных доноров.
Восстановление гемопоэза и иммунитета после гаплоидентичной ТКМ и селективной деплеции клеток, чувствительных к апоптозу,
БТПХ подавляет функцию трансплантата и основательно подавляет иммунные ответы реципиентов. Чтобы определить, как деплеция донорских спленоцитов, чувствительных к Fas, влияет на восстановление гемопоэза и иммунитета после трансплантации, у хозяев оценивали выживаемость трансплантированных донорских лимфоцитов, химеризм и иммунологическую реактивность на неродственные антигены. У реципиентов самой высокой дозы (4,5×106) неманипулированных донорских спленоцитов, предварительно инкубированных в контрольной среде, отмечалось снижение уровней химеризма через 3 недели и 6 недель после трансплантации (Фигура 13А) по сравнению с реципиентами спленоцитов, предварительно обработанных FasL, что продемонстрировало, что предварительная обработка донорских лимфоцитов отменяет супрессорное влияние трансплантата, опосредованное необработанными клетками.
Из-за того, что зрелые лимфоциты, которые вводят посредством инфузии во время трансплантации, конкурируют с лимфоцитами, которые образуются в трансплантате, полученном из клеток костного мозга, за гомеостатическую периферическую экспансию и влияют на итоговое количество лимфоцитов, изучали влияние инфузий лимфоцитов, обработанных FasL, на восстановление количества лимфоцитов. Для того чтобы на протяжении нескольких недель после трансплантации можно было различать у реципиентов спленоциты, введенные посредством инфузии, и Т-клетки, полученные из трансплантата ВМС, воспользовались небольшим различием по антигенам CD45 (CD45.1 по сравнению с CD45.2). Несмотря на то, что у реципиентов F1 не происходило отторжение родительских иммунокомпетентных клеток, через 3 недели после трансплантации в периферических лимфоидных органах реципиентов было обнаружено незначительное количество вводимых посредством инфузии спленоцитов (H2Kb, CD45.2+GFP+), независимо от того, инкубировали ли их с FasL или в контрольной среде (Фигура 13В). В качественном отношении у реципиентов контрольных и предварительно обработанных FasL спленоцитов отмечалась иммунологическая неотвечаемость как на антигены донора, так и хозяина, как и следовало ожидать в случае химерных мышей, и в равной степени выраженный ответ на посторонние антигены (H2Kk) (Фигура 13С). Таким образом, похоже, что обработка лимфоцитов ex vivo с помощью FasL не влияет ни на количественное, ни на функциональное восстановление иммунной системы после трансплантации, все же предотвращение БТПХ улучшает приживление донорских клеток.
Мы задались вопросом, вызвано ли предотвращение БТПХ за счет предварительной инкубации нестимулированных лимфоцитов с помощью FasL ухудшением миграционной способности клеток. Эту вероятность оценили посредством инфузии жизнеспособных спленоцитов сублетально облученным реципиентам F1 (H2Kb-GFP→H2Kb/d) и демонстрации того, что лимфоциты возвращались в селезенку и мезентеральные лимфатические узлы реципиента через 24 часа после инфузии, независимо от того, подвергались ли клетки предварительному воздействию FasL или контрольной среды (Фигура 13D). Эти данные демонстрируют, что предварительное воздействие на лимфоциты обеспечивает деплецию потенциальных эффекторов БТПХ без ухудшения их миграционной способности.
Подвергание гемопоэтических клеток воздействию лигандов рецепторов смерти улучшает приживление
Деплеция всех Т-клеток из трансплантатов, состоящих из гемопоэтических клеток, обеспечивает удаление клеток, способствующих приживлению, что требует компенсаторной инфузии больших доз стволовых клеток. Предварительная инкубация трансплантата с FasL в течение 24 часов (Фигура 14А) не влияла на уровни донорского химеризма, индуцированного с помощью трансплантации 2×106 аллогенных (H2Kd→H2Kb) клеток цельного костного мозга облученным хозяевам (Фигура 14В). Таким образом, приживление сохраняется, когда гетерогенные популяции клеток цельного костного мозга подвергаются воздействию лигандов рецепторов смерти до трансплантации.
Т-клетки поддерживают приживление гемопоэтических клеток-предшественников посредством двух механизмов: а) донорские Т-клетки могут препятствовать отторжению трансплантата за счет остаточного донорского иммунитета и b) Т-клетки совместно с клетками-предшественниками находятся в местах оседания в костном мозге и поддерживают приживление посредством неидентифицированных неиммуногенных механизмов. Чтобы определить, приводит ли деплеция апоптоз-чувствительных Т-клеток, используемых для предотвращения отторжения, также и к удалению клеток, способствующих приживлению, облученным мышам H2Kb трансплантировали 106 жизнеспособных неманипулированных H2Kd lin- клеток-предшественников. В случае данной модели использовали спленоциты, полученные от доноров F1 (H2Kb/d), которые не приводят к развитию БТПХ. Инфузия 106 жизнеспособных спленоцитов F1 после инкубации в контрольной среде или с FasL повышала уровни донорского химеризма (Фигура 14С), что продемонстрировало, что деплеция спленоцитов, чувствительных к Fas, не обеспечивает удаление клеток, которые способствуют приживлению HSPC.
Предварительное подвергание донорских лимфоцитов воздействию лигандов рецепторов смерти сохраняет эффект замедленной инфузии донорских лимфоцитов, поддерживающий приживление
Посттрансплантационная инфузия донорских лимфоцитов (DLI) является эффективным подходом к улучшению донорского химеризма, повышению отвечаемости на инфекции и вызову реакции «трансплантат против опухоли». Хотя замедленная DLI лучше переносится и вызывает менее выраженную БТПХ, чем инфузия лимфоцитов кондиционированным реципиентам во время трансплантации гемопоэтических клеток, лимфоциты обладают способностью к увеличению тяжести БТПХ. Замедленная инфузия донорских лимфоцитов в модели смешанного химеризма (Фигура 14Е) была эффективной в отношении увеличения уровней донорского химеризма независимо от дотрансплантационного подвергания лимфоцитов воздействию FasL (Фигура 14F). Таким образом, деплеция нестимулиованных лимфоцитов, чувствительных к Fas, не ухудшает эффективность посттрансплантационной DLI.
FasL ослабляет развитие БТПХ без ослабления реакции ТПО
Общий успех трансплантации стволовых клеток, которую осуществляют для лечения злокачественной болезни, может зависеть от результатов реакции «трансплантат против опухоли» (ТПО). Тотальная деплеция Т-клеток (либо ex vivo, либо посредством применения сильной иммуносупрессии in vivo) может ослабить влияние ТПО и повысить частоту посттрансплантационных рецидивов в различных клинических ситуациях. В двух экспериментальных моделях исследовали вероятность того, отменяет ли инкубации с FasL эффекты ТПО параллельно с его положительным влиянием на БТПХ. В первой модели оценивали сохранение ТПО-реактивности совместимых с хозяином лимфоцитов после FasL-опосредованной деплеции, направленной против аллогенной опухоли. Клетки карциномы толстой кишки СТ26 (H2Kd) имплантировали мышам NOD.SCID (H2Kg7) после адоптивного переноса 1,5×107 спленоцитов, полученных от (иммунокомпетентных) доноров NOD (H2Kg7, Фигура 15А). Спленоциты, предварительно инкубированные с FasL в течение 24 часов, подавляли рост опухолевых имплантатов в той же степени, что и спленоциты, инкубированные в контрольной среде (Фигура 15В), что подтверждает тот факт, что инкубация с FasL сохраняет опосредованную лимфоцитами ТПО-реактивность. Во второй модели мышей H2Ka, имеющих нейробластомы, совместимые по ГКГС (Neuro-2a, H2Ka), подвергали сублетальному облучению, и им трансплантировали 2×106 lin- клеток-предшественников, полученных от аллогенных доноров (H2Kb, Фигура 15С). Инфузия лимфоцитов, полученных от доноров F1 (H2Kb/d), которые распознают опухоль как свое, не оказывала значительного влияния на опухолевый рост (Фигура 15D). В отличие от этого, инфузия аллогенных спленоцитов (H2Kb) снижала показатели опухолевого роста, однако 80% мышей погибло в пределах 3 недель вследствие тяжелой формы БТПХ. В Fas-опосредованная деплеция нестимулированных донорских спленоцитов (H2Kb) ex vivo имела равноценный эффект, обеспечивающий подавление опухолевого роста, при этом происходило ослабление развития летальной БТПХ и все мыши дожили до экспериментальной конечной точки. Эти данные подтверждают, что БТПХ не является предпосылкой для проявления эффектов ТПО, таким образом, предотвращение БТПХ не ухудшает противоопухолевую активность лимфоцитов после селективной деплеции.
FasL-опосредованное удаление злокачественных клеток
Несмотря на значительные преимущества и безопасность аутологичных трансплантаций после агрессивной химиолучевой терапии, контаминация трансплантата остаточными злокачественными клетками представляет собой фактор риска рецидива болезни. Для оценки такой вероятности мышам проводили инфузию смеси клеток костного мозга и В-клеточной лимфомы А20 в дозе, которая является летальной для мышей изогенной линии BALB/c (Фигура 15Е). Тогда как инфузия смесей клеток, инкубированных в среде, приводила к гибели 80% реципиентов в пределах 4 недель, все мыши, которым проводили инфузию трансплантатов, подверженных воздействию FasL, пережили этот период (Фигура 15F). Эти данные подтверждают эффективность краткой инкубации с лигандами смерти для деплеции апоптоз-чувствительных злокачественных клеток из трансплантата из гемопоэтических клеток.
Деплеция клеток, чувствительных к Fas, предотвращает адоптивный перенос аутоиммунитета при аутологичных трансплантациях
Сахарный диабет 1 типа представляет собой аутоиммунную реакцию, которая разрушает инсулин-продуцирующие β-клетки в островках Лангерганса поджелудочной железы. Это нарушение обычно моделируют на мышах, страдающих диабетом, который не вызван ожирением (NOD), причем частота заболевания составляет выше 80% у самок в возрасте 30 недель. Развития аутоиммунного диабета можно добиться путем адоптивного переноса Т-клеток, но трансплантация клеток цельного костного мозга не является столь же эффективным методом. В предварительных экспериментах определили, что 5×107 клеток костного мозга обеспечивали перенос болезни у ~50% сублетально облученных мышей NOD.SCID (Фигура 15G), причем обе линии имели одинаковый гаплотип (H2Kg7). В дополнение к этому, у мышей NOD.SCID не происходило отторжение донорских клеток, поскольку у них отсутствуют компетентные Т- и В-лимфоциты. Тогда как адоптивный перенос ВМС, инкубированных в среде в течение 48 часов, обеспечивал эффективный перенос болезни, ни у одного из реципиентов ВМС, подверженных воздействию FasL, не отмечалась гипергликемия (Фигура 15Н). Таким образом, подвергание гемопоэтических клеток воздействию лигандов смерти ex vivo приводит к удалению клеток с аутоиммунной реактивностью.
