Перекрестная ссылка на родственные заявки
Данная заявка притязает согласно разделу 35 Кодекса законов США на приоритет предварительной заявки на патент США № 60/772614, озаглавленной “Modified Gene Silencing”, которая была подана 13 февраля 2006 г.; предварительной заявки на патент США № 60/781519, озаглавленной “Soybean Seed and Oil Compositions and Method for Making Same”, которая была подана 10 марта 2006 г.; и заявки на патент США № 11/376328, озаглавленной “Nucleic Acid Constructs and Methods for Producing Altered Seed Oil Compositions”, которая была подана 16 марта 2006 г.
Включение списка последовательностей
В настоящее описание изобретения в качестве ссылки включена бумажная копия списка последовательностей и компьютерная форма списка последовательностей на дискете, содержащей файл с именем “Omni2 AS FILED.txt” размером 60690 байт (измерен в MS-DOS), который был записан 25 сентября 2003 г. и подан в заявке на патент США № 10/669888. В настоящее описание изобретения в качестве ссылки включена бумажная копия списка последовательностей и компьютерная форма списка последовательностей на дискете, содержащей файл с именем “OmniChild.txt” размером 61434 байт (измерен в MS-DOS), который был записан 15 марта 2006 г.
Область техники, к которой относится изобретение
Настоящее изобретение относится к молекулам рекомбинантной нуклеиновой кислоты, конструкциям и другим агентам, ассоциированным с координированной манипуляцией многими генами в пути синтеза жирных кислот. В частности, агенты по настоящему изобретению ассоциированы с одновременным повышением экспрессии определенных генов в пути синтеза жирных кислот и подавлением экспрессии других генов в том же пути. Настоящее изобретение относится также к растениям, содержащим такие агенты, и, в частности, к растениям, содержащим такие конструкции, которые характеризуются измененными составами масла из семян.
Уровень техники
Растительные масла применяются в разных областях. Необходимы новые составы растительных масел и более совершенные методы получения составов масла из биосинтезированных или природных растительных источников. В зависимости от предполагаемого применения масла желательны разные составы жирных кислот. Растения, в частности виды, синтезирующие большое количество масла в семенах, являются важным источником масел как для пищевого, так и для промышленного применения. Масла из семян почти полностью состоят из триацилглицеринов, в которых жирные кислоты этерифицированы в три гидроксильные группы глицерина.
Соевое масло обычно содержит примерно 16-20% насыщенных жирных кислот: 13-16% пальмитата и 3-4% стеарата. См. публикацию Gunstone et al., The Lipid Handbook, Chapman & Hall, London (1994). Соевые масла были модифицированы разными методами селекции в соответствии с требованиями конкретных рынков сбыта. Однако до сих пор не существует соевого масла, которое отвечало бы требованиям основных потребителей, таких как пищевая промышленность, где используется масло для заправки салатов, масло для выпечки и масло для жарения, и промышленные рынки, где соевое масло служит для производства биологического дизельного топлива и биологических смазочных масел. Ранее использовавшиеся соевые масла были или слишком дороги, или у них отсутствовало важное свойство пищевого качества, такое как устойчивость к окислению, хороший вкус жареной пищи или содержание насыщенных жиров, либо важное свойство биологического топлива, такое как требуемое выделение оксида азота, стойкость в холодном состоянии или текучесть на холоде.
Высшие растения синтезируют жирные кислоты в соответствии с обычным путем метаболизма, каким является путь жирная кислота-синтетаза (FAS), локализованный в пластидах. β-Кетоацил-АСР-синтазы являются важными ограничивающими синтез ферментами в FAS растительных клеток и существуют в нескольких вариантах. β-Кетоацил-АСР-синтаза I катализирует удлинение цепи до образования пальмитоил-АСР (С16:0), а то время как β-кетоацил-АСР-синтаза II катализирует удлинение цепи до образования стеароил-АСР (С18:0). β-Кетоацил-АСР-синтаза IV является вариантом β-кетоацил-АСР-синтазы II и может также катализировать удлинение цепи до 18:0-АСР. В сое основными продуктами FAS являются 16:0-АСР и 18:0-АСР. Десатурация 18:0-АСР с образованием 18:1-АСР катализируется локализованной в пластидах растворимой дельта-9-десатуразой (именуемой также “стеароил-АСР-десатураза”). См. публикацию Voelker et al., 52 Annu. Rev. Plant Physiol. Plant Mol. Biol. 335-61 (2001).
Продукты FAS и дельта-9-десатуразы в пластидах, 16:0-АСР, 18:-АСР и 18:1-АСР, гидролизуются специфичными тиоэстеразами (FAT). Растительные тиоэстеразы могут быть отнесены к двум семействам генов на основании гомологии последовательностей и предпочтения субстрата. Первое семейство, FATA, включает ацил-АСР-тиоэстеразы с длинными цепями, обладающие главным образом активностью 18:1-АСР. Ферменты второго семейства, FATB, обычно используют 16:0-АСР (пальмитоил-АСР), 18:0-АСР (стеароил-АСР) и 18:1-АСР (олеоил-АСР). Такие тиоэстеразы играют важную роль в определении длины цепи в процессе биосинтеза новых жирных кислот в растениях, поэтому указанные ферменты могут быть использованы для разных модификаций составов жирных кислот, в частности, для определения относительных соотношений групп разных жирных кислот, присутствующих в маслах из семян.
Продукты реакций FATA и FATB, свободные жирные кислоты, выводятся из пластид и превращаются в соответствующие сложные эфиры ацил-СоА. Ацил-СоА представляют собой субстраты для пути биосинтеза липидов (путь Кеннеди), который локализован в эндоплазматической сети (ER). Данный путь отвечает за образование мембранных липидов, а также за биосинтез триацилглицеринов, образующих масло из семян. В эндоплазматической сети находятся дополнительные мембранообразующие десатуразы, которые могут далее десатурировать 18:1 в полиненасыщенные жирные кислоты. Дельта-12 десатураза (FAD2) катализирует введение двойной связи в 18:1 с образованием линолевой кислоты (18:2). Дельта-15 десатураза (FAD3) катализирует введение двойной связи в 18:2 с образованием линоленовой кислоты (18:3).
Многие комплексные биохимические пути в настоящее время поддаются генетической манипуляции обычно путем подавления или сверхэкспрессии отдельных генов. Дальнейшее использование возможности генетической манипуляции растениями требует координированной манипуляции многими генами в пути. Были использованы разные методы объединения трансгенов в одном растении, которые включают скрещивание, ретрансформацию, котрансформацию и применение связанных трансгенов. Для координированного подавления многих эндогенных генов растения может быть использован химерный трансген со связанными неполными последовательностями генов. Конструкции, созданные на основе вирусных полипротеинов, могут быть использованы для одновременного введения нескольких кодирующих генов в растительные клетки. См. публикацию Halpin et al., Plant Mol. Biol. 47:295-310 (2001).
Таким образом, желаемый фенотип растения может потребовать экспрессии одного или нескольких генов и одновременного снижения экспрессии другого гена или генов. Поэтому существует потребность в одновременной сверхэкспрессии одного или нескольких генов и подавлении или снижении экспрессии другого гена или генов в растениях с использованием одной трансгенной конструкции.
Сущность изобретения
Настоящее изобретение относится к одной или нескольким молекулам рекомбинантной нуклеиновой кислоты, которые при введении в клетку или организм способны подавлять, по крайней мере частично снижать, сокращать, значительно сокращать или эффективно устранять экспрессию по крайней мере одной или нескольких эндогенных РНК FAD2, FAD3 или FATB и в то же время коэкспрессировать, одновременно экспрессировать или координированно продуцировать одну или несколько РНК или белков, транскрибированных из гена, кодирующего бета-кетоацил-АСР-синтазу I, бета-кетоацил-АСР-синтазу IV, дельта-9-десатуразу или СР4 EPSPS. Настоящее изобретение относится также к растительным клеткам и растениям, трансформированным одной или несколькими молекулами нуклеиновой кислоты, к семенам, маслу и другим продуктам, получаемым из трансформированных растений.
Настоящее изобретение относится также к молекуле рекомбинантной нуклеиновой кислоты, содержащей первую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна подавлять эндогенную экспрессию по крайней мере одного, предпочтительно двух генов, выбираемых из группы, включающей гены FAD2, FAD3 и FATB; и вторую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна повышать эндогенную экспрессию по крайней мере одного гена, выбираемого из группы, включающей ген бета-кетоацил-АСР-синтазы I, ген бета-кетоацил-АСР-синтазы IV, ген дельта-9-десатуразы и СР4 EPSPS.
Настоящее изобретение далее относится к молекуле рекомбинантной нуклеиновой кислоты, содержащей первую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна образовывать конструкцию дцРНК и подавлять эндогенную экспрессию по крайней мере одного, предпочтительно двух генов, выбираемых из группы, включающей гены FAD2, FAD3 и FATB, при этом первая совокупность последовательностей ДНК включает первую некодирующую последовательность, экспрессирующую первую последовательность РНК, которая по крайней мере на 90% идентична некодирующей области гена FAD2, первую антисмысловую последовательность, экспрессирующую первую антисмысловую последовательность РНК, способную образовывать молекулу двухцепочечной РНК с первой последовательностью РНК, вторую некодирующую последовательность, экспрессирующую вторую последовательность РНК, которая по крайней мере на 90% идентична некодирующей области гена FATB, и вторую антисмысловую последовательность, экспрессирующую вторую антисмысловую последовательность РНК, способную образовывать молекулу двухцепочечной РНК со второй последовательностью РНК; и вторую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способа повышать эндогенную экспрессию по крайней мере одного гена, выбираемого из группы, включающей ген бета-кетоацил-АСР-синтазы I, ген бета-кетоацил-АСР-синтазы IV, ген дельта-9-десатуразы и СР4 EPSPS.
Настоящее изобретение относится к способам трансформации растений при помощи указанных молекул рекомбинантной нуклеиновой кислоты. Данные способы включают способ создания трансформированного растения с повышенным содержанием олеиновой кислоты, пониженным содержанием насыщенных жирных кислот и пониженным содержанием полиненасыщенных жирных кислот в семени, который включает (А) трансформацию растительной клетки молекулой рекомбинантной нуклеиновой кислоты, содержащей первую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна подавлять эндогенную экспрессию по крайней мере одного, предпочтительно двух генов, выбираемых из группы, включающей гены FAD2, FAD3 и FATB, и вторую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна повышать эндогенную экспрессию по крайней мере одного гена, выбираемого из группы, включающей ген бета-кетоацил-АСР-синтазы I, ген бета-кетоацил-АСР-синтазы IV, ген дельта-9-десатуразы и СР4 EPSPS; и (В) выращивание трансформированного растения, образующего семя с повышенным содержанием олеиновой кислоты, пониженным содержанием насыщенных жирных кислот и пониженным содержанием полиненасыщенных жирных кислот по сравнению с семенем растения, имеющего подобную генетическую среду, но не содержащего молекулу рекомбинантной нуклеиновой кислоты.
Настоящее изобретение далее относится к способам трансформации растительных клеток молекулами рекомбинантной нуклеиновой кислоты. Указанные способы включают способ изменения состава масла растительной клетки, который включает (А) трансформацию растительной клетки молекулой рекомбинантной нуклеиновой кислоты, включающей первую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна подавлять эндогенную экспрессию по крайней мере одного, предпочтительно двух генов, выбираемых из группы, включающей гены FAD2, FAD3 и FATB, и вторую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна повышать эндогенную экспрессию по крайней мере одного гена, выбираемого из группы, включающей ген бета-кетоацил-АСР-синтазы I, гена бета-кетоацил-АСР-синтазы IV, ген дельта-9-десатуразы и СР4 EPSPS; и (В) выращивание растительной клетки в условиях инициации транскрипции первой совокупности последовательностей ДНК и второй совокупности последовательностей ДНК, в результате чего изменяется состав масла по сравнению с растительной клеткой, имеющей подобную генетическую среду, но не содержащей молекулу рекомбинантной нуклеиновой кислоты.
Настоящее изобретение относится также к трансформированному растению, содержащему молекулу рекомбинантной нуклеиновой кислоты, включающую первую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна подавлять эндогенную экспрессию по крайней мере одного, предпочтительно двух генов, выбираемых из группы, включающей гены FAD2, FAD3 и FATB, и вторую совокупность последовательностей ДНК, которая при экспрессии в клетке-хозяине способна повышать эндогенную экспрессию по крайней мере одного гена, выбираемого из группы, включающей ген бета-кетоацил-АСР-синтазы I, ген бета-кетоацил-АСР-синтазы IV, ген дельта-9-десатуразы и СА4 EPSPS. Настоящее изобретение далее относится к трансформированному растению сои с семенем, в состав масла которого входит 55-80 мас.% олеиновой кислоты, 10-40 мас.% линолевой кислоты, 6 мас.% или меньше линоленовой кислоты и 2-8 мас.% насыщенных жирных кислот, к кормовому продукту, частям растения и семени, полученным из данного растения. Другой вариант осуществления настоящего изобретения относится к трансформированному растению сои с семенем, в состав масла которого входит около 65-80% олеиновой кислоты, около 3-8% насыщенных жирных кислот и около 12-32% полиненасыщенных жирных кислот. Настоящее изобретение относится также к кормовому продукту, частям растения и семени, полученным из такого растения. Другой вариант осуществления настоящего изобретения относится к трансформированному растению сои с семенем, в состав масла которого входит около 65-80% олеиновой кислоты, около 2-3,5% насыщенных жирных кислот и около 16,5-33% полиненасыщенных жирных кислот. Настоящее изобретение относится также к кормовому продукту, частям растения и семени, полученным из такого растения.
Настоящее изобретение относится к семени сои, в состав масла которого входит 55-80 мас.% олеиновой кислоты, 10-40 мас.% линолевой кислоты, 6 мас.% или меньше линоленовой кислоты и 2-8 мас.% насыщенных жирных кислот, а также относится к семени сои, в состав масла которого входит 65-80 мас.% олеиновой кислоты, 10-30 мас.% линолевой кислоты, 6 мас.% или меньше линоленовой кислоты и 2-8 мас.% насыщенных жирных кислот. Другой вариант осуществления настоящего изобретения относится к семени сои, в состав масла которого входит около 65-80% олеиновой кислоты, около 3-8% насыщенных жирных кислот и около 12-32% полиненасыщенных жирных кислот. Другой вариант осуществления настоящего изобретения относится к семени сои, в состав масла которого входит около 65-80% олеиновой кислоты, около 2-3,5% насыщенных жирных кислот и около 16,5-33% полиненасыщенных жирных кислот.
Настоящее изобретение относится также к продуктам сои, в состав масла которых входит 69-73 мас.% олеиновой кислоты, 21-24 мас.% линолевой кислоты, 0,5-3 мас.% линолевой кислоты и 2-3 мас.% насыщенных жирных кислот.
Неочищенное соевое масло по настоящему изобретению имеет состав, включающий 55-80 мас.% олеиновой кислоты, 10-40 мас.% линолевой кислоты, 6 мас.% или меньше линоленовой кислоты и 2-8 мас.% насыщенных жирных кислот. Другое неочищенное соевое масло по настоящему изобретению имеет состав, включающий 65-80 мас.% олеиновой кислоты, 10-30 мас.% линолевой кислоты, 6 мас.% или меньше линоленовой кислоты и 2-8 мас.% насыщенных жирных кислот. В другом варианте осуществления изобретения неочищенное соевое масло по настоящему изобретению имеет состав, включающий около 65-80% олеиновой кислоты, около 3-8% насыщенных жирных кислот и около 12-32% полиненасыщенных жирных кислот. В другом варианте осуществления изобретения неочищенное соевое масло по настоящему изобретению имеет состав, включающий около 65-80% олеиновой кислоты, около 2-3,5% насыщенных жирных кислот и около 16,5-33% полиненасыщенных жирных кислот.
Настоящее изобретение относится также к семени сои, в состав масла которого входит от около 42 мас.% до около 85 мас.% олеиновой кислоты и от около 8 мас.% до около 1,5 мас.% насыщенных жирных кислот. В другом варианте осуществления изобретения семя сои по настоящему изобретению имеет состав масла, включающий от около 42 мас.% до около 85 мас.% олеиновой кислоты, от около 8 мас.% до около 1,5 мас.% насыщенных жирных кислот, менее 35 мас.% линоленовой кислоты, при этом объединенное количество олеиновой кислоты и линоленовой кислоты составляет от около 65 мас.% до около 90 мас.% от общего состава масла; и указанное семя содержит в клетке-хозяине молекулу рекомбинантной нуклеиновой кислоты с последовательностью ДНК, включающей фрагмент интрона FAD2-1 длиной от около 50 до около 400 последовательных нуклеотидов, 3'-UTR FATB и 5'-UTR FATB, гетерологичную бета-кетоацил-АСР-синтазу IV и гетерологичную дельта-9-десатуразу.
Семя сои по настоящему изобретению может иметь состав масла, включающий от около 50 мас.% до около 80 мас.% олеиновой кислоты, от около 8 мас.% до около 1,5 мас.% насыщенных жирных кислот, от около 2 мас.% до около 45 мас.% линолевой кислоты, от около 4 мас.% до около 14 мас.% линоленовой кислоты, при этом объединенное количество олеиновой кислоты и линоленовой кислоты составляет от около 65 мас.% до около 90 мас.% от общего состава масла, и указанное семя содержит молекулу рекомбинантной нуклеиновой кислоты с последовательностью ДНК, включающей фрагмент интрона FАD2-1 длиной от около 50 до около 400 последовательных нуклеотидов, кодирующую область СТР FATB и 42 последовательных нуклеотида 5'-UTR FATB. В другом варианте осуществления изобретения семя сои может содержать молекулу рекомбинантной нуклеиновой кислоты с последовательностью ДНК, подавляющей эндогенную экспрессию FAD2 и FATB, при этом указанное семя имеет состав масла, включающий 46-75 мас.% олеиновой кислоты, 1,5-8,5 мас.% насыщенных жирных кислот, 2,5-38 мас.% линолевой кислоты и 4,5-17,5 мас.% линоленовой кислоты.
Настоящее изобретение относится также к способу уменьшения величины подавления гена FAD2 по сравнению с величиной подавления гена FAD2, достигаемой в результате экспрессии конструкции дцРНКи, содержащей рекомбинантную последовательность FAD2, включающую весь интрон FAD2 или всю UTR FАD2, путем: i) экспрессии рекомбинантной последовательности FAD2 в растительной клетке, при этом рекомбинантная последовательность FAD2, выделяемая из эндогенного гена FAD2 в растительной клетке, содержит фрагмент интрона FAD2 или фрагмент UTR FAD2, и ii) подавления эндогенного гена FAD2 рекомбинантной последовательностью FAD2, при этом величина подавления гена FAD2 меньше величины экспрессии гена, достигаемой в результате экспрессии конструкции дцРНКи, содержащей рекомбинантную последовательность FAD2, включающую весь интрон FAD2 или всю UTR FAD2.
Настоящее изобретение относится также к способам изменения состава масла в растительной клетке путем: трансформации растительной клетки рекомбинантной последовательностью FAD2, выделенной из части эндогенного гена FAD2, которая включает фрагмент интрона FAD2 или фрагмент UTR FAD2, и выращивания растительной клетки в условиях инициации транскрипции рекомбинантной последовательности FAD2, в результате чего состав масла изменяется по сравнению с растительной клеткой, имеющей подобную генетическую среду, но не содержащей рекомбинантную последовательность FAD2. Другой вариант осуществления изобретения относится к способу повышения содержания олеиновой кислоты и уменьшения содержания насыщенных жирных кислот в семени растения путем: i) укорачивания длины первой рекомбинантной последовательности FAD2 по крайней мере до частичного уменьшения величины подавления гена FAD2 в растении, трансформированном первой рекомбинантной последовательностью FAD2, по сравнению с величиной подавления гена FAD2 в растительной клетке, имеющей подобную генетическую среду и вторую рекомбинантную последовательность FAD2, которая состоит из более эндогенной последовательности FAD2, чем первая рекомбинантная последовательность FAD2; ii) экспрессии рекомбинантной последовательности FATB, способной по крайней мере частично уменьшать экспрессию гена FATB в растительной клетке по сравнению с подавлением FATB в растительной клетке, имеющей подобную генетическую среду, но не содержащей рекомбинантную последовательность FATB; iii) выращивания растения с молекулой рекомбинантной нуклеиновой кислоты, содержащей первую рекомбинантную последовательность FAD2 и рекомбинантную последовательность FATB, и iv) культивирования растения, образующего семя с меньшим содержанием насыщенных жирных кислот по сравнению с семенем растения, имеющего подобную генетическую среду, но не содержащего первую рекомбинантную последовательность FAD2 и рекомбинантную последовательность FATB.
Другой вариант осуществления настоящего изобретения относится к способу получения трансформированного растения, семя которого характеризуется меньшим содержанием насыщенных жирных кислот, путем: трансформации растительной клетки молекулой рекомбинантной нуклеиновой кислоты, содержащей последовательность рекомбинантной ДНК, подавляющей эндогенную экспрессию FAD2 и FATB, которая включает последовательность нуклеиновой кислоты рекомбинантного гена FAD2 и рекомбинантного гена FATB, при этом последовательность FAD2 содержит не всю последовательность интрона FAD2; и выращивания трансформированного растения, образующего семя с пониженным содержанием насыщенных жирных кислот, по сравнению с семенем растения, имеющего подобную генетическую среду, но не содержащего последовательность рекомбинантной ДНК.
Другой вариант осуществления настоящего изобретения относится к способу модуляции состава жирных кислот в масле из семян масличной культуры умеренного пояса путем выделения генетического элемента длиной, равной по крайней мере 40 нуклеотидам, способного подавлять экспрессию эндогенного гена в пути синтеза жирных кислот; создания нескольких укороченных фрагментов генетического элемента; введения всех укороченных фрагментов в растительную клетку масличной культуры умеренного пояса с целью создания трансгенного растения и отбора трансгенного растения, содержащего укороченный фрагмент определенной длины и последовательность, обеспечивающую требуемое изменение в составе жирных кислот масла из семян.
Настоящее изобретение относится также к семени сои, состав масла которого характеризуется значительно меньшим содержанием насыщенных жирных кислот и умеренно повышенным содержанием олеиновой кислоты и которое содержит последовательность ДНК, подавляющую эндогенную экспрессию FAD2 в растительной клетке, при этом последовательность ДНК имеет рекомбинантную последовательность FAD2, включающую фрагмент интрона FAD2. Другой вариант осуществления настоящего изобретения относится к молекуле нуклеиновой кислоты, содержащей последовательность интрона FAD2-1A длиной от около 60 до около 320 последовательных нуклеотидов. Альтернативный вариант осуществления настоящего изобретения относится также к семени сои, содержащему первую последовательность рекомбинантной ДНК, которая подавляет экспрессию эндогенного гена FAD2-1 сои, включающего интрон FAD2-1 сои, и вторую последовательность рекомбинантной ДНК, которая экспрессирует повышенные уровни гена, выбираемого из группы, включающей ген KAS I, дельта-9-десатуразы, KAS IV и их комбинации.
Настоящее изобретение относится также к растительной клетке семени сои, характеризующегося составом жирных кислот масла из семян, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет менее 8 мас.% от общего содержания жирных кислот. Настоящее изобретение относится также к растительной клетке семени сои, характеризующегося составом масла, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот и содержание линоленовой кислоты составляет менее примерно 3 мас.% от общего содержания жирных кислот.
Настоящее изобретение относится также к молекуле нуклеиновой кичслоты с последовательностью интрона FAD2-1A длиной от около 60 до около 320 последовательных нуклеотидов. В объем настоящего изобретения входит также конструкция рекомбинантной ДНК, включающая фрагмент интрона FAD2-1 сои длиной от около 20 до около 420 последовательных нуклеотидов и фрагмент гена FATB сои длиной от около 40 до около 450 последовательных нуклеотидов. Другой вариант осуществления изобретения относится к молекуле рекомбинантной нуклеиновой кислоты, содержащей первую последовательность ДНК, подавляющую эндогенную экспрессию FAD2-1 и FATB сои, которая включает фрагмент интрона FAD2-1 длиной от около 20 до около 420 последовательных нуклеотидов, 3'-UTR FATB сои, 5'-UTR FATB сои или кодирующую область СТР, и вторую последовательность рекомбинантной ДНК, которая повышает экспрессию по крайней мере одного из генов, выбираемых из группы, включающей бета-кетоацил-АСР-синтазу IV и дельта-9-десатуразу.
Настоящее изобретение относится также к несмешанному соевому маслу, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет от около 1,5 мас.% до около 8 мас.% от общего содержания жирных кислот; к несмешанному соевому маслу, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет около 8 мас.% или меньше от общего содержания жирных кислот; к несмешанному соевому маслу, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот и содержание линоленовой кислоты составляет менее 3 мас.% от общего содержания жирных кислот; и к несмешанному соевому маслу, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот, содержание насыщенных жирных кислот составляет около 8 мас.% или меньше от общего содержания жирных кислот и содержание линоленовой кислоты составляет около 1,5 мас.% или меньше от общего содержания жирных кислот.
Настоящее изобретение относится также к соевой муке, получаемой из семени сои, характеризующегося составом жирных кислот в масле из семян, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет менее 8 мас.% от общего содержания жирных кислот. В объем настоящего изобретения входит также соевая мука, получаемая из семени сои, характеризующегося составом жирных кислот в масле из семян, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот и содержание линоленовой кислоты составляет менее примерно 3 мас.% от общего содержания жирных кислот.
Настоящее изобретение относится также к способу уменьшения величины подавления гена FAD2 по сравнению с величиной подавления гена FAD2, достигаемой в результате экспрессии конструкции дцРНКи, содержащей гетерологичную последовательность FAD2, включающую весь интрон FAD2 или всю UTR FAD2, путем: i) экспрессии гетерологичной последовательности FAD2 в растительной клетке, при этом гетерологичная последовательность FAD2, выделяемая из эндогенного гена FAD2 растительной клетки, состоит из фрагмента интрона FAD2 или фрагмента UTR FAD2, и ii) подавления эндогенного гена FAD2 гетерологичной последовательностью FAD2, при этом величина подавления гена FAD2 меньше величины экспрессии гена, достигаемой в результате экспрессии гетерологичной последовательности FAD2, включающей весь интрон FAD2 или всю UTR FAD2.
Настоящее изобретение относится также к способу изменения состава масла растительной клетки путем трансформации растительной клетки гетерологичной последовательностью FAD2, выделенной из части эндогенного гена FAD2, которая включает фрагмент интрона FAD2 или фрагмент UTR FAD2, и выращивания растительной клетки в условиях инициации транскрипции гетерологичной последовательности FAD2, в результате чего происходит изменение состава масла по сравнению с растительной клеткой, имеющей подобную генетическую среду, но не содержащей гетерологичную последовательность FAD2.
Настоящее изобретение относится также к способу увеличения содержания олеиновой кислоты и уменьшения содержания насыщенных жирных кислот в семени растения, который включает: i) укорачивание длины первой гетерологичной последовательности FAD2 по крайней мере до частичного уменьшения величины подавления гена FAD2 в растении, трансформированном первой гетерологичной последовательностью FAD2, по сравнению с величиной подавления гена FAD2 в растительной клетке, имеющей подобную генетическую среду и вторую гетерологичную последовательность FAD2, которая состоит из более эндогенной последовательности FAD2, чем первая гетерологичная последовательность FAD2, ii) экспрессию гетерологичной последовательности FATB, способной по крайней мере частично уменьшать экспрессию гена FATB в растительной клетке по сравнению с подавлением FATB в растительной клетке, имеющей подобную генетическую среду, но не содержащей гетерологичную последовательность FATB, iii) выращивание растения, включающего геном с первой гетерологичной последовательностью FAD2 и гетерологичной последовательностью FATB, и iv) культивирование растения, образующего семя с пониженным содержанием насыщенных жирных кислот по сравнению с семенем растения, имеющего подобную генетическую среду, но не содержащего первую гетерологичную последовательность FAD2 и гетерологичную последовательность FATB.
Настоящее изобретение относится также к способу модуляции состава жирных кислот в масле из семян масличной культуры умеренного пояса, выделения фрагмента генетического элемента длиной, равной по крайней мере 40 нуклеотидам, способного подавлять экспрессию эндогенного гена в пути синтеза жирных кислот; введения указанного генетического элемента в растительную клетку масличной культуры умеренного пояса; создания трансгенного растения и отбора семени трансгенного растения, содержащего указанный генетический элемент, модулирующий состав жирных кислот в масле из семян.
Другой вариант осуществления настоящего изобретения относится к клетке семени сои, характеризующегося составом жирных кислот в масле из семян, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет менее 8 мас.% от общего содержания жирных кислот.
Настоящее изобретение относится также к молекуле гетерологичной нуклеиновой кислоты, включающей фрагмент интрона FAD2-1 сои длиной от около 20 до около 420 последовательных нуклеотидов и фрагмент гена FATB сои длиной от около 40 до около 450 последовательных нуклеотидов. Другой вариант осуществления настоящего изобретения относится к молекуле гетерологичной нуклеиновой кислоты, включающей последовательность нуклеиновой кислоты, содержащей фрагмент интрона FAD2-1 сои длиной от около 20 до около 420 нуклеотидов, фрагмент гена FATB сои длиной от около 40 до около 450 нуклеотидов и последовательность нуклеиновой кислоты, которая повышает экспрессию бета-кетоацил-АСР-синтазы IV или дельта-9-десатуразы либо обеих вместе.
Настоящее изобретение относится также к способу уменьшения содержания линоленовой кислоты в семени сои путем: i) введения в клетку сои молекулы гетерологичной нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты по крайней мере из двух членов семейства генов FАD3; ii) экспрессии последовательности нуклеиновой кислоты из гена FAD2, способной по крайней мере частично уменьшать экспрессию эндогенного гена FAD3 в растительной клетке; iii) выращивания растительной клетки, включающей геном с последовательностью нуклеиновой кислоты по крайней мере из двух членов семейства генов FAD3; и iv) культивирования указанной растительной клетки с уменьшенным содержанием линоленовой кислоты по сравнению с растительной клеткой, имеющей подобную генетическую среду, но не содержащей по крайней мере два члена семейства генов FAD3. Настоящее изобретение относится также к конструкции рекомбинантной ДНК с фрагментами ДНК по крайней мере из двух членов семейства генов FAD3.
Настоящее изобретение относится также к несмешанному соевому маслу, характеризующемуся составом жирных кислот, в котором содержание олеиновой кислоты составляет от около 42 мас.% до около 85 мас.% от общего содержания жирных кислот, содержание насыщенных жирных кислот составляет около 8 мас.% или меньше от общего содержания жирных кислот и содержание линоленовой кислоты составляет около 1,5 мас.% или меньше от общего содержания жирных кислот.
Краткое описание фигур
На фиг.1-4 изображены типичные конфигурации молекулы нуклеиновой кислоты.
На фиг.5(а)-(d) и 6(а)-(с) изображены иллюстративные конфигурации первой совокупности последовательностей ДНК.
На фиг.7-20 изображены молекулы нуклеиновой кислоты по настоящему изобретению.
На фиг.21 изображена конструкция pMON68537.
На фиг.22 изображена конструкция pMON68539.
Подробное описание изобретения
Описание последовательностей нуклеиновых кислот
SEQ ID NO:1 является последовательностью нуклеиновой кислоты интрона 1 FAD2-1A.
SEQ ID NO:2 является последовательностью нуклеиновой кислоты интрона 1 FAD2-1B.
SEQ ID NO:3 является последовательностью нуклеиновой кислоты промотора FAD2-1B.
SEQ ID NO:4 является последовательностью нуклеиновой кислоты геномного клона FAD2-1A.
SEQ ID NO:5 и 6 являются последовательностями нуклеиновой кислоты соответственно 3'-UTR и 5'-UTR FAD2-1A.
SEQ ID NO:7-13 являются последовательностями нуклеиновой кислоты соответственно интронов 1, 2, 3А, 4, 5, 3В и 3С FAD3-1A.
SEQ ID NO:14 является последовательностью нуклеиновой кислоты интрона 4 FAD3-1C.
SEQ ID NO:15 является последовательностью нуклеиновой кислоты неполного геномного клона FAD3-1A.
SEQ ID NO:16 и 17 являются последовательностями нуклеиновой кислоты соответственно 3'-UTR и 5'-UTR FAD3-1A.
SEQ ID NO:18 является последовательностью нуклеиновой кислоты неполного геномного клона FAD3-1B.
SEQ ID NO:19-25 являются последовательностями нуклеиновой кислоты соответственно интронов 1, 2, 3А, 3В, 3С, 4 и 5 FAD3-1B.
SEQ ID NO:26 и 27 являются последовательностями нуклеиновой кислоты соответственно 3'-UTR и 5'-UTR FAD3-1B.
SEQ ID NO:28 является последовательностью нуклеиновой кислоты геномного клона FATB-1.
SEQ ID NO:29-35 являются последовательностями нуклеиновой кислоты соответственно интронов I, II, III, IV, V, VI и VII FATB-1.
SEQ ID NO:36 и 37 являются последовательностями нуклеиновой кислоты соответственно 3'-UTR и 5'-UTR FATB-1.
SEQ ID NO:38 является последовательностью нуклеиновой кислоты гена KAS I Cuphea pulcherrima.
SEQ ID NO:39 является последовательностью нуклеиновой кислоты гена KAS IV Cuphea pulcherrima.
SEQ ID NO:40 и 41 являются последовательностями нуклеиновой кислоты генов дельта-9-десатуразы соответственно Ricinus communis и Simmondsia chinensis.
SEQ ID NO:42 является последовательностью нуклеиновой кислоты кДНК FATB-2.
SEQ ID NO:43 является последовательностью нуклеиновой кислоты геномного клона FATB-2.
SEQ ID NO:44-47 являются последовательностями нуклеиновой кислоты соответственно интронов I, II, III и IV FATB-2.
SEQ ID NO:48-60 являются последовательностями нуклеиновой кислоты праймеров для полимеразной цепной реакции (ПЦР).
SEQ ID NO:61 и 62 являются последовательностями нуклеиновой кислоты соответственно 3'-UTR и 5'-UTR FAD3-1 сои.
Определения терминов
“АСР” означает ацилпереносящий белок. “Измененный состав масла из семян” означает состав масла из семян трансгенного или трансформированного растения по настоящему изобретению с измененными или модифицированными уровнями жирных кислот по сравнению с маслом из семян растения, имеющего сходную генетическую среду, но не подвергшегося трансформации.
“Подавление антисмысловой последовательностью” означает генспецифичный сайленсинг, индуцированный введением молекулы антисмысловой РНК.
“Коэкспрессия более чем одного агента, такого как мРНК или белок” означает одновременную экспрессию агента в перекрывающихся временных рамках и в одной и той же клетке или ткани вместе с другим агентом. “Координированная экспрессия более чем одного агента” означает коэкспрессию нескольких агентов, которые продуцируют транскрипты и белки при использовании общего или идентичного промотора.
“Комплемент” последовательности нуклеиновой кислоты означает комплемент последовательности на всем протяжении ее длины.
“Косупрессия” означает снижение уровней экспрессии, обычно на уровне РНК, конкретного эндогенного гена или семейства генов в результате экспрессии гомологичной смысловой конструкции, способной транскрибировать мРНК с таким же числом цепей, что и транскрипт эндогенного гена. Napoli et al., Plant Cell 2:279-289 (1990); van der Krol et al., Plant Cell 2:291-299 (1990).
“Неочищенное соевое масло” означает соевое масло, экстрагированное из семян сои, которое не было очищено, обработано или смешано, хотя из такого масла могут быть удалены смолистые вещества.
“СТР” означает транзитный пептид хлоропласта, кодированный “кодирующей последовательностью транзитного пептида хлоропласта”.
Применительно к белкам и нуклеиновым кислотам термин “выделенный” означает прямое (например, определение последовательности известного белка или нуклеиновой кислоты и создание белка или нуклеиновой кислоты с последовательностью, подобной по крайней мере частично последовательности известного белка или нуклеиновой кислоты) или непрямое (например, получение белка или нуклеиновой кислоты из организма, родственного известному белку или нуклеиновой кислоте) получение белка или нуклеиновой кислоты из известного белка или нуклеиновой кислоты. Специалисту в данной области должны быть известны другие методы “выделения” белка или нуклеиновой кислоты из известного белка или нуклеиновой кислоты.
Двухцепочечная РНК (“дцРНК”), интерферирующая двухцепочечная РНК (“дцРНКи”) и интерферирующая РНК (“РНКи”) служат для определения генспецифического сайленсинга, вызванного введением конструкции, способной транскрибировать молекулу по крайней мере частично двухцепочечной РНК. “Молекула дцРНК” и “молекула РНКи” означают область молекулы РНК, содержащую сегменты с комплементарными нуклеотидными последовательностями, которые могут гибридизироваться друг с другом и образовывать двухцепочечную РНК. Такие молекулы двухцепочечной РНК при введении в клетку или организм способны по крайней мере частично уменьшать уровень мРНК, присутствующей в клетке или клетке организма. Кроме того, дцРНК может быть создана в результате сборки in vivo соответствующих фрагментов ДНК путем незаконной рекомбинации и сайт-специфической рекомбинации, описанной в международной заявке на патент № РСТ/US2005/004681, поданной 11 февраля 2005 г., которая полностью включена в настоящее описание изобретения в виде ссылки.
“Экзон” означает сегмент молекул нуклеиновой кислоты, обычно ДНК, который кодирует часть или весь экспрессированный белок.
“Жирная кислота” означает свободные жирные кислоты и ацильные группы жирных кислот.
“Ген” означает последовательность нуклеиновой кислоты, которая включает 5'-концевую промоторную область, ассоциированную с экспрессией генного продукта, любые области интрона и экзона, 3'- или 5'-концевые нетранслируемые области, ассоциированные с экспрессией генного продукта.
“Сайленсинг гена” означает подавление экспрессии гена или снижение экспрессии гена.
“Семейство генов” означает два или более генов в организме, кодирующих белки, обладающие подобными функциональными свойствами, и “член семейства генов” означает любой ген семейства генов, обнаруженный в генетическом материале растения, например “член семейства генов FAD2” является любым геном FAD2, обнаруженным в генетическом материале растения. Примером двух членов семейства генов являются FAD2-1 и FAD2-2. Семейство генов может быть дополнительно классифицировано по сходству последовательностей нуклеиновой кислоты. Ген FAD2, например, включает аллели в данном локусе. Член семейства генов предпочтительно характеризуется по крайней мере 60%, более предпочтительно по крайней мере 70%, более предпочтительно по крайней мере 80% идентичностью последовательности нуклеиновой кислоты в кодирующей последовательности гена.
“Гетерологичный” означает не существующий вместе в естественных условиях.
