ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет на основании предварительной патентной заявки США №61/697886, поданной 7 сентября 2012 г., полное содержание которой, таким образом, включено в настоящий документ посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится, в целом, к композициям и способам для применения в технологии рекомбинантных растений (например, для получения трансгенного растения). Более конкретно, настоящее изобретение относится к растительным клеткам и растениям, содержащим локусы в их геномах, которые можно использовать для сайт-специфического введения любой интересующей нуклеиновой кислоты.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Многие растения генетически трансформируют экзогенными нуклеиновыми кислотами (например, трансгенами) для введения желательных признаков, например, для повышения сельскохозяйственной ценности. Примеры усовершенствований для повышения сельскохозяйственной ценности, которые могут быть достигнуты благодаря генетической трансформации, включают: улучшение питательных качеств, повышение урожайности, устойчивость к вредителям или болезням, устойчивость к засухе и стрессам, улучшение качества плодоовощной продукции (например, улучшение пигментации и/или роста), устойчивость к гербицидам, производство промышленно полезных соединений и/или материалов из растений, и/или производство фармацевтических продуктов. Введение клонированных генов в клетки растений и получение стабильных плодородных трансгенных растений можно использовать для создания генетической модификации растения, стабильной на протяжении нескольких поколений, и таким образом, становится возможной генетическая инженерия сельскохозяйственных культур.
В методах генетической трансформации и получения трансгенных растений экзогенную ДНК, как правило, случайным образом вводят в ядерную или пластидную ДНК эукариотической растительной клетки, с последующим выделением клеток, содержащих интегрированную экзогенную ДНК, и дальнейшей регенерацией стабильно трансформированного растения. Трансгенные растения, как правило, получали при помощи технологии опосредованной Agrobacterium трансформации. Успехи, достигнутые при использовании данных методов, стимулировали разработку других методов для введения интересующей молекулы нуклеиновой кислоты в геном растения, таких как опосредованное ПЭГ поглощение ДНК протопластами, бомбардировка микрочастицами и опосредованная силиконовыми нитями трансформация.
Однако во всех этих методах трансформации растений экзогенные нуклеиновые кислоты, вводимые в геном растений, интегрируются случайным образом в геном растительной клетки, и в непредсказуемом количестве копий. Terada et al. (2002) Nat Biotechnol 20(10): 1030, Terada et al. (2007) Plant Physiol 144(2): 846, D'Halluin et al. (2008) Plant Biotechnology J. 6(1): 93. Например, трансгены часто интегрируются в форме повторяющихся последовательностей, либо всего трансгена, либо его частей. Такая сложная схема интеграции обычно отрицательно влияет на уровень экспрессии интегрированной нуклеиновой кислоты (например, путем разрушения транскрибированной РНК за счет механизмов посттранскрипционного выключения генов или в результате индукции метилирования интегрированной ДНК). Кроме того, место расположения сайта интеграции обычно влияет на уровень экспрессии интегрированной нуклеиновой кислоты. Более того, интегрирование экзогенной ДНК может оказывать разрушительное воздействие на область генома, в которой происходит интеграция, и тем самым влиять или нарушать нормальное функционирование этой области-мишени, вызывая нежелательные побочные эффекты. Сочетание факторов, включая вышеизложенные, приводит к вариациям в широких пределах уровня экспрессии трансгена или экзогенный ДНК (и в целом, агрономического качества) среди различных трансгенных растительных клеток и линий растений, даже тех, которые получены одними и теми же методами. Поскольку интегрирование происходит случайным образом, специалист-практик не может контролировать эти эффекты, когда он или она пытается получить новое растение с желаемыми характеристиками.
С учетом вышеизложенных соображений, при исследовании эффектов от введения конкретной экзогенной нуклеиновой кислоты в растение необходимо создавать большое число линий трансгенных растений и анализировать их для получения значимых результатов. Аналогично, при создании трансгенного растения, содержащего конкретную интегрированную нуклеиновую кислоту для придания трансгенному растению желаемого фенотипа, необходимо получать большую популяцию независимо созданных линий трансгенных растений, чтобы иметь возможность отбора линии растения с оптимальной экспрессией нуклеиновой кислоты и с минимальным или отсутствующим побочным влиянием на общий фенотип и характеристики трансгенного растения. Эти практические соображения приобретают дополнительную важность в случае трансгенных растений, полученных путем включения нескольких экзогенных нуклеиновых кислот (то есть, пакетирования генов). В таких растениях может быть усилено такое явление, как посттранскрипционное выключение гена.
Разработано несколько способов в попытке контролировать вставки трансгенов в растения. Смотри, например, Kumar and Fladung (2001) Trends Plant Sci. 6: 155-9. Такие способы опираются на основанную на гомологичной рекомбинации интеграцию трансгенов, которая была успешно использована как в случае прокариотов, так и низших эукариотов. Paszkowski et al. (1988) EMBO J. 7: 4021-6. Однако до недавнего времени в растениях преобладающий механизм интеграции трансгенов имел в основе незаконную рекомбинацию, которая подразумевает небольшую гомологию между рекомбинирующими цепями ДНК. Таким образом, основной задачей в данной области является обнаружение и избирательное создание редких событий гомологичной рекомбинации, которые маскируются гораздо более эффективными интеграционными событиями за счет незаконной рекомбинации. Более того, даже в случае достижения избирательного создания и обнаружения событий направленной гомологичной рекомбинации, событие должно быть направлено в желаемый участок генома хозяина для того, чтобы извлечь максимальную выгоду из данной стратегии.
Например, предполагаемым преимуществом направленной генетической трансформации является снижение вариабельности экспрессии трансгена от события к событию, по сравнению с событиями трансформации, которые получают в результате случайного интегрирования. Другим предполагаемым преимуществом является значительное сокращение числа событий, необходимых для скрининга введенных нуклеиновых кислот, сортировки трансформационных конструктов и получения событий, которые вносят вклад в желаемые общие характеристики полученного трансгенного растения. Критическим фактором, необходимым для реализации этих преимуществ, является идентификация определенных мест в геноме, где функционирование трансгена будет соответствующим и, по возможности, где неблагоприятные эффекты на растение-хозяина будут устранены или сведены к минимуму.
В последнее время были описаны методы и композиции для направленного расщепления геномной ДНК. Такие направленные события расщепления можно использовать, например, для индукции направленного мутагенеза, индукции направленных делеций последовательностей клеточной ДНК и содействия направленной рекомбинации и интеграции в заданный хромосомный локус. Смотри, например, Urnov et al. (2010) Nature 435(7042): 646-51, патентные публикации США 20030232410, 20050208489, 20050026157, 20050064474, 20060188987, 20090263900, 20090117617, 20100047805, 20110207221, 20110301073, 2011089775, 20110239315, 20110145940 и международную публикацию WO 2007/014275, содержание которых включено в настоящий документ посредством ссылки в полном объеме и для всех целей. Расщепление может происходить за счет использования определенных нуклеаз, таких как сконструированные нуклеазы с «цинковыми пальцами» (ZFN), подобные активаторам транскрипции эффекторные нуклеазы (TALENs), или использования системы CRISPR/Cas с разработанной crRNA/tracr РНК («одиночной направляющей РНК») для направления специфического расщепления. В патентной публикации США №20080182332 описано использование неканонических нуклеаз с «цинковыми пальцами» (ZFNs) для направленной модификации геномов растений, в патентной публикации США №20090205083 описана опосредованная ZFN направленная модификация локуса EPSPS растения, в патентной публикации США №20100199389 описана направленная модификация локуса Zp15 растения и в патентной публикации США №20110167521 описана направленная модификация растительных генов, вовлеченных в биосинтез жирных кислот. Кроме того, в Moehle et al. (2007) Proc. Natl. Acad, Sci. USA 104(9): 3055-3060 описано использование разработанных ZFNs для направленного добавления гена в заданный локус. В патентной публикации США 20110041195 описаны способы получения гомозиготных диплоидных организмов.
