Область техники
Заявляемая группа изобретений относится к получению гетерогенных биокатализаторов для ферментативного синтеза аминобета-лактамного антибиотика.
Уровень техники
Гидролазами эфиров альфа-аминокислот (АЭГ) называют ферменты, катализирующие гидролиз эфиров альфа-аминокислот, гидролиз ациламидной связи различных цефалоспоринов и пенициллинов, а также N-ацилирование 7-аминоцефема и 6-аминопенама производными альфа-аминокислот (эфирами или амидами). Подобная субстратная специфичность позволяет использовать эти ферменты в качестве биокатализаторов процессов синтеза цефалоспоринов и пеницициллинов, содержащих в боковой цепи аминогруппу в альфа-положении аминоцефалоспоринов и аминопенициллинов, составляющих группу аминобета-лактамных антибиотиков.
Исследования структуры АЭГ показали, что эти ферменты относятся к классу α/β-гидролаз и содержат в активном центре каталитическую триаду Ser-His-Asp.По структуре, строению активного центра, субстратной специфичности и каталитическим свойствам АЭГ принципиально отличаются от таких ацилаз бета-лактамов как, например, пенициллин G ацилаза, также используемых для синтеза бета-лактамных антибиотиков. Эти ферменты структурно относятся к классу N-концевых гидролаз, содержащих в активном центре N-концевой остаток β-цепи, выступающий нуклеофилом при каталитическом акте.
Применительно к синтезу аминобета-лактамных антибиотиков известны ферменты из бактерий семейства Pseudomonadaceae, являющиеся гидролазами эфиров альфа-аминокислот [GB 1382255, Jpn. J.Antibiot Suppl. 30S: 230 (1977), CA 1044627, RU 2136759, KR 20010069097]. Наиболее полно изучены биохимические, физико-химические и каталитические свойства АЭГ из Acetobacter turbidans [Enzyme Microb. Technol. 9:339 (1987)], Xanthomonas citri [Agric. Biol. Chem. 44: 1069 (1980); Agric. Biol. Chem. 44: 1663 (1980)], Xanthomonas rubrilineans [Биохимия 55 (12): 2226 (1990); Enzyme Microb. Technol. 15: 965 (1993)], Pseudomonas melanogenum [BBA - Protein Struct.& Molecul. Enzymol. 1040 (1): 12 (1990)]. Показано, что эти ферменты обладают субъединичной структурой. Фермент из Pseudomonas melanogenum состоит из двух субъединиц с молекулярной массой 70000-72000 каждая, ферменты из Acetobacter turbidans, Xanthomonas citri и Xanthomonas rubrilineans состоят из четырех субъединиц такой же молекулярной массы. Биохимические и каталитические свойства АЭГ из различных источников близки. Установлена кристаллическая структура, определен состав активного центра и клонированы гены для ферментов из Xanthomonas citri [WO 02086111; J. Biol. Chem. 278: 23076 (2003); WO 02086127] и Acetobacter turbidans [WO 02086111; J.Biol. Chem. 277: 28474 (2002)].
Разработка методов ферментативного синтеза антибиотиков с использованием АЭГ требует создания технологических гетерогенных биокатализаторов, в том числе путем иммобилизации выделенных из биомассы клеток ферментов. Процедуры получения биокатализаторов на основе АЭГ состоят из трех основных этапов: (1) выделение фермента из биомассы клеток путем ее разрушения и экстракции фермента в водную фазу (получение бесклеточного экстракта); (2) получение ферментного препарата той или иной степени очистки; (3) иммобилизация фермента на каком-либо носителе.
Известны способы разрушения биомассы бактериальных клеток, содержащих АЭГ, под давлением (French Press) [Enzyme Microb. Technol. 15: 965 (1993); Enzyme Microb. Technol. 25: 336 (1999)] или путем гомогенизации [BBA - Protein Struct.& Molecul. Enzymol. 1040 (1): 12 (1990)]. Эти способы приводят к тотальному разрушению клеточных структур и позволяют практически полностью экстрагировать целевой фермент из разрушенной клеточной биомассы, обеспечивая высокий выход активности на стадии получения бесклеточного экстракта (до 100%). Однако белки и другие водорастворимые вещества также легко переходят в водную фазу из полностью разрушенных клеток, поэтому получаемый бесклеточный экстракт содержит много балластных веществ и имеет низкую удельную активность. Описан способ разрушения биомассы бактериальных клеток, содержащих АЭГ, с помощью ультразвука в присутствии поверхностно-активных веществ [Biotechnol. Bioeng. 23 (2): 361 (1981)]. При использовании этого относительно мягкого способа клетки разрушаются не полностью, что приводит к снижению выхода активности при экстракции фермента, однако обеспечивает более высокую чистоту беклеточного экстракта. Способ разрушения биомассы бактериальных клеток, содержащих АЭГ, под действием несмешивающегося с водой органического растворителя бутилацетата [Биохимия 55 (12): 2226 (1990); Zaslavskaya P.L.,. Biotechnol. Appl. Biochem. 18: 299 (1993)] действует более избирательно, чем обсуждавшиеся выше, однако его эффективность характеризуется недостаточно высоким выходом активности.
Сопоставление различных способов разрушения биомассы клеток бактерий рода Xanthomonas - продуцентов АЭГ
В таблице 1 сопоставлены различные способы разрушения биомассы клеток бактерий рода Xanthomonas с целью экстракции из них фермента АЭГ. Величины удельной активности бесклеточного экстракта даны в международных единицах (ME)* (* - За международную единицу (ME) ферментативной активности препарата в отношении какой-либо биокаталитической трансформации принимают такое количество этого препарата, которое катализирует превращение 1 мкмоля субстрата (или образование 1 мкмоля продукта) в выбранных условиях за 1 минуту), отнесенных к миллиграмму белка, и во всех случаях приведены к активности по синтезу антибиотика цефалексина с использованием экспериментальных соотношений между гидролазными и синтетазной активностью.
Известны способы получения АЭГ из различных бактериальных источников в водорастворимой форме, направленные на получение высокоочищенных ферментных препаратов, вплоть до получения гомогенных белков [Enzyme Microb. Technol. 9: 339 (1987); BBA - Protein Struct.& Molecul. Enzymol. 1040 (1): 12 (1990); Биохимия 55 (12): 2226 (1990); Enzyme Microb. Technol. 15: 965 (1993)]. Эти способы обязательно включают в себя несколько последовательных стадий хроматографической очистки фермента на различных сорбентах и непригодны для использования при получении технологических биокатализаторов из-за трудоемкости и продолжительности процедуры, а также низких выходов активности.
Известны способы получения гетерогенных биокатализаторов на основе АЭГ из Acetobacter turbidans путем выделения фермента из биомассы клеток, его частичной очистки и последующей иммобилизации. Способ выделения АЭГ, состоящий в механическом разрушении клеточной биомассы под давлением, экстракции фермента в водную фазу и частичной очистке АЭГ путем обработки водного экстракта стрептомицином с последующим диализом [Enzyme Microb.Technol. 25: 336 (1999)] не охарактеризован с точки зрения выхода активности процедуры. Получаемый грубоочищенный препарат АЭГ имеет очень низкую удельную активность, определяемую в отношении гидролиза бензилпенициллина, - 0,06 МЕ/мг белка. Способ ковалентной иммобилизации получаемого таким способом грубоочищенного фермента на модифицированной глутаровым альдегидом агарозе [Enzyme Microb.Technol. 25: 336 (1999)], а также усовершенствованный способ его иммобилизации, включающий дополнительную модификацию иммобилизованного на агарозе фермента альдегид декстраном [J. Mol. Catal. В: Enzym. 11: 633 (2001)], трудоемки и требуют специальной подготовки носителя. Указанные способы не охарактеризованы с точки зрения выхода активности процедуры иммобилизации и абсолютной активности биокатализатора в отношении синтеза какого-либо антибиотика (синтетазной активности). Известен также способ более глубокой очистки АЭГ из Acetobacter turbidans и его ковалентной иммобилизации на агарозе с последующей модификацией альдегид декстраном [Biomacromolecules 2(1): 95 (2001)]. Грубоочищенный водный экстракт с удельной активностью 0,1 МЕ/мг белка, определяемой в отношении гидролиза метилового эфира Д-фенилглицина (МЭФГ), подвергают двухстадийной хроматографической очистке на СМ-52 и фенил-сефарозе. Выход активности процедуры очистки составляет 62% по отношению к грубоочищенному экстракту, однако общий выход, по отношению к биомассе клеток не известен. АЭГ очищают в 420 раз по сравнению с грубоочищенным водным экстрактом, достигая удельной активности 42 МЕ/мг белка. Процедура иммобилизации очищенной таким образом АЭГ не охарактеризована с точки зрения выхода активности и абсолютной синтетазной активности продукта. Данный способ получения биокатализатора очень трудоемок и малоэффективен из-за применения процедуры глубокой очистки фермента перед иммобилизацией и сложной многостадийной процедуры иммобилизации.