Молекула нуклеиновой кислоты считается “введенной”, если она введена в клетку или организм в результате манипуляции человеком независимо от метода введения. Примеры введенных молекул нуклеиновой кислоты включают, не ограничиваясь ими, нуклеиновые кислоты, которые были введены в клетки путем трансформации, трансфекции, инъекции и проецирования, и нуклеиновые кислоты, введенные в организм методами, которые включают, не ограничиваясь ими, конъюгирование, эндоцитоз и фагоцитоз.
“Интрон” означает сегмент молекул нуклеиновой кислоты, обычно ДНК, который не кодирует часть или весь экспрессированный белок и который в эндогенных условиях транскрибируется в молекулы РНК, но сплайсируется из эндогенной РНК, прежде чем РНК транслируется в белок. “Молекула дцРНК интрона” и “молекула РНКи интрона” означают молекулу двухцепочечной РНК, которая при введении в клетку или организм способна по крайней мере частично уменьшать уровень мРНК, присутствующей в клетке или клетке организма, при этом молекула двухцепочечной РНК в достаточной степени идентична интрону гена, присутствующего в клетке или организме, для снижения уровня мРНК, содержащей данную последовательность интрона.
Состав масла с “низким содержанием насыщенных жирных кислот” включает от 3,6% до 8% насыщенных жирных кислот.
“Семя сои со средним содержанием олеиновой кислоты” означает семя, содержащее от 50% до 85% олеиновой кислоты в составе масла семени.
Состав масла с “низким содержанием линоленовой кислоты” включает менее примерно 3 мас.% линоленовой кислоты от общего содержания жирных кислот.
Термин “некодирующая” служит для определения последовательностей молекул нуклеиновой кислоты, которые не кодируют часть или весь экспрессированный белок. Некодирующие последовательности включают, не ограничиваясь интронами, промоторные области, 3'-концевые нетранслируемые области (3'-UTR) и 5'-концевые нетранслируемые области (5'-UTR).
Термин “состав масла” служит для определения содержания жирных кислот.
Промотор, который “функционально связан” с одной или несколькими последовательностями нуклеиновой кислоты, способен вызывать экспрессию одной или нескольких последовательностей нуклеиновой кислоты, включая многие кодирующие или некодирующие последовательности нуклеиновой кислоты, имеющие полицистронную конфигурацию.
“Физически связанные” последовательности нуклеиновой кислоты являются последовательностями нуклеиновой кислоты, находящимися в одной молекуле нуклеиновой кислоты.
“Растение” означает целые растения, органы растения (например, листья, стебли, корни и т.д.), семена, растительные клетки и их потомство.
Термин “растительная клетка” означает, не ограничиваясь ими, суспензионные культуры семян, зародыши, меристематические области, ткань каллюса, листья, корни, побеги, гаметофиты, спорофиты, пыльцу и микроспоры.
“Растительные промоторы” означают, не ограничиваясь ими, вирусные промоторы растений, промоторы, выделенные из растений, и синтетические промоторы, способные функционировать в растительной клетке, стимулируя экспрессию мРНК.
“Полицистронный ген” или “полицистронная мРНК” означает любой ген или мРНК, содержащую транскрибированные последовательности нуклеиновой кислоты, которые соответствуют последовательностям нуклеиновой кислоты более чем одного гена, подвергаемого направленному воздействию с целью подавления или экспрессии. Известно, что такие полицистронные гены или мРНК могут содержать последовательности, соответствующие интронам, 5'-UTR, 3'-UTR, кодирующим последовательностям транзитных пептидов, экзонам или их комбинациям, и что рекомбинантный полицистронный ген или мРНК может содержать, не ограничиваясь ими, последовательности, соответствующие одной или нескольким UTR из одного гена и одному или нескольким интронам из второго гена.
“Семяспецифичный промотор” означает промотор, который предпочтительно или исключительно активен в семени. “Предпочтительная активность” означает активность промотора, которая значительно выше в семени, чем в других тканях, органах или органеллах растения. Термин “семяспецифичный” означает, не ограничиваясь ими, активность в алейроновом слое, эндосперме и/или зародыше семени.
“Подавление смысловым интроном” означает сайленсинг гена, вызванный введением смыслового интрона или его фрагмента. Подавление смысловым интроном описано, например, Fillatti в публикации РСТ WO 01/14538 А2.
“Одновременная экспрессия” нескольких агентов, таких как мРНК или белок, означает экспрессию агента одновременно с другим агентом. Такая экспрессия может перекрываться только частично и может также происходить в разной ткани или на разных уровнях.
“Общее содержание в масле” означает общее содержание жирной кислоты независимо от типа жирной кислоты. В используемом здесь значении в определение термина “общее содержание в масле” не входит глицериновый остов.
“Трансген” означает последовательность нуклеиновой кислоты, ассоциированную с экспрессией гена, введенного в организм. Трансген включает, не ограничиваясь ими, эндогенный ген или ген, отсутствующий в организме в естественных условиях. “Трансгенное растение” является любым растением, устойчиво содержащим трансген с возможностью его передачи растением путем полового или вегетативного размножения.
Состав масла с “нулевым содержанием насыщенных жирных кислот” включает менее 3,6% насыщенных жирных кислот.
Применительно к белкам и нуклеиновым кислотам в настоящем описании изобретения обычные заглавные буквы, например “FAD2”, служат для обозначения фермента, белка, полипептида или пептида, и заглавные буквы, напечатанные курсивом, например “FAD2”, служат для обозначения нуклеиновых кислот, включающих, не ограничиваясь ими, гены, кДНК и мРНК. Клетка или организм может содержать семейство из нескольких генов, кодирующих определенный фермент, и заглавная буква, следующая за обозначением гена (А, В, С), означает члена семейства, то есть FAD2-1A является членом семейства генов, отличным от FAD2-1B.
В используемом здесь значении в любой указанный диапазон входят конечные значения данного диапазона за исключением особо оговоренных случаев.
А. Агенты
Агенты по настоящему изобретению предпочтительно являются “биологически активными” в отношении структурного признака, такого как способность молекулы нуклеиновой кислоты гибридизироваться с другой молекулой нуклеиновой кислоты или способность белка связываться антителом (или конкурировать с другой молекулой за такое связывание). Альтернативно такой признак может быть определен как каталитически активный и, таким образом, может включать способность агента опосредовать химическую реакцию или ответ. Указанные агенты предпочтительно являются “по существу чистыми”. Термин “по существу чистый” в используемом здесь значении означает молекулу, отделенную по существу от всех других молекул, ассоциированных с ней в естественных окружающих условиях. Более предпочтительно по существу чистой молекулой является преобладающая разновидность, присутствующая в препарате. По существу чистая молекула может быть более чем на 60%, более чем на 75%, предпочтительно более чем на 90% и наиболее предпочтительно более, чем на 95% очищена от других молекул (за исключением растворителя), присутствующих в естественной смеси. В определение термина “по существу чистый” не входят молекулы, находящиеся в естественных окружающих условиях.
Агенты по настоящему изобретению могут быть также рекомбинантными. В используемом здесь значении термин “рекомбинантный” означает любой агент (например, включающий, не ограничиваясь ими, ДНК или пептид), который получен, однако, непрямым методом, в результате манипуляции молекулой нуклеиновой кислоты человеком. Кроме того, агенты по настоящему изобретению могут быть помечены реагентами, облегчающими обнаружение данного агента, например флуоресцентными метками, химическими метками и/или модифицированными основаниями.
Агенты по настоящему изобретению включают молекулы ДНК, содержащие нуклеотидную последовательность, которая может быть транскрибирована в смысловой или антисмысловой ориентации, в результате чего образуется по крайней мере одна молекула РНК, которая по крайней мере частично является двухцепочечной. В предпочтительном варианте осуществления изобретения агент по настоящему изобретению является молекулой двухцепочечной РНК, содержащей нуклеотидную последовательность, которая является фрагментом FAD2, FATB или FAD2 и FATB. В другом варианте осуществления изобретения агент по настоящему изобретению является молекулой ДНК, которая может быть транскрибирована с образованием нуклеотидной последовательности в смысловой или антисмысловой ориентации в клетке-хозяине. В другом варианте осуществления изобретения молекула нуклеиновой кислоты может содержать нуклеотидную последовательность в смысловой и антисмысловой ориентации, или в другом варианте осуществления изобретения молекула нуклеиновой кислоты может содержать нуклеотидную последовательность в смысловой или антисмысловой ориентации. Такие нуклеотидные последовательности могут быть функционально связаны с одним и тем же промотором, разными промоторами, одним промотором или несколькими промоторами. Такие нуклеотидные последовательности могут находиться в одной молекуле ДНК или в нескольких молекулах ДНК.
Агенты по настоящему изобретению включают молекулы нуклеиновой кислоты, содержащие последовательность ДНК, которая на протяжении всей длины по крайней мере на 50%, 60% или 70% идентична кодирующей или некодирующей области растения либо последовательности нуклеиновой кислоты, комплементарной кодирующей или некодирующей области растения. Более предпочтительными являются последовательности ДНК, которые на протяжении всей длины по крайней мере на 80%, по крайней мере на 85%, по крайней мере на 90%, по крайней мере на 95%, по крайней мере на 97%, по крайней мере на 98%, по крайней мере на 99% или на 100% идентичны кодирующей или некодирующей области растения, либо последовательности нуклеиновой кислоты, комплементарной кодирующей или некодирующей области растения.
Термин “идентичность”, как хорошо известно в данной области, означает взаимосвязь между двумя или более полипептидными последовательностями либо между двумя или более последовательностями молекулы нуклеиновой кислоты, определяемую путем сравнения указанных последовательностей. В данной области термин “идентичность” означает также степень родства между полипептидными последовательностями и последовательностями молекулы нуклеиновой кислоты, определяемую при сравнении цепей таких последовательностей. “Идентичность” можно легко высчитать известными методами, которые включают, не ограничиваясь ими, методы, описанные в публикациях Computational Molecular Biology, Lesk, ed., Oxford University Press, New York 1988; Biocoinputing: Informatics and Genome Projects, Smith, ed., Academic Press, New York 1993; Computer Analysis of Sequence Data, Part I, Griffin and Griffin, eds., Humana Press, New Jersey 1994; Sequence Analysis in Molecular Biology, von Heinje, Academic Press 1987; Sequence Analysis Primer, Gribskov and Devereux, cds., Stockton Press, New York 1991; and Carillo and Lipman, SIAM J Applied Math, 48:1073 1988.
Методы определения идентичности предназначены для выявления наибольшего соответствия между анализируемыми последовательностями. Кроме того, методы определения идентичности кодифицированы в общедоступных программах. Компьютерные программы, которые могут быть использованы для определения идентичности двух последовательностей, включают, не ограничиваясь ими, GCG; пакет из пяти программ BLAST, из которых три программы предназначены для запроса нуклеотидных последовательностей (BLASTN, BLASTX и TBLASTX) и две программы предназначены для запроса белковых последовательностей (BLASTP и TBLASTN). Программу BLASTX можно приобрести в NCBI и из других источников, например BLAST Manual, Altschul et al., NCBI NLM NIH, Bethesda, MD 20894; Altschul et al., J. Mol. Biol. 215:403-410 (1990). Для определения идентичности можно также использовать хорошо известный алгоритм Смита-Ватермана.
При сравнении полипептидных последовательностей обычно используются следующие параметры: алгоритм: Needleman and Wunsch, J. Mol. Biol. 48:443-453 (1970); матрица сравнения: BLOSSUM62 из публикации Hentikoff and Hentikoff, Proc. Natl. Acad. Sci. USA 89:10915-10919 (1992); штраф за разрыв: 12; штраф за длину разрыва: 4. Программа, которая может быть использована с вышеуказанными параметрами, является общедоступной программой, известной как программа “разрывов” компании Genetics Computer Group (“GCG”), Madison, Wisconsin. Вышеуказанные параметры наряду с отсутствием штрафа за концевой разрыв являются параметрами по умолчанию для сравнения пептидов.
При сравнении последовательностей молекул нуклеиновой кислоты используются следующие параметры: алгоритм: Needleman and Wunsch, J. Mol. Biol. 48:443-453 (1970); матрица сравнения: совпадения - +10; несовпадения = 0; штраф за разрыв: 50; штраф за длину разрыва: 3. В используемом здесь значении “% идентичности” определяют, используя вышеуказанные параметры в качестве параметров по умолчанию для сравнения последовательностей молекул нуклеиновой кислоты и программу “разрывов” компании GCG, версия 10.2.
Подмножества последовательностей нуклеиновой кислоты по настоящему изобретению включают фрагментарные молекулы нуклеиновой кислоты. “Фрагментарная молекула нуклеиновой кислоты” означает часть более крупной молекулы нуклеиновой кислоты и может состоять из значительной части или наибольшей части более крупной молекулы нуклеиновой кислоты. Фрагментарная молекула нуклеиновой кислоты может включать олигонуклеотид меньшей длины от около 15 до около 400 последовательных нуклеотидов и более предпочтительно от около 15 до около 45 последовательных нуклеотидов, от около 20 до около 45 последовательных нуклеотидов, от около 15 до около 30 последовательных нуклеотидов, от около 21 до около 30 последовательных нуклеотидов, от около 21 до около 25 последовательных нуклеотидов, от около 21 до около 24 последовательных нуклеотидов, от около 19 до около 25 последовательных нуклеотидов или около 21 последовательного нуклеотида. Фрагментарные молекулы нуклеиновой кислоты могут состоять из значительной части или наибольшей части кодирующей или некодирующей области растения либо альтернативно могут включать олигонуклеотиды меньшей длины. В предпочтительном варианте осуществления изобретения фрагмент характеризуется 100% идентичностью с кодирующей или некодирующей областью растения. В другом предпочтительном варианте осуществления изобретения фрагмент включает часть более крупной последовательности нуклеиновой кислоты. В другом варианте осуществления изобретения фрагментарная молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая включает по крайней мере 15, 25, 50, 100, 200, 300 или 400 последовательных нуклеотидов молекулы нуклеиновой кислоты по настоящему изобретению. В предпочтительном варианте осуществления изобретения молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая включает по крайней мере 15, 25, 50, 100, 200, 300 или 400 последовательных нуклеотидов кодирующей или некодирующей области растения. В наиболее предпочтительном варианте осуществления изобретения молекула нуклеиновой кислоты содержит последовательность нуклеиновой кислоты, которая включает примерно 1, 2, 5, 10, 20, 30, 40, 50, 60, 70, 80 или 90% последовательных нуклеотидов всей кодирующей или некодирующей области. В предпочтительном варианте осуществления изобретения вся кодирующая или некодирующая область может быть генетическим элементом, выбираемым из всего гена, одного экзона, одного интрона, сигнальной последовательности или нетранслируемой области (UTR). Генетический элемент, который не содержит полной последовательности всего генетического элемента, может быть фрагментом генетического элемента. В предпочтительном варианте осуществления настоящего изобретения генетический элемент имеет длину, равную по крайней мере 40 нуклеотидам. В одном варианте осуществления настоящего изобретения фрагмент гена является частью всего генетического элемента, и такой фрагмент содержит последовательные нуклеотиды примерно 1, 2, 5, 10, 20, 30, 40, 50, 60, 70, 80 или 90% всего генетического элемента. В одном варианте осуществления настоящего изобретения фрагментарная молекула нуклеиновой кислоты соответствует примерно 5%-80%, примерно 10%-70%, примерно 10%-60%, примерно 10%-50%, примерно 25%-60%, примерно 25%-50%, примерно 40%-60%, примерно 40%-80%, примерно 50%-90% длины всего генетического элемента.
В предпочтительном варианте осуществления изобретения фрагмент интрона FAD2-1 включает от около 20 до около 420, от около 30 до около 420, от около 40 до около 320, от около 50 до около 200, от около 50 до около 400, от около 50 до около 420,от около 60 до около 320, от около 70 до около 220, от около 100 до около 200, от около 100 до около 320, от около 150 до около 200, от около 150 до около 220, от около 150 до около 400, от около 200 до около 300 или от около 300 до около 400 последовательных нуклеотидов. В другом предпочтительном варианте осуществления изобретения фрагмент интрона FAD2-1 имеет длину, равную примерно 100, примерно 150, примерно 200, примерно 220, примерно 250, примерно 300, примерно 320 или примерно 350 последовательным нуклеотидам. В другом предпочтительном варианте осуществления изобретения длина фрагмента интрона FAD2-1 уменьшена примерно на 20, примерно на 40, примерно на 60, примерно на 80, примерно на 100, примерно на 120, примерно на 140, примерно на 160, примерно на 180, примерно на 200, примерно на 220, примерно на 240, примерно на 260, примерно на 280, примерно на 290, примерно на 300, примерно на 320, примерно на 340, примерно на 360, примерно на 380, примерно на 400 последовательных нуклеотидов по сравнению с длиной SEQ ID NO:1. Во всех указанных фрагментах интрона FAD2-1 усечение или делеция может быть начата у 5'-конца, 3'-конца или внутри интрона FAD2-1. Во всех указанных фрагментах интрона FAD2-1 последовательность интрона FAD2-1 может представлять собой SEQ ID NO:1.
В предпочтительном варианте осуществления изобретения фрагмент гена FATB включает от около 80 до около 450, от около 100 до около 500, от около 70 до около 500, от около 200 до около 400, от около 150 до около 300, от около 250 до около 350, от около 200 до около 350 последовательных нуклеотидов гена FATB. В предпочтительном варианте осуществления изобретения фрагмент FATB выделяют из половины всех нуклеотидов FATB, начиная с 5'-конца. Во всех указанных фрагментах FATB усечение или делеция может быть начата у 5'-конца, 3'-конца или внутри FATB. В предпочтительном варианте осуществления изобретения фрагмент FATB выделяют из половины всех нуклеотидов FATB, начиная с 5'-конца FATB, из третьей части всех нуклеотидов FATB, расположенных ближе всего к 5'-концу. В особенно предпочтительном варианте осуществления изобретения фрагмент FATB содержит кодирующую последовательность транзитного пептида, которая предпочтительно кодирует транзитный пептид хлоропласта. В особенно предпочтительном варианте осуществления изобретения фрагмент FATB является фрагментом кодирующей последовательности транзитного пептида, которая предпочтительно кодирует транзитный пептид хлоропласта. В другом, особенно предпочтительном, варианте осуществления изобретения фрагмент FATB дополнительно включает примерно 20, примерно 25, примерно 30, примерно 35, 38, 39, 40, 41, 42, 43, примерно 45, примерно 50, примерно 55 или примерно 60 последовательных нуклеотидов 5'-UTR FATB. В наиболее предпочтительном варианте осуществления изобретения фрагмент включает комбинацию двух или более несмежных фрагментов или отдельных генетических элементов, таких как 3'-UTR FATB, слитая с 5'-UTR FATB. Агенты по настоящему изобретению включают молекулы нуклеиновой кислоты. Например, в одном варианте осуществления изобретения молекула нуклеиновой кислоты по настоящему изобретению включает, не ограничиваясь ими, последовательности интрона SEQ ID NO:19, 20, 21, 22, 23, 25, 32, 33, 34, 35, 44, 45, 46 или 47 либо их фрагменты или комплементы. В другом варианте осуществления изобретения молекула нуклеиновой кислоты включает последовательность нуклеиновой кислоты, которая при введении в клетку или организм способна подавлять продуцирование РНК или белка при одновременной экспрессии, коэкспрессии или координированной экспрессии другой РНК или белка. В одном варианте осуществления изобретения молекула нуклеиновой кислоты включает последовательность нуклеиновой кислоты, которая при введении в клетку или организм способна подавлять, по крайней мере частично снижать, уменьшать, значительно сокращать или эффективно устранять экспрессию эндогенной РНК FAD2, FAD3 и/или FATB при одновременной экспрессии, коэкспрессии или координированной экспрессии по крайней мере одного продукта РНК или белка бета-кетоацил-АСР-синтазы I, бета-кетоацил-АСР-синтазы IV, дельта-9-десатуразы и/или СР4 EPSPS.
В результате подавления, по крайней мере частичного снижения, уменьшения, значительного сокращения или эффективного устранения экспрессии по крайней мере одного или нескольких эндогенных генов в растительной клетке уменьшается количество FAD2 и/или FAD3, то есть снижаются уровни белка в устойчивом состоянии, и может быть уменьшено процентное содержание полиненасыщенных жирных кислот, таких как линолеат (С18:2) и линоленат (С18:3). Модификация состава жирных кислот, пригодных для включения в триацилглицерины, может влиять на состав масел в растительной клетке. Таким образом, уменьшение экспрессии FAD2 и/или FAD3 может вызывать увеличение доли мононенасыщенных жирных кислот, таких как олеат (С18:1). При уменьшении количества FATB в растительной клетке происходит сокращение количества насыщенных жирных кислот, таких как пальмитат и стеарат. Таким образом, уменьшение экспрессии FATB может вызывать увеличение доли ненасыщенных жирных кислот, таких как олеат (18:1). Одновременное подавление экспрессии FAD2, FAD3 и FATB направляет путь FAS в сторону увеличения мононенасыщенных жирных кислот, содержащих 18 атомов углерода, таких как олеат (С18:1). См. патент США № 5955650.
В результате увеличения количества бета-кетоацил-АСР-синтазы I (KAS I) и/или бета-кетоацил-АСР-синтазы IV (KAS IV) в растительной клетке может быть уменьшено процентное содержание 16:0-АСР, что вызывает увеличение процентного содержания 18:0-АСР. Большее количество 18:0-АСР в сочетании с одновременным подавлением одного или нескольких генов FAD2, FAD3 и FATB позволяет увеличить содержание олеата (С18:1) в составе масла. В результате увеличения количества дельта-9-десатуразы в растительной клетке может быть увеличено процентное содержание ненасыщенных жирных кислот, что вызывает общее уменьшение стеарата и общего содержания насыщенных жирных кислот.
Указанными комбинациями повышенной и пониженной экспрессии ферментов можно манипулировать для получения составов масла, включающих жирных кислоты, с повышенным уровнем олеата, пониженными уровнями линолеата, линолената, стеарата и/или пальмитата и пониженным общим содержанием насыщенных жирных кислот. Экспрессия гена в растениях может быть усилена путем введения дополнительных копий кодирующих последовательностей генов в растительную клетку или предпочтительно путем введения дополнительных копий кодирующих последовательностей гена в геном растения. Сверхэкспрессия может быть также достигнута в результате увеличения активности регуляторных механизмов, которые регулируют экспрессию генов, то есть повышают экспрессию генов.
Продуцирование СР4 EPSPS в растительной клетке сообщает растительной клетке устойчивость или толерантность к глифосату, что является удобным методом идентификации успешных трансформантов на основании устойчивости к глифосату.
Подавление экспрессии гена в растениях, известное также как сайленсинг гена, происходит как на транскрипционном уровне, так и на посттранскрипционном уровне. Существуют разные методы подавления экспрессии эндогенных последовательностей в клетке-хозяине, которые включают, не ограничиваясь ими, подавление антисмысловой последовательностью, косупрессию, рибозимы, комбинации смысловой и антисмысловой двухцепочечной интерферирующей РНК, сайленсинг промотором и ДНК-связывающие белки, такие как белки цинксодержащей пальцеобразной области. (См., например, WO 98/53083, WO 01/14538 и патент США № 5759829 (Shewmaker)). Некоторые из указанных механизмов ассоциированы с гомологией нуклеиновых кислот на уровне ДНК или РНК. Такая гомология означает сходство ДНК или белковых последовательностей в одном виде или в разных видах. Сайленсинг гена происходит в том случае, если последовательность ДНК, введенная в клетку-хозяина, настолько гомологична эндогенному гену, что транскрипция введенной последовательности ДНК вызывает транскрипционный или посттранскрипционный сайленсинг эндогенного гена. Гомология, достаточная для подавления экспрессии в устойчивом состоянии, может соответствовать по крайней мере 50%, примерно 60% или примерно 70% идентичности всей последовательности ДНК кодирующей или некодирующей области растения либо последовательности нуклеиновой кислоты, комплементарной кодирующей или некодирующей области растения. Более предпочтительными являются последовательности ДНК, которые по крайней мере на 80%, по крайней мере на 85%, по крайней мере на 90%, по крайней мере на 95%, по крайней мере на 97%, по крайней мере на 98%, по крайней мере на 99% или 100% идентичны кодирующей или некодирующей области растения, либо последовательности нуклеиновой кислоты, комплементарной кодирующей или некодирующей области растения. В растениях специфичный для последовательности сайленсинг могут вызывать молекулы двухцепочечной РНК. Сайленсинг гена часто определяется как интерференция двухцепочечной РНК (“дцРНКи”) в растениях, как интерференция РНК или РНКи в Caenorhabditis elegans и у животных и как подавление в грибах.
В предпочтительном варианте осуществления изобретения молекула нуклеиновой кислоты по настоящему изобретению включает первую совокупность последовательностей ДНК, каждая из которых обладает достаточной гомологией с одной или несколькими кодирующими или некодирующими последовательностями гена растения, благодаря чему в случае экспрессии такая последовательность может эффективно устранять, значительно сокращать или по крайней мере частично снижать уровень транскрипта или белка мРНК, кодированного геном, из которого была выделена кодирующая или некодирующая последовательность, или любого гена, гомологичного кодирующей или некодирующей последовательности-мишени.
В предпочтительном варианте осуществления изобретения молекула нуклеиновой кислоты по настоящему изобретению включает: (а) первую совокупность последовательностей ДНК, каждая из которых обладает достаточной гомологией с одной или несколькими кодирующими или некодирующими последовательностями гена растения, благодаря чему в случае экспрессии такая последовательность может эффективно устранять, значительно сокращать или по крайней мере частично снижать уровень транскрипта или белка мРНК, кодированного геном, из которого была выделена кодирующая или некодирующая последовательность, или любого гена, гомологичного некодирующей последовательности-мишени, и (b) вторую совокупность последовательностей ДНК, каждая из которых обладает достаточной гомологией с геном растения, благодаря чему в случае экспрессии такая последовательность может по крайней мере частично усиливать, увеличивать или значительно повышать уровень транскрипта или белка мРНК, кодированного данным геном.
В используемом здесь значении “совокупность” последовательностей ДНК может представлять собой одну или несколько последовательностей, кодирующих или не кодирующих белок. Например, первая совокупность последовательностей ДНК может включать только промотор, некодирующую область и терминатор. Вторая совокупность последовательностей ДНК может или не может присутствовать после или до первой совокупности последовательностей ДНК.
В используемом здесь значении “уменьшение” уровня или количества агента, такого как белок или мРНК, означает снижение уровня или количества по сравнению с клеткой или организмом, в котором отсутствует последовательность ДНК, способная уменьшать содержание данного агента. Например, “по крайней мере частичное уменьшение” означает сокращение по крайней мере на 25%, “значительное сокращение” означает уменьшение по крайней мере на 75%, и “эффективное устранение” означает уменьшение более чем на 95%, при этом все уменьшения уровней или количества агента определяются по сравнению с клеткой или организмом, в котором отсутствует последовательность ДНК, способная уменьшать содержание данного агента.
В используемом здесь значении “повышенный” или “увеличенный” уровень или количество агента, такого как белок или мРНК, означает, что данный уровень или количество выше уровня или количества агента, присутствующего в клетке, ткани или растении с подобной генетической средой, но при отсутствии введенной молекулы нуклеиновой кислоты, кодирующей данный белок или мРНК. Например, “по крайней мере частично повышенный” уровень означает увеличение по крайней мере на 25%, “значительно повышенный” уровень означает увеличение по крайней мере на 100%, при этом все увеличения уровней или количества агента определяют по сравнению с уровнем или количеством агента, присутствующего в клетке, ткани или растении с подобной генетической средой, но при отсутствии введенной молекулы нуклеиновой кислоты, кодирующей данный белок или мРНК. В предпочтительном варианте осуществления изобретения повышение экспрессии может быть любой экспрессией, когда белок является гетерологичным для данной системы. Например, экспрессия СР4 EPSPS может быть увеличением экспрессии, если в растении отсутствовала экспрессия до введения молекулы нуклеиновой кислоты, кодирующей данный белок.
Сравнение уровней агента предпочтительно выполняют в организмах с подобной генетической средой. Подобная генетическая среда предпочтительно имеет место в том случае, когда в сравниваемых организмах последовательности характеризуются 50% или большей, более предпочтительно 75% или большей и еще предпочтительнее 90% или большей идентичностью ядерного генетического материала. В другом предпочтительном варианте осуществления изобретения подобная генетическая среда имеет место в том случае, когда сравниваемые организмы являются растениями и указанные растения являются изогенными за исключением любого генетического материала, первоначально введенного методами трансформации растения. Уровень или количество агента может быть измерено любым приемлемым методом, неограничивающие примеры которого включают сравнение уровней транскрипта мРНК, уровней белка или пептида и/или фенотипа, особенно состава масла. В используемом здесь значении транскрипты мРНК включают процессированные и непроцессированные транскрипты мРНК, и белки или пептиды включают белки или пептиды с посттрасляционной модификацией или без нее.
Последовательности ДНК первой совокупности последовательностей ДНК могут быть кодирующими последовательностями, последовательностями интронов, последовательностями 3'-UTR, последовательностями 5'-UTR, промоторными последовательностями, другими некодирующими последовательностями или любой комбинацией указанных последовательностей. Первая совокупность последовательностей ДНК кодирует одну или несколько последовательностей, которые в случае экспрессии способны избирательно снижать уровень белка или транскрипта или как белка, так и транскрипта, кодированных геном, выбираемым из группы, включающей FAD2, FAD3 и FATB. В предпочтительном варианте осуществления изобретения первая совокупность последовательностей ДНК способна экспрессировать антисмысловую РНК, в которой отдельные антисмысловые последовательности могут быть связаны в одном транскрипте или могут представлять собой несвязанные отдельные транскрипты. В другом предпочтительном варианте осуществления изобретения первая совокупность последовательностей ДНК представляет собой физически связанные последовательности, способные экспрессировать одну молекулу дцРНК. В другом предпочтительном варианте осуществления изобретения первая совокупность последовательностей ДНК способна экспрессировать смысловую косупрессорную РНК, в которой отдельные смысловые последовательности могут быть связаны в одном транскрипте или могут представлять собой несвязанные отдельные транскрипты. Типичные варианты первой совокупности последовательностей ДНК приведены в части В подробного описания изобретения и в примерах.
Вторая совокупность последовательностей ДНК кодирует одну или несколько последовательностей, которые в случае экспрессии способны повышать уровень белка или транскрипта или как белка, так и транскрипта, кодированных геном, выбираемым из группы, включающей бета-кетоацил-АСР-синтазу I (KAS I), бета-кетоацил-АСР-синтазу IV (KAS IV), дельта-9-десатуразу и СП4 EPSPS. Последовательности ДНК второй совокупности последовательностей ДНК могут быть физически связанными последовательностями. Типичные варианты второй совокупности последовательностей ДНК приведены ниже в частях С и D подробного описания изобретения.
Таким образом, настоящее изобретение относится к способам изменения состава жирных кислот и соединений, содержащих такие жирных кислоты, таких как масла, воски и жиры. Настоящее изобретение относится также к способам продуцирования определенных жирных кислот в растительных клетках-хозяевах. Такие способы предполагают использование полигенных экспрессирующих кластеров, представленных в настоящем описании изобретения, для модификации пути FAS в растительной клетке-хозяине.
В. Первая совокупность последовательностей ДНК
В одном варианте осуществления настоящего изобретения молекула нуклеиновой кислоты включает первую совокупность последовательностей ДНК, которая при введении в клетку или организм экспрессирует одну или несколько последовательностей, способных эффективно устранять, значительно сокращать или по крайней мере частично уменьшать уровни транскриптов или белков мРНК, кодированных одним или несколькими генами. Предпочтительные варианты осуществления изобретения включают в качестве мишени эндогенный ген, ген растения и невирусный ген. В одном варианте осуществления настоящего изобретения ген является геном FAD2, FAD3 или FATB.
В одном варианте осуществления изобретения молекула нуклеиновой кислоты по настоящему изобретению включает последовательность ДНК, обладающую значительной гомологией с одной или несколькими кодирующими или некодирующими последовательностями из гена растения, которая при введении и экспрессии в растительной клетке или растении может эффективно устранять, значительно сокращать или по крайней мере частично снижать уровень транскрипта или белка мРНК, кодированного геном, из которого была выделена кодирующая или некодирующая последовательность. Последовательности ДНК в первой совокупности последовательностей ДНК транскрибируют последовательности РНК или фрагменты РНК, которые по крайней мере на 90%, предпочтительно по крайней мере на 95%, более предпочтительно по крайней мере на 98% и наиболее предпочтительно на 100% идентичны кодирующей или некодирующей области, выделенной из подавляемого гена. Такая процентная идентичность может быть сравнима с другим фрагментом нуклеиновой кислоты.
Некодирующая последовательность предпочтительно является 3'-UTR, 5'-UTR, частью последовательности, кодирующей белок или интрон из гена растения. Более предпочтительно некодирующая последовательность является промоторной последовательностью, 3'-UTR, 5'-UTR или интроном из гена растения. Интрон может быть локализован между экзонами, в 5'-UTR или 3'-UTR гена растения. Кодирующая последовательность предпочтительно является частью кодирующей белок рамки.
Одна или несколько последовательностей в первой совокупности последовательностей ДНК могут быть предназначены для продуцирования дцРНК, смысловой супрессорной РНК, антисмысловой РНК или любого другого супрессорного транскрипта для достижения требуемого эффекта при введении в растительную клетку или растение. Такая последовательность ДНК может быть фрагментарной молекулой нуклеиновой кислоты.
Растительный интрон может быть любым растительным интроном из эндогенного или введенного гена. Последовательности нуклеиновой кислоты таких интронов из организмов могут быть получены или выделены из многих источников, которые включают, не ограничиваясь ими, базы данных, такие как EMBL и банк генов Genbank, которые могут быть найдены в Интернете по адресам ebi.ac.uk/swisprot/; expasy.ch/; embl-heidelberg.de/ и ncbi.nlm.nih.gov. Последовательности нуклеиновой кислоты таких интронов могут быть также получены без каких-либо ограничений из таких источников, как программа GENSCAN, которая может быть найдена в Интернете по адресу genes.mit.edu/GENSCAN.html.
Дополнительные интроны могут быть также получены методами, которые включают, не ограничиваясь ими, скрининг геномной библиотеки зондом последовательностей известного экзона или интрона, сравнение геномной последовательности с соответствующей последовательностью кДНК или клонирование интрона, такого как кДНК сои, путем сравнительного анализа с геномной последовательностью из другого организма, такого как, например, Arabidopsis. Кроме того, специалисту в данной области должны быть известны другие последовательности нуклеиновой кислоты интронов. Описанные выше методы могут быть также использованы для выделения и получения других некодирующих последовательностей, которые включают, не ограничиваясь ими, промоторные последовательности, последовательности 3'-UTR и последовательности 5'-UTR.
Ген “FAD2”, “Δ12 десатуразы” или “омега-6-десатуразы” кодирует фермент (FAD2), способный катализировать введение двойной связи в ацильную часть жирной кислоты в двенадцатом положении от карбоксильного конца. Термин “FAD2-1” служит для обозначения гена FAD2, который специфично экспрессирован в естественных условиях в ткани семени, и термин “FAD2-2” служит для обозначения гена FAD2, который: (а) отличается от гена FAD2-1 и (b) экспрессирован в естественных условиях во многих тканях, включая семя. Типичные последовательности FAD2 включают, не ограничиваясь ими, последовательности, представленные в заявке на патент США № 10/176149, поданной 21 июня 2002 г., и в SEQ ID NO:1-6.
Ген “FAD3”, “Δ15 десатуразы” или “омега-3-десатуразы” кодирует фермент (FAD3), способный катализировать введение двойной связи в ацильную часть жирной кислоты в пятнадцатом положении от карбоксильного конца. Термины “FAD3-1, FAD3-A, FAD3-B и FAD3-C” служат для обозначения членов семейства генов FAD3, которые экспрессированы в естественных условиях во многих тканях, включая семя. Типичные последовательности FAD3 включают, не ограничиваясь ими, последовательности, представленные в заявке на патент США № 10/176149, поданной 21 июня 2002 г., и в SEQ ID NO:7-27.
Ген “FATB” или “пальмитоил-АСР-тиоэстеразы” кодирует фермент (FATB), способный катализировать гидролитическое расщепление сложной тиоэфирной связи углерод-сера в пантотенпростетиновой группе пальмитоил-АСР в качестве предпочтительной реакции. Указанный фермент может также катализировать гидролиз других сложных тиоэфиров АСР жирной кислоты. Типичные последовательности FATB-1 включают, не ограничиваясь ими, последовательности, представленные в предварительной заявке на патент США № 60/390185, поданной 21 июня 2002 г.; в патентах США № 5955329, 5723761, 5955650 и 6331664 и в SEQ ID NO:28-37. Типичные последовательности FATB-2 включают, не ограничиваясь ими, последовательности, представленные в SEQ ID NO:42-47.
С. Вторая совокупность последовательностей ДНК
В одном варианте осуществления настоящего изобретения молекула нуклеиновой кислоты содержит вторую совокупность последовательностей ДНК, которая при введении в клетку или организм способна частично повышать, увеличивать, повышать или значительно повышать уровни транскриптов или белков мРНК, кодированных одним или несколькими генами. В одном варианте осуществления настоящего изобретения ген является эндогенным геном. В другом варианте осуществления настоящего изобретения ген может быть гетерологичным геном. В предпочтительном варианте осуществления изобретения гетерологичные и эндогенные гены могут присутствовать в одной и той же молекуле нуклеиновой кислоты. В одном варианте осуществления настоящего изобретения ген является геном растения. В другом варианте осуществления настоящего изобретения ген является усеченным геном, при этом усеченный ген способен катализировать реакцию, катализируемую полным геном. В одном варианте осуществления настоящего изобретения ген является геном бета-кетоацил-АСР-синтазы I, геном бета-кетоацил-АСР-синтазы IV, геном дельта-9-десатуразы, геном СР4 EPSPS или комбинацией указанных генов.
Ген по настоящему изобретению может быть любым геном, эндогенным или введенным. Последовательности нуклеиновой кислоты таких генов могут быть получены из многих источников, которые включают, не ограничиваясь ими, такие базы данных, как EMBL и Genbank, которые могут быть найдены в Интернете по адресам ebi.ac.uk/swisprot/; expasy.ch/; embl-heidelberg.de/ и ncbi.nlm.nih.gov. Последовательности нуклеиновой кислоты таких генов могут быть также получены из таких источников, как программа GENSCAN, которая может быть найдена в Интернете по адресу genes.mit.edu/GENSCAN.html.
Дополнительные гены могут быть также получены методами, которые включают, не ограничиваясь ими, скрининг геномной библиотеки или библиотеки кДНК зондом последовательностей известных генов, клонирование гена путем сравнительного анализа с геном или зондом из другого организма, такого как, например, Arabidopsis. Кроме того, специалисту в данной области должны быть известны другие последовательности нуклеиновой кислоты генов. Дополнительные гены могут быть, например, амплифицированы при помощи полимеразной цепной реакции (ПЦР) и использованы в одном варианте осуществления настоящего изобретения. Кроме того, специалисту в данной области должны быть известны другие последовательности нуклеиновой кислоты генов.
Автоматические синтезаторы нуклеиновой кислоты могут быть использованы для данной цели, а также для создания молекулы нуклеиновой кислоты, имеющей последовательность, обнаруженную в клетке или организме. Помимо такого синтеза молекулы нуклеиновой кислоты могут быть использованы для определения пар праймеров, которые могут быть использованы при выполнении ПЦР для амплификации и получения любой требуемой молекулы нуклеиновой кислоты или фрагмента первого гена.
Ген “KAS I” или “бета-кетоацил-АСР-синтазы I” кодирует фермент (KAS I), способный катализировать удлинение ацильной части жирной кислоты до образования пальмитоил-АСР (С16:0). Типичные последовательности KAS I включают, не ограничиваясь ими, последовательности, представленные в патенте США № 5475099, публикации РСТ WO 94/10189 и SEQ ID NO:38.
Ген “KAS IV” или “бета-кетоацил-АСР-синтазы IV” кодирует фермент (KAS IV), способный катализировать конденсацию ацил-АСР со средней длиной цепи и увеличивать продуцирование 18:0-АСР. Типичные последовательности KAS IV включают, не ограничиваясь ими, последовательности, представленные в публикации РСТ WO 98/46776 и SEQ ID NO:39.
Ген “дельта-9-десатуразы”, “стеароил-АСР-десатуразы” или “омега-9-десатуразы” кодирует фермент, способный катализировать введение двойной связи в ацильную часть жирной кислоты в девятом положении от карбоксильного конца. Предпочтительная дельта-9-десатураза по настоящему изобретению является растительной или цианобактериальной дельта-9-десатуразой и более предпочтительно дельта-9-десатуразой, которая также обнаружена в организме, выбираемом из группы, включающей Cartharmus tinctorius, Ricinus communis, Simmonsia chinensis и Brassica campestris. Типичные последовательности дельта-9-десатуразы включают, не ограничиваясь ими, последовательности, представленные в патенте США № 5723595 и SEQ ID NO:40-41.
Ген “CP4 EPSPS” или “СР4 5-енолпирувилшикимат-3-фосфатсинтазы” кодирует фермент (СР4 EPSPS), способный сообщать значительную степень устойчивости к глифосату полученной растительной клетке или растениям. Последовательность СР4 EPSPS может быть последовательностью СР4 EPSPS, полученной из СР4 Agrobacterium tumefaciens sp., ее вариантом или синтетической формой, описанной в патенте США № 5633435. Типичные последовательности СР4 EPSPS включают, не ограничиваясь ими, последовательности, представленные в патентах США № 5627061 и 5633435.
D. Рекомбинантные векторы и конструкции
Для трансформации или трансфекции растений могут быть использованы одна или несколько конструкций нуклеиновой кислоты по настоящему изобретению. Уровни продуктов, таких как транскипты или белки, могут быть повышены или снижены в таком организме, как растение, или такие продукты могут быть локализованы в одном или нескольких специфичных органах или тканях организма. Например, уровни продуктов могут быть повышены или понижены в одной или нескольких тканях и органах растения, которые включают, не ограничиваясь ими, корни, клубни, стебли, листья, побеги, фрукты, ягоды, орехи, кору, стручки, семена и цветы. Предпочтительным органом является семя. Например, экзогенный генетический материал может быть перенесен в растительную клетку, которая регенерирует с образованием цельного, плодородного или стерильного растения или части растения.
“Экзогенный генетический материал” может быть любым генетическим материалом, встречающимся в естественных условиях или полученным из любого источника, пригодного для введения в любой организм. Такой экзогенный генетический материал включает, не ограничиваясь ими, молекулы и конструкции нуклеиновой кислоты по настоящему изобретению. Экзогенный генетический материал может быть перенесен в клетку-хозяина при помощи вектора на основе ДНК или конструкции, созданной для такой цели. Аналогичным образом экзогенный генетический материал может быть перенесен в клетку-хозяина вирусом. Экзогенный генетический материал может содержать последовательность ДНК, идентичную эндогенному гену, но повторно введенную в клетку-хозяина при помощи вектора на основе ДНК или конструкции, предназначенной для подавления экспрессии эндогенного гена. Создание такого вектора описано в данной области (см., например, публикацию Plant Molecular Biology: A Laboratory Manual, Clark (ed.), Springer, New York (1997)). В предпочтительном варианте осуществления изобретения экзогенный генетический материал является рекомбинантной ДНК.
Конструкция или вектор может включать промотор, функционирующий в растительной клетке, или растительный промотор для экспрессии выбранной молекулы нуклеиновой кислоты. В научной литературе описан ряд промоторов, являющихся активными в растительных клетках, причем в растениях предпочтительно используют промоторы CaMV 35S и FMV. Другие примеры предпочтительных промоторов включают арцелин фасоли и 7S альфа. Дополнительные предпочтительные промоторы являются усиленными или дуплицированными вариантами промоторов CaMV 35S и FMV 35S. Odell et al., Nature 313:810-812 (1985); патент США № 5378619. Дополнительные промоторы, пригодные для использования, описаны, например, в патентах США № 5378619, 5391725, 5428147, 5447858, 5608144, 5614399, 5633441, 5633435 и 4633436. Кроме того, может быть использован тканеспецифичный энхансер.
Особенно предпочтительные промоторы могут быть также использованы для экспрессии молекулы нуклеиновой кислоты по настоящему изобретению в семенах или фруктах. В предпочтительном варианте осуществления изобретения используемым промотором является семяспецифичный промотор. Примеры таких промоторов включают 5'-концевые регуляторные области из таких генов, как напин (Kridl et al., Seed Sci. Res. 1:209-219 (1991)), фазеолин, стеароил-АСР-десатураза, 7Sα, 7Sα' (Chen et al., Proc. Natl. Acad. Sci., 83:8560-8564 (1986), USP, арцелин и олеозин. Предпочтительными промоторами для экспрессии в семени являются промоторы 7Sα, 7Sα', напина и FAD2-1A.
Конструкции или векторы могут также содержать другие генетические элементы, которые включают, не ограничиваясь ими, 3'-концевые терминаторы транскрипции, 3'-концевые сигналы полиаденилирования, другие нетранслируемые последовательности нуклеиновой кислоты, последовательности переноса или направленного воздействия, селектируемые или отбираемые маркеры, промоторы, энхансеры и операторы. Конструкции или векторы могут также содержать ген без промотора, который после введения может использовать эндогенный промотор.
Молекулы нуклеиновой кислоты, которые могут быть использованы для трансформации или трансфекции растений, могут быть любыми молекулами нуклеиновой кислоты по настоящему изобретению. Настоящее изобретение не ограничивается иллюстрированными вариантами осуществления. Типичные молекулы нуклеиновой кислоты описаны в части А подробного описания изобретения, и другие не ограничивающие объем изобретения типичные молекулы нуклеиновой кислоты описаны ниже и иллюстрированы на фиг.1-4 и в примерах.
Варианты молекулы нуклеиновой кислоты по настоящему изобретению показаны на фиг.1-4. Как было указано выше, молекула нуклеиновой кислоты содержит (а) первую совокупность последовательностей ДНК и (b) вторую совокупность последовательностей ДНК, которые локализованы в одной или нескольких областях Т-ДНК, каждая из которых фланкирована правым краем и левым краем. Направление транскрипции в областях Т-ДНК показано стрелками. Описанные молекулы нуклеиновой кислоты могут содержать последовательности ДНК, расположенные с образованием моноцистронной или полицистронной конфигурации. Предпочтительные конфигурации включают конфигурацию, в которой первая совокупность последовательностей ДНК и вторая совокупность последовательностей ДНК расположены в одной области Т-ДНК.
В каждом иллюстрированном варианте осуществления изобретения первая совокупность последовательностей ДНК включает одну или несколько последовательностей, которые в случае экспрессии способны избирательно уменьшать содержание одного, двух или всех белков и транскриптов, кодированных геном, выбираемым из группы, включающей FAD2, FAD3 и FATB. Каждая последовательность в первой совокупности последовательностей ДНК предпочтительно способна в случае экспрессии подавлять экспрессию другого гена, включающего, не ограничиваясь ими, других членов семейства генов. Данные последовательности могут включать кодирующие последовательности, последовательности интронов, последовательности 3'-UTR, последовательности 5'-UTR, другие некодирующие последовательности и любую комбинацию указанных последовательностей. Первая совокупность последовательностей ДНК может быть экспрессирована в любой приемлемой форме, например в виде конструкции дцРНК, смысловой косупрессорной конструкции или антисмысловой конструкции. Первая совокупность последовательностей ДНК функционально связана по крайней мере с одним промотором, который вызывает экспрессию последовательностей и может быть любым промотором, функционирующим в растении, или любым растительным промотором. Предпочтительные промоторы включают, не ограничиваясь ими, промотор напина, промотор 7Sα, промотор 7Sα', промотор арцелина или промотор FAD2-1A.
Вторая совокупность последовательностей ДНК включает кодирующие последовательности, каждая из которых является последовательностью ДНК, кодирующей последовательность, которая в случае экспрессии способна увеличивать содержание белка или транскрипта либо как белка, так и транскрипта, кодированных геном, выбираемым из группы, включающей KAS I, KAS IV, дельта-9-десатуразу и СР4 EPSPS. Каждая кодирующая последовательность ассоциирована с промотором, который может быть любым промотором, функционирующим в растении, или любым растительным промотором. Предпочтительными промоторами, пригодными для использования во второй совокупности последовательностей ДНК, являются промотор FMV и/или семяспецифичные промоторы. Особенно предпочтительные семяспецифичные промоторы включают, не ограничиваясь ими, промотор напина, промотор 7Sα, промотор 7Sα', промотор арцелина, промотор дельта-9-десатуразы или промотор FAD2-1A.
В вариантах осуществления изобретения, показанных на фиг.1 и 2, первая совокупность последовательностей ДНК в случае экспрессии может образовывать молекулу дцРНК, способную подавлять экспрессию белка или транскрипта либо как белка, так и транскрипта, кодированных геном или транскрибированных из гена, выбираемого из группы, включающей FAD2, FAD3 и FATB. Первая совокупность последовательностей ДНК, показанная на фиг.1, включает три некодирующие последовательности, каждая из которых экспрессирует последовательность РНК (не показана), идентичную некодирующей области гена, выбираемого из группы, включающей гены FAD2, FAD3 и FATB. Некодирующие последовательности экспрессируют последовательность РНК, которая по крайней мере на 90% идентична некодирующей области гена, выбираемого из группы, включающей гены FAD2, FAD3 и FATB. Первая совокупность последовательностей ДНК включает также три антисмысловые последовательности, каждая из которых экспрессирует антисмысловую последовательность РНК (не показана), способную образовывать молекулу двухцепочечной РНК с соответствующей последовательностью РНК (экспрессированной некодирующими последовательностями).
Некодирующие последовательности могут быть отделены от антисмысловых последовательностей спейсерной последовательностью, предпочтительно последовательностью, которая стимулирует образование молекулы дцРНК. Примеры таких спейсерных последовательностей включают последовательности, представленные в публикациях Wesley et al., Plant J., 27(6):581-90 (2001) и Hamilton et al., Plant J., 15:737-746 (1988). В предпочтительном варианте осуществления изобретения спейсерная последовательность способна образовывать шпилькообразную структуру, представленную в приведенной выше публикации Wesley et al. Особенно предпочтительными спейсерными последовательностями в данном контексте являются растительные интроны или их части. Особенно предпочтительным растительным интроном является сплайсируемый интрон. Сплайсируемые интроны включают, не ограничиваясь ими, интрон, выбираемый из группы, включающей интрон PDK, интрон №5 FAD3-1A или FAD3-IB, интрон №1 FAD3, интрон №3А FAD3, интрон №3В FAD3, интрон №3С FAD3, интрон №4 FAD3, интрон №5 FAD3, интрон №1 FAD2 и интрон FAD2-2. Предпочтительные сплайсируемые интроны включают, не ограничиваясь ими, интрон, выбираемый из группы, включающей интрон №1 FAD3, интрон №3А FAD3, интрон №3В FAD3, интрон №3С FAD3 и интрон №5 FAD3. Другие предпочтительные сплайсируемые интроны включают, не ограничиваясь ими, сплайсируемый интрон длиной от около 0,75 т.п.о. до около 1,1 т.п.о, способный облегчать образование шпилькообразной структуры РНК. Одним неограничивающим примером особенно предпочтительного сплайсируемого интрона является интрон №5 FAD3.
Некодирующие молекулы со смысловой ориентацией могут быть необязательно отделены от соответствующих молекул с антисмысловой ориентацией спейсерным сегментом ДНК. Спейсерный сегмент может быть фрагментом гена или искусственной ДНК. Спейсерный сегмент может быть коротким для облегчения образования шпилькообразной структуры дцРНК или длинным для облегчения образования дцРНК без шпилькообразной структуры. Спейсер может быть получен путем удлинения цепи одной из смысловых или антисмысловых молекул, описанных в заявке на патент США 2005/0176670 А1. Альтернативно может быть создана последовательность с ориентацией правый край-правый край (“RB-RB”) после введения в геном растения, как описано в заявке на патент США 2005/0183170.
Как показано на фиг.1, молекула нуклеиновой кислоты включает две области Т-ДНК, каждая из которых фланкирована правым краем и левым краем. Первая область Т-ДНК включает первую совокупность последовательностей ДНК, функционально связанную с промотором, при этом первая область Т-ДНК дополнительно включает первую часть второй совокупности последовательностей ДНК, которая содержит первый промотор, функционально связанный с первой кодирующей последовательностью, и второй промотор, функционально связанный со второй кодирующей последовательностью. Вторая область Т-ДНК включает вторую часть второй совокупности последовательностей ДНК, которая содержит третий промотор, функционально связанный с третьей кодирующей последовательностью. В предпочтительном варианте осуществления изобретения, показанном на фиг.2, молекула нуклеиновой кислоты включает первую область Т-ДНК, которая фланкирована правым краем и левым краем. Первая и вторая совокупности последовательностей ДНК расположены в одной области Т-ДНК.
В вариантах осуществления изобретения с экспрессией дцРНК, показанных на фиг.1 и 2, порядок последовательностей может быть изменен по сравнению с иллюстрированным и описанным порядком, однако некодирующие последовательности и антисмысловые последовательности предпочтительно расположены вокруг спейсерной последовательности таким образом, что в случае экспрессии первая некодирующая последовательность может гибридизироваться с первой антисмысловой последовательностью, вторая некодирующая последовательность может гибридизироваться со второй антисмысловой последовательностью и третья некодирующая последовательность может гибридизироваться с третьей антисмысловой последовательностью таким образом, что может быть образована одна молекула дцРНК. Некодирующие последовательности предпочтительно имеют смысловую ориентацию, и антисмысловые последовательности имеют антисмысловую ориентацию относительно промотора. Число некодирующих, антисмысловых и кодирующих последовательностей и их относительные положения в одной или нескольких областях Т-ДНК могут быть также изменены любым образом, приемлемым для достижения целей настоящего изобретения.
Как показано на фиг.3 и 4, иллюстрированная молекула нуклеиновой кислоты включает область Т-ДНК, фланкированную правым краем и левым краем, у которых расположены первая и вторая совокупности последовательностей ДНК. Первая совокупность последовательностей ДНК функционально связана с промотором и сигналом терминации транскрипции. Вторая совокупность последовательностей ДНК включает первый промотор, функционально связанный с первой кодирующей последовательностью, второй промотор, функционально связанный со второй кодирующей последовательностью, и третий промотор, функционально связанный с третьей кодирующей последовательностью. Сигнал терминации транскрипции может быть любым сигналом терминации транскрипции, функционирующим в растении, или любым растительным сигналом терминации транскрипции. Предпочтительные сигналы терминации транскрипции включают, не ограничиваясь ими, 3'-концевую последовательность гороха Rubisco Е9, 3'-концевую последовательность напина Brassica, 3'-концевую последовательность tml и 3'-концевую последовательность nos.
В варианте осуществления изобретения, показанном на фиг.3, первая совокупность последовательностей ДНК в случае экспрессии может образовывать смысловую косупрессорную конструкцию, способную подавлять экспрессию одного или нескольких белков или транскриптов, кодированных или выделенных из гена, выбираемого из группы, включающей FAD2, FAD3 и FATB. Первая совокупность последовательностей ДНК включает три некодирующие последовательности, каждая из которых экспрессирует последовательность РНК (не показана), достаточно идентичную одной или нескольким некодирующими областям гена, выбираемого из группы, включающей гены FAD2, FAD3 и FATB. Некодирующие последовательности экспрессируют последовательность РНК, которая по крайней мере на 90% идентична одной или нескольким некодирующим областям гена, выбираемого из группы, включающей гены FAD2, FAD3 и FATB. Порядок расположения некодирующих последовательностей в первой совокупности последовательностей ДНК может быть изменен по сравнению с иллюстрированным и описанным в настоящем описании изобретения, но некодирующие последовательности имеют смысловую ориентацию относительно промотора.
На фиг.4 показан вариант осуществления изобретения, в котором первая совокупность последовательностей ДНК в случае экспрессии может образовывать антисмысловую конструкцию, способную подавлять экспрессию одного или нескольких белков или транскриптов, кодированных или выделенных из гена, выбираемого из группы, включающей FAD2, FAD3 и FATB. Первая совокупность последовательностей ДНК включает три антисмысловые последовательности, каждая из которых экспрессирует антисмысловую последовательность РНК (не показано), идентичную одной или нескольким некодирующим областям гена, выбираемого из группы, включающей гены FAD2, FAD3 и FATB. Антисмысловые последовательности экспрессируют антисмысловую последовательность РНК, которая по крайней мере на 90% идентична одной или нескольким некодирующим областям гена, выбираемого из группы, включающей гены FAD2, FAD3 и FATB. Порядок расположения антисмысловых последовательностей в первой совокупности последовательностей ДНК может быть изменен по сравнению с иллюстрированным и описанным в настоящем описании изобретения, но антисмысловые последовательности имеют антисмысловую ориентацию относительно промотора.
Описанные выше молекулы нуклеиновой кислоты являются предпочтительными, так как они обеспечивают достижение целей, обладают отличительными признаками и преимуществами настоящего изобретения. Не следует считать, что настоящее изобретение ограничено иллюстрированными вариантами осуществления изобретения. Расположение последовательностей в первой и второй совокупностях последовательностей ДНК в молекуле нуклеиновой кислоты не ограничено иллюстрированными и описанными расположениями и может быть изменено любым образом, приемлемым для достижения целей и обеспечения отличительных признаков и преимуществ настоящего изобретения, рассмотренных в настоящем описании изобретения и иллюстрированных на прилагаемых чертежах.
Е. Трансгенные организмы и способы их получения
Любые молекулы и конструкции нуклеиновой кислоты по настоящему изобретению могут быть постоянно или временно введены в растение или растительную клетку. Предпочтительные молекулы и конструкции нуклеиновой кислоты по настоящему изобретению описаны выше в частях А-D подробного описания изобретения и в примерах. Другой вариант осуществления настоящего изобретения относится к способу создания трансгенных растений, который обычно включает стадии отбора приемлемого растения или растительной клетки, трансформации растения или растительной клетки рекомбинантным вектором и получения трансформированной клетки-хозяина.
В предпочтительном варианте осуществления изобретения растение или клетка, выделенная из растения, продуцирует растительные масла для пищевого и промышленного применения. Особенно предпочтительными являются масличные культуры умеренного пояса. Представляющие интерес растения включают, не ограничиваясь ими, рапс (канола и сорта с высоким содержанием эруковой кислоты), кукуруза, соя, крамб, горчица, клещевина обыкновенная, арахис, кунжут, хлопчатник, лен, сафлор, масличная пальма, льнянка, подсолнечник и кокосовая пальма. Настоящее изобретение в равной степени относится к однодольным или двудольным видам и применимо к новым и/или усовершенствованным методам трансформации и регуляции.
Способы и методы введения ДНК в растительные клетки хорошо известны специалистам в данной области, и в настоящем изобретении могут быть использованы фактически любые методы введения молекул нуклеиновой кислоты в клетку. Неограничивающие примеры приемлемых методов включают: химические методы; физические методы, такие как микроинъекция, электропорация, выстреливание генов, бомбардировка микрочастицами и вакуумная инфильтрация; вирусные векторы и опосредуемые рецепторами механизмы. Могут быть использованы другие методы трансформации клетки, которые включают, не ограничиваясь ими, введение ДНК в растения путем прямого переноса ДНК в пыльцу, прямого инъецирования ДНК в репродуктивные органы растения или прямого инъецирования ДНК в клетки незрелых зародышей с последующей регидратацией высушенных зародышей.
Широко применяемой системой для введения генов в растительные клетки является перенос, опосредуемый Agrobacterium. См., например, публикации Fraley et al., Bio/Technology 3:629-635 (1985); Rogers et al., Methods Enzymol. 153:253-277 (1987). Переносимая область ДНК определяется краевыми последовательностями, при этом в геном растения обычно вводят проникающую ДНК. Spielmann et al., Mol. Gen. Genet. 205:34 (1986). Современные трансформирующие векторы на основе Agrobacterium могут реплицировать в E. coli, а также в Agrobacterium, обеспечивая удобное манипулирование. Klee et al., In: Plant DNA Infectious Agents, Hohn and Schell (eds.), Springer-Verlag, New York, pp. 179-203 (1985).
Регенерация, развитие и культивирование растений из отдельных трансформантов протопласта растения и из разных трансформированных эксплантатов хорошо известны в данной области. См. публикации Maliga et al., Methods in Plant Molecular Biology, Cold Spring Harbor Press (1995); Weissbach and Weissbach, In: Methods for Plant Molecular Biology, Academic Press, San Diego, CA (1988). Растения по настоящему изобретению могут быть частью программы селекции и могут быть также репродуцированы путем апомиксиса. Методы получения апомиктических растений хорошо известны в данной области. См., например, патент США № 5811636.
В предпочтительном варианте осуществления изобретения растение по настоящему изобретению, содержащее последовательности нуклеиновой кислоты, которые в случае экспрессии способны избирательно снижать уровни белка FAD2, FAD3 и/или FATB и/или транскрипта FAD2, FAD3 и/или FATB, скрещивают с другим растением по настоящему изобретению, содержащим последовательности нуклеиновой кислоты, которые в случае экспрессии способны сверхэкспрессировать другой фермент. Вышеуказанный другой фермент предпочтительно выбирают из группы, включающей бета-кетоацил-АСР-синтазу I, бета-кетоацил-АСР-синтазу IV, дельта-9-десатуразу и СР4 EPSPS.
В другом варианте осуществления изобретения растение по настоящему изобретению может быть скрещено с другим трансгенным или нетрансгенным растением. Растение по настоящему изобретению может быть скрещено с другим растением, в котором состав масла характеризуется модифицированными уровнями жирных кислот, например, в качестве неограничивающего примера можно привести сорт с пониженным уровнем линоленовой кислоты в составе масла. В предпочтительном варианте осуществления изобретения растение по настоящему изобретению скрещивают с сортом, содержащим менее 3 мас.% линоленовой кислоты, или в другом варианте осуществления изобретения растение по настоящему изобретению скрещивают с другим растением, содержащим более 20 мас.% стеариновой кислоты. Растения с модифицированными уровнями жирных кислот известны в данной области и описаны, например, в публикации Hawkins and Kridl (1998) Plant Journal 13(6):743-752 и в патенте США № 6365802.
F. Продукты по настоящему изобретению
Растения по настоящему изобретению могут быть использованы полностью или частично. Предпочтительные части растения включают репродуктивные или запасающие части. Термин “части растения” в используемом здесь значении включает, не ограничиваясь ими, семя, эндосперм, семяпочку, пыльцу, корни, клубни, стебли, листья, побеги, фрукты, ягоды, орехи, кору, стручки, семена и цветы. В особенно предпочтительном варианте осуществления настоящего изобретения частью растения является семя.
Любые растения или их части по настоящему изобретению могут быть подвергнуты обработке для получения корма, муки, белка или масляных продуктов. В предпочтительном варианте осуществления настоящего изобретения используют растение по настоящему изобретению, содержащее масло с составом жирных кислот по настоящему изобретению. Особенно предпочтительной частью растения для указанной цели является семя. В предпочтительном варианте осуществления изобретения корм, мука, белок или масляный продукт предназначены для питания скота, рыбы или человека. В данной области известны методы получения корма, муки, белка и масляных продуктов. См., например, патенты США № 4957748, 5100679, 5219596, 5936069, 6005076, 6146669 и 6156227. В предпочтительном варианте осуществления изобретения белковый продукт является продуктом с высоким содержанием белка. Такой продукт с высоким содержанием белка предпочтительно содержит белок в количестве более 5% мас./об., более предпочтительно в количестве 10% мас./об. и еще предпочтительнее в количестве 15% мас./об.
В предпочтительном варианте осуществления изобретения масляный продукт является продуктом с высоким содержанием масла, в котором содержание масла, полученного из растения или его части по настоящему изобретению, превышает 5% мас./об., более предпочтительно 10% мас./об. и еще предпочтительнее 15% мас./об. В предпочтительном варианте осуществления изобретения масляный продукт является жидкостью с объемом более 1, 5, 10 или 50 литров. Настоящее изобретение относится к маслу, полученному из растений по настоящему изобретению или способом по настоящему изобретению. Такое масло может характеризоваться повышенной устойчивостью к окислению. Кроме того, такое масло может быть второстепенным или главным компонентом любого полученного продукта.
Кроме того, такое масло может быть смешано с другими маслами. В предпочтительном варианте осуществления изобретения масло, полученное из растений по настоящему изобретению или способом по настоящему изобретению, составляет более 0,5%, 1%, 5%, 10%, 25%, 50%, 75% или 90% от объема или массы масляного компонента любого продукта. В другом варианте осуществления изобретения масляный продукт может быть смешанным и может составлять более 10%, 25%, 35%, 50% или 75% от объема смеси. Масло, полученное из растения по настоящему изобретению, может быть смешано с одним или несколькими органическими растворителями или нефтяными дистиллятами.
Семена растений могут быть помещены в контейнер. В используемом здесь значении контейнер является любым объектом, пригодным для хранения таких семян. Контейнер предпочтительно содержит больше примерно 500, 1000, 5000 или 25000 семян, из которых по крайней мере около 10%, 25%, 50%, 75% или 100% семян получены из растения по настоящему изобретению. Настоящее изобретение относится также к контейнеру, рассчитанному более чем примерно на 10000, более предпочтительно примерно на 20000 и еще предпочтительнее примерно на 40000 семян, из которых больше примерно 10%, более предпочтительно примерно 25%, более предпочтительно 50% и еще предпочтительнее примерно 75% или 90% семян являются семенами, полученными из растения по настоящему изобретению. Настоящее изобретение относится также к контейнеру, содержащему больше примерно 10 кг, более предпочтительно примерно 25 кг и еще предпочтительнее примерно 50 кг семян, при этом больше примерно 10%, более предпочтительно примерно 25%, более предпочтительно примерно 50% и еще предпочтительнее примерно 75% или 90% семян являются семенами, полученными из растения по настоящему изобретению.
G. Составы масла
Для многих применений масла содержание насыщенных жирных кислот предпочтительно составляет менее 8 мас.% и более предпочтительно около 2-3 мас.%. Насыщенные жирные кислоты имеют высокие температуры плавления, которые нежелательны во многих применениях. При использовании в качестве корма или топлива насыщенные жирные кислоты вызывают помутнение при низких температурах и сообщают топливу плохие свойства текучести на холоде, такие как потеря текучести и забивание фильтра в холодном состоянии. Масляные продукты с низким содержанием насыщенных жирных кислот являются более предпочтительными для потребителей и пищевой промышленности, так как они считаются более полезными и/или могут быть маркированы как “продукты без насыщенных жиров” в соответствии с правилами FDA. Кроме того, масла с низким содержанием насыщенных жирных кислот уменьшают или устраняют необходимость удаления твердых компонентов из масла путем охлаждения для использования в пище, например, в качестве масла для заправки салатов. В применениях, связанных с биологическим дизельным топливом и смазочными маслами, масло с низким содержанием насыщенных жирных кислот сообщает улучшенные свойства текучести на холоде и не мутнеет при низких температурах.
Факторы, определяющие физические свойства конкретного масла, носят сложный характер. Пальмитиновая, стеариновая и другие насыщенные жирные кислоты обычно являются твердыми при комнатной температуре в отличие от ненасыщенных жирных кислот, которые остаются жидкими. Так как насыщенные жирные кислоты не имеют двойных связей в ацильной цепи, они остаются устойчивыми к окислению при повышенных температурах. Насыщенные жирные кислоты являются важными компонентами в составе маргаринов и шоколада, но для многих применений в пищевой промышленности желательны более низкие уровни насыщенных жирных кислот.
Олеиновая кислота имеет одну двойную связь, но остается относительно устойчивой при высоких температурах, и масла с высокими уровнями олеиновой кислоты пригодны для приготовления пищи и других применений, требующих нагревания. Недавно было рекомендовано употреблять больше масла с высоким содержанием олеиновой кислоты, так как олеиновая кислота, по-видимому, снижает в крови уровни липопротеинов низкой плотности (“LDL”), не влияя на уровни липопротеинов высокой плотности (“HDL”). Однако желательно несколько ограничивать содержание олеиновой кислоты, так как, разлагаясь при высоких температурах, олеиновая кислота образует соединения с неприятными запахами и уменьшает количество соединений с приятными запахами, образующихся в результате окисления линолевой кислоты. Neff et al., JAOCS, 77:1303-1313 (2000); Warner et al., J. Agric. Food Chem. 49:899-905 (2001). Предпочтительные масла содержат олеиновую кислоту в количестве 65-85 мас.% или меньше, что ограничивает образование неприятных запахов при приготовлении пищи, например у масла для жарения и жареной пищи. Другие предпочтительные масла содержат олеиновую кислоту в количестве более 55 мас.% для улучшения устойчивости при окислении.
Линолевая кислота является главной полиненасыщенной жирной кислотой в пищевых продуктах и важным питательным веществом для человека. Линолевая кислота является желательным компонентом во многих применениях, связанных с приготовлением пищи, будучи главным предшественником веществ, вызывающих запах жареной пищи, таких как 2,4-декадиенал, которые придают хороший вкус жареным продуктам. Однако линолевая кислота характеризуется плохой устойчивостью при нагревании. В предпочтительных пищевых маслах содержание линолевой кислоты составляет 10 мас.% или больше для усиления образования веществ, придающих запах жареной пище, а также 25 мас.% или меньше для уменьшения образования неприятных запахов. Линолевая кислота обладает свойствами, снижающими уровень холестерина, хотя избыток линолевой кислоты в рационе питания может снизить способность человеческих клеток защищать себя от окисления, в результате чего повышается риск возникновения сердечно-сосудистых заболеваний. Toborek et al., Am. J. Clin. J. 75:119-125 (2002). См. публикацию Flavor Chemistry of Lipid Foods, editors D.B. Min & T.H. Smouse, Am. Oil Chem. Soc., Champaign, IL (1989).
Линолевая кислота, которая имеет более низкую температуру плавления, чем олеиновая кислота, далее способствует улучшению текучести на холоде, требуемому в применениях, связанных с биологическим дизельным топливом и биологическими смазочными маслами. Предпочтительные масла для большинства применений характеризуются содержанием линолевой кислоты, равным 30 мас.% или меньше, так как окисление линолевой кислоты ограничивает время хранения и применения масла для жарения, корма, пищевых продуктов, топлива и смазочных масел. См. публикации Physical Properties of Fats, Oils, and Emulsifiers, ed. N. Widlak, AOCS Press (1999); Erhan & Asadauskas, Lubricant Basestocks from Vegetable Oils, Industrial Crops and Products, 11:277-282 (2000). Кроме того, высокое содержание линолевого масла в корме для скота может стать причиной нежелательно высокого содержания линолевой кислоты в молоке молочного скота и, следовательно, плохой устойчивости к окислению и неприятного запаха. Timmons et al., J. Dairy Sci. 84:2440-2449 (2001). В широко используемом составе масла содержание линолевой кислоты составляют 10-25 мас.%.
Линоленовая кислота также является важным компонентом питания человека. Данная кислота используется для синтеза длинноцепочечных жирных кислот семейства ω-3 и выделяемых из них простагландинов. Однако двойные связи указанной кислоты весьма подвержены окислению, поэтому масла с высоким содержанием линоленовой кислоты быстро портятся под воздействием воздуха, особенно при высоких температурах. Часто необходимо частичное гидрирование таких масел до их использования в пищевых продуктах для замедления образования неприятных запахов и прогорклости при нагревании масла, но гидрирование вызывает образование вредных жирных транс-кислот, которые могут способствовать возникновению сердечно-сосудистых заболеваний. Для улучшения устойчивости к окислению и уменьшения потребности в гидрировании масла предпочтительные масла содержат линоленовую кислоту в количестве 8 мас.% или меньше, 6% или меньше, 4% или меньше, менее примерно 3% и более предпочтительно в количестве 0,5-2 мас.% от общего содержания жирных кислот в масле по настоящему изобретению.
Масло из сои по настоящему изобретению может быть также использовано в качестве источника смешивания для изготовления смешанного масляного продукта. Источник смешивания означает, что масло из сои по настоящему изобретению может быть смешано с другими растительными маслами для улучшения таких характеристик, как состав жирных кислот, запах и устойчивость к окислению других масел. Используемое количество масла из сои по настоящему изобретению зависит от требуемых свойств, которые должны быть получены в конечном смешанном масляном продукте. Примеры смешанных масляных продуктов включают, не ограничиваясь ими, маргарины, шортенинги, масла для жарения, масла для заправки салатов и т.д. Масло из сои по настоящему изобретению может быть смешанным маслом, синтезированным маслом или в предпочтительном варианте осуществления изобретения маслом, полученным из масличной культуры с соответствующим составом масла. Масло, полученное непосредственно из масличной культуры, является несмешанным маслом. В другом варианте осуществления изобретения масло получают непосредственно из созревшей масличной культуры. В одном варианте осуществления изобретения созревшая масличная культура определяется как культура, которую собирают на поле для коммерческих сельскохозяйственных целей, таких как продажа для приготовления пищевых продуктов. В предпочтительном варианте осуществления изобретения масло является соевым маслом. Масло может быть неочищенным маслом, например неочищенным соевым маслом, или может быть переработанным маслом, например масло может быть очищено, обесцвечено, дезодорировано и/или подвергнуто удалению твердых компонентов путем охлаждения. В используемом здесь значении термин “очистка” означает процесс обработки естественного или переработанного жира или масла для удаления загрязняющих примесей, при выполнении которого жир или масло обрабатывают каустиком, затем центрифугируют, промывают водой и нагревают в вакууме. “Обесцвечивание” означает процесс обработки жира или масла для удаления или уменьшения содержания в жире или масле окрашивающих веществ. Обесцвечивание может производиться путем обработки жира или масла активированным углем или диатомовой землей. “Дезодорация” означает процесс удаления из жира или масла компонентов, сообщающих конечному продукту неприятные запахи, который может быть выполнен при помощи высокого вакуума и промывки перегретым паром. “Удаление твердых компонентов путем охлаждения” означает процесс удаления из масла насыщенных глицеридов, который может быть выполнен путем охлаждения масла и удаления отвержденных частей жира.
Предпочтительное масло по настоящему изобретению характеризуется низким содержанием насыщенных жирных кислот или отсутствием насыщенных жирных кислот. В других предпочтительных вариантах осуществления изобретения масла по настоящему изобретению характеризуются повышенным содержанием олеиновой кислоты, пониженным содержанием насыщенных жирных кислот и пониженным содержанием полиненасыщенных жирных кислот. В других предпочтительных вариантах осуществления изобретения масла по настоящему изобретению характеризуются повышенным содержанием олеиновой кислоты и пониженным содержанием насыщенных жирных кислот. В предпочтительном варианте осуществления изобретения масло является соевым маслом. Процентное содержание жирных кислот или уровни жирных кислот выражены в массовых процентах.
В первом варианте осуществления изобретения масло по настоящему изобретению предпочтительно содержит 55-80% олеиновой кислоты, примерно 12-43% полиненасыщенных жирных кислот и 2-8% насыщенных жирных кислот; более предпочтительно масло содержит 55-80% олеиновой кислоты, примерно 14-42% полиненасыщенных жирных кислот и 3-6% насыщенных жирных кислот; и еще предпочтительнее масло содержит 55-80% олеиновой кислоты, примерно 16,5-43% полиненасыщенных жирных кислот и 2-3,6% насыщенных жирных кислот.
Во втором варианте осуществления изобретения масло по настоящему изобретению предпочтительно содержит 65-80% олеиновой кислоты, примерно 12-33% полиненасыщенных жирных кислот и 2-8% насыщенных жирных кислот; более предпочтительно масло содержит 65-80% олеиновой кислоты, примерно 14-32% полиненасыщенных жирных кислот и 3-6% насыщенных жирных кислот; еще предпочтительнее масло содержит 65-80% олеиновой кислоты, примерно 16,5-33% полиненасыщенных жирных кислот и 2-3,6% насыщенных жирных кислот.
В третьем варианте осуществления изобретения масло по настоящему изобретению предпочтительно содержит от около 42% до около 85% олеиновой кислоты и от около 8% до около 1,5% насыщенных жирных кислот; более предпочтительно масло содержит объединенное количество олеиновой кислоты и линоленовой кислоты от около 65% до около 95 мас.% от общего состава масла. Еще предпочтительнее масло по настоящему изобретению содержит объединенное количество олеиновой кислоты и линоленовой кислоты от около 75% до около 90%, от около 75% до около 95%, от около 75% до около 85%, от около 65% до около 90%, от около 70% до около 90 мас.% от общего состава масла.
В четвертом варианте осуществления изобретения масло по настоящему изобретению содержит от около 42% до около 85% олеиновой кислоты, от около 8% до около 1,5% насыщенных жирных кислот, от около 6% до около 15 мас.% линоленовой кислоты; более предпочтительно масло содержит от около 42% до около 85% олеиновой кислоты, от около 8% до около 1,5% насыщенных жирных кислот, менее 35 мас.% линоленовой кислоты; еще предпочтительнее масло содержит от около 42% до около 85% олеиновой кислоты, от около 8% до около 1,5% насыщенных жирных кислот, около 9 мас.% линоленовой кислоты.