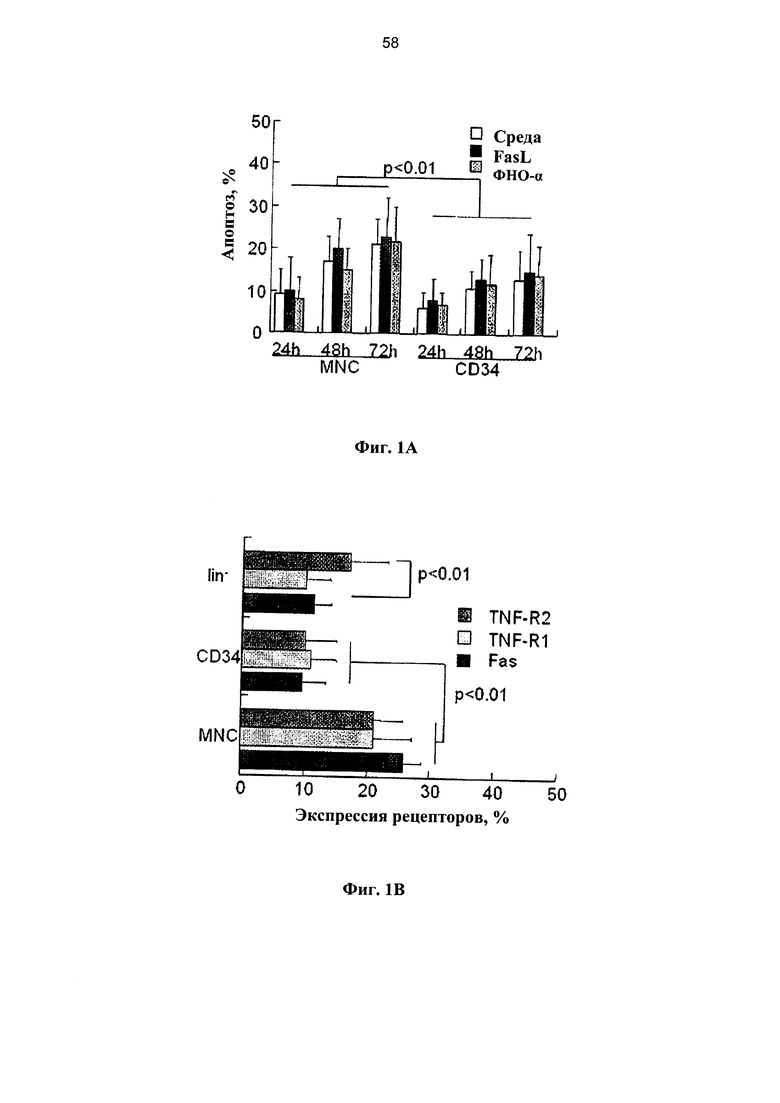
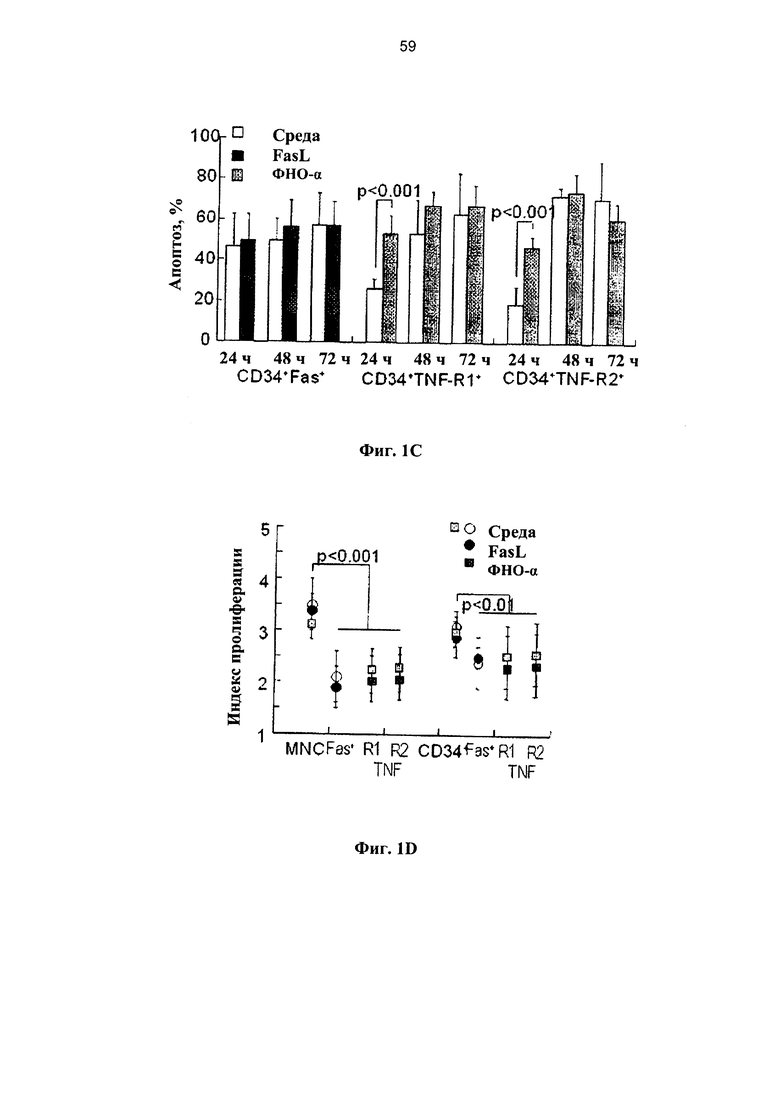
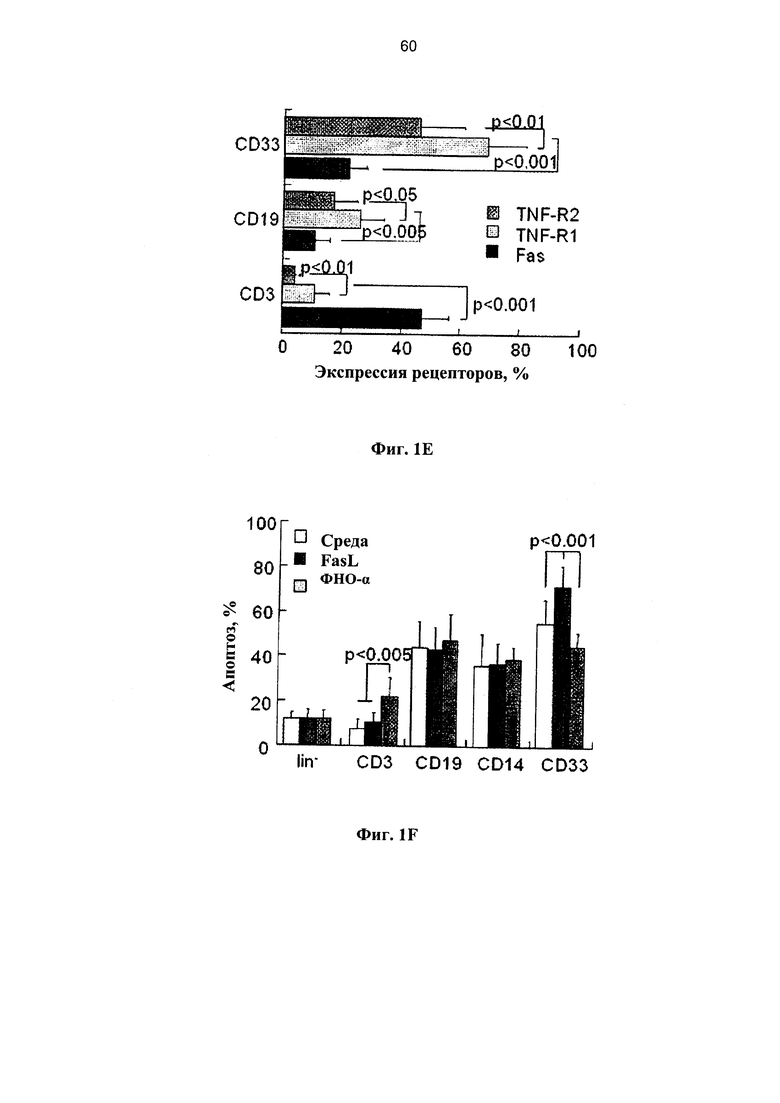
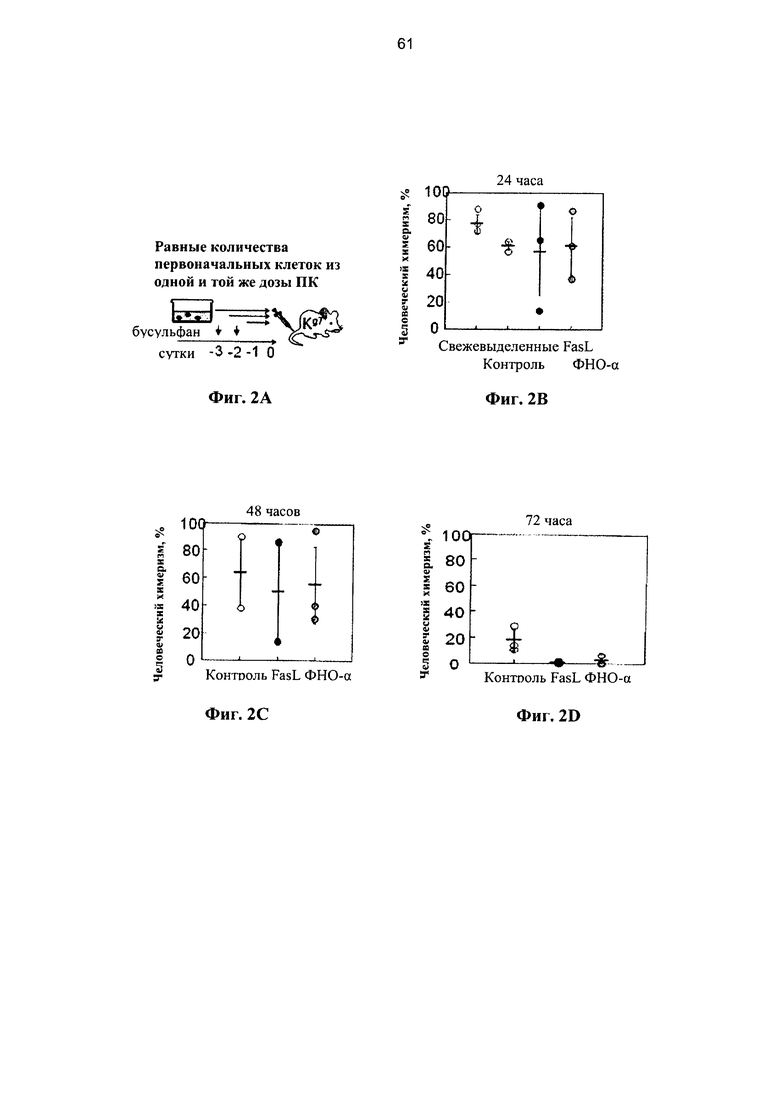
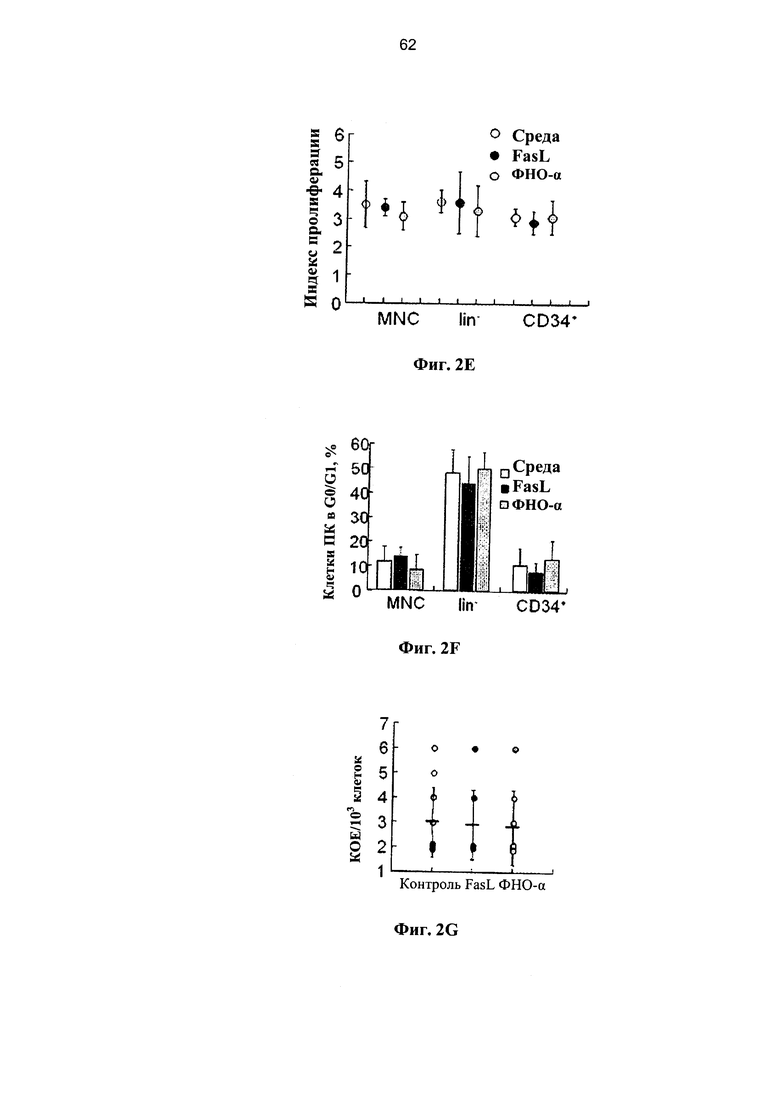
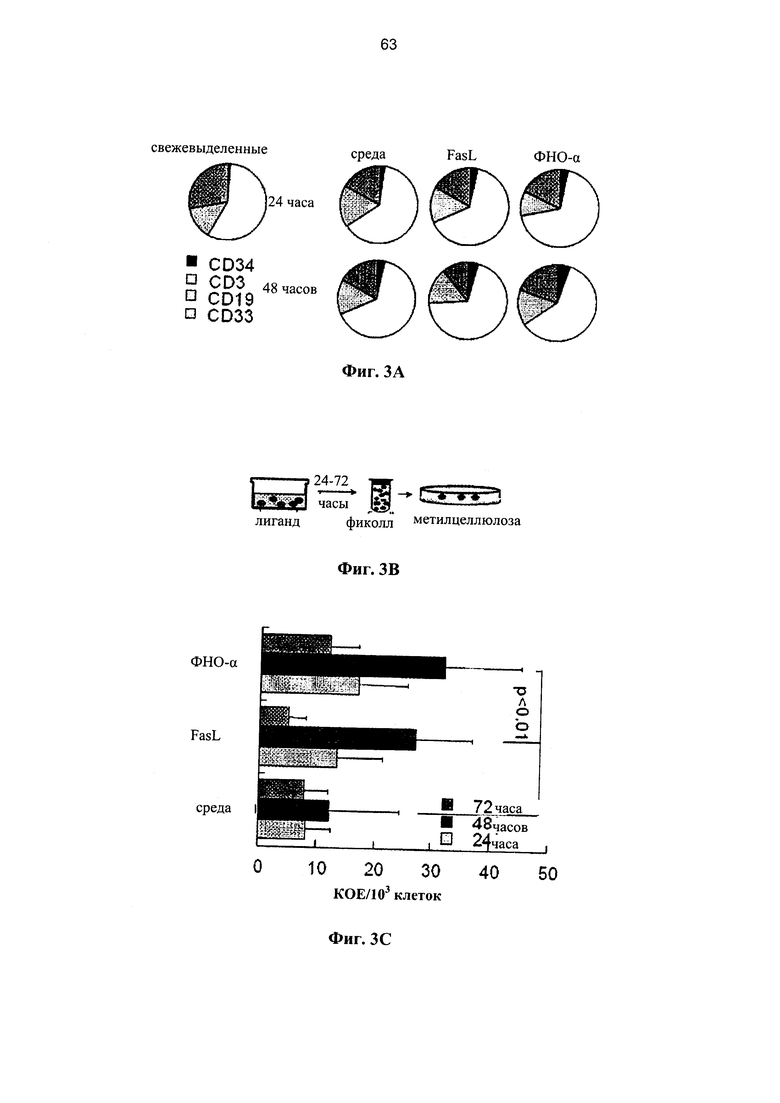
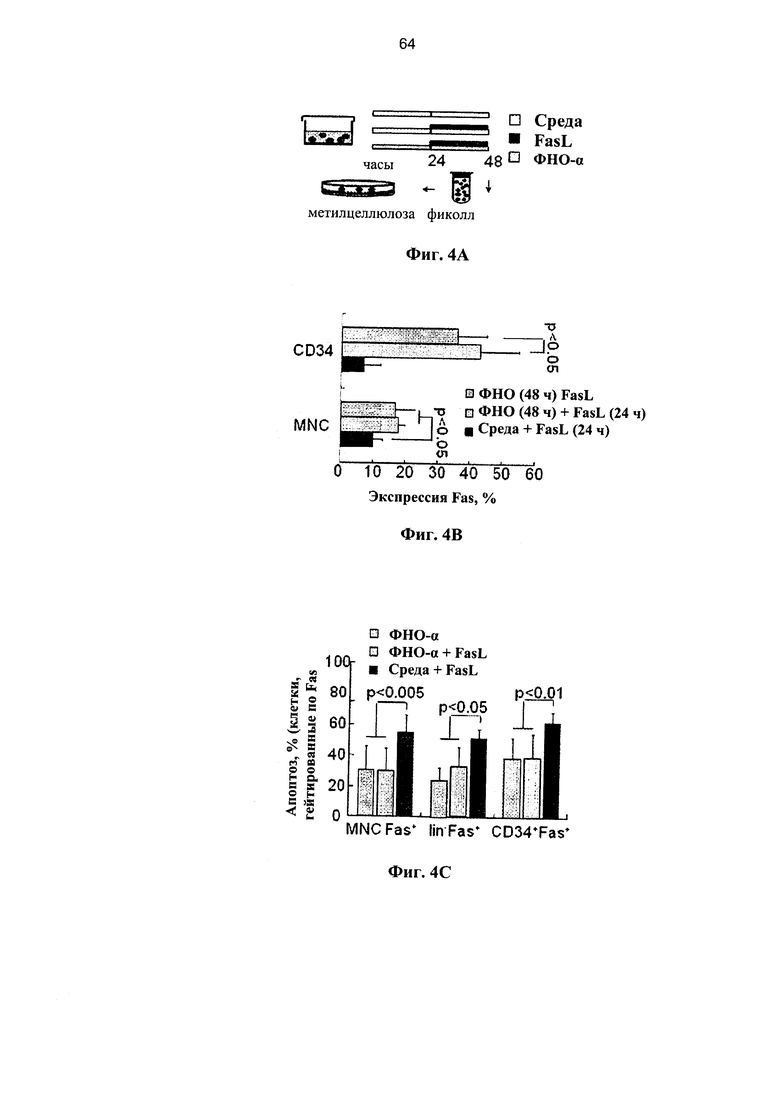
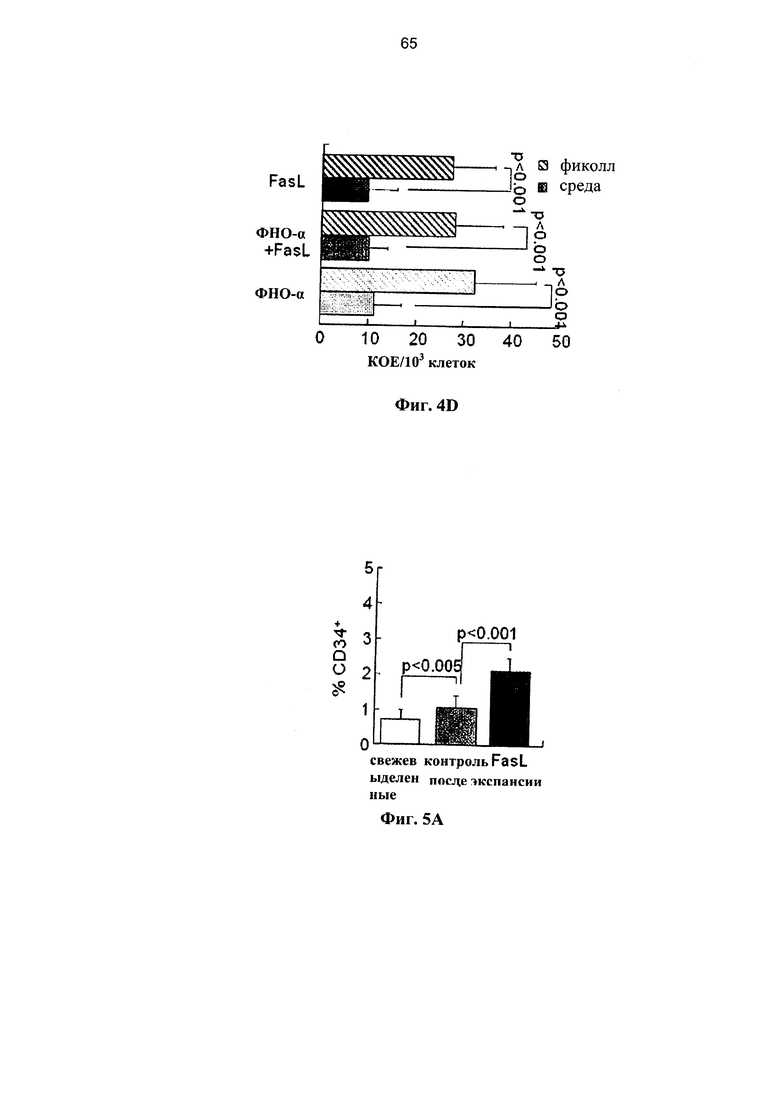
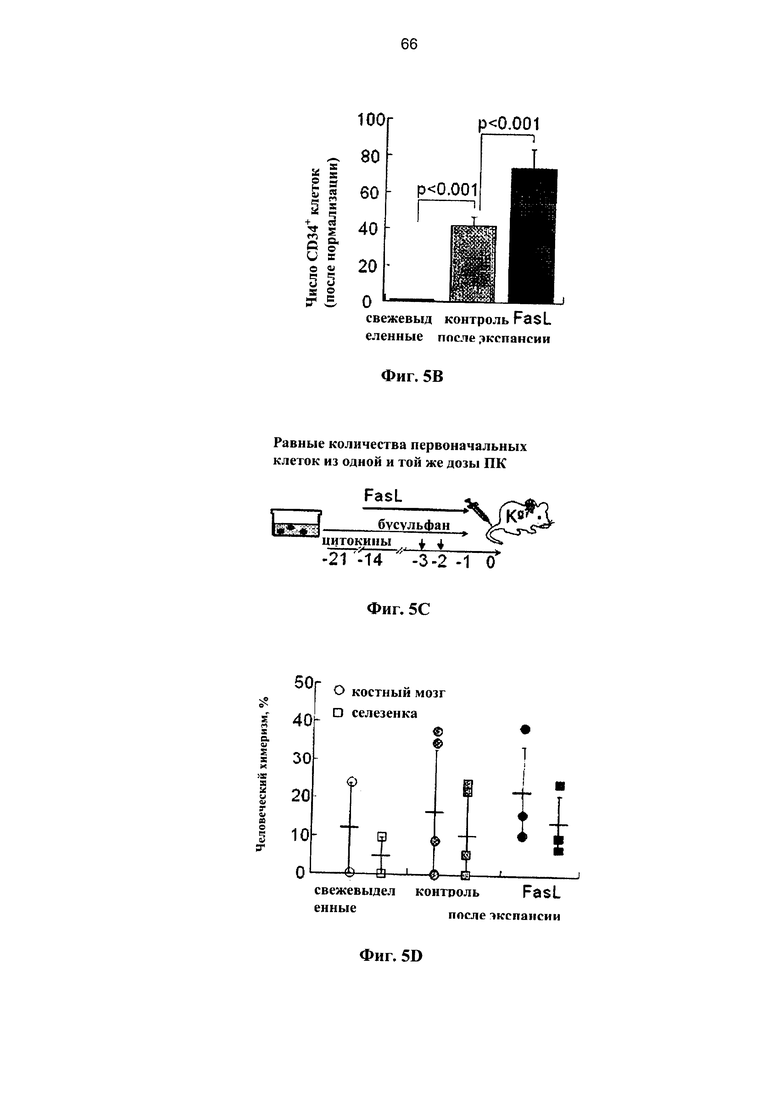
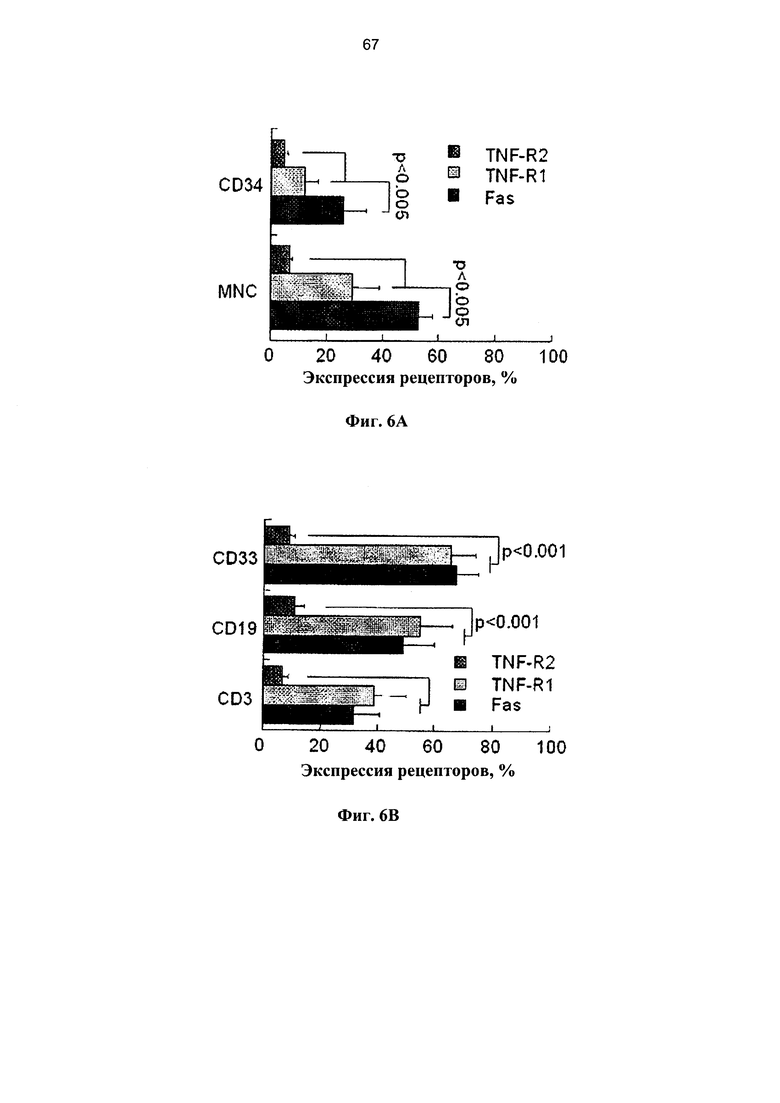
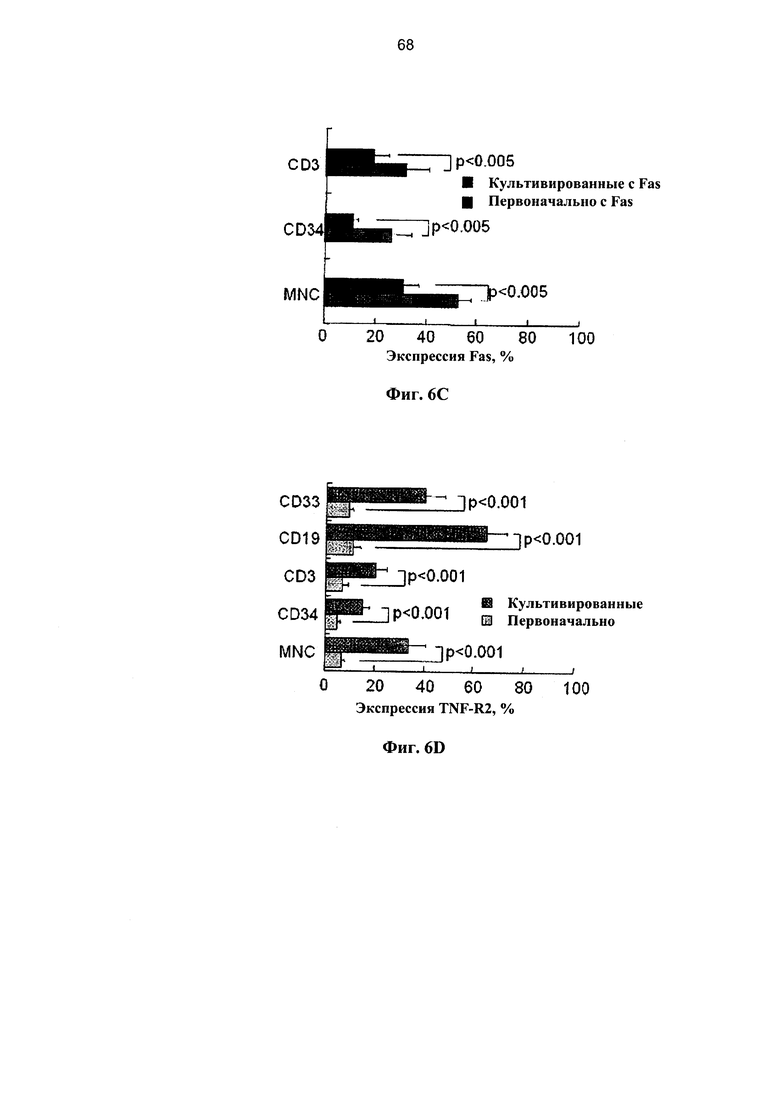
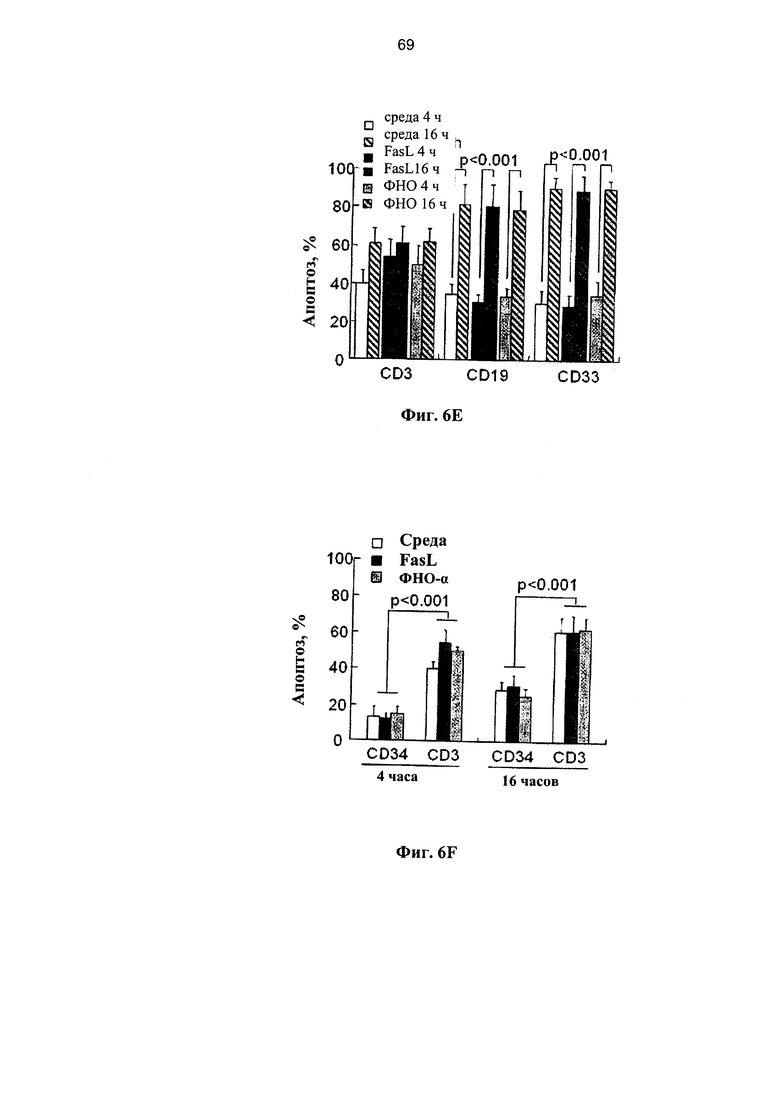
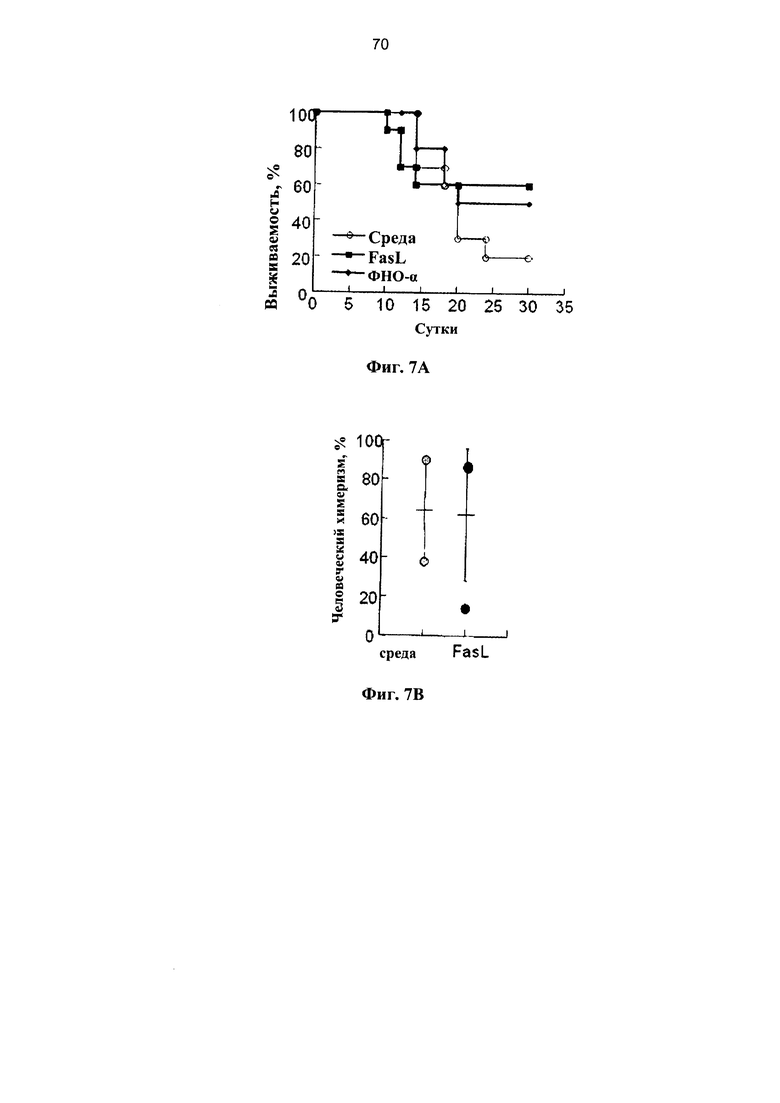
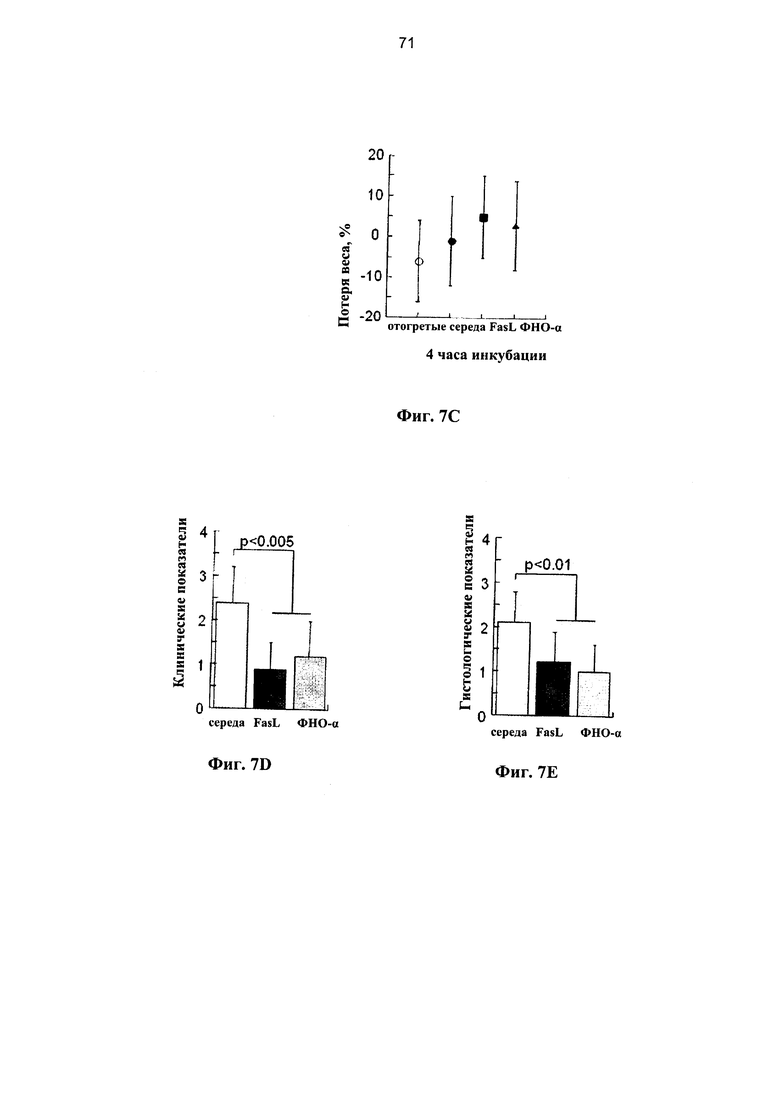
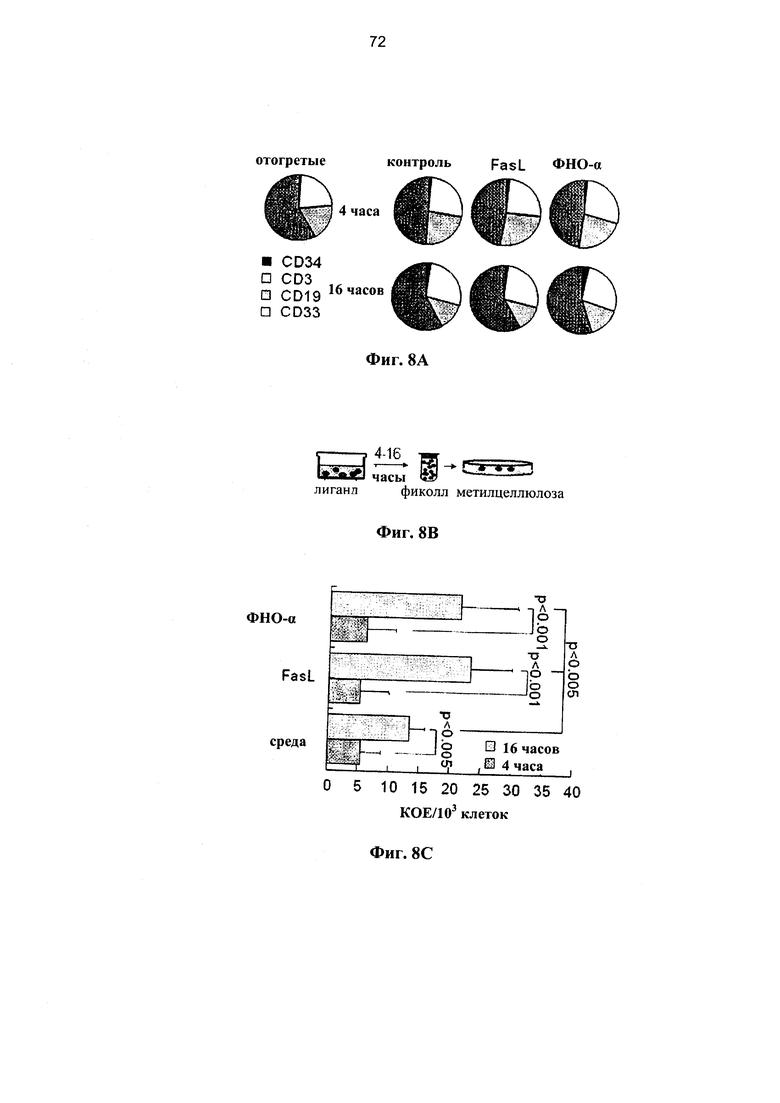
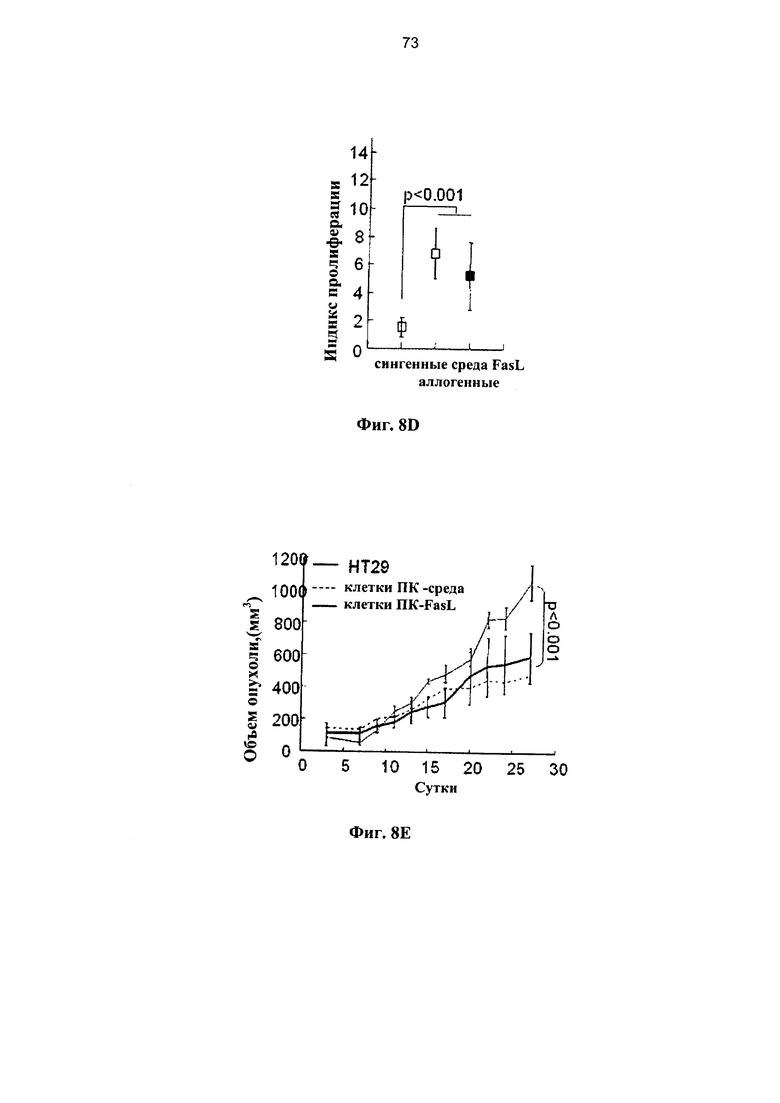
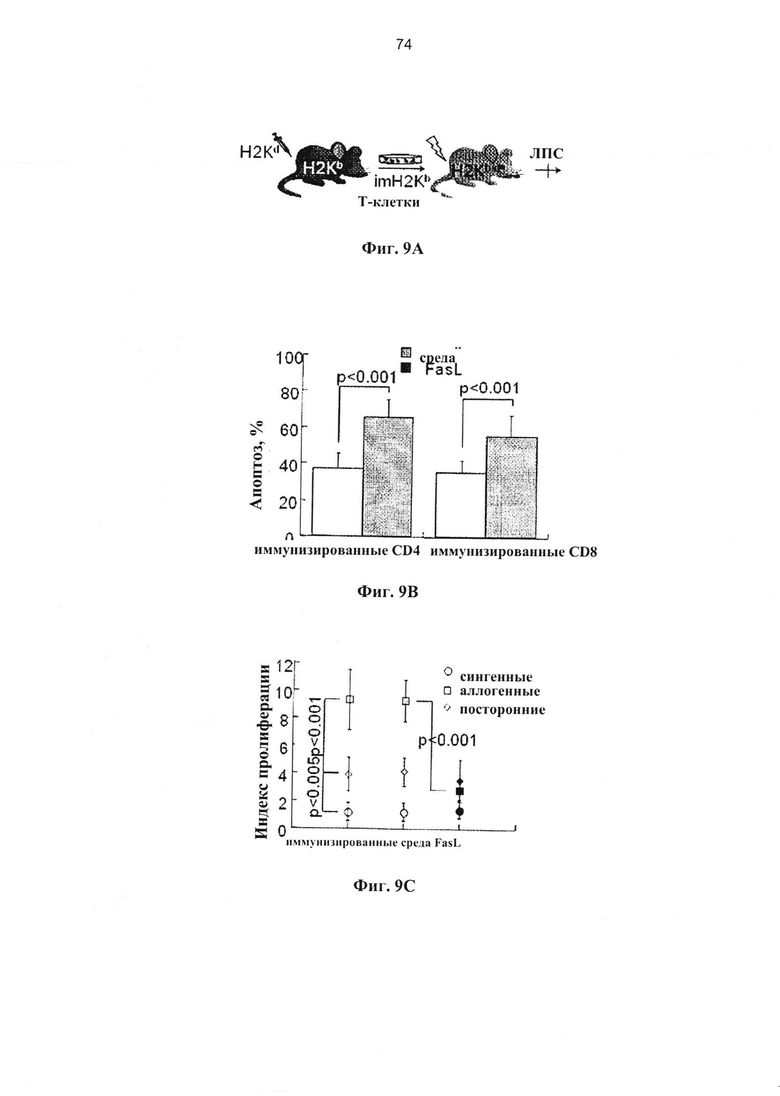
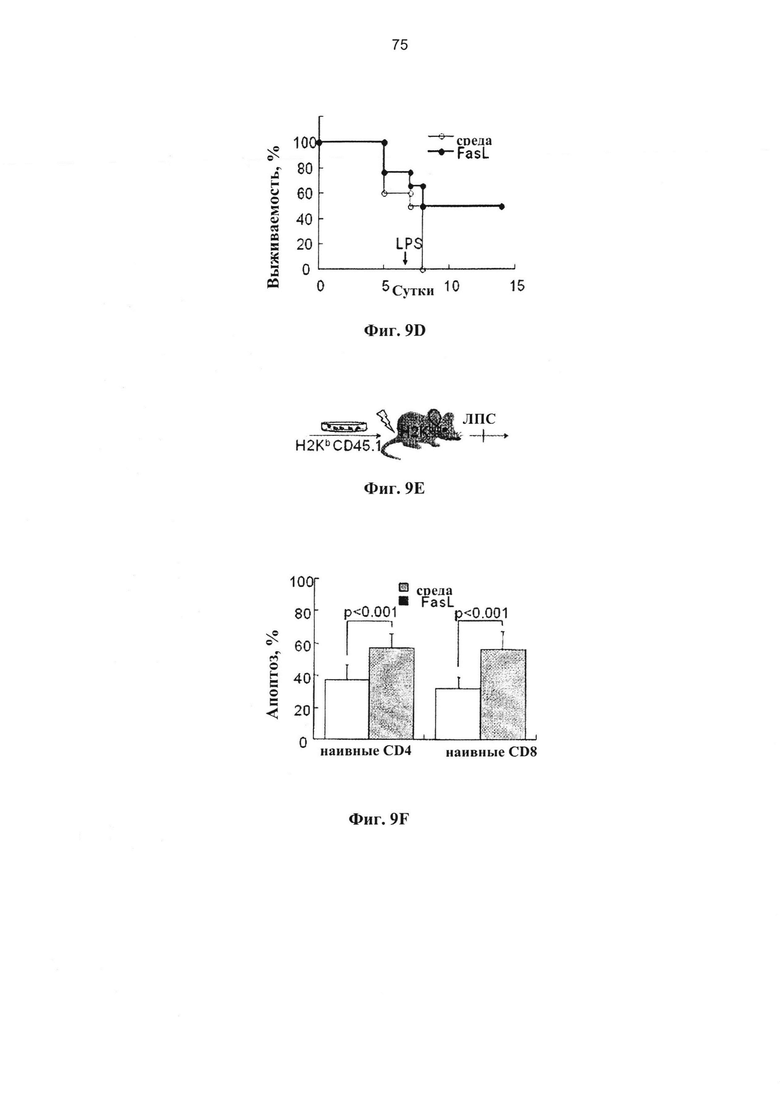
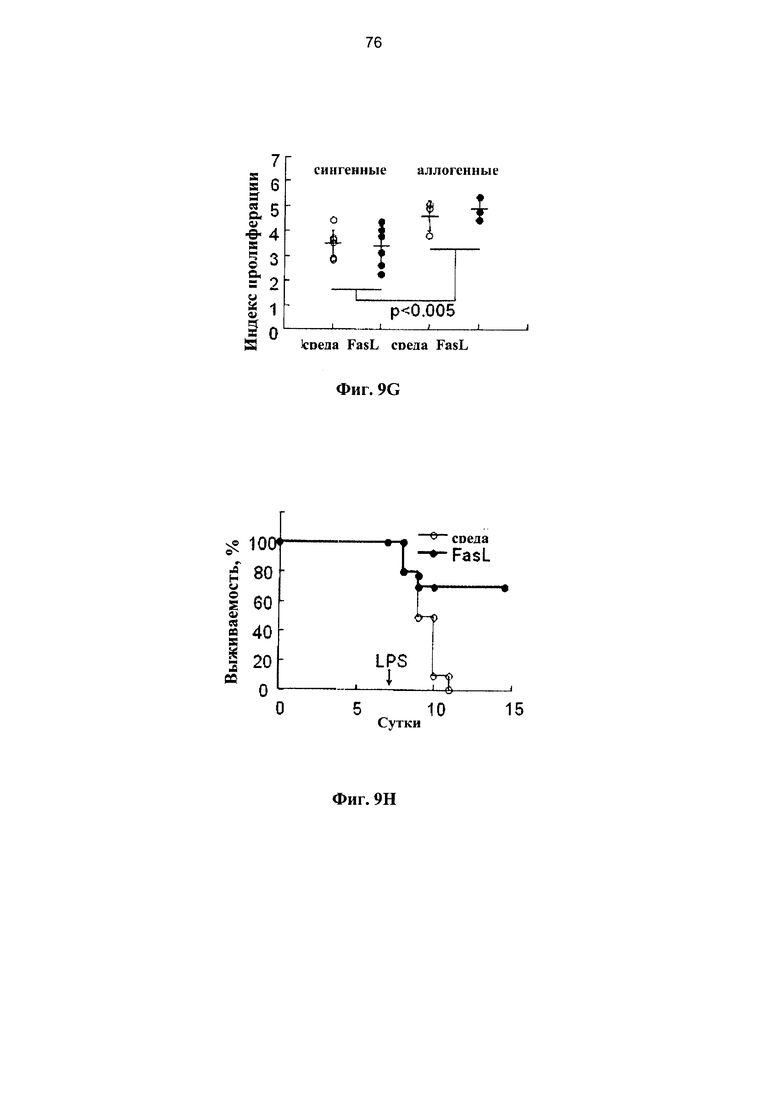
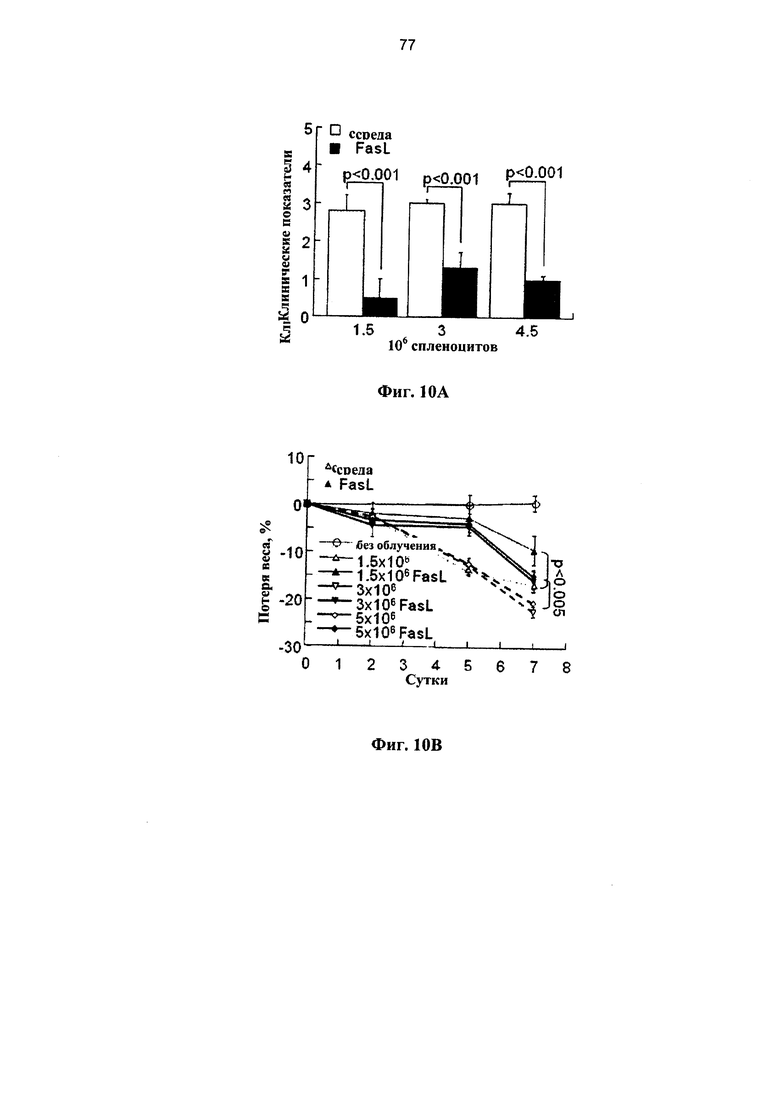
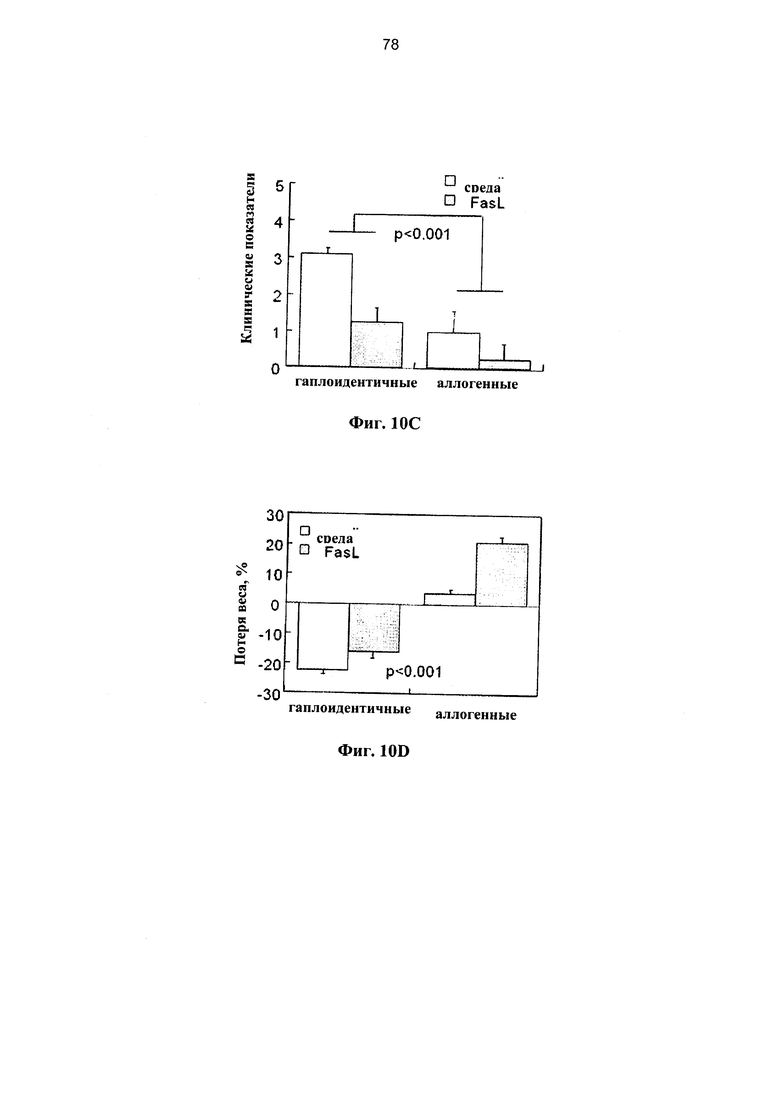
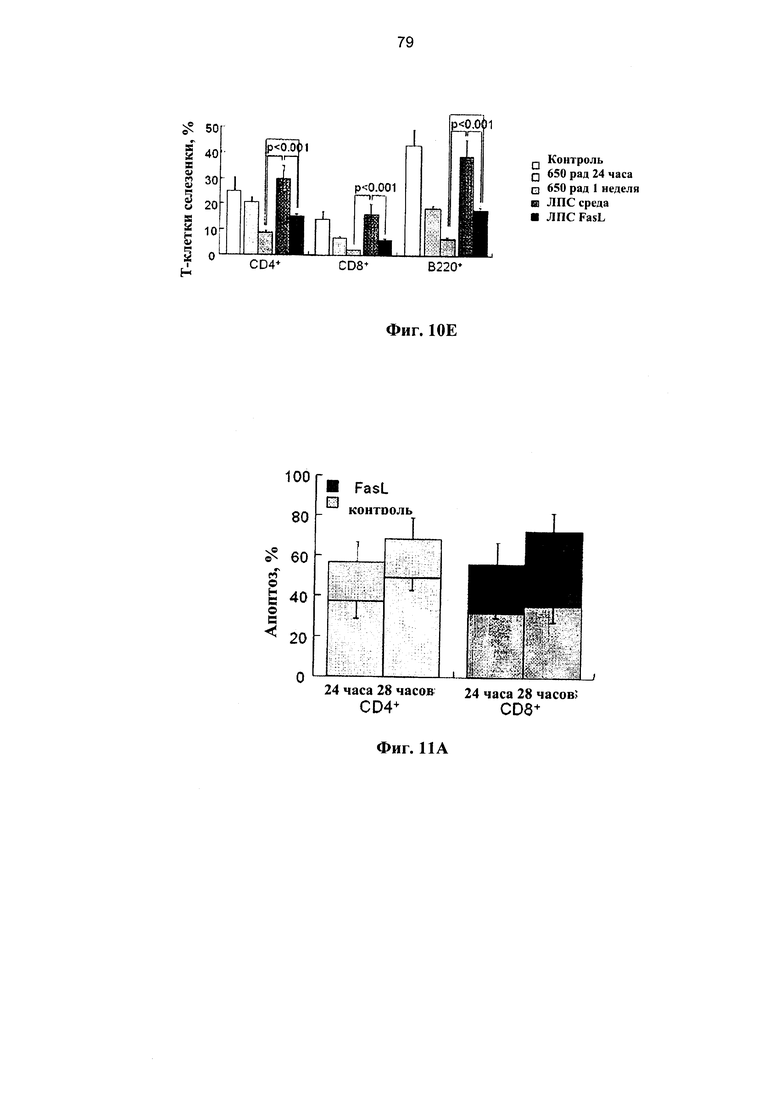
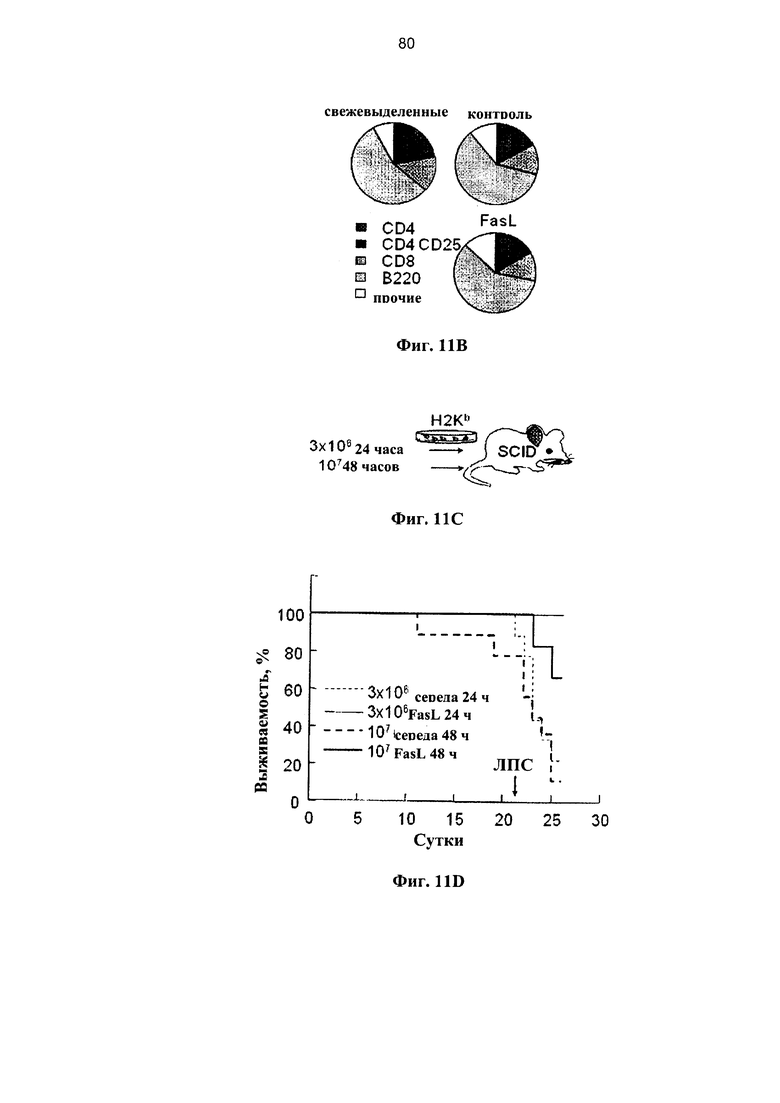
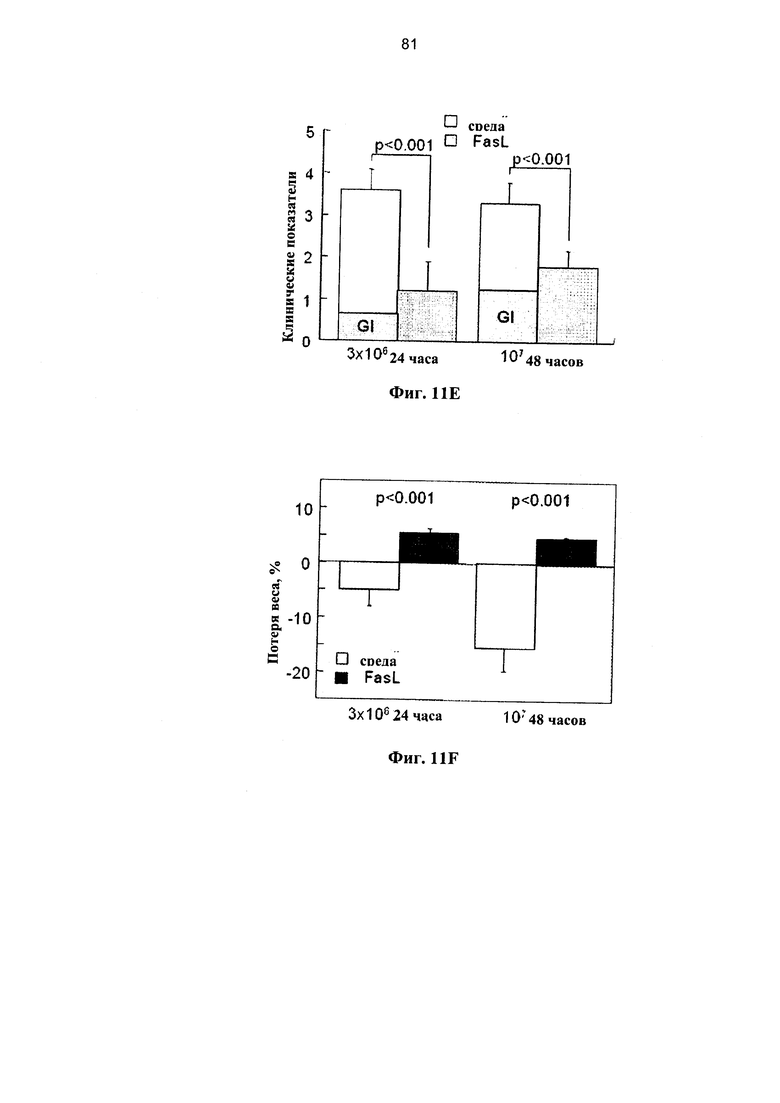
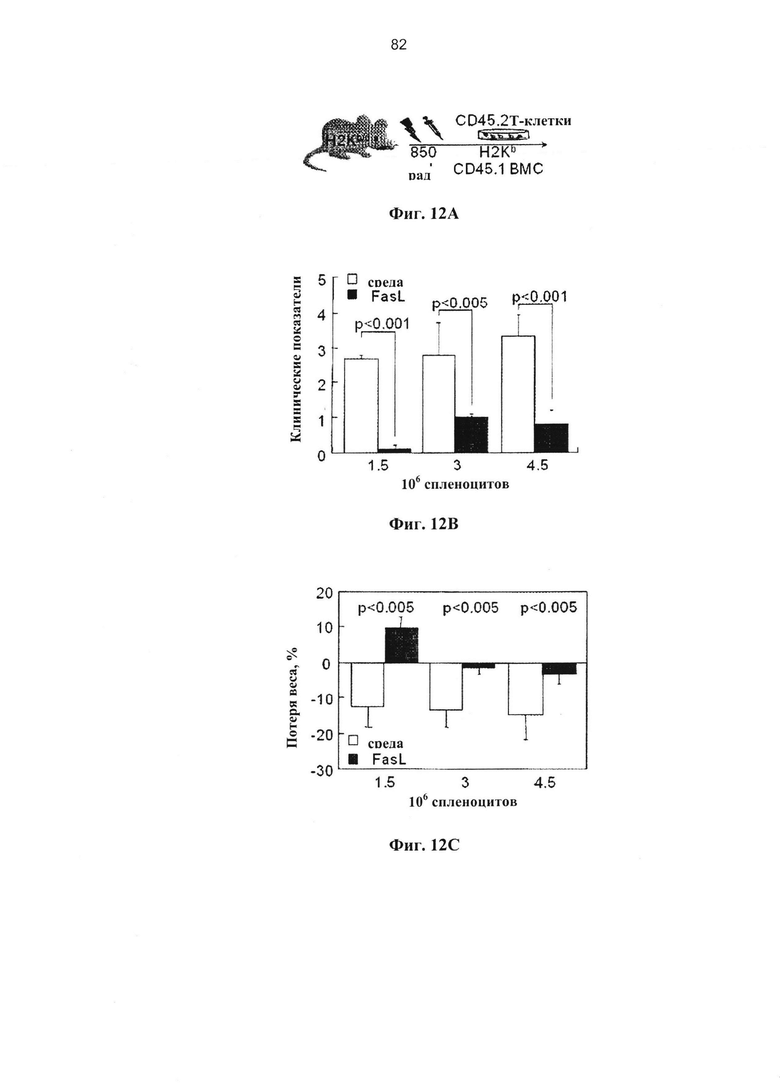
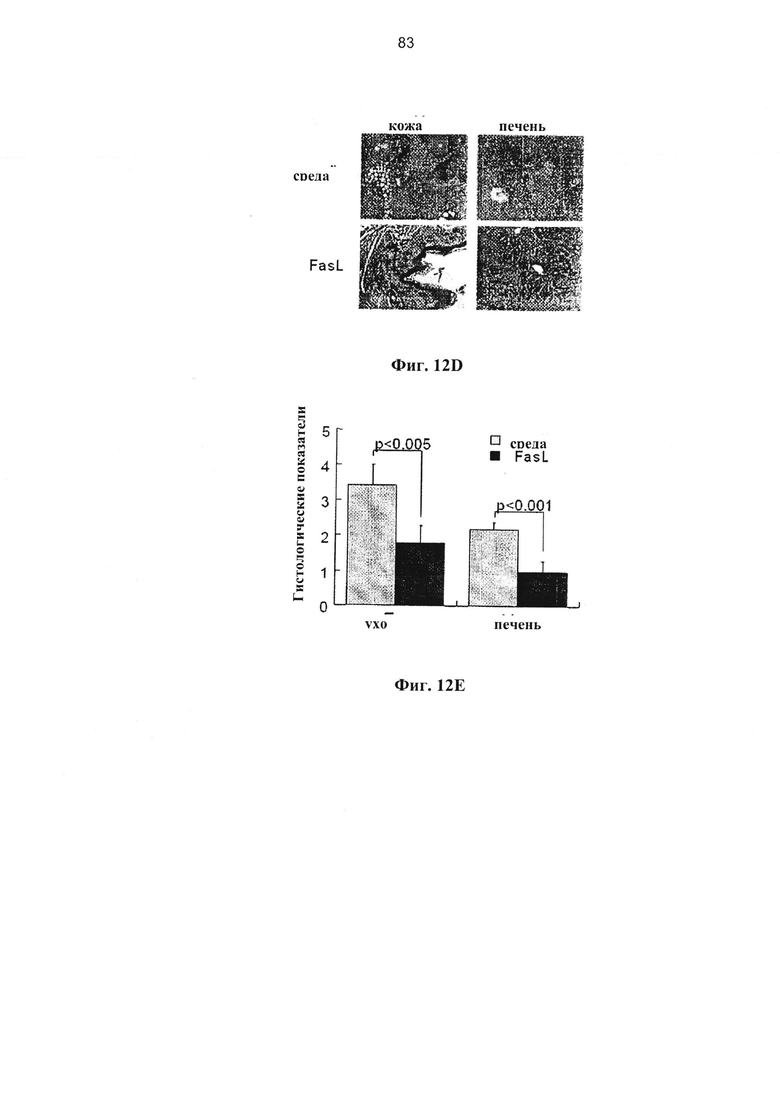
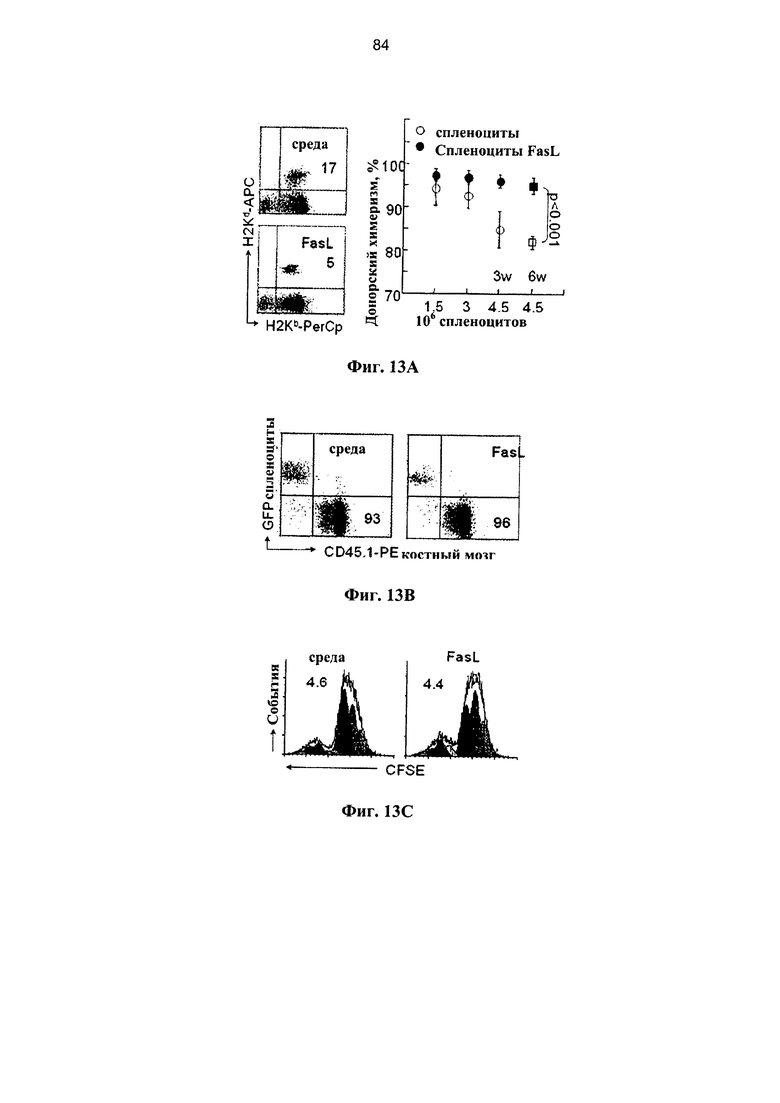
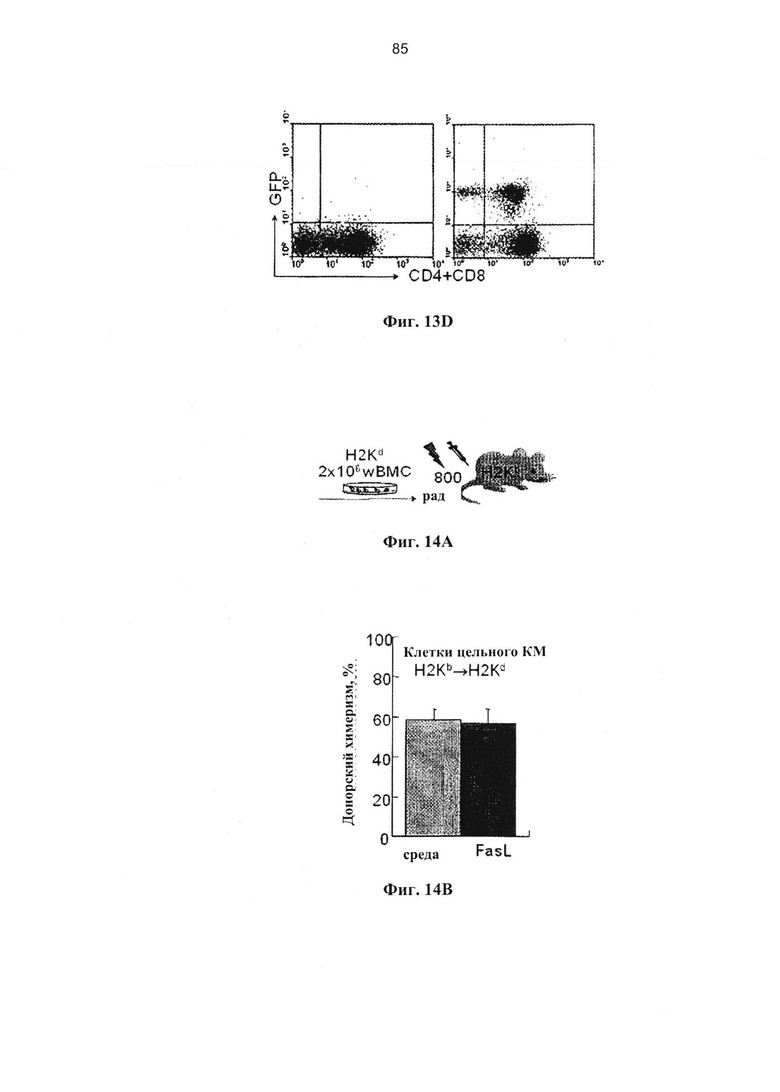
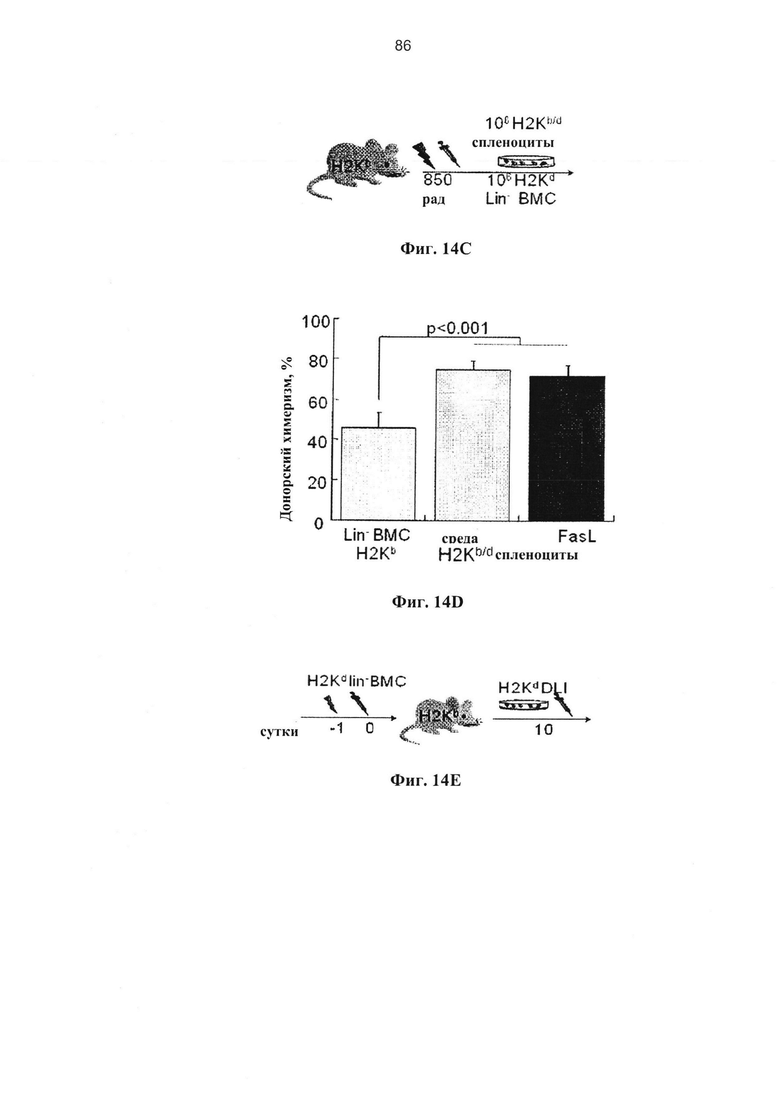
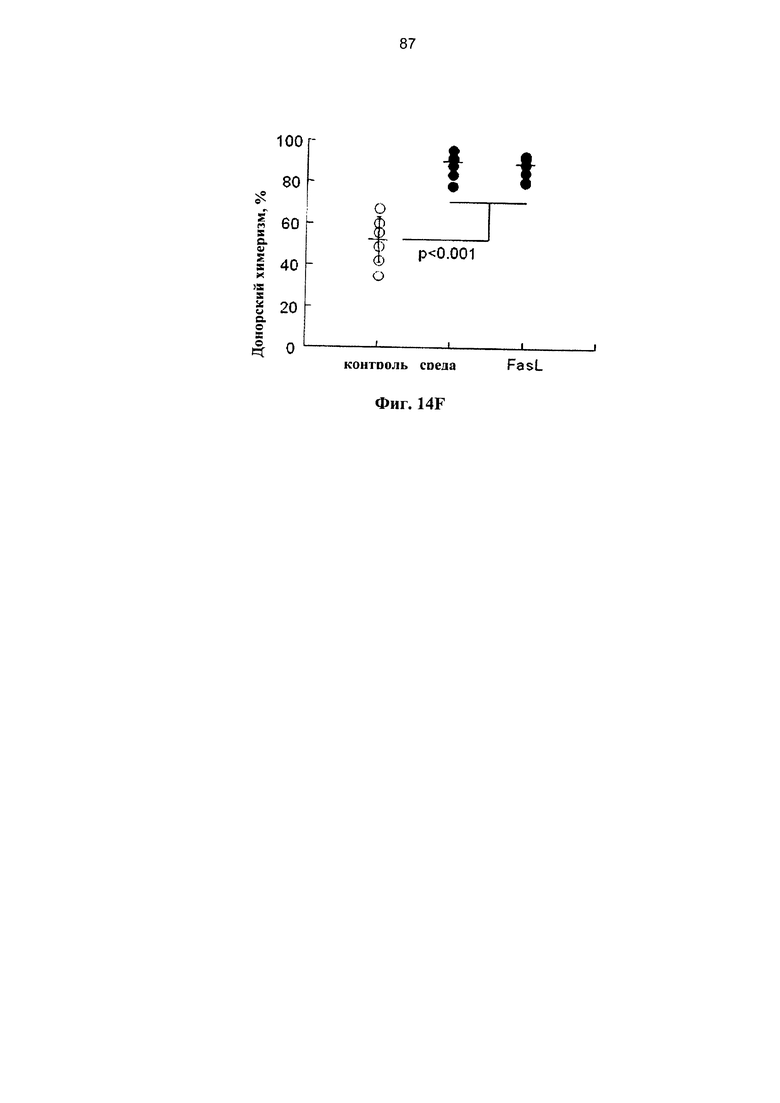
Группа изобретений относится к области биохимии. Предложен способ, устройство и набор для отбора клеток, а также способ улучшения клинического исхода трансплантации гемопоэтических стволовых клеток и клеток-предшественников, способ удаления злокачественной клетки в композиции, содержащей трансплантат