Группа изобретений относится к биотехнологии, в частности, к биосинтезу гидролазы эфиров альфа-аминокислот и представляет собой рекомбинантный штамм бактерий Escherichia coli (E.coli), содержащий ген гидролазы эфиров альфа-аминокислот, и способный с высокой эффективностью синтезировать гидролазу эфиров альфа-аминокислот, а также способ микробиологического синтеза гидролазы эфиров альфа-аминокислот на основе этого штамма.
Гидролазы эфиров альфа-аминокислот (alpha-amino acid esther hydrolase, AEH) могут быть использованы в качестве биокатализаторов процессов синтеза цефалоспоринов и пеницициллинов, содержащих в боковой цепи аминогруппу в альфа-положении - аминоцефалоспоринов и аминопенициллинов, составляющих группу аминобета-лактамных антибиотиков [1].
Как и другие полусинтетические бета-лактамные антибиотики, аминопенициллины и аминоцефалоспорины в промышленных масштабах получают преимущественно химическим ацилированием соответствующих ключевых полупродуктов, однако, мировой тенденцией является переход к экологически безопасным энергосберегающим методам биокаталитического синтеза [2, 3]. Успехи генетической инженерии по созданию рекомбинантных штаммов-продуцентов различных ферментов трансформации бета-лактамов создают основу для внедрения биокатализа в промышленное производство полусинтетических пенициллинов и цефалоспоринов.
В источниках информации имеются сведения о микроорганизмах - продуцентах внутриклеточной AEH, представляющих собой природные штаммы или штаммы, полученные методами селекции: Acetobacter turbidans [4-6], Pseudomonas melanogenum [7-13], Flavobacterium [14], Xanthomonas citri [15-19], Xanthomonas rubrilineans [20]. Описаны также полученные методами генной инженерии штаммы E.coli - продуценты внутриклеточных рекомбинантных AEH из Acetobacler turbidans, Zymomonas mobilise, Xanthomonas citri, Xanthomonas campestris pv. campestris, Xanthomonas rubrilineans [21-29]. В различных источниках количественную характеристику продуцентов проводили по различным видам ферментативной активности (гидролиз эфиров; гидролиз аминобета-лактамов, например, цефалексина, синтез антибиотиков, как правило, цефалексина). Сопоставление имеющихся данных показало, что в большинстве описанных случаев удельная (в пересчете на содержание белка) ферментативная активность биомассы клеток по синтезу цефалексина из 7-аминодезацетоксицефалоспорановой кислоты и метилового эфира D-фенилглицина не превышает 2 МЕ/мг белка.* (За международную единицу (ME) ферментативной активности препарата в отношении какой-либо биокаталитической трансформации принимают такое количество этого препарата, которое катализирует превращение 1 мкмоля субстрата (или образование 1 мкмоля продукта) в стандартных условиях за 1 минуту.) Сопоставление проведено с учетом приблизительного соотношения скоростей перечисленных выше ферментативных реакций [16], а также того факта, что при применяемых мягких методах разрушения клеток удельная активность исходной клеточной биомассы не выше удельной активности получаемого из нее бесклеточного экстракта. Исключение составляет рекомбинантный штамм Е. coli - продуцент АЕН из Acetobacter turbidans, на основе которого получен бесклеточный экстракт с удельной активностью 17,6 МЕ/мг белка [22, 24].
Известные работы по получению методами генной инженерии штаммов Escherichia coli - продуцентов АЕН направлены, в первую очередь, на наработку чистого фермента с целью изучения его структуры и свойств и не привели к существенному увеличению продуктивности рекомбинантных штаммов по сравнению с исходными продуцентами. Например [21], при разрушении клеток ультразвуком и извлечении АЕН из Acetobacter turbidans ATCC 9325 получают бесклеточный экстракт с удельной активностью 1,3 МЕ/мг белка, а созданный на основе Е. coli генно-инженерный продуцент АЕН из Acetobacter turbidans обеспечивает удельную активность лишь 2 МЕ/мг белка.
Повышение уровня продукции гидролаз эфиров альфа-аминокислот является необходимым этапом создания высокоэффективных технологических биокатализаторов для промышленных процессов производства полусинтетических аминопенициллинов и аминоцефалоспоринов и может быть достигнуто путем использования гетерологичных микроорганизмов, на что и направлена заявляемая группа изобретений.
Ближайшим аналогом заявляемого штамма является штамм Е. coli BL21 (DE3), содержащий на плазмиде рЕТ-28а ген aeh из Xanlhomonas rubrilineans CPCC 140817, содержащий мутацию V131S, полученную путем сайт-специфического мутагенеза и приводящую к замене серина, находящегося в 131 положении, на валин. Целью мутагенеза являлось не повышение продуктивности штамма, а увеличение специфичности фермента в сторону синтеза цефалоспоринов, содержащих в боковой цепи пара-гидроксифенильную группу по сравнению с синтезом антибиотиков, содержащих фенильную группу [29]. Несмотря на использование экспрессионной системы рЕТ (Novagen), полученный штамм Е.coli не является суперпродуцентом активной АЕН, поскольку синтезирующийся рекомбинантный белок содержит на N-конце последовательность из 33 аминокислот, предшествующую лидерному пептиду и содержащую полигистидиновый «хвост» (His-Tag) и фрагмент гена 10 бактериофага Т7 (T7-Tag). Данная последовательность существенно затрудняет посттрансляционный транспорт белка в периплазму, в процессе которого происходит созревание белка вследствие отщепления лидерного пептида. В результате, лишь небольшая часть синтезированного белка превращается в активный фермент [30].
