Данная заявка притязает на преимущества приоритета по Американской предварительной заявке 61/193,137, зарегистрированной 30 октября 2008 года и 61/213,482, зарегистрированной 12 июня 2009 года, каждая из которых полностью включена здесь путем отсылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение, в некоторых его вариантах, относится к не индуцирующим заболевание «трансплантат против хозяина» (GVHD) клеткам против третьей стороны, имеющим фенотип Т-лимфоцитов центральной памяти, и, в частности, к их получению и применению в трансплантации и лечении заболеваний.
УРОВЕНЬ ТЕХНИКИ
Трансплантация костного мозга (ВМ) предлагает радикальное лечение для многих пациентов с гемобластозами и другими гематологическим нарушениям. Однако, трансплантат ВМ содержит Т-клетки донора, которые реагируют на антигены клетки-хозяина (Ag) и вызывают мультисистемное заболевание «трансплантат против хозяина» (GVHD). В начале 80-х годов трансплантация костного мозга (ВМТ) без вредного действия GVHD была продемонстрирована в идентичных по гаплотипу (три несовпадающих локуса HLA) условиях у пациентов с тяжелым комбинированным иммунодефицитом (SCID). Проблема GVHD, которая является почти всегда летальной в таких условиях, была полностью предотвращена истощением Т-клеток.
Однако, у пациентов с лейкозом клинические результаты истощенного Т-клетками ВМ оказались неутешительными, так как эффект предотвращения GVHD был скомпенсирован заметно увеличенной скоростью отторжения трансплантата. Было показано, что отторжение опосредовано устойчивыми к химиолучевой терапии Т-клетками хозяина [Reisner et al., Proc Nati Acad Sci USA. (1986) 83:4012-4015]. Одним из способов решения этой проблемы является выполнение ВМТ после сверхлетального кондиционирования и функциональной инактивации Т-клеток хозяина с помощью иммуносупрессивных средств. Тем не менее, данной стратегии препятствуют оппортунистические инфекции вследствие медленного восстановления иммунитета и значительной токсичности иммуносупрессантов.
Такая смертность, связанная с процедурой трансплантации, может быть допустима для пациентов с высокой степенью риска лейкоза, в отличие от пациентов с ожидаемой продолжительной жизнью. Таким образом, оправданным является использование кондиционирования с пониженной интенсивностью, с менее тяжелым подавлением иммунитета для облегчения приживления истощенного Т-клетками трансплантата ВМ (TDBM), что связано с пониженным риском GVHD. Установление донорского химеризма при таком пониженном кондиционировании является самой желательной целью в трансплантационной биологии, так как в целом это связано с продолжительной толерантностью по отношению к клеткам или тканям оригинального донора. Тем не менее, заметные уровни иммуноцитов хозяина, выдерживающих умеренные подготовительные режимы, являются сложным препятствием для приживления донорских клеток.
Одним подходом к преодолению отторжения аллогенных гематопоэтических стволовых клеток является использование больших доз клеток ВМ. Впервые на моделях грызунов было продемонстрировано, что «мегадоза» трансплантата TDBM может победить опосредованное Т-клетками отторжение трансплантата [Lapidot et al., Blood (1989) 73:2025-2032; Bachar-Lustig et al., Nat Med. (1995) 1:1268-1273; Uharek et al., Blood (1992) 79:1612-1621]. Однако, значительное увеличение в инокуляте ВМ было трудно достигнуть у человека. Для решения этой проблемы гранулоцитарный колониестимулирующий фактор (G-CSF), который способствует мобилизации гемопоэтических стволовых клеток (HSC, CD34+клетки человека) из ВМ, использовали для увеличения выхода HSC, собранных из крови, и данные HSC были добавлены к обычному TDBM [Aversa et al., N Engi J Med. (1998) 339:1186-1193; Aversa et al., J Clin Oncol. (2005) 23:3447-3454; Reisner and Martelli, Immunol Today (1999) 20:343-347; Handgretinger et al., Bone Marrow Transplant. (2001) 27:777-783].
Трансплантаты с «мегадозой» CD34 подняли интересные вопросы относительно того, как эти клетки преодолевают барьер, показанный предшественниками цитотоксических Т-лимфоцитов (CTL-p) хозяина. На данный вопрос был получен ответ, отчасти, с помощью открытия того, что клетки во фракции CD34 наделены сильной вето-активностью [Gur et al.. Blood (2005) 105:2585-2593; Gur et al., Blood (2002) 99:4174-4181; Rachamim et al., Transplantation (1998) 65:1386-1393]. Было обнаружено, что другие типы клеток также опосредуют вето-активность, включающие Т-лимфоциты (например, CD8+ CTL), естественные клетки-киллеры и дендритные клетки. Прямое сравнение вето реактивности разных типов клеток показало что CTL имеют самый сильный вето эффект [Reich-Zeliger et al., J Immunol. (2004) 173:6654-6659].
Один подход, разработанный для генерации вето CTL без GVH реактивности, был описан Reisner и сотрудниками, в котором CTL были стимулированы против сторонних стимуляторов в отсутствии экзогенного IL-2. Данный подход был основан на наблюдении, что только активированные CTLp были способны перенести депривацию IL-2 в первичной культуре. Было показано, что данный способ in vitro и in vivo уменьшает GVH реактивность вето против третьей стороны CTL [PCT Publication No. WO 2001/049243, Bachar-Lustig et al., Blood. 2003; 102:1943-1950; Aviner et al., Hum Immunol. (2005) 66:644-652]. Введение этих вето против третьей стороны CTL реципиенту (вместе с трансплантатом) предотвращало отторжение трансплантата без индуцирования GVHD (публикация заявки PCT WO 2001/049243).
Предусмотрены разные подходы к трансплантации без отторжения трансплантата и/или заболевания «трансплантат против хозяина», некоторые обобщены далее.
Публикация заявки PCT WO 2007/023491 раскрывает применение толерогенных клеток для уменьшения или предотвращения отторжения несингенного трансплантата у субъекта. Раскрытые толерогенные клетки (например, CD4+ CD25+ клетки) могут быть получены от любого донора, который является несингенным как по отношению субъекту, так и по отношению к трансплантату («сторонние» толерогенные клетки). Трансплантат (например, костный мозг) может быть получен от любого донора, который является аллогенным или ксеногенным по отношению к субъекту.
Публикация заявки PCT W02002/102971 раскрывает применение культивированных гематопоэтических клеток-предшественников (НРС), имеющих повышенную вето-активность для индукции толерантности к трансплантату, пересаженному от донора реципиенту. Раскрытые толерогенные клетки предпочтительно экспрессируют CD33 и вводятся до, одновременно или после трансплантации (например, трансплантата клетки или органа).
Публикация заявки PCT WO 2002/043651 раскрывает применение не индуцирующей GVHD популяции иммунных эффекторных клеток для лечения заболевания. Для получения не индуцирующей GVHD популяции иммунных эффекторных клеток первая клеточная популяция (например, Т-лимфоциты) совместно культивируются со второй клеточной популяцией, являющейся несингенной по отношению к субъекту и несингенной по отношению к первой клеточной популяции (например, EBV-инфицированные В-лимфоциты) в условиях, которые включают недостаточность IL-2 с последующим добавлением IL-2. Полученные иммунные эффекторные клетки могут быть использованы для лечения заболеваний, таких как злокачественные опухоли, вирусные заболевания и аутоиммунные заболевания.
Патент US 6,759,035 раскрывает способы ингибирования отторжения трансплантата и индукцию толерантности Т-клеток в трансплантате солидного органа реципиента. Раскрытые способы включают удаление мононуклеарных клеток периферической крови (РВМС) из донора и реципиента, культивирование клеток донора и реципиента вместе в присутствии соединения, которое индуцирует супрессорную активность Т-клеток (например, TGF-β, IL-15 и IL-2), и введение реципиенту супрессорных Т-клеток реципиента вместе с трансплантатом для предотвращения гибели донорских клеток Т-клетками реципиента, индуцируя таким образом толерантность и продолжительную выживаемость трансплантата.
Патент US 6,803,036 раскрывает способы обработки клеток донора для ослабления заболевания «трансплантат против хозяина» у реципиента-пациента. Раскрытые способы включают удаление РВМС из донора и обработку клеток супрессивной композицией (например, IL-10, IL-2, IL-4, IL-15 и TGF-β) в течение времени, достаточного для индукции толерантности Т-клеток. Клетки затем вводятся реципиенту-пациенту. Обработанные клетки могут быть добавлены в стволовые клетки донора до введения пациенту.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Согласно одному аспекту некоторых вариантов настоящего изобретения обеспечена изолированная популяция клеток, включающая не индуцирующие GVHD клетки против третьей стороны, имеющие фенотип Т-лимфоцитов центральной памяти (Tcm), при этом клетки представляют собой клетки, индуцирующие толерантность и способные к хомингу в лимфоузлы после трансплантации.
Согласно одному аспекту некоторых вариантов настоящего изобретения, обеспечено применение изолированной популяции клеток по п.1 для лечения заболевания у субъекта, нуждающегося в этом.
Согласно одному аспекту некоторых вариантов настоящего изобретения, обеспечено применение изолированной популяции клеток по п.1 в качестве адъювантного лечения при проведении трансплантации клеток или тканей субъекту, при этом субъект нуждается в клеточной или тканевой трансплантации.
Согласно одному аспекту некоторых вариантов настоящего изобретения обеспечен способ лечения субъекта, нуждающегося в клеточной или тканевой трансплантации, при этом способ включает: (а) трансплантацию клеток или органа субъекту; и (b) введение субъекту терапевтически эффективного количества изолированной популяции клеток по п.1, производя тем самым лечение субъекта.
Согласно некоторым вариантам изобретения способ, кроме того, включает кондиционирование субъекта в сублетальных, летальных или сверхлетальных условиях перед трансплантацией.
Согласно одному аспекту некоторых вариантов настоящего изобретения, обеспечен способ генерации изолированной популяции клеток по п.1, при этом способ включает: (а) контактирование несингенных мононуклеарных клеток периферической крови (РВМС) со сторонним антигеном или антигенами в условиях, которые делают возможной элиминацию GVH реактивных клеток; и (b) культивирование клеток, полученных на стадии (а), в присутствии IL-15 в условиях, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm).
Согласно некоторым вариантам изобретения, условия, которые делают возможной элиминацию GVH реактивных клеток, включают культуру, не содержащую цитокины.
Согласно некоторым вариантам изобретения, условия, которые делают возможной элиминацию GVH реактивных клеток, включают, по меньшей мере, 2 дня культивирования.
Согласно некоторым вариантам изобретения, условия, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm), включают IL-7 и/или IL-21.
Согласно некоторым вариантам изобретения, условия, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm), включают среду, свободную от антигена.
Согласно некоторым вариантам изобретения, условия, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm), включают, по меньшей мере, 14 дней культивирования.
Согласно некоторым вариантам изобретения, посторонний антиген или антигены являются выбранными из группы, состоящей из сторонних клеток, клеточного антигена, вирусного антигена, бактериального антигена, белкового экстракта, очищенного белка и синтетического пептида, представленного аутологичными или неаутологичными презентирующими клетками.
Согласно некоторым вариантам изобретения, сторонние клетки являются аллогенными или ксеногенными клетками по отношению к субъекту и/или донору.
Согласно некоторым вариантам изобретения, аллогенные клетки являются стимулирующими клетками, выбранными из группы, состоящей из клеток, очищенных из лимфоцитов периферической крови, селезенки или лимфоузлов, цитокин-активированных лимфоцитов периферической крови (PBL) и размноженных in vitro антигенпрезентирующий дендритных клеток (АРС).
Согласно некоторым вариантам изобретения, несингенные мононуклеарные клетки периферической крови (РВМС) являются аллогенными по отношению к субъекту.
Согласно некоторым вариантам изобретения, несингенные мононуклеарные клетки периферической крови (РВМС) являются ксеногенными по отношению к субъекту.
Согласно некоторым вариантам изобретения, заболевание является выбранным из группы, состоящей из злокачественной опухоли, вирусного заболевания и аутоиммунного заболевания.
Согласно некоторым вариантам изобретения, изолированная популяция клеток вводится до, одновременно или после клеточного или тканевого трансплантата.
Согласно некоторым вариантам изобретения, фенотип Т-лимфоцитов центральной памяти (Tcm) включает CD8+, CD62L+, CD45RA-, CD45 RO+ сигнатуру.
Согласно некоторым вариантам изобретения, фенотип Т-лимфоцитов центральной памяти (Tcm) включает CD8+/CD62L+/CD45RO+/L-целестин+/CD45RA-.
Согласно некоторым вариантам изобретения, по меньшей мере, 50% изолированной популяции клеток имеет сигнатуру.
Согласно некоторым вариантам изобретения, лимфоузлы включают периферические лимфоузлы.
Согласно некоторым вариантам изобретения, лимфоузлы включают мезентериальные лимфоузлы.
Согласно некоторым вариантам изобретения, субъектом является человек, Согласно некоторым вариантам изобретения, клеточный или тканевый трансплантат получен от донора, выбранного из группы, состоящей из HLA-идентичного аллогенного донора, HLA-неидентичного аллогенного донора и ксеногенного донора.
Согласно некоторым вариантам изобретения, субъектом и донором является человек.
Согласно некоторым вариантам изобретения, изолированную популяцию клеток, содержащую не индуцирующие GVHD клетки против третьей стороны, имеющие фенотип Т-лимфоцитов центральной памяти (Tcm), генерируют с помощью:
(а) контактирования несингенных мононуклеарных клеток периферической крови (РВМС) со сторонним антигеном или антигенами в условиях, которые делают возможной элиминацию GVH реактивных клеток; и (b) культивирования клеток, полученных на стадии (а), в присутствии IL-15 в условиях, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm).
Согласно некоторым вариантам изобретения, условия, которые делают возможной элиминацию GVH реактивных клеток, включают культуру, не содержащую цитокины.
Согласно некоторым вариантам изобретения, условия, которые делают возможной элиминацию GVH реактивных клеток, включают, по меньшей мере, 2 дня культивирования.
Согласно некоторым вариантам изобретения, условия, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm), включают IL-7 и/или IL-21.
Согласно некоторым вариантам изобретения, условия, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm), включают, по меньшей мере, 14 дней культивирования.
Согласно некоторым вариантам изобретения, условия, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm), включают среду, свободную от антигена.
Согласно некоторым вариантам изобретения, посторонний антиген или антигены являются выбранными из группы, состоящей из сторонних клеток, клеточного антигена, вирусного антигена, бактериального антигена, экстракта белка, очищенного белка и синтетического пептида, представленного аутологичными или неаутологичными представляющими клетками.
Согласно некоторым вариантам изобретения, сторонние клетки являются аллогенными или ксеногенными клетками по отношению к субъекту и/или донору.
Согласно некоторым вариантам изобретения, аллогенные или ксеногенные клетки являются стимулирующими клетками, выбранными из группы, состоящей из клеток, очищенных из лимфоцитов периферической крови, селезенки или лимфоузлов, цитокин-активированных лимфоцитов периферической крови (PBL) и размноженных in vitro антигенпрезентирующий дендритных клеток (АРС)
Согласно некоторым вариантам изобретения, несингенные мононуклеарные клетки периферической крови (РВМС) являются аллогенными по отношению к субъекту.
Согласно некоторым вариантам изобретения, несингенные мононуклеарные клетки периферической крови (РВМС) являются ксеногенными по отношению к субъекту.
Согласно некоторым вариантам изобретения, изолированная популяция клеток является генерированной с помощью: (а) контактирования несингенных мононуклеарных клеток периферической крови (РВМС) с посторонним антигеном или антигенами, по меньшей мере, в течение двух дней в культуре, не содержащей цитокины, что делает возможной элиминацию GVH реактивных клеток; и (b) культивирования клеток, полученных на стадии (а), в присутствии IL-15 в течение, по меньшей мере, 14 дней в среде, свободной от антигена, что делает возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm).
Согласно некоторым вариантам изобретения, культивирование клеток, полученных на стадии (а), включает IL-7 и/или IL-21.
Если не указано иное, все технические и/или научные термины, использованные здесь, имеют такое же значение, которое понимается специалистами в области, к которой относится изобретение. Хотя способы и материалы, аналогичные или эквивалентные описанным здесь, могут применяться при осуществлении или проверке вариантов изобретения, ниже представлено описание иллюстративных способов и/или материалов. В случае противоречия, описание к патенту, включающее определения, будет являться контролем. К тому же, материалы, способы и примеры являются только иллюстративными и не рассматриваются в качестве обязательного ограничения.
КРАТКОЕ ОПИСАНИЕ ФИГУР
В качестве примера дано описание некоторых вариантов изобретения со ссылкой на прилагающиеся фигуры. При более подробном обращении к фигурам подчеркивается, что показанные подробности приведены только в качестве примера идля иллюстративного обсуждения вариантов изобретения. Благодаря этому, описание, взятое вместе с фигурами, делает очевидным для опытных специалистов в данной области осуществление вариантов изобретения на практике.
На фигурах:
Фиг.1A-D представляют собой фотографии, отражающие паттерны хоуминга вето CTL против третьей стороны в сравнении с сигненными и аллогенными наивными Т-клетками через 36 часов после инокуляции. Летально облученные мыши FVB (Н-2q) получали 5×106 истощенного Т-клетками аллотрансплантата костного мозга мышей BALB/C (Н-2d) (Фигура 1А) и 104 сингенных Т-клеток (Фигуры 1C-D). Дополнительно, реципиентная мышь получала 2×106 сингенных DIR-меченных наивных НТС (Фигура 1В), 7×106 наивных аллогенных DIR-меченных Т-клеток (Фигура 1C) или 7×106 DIR-меченных аллогенных активированных вето CTL против третьей стороны (Фигура ID).
Фиг.2А-В представляют собой диаграммы, показывающие, что депривация цитокинов с короткой цепью т vitro способствует индукции Т-клеток центральной памяти (Tcm) с помощью IL-15. Спленоциты мышей BALB/C или СВ6 стимулировали облученными спленоцитами мышей FVB в течение 60 часов (2 дней) или 6 дней в отсутствии цитокинов и оценивали на экспрессию CD62L CD8+ Т-клетками с использованием FACS-анализа (Фигура 2А). Последовательно проводили положительную селекцию CD8+ Т-клеток, культивировали с rhIL-2 (40 IU/мл) и реактивировали спленоцитами мышей FVB, rhIL-2 (40 IU/мл) или rhIL-15 (20 нг/мл) в Ag-свободной среде. Свежую среду и цитокины добавляли через день. На 15 день культивирования клетки оценивали на процентное содержание Tcm клеток (CD44+CD62L+) с использованием FACS-анализа (Фигура 2В).