В пятом варианте осуществления изобретения масло по настоящему изобретению содержит от около 50% до около 85% олеиновой кислоты и от около 8% до около 1,5% насыщенных жирных кислот; более предпочтительно от около 50% до около 85% олеиновой кислоты, от около 8% до около 1,5% насыщенных жирных кислот, от около 4% до около 14 мас.% линоленовой кислоты; более предпочтительно масло содержит от около 50% до около 85% олеиновой кислоты, от около 8% до около 1,5% насыщенных жирных кислот, менее 35 мас.% линоленовой кислоты; еще предпочтительнее масло содержит от около 42% до около 85% олеиновой кислоты, от около 8% до около 1,5% насыщенных жирных кислот, от около 2% до около 45 мас.% линоленовой кислоты.
В другом варианте осуществления изобретения масло по настоящему изобретению содержит около 65-80% олеиновой кислоты, около 3-8% насыщенных жирных кислот и около 12-32% полиненасыщенных жирных кислот. В другом варианте осуществления изобретения масло по настоящему изобретению содержит около 65-80% олеиновой кислоты, около 2-3,5% насыщенных жирных кислот и около 16,5-33% полиненасыщенных жирных кислот.
В особенно предпочтительном варианте осуществления изобретения масло по настоящему изобретению содержит около 47-83% олеиновой кислоты и около 5% насыщенных жирных кислот; около 60-80% олеиновой кислоты и около 5% насыщенных жирных кислот; около 50-85% олеиновой кислоты и около 2-7% насыщенных жирных кислот; около 55-85% олеиновой кислоты и около 2,5-7% насыщенных жирных кислот; около 47-88% олеиновой кислоты и около 3-7% насыщенных жирных кислот; около 43-85% олеиновой кислоты и около 5-7% насыщенных жирных кислот; около 81-85% олеиновой кислоты и около 5% насыщенных жирных кислот; около 74-83% олеиновой кислоты и около 6% насыщенных жирных кислот; около 65-87% олеиновой кислоты и около 6% насыщенных жирных кислот; около 66-80% олеиновой кислоты и около 6% насыщенных жирных кислот; около 42-77% олеиновой кислоты с около 5-8% насыщенных жирных кислот; около 60-77% олеиновой кислоты и около 6% насыщенных жирных кислот; около 70-81% олеиновой кислоты и около 5-7% насыщенных жирных кислот; около 52-71% олеиновой кислоты и около 5-7% насыщенных жирных кислот; около 44-71% олеиновой кислоты и около 6% насыщенных жирных кислот; около 61-71% олеиновой кислоты и около 8% насыщенных жирных кислот; около 57-71% олеиновой кислоты и около 7% насыщенных жирных кислот; около 23-58% олеиновой кислоты и около 8-14% насыщенных жирных кислот; около 20-70% олеиновой кислоты и около 6% насыщенных жирных кислот; около 21-35% олеиновой кислоты и около 5-6% насыщенных жирных кислот или около 19-28% олеиновой кислоты и около 5% насыщенных жирных кислот.
В других вариантах осуществления изобретения процентное содержание олеиновой кислоты составляет 50% или больше, 55% или больше, 60% или больше, 65% или больше, 70% или больше, 75% или больше или 80% или больше либо находится в диапазоне от 50% до 80%, от 55% до 80%, от 55% до 75%, от 55% до 65%, от 60% до 85%, от 60% до 80%, от 60% до 75%, от 60% до 70%, от 65% до 85%, от 65% до 80%, от 65% до 75%, от 65% до 70% или от 69% до 73%. Приемлемые диапазоны процентного содержания олеиновой кислоты в маслах по настоящему изобретению включают также диапазоны, в которых нижний предел выбирают из следующих процентных значений: 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79 или 80%; и верхний предел выбирают из следующих процентных значений: 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89 или 90%.
В указанных других вариантах осуществления изобретения процентное содержание линолевой кислоты в масле по настоящему изобретению находится в диапазоне от 10% до 40%, от 10% до 39%, от 10% до 30%, от 10% до 29%, от 10% до 28%, от 10% до 25%, от 10% до 21%, от 10% до 20%, от 11% до 30%, от 12% до 30%, от 15% до 25%, от 20% до 25%, от 20% до 30% или от 21% до 24%. Приемлемые диапазоны процентного содержания линолевой кислоты в маслах по настоящему изобретению включают также диапазоны, в которых нижний предел выбирают из следующих процентных значений: 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30%; и верхний предел выбирают из следующих процентных значений: 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39 или 40%.
В указанных других вариантах осуществления изобретения процентное содержание линоленовой кислоты в масле по настоящему изобретению составляет 10% или меньше, 9% или меньше, 8% или меньше, 7% или меньше, 6% или меньше, 5% или меньше, 4,5% или меньше, 4% или меньше, 3,5% или меньше, 3% или меньше, 2,5% или меньше, 2% или меньше; либо находится в диапазоне от 0,5% до 2%, от 0,5% до 3%, от 0,5% до 4,5%, от 0,5% до 6%, от 3% до 5%, от 3% до 6%, от 3% до 8%, от 1% до 2%, от 1% до 3% или от 1% до 4%. В указанных других вариантах осуществления изобретения процентное содержание насыщенных жирных кислот в составе масла по настоящему изобретению составляет 15% или меньше, 14% или меньше, 13% или меньше, 12% или меньше, 11% или меньше, 10% или меньше, 9% или меньше, 8% или меньше, 7% или меньше, 6% или меньше, 5% или меньше, 4% или меньше, 3,6% или меньше или находится в диапазоне от 2% до 3%, от 2% до 3,6%, от 2% до 4%, от 2% до 8%, от 3% до 15%, от 3% до 10%, от 3% до 8%, от 3% до 6%, от 3,6% до 7%, от 5% до 8%, от 7% до 10% или от 10% до 15%.
В других вариантах осуществления изобретения насыщенные жирные кислоты в масле по настоящему изобретению включают комбинацию пальмитиновой и стеариновой жирных кислот. В одном варианте осуществления изобретения процентное содержание насыщенных жирных кислот составляет около 10% или меньше, около 9% или меньше, около 8% или меньше, около 7% или меньше, около 6% или меньше, около 5% или меньше, около 4,5% или меньше, около 4% или меньше, около 3,5% или меньше, около 3% или меньше, около 2,5% или меньше; около 2% или меньше; либо находится в диапазоне от 0,5% до 2%, от 0,5% до 3%, от 0,5% до 4,5%, от 0,5% до 6%, от 0,5% до 7%, от 0,5% до 8%, от 0,5% до 9%, от 1% до 4%, от 1% до 5%, от 1% до 6%, от 1% до 7%, от 1% до 8%, от 1% до 9%, от 1,5% до 5%, от 1,5% до 6%, от 1,5% до 7%, от 1,5% до 8%, от 1,5% до 9%, от 2% до 5%, от 2% до 6%, от 2% до 7%, от 2% до 8%, от 2% до 9%, от 3% до 5%, от 3% до 6%, от 3% до 7%, от 3% до 8%, от 3% до 9%, от 4% до 7%, от 4% до 8%, от 4% до 9%, от 5% до 7%, от 5% до 8% и от 5% до 9%. В указанных вариантах осуществления изобретения приемлемые диапазоны процентного содержания насыщенных жирных кислот в маслах по настоящему изобретению включают также диапазоны, в которых нижний предел выбирают из следующих процентных значений: 0,5, 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 5,5, 6 или 6,5%; и верхний предел выбирают из следующих процентных значений: 11, 10, 9, 8, 7, 6, 5, 4,5, 4, 3,5, 3, 2,5, 2, 1,5, 1 или 0,5%.
В других вариантах осуществления изобретения процентное содержание пальмитиновой жирной кислоты в составе масла по настоящему изобретению составляет 6% или меньше, 5% или меньше, 4,5% или меньше, 4% или меньше, 3,5% или меньше, 3% или меньше, 2,5% или меньше, 2% или меньше или находится в диапазоне от 0,5% до 2%, от 0,5% до 3%, от 0,5% до 4,5%, от 0,5% до 6%, от 1% до 3%, от 1% до 4%, от 1% до 5%, от 1% до 6%, от 1,5% до 2%, от 1,5% до 3%, от 1,5% до 4%, от 1,5% до 4,5%, от 1,5% до 5,5%, от 1,5% до 6%, от 1,5% до 6,5%, от 1,5% до 7%, от 2% до 3%, от 2% до 3,5%, от 2% до 4%, от 2% до 4,5%, от 2% до 5%, от 2% до 6%, от 2% до 7%, от 2% до 8%, от 3% до 5%, от 3% до 6%, от 3% до 7%, от 3% до 8%, от 3% до 9%. В указанных вариантах осуществления изобретения приемлемые диапазоны процентного содержания линолевой кислоты в маслах по настоящему изобретению включают также диапазоны, в которых нижний предел выбирают из следующих процентных значений: 0,5, 1, 1,5, 2, 2,5, 3, 3,5, 4, 4,5, 5, 5,5, 6, 6,5, 7 или 7,5%; и верхний предел выбирают из следующих процентных значений: 11, 10, 9, 8, 7, 6, 5, 4,5, 4, 3,5, 3 или 2%.
В других вариантах осуществления изобретения процентное содержание стеариновой жирной кислоты в составе масла по настоящему изобретению составляет 3% или меньше, 2,5% или меньше, 2% или меньше или находится в диапазоне от 0,5% до 1%, от 0,5% до 1,5%, от 0,5% до 2%, от 0,5% до 2,5%, от 0,5% до 3%, от 0,5% до 4%, от 1% до 2%, от 1% до 3%, от 1% до 4%, от 1,5% до 2%, от 1,5% до 3% или от 1,5% до 4%. В указанных вариантах осуществления изобретения приемлемые диапазоны процентного содержания линолевой кислоты в маслах по настоящему изобретению включают также диапазоны, в которых нижний предел выбирают из следующих процентных значений: 0,5, 1, 1,5, 2, 2,5, 3 или 3,5%; и верхний предел выбирают из следующих процентных значений: 3,5, 3, 2,5, 2 или 1,5%.
Масло по настоящему изобретению особенно пригодно для использования в качестве масла для выпечки или жарения. Благодаря пониженному содержанию полиненасыщенных жирных кислот масло по настоящему изобретению не требует продолжительной обработки, необходимой для обычных масел, так как в указанном масле присутствует меньше соединений, вызывающих неприятный запах или цвет. Кроме того, низкое содержание насыщенных жирных кислот в масле по настоящему изобретению улучшает свойства текучести масла на холоде и устраняет необходимость нагревания хранящегося масла для предотвращения его кристаллизации или отверждения. Улучшенная текучесть на холоде также увеличивает стекание масла с жареных продуктов после их удаления из масла для жарения, что позволяет получить продукт с более низким содержанием жира. См. публикацию Bouchon et al., J. Food Science 66:918-923 (2001). Низкое содержание линоленовой кислоты в масле по настоящему изобретению является особенно благоприятным при жарении для уменьшения неприятных запахов.
Масло по настоящему изобретению хорошо подходит для использования в качестве масла для заправки салата (масло, которое остается прозрачным при температурах в холодильнике, равных 40-50°F). Улучшенная прозрачность при температурах в холодильнике благодаря низкому содержанию насыщенных жирных кислот и умеренному содержанию линолевой кислоты уменьшает или устраняет потребность в удалении из масла твердых компонентов путем охлаждения до использования в качестве масла для заправки салата.
Кроме того, умеренное содержание линолевой кислоты и низкое содержание линоленовой кислоты в масле по настоящему изобретению делает его пригодным для получения шортенинга, маргарина и других полутвердых растительных жиров, используемых в пищевых продуктах. Производство таких жиров обычно включает гидрирование ненасыщенных масел, таких как соевое масло, кукурузное масло или каноловое масло. Повышенная устойчивость масла по настоящему изобретению к окислению и запахам означает, что данное масло не требует гидрирования в такой же степени, что и обычное растительное масло, предназначенное для использования в качестве маргарина или шортенинга, что позволяет сократить производственные затраты и получение вредных транс-изомеров.
Масло по настоящему изобретению пригодно также для использования в качестве сырья для производства биологического дизельного топлива, в частности, благодаря тому, что биологическое дизельное топливо, изготовленное из такого масла, характеризуется улучшенной текучестью на холоде, лучшей воспламеняемостью (цетановое число), лучшей устойчивостью к окислению и меньшими эмульсиями оксида азота. Биологическое дизельное топливо является альтернативным дизельным топливом, обычно содержащим сложные метиловые эфиры насыщенных, мононенасыщенных или полиненасыщенных С16-С22 жирных кислот. Цетановое число является единицей измерения воспламеняемости - чем короче время задержки воспламенения топлива в двигателе, тем выше цетановое число. Стандарт ASTM для биологического дизельного топлива (D 6751-02) требует минимального цетанового числа, равного 47.
Применение биологического дизельного топлива в обычных дизельных двигателях значительно уменьшает количество загрязняющих веществ, таких как сульфаты, оксид углерода и частицы, по сравнению с нефтяным дизельным топливом, и его применение в школьных автобусах может значительно уменьшить воздействие на детей токсичных выхлопов дизельного топлива. Ограничением для применения 100% обычного биологического дизельного топлива является высокая температура помутнения стандартного биологического дизельного топлива из сои (2°С) по сравнению с дизельным топливом номер 2 (-16°С). Dunn et al., Recent. Res. Devel. in Oil Chem., 1:31-56 (1997). Биологическое дизельное топливо, изготовленное из масла по настоящему изобретению, характеризуется лучшей (более низкой) температурой помутнения и забивания фильтра в холодном состоянии и может быть также использовано в смесях для улучшения низкотемпературных свойств биологического дизельного топлива, изготовленного из недорогих, но высоконасыщенных источников жира, таких как животные жиры (говяжий жир, свиное топленое сало, куриный жир) и пальмовое масло. Биологическое дизельное топливо может быть также смешано с нефтяным дизельным топливом в любой пропорции.
Биологическое дизельное топливо обычно получают путем экстракции, фильтрования и очистки соевого масла для удаления свободных жиров и фосфолипидов с последующей переэтерификацией масла метанолом для образования сложных метиловых эфиров жирных кислот. См., например, патент США № 5891203. Полученные сложные метиловые эфиры соевого масла обычно определяются как “биологическое дизельное топливо”. Масло по настоящему изобретению может быть также использовано в качестве дизельного топлива без образования сложных метиловых эфиров, например, путем смешивания ацеталей с данным маслом. См., например, патент США № 6013114. Благодаря улучшенной текучести на холоде и устойчивости к окислению масло по настоящему изобретению может быть также использовано в качестве смазочного масла и присадки к дизельному топливу. См., например, патенты США № 5888947, 5454842 и 4557734.
Соевые бобы и масла по настоящему изобретению пригодны также для использования в разных соевых продуктах, изготовленных из цельных соевых бобов, таких как соевое молоко, соевое масло, натто и темпех, и в соевых продуктах, изготовленных из переработанных соевых бобов и соевого масла, включающих соевую крупу, соевую муку, концентрат соевого белка, вытяжки из соевого белка, текстурированный концентрат соевого белка, гидролизованный соевый белок, взбитую заправку, масло для выпечки, масло для заправки салата, шортенинг и лецитин. Цельные соевые бобы также являются съедобными и обычно продаются покупателям в сыром, жареном виде или как эдамаме (зеленые соевые бобы). Соевое молоко, которое обычно получают путем вымачивания и измельчения цельных соевых бобов, может быть использовано в том виде, как есть, высушено распылением или подвергнуто обработке для получения соевого йогурта, соевого сыра, тофу или юба. Соя или масло по настоящему изобретению могут быть успешно использованы в указанных и других соевых продуктах благодаря улучшенной устойчивости к окислению, меньшему содержанию предшественников веществ, вызывающих неприятные запахи, и низкому содержанию насыщенных жирных кислот.
G. Модуляция подавления
Другой вариант осуществления изобретения относится к способу модуляции уровней подавления гена. Модуляция подавления гена может вызывать большее или меньшее подавление гена. Подавление генного продукта может быть достигнуто в результате введения в геном растения конструкции по настоящему изобретению. Аналогичным образом подавление гена можно модулировать путем введения в геном растения конструкции по настоящему изобретению. Другие примеры методов модуляции подавления гена включают, не ограничиваясь ими, методы подавления антисмысловыми последовательностями, косупрессию, введение интерферирующей РНК (дцРНКи), использование трансгенных животных, создание гибридов и рибозим с помощью конструкции по настоящему изобретению. Нижеследующие примеры приведены в качестве иллюстрации и не ограничивают объем настоящего изобретения.
Подавление гена можно модулировать путем изменения длины транскрибируемой ДНК, используемой для подавления, последовательность которой выделена из гена, предназначенного для подавления. Для подавления гена при помощи механизмов посттранскрипционного сайленсинга гена могут быть использованы многие методы. Не ограничивая себя теорией, можно предположить, что указанные методы включают экспрессию молекулы РНК, которая гибридизуется с другой молекулой РНК. Весьма удивительным является то, что благоприятные результаты достигаются при использовании молекулы РНК определенной длины для модуляции или умеренного подавления уровня устойчивости экспрессии эндогенного гена-мишени.
Подавление гена FAD2-1 вызывает повышение уровней олеиновой кислоты и снижение уровней линолевой кислоты. При сильном подавлении FAD2-1 уровни олеиновой кислоты могут превышать 65%, что вызывает снижение уровней пальмитиновой кислоты и линоленовой кислоты. Например, при подавлении FAD2-1 уровни олеиновой кислоты могут достигать 85%, при этом объединенные уровни пальмитиновой и стеариновой кислот снижаются примерно до 10%. Аналогичным образом подавление FATB приводит к уменьшению уровней насыщенных жирных кислот, главным образом пальмитата. При подавлении FAD2 и FATB таким образом, что уровни олеиновой кислоты равны примерно 85%, содержание насыщенных жирных кислот составляет примерно 10%. При подавлении FAD2 и FATB таким образом, что уровни олеиновой кислоты превышают 85%, содержание насыщенных жирных кислот может быть ниже 10%.
В соответствии с настоящим изобретением уровни насыщенных жирных кислот могут быть снижены до менее 10% без увеличения содержания олеиновых кислот выше 85%. В одном варианте осуществления изобретения подавление FAD2 модулируют путем уменьшения длины интрона FAD2-1, введенного в растение. Меньшее подавление FAD2 соответствует средним уровням олеиновой кислоты, равным примерно 40-85% олеиновой кислоты. Подавление FAD2 снижается по мере уменьшения длины введенного фрагмента интрона FAD2-1. Например, интрон FAD2-1, длина которого уменьшена по крайней мере на 100 последовательных нуклеотидов, снижает подавление FAD2 и соответствующее увеличение уровня олеиновой кислоты и уменьшение уровней линолевой кислоты.
Взаимосвязь между снижением подавления эндогенного гена и уменьшением длины гомологичной ДНК можно определить эмпирически путем введения ДНК разной длины. Например, величину снижения подавления, достигаемую при уменьшении длины введенной гомологичной ДНК, можно определить путем удаления удлиняющих частей вводимой гомологичной ДНК и анализа экспрессии гена-мишени.
В объем настоящего изобретения входит способ умеренного подавления FAD2 при сильном снижении уровней насыщенных жирных кислот в растении. В таких растениях уровни олеиновой кислоты могут составлять 40-85%. Аналогичным образом неполное подавление FATB имеет место при введении объединенных 3'-концевых и 5'-концевых нетранслируемых областей по сравнению с введением в клетку-хозяина непроцессированного гена FATB. Аналогичным образом уровни подавления FATB снижаются при введении в клетку-хозяина 5'-концевой части открытой рамки считывания, которая кодирует в основном транзитный пептид хлоропласта. В клетках, содержащих FAD2 и FATB, подавляемые способами по настоящему изобретению, уровни олеиновой кислоты могут быть равны 40-85%, в то время как уровни насыщенных жирных кислот могут составлять 1-9%.
Один вариант осуществления настоящего изобретения относится к способу модуляции подавления гена, позволяющему уменьшить подавление по сравнению с подавлением генетического элемента, являющегося полным геном, полным экзоном, полным интроном, полной сигнальной последовательностью или полной UTR, который включает создание молекулы рекомбинантной нуклеиновой кислоты, содержащей фрагмент эндогенной последовательности из данного генетического элемента, инициацию экспрессии молекулы рекомбинантной нуклеиновой кислоты в клетке-хозяине и подавление эндогенного гена молекулой рекомбинантной нуклеиновой кислоты. Подавляемый ген может быть любым геном, включая FAD2 и FATB. Один вариант осуществления изобретения относится к способу модуляции подавления FAD2 или FATB, который включает: экспрессию неполной последовательности генетического элемента FAD2 или FATB в клетке-хозяине, при этом генетический элемент FAD2 или FATB получают из эндогенного гена FAD2 или FATB в клетке-хозяине, последовательность генетического элемента FAD2 или FATB может быть геном FAD2 или FATB, экзоном FAD2 или FATB, интроном FAD2 или FATB, кодирующей областью транзитного пептида FAD2 или FATB или UTR FAD2 или FATB, и неполная последовательность генетического элемента FAD2 или FATB меньше полной последовательности генетического элемента FAD2 или FATB; и подавление эндогенного гена FAD2 или FATB неполной последовательностью генетического элемента FAD2 или FATB, при этом уровни подавления эндогенного гена FAD2 или FATB в клетке-хозяине меньше уровней подавления эндогенного гена FAD2 или FATB в клетке-хозяине с подобной генетической средой и второй последовательностью нуклеиновой кислоты FAD2 или FATB, содержащей всю последовательность генетического элемента FAD2 или FATB.
Другой вариант осуществления настоящего изобретения относится к способу изменения состава масла растительной клетки, который включает трансформацию растительной клетки молекулой рекомбинантной нуклеиновой кислоты, содержащей последовательность ДНК, которая подавляет эндогенную экспрессию FAD2, FATB или FAD2 и FATB, при этом последовательность ДНК включает последовательность нуклеиновой кислоты FAD2, FATB или FAD2 и FATB, которая короче полной последовательности всего генетического элемента, выбираемого из гена, экзона, интрона, кодирующей области транзитного пептида, 3'-UTR, 5'-UTR и открытой рамки считывания; и выращивание растительной клетки в условиях инициации транскрипции указанной последовательности ДНК, в результате чего происходит изменение состава масла по сравнению с растительной клеткой, имеющей подобную генетическую среду, но не содержащей молекулу рекомбинантной нуклеиновой кислоты. Генетический элемент FAD2 или FATB может быть короче на 50, 75, 100, 150, 175, 200, 250, 300, 350, 400, 450, 500, 600, 800, 1000, 2000, 3000 или 4000 нуклеотидов. Длина генетического элемента FAD2 или FATB может быть равна 50, 75, 100, 150, 175, 200, 220, 250, 300, 320, 350, 400, 420, 450, 500, 550, 600, 800 или 1000 нуклеотидам.
Другой вариант осуществления настоящего изобретения относится к способу повышения содержания олеиновой кислоты и уменьшения содержания насыщенных жирных кислот в семени растения, который включает: i) укорачивание длины последовательности ДНК экзогенного гена FAD2 в клетке-хозяине так, чтобы по крайней мере частично сократить величину подавления экспрессии FAD2 в трансформированном растении по сравнению с подавлением экспрессии FAD2 в клетке-хозяине с подобной генетической средой и полной последовательностью ДНК экзогенного гена FAD2; и ii) выращивание растения с молекулой нуклеиновой кислоты, включающей укороченную последовательность ДНК FAD2, при этом укороченная последовательность ДНК FAD2 по крайней мере частично подавляет эндогенную экспрессию FAD2; и iii) культивирование растения, продуцирующего семя с пониженным содержанием насыщенных жирных кислот по сравнению с семенем растения, имеющего подобную генетическую среду, но не содержащего укороченную последовательность ДНК FAD2. Величина, на которую должна быть укорочена последовательность ДНК экзогенного гена FAD2, чтобы по крайней мере частично уменьшить подавление эндогенного гена FAD2, можно определить эмпирически путем введения ДНК разной длины. Например, величину уменьшения подавления, достигаемого в результате уменьшения длины введенной гомологичной ДНК, можно определить путем удаления удлиняющих частей вводимой гомологичной ДНК и анализа экспрессии гена-мишени. Величина подавления экспрессии FAD2 может быть определена в виде среднего значения для трех или более, шести или более, десяти или более, пятнадцати или более, двадцати или более семян растения.
Другой вариант осуществления настоящего изобретения относится к способу получения трансформированного растения, содержащего семя с пониженным содержанием насыщенных жирных кислот, который включает трансформацию растительной клетки молекулой рекомбинантной нуклеиновой кислоты, содержащей последовательность ДНК, подавляющую эндогенную экспрессию FAD2 и FATB, при этом последовательность ДНК содержит последовательность нуклеиновой кислоты FAD2, которая короче полной последовательности всего генетического элемента, выбираемого из гена, экзона, интрона, кодирующей области транзитного пептида и UTR; и выращивание трансформированного растения, образующего семя с пониженным содержанием насыщенных жирных кислот по сравнению с семенем растения, имеющего подобную генетическую среду, но не содержащего указанную молекулу рекомбинантной нуклеиновой кислоты.
Другой вариант осуществления настоящего изобретения относится к способу модуляции состава жирных кислот масла, полученного из семян масличной культуры умеренного пояса, который включает выделение генетического элемента длиной, равной по крайней мере 40 нуклеотидам, способного подавлять экспрессию эндогенного гена в пути синтеза жирных кислот; создание нескольких укороченных фрагментов генетического элемента; введение всех укороченных фрагментов в растительную клетку масличной культуры умеренного пояса с целью создания трансгенных растений и отбор трансгенного растения, содержащего укороченный фрагмент определенной длины и последовательность, вызывающую требуемое изменение состава жирных кислот в масле из семян. В предпочтительном варианте осуществления изобретения вышеуказанный способ включает также создание конструкции рекомбинантной ДНК, содержащей по крайней мере два укороченных фрагмента двух разных эндогенных генов, вызывающих разные требуемые изменения в составе жирных кислот масла из семян; введение конструкции рекомбинантной ДНК в растительную клетку масличной культуры умеренного пояса с целью создания трансгенных растений и отбор трансгенного растения, содержащего по крайней мере два укороченных фрагмента, в котором состав жирных кислот в масле из семян имеет несколько требуемых изменений, вызываемых по крайней мере двумя укороченными фрагментами.
Другой вариант осуществления настоящего изобретения относится к семени сои, в котором состав масла характеризуется значительно меньшим содержанием насыщенных жирных кислот и умеренно повышенным содержанием олеиновой кислоты и которое содержит последовательность ДНК, подавляющую эндогенную экспрессию FAD2 в клетке-хозяине, при этом последовательность ДНК имеет последовательность нуклеиновой кислоты FAD2, которая короче полной последовательности всего генетического элемента, выбираемого из гена, экзона, интрона, кодирующей области транзитного пептида и UTR.
Приведенные ниже примеры имеют иллюстративный характер и не ограничивают объем настоящего изобретения.
Все публикации, патенты и заявки на патент, приведенные в данном описании изобретения, включены в него в качестве ссылки, как если бы каждая отдельная публикация, патент или заявка на патент была отдельно включена в качестве ссылки.
ПРИМЕРЫ
Пример 1. Выделение последовательностей FATB-2
Ткань листа, взятую у сои Asgrow сорта А3244, измельчают в жидком азоте и хранят при -80°С до использования. Шесть мл натрий-додецилсульфатного (SDS) буфера для экстракции (650 мл стерильной ddH2O, 100 мл 1 М трис-Сl рН 8, 100 мл 0,25 М EDTA, 50 мл 20% SDS, 100 мл 5 М NaCl, 4 мкл бета-меркаптоэтанола) добавляют к 2 мл замороженной/измельченной ткани листа и смесь инкубируют при 65°С в течение 45 минут. Образец встряхивают через каждые 15 минут. К образцу добавляют 2 мл охлаждаемого льдом 5 М раствора ацетата калия, образец встряхивают и инкубируют на льду в течение 20 минут. К образцу добавляют 3 мл CHCl3 и встряхивают в течение 10 минут.
Образец центрифугируют со скоростью 10000 оборотов/мин в течение 20 минут и собирают супернатант. К супернатанту добавляют 2 мл изопропанола и смешивают. Образец центрифугируют со скоростью 10000 оборотов/мин в течение 20 минут и супернатант сливают. Осадок ресуспендируют в 200 мкл РНКазы и инкубируют при 65°С в течение 20 минут. Добавляют 300 мкл ацетата аммония/изопропанола (1:7) и смешивают. Затем образец центрифугируют со скоростью 10000 оборотов/мин в течение 15 минут и супернатант выбрасывают. Осадок промывают 500 мкл 80% этанола и подвергают воздушной сушке. Осадок геномной ДНК ресуспендируют в 200 мкл Т10Е1 (10 мМ трис:1 мМ EDTA).
Последовательность кДНК FATB-2 сои (SEQ ID NO:42) используют для создания тринадцати олигонуклеотидов, образующих данный ген: F1 (SEQ ID NO:48), F2 (SEQ ID NO:49), F3 (SEQ ID NO:50), F4 (SEQ ID NO:51), F5 (SEQ ID NO:52), F6 (SEQ ID NO:53), F7 (SEQ ID NO:54), R1 (SEQ ID NO:55), R2 (SEQ ID NO:56), R3 (SEQ ID NO:57), R4 (SEQ ID NO:58), R5 (SEQ ID NO:59) и R6 (SEQ ID NO:60). Указанные олигонуклеотиды используют попарно для амплификации методом ПЦР из выделенной геномной ДНК сои: пара 1 (F1 + R1), пара 2 (F2 + R1), пара 3 (F3 + R2), пара 4 (F4 + R3), пара 5 (F5 + R4), пара 6 (F6 + R5) и пара 7 (F7 + R6). Амплификацию методом ПЦР для пары 5 выполняют следующим образом: 1 цикл, 95°С в течение 10 минут; 30 циклов, 95°С в течение 15 секунд, 43°С в течение 30 секунд, 72°С в течение 45 секунд; 1 цикл, 72°С в течение 7 минут. Для всех других пар олигонуклеотидов амплификацию методом ПЦР выполняют следующим образом: 1 цикл, 95°С в течение 10 минут; 30 циклов, 95°С в течение 15 секунд, 48°С в течение 30 секунд, 72°С в течение 45 секунд; 1 цикл, 72°С в течение 7 минут. Положительные фрагменты получают при использовании пар праймеров 1, 2, 4, 5, 6 и 7. Каждый фрагмент клонируют в векторе рCR2.1 (Invitrogen). Фрагменты 2, 4, 5 и 6 подтверждают и секвенируют. Четыре последовательности объединяют с образованием геномной последовательности для гена FATB-2 (SEQ ID NO:43).
При сравнении геномной последовательности с последовательностью кДНК в гене FATB-2 сои идентифицированы четыре интрона: интрон I (SEQ ID NO:44) расположен от основания 119 до основания 1333 геномной последовательности (SEQ ID NO:43) и имеет длину, равную 1215 п.о.; интрон II (SEQ ID NO:45) расположен от основания 2231 до основания 2568 геномной последовательности (SEQ ID NO:43) и имеет длину, равную 338 п.о.; интрон III (SEQ ID NO:46) расположен от основания 2702 до основания 3342 геномной последовательности (SEQ ID NO:43) и имеет длину, равную 641 п.о.; и интрон IV (SEQ ID NO:47) расположен от основания 3457 до основания 3823 геномной последовательности (SEQ ID NO:43) и имеет длину, равную 367 п.о.
Пример 2. Супрессорные конструкции
2А. Конструкции FAD2-1
Интрон № 1 FAD2-1A (SEQ ID NO:1) клонируют в полигенном экспрессирующем кластере рСGN3892 в смысловой и антисмысловой ориентации. Вектор pCGN3892 содержит промотор 7S сои и 3'-конец rbcS гороха. Оба слитых гена затем отдельно лигируют с вектором pCGN9372, который содержит ген CP4 EPSPS, регулируемый промотором FMV. Полученные экспрессирующие конструкции (смысловая конструкция pCGN5469 и антисмысловая конструкция pCGN5471) используют для трансформации сои.
Интрон FAD2-1B (SEQ ID NO:2) сливают с 3'-концом интрона № 1 FAD2-1A в плазмиде pCGN5468 (содержит промотор 7S сои, слитый с интроном FAD2-1A (смысловой), и 3'-конец rbcS гороха) или в плазмиде pCGN5470 (содержит промотор 7S сои, слитый с интроном FAD2-1A (антисмысловой), и 3'-концом rbc гороха) соответственно в смысловой и антисмысловой ориентации. Полученные слитые комбинации интронов затем отдельно лигируют с вектором pCGN9372, который содержит ген СР4 EPSPS, регулируемый промотором FMV. Полученные экспрессирующие конструкции (pCGN5485, смысловой интрон FAD2-1A, FAD2-1B и pCGN5486, антисмысловой интрон FAD2-1A и FAD2-1B) используют для трансформации сои.
2В. Конструкции FAD3-1
Интроны №1, №2, №4 и №5 (соответственно SEQ ID NO:7, 8, 10 и 11) FAD3-1A, интроны №3С (SEQ ID NO:23) и №4 (SEQ ID NO:24) FAD3-1B отдельно лигируют с pCGN3892 в смысловой или антисмысловой ориентации. pCGN3892 содержит промотор 7S сои и 3'-конец rbcS гороха. Указанные слитые интроны лигируют с вектором pCGN9372, который содержит ген СР4 EPSPS, регулируемый промотором FMV, для введения в сою. Полученные экспрессирующие конструкции (pCGN5455, смысловой интрон № 4 FAD3-1А; pСGN5459, антисмысловой интрон №4 FAD3-1A; pCGN5456, смысловой интрон №5 FAD3; pCGN5460, антисмысловой интрон №5 FAD3-1A; pCGN5466, антисмысловой интрон №2 FAD3-1A; pCGN5473, антисмысловой интрон №1 FAD3-1A) используют для трансформации сои.
2С. Конструкции FATB
Последовательность интрона II FATB сои (SEQ ID NO:30) амплифицируют методом ПЦР, используя неполный геномный клон FATB-1 в качестве матрицы. Амплификацию методом ПЦР выполняют следующим образом: 1 цикл, 95°С в течение 10 минут; 25 циклов, 95°С в течение 30 секунд, 62°С в течение 30 секунд, 72°С в течение 30 секунд; 1 цикл, 72°С в течение 7 минут. В результате амплификации методом ПЦР получают продукт длиной 854 п.о., содержащий у обоих концов трансформированные сайты рестрикции. Продукт ПЦР клонируют непосредственно в полигенном экспрессирующем кластере pCGN3892 в смысловой ориентации, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР, с образованием pMON70674. Вектор pCGN3892 содержит промотор 7S сои и 3'-конец rbcS гороха. pMON70674 затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV. Полученную экспрессирующую ген конструкцию pMON70678 используют для трансформации сои методами с использованием Agrobacterium.
Создают две другие экспрессирующие конструкции, содержащие последовательность интрона II FATB-1 сои (SEQ ID NO:30). Вектор pMON70674 разрезают NotI и лигируют с вектором pMON70675, содержащим ген СР4 EPSPS, регулируемый промотором FMV, и ген KAS IV, регулируемый промотором напина, получая при этом pMON70680. Экспрессирующий вектор pMON70680 затем разрезают SnaBI и лигируют с геном, слитым с геном дельта-9-десатуразы гиогиоба (SEQ ID NO:41) в смысловой ориентации, регулируемым промотором 7S. Экспрессирующие конструкции pMON70680 и pMON70681 используют для трансформации сои методами с использованием Agrobacterium.
2D. Комбинированные конструкции
Создают экспрессирующие конструкции, содержащие разные пермутации первой совокупности последовательностей ДНК. Первая совокупность последовательностей ДНК включает любые описанные последовательности, или иллюстрированные на фиг.5 и 6, или любую другую совокупность последовательностей ДНК, содержащих разные комбинации некодирующих или кодирующих областей смысловых, антисмысловой или смысловых и антисмысловых последовательностей FAD2, FAD3 и FATB, способные образовывать конструкции дцРНК, смысловые косупрессорные конструкции, антисмысловые конструкции или разные комбинации вышеуказанных конструкций.
На фиг.5(а)-(с) показано несколько первых совокупностей последовательностей ДНК, способных экспрессировать смысловые косупрессорные или антисмысловые конструкции по настоящему изобретению, некодирующие последовательности которых представлены в приведенных ниже таблицах 1 и 2. Некодирующие последовательности могут представлять собой отдельные последовательности, комбинации последовательностей (например, 5'-UTR, связанная с 3'-UTR) или любую комбинацию вышеуказанных последовательностей. Для экспрессии смысловой косупрессорной конструкции все некодирующие последовательности являются смысловыми последовательностями, и для экспрессии антисмысловой конструкции все некодирующие последовательности являются антисмысловыми последовательностями. На фиг.5(d) показана первая совокупность последовательностей ДНК, которая способна экспрессировать смысловые и антисмысловые конструкции по настоящему изобретению.
На фиг.6(а)-(с) показано несколько первых совокупностей последовательностей ДНК, способных экспрессировать конструкции дцРНК по настоящему изобретению, некодирующие последовательности которых представлены в приведенных ниже таблицах 1 и 2. Первая совокупность последовательностей ДНК, показанная на фиг.6, содержит пары родственных смысловых и антисмысловых последовательностей, расположенных так, что, например, РНК, экспрессированная первой смысловой последовательностью, способна образовывать двухцепочечную РНК с антисмысловой РНК, экспрессированной первой антисмысловой последовательностью. Например, как показано на фиг.6(а) и в иллюстративной комбинации №1 (таблицы 1), первая совокупность последовательностей ДНК включает смысловую последовательность FAD2-1, смысловую последовательность FAD3-1, антисмысловую последовательность FAD2-1 и антисмысловую последовательность FAD3-1. Обе антисмысловые последовательности соответствуют смысловым последовательностям, поэтому продукты экспрессии первой совокупности последовательностей ДНК могут образовывать друг с другом двухцепочечную РНК. Смысловые последовательности могут быть отделены от антисмысловых последовательностей спейсерной последовательностью, предпочтительно последовательностью, которая стимулирует образование молекулы дцРНК. Примеры таких спейсерных последовательностей включают последовательности, представленные в приведенной выше публикации Wesley et al. и в публикации Hamilton et al., Plant J., 15:737-746 (1988). Промотор является любым промотором, функционирующим в растении, или любым растительным промотором. Неограничивающие примеры приемлемых промоторов приведены в части D подробного описания изобретения.