Ближайшим аналогом заявляемого способа микробиологического синтеза гидролазы эфиров альфа-аминокислот из Xanthomonas rubrilineans является способ с использованием мутантного штамма A" rubrilineans ВКПМ В-9915, продуцирующего данный фермент [31, 32]. Ферментативная активность биомассы клеток по синтезу цефалексина из 7-аминодезацетоксицефалоспорановой кислоты (7-АДЦК), взятой в концентрации 0,04 М, и метилового эфира D-фенилглицина (МЭФГ), взятого в концентрации 0,08 М, при 40°С и рН 6.0 составляет 83,3 МЕ/г влажн., 450 МЕ/г сух. Удельная активность клеток может быть примерно оценена как 1 МЕ/мг белка, учитывая, что суммарный белок составляет примерно половину сухого веса биомассы данного штамма.
Задача заявляемой группы изобретений состоит в разработке способа микробиологического синтеза гидролазы эфиров альфа-аминокислот из X.rubrilineans ВКПМ В-9915 с повышенной продуктивностью.
Задачу решают путем:
- конструирования рекомбинантной плазмидной ДНК (плазмиды) pAEH-OZ, соответствующей нуклеотидной последовательности SEQ ID NO 1 и содержащей ген aehR гидролазы эфиров альфа-аминокислот из Xanlhomonas rubrilineans ВКПМ В-9915, не модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем промотора Т7, а также ген lacI, ген устойчивости к канамицину Кап и участки инициации репликации pBR322 и f1;
- конструирования рекомбинантного штамма бактерий Escherichia coli ВКПМ В-11271 - продуцента гидролазы эфиров альфа-аминокислот из Xanthomonas rubrilineans ВКПМ В-9915 путем трансформации штамма-реципиента Escherichia coli BL21 (DE3) плазмидой pAEH-OZ;
- разработки способа микробиологического синтеза гидролазы эфиров альфа-аминокислот из Xanthomonas rubrilineans ВКПМ В-9915 путем культивирования рекомбинантного штамма-продуцента Escherichia coli ВКПМ В-11271 в подходящей питательной среде с индукцией синтеза целевого продукта добавлением изопропил-β-D-тиогалактозида (ИПТГ) до конечной концентрации 1 мМ после достижения культурой плотности 0,8-1,0 ОЕ.
Работа включает:
- клонирование гена aehR, кодирующего гидролазу эфиров альфа-аминокислот из X. rubrilineans ВКПМ В-9915;
- конструирование рекомбинантной плазмидной ДНК (плазмиды) рАЕН, содержащей ген гидролазы эфиров альфа-аминокислот из X. rubrilineans ВКПМ В-9915 {aehR), находящийся под контролем Т7 промотора и модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост»;
- конструирование рекомбинантного штамма бактерий E. coli BL21 (DE3)/pAEH, содержащего плазмиду рАЕН и способного синтезировать гидролазу эфиров альфа-аминокислот из X. rubrilineans ВКПМ В-9915;
- конструирование, на основе плазмиды рАЕН рекомбинантной плазмиды pAEH-OZ, содержащей ген гидролазы эфиров альфа-аминокислот из X. rubrilineans ВКПМ В-9915 (aehR), не модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем промотора Т7;
- конструирование рекомбинантного штамма бактерий E.coli BL21 (DE3)/pAEH-OZ, содержащего плазмиду pAEH-OZ и способного с высокой эффективностью синтезировать гидролазу эфиров альфа-аминокислот из Х. rubrilineans ВКПМ В-9915. Экспрессируемый плазмидой pAEH-OZ рекомбинантный белок не модифицирован на N-конце полигистидиновой последовательностью, что облегчает процесс его пострансляционного созревания и повышает уровень продукции активного фермента АЕН штаммом BL21 (DE3)/pAEH-OZ по сравнению со штаммом BL21 (DE3)/pAEH.
- разработку способа микробиологического синтеза гидролазы эфиров альфа-аминокислот из X.rubrilineans ВКПМ В-9915 на основе рекомбинантных бактерий Е. coli BL21 (DE3)/pAEH-OZ, обладающих повышенным уровнем продукции целевого фермента.
Процесс конструирования заявляемого штамма состоит из нескольких этапов.
Этап 1. Конструирование рекомбинантной плазмидной ДНК рАЕН, содержащей ген aehR, кодирующий гидролазу эфиров альфа-аминокислот из X.rubrilineans ВКПМ В-9915
Источником гена гидролазы эфиров альфа-аминокислот является штамм X.rubrilineans ВКПМ В-9915, геномную ДНК которого, выделенную из биомассы штамма X.rubrilineans ВКПМ В-9915, используют в качестве матрицы для амплификации кодирующей части гена aehR.
Плазмиду рАЕН конструируют путем клонирования ПЦР-фрагмента, содержащего кодирующую область гена aehR, в экспрессионный вектор рЕТ-28а (Novagen) по сайтам рестрикции NdeI и XhoI. Плазмида рАЕН размером 7210 пар оснований содержит ген aehR модифицированный с 5' конца последовательностью, кодирующей 6 аминокислотных остатков гистидина - полигистидиновый «хвост» (His-Tag) и находящийся под контролем Т7 промотора и терминатора; кодирующую область гена lacI; кодирующую область гена Кап, обеспечивающую устойчивость штаммов Е.coli к канамицину; участок инициации репликации pBR322; участок инициации репликации f1 (Фиг.1).
Этап 2. Конструирование рекомбинантной плазмидной ДНК pAEH-OZ, содержащей ген гидролазы эфиров альфа-аминокислот из X. rubrilineans ВКПМ В-9915 (aehR), не модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост».
Плазмиду pAEH-OZ конструируют на основе плазмиды рАЕН путем удаления из нее фрагмента ДНК, расположенного между сайтами рестрикции NdeI и NcoI.
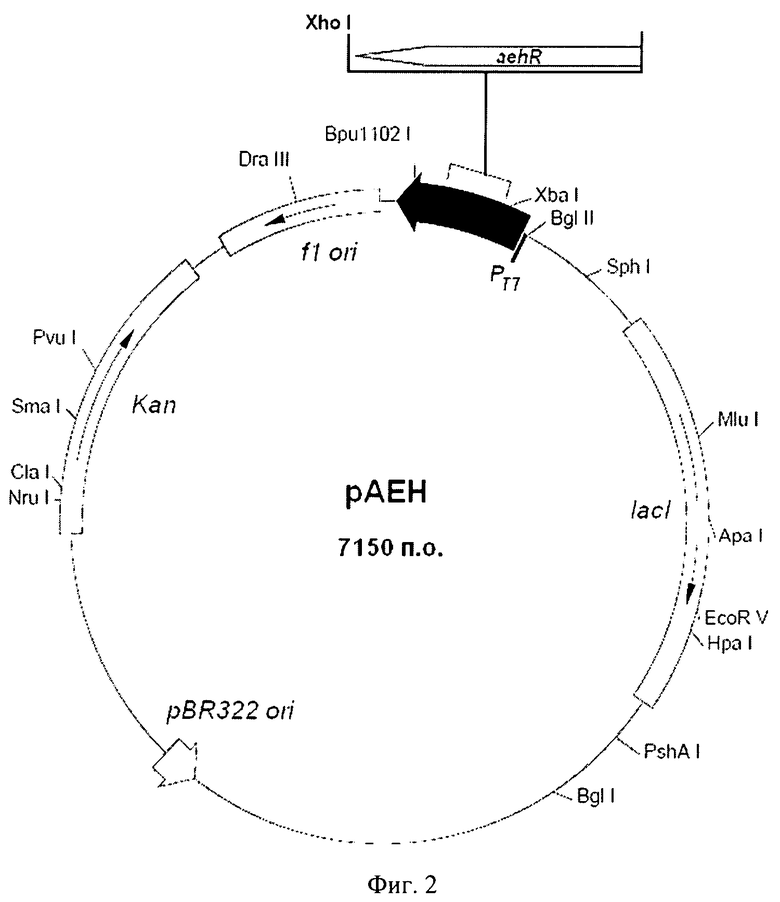
Рекомбинантная плазмидная ДНК pAEH-OZ (SEQ ID NO 1) размером 7150 пар оснований, содержит все элементы вектора рАЕН, кроме фрагмента ДНК, расположенного между сайтами рестрикции NdeI и NcoI и включающего нуклеотидную последовательность, граничащую с 5'-концом гена aehR и кодирующую полигистидиновый «хвост» (Фиг.2). Экспрессируемый данной плазмидой рекомбинантный белок АЕН не модифицирован полигистидиновой последовательностью.
Этап 3. Трансформация плазмидой pAEH-OZ штамма Е. coli BL21 (DE3)
Компетентные клетки штамма Е. coli BL21 (DE3) трансформируют плазмидой pAEH-OZ. В результате получают рекомбинантный штамм Е. coli BL21 (DE3)/pAEH-OZ, который содержит ген гидролазы эфиров альфа-аминокислот из X. rubrilineans ВКПМ В-9915 (aehR), не модифицированный на 5'-конце последовательностью, кодирующей полигистидиновый «хвост», вследствие чего рекомбинантный штамм с высокой эффективностью синтезирует гидролазу эфиров альфа-аминокислот из X. rubrilineans ВКПМ В-9915 (Фиг.3).
Штамм Е. =coli BL21 (DE3)/pAEH-OZ депонирован во Всероссийской коллекции промышленных микроорганизмов как Е.coli ВКПМ В-11271.
Заявляемый штамм ВКПМ В-11271 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки
Клетки прямые, палочковидные, подвижные, грамотрицателиные, неспорообразующие. При выращивании в течение 24-72 час при температуре 30-37°С на агаризованных средах (LB, 2YT), содержащих канамицин (17 мг/л), колонии гладкие, круглые, блестящие, край ровный.
Состав среды LB, мас.%:
Бакто-триптон - 1
дрожжевой экстракт - 0,5
NaCl - 1,
вода - остальное.
Состав среды 2YT, мас.%:
Бакто-триптон - 1,6
дрожжевой экстракт - 1,0
NaCl - 0,5
вода - остальное.
Физиолого-биохимические признаки
Штамм растет при температуре от 25 до 40°С (оптимум 37°С). В качестве источника азота использует органический азот в виде пептона, аминокислот.