Тем не менее, сохраняется потребность в композициях и способах для модифицирования и/или модулирования экспрессии генов FAD2 в растениях, включая создание растений с направленными вставками нужных трансгенов в локус FAD2.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композициям и способам для модулирования экспрессии генов FAD2 (например, в растениях, водорослях и грибах) и использованию данных локусов в качестве сайтов для направленного интегрирования интересующей последовательности нуклеиновой кислоты (например, экзогенной последовательности нуклеиновой кислоты) в клетку-хозяина. В некоторых вариантах осуществления клетка-хозяин может содержать один или более геномов с одной или более последовательностями FAD2 (например, гомологами или паралогами), любая или все из которых могут быть избирательно модифицированы и/или разрушены. В конкретных примерах настоящего изобретения описаны гены FAD2 2.3 и FAD2 2.6, а также соответствующие гомологи и паралоги в Glycine max (например, G. max культиваров Jack, Williams 82, X5, Westag и Maverick) и их использование в качестве локусов для направленного интегрирования интересующей последовательности нуклеиновой кислоты. Как описано в настоящем документе, хотя гены FAD2 вовлечены в биосинтез жирных кислот в организме хозяина, их модифицирование или разрушение (например, в результате интегрирования экзогенной нуклеиновой кислоты в кодирующую последовательность FAD2) неожиданно может не оказывать никакого или оказывать минимальное неблагоприятное воздействие на полученный организм-хозяина.
В настоящем документе также описано использование одного или более конкретных локусов FAD2 в тандеме с полипептидом, способным осуществлять расщепление и/или интегрирование определенных последовательностей нуклеиновой кислоты в локусах FAD2. Примеры использования локусов FAD2 в тандеме с полипептидом, способным осуществлять расщепление и/или интегрирование в локусах FAD2, включают полипептид, выбранный из группы, состоящей из белков с «цинковыми пальцами», мегануклеаз, доменов TAL, TALENs, РНК-направляемых CRISPR-Cas9, рекомбиназ, лейциновых «молний», CRISPr/Cas и других, известных специалистам в данной области. Конкретные примеры включают димерный («слитый») белок, содержащий полипептид сайт-специфического ДНК-связывающего домена и полипептид расщепляющего домена (например, нуклеазы), такой как белок ZFN, содержащий полипептид «цинковый палец» и полипептид нуклеазы FokI. Например, в настоящем документе описана демонстрация in vitro и in vivo эффективности и специфичности конкретных ZFNs, разработанных для связывания и индукции двухцепочечных разрывов в генах FAD2 2.3 и FAD2 2.6, и в их сочетаниях, без расщепления соответствующих гомологов или паралогов. В некоторых вариантах осуществления конкретные локусы FAD2 можно использовать с любым из вышеуказанных полипептидов для осуществления сайт-специфического интегрирования интересующей нуклеиновой кислоты, которая впоследствии экспрессируется в хозяине, причем с минимальным неблагоприятным воздействием на агрономические характеристики хозяина.
В некоторых аспектах в настоящем документе описаны полипептиды, содержащие ДНК-связывающий домен, который специфически связывается с геном FAD2. В некоторых вариантах осуществления такой полипептид может также содержать домен или полудомен нуклеазы (расщепляющий) (например, ZFN, рекомбиназы, транспозазы или хоминг-эндонуклеазы, включая хоминг-эндонуклеазу с модифицированным ДНК-связывающим доменом, домены TAL, TALENs, РНК-направляемые CRISPR-Cas9) и/или домен лигазы, так что полипептид может индуцировать направленный двухцепочечный разрыв и/или способствовать рекомбинации интересующей нуклеиновой кислоты в участке разрыва. В конкретных вариантах осуществления ДНК-связывающий домен, который направлен на локус FAD2, может представлять собой ДНК-расщепляющий функциональный домен. Вышеуказанные полипептиды можно использовать в некоторых вариантах осуществления для включения экзогенной нуклеиновой кислоты в геном организма-хозяина (например, из видов растений или животных) в одном или более локусах FAD2. В некоторых вариантах осуществления ДНК-связывающие домены содержат белок с «цинковыми пальцами», имеющий один или более «цинковых пальцев» (например, 2, 3, 4, 5, 6, 7, 8, 9 или более «цинковых пальцев»), сконструированный (не существующий в природе) для связывания с любой последовательностью в гене FAD2. Любой из белков с «цинковыми пальцами», описанных в настоящем документе, может связываться с сайтом-мишенью в кодирующей последовательности гена-мишени или в прилегающих последовательностях (например, промоторе или других элементах экспрессии). В некоторых вариантах осуществления белок с «цинковыми пальцами» связывается с сайтом-мишенью в гене FAD2, например, как показано в таблице 1. Спиральные области узнавания иллюстративных FAD2-связывающих «цинковых пальцев» приведены в таблице 2. Один или более из компонентов связывающих доменов типа «цинковые пальцы» белка с «цинковыми пальцами» может быть каноническим (C2H2) «цинковым пальцем» или неканоническим (например, C3H) «цинковым пальцем» (например, N-концевой и/или C-концевой «цинковый палец» может быть неканоническим «пальцем»).
В настоящем документе также описаны способы разрушения или корректирования гена FAD2. Кроме того, в настоящем документе описаны генетически модифицированные организмы-хозяева (например, трансгенные растения), полученные способами в соответствии с вариантами осуществления изобретения. В конкретных примерах трансгенный организм, полученный способом в соответствии с одним из вариантов осуществления изобретения, может представлять собой, без ограничения, водоросль, гриб, однодольное растение, двудольное растение и так далее. В некоторых конкретных вариантах осуществления двудольное растение может представлять собой растение сои (Glycine max).
Гены FAD2, описанные в настоящем документе, могут включать гены любого растения, водорослей или грибов, которые имеют один или более генов FAD2.
Вышеуказанные и другие признаки станут более очевидными из следующего далее подробного описания некоторых вариантов осуществления, в котором содержатся ссылки на сопроводительные фигуры.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 приведено выравнивание кодирующих последовательностей FAD2 2.3 из Williams 82 (SEQ ID NO: 4), Westag (SEQ ID NO: 5), X5 (SEQ ID NO: 6), Jack (SEQ ID NO: 7) и Maverick (SEQ ID NO: 8).
На фигуре 2 приведено выравнивание кодирующих последовательностей FAD2 2.6 из Williams 82 (SEQ ID NO: 9), Westag (SEQ ID NO: 10), X5 (SEQ ID NO: 11), Jack (SEQ ID NO: 12) и Maverick (SEQ ID NO: 13).
На фигуре 3 показана активность ZFNs, разработанных для генов FAD2 2.3 и 2.6, в анализе DLSSA. ZFNs, разработанные для локусов FAD2 2.3 и 2.6, оценивали на расщепляющую активность в отношении последовательностей FAD2 2.3 и 2.6, которые были клонированы в клетки млекопитающих в качестве репортеров.
На фигуре 4 приведена плазмидная карта pDAB115620.
На фигуре 5 приведена плазмидная карта pDAB115622.
На фигуре 6 приведена плазмидная карта pDAB7221.
На фигуре 7 схематично изображены зонды/праймеры для анализа разрушения локуса. Указаны сайты связывания F2 ZFN для генов FAD2 2.3 и 2.6 и праймеры, используемые для анализа разрушения локуса.
На фигуре 8 приведена последовательность продуктов «внутренней-внешней» (In-Out) ПЦР, полученных в результате направления на мишень с помощью NHEJ донорской последовательности при использовании нуклеазы с «цинковыми пальцами» F2 ZFN2 в локусе FAD2 2.3. Референсная последовательность (на фигуре сверху) представляет собой конфигурацию направленной вставки вектора донора в обратной ориентации. Одноцепочечные концы ДНК, возникающие в результате расщепления FokI, были заполнены для создания референсной последовательности. Приведены последовательности, секвенированные по методу Сэнгера. Последовательности связывания ZFN для F2 ZFN2 подчеркнуты. Плазмидные клоны с последовательностью, аналогичной указанной последовательности, перечислены справа.