из клеток-предшественников, и способ предотвращения болезни «трансплантат против хозяина» с сохранением эффекта «трансплантат против опухоли», причем все предложенные способы осуществляют с использованием вышеуказанного устройства для отбора клеток. Устройство для отбора клеток содержит контейнер из биосовместимого материала и биологически активный апоптоз-индуцирующий лиганд. Причем указанный лиганд иммобилизован на внутренней поверхности контейнера или на поверхности находящихся внутри контейнера микрогранул. Набор содержит вышеуказанное устройство и раствор для поддержания сохранности и активности апоптоз-индуцирующего лиганда. Все предложенные способы осуществляют с использованием вышеуказанного устройства для отбора клеток. Изобретения обеспечивают эффективный способ селективной деплекции Т-клеток, которые индуцируют болезнь «трансплантат против хозяина», с сохранением субпопуляций клеток, которые поддерживают эффект «трансплантат против опухоли» и приживление трансплантата. 6 н. и 26 з.п. ф-лы, 74 ил., 4 пр.
1. Устройство для отбора клеток, содержащее: контейнер, состоящий из биосовместимого материала, и биологически активный апоптоз-индуцирующий лиганд, иммобилизованный на поверхности, притом что указанное устройство выполнено с возможностью отбора клеток посредством апоптоза чувствительных к апоптозу клеток, где указанный иммобилизованный апоптоз-индуцирующий лиганд представлен группой, включающей фактор некроза опухоли α (ФНО-α), Fas-лиганд (FasL), Trail, Tweak или любую их комбинацию, и где указанная поверхность является внутренней поверхностью указанного контейнера или поверхностью микрогранул, находящихся внутри указанного контейнера.
2. Устройство по п. 1, отличающееся тем, что указанный контейнер представлен группой, включающей пакет, колонку, трубку, бутылку, пузырек и флакон.
3. Устройство по п. 1, отличающееся тем, что указанный биосовместимый материал представлен группой, включающей: полипропилен, полистирол, силикон, поливинилхлорид или их комбинацию.
4. Устройство по п. 1, отличающееся тем, что указанный иммобилизованный апоптоз-индуцирующий лиганд представляет собой Fas-лиганд (FasL).
5. Способ отбора клетки, устойчивой к апоптотическим сигналам, из клеточной популяции, при котором указанная клеточная популяция содержит клетку, устойчивую к апоптотическим сигналам, и клетку, чувствительную к апоптотическим сигналам, включающий этапы, на которых:
(a) образец, содержащий клеточную популяцию, вводят в устройство по п. 1; и
(b) указанные клетки инкубируют внутри указанного устройства, причем инкубацию проводят до момента выживания клеток, устойчивых к апоптозу, и апоптотической гибели типов клеток, чувствительных к апоптозу.
6. Способ по п. 5, отличающийся тем, что указанная клетка, устойчивая к апоптотическим сигналам, включает стволовую клетку, иммунокомпетентную клетку, нечувствительную к гибели клеток, индуцированной активацией (AICD), клетку-предшественник или любую их комбинацию.