Штамм Е.coli ВКПМ В-11271 синтезирует гидролазу эфиров альфа-аминокислот из X.rubrilineans после индукции 1 мМ ИПТГ (изопропил-β-D-тиогалактозидом).
Генотипические признаки
Штамм Е.coli ВКПМ В-11271 устойчив к канамицину (17 мг/л).
Штамм Е.coli ВКПМ В-11271 содержит на плазмиде pAEH-OZ ген гидролазы эфиров альфа-аминокислот (aehR) из Х. rubrilineans ВКПМ В-9915, не модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем Т7 промотора.
Способ микробиологического синтеза гидролазы эфиров альфа-аминокислот в общем виде
Посевной материал, представляющий собой клетки рекомбинантного штамма-продуцента Е.coli ВКПМ В-11271, подготавливают путем инкубации в течение 16 часов при температуре 37°С на среде LB, содержащей 17 мг/мл канамицина. Выросшую культуру переносят в соотношении 1:200 (по объему) в среду LB, содержащую 17 мг/мл канамицина. Процесс культивирования ведут при температуре 37°С с аэрацией. Синтез целевого белка индуцируют путем добавления ИПТГ при OD600=0,8-1,0 ОЕ. Биомассу клеток отделяют центрифугированием, промывают фосфатным буфером и повторно центрифугируют.
Уровень синтетазной активности гидролазы эфиров альфа-аминокислот в биомассе клеток составляет не менее чем 4100 МЕ/г влажн., 22000 МЕ/г сух.
Заявляемый способ позволяет повысить уровень продукции гидролазы эфиров альфа-аминокислот до уровня не менее 50 МЕ/мг белка, что в 50 раз превышает продукцию ближайшего аналога.
Изобретение проиллюстрировано следующими фигурами графических изображений:
Фиг.1. Схема рекомбинантной плазмидной ДНК рАЕН.
aehR - ген гидролазы эфиров альфа-аминокислот, модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем Т7 промотора; lacI - регулятор транскрипции; Кап - ген устойчивости к канамицину; pBR322 ori и f1 ori участки инициации репликации.
Фиг.2. Схема рекомбинантной плазмидной ДНК pAEH-OZ.
aehR - ген гидролазы эфиров альфа-аминокислот, не модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем Т7 промотора; lacI - регулятор транскрипции; Kan - ген устойчивости к канамицину; pBR322 ori и/7 ori участки инициации репликации.
Фиг.3 Электрофоретический анализ внутриклеточных белков штаммов Е.coli ВКПМ В-11246 (АЕН) и Е.coli ВКПМ В-11271 (AEH-OZ) после индукции ИПТГ. М - маркер молекулярного веса Prestained Protein Ladder (Fermentas). Электрофоретическое разделение белков проводили в денатурирующих условиях в 15% ПААГ. Окрашивание гелей серебром проводили, используя реактивы набора К0681 и протокол фирмы Fermentas.
Пример 1. Конструирование рекомбинантной плазмидной ДНК рАЕН, содержащей ген гидролазы эфиров альфа-аминокислот из X.rubrllineans ВКПМ В-9915 (aehR), модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем Т7 промотора.
В качестве источника гена aehR используют геномную ДНК, выделенную из биомассы штамма X.rubrilineans ВКПМ В-9915, полученного из музея культур ВКПМ. Все стандартные генно-инженерные и микробиологические манипуляции проводят по известным методикам [33].
Культуру штамма X.rubrilineans ВКПМ В-9915 выращивают при 27°С в течение 28 часов. Из полученной биомассы выделяют геномную ДНК, которую используют в качестве матрицы для ПЦР.
Для амплификации кодирующей части гена aehR используют праймеры, содержащие сайты рестрикции NdeI и XhoI (PrAEH_for 5'-gcatatgcgccgcatcgctccctg-3' и PrAEH_rev 5'-cctcgagtcagtacaccggcagactgatgaaactgg-3'), сконструированные на основе последовательности гена aehR из X.rubrilineans, и Pfu ДНК-полимеразу повышенной точности (Fermentas). Полимеразную цепную реакцию проводят согласно методике компании Fermentas. Полученный ПЦР-продукт включает кодирующую область гена aehR размером 1914 п.о., в том числе и лидерный пептид.
С целью получения плазмидной конструкции для экспрессии гидролазы эфиров альфа-аминокислот ПЦР-продукт, содержащий кодирующую область гена aehR, клонируют в экспрессионный вектор рЕТ-28а (Novagen). Для этого вектор рЕТ-28а и ПЦР-продукт обрабатывают рестриктазами NdeI и XhoI. Из рестрикционной смеси выделяют фрагмент вектора размером 5289 п.о. и фрагмент ПЦР-продукта размером 1921 п.о. Выделенные фрагменты смешивают и лигируют; полученной лигазной смесью трансформируют клетки Е.coli DH5a. Селекцию клонов проводят на агаризованной среде LB с канамицином. Правильность клонирования и отсутствие ошибок в последовательности гена aehR подтверждают секвенированием плазмидной вставки. Рекомбинантная плазмидная ДНК рАЕН имеет размер 7210 пар нуклеотидов (п.н.) и содержит ген гидролазы эфиров альфа-аминокислот (aehR) из X. rubrilineans ВКПМ В-9915, модифицированный на 5'-конце последовательностью, кодирующей полигистидиновый «хвост», и регулируемый Т7 промотором и терминатором; ген Кап, обеспечивающий устойчивость Е.coli к канамицину; участки инициации репликации pBR322 и f1; кодирующую область гена lacI (Фиг.1).