ПОСЛЕДОВАТЕЛЬНОСТИ
Нуклеотидные последовательности изображены с использованием стандартных буквенных сокращений для нуклеотидных оснований, как определено в 37 C.F.R. § 1.822. Показана только одна цепь каждой нуклеотидной последовательности, однако следует иметь в виду, что комплементарная цепь включена при каждой ссылке на изображенную цепь.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. Обзор нескольких вариантов осуществления
Варианты осуществления изобретения относятся к подходу для направленного интегрирования экзогенных нуклеиновых кислот (например, трансгенов) в геном хозяина без существенного негативного влияния на другие характеристики фенотипа хозяина, помимо тех, которые затронуты интегрированием нуклеиновой кислоты. Некоторые варианты осуществления можно использовать для «пакетирования» нескольких нуклеиновых кислот в геноме одного хозяина. Такой подход требует разработки и применения четырех взаимосвязанных технологий: технологий направления на цель, позволяющих вносить двухцепочечные разрывы в определенные участки геномной ДНК (смотри, например, Puchta et al. (1993) Nucleic Acids Res. 21: 5034-40, Siebert and Puchta (2002) Plant Cell 14: 1121-31, D'Halluin et al. (2008) Plant Biotechnol. J. 6(1): 93-102, Cai et al. (2009) Plant Mol. Biol. 69(6): 699-709, Shukla et al. (2009) Nature 459(7245): 437-41), Shan et al. (2103) Nature Biotechnol. 31: 686-680, Le et al. (2013) Nature Biotechnol 31: 688-691, Nekrasov et al. (2013) Nature Biotechnol. 31: 691-693, Ainely et al. (2013) Plant Biotechnol. J. (онлайн 19 августа); технологий доставки, делающих возможной доставку оптимизированной экзогенной (донорской) нуклеиновой кислоты (Bibikova et al. (2003) Science 300(5620): 764); технологий интегрирования, включающих модификацию генов хозяина (с упором либо на гомологичную рекомбинацию, либо на пути NHEJ), с тем, чтобы увеличить частоту HDR или NHEJ для направленных вставок донорской ДНК; аналитических инструментов для улучшения и характеризации событий направленного интегрирования; и определенных нужных участков генома хозяина («функциональных локусов»), которые генетически хорошо изучены и которые поддерживают стабильную экспрессию гена из поколения в поколение без значительного ущерба для трансформированного организма хозяина. Смотри также патентные публикации США 20030232410, 20050208489, 20050026157, 20050064474, 20060188987, 20090263900, 20090117617, 20100047805, 20110207221, 20110301073, 2011089775, 20110239315, 20110145940, 20080182332, 20090205083, 20100199389, 20110167521. Например, в растениях функциональный локус представляет собой локус, в котором негативное влияние на агрономические или качественные характеристики трансгенного растения, в локус которого был введен трансген, является незначительным или отсутствует.
В вариантах осуществления, описанных в настоящем документе, использованы преимущества того неожиданного факта, что гены FAD2 растения являются функциональными локусами для направленной вставки экзогенных нуклеиновых кислот (например, гена(ов), некодирующих последовательностей ДНК, таких как сконструированные посадочные площадки (Engineered Landing Pads, ELPs) (патентная заявка США 12/011735) и сконструированные платформы для вставки трансгена (Engineered Transgene Insertion Platform, ETIP) (находящаяся на рассмотрении патентная заявка США №61/697882), а также трансформационной единицы(единиц) растения). Повсеместная распространенность локусов FAD2 в растениях и свидетельства того, что изменение или нокаут FAD2 в каноле, кукурузе, подсолнечнике, пшенице, хлопке и соевых бобах не несет отрицательных последствий для агрономических или качественных характеристик, делает локусы FAD2 широко используемым классом функциональных локусов в коммерчески используемых видах растений.
В некоторых вариантах осуществления используют сайт-специфическое расщепление двухцепочечной ДНК в локусе FAD2, например, в результате доставки и экспрессии специфического для сайта-мишени узнающего и расщепляющего ДНК белка. В конкретных примерах таким FAD2-специфичным узнающим и расщепляющим ДНК белком может быть, например, без ограничения, ZFN, TALEN, РНК-направляемая CRISPR-Cas9, рекомбиназа (например, рекомбиназы Cre, Hin, RecA, Tre и FLP), мегануклеаза и сконструированный белок, полученный из любого из вышеперечисленных или их эквивалентов. Расщепление может также осуществляться с использованием системы CRISPR/Cas с разработанной crRNA/tracr РНК («одиночной направляющей РНК») для направления специфического расщепления. В некоторых вариантах осуществления такой двухцепочечный разрыв может быть восстановлен путем интегрирования донорской нуклеиновой кислоты в сайт расщепления в функциональном локусе FAD2, например, путем управляемой гомологией репарации (Homology Directed Repair, HDR) или путем негомологичного соединения концов (Non-Homologous End Joining, NHEJ).
Данное изобретение иллюстрирует полезность локусов FAD2 в качестве функциональных локусов, например, путем описания локусов FAD2 2.3 и FAD2 2.6 в сое (Glycine max) и соответствующих FAD2-специфичных ZFNs, которые можно использовать для интегрирования экзогенной нуклеиновой кислоты в локус FAD2 2.3 и/или FAD2 2.6.
Варианты осуществления настоящего изобретения направлены на решение многих нерешенных проблем в данной области. Например, избирательность подхода направленного интегрирования, описанного в настоящем документе, может уменьшать или устранять необходимость повторных полевых испытаний, нужных для устранения нежелательных трансгенных событий, при том, что данные исследования являются дорогостоящими из-за задействованных ресурсов и обременительных нормативных требований в данной области. Кроме того, подходы направленной вставки ДНК, описанные в настоящем документе, могут быть особенно полезными в случае пакетирования трансгенов.
Хотя природную нуклеотидную последовательность в эндогенном локусе FAD2 можно использовать для прямого направления на цель интересующей нуклеиновой кислоты, в некоторых вариантах осуществления нуклеиновую кислоту можно сначала направлять на по меньшей мере один локус FAD2 в организме хозяина, так что интегрирование других интересующих молекул нуклеиновой кислоты в организм хозяина облегчается. В других примерах можно использовать нуклеотидные последовательности, не гомологичные природным последовательностям организма хозяина (например, образованные практически случайным образом последовательности нуклеиновой кислоты), которые фланкируют сайт узнавания ДНК (например, сайт узнавания для «цинковых пальцев»).
II. Термины
При использовании в данной заявке, включая формулу изобретения, термины в форме единственного числа включают форму их множественного числа, если из контекста явно не следует иное. Таким образом, например, ссылка на «растение» также относится и к множеству растений. Кроме того, в зависимости от контекста используемый термин «растение» может также относиться к генетически аналогичному или идентичному потомству данного растения. Аналогично, термин «нуклеиновая кислота» может относиться к множеству копий молекулы нуклеиновой кислоты. Подобным же образом, термин «зонд» может относиться ко многим аналогичным или идентичным молекулам зонда.
Числовые диапазоны включают числа, ограничивающие диапазон, и определенно включают каждое целое и дробное число в пределах заданного диапазона. Если не указано иное, все технические и научные термины, используемые в настоящем документе, имеют то же значение, которое им обычно придают специалисты в данной области.
Для облегчения обзора различных вариантов осуществления, описанных в данном документе, приводятся следующие объяснения специальных терминов:
Выделенный: «выделенный» биологический компонент (такой как нуклеиновая кислота или белок) был в значительной степени отделен, произведен отдельно от, или очищен от других биологических компонентов в клетке организма, в которой компонент естественным образом присутствует (то есть, другой хромосомной и внехромосомной ДНК и РНК, и белков), при осуществлении химического или функционального изменения компонента (например, нуклеиновую кислоту можно выделять из хромосомы, разрушая химические связи, соединяющие нуклеиновую кислоту с остальной ДНК в хромосоме). Молекулы нуклеиновых кислот и белки, которые были «выделены», включают молекулы нуклеиновых кислот и белки, очищенные стандартными методами очистки. Термин также охватывает нуклеиновые кислоты и белки, полученные методом рекомбинантной экспрессии в клетке-хозяине, а также химически синтезированные молекулы нуклеиновой кислоты, белки и пептиды.
Скрещивание: используемый в настоящем документе применительно к растениям термин «скрещивание» или «скрещенные» означает слияние гамет путем опыления для получения потомства (например, клеток, семян и растений). Этот термин включает как половое скрещивание (то есть, опыление одного растения другим), так и самооплодотворение (то есть, самоопыление, например, при использовании пыльцы и семяпочки от одного и того же растения).
