Изобретение относится к области биотехнологии, а именно к получению рекомбинантного проинсулина человека, и может быть использовано в медицине.
Проинсулин синтезируется в шероховатом эндоплазматическом ретикулуме b-клеток островков Лангерганса поджелудочной железы в виде предшественника - препроинсулина (молекулярная масса 11500 Да). Лидерная последовательность, состоящая из 23 аминокислотных остатков, направляет молекулу-предшественник в аппарат Гольджи и там отщепляется. В результате образуется молекула проинсулина (молекулярная масса около 9000 Да), принимающая конформацию, необходимую для правильного образования дисульфидных мостиков. Затем проинсулин расщепляется в нескольких специфических участках с образованием зрелого инсулина и С-пептида, которые депонируются в секреторных гранулах.
Последние медицинские исследования демонстрируют, что проинсулин может блокировать разрушение b-клеток поджелудочной железы, действуя в качестве локального протектора, а также стимулировать их регенерацию. Поэтому разработка способов получения рекомбинантного проинсулина человека для профилактики и лечения сахарного диабета крайне актуальна.
Известен способ получения рекомбинантного проинсулина человека [1-3], который включает лизис клеток Е.coli, экспрессирующих гибридные белки с проинсулином человека в форме телец включения, растворение телец включения в буферном растворе, содержащем мочевину или гуанидин хлорид, сульфирование гибридных белков сульфонатом натрия и тетратионатом натрия, центрифугирование гибридного белка S-сульфоната проинсулина, отщепление S-сульфоната проинсулина от лидерного пептида путем обработки бромистым цианом, очистку S-сульфоната проинсулина анионообменной хроматографией, повторную укладку цепи S-сульфоната проинсулина и очистку проинсулина человека адсорбционной хроматографией.
Недостатками описанного способа являются многостадийность технологического процесса и использование при расщеплении гибридного белка высокотоксичного реагента - бромистого циана.
Поэтому в настоящее время существует необходимость в разработке усовершенствованных способов получения рекомбинантного проинсулина человека, которые были бы высокопроизводительными и нетоксичными.
Для получения рекомбинантных белков из гибридных полипептидов известны подходы, в которых отщепление конечного продукта проводится при помощи высокоспецифических протеолитических ферментов. Такие способы предусматривают конструирование гибридных белков, содержащих сайт узнавания специфической протеиназы между белком-носителем (лидерным пептидом) и целевым белком, очистку синтезированного гибридного белка и его ферментативное расщепление. Одной из высокоспецифических протеиназ, используемых для этой цели, является энтерокиназа (энтеропептидаза, КФ 3.4.21.9), которая расщепляет пептидную связь после последовательности (Asp)4Lys.
Описание изобретения
Настоящее изобретение относится к гибридному белку - предшественнику проинсулина человека с последовательностью, представленной на фиг.2, включающей аминокислотные последовательности лидерного пептида, представляющего собой N-концевой фрагмент гамма-интерферона человека (SEQ ID NO:1), пептидного линкера (SEQ ID NO:2), содержащего сайт расщепления энтерокиназы и проинсулина человека (SEQ ID NO:3).
Кроме того, изобретение относится к ДНК, кодирующей указанный гибридный белок, с последовательностью нуклеотидов, представленной на фиг.2.
Изобретение относится к рекомбинантной плазмиде, содержащей указанную ДНК, кодирующей указанный гибридный белок с проинсулином человека, а также включающей регуляторные элементы. В качестве регуляторных элементов и генетических маркеров, которые используются в плазмиде по настоящему изобретению, можно привести следующие неограничивающие элементы и маркеры: tac-промотор транскрипции, терминатор транскрипции рибосомного оперона Е.coli, ген β-лактамазы (bla), использующийся в качестве генетического маркера, определяющий устойчивость трансформированных плазмидой pHINS21 клеток бактерий к ампициллину.