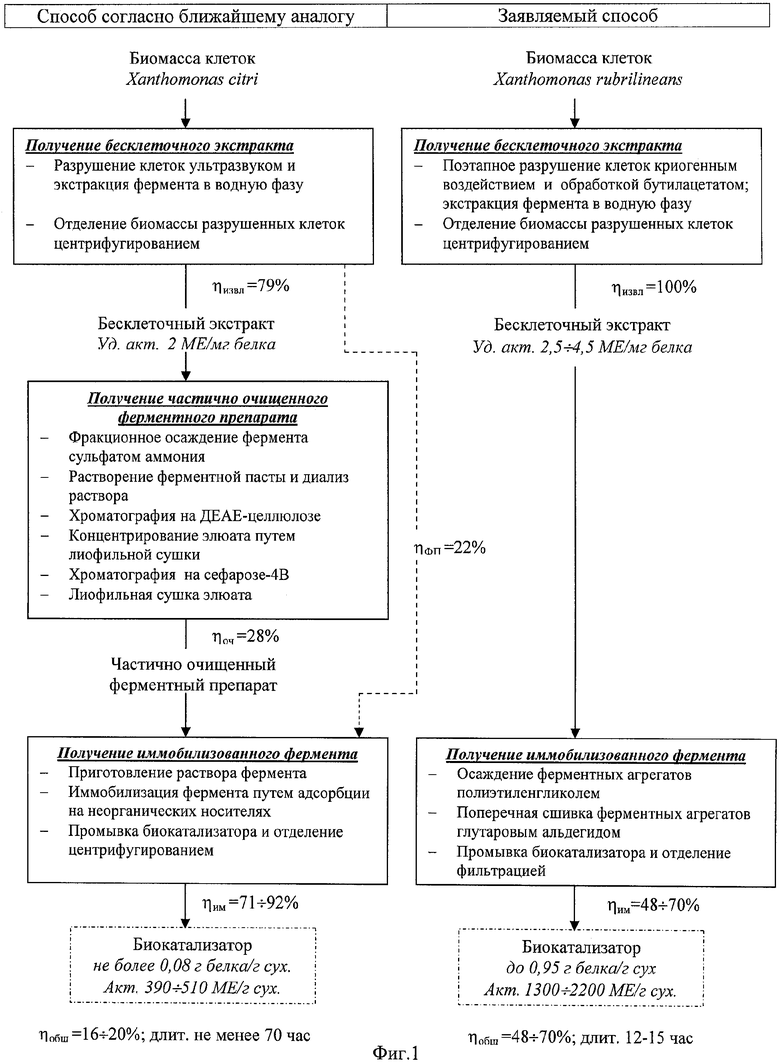
Наиболее близким к заявляемому способу получения гетерогенного биокатализатора на основе внутриклеточной гидролазы эфиров альфа-аминокислот из бактерий рода Xanthomonas является способ получения биокатализатора на основе фермента из микроорганизма того же рода, а именно из Xanthomonas citri [Biotechnol. Bioeng. 23 (2): 361 (1981)]. Как и заявляемый способ, ближайший аналог отражает полную процедуру получения гетерогенного биокатализатора, исходя из биомассы клеток (фиг.1).
В качестве ближайшего аналога для заявляемого гетерогенного биокатализатора на основе внутриклеточной гидролазы эфиров альфа-аминокислот из бактерий рода Xanthomonas выбран гетерогенный биокатализатор на основе АЭГ из Xanthomonas citri, иммобилизованной на каолине, получаемый по способу [Biotechnol. Bioeng. 23 (2): 361 (1981)], содержащий не более 0,08 г белка/г сухого биокатализатора и характеризующийся синтезной активностью 510 МЕ/г сухого биокатализатора.
В качестве ближайшего аналога для заявляемого способа ферментативного синтеза аминобета-лактамного антибиотика путем ацилирования ключевого бета-лактамного соединения соответствующим производным эфиров Д-аминофенилуксусной кислоты выбран способ синтеза аминоцефалоспоринового антибиотика цефалексина путем ацилирования 7-аминодезацетоксицефалоспорановой кислоты (7-АДЦК) метиловым эфиром Д-аминофенилуксусной кислоты под действием гетерогенного биокатализатора на основе АЭГ из Xanthomonas citri [Biotechnol. Bioeng. 23 (2): 361 (1981)].
Способ получения гетерогенного биокатализатора согласно ближайшему аналогу состоит в следующем. Биомассу клеток подвергают ультразвуковой дезинтеграции в присутствии поверхностно-активных веществ и экстрагируют фермент в водную фазу. Получают бесклеточный экстракт с удельной активностью 2 МЕ/мг белка, определяемой по синтезу цефалексина. Выход активности на стадии получения бесклеточного экстракта (ηизвл) достигает 79%. АЭГ концентрируют из бесклеточного экстракта фракционным осаждением сульфатом аммония. После растворения ферментного осадка препарат обессоливают путем диализа и подвергают двухстадийной хроматографической очистке сначала на ДЕАЕ-целлюлозе, а затем на сефарозе 4-В. После каждой из хроматографических стадий ферментный препарат лиофильно сушат. Выход синтетазной активности на стадиях концентрирования и очистки АЭГ (ηоч) составляет 28%, при этом степень очистки препарата достигает 30 раз. Выход процедуры получения ферментного препарата, пригодного для иммобилизации (ηФП), определяется выходами стадий извлечения и очистки фермента и составляет 22%: (ηФП=ηизвл·ηоч).
Полученный частично очищенный препарат АЭГ с удельной активностью в отношении синтеза цефалексина 60 МЕ/мг белка иммобилизуют путем адсорбции на бентоните или каолине. Выход активности процедуры иммобилизации (ηим) составляет 71÷92% в зависимости от используемого носителя. Общий выход процесса получения гетерогенного биокатализатора от биомассы клеток (ηобщ) согласно ближайшему аналогу - 16-20%, продолжительность процедуры - не менее 70 часов.
Лучшим из биокатализаторов, полученных по способу согласно ближайшему аналогу, является АЭГ, иммобилизованная на каолине. Этот гетерогенный биокатализатор содержит не более 0,08 г белка/г сухого биокатализатора и обладает синтетазной активностью (в отношении синтеза цефалексина), отнесенной к массе сухого носителя, 510 МЕ/г сух. Операционные испытания его в процессе синтеза цефалексина при 30°С с использованием низкой (4 мг/мл) начальной концентрации субстрата 7-АДЦК показывают, что через 10 циклов остаточная активность биокатализатора составляет 85% от исходной. Период полуинактивации биокатализатора, рассчитанный по этим данным, составляет τ1/2=42 цикла. Однако обработка представленных в ближайшем аналоге экспериментальных данных по зависимости операционной стабильности биокатализатора от количества циклов показывает, что инактивация биокатализатора не подчиняется кинетическим закономерностям первого порядка и в первых пяти циклах протекает в 4÷5 раз быстрее, чем в пяти последующих. Это может свидетельствовать о частичном смывании фермента с носителя в процессе синтеза даже при использованных очень низких концентрациях субстратов. Повышение температуры до 35°С снижает операционную стабильность иммобилизованной на каолине АЭГ в 8 раз (τ1/2=5 циклов).