Обратное скрещивание: методы обратного скрещивания можно использовать для введения последовательности нуклеиновой кислоты в растение. Этот метод широко используется в течение многих десятилетий для введения новых признаков в растения. Jensen, N., Ed. Plant Breeding Methodology, John Wiley & Sons, Inc., 1988. В типичном протоколе обратного скрещивания растение исходного интересующего сорта (рекуррентный родитель) скрещивают с растением второго сорта (нерекуррентный родитель), которое несет интересующую последовательность нуклеиновой кислоты, которую предстоит переносить. Полученное от этого скрещивания потомство затем снова скрещивают с рекуррентным родителем, и процесс повторяют до тех пор, пока не будет получено растение, в котором практически все желаемые морфологические и физиологические характеристики рекуррентного родителя будут восстановлены в преобразованном растении, в дополнение к перенесенной последовательности нуклеиновой кислоты от нерекуррентного родителя.
Интрогрессия: используемый в настоящем документе термин «интрогрессия» означает передачу аллеля (или модифицированного аллеля, содержащего экзогенную нуклеиновую кислоту) в генетический фон в определенном локусе. В некоторых вариантах осуществления интрогрессия специфического аллеля в локусе может происходить путем передачи аллеля по меньшей мере одному потомству через половое скрещивание двух родителей, относящихся к одному и тому же виду, при этом по меньшей мере один из родителей имеет специфическую аллельную форму в своем геноме. Потомство, содержащее специфический аллель, можно повторно обратно скрещивать с растениями линии, имеющей желаемый генетический фон. Потомство от обратного скрещивания можно отбирать на наличие специфической аллельной формы, с тем, чтобы создать новый сорт, в котором специфическая аллельная форма зафиксирована в генетическом фоне. В некоторых вариантах осуществления интрогрессия специфического аллеля может происходить путем рекомбинации между двумя донорскими геномами (например, в слитом протопласте), при этом по меньшей мере один из донорских геномов имеет специфическую аллельную форму в своем геноме. Интрогрессия может включать передачу специфической аллельной формы, которая может представлять собой, например, и без ограничения, разрушенный или модифицированный аллель, трансген, PTU и ELP.
Зародышевая плазма: используемый в настоящем документе термин «зародышевая плазма» означает генетический материал отдельного растения, группы растений (например, линии, сорта или семейства растений) и клона, полученного из растения или группы растений. Зародышевая плазма может быть частью организма или клетки, или она может быть отделена (например, выделена) от организма или клетки. Как правило, зародышевая плазма обеспечивает генетический материал определенной молекулярной моделью, которая является основой для наследственных качеств растения. Используемый в настоящем документе термин «зародышевая плазма» относится к клеткам конкретного растения, семенам, ткани конкретного растения (например, ткани, из которой могут быть выращены новые растения) и не семенным частям конкретного растения (например, листьям, стеблям, пыльце и клеткам). Используемый в настоящем документе термин «зародышевая плазма» является синонимом термина «генетический материал» и его можно использовать для обозначения семени (или другого растительного материала), из которого растение может быть размножено. Термин «банк зародышевой плазмы» может относиться к организованной коллекции различных семян или другого генетического материала (где каждый генотип однозначно идентифицирован), из которого известный культивар может быть культивирован и из которого новый культивар может быть создан.
Ген: используемый в настоящем документе термин «ген» (или «генетический элемент») может означать последовательность наследуемой геномной ДНК, имеющую функциональное значение. Ген может представлять собой природную нуклеиновую кислоту или нуклеиновую кислоту, которая была интегрирована в геном. Термин «ген» можно также использовать для обозначения, например, и без ограничения, кДНК и/или мРНК, кодируемую последовательностью наследуемой геномной ДНК.
Молекула нуклеиновой кислоты: используемый в настоящем документе термин «молекула нуклеиновой кислоты» может означать полимерную форму нуклеотидов (то есть, рибонуклеотидов, дезоксирибонуклеотидов и/или модифицированной формы любых из перечисленных выше). Используемый в настоящем документе термин «молекула нуклеиновой кислоты» является синонимом терминов «нуклеиновая кислота» и «полинуклеотид». Термин включает как смысловые, так и антисмысловые цепи РНК, кДНК, геномной ДНК, а также их синтетические формы и смешанные полимеры. Термин включает любую топологическую конформацию, в том числе одноцепочечную, двухцепочечную, частично дуплексную, триплексную, шпилечную, кольцевую и запертую конформации. Молекула нуклеиновой кислоты может включать или природные или модифицированные нуклеотиды, либо и те и другие. Такие нуклеотиды могут быть связаны друг с другом естественными и/или искусственными нуклеотидными связями.
Молекулы нуклеиновой кислоты могут быть модифицированы химическими или биохимическими методами, или могут содержать дериватизированные нуклеотидные основания, как легко понятно специалистам в данной области. Такие модификации включают, например, и без ограничения: метки, метилирование, замену одного или более из природных нуклеотидов аналогом и межнуклеотидные модификации (например, незаряженные связи, например, метилфосфонаты, фосфотриэфиры, фосфорамидаты и карбаматы, заряженные связи, например, фосфоротиоаты и фосфородитиоаты), подвешенные фрагменты, например, пептиды, интеркаляторы, например, акридин и псорален, хелаторы, алкиляторы, и модифицированные связи, например, альфа аномерные нуклеиновые кислоты).
Экзогенные: «экзогенная» молекула представляет собой молекулу, которая не является родной для определенной системы (например, зародышевой плазмы, сорта, элитного сорта и/или растения), применительно к нуклеотидной последовательности и/или местоположению в геноме (то есть, локусу) в случае полинуклеотида (и применительно к аминокислотной последовательности и/или клеточной локализации в случае полипептида). В вариантах осуществления экзогенные или гетерологичные полинуклеотиды или полипептиды могут представлять собой молекулы, которые были искусственно введены в биологическую систему (например, клетку растения, ген растения, конкретный вид или сорт растения и/или хромосому растения) и не являются родными для данной конкретной биологической системы. Таким образом, обозначение нуклеиновой кислоты как «экзогенной» может указывать на то, что нуклеиновая кислота происходит из иного источника, нежели природный источник, или это может указывать на то, что нуклеиновая кислота имеет неестественную конфигурацию, генетическое местоположение или расположение элементов.
Напротив, например, «родная» или «эндогенная» нуклеиновая кислота представляет собой нуклеиновую кислоту (например, ген), которая не содержит иной нуклеотидный элемент, нежели те, которые обычно присутствуют в хромосоме или другом генетическом материале, в котором нуклеиновая кислота обычно находится в природе. Эндогенный генный транскрипт кодируется нуклеотидной последовательностью в ее естественном хромосомном локусе, а не введен искусственно в клетку.
Функционально связанные: первая последовательность нуклеиновой кислоты является функционально связанной со второй последовательностью нуклеиновой кислоты, когда первая последовательность нуклеиновой кислоты находится в функциональном взаимоотношении со второй последовательностью нуклеиновой кислоты. Например, промотор является функционально связанным с кодирующей последовательностью, когда промотор влияет на транскрипцию или экспрессию кодирующей последовательности. При получении рекомбинантными методами функционально связанные последовательности нуклеиновой кислоты, как правило, являются смежными и, если необходимо соединить две кодирующие белок области, в одной и той же рамке считывания. Однако элементы не обязательно должны быть смежными, чтобы быть функционально связанными.
Промотор: промотор представляет собой область ДНК, которая, как правило, расположена выше (ближе к 5'-области) в нуклеиновой кислоте и которая стимулирует транскрипцию нуклеиновой кислоты. Промоторы делают возможной надлежащую активацию или репрессию нуклеиновой кислоты(кислот), с которой они функционально связаны. Промотор содержит определенные последовательности, которые узнаются факторами транскрипции. Эти факторы связываются с промоторными последовательностями ДНК, что приводит к рекрутированию РНК-полимеразы, фермента, который синтезирует РНК с кодирующей области нуклеиновой кислоты.
Трансформированные: вектор «трансформирует» или «трансдуцирует» клетку, когда он переносит молекулы нуклеиновой кислоты в клетку. Клетка является «трансформированной» молекулой нуклеиновой кислоты, когда молекула нуклеиновой кислоты становится стабильно реплицируемой в клетке, либо за счет встраивания молекулы нуклеиновой кислоты в клеточный геном, либо за счет эписомной репликации. Используемый в настоящем документе термин «трансформация» охватывает все методы, с помощью которых молекула нуклеиновой кислоты может быть введена в клетку. Примеры включают, но не ограничиваются ими: трансфекцию вирусными векторами, трансформацию плазмидными векторами, электропорацию (Fromm et al. (1986) Nature 319: 791-3), липофекцию (Felgner et al. (1987) Proc. Natl. Acad. Sci. USA 84: 7413-7), микроинъекцию (Mueller et al. (1978) Cell 15: 579-85), опосредованный Agrobacterium перенос (Fraley et al. (1983) Proc. Natl. Acad. Sci. USA 80: 4803-7), прямое поглощение ДНК и бомбардировку микрочастицами (Klein et al. (1987) Nature 327: 70).