Первую совокупность последовательностей ДНК вводят в экспрессирующую конструкцию в смысловой или антисмысловой ориентации разными методами манипуляции ДНК. При наличии в последовательностях ДНК удобных сайтов рестрикции такие сайты вводят в экспрессирующую конструкцию путем расщепления рестрикционными эндонуклеазами и лигирования с конструкцией, расщепленной у одного или нескольких имеющихся сайтов клонирования. При отсутствии в последовательностях ДНК удобных сайтов рестрикции ДНК в конструкции или последовательности ДНК модифицируют разными способами для облегчения клонирования последовательностей ДНК в конструкции. Примеры методов модификации ДНК включают ПЦР, лигирование с синтетическим линкером или адаптером, сайтнаправленный мутагенез in vitro, удлинение или укорачивание выступающих 5'- или 3'-концов и тому подобные. Специалистам в данной области хорошо известны вышеуказанные и другие методы манипуляции ДНК.
Вектор pMON97552 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 140 последовательных нуклеотидов от 3'-конца, функционально связанного с 42 последовательными нуклеотидами 5'-UTR FATB-1α, за которой следует кодирующая область СТР FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с кодирующей областью СТР FATB-1α в антисмысловой ориентации, за которой следуют 42 последовательных нуклеотида 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 140 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB). Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON93758 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 160 последовательных нуклеотидов от 5'-конца и лигированным с 3'-UTR FATB-1α, за которой следует 5'-UTR FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует 3'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 160 последовательных нуклеотидов от 5'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON97553 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 200 последовательных нуклеотидов от 3'-конца и лигированным с 42 последовательными нуклеотидами 5'-UTR FATB-1α, за которой следует кодирующая область СТР FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с кодирующей областью СТР FATB-1α в антисмысловой ориентации, за которой следуют 42 последовательных нуклеотида 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A (SEQ ID NO:1), уменьшенный на 200 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON93770 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 240 последовательных нуклеотидов от 3'-конца и лигированным с 3'-UTR FATB-1α, за которой следует 5'-UTR FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует 3'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 240 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON93759 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 240 последовательных нуклеотидов от 5'-конца и лигированным с 3'-UTR FATB-1α, за которой следует 5'-UTR FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует 3'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 240 последовательных нуклеотидов от 5'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco E9, при этом все последовательности фланкированы правый краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON97554 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 260 последовательных нуклеотидов от 3'-конца и лигированным с 42 последовательными нуклеотидами 5'-UTR FATB-1α, за которой следует кодирующая область СТР FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с кодирующей областью СТР FATB-1α в антисмысловой ориентации, за которой следуют 42 последовательных нуклеотида 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 260 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON93771 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 300 последовательных нуклеотидов от 3'-конца и лигированным с 3'-UTR FATB-1α, за которой следует 5'-UTR FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует 3'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 300 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON97555 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с 42 последовательными нуклеотидами 5'-UTR FATB-1α, за которой следует кодирующая область СТР FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с кодирующей областью СТР FATB-1α в антисмысловой ориентации, за которой следуют 42 последовательных нуклеотида 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A (SEQ ID NO:1), уменьшенный на 320 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON93760 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 5'-конца и лигированным с 3'-UTR FATB-1α, за которой следует 5'-UTR FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует 3'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 320 последовательных нуклеотидов от 5'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON93772 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 360 последовательных нуклеотидов от 3'-конца и лигированным с 3'-UTR FATB-1α, за которой следует 5'-UTR FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с 5'-UTR FATB-1α в антисмысловой ориентации, за которой следует 3'-UTR FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 360 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON97556 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 380 последовательных нуклеотидов от 3'-конца и лигированным с 42 последовательными нуклеотидами 5'-UTR FATB-1α, за которой следует кодирующая область СТР FATB-1α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с кодирующей областью СТР FATB-1α в антисмысловой ориентации, за которой следуют 42 последовательных нуклеотида 5'-UTR FATB-1α в антисмысловой ориентации, функционально связанной с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 380 последовательных нуклеотидов от 3'-конца и находящимся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON93764 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 400 последовательных нуклеотидов от 3'-конца и лигированным с кодирующей областью СТР FATB-1α, за которой следует кодирующая область СТР FATB-2α, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с кодирующей областью СТР FATB-2α в антисмысловой ориентации, за которой следует кодирующая область СТР FATB-1α в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 400 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с кодирующей областью СТР FATB-2α в антисмысловой ориентации, за которой следуют 42 последовательных нуклеотида 5'-UTR FATB-2α в антисмысловой ориентации, функционально связанной с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Пример 3. Комбинированные конструкции
На фиг.7-15 промоторы показаны стрелками, кодирующие последовательности (как кодирующие, так и некодирующие) показаны пятиугольниками, обращенными в направлении транскрипции, смысловые последовательности обозначены обычным текстом и антисмысловые последовательности обозначены перевернутым текстом. На указанных фигурах использованы следующие аббревиатуры: 7Sa = промотор 7Sα; 7Sa' = промотор 7Sα'; Br napin = промотор напина Brassica; FMV = промотор FMV; ARC = промотор арцелина; RBC E9 3' = сигнал терминации Rubisco Е9; Nos 3' = сигнал терминации nos; TML 3' = сигнал терминации tml; napin 3'= сигнал терминации напина; 3' (в той же рамке, что FAD или FAT) = 3'-UTR; 5' (в той же рамке, что FAD или FAT) = 5'-UTR; Cr = Cuphea pulcherrima; Gm = Глицин макс.; Rc = Ricinus communis; FAD2 = аллель FAD2 гена дельта-9-стеароилдесатуразы; и Intr или Inr = интрон.
3А. Конструкции дцРНК
На фиг.7-9 показаны молекулы нуклеиновой кислоты по настоящему изобретению, в которых первые совокупности последовательностей ДНК могут экспрессировать конструкции дцРНК. Первая совокупность последовательностей ДНК, показанная на фиг.7-9, включает пары родственных смысловых и антисмысловых последовательностей, расположенные таким образом, что, например, РНК, экспрессированная первой смысловой последовательностью, может образовывать двухцепочечную РНК с антисмысловой РНК, экспрессированной первой антисмысловой последовательностью. Смысловые последовательности могут быть расположены рядом с антисмысловыми последовательностями или могут быть отделены от антисмысловых последовательностей спейсерной последовательностью, как показано на фиг.9.
Вторая совокупность последовательностей ДНК включает кодирующие последовательности, каждая из которых является последовательностью ДНК, кодирующей последовательность, которая в случае экспрессии может увеличивать белок или транскрипт либо как белок, так и транскрипт, кодированные геном, выбираемым из группы, включающей KAS I, KAS IV, дельта-9-десатуразу и СР4 EPSPS. Каждая кодирующая последовательность ассоциирована с промотором, который может быть любым промотором, функционирующим в растении, или любым растительным промотором и может быть промотором FMV, промотором напин, промотором 7S (7Sα или 7Sα'), промотором арцелин, промотором дельта-9-десатураза или промотором FAD2-1A.
Как показано на фиг.7, последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), 3'-UTR FAD3-1A (SEQ ID NO:16) или 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, которые включают повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1A сои (SEQ ID NO:11) в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Векторы, содержащие ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, и ген дельта-9-десатуразы (FAB2) R. communis (SEQ ID NO:40), регулируемый промотором FAD2 сои и 3'-концевой терминирующей последовательностью nos, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON68539, показанную на фиг.7, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), интрона 4 FAD3-1A (SEQ ID NO:10) и интрона II FATB-1 (SEQ ID NO:30) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1A сои (SEQ ID NO:11) в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON68540, показанную на фиг.7, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), интрона 4 FAD3-1A (SEQ ID NO:10) и интрона II FATB-1 (SEQ ID NO:30) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1A сои (SEQ ID NO:11) с вектором, содержащим промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON68544, показанную на фиг.7, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), интрона 4 FAD3-1A (SEQ ID NO:10), интрона II FATB-1 (SEQ ID NO:30) и интрона 4 FAD3-1B (SEQ ID NO:24) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1A сои (SEQ ID NO:11) в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON68546, показанную на фиг.7, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.8, последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1A сои (SEQ ID NO:11) в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON68536, показанную на фиг.8, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1А сои (SEQ ID NO:11) в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Вектор, содержащий ген дельта-9-десатуразы (FAВ2) R. communis (SEQ ID NO:40), регулируемый промотором FAD2 сои и 3'-концевой терминирующей последовательностью nos, разрезают соответствующими рестрикционными ферментами и лигируют вверху от 3'-концевой терминирующей последовательности tml. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON68537, показанную на фиг.8, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой или антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1A сои (SEQ ID NO:11) в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген KAS IN C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON68538, показанную на фиг.8, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.9, последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FATB-1 (SEQ ID NO:36), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FAD3-1B (SEQ ID NO:26) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1A сои (SEQ ID NO:11) в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемым промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON80622, показанную на фиг.9, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FATB-1 (SEQ ID NO:36) и 3'-UTR FAD3-1A (SEQ ID NO:16) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации при разделении сплайсируемым интроном 5 FAD3-1A сои (SEQ ID NO:11) в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемым промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON80623, показанную на фиг.9, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 5'-UTR-3'-UTR FAD2-1 (SEQ ID NO:6 и 5, лигированные друг с другом), 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом), 3'-UTR FAD3-1A (SEQ ID NO:16) и 5'-UTR-3'-UTR FAD3-1B (SEQ ID NO:27 и 26, лигированные друг с другом) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию О5, показанную на фиг.9, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 5'-UTR-3'-UTR FAD2-1 (SEQ ID NO:6 и 5, лигированные друг с другом), 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом) и 3'-UTR FAD3-1A (SEQ ID NO:16) сои амплифицирцуют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемым промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubiscо Е9. Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию О6, показанную на фиг.9, используют для трансформации методами, представленными в настоящем описании изобретения.
3В. Смысловые косупрессорные конструкции
На фиг.10-13 и 19-20 показаны молекулы нуклеиновой кислоты по настоящему изобретению, в которых первые совокупности последовательностей ДНК способны экспрессировать смысловые косупрессорные конструкции. Вторая совокупность последовательностей ДНК включает кодирующие последовательности, каждая из которых является последовательностью ДНК, кодирующей последовательность, которая в случае экспрессии способна повышать уровень белка или транскрипта либо как белка, так и транскрипта, кодированного геном, выбираемым из группы, включающей KAS I, KAS IV, дельта-9-десатуразу и СР4 EPSPS. Каждая кодирующая последовательность ассоциирована с промотором, который является любым промотором, функционирующим в растении, или любым растительным промотором и может быть промотором FMV, промотором напина, промотором 7S (7Sα или 7Sα'), промотором арцелина, промотором дельта-9-десатуразы или промотором FAD2-1A.
Как показано на фиг.10, последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), интрона 4 FAD3-1C (SEQ ID NO:14), интрона II FATB-1 (SEQ ID NO:30), интрона 4 FAD3-1A (SEQ ID NO:10) и интрона 4 FAD3-1B (SEQ ID NO:24) сои амприфицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность гороха Rubisco Е9, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON68522, показанную на фиг.10, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), интрона 4 FAD3-1A (SEQ ID NO:10), интрона 4 FAD3-1B (SEQ ID NO:24) и интрона II FATB-1 (SEQ ID NO:30) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Векторы, содержащие ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, и ген дельта-9-десатуразы R. communis (FAB2) (SEQ ID NO:40), регулируемый промотором FAD2 сои и 3'-концевой терминирующей последовательностью nos, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON80614, показанную на фиг.10, используют для трансформации методами, представленными в настоящем описании изобретении.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), 3'-UTR FAD-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON68531, показанную на фиг.10, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов PCR, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты PCR клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Векторы, содержащие ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, и ген дельта-9-десатуразы (FAB2) R. communis (SEQ ID NO:40), регулируемый промотором FAD2 сои и 3'-концевой терминирующей последовательностью nos, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON68534, показанную на фиг.10, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукт ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген дельта-9-десатуразы (FAB2) R. communis (SEQ ID NO:40), регулируемый промотором FAD2 сои и 3'-концевой терминирующей последовательностью nos, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON68535, показанную на фиг.10, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.11, последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором рMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON80605, показанную на фиг.11, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON80606, показанную на фиг.11, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплиицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген дельта-9-десатуразы (FAB2) R. communis (SEQ ID NO:40), регулируемый промотором FAD2 сои и 3'-концевой терминирующей последовательностью nos, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON80607, показанную на фиг.11, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FAD3-1A (SEQ ID NO:16) и 3'-UTR FATB-1 (SEQ ID NO:36) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sа' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью Rubisco Е9. Векторы, содержащие ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, и ген дельта-9-десатуразы R. communis (FAB2) (SEQ ID NO:40), регулируемый промотором FAD2 сои и 3'-концевой терминирующей последовательностью FAD2, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON80614, показанную на фиг.11, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.12, последовательноcти 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FAТВ-1 (SEQ ID NO:36) и 3'-UTR FAD3-1A (SEQ ID NO:16) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON80629, показанную на фиг.12, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности интрона 1 FAD2-1 (SEQ ID NO:1 или 2), интрона 4 FAD3-1A (SEQ ID NO:10), интрона II FATB-1 (SEQ ID NO:30) и интрона 4 FAD3-1A (SEQ ID NO:10) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию рMON81902, показанную на фиг.12, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 5'-UTR-3'-UTR FAD2-1 (SEQ ID NO:6 и 5, лигированные друг с другом), 5'-UTR-3'-UTR FAD3-1 (SEQ ID NO:17 и 16, лигированные друг с другом, или 27 и 26, лигированные друг с другом) и 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукт ПЦР FAD2-1 клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Аналогичным образом продукт ПЦР FAD3-1 клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα сои и 3'-концевую терминирующую последовательностью tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Продукт ПЦР FATB-1 клонируют в смысловой ориентации в векторе, содержащем промотор арцелина и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данные векторы разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемым промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию О1, показанную на фиг.12, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 5'-UTR-3'-UTR FAD2-1 (SEQ ID NO:6 и 5, лигированные друг с другом), 5'-UТR-3'-UTR FAD3-1 (SEQ ID NO:17 и 16, лигированные друг с другом, или 27 и 26, лигированные друг с другом) и 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукт ПЦР FAD2-1 клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Аналогичным образом продукт ПЦР FAD3-1 клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Продукт ПЦР FATB-1 клонируют в смысловой ориентации в векторе, содержащем промотор арцелина и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данные векторы разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемым промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisсo Е9. Вектор, содержащий ген KAS IN C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию О2, показанную на фиг.12, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.13, последовательности 5'-UTR-3'-UTR FAD2-1 (SEQ ID NO:6 и 5, лигированные друг с другом), 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом), 3'-UTR FAD3-1A (SEQ ID NO:16) и 5'-UTR-3'-UTR FAD3-1B (SEQ ID NO:27 и 26, лигированные друг с другом) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данные векторы затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют в вектором pMON41164. Полученную экспрессирующую ген конструкцию О7, показанную на фиг.13, используют для трансформации методами, представленными в настоящем описании изобретения.
Интрон 1 FAD2-1 сои (SEQ ID NO: 1 или 2) амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Последовательности 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом), 3'-UTR FAD3-1A и (SEQ ID NO:16) и 5'-UTR-3'-UTR FAD3-1B (SEQ ID NO:27 и 26, лигированные друг с другом) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα сои и 3-концевую терминирующую последовательность nos, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данные векторы затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген KAS IV С. рulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию О9, показанную на фиг.13, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.19, некодирующие последовательности FATB-2 (SEQ ID NO:44-47), некодирующие последовательности FAD2-1 (SEQ ID NO:1 и 5-6) и некодирующие последовательности FATB-1 (SEQ ID NO:29-37) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции с обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данные векторы затем разрезают NotI и лигируют с вектором pMON80612, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию, показанную на фиг.19-А, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательность ДНК, содержащую ген дельта-9-десатуразы, регулируемую промотором 7Sальфа и 3'-концевой терминирующей последовательностью TML, разрезают соответствующими рестрикционными ферментами, и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую конструкцию, показанную на фиг.19-В, используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором арцелина фасоли и 3'-концевой терминирующей последовательностью напина, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую ген конструкцию, показанную на фиг.19-С, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.20, некодирующие последовательности FATB-2 (SEQ ID NO:44-47), некодирующие последовательности FAD2-1 (SEQ ID NO:1 и 5-6), некодирующие последовательности FATB-1 (SEQ ID NO:29-37), некодирующие последовательности FAD3-1A (SEQ ID NO: 7-13 и 16-17) и некодирующие последовательности FAD3-1B (SEQ ID NO:19-27) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данные векторы затем разрезают NotI и лигируют с вектором pMON80612, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию, показанную на фиг.20-А, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательность ДНК, содержащую ген дельта-9-десатуразы, регулируемую промотором 7Sальфа и 3'-концевой терминирующей последовательностью TML, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую конструкцию, показанную на фиг.20-В, используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор, содержащий ген КAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором арцелина фасоли Brassica и 3'-концевой терминирующей последовательностью напина, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую ген конструкцию, показанную на фиг.20-С, используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор pMON93501 содержит интрон 1 FAD2-1A сои (SEQ ID NO:1), функционально связанный с промотором 7Sα' сои и 3'-концевой терминирующей последовательностью Н6, ген KAS IV C. pulcherrima (SEQ ID NO:39), функционально связанный с промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, ген дельта-9-десатуразы Ricinus communis (публикация заявки на патент США № 2003/00229918 А1), функционально связанный с промотором 7SA сои и 3'-концевой терминирующей последовательностью nos, и ген СР4 EPSPS, функционально связанный с промотором EFMV (конститутивный промотор, полученный из вируса мозаики норичника) и 3'-концевой терминирующей последовательностью гороха Rubisco, при этом все последовательности фланкированы краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
3С. Антисмысловые конструкции
На фиг.14 показаны молекулы нуклеиновой кислоты по настоящему изобретению, в которых первые совокупности последовательноcтей ДНК способны экспрессировать антисмысловые конструкции, и на фиг.15-18 показаны молекулы нуклеиновой кислоты по настоящему изобретению, в которых первые совокупности последовательностей ДНК способны экспрессировать комбинации смысловых и антисмысловых конструкций. Вторая совокупность последовательностей ДНК включает кодирующие последовательности, каждая из которых является последовательностью ДНК, кодирующей последовательность, которая в случае экспрессии способна повышать уровень белка или транскрипта либо как белка, так и транскрипта, кодированного геном, выбираемым из группы, включающей KAS I, KAS IV, дельта-9-десатуразу и СР4 EPSPS. Каждая кодирующая последовательность ассоциирована с промотором, который является любым промотором, функционирующим в растении, или любым растительным промотором и может быть промотором FMV, промотором напина, промотором 7S (7Sα или 7Sα'), промотором арцелина, промотором дельта-9-десатуразы или промотором FAD2-1A.
Как показано на фиг.14, последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FATB-1 (SEQ ID NO:36) и 3'-UTR FAD3-1A (SEQ ID NO:16) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubiscо Е9. Полученную экспрессирующую ген конструкцию pMON80615, показанную на фиг.14, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FATB-1 (SEQ ID NO:36) и 3'-UTR FAD3-1A (SEQ ID NO:16) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию pMON80616, показанную на фиг.14, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FATB-1 (SEQ ID NO:36) и 3'-UTR FAD3-1A (SEQ ID NO:16) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген дельта-9-десатуразы (FAD2) R. сommunis (SEQ ID NO:40), регулируемый промотором FAD2 сои и 3'-концевой терминирующей последовательностью nos, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMOB41164. Полученную экспрессирующую ген конструкцию pMON80617, показанную на фиг.14, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 3'-UTR FAD2-1 (SEQ ID NO:5), 3'-UTR FATB-1 (SEQ ID NO:36) и 3'-UTR FAD3-1A (SEQ ID NO:16) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в антисмысловой ориентации в векторе, содержащем промотор 7Sα сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию pMON80630, показанную на фиг.14, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 5'-UTR-3'-UTR FAD2-1 (SEQ ID NO:6 и 5, лигированные друг с другом), 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом). 3'-UTR FAD3-1A (SEQ ID NO:16) и 5'-UTR-3'-UTR FAD3-1B (SEQ ID NO:27 и 26, лигированные друг с другом) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в антимысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию О8, показанную на фиг.14, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.15, последовательности 5'-UTR-3'-UTR FAD2-1 (SEQ ID NO:6 и 5, лигированные друг с другом), 5'-UTR-3'-UTR FAD3-1A (SEQ ID NO:17 и 16, лигированные друг с другом) и 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, при наличии дополнительного промотора 7Sα сои между смысловыми и антисмысловыми последовательностями, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию О3, показанную на фиг.15, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательности 5'-UTR-3'-UTR FAD2-1 (SEQ ID NO:6 и 5, лигированные друг с другом), 5'-UTR-3'-UTR FAD3-1A (SEQ ID NO:27 и 26, лигированные друг с другом) и 5'-UTR-3'-UTR FATB-1 (SEQ ID NO:37 и 36, лигированные друг с другом) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, при наличии дополнительного промотора 7Sα сои между смысловыми и антисмысловыми последовательностями, используя сайты XhoI, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают NotI и лигируют с вектором pMON41164, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором напина Brassica и 3'-концевой последовательностью напина Brassica, разрезают соответствующими рестрикционными ферментами и лигируют с вектором pMON41164. Полученную экспрессирующую ген конструкцию О4, показанную на фиг.15, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.16, некодирующие последовательности FATB-2 (SEQ ID NO:44-47), некодирующие последовательности FATB-1 (SEQ ID NO:29-37) и некодирующие последовательности FAD2-1 (SEQ ID NO:1 и 5-6) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml. Данный вектор затем разрезают соответствующей рестрикционной эндонуклеазой и лигируют с вектором pMON80612, содержащим ген СР4 EPSPS, регулируемым промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию, показанную на фиг.16-А, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательность ДНК, содержащую ген дельта-9-десатуразы, регулируемую промотором 7Sальфа и 3'-концевой терминирующей последовательностью TML, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую конструкцию, показанную на фиг.16-В, используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором арцелина фасоли и 3'-концевой терминирующей последовательностью напина, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую ген конструкцию, показанную на фиг.16-С, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.17, некодирующие последовательности FATB-2 (SEQ ID NO:44-47), некодирующие последовательности FATB-1 (SEQ ID NO:29-37), некодирующие последовательности FAD2-1 (SEQ ID NO:1 и 5-6) и некодирующие последовательности FAD3-1A (SEQ ID NO:7-13 и 16-17) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, содержащих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательности tml. Данный вектор затем разрезают соответствующей рестрикционной эндонуклеазой и лигируют с вектором pMON80612, содержащим ген СР4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9. Полученную экспрессирующую ген конструкцию, показанную на фиг.17-А, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательность ДНК, содержащую ген дельта-9-десатуразы, регулируемую промотором 7Sальфа и 3'-концевой терминирующей последовательностью TML, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанными экспрессирующими конструкциями. Полученную экспрессирующую конструкцию, показанную на фиг.17-В, используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором арцелина фасоли и 3'-концевой терминирующей последовательностью напина, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую ген конструкцию, показанную на фиг.17-С, используют для трансформации методами, представленными в настоящем описании изобретения.
Как показано на фиг.18, некодирующие последовательности FATB-2 (SEQ ID NO:44-47), некодирующие последовательности FATB-1 (SEQ ID NO:29-37), некодирующие последовательности FAD2-1 (SEQ ID NO:1 и 5-6), некодирующие последовательности FAD3-1A (SEQ ID NO:7-13 и 16-17) и некодирующие последовательности FAD3-1B (SEQ ID NO:19-27) сои амплифицируют методом ПЦР с образованием продуктов ПЦР, включающих повторно созданные сайты рестрикции у обоих концов. Продукты ПЦР клонируют в смысловой и антисмысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml. Данный вектор затем разрезают соответствующей рестрикционной эндонуклеазой и лигируют с вектором pMON80612, содержащим ген СР4 EPSPE, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco. Полученную экспрессирующую ген конструкцию, показанную на фиг.18-А, используют для трансформации методами, представленными в настоящем описании изобретения.
Последовательность ДНК, содержащую ген дельта-9-десатуразы, регулируемую промотором 7Sальфа и 3'-концевой терминирующей последовательностью TML, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую конструкцию, показанную на фиг.18-В, используют для трансформации методами, представленными в настоящем описании изобретения.
Вектор, содержащий ген KAS IV C. pulcherrima (SEQ ID NO:39), регулируемый промотором арцелина фасоли и 3'-концевой терминирующей последовательностью напина, разрезают соответствующими рестрикционными ферментами и лигируют с вышеуказанной экспрессирующей конструкцией. Полученную экспрессирующую ген конструкцию, показанную на фиг.18-С, используют для трансформации методами, представленными в настоящем описании изобретения. Вышеуказанные молекулы нуклеиновой кислоты являются предпочтительными вариантами осуществления изобретения, позволяющими реализовать цели, отличительные признаки и преимущества настоящего изобретения. Не следует считать, что настоящее изобретение ограничено иллюстрированными вариантами изобретения. Расположение последовательностей в первой и второй совокупностях последовательностей ДНК в молекуле нуклеиновой кислоты не ограничено иллюстрированными и описанными расположениями и может изменяться любым образом, приемлемым для реализации целей, отличительных признаков и преимуществ настоящего изобретения, представленного в настоящем описании изобретения, иллюстрированного на прилагаемых чертежах и охватываемого формулой изобретения.
3D. Сборка in vivo
Одним объектом настоящего изобретения является конструкция ДНК, используемая для сборки рекомбинантной единицы транскрипции в хромосоме растения in planta, которая способна образовывать двухцепочечную РНК. Сборка таких конструкций и способы сборки in vivo рекомбинантной единицы транскрипции для подавления гена описаны в международной заявке на патент № РСТ/US2005/00681, которая полностью включена в настоящее описание изобретения в виде ссылки.
pMON93505 представляет собой конструкцию, используемую для сборки in vivo, которая содержит два сегмента Т-ДНК, фланкированных краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Первый сегмент Т-ДНК содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 100 последовательных нуклеотидов от 3'-конца и лигированным с 3'-UTR FATB-1a, за которым следует 5'-UTR FATB-1a, ген KAS IV C. pulcherrima (SEQ ID NO:39), функционально связанный с промотором напина Brassica и 3'-концевой терминирующей последовательностью напина Brassica, ген дельта-9-десатуразы Ricinus communis (публикация заявки на патент США № 2003/00229918 А1), функционально связанный с промотором 7SA сои и 3'-концевой терминирующей последовательностью nos, и ген СР EPSPS, функционально связанный с промотором еFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). В той же конструкции во втором сегменте Т-ДНК, фланкированном другим RB и LB, находится 3'-концевая терминирующая последовательность Н6, функционально связанная с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 100 последовательных нуклеотидов от 3'-конца и лигированным с 3'-UTR FATB-1a, за которой следует 5'-UTR FATB-1a. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Когда два сегмента Т-ДНК вышеуказанной конструкции введены в один локус хромосомы организма-хозяина в ориентации RB-RB, собранная единица транскрипции содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1А и фрагментами ДНК FATB-1a сои в смысловой или антисмысловой ориентации. Будучи транскрибированными, функционально связанные последовательности РНК в смысловой и антисмысловой ориентации способны образовывать двухцепочечную РНК, эффективно подавляющую FAD2-1A и FATB.
pMON93506 представляет собой конструкцию, используемую для сборки in vivo, которая содержит два сегмента Т-ДНК, фланкированных краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Первая Т-ДНК содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A (SEQ ID NO:1), уменьшенным на 100 последовательных нуклеотидов от 3'-конца и лигированным с 3'-UTR FATB-1a, за которой следует 5'-UTR FATB-1a, ген дельта-9-десатуразы Ricinus communis (публикация заявки на патент США № 2003/00229918 А1), функционально связанный с промотором 7SA сои и 3'-концевой терминирующей последовательностью nos, и ген СР4 EPSPS, функционально связанный с промотором eFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы LB и RB. В том же векторе во втором сегменте Т-ДНК находится 3'-концевая терминирующая последовательность, функционально связанная с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 100 последовательных нуклеотидов от 3'-конца и лигированным с 3'-UTR FATB-1a, за которой следует 5'-UTR FATB-1a, фланкированная другим RB и LB. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Когда два сегмента Т-ДНК вышеуказанной конструкции введены в один локус хромосомы организма-хозяина в ориентации RB-RB, собранная единица транскрипции содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A и фрагментами ДНК FATB-1a сои в смысловой или антисмысловой ориентации. Будучи транскрибированными, функционально связанные последовательности РНК в смысловой и антимысловой ориентации способны образовывать двухцепочечную РНК, эффективно подавляющую FAD2-1A и FATB.
pMON95829 представляет собой конструкцию, используемую для сборки in vivo, которая содержит два сегмента Т-ДНК, фланкированных краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Первая Т-ДНК содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 100 последовательных нуклеотидов от 3'-конца и лигированным с 42 последовательными нуклеотидами 5'-UTR FATB-1a, за которой следует кодирующая область транзитного пептида хлоропласта (“CTP”) FATB-1, и ген СР2 EPSPS, функционально связанный с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). В том же векторе во втором сегменте Т-ДНК, фланкированном другим RB и LB, находится 3'-концевая терминирующая последовательность Н6, функционально связанная с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 100 последовательных нуклеотидов от 3'-конца и лигированным с 42 последовательными нуклеотидами 5'-UTR FATB-1a, за которой следует кодирующая область транзитного пептида хлоропласта FATB-1 (“CTP”). Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Когда два сегмента Т-ДНК вышеуказанной конструкции введены в один локус хромосомы организма-хозяина в ориентации RB-RB, собранная единица транскрипции содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1А и фрагментами ДНК FATB-1a сои в смысловой или антисмысловой ориентации. Будучи транскрибированными, функционально связанные последовательности РНК в смысловой и антисмысловой ориентации способны образовывать двухцепочечную РНК, эффективно подавляющую FAD2-1A и FATB.
pMON97595 представляет собой конструкцию, используемую для сборки in vivo, которая содержит два сегмента Т-ДНК, фланкированных краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Первый сегмент Т-ДНК содержит промотор 7Sα'сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с 42 последовательными нуклеотидами 5'-UTR FATB-1a, за которой следует кодирующая область транзитного пептида хлоропласта (“CTP”) FATB-1a, и ген СР4 EPSPS, функционально связанный с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы кривыми элементами Т-ДНК, то есть ДНК правого края (RB) и ДНК левого края (LB). Во втором сегменте Т-ДНК, фланкированном другим RB и LB, находится 3'-концевая терминирующая последовательность Н6, функционально связанная с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с 42 последовательными нуклеотидами 5'-UTR FATB-1a, за которой следует кодирующая область СТР FATB-1a. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Когда два сегмента Т-ДНК вышеуказанной конструкции введены в один локус хромосомы организма-хозяина в ориентации RB-RB, собранная единица транскрипции содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1А и фрагментами ДНК FATB-1a сои в смысловой или антисмысловой ориентации. Будучи транскрибированными, функционально связанные последовательности РНК в смысловой и антисмысловой ориентации способны образовывать двухцепочечную РНК, эффективно подавляющую FAD2-1A и FATB.
pMON97581 представляет собой конструкцию, используемую для сборки in vivo, которая содержит два сегмента Т-ДНК, фланкированных краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Первый сегмент Т-ДНК содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с кодирующей областью транзитного пептида хлоропласта (“CTP”) FATB-1a, и ген СР4 EPSPS, функционально связанный с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). В той же конструкции во втором сегменте Т-ДНК, фланкированном другим RB и LB, находится 3'-концевая терминирующая последовательность Н6, функционально связанная с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с кодирующей областью СТР FATB-1a. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Когда два сегмента Т-ДНК вышеуказанной конструкции введены в один локус хромосомы организма-хозяина в ориентации RB-RB, собранная единица транскрипции содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1А и фрагментами ДНК FATB-1a сои в смысловой или антисмысловой ориентации. Будучи транскрибированными, функционально связанные последовательности РНК в смысловой и антисмысловой ориентации способны образовывать двухцепочечную РНК, эффективно подавляющую FAD2-1A и FATB.
pMON97596 представляет собой конструкцию, используемую для сборки in vivo, которая содержит два сегмента Т-ДНК, фланкированных краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Первый сегмент Т-ДНК содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с 180 п.о. 5'-конца кодирующей области транзитного пептида хлоропласта (“CTP”) FATB-1, и ген СР4 EPSPS, функционально связанный с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). В той же конструкции во втором сегменте Т-ДНК, фланкированном другим RB и LB, находится 3'-концевая терминирующая последовательность Н6, функционально связанная с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с 180 п.о. 5'-конца кодирующей области СТР FATB-1a. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Когда два сегмента Т-ДНК вышеуказанной конструкции введены в один локус хромосомы организма-хозяина в ориентации RB-RB, собранная единица транскрипции содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1А и фрагментами ДНК FATB-1а сои в смысловой или антисмысловой ориентации. Будучи транскрибированными, функционально связанные последовательности РНК в смысловой и антисмысловой ориентации способны образовывать двухцепочечную РНК, эффективно подавляющую FAD2-1A и FATB.
рMON97597 представляет собой конструкцию, используемую для сборки in vivo, которая содержит два сегмента Т-ДНК, фланкированных краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Первый сегмент Т-ДНК содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с 120 п.о. 5'-конца кодирующей области транзитного пептида хлоропласта (“CTP”) FATB-1a, и ген СР4 EPSPS, функционально связанный с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). В той же конструкции во втором сегменте Т-ДНК, фланкированном другим RB и LB, находится 3'-концевая терминирующая последовательность Н6, функционально связанная с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 320 последовательных нуклеотидов от 3'-конца и лигированным с 120 п.о. 5'-конца кодирующей области СТР FATB-1a. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Когда два сегмента Т-ДНК вышеуказанной конструкции введены в один локус хромосомы организма-хозяина в ориентации RB-RB, собранная единица транскрипции содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1А и фрагментами ДНК FATB-1a сои в смысловой или антисмысловой ориентации. Будучи транскрибированными, функционально связанные последовательности РНК в смысловой и антисмысловой ориентации способны образовывать двухцепочечную РНК, эффективно подавляющую FAD2-1A и FATB.
pMON97598 представляет собой конструкцию, используемую для сборки in vivo, которая содержит два сегмента Т-ДНК, фланкированных краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). Первый сегмент Т-ДНК содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 340 последовательных нуклеотидов от 3'-конца и лигированным с кодирующей областью транзитного пептида хлоропласта (“CTP”) FATB-1a, и ген СР4 EPSPS, функционально связанный с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы краевыми элементами Т-ДНК Agrobacterium, то есть ДНК правого края (RB) и ДНК левого края (LB). В той же конструкции во втором сегменте Т-ДНК, фланкированном другим RB и LB, находится 3'-концевая терминирующая последовательность Н6, функционально связанная с интроном 1 FAD2-1A сои (SEQ ID NO:1), уменьшенным на 340 последовательных нуклеотидов от 3'-конца и лигированным с кодирующей областью СТР FATB-1a. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Когда два сегмента Т-ДНК вышеуказанной конструкции введены в один локус хромосомы организма-хозяина в ориентации RB-RB, собранная единица транскрипции содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1А и фрагментами ДНК FATB-1a сои в смысловой или антисмысловой ориентации. Будучи транскрибированными, функционально связанные последовательности РНК в смысловой и антисмысловой ориентации способны образовывать двухцепочечную РНК, эффективно подавляющую FAD2-1A и FATB.
Пример 4. Трансформация растений и анализ
Конструкции по примерам 2 и 3 устойчиво вводят в сою (например, Asgrow сорта А4922, Asgrow сорта А3244 или Asgrow сорта А3525) описанными выше методами, в том числе методами, представленными в публикации McCabe et al., Bio/Technology 6:923-926 (1988), или методом трансформации, опосредованной Agrobacterium. Трансформированные растения сои идентифицируют путем отбора на среде, содержащей глифосат. Составы жирных кислот анализуют газовой хроматографией в семенах линий сои, трансформированных вышеуказанными конструкциями. Кроме того, любые конструкции могут содержать другие представляющие интерес последовательности, а также разные комбинации промоторов.
Для некоторых применений модифицированные составы жирных кислот определяют в развивающихся семенах, в то время как в других случаях, например при выполнении анализа профиля масла, определении модификаций жирных кислот, происходящих позднее в пути FAS, или обнаружении незначительных модификаций в составе жирных кислот, предпочтительно используют зрелые семена. Кроме того, может быть желательно выполнить анализ масла и/или содержания жирных кислот в отдельных семенах, особенно при определении модификации масла в расщепляющихся популяциях семян R1. В используемом здесь значении поколение R0 означает растение, получаемое методами трансформации/регенерации, описанными в настоящем описании изобретения, и поколение R1 означает семена, полученные у “своего” трансгенного растения R0. Растения R1 выращены из семян R1.
Составы жирных кислот определяют для семени линий сои, трансформированных конструкциями по примеру 3. От одного до десяти семян трансгенных и контрольных линий сои измельчают отдельно, используя гомогенизатор ткани (Pro Scientific) для экстракции масла. Масло из измельченного семени сои экстрагируют в течение ночи в 1,5 мл гептана, содержащего тригептадеканоин (0,50 мг/мл). Из аликвот 200 мкл экстрагированного масла получают сложные метиловые эфиры, добавляя 500 мкл метоксида натрия в абсолютном метаноле. Реакцию образования производного соединения осуществляют в течение 20 минут при 50°С. Указанную реакцию прекращают в результате одновременного добавления 500 мкл 10% (мас./об.) хлорида натрия и 400 мкл гептана. Полученные сложные метиловые эфиры жирных кислот, экстрагированные гексаном, исследуют газовой хроматографией (GC) в газовом хроматографе модели 6890 компании Hewlett-Packard (Palo Alto, CA). Указанный газовый хроматограф оснащен колонкой Supercowax 250 (30 м, внутренний диаметр 0,25 мм, толщина пленки 0,25 микрона) (Supelco, Bellefonte, PA). Температура колонки во время впрыскивания равна 175°С, при этом запрограммировано изменение температуры от 175°С до 245°С до 175°С со скоростью 40°С/мин. Температуры дозатора и детектора равны соответственно 250°С и 270°С.
Пример 5. Синтезированное жидкое топливо с улучшенными свойствами биологического дизельного топлива
В составе жирных кислот синтезированного жидкого топлива присутствуют следующие смеси сложных метиловых эфиров жирных кислот: 73,3% олеиновой кислоты, 21,4% линолевой кислоты, 2,2% пальмитиновой кислоты, 2,1% линоленовой кислоты и 1,0% стеариновой кислоты (все значения выражены в массовых процентах). Очищенные сложные метиловые эфиры жирных кислот приобретены в компании Nu-Chek Prep., Ind., Elysian, MN, USA. Цетановое число и задержка воспламенения указанной композиции определены в юго-западном научно-исследовательском институте при помощи анализатора качества воспламенения (“IQT”) 613 (Southwest Research Institute, San Antonio, Texas, USA).