7. Способ по п. 6, отличающийся тем, что указанную стволовую клетку выбирают из группы, включающей стволовую клетку пуповинной крови, мобилизованную стволовую клетку периферической крови, стволовую клетку костного мозга и нейральную стволовую клетку.
8. Способ по п. 6, отличающийся тем, что указанная стволовая клетка представляет собой стволовую клетку костного мозга.
9. Способ по п. 6, отличающийся тем, что указанная иммунокомпетентная клетка, нечувствительная к гибели клеток, индуцированной активацией, представляет собой Т-клетку.
10. Способ по п. 5, отличающийся тем, что указанную клеточную популяцию получают из: костного мозга, периферической крови после мобилизации клеток-предшественников или пуповинной крови (ПК).
11. Способ по п. 5, отличающийся тем, что указанная инкубация представляет собой инкубацию от 2 часов до 72 часов.
12. Набор для отбора клеток, содержащий: (а) устройство для отбора клеток по п. 1; и (b) раствор для поддержания сохранности и активности апоптоз-индуцирующего лиганда внутри указанного устройства.
13. Набор по п. 12, который дополнительно содержит дополнительный апоптоз-индуцирующий лиганд.
14. Набор по п. 12, отличающийся тем, что указанный раствор представляет собой буфер или среду.
15. Набор по п. 13, отличающийся тем, что указанный апоптоз-индуцирующий лиганд, находящийся внутри указанного контейнера, представляет собой фактор некроза опухолей α (ФНО-α), Fas-лиганд (FasL), Trail, Tweak или любую их комбинацию.
16. Способ улучшения клинического исхода трансплантации гемопоэтических стволовых клеток и клеток-предшественников (HSPC), включающий этапы, на которых:
(a) получают образец, содержащий клеточную популяцию, причем указанная клеточная популяция содержит стволовые и клетки-предшественники;
(b) обеспечивают контакт указанной клеточной популяции с апоптоз-индуцирующим лигандом и инкубируют указанную клеточную популяцию с апоптоз-индуцирующим лигандом, причем указанные контактирование и инкубацию осуществляют с использованием устройства по п. 1;
(c) выделяют выжившие клетки; и
(d) выделенные на этапе (с) клетки используют для трансплантации; и
посредством этого улучшают клинический исход трансплантации гемопоэтических стволовых клеток и клеток-предшественников (HSPC).
17. Способ по п. 16, отличающийся тем, что указанную клеточную популяцию получают из: костного мозга, периферической крови после мобилизации клеток-предшественников или пуповинной крови (ПК).
18. Способ по п. 16, отличающийся тем, что указанную стволовую клетку выбирают из группы, включающей стволовую клетку пуповинной крови, мобилизованную стволовую клетку периферической крови, стволовую клетку костного мозга и нейральную стволовую клетку.
19. Способ по п. 16, отличающийся тем, что указанная стволовая клетка представляет собой стволовую клетку костного мозга.
20. Способ по п. 16, отличающийся тем, что указанный апоптоз-индуцирующий лиганд представляет собой Fas-лиганд (FasL) и фактор некроза опухолей α (ФНО-α).
21. Способ по п. 20, отличающийся тем, что указанную клеточную популяцию инкубируют с Fas-лигандом (FasL) в течение периода от 1 до 24 часов.