Введенные: используемый в настоящем документе термин «введенная», когда речь идет о перемещении экзогенной нуклеиновой кислоты в клетку, означает включение нуклеиновой кислоты в клетку с использованием любой методологии, доступной в данной области. Этот термин охватывает методы введения нуклеиновой кислоты, включая, например, и без ограничения, трансфекцию, трансформацию и трансдукцию.
Трансген: используемый в настоящем документе термин «трансген» означает экзогенную нуклеиновую кислоту, кодирующую интересующую последовательность. Например, трансген может кодировать полезное для применения в промышленности или фармацевтике соединение, или продукт экспрессии, который способствует приобретению желаемого сельскохозяйственного признака (например, устойчивости к гербицидам или устойчивости к вредителям). В другом примере трансген может представлять собой антисмысловую нуклеиновую кислоту, при этом экспрессия антисмысловой нуклеиновой кислоты ингибирует экспрессию нуклеотидной последовательности-мишени. Трансген может содержать регуляторные последовательности, функционально связанные с трансгеном (например, промотор). В некоторых вариантах осуществления интересующая молекула нуклеиновой кислоты, предназначенная для введения путем сайт-специфического направления в локус FAD2, представляет собой трансген. Однако в других вариантах осуществления интересующая молекула нуклеиновой кислоты может представлять собой PTU, ELP, ETIP или эндогенную нуклеотидную последовательность (например, если нужны дополнительные, экзогенные геномные копии эндогенной нуклеотидной последовательности).
Элементы могут также включать ДНК, кодирующую структурную РНК, такую как кшРНК. Такая РНК может модифицировать экзогенные или эндогенные гены, включая, но без ограничения, влияние на расположение или придание устойчивости к гербицидам.
Рекомбинантные: используемый в настоящем документе термин «рекомбинантный» относится к материалу (например, нуклеиновой кислоте, гену, полинуклеотиду и/или полипептиду), который был изменен в результате вмешательства человека. Например, расположение частей или элементов рекомбинантной молекулы может быть неестественным расположением и/или первичная последовательность рекомбинантной молекулы может быть изменена относительно ее природной последовательности, например, для оптимизации ее экспрессии и/или активности. Материал можно изменять для получения рекомбинантного материала в пределах или за пределами его естественного окружения или состояния. В качестве одного из примеров, открытая рамка считывания нуклеиновой кислоты является рекомбинантной, если нуклеотидная последовательность открытой рамки считывания была удалена из ее естественного окружения и клонирована в искусственную молекулу нуклеиновой кислоты (например, вектор). Протоколы и реагенты для получения рекомбинантных молекул (например, рекомбинантных нуклеиновых кислот) общеизвестны в данной области, и их использование является обычным. Термин «рекомбинантные» в настоящем документе также может относиться к клетке или организму, которые содержат рекомбинантный материал (например, растение и/или клетка растения, которые содержат рекомбинантную нуклеиновую кислоту). В некоторых примерах рекомбинантный организм является трансгенным организмом.
Вектор: используемый в настоящем документе термин «вектор» относится к полинуклеотиду или другой молекуле, которая способна переносить по меньшей мере один сегмент(ы) нуклеиновой кислоты в клетку. Вектор может необязательно содержать компоненты/элементы, способствующие сохранению вектора и/или делающие возможным его намеченное использование (например, последовательности, необходимые для репликации, гены, придающие устойчивость к лекарственному средству или антибиотику, сайт множественного клонирования и/или функционально связанные элементы промотора/энхансера, которые делают возможной экспрессию клонированного гена). Векторы могут быть получены, например, из плазмид, бактериофагов или вирусов растений или животных. «Клонирующий вектор», «челночный вектор» или «субклонирующий вектор», как правило, содержит функционально связанные элементы для облегчения этапов клонирования или субклонирования (например, сайт множественного клонирования, содержащий несколько сайтов для эндонуклеаз рестрикции).
Экспрессионный вектор: используемый в настоящем документе термин «экспрессионный вектор» относится к вектору, содержащему функционально связанные полинуклеотидные последовательности, которые могут способствовать экспрессии кодирующей последовательности в конкретном организме-хозяине. Например, бактериальный экспрессионный вектор может способствовать экспрессии кодирующей последовательности в бактерии. Аналогично, растительный экспрессионный вектор может способствовать экспрессии кодирующей последовательности в растительной клетке. Полинуклеотидные последовательности, которые способствуют экспрессии в прокариотах, могут включать, например, и без ограничения, промотор, оператор и сайт связывания рибосом. Эукариотические экспрессионные векторы (например, растительный экспрессионный вектор) могут содержать, например, промоторы, энхансеры, сигналы терминации и сигналы полиаденилирования (а также другие последовательности), которые, как правило, отличаются от тех, которые используют в прокариотических экспрессионных векторах.
Идентичность последовательностей: используемый в настоящем документе термин «идентичность последовательностей» или «идентичность» в контексте двух нуклеотидных или полипептидных последовательностей относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для максимального соответствия в определенном окне сравнения. Величину идентичности последовательностей можно определять путем сравнения двух оптимально выровненных последовательностей (например, нуклеотидных последовательностей и аминокислотных последовательностей) в окне сравнения, при этом часть последовательности в окне сравнения может содержать добавления или делеции (то есть, разрывы) по сравнению с референсной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания двух последовательностей. Идентичность последовательностей рассчитывают в процентах путем определения числа положений, в которых идентичные нуклеотиды или аминокислотные остатки находятся в обеих последовательностях, с получением числа совпадающих положений, деления числа совпадающих положений на общее число положений в окне сравнения и умножения полученного результата на 100 для получения процента идентичности последовательностей.
Методы выравнивания последовательностей для сравнения хорошо известны в данной области. Различные программы и алгоритмы выравнивания описаны, например, в: Smith and Waterman (1981) Adv. Appl. Math. 2: 482, Needleman and Wunsch (1970) J. Mol. Biol. 48: 443, Pearson and Lipman (1988) Proc. Natl. Acad. Sci. U.S.A. 85: 2444, Higgins and Sharp (1988) Gene 73: 237-44, Higgins and Sharp (1989) CABIOS 5: 151-3, Corpet et al. (1988) Nucleic Acids Res. 16: 10881-90, Huang et al. (1992) Comp. Appl. Biosci. 8: 155-65, Pearson et al. (1994) Methods Mol. Biol. 24: 307-31, Tatiana et al. (1999) FEMS Microbiol. Lett. 174: 247-50. Подробное рассмотрение методов выравнивания последовательностей и расчета гомологии можно найти в Altschul et al. (1990) J. Mol. Biol. 215: 403-10.
Для выравнивания последовательностей можно использовать базовый инструментарий поиска локальных блоков (Basic Local Alignment Search Tool, BLAST™, Altschul et al. (1990)) Национального центра биотехнологической информации (NCBI), и он доступен из нескольких источников, включая Национальный центр биотехнологической информации (Bethesda, MD), а также в интернете, для использования совместно с несколькими программами анализа последовательностей. Описание того, как определять идентичность последовательностей с использованием данной программы, доступно в интернете в разделе «помощь» для BLAST™. Для сравнения нуклеотидных последовательностей можно использовать функцию «Blast 2 последовательности» программы BLAST™ (Blastn) с параметрами по умолчанию. Нуклеотидные последовательности, обладающие большим сходством с референсными последовательностями, будут демонстрировать больший процент идентичности при оценке данным методом.
Используемый в настоящем документе термин «в значительной степени идентичные» может относиться к нуклеотидным последовательностям, которые идентичны более чем на 80%. Например, в значительной степени идентичная нуклеотидная последовательность может быть на по меньшей мере 85%, по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или по меньшей мере 99,5% идентична референсной последовательности.