Способ синтеза аминоцефалоспоринового антибиотика цефалексина согласно ближайшему аналогу состоит в ацилировании 7-АДЦК, взятой в концентрации 4 мг/мл, метиловым эфиром Д-фенилглицина при молярном соотношении ацилирующего агента (МЭФГ) и 7-АДЦК в исходной реакционной смеси от 1 до 3. Реакцию синтеза проводят в течение 2÷3 часов при рН 6,2 и температуре 35°С в водной среде под действием гетерогенного биокатализатора на основе АЭГ из Xanthomonas citri, иммобилизованной на каолине. Лучший выход цефалексина (85% по 7-АДЦК) достигнут при использовании молярного соотношения субстратов 3:1. Данные по синтезу других аминоцефалоспоринов или аминопенициллинов в ближайшем аналоге отсутствуют.
Основные недостатки гетерогенного биокатализатора, а также способов его получения и использования согласно ближайшему аналогу следующие:
1. Сложность и многостадийность используемой процедуры выделения и очистки фермента; связанные с этим большие потери ферментативной активности, обуславливающие низкий выход активности процедуры получения биокатализатора в целом.
2. Получение биокатализатора с низким содержанием белка (не более 0,08 г белка/г сухого биокатализатора) из-за использования метода иммобилизации фермента на носителе, являющемся балластом.
3. Низкий уровень активности конечного продукта - гетерогенного биокатализатора.
4. Низкий уровень операционной стабильности биокатализатора, обусловленный использованием метода адсорбционной иммобилизации, не обеспечивающего многоточечного, прочного и необратимого связывания фермента с носителем; следствием этого является низкая термическая стабильность иммобилизованного фермента, а также возможная десорбция фермента и других белков раствором субстратов (особенно при высоких концентрациях субстратов).
5. Необходимость использования низких концентраций субстратов при синтезе антибиотика для уменьшения опасности загрязнения целевого продукта примесями белковой природы.
Раскрытие изобретения
Заявляемая группа изобретений направлена на решение следующих задач:
- повышение эффективности способа получения гетерогенного биокатализатора на основе гидролазы эфиров альфа-аминокислот за счет упрощения процедуры получения биокатализатора и увеличения выхода ферментативной (синтетазной) активности этой процедуры;
- повышение активности и стабильности получаемого биокатализатора;
- разработка способа ферментативного синтеза аминобета-лактамного антибиотика с использованием получаемого гетерогенного биокатализатора.
Поставленные задачи решают путем совокупного использования в процессе получения гетерогенного биокатализатора ряда приемов, осуществляемых в выбранных оптимизированных условиях. Для выделения АЭГ используют биомассу клеток бактерий Xanthomonas rubrilineans. Клетки, содержащие фермент, разрушают поэтапно в мягких условиях, сначала нарушая их целостность криогенным воздействием, а затем обрабатывая водную суспензию размороженных клеток бутилацетатом. Из разрушенной клеточной биомассы извлекают фермент путем его экстракции в водную фазу. Получаемый после отделения твердой фазы бесклеточный экстракт без какой-либо очистки используют для иммобилизации фермента. Для получения такого бесклеточного экстракта, который непосредственно пригоден для иммобилизации, процедуру разрушения биомассы клеток и извлечения фермента осуществляют в мягких условиях, оптимизированных по концентрации биомассы клеток в суспензии, концентрации бутилацетата, температуре, рН и моменту остановки процесса. Экстракцию ведут сначала в условиях спонтанно устанавливающегося градиента рН, а затем при постоянном значении рН, поддерживаемом путем добавления титранта, расход которого регистрируют.
Критерием выбора момента остановки экстракции служит изменение скорости расхода титранта. Получение бесклеточного экстракта требуемой чистоты при достижении (100±5)% выхода активности в процессе извлечения фермента обеспечивается остановкой процедуры в момент, когда скорость расхода титранта снижается в определенное число раз по сравнению с первоначальным значением. Степень снижения расхода титранта, обеспечивающая оптимальные результаты (Копт), зависит от используемого штамма Xanthomonas rubrilineans, а также от условий биосинтеза культуры и устанавливается в серии предварительных экспериментов (см. таблицу 2 в Примере 3).
Иммобилизацию осуществляют путем осаждения АЭГ в виде ферментных агрегатов из бесклеточного экстракта под действием полиэтиленгликоля с выбранной молекулярной массой, взятого в количестве, необходимом для полного осаждения целевого фермента, и последующей поперечной сшивки этих агрегатов глутаровым альдегидом, взятым в оптимизированной концентрации, рассчитываемой по отношению к содержанию белка в бесклеточном экстракте (см. Пример 4, №1-4 в таблице 3). Получение бесклеточного экстракта в оптимизированных для каждого штамма условиях обеспечивает стандартизованный исходный материал для процедуры иммобилизации, что видно при сравнении №2, 3 и 5 таблицы 3 в Примере 4.
Заявляемый способ получения гетерогенного биокатализатора схематически представлен на фиг.1 и заключается в следующем. Биомассу клеток бактерий Xanthomonas rubrilineans, содержащих фермент АЭГ, помещают в морозильную камеру с температурой -25°С или ниже, предпочтительно от -25 до -30°С, и выдерживают в течение времени, необходимого для промораживания всего объема биомассы до заданной температуры. Затем клеточную биомассу размораживают, выдерживая необходимое время при комнатной температуре, и суспендируют в буфере Трис-HCl, 0,05 моль/л, рН 7,8. Концентрация биомассы клеток в суспензии составляет 20÷100 мг сух/мл, предпочтительно 40÷50 мг сух/мл. К полученной суспензии добавляют бутилацетат в концентрации 2÷6 об.%, предпочтительно 4,5 об.%. Процедуру разрушения клеточной биомассы и экстракции фермента в водную фазу осуществляют при интенсивном перемешивании при температуре 25÷35°С, предпочтительно 30°С. Экстракцию ведут сначала в условиях спонтанно устанавливающегося градиента рН, а затем при постоянном значении рН, поддерживаемом путем добавления щелочного титранта, и регистрируют его расход. В качестве титранта используют водный раствор аммиака либо другие титранты, такие как водный раствор гидроокиси натрия или гидроокиси калия.
Диапазон рН, обеспечивающий оптимальные результаты, зависит от используемого штамма Xanthomonas rubrilineans, а также от условий биосинтеза культуры и устанавливается в серии предварительных экспериментов. Процедуру экстракции фермента прекращают, когда скорость расхода раствора аммиака сократится в установленное экспериментально число раз Копт. Время достижения Копт, то есть продолжительность процедуры разрушения клеточной биомассы и экстракции фермента не нормируется и составляет от 3 до 7 часов, в зависимости от биохимических параметров исходной биомассы клеток, а также от выбранных условий проведения процесса по температуре и концентрациям биомассы клеток и бутилацетата.
Разрушенную клеточную биомассу отделяют от бесклеточного экстракта любым доступным эффективным способом, например центрифугированием, и промывают натрий-ацетатным буфером, 0,1 моль/л, рН 6,5, содержащим 0,005 моль/л ацетата кальция. Буфер используют в количестве 20% от исходного объема клеточной суспензии. Промывные воды отделяют от разрушенной клеточной биомассы и присоединяют к бесклеточному экстракту. Получают прозрачный бесклеточный экстракт с удельной активностью 2,5÷4,5 МЕ/мг белка, определяемой по синтезу цефалексина. Выход активности на стадии извлечения фермента по отношению к активности исходной биомассы клеток составляет (100±5)%, то есть является количественным в пределах ошибки эксперимента.
Полученный бесклеточный экстракт используют без какой-либо очистки для процедуры иммобилизации фермента АЭГ путем получения ферментных агрегатов и их поперечной сшивки. Таким образом, выход процедуры получения ферментного препарата, пригодного для иммобилизации, тождественен выходу процедуры извлечения фермента:
ηФП=ηизвл=(100±5) %.