Анализатор качества воспламенения состоит из камеры горения постоянного объема с электрическим нагреванием, системы впрыскивания топлива и компьютера, используемого для контроля за ходом эксперимента, регистрации и интерпретации данных. Система впрыскивания топлива включает топливную форсунку, образующую входное отверстие в камеру горения. Игольчатый датчик в топливной форсунке детектирует поток топлива в камеру горения. Датчик давления, присоединенный к камере горения, измеряет давление в цилиндре и давление в камере горения. Анализатор качества воспламенения служит главным образом для измерения времени от начала впрыскивания топлива в камеру горения до начала горения. Термодинамические условия в камере горения контролируются с высокой точностью для согласованного измерения времени задержки воспламенения.
Для определения цетанового числа и задержки воспламенения испытуемое топливо фильтруют через 5-микронный фильтр. Топливный резервуар, линию впрыскивания и форсунку продувают подаваемым под давлением азотом. Топливный резервуар затем очищают неволокнистой тканью. Часть испытуемого топлива используют для промывки топливного резервуара, линии впрыскивания и форсунки. Резервуар заполняют испытуемым топливом и из системы удаляют весь воздух. В резервуаре создают давление, равное 3,5 атм (50 фунтов/кв.дюйм). Данный метод состоит в инъецировании под высоким давлением точно отмеренного количества испытуемого топлива в камеру горения, которая заполнена воздухом для достижения требуемого давления и температуры. Измерение состоит в определении времени от начала впрыскивания до начала горения, которое часто определяется как время задержки воспламенения. Данное определение основано на измеренном подъеме иглы и давлении в камере горения. Обычная процедура определения цетанового числа требует создания поверхностной температуры, равной 567,5°С, и давления воздуха, равного 2,1 МПа.
Топливо с известной задержкой впрыскивания подают в камеру горения IQT в начале дня для гарантии того, что в данном устройстве созданы нормальные параметры. Затем исследуют испытуемое синтетическое топливо. Затем снова исследуют известное топливо для проверки правильности установки параметров системы. Сразу же после присоединения топливного резервуара к насосу впрыскивания топлива контроллер компьютера инициирует процесс испытания. Компьютер контролирует весь процесс, включая подачу воздуха, впрыскивание топлива и удаление продуктов горения. Сжигание топлива повторяется 32 раза.
Время задержки воспламенения является временем от начала впрыскивания до начала горения. Данный параметр определяют на основании данных подъема иглы и давления в цилиндре. Подъем иглы впрыскивания свидетельствует о начале впрыскивания. Давление в цилиндре немного понижается вследствие охлаждающего эффекта испарения топлива. Начало горения определяют как время восстановления давления в цилиндре, которое повышается вследствие сгорания до достижения давления, предшествующего впрыскиванию топлива.
Измеренное время задержки воспламенения используют для определения цетанового числа на основании калибровочной кривой, которую вводят в программу сбора и предварительной обработки данных. Калибровочную кривую, определяющую цетановое число в зависимости от времени задержки воспламенения, получают, используя смеси эталонных топлив и контрольных топлив NEG. В случае испытуемых топлив, которые являются жидкими при комнатной температуре, калибровочную кривую ежедневно проверяют, используя по крайней мере одно контрольное топливо с известных цетановым числом (Ryan, "Correlation of Physical and Chemical Ignition Delay to Cetane Number", SAE Paper 852103 (1985); Ryan, "Diesel Fuel Ignition Quality as Determined in a Constant Volume Combustion Bomb", SAE Paper 870586 (1986); Ryan, "Development of a Portable Fuel Cetane Quality Monitor", Belvoir Fuels and Lubricants Research Facility Report No. 277, May (1992); Ryan, "Engine and Constant Volume Bomb Studies of Diesel Ignition and Combustion", SAE Paper 881616 (1988); and Allard et al. "Diesel Fuel Ignition Quality as Determined in the Ignition Quality Tester ("IQT")", SAE Paper 961182 (1996)). Как показано в таблице 3, синтезированное масло имеет цетановые числа и значения времени задержки воспламенения, пригодные для использования масла с данным составом без каких-либо ограничений в качестве биологического дизельного топлива.
испытаний
Плотность (ASTM D-4052), температуру помутнения (ASTM D-2500), температуру потери текучести (ASTM D-97) и температуру забивания фильтра в холодном состоянии (IP 309/ASTM D-6371) определяют для синтезированного масла в соответствии с протоколами испытаний ASTM D. Протоколы испытаний ASTM D предоставлены Американским обществом по испытанию материалов, 100 Barr Harbor Drive, West Conshohocken, PA, USA. Результаты указанных испытаний приведены в таблице 4. Как показано в таблице 4, синтезированное масло имеет показатели, пригодные для использования масла с данным составом без каких-либо ограничений в качестве биологического дизельного топлива.
Уровни выделения оксида азота оценивают, определяя уровни ненасыщенности биологического топлива путем измерения плотности топлива и вычисления расчетных уровней выделения или путем прямого измерения. Кроме того, существуют стандартные методы, предназначенные для прямого измерения уровней выделения оксида азота. На основании оценки общего уровня ненасыщенности синтезированного масла установлено, что синтезированное масло характеризуется более низкими уровнями выделения оксида азота по сравнению со сложными метиловыми эфирами жирных кислот, полученными из обычного соевого масла. Масла, содержащие большее число двойных связей, то есть имеющие более высокую степень ненасыщенности, выделяют больше оксида азота. Данное масло имеет в общей сложности 123 двойные связи по сравнению с 153 двойными связями обычного соевого масла, как показано в таблице 5.
По данных Национальной лаборатории возобновляемой энергии, контракт № ACG-8-17106-02 Final Report, The Effect Of Biodiesel Composition On Engine Emissions From A DDC Series 60 Diesel Engine, (June 2000) выделение азотной кислоты биологическим дизельным топливом можно вычислить по формуле у = 46,959х - 36,388, где у означает выделение оксида в граммах/мощность торможения в лошадиных силах × час (г/лс·час); и х означает плотность биологического дизельного топлива. Данная формула создана на основе регрессионного анализа данных выделения азотной кислоты при испытании 16 биологических дизельных топлив. При выполнении указанного испытания использован калибровочный двигатель производственной серии 1991 модели 60 Detroit Diesel Corporation.
Плотность синтезированного масла определена в юго-западном научно-исследовательском институте методом ASTM D4052. Результат, приведенный в таблице 4, использован в вышеуказанном уравнении для прогнозирования значения выделения оксида азота, равного 4,89 г/лс·час. Полученный результат сравним с показателем контрольного соевого продукта. В отчете Национальной лаборатории возобновляемой энергии приведены данные плотности и выделения оксида азота для контрольного биологического дизельного топлива из сои (сложный метиловый эфир сои IGT). Плотность контрольного биологического дизельного топлива равна 0,8877 г/мл, поэтому вычисленное значение выделения оксида азота равно 5,30 г/лс·час. Вычисленное значение выделения аналогично экспериментальному значению выделения оксида азота, равному 5,32 г/лс·час. Синтетическое масло характеризуется лучшими значениями по сравнению с контрольным топливом и пригодно для использования без каких-либо ограничений в качестве биологического дизельного топлива.
Пример 6. Оптимальный состав жирных кислот, необходимый для обеспечения нормальных уровней липидов в сыворотке
Определяли свойства составов растительных масел в отношении снижения уровня холестерина, чтобы идентифицировать составы жирных кислот, оказывающие более благоприятное воздействие на содержание липидов в сыворотке по сравнению с обычным соевым маслом (то есть более низкое содержание холестерина LDL и более выcокое содержание холестерина HDL). Для сравнения воздействия на уровни липидов в сыворотке человека новых составов масла из семян с обычным соевым маслом использованы опубликованные уравнения, выведенные на основе 27 клинических испытаний (Mensink, R.P. and Katan, M.B. Arteriosclerosis and Thrombosis, 12:911-919 (1992)).
В приведенной ниже таблице 6 представлены результаты изменения содержания липидов в сыворотке при замене липидами 30% энергии пищи, создаваемой углеводами. Полученные результаты показывают, что соевое масло уже оказывает благоприятное воздействие на уровни липидов в сыворотке при замене им углеводов в рационе питания. Состав указанного масла можно улучшить путем снижения уровней насыщенного жира и доведения уровня линолевой кислоты до 10-30% от общего содержания жирных кислот, предпочтительно примерно до 15-25% от общего содержания жирных кислот. Когда доля линолевой кислоты составляет менее 10% от общего содержания жирных кислот, новый состав повышает уровень холестерина LDL по сравнению с контрольным соевым маслом, даже если содержание насыщенного жира составляет 5% от общего содержания жирных кислот. При увеличении доли линолевой кислоты способность композиции повышать уровни HDL в сыворотке снижается. Поэтому предпочтительное содержание линолевой кислоты в составе масла должно быть равно примерно 15-25% от общего содержания жирных кислот.
(С20:1)
Пример 7
Следующие четырнадцать стадий иллюстрируют создание вектора pMON68537, предназначенного для трансформации растения с целью подавления генов FAD2, FAD3 и FATB и сверхэкспрессии дельта-9-десатуразы в сое. В частности, данная конструкция включает промотор 7Sальфа, функционально связанный с интроном и 3'-UTR сои в смысловой ориентации, то есть интрон № 1 FAD2-1A, 3'-UTR FAD3-1A, 3'-UTR FATB-1, шпилькообразный петлеобразующий сплайсируемый интрон и комплементарную серию интрона и 3'-UTR сои в антисмысловой ориентации, то есть 3'-UTR FATB-1, 3'-UTR FAD3-1A и интрон № 1 FAD2-1A, и промотор FAD2 сои, стимулирующий дельта-9-десатуразу.
Стадия 1. Интрон № 5 FAD3-1A сои, который является частью сплайсируемого интрона конструкции дцРНКи, амплифицируют методом ПЦР, используя геномную ДНК сои в качестве матрицы, со следующими праймерами:
5'-концевой праймер = 19037 = ACTAGTATATTGAGCTCATATTCCACTGCAGTGGATATT
GTTTAAACATAGCTAGCATATTACGCGTATATTATACAAGCTTATATTCCCGGG
ATATTGTCGACATATTAGCGGTACATTTTATTGCTTATTCAC
3'- концевой праймер = 19045 =
ACTAGTATATTGAGCTCATATTCCTGCAGGATATTCTCGAG
ATATTCACGGTAGTAATCTCCAAGAACTGGTTTTGCTGCTTGTGTCTGCAGTGAATC
Указанные праймеры добавляют сайты клонирования к 5'- и 3'-концам. К 5'-концу: SleI, SacI, BstXI, PmeI, NheI, MluI, HindIII, XmaI, SmaI, SalI. К 3'-концу: SрeI, SacI, Sse8387I, XhoI. Интрон № 5 FAD3-1A сои, являющийся продуктом ПЦР, клонируют в pCT2.1, создавая KAWHIT03.0065. Затем KAWHIT03.0065 расщепляют SpeI и концы заполняют полимеразой Pfu и pMON68526 (пустой вектор, устойчивый к хлорамфениколу (далее именуемому СМ), расщепляют HindIII и концы заполняют полимеразой Pfu. KAWHIT03.0065 и pMON68526 лигируют с образованием pMON68541 (интрон № 5 FAD3-1A сои с сайтами множественного клонирования в векторе, устойчивом к амфениколу).
Стадия 2. 3'-UTR FATB-1 сои амплифицируют со следующими праймерами:
18662 = TTTTAATTACAATGAGAATGAGATTTACTGC (добавление Bsp120I к 5'-концу) и 18661 = GGGCCCGATTTGAAATGGTTAACG. Продукт ПЦР лигируют с pCR2.1, создавая KAWHIT03.0036.
Стадия 3. KAWHIT03.0036 расщепляют Bsp120I и EcoRI и клонируют в KAWHIT03.0032 (пустой СМ-устойчивый вектор с сайтом множественного клонирования), создавая KAWHIT03.0037 (3'-UTR FATB-1 в пустом СМ-устойчивом векторе).
Стадия 4. 3'-UTR FAD3-1A сои амплифицируют со следующими праймерами:
18639 = GGGCCCGTTTCAAACTTTTTGG (добавление Bsp120I к 5'-концу) и 18549 = TGAAACTGACAATTCAA. Продукт ПЦР лигируют с pCR2.1, создавая KAWHIT03.0034.
Стадия 5. KAWHIT03.0034 расщепляют Bsp120I и EcoRI и лигируют с KAWHIT03.0032 (пустой СМ-устойчивый вектор с сайтом множественного клонирования), создавая KAWHIT03.0035 (3'-UTR FAD3-1A в пустом СМ-устойчивом векторе).
Стадия 6. Интрон № 1 FAD2-1A сои амплифицируют методом ПЦР, используя геномную ДНК сои в качестве матрицы, со следующими праймерами: 5'-концевой праймер = 18663 = GGGCCCGGTAAATTAAATTGTGC (добавление сайта Bsp120I к 5'-концу); и 3'-концевой праймер = 18664 = CTGTGTCAAAGTATAAACAAGTTCAG. Полученный продукт ПЦР клонируют в pCR2.1, создавая KAWHIT03.0038.
Стадия 7. Интрон № 1 FAD2-1A сои, являющийся продуктом ПЦР, в KAWHIT03.0038, клонируют в KAWHIT03.0032 (пустой СМ-устойчивый вектор с сайтом множественного клонирования), используя сайты рестрикции Bsp120I и EcoRI. Полученная плазмида представляет собой KAWHIT03.0039 (интрон № 1 FAD2-1A в пустом СМ-устойчивом векторе).
Стадия 8. KAWHIT03.0039 расщепляют AscI и HindIII и pMON68541 (АМР-устойчивый базовый вектор с дцРНКи интрона № 5 FAD3-1A) расщепляют MluI и HindIII. Интрон № 1 FAD2-1A сои направленно клонируют в pMON68541, создавая KAWHIT03.0071 (интрон № 1 FAD2-1A сои с интроном № 5 FAD3-1A сои).
Стадия 9. KAWHIT03.0035 (3'-UTR FAD3-1A в СМ-устойчивом векторе) расщепляют AscI и HindIII и KAWHIT03.0071 (АМР-устойчивый базовый вектор с дцРНКи интрона FAD2-1A и интрона № 5 FAD3-1A) расщепляют MluI и HindIII. 3'-UTR FAD3-1A сои направленно клонируют в KAWHIT03.0071, создавая KAWHIT03.0072 (интрон № 1 FAD2-1A и 3'-UTR FAD3-1A с интроном № 5 FAD3-1A сои).
Стадия 10. KAWHIT03.0037 (3'-UTR FATB-1 в СМ-устойчивом векторе) расщепляют AscI и HindIII и KAWHIT03.0072 расщепляют MluI и HindIII. 3'-UTR FATB-1 направленно клонируют в KAWHIT03.0072, создавая KAWHIT03.0073 (интрон FAD2-1A, 3'-UTR FAD3-1A, 3'-UTR FATB-1 сои с интроном № 5 FAD3-1A).
Стадия 11. KAWHIT03.0073 расщепляют BstXI и SаlI и фрагмент, содержащий интрон FAD2-1A, 3'-UTR FAD3-1A и 3'-UTR FATB-1, очищают гелем. В другой пробирке KAWHIT03.0073 расщепляют XhoI и Sse83871. Фрагмент интрон/3'-UTR клонируют в KAWHIT03.0073 в противоположной ориентации в другом сайте интрона № 5 FAD3-1A сои, создавая KAWHIT03.0074 (смысловой интрон № 1 FAD2-1A сои, смысловая 3'-UTR FAD3-1A сои, смысловая 3'-UTR FATB-1 сои, сплайсируемый интрон № 5 FAD3-1A сои, антисмысловая 3'-UTR FATB-1 сои, антисмысловая 3'-UTR FAD3-1A сои, антисмысловой интрон № 1 FAD2-1A сои).
Стадия 12. Чтобы связать конструкцию дцРНКи с промотором 7Sальфа и 3'-концевой последовательностью TML, KAWHIT03.0074 и pMON68527 (кластер 7Sа'/3'-концевая последовательность TML) расщепляют SacI и лигируют друг с другом, создавая pMON68563 (промотор 7Sальфа - смысловой интрон № 1 FAD2-1A, смысловая 3'-UTR FAD3-1A сои, смысловая 3'-UTR FATB-1 сои, сплайсируемая антисмысловая 3'-UTR FATB-1 сои, антисмысловая 3'-UTR FAD3-1A сои, антисмысловой интрон № 1 FAD2-1A сои - 3'-концевая последовательность TML).
Стадия 13. Чтобы ввести собранную конструкцию дцРНКи в pMON70682, векторы pMON68563 и pMON70682 расщепляют NotI и лигируют друг с другом, создавая pMON68536, включающий промотор 7Sальфа, функционально связанный с образующей двухцепочечную РНК конструкцией смыслового интрона № 1 FAD2-1A, смысловой 3'-UTR FAD3-1A сои, смысловой 3'-UTR FATB-1 сои, сплайсируемого интрона № 5 FAD3-1A сои, антисмысловой 3'-UTR FATB-1 сои, антисмысловой 3'-UTR FAD3-1A сои, антисмыслового интрона № 1 FAD2-1A сои и терминатора 3'-концевой последовательности TML).
Стадия 14. pMON68536 расщепляют AscI и RsrII и pMON68529 (который содержит селектируемый маркер СР4, слитый с промотором FMV и 3'-концом RBCS, и промотор FAD2 сои, стимулирующий дельта-9-десатуразу) расщепляют SanDI и AscI. Часть дцРНКи вектора pMON68536 направленно клонируют в pMON68529, создавая pMON68537 (промотор 7Sальфа, функционально связанный с образующей двухцепочечную РНК конструкцией смыслового интрона № 1 FAD2-1a, смысловой 3'-UTR FAD3-1A сои, смысловой 3'-UTR FATB-1 сои, сплайсируемого интрона № 5 FAD3-1A сои, антисмысловой 3'-UTR FATB-1 сои, антисмысловой 3'-UTR FAD3-1A сои, антисмыслового интрона № 1 FAD2-1A сои и терминатора 3'-концевой последовательности TML, промотор FAD2 сои, стимулирующий дельта-9-десатуразу).
Пример 8
Следующие пятнадцать стадий иллюстрируют создание вектора pMON68539 (фиг.22), предназначенного для трансформации растений с целью подавления генов FAD2, FAD3 и FATB и сверхэкспрессии дельта-9-десатуразы и фермента KAS IV в сое. В частности, данная конструкция включает промотор 7Sальфа, функционально связанный с интроном и 3'-UTR сои в смысловой ориентации, то есть интрон № 1 FAD2-1A, 3'-UTR FAD3-1A, 3'-UTR FATB-1, шпилькообразный петлеобразующий сплайсируемый интрон, комплементарную серию интрона и 3'-UTR сои в антисмысловой ориентации, то есть 3'-UTR FATB-1, 3'-UTR FAD3-1A и интрон № 1 FAD2-1A, промотор FAD2 сои, стимулирующий дельта-9-десатуразу, и промотор напина, стимулирующий KAS IV.
Стадия 1. Интрон № 5 FAD3-1A сои, являющийся частью сплайсируемого интрона в конструкции дцРНКи, амплифицируют методом ПЦР, используя геномную ДНК сои в качестве матрицы, со следующими праймерами:
5'-концевой праймер = 19037 =
ACTAGTATATTGAGCTCATATTCCACTGCAGTGGATATTG
TTTAAACATAGCTAGCATATTACGCGTATATTATACAAGCTTATATTCCCGGGA
TATTGTCGACATATTAGCGGTACATTTTATTGCTTATTCAC
3'-концевой праймер = 19045 =
ACTAGTATATTGAGCTCATATTCCTGCAGGATATTCTCGAG
ATATTCACGGTAGTAATCTCCAAGAACTGGTTTTGCTGCTTGTGTCTGCAGTGAATC
Указанные праймеры добавляют сайты клонирования к 5'- и 3'-концам. К 5'-концу: SpeI, SacI, BstXI, PmeI, NheI, MluI, HindIII, XmaI, SmaI, SalI. К 3'-концу: SpeI, SacI, Sse8387I, XhoI. Интрон № 5 FAD3-1A сои, являющийся продуктом ПЦР, клонируют в pCR2.1, создавая KAWHIT03.0065. Затем KAWHIT03.0065 расщепляют SpeI и концы заполняют полимеразой Pfu, pMON68526 (пустой СМ-устойчивый вектор) расщепляют HindIII и концы заполняют полимеразой Pfu. KAWHIT03.0065 и pMON68526 лигируют, создавая pMON68541 (интрон № 5 FAD3-1A сои с сайтами множественного клонирования в АМР-устойчивом векторе).
Стадия 2. 3'-UTR FATB-1 сои амплифицируют со следующими праймерами:
18662 = TTTTAATTACAATGAGAATGAGATTTACTGC (добавление Bsp120I к 5'-концу) и 18661 = GGGCCCGATTTGAAATGGTTAACG. Продукт ПЦР затем лигируют с pCR2.1, создавая KAWHIT03.0036.
Стадия 3. KAWHIT03.0036 расщепляют Bsp120I и EcoRI и клонируют в KAWHIT03.0032 (пустой СМ-устойчивый вектор с сайтом множественного клонирования), создавая KAWHIT03.0037 (3'-UTR FATB-1 в пустом СМ-устойчивом векторе).
Стадия 4. 3'-UTR FAD3-1A сои амплифицируют со следующими праймерами:
18639 = GGGCCCGTTTCAAACTTTTTGG (добавление Bsp120I к 5'-концу) и 18549 = TGAAACTGACAATTCAA. Продукт ПЦР лигируют с pCR2.1, создавая KAWHIT03.0034.
Стадия 5. KAWHIT03.0034 расщепляют Bsp120I и EcoRI и лигируют с KAWHIT03.0032 (пустой СМ-устойчивый вектор с сайтом множественного клонирования), получая при этом KAWHIT03.0035 (3'-UTR FAD3-1A в пустом СМ-устойчивом векторе).
Стадия 6. Интрон № 1 FAD2-1A сои амплифицируют методом ПЦР, используя геномную ДНК сои в качестве матрицы, со следующими праймерами: 5'-концевой праймер = 18663 = GGGCCCGGTAAATTAAATTGTGC (добавление сайта Вsp120I к 5'-концу); и 3'-концевой праймер = 18664 = CTGTGTCAAAGTATAAACAAGTTCAG. Полученный продукт ПЦР клонируют в pCR2.1, создавая KAWHIT03.0038.
Стадия 7. Интрон № 1 FAD2-1A сои, являющийся продуктом ПЦР, в KAWHIT03.0038, клонируют в KAWHIT03.0032 (пустой СМ-устойчивый вектор с сайтом множественного клонирования), используя сайты рестрикции Bsp120I и EcoRI. Полученная плазмида представляет собой KAWHIT03.0039 (интрон № 1 FAD2-1A сои в путом СМ-устойчивом векторе).
Стадия 8. KAWHIT03.0039 расщепляют AscI и HindIII и вектор pMON68541 (АМР-устойчивый базовый вектор с дцРНКи интрона № 5 FAD3-1A) расщепляют MluI и HindIII. Интрон № 1 FAD2-1A сои направленно клонируют в pMON68541 (интрон № 5 FAD3-1A в АМР-устойчивом векторе с сайтами множественного клонирования), создавая KAWHIT03.0071 (интрон № 1 FAD2-1A сои с интроном № 5 FAD3-1A сои).
Стадия 9. KAWHIT03.0035 (3'-UTR FAD3-1A в СМ-устойчивом векторе) расщепляют AscI и HindIII и KAWHIT03.0071 (АМР-устойчивый базовый вектор с дцРНКи интрона FAD2-1A и интрона № 5 FAD3-1A) расщепляют МluI и HindIII. 3'-UTR FAD3-1A сои направленно клонируют в KAWHIT03.0071, создавая KAWHIT03.0072 (интрон № 1 FAD2-1A сои и 3'-UTR FAD3-1A с интроном № 5 FAD3-1A сои).
Стадия 10. KAWHIT03.0037 (3'-UTR FATB-1 в СМ-устойчивом векторе) расщепляют AscI и HindIII и KAWHIT03.0072 расщепляют MluI и HindIII. 3'-UTR FATB-1 направленно клонируют в KAWHIT03.0072, создавая KAWHIT03.0073 (интрон FAD2-1A, 3'-UTR FAD3-1A, 3'-UTR FATB-1 сои с интроном № 5 FAD3-1A).
Стадия 11. KAWHIT03.0073 расщепляют BstXI и SalI и фрагмент, содержащий интрон FAD2-1A, 3'-UTR FAD3-1A и 3'-UTR FATB-1, очищают гелем. В другой пробирке KAWHIT03.0073 расщепляют XhoI и Sse83871. Фрагмент интрон/3'-UTR клонируют в KAWHIT03.0073 в противоположной ориентации в другом сайте интрона № 5 FAD3-1A сои, создавая KAWHIT03.0074 (смысловой интрон № 1 FAD2-1A сои, смысловая 3'-UTR FAD3-1A сои, смысловая 3'-UTR FATB-1 сои, сплайсируемый интрон № 5 FAD3-1A сои, антисмысловая 3'-UTR FATB-1 сои, антисмысловая 3'-UTR FAD3-1A сои, антисмысловой интрон № 1 FAD2-1A сои).
Стадия 12. Чтобы связать конструкцию дцРНКи с промотором 7Sальфа и 3'-концевой последовательностью TML, KAWHIT03.0074 и pMON68527 (кластер 7Sа'/3'-концевая последовательность TML) расщепляют SacI и лигируют друг с другом, создавая pMON68563 (промотор 7Sальфа - смысловой интрон № 1 FAD2-1A, смысловая 3'-UTR FAD3-1A сои, смысловая 3'-UTR FATB-1 сои, сплайсируемая антисмысловая 3'-UTR FATB-1 сои, антисмысловая 3'-UTR FAD3-1A сои, антисмысловой интрон № 1 FAD2-1A сои - 3'-концевая последовательность TML).
Стадия 13. Чтобы ввести собранную конструкцию дцРНКи в pMON70682, векторы pMON68563 и pMON70682 расщепляют NotI и лигируют друг с другом, создавая pMON68536, включающий промотор 7Sальфа, функционально связанный с образующей двухцепочечную РНК конструкцией смыслового интрона № 1 FAD2-1A, смысловой 3'-UTR FAD3-1A сои, смысловой 3'-UTR FATB-1 сои, сплайсируемого интрона № 5 FAD3-1A сои, антисмысловой 3'-UTR FATB-1 сои, антисмысловой 3'-UTR FAD3-1A сои, антисмыслового интрона № 1 FAD2-1A сои и терминатора 3'-концевой последовательности TML).
Стадия 14. pMON68536 расщепляют AscI и RsrII и pMON68529 (который содержит селектируемый маркер СР4, слитый с промотором FMV и 3'-концом RBCS, и промотор FAD2 сои, стимулирующий дельта-9-десатуразу) расщепляют SanDI и AscI. Часть дцРНКи вектора pMON68536 направленно клонируют в pMON68529, создавая pMON68537 (промотор 7Sальфа, функционально связанный с образующей двухцепочечную РНК конструкцией смыслового интрона № 1 FAD2-1А, смысловой 3'-UTR FAD3-1A сои, смысловой 3'-UTR FATB-1 сои, сплайсируемого интрона № 5 FAD3-1A сои, антисмысловой 3'-UTR FATB-1 сои, антисмысловой 3'-UTR FAD3-1A сои, антисмыслового интрона № 1 FAD2-1A сои и терминатора 3'-концевой последовательности TML, промотор FAD2 сои, стимулирующий дельта-9-десатуразу).
Стадия 15. рMON68537 расщепляют SanDI и AscI и pMON70683 (напин, стимулирующий Kas IV) расщепляют AscI и RsrII. Фрагмент напин/Kas IV направленно клонируют в pMON68537, создавая pMON68539 (промотор 7Sальфа, функционально связанный с образующей двухцепочечную РНК конструкцией смыслового интрона № 1 FAD2-1A, смысловой 3'-UTR FAD3-1A сои, смысловой 3'-UTR FATB-1 сои, сплайсируемого интрона № 5 FAD3-1A сои, антисмысловой 3'-UTR FATB-1 сои, антисмысловой 3'-UTR FAD3-1A сои, антисмыслового интрона № 1 FAD2-1A сои и терминатора 3'-концевой последовательности TML, промотор FAD2 сои, стимулирующий дельта-9-десатуразу, и промотор напин, стимулирующий Kas IV).
Пример 9
Данный пример иллюстрирует трансформацию растений для получения растений сои с подавленными генами.
Трансформирующий вектор pMON68537, полученный в примере 7, используют для введения в сою образующей двухцепочечную РНК конструкции интрона/3'-UTR с целью подавления генов Δ12 десатуразы, Δ15 десатуразы и FATB. Вектор pMON68537 содержит также гены дельта-9-десатуразы (FAB2) и СР4. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм АВI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных дцРНКи-экспрессирующими конструкциями с интроном/3'-UTR. Составы масла в семенном пуле R1 и отдельном семени R1 свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 7). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты; подавление FAD3 позволяет получить растения с пониженным содержанием соединений сложного эфира линоленовой кислоты; и подавление FATB позволяет получить растения с пониженным содержанием соединений сложного эфира насыщенных жирных кислот, например пальмитатов и стеаратов. Отбор можно производить из таких линий в зависимости от требуемого относительного состава жирных кислот. Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкциями.
Пример 10
Данный пример иллюстрирует трансформацию растений для получения растений сои с подавленными генами.
Трансформирующий вектор pMON68539, полученный в примере 3, используют для введения в сою образующей двухцепочечную РНК конструкции интрона/3'-UTR с целью подавления генов Δ12 десатуразы, Δ15 десатуразы и FATB. Вектор pMON68539 содержит также гены Kas IV и СР4. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм АВI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных дцРНКи-экспрессирующими конструкциями с интроном/3'-UTR. Составы масла в семенном пуле R1 и отдельном семени R1 свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 8). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты; подавление FAD3 позволяет получить растения с пониженным содержанием соединений сложного эфира линоленовой кислоты; и подавление FATB позволяет получить растения с пониженным содержанием соединений сложного эфира насыщенных жирных кислот, например пальмитатов и стеаратов. Отбор можно производить из таких линий в зависимости от требуемого относительного состава жирных кислот. Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкциями.
Таблица 7
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON68537
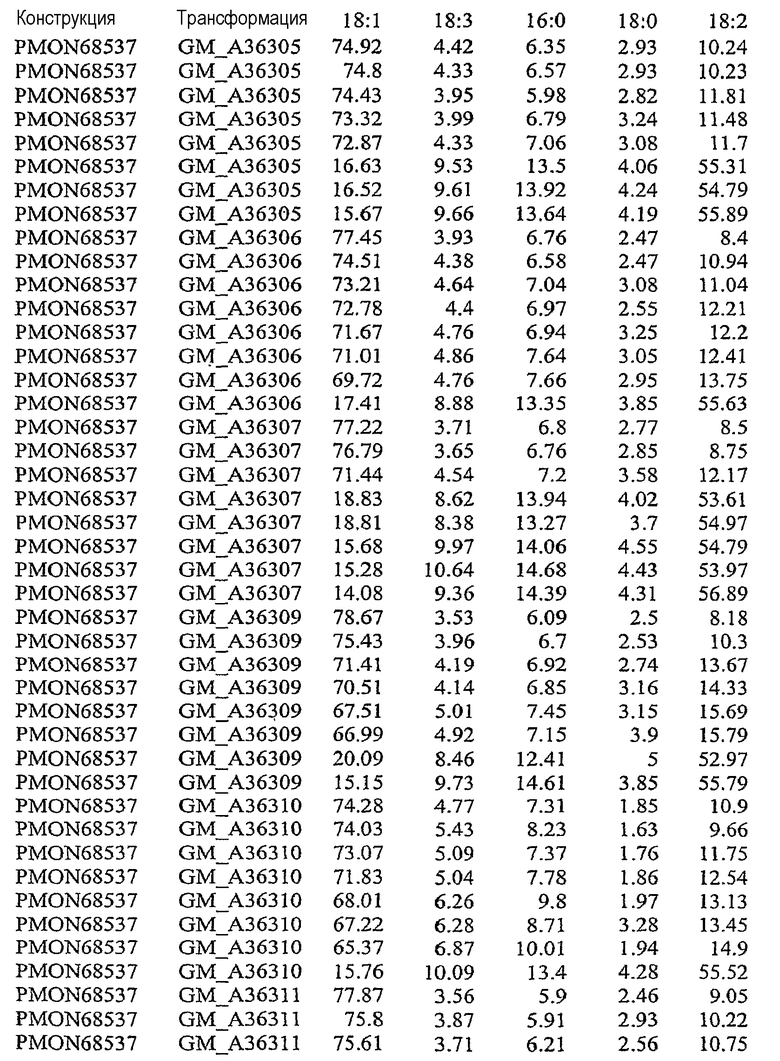
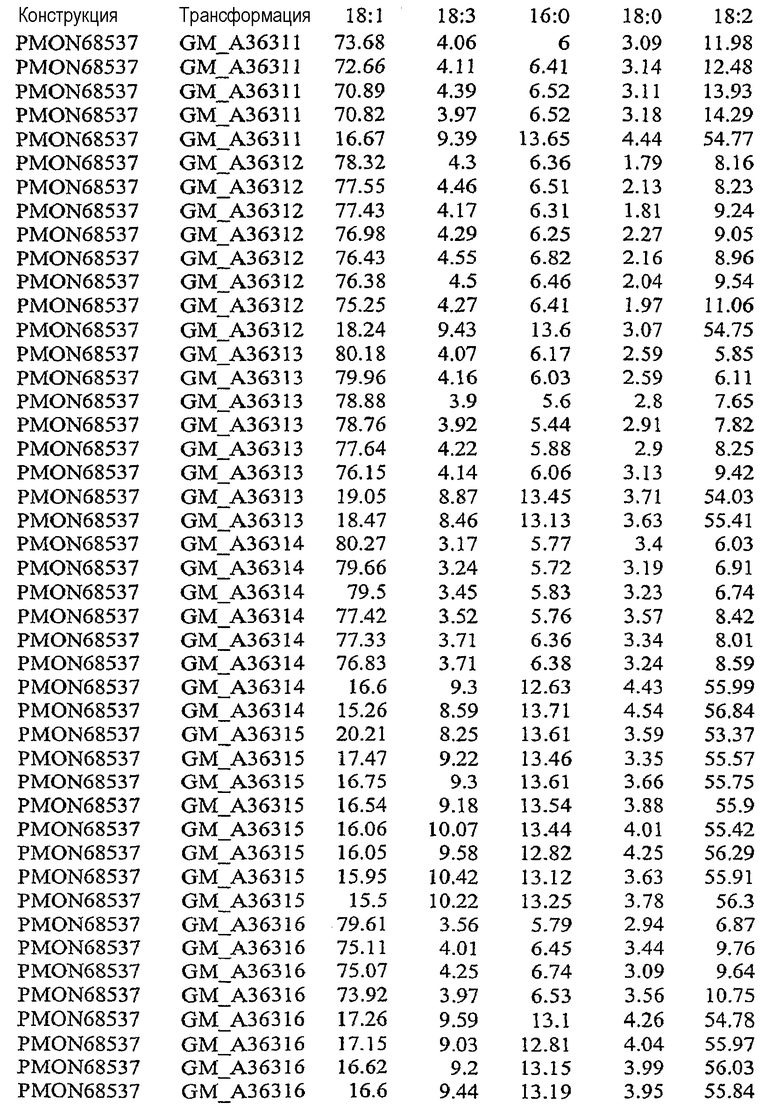
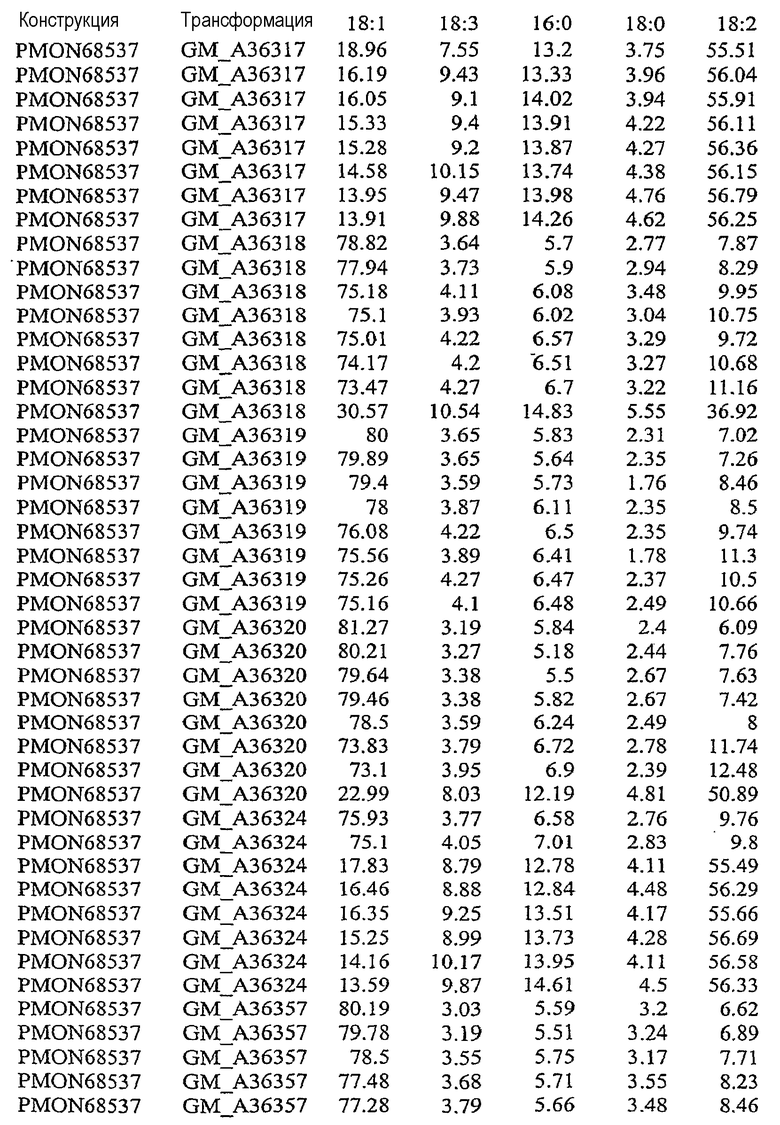
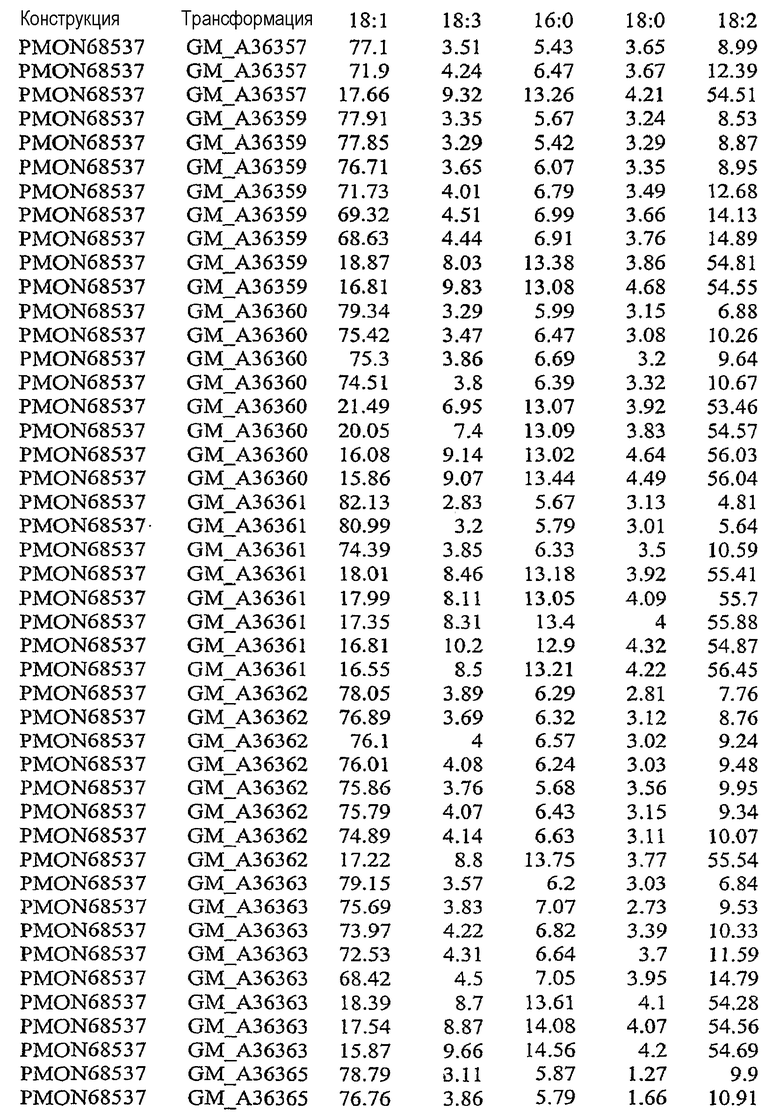
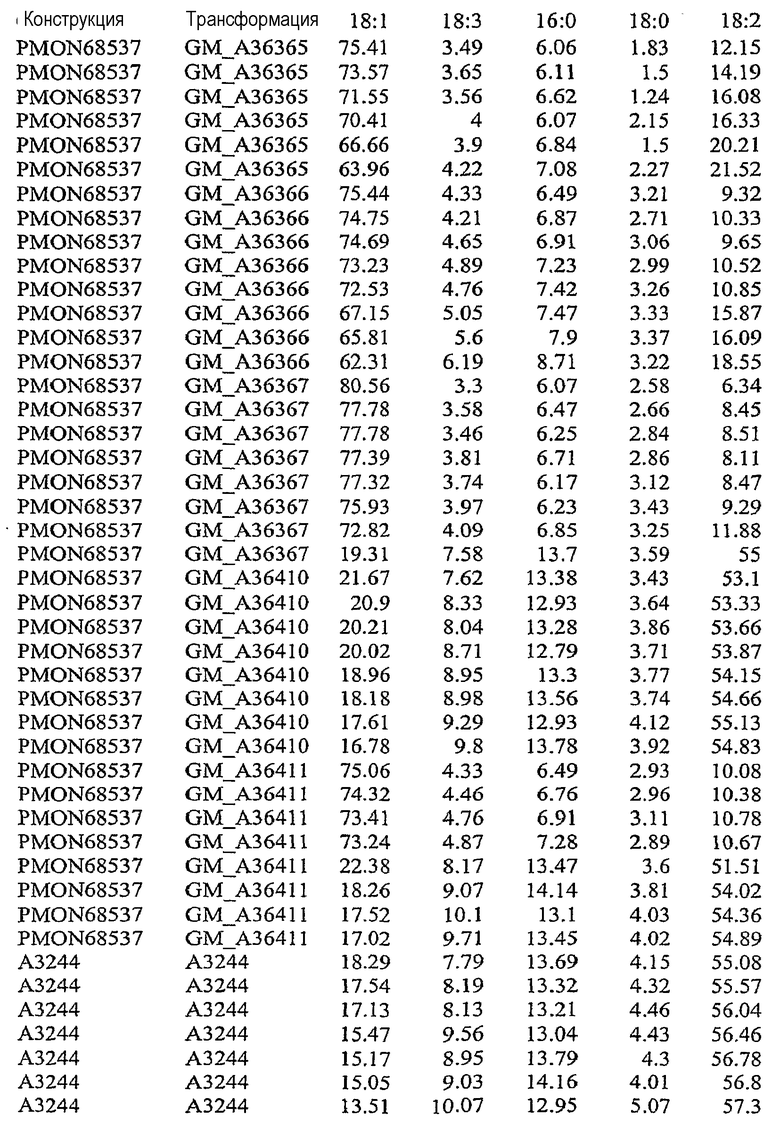

Таблица 8
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON68539
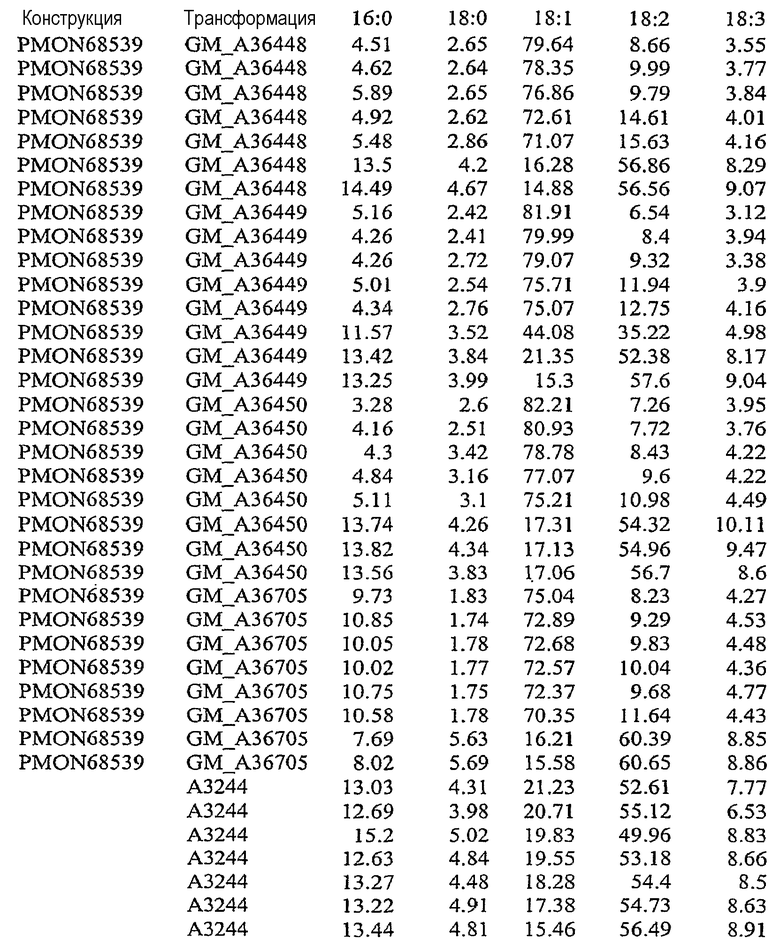
Пример 11
Конструкцию pMON95829, аналогичную описанной в примере 3D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1 с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат. Затем геномы трансформированных растений исследуют в отношении одновременного последовательного введения первой Т-ДНК и второй Т-ДНК, то есть со сборкой “правый край с правым краем”. Исследование выполняют методами картирования гибридизации с использованием саузерн-блоттинга. Трансформированные растения сои с предпочтительной конфигурацией в геноме переносят в теплицу для получения семян.