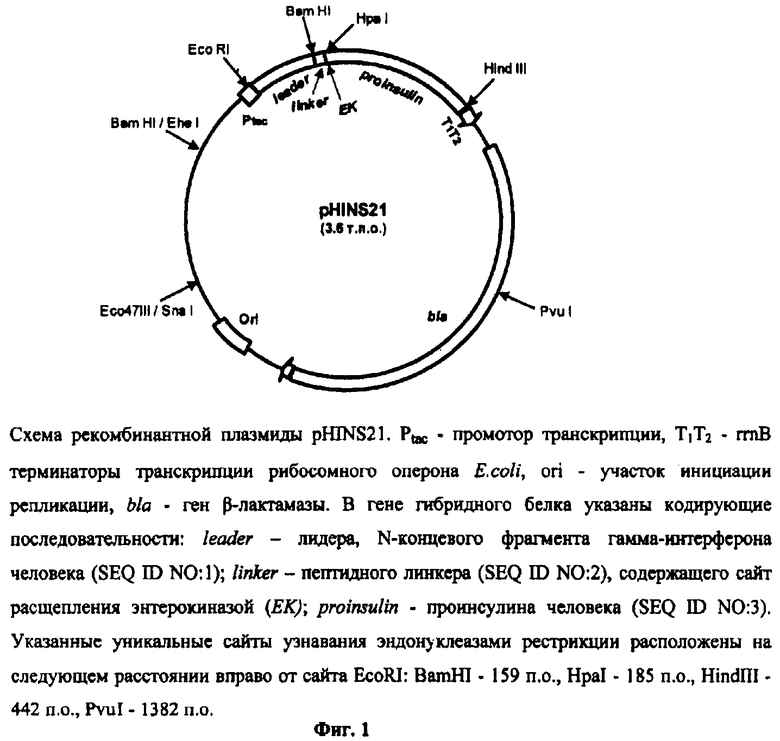
В частном случае, изобретение относится к рекомбинантной плазмиде pHINS21 (фиг.1), содержащей указанную ДНК, кодирующую гибридный белок (фиг.2), в котором аминокислотные последовательности лидерного пептида, представляющего собой N-концевой фрагмент гамма-интерферона человека, и проинсулина человека соединены пептидным линкером, содержащим сайт расщепления энтерокиназы, при этом указанная плазмида имеет молекулярную массу 2,36 МДа и включает уникальные сайты узнавания эндонуклеазами рестрикции, расположенные на следующем расстоянии вправо от сайта ЕсоRI: BamHI - 159 п.о., HpaI - 185 п.о., HindIII - 442 п.о., PvuI - 1382 п.о.
Новый гибридный белок обеспечивает высокий выход целевого продукта - проинсулина человека за счет эффективной ренатурации и эффективного расщепления энтерокиназой указанного гибридного белка.
Также изобретение относится к трансформированным клеткам Escherichia coli, содержащим указанную рекомбинантную плазмиду, продуцент гибридного белка проинсулина человека. В частном случае, изобретение относится к штамму бактерий Escherichia coli JM109/pHINS21 - продуценту гибридного белка проинсулина человека.
Кроме того, заявленный гибридный белок с проинсулином человека, например, продуцируемый штаммом E.coli JM109/pHINS21, может быть использован в способе получения инсулина человека. При этом новый гибридный белок трансформируют в инсулин путем ферментативного расщепления указанного гибридного белка энтерокиназой, трипсином и карбоксипептидазой Б.
Изобретение относится к новому способу получения проинсулина человека. Заявляемый способ получения проинсулина человека включает культивирование штамма-продуцента Escherichia coli, разрушение бактериальных клеток, выделение телец включения, их растворение в буфере, содержащем мочевину и дитиотреитол, ренатурацию и очистку ренатурированного гибридного белка с проинсулином человека, его расщепление специфической протеиназой с последующей очисткой и получением целевого продукта. В частном случае, способ включает культивирование нового штамма E.coli JM109/pHINS21, несущего новую плазмиду pHINS21, расщепление гибридного белка проводят энтерокиназой (или ее каталитической субъединицей), а полученный после ферментативного расщепления проинсулин очищают ионообменной хроматографией на SP-сефарозе с последующей высокоэффективной жидкостной хроматографией на обращенных фазах.
Преимуществом заявленного способа получения проинсулина человека является упрощение технологического процесса и исключение из технологии высокотоксичного реагента.
Подробное описание изобретения
Была создана новая рекомбинантная плазмида pHINS21, которая определяет синтез гибридного белка с молекулярной массой около 15,85 кДа, в котором аминокислотные последовательности лидерного пептида, представляющего собой N-концевой фрагмент гамма-интерферона человека, и проинсулина человека соединены пептидным линкером, содержащим сайт расщепления энтерокиназы. Новый гибридный белок, свойства которого определяют его эффективную ренатурацию и эффективное ферментативное расщепление энтерокиназой, обеспечивает высокий выход целевого продукта - проинсулина человека.
Оптимальная длина линкера и его аминокислотный состав были определены экспериментально.