Локус: используемый в настоящем документе термин «локус» означает местоположение в геноме, которое соответствует поддающейся измерению характеристике (например, признаку). В некоторых вариантах осуществления локус, представляющий особый интерес, является местоположением в геноме гена FAD2, где разрушение гена уменьшает или устраняет экспрессию мРНК, транскрибированной с гена дикого типа. Локус можно определять с помощью зонда, который гибридизуется с уникальной нуклеотидной последовательностью, содержащейся внутри локуса, либо при гибридизации по Саузерну, либо при ПЦР.
Маркер: используемый в настоящем документе термин «маркер» означает ген или нуклеотидную последовательность, которые можно использовать для идентификации растений, которые могут иметь конкретный аллель и/или проявлять конкретный признак или фенотип. Маркер можно описать как вариацию в конкретном геномном локусе. Генетический маркер может представлять собой короткую последовательность ДНК, такую как последовательность вокруг одной измененной пары оснований (однонуклеотидный полиморфизм, или «SNP»), или длинную последовательность, например, минисателлит/повтор простой последовательности («SSR»). «Аллель маркера» означает вариант маркера, присутствующий в конкретном растении. Используемый в настоящем документе термин «маркер» может относиться к клонированному сегменту хромосомной ДНК растения (например, сегменту, содержащему локус FAD2 или модифицированный и/или разрушенный локус FAD2) и может также или альтернативно относиться к молекуле ДНК, которая комплементарна клонированного сегменту хромосомной ДНК растения. Как известно специалистам в данной области, процесс получения дополнительной непрерывной нуклеотидной последовательности для включения в маркер можно повторять почти до бесконечности (ограничение накладывается только длиной хромосомы), тем самым идентифицируя дополнительные маркеры вдоль хромосомы. Любой и все из вышеописанных разновидностей маркеров можно использовать в некоторых вариантах осуществления настоящего изобретения.
В некоторых вариантах осуществления присутствие трансгена или маркера (который характеризуется последовательностью-«мишенью») в зародышевой плазме можно обнаруживать при помощи нуклеотидного зонда, например, олигонуклеотида. Зонд может представлять собой молекулу ДНК или молекулу РНК. Олигонуклеотидный зонд можно получать синтетическими методами или путем клонирования. Подходящие клонирующие векторы хорошо известны специалистам в данной области. РНК зонды можно синтезировать методами, известными в данной области, например, с использованием молекулы-матрицы ДНК.
Олигонуклеотидный зонд может быть меченым или немеченым. Множество разнообразных методов существует для мечения молекул нуклеиновой кислоты, включая, например, и без ограничения, радиоактивное мечение ник-трансляцией, случайное праймирование и наращивание цепи с помощью концевой дезокситрансферазы, при этом используемые нуклеотиды являются мечеными, например, радиоактивным изотопом 32P. Другие метки, которые можно использовать, включают, например, и без ограничения, флуорофоры, ферменты, субстраты ферментов, кофакторы ферментов и ингибиторы ферментов. Альтернативно, использование метки, создающей обнаруживаемый сигнал самостоятельно или в сочетании с другими реакционноспособными веществами, можно заменять лигандами, с которыми рецепторы связываются, при этом рецепторы являются мечеными (например, вышеуказанными метками) для создания обнаруживаемых сигналов либо самостоятельно, либо в сочетании с другими реагентами. Смотри, например, Leary et al. (1983) Proc. Natl. Acad. Sci. USA 80: 4045-9.
Зонд может быть точной копией трансгена или маркера, который предстоит обнаруживать. Зонд также может представлять собой молекулу нуклеиновой кислоты, содержащую или состоящую из нуклеотидной последовательности, которая в значительной степени идентична клонированному сегменту хромосомной ДНК, содержащему трансген или маркер, который предстоит обнаруживать. Кроме того, зонд может содержать дополнительные нуклеотидные последовательности, например, промоторы, сигналы транскрипции и/или последовательности вектора.
Зонд может содержать всю или часть нуклеотидной последовательности-мишени и дополнительную смежную нуклеотидную последовательность из генома. В настоящем документе это называется «смежным зондом». Дополнительная смежная нуклеотидная последовательность называется расположенной «выше» или «ниже» оригинальной мишени, в зависимости от того, находится ли смежная нуклеотидная последовательность из хромосомы с 5' или с 3'-стороны от оригинального маркера, в обычном понимании. Зонд может также содержать нуклеотидную последовательность, которая не является смежной для оригинальной мишени, такой зонд в настоящем документе называется «несмежным зондом». Последовательность несмежного зонда может быть расположена достаточно близко к последовательности оригинальной мишени на хромосоме, так что несмежный зонд является связанным с оригинальным маркером или трансгеном.
В некоторых вариантах осуществления зонд представляет собой молекулу нуклеиновой кислоты, которая является «специфически гибридизуемой» или «специфически комплементарной» с точной копией мишени, которую предстоит обнаруживать. «Специфически гибридизуемая» и «специфически комплементарная» являются терминами, которые указывают на достаточную степень комплементарности, так что между молекулой нуклеиновой кислоты и мишенью существует стабильное и специфическое связывание. Молекула нуклеиновой кислоты не обязательно должна быть на 100% комплементарной своей последовательности-мишени, чтобы быть специфически гибридизуемой. Молекула нуклеиновой кислоты является специфически гибридизуемой, когда существует достаточная степень комплементарности, чтобы избежать неспецифического связывания нуклеиновой кислоты с не являющимися мишенью последовательностями в условиях, когда желательно специфическое связывание, например, в строгих условиях гибридизации.
Условия гибридизации, соответствующие конкретной степени строгости, будут варьироваться в зависимости от характера выбранного метода гибридизации, а также состава и длины гибридизующихся нуклеотидных последовательностей. Как правило, температура гибридизации и ионная сила (особенно концентрация Na+ и/или Mg++) буфера гибридизации определяют строгость условий гибридизации, хотя на строгость также влияет количество промываний. Расчеты, касающиеся условий гибридизации, необходимые для достижения конкретных степеней строгости, известны специалистам в данной области и обсуждаются, например, в Sambrook et al. (ed.) Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989, chapters 9 and 11, и Hames and Higgins (eds.) Nucleic Acid Hybridization, IRL Press, Oxford, 1985. Более подробные инструкции и руководство по гибридизации нуклеиновых кислот можно найти, например, в Tijssen, «Overview of principles of hybridization and the strategy of nucleic acid probe assays», в Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, Part I, Chapter 2, Elsevier, NY, 1993, и Ausubel et al., Eds., Current Protocols in Molecular Biology, Chapter 2, Greene Publishing и Wiley-Interscience, NY, 1995.
Используемый в настоящем документе термин «строгие условия» охватывает условия, в которых гибридизация будет происходить только в случае наличия менее 25% несовпадений между гибридизующейся молекулой и ДНК-мишенью. «Строгие условия» включают дополнительные конкретные уровни строгости. Так, при использовании в настоящем документе условия «умеренной строгости» означают такие условия, в которых молекулы с более чем 25% несовпадений последовательностей не будут гибридизоваться, условия «средней строгости» являются такими, в которых молекулы с более чем 15% несовпадений не будут гибридизоваться, и условия «высокой строгости» являются такими, в которых последовательности с более чем 10% несовпадений не будут гибридизоваться. Условия «очень высокой строгости» являются такими, в которых последовательности с более чем 6% несовпадений не будут гибридизоваться.
В конкретных вариантах осуществления строгие условия соответствуют гибридизации при 65°C в 6x буфере SSC (солевой раствор-цитрат натрия), 5x растворе Денхардта, 0,5% SDS и 100 мкг фрагментированной ДНК спермы лосося, с последующими 15-30-минутными последовательными промываниями при 65°C в 2x буфере SSC и 0,5% SDS, затем в 1x буфере SSC и 0,5% SDS, и наконец, в 0,2x буфере SSC и 0,5% SDS.
(Не)равновесие сцепления: используемый в настоящем документе термин «равновесие сцепления» относится к ситуации, когда маркер и вторая нуклеиновая кислота (например, трансген, PTU и второй маркер) независимо сегрегируют, то есть, маркер и вторая нуклеиновая кислота распределяются случайным образом среди потомства. Нуклеиновые кислоты, которые демонстрируют равновесие сцепления, считаются несцепленными (лежат ли они или не лежат на одной и той же хромосоме). Используемый в настоящем документе термин «неравновесие сцепления» относится к ситуации, когда маркер и вторая нуклеиновая кислота сегрегируют неслучайным образом, то есть, нуклеиновые кислоты имеют частоту рекомбинации менее 50% (и таким образом, по определению, разделены менее чем 50 сМ на одной и той же группе сцепления). В некоторых примерах нуклеиновые кислоты, которые демонстрируют неравновесие сцепления, считаются сцепленными.