Фиг.2С представляет собой блок-схему, на которой показаны стадии, выполненные для индукции фенотипа Tcm клеток настоящего изобретения. Мышиные спленоциты стимулировали облученными посторонними стимуляторами в течение 60 часов без добавления цитокинов. Последовательно проводили положительную селекцию выживших CD8+ Т-клеток и затем культивировали с rhIL-15 (20 нг/мл) в свободной от антигена среде в течение 14 дней. Через 14 дней культура содержала примерно 70% Tcm клеток, как показано на Фигуре 2 В.
На Фиг.3А-В представлены фотографии, отражающие присутствие CD8+ CD62L ТСМ клеток в лимфоузлах хозяина и в более высоком количестве по сравнению с вето CTL клетками (через 60 часов после адоптивной трансплантации). Летально облученные мыши BALB/C (Н-2d) получали 5×106 аллотрансплантата костного мозга мышей nude C57BL/6 (Н-2b) и 104 сингенных Т-клеток. Мышам затем трансплантировали 10×106 DIR-меченных CD8+ ТСМ (Фигура 3А) или вето CTL клетки (Н-2bd, Фигура 3В). Через 60 часов выбранных реципиентов умерщвляли. Изображения сделаны ex-vivo с использованием IVIS.
Фиг.4A-D представляют собой диаграммы, отражающие обнаружение CD8+ CD62L+ ТСМ клеток как в периферических, так и в мезентриальных лимфоузлах, и в более высоком количестве по сравнению с вето CTL клетками (через 60 часов после адоптивной трансплантации). Летально облученные мыши BALB/C (Н-2d) получали 5×106 аллотрансплантата костного мозга мышей nude C57BL/6 (Н-2b) и 104 сингенных Т-клеток. Мышам затем трансплантировали 10×106 DIR-меченых CD8+ ТСМ (Фигуры 4А-В) или вето CTL клетки (H-2bd, Фигура 4C-D). Через два дня выбранных реципиентов умерщвляли, после чего периферические лимфоузлы (Фигуры 4А и 4С) и мезентриальные лимфоузлы (Фигуры 4В и 4D) экстрагировали, и растирали. Клеточные суспензии анализировали с помощью FACS и гейтировали по CD8.
Фиг.5А-С представляют собой фотографии, отражающие присутствие CD62L+ Tcm клеток во всех вторичных лимфоидных органах (SLO) и в высоком количестве, в отличие от CD8+ вето CTL клеток, которые сконцентрированы главным образом в печени и селезенке (через 6.5 дней после адоптивной трансплантации). Летально облученные мыши BALB/C (Н-2d) получали 5×106 аллотрансплантата костного мозга мышей nude C57BL/6 (Н-2b) и 104 сингенных Т-клеток (контроль. Фигура 5А). Мышам затем трансплантировали 10×106 DIR-меченных CD8+ ТСМ (Фигура 5 В) или вето CTL клетки (Н-2bd, Фигура 5С). Через 6.5 дней выбранных реципиентов умерщвляли и изображения делали ex-vivo с использованием IVIS.
Фиг.6А-K представляют собой диаграммы, отражающие присутствие CD62L+ Tcm клеток во всех SLO и в высоком количестве, в отличие от CD8+ вето CTL клеток (через 6.5 дней после адоптивной трансплантации). Летально облученные мыши BALB/C (Н-2d) получали 5×10 аллотрансплантата костного мозга мышей nude C57BL/6 (Н-2b) и 104 сингенных Т-клеток. Мышам затем трансплантировали 10×106 DIR-меченных CD8+ ТСМ (Фигуры 6А-Е) или вето CTL клетки (Н-2bd, Фигуры 6F-J). Через 6.5 дней выбранных реципиентов умерщвляли, внутренние органы экстрагировали, и очищенные клетки анализировали с использованием FACS. Числа отражают общее содержание Н-2bd клеток, гейтированных по CD8, собранных за фиксированное время. Общее количество Tcm или CTL, собранных из всех тестируемых органов через 2 дня (черные столбики) или 6.5 дней (серые столбики) после трансплантации, показаны на Фигуре 6K.
Фиг.7 представляет собой диаграмму, отражающую использование адоптивного переноса Tcm клеток для преодоления отторжения истощенных Т-клетками аллотрансплантатов костного мозга (TDBM). Летально облученные мыши С3Н (Н-2K) получали 1.25×104 сингенных Т-клеток. Мышам затем трансплантировали 3×10 клеток ВМ мышей nude BALB/C (Н-2d) с добавлением или без добавления 107 CD8+ ТСМ клеток мышей (H-2kd) (C3H*BALB/C)F1.
Фиг.8 представляет собой диаграмму, отражающую процент донорского химеризма (через 80 дней после трансплантации ВМ). Летально облученным мышам С3Н (Н-2K) трансплантировали 3×106 аллотрансплантата ВМ мышей nude BALB/C (Н-2d) (колонка А), или получали дополнительно 1.25×104 сингенных Т-клеток и 107 CD8+ ТСМ (Н-2kd) мышей (C3H*BALB/C)F1 (колонка В). Через 80 дней после трансплантации процент донорского химеризма в периферической крови мышей анализировали с использованием FACS.
Фиг.9А-В представляют собой диаграммы, отражающие уровни в периферической крови инфузированных Tcm клеток через 80 дней после трансплантации ВМ. Летально облученным мышам С3Н (Н-2K) трансплантировали 3×106 клеток мышей nude BALB/C ВМ (Н-2d) (Фигура 9А), или вводили дополнительно 1.25×104 сингенных Т-клеток и 107 CD8+ ТСМ клеток мышей (Н-2kd) (C3H*BALB/C)F1 (Фигура 9 В). Уровни содержания в периферической крови Tcm клеток анализировали с помощью FACS-анализа, измеряя H2KkH2Dd двойные положительные клетки в CD8+ гейте. Фигуры показывают характерных мышей из каждой группы.
Фиг.10А-С представляют собой диаграммы, на которых показано, что Tcm обладают значительными способностями индуцировать толерантность, и сохраняются in-vivo в течение, по меньшей мере, 1 года после ВМТ. Фигура 10А - Летально облученные (10Gy) мыши С3Н (Н-2K) получали 1.25×104 сингенных НТС. Мышам затем трансплантировали 3×106 клеток ВМ мышей BALB/c-NUDE (Н-2d,"BA-NU ВМ") в присутствии или отсутствии разных доз очищенных CD8+ 'Tcm' (C3HXBALB/c)F1 (Н-2Kd,"C3BF1"). Данные представляют группу только облучения из n=22 животных, группу облучение +ВМ из 30 животных, группу облучение+ВМ+НТС из 59 животных, и следующие группы мышей из n=16, 23 или 18 животных, соответственно: облучение+ВМ+НТС+(2*106) или (5*106) или (107) 'Tcm'. Данные собирали из шеста независимых экспериментов. Фигура 10В - Модель отторжения трансплантата установлена аналогично 9С. Мышам трансплантировали 3×106 клеток ВМ мышей BALB/c-NUDE (H-2d, ”BA-NU ВМ") в присутствии или отсутствии 5×106 очищенных 'Tcm' (C3H*BALB/c)F1 (H-2Kd, “CSBF1") или 107 'CTL' (C3H*BALB/c)F1. Данные представляют группу только облучения из n=14 животных, группу облучение+ВМ из 12 животных, группу облучение+ВМ+НТС из 21 животных и группу из n=23 или 16 мышей, которые получали облучение+ВМ+НТС+(5×106)'Tcm' или (107)'CTL', соответственно. Данные собирали из пяти независимых экспериментов. Фигура 10С - Уровни 'Тст' в периферической крови анализировали через 1 год после ВМТ с использованием FACS, измеряя H2KkH2Dd двойных положительных клеток в CD8+ гейте. Фигура показывает характерную мышь из семи мышей, которым трансплантировали только ВМ ("ВМ только") или семь мышей, которым трансплантировали ВМ+НТС+5×106 CD8+ 'Tcm' (C3H*BALB/c)F1 ("BM+Tcm").
Фиг.11А-С представляют собой диаграммы, на которых показано, что полностью аллогенные 'Tcm' против третьей стороны лишены GVH реактивности и способствуют приживлению аллотрансплантата TDBM. Фигуры 11А-В - Сверхлетально облученные (11Gy) мыши С3Н были радиозащищены 5×106 клетками ВМ мышей BALB/c-NUDE в присутствии или отсутствии 5×106 или 2×106 CD8+ очищенных 'Тст', полученных от мышей BLAB/c, ("ВМ+Tcm 5" или "ВМ+Tcm 2", соответственно), или наивных клеток ("ВМ+наивные 5" или "ВМ+наивные 2", соответственно). GVH реактивность 'Tcm' или наивных клеток выражена процентом выживаемости (Фигура 11А) или средним изменением массы (Фигура 11В) в течение 100 дней после трансплантации. Данные представляют средние показатели трех независимых экспериментов, по меньшей мере, с пятью мышами в каждой группе в каждом эксперименте. (**) - Представляет собой р-величину 0.01, по сравнению с группой мышей, которые получили только ВМ. Фигура 11С - Летально облученные (10Gy) мыши С3Н получали 1.25×104 сингенных НТС. Мышам затем трансплантировали 3×106 клеток ВМ мышей BALB/c-NUDE ("BA-NU КМ") в присутствии или отсутствии 5×106 очищенных 'Tcm' мышей BALB/c CD8+ или 107 'CTL' мышей BALB/c. Данные представляют группу только облучения из n=17 мышей, группу облучение+ВМ из 11 мышей, группу облучение+ВМ+НТС из 24 мышей и группы из n=27 или 25 мышей, которые получали облучение+ВМ+НТС+107 CTLs' или 5×10 'Tcm', соответственно. Данные собирали из трех независимых экспериментов.
Фиг.12А-Е представляют собой диаграммы, на которых показано, что Tcm' проявляют низкую вето-активность in vitro, но при реактивации приобретают эффекторный фенотип, который ассоциирован с сильной и специфичной вето-активностью. Фигура 12А-2 с показывает спленоциты, которые были стимулированы облученными спленоцитами мышей BALB/c (Н-2d) в присутствии или отсутствии очищенных 'Tcm', полученных от мышей СВ6 (Н-2bd), 'CTL' или очищенных 'Tcm', реактивированных in-vitro их когнитивными сторонними FVB стимуляторами в течение 60 часов ("R' Tcm"). Вето клетки добавляли при указанных соотношениях вето/эффектор. Вето-активность анализировали с использованием FACS-анализа через 3 дня после иницирования MLR для наблюдения ингибирования размножения CD8+1B2 2с клеток. Результаты представлены как среднее±Станд. откл. процента ингибирования из пяти независимых экспериментов. Фигура 12В представляет FACS-анализ уровней аннексина V на живых (7AAD-) CD8+1B2+2 с клетках, в конце MLR, засеянные или не засеянные вето клетками при соотношении вето/эффектор, равном 0.2. Результаты представлены как среднее±Станд. откл. процента аннексина V в семи независимых экспериментах. Фигура 12С - MLR была установлена аналогично А. Ингибирование роста клеток 2 с оценивали, когда вето клетки, полученные от "специфичных" мышей СВ6 (Н-2bd, ”spe'") или "неспецифичных" мышей C3B6F1 (Н-2bk,"non-spe'"), добавляли при соотношении вето/эффектор, равном 0.02. Результаты представлены как среднее±Станд. откл. процента ингибирования из четырех независимых экспериментов. Клетки 2 с также анализировали на уровни аннексина V (Фигура 12D). Фигура 12Е - Летально облученные (8Gy) мыши BALB/c получали 4×106 клеток ВМ мышей C57BL/6-NUDE и 1.25×104 сингенных Т-клеток. Мышам затем трансплантировали 2-10×106 очищенных CD8+ 'Tcm' мышей СВ6, которых анализировали на CD62L фенотип перед адоптивным переносом (левая панель, "Tcm"). Мышей умерщвляли через 4 дня после адоптивного переноса, LN собирали и растирали; экспрессию CD62L на CD8+ Т-клетках мышей СВ6, изолированных из LN, анализировали с использованием FACS (правая панель, "LN Tcm"). Показан характерный результат 'Tcm', изолированных из LN одной из 20 тестированных мышей в четырех независимых экспериментах. На Фигурах 12A-F, (***) представляет собой р-величину <0.001, (**) представляет собой р-величину <0.01 и (*) представляет собой р-величину <0.05.
Фиг.13A-D представляют собой диаграммы, показывающие, что Tcm специфично удаляет анти-донорские Т-клетки in-vivo. Фигуры 13А-В - Летально облученные (10Gy) мыши C57BL/6 получали 1×105 очищенных CD8+ 2с клеток и 5×105 облученных спленоцитов мышей BALB/c. На следующий день мыши были трансплантированы 1×106 клетками ВМ мышей C57BL/6-NUDE или получали дополнительно 5×106 "специфичных", очищенных 'Tcm', полученных от мышей СВ6 (черные столбики), или "неспецифичных", полученных от мышей C57BL/6 (серые столбики). Реципиентов умерщвляли через 8 дней после трансплантации, их селезенки собирали, и удаление 2 с Т-клеток регистрировали с помощью FACS. (А) Иллюстративный результат, демонстрирующий уровень выживания 2 с клеток (7AAD-) в отсутствии (левая панель, "2с только") или в присутствии "специфических" 'Tcm' (правая панель "2с + специфические Tcm"). (В) Количественная оценка результатов, измеряющих ингибирование 2 с клеток с помощью "специфических" и "неспецифических" 'Tcm'. Данные представляют среднее±Станд. откл. процента ингибирования, по меньшей мере, от 10 животных в каждой группе, собранных в результате независимых экспериментов. Фигуры 13C-D - Сингенная модель ВМТ была установлена как в А-В, но было введено 5×105 очищенных CD8+ 2с клеток и 2.5×106 облученных спленоцитов мышей BALB/c. Реципиентов умерщвляли через 8 дней после трансплантации, их селезенки собирали и проводили FACS-анализ уровней аннексина V на живых (7AAD-) CD8+1B2+2 с клетках. Фигура 13С - Типичный результат, демонстрирующий уровни аннексина V на 2с клетках в отсутствии (левая панель, "2с только") или присутствии "специфических" 'Tcm' (правая панель "2с + специфические Tcm"). Фигура 13D - Количественная оценка результатов измерения уровней аннексина V на 2с клетках с последующими взаимодействиями с "специфическими" или "неспецифическими" 'Tcm'. Данные представляют среднее±Станд. откл. процентного содержания аннексина V, по меньшей мере, у четырех животных из каждой группы, в одном типичном эксперименте из трех выполненных. (**) Представляет собой р-величину <0.01 (***) Представляет собой р-величину <0.001.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение, в некоторых его вариантах, относится к не индуцирующим GVHD клеткам против третьей стороны, имеющим фенотип Т-лимфоцитов центральной памяти и, в частности, к их генерации и использованию в трансплантации и лечении заболеваний.
Принципы и действие настоящего изобретения можно лучше понять при обращении к чертежам и сопровождающим описаниям.
Перед подробным объяснением, по меньшей мере, одного варианта изобретения должно быть понятно, что изобретение не обязательно ограничивается деталями, изложенными в последующем описании, или приведенными для иллюстрации с помощью Примеров. Возможны другие варианты изобретения, которые можно осуществить практически или выполнить разными способами. Также, должно быть понятно, что фразеология и терминология, которая используется здесь, предназначена для описания и не должна рассматриваться в качестве ограничения.
При осуществлении настоящего изобретения на практике, автор настоящего изобретения раскрывает новую популяцию Т-клеток центральной памяти против третьей стороны (Tcm), которые мигрируют в лимфоузлы после трансплантации и вызывают толерантность без индуцирования реакции «трансплантат против хозяина» (GVH).
Как показано здесь далее и в последующем разделе «Примеры», автор настоящего изобретения генерировал новую популяцию индуцирующих толерантность Tcm клеток вначале воздействием на CD8+ Т-клетки стимулов против третьей стороны в отсутствии цитокинов в течение 2 дней и последующим культивированием полученных клеток в присутствии IL-15 в свободной от антигена среде в течение 2 недель (смотри Фигуру 2С). Tcm клетки, генерированные автором настоящего изобретения, имели фенотип Tcm (например, CD44+CD62L+ экспрессирующие клетки, Фигура 2В) и мигрировали в лимфоузлы после трансплантации (Фигуры 3А-В). Более того, эти клетки были четко видны в лимфоузлах через несколько дней после трансплантации (Фигуры 5В и 6А-Е). Значительный эффект толерантности Tcm клеток настоящего изобретения был очевиден после сотрансплантации истощенного Т-клетками костного мозга (TDBM) с Tcm клетками против третьей стороны реципиенту без дальнейшего лечения (Фигуры 7, 10 и 11). Данные Tcm клетки проявили сильную пролиферацию и продолжительную устойчивость у реципиентов с трансплантатом ВМ (Фигура 6K и Фигуры 9С-Е). Использование Tcm клеток против третьей стороны делает возможным приживление трансплантата без болезни «трансплантат против хозяина» (GVHD), как показано на Фигурах 12 и 13. Взятые вместе, все эти открытия подтверждают применение Tcm клеток против третьей стороны в качестве клеток, способствующих длительному выживанию трансплантатов.
Таким образом, согласно одному аспекту настоящего изобретения, обеспечена изолированная популяция клеток, включающая не индуцирующие GVHD клетки против третьей стороны, имеющие фенотип Т-лимфоцитов центральной памяти (Tcm), клетки, индуцирующие толерантность, и способные к хоумингу в лимфоузлы после трансплантации.
Использующееся здесь выражение «изолированная популяция клеток» относится к клеткам, которые были изолированы из их естественной среды (например, тела человека).
Использующийся здесь термин «не-GVHD» означает практически не имеющие реактивности «трансплантат против хозяина». Таким образом, клетки настоящего изобретения являются генерированными как практически не вызывающие болезнь «трансплантат против хозяина» (GVHD), что подтверждается выживаемостью, весом и общим видом трансплантированных мышей через 100 дней после трансплантации.
Использующееся здесь выражение «клетки против третьей стороны» относится к лимфоцитам (например, Т-лимфоцитам), которые направлены (например, с помощью Т-клеточного распознавания) против постороннего антигена или антигенов.
Использующееся здесь выражение «посторонний антиген или антигены» относится к растворимому или нерастворимому (такому как мембраносвязанный) антигену или антигенам, которые не присутствуют ни у донора, ни у реципиента, как подробно отражено далее.
Например, посторонние антигены могут быть сторонними клетками, антигенами вирусов, такими как, например, Эпштейн-Барр вирус (EBV) или цито-мегало-вирус (CMV), или антигенами бактерий, такими как флагеллин. Вирусные или бактериальные антигены могут быть представлены инфицированными ими клетками (например, клеточной линией) или приготовленными иным способом для экспрессирования вирусных/бактериальных белков. Аутологичные или неаутологичные антигенпрезентирующие клетки могут быть использованы для презентации коротких синтетических пептидов, конденсированных или нагруженных в них. Такие короткие пептиды могут представлять собой пептиды, извлеченные из вируса, или пептиды, представляющие любой другой антиген.
Специализированное программное обеспечение может быть использовано для анализа вирусной или другой последовательностей для идентификации иммуногенетических коротких пептидов, т.е. пептидов, представленных в контексте молекулы МНС класса I или молекулы МНС класса II.
Сторонние клетки могут быть аллогенными или ксеногенными по отношению к реципиенту (более подробное объяснение представлено далее). В случае аллогенных сторонних клеток, такие клетки имеют HLA-антигены, отличные от донорских, но которые не являются перекрестие реактивными с HLA-антигенами реципиента, так, что клетки против третьей стороны, генерированные против таких клеток, не являются реактивными против трансплантата или антигенов реципиента.
Согласно одному варианту настоящего изобретения, аллогенные или ксеногенные сторонние клетки являются стимулирующими клетками, выбранными из группы, состоящей из клеток, очищенных из лимфоцитов периферической крови (PBL), селезенки или лимфоузлов, цитокин-активированных PBL и размноженных in vitro антигенпрезентирующих дендритных клеток (АРС).
Посторонние антигены могут быть представлены на клеточных, вирусных или бактериальных поверхностях, или из них извлекаться и/или очищаться. К тому же, вирусный или бактериальный антиген может быть расположен на инфицированной клетке и клеточный антиген может быть расположен на искусственном носителе, таком как липосома.
К тому же, посторонними антигенами могут являться, например, белки, экстрагированные или очищенные из разных источников. Примером очищенного белка, который может служить в качестве постороннего антигена согласно настоящему изобретению, может являться овальбумин. Другие примеры представляются возможными.
Используемые клетки, вирусы, бактерии, инфицированные вирусом, инфицированные бактериями, вирусные пептиды или бактериальные пептиды, представляющие клетки в качестве посторонних антигенов, являются особенно эффективными, поскольку такие посторонние антигены включают множество различных антигенных детерминант и, таким образом, направляют образование клеток разных популяций против третьей стороны, которые могут затем служить для более быстрого восстановления Т-клеток в случаях, когда такое восстановление требуется, например, после летального или сублетального облучения, или химиотерапии.
Более того, когда клетки против третьей стороны направлены против посторонних антигенов, можно получить, по меньшей мере, некоторую «трансплантат против заболевания" клеточную (например, раковых клеток, такую как «трансплантат против лейкоза») активность благодаря TCR-независимому киллингу, опосредованному LFA1-I/CAM1 связыванием [Arditti et al., Blood (2005) 105(8):3365-71. Epub 2004 Jul 6].
Согласно некоторым вариантам, клетки против третьей стороны настоящего изобретения включают фенотип Т-лимфоцита центральной памяти (Tcm).
Использующееся здесь выражение «фенотип Т-лимфоцита центральной памяти (Tern)» относится к субпопуляции цитотоксических Т-клеток, которые мигрируют в лимфоузлы. У человека клетки, имеющие фенотип Tcm, обычно включают CD8+/CD62L+/CD45RO+/L-селектин+/CD45RA-. Согласно более конкретному варианту, фенотип Tcm включает CD8+/CD62L+/CCR7+/CD45RO+/L-селектин+/CD45KA. Предпочтительно, чтобы Tcm клетки могли экспрессировать все маркеры на одной клетке или могли экспрессировать только часть маркеров на одной клетке.
Предпочтительно, чтобы, по меньшей мере, 30%, по меньшей мере 40%, 50%, по меньшей мере 55%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 100% изолированной популяции клеток содержало клетки, имеющие сигнатуру Тст клетки.
Как отмечалось, Tcm клетки обычно мигрируют в лимфоузлы после трансплантации. Согласно некоторым вариантам, Tcm клетки против третьей стороны настоящего изобретения могут мигрировать в любой из лимфоузлов после трансплантации, например в периферические лимфоузлы и мезентриальные лимфоузлы (как показано подробно в Примере 4 последующего раздела «Примеры»). Природа хоуминга данных клеток позволяет им проявить свой эффект толерантности эффективно и быстро.
Таким образом, Tcm клетки против третьей стороны настоящего изобретения являются индуцирующими толерантность клетками.
Использующееся здесь выражение «индуцирующие толерантность клетки» относится к клеткам, которые вызывают слабую отвечаемость клеток реципиента (например, Т-клеток реципиента) при контакте с ними. Индуцирующие толерантность клетки включают вето клетки (т.е. Т-клетки, которые приводят к апоптозу Т-клеток хозяина при контакте с ними), как было описано ранее в РСТ публикации WO 2001/049243 и WO 2002/102971.
Согласно некоторым вариантам, Tcm клетки настоящего изобретения могут быть наивными клетками (например, негенетически модифицированными) или генетически модифицированными клетками (например, клетками, которые созданы генетически для экспрессирования или не экспрессирования специфических генов, маркеров или пептидов, или для секреции или не секреции специфических цитокинов). Любой известный в данной области способ может быть реализован с помощью генетического инжениринга клеток, например, инактивацией соответствующего гена/ов, или внедрения антисмысловой РНК, препятствующей экспрессии полипептида (смотри, например, WO/2000/039294, который включен здесь в виде ссылки).
Согласно некоторым вариантам изобретения, обеспечен способ получения изолированной популяции клеток, при этом способ включает: (а) контактирование несингенных мононуклеарных клеток периферической крови (РВМС) с посторонним антигеном или антигенами в условиях, которые делают возможной элиминацию GVH реактивных клеток; и (b) культивирование клеток, полученных на стадии (а), в присутствии IL-15 в условиях, которые делают возможной пролиферацию клеток, имеющих фенотип Т-лимфоцитов центральных памяти (Tcm).
Определенные варианты описанного выше способа подробно обеспечены в Примерах 1 и 3 здесь ниже. Таким образом, согласно иллюстративному варианту изобретения, клетки против третьей стороны получают сначала контактированием несингенных мононуклеарных клеток периферической крови (РВМС) с посторонним антигеном или антигенами в культуре, не содержащей цитокины (например, без добавления цитокинов). Данную стадию обычно проводят в течение примерно 2 дней и делают возможной элиминацию GVH реактивных клеток (например, Т-клеток). Затем, клетки против третьей стороны культивируют в присутствии IL-15 в течение примерно 14 дней в свободной от антигена среде. Культуру можно, кроме того, получить в присутствии дополнительных цитокинов, таких как IL-7 и/или IL-21. Данный процесс делает возможной пролиферацию клеток против третьей стороны, имеющих фенотип Т-лимфоцитов центральной памяти (Tcm) и не имеющих GVHD реактивности.
Предпочтительно, чтобы дополнительную стадию, позволяющую произвести селекцию CD8+ клеток, можно было выполнить, например, с использованием магнитных бусинок MACS перед культивированием клеток в присутствии IL-15. Такая стадия может быть полезной для повышения чистоты CD8+ клеток внутри культуры (т.е. элиминации других лимфоцитов внутри культуры клетки, например, CD4+ Т-клеток) или для повышения количества CD8+ Т-клеток.
Согласно некоторым вариантам изобретения, несингенные РВМС настоящего изобретения могут быть аллогенными или ксеногенными по отношению к субъекту (подробное объяснение представлено далее). Источник РВМС будет определяться в отношении предполагаемого использования клеток (смотри дальнейшие детали ниже) и находится в компетенции опытного специалиста в данной области, особенно в свете подробного раскрытия, обеспеченного здесь.
Предпочтительно, чтобы сингенные РВМС (например, от субъекта) могли также быть использованы в соответствии с современными учениями (т.е. в ситуациях, когда сингенные Tcm клетки могут быть полезными для лечения, смотри детали ниже). Синтез таких клеток можно выполнить, как описано подробно для несингенных клеток.
Использование индуцирующих толерантность клеток настоящего изобретения является особенно полезным в ситуациях, когда требуется устранить отторжение трансплантата и преодолеть болезнь «трансплантат против хозяина» (GVHD), например, при трансплантации аллогенных или ксеногенных клеток или тканей.
Таким образом, согласно другому аспекту настоящего изобретения, обеспечен способ лечения субъекта, нуждающегося в клеточной или тканевой трансплантации, при этом способ включает трансплантацию клетки или органа субъекту и введение субъекту терапевтически эффективного количества изолированной популяции клеток.
Использующийся здесь термин «лечение» включает отмену, значительное ингибирование, замедление или реверсию прогрессирования состояния, значительное ослабление клинических или внешних симптомов состояния, или значительное предотвращение появления клинических или внешних симптомов состояния.
Использующийся здесь термин «субъект» или «нуждающийся субъект» относится к млекопитающим, предпочтительно человеку, мужчине или женщине любого возраста, которым требуется клеточная или тканевая трансплантация. Обычно, субъект нуждается в клеточной или тканевой трансплантации (также называемый здесь как реципиент) из-за нарушения патологического или нежелательного состояния, заболевания или синдрома, или физического, морфологического или физиологического отклонения, которое восприимчиво к лечению с помощью клеточной или тканевой трансплантации. Примеры таких нарушений представлены здесь далее.
Использующееся здесь выражение «клеточная или тканевая трансплантация» относится к клетке тела (например, одной клетке или группе клеток) или тканям (например, твердые ткани или мягкие ткани, которые могут быть трансплантированы полностью или частично). Примеры тканей, которые могут быть трансплантированы согласно настоящему изобретению, включают, но без ограничения, ткани печени, поджелудочной железы, селезенки, почки, сердца, легких, кожи, кишечника и лимфоидные/кроветворные ткани (например, лимфоузлы, Пейеровы бляшки тимуса или костный мозг). Примеры клеток, которые могут быть трансплантированы согласно современным учениям, включают, но без ограничения, незрелые кроветворные клетки.
Более того, настоящее изобретение также предполагает трансплантацию целых органов, таких как, например, почки, сердце, печень или кожа.
В зависимости от применения, способ может быть осуществлен с использованием клеток или тканей, которые являются сингенными или несингенными по отношению к субъекту.
Использующийся здесь термин «сингенный» относится к клетке или ткани, которая получена от индивида, который является в значительной степени генетически идентичным субъекту. Обычно, в основном полностью инбредные млекопитающие, клоны млекопитающих или гомозиготные млекопитающие-близнецы являются сингенными.
Примеры сингенных клеток или тканей включают клетки или ткани, полученные от субъекта (также называемые в данной области «аутологическими»), клона субъекта или гомозиготного близнеца субъекта.
Использующийся здесь термин «несингенный» относится к клетке или ткани, которая получена от индивида, который является аллогенным или ксеногенным по отношению к лимфоцитам субъекта.
Использующийся здесь термин «аллогенный» относится к клетке или ткани, которая получена от донора, который принадлежит к такой же линии, что и субъект, но является по существу не клональным в отношении субъекта. Обычно, аутбредные, незиготные млекопитающие-близнецы одной линии являются аллогенными друг с другом. Предпочтительно, чтобы аллогенный донор был HLA - идентичным или HLA-неидентичным в отношении субъекта.
Использующийся здесь термин «ксеногенный» относится к клетке или ткани, которая главным образом экспрессирует антигены разных линий по отношению к линиям основного соотношения лимфоцитов субъекта. Обычно, аутбредные млекопитающие разных линий являются ксеногенными друг с другом.
Настоящее изобретение предусматривает, что ксеногенные клетки или ткани получены от разных линий, например, но без ограничения, крупного рогатого скота (например, корова), непарнокопытных (например, лошадь), свиней (например, поросенок), козьих (например, коза, овца), кошачьих (например, Felis domestica), псовых (например, домашние собаки), грызунов (например, мыши, крысы, кролики, морская свинка, песчанка, хомяк) или приматов (например, шимпанзе, макака-резус, макака, мартышка).
Клетки или ткани ксеногенного происхождения (например, свиного происхождения) предпочтительно получены от источника, свободного от зоонозов, таких как свиной эндогенный ретровирус. Аналогично, клетки или ткани человека предпочтительно получены главным образом от непатогенных источников.
Согласно варианту настоящего изобретения, субъектом и донором является человек.
В зависимости от применения и доступных источников, клетки или ткани настоящего изобретения могут быть получены из пренатального организма, постнатального организма, взрослого или трупного донора. Более того, в зависимости от применения, клетки или ткани могут быть наивными или генетически модифицированными. Такие детерминации находятся в пределах компетенции обычного специалиста в данной области.
Любой известный в данной области способ может использоваться для получения клетки или ткани (например, для трансплантации).
Трансплантацию клетки или ткани субъекту можно осуществить разными способами, в зависимости от разных параметров, таких как, например, тип клетки или ткани; тип, стадия или тяжесть заболевания реципиента (например, органная недостаточность); физических или физиологических параметров, специфичных для субъекта; и/или требуемого терапевтического результата.
Пересадку клеточного или тканевого трансплантата настоящего изобретения можно осуществить путем пересадки клеточного или тканевого трансплантата в любую анатомическую локализацию в зависимости от применения. Клеточный или тканевый трансплантат можно пересадить в гомотопную анатомическую локализацию (нормальная анатомическая локализация для трансплантата) или в эктопическую анатомическую локализацию (патологическая анатомическая локализация для трансплантата). В зависимости от применения, клеточный или тканевый трансплантат можно преимущественно имплантировать под почечную капсулу или в почки, тестикулярную жировую клетчатку, под кожу, сальник, воротную вену, печень, селезенку, полость сердца, сердце, грудную полость, легкие, кожу, поджелудочную железу и/или внутрибрюшинное пространство.
Например, ткань печени согласно настоящему изобретению может быть трансплантирована в печень, воротную вену, под почечную капсулу, под кожу, в сальник, селезенку и внутрибрюшинное пространство. Трансплантация печени в разные анатомические локализации, такие как указанные, широко практикуется в данной области для лечения заболеваний, восприимчивых к лечению с помощью пересадки печени (например, печеночной недостаточности). Аналогично, трансплантация ткани поджелудочной железы по настоящему изобретению может быть успешно выполнена путем пересадки ткани в воротную вену, печень, поджелудочную железу, тестикулярную жировую клетчатку, подкожно, в сальник, кишечную петлю (субсероза U петли тонкого кишечника) и/или внутрибрюшинное пространство. Трансплантация ткани поджелудочной железы может быть использована для лечения заболеваний, восприимчивых к лечению путем пересадки поджелудочной железы (например, диабет). Аналогично, трансплантацию тканей, таких как почки, сердце, легкие или ткани кожи можно осуществить в разные анатомические локализации, описанные выше для лечения реципиентов, страдающих, например, от почечной недостаточности, сердечной недостаточности, легочной недостаточности или повреждения кожи (например, ожоги).
Способ настоящего изобретения можно также использовать, например, для лечения реципиента, страдающего от заболевания, требующего трансплантации незрелых кроветворных клеток.
В последнем случае, незрелые аллогенные или ксеногенные кроветворные клетки (включающие стволовые клетки), которые могут быть получены, например, из костного мозга, мобилизированной периферической крови (путем, например, лейкафереза), фетальной печени, желточного мешка и/или пуповинной крови донора, и которые предпочтительно являются истощенными по Т-клеткам незрельми кроветворными CD34+ клетками, могут быть трансплантированы реципиенту, страдающему от заболевания. Такое заболевание включает, но без ограничения, лейкемию, такую как острый лимфобластный лейкоз (ALL), острый нелимфобластный лейкоз (ANLL), острый миелоцитарный лейкоз (AML) или хронический миелоцитарный лейкоз (CML), тяжелый комбинированный иммунодефицит (SCID), включающий аденозиндеаминазу (ADA), остеопетроз, апластическую анемию, болезнь Гоше, талассемию и другие врожденные или наследственные кроветворные нарушения.
Предпочтительно, чтобы незрелые аллогенные или ксеногенные гематопоэтические клетки настоящего изобретения можно было трансплантировать реципиенту любым способом, известным в данной области для трансплантации клетки, таким как, но без ограничения, инфузия клеток (например, внутривенно I.V.) или внутрибрюшинно.
Факультативно, при трансплантации клеточного или тканевого трансплантата по настоящему изобретению пациенту, имеющему поврежденный орган, может быть предпочтительным вначале, по меньшей мере, частично удалить поврежденный орган у субъекта для того, чтобы обеспечить оптимальное развитие трансплантата и его структурную/функциональную интеграцию с анатомией/физиологией субъекта.
Способ настоящего изобретения также предусматривает сотрансплантацию нескольких органов (например, сердечных и легочных тканей) в случае, если такая процедура может оказать благотворное воздействие на субъекта.
После трансплантации клеточного или тканевого трансплантата субъекту, согласно современным учениям, желательно, в соответствии со стандартной медицинской практикой, наблюдать за ростом функциональных характеристик и иммуносовместимостью органа согласно любой стандартной методике, существующей в данной области. Например, функциональное состояние тканевого трансплантата поджелудочной железы можно наблюдать после трансплантации с помощью стандартных тестов функции поджелудочной железы (например, анализ уровней инсулина в сыворотке). Аналогично, трансплантат ткани печени можно наблюдать после трансплантации с помощью стандартных тестов функции печени (например, анализ уровней альбумина в сыворотке, общего белка, ALT, AST и билирубина, и анализ времени свертывания крови). Структурное развитие клеток или тканей можно наблюдать с помощью компьютерной томографии или ультразвуковой визуализации.
В контексте трансплантации, для способствования приживлению клеточного или тканевого трансплантата способ может, кроме того, успешно включать кондиционирование субъекта иммуносупрессивной терапией до, одновременно или после трансплантации клеточного или тканевого трансплантата.
Примеры приемлемых схем иммуносупрессии включают введение иммуносупрессивных препаратов, вызывающих толерантность клеточных популяций (как подробно описано далее), и/или иммуносупрессивное облучение.
Обширное руководство по выбору и введению приемлемых иммунодепрессивных схем для трансплантации обеспечено в литературе данной области (например, см.: Kirkpatrick CH. and Rowlands DT Jr., 1992. JAMA. 268, 2952; Higgins RM. et al, 1996. Lancet 348, 1208; Suthanthiran M. and Strom ТВ., 1996. New Engl. J. Med. 331, 365; Midthun DE. et al., 1997. Mayo Clin Proc. 72, 175; Momson VA. et al., 1994. Am J Med. 97, 14; Hanto DW, 1995. Annu Rev Med. 46, 381; Senderowicz AM. et al., 1997. Ann Intern Med. 126, 882; Vincenti F. et al, 1998. New Engl. J. Med. 338, 161; Dantal J. et al. 1998. Lancet 351, 623).
Предпочтительно, чтобы иммуносупрессивный режим состоял из введения, по меньшей мере, одного иммуносупрессивного препарата субъекту.
Примеры иммуносупрессивных препаратов включают, но без ограничения, метотрексат, циклофосфамид, циклоспорин, циклоспорин А, хлорохин, гидроксихлорохин, сульфасалазин (сульфасалазопирин), золото-хлористоводородный натрий, D-пеницилламин, лефлуномид, азатиоприн, анакинра, инфликсимаб (РЕМИКЕЙД), этанерцепт, блокаторы TNF-альфа, биологический агент, который воздействует на воспалительный цитокин, и нестероидные противовоспалительные препараты (NSAID). Примеры NSAID включают, но без ограничения, ацетилсалициловую кислоту, магния салицилат-холин, дифлунизал, салицилат магния, салсалат, салицилат натрия, диклофенак, этодолак, фенопрофен, флурбипрофен, индометацин, кетопрофен, кеторолак, меклофенамат, напроксен, набуметон, фенилбутазон, пироксикам, сулиндак, толметин, ацетаминофен, ибупрофен, ингибиторы Сох-2, трамадол, рапамицин (сиролимус) и аналоги рапамицина (такие как CCI-779, RAD001, АР23573). Данные средства могут вводиться индивидуально или в комбинации.
Независимо от типа трансплантата, для предотвращения отторжения трансплантата и болезни «трансплантат против хозяина», способ настоящего изобретения использует новые Tcm клетки против третьей стороны (как описано подробно здесь далее).
Согласно способу настоящего изобретения, данные Tcm клетки против третьей стороны вводятся одновременно, до или после трансплантации клеточного или тканевого трансплантата.
Tcm клетки против третьей стороны могут вводиться любьм способом, известным в области трансплантации клеток, таким как, но без ограничения, инфузия клеток (например, внутривенно I.V.) или внутрибрюшинное введение.
Без привязки к теории, терапевтически эффективное количество представляет собой количество Tcm клеток против третьей стороны, достаточное для толеризации, противоракового действия и/или восстановления иммунитета без индукции GVHD. Так как Tcm клетки настоящего изобретения мигрируют в лимфоузлы после трансплантации, для достижения благоприятного действия/ий клеток (например, толеризации, противоопухолего действия и/или восстановления иммунитета) может потребоваться меньшее количество клеток (по сравнению с дозой клеток, используемых ранее, смотри, например, WO 2001/049243). Следует учесть, что могут потребоваться более низкие уровни иммуносупрессивных препаратов в сочетании с Tcm клетками настоящего изобретения (например, исключение рапамицина из терапевтического протокола).
Определение терапевтически эффективного количества находится в компетенции опытного специалиста в данной области, особенно в свете подробного раскрытия, обеспеченного здесь.
Для любого состава, который используется в способах изобретения, терапевтически эффективное количество или доза может быть установлена на основании анализов in vitro и культуры клеток. Например, доза может быть составлена на животных моделях для достижения требуемой концентрации или титра. Такую информацию можно использовать для более тщательного определения эффективных доз для человека.
Например, в случае трансплантации ткани количество Tcm клеток против третьей стороны, инфузируемых реципиенту, должно быть больше, чем 1×104/кг массы тела. Число Tcm клеток против третьей стороны, инфузируемых реципиенту, обычно находится в диапазоне от 1×104 /кг массы тела до 1×108 /кг массы тела.
Таким образом, новые Tcm клетки против третьей стороны настоящего изобретения могут быть использованы в качестве адъювантной терапии для клеточного или тканевого трансплантата (как описано здесь далее). К тому же, новые Tcm клетки настоящего изобретения наделены активностью «трансплантат против пораженной клетки» (описано подробно выше) и, таким образом, могут быть использованы per se для лечения заболевания.
Согласно определенным вариантам, для получения активности «трансплантат против пораженной клетки» (например, противоопухолевое действие, такое как антилейкозная терапия) могут использоваться сингенные, а также несингенные клетки.
Таким образом, способ настоящего изобретения может применяться для лечения любого заболевания, такого как, но без ограничения, злокачественное заболевание, заболевание, связанное с пересадкой трансплантата, инфекционное заболевание, такое как вирусное заболевание или бактериальное заболевание, воспалительное заболевание и/или аутоиммунное заболевание.
Заболевания, которые можно лечить с использованием способов настоящего изобретения, включают, но без ограничения, злокачественные заболевания, такие как лейкоз (например, острый лимфатический, острый лимфобластный, пре-В-клеточный острый лимфобластный, острый лимфобластный Т-клеточный лейкоз, острый-мегакариобластный, моноцитарный, острый миелогенный, острый миелоидный, острый миелоидный с эозинофилией, В-клеточный, базофильный, хронический миелоидный, хронический, эозинофильный, Фрейнда, гранулоцитарный или миелоцитарный, волосатоклеточный, лимфоцитарный, мегакариобластный, моноцитарный, моноцитарно-макрофагальный, миелобластный, миелоидный, миеломоноцитарный, плазмоклеточный, пре-В-клеточный, промиелоцитарный, подострый, Т-клеточный, лимфосаркому, предрасположенность к миелоидной злокачественной опухоли, острый нелимфоцитарный лейкоз, Т-клеточный острый лимфоцитарный лейкоз (T-ALL) и В-клеточный хронический лимфоцитарный лейкоз (B-CLL)], лимфому (например, болезнь Ходжкина, неходжкинскую лимфому, В-клеточную, Беркитта, кожную Т-клеточную, ретикулоклеточную, лимфобластную, Т-клеточную, тимусную), карциному, бластому и саркому; заболевания, связанные с пересадкой трансплантата (например, отторжение трансплантата, хроническое отторжение трансплантата, подострое отторжение трансплантата, гиперострое отторжение трансплантата, острое отторжение трансплантата и реакция «трансплантат против хозяина»); инфекционные заболевания, включающие, но без ограничения, хронические инфекционные заболевания, подострые инфекционные заболевания, острые инфекционные заболевания, вирусные заболевания (например, EBV, CMV, HIV), бактериальные заболевания, протозойные заболевания, паразитарные заболевания, микозы, микоплазменные заболевания и прионные заболевания; воспалительные заболевания (например, хронические воспалительные заболевания и острые воспалительные заболевания); и аутоиммунные заболевания (например, сердечно-сосудистые заболевания, ревматоидные заболевания, эндокринные заболевания, желудочно-кишечные заболевания, кожные заболевания, болезни печени, неврологические заболевания, мышечные заболевания, заболевания почек, заболевания, относящиеся к репродуктивной системе, заболевания соединительных тканей и системные заболевания).
Таким образом, способ настоящего изобретения может, кроме того, успешно применяться в лечении заболевания у субъекта, одновременно способствуя приживлению трансплантата клеток или тканей, сингенных по отношению к Tcm клеткам против третьей стороны (например, в ситуациях, когда клеточный или тканевый трансплантат и клетки против третьей стороны получены от одного донора).
Использующийся здесь термин «примерно» означает±10%.
Термины «содержит», «содержащий», «включает», «включая», «имеет» и их конъюгаты означают «включающий, но не ограничивающийся».
Термин «состоящий из» означает «включающий и ограничивающийся».
Термин «состоящий главным образом из» означает, что композиция, способ или структура может включать дополнительные ингредиенты, стадии и/или части, но только в случае, если дополнительные ингредиенты, стадии и/или части существенно не изменяют базовые и новые характеристики заявленной композиции, способа или структуры.
Использующаяся здесь форма единственного числа «a», «an» и «the» включает множественное число, если в контексте четко не указано иное. Например, термин «соединение» или «по меньшей мере, одно соединение» может включать множество соединений, включая их смеси.
По тексту данной заявки разные варианты настоящего изобретения могут быть представлены в формате диапазона. Должно быть понятно, что описание в формате диапазона представлено для удобства и краткости, и не должно рассматриваться как ограничение объема изобретения. Соответственно, предполагается, что описание диапазона должно содержать конкретные раскрытые все возможные поддиапазоны, а также отдельные численные значения внутри этого диапазона. Например, описание диапазона, такого как от 1 до 6, должно рассматриваться как имеющее конкретные раскрытые субдиапазоны, такие как от 1 до 3, от 1 до 4, от 1 до 5, от 2 до 4, от 2 до 6, от 3 до 6 и т.д., а также отдельные значения внутри этого диапазона, например, 1, 2, 3, 4, 5 и 6. Это применяется независимо от ширины диапазона.
Во всех случаях, когда здесь указан численный диапазон, это означает включение любого числа (дробного или целого) внутри указанного диапазона. Выражения «изменяясь/изменяется в диапазоне», где первый указывает число и второй указывает число, и «изменяясь/изменяется от», где первый указывает на число «до» второго числа, используются здесь взаимозаменяемо и означают включение первого и второго указанных чисел и всех дробных и целых числа между ними.
Использующийся здесь термин «способ» означает способы, средства, методики и процедуры, предназначенные для выполнения поставленной задачи, при этом термин включает, но без ограничения, такие способы, средства, методики и процедуры, которые являются известными или легко разрабатываемыми из известных способов, средств, методик и процедур практикующими врачами в области химии, фармакологии, биологии, биохимии и медицины.
Предпочтительно, чтобы определенные признаки изобретения, которые для ясности описаны в контексте отдельных вариантов, могли быть также обеспечены в комбинации в одном варианте. Напротив, разные признаки изобретения, которые для краткости описаны в контексте одного варианта, могут также быть обеспечены отдельно или в любой приемлемой подкомбинации, или в любом другом описанном варианте изобретения. Определенные признаки, описанные в контексте разных вариантов, не должны рассматриваться как главные признаки тех вариантов, если вариант является неработоспособным без этих элементов.
Разные варианты и аспекты настоящего изобретения, как подчеркнуто здесь выше, и как заявлено в разделе формулы ниже, находит экспериментальное подтверждение в следующих примерах.
ПРИМЕРЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Последующие примеры, вместе с представленными выше описаниями, иллюстрируют изобретение неограничивающим образом.
В целом, используемая здесь номенклатура и лабораторные методики, используемые в настоящем изобретении, включают молекулярные, биохимические, микробиологические и рекомбинантные ДНК методики. Такие методики всесторонне описаны в литературе. Смотри, например, "Molecular Cloning: A laboratory Manual" Sambrook et al., (1989); "Current Protocols in Molecular Biology" Volumes I-III Ausubel, R. M., ed. (1994); Ausubel et al., "Current Protocols in Molecular Biology", John Wiley and Sons, Baltimore, Maryland (1989); Perbal, "A Practical Guide to Molecular Cloning", John Wiley & Sons, New York (1988); Watson et al., "Recombinant DNA", Scientific American Books, New York; Birren et al. (eds) "Genome Analysis: A Laboratory Manual Series", Vols. 1-4, Cold Spring Harbor Laboratory Press, New York (1998); methodologies as set forth in U.S. Pat. Nos. 4,666,828; 4,683,202; 4,801,531; 5,192,659 and 5,272,057; "Cell Biology: A Laboratory Handbook", Volumes I-III Cellis, J. E., ed. (1994); "Current Protocols in Immunology" Volumes I-III Coligan J. E., ed. (1994); Stites et al. (eds), "Basic and Clinical Immunology" (8th Edition), Appleton & Lange, Norwalk, CT (1994); Mishell and Shiigi (eds), "Selected Methods in Cellular Immunology", W. H. Freeman and Co., New York (1980); available immunoassays are extensively described in the patent and scientific literature, see, for example, U.S. Pat. Nos. 3,791,932; 3,839,153; 3,850,752; 3,850,578; 3,853,987; 3,867,517; 3,879,262; 3,901,654; 3,935,074; 3,984,533; 3,996,345; 4,034,074; 4,098,876; 4,879,219; 5,011,771 and 5,281,521; "Oligonucleotide Synthesis" Gait, M. J., ed. (1984); "Nucleic Acid Hybridization" Hames, B. D., and Higgins S. J., eds. (1985); "Transcription and Translation" Hames, B. D., and Higgins S. J., Eds. (1984); "Animal Cell Culture" Freshney, R. I., ed. (1986); "Immobilized Cells and Enzymes" IRL Press, (1986); "A Practical Guide to Molecular Cloning" Perbal, В., (1984) and "Methods in Enzymology" Vol.1-317, Academic Press; "PCR Protocols: A Guide To Methods And Applications", Academic Press, San Diego, CA (1990); Marshak et al., "Strategies for Protein Purification and Characterization - A Laboratory Course Manual" CSHL Press (1996); все из которых включены здесь полностью в виде ссылки, представленной здесь. Другие общие ссылки обеспечены по всему этому документу. Предполагается, что указанные здесь методики хорошо известны в данной области и приведены для удобства читателя. Вся содержащаяся здесь информация включена здесь в виде ссылки.
ПРИМЕР 1
Общие материалы и методика эксперимента
Животные
6-12 недельные самки мышей линии BALB/c (Н-2d), СВ6 (H-2bd), FVB (Н-2q), SJL (Н-2s), C57BL/6 (Н-2b), С3Н (Н-2k), Nude-BALB/C и Nude-C57BL/6 были получены от компании Weizmann Institute Animal Center (Rehovot, Израиль). Мышей линии С3Н и BALB/C скрещивали для генерации мышей F1 (C3HxBALB/C). Скрещенная пара трансгенных (Tg) мышей Н-2b, экспрессирующих TCR из CTL клона 2с со специфичностью к H-2Ld, были представлены компанией Janko Nikolic-Zugic (Sloan-Kettering, NY). Потомство данных Tg мышей скрещивали в Weizmann Institute Animal Breeding Center. Всех мышей хранили в маленьких клетках (пять животных в клетке), и кормили стерильной пищей и кислой водой.
Приготовление нереактивных по отношению к хозяину донорских CTL против третьей стороны CTL против третьей стороны были приготовлены, как было описано ранее Bachar et at. [Bachar-Lustig et al., Blood. (2003) 102:1943-1950]. Вкратце, спленоциты мышей-доноров культивировали в присутствии облученных (20 Gy) спленоцитов мышей FVB. Респондеры (4×106/мл) и стимуляторы (4×106/мл) культивировали в течение 6 дней в полной среде RPMI для культивирования тканей (СМ) при 37°C в атмосфере 5% CO2/воздух-инкубаторе. Через 6 дней после инициации культивирования клетки разделяли на фракции на Ficoll-Paque (Amersham Pharmacia Biotech AB) и фракцию лимфоцитов подвергали положительной селекции CD8+ клеток с использованием BD IMag™ CD8 Magnetic Particles (BD Pharmingen). Изолированные клетки (1×106/мл) рестимулировали облученными (20 Gy) сторонними спленоцитами (4×106/мл) и добавляли человеческий rIL-2 (40 U/мл, Eurocetus) в реакцию смешанной культуры лимфоцитов через день. На 16 день MLR культуры собирали, фракционировали на Ficoll-Paque и анализировали с помощью FACS на их CD8 чистоту.
Приготовление Tcm клеток против третьей стороны
Спленоциты мышей-доноров культивировали в присутствии облученных (20 Gy) сторонних спленоцитов мышей FVB. Респондеры (4×106/мл) и стимуляторы (4×106/мл) культивировали в течение 60 часов в полной среде RPMI культуры тканей (СМ) при 37°C в 5% СО2/воздух-инкубаторе. Через 60 часов после инициации культивирования клетки фракционировали на Ficoll-Paque (Amersham Pharmacia Biotech AB) и фракцию лимфоцитов подвергали положительной селекции CD8+ клеток с использованием BD IMag™ CD8 Magnetic Particles (BD Pharmingen). Изолированные клетки (1×106/мл) засевали с rIL-15 (20 нг/мл, R&D). Среду и цитокины заменяли через день. На 16 день клетки собирали, фракционировали на Ficoll-Paque и анализировали с помощью FACS на их чистоту Tcm (CD8, CD62L и CD44 экспрессия).
Модель отторжения аллотрансплантата, опосредованная Т-клетками
Самок мышей-хозяев (13-14 недельных) подвергали однократному облучению в дозе 10 Gy (сверхлетальное кондиционирование) TBI на день -2. На следующий день мыши получали внутривенно 1.25×104 неразделенных Т-клеток хозяина. Трансплантацию 3×106 аллогенных клеток ВМ мышей Nude выполняли на день О совместно с вето клетками, подлежащими оценке.
Приготовление Т-клеток хозяина
Спленоциты мышей-хозяев фракционировали на Ficoll/Paque и изолированные мононуклеарные клетки подвергали положительной селекции Т-клеток (CD4+ плюс CD8) с помощью магнитного сортера клеток (MACS).
Изображение In vivo
1×107 клеток-мишеней инкубировали с липофильным карбоцианиновым красителем в ближней ИК-области (DIR, Invitrogen) в 10 мл фосфатно-буферного солевого раствора (PBS), содержащего 1.5 мкг/мл DIR красителя и 0.5% этанола, в течение 60 минут при комнатной температуре. Затем клетки отмывали дважды PBS, и жизнеспособность меченых клеток проверяли по окрашиванию трипановым синим. DIR меченые клетки затем трансплантировали вместе с истощенным по Т-клеткам ВМ сверхлетально облученным мышам-хозяевам, которым ранее инфузировали очищенные Т-клетки хозяина в качестве индукторов отторжения трансплантата (как подробно указано выше). В заданное время после трансплантации мышей наблюдали с помощью оптической системы изображения всего тела (IVIS® 100, Xenogen), сопряженной с камерой Pixelfly QE (PCO, Kelheim, Германия) с зарядовой связью (CCD). Фильтры возбуждения и эмиссии в IVIS были от 710 до 760 нм и от 810 до 860 нм, соответственно. Получение изображения и анализ данных выполняли с использованием программного обеспечения Living Image 2.5.