22. Способ по п. 20, отличающийся тем, что указанную клеточную популяцию инкубируют с фактором некроза опухолей α (ФНО-α) в течение периода от 24 до 48 часов.
23. Способ по п. 16, отличающийся тем, что указанные выделенные клетки используют либо для аутологичных, аллогенных, либо гаплоидентичных трансплантаций.
24. Способ по п. 16, отличающийся тем, что клетки, выделенные на этапе (с), дополнительно содержат иммунокомпетентную клетку, нечувствительную к гибели клеток, индуцированной активацией (AICD).
25. Способ по п. 24, отличающийся тем, что указанная иммунокомпетентная клетка, нечувствительная к гибели клеток, индуцированной активацией, представляет собой Т-клетку.
26. Способ удаления злокачественной клетки в композиции, содержащей трансплантат, состоящий из клеток-предшественников, включающий этапы, на которых:
(a) получают композицию, содержащую трансплантат, состоящий из клеток-предшественников; и
(b) обеспечивают контакт указанной композиции с апоптоз-индуцирующим лигандом, где указанное контактирование осуществляют с использованием устройства по п. 1, причем указанное контактирование осуществляют до момента удаления злокачественных клеток из композиции и выживания клеток-предшественников, устойчивых к апоптозу.
27. Способ по п. 26, отличающийся тем, что указанный апоптоз-индуцирующий лиганд представляет собой Fas-лиганд (FasL).
28. Способ по п. 26, отличающийся тем, что указанный трансплантат, состоящий из клеток-предшественников, инкубируют с указанным лигандом в течение периода около 24 часов.
29. Способ по п. 26, отличающийся тем, что указанный трансплантат, состоящий из клеток-предшественников, используют в качестве аутологичного трансплантата.
30. Способ предотвращения болезни «трансплантат против хозяина» (БТПХ) с сохранением эффекта «трансплантат против опухоли» (ТПО), включающий этапы, на которых:
(a) получают образец, содержащий клеточную популяцию, причем указанная клеточная популяция содержит HSPC и иммунокомпетентные клетки;
(b) обеспечивают контакт указанной клеточной популяции с апоптоз-индуцирующим лигандом, где указанное контактирование осуществляют с использованием устройства по п. 1;
(c) выделяют выжившие клетки; и
(d) выделенные на этапе (с) клетки используют для трансплантации; и
посредством этого предотвращают болезнь «трансплантат против хозяина» (БТПХ) с сохранением эффекта «трансплантат против опухоли» (ТПО).
31. Способ по п. 30, отличающийся тем, что указанный апоптоз-индуцирующий лиганд представляет собой Fas-лиганд (FasL).
32. Способ по п. 30, отличающийся тем, что указанные клетки инкубируют с указанным лигандом в течение 2-16 часов.
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| SASHCHENKO L.P | |||
| ET AL., Cytotoxic T lymphocytes carrying a pattern recognition protein Tag7 can detect evasive, HLA- negative but Hsp70-exposing tumor cells, thereby ensuring FasL/ Fas-mediated contact killing // BLOOD, 15.09.2007, vol | |||
| Прибор, автоматически записывающий пройденный путь | 1920 |
|
SU110A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| МИКРООРГАНИЗМЫ И КЛЕТКИ ДЛЯ ДИАГНОСТИКИ И ЛЕЧЕНИЯ ОПУХОЛЕЙ | 2002 |
|
RU2357754C2 |