Пример 2. Трансформация штамма Е.coli BL21 (DE3) плазмидой рАЕН
С целью получения рекомбинантного штамма Е.coli - продуцента гидролазы эфиров альфа-аминокислот, клетки штамма Е. coli BL21 (DE3) трансформируют плазмидой рАЕН методом химической трансформации. Селекцию трансформантов проводят на LB-агаре, содержащем канамицин (17 мг/л). В результате получают штамм Е.coli, синтезирующий гидролазу эфиров альфа-аминокислот из X.rubrilineans ВКПМ В-9915, модифицированную на N-конце полигистидиновым «хвостом». His-Tag снижает эффективность транспорта синтезированного белка в периплазму, в процессе которого происходит отщепление лидерного пептида и созревание белка. Поэтому лишь часть рекомбинантной АЕН переходит в активную форму (Фиг.3), вследствие чего уровень синтеза активного фермента снижается. Поэтому следующим этапом работы стало удаление из плазмидной конструкции рАЕН нуклеотидной последовательности, кодирующей полигистидиновый «хвост».
Штамм Е.coli BL21 (DE3)/pAEH депонирован во Всероссийской коллекции промышленных микроорганизмов как Е.coli ВКПМ В-11246.
Пример 3. Конструирование рекомбинантной плазмидной ДНК pAEH-OZ, содержащей ген гидролазы эфиров альфа-аминокислот из X.rubrilineans ВКПМ В-9915 (aehR), не модифицированный с 5' конца последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем Т7 промотора.
Плазмиду pAEH-OZ конструируют на основе плазмиды рАЕН. С целью удаления последовательности, кодирующей 6 His с 5'-конца открытой рамки считывания, включающей кодирующую часть гена aehR., плазмидный вектор рАЕН обрабатывают рестриктазой NcoI, после чего удаляют образовавшиеся выступающие 5'-концы линеаризованной молекулы ДНК с помощью нуклеазы S1. Плазмидную ДНК очищают на колонке Quiagen и обрабатывают рестриктазой NdeI, после чего заполняют образовавшиеся выступающие 5'-концы с помощью Фрагмента Кленова. Обработку плазмидной ДНК перечисленными ферментами производят в соответствии с рекомендациями производителя (Fermentas, Латвия). Из рестрикционной смеси выделяют фрагмент ДНК размером 7150 п.о. и циркуляризируют его путем лигирования. Полученной лигазной смесью трансформируют клетки Е. coli DH5α. Селекцию клонов проводят на агаризованной среде LB с канамицином. В результате получают рекомбинантный штамм Е. coli DH5α/pAEH-OZ, содержащий плазмиду pAEH-OZ. Правильность клонирования подтверждают рестрикционным картированием. Штамм Е.coli DH5α/pAEH-OZ не синтезирует гидролазу эфиров альфа-аминокислот, из-за отсутствия в клетках штамма Е.coli DH5a Т7 РНК полимеразы.
Рекомбинантная плазмидная ДНК pAEH-OZ (SEQ ID NO 1) имеет размер 7150 пар нуклеотидов (п.н.) и содержит ген гидролазы эфиров альфа-аминокислот (aehR) из X.rubrilineans ВКПМ В-9915, регулируемый Т7 промотором и терминатором; ген Kan, обеспечивающий устойчивость Е.coli к канамицину; участки инициации репликации pBR322 и f1; кодирующую область гена lacI. На 5'-конце открытой рамки считывания, включающей кодирующую часть гена aehR, отсутствует последовательность, кодирующая 6 His (Фиг.2).
Пример 4. Получение заявляемого штамма Е.coli ВКПМ В-11271
С целью получения рекомбинантного штамма Е.coli - продуцента гидролазы эфиров альфа-аминокислот, клетки штамма Е.coli BL21 (DE3) трансформируют плазмидой pAEH-OZ методом химической трансформации. Селекцию трансформантов проводят на LB-агаре, содержащем канамицин (17 мг/л). В результате получают заявляемый штамм Е.coli ВКПМ В-11271, синтезирующий гидролазу эфиров альфа-аминокислот из X. rubrilineans ВКПМ В-9915, не модифицированную на N-конце полигистидиновой последовательностью, что облегчает посттрансляционное созревание экспрессирующегося белка и повышает выход активного фермента (Фиг.3), (пример 5).
Пример 5. Биосинтез гидролазы эфиров альфа-аминокислот с использованием штамма Е.coil ВКПМ В-11271 и добавлением индуктора ИПТГ
Исходным посевным материалом служит культура Е. coli ВКПМ В-11271, выращенная на чашках Петри с агаризованной средой. Посевной материал выращивают в путем инкубации клеток при температуре 37°С в течение 15-17 часов на среде LB, содержащей 17 мг/мл канамицина.