Отслеживание in-vivo CD8+ клеток с использованием цитофлуориметрического анализа
Лимфоузлы, ВМ, печень и селезенку собирали от мышей-хозяев через шестьдесят часов и 6.5 дней после трансплантации ВМ и адоптивного переноса клеток-мишеней (как описано выше для in-vivo протокола радиологического исследования). Суспензию отдельных клеток готовили из каждого органа, и клетки фракционировали на Ficoll. Клетки затем окрашивали анти-Н-2kb, анти-Н-2Dd, анти-CD8 и анти-CD62L Abs (BD bioscience), и суспендировали в постоянном объеме PBS. FACS-анализ выполняли в установленное для каждого образца время.
Анализ вето-активности в TCR Tg 2с мышиной модели
Клетки селезенки мышей 2с Tg Н-2b, экспрессирующих TCR со специфичностью к H-2Ld мышам, использовали в качестве эффекторных клеток в вето анализе. Клетки селезенки собирали, лизировали в холодном лизирующем эритроциты (RBC) буфере (АСК буфер) для удаления эритроцитов и доводили до концентрации 2×106 клеток/мл в полной среде RPMI культуры тканей (СТМС). Клетки затем стимулировали облученными (20 Gy) спленоцитами мышей BALB/c (2×106/мл) в присутствии вето клеток, подлежащих оценке (предшественники СВ6 Н-2bd) при заданных концентрациях. Культуры инкубировали в течение 72 часов в U-образных 96-луночных планшетах. Делецию специфических эффекторных Т-клеток наблюдали с помощью цитофлуорометрического анализа, измеряя уровень живых [клетки отрицательные по отношению к 7-аминоактиномицину D (7AAD; Invitrogen)] CD8+ 2C клеток, специфически окрашенных 1В2 Ab, направленных против клонотипичных анти-Н-2Ld TCR. Активность ингибирования рассчитывали по следующей формуле:
(1-Кол-во 1B2+CDS+ клеток в анализируемой ячейке/ Кол-во 1B2+CD8+ клеток в контрольной ячейке) × 100
Оценка апоптоза с помощью проточной цитометрии с использованием аннексина V
Клетки (1-2×105) окрашивали Fitc-аннексин V (MBL Medical & Biological Laboratories, Naka-ku Nagoya, Япония) согласно протоколу производителя. Вкратце, клетки суспендировали в 100 мкл связующего буфера (10 мМ HEPES/NaOH, pH 7.4; 140 мМ NaCl; 2.5 мМ CaCl2) и инкубировали в течение 10 минут. Клетки затем отмывали связующим буфером и анализировали с использованием проточной цитометрии.
Анализ химеризма
Химеризм определяли с использованием цитофлуометрии. Клетки периферической крови фракционировали на Ficoll-Paque plus и изолированные мононуклеарные клетки каждой мыши дважды окрашивали методом прямой иммунофлуоресценции моноклональным антителом анти-H2d, специфичным к донору и анти-H2k специфичным к хозяину.
Проточно-цитометрический анализ
FACS-анализ выполняли с применением модифицированного FACScan проточного цитометра (Becton Dickinson). Клетки окрашивали мечеными антителами, специфичными к CD8α-PE/FITC/APC, CD3-PE/FITC/APC, CD62L-PE/FITC/APC, CD44-PE/FITC/APC, H2Kb-РЕ/FITC, H2Dd-PE/FITC, H2Kk-РЕ/FITC (BD Pharmigen), биотинилированному 1В2 и стрептавидину АРС (Jackson Laboratories). Окрашивание аннексином V и 7AAD проводили согласно инструкциям производителя (BD Pharmigen).
Анализ in-vitro вето-активности в 2с TCR Tg мышиной модели
Анализ in-vitro вето-активности выполняли, как описано выше30. Вкратце, 2 с спленоциты стимулировали облученными спленоцитами мышей BALB/c в присутствии вето клеток, подлежащих оценке при заданных концентрациях. Культуры инкубировали в течение 72 ч в 96-луночных планшетах. Делецию специфических эффекторных Т-клеток наблюдали с помощью FACS-анализа измерением уровня выживаемости (7AAD-), CD8+ 2С клеток, специфично окрашенных 1В2 Ab. Активность ингабирования рассчитывали по следующей формуле:
(1-Кол-во 1B2+CD8+ клеток в анализируемой ячейке/ Кол-во 1B2+CD8+ клеток в контрольной ячейке) × 100
Анализ in-vivo вето активности в 2с TCR Tg мышиной модели.
Летально облученные (10Gy) мыши C57BL/6 получали 1×105 очищенных CD8+2 с клеток (MACS) и 5×105 облученных (20Gy) спленоцитов мышей BALB/c. На следующий день мышам трансплантировали 1×106 клеток ВМ мышей C57BL/6-NUDE и 5×106 Tcm. Реципиентов умерщвляли через 8 дней после трансплантации, их селезенки собирали и делецию 2с Т-клеток наблюдали с помощью FACS, как описано в предыдущем разделе для анализа in-vitro.
Статистический анализ
Анализ данных по выживаемости выполняли с использованием кривых Kaplan-Meier (log-rank тест). Сравнение средних значений показателей производили с использованием t-критерия Стьюдента.
ПРИМЕР 2
Характер хоуминга вето CTL против третьей стороны (описанных ранее) по сравнению с сингенными и аллогенными наивными Т-клетками
При получении нереактивных по отношению к хозяину вето CTL, клетки подвергались продолжительной ex-vivo активации против посторонних стимуляторов в присутствии IL-2. В результате, клетки, по-видимому, превратились в эффекторные CTL, которые, как было показано ранее, проявляют распознающий воспаление фенотип с потерей способностей к хоумингу в лимфоузлы (LN) [Reinhardt et al., Nature (2001) 410:101-105; Weninger et al., J Exp Med. (2001) 194:953-966; Masopust et al., Science (2001) 291:2413-2417]. Такой механизм миграции может являться одной из причин несоответствия между эффективностью вето CTL против третьей стороны in vitro и in vivo. Для проверки данной вероятности автор настоящего изобретения использовал in-vivo модель проверки эффективности вето CTL против третьей стороны. Эта точная модель включала летально облученных мышей, которые получали истощенный Т-клетками аллогенный костный мозг (TDBM) в присутствии или отсутствии дозированного количества Т-клеток хозяина, которые индуцировали отторжение и фатальную анемию на третьей неделе после трансплантации костного мозга (ВМТ).
Вкратце, сверхлетально облученные мыши FVB были радиозащищены аллогенным TDBM мышей Ba1b/c в присутствии или отсутствии адоптивно перенесенных очищенных Т-клеток хозяина (НТС), которые индуцировали отторжение аллотрансплантата. Данные реципиенты ВМ (подвергающиеся отторжению аллотрансплантата ВМ) были дополнительно инфузированы разными типами Т-клеток, меченых DIR, которые могли быть обнаружены с помощью in-vivo системы изображений (IVIS).
Как видно на Фигурах 1A-D, через 36 часов после инфузии меченых клеток, сингенные (НТС) и аллогенные наивные Т-клетки проявляли значительный хоуминг в LN, в то время как аллогенные вето CTL против третьей стороны (описанные ранее) были сконцентрированы в печени, легких и ВМ, и были исключены из LN (Фигура 1D). Способность аллогенных наивных Т-клеток солокализоваться с НТС указывает на то, что аллогенные Т-клетки, проявляющие соответствующий фенотип, могут мигрировать в LN в контексте специальной среды модели отторжения трансплантата.
ПРИМЕР 3
Индукция Tcm клеток против третьей стороны in vitro
Автор настоящего изобретения ранее разработал CTL против третьей стороны, которые проявляют заметную вето-активность (благодаря их FasL экспрессии) и которые лишены GVHD реактивности по причине их продолжительной активации против третьей стороны [Bachar-Lustig et al.. Blood. (2003) 102:1943-1950]. Однако, как указано в Примере 2 выше, эти клетки проявляли слабый LN хоуминг. Данные результаты привели автора настоящего изобретения к попытке дальнейшего улучшения реактивности in-vivo этих клеток путем индукции фенотипа Т-клеток центральной памяти (Tcm) в этих клетках при их активации против третьей стороны. Ранее было показано, что фенотип Tcm связан с активированными клетками (CD44+), которые экспрессируют, помимо других поверхностных антигенов, CD62L и CCR7, что делает возможным их хоуминг в LN [Sallusto et al.. Nature (1999) 401:708-712; Weninger et al, supra; Masopust et al., supra].
Таким образом, автор настоящего изобретения проверил способность IL-2 или IL-15 индуцировать фенотип Tcm на CD8+ Т клетках против третьей стороны. Так как для ослабления GVH реактивности требовался недостаток цитокинов до размножения в присутствии тестируемых цитокинов, было произведено сравнение разных протоколов после короткого и продолжительного периодов депривации цитокинов. Таким образом, CD8+ Т клетки подвергали 2 дням или 6 дням стимуляции облученными сторонними стимуляторами в отсутствии цитокинов и затем культивировали в присутствии IL-2 или IL-15, и наблюдали на экспрессию CD62L и CD44 (как отражено подробно в Примере 1 здесь выше).
Как показано на Фигуре 2А, через 60 часов стимуляции сторонними стимуляторами в отсутствии цитокинов, CD8+ Т клетки сохранили экспрессию CD62L (90±5% клеток, экспрессирующих CD62L), в то время как через 6 дней стимуляции только 18±6% CD8+ Т клеток сохранили экспрессию CD62L (р-величина <0.001). Согласно этому наблюдению, когда CD8+ T клетки затем культивировали с IL-15 в Ag-свободной среде после периода стимуляции, значительно большее количество клеток с фенотипом CD44+CD62L+ Tcm было индуцировано на 15 день культивирования, когда клетки подвергали 60 часам стимуляции (Фигуры 2В-С), в противоположность клеткам, которых подвергали 6 дням стимуляции (р-величина = 0.005). Использование IL-2 не способствовало значительной индукции Tcm клеток против третьей стороны после короткой (2 дня) сторонней стимуляции клеток (Фигура 2В, р-величина = 0.007).
Более того, по окончании периода культивирования, CTL против третьей стороны, которых подвергали 6 дням сторонней стимуляции, последовательно культивировали с IL-2 и реактивировали их когнатным Ag, в большинстве случаев проявили эффекторный фенотип, и только 4±2% данных клеток проявили фенотип Tcm (Фигура 2В), при этом полученные результаты соответствовали характеру периферического хоуминга данных клеток, описанному в Примере 2 выше (Фигура 1D).
Представленные результаты, которые указывают на то, что CD8+ Т-клетки (которые подвергаются протоколу длительной активации, например, 6 дней) менее склонны к изменению на фенотип Tcm, согласуются с ранними исследованиями, согласно которым существует временное окно, в котором Ag-активированные Т-клетки могут направляться IL-15 в фенотип Tcm [Manjunath et al., J Clin Invest. (2001) 108:871-878; Carrio et al., J Immunol. (2004) 172:7315-7323].
ПРИМЕР 4
Сравнение LN хоуминга Tcm клеток и вето CTL
Как показано в Примере 3 выше, результаты настоящего изобретения ясно продемонстрировали, что Tcm-подобный фенотип может быть индуцирован кратковременной аллоактивацией и культивированием клеток в Ag-свободной среде в присутствии IL-15. Однако, было важным проверить в условиях модели отторжения трансплантата, что клетки, экспрессирующие этот Tcm-подобный фенотип, проявляют отличный LN хоуминг по сравнению с вето CTL.
Таким образом. Tcm клетки (описанные здесь) и вето CTL (представленные ранее) были получены, как описано в Примере 1 здесь выше. Через 2 недели in vitro размножения в присутствии IL-15, образующие клетки прошли положительную селекцию на экспрессию CD62L для создания чистой популяции Tern клеток (>96% CD62L+ чистоты), в отличие от вето CTL, которые проявили низкую экспрессию CD62L (>95% CD62L- чистоты). Клетки были мечены DIR и адоптивно перенесены в мышей-хозяев BALB/C в контексте модели отторжения трансплантата, описанной в Примерах 1 и 2 здесь выше. Так как Tern клетки подвергали недостаточности IL-2 только в течение 60 часов вместо 6 дней согласно методу, который ранее показал in vitro и in vivo уменьшение GVHD активности из вето CTL [Bachar-Lustig et al., supra], в настоящем эксперименте использовали мышей СВ6 [(C57BL/6*BALB/b)F1, Н2-bd] в качестве источника адоптивно перенесенных клеток. Через два дня (Фигуры 3А-В и Фигуры 4A-D) или 6.5 дней (Фигуры 5А-С и Фигуры 6A-J) после адоптивного переноса мышей умерщвляли, и распределение клеток СВ6 происхождения в разных органах определяли с использованием ex-vivo визиографии всего тела (Фигуры 3А-В и Фигуры 5А-С) и с помощью FACS-анализа клеточных суспензий, полученных после растирания органов (Фигуры 4A-D и Фигуры 6A-J).
Как видно из результатов, через шестьдесят часов после трансплантации оба анализа, IVIS (Фигуры 3А-В) и FACS (Фигуры 4A-D), уверенно продемонстрировали четко выраженные различия в характерах миграции между двумя клеточными популяциями. В то время как Tcm клетки были четко видны в периферических и мезентериальных LN (Фигуры 3А и 4А-В), вето CTL с трудом определялись в этих органах и были обнаружены в основном во внутренних органах (печень, легкие, селезенка) и Вм (Фигуры 3В и 4C-D).
Аналогичный паттерн был также обнаружен через 6.5 дней, когда Tcm клетки, в противоположность вето CTL, четко наблюдались в LN (Фигура 5В), С использованием FACS-анализа и прямого подсчета клеток в растертых органах (Фигуры 6A-J) было очевидно, что вето CTL локализованы главным образом в селезенке, печени и ВМ (Фигуры 6F-J), в то время как через 6.5 дней после ВМТ в LN было обнаружено в 23 раза больше Tern клеток, чем вето CTL (Фигуры 6А-Е). Аналогичные результаты были получены уже после 2 дней ВМТ (не показано). Более того, было обнаружено, что общее количество 'Tcm', собранных из всех тестируемых органов, увеличилось в 9 раз в период от 2 до 6.5 дней после ВМТ, в противоположность 'CTL', которые проявили незначительную пролиферацию (Фигура 6К). Таким образом, было сделано заключение о том, что 'Тст' не только мигрируют в LN ВМТ-реципиентов, но также интенсивно пролиферируют на раннем пост-трансплантационном периоде.
ПРИМЕР 5
Влияние адоптивного переноса Tcm клеток на выживаемость реципиентов с аллотрансплантатом TDBM
Так как Tcm клетки эффективно мигрируют в LN и проявляют вето ассоциированный фенотип (данные не показаны), который делает возможной толеризацию солокализованных Т-клеток хозяина (НТС), автор настоящего изобретения оценил способность адоптивно перенесенных Tcm клеток против третьей стороны усиливать выживаемость реципиентов с аллотрансплантатом TDBM без дальнейшего лечения. Данное исследование проводили на ранее описанной модели отторжения трансплантата (смотри Пример 1 здесь выше), которая является специально разработанной для измерения опосредованного Т-клетками отторжения трансплантата TDBM без вмешательства конкуренции стволовых клеток, которая может возникнуть у мышей, подвергшихся воздействию кондиционирования со сниженной интенсивностью.
Таким образом, мыши С3Н были летально облучены и инфузированы НТС перед пересадкой аллотрансплантата TDBM от мышей-доноров nude BALB/C в присутствии или отсутствии адоптивно перенесенных очищенных Tcm клеток против третьей стороны (>95% чистота). Мышей (C3HxBALB/C)F1 затем использовали в качестве источника Tcm клеток для исключения возможного усиления приживаемости трансплантата или летальности, вызванной GVHD и/или аллореактивностью Tcm клеток.
Как видно на Фигуре 7, все мыши в облученной контрольной группе, не радиозащищенные трансплантатом ВМ, быстро умирали после лучевой терапии. Напротив, все мыши, которые получали трансплантат ВМ, выжили, не смотря на то, что добавление НТС приводило к отторжению трансплантата и летальности.
Удивительно, что отторжение, опосредованное НТС, преодолевалось при адоптивном переносе 107 Tcm клеток без дальнейшего лечения (7/8 мышей выжили).
Для оценки неудачи НТС отторгнуть трансплантат ВМ при введении в сочетании с Tcm клетками против третьей стороны, химеризм реципиентных мышей проверяли через 80 дней после трансплантации с помощью FACS-анализа периферической крови. Как показано на Фигуре 8, мыши, которые получали только трансплантаты ВМ, без НТС для индуцирования отторжения (колонка А), проявили сравнимые уровни донорского химеризма, как и мыши, которые получали, дополнительно к ВМ, НТС и 107 Tcm клеток (колонка В, >90% химеризма донора). Таким образом, Tcm клетки обеспечивают успешное приживление трансплантата, несмотря на присутствие аллореактивных НТС.
Более того, через 80 дней после трансплантации, Tcm клетки легко обнаруживали в периферической крови реципиента и содержали 44±6% общего количества CD8 клеток в периферии (Фигуры 9А-В).
Эта высокая устойчивость Tcm клеток в аллотрансплантате TDBM реципиента указывала на то, что дополнительно к их заметной способности индуцировать толерантность, эти клетки могут внести важный вклад в восстановление иммунитета после тяжелого кондиционирования, которое применяется для реципиентов при трансплантации ВМ.
ПРИМЕР 6
Прямое сравнение способностей индуцировать толерантность Tcm клеток и вето CTL
Для прямого сравнения способностей Tcm клеток против третьей стороны настоящего изобретения и описанных ранее вето CTL против третьей стороны индуцировать толерантность, автор настоящего изобретения адоптивно переносил 107 Tcm или вето CTL, полученных от мышей (C3HxBALB/C)F1, в мышей С3Н, реципиентов аллотрансплантата TDBM мышей BALB/C, в контексте ранее описанной модели отторжения трансплантата (смотри Пример 1, здесь выше).
Для оценки продолжительной способности адоптивно перенесенных донорского типа 'Tcm' против третьей стороны победить отторжение аллотрансплантата ВМ, использовали модель отторжения трансплантата, описанную выше, которая является специально разработанной для измерения опосредованного Т-клетками отторжения аллотрансплантата TDBM без вмешательства конкуренции стволовых клеток, которая может возникнуть у мышей, подвергшихся RJC. Таким образом, мышей С3Н летально облучали и инфузировали НТС перед пересадкой аллотрансплантата TDBM от BALB/c-NUDE доноров в присутствии или отсутствии очищенных 'Tcm'. Мышей (C3HxBALB/c)F1 использовали в качестве источника 'Tcm' для исключения возможного усиления приживления трансплантата или летальности, вызванной GVHD, и/или остаточной аллореактивностью 'Tcm'. Как видно на Фигуре 10А, все мыши в облученной контрольной группе, не радиозащищенные трансплантатом ВМ, быстро умирали после облучения. Напротив, 29 из 30 мышей, получающих TDBM, выживали, в то время как добавление НТС приводило к отторжению трансплантата и летальности всех 59 мышей, которые получали данное лечение. Удивительно, что данное НТС-опосредованное отторжение было преодолено у 16/18 мышей и у 19/23 мышей при адоптивном переносе 1×107 или 5×106 Tcm', соответственно, в то время как более низкое количество 'Tcm' было менее эффективным (Фигура 10А). Все выжившие мыши проявили устойчивый донорский химеризм (>90%), который присутствовал в течение более 1 года (данные не показаны, CD8 химеризм показан на Фигуре 10С). Конечно, как видно на Фигуре 10 В, только 3/16 из реципиентных мышей, получивших 1×107 CTL', без рапамицина, выжили (р<0.0001). Более того, 'Tcm' проявили удивительную устойчивость in-vivo и могли быть обнаружены в периферической крови реципиента в течение продолжительно периода времени. Таким образом, даже через 1 год после трансплантации H2kd положительные CD8 Т клетки (т.е. потомство инфузированных 'Tcm') содержали 19±6% общего количества CD8 Т-клеточного компартмента в периферической крови (Фигура 10С).
ПРИМЕР 7
Полностью аллогенные 'Tcm' против третьей стороны лишены GVH реактивности и способствуют примсивлению аллотрансплантатов TDBM
После демонстрации способности 'Tcm' побеждать опосредованное Т-клетками отторжение аллотрансплантата ВМ с использованием (HostxDonor) F1 доноров, лишенных аллореактивности, дальнейшую оценку полного потенциала данных клеток проводили с использованием полностью МНС несовместимых аллогенных доноров. В частности, полагая, что 'Tcm' против третьей стороны генерируются посредством начальной аллогенной стимуляции только в течение 60 часов в условиях депривации цитокинов (для того, чтобы избежать даун-регуляции CD62L) вместо обычных 6 дней метода депривации19 для генерации 'CTL' против третьей стороны, было важно оценить GVH реактивность полностью аллогенных 'Tcm'.
Таким образом, очищенные 'Tcm' против третьей стороны, полученные от мышей BALB/c, сравнивали с наивными CD8+ клетками, полученными от мышей BALB/c, на их GVH реактивность при адоптивном переносе в сверхлетально облученных С3Н реципиентов, также трансплантированных ВМ мышей BALB/c-NUDE. Мышей наблюдали на выживаемость и признаки GVHD: взъерошенная шерсть, согнутая спина и сниженная масса тела. Как видно на Фигуре 11А, инфузия 5×10 или 2×106 'Tcm' не индуцировала летальной GVHD, и 14/17 мышей в каждой группе выживало, аналогично реципиентам NUDE-BMT без дополнительных Т-клеток (18/20 мышей выживало). В противоположность, только 1/15 и 2/22 мышей, получивших 5×106 или 2×106 наивных CD8+ Т-клеток, соответственно, выжили. Более того, внешний вид мышей, получавших 'Tcm', а также их вес, значительно не отличались от контрольных мышей, трансплантированных только NUDE-KM (Фигура 11В). Это тоже находится в контрасте с результатами у мышей, получавших наивные Т-клетки, которые показали значительную потерю веса (р<0.05) и имели внешний вид, сравнимый с GVHD.
В заключение, способность данных полностью аллогенных 'Tcm' против третьей стороны, лишенных GVH реактивности, победить опосредованное Т клетками отторжение, оценивали в контексте ранее описанной модели отторжения трансплантата. Как видно на Фигуре 11С, 21/25 мышей, которые получали 5×106 'Tcm', полученных от мышей BALB/c, выживали после трансплантации, и показали стабильный донорский химеризм (>95%). Напротив, инфузия 1×107 'CTL', полученных от мышей BALB/c, могла сохранить только 7/27 реципиентных мышей (р<0.0001), и меньшее количество 'CTL' (5×106 или 2×106) было полностью неэффективным для предотвращения данного НТС-опосредованного отторжения (данные не показаны).
ПРИМЕР 8
'Tcm' проявляют низкую вето-активность in-vitro, но при реактивации приобретают эффекторный фенотип, сопровождаемый сильной и специфичной вето-активностью
Несмотря на то, что 'Tcm' против третьей стороны эффективно мигрируют в LN, индукция толерантности этими клетками также требует, чтобы эти клетки были наделены сильной вето-активностью. Ранее было показано, что в целом, CD8+ Т клетки, несущие фенотип Tcm, проявляют слабые эффекторные механизмы CTL, но их вето-активность никогда не была описана.
Для оценки вето активности 'Tcm' использовали анализ TCR трансгенных CD8 эффекторных клеток (2с модель). 2 с CD8+ клетки экспрессируют трансгенные TCR против Н-2d и, следовательно, являются предрасположенными к вето делеции только вето 'CTL' Н-2d происхождения, но не неспецифичными 'CTL' разного Н-2 типа, которые не распознаются 2с клетками19 2с клетки могут быть специфично идентифицированы с помощью клонотипичного 1В2 антитела. Мышей СВ6 использовали в качестве источника вето клеток для исключения гипотетической остаточной аллореактивности против 2с клеток. Согласно более ранним исследованиям, 'CTL', несущие эффекторный фенотип, проявили эффективное ингибирование размножения 2с клеток (Фигура 12А), вызванное апоптозом, на что указывает окрашивание аннексином V (Фигура 12 В). Напротив, 'Tcm' проявили слабую ингибирующую реактивность (Фигура 12А) и не индуцировали апоптоз в отношении 2с клеток, на что указывают более низкие уровни окрашивания аннексином V (Фигура 12В).
Однако, короткая реактивация 'Tcm' приводила к даун-регуляции экспрессии CD62L (данные не показаны) и к заметно усиленной супрессорной активности (Фигура 12А), по-видимому, опосредованной механизмом, основанным на делеции, на что указывает окрашивание аннексином V (Фигура 12В). Более того, реактивированные 'Tcm', аналогично 'CTL', проявили специфичную вето-активность, как показано при сравнении реактивированных "специфичных" 'Tcm' или 'CTL', полученных от мышей СВ6 (H2bd) или "неспецифичных" (H2bk) CD8+ Т-клеток мышей (C57xC3H)F1, которые не экспрессируют молекулу Н-2d и не распознаются 2с CD8+ Т-клетками. Таким образом, только "специфичные" реактивированные 'Tcm' или 'CTL' эффективно ингибировали размножение 2с клеток (Фигура 12С) и были способны индуцировать экспрессию аннексина V в 2с эффекторных клетках (Фигура 12D). В совокупности, данные результаты предполагают, что 'Tcm' не проявляют истинной вето активности, но при реактивации приобретают эффекторный фенотип и вето-активность. Это согласуется с быстрой индукцией эффекторного фенотипа и функции, описанной ранее для CD8 Т-клеток, несущих фенотип Tcm при реактивации. В совокупности, описанные выше данные предполагают, что усиленный LN хоуминг и эффективная вето-активность in-vitro являются взаимоисключающими. При этом, поскольку реактивация 'Tcm' существенно улучшала их вето-активность, представляло интерес определить, могут ли адоптивно перенесенные 'Tcm' быть реактивированы в LN трансплантата ВМ реципиентов, где адоптивно перенесенные Т-клетки хозяйского типа могут преодолевать ответные реакции анти-донора. Фактически, через 4 дня после адоптивного переноса очищенных 'Tcm', значительное количество инфузированных клеток, восстановленных из LN, проявило заметно сниженную экспрессию CD62L (Фигура 12Е), указывая на то, что реактивация может возникнуть, возможно, в результате стимула, индуцированного летальным кондиционированием и/или аллореактивностью НТС.
ПРИМЕР 9
'Tcm' специфично удаляют анти-донорские Т-клетки in-vivo
Предполагая, что 'Tcm' эффективно мигрируют в LN реципиентной мыши и проявляют вето-ассоциированный фенотип, который дает возможность толеризации солокализованных НТС, была оценена способность адоптивно перенесенных 'Tcm' подавлять Ag-специфичные Т-клетки in-vivo. Для непосредственной оценки толерогенной активности 'Tcm' без вмешательства документально подтвержденной вето активности клеток ВМ и извлеченных клеток13, 15, 18 ВМ, была установлена сингенная модель ВМТ, в которой летально облученные мыши-хозяева C57BL/6 (H2b) были радиозащищены сингенными клетками ВМ мышей C57BL/6-NUDE. К тому же, реципиентные мыши получали 2С (H2b) CD8+Т-клетки и облученные спленоциты мышей BALB/c (H2d), которые индуцируют размножение 2с клеток in-vivo. Через 1 день после переноса 2с клеток и их стимуляторов, реципиенты получали либо "специфичные" очищенные 'Tcm' (H2bd), полученные от мышей СВ6, либо "неспецифичные" очищенные 'Tcm', полученные от мышей C57BL/6, которые не экспрессируют молекулу Н-2d и, следовательно, не распознаются 2с CD8+ Т-клетками. Селезенки реципиентов собирали и оценивали через 8 дней после трансплантации на присутствие 2с клеток. Удивительно, что "специфичные" 'Tcm' эффективно ингибировали (71% ингибирования) размножение 2с клеток (Фигуры 13А-В) по сравнению с "неспецифичными" 'Tcm', которые проявили только 20% ингибирования. Более того, окрашивание аннексином V показало, что механизм ингибирования, проявленный "специфичными" 'Tcm', был опосредован индукцией апоптоза при разпознавании 2 с клеток (Фигуры 13C-D).
Несмотря на то, что изобретение было описано в отношении его определенных вариантов, очевидно, что существует много альтернатив, модификаций и вариантов, которые будут очевидны опытным в данной области специалистам. Соответственно, предполагается включение всех таких альтернатив, модификаций и вариантов, которые находятся в пределах сущности и широкого объема прилагаемой формулы.
Все публикации, патенты и патентные заявки, упомянутые в данном описании, включены полностью в виде ссылки в том же объеме, как если бы они были отдельной публикацией, патентом или патентной заявкой. К тому же, размещение или идентификация любой ссылки в данной заявке не должно рассматриваться как признание того, что такая ссылка существует в качестве прототипа настоящего изобретения. В том смысле, в котором используются заголовки разделов, они не должны рассматриваться в качестве обязательного ограничения.
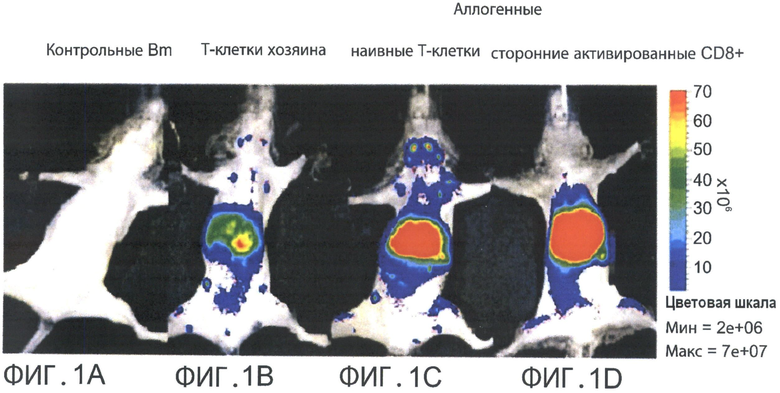
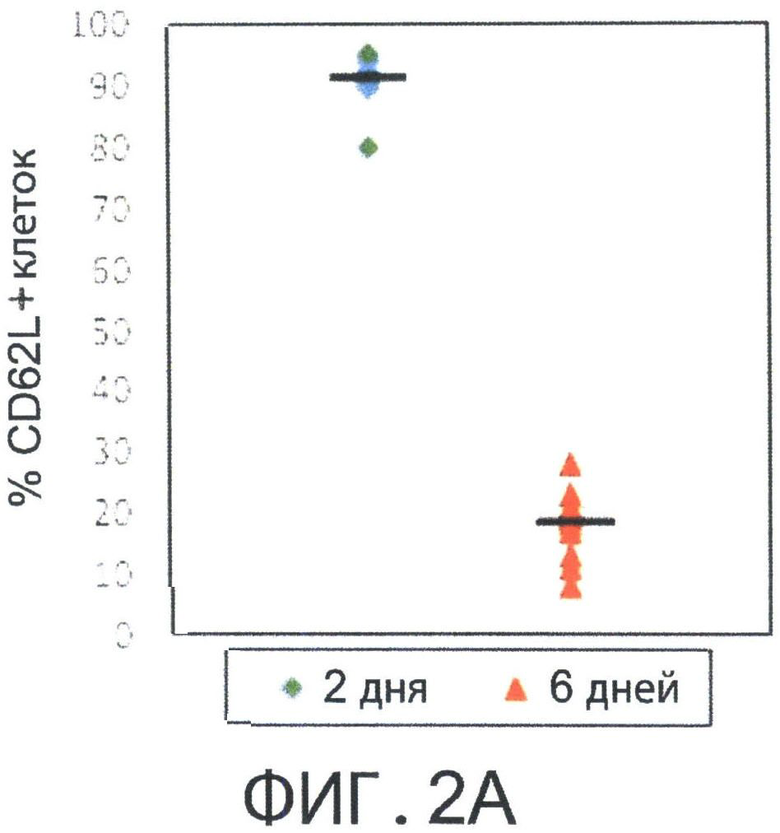
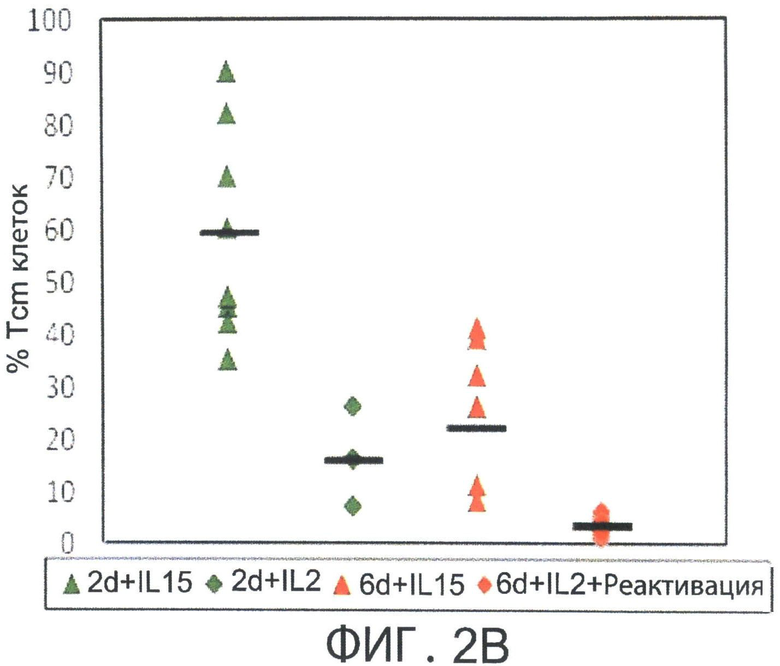
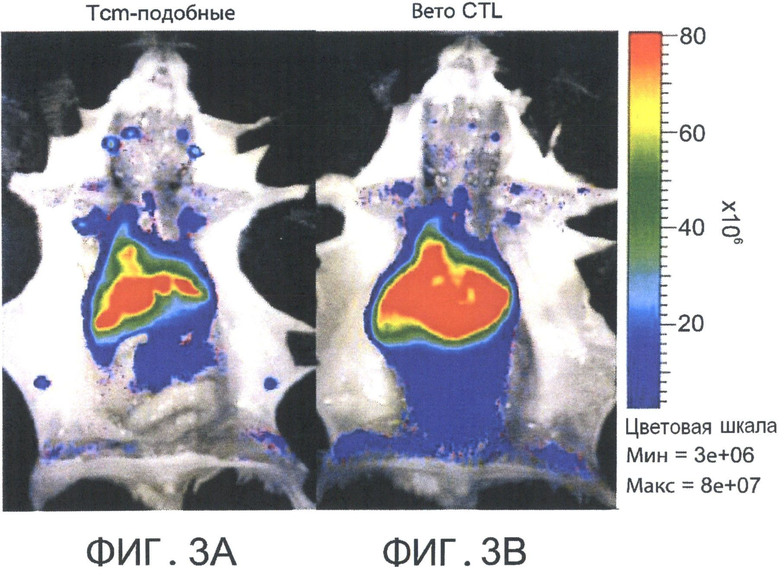
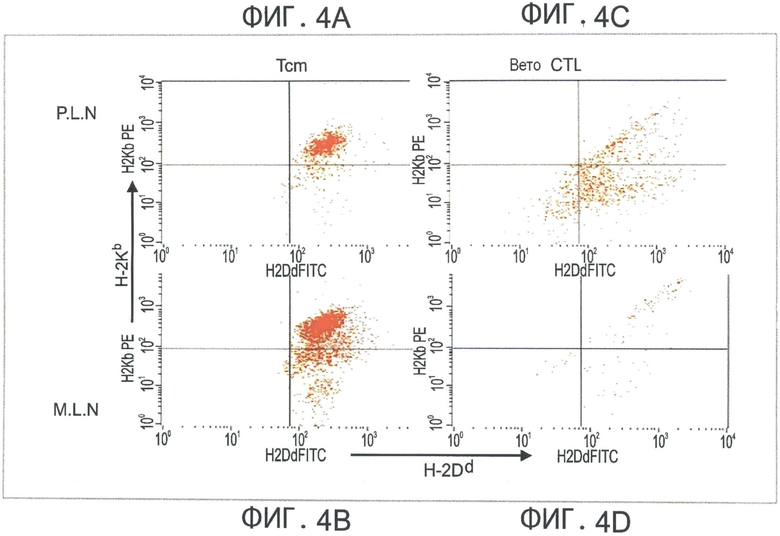
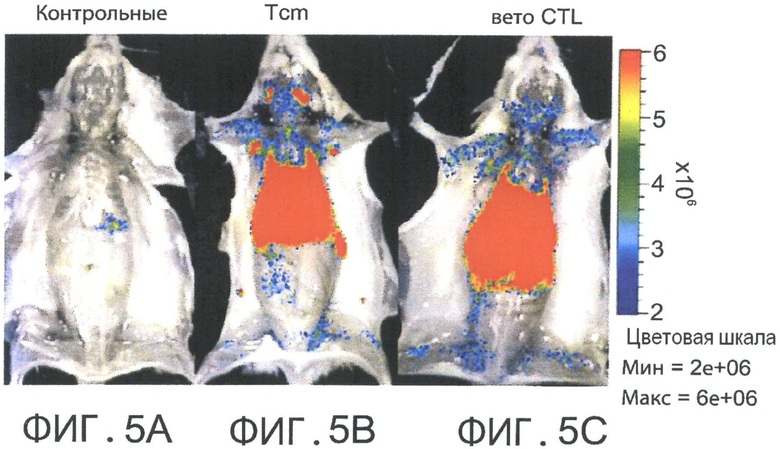
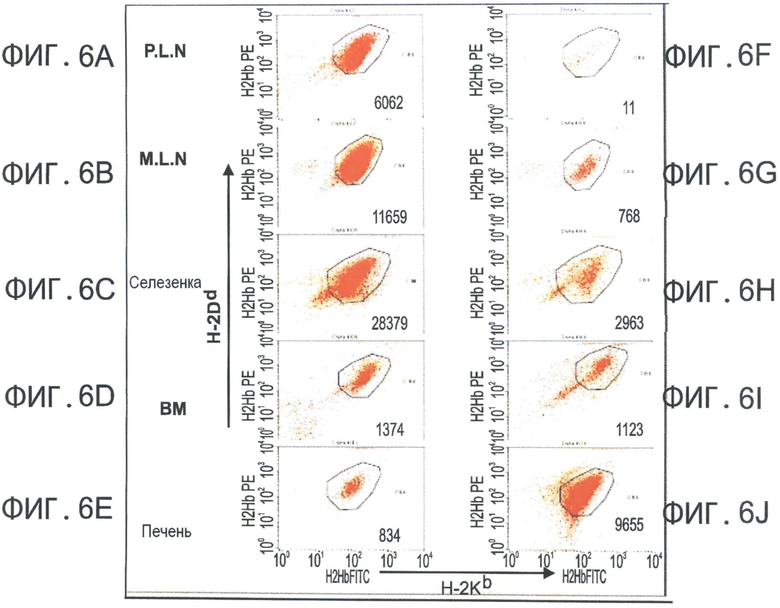
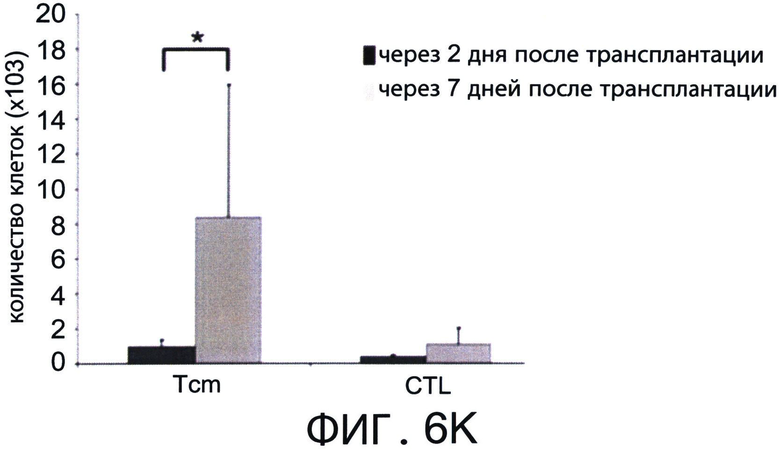
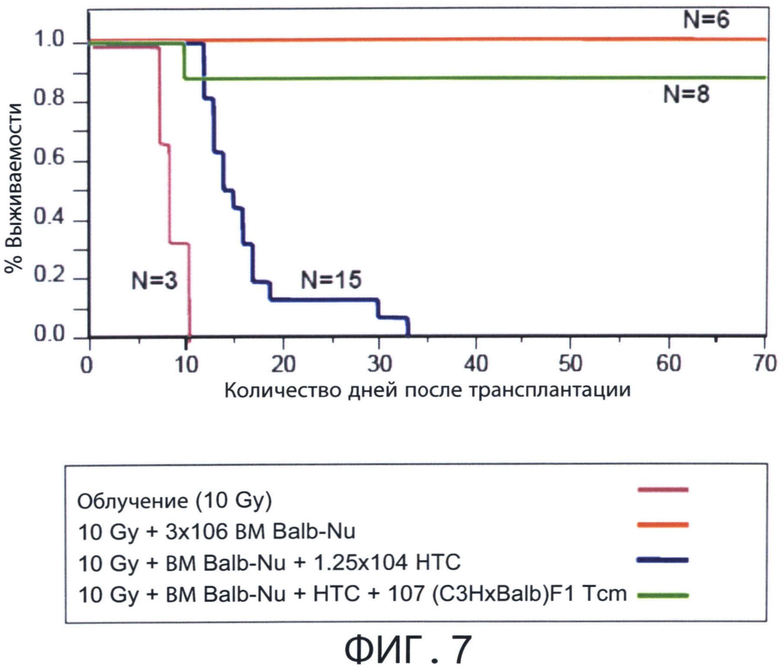
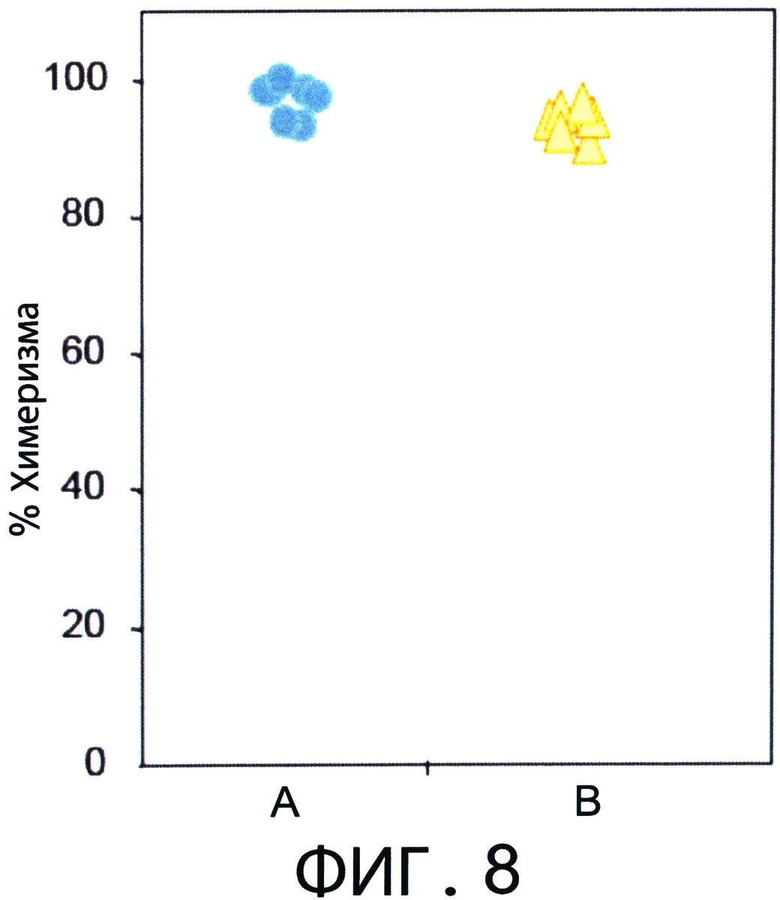
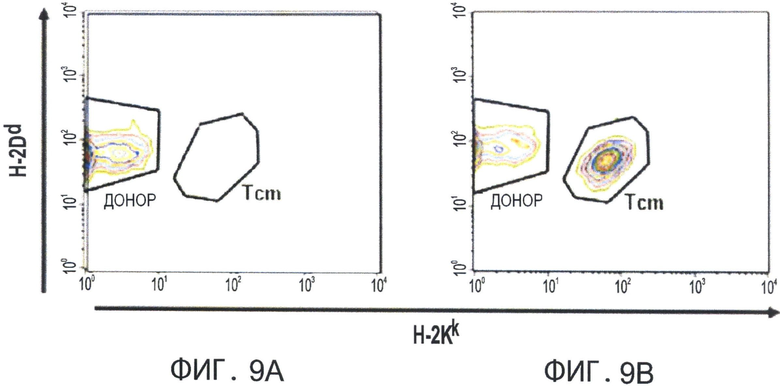
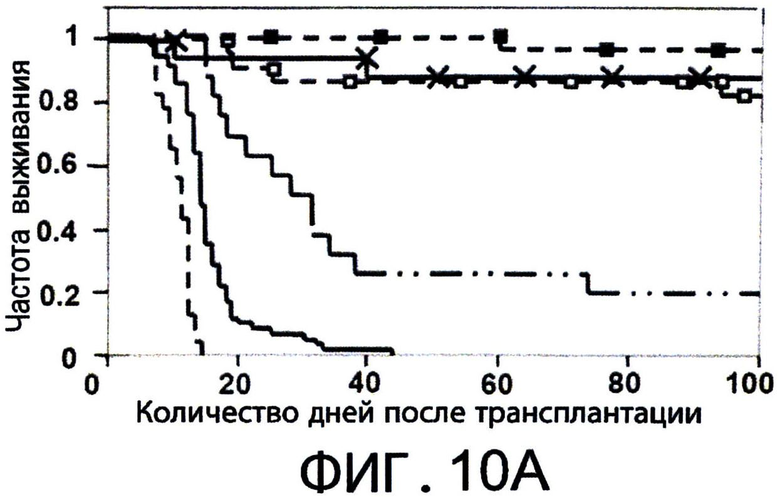
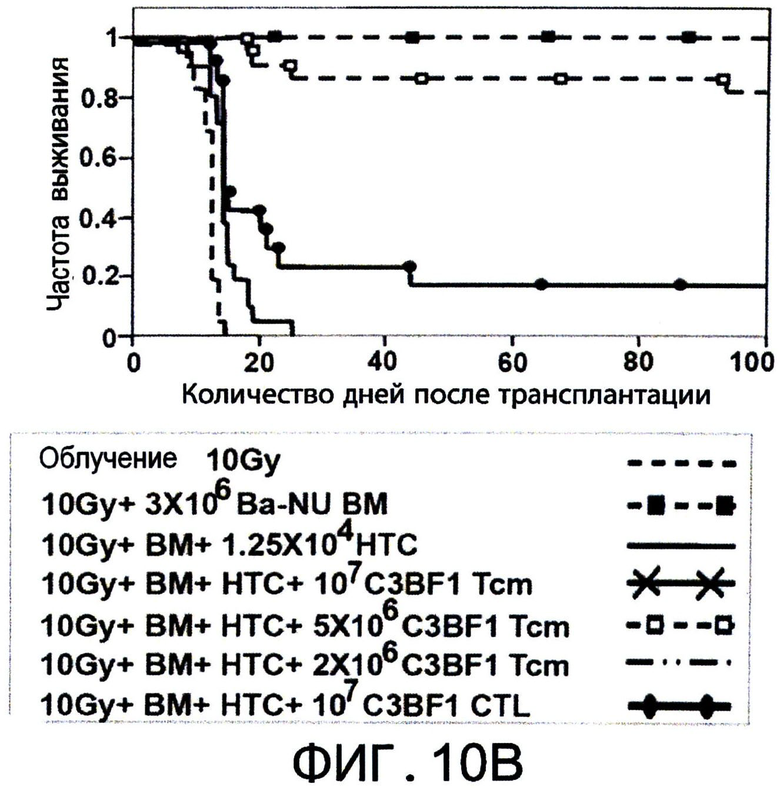
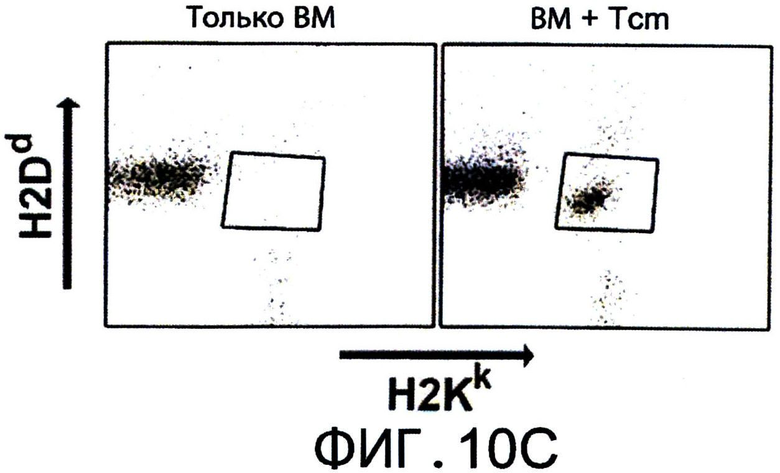
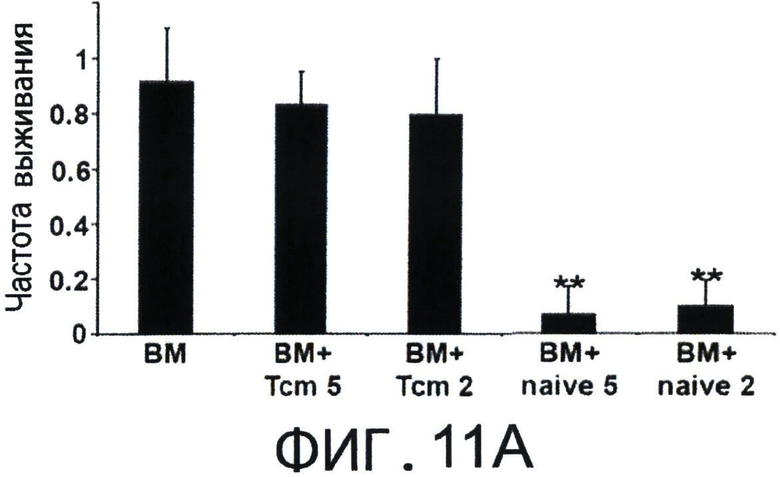
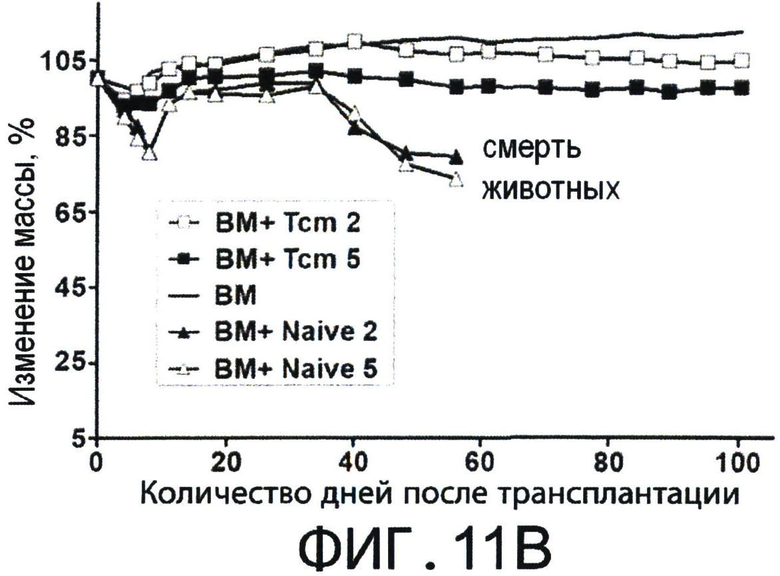
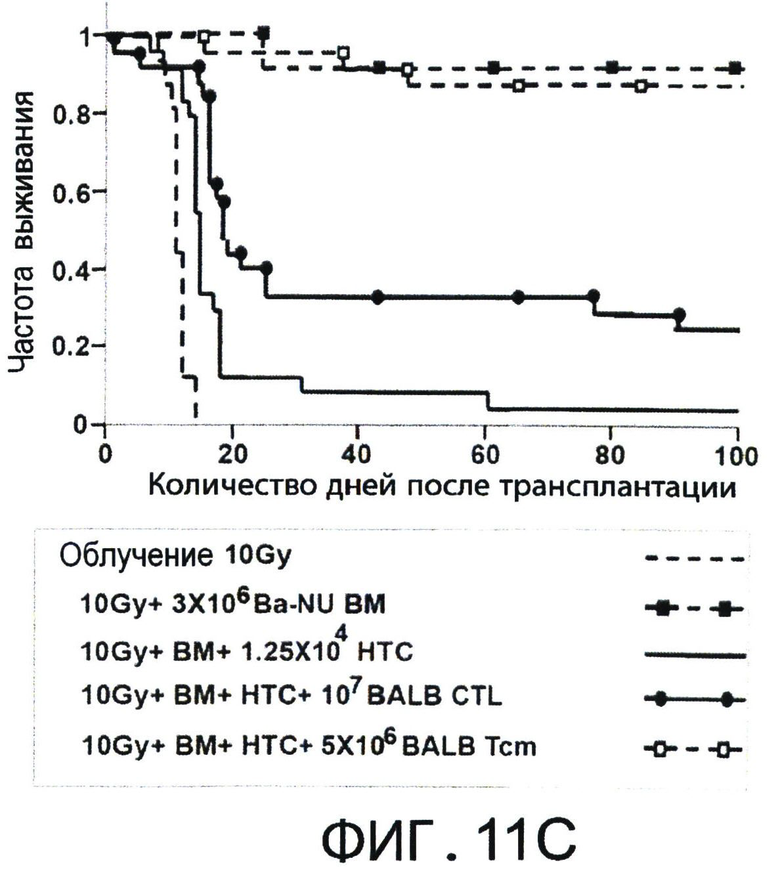

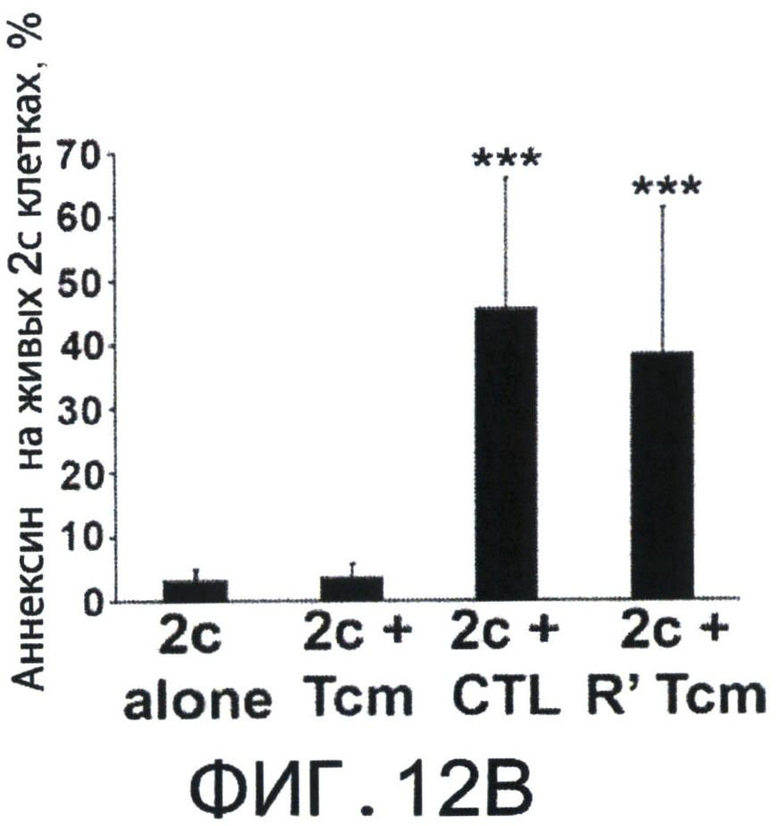
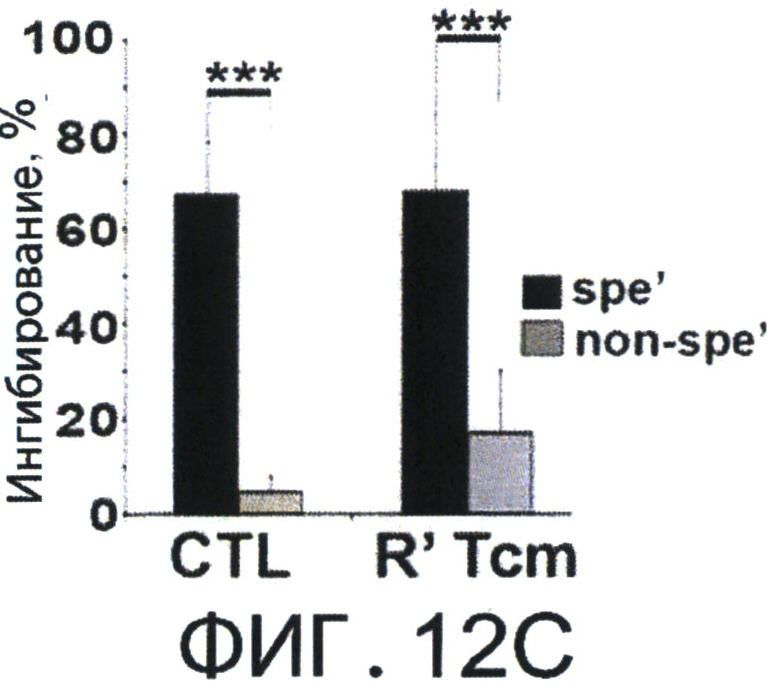
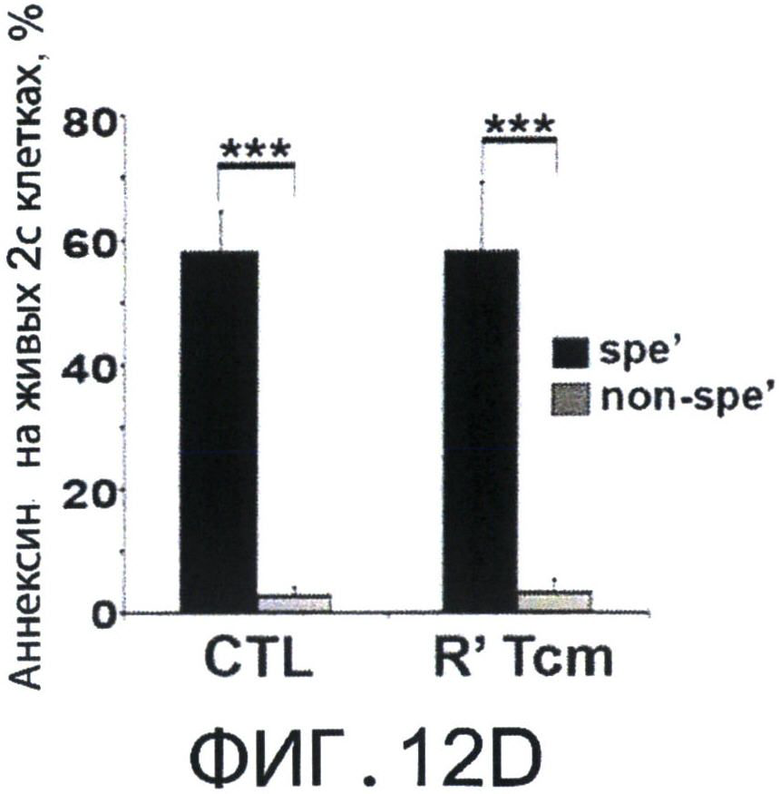
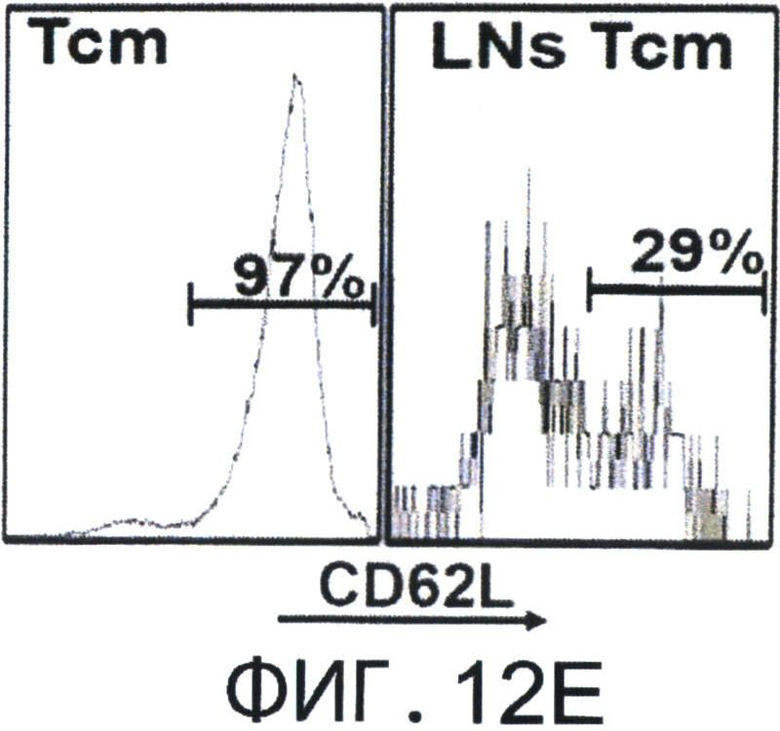

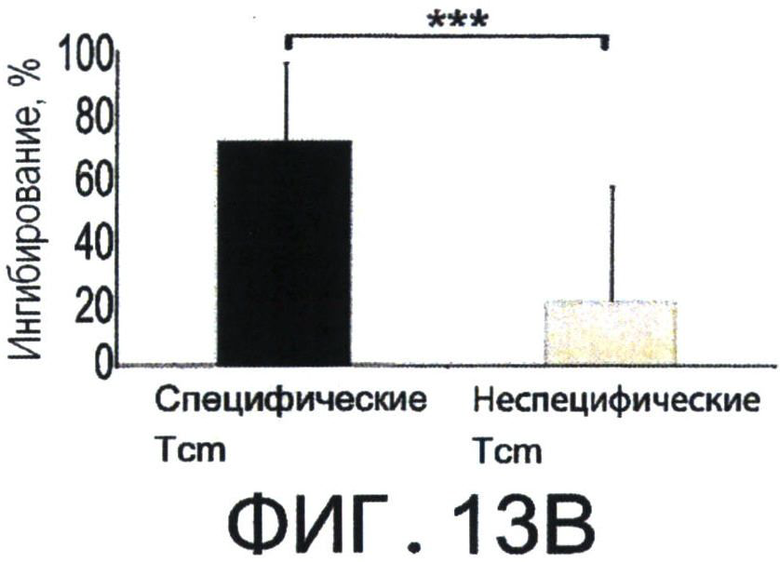
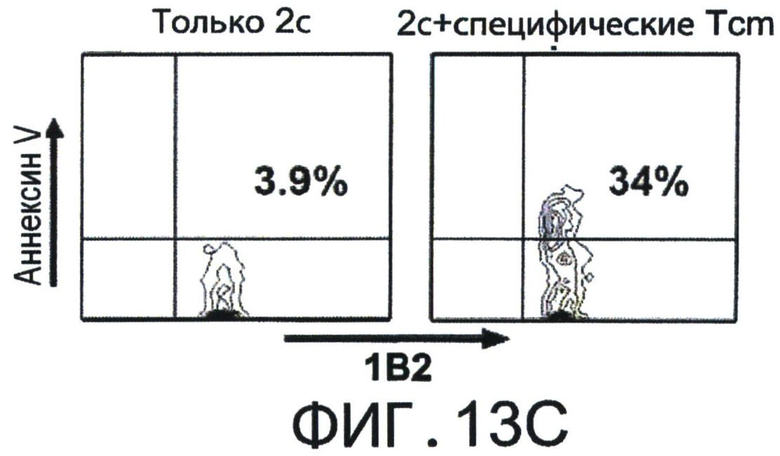
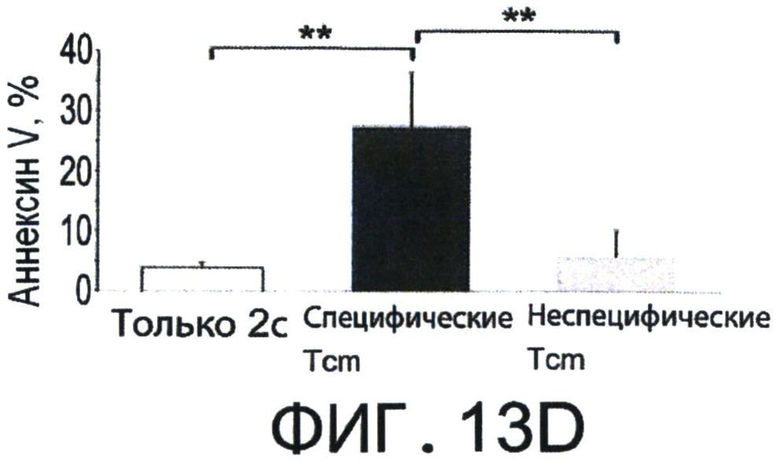
Изобретение относится к области биотехнологии, конкретно к получению популяции Т-лимфоцитов центральной памяти, и может быть использовано в медицине. Получают популяцию клеток, имеющих фенотип Т-лимфоцитов центральной памяти (Тcm) и не индуцирующих GVHD против третьей стороны, в которой по меньшей мере 50% клеток имеет сигнатуру CD8+, CD62L+, CD45RA-, CD45 RO+. Изобретение позволяет получить популяцию индуцирующих толерантность клеток, которые способны к хоумингу в лимфоузлы после трансплантации. 2 н. и 13 з.п. ф-лы, 13 ил., 9 пр.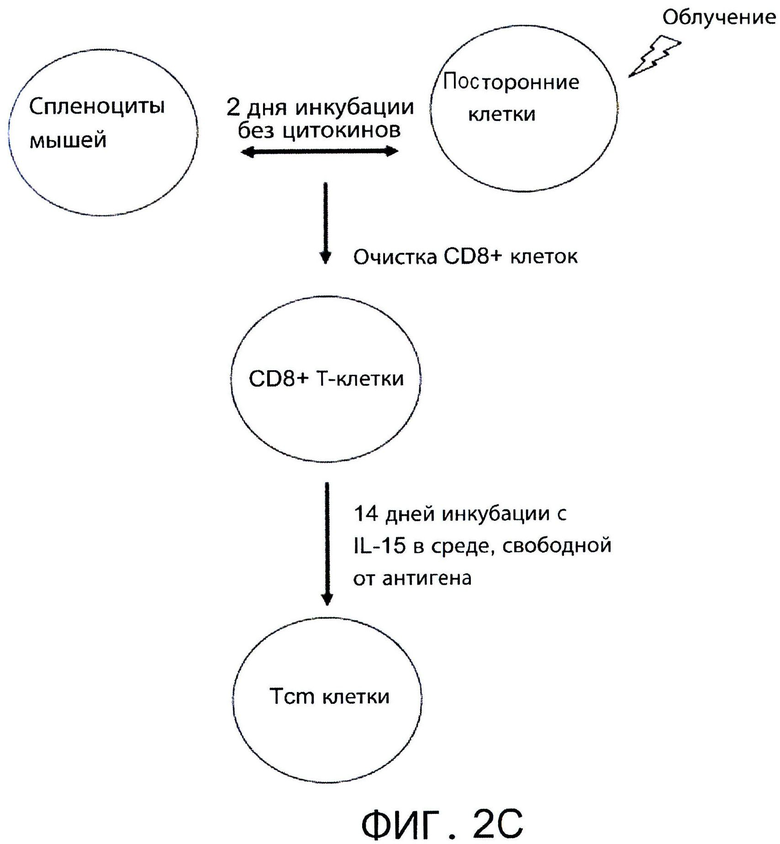
1. Изолированная популяция клеток, включающая не индуцирующие GVHD клетки против третьей стороны, имеющие фенотип Т-лимфоцитов центральной памяти (Тcm), причем указанные Т-лимфоциты центральной памяти (Tcm) содержат сигнатуру CD8+, CD62L+, CD45RA-, CD45 RO+, и причем, по меньшей мере, 50% изолированной популяции клеток имеет указанную сигнатуру, причем указанные клетки являются клетками, индуцирующими толерантность и способными к хоумингу в лимфоузлы после трансплантации.
2. Изолированная популяция клеток по п.1 для применения при лечении человеческого субъекта, нуждающегося в этом, причем указанное заболевание является выбранным из группы, состоящей из злокачественного заболевания, вирусного заболевания и аутоиммунного заболевания.
3. Изолированная популяция клеток по п.1 для применения в качестве адъювантного лечения при клеточной или тканевой трансплантации человеческому субъекту, нуждающемуся в клеточной или тканевой трансплантации.
4. Изолированная популяция клеток по п.3, где указанную изолированную популяцию клеток вводят до, одновременно или после указанного клеточного или тканевого трансплантата.
5. Изолированная популяция клеток по п.3, где указанная изолированная популяция клеток является сингенной по отношению к субъекту.
6. Изолированная популяция клеток по п.3, где указанная изолированная популяция клеток является несингенной по отношению к субъекту.
7. Изолированная популяция клеток по п.3, где указанный клеточный или тканевый трансплантат получен от донора, выбранного из группы, состоящей из HLA-идентичного аллогенного донора, HLA-неидентичного аллогенного донора и ксеногенного донора.
8. Изолированная популяция клеток по п.3, где указанный клеточный или тканевый трансплантат является сингенным по отношению к субъекту.
9. Изолированная популяция клеток по п.3, где указанный клеточный или тканевый трансплантат содержит незрелые гематопоэтические клетки.
10. Изолированная популяция клеток по п.3, где указанный клеточный или тканевый трансплантат включает сотрансплантацию нескольких органов.
11. Изолированная популяция клеток по п.3, где указанный клеточный или тканевый трансплантат и указанная изолированная популяция клеток являются полученными от одного донора.
12. Изолированная популяция клеток по любому из пп.1, 2 или 3, где указанную изолированную популяцию клеток, включающую не индуцирующие GVHD клетки против третьей стороны, имеющие фенотип Т-лимфоцитов центральной памяти (Тcm), получают с помощью: (а) контактирования несингенных мононуклеарных клеток периферической крови (РВМС) с посторонним антигеном или антигенами в условиях, которые делают возможной элиминацию GVH реактивных клеток; и (b) культивирования указанных клеток, полученных на стадии (а), в присутствии IL-15 в условиях, которые делают возможной пролиферацию клеток, имеющих указанный фенотип Т-лимфоцитов центральной памяти (Тcm).
13. Способ получения изолированной популяции клеток по п.1, включающий:
(а) контактирование несингенных мононуклеарных клеток периферической крови (РВМС) с посторонним антигеном или антигенами в условиях, которые делают возможной элиминацию GVH реактивных клеток; и
(b) культивирование указанных клеток, полученных на стадии (а), в присутствии IL-15 в условиях, которые делают возможной пролиферацию клеток, имеющих указанный фенотип Т-лимфоцитов центральной памяти (Тcm).
14. Способ по п.13, где указанные условия, которые делают возможной элиминацию GVH реактивных клеток, включают:
(i) культивирование в отсутствие цитокинов; и/или
(ii) по меньшей мере, 2 дня культивирования.
15. Способ по п.13, где указанные условия, которые делают возможной пролиферацию клеток, имеющих указанный фенотип Т-лимфоцитов центральной памяти (Тcm), включают:
(i) IL-7 и/или IL-21
(ii) по меньшей мере, 14 дней указанного культивирования; и/или
(iii) среду, свободную от антигена.
| US 20060062763 A1, 23.03.2006 | |||
| KAWAI Т | |||
| ЕТ AL | |||
| HLA-mismatched renal transplantation without maintenance immunosupression, THE NEW ENGLAND JOURNAL OF MEDICINE, 2008, v.358, № 4, p.353-361 | |||
| SCANDLING J | |||
| D | |||
| et al | |||
| Tolerance and chimerism after renal and hematopoietic-cell transplantation, THE NEW ENGLAND JOURNAL OF MEDICINE, 2008, v.358, № |