С целью осаждения ферментных агрегатов к бесклеточному экстракту добавляют полиэтиленгликоль с молекулярной массой 4000÷10000, предпочтительно 6000÷8000, взятый в количестве, необходимом для полного осаждения целевого фермента, а именно 0,25÷0,30 г/мл бесклеточного экстракта, в зависимости от молекулярной массы полиэтиленгликоля. Процедуру осуществляют при температуре 2÷4°С при перемешивании в течение времени, необходимого для полного растворения полиэтиленгликоля (15÷30 мин). Затем для поперечной сшивки ферментных агрегатов в реакционную смесь вносят глутаровый альдегид в количестве, необходимом для создания его относительной концентрации 1·10-2÷3·10-2 моль/мг белка, и проводят модификацию и сшивку белков в течение 2 часов при слабом перемешивании при температуре 2÷4°С. Гетерогенный биокатализатор в виде поперечно-сшитых ферментных агрегатов отделяют от реакционной смеси вакуумной фильтрацией и отмывают на пористом стеклянном фильтре до бесцветных промывных вод натрий-ацетатным буфером, 0,1 моль/л, рН 6,5, содержащим 0,005 моль/л ацетата кальция.
Выход активности процедуры иммобилизации АЭГ составляет 48÷70%. Так как выделение фермента осуществляют без потерь активности, то общий выход процесса получения гетерогенного биокатализатора (от биомассы клеток) по заявляемому способу - 48÷70%, продолжительность процедуры - 12÷15 часов.
Характеристика заявляемого биокатализатора. Гетерогенный биокатализатор, иммобилизованная АЭГ, не содержит балластного носителя и состоит преимущественно из белка (до 0,95 г белка/г сух). Иммобилизованная АЭГ обладает активностью (в отношении синтеза цефалексина), отнесенной к массе сухого биокатализатора, 1300÷2200 МЕ/г сух. Диапазон относительной концентрации глутарового альдегида в реакционной смеси при проведении поперечной сшивки ферментных агрегатов 1·10-2÷5·10-2 моль/мг белка обеспечивает получение иммобилизованного фермента, сочетающего указанный уровень синтетазной активности с операционной стабильностью, характеризующейся сохранением 95% исходной активности после использования биокатализатора в 10 циклах синтеза цефалексина (см. Пример 4, табл.3).
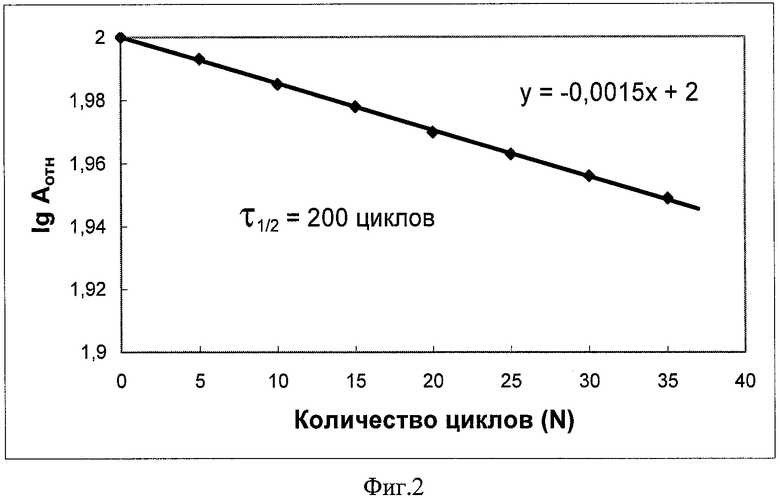
Более продолжительные операционные испытания биокатализатора в процессе синтеза цефалексина при 30°С и начальной концентрации субстрата 7-АДЦК 20 мг/мл показывают, что инактивация иммобилизованного фермента подчиняется кинетическим закономерностям первого порядка. Это свидетельствует об отсутствии вымывания АЭГ из биокатализатора даже при использованнии относительно высоких концентраций субстратов (в 5 раз выше, чем описано в ближайший аналоге). Период полуинактивации биокатализатора, определенный по 35 циклам использования в процессе синтеза цефалексина, составляет τ1/2=200 циклов (см. Пример 10 и фиг.2).
Заявляемый способ синтеза аминобета-лактамного антибиотика в общем виде заключается в следующем. С использованием полученного биокатализатора проводят ферментативный синтез различных аминопенициллинов, например ампициллина, амоксициллина, и аминоцефалоспоринов например, цефалексина цефадроксила, цефрадина, цефаклора, цефалоглицина, путем ацилирования ключевого бета-лактамного соединения, взятого в концентрации 5÷40 мг/мл, соответствующими производными эфиров Д-аминофенилуксусной кислоты, например, метиловыми эфирами Д-фенилглицина, Д-пара-гидроксифенилглицина, Д-2,5-дигидрофенилглицина, взятыми в молярном избытке 1,5÷4,0. Реакцию синтеза проводят при рН 6,0÷6,2 и температуре 1÷40°С в водной среде или в смесях воды с многоатомным спиртом этиленгликолем при содержании последнего в исходной смеси 30÷50%. Содержание гетерогенного биокатализатора в реакционной смеси варьируют от 5 до 50 МЕ/мл в зависимости от выбранных условий проведения реакции синтеза так, чтобы длительность процесса не превышала трех часов.
Преимущества заявляемой группы изобретений:
1. Получение гетерогенного биокатализатора по простейшей возможной схеме (см. фиг.1), включающей лишь этапы выделения фермента из биомассы клеток и его иммобилизации (непосредственно из бесклеточного экстракта), в отличие от способа согласно ближайшему аналогу, включающего еще и многостадийный этап получения очищенного ферментного препарата; соответствующее сокращение общего времени процедуры до 12÷15 часов, вместо, по крайней мере, 70 часов по прототипу.
2. Повышение общего выхода активности процедуры получения биокатализатора более чем в 3 раза (48÷70% по заявляемому способу; 16÷20% согласно ближайшему аналогу).
3. Получение биокатализатора, не содержащего балластного носителя и состоящего преимущественно из белка (в заявляемом биокатализаторе до 0,95 г белка/г сух; в ближайшем аналоге - не более чем 0,08 г белка/г сух.).
4. Повышение синтетазной активности биокатализатора (в отношении синтеза цефалексина) в 3,3÷4,3 раза (в заявляемом биокатализаторе 1300÷2200 МЕ/г сух.; в ближайшем аналоге - 510 МЕ/г сух.).
5. Повышение операционной стабильности биокатализатора, по крайней мере, в 5 раз (при 30°С: τ1/2=200 циклов для заявляемого биокатализатора; не более чем τ1/2=42 цикла согласно ближайшему аналогу).
6. Отсутствие вымывания фермента из биокатализатора в процессе его эксплуатации даже при начальных концентрациях субстрата, в 5 раз превышающих концентрации, использованные при операционных испытаниях.
7. Возможность использования биокатализатора для ферментативного синтеза аминобета-лактамного антибиотика в широком диапазоне температур и концентраций исходных соединений в водной среде или в смесях воды с многоатомным спиртом этиленгликолем, что позволяет достигать более высоких выходов целевого антибиотика (до 98%, см. Пример 6) по сравнению с 85% согласно ближайшему аналогу.
Заявляемая группа изобретений проиллюстрирована следующими фигурами графического изображения:
Фиг.1 Схема способов получения биокатализатора на основе АЭГ.
Фиг.2 Операционная стабильность биокатализатора в процессе синтеза цефалексина (по данным таблицы 4).
Осуществление изобретения
Изобретение иллюстрируется следующими примерами осуществления способа получения биокатализатора и способа синтеза аминобета-лактамного антибиотика под действием полученного гетерогенного биокатализатора. Во всех описанных примерах приведена активность ферментных препаратов по синтезу цефалексина. Ее определяют по начальной скорости образования целевого продукта в следующих условиях: рН 6,0÷6,2, температура (40±1)°С, концентрации субстратов - (0,040±0,002) моль/л 7-АДЦК и (0,080±0,004) моль/л МЭФГ. Содержание цефалексина в реакционной смеси определяют методом высокоэффективной жидкостной хроматографии (ВЭЖХ). Содержание белка в препаратах определяют традиционными методами [Biotechnol. Appl. Biochem. 29: 99 (1999)], а содержание сухих веществ - весовым методом после высушивания препарата при (105±1)°С до постоянной массы.
Пример 1. Осуществление заявляемого способа получения биокатализатора с использованием штамма Xanthomonas rubrilineans VKPM B-9915.
В качестве исходного материала для получения гетерогенного биокатализатора использована биомасса клеток штамма Xanthomonas rubrilineans VKPM B-9915 из Всероссийской коллекции промышленных микроорганизмов (VKPM). Культивирование продуцента проведено в течение 22 часов в колбах Эрленмейера на среде, состоящей из кукурузного экстракта (2% по сух.), пептона (1%) и глюкозы (1%) при 28°С на круговой качалке при 240-260 об/мин. Биомасса клеток отделена центрифугированием при 7000 об/мин.
Взято 273 г влажной биомассы клеток с содержанием сухих веществ 18,5%, синтетазной активностью 83,3 МЕ/г влажн, 450 МЕ/г сух.
Для осуществления криогенного воздействия биомассу клеток помещают в пластиковый сосуд и замораживают в морозильной камере при температуре - 28°С. Через 3 часа сосуд извлекают из морозильной камеры, биомассу размораживают при комнатной температуре и переносят в термостатируемый реактор объемом 2,5 л, снабженный механической мешалкой и системой контроля и поддержания рН.
Процедуру обработки клеток бутилацетатом и извлечения фермента в водную фазу осуществляют в этом реакторе в следующих условиях: концентрация биомассы клеток в суспензии 40 мг сух/мл; концентрация бутилацетата 4,5 об.%; температура 30°С; градиент рН, постоянное перемешивание. Размороженную биомассу клеток суспендируют в буфере Трис-HCl, 0,05 моль/л, рН 7,8, взятом в количестве, необходимом для получения суспензии клеток требуемой концентрации, с учетом влаги, содержащейся в биомассе клеток. При проведении данной процедуры для получения 1260 мл суспензии используют 1040 мл буфера. Размороженную биомассу суспендируют в буфере при температуре не выше 20°С, добавляют 150 мкл меркаптоэтанола для стабилизации фермента и вносят 57 мл бутилацетата. Момент добавления бутилацетата считают началом процедуры. В начальный момент времени включают обогрев реактора для поддержания температуры 30°С и контролируют исходное значение рН, величина которого не должна превышать 8,0. По ходу процесса рН снижается и процедура протекает в спонтанно устанавливающемся градиенте его значений, вплоть до рН 6,8. При достижении значения рН 6,8 включают систему автоматической подтитровки и поддерживают рН 6,8±0,1 путем добавления в реакционную смесь водного раствора аммиака с концентрацией 12,5%, фиксируя расход этого раствора по ходу процесса. Процедуру извлечения фермента прекращают через 4 часа 30 минут, когда скорость расхода раствора аммиака сократится в 2,5 раза.
Реакционную смесь быстро охлаждают до температуры 15÷20°С и обрабатывают флокулянтом для облегчения отделения разрушенной биомассы клеток. В качестве флокулянта используют водорастворимый полимерный анионит, содержащий пиридиниевые или четвертичные аммониевые основания, например Флокатон ППС-400. Разрушенную биомассу клеток отделяют от бесклеточного экстракта центрифугированием и промывают 250 мл натрий-ацетатного буфера, 0,1 моль/л, рН 6,5, содержащего 0,005 моль/л ацетата кальция. Промывные воды, отделенные центрифугированием, присоединяют к бесклеточному экстракту. В полученном растворе контролируют рН и устанавливают значение рН 6,8÷7,2.
Получают 1440 мл прозрачного бесклеточного экстракта с синтетазной активностью 16,0 МЕ/мл, содержанием белка 4,2 мг/мл, удельной активностью 3,8 МЕ/мг белка. Выход активности процесса извлечения фермента из биомассы клеток составляет ηизвл=101%.
Процедуру иммобилизации фермента осуществляют в реакторе объемом 2 л, снабженном механической мешалкой и помещенном в ледяную баню. 1440 мл бесклеточного экстракта помещают в этот реактор, охлаждают до 2÷4°С и при постоянном перемешивании добавляют к нему полиэтиленгликоль с молекулярной массой 6000, взятый в количестве 403 г (0,28 г/мл бесклеточного экстракта), что обеспечивает полное осаждение целевого фермента. Перемешивают реакционную смесь в течение 15 мин (до полного растворения осадителя) и добавляют 76,7 мл 25% водного раствора глутарового альдегида (3,5·10-2 ммоль/мг белка). Реакционную смесь инкубируют при слабом перемешивании при температуре 2÷4°С в течение 2 часов. Полученные твердые частицы отделяют вакуумной фильтрацией и отмывают на пористом стеклянном фильтре до бесцветных промывных вод натрий-ацетатным буфером, 0,1 моль/л, рН 6,5, содержащим 0,005 моль/л ацетата кальция. На промывку расходуют 1500 мл этого буфера.
Получают 38,6 г влажного гетерогенного биокатализатора в виде поперечно-сшитых ферментных агрегатов АЭГ с содержанием сухих веществ 20,4%. Синтетазная активность продукта составляет 370 МЕ/г влажного или 1810 МЕ/г сухого биокатализатора. Содержание белка в продукте составляет 0,93 г белка/г сух. Выход активности процедуры иммобилизации АЭГ составляет ηим=59,1%, общий выход активности процесса получения гетерогенного биокатализатора (от биомассы клеток) ηoбщ=59,8%. Период полуинактивации биокатализатора, определенный по 35 циклам его использования в процессе синтеза цефалексина, составляет τ1/2=200 циклов (см. Пример 10).
Пример 2. Осуществление заявляемого способа получения биокатализатора с использованием штамма Xanthomonas rubrilineans CGMCC №2339
В качестве исходного материала для получения гетерогенного биокатализатора использована биомасса клеток штамма Xanthomonas rubrilineans CGMCC №. 2339 из Главной китайской коллекции культур микроорганизмов (CGMCC). Биомасса получена в условиях, приведенных в примере 1.
Взято 91,2 г влажной биомассы клеток с содержанием сухих веществ 19,5%, синтетазной активностью 70,2 МЕ/г влажн, 360 МЕ/г сух.
Криогенное воздействие на биомассу клеток осуществляют по методике, описанной в примере 1, но при температуре -30°С и в течение 5 часов.
Процедуру обработки клеток бутилацетатом и извлечения фермента в водную фазу осуществляют согласно примеру 1 в следующих условиях: концентрация биомассы клеток в суспензии 50 мг сух/мл; концентрация бутилацетата 4,0 об.%; температура 28°С; градиент рН, постоянное перемешивание. Используют следующие количества реагентов: 280 мл буфера, 50 мкл меркаптоэтанола, 14 мл бутилацетата. Объем суспензии составляет 355 мл. Процедуру извлечения фермента осуществляют в течение 6 часов 10 минут и останавливают в момент достижения снижения скорости расхода титранта в 2,8 раза по сравнению с ее исходным значением.
Обработку реакционной смеси флоккулянтом и получение бесклеточного экстракта осуществляют согласно примеру 1. Получают 420 мл прозрачного бесклеточного экстракта с синтетазной активностью 15,7 МЕ/мл, содержанием белка 3,5 мг/мл, удельной активностью 4,5 МЕ/мг белка. Выход активности процесса извлечения фермента из биомассы клеток составляет ηизвл=103 %.
Процедуру иммобилизации фермента осуществляют по методике, описанной в примере 1, с той разницей, что используют 420 мл бесклеточного экстракта, 105 г полиэтиленгликоля с молекулярной массой 8000 (0,25 г/мл бесклеточного экстракта) и 8,8 мл 25% водного раствора глутарового альдегида (1,5·10-2 ммоль/мг белка).
Получают 11,1 г влажного гетерогенного биокатализатора в виде поперечно-сшитых ферментных агрегатов АЭГ с содержанием сухих веществ 20,2%. Синтетазная активность продукта составляет 380 МЕ/г влажного или 1880 МЕ/г сухого биокатализатора. Содержание белка в продукте составляет 0,92 г белка/г сух. Выход активности процедуры иммобилизации АЭГ составляет ηим=64,0%, общий выход активности процесса получения гетерогенного биокатализатора (от биомассы клеток) ηобщ=65,9%. Относительная остаточная активность биокатализатора после 10 циклов синтеза цефалексина (см. Пример 10) составляет 96,3%.
Пример 3. Осуществление заявляемого способа получения биокатализатора при варьировании момента остановки процесса извлечения фермента
В таблице 2 (№1-3) приведены данные, полученные с использованием культуры штамма Xanthomonas rubrilineans VKPM В-9915. Процедуры криогенного воздействия на биомассу клеток, ее обработки бутилацетатом и извлечения фермента в водную фазу осуществляют согласно примеру 1, с той разницей, что варьируют момент остановки процесса извлечения фермента, соотнося его со скоростью расхода раствора аммиака. Бесклеточные экстракты используют для иммобилизации фермента согласно процедуре, описанной в примере 1.
Представленные результаты отражают влияние момента остановки процесса на свойства бесклеточных экстрактов и биокатализаторов, а также на эффективность процессов извлечения и иммобилизации фермента. В таблице 2 (№4) приведены также данные примера 2 по получению биокатализатора, исходя из биомассы клеток штамма Xanthomonas rubrilineans CGMCC №2339, демонстрирующие зависимость оптимального момента остановки процесса экстракции фермента от используемого штамма.
Влияние момента остановки процесса извлечения фермента на свойства бесклеточного экстракта и гетерогенного биокатализатора, а также на эффективность их получения
ηим, %
B-9915
Пример 1
B-9915
B-9915
Пример 2
Пример 4. Осуществление заявляемого способа получения биокатализатора при различных концентрациях глутарового альдегида
В таблице 3 (№1-4) приведены данные, полученные с использованием культуры штамма Xanthomonas rubrilineans VKPM B-9915. Процедуры криогенного воздействия на биомассу клеток, ее обработки бутилацетатом и извлечения фермента в водную фазу осуществляют согласно примеру 1, то есть в оптимизированных для данного штамма условиях. Полученные бесклеточные экстракты используют для иммобилизации фермента согласно процедуре, описанной в примере 1, с той разницей, что варьируют относительную концентрацию глутарового альдегида в реакционной смеси при поперечной сшивке ферментных агрегатов.
В качестве критерия для сопоставления стабильности иммобилизованных ферментов, полученных в различных условиях, используют остаточную ферментативную активность биокатализатора после его эксплуатации в 10 циклах синтеза цефалексина, осуществляемого согласно примеру 10. Представленные результаты отражают влияние концентрации глутарового альдегида на свойства биокатализаторов и эффективность процесса иммобилизации.
Влияние относительной концентрации глутарового альдегида в реакционной смеси на свойства гетерогенного биокатализатора и эффективность процесса иммобилизации фермента
Пример 1
Пример 2
№2339
В таблице 3 (№5) приведены также характеристики биокатализатора, полученного исходя из биомассы клеток штамма Xanthomonas rubrilineans CGMCC №2339. Согласно примеру 2 бесклеточный экстракт получают в условиях, оптимизированных для данного штамма, затем осаждают ферментные агрегаты и осуществляют поперечную сшивку в условиях, попадающих в оптимальный для штамма VKPM В-9915 диапазон концентраций глутарового альдегида, достигая при этом аналогичных характеристик конечного биокатализатора.
Пример 5. Синтез цефалексина в водной среде
Процесс ферментативного синтеза цефалексина проводят в термостатируемом стеклянном реакторе вместимостью 100 мл, снабженном механической мешалкой и устройством для измерения и поддержания рН, в следующих условиях: водная среда; температура 2°С; рН 6,0÷6,2; содержание фермента (25±1)МЕ/мл; начальная концентрация 7-АДЦК 25 мг/мл; молярное соотношение МЭФГ и 7-АДЦК в исходной реакционной смеси 3,0:1. Расчет концентраций реагентов в исходном растворе проводят с учетом объема влаги, вносимой с навеской биокатализатора.
Для приготовления раствора субстратов 1,25 г (5,84 ммолей) 7-АДЦК помещают в химический стакан вместимостью 100 мл, добавляют 40 мл дистиллированной воды. При перемешивании на магнитной мешалке и контроле рН к суспензии порциями добавляют 12,5% раствор аммиака до растворения навески 7-АДЦК. При этом рН не должно превышать значения 8,2. Затем к раствору при перемешивании добавляют порциями 3,53 г (17,52 ммолей) гидрохлорида МЭФГ, не допуская снижения рН раствора ниже 6,0 путем добавления 12,5% раствора аммиака. После растворения МЭФГ раствор количественно переносят в мерный цилиндр и объем раствора доводят водой до 50 мл.
Приготовленный раствор субстратов переносят в реактор, устанавливают температуру 2°С и при интенсивном перемешивании добавляют 3,4 г биокатализатора, полученного по примеру 1, с активностью 370 МЕ/г влажн биокатализатора и содержанием сухих веществ 20,4%. Момент добавления биокатализатора считается началом реакции синтеза. Процесс проводят в течение 70 минут при температуре 2°С и значении рН 6,0÷6,2, которое поддерживают путем добавления 12,5% раствора аммиака в автоматическом режиме. По окончании процесса биокатализатор отделяют от реакционной смеси вакуумной фильтрацией, промывают 10 мл воды и в фильтрате, объединенном с промывкой, определяют содержание цефалексина методом ВЭЖХ. Выход цефалексина по 7-АДЦК составляет 85,7%.
Пример 6. Синтез цефалексина в смеси воды с этиленгликолем
Процесс ферментативного синтеза цефалексина проводят в реакторе, описанном в примере 5, в следующих условиях: содержание этиленгликоля в исходной реакционной смеси 50 об.%; температура 5°С; рН 6,0÷6,2; содержание фермента (40±1) МЕ/мл; начальная концентрация 7-АДЦК 30 мг/мл; молярное соотношение МЭФГ и 7-АДЦК в исходной реакционной смеси 2,5:1.
Приготовление раствора субстратов из 1,50 г (7,01 ммолей) 7-АДЦК и 3,53 г (17,52 ммолей) гидрохлорида МЭФГ осуществляют по методике, описанной в примере 5, с той разницей, что вместо 40 мл воды для растворения навески 7-АДЦК используют смесь 15 мл дистиллированной воды и 25 мл этиленгликоля.
Синтез цефалексина осуществляют согласно примеру 5 с той разницей, что процесс ведут в течение 180 минут при температуре 5°С, используя 5,41 г биокатализатора, полученного по примеру 1, с активностью 370 МЕ/г влажн биокатализатора и содержанием сухих веществ 20,4%. Выход цефалексина по 7-АДЦК составляет 98,3%.
Пример 7. Синтез цефаклора
Процесс ферментативного синтеза цефаклора проводят в реакторе, описанном в примере 5, в следующих условиях: содержание этиленгликоля в исходной реакционной смеси 45 об.%; температура 10°С; рН 6,0-6,2; содержание фермента (40±1)МЕ/мл; начальная концентрация 3-хлор-7-аминоцефалоспорановой кислоты (7-АХЦК) 15 мг/мл; молярное соотношение МЭФГ и 7-АХЦК в исходной реакционной смеси 4:1.
Приготовление раствора субстратов из 0,75 г (3,20 ммолей) 7-АХЦК и 2,58 г (12,80 ммолей) гидрохлорида МЭФГ осуществляют по методике, описанной в примере 5, с той разницей, что вместо 40 мл воды для растворения навески 7-АХЦК используют 17,5 мл дистиллированной воды и 22,5 мл этиленгликоля.
Синтез цефаклора осуществляют согласно примеру 5 с той разницей, что процесс ведут в течение 100 минут при температуре 10°С, используя 5,41 г биокатализатора, полученного по примеру 1, с активностью 370 МЕ/г влажн биокатализатора и содержанием сухих веществ 20,4%. Выход цефаклора по 7-АХЦК составляет 92,4%.
Пример 8. Синтез ампициллина
Процесс ферментативного синтеза ампициллина проводят в реакторе, описанном в примере 5, в следующих условиях: содержание этиленгликоля в исходной реакционной смеси 30 об.%; температура 20°С; рН 6,0÷6,2; содержание фермента (30±1) МЕ/мл; начальная концентрация 6-аминопенициллановой кислоты (6-АПК) 40 мг/мл; молярное соотношение МЭФГ и 6-АПК в исходной реакционной смеси 3:1.
Приготовление раствора субстратов из 2,00 г (9,26 ммолей) 6-АПК и 5,60 г (27,7 ммолей) гидрохлорида МЭФГ осуществляют по методике, описанной в примере 5, с той разницей, что вместо 40 мл воды для растворения навески 6-АПК используют 25 мл дистиллированной воды и 15 мл этиленгликоля, а рН раствора при растворении 6-АПК не превышает значения 7,5.
Синтез ампициллина осуществляют согласно примеру 5 с той разницей, что процесс ведут в течение 75 минут при температуре 20°С, используя 3,97 г биокатализатора, полученного по примеру 1, с активностью 370 МЕ/г влажн биокатализатора и содержанием сухих веществ 20,4%. Выход ампициллина по 6-АПК составляет 88,6%.
Пример 9. Синтез амоксициллина
Процесс ферментативного синтеза амоксициллина проводят в реакторе, описанном в примере 5, в следующих условиях: водная среда; температура 40°С; рН 6,0÷6,2; содержание фермента (6,5±0,5) МЕ/мл; начальная концентрация 6-АПК 10 мг/мл; молярное соотношение метилового эфира Д-пара-гидроксифенилглицина (МЭГФГ) и 6-АПК в исходной реакционной смеси 1,7:1.
Для приготовления раствора субстратов 0,720 г (3,97 ммолей) МЭГФГ помещают в химический стакан вместимостью 100 мл, добавляют 40 мл дистиллированной воды. При перемешивании на магнитной мешалке и контроле рН к суспензии порциями добавляют 20% раствор соляной кислоты до растворения навески МЭГФГ. Затем рН раствора доводят до 6 путем добавления 12,5% раствора аммиака и сразу же вносят в раствор 0,500 г (2,34 ммолей) 6-АПК. При перемешивании к суспензии добавляют порциями 12,5% раствор аммиака до растворения навески 6-АПК, следя, чтобы рН не превышало значения 6,5. После растворения 6-АПК раствор количественно переносят в мерный цилиндр и объем раствора доводят водой до 50 мл.
Синтез амоксициллина осуществляют согласно примеру 5 с той разницей, что процесс ведут в течение 60 минут при температуре 40°С, используя 0,86 г биокатализатора, полученного по примеру 2, с активностью 380 МЕ/г влажн биокатализатора и содержанием сухих веществ 20,2%. Выход амоксициллина по 6-АПК составляет 72,8%.
Пример 10. Операционная стабильность биокатализатора в процессе синтеза цефалексина
Для изучения операционной стабильности биокатализатора проводят ряд последовательных циклов синтеза цефалексина с использованием одной и той же порции биокатализатора в следующих условиях: содержание этиленгликоля в исходной реакционной смеси 40 об.%, температура 30°С, рН 6,0÷6,2, содержание фермента (15±1) МЕ/мл, начальная концентрация 7-АДЦК 20 мг/мл; молярное соотношение МЭФГ и 7-АДЦК в исходной реакционной смеси 2,5:1.
Первый цикл синтеза цефалексина проводят по методике, описанной в примере 5, но при температуре процесса 30°С и в объеме реакционной смеси 100 мл. По окончании реакции биокатализатор отделяют от реакционной смеси вакуумной фильтрацией, промывают его 10 мл воды и используют в следующем цикле синтеза цефалексина. В объединенном фильтрате определяют содержание цефалексина методом ВЭЖХ и рассчитывают выход цефалексина по 7-АДЦК.
Последующие циклы синтеза проводят аналогичным образом. После каждых пяти циклов определяют остаточную активность биокатализатора и содержание в нем сухих веществ. Для создания выбранных стандартных условий процесса по концентрации биокатализатора объем реакционной массы соответствующего цикла корректируют с учетом снижения активности биокатализатора и уменьшения его массы из-за расхода на аналитические процедуры. Проводят необходимое количество последовательных циклов синтеза цефалексина. Результаты экспериментов, проведенных с использованием биокатализатора, полученного по примеру 1, представлены в таблице 4.
Обработка экспериментальных данных таблицы 4 показывает, что инактивация биокатализатора в процессе синтеза цефалексина подчиняется кинетическим закономерностям первого порядка с периодом полуинактивации τ1/2=200 циклов (см. фиг.2).
Операционная стабильность биокатализатора, полученного по примеру 1, в процессе синтеза цефалексина при температуре 30°С
Источники информации
1. GB 1382255
2. Takahashi Т., Kato К, Yamazaki Y., et al.: Synthesis of cephalosporins and penicillins by enzymatic acylation. Japanese Journal of Antibiotics (Suppl.) 30S: 230-238 (1977)
3. CA 1044627
4. RU 2136759
5. KR 20010069097
6. Ryu Y.W., Ryu D.D.Y., Semisynthetic β-lactam antibiotics synthesizing enzyme from Acetobacter turbindance: purification and propeties. Enzyme Microb. Technol. 9: 339-344 (1987)
7. Kato К., Kawahara К, Takahashi Т., Kakinuma A.: Purification of α-amino acid ester hydrolase from Xanthomonas citri. Agric. Biol. Chem. 44: 1069-1074 (1980)
8. Kato К., Kawahara К, Takahashi Т. et al. α-amino acid ester hydrolase from Xanthomonas citri. Agric. Biol. Chem. 44: 1663-1664 (1980)
9. Крестьянова И.Н., Уваров Н.Н., Руденская Г.Н. и др.: Внутриклеточная аминопептидаза из Xanthomonas rublineans, гидролизующая эфиры α-аминокислот и цефалексин. Биохимия 55 (12): 2226 (1990)
10. Blinkovsky A.M., Markaryan A.N., Synthesis of β-lactam antibiotics containing α-aminophenylacetyl group in the acyl moiety catalyzed by D-(-)-phenylglycyl-β-lactamide amidohydrolase. Enzyme Microb. Technol. 15: 965-973 (1993)
11. Kim D.J., Byun S.M., Purification and properties of ampicillin acylase from Pseudomonas melanogenum. Biochim Biophys Acta 1040 (1): 12-18 (1990)
12. WO 02086111
13. Barends T.R.M., Polderman-Tijmes J.J. et al., The sequence and crystal structure of the α-amino acid ester hydrolase from Xanthomonas citri define a new family of β-lactam antibiotic acylases. J. Biol. Chem. 278: 23076-23084 (2003)
14. WO 02086127
15. Polderman-Tijmes J.J., Jekel P.A. et al.: Identification of the catalytic residues of α-amino acid ester hydrolase from Acetobacter turbindance by labeling and site-directed mutagenesis. J. Biol. Chem. 277, No32: 28474-28482 (2002)
16. Hernandez-Justiz O., Terreni M., Pagani G. et al.: Evaluation of different enzymes as catalysts for the production of b-lactam antibiotics following a kinetically controlled strategy. Enzyme and Microb. Technol., 25, 336 (1999)
17. Choi W.G., Lee S.B., Ryu D.D.Y, Cephalexin synthesis by paratially purified and immobilized enzymes. Biotechnol. Bioeng. 23 (2): 361-371 (1981)
18. Zaslavskaya P.L., Chekalina IV., Igans D.N. et al.: Interrelated approach to optimization of biosynthesis and chemical isolation of biologically active substances: the production of penicillinamidase by Escherichia coli and peptidohydrolase by Xanthomonas sp. Biotechnol. Appl. Biochem. 18: 299-309 (1993)
19. Fernandez-Lafuente R., Hernandez-Justiz O., et al.: Stabilization of tetrameric enzyme (α-amino acid ester hydrolase from Acetobacter turbindance) enables a very improved perfomance of ampicillin syntesis. J. Mol. Catal. B: Enzym. 11: 633-638 (2001)
20. Fernandez-Lafuente R., Hernandez-Justiz O., et al.: Biotransformations catalyzed by multimeric enzymes: stabilization of tetrameric ampicillin acylase permits the optimization of ampicillin synthesis under dissotiation conditions. Biomacromolecules 2 (1): 95-104 (2001)
21. Sapan C.V., Lundblad R.L., Price C.: Review. Colorimertric protein assay technoques. Biotechnol. Appl. Biochem. 29: 99-108 (1999)
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОГЕННОГО БИОКАТАЛИЗАТОРА НА ОСНОВЕ ГИДРОЛАЗЫ ЭФИРОВ АЛЬФА-АМИНОКИСЛОТ, ГЕТЕРОГЕННЫЙ БИОКАТАЛИЗАТОР, ПОЛУЧЕННЫЙ ТАКИМ СПОСОБОМ, И СПОСОБ СИНТЕЗА АМИНОБЕТА-ЛАКТАМНОГО АНТИБИОТИКА ПОД ДЕЙСТВИЕМ ЭТОГО ГЕТЕРОГЕННОГО БИОКАТАЛИЗАТОРА | 2013 |
|
RU2535893C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli - ПРОДУЦЕНТ ГИДРОЛАЗЫ ЭФИРОВ АЛЬФА-АМИНОКИСЛОТ ИЗ Xanthomonas rubrilineans И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ГИДРОЛАЗЫ ЭФИРОВ АЛЬФА-АМИНОКИСЛОТ НА ОСНОВЕ ЭТОГО ШТАММА | 2012 |
|
RU2499830C1 |
| ШТАММ Escherichia coli - ПРОДУЦЕНТ ГИДРОЛАЗЫ ЭФИРОВ АЛЬФА-АМИНОКИСЛОТ ИЗ Xanthomonas rubrilineans И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ГИДРОЛАЗЫ ЭФИРОВ АЛЬФА-АМИНОКИСЛОТ НА ОСНОВЕ ЭТОГО ШТАММА | 2012 |
|
RU2502797C1 |
| СПОСОБ ПОЛУЧЕНИЯ БИОКАТАЛИЗАТОРА, ОБЛАДАЮЩЕГО АКТИВНОСТЬЮ В ОТНОШЕНИИ СИНТЕЗА ЦЕФАЛОСПОРИНОВ-КИСЛОТ | 2010 |
|
RU2420581C1 |
| ПРОМЫШЛЕННЫЙ СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ФЕРМЕНТА ПЕНИЦИЛЛИН G АЦИЛАЗЫ ESCHERICHIA COLI | 2020 |
|
RU2729410C1 |
| Способ получения гетерогенного биокатализатора на основе липазы дрожжей Candida antarctica фракции В | 2016 |
|
RU2650668C1 |
| СПОСОБ ПОЛУЧЕНИЯ ИММОБИЛИЗОВАННОГО БИОКАТАЛИЗАТОРА И СПОСОБ ПОЛУЧЕНИЯ ВОДНЫХ РАСТВОРОВ АМИДОВ С ИСПОЛЬЗОВАНИЕМ ЭТОГО БИОКАТАЛИЗАТОРА | 2007 |
|
RU2352635C2 |
| ИММОБИЛИЗОВАННЫЙ БИОКАТАЛИЗАТОР, СПОСОБ ЕГО ПОЛУЧЕНИЯ И СПОСОБ ПОЛУЧЕНИЯ МОЛОЧНОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО БИОКАТАЛИЗАТОРА | 2002 |
|
RU2253677C2 |
| СПОСОБ ПОЛУЧЕНИЯ БИОКАТАЛИЗАТОРА И БИОКАТАЛИЗАТОР ДЛЯ ДЕТОКСИКАЦИИ ФОСФОРОРГАНИЧЕСКИХ НЕЙРОТОКСИЧНЫХ СОЕДИНЕНИЙ В ПРОТОЧНЫХ СИСТЕМАХ | 2006 |
|
RU2315103C1 |
| ИММОБИЛИЗОВАННЫЙ БИОКАТАЛИЗАТОР ДЛЯ БИОЛОГИЧЕСКОЙ ОЧИСТКИ ЖИРОСОДЕРЖАЩИХ СТОЧНЫХ ВОД И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2006 |
|
RU2315102C1 |
Изобретение относится к биотехнологии. Способ получения гетерогенного биокатализатора для ферментативного синтеза аминобета-лактамного антибиотика на основе внутриклеточной гидролазы эфиров альфа-аминокислот из бактерий рода Xanthomonas, включающий разрушение биомассы клеток Xanthomonas rubrilineans, экстракцию из нее фермента в водную фазу и последующую его иммобилизацию. Причем разрушение клеток осуществляют поэтапно, сначала нарушая их целостность криогенным воздействием, а затем обрабатывая водную суспензию размороженных клеток бутилацетатом. Экстракцию фермента в водную фазу проводят при нагревании обработанной клеточной суспензии сначала в условиях спонтанно устанавливающегося градиента рН, а затем при постоянном значении рН. Получаемый бесклеточный экстракт используют для процедуры иммобилизации фермента путем его осаждения полиэтиленгликолем в виде ферментных агрегатов и их последующей поперечной сшивки в присутствии глутарового альдегида. Полученный таким образом гетерогенный биокатализатор содержит до 0,95 г белка/г сухого биокатализатора и обладает синтетазной активностью до 2200 МЕ/г сухого биокатализатора. Изобретение позволяет повысить эффективность процедуры получения биокатализатора, повысить активность и стабильность получаемого биокатализатора. 3 н. и 3 з.п. ф-лы, 2 ил., 4 табл.
1. Способ получения гетерогенного биокатализатора для ферментативного синтеза аминобета-лактамного антибиотика на основе внутриклеточной гидролазы эфиров альфа-аминокислот из бактерий рода Xanthomonas, включающий разрушение биомассы клеток, экстракцию из нее фермента в водную фазу и последующую его иммобилизацию, отличающийся тем, что используют биомассу клеток бактерий Xanthomonas rubrilineans; разрушение клеток осуществляют поэтапно, сначала нарушая их целостность криогенным воздействием, замораживая биомассу до температуры -25°С или ниже, предпочтительно от -25°С до -30°С, а затем, обрабатывая водную суспензию размороженных клеток с концентрацией биомассы от 20 до 100 мг сух./мл бутилацетатом в концентрации 2÷6 об.%; экстракцию фермента в водную фазу проводят при нагревании обработанной клеточной суспензии сначала в условиях спонтанно устанавливающегося градиента рН, а затем при постоянном значении рН, поддерживаемом путем добавления раствора титранта, вплоть до прекращения процедуры экстракции, используя в качестве критерия выбора момента остановки степень снижения скорости расхода раствора титранта; бесклеточный экстракт, получаемый после отделения разрушенной клеточной биомассы, используют для процедуры иммобилизации фермента путем его осаждения полиэтиленгликолем в виде ферментных агрегатов и их последующей поперечной сшивки в присутствии глутарового альдегида.
2. Способ по п.1, отличающийся тем, что экстракцию фермента осуществляют при температуре 25÷35°С.
3. Способ по п.1, отличающийся тем, что для осаждения ферментных агрегатов из бесклеточного экстракта используют полиэтиленгликоль с молекулярной массой 4000÷10000 в количестве, необходимом для полного осаждения фермента.
4. Способ по п.1, отличающийся тем, что для поперечной сшивки ферментных агрегатов используют глутаровый альдегид в относительной концентрации
1·10-2÷5·10-2 ммоль/мг белка.
5. Гетерогенный биокатализатор для ферментативного синтеза аминобета-лактамного антибиотика на основе внутриклеточной гидролазы эфиров альфа-аминокислот из бактерий рода Xanthomonas, характеризующийся тем, что он получен способом по п.1, содержит до 0,95 г белка/г сухого биокатализатора и обладает синтетазной активностью до 2200 МЕ/г сухого биокатализатора.
6. Способ ферментативного синтеза аминобета-лактамного антибиотика путем ацилирования ключевого бета-лактамного соединения соответствующими производными эфиров Д-аминофенилуксусной кислоты, отличающийся тем, что ферментативную реакцию синтеза проводят под действием гетерогенного биокатализатора по п.5.
| RU 2000102705 A, 27.08.2002 | |||
| RU 22210446 C2, 10.01.2004 | |||
| КУРОЧКИНА В.Б | |||
| и др | |||
| Ферментативный синтез беталактамных антибиотиков, II Аминоцефалоспорины | |||
| - Антибиотики и химиотерапия, №8, 1999, с.6-11 | |||
| Взрывозащищенная электрическая машина | 1984 |
|
SU1179482A1 |