Рекомбинантную плазмиду pHINS21 конструировали на основе известной плазмиды pHINS05 [4]. ДНК плазмиды pHINS05 была подвергнута полному гидролизу рестриктазами BclI и HpaI, а полученный фрагмент BclI-HpaI (4,2 т.п.о.) лигировали с олигонуклеотидным дуплексом, полученным в результате отжига синтетических олигонуклеотидов: смыслового, представленного в последовательности SEQ ID NO:4, и антисмыслового, представленного в последовательности SEQ ID NO:5. В результате в ген гибридного белка перед ДНК проинсулина человека встраивается нуклеотидная последовательность, кодирующая сайт расщепления энтерокиназы, а именно пептидный фрагмент (Asp)4Lys.
Лигированную смесь использовали для трансформации компетентных клеток Е.coli JM109. В результате получали рекомбинантную плазмиду pHINS20, строение которой подтверждали рестрикционным анализом. Структура гена (фиг.2), кодирующего гибридный белок, была подтверждена секвенированием.
Для увеличения числа копий полученной плазмиды pHINS20 в бактериальной клетке ее делетировали по rop-гену (негативному регулятору копийности) [5]. С этой целью ДНК плазмиды pHINS20 подвергали полному гидролизу рестриктазами Eco47III и SnaI, а полученный фрагмент Eco47III-SnaI (3,6 т.п.о.) лигировали. В результате получали рекомбинантную плазмиду pHINS21, строение которой подтверждали рестрикционным анализом и секвенированием.
Новая рекомбинантная плазмида pHINS21 кодирует гибридный белок размером 143 аминокислоты с молекулярной массой 15,85 кДа, в котором аминокислотные последовательности лидерного пептида, представляющего собой N-концевой фрагмент гамма-интерферона человека (SEQ ID NO:1), и проинсулина человека (SEQ ID NO:3) соединены пептидным линкером (SEQ ID NO:2), содержащим сайт расщепления энтерокиназы.
Указанная плазмида состоит из фрагмента BamHI-EcoRI плазмиды рКК223-3 [6], содержащего промотор транскрипции tac; фрагмента EcoRI-HindIII, включающего ген гибридного белка с последовательностью нуклеотидов, представленной на фиг.2, кодирующий N-концевой фрагмент гамма-интерферона человека, пептидный линкер с сайтом расщепления энтерокиназы и аминокислотную последовательность проинсулина человека; фрагмента HindIII-SnaI плазмиды рКК223-3, содержащего терминатор транскрипции рибосомного оперона Е.coli, ген β-лактамазы (bla), участок инициации репликации (ori); и Eco47III-EheI фрагмента плазмиды рКК223-3; уникальные сайты узнавания рестрикционными эндонуклеазами, расположенные на следующем расстоянии вправо от сайта ЕсоRI: BamHI - 159 п.о., HpaI - 185 п.о., HindIII - 442 п.о., PvuI - 1382 п.о.
Штамм-продуцент E.coli JM109/pHINS21 получают путем трансформации клеток E.coli штамма JM109 плазмидой pHINS21. После трансформации отбирают колонии, выращенные на среде с ампициллином, выделяют из них плазмиды и подвергают их рестрикционному анализу и секвенированию. Линию клеток, несущую плазмиду pHINS21, несколько раз пересевают на среду с агарозой с добавлением ампициллина и полученной моноклоновой культурой инокулируют 5 мл жидкой среды с ампициллином. Культуру проверяют на наличие индуцируемой экспрессии гибридного белка, фасуют, добавляют глицерин и хранят при минус 40°C.
Новый штамм Escherichia coli JM109/pHINS21, несущий плазмиду pHINS21, является продуцентом гибридного белка, содержащего аминокислотную последовательность проинсулина человека, и характеризуется следующими признаками.
Морфологические признаки: клетки мелкие, палочковидной формы, грамотрицательные, неспороносные, размером 1Ч3,5 мкм, подвижные, с хорошо различимыми тельцами включения после индукции синтеза гибридного белка.
Культуральные признаки: при росте на агаризованной среде LB колонии круглые, гладкие, полупрозрачные, блестящие, серые. Край ровный, диаметр колоний 1-3 мм, консистенция пастообразная. Рост в жидких средах (LB, минимальная среда с глюкозой) характеризуется ровным помутнением.
Физиолого-биохимические признаки: клетки растут при температуре 4-42°С, оптимум рН 6,8-7,6. В качестве источника азота используют как минеральные соли аммония, так и органические соединения: аминокислоты, пептон, триптон, дрожжевой экстракт. В качестве источника углерода при росте на минимальной среде используют глицерин, углеводы, аминокислоты.
Устойчивость к антибиотикам: клетки штамма-продуцента проявляют устойчивость к ампициллину (до 500 мг/мл), обусловленную наличием в плазмиде гена β-лактамазы (bla).
Стабильность плазмиды в штамме. При поддержании клеток в течение нескольких месяцев на агаризованной среде LB, содержащей ампициллин, не наблюдаются потери или перестройки плазмиды, влияющие на экспрессию гибридного белка.
B новом штамме гибридный белок после индуцированной экспрессии накапливается в виде телец включения, и его содержание составляет не менее 30% от общего белка клетки.
Полученный штамм-продуцент E.coli JM109/pHINS21 депонирован в коллекции микроорганизмов ОАО "Национальные Биотехнологии" под номером 21-06.
Новая рекомбинантная плазмида pHINS21 обеспечивает эффективный биосинтез гибридного белка, содержащего проинсулин человека, и в других бактериальных штаммах Escherichia coli, например в штамме-хозяине E.coli BL21, как описано ниже.
В качестве неограничивающего примера в описании приведен способ получения проинсулина человека с использованием E.coli JM109/pHINS21. Условия культивирования штамма-продуцента и выделение телец включения не являются определяющими, и специалист в данной области на основании общих знаний подберет аналогичные условия, не влияющие на достижение технического результата.
Выращивание культуры штамма-продуцента E.coli JM109/pHINS21 проводят на питательной среде на основе гидролизата казеина и экстракта дрожжей Saccharomyces cerevisiae. Для индукции синтеза гибридного белка в середине логарифмической фазы роста вносят 1-изопропил-β-D-1-тиогалактопиранозид (ИПТГ). Культивирование продолжают до образования желательного уровня внутриклеточных включений гибридного белка (телец включения). После окончания выращивания культуру осаждают, клетки разрушают одним из приемлемых методов и тельца включения отделяют центрифугированием. Осадок телец включения (паста) содержит около 5-7% гибридного белка проинсулина.
Выделение гибридного белка из телец включения, его ферментативное расщепление и очистку проинсулина проводят по следующей схеме:
• тельца включения растворяют в 0,1М буфере трис-HCl, рН 8,0, содержащем 8М мочевину, с последующим добавлением дитиотреитола до конечной концентрации 10 мМ;
• гибридный белок ренатурируют в 10-кратном объеме 0,1М глицинового буфера, рН 9-11 при температуре 10-14°С в течение 20-24 час;
• очистку ренатурированного гибридного белка проводят хроматографией на DEAE-сефарозе FF, уравновешенной 0,05М трис-НСl буфером, рН 7,0-7,5, содержащим 0,15М NaCl. Белок элюируют линейным градиентом 0,15-0,4M хлористого натрия в уравновешивающем буфере;
• очищенный гибридный белок расщепляют энтерокиназой (или ее каталитической субъединицей) в течение от 12 до 24 час при рН 7,0-7,5 и температуре 16-26°C. Реакцию останавливают, подкисляя гидролизат соляной кислотой до рН 4,6-5,0, а образовавшийся осадок примесных белков отделяют центрифугированием;
• проинсулин очищают ионообменной хроматографией на SP-сефарозе FF в 0,05-0,2M аммоний-ацетатном буфере, рН 3,0-6,0, содержащем 1-2М мочевину. Сорбированный белок элюируют линейным градиентом хлористого калия от 0 до 0,5М в уравновешивающем буфере;
• дальнейшую очистку проинсулина проводят методом обращенно-фазовой высокоэффективной жидкостной хроматографии (ОФ ВЭЖХ).
Краткое описание фигур
На фиг.1 представлена физическая карта рекомбинантной плазмиды pHINS21.
На фиг.2 представлены нуклеотидная последовательность гена гибридного белка с происулином человека в составе плазмиды pHINS21 и кодируемая им аминокислотная последовательность.
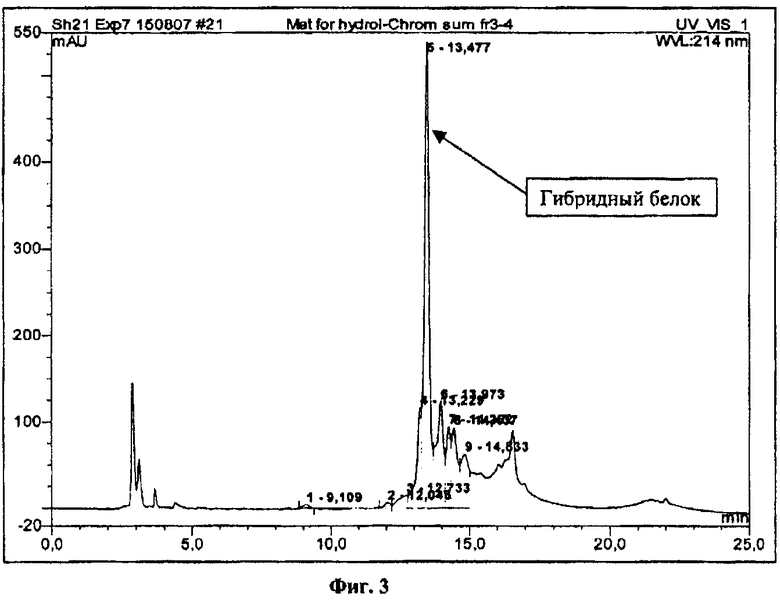
На фиг.3 показан ВЭЖХ анализ материала, полученного после очистки ренатурированного гибридного белка с проинсулином человека на DEAE-сефарозе.