Процесс биосинтеза ведут к колбах Эрленмейера объемом 750 мл, содержащих 100 мл среды LB с канамицином (17 мг/л). Посевной материал вносят в ферментационную среду в количестве, необходимом для создания его концентрации около 0,5 об.%. Процесс культивирования продуцента ведут при температуре 37°С на круговой качалке со скоростью вращения 200-220 об/мин до достижения оптической ОD600=0,8-1,0 ОЕ (около 3 часов культивирования), после чего в культуральную жидкость добавляют ИПТГ до концентрации 1 мМ. Ферментацию продолжают при тех же условиях в течение еще 3 часов.
Биомассу клеток отделяют центрифугированием при 5000 об/мин, промывают 0,1 М фосфатным буфером, рН 7.5 и вновь центрифугируют.
Из 2 л ферментационной среды (20 колб) получают 9,73 г влажной биомассы.
Активность ферментных препаратов по синтезу цефалексина определяют по начальной скорости образования целевого продукта в следующих условиях: рН 6,0-6,2, температура (40±1)°С, концентрации субстратов - (0,040±0,002) М 7-АДЦК и (0,080±0,004) М МЭФГ. Содержание цефалексина в реакционной смеси определяют методом высокоэффективной жидкостной хроматографии (ВЭЖХ). Содержание сухих веществ определяют весовым методом после высушивания препарата при (105±1)°С до постоянной массы. Содержание белка в биомассе клеток определяют методом Лоури [34]. Показано, что этот показатель не зависит от степени разрушения клеток ультразвуком, то есть в нативных клетках он определен корректно.
Полученная биомасса клеток характеризуется следующими показателями:
- содержание сухих веществ: 19,4%;
- синтетазная активность: 4880 МЕ/г влажн.; 25140 МЕ/г сух.;
- удельная активность: 53,4 МЕ/мг белка.
Пример 6. Биосинтез гидролазы эфиров альфа-аминокислот с использованием штамма Е.coil ВКПМ В-11271 без добавления индуктора
Способ осуществляют по примеру 5, с той разницей, что культивирование ведут в течение 6 часов без добавления индуктора ИПТГ.
Из 200 мл культуральной жидкости получают 0,93 г влажной биомассы.
Полученная биомасса клеток характеризуется следующими показателями:
- содержание сухих веществ -18,0%;
- синтетазная активность: 190 МЕ/г влажн.; 1070 МЕ/г сух.;
- удельная активность: 2,0 МЕ/мг белка.
Пример 7 (Контроль). Биосинтез гидролазы эфиров альфа-аминокислот с использованием штамма Е. coli ВКПМ В-11246
Способ осуществляют по примеру 5, используя для ферментации культуру штамма Е.coli ВКПМ В-11246. В отличие от заявляемого штамма Е.coli ВКПМ В-11271, штамм штамма Е.coli ВКПМ В-11246 экспрессирует гидролазу эфиров альфа-аминокислот из X.rubrilineans, модифицированную на N-конце полигистидиновой последовательностью.
Из 2 л ферментационной среды (20 колб) получают 7,45 г влажной биомассы.
Полученная биомасса клеток характеризуется следующими показателями:
- содержание сухих веществ: 21,4%;
- синтетазная активность: 2020 МЕ/г влажн.; 9440 МЕ/г сух.;
- удельная активность: 23,8 МЕ/мг белка.
Высокий уровень синтетазной активности гидролазы эфиров альфа-аминокислот в биомассе рекомбинантного штамма Е.coli ВКПМ В-11271 после индукции экспрессии, по сравнению с неиндуцированной культурой, подтверждает экспрессию гена aehR из X.rubrilineans в Е. coli (Табл.1).
Уровень синтеза активной гидролазы эфиров альфа-аминокислот у заявляемого рекомбинантного штамма Е.coli ВКПМ В-11271 в 2,2 раза превосходит таковой у штамма Е.coli ВКПМ В-11246, экспрессирующего гидролазу эфиров альфа-аминокислот из X.rubrilineans, модифицированную на N-конце полигистидиновой последовательностью (Табл.1).
Уровень синтеза гидролазы эфиров альфа-аминокислот у заявляемого рекомбинантного штамма Е.coli ВКПМ В-11271 в 50 раз превосходит таковой у ближайшего аналога-штамма X. rubrilineans ВКПМ В-9915 (Табл.1).

Результаты биосинтеза АЕН различными штаммами-продуцентами данного фермента сопоставлены в Таблице 1.
Таким образом, в заявляемых результатах продемонстрировано, что генетически модифицированные бактерии Е.coli, способны экспрессировать гидролазу эфиров альфа-аминокислот из X.rubrilineans, причем высокий уровень продукции активного фермента обусловлен, в частности, отсутствием на N-конце продуцируемого белка полигистидинового «хвоста», существенно затрудняющего посттрансляционный транспорт белка в периплазму, в процессе которого происходит созревание белка вследствие отщепления лидерного пептида. Экспрессируемый плазмидой pAEH-OZ рекомбинантный белок не модифицирован на N-конце полигистидиновой последовательностью, что облегчает процесс его пострансляционного созревания и повышает уровень продукции активного фермента АЕН штаммом BL21 (DE3)/pAEH-OZ по сравнению со штаммом BL21 (DE3)/pAEH.
Высокое содержание целевого белка в клеточной биомассе, получаемой по заявляемому способу, создает возможности для разработки простых методов получения высокоэффективных технологических биокатализаторов процессов синтеза аминопенициллинов и аминоцефалоспоринов путем иммобилизации целых клеток, исключая стадии выделения, концентрирования и очистки гидролазы эфиров альфа-аминокислот.
Источники информации
1. Deaguero A.L., Blum J.K., Bommarius A.S. Biocatalytic synthesis of β-lactam antibiotics. // In: Flickinger M.C. (Ed) Encyclopedia of Industrial Biotechnology: bioprocess, bioseparation, and cell technology. John Wiley & Sons. - 2010. - Inc.: 1-32
2. Volpato G., Rodriges R.C., Femandez-Lafuente R. Use of enzymes in production of semi-synthetic penicillins and cephalosporins: drawbacks and perspectives. // Curr Med Chem. - 2010. - V.17. - P.3855-3873
3. ElanderR.P. Industrial production of β-lactam antibiotics. // Appl. Microbiol. Biotechnol. - 2003. - V.61. - P.385-392
4. Takahashi Т, Yamazaki Y, Kato K (1974) Substrate specificity of an alpha-amino acid ester hydrolase produced by Acetobacter turbidans ATCC 9325. Biochem J 137(3): 497-503
5. Ryu YW, Ryu DY (1987) Semisynthetic β-lactam antibiotics synthesizing enzyme from Acetobacter turbidans: purification and properties. Enzyme Microb Technol 9: 339-344
6. Ryu YW, Ryu DY (1988) Semisynthetic β-lactam antibiotics synthesizing enzyme from Acetobacter turbidans: catalytic properties. Enzyme Microb Technol 10: 239-245
7. Okachi R, Kato К, Miyamura Y, Nara Т (1973) Selection of Pseudomonas melanogenum KY 3987 as a new ampicillin producing bacteria. Agric Biol Chem 37 (8): 1953-1957
8. Okachi R, Nara Т (1973) Penicillin acylase of Pseudomonas melanogenum KY 3987. Agric Biol Chem 37 (12): 2797-2804
9. Shimizu M, Masuike Т, Fujita H, Kimura К, Okachi R, Nara Т (1975) Search for microorgamisms producing cephalosporin acylase and enzymatic synthesis of cephalosporins. Agric Biol Chem 39 (6): 1225-1232
10. Kawamori M, Hashimoto Y, Katsumata R, Okachi R, Takayama К (1983) Enzymatic production ofamoxicillin by β-Lactamase-deficient mutants of Pseudomonas melanogenum KY3987. Agric Biol Chem 47 (11): 2503-2509
11. Kim DJ, Byun SM (1990) Purification and properties of ampicillin acylase from Pseudomomas melanogenum. Bichim Biophis Acta - Prot Structure and Mol Enzymol 1040:12-18
12. Kim DJ, Byun SM (1990) Evidence for involvement of 2 histiddine residues in the reaction of ampicillin acylase. Bichem Biophys Res Comm 166 (2): 904-908
13. Wang M, Wang Z, Yue H, Han W, Jiao Q (1990) Screening of alpha-amino acid ester hydrolase producing strain and synthesis ofcephalexin by Pseudomonas aeruginosa. Wei Sheng Wu Xue Bao 30: 238-241
14. US 3716454 (A) (1973) Process for the production of α-aminobenzilpenicillin
15. Kato K, Kavahara K, Takahashi T, Kahinuma A (1980) Purification of α-Amino Acid Ester Hydrolase from Xanthomonas citri. Agric Biol Chem 44 (5): 1069-1074
16. Kato К, Kavahara К, Takahashi T, Kahinuma A (1980) Substrate Specificity of α-Amino Acid Ester Hydrolase from Xanthomonas citri. Agric Biol Chem 44 (5): 1075-1081
17. Kato K, Kavahara K (1980) Dissociation and reassociation of Xanthomonas a-amino acid ester hydrolase. Agric Biol Chem 44 (7): 1663-1664
18. Nam DH, Kim С (1985) Reaction kinetics ofcephalexin synthesizing enzyme from Xanthomonas citri. Biothechnol Bioeng 27: 953-960
19. Kato K, Kavahara К, Takahashi Т (1980) Enzymatic synthesis ofamoxicillin by the cell-bound α-amino acid ester hydrolase of Xanthomonas citri. Agric Biol Chem 44(4):821-825
20. Крестьянова И.Н., Уваров Н.Н., Руденская Г.Н. и др. (1990) Внутриклеточная аминопептидаза из Xanthomonas rublineans, гидролизующая эфиры α-аминокислот и цефалексин. Биохимия 55(12): 2226-2238.
21. Polderman-Tijmes JJ, Jekel PA, van Merode A, Floris TAG, van der Laan JM, Sonke T, Janssen DB (2002) Cloning, sequence and expression in Escherichia coli of the gene encoding a-amino acid ester hydrolase from Acetobacter turbindance. Appi Environ Microbiol 68: 211-218 A
22. Polderman-Tijmes JJ., Jekel PA, Jeronimus-Stratingh CM, Bruins AP, van der Laan J-M, Sonke T, Janssen DB (2002) Identification of the catalytic residues of α-amino acid ester hydrolase from Acetobacter turbindance by labeling and site-directed mutagenesis. J Biol Chem 277 (32): 28474-28482
23. Barends RM, Polderman-Tijmes JJ, Jekel PA, Willams C, Wybenga G, Janssen DB, Dijkstra BW (2006) Acetobacter turbidans a-amino acid ester hydrolase: how a single mutation improves an antibiotic-producing enzyme. J Biol Chem 281(9): 5804-5810
24. WO 02086111 (2002) Recombinant alpha-amino ester hydrolases and uses thereof
25. WO 02086127 (2002) Acylase gene
26. Barends TRM, Polderman-Tijmes JJ, Jekel PA, Hensgens CMH, de Vries EJ, Janssen DB, Dijkstra BW (2003) The sequence and crystal structure of the α-amino acid ester hydrolase from Xanthomonas citri define a new family of β-lactam antibiotic acylases. J Biol Chem 278: 23076-23084
27. Blum JK, Bommarius AS (2010) Amino ester hydrolase from Xanthomonas campestris pv. campestris, ATCC 33913 for enzymatic synthesis of ampicillin. J Mol Catal B: Enzym 67:21-28
28. Wang L, Ye LJ, Pan Y, Cao Y (2012) Two plate-based colorimetric assays for screening α-amino acid ester hydrolase with high synthesis/hydrolysis ratio. Enzyme MicrobTechnol51:107-12
29. Ye LJ, Wang L, Pan Y, Cao Y (2012) Changing the specificity of α-amino acid ester hydrolase toward para-hydroxyl cephalosporins synthesis by site-directed saturation mutagenesis. Biotechnol Lett 34:1719-24.
30. J.H. Choi. S.Y. Lee (2004) Secretory and extracellular production of recombinant proteins using Escherichia coli Appi Microbiol Biotechnol 64: 625-635
31. RU 2381273 (2009) Способ получения гетерогенного биокатализатора, биокатализатор на основе гидролазы эфиров альфа-аминокислот и способ синтеза аминобета-лактамного антибиотика под действием этого биокатализатора
32. CN 101525603 (2009) Immobilized alpha-amino-acid ester hydrolase, preparation and application thereof
33. Sambrook J, Fritsch EF, Maniatis Т (1989) Molecular Cloning: a Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor
34. Sapan CV, Lundblad RL, Price С (1999) Review. Colorimertric protein assay technoques. Biotechnol. Appl. Biochem. 29: 99-108





Изобретение относится к биотехнологии, а именно к способу получения гидролазы эфиров альфа-аминокислот путем биосинтеза рекомбинантными бактериями. Для осуществления способа сконструирован рекомбинантный штамм бактерий Escherichia coli ВКПМ В-11271 - продуцент гидролазы эфиров альфа-аминокислот из Xanthomonas rubrilineans ВКПМ В-9915, полученный путем трансформации штамма-реципиента Escherichia coli BL21(DE3) плазмидной ДНК, соответствующей нуклеотидной последовательности SEQ ID NO 1, содержащей ген aehR гидролазы эфиров альфа-аминокислот из Xanthomonas rubrilineans ВКПМ В-9915, не модифицированный на 5′-конце последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем промотора Т7, а также ген устойчивости к канамицину Kan. Изобретение позволяет получать гидролазу эфиров альфа-аминокислот с высокой степенью эффективности. 2 н.п. ф-лы, 3 ил., 1 табл., 7 пр.
1. Рекомбинантный штамм бактерий Escherichia coli ВКПМ В-11271 - продуцент гидролазы эфиров альфа-аминокислот из Xanthomonas rubrilineans ВКПМ В-9915, полученный путем трансформации штамма- реципиента Escherichia coli BL21 (DE3) плазмидной ДНК, соответствующей нуклеотидной последовательности SEQ ID NO 1, содержащей ген aehR гидролазы эфиров альфа-аминокислот из Xanthomonas rubrilineans ВКПМ В-9915, не модифицированный на 5′-конце последовательностью, кодирующей полигистидиновый «хвост», и находящийся под контролем промотора Т7, а также ген устойчивости к канамицину Каn.
2. Способ микробиологического синтеза гидролазы эфиров альфа-аминокислот, не модифицированной на N-конце полигистидиновой последовательностью, предусматривающий культивирование бактерий, содержащих ген aehR из Xanthomonas rubrilineans ВКПМ В-9915, в подходящей питательной среде, отличающийся тем, что в качестве продуцента используют рекомбинантный штамм по п.1, в состав среды включают канамицин, а экспрессию целевого продукта осуществляют добавлением изопропил-β-D-тиогалактозида в качестве индуктора.
| ПЕРИСТАЛЬТИЧЕСКИЙ НАСОС | 1995 |
|
RU2086111C1 |
| YE LJ Changing the specificity of alfa-amino acid ester hydrolase toward para-hydroxyl cephalosporins synthesis by site-directed saturation mutagenesis | |||
| Biotechnol Lett | |||
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| CN 102653726 A, 05.09.2012 | |||
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОГЕННОГО БИОКАТАЛИЗАТОРА, БИОКАТАЛИЗАТОР НА ОСНОВЕ ГИДРОЛАЗЫ ЭФИРОВ АЛЬФА-АМИНОКИСЛОТ И СПОСОБ СИНТЕЗА АМИНОБЕТА-ЛАКТАМНОГО АНТИБИОТИКА ПОД ДЕЙСТВИЕМ ЭТОГО БИОКАТАЛИЗАТОРА | 2008 |
|
RU2381273C2 |