Например, у растений R0, трансформированных конструкцией pMON95829, берут ткань листа и анализируют методом саузерн-блоттинга. Зонды и гидролизаты, полученные с помощью рестрикционных ферментов, используют для идентификации трансформаций со сборкой обеих Т-ДНК с ориентацией правый край - правый край (“RB-RB”). Обычно примерно 25% всех трансформантов содержат правильно собранные Т-ДНК с ориентацией RB-RB.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON95829, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON95829, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 9). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 9
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON95829
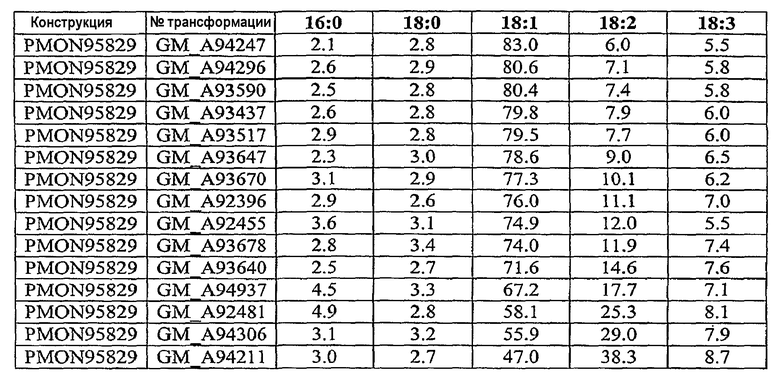
Пример 12
Конструкцию pMON93505, аналогичную описанной в примере 3D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат. Затем геномы трансформированных растений исследуют в отношении одновременной последовательной вставки первой Т-ДНК и второй Т-ДНК, то есть со сборкой “правый край с правым краем”. Исследование выполняют методами картирования гибридизации с использованием саузерн-блоттинга. Трансформированные растения сои с предпочтительной конфигурацией в геноме переносят в теплицу для получения семян.
Например, у растений R0, трансформированных конструкцией pMON93505, берут ткань листа и анализируют методом саузерн-блоттинга. Зонды и гидролизаты, полученные с помощью рестрикционных ферментов, используют для идентификации трансформаций со сборкой обеих Т-ДНК с ориентацией правый край - правый край (“RB-RB”). Обычно примерно 25% всех трансформантов содержат правильно собранные Т-ДНК с ориентацией RB-RB.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93505, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93505, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 10). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 10
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93505
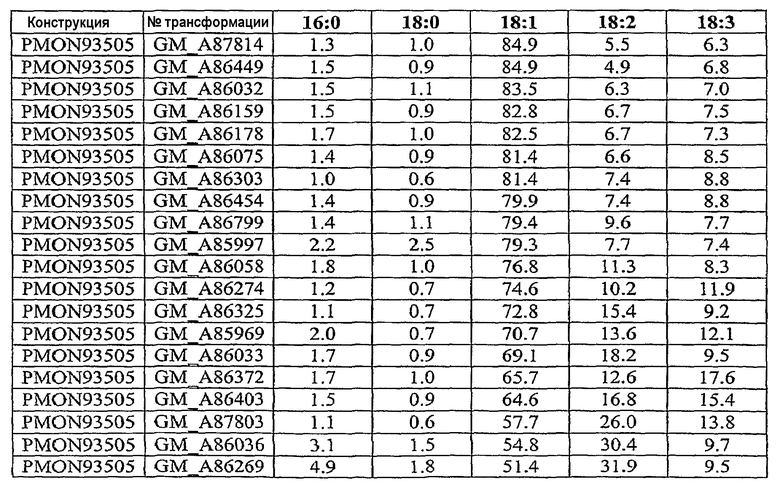
Пример 13
Конструкцию pMON93506, аналогичную описанной в примере 3D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат. Затем геномы трансформированных растений исследуют в отношении одновременной последовательной вставки первой Т-ДНК и второй Т-ДНК, то есть со сборкой “правый край с правым краем”. Исследование выполняют методами картирования гибридизации с использованием саузерн-блоттинга. Трансформированные растения сои с предпочтительной конфигурацией в геноме переносят в теблицу для получения семян.
Например, у растений R0, трансформированных конструкцией pMON93506, берут ткань листа и анализируют методом саузерн-блоттинга. Зонды и гидролизаты, полученные с помощью рестрикционных ферментов, используют для идентификации трансформаций со сборкой обеих Т-ДНК с ориентацией правый край - правый край (“RB-RB”). Обычно примерно 25% всех трансформантов содержат правильно собранные Т-ДНК с ориентацией RB-RB.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93506, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93506, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 11). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 11
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93506
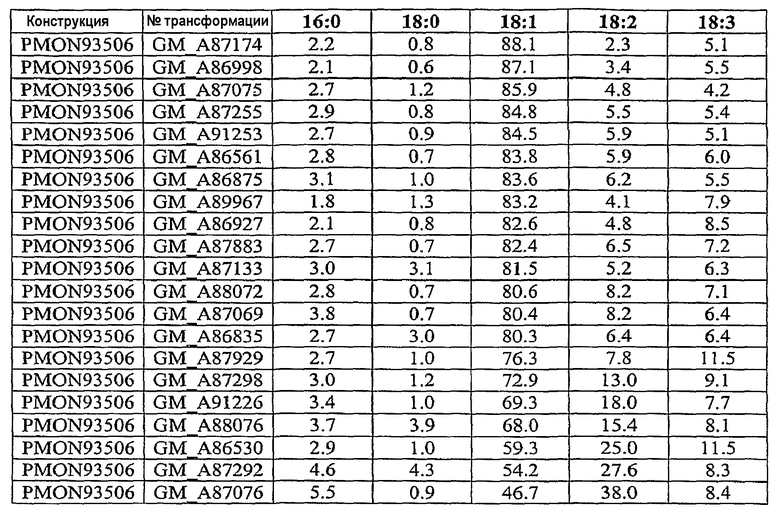
Пример 14
Конструкцию pMON93501, аналогичную описанной в примере 3В, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93501, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93501, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 12). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 12
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93501
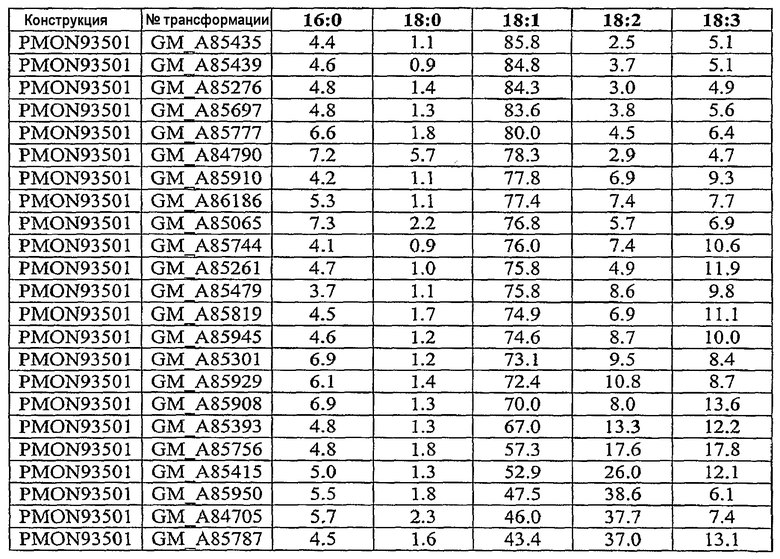
Пример 15
Конструкцию pMON97552, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON97552, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON97552, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 13). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 13
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON97552
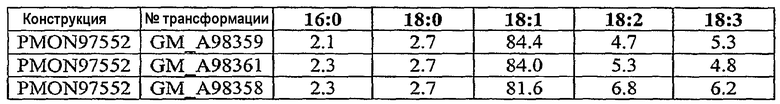
Пример 16
Конструкцию pMON93758, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93758, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93758, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 14). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 14
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93758
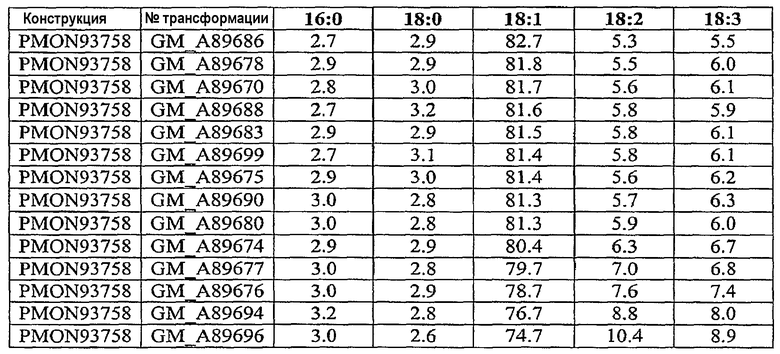
Пример 17
Конструкцию pMON97553, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON97553, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON97553, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 15). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 14
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON97553
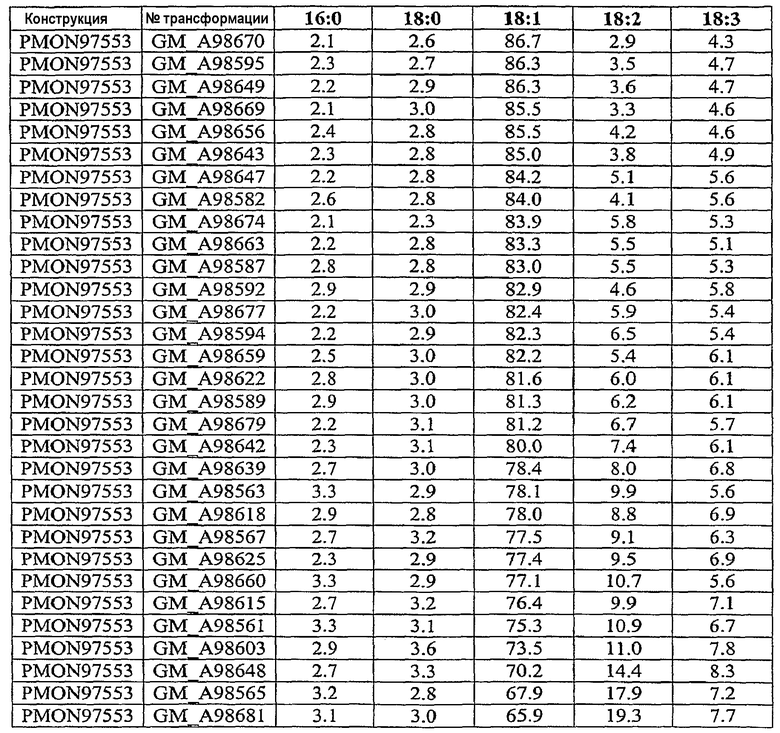
Пример 18
Конструкцию pMON93770, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93770, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93770, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 16). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 16
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93770
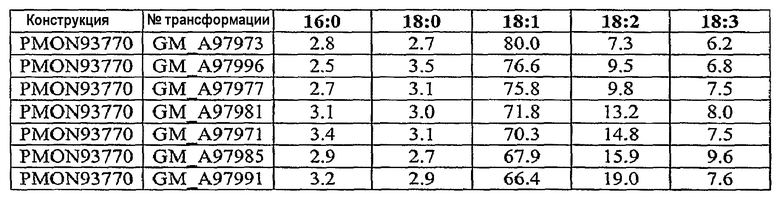
Пример 19
Конструкцию pMON93759, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93759, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93759, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 17). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 17
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93759
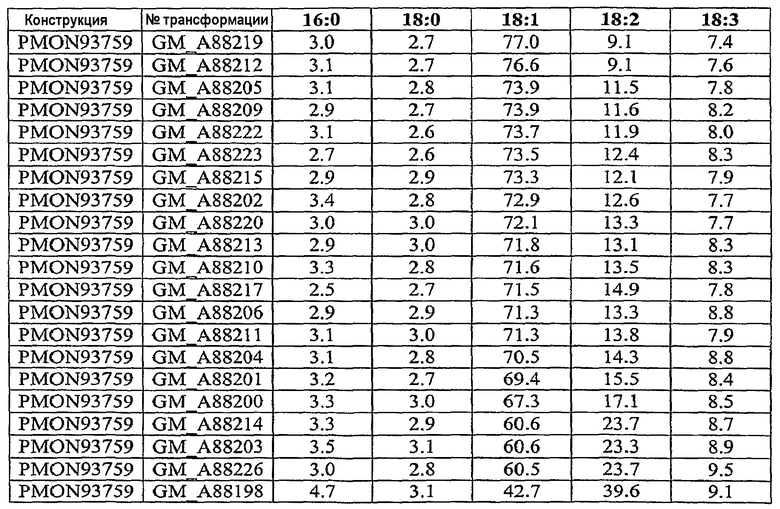
Пример 20
Конструкцию pMON97554, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON97554, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON97554, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 18). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 18
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON97554
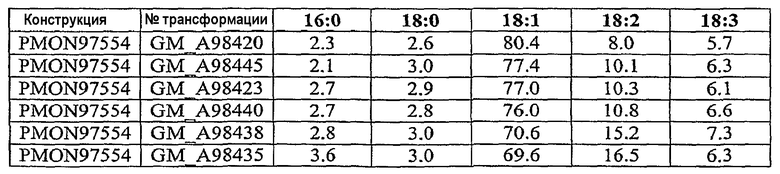
Пример 21
Конструкцию pMON93771, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93771, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93771, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 19). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 19
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93771
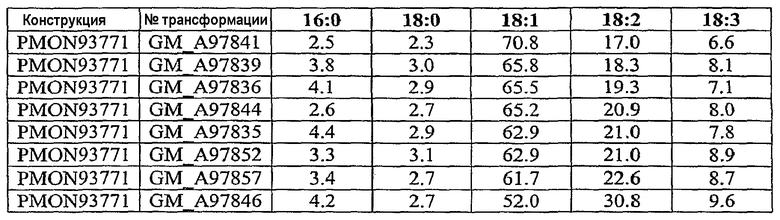
Пример 22
Конструкцию pMON97555, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON97555, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON97555, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 20). Например, подавление FAD2 позволяет получить растения с повышенным содержанием соединений сложного эфира олеиновой кислоты.
Таблица 20
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON97555
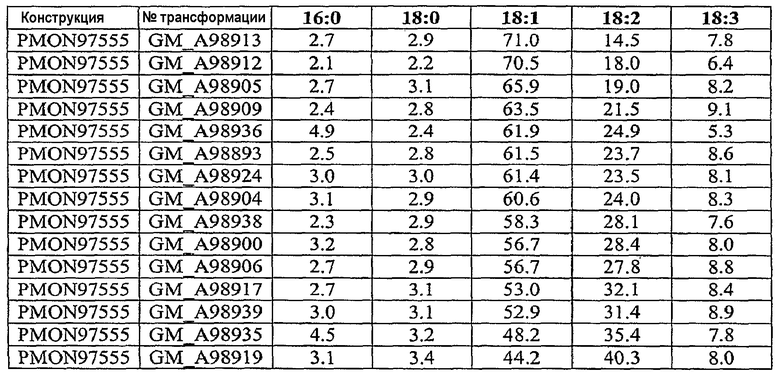
Пример 23
Конструкцию pMON93760, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93760, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93760, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 21). Например, интрон FAD2-1, уменьшенный на 320 последовательных нуклеотидов от 5'-конца (SEQ ID NO:1) и способный образовывать дцРНК, по крайней мере частично подавляет FAD2.
Таблица 21
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93760
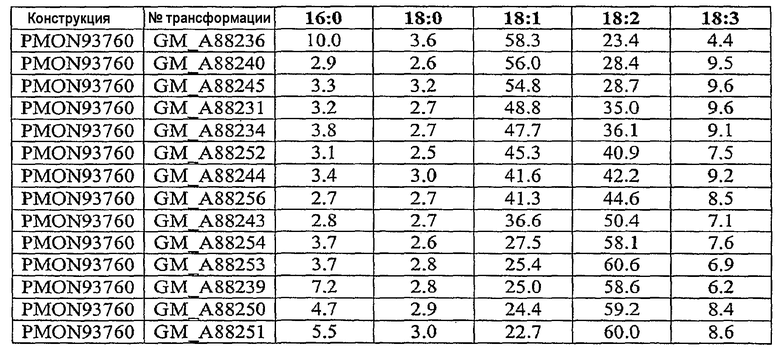
Пример 24
Конструкцию pMON93772, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93772, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93772, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 22). Например, интрон FAD2-1, уменьшенный на 360 последовательных нуклеотидов от 3'-конца (SEQ ID NO:1) и способный образовывать дцРНК, по крайней мере частично подавляет FAD2 в некоторых трансформациях.
Таблица 22
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93772
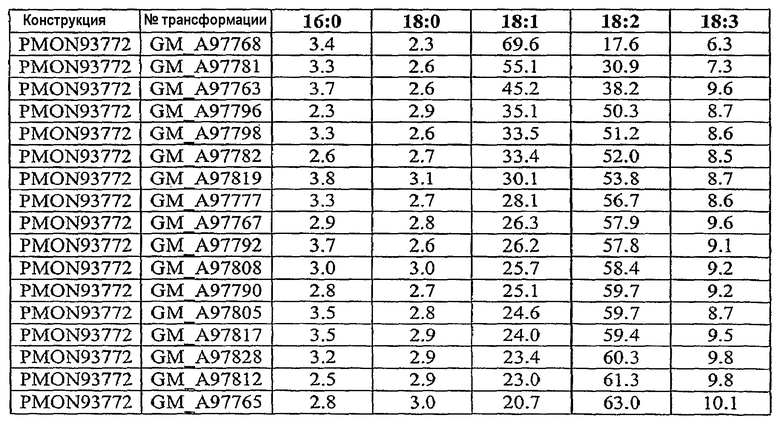
Пример 25
Конструкцию pMON97556, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON97556, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON97556, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов моно- и полиненасыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 23). Например, интрон FAD2-1, уменьшенный на 200 последовательных нуклеотидов от 3'-конца (SEQ ID NO:1) и способный образовывать дцРНК, по крайней мере частично подавляет FAD2.
Таблица 23
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON97556

Пример 26
Конструкцию pMON93764, аналогичную описанной в примере 2D, используют для введения в сою образующей двухцепочечную РНК конструкции интрона FAD2-1A с целью подавления гена FAD2. Указанный вектор устойчиво вводят в сою (Asgrow сорта А4922), используя штамм ABI Agrobacterium tumefaciens (Martinell, патент США № 6384301). Селектируемый маркер СР4 позволяет идентифицировать трансформированные растения сои путем отбора на среде, содержащей гербицид глифосат.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных конструкцией pMON93764, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных конструкцией pMON93764, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов насыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои (см. таблицу 24). Например, интрон FAD2-1, уменьшенный на 400 последовательных нуклеотидов от 3'-конца (SEQ ID NO:1) и способный образовывать дцРНК, по существу не уменьшает экспрессию FAD2.
Таблица 24
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON93764
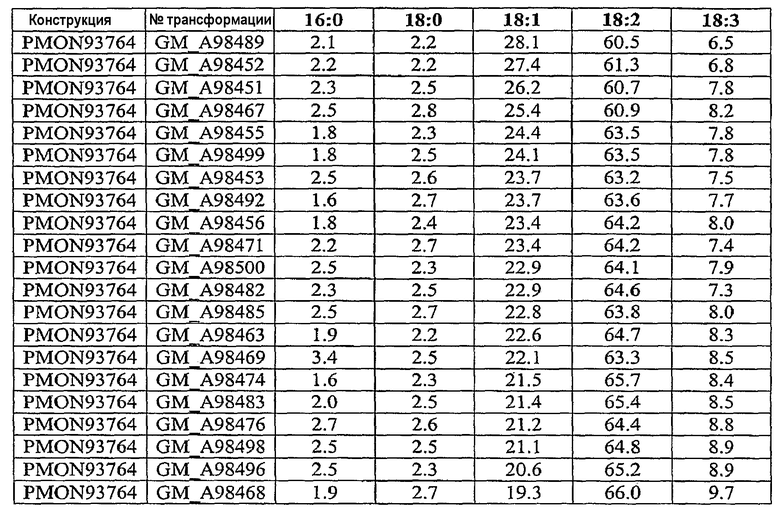
Пример 27
Анализ TaqMan предназначен для количественного определения нуклеиновых кислот путем избирательной амплификации и измерений флуоресценции в реальном времени (именуемый также ПЦР в реальном времени). Указанный метод используют для определения степени подавления транскрипта-мишени в развивающихся трансгенных семенах. Для определения абсолютных уровней транскрипта мРНК-мишени в образце при выполнении каждого эксперимента TaqMan строят стандартную кривую. Для указанной цели разные количества клонированной последовательности гена-мишени сои, разведенной в 20 нг общей РНК из канолы, амплифицируют параллельно с образцами неизвестных количеств мишени. Точность числа копий транскрипта, определенного данным методом, имеет предел погрешности, равный 25%.
Для получения материала матрицы общую РНК экстрагируют в устройстве для получения нуклеиновой кислоты ABI 6100 и используют 20 нг для каждого образца TaqMan. Образцы анализируют в устройстве для обнаружения последовательностей ABI 700 с помощью химии получения маточной смеси путем одностадийной ПЦР с обратной транскриптазой и призмы ABI. Затем строят график значений подсчета (Ct) методом TaqMan, полученных в результате выполнения реакции ПЦР TaqMan, в зависимости от известного количества синтетической последовательности-мишени для вычисления линейной регрессии с целью определения количества ДНК-мишени FAD2-1 в неизвестном образце на основании значений подсчета методом TaqMan, полученных в конце каждой реакции ПЦР TaqMan.
Растения трансформировали конструкциями pMON68540, pMON68546 или pMON80623, подавляющими FAD2-1A (см. раздел 3А и фиг.7 для ознакомления с описаниями конструкций).
Общую РНК получают из нулевых и трансформированных растений, используя устройство для получения нуклеиновой кислоты ABI 6100. Трансформированные растения являются гомозиготными растениями в третьем поколении и содержат олеиновую кислоту в количестве более 50%. Праймеры для FAD2-1A, праймеры для FAD2-1B или праймеры для FAD2-2A добавляют в отдельных образцах TaqMan к общей РНК из каждого испытуемого растения. Образцы анализируют в устройстве для обнаружения последовательностей ABI 700 с помощью химии получения маточной смеси путем одностадийной ПЦР с обратной транскриптазой и призмы ABI.
Все трансгенные растения в значительной степени снижают уровни транскриптов FAD2-1A и FAD2-1B. Ни одно из трансгенных растений даже частично не снижает уровни FAD2-2A или FAD2-2B.
Сравнение уровней транскриптов FAD2-1A в нулевых растениях позволяет выявить естественные расхождения между растениями. мРНК FAD2-1A в развивающихся семенах анализируют, используя праймеры для ПЦР, которые образуют последовательность зонда во многих растениях. Семена с массой сырой ткани 0,2 г получают из четырех разных нулевых растений-сегрегантов R2, относящихся к разным линиям. Испытанию подвергают пулы семян R2, имеющих одинаковый размер и полученных из четырех разных нулевых сегрегантов. Реакции ПЦР выполняют трижды и результаты нормализуют, сравнивая с количеством РНК 18S в каждом образце. Биологическая вариабельность транскриптов FAD2-1A у разных растений является низкой. Три из четырех образцов имеют нормализованное значение подсчета (Ct) методом TaqMan, равное примерно 65, и один из образцов имеет нормализованное значение подсчета методом TaqMan, равное примерно 50.
Пример 28
Фрагмент из 200 последовательных нуклеотидов последовательности интрона 1 FAD2-1 сои (SEQ ID NO:1) амплифицируют методом ПЦР, получая при этом продукты ПЦР, содержащие первые 200 нуклеотидов SEQ ID NO:1, начиная с 5'-конца SEQ ID NO:1. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты рекстрикции, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают рестрикционным ферментом и лигируют с вектором, содержащим ген CP4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco E9. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных указанной конструкцией, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных указанной конструкцией, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов насыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои.
Пример 29
Фрагмент из 180 последовательных нуклеотидов последовательности интрона 1 FAD2-1 сои (SEQ ID NO:1) амплифицируют методом ПЦР, получая при этом продукты ПЦР, содержащие первые 180 нуклеотидов SEQ ID NO:1, начиная с 3'-конца SEQ ID NO:1. Продукты ПЦР клонируют в смысловой ориентации в векторе, содержащем промотор 7Sα' сои и 3'-концевую терминирующую последовательность tml, используя сайты рекстрикции, созданные у 5'-концов праймеров для ПЦР. Данный вектор затем разрезают рестрикционным ферментом и лигируют с вектором, содержащим ген CP4 EPSPS, регулируемый промотором FMV и 3'-концевой терминирующей последовательностью гороха Rubisco E9. Полученную экспрессирующую ген конструкцию используют для трансформации методами, представленными в настоящем описании изобретения.
Составы жирных кислот анализируют газовой хроматографией в семени линий сои, трансформированных указанной конструкцией, в соответствии с описанием, приведенным в примере 4, для идентификации соединений сложных метиловых эфиров жирных кислот, экстрагированных из семян. Сначала собирают шесть семян R1 из растений сои, трансформированных указанной конструкцией, и определяют состав жирных кислот в каждом отдельном семени. Так как растения R1, полученные в результате каждой трансформации, расщепляются на трансгены, то они дают семена как с обычным составом, так и модифицированные варианты. Положительные семена собирают и усредняют для каждой трансформации. Собранные положительные усредненные семена свидетельствуют об изменении составов насыщенных жирных кислот в масле из семян линий трансгенной сои по сравнению с маслом из семян нетрансформированной сои.
Пример 30
pMON97562 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), который уменьшен на 100 последовательных нуклеотидов от 3'-конца и связан с 5'-UTR FAD3-1A, за которой следует 3'-UTR FAD3-1A, связанная с 5'-UTR FAD3-1В, за которой следует 3'-UTR FAD3-1B, за которой следует 5'-UTR FATB-1a, за которой следует 3'-UTR FATB-1a, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с 3'-UTR FATB-1a в антисмысловой ориентации, за которой следует 5'-UTR FATB-1a в антисмысловой ориентации, связанная с 3'-UTR FAD3-1B в антисмысловой ориентации, за которой следует 5'-UTR FAD3-1B в антисмысловой ориентации, связанная с 3'-UTR FAD3-1A в антисмысловой ориентации, за которой следует 5'-UTR FAD3-1A в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 100 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3'-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco Е9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации сои методами, представленными в настоящем описании изобретения. Составы жирных кислот определяют газовой хроматографией в семени линий сои, трансформированных указанной конструкцией, в соответствии с описанием, приведенным в примере 4. В таблице 25 приведены составы типичных семян. Уровень 18:3 снижен примерно на 1%.
Таблица 25
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON97562

Пример 31
pMON97563 содержит промотор 7Sα' сои, функционально связанный с интроном 1 FAD2-1A сои (SEQ ID NO:1), который уменьшен на 100 последовательных нуклеотидов от 3'-конца и связан с 5'-UTR FAD3-1A, за которой следует 3'-UTR FAD3-1A, связанная с 5'-UTR FAD3-1B, за которой следует 3'-UTR FAD3-1B, связанная с 5'-UTR FAD3-1C, за которой следует 3'-UTR FAD3-1C, за которой следует кодирующая область СТР FATB-1a, за которой следует кодирующая область СТР FATB-2a, функционально связанная с 70 нуклеотидами интрона 4 FAD3-1A, функционально связанного с кодирующей областью СТР FATB-2a в антисмысловой ориентации, за которой следует кодирующая область СТР FATB-1a в антисмысловой ориентации, связанная с 3'-UTR FAD3-1C в антисмысловой ориентации, за которой следует 5'-UTR FAD3-1C в антисмысловой ориентации, связанная с 3'-UTR FAD3-1B в антисмысловой ориентации, за которой следует 5'-UTR FAD3-1B в антисмысловой ориентации, связанная с 3'-UTR FAD3-1А в антисмысловой ориентации, за которой следует 5'-UTR FAD3-1A в антисмысловой ориентации, за которой следует интрон 1 FAD2-1A сои (SEQ ID NO:1), уменьшенный на 100 последовательных нуклеотидов от 3'-конца и находящийся в антисмысловой ориентации, который функционально связан с 3-концевым сегментом полиаденилирования Н6 с геном СР4 EPSPS, функционально связанным с промотором EFMV и 3'-концевой терминирующей последовательностью гороха Rubisco E9, при этом все последовательности фланкированы правым краем (RB) и левым краем (LB) в одной молекуле ДНК. Полученную экспрессирующую ген конструкцию используют для трансформации растений методами, представленными в настоящем описании изобретения. Составы жирных кислот определяют газовой хроматографией в семени линий сои, трансформированных указанной конструкцией, в соответствии с описанием, приведенным в примере 4. В таблице 26 приведены составы типичных семян. Уровень 18:3 снижен примерно на 1%.
Таблица 26
Состав жирных кислот в отдельных семенах R1, полученных в результате трансформаций pMON97563
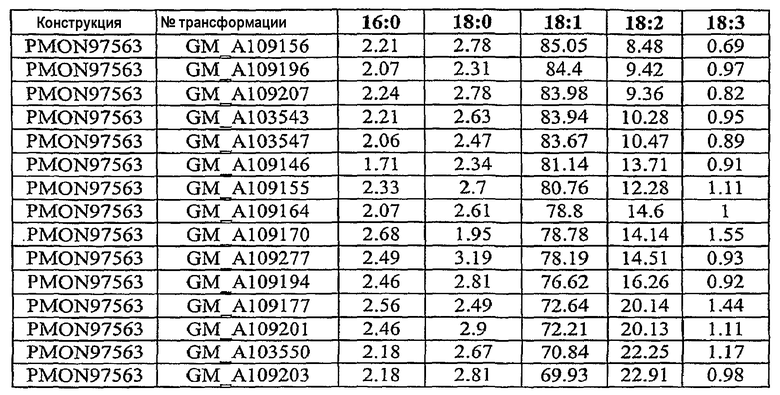
Все составы, представленные в настоящем описании изобретения и формуле изобретения, могут быть получены и/или способы получения указанных составов могут быть выполнены без ненужного экспериментирования с учетом настоящего описания изобретения. Несмотря на то что составы по настоящему изобретению и способы их получения описаны в виде предпочтительных вариантов осуществления изобретения, специалистам в данной области должно быть очевидно, что в составы и/или способы, а также в стадии или последовательность стадий способов по настоящему изобретению могут быть внесены изменения, не выходящие за пределы объема и существа изобретения. В частности, очевидно, что вместо агентов, рассмотренных в настоящем описании изобретения, могут быть использованы другие агенты, родственные в химическом и физиологическом отношении, при достижении аналогичных или подобных результатов. Все подобные замены и модификации, очевидные специалистам в данной области, находятся в пределах существа, объема и идеи изобретения, определяемых в прилагаемой формуле изобретения.

































| название | год | авторы | номер документа |
|---|---|---|---|
| СКОНСТРУИРОВАННАЯ СПОСОБАМИ ИНЖЕНЕРИИ ПЛАТФОРМА ДЛЯ ВСТРАИВАНИЯ ТРАНСГЕНА (ETIP) ДЛЯ НАЦЕЛИВАНИЯ ГЕНОВ И СТЭКИНГА ПРИЗНАКОВ | 2013 |
|
RU2666916C2 |
| КОНСТРУИРУЕМЫЕ БЕЛКИ С ЦИНКОВЫМИ ПАЛЬЦАМИ, НАПРАВЛЕННЫЕ НА ГЕНЫ РАСТЕНИЙ, ВОВЛЕЧЕННЫЕ В БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ | 2010 |
|
RU2712833C2 |
| КОНСТРУИРУЕМЫЕ БЕЛКИ С ЦИНКОВЫМИ ПАЛЬЦАМИ, НАПРАВЛЕННЫЕ НА ГЕНЫ РАСТЕНИЙ, ВОВЛЕЧЕННЫЕ В БИОСИНТЕЗ ЖИРНЫХ КИСЛОТ | 2010 |
|
RU2568056C2 |
| ОБОГАЩЕНИЕ АКТИВИРУЕМОЙ ФЛУОРЕСЦЕНЦИЕЙ СОРТИРОВКИ КЛЕТОК (FACS) ДЛЯ СОЗДАНИЯ РАСТЕНИЙ | 2013 |
|
RU2679510C2 |
| Brassica juncea КАЧЕСТВА ОМЕГА-9 | 2009 |
|
RU2557316C2 |
| КАНОЛА HO/LL С УСТОЙЧИВОСТЬЮ К ЗАБОЛЕВАНИЮ КИЛОЙ КРЕСТОЦВЕТНЫХ | 2012 |
|
RU2618846C2 |
| КАНОЛА HO/LL С УСТОЙЧИВОСТЬЮ К ЗАБОЛЕВАНИЮ КИЛОЙ КРЕСТОЦВЕТНЫХ | 2012 |
|
RU2711934C2 |
| ЛОКУСЫ FAD3 ДЛЯ ВЫПОЛНЕНИЯ ОПЕРАЦИЙ И СООТВЕТСТВУЮЩИЕ СВЯЗЫВАЮЩИЕСЯ СО СПЕЦИФИЧЕСКИМИ САЙТАМИ-МИШЕНЯМИ БЕЛКИ, СПОСОБНЫЕ К ВЫЗОВУ НАПРАВЛЕННЫХ РАЗРЫВОВ | 2013 |
|
RU2665811C2 |
| ЛОКУСЫ ФУНКЦИОНАЛЬНОСТИ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ К УЧАСТКУ-МИШЕНИ СВЯЗЫВАЮЩИЕ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656159C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
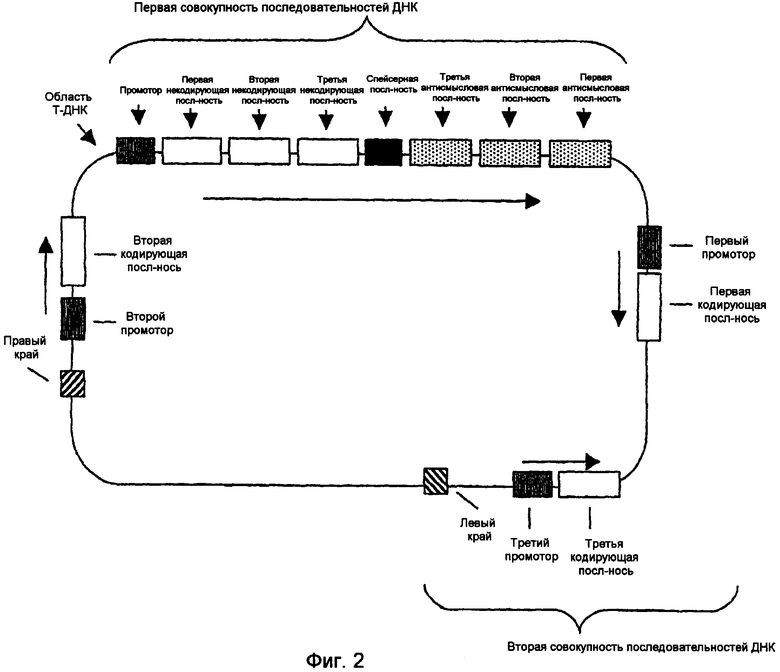
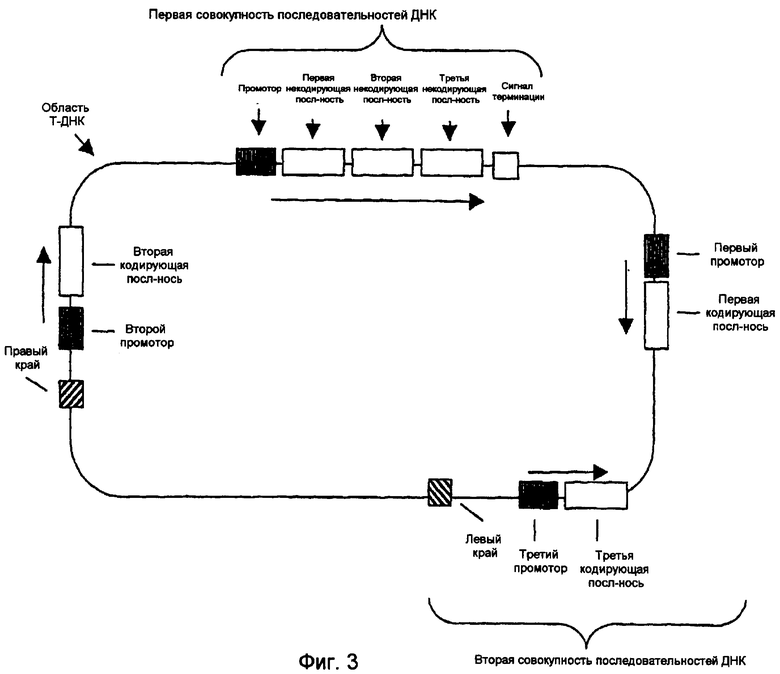
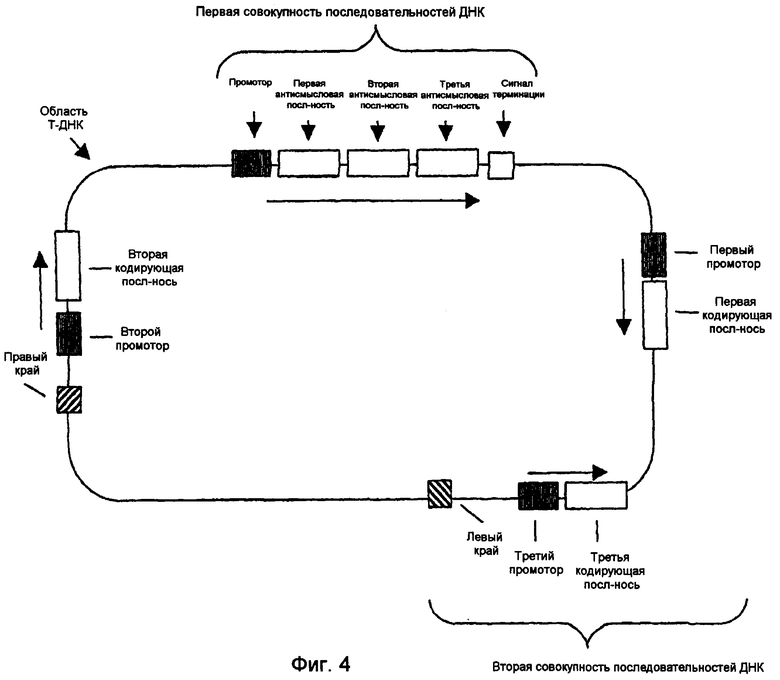
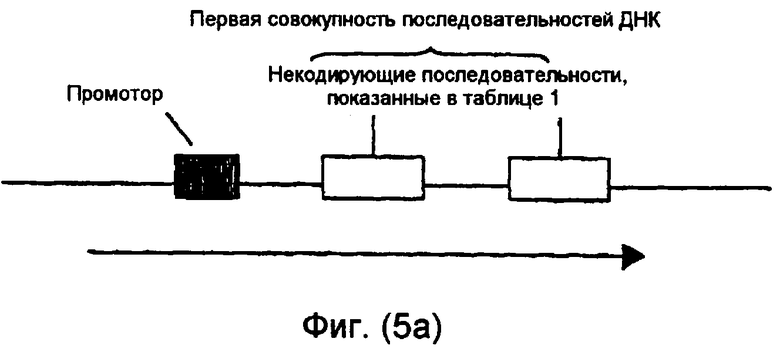
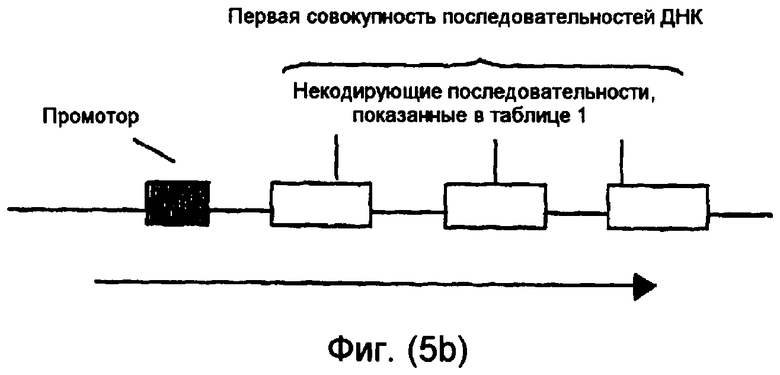

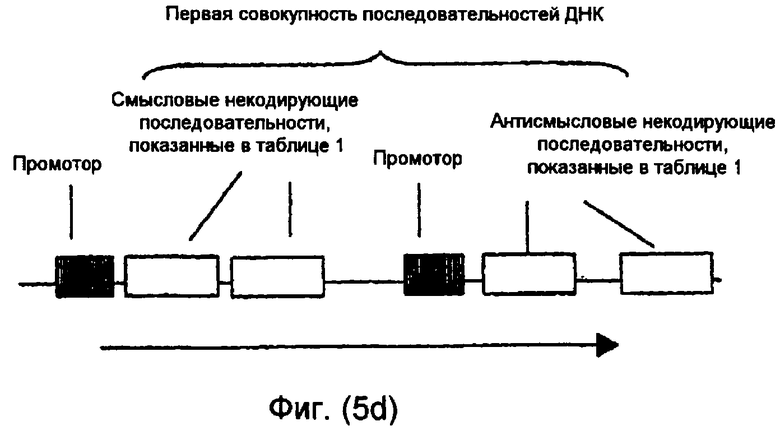
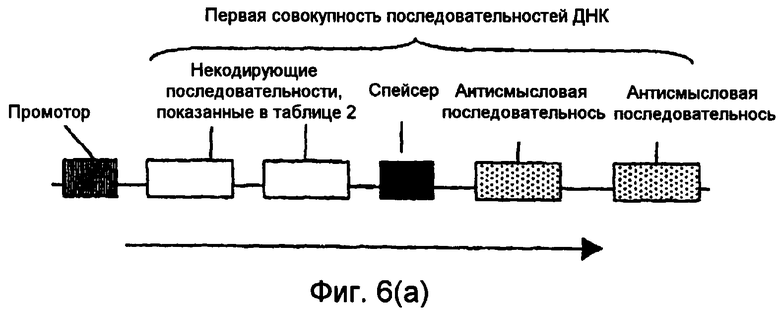
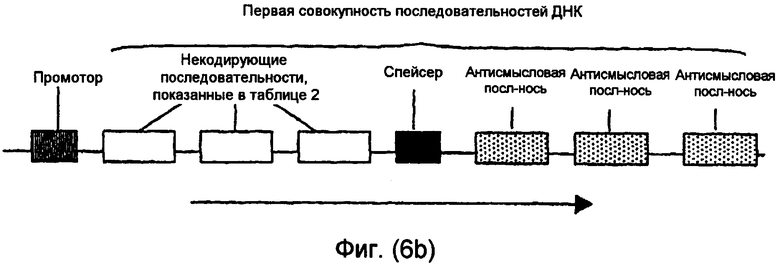
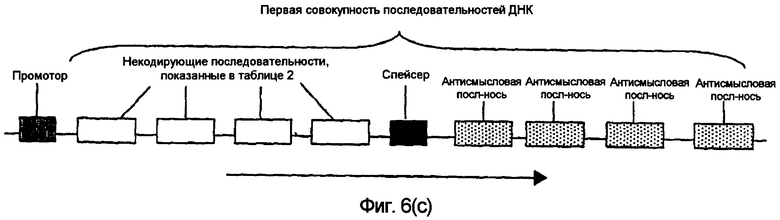
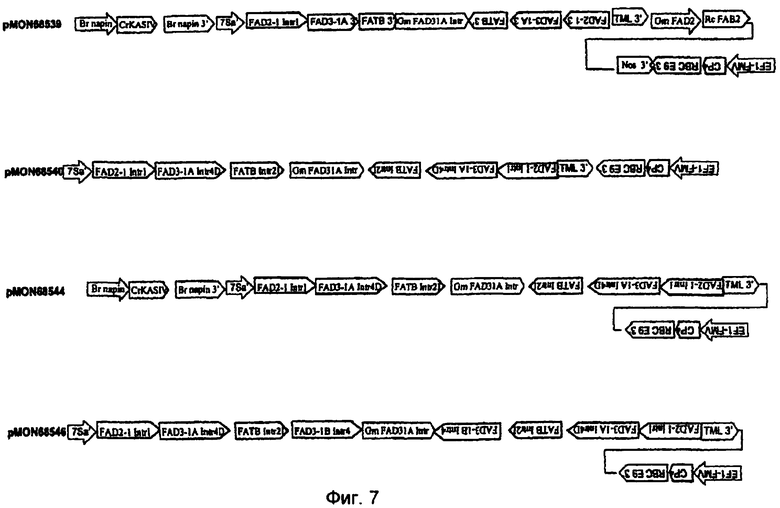



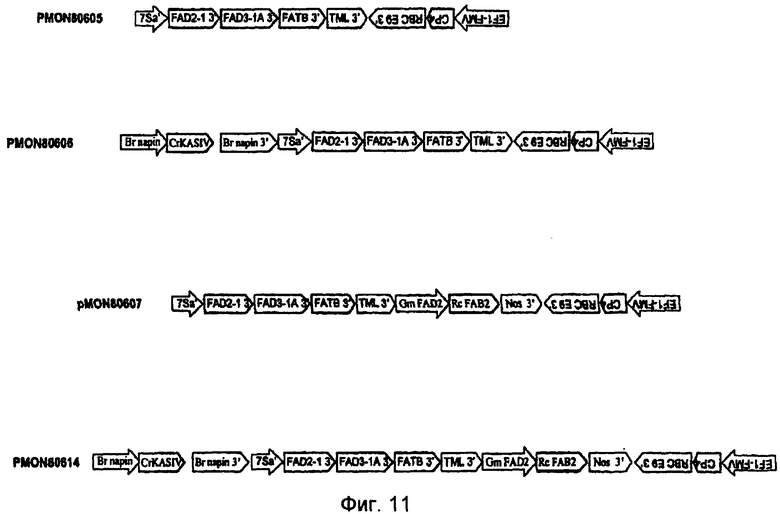

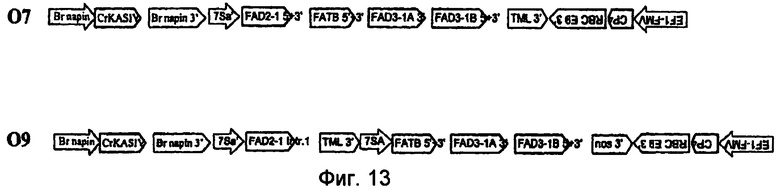
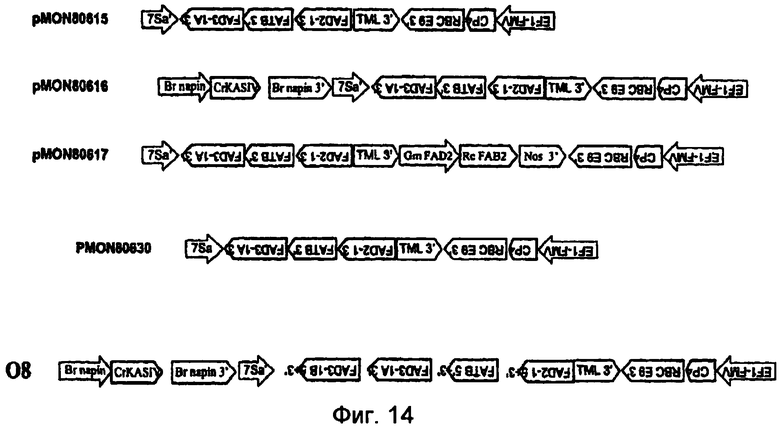
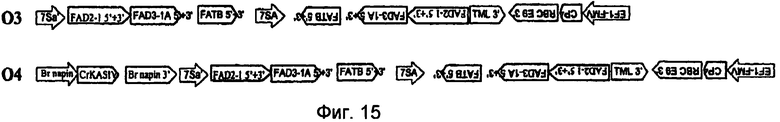

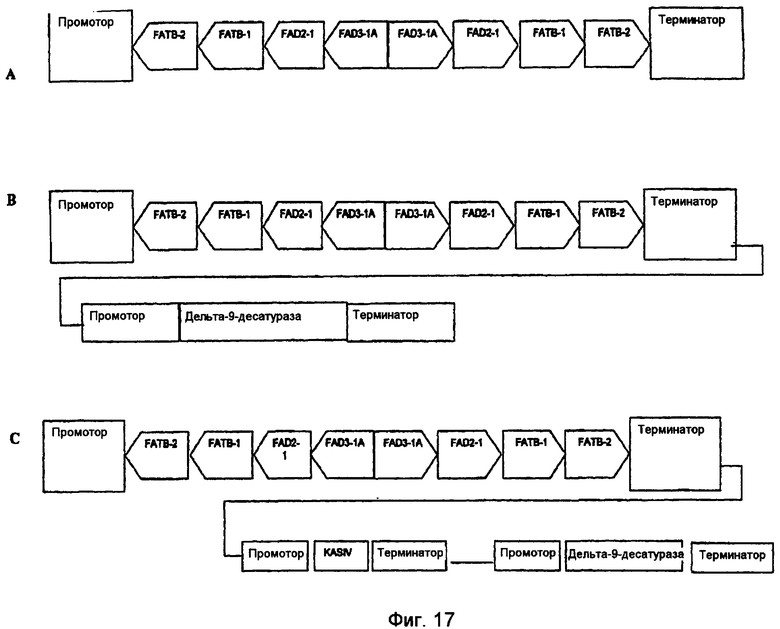
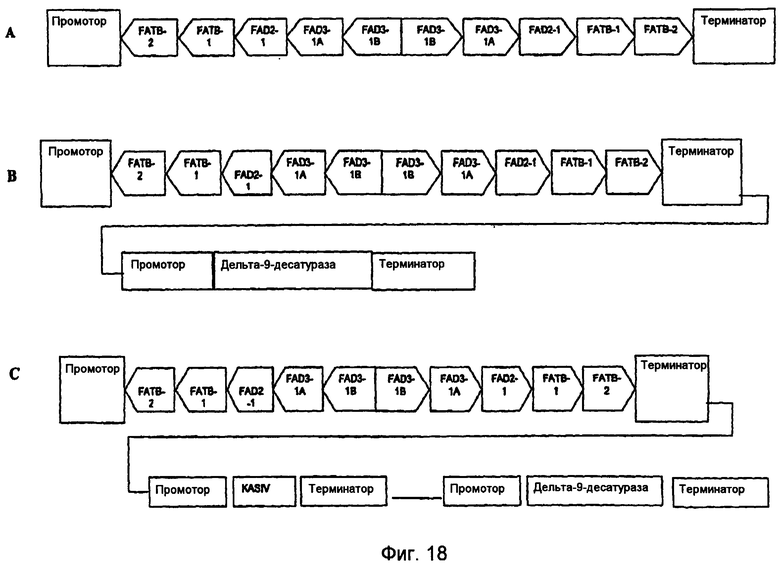
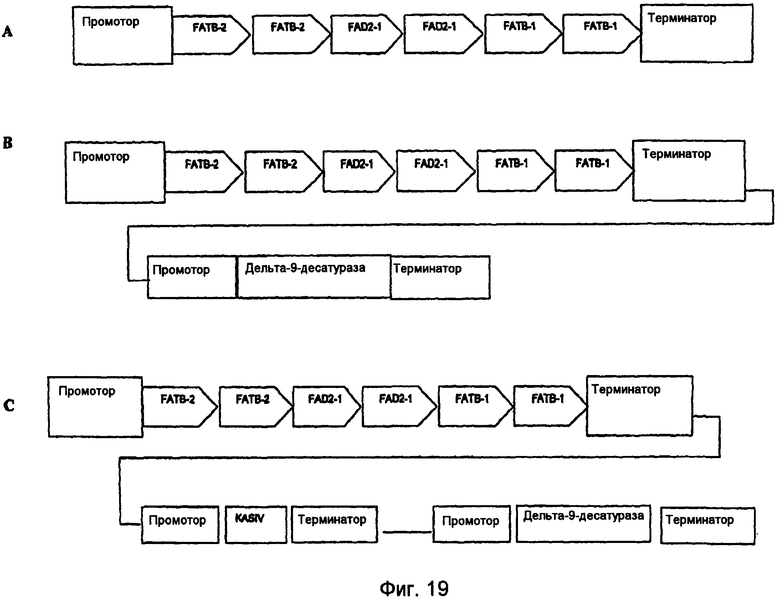
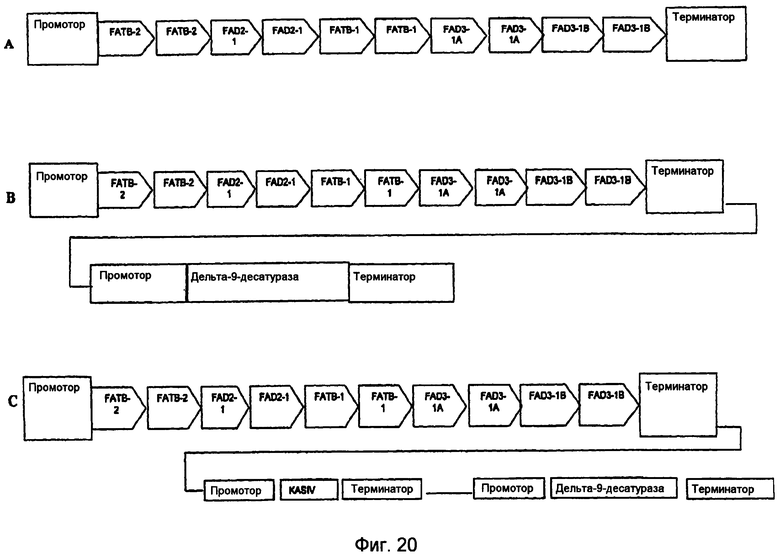
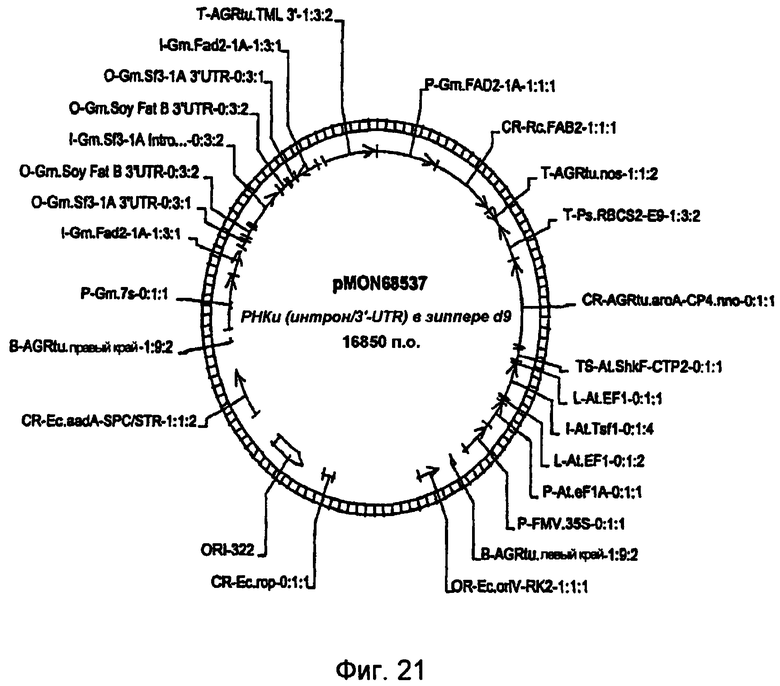
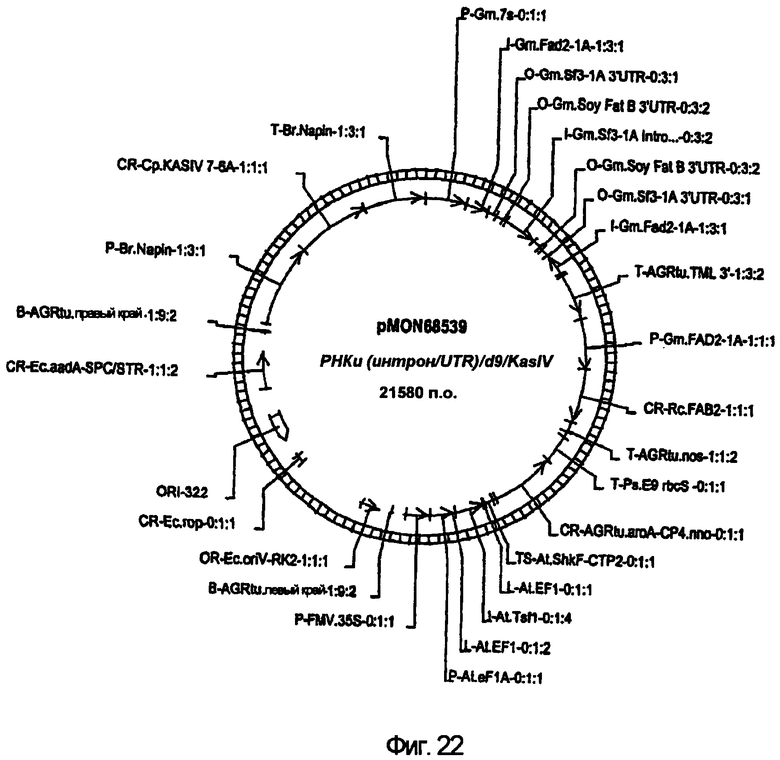
Трансформированное растение сои содержит последовательность нуклеиновой кислоты, идентичную фрагменту интрона FAD2-1A сои, и последовательность нуклеиновой кислоты, идентичную фрагменту гена FADB сои. Семя трансформированного растения, а также полученные из него масло и мука содержат олеиновую кислоту в количестве от примерно 42 мас.% до примерно 85 мас.% от общего содержания жирных кислот, при этом содержание насыщенных жирных кислот от общего их количества составляет менее 8 мас.%. 5 н. и 23 з.п. ф-лы, 22 ил., 26 табл.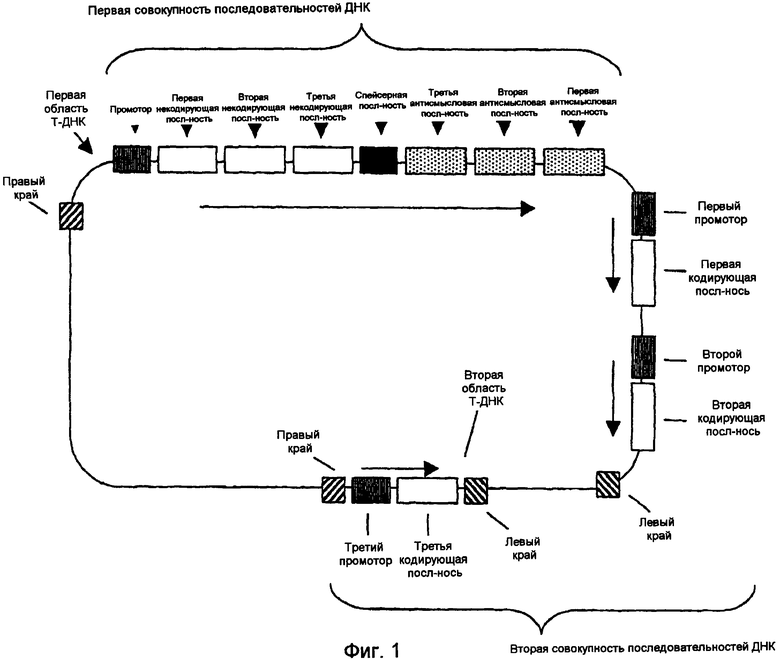
1. Семя сои, характеризующееся составом жирных кислот в масле из семян, в котором содержание олеиновой кислоты составляет от около 42 до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет менее 8 мас.% от общего содержания жирных кислот, где указанное растение дополнительно содержит геном, включающий:
(i) первую последовательность нуклеиновой кислоты, по меньшей мере, на 90% идентичную фрагменту интрона FAD2-1A сои длиной между 20 и 420 последовательных нуклеотидов, и
(ii) вторую последовательность нуклеиновой кислоты, по меньшей мере, на 90% идентичную фрагменту гена FADB сои длиной между 40 и 450 последовательных нуклеотидов.
2. Семя сои по п.1, где указанный геном дополнительно включает последовательность нуклеиновой кислоты, повышающую экспрессию бета-кероацил-АСР-синтазы IV или дельта-9-десатуразы либо обеих вместе.
3. Семя сои по п.1, в котором указанный фрагмент указанного интрона FAD2-1 сои и указанный фрагмент указанного гена FATB сои транскрибированы в смысловой и антисмысловой ориентации с образованием РНК, которая по крайней мере частично является двухцепочечной.
4. Семя сои по п.1, в котором указанная последовательность нуклеиновой кислоты, подавляющая экспрессию эндогенных гена FAD2-1 и гена FATB сои, собирается в виде функциональной единицы транскрипции после введения в хромосому растения.
5. Семя сои по п.4, в котором указанный фрагмент указанного интрона FAD2-1 сои и указанный фрагмент указанного гена FATB сои транскрибированы в смысловой и антисмысловой ориентации с образованием РНК, которая по крайней мере частично является двухцепочечной.
6. Семя сои по п.1, где первая последовательность нуклеиновой кислоты, по меньшей мере, на 90% идентичная фрагменту интрона FAD2-1А сои длиной между 20 и 420 последовательных нуклеотидов, подавляет экспрессию эндогенного FAD2-1 сои и указанный геном дополнительно содержит последовательность нуклеиновой кислоты, повышающую экспрессию бета-кетоацил-АСР-синтазы IV или дельта-9-десатуразы либо обеих вместе.
7. Клетка семени сои, характеризующегося составом жирных кислот в масле из семян, в котором содержание олеиновой кислоты составляет от около 42 до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет менее 8 мас.% от общего содержания жирных кислот, где указанное семя содержит:
(i) последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичную фрагменту интрона FAD2-1А сои длиной между 20 и 420 последовательных нуклеотидов, и
(ii) последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичную фрагменту гена FADB сои длиной между 40 и 450 последовательных нуклеотидов.
8. Несмешанное соевое масло, полученное из семени, характеризующегося составом жирных кислот, в котором содержание олеиновой кислоты составляет от около 42 до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет около 8 мас.% или меньше от общего содержания жирных кислот, где указанное семя дополнительно содержит геном, включающий:
(i) последовательность нуклеиновой кислоты, по меньшей мере, на 90% идентичную фрагменту интрона FAD2-1A сои длиной между 20 и 420 последовательных нуклеотидов, и
(ii) последовательность нуклеиновой кислоты, по меньшей мере, на 90% идентичную фрагменту гена FADB сои длиной между 40 и 450 последовательных нуклеотидов.
9. Соевая мука, полученная из семени сои, характеризующегося составом жирных кислот в масле из семян, в котором содержание олеиновой кислоты составляет от около 42 до около 85 мас.% от общего содержания жирных кислот и содержание насыщенных жирных кислот составляет менее 8 мас.% от общего содержания жирных кислот, где указанное семя дополнительно содержит геном, включающий:
(i) последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичную фрагменту интрона FAD2-1A сои длиной между 20 и 420 последовательных нуклеотидов, и
(ii) последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичную фрагменту гена FADB сои длиной между 40 и 450 последовательных нуклеотидов.
10. Несмешанное соевое масло, характеризующееся составом жирных кислот, в котором содержание олеиновой кислоты составляет от около 42 до около 85 мас.% от общего содержания жирных кислот, содержание насыщенных жирных кислот составляет около 8 мас.% или меньше от общего содержания жирных кислот и содержание линоленовой кислоты составляет около 1,5 мас.% или меньше от общего содержания жирных кислот.
11. Семя сои по п.1, где содержание олеиновой кислоты составляет от около 50 до около 80 мас.% от общего содержания жирных кислот.
12. Семя сои по п.1, где содержание олеиновой кислоты составляет от около 46 до около 75 мас.% от общего содержания жирных кислот.
13. Семя сои по п.2, где содержание олеиновой кислоты составляет от около 50 до около 80 мас.% от общего содержания жирных кислот.
14. Семя сои по п.2, где содержание олеиновой кислоты составляет от около 46 до около 75 мас.% от общего содержания жирных кислот.
15. Семя сои по п.6, где содержание олеиновой кислоты составляет от около 50 до около 80 мас.% от общего содержания жирных кислот.
16. Семя сои по п.6, где содержание олеиновой кислоты составляет от около 46 до около 75 мас.% от общего содержания жирных кислот.
17. Семя сои по п.1, где указанная первая последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичная фрагменту интрона FAD2-1A сои, составляет от приблизительно 100 до приблизительно 320 нуклеотидов в длину, и указанная комбинация двух последовательностей нуклеиновой кислоты содержит фрагмент FATB-1 5' UTR.
18. Семя сои по п.1, где указанная первая последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичная фрагменту интрона FAD2-1A сои, выбрана из группы, состоящей из: от приблизительно 30 до приблизительно 420, от приблизительно 40 до приблизительно 320, от приблизительно 50 до приблизительно 200, от приблизительно 50 до приблизительно 400, от приблизительно 50 до приблизительно 420, от приблизительно 60 до приблизительно 320, от приблизительно 70 до приблизительно 220, от приблизительно 100 до приблизительно 200, от приблизительно 150 до приблизительно 200, от приблизительно 150 до приблизительно 220, от приблизительно 150 до приблизительно 400, от приблизительно 200 до приблизительно 300 и от приблизительно 300 до приблизительно 400 нуклеотидов в длину.
19. Семя сои по п.2, где указанная первая последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичная фрагменту интрона FAD2-1A сои, составляет от приблизительно 100 до приблизительно 320 нуклеотидов в длину.
20. Семя сои по п.2, где указанная первая последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичная фрагменту интрона FAD2-1A сои, выбрана из группы, состоящей из: от приблизительно 30 до приблизительно 420, от приблизительно 40 до приблизительно 320, от приблизительно 50 до приблизительно 200, от приблизительно 50 до приблизительно 400, от приблизительно 50 до приблизительно 420, от приблизительно 60 до приблизительно 320, от приблизительно 70 до приблизительно 220, от приблизительно 100 до приблизительно 200, от приблизительно 150 до приблизительно 200, от приблизительно 150 до приблизительно 220, от приблизительно 150 до приблизительно 400, от приблизительно 200 до приблизительно 300 и от приблизительно 300 до приблизительно 400 нуклеотидов в длину.
21. Семя сои по п.6, где указанная первая последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичная фрагменту интрона FAD2-1A сои, составляет от приблизительно 100 до приблизительно 320 нуклеотидов в длину.
22. Семя сои по п.6, где указанная первая последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентичная фрагменту интрона FAD2-1A сои, выбрана из группы, состоящей из: от приблизительно 30 до приблизительно 420, от приблизительно 40 до приблизительно 320, от приблизительно 50 до приблизительно 200, от приблизительно 50 до приблизительно 400, от приблизительно 50 до приблизительно 420, от приблизительно 60 до приблизительно 320, от приблизительно 70 до приблизительно 220, от приблизительно 100 до приблизительно 200, от приблизительно 150 до приблизительно 200, от приблизительно 150 до приблизительно 220, от приблизительно 150 до приблизительно 400, от приблизительно 200 до приблизительно 300 и от приблизительно 300 до приблизительно 400 нуклеотидов в длину.
23. Семя сои по п.1, где указанная первая последовательность нуклеиновой кислоты содержит фрагмент, на 100% идентичный фрагменту интрона FAD2-1A сои длиной между 20 и 420 последовательных нуклеотидов, и указанная комбинация двух последовательностей на 100% идентична FATB-1 сои и составляет от приблизительно 40 до приблизительно 450 нуклеотидов в длину, и указанный геном дополнительно содержит последовательность нуклеиновой кислоты, повышающую экспрессию бета-кетоацил-АСР-синтазы IV или дельта-9-десатуразы, либо обеих вместе.
24. Семя сои по п.2, где указанная первая последовательность нуклеиновой кислоты содержит фрагмент, на 100% идентичный фрагменту интрона FAD2-1A сои длиной между приблизительно 20 и приблизительно 420 нуклеотидов и последовательности FATB-1 сои длиной от приблизительно 40 до приблизительно 450 нуклеотидов в длину, где указанная последовательность FATB-1 содержит фрагмент UTR.
25. Семя сои по п.6, где указанная последовательность нуклеиновой кислоты, подавляющей экспрессию эндогенного FAD2-1 сои, содержит фрагмент на 100% идентичный фрагменту интрона FAD2-1A сои длиной между приблизительно 20 и приблизительно 420 нуклеотидов.
26. Семя сои по п.1, где первая последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентична фрагменту интрона FAD2-1А сои, а вторая последовательность нуклеиновой кислоты, по меньшей мере, на 95% идентична фрагменту гена FADB сои.
27. Семя сои по п.1, где указанный фрагмент гена FATB сои представляет собой кодирующую последовательность транзитного пептида FATB сои и приблизительно 45 последовательных нуклеотидов указанного FATB-1 5' UTR сои.
28. Семя сои по п.1, где указанный фрагмент гена FATB сои представляет собой кодирующую последовательность транзитного пептида FATB сои и приблизительно 42 последовательных нуклеотида указанного FATB-1 5' UTR сои.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |