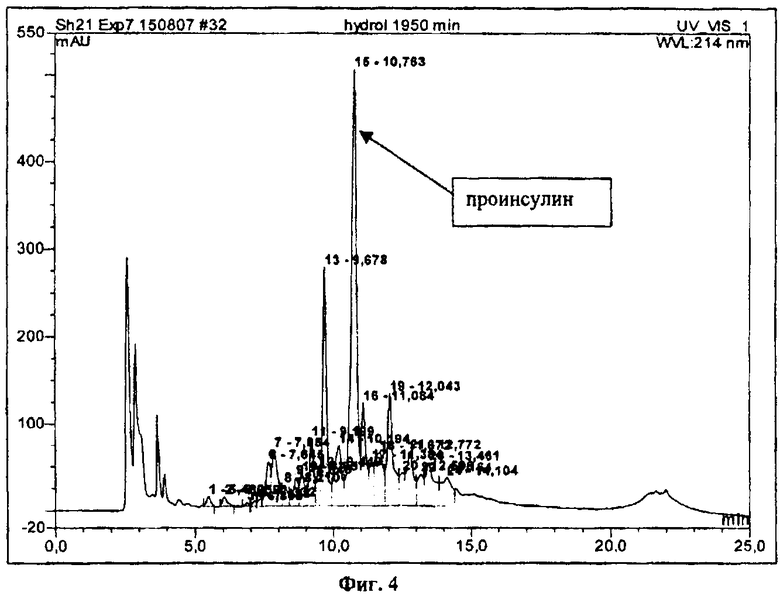
На фиг.4 показан ВЭЖХ анализ продуктов расщепления гибридного белка с проинсулином человека энтерокиназой.
Примеры
Изобретение иллюстрируется следующими не ограничивающими его примерами.
Пример 1. Конструирование плазмиды pHINS21
Плазмиду pHINS21 конструируют на основе известной плазмиды pHINS05 [4]. Плазмидную ДНК pHINS05 подвергают полному гидролизу рестриктазами BclI и HpaI. Для этого 5 мкг плазмидной ДНК рHINS05 в 20 мкл буфера, содержащего 33 мМ трис-ацетата, рН 7,9, 66 мМ К-ацетата, 10 мМ Mg-ацетата и 0,1 мг/мл БСА (буфер 1) и 10 ЕД рестриктазы BclI и HpaI инкубируют 1,0 час при 37°С. Из полученного гидролизата выделяют ДНК фрагмент BclI-HpaI размером около 4,2 т.п.о. при помощи электрофореза в 0,8% геле легкоплавкой агарозы. Далее ДНК депротеинизируют фенолом, смесью фенола с хлороформом (1:1), хлороформом, осаждают этиловым спиртом, растворяют в 20 мкл воды. Полученный фрагмент BclI-HpaI, содержащий ген гибридного белка и векторную часть плазмиды рHINS05, лигируют с 20-кратным молярным избытком олигонуклеотидного дуплекса, полученного в результате отжига синтетических олигонуклеотидов: смыслового, представленного в последовательности SEQ ID NO:4, и антисмыслового, представленного в последовательности SEQ ID NO:5.
В результате в ген гибридного белка перед ДНК проинсулина человека встраивается нуклеотидная последовательность, кодирующая сайт расщепления энтерокиназы.
Компетентные клетки штамма E.coli JM109 трансформируют лигированной смесью (10 мкл) и высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Из выросших колоний выделяют плазмидную ДНК и секвенируют между сайтами рестриктаз EcoRI и HpaI для подтверждения вставки в линкерную часть гена гибридного белка. В результате получают плазмиду рHINS20.
Для получения плазмиды с делецией по rop-гену (негативного регулятора копийности) к 5 мкг плазмидной ДНК рHINS20 в 20 мкл буфера 1 добавляют 10 ЕД рестриктазы Eco47III и SnaI, генерирующие ”тупые” концы, и смесь инкубируют 1 час при 37°С. Далее ДНК депротеинизируют фенолом, смесью фенола с хлороформом (1:1), хлороформом, осаждают этиловым спиртом, растворяют в 20 мкл воды. Полученную таким образом ДНК лигируют в 30 мкл буфера для лигирования в присутствии 5 Ед ДНК лигазы фага Т4 в течение 16 час при 8°С. Лигированной смесью (10 мкл) трансформируют компетентные клетки штамма E.coli JM109 и высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Отбирают бактериальные клоны, несущие плазмидную ДНК размером 3,6 т.п.о. Выделенные плазмиды подвергают рестрикционному анализу и секвенируют по методу Сенгера. В результате получают плазмиду pHINS21.
Пример 2. Получение штамма E.coli JM109/pHINS21 и определение уровня его продуктивности
Плазмидой pHINS21 трансформируют компетентные клетки штамма E.coli JM109 и высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Отдельно локализованную колонию трижды пересевают на чашки с LB-агаром, содержащим 100 мкг/мл ампициллина. Полученной моноклоновой культурой инокулируют 5 мл жидкой среды LB с ампициллином и инкубируют в течение ночи при интенсивном встряхивании при 37°C.
Полученный штамм-продуцент E.coli JM109/pHINS21 хранят в 20% глицерине при минус 40°C.
Для определения уровня индуцируемой экспрессии гибридного белка ночную культуру засевают в разведении 1:50 в 5 мл жидкой среды LB, содержащей 100 мкг/мл ампициллина, и растят до мутности 0,8 при 37°С на качалке при 200 об/мин. К культуре добавляют ИПТГ до концентрации 1,0 мМ и продолжают инкубацию в тех же условиях в течение 3 час. Клетки собирают центрифугированием, осадок суспендируют в буфере, содержащем 62,5 мМ трис-HCl, рН 6,8, 3% додецилсульфата натрия, 5% 2-меркаптоэтанола, 10% глицерина и 0,01% бромфенолового синего и прогревают 3 мин на кипящей водяной бане. Полученный лизат клеток анализируют электрофорезом в 18% полиакриламидном геле с додецилсульфатом натрия [7]. Гель окрашивают Coomassie R-250, сканируют и проводят его денситометрию. По данным денситометрии содержание гибридного белка составляет не менее 30% от общего белка клетки.
Пример 3. Получение штамма E.coli BL21/pHINS21 и определение уровня его продуктивности
Компетентные клетки штамма E.coli BL21 трансформируют плазмидной ДНК pHINS21 и высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Отдельно локализованную колонию трижды пересевают на чашки с LB-агаром, содержащим 100 мкг/мл ампициллина. Полученной моноклоновой культурой инокулируют 5 мл жидкой среды LB и инкубируют в течение ночи при интенсивном встряхивании при 37°C.
Полученный штамм-продуцент E.coli BL21/pHINS21 хранят в 20% глицерине при минус 40°C.
Определение продуктивности штамма Е.coli BL21/pHINS21 проводят таким же методом, как и для штамма Е.coli JM109/pHINS21, описанным в примере 2. По данным денситометрии содержание гибридного белка в индуцированных клетках полученного штамма составляет не менее 30% от общего белка клетки.
Пример 4. Получение проинсулина человека при использовании штамма-продуцента E.coli JM109/pHINS21
Этап 1. Выделение телец включения, содержащих гибридный белок с проинсулином человека
Выращивают культуру штамма E.coli JM109/pHINS21 на питательной среде на основе гидролизата казеина и экстракта пекарских дрожжей. Индукцию биосинтеза гибридного белка проводят внесением индуктора ИПТГ в середине логарифмической фазы роста культуры. Культивирование продолжают до образования внутриклеточных включений гибридного белка. После завершения выращивания культуру осаждают и используют для выделения телец включения, содержащих гибридный белок проинсулина.
Клетки суспендируют в буфере А, содержащем 50 мМ натрия фосфорнокислого двузамещенного, 1 мМ ЭДТА, 0,2М хлорида натрия из расчета на 1 г биомассы 5-10 мл буфера и разрушают на дезинтеграторе Гаулина. Разрушенные клетки центрифугируют в течение 20 мин на Beckman J-30I при 8000 об/мин. Полученный осадок ресуспендируют в буфере А, содержащем 1% Triton X-100, инкубируют в течение 1 час и суспензию центрифугируют в течение 20 мин на Beckman J-30I при 8000 об/мин. Осадок повторно ресуспендируют в буфере А и осаждают на центрифуге. Отмытый осадок телец включения (паста), содержащий около 5% гибридного белка проинсулина, фасуют, замораживают и хранят при минус 40°С.
Этап 2. Ренатурация и выделение гибридного белка
5 г размороженной пасты телец включения растворяют в 100 мл буферного раствора, содержащего 0,1М трис-HCl, рН 8,0, 10 мМ дитиотреитол и 8М мочевину, и инкубируют при перемешивании в течение 18-24 час при 14°С. Раствор центрифугируют в течение 20 мин на Beckman J-30I при скорости 8000 об/мин при 14°С. Ренатурацию гибридного белка, содержащегося в супернатанте, проводят 10-кратным разбавлением в 0,1М глициновом буфере, рН 9-11. Раствор гибридного белка инкубируют в течение 20-24 час, перемешивая и поддерживая температуру 10-14°С. Процесс ренатурации гибридного белка контролируют методом ОФ ВЭЖХ. После образования правильно замкнутых S-S связей у 50-70% молекул гибридного белка раствор подкисляют 2 н. соляной кислотой до рН 7,0-7,2, а образующийся осадок отделяют центрифугированием.
Содержание ренатурированного гибридного белка в супернатанте составляет 112 мг.
Раствор гибридного белка наносят на колонку объемом 100 мл, заполненную DEAE-сефарозой FF, предварительно уравновешенную 0,05М трис-НСl буфером, рН 7,0-7,5. Колонку промывают уравновешивающим буфером, содержащим 0,15М NaCl. Сорбированный белок элюируют линейным градиентом 0,15-0,4M хлорида натрия в уравновешивающем буфере. Фракции, содержащие гибридный белок, анализируют методом ОФ ВЭЖХ (фиг.3). В результате получают 95 мг очищенного гибридного белка.
Соответствие полученного гибридного белка с проинсулином человека подтверждали методом масс-спектрометрии по совпадению с расчетной молекулярной массой указанного белка.
Этап 3. Ферментативное расщепление гибридного белка
Реакцию ферментативного гидролиза проводят при температуре 25°С при постоянном перемешивании. К раствору очищенного гибридного белка добавляют CaCl2 до концентрации 1 мМ. Затем в реакционную смесь вносят раствор рекомбинантной энтерокиназы (каталитической субъединицы энтерокиназы быка) (Novagen) из расчета 1 ЕД на 4 мг белка и инкубируют в течение 20 час. Продукты гидролиза гибридного белка анализируют методом ОФ ВЭЖХ. На фиг.4 представлен ВЭЖХ анализ продуктов расщепления гибридного белка энтерокиназой после 20 час инкубации. Реакцию расщепления останавливают, подкисляя материал 2 н. соляной кислотой до рН 4,0-5,0. Образовавшийся осадок примесных белков отделяют центрифугированием.
Этап 4. Очистка проинсулина человека
Осветленный раствор, содержащий проинсулин, наносят на хроматографическую колонку объемом 20 мл, заполненную SP-сефарозой FF, предварительно уравновешенную 0,03М аммоний-ацетатным буфером, рН 5,0, с 2М мочевиной. Колонку промывают уравновешивающим буфером до достижения базовой линии контрольного проточного денситометра. Элюцию сорбированного проинсулина проводят линейным градиентом хлористого калия от 0 до 0,5М в уравновешивающем буфере. Содержание и чистоту проинсулина человека в собираемых фракциях определяют методом ОФ ВЭЖХ. Объединенные фракции содержат 54 мг проинсулина человека с чистотой не менее 85%.
Дальнейшую очистку проинсулина проводят методом ОФ ВЭЖХ на препаративном хроматографе ”Armen” (Франция). Колонку объемом 200 мл, заполненную Кромасилом С18, уравновешивают 20%-ным изопропанолом с 0,1% трифторуксусной кислотой и подают раствор проинсулина с предыдущей стадии очистки в количестве 54 мг белка. Белок элюируют градиентом изопропанола (с 0,1% трифторуксусной кислотой) от 20 до 40%. Собранные фракции основного пика проинсулина анализируют на содержание примесей методом ОФ ВЭЖХ.
После очистки получают 46 мг высокоочищенного проинсулина человека с содержанием основного вещества 98%.
Подлинность полученного целевого продукта подтверждена по совпадению расчетной молекулярной массы проинсулина человека с молекулярной массой целевого продукта, определенной методом масс-спектрометрии.
Пример 5
Полученный в примере 4 (этап 2) гибридный белок с проинсулином человека расщепляли энтерокиназой из природного источника.
Реакцию ферментативного расщепления гибридного белка проводят при температуре 25°С при постоянном перемешивании. К раствору очищенного гибридного белка добавляют CaCl2 до концентрации 1 мМ. Затем в реакционную смесь вносят раствор энтерокиназы (Sigma, США) из расчета 1 ЕД на 4 мг белка и инкубируют в течение 16 час. Продукты гидролиза гибридного белка анализируют методом ОФ ВЭЖХ. Реакцию расщепления останавливают, подкисляя материал 2 н. соляной кислотой до рН 4,0-5,0.
Источники информации
1. Патент РФ 2203949. Способ получения проинсулина человека.
2. Патент США 5952461. Process for preparing human proinsulin.
3. Европейский патент EP1042479. A process for preparing human proinsulin.
4. Патент РФ 2263147 C1, 27.10.2005. Бюл. № 30.
5. Twigg A.J. and Sherrat D.//Nature, 1980, v.283, p.216-218.
6. Brosius J., Dull T.J., Sleeter D.D., Noller H.F.//J. Mol. Biol., 1981, v.148, p.107-127.
7. Laemmli U.K.//Nature, 1970, v.227, p.680-687.


Изобретение относится к области биотехнологии, а именно к получению рекомбинантного проинсулина человека, и может быть использовано в биомедицинской промышленности. Получен рекомбинантный штамм Escherichia coli, содержащий плазмиду pHINS21 (Escherichia coli JM109/ pHINS21), которая определяет синтез гибридного белка, состоящего из N-концевого фрагмента гамма-интерферона человека и проинсулина человека, соединенных пептидным линкером, содержащим сайт расщепления энтерокиназой (Asp4Lys). Выход включающего проинсулин гибридного продукта, обеспечиваемый новым штаммом-продуцентом, составляет не менее 30% от суммарного количества клеточного белка. Предложен способ получения проинсулина человека, включающий культивирование штамма-продуцента Escherichia coli JM109/pHINS21, выделение телец включения и их растворение, ренатурацию гибридного белка и его очистку ионообменной хроматографией, расщепление гибридного белка энтерокиназой или ее каталитической субъединицей и очистку проинсулина ионообменной хроматографией на сорбентах с сульфпропильными группами. Изобретение упрощает технологический процесс получения рекомбинантного проинсулина человека и улучшает условия его проведения с точки зрения техники безопасности. 2 н. и 2 з.п. ф-лы, 4 ил.
1. Штамм бактерий Escherichia coli JM109/pHINS21 - продуцент гибридного белка, содержащего проинсулин человека.
2. Способ получения проинсулина человека, включающий
культивирование штамма-продуцента по п.1,
разрушение бактериальных клеток,
выделение телец включения, их растворение в буфере, содержащем мочевину и дитиотреитол,
ренатурацию и очистку ренатурированного гибридного белка с проинсулином человека,
расщепление гибридного белка энтерокиназой или ее каталитической субъединицей,
очистку и получение целевого продукта.
3. Способ по п.2, отличающийся тем, что
полученный после ферментативного расщепления проинсулин очищают ионообменной хроматографией на сорбентах с сульфопропильными группами.
4. Способ по п.3, отличающийся тем, что
после ферментативного расщепления гибридного белка очистку проинсулина проводят хроматографией на SP-сефарозе FF в 0,05-0,2М аммоний-ацетатном буфере, рН 3,0-6,0, содержащем 1-2М мочевину, а сорбированный белок элюируют линейным градиентом хлористого калия от 0 до 0,5М в уравновешивающем буфере.
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pHINS05, КОДИРУЮЩАЯ ГИБРИДНЫЙ ПОЛИПЕПТИД, СОДЕРЖАЩИЙ ПРОИНСУЛИН ЧЕЛОВЕКА, КЛЕТКА ESCHERICHIA COLI, ТРАНСФОРМИРОВАННАЯ РЕКОМБИНАНТНОЙ ПЛАЗМИДНОЙ ДНК pHINS05, И ШТАММ БАКТЕРИЙ ESCHERICHIA COLI JM109/pHINS05 - ПРОДУЦЕНТ ГИБРИДНОГО ПОЛИПЕПТИДА, СОДЕРЖАЩЕГО ПРОИНСУЛИН ЧЕЛОВЕКА | 2004 |
|
RU2263147C1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| AKIYOSHI-SHIBATA M | |||
| ET AL., Eur | |||
| J | |||
| Biochem., 224, 335-343, 1994. | |||