Сцепленные, тесно сцепленные и очень тесно сцепленные: описанное в настоящем документе сцепление между маркером и второй нуклеиновой кислотой (например, трансгеном, PTU и вторым маркером) может относиться к явлению, при котором нуклеиновые кислоты на хромосоме имеют ощутимую вероятность быть переданными совместно индивидуумам в следующем поколении. Таким образом, сцепление одного маркера со второй нуклеиновой кислотой можно количественно определять и/или выражать в виде частоты рекомбинации. Чем ближе две нуклеиновые кислоты находятся друг к другу, тем данная вероятность становится ближе к «1». Таким образом, термин «сцепленные» может относиться к одному или более генам или маркерам, которые передаются вместе со второй нуклеиновой кислотой с вероятностью, превышающей 0,5 (что ожидается при независимом распределении, когда маркеры/гены расположены на разных хромосомах). Если присутствие гена (например, трансгена) отражается на фенотипе индивидуума, можно сказать, что маркеры, которые сцеплены с данным геном, сцеплены с данным фенотипом. Таким образом, термин «сцепленные» может относиться к взаимосвязи между маркером и геном или между маркером и фенотипом.
Относительное генетическое расстояние (определяемое частотой кроссинговера и измеряемое в сантиморганах (cM)), как правило, пропорционально физическому расстоянию (измеряемому в парах оснований), которым два сцепленных маркера или гена разделены друг от друга на хромосоме. Один сантиморган определяют как расстояние между двумя генетическими маркерами, которые демонстрируют 1% частоту рекомбинации (то есть, событие кроссинговера происходит между двумя маркерами один раз в 100 клеточных делений). Как правило, чем ближе находится один маркер к другому маркеру или гену (измеряют ли расстояние между ними в терминах генетического расстояния или физического расстояния), тем более тесно они связаны. Поскольку хромосомное расстояние приблизительно пропорционально частоте событий рекомбинации между признаками, существует приблизительное физическое расстояние, которое коррелирует с частотой рекомбинации. Эта корреляция, как правило, известна или легко определяема у основных сельскохозяйственных культур (Helentjaris and Burr (eds.) (1989) Development and Application of Molecular Markers to Problems in Plant Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, Gresshoff (ed.) (1994) Plant Genome Analysis. CRC Press, Boca Raton, FL, Lander et al. (1987) Genomics 1: 174-81, Tanksley et al. (1988) «Molecular mapping of plant chromosomes», в Chromosome Structure and Function. Gustafson and Appels (eds.) Plenum Press, NY, pp. 157-73) и многих других организмов. Например, 1 сМ соответствует примерно 2,5-3,0 т.п.н. у дрожжей, примерно 140 т.п.н. у Arabidopsis, примерно 400 т.п.н. у подсолнечника и примерно 350 т.п.н. у Eucalyptus.
Термин «сцепленные» в настоящем документе может относиться к одной или более нуклеиновым кислотам, которые демонстрируют частоту рекомбинации менее 50% (то есть, менее 50 сМ). Например, «сцепленные» нуклеиновые кислоты могут рекомбинировать с частотой примерно 45% или менее, примерно 40% или менее, примерно 35% или менее, примерно 30% или менее, примерно 25% или менее, примерно 20% или менее, примерно 15% или менее и примерно 10% или менее. Физические расстояния между такими нуклеиновыми кислотами на одной и той же хромосоме (ожидается, что нуклеиновые кислоты на разных хромосомах находятся в равновесии сцепления), которые соответствуют вышеуказанным частотам рекомбинации, зависят от генома хозяина, и могут быть легко рассчитаны, как указано выше.
Используемый в настоящем документе термин «тесно сцепленные» может относиться к одной или более нуклеиновым кислотам, которые демонстрируют частоту рекомбинации примерно 20% или менее (то есть, примерно 20 сМ или менее). Например, «тесно сцепленные» нуклеиновые кислоты могут рекомбинировать с частотой 22% или менее, примерно 18% или менее, примерно 16% или менее, примерно 14% или менее, примерно 12% или менее, примерно 10% или менее, примерно 8% или менее, примерно 6% или менее, примерно 4% или менее и примерно 2% или менее.
Используемый в настоящем документе термин «очень тесно сцепленные» может относиться к одной или более нуклеиновым кислотам, которые демонстрируют частоту рекомбинации примерно 10% или менее (то есть, примерно 10 сМ или менее). Например, «очень тесно сцепленные» нуклеиновые кислоты могут рекомбинировать с частотой 11% или менее, примерно 9% или менее, примерно 8% или менее, примерно 7% или менее, примерно 6% или менее, примерно 5% или менее, примерно 4% или менее, примерно 3% или менее, примерно 2% или менее и примерно 1% или менее.
Чем ближе конкретная нуклеиновая кислота находится к гену, кодирующему полипептид, который вносит вклад в конкретный фенотип (при измерении расстояния либо как генетического, либо как физического расстояния), тем более тесно сцепленной является конкретная нуклеиновая кислота с фенотипом. В свете вышеизложенного, следует иметь в виду, что нуклеиновые кислоты, сцепленные с конкретным геном или фенотипом, включают те нуклеиновые кислоты, которые тесно сцеплены, и те нуклеиновые кислоты, которые очень тесно сцеплены, с геном или фенотипом. В некоторых вариантах осуществления чем ближе конкретная нуклеиновая кислота находится к локусу FAD2 (например, модифицированному или разрушенному локусу FAD2) при измерении расстояния либо как генетического, либо как физического расстояния, тем более тесно сцепленной является конкретная нуклеиновая кислота с любым признаком/фенотипом, придаваемым экзогенной нуклеиновой кислотой, интегрированной в локусе FAD2 (или с фенотипом FAD2 дикого типа в случае немодифицированного локуса). Таким образом, генетические маркеры, которые сцеплены, тесно сцеплены и/или очень тесно сцеплены с локусом FAD2, содержащим интегрированную экзогенную нуклеиновую кислоту, могут быть полезны в программе MAS для идентификации организмов (например, растений и сортов растений), содержащих интегрированную нуклеиновую кислоту, для идентификации организмов, обладающих фенотипом, придаваемым интегрированной нуклеиновой кислотой, и для передачи при размножении такой интегрированной нуклеиновой кислоты и/или фенотипа, придаваемого интегрированной нуклеиновой кислотой, в другие совместимые организмы.
Селекция с помощью маркеров: используемый в настоящем документе термин «селекция с помощью маркеров» может означать подход к селекции растений непосредственно в отношении одного или более признаков (например, полигенного признака). В современной практике селекционеры растений пытаются определить легко обнаруживаемые признаки, такие как окраска цветка, внешний вид семенной оболочки или варианты изозимов, которые сцеплены с желаемым агрономическим признаком. Затем селекционеры отслеживают агрономический признак в сегрегирующих, размножающихся популяциях, следя за сегрегацией легко обнаруживаемого признака. Однако существует очень немного таких отношений сцепления между интересующими признаками и легко обнаруживаемыми признаками, доступных для использования в селекции растений. В некоторых вариантах осуществления изобретения селекция с помощью маркеров включает идентификацию одного или более генетических маркеров (например, SNP, изозима и/или SSR маркеров), которые сцеплены с локусом FAD2, в который экзогенная нуклеиновая кислота, обуславливающая интересующий признак, была интегрирована, и отслеживание интересующего признака в сегрегирующей размножающейся популяции путем отслеживания сегрегации одного или более генетических маркеров. В некоторых примерах сегрегацию одного или более генетических маркеров можно определять с использованием зонда для одного или более генетических маркеров, анализируя генетический образец от растения-потомка на присутствие одного или более генетических маркеров. Селекция с помощью маркеров является быстрым и экономичным способом улучшения сортов растений.
Признак или фенотип: термины «признак» и «фенотип» в настоящем документе используют взаимозаменяемо. Для целей настоящего изобретения признаки, представляющие особый интерес, включают важные агрономические признаки, которые могут экспрессироваться, например, в культурном растении, и производство продуктов экспрессии трансгена в результате события направленного интегрирования. Термин «молекулярный фенотип» может означать фенотип, который можно обнаружить на уровне популяции из (одной или более) молекул. В некоторых примерах молекулярный фенотип можно обнаружить только на молекулярном уровне. Обнаруживаемыми молекулами фенотипа могут быть нуклеиновые кислоты (например, геномная ДНК или РНК), белки и/или метаболиты. Например, молекулярный фенотип может представлять собой профиль экспрессии одного или более генных продуктов (например, на определенной стадии развития растения или в ответ на состояние окружающей среды или стресс).
Локус количественного признака: признаки, которые постоянно меняются вследствие генетического (аддитивные, доминантные и эпистатические) и внешнего влияния, обычно называют «количественными признаками». Количественные признаки можно отличать от «качественных» или «дискретных» признаков на основании двух факторов: влияния окружающей среды на экспрессию гена, которое приводит к непрерывному распределению фенотипов, и комплексного паттерна сегрегации, возникающего вследствие мультигенного наследования. При идентификации одной или более областей генома, связанных с экспрессией количественного признака, такие области определяют как локусы количественных признаков (Quantitative Trait Loci, «QTL»).
Растение: используемый в настоящем документе термин «растение» может означать целое растение, клеточную или тканевую культуру, полученную из растения, и/или любую часть любого из вышеперечисленного. Таким образом, термин «растение» охватывает, например, и без ограничения, целые растения, компоненты и/или органы растения (например, листья, стебли и корни), ткань растения, семя и клетку растения. Клетка растения может представлять собой, например, и без ограничения, клетку в растении и/или клетку растения, клетку, выделенную из растения, и клетку, полученную путем культивирования клетки, выделенной из растения.
«Трансгенное растение» представляет собой растение, содержащее, по меньшей мере в одной из его клеток, экзогенный полинуклеотид. Используемый в настоящем документе термин «трансгенные» относится к любой клетке, линии клеток, каллюсу, ткани, части растения или растению, генотип которых был изменен за счет присутствия экзогенной нуклеиновой кислоты. Таким образом, данный термин охватывает трансгенные организмы и клетки, которые были исходно изменены для содержания экзогенного полинуклеотида, и те организмы и клетки, которые были созданы в результате скрещивания или бесполого размножения исходного трансгенного организма или клетки. Используемый в настоящем документе термин «трансгенные» не относится к изменениям генома (хромосомным или внехромосомным), внесенным путем общепринятых методов селекции растений (например, скрещиваний только не трансгенных организмов) или путем естественных событий (например, случайного перекрестного опыления, не рекомбинантной вирусной инфекции, не рекомбинантной бактериальной трансформации, не рекомбинантной транспозиции и спонтанной мутации).
«Линия», «сорт» или «штамм» растения представляет собой группу отдельных растений, имеющих одинаковое происхождение. Растения линии, как правило, являются инбредными до некоторой степени и, как правило, являются гомозиготными и гомогенными в большинстве генетических локусов (например, локусе FAD2). Термин «сублиния» может означать инбредное подмножество потомков от общего предка, которые генетически отличаются от других аналогичных инбредных подмножеств, происходящих от того же предка. В некоторых вариантах осуществления «сублинию» можно получать путем инбридинга семян от отдельного трансгенного растения, отобранного в поколении F3-F5, до тех пор, пока остаточные сегрегирующие локусы не станут гомозиготными в большинстве своем или все.
«Связывающий белок» представляет собой белок, который способен связываться с другой молекулой. Связывающий белок способен связываться, например, с молекулой ДНК (ДНК-связывающий белок), молекулой РНК (РНК-связывающий белок) и/или с молекулой белка (белок-связывающий белок). В случае белок-связывающего белка, он может связываться с самим собой (образуя гомодимеры, гомотримеры и так далее) и/или он может связываться с одной или более молекулами другого белка или белков. Связывающий белок может иметь активность связывания более одного типа. Например, белки с «цинковыми пальцами» имеют ДНК-связывающую, РНК-связывающую и белок-связывающую активность.
«ДНК-связывающий белок с «цинковыми пальцами»» (или его связывающий домен) представляет собой белок или домен в составе более крупного белка, который связывает ДНК специфичным для последовательности образом через один или более «цинковых пальцев», которые представляют собой области аминокислотной последовательности в связывающем домене, структура которых стабилизирована за счет координации ионов цинка. ДНК-связывающий белок с «цинковыми пальцами» часто сокращенно называют белком с «цинковыми пальцами» или ZFP.
«ДНК-связывающий домен TALE» или «TALE» представляет собой полипептид, содержащий один или более повторяющихся доменов/единиц TALE. Повторяющиеся домены вовлечены в связывание TALE с его узнаваемой последовательностью-мишенью ДНК. Одна «повторяющаяся единица» (также называемая «повтором») имеет, как правило, 33-35 аминокислот в длину и демонстрирует по меньшей мере некоторую гомологию последовательностей с другими повторяющимися последовательностями TALE в природном белке TALE.
Домены «цинковые пальцы» и связывающие домены TALE можно «конструировать» для связывания с заранее определенной нуклеотидной последовательностью, например, путем конструирования (изменения одной или более аминокислот) спиральной области узнавания природного белка с «цинковыми пальцами» или TALE. Таким образом, сконструированные ДНК-связывающие белки (с «цинковыми пальцами» или TALEs) являются белками, не существующими в природе. Неограничивающие примеры методов конструирования ДНК-связывающих белков включают проектирование и отбор. Спроектированный ДНК-связывающий белок представляет собой белок, не существующий в природе, конструкция/состав которого является результатом использования преимущественно рациональных критериев. Рациональные критерии для проектирования включают применение правил замещения и компьютеризированных алгоритмов для обработки информации в базе данных, хранящей информацию о существующих данных, относящихся к конструкциям и связыванию ZFP и/или TALE. Смотри, например, патенты США 6140081, 6453242 и 6534261, смотри также WO 98/53058, WO 98/53059, WO 98/53060, WO 02/016536 и WO 03/016496, а также патентную публикацию США №20110301073.
«Отобранный» белок с «цинковыми пальцами» или TALE представляет собой белок, не встречающийся в природе, полученный, главным образом, эмпирическим методом, таким как фаговый дисплей, ловушка взаимодействия или отбор гибридов. Смотри, например, US 5789538, US 5925523, US 6007988, US 6013453, US 6200759, WO 95/19431, WO 96/06166, WO 98/53057, WO 98/54311, WO 00/27878, WO 01/60970, WO 01/88197, WO 02/099084 и патентную публикацию США №20110301073.
«Расщепление» означает разрыв связанного ковалентными связями каркаса молекулы ДНК. Расщепление можно инициировать различными методами, включая, но не ограничиваясь ими, ферментативный или химический гидролиз фосфодиэфирной связи. Возможно как одноцепочечное расщепление, так и двухцепочечное расщепление, и двухцепочечное расщепление может происходить в результате двух отдельных событий одноцепочечного расщепления. Расщепление ДНК может приводить к образованию либо тупых концов, либо ступенчатых концов. В конкретных вариантах осуществления для направленного расщепления двухцепочечной ДНК используют слитые полипептиды.
«Расщепляющий полудомен» представляет собой полипептидную последовательность, которая в сочетании со вторым полипептидом (либо идентичным, либо отличающимся) образует комплекс, обладающий способностью к расщеплению (предпочтительно способностью к двухцепочечному расщеплению). Термины «первый и второй расщепляющие полудомены» «+ и - расщепляющие полудомены» и «правый и левый расщепляющие полудомены» используются взаимозаменяемо для обозначения пар расщепляющих полудоменов, которые димеризуются.
«Сконструированный расщепляющий полудомен» представляет собой расщепляющий полудомен, который был модифицирован для образования облигатных гетеродимеров с другим расщепляющим полудоменом (например, другим сконструированным расщепляющим полудоменом). Смотри, также, патентные публикации США №№2005/0064474, 20070218528, 2008/0131962 и 2011/0201055, полное содержание которых включено в настоящий документ посредством ссылки.
Средства для создания двухцепочечных разрывов ДНК: используемый в настоящем документе термин «средства для создания двухцепочечных разрывов ДНК» подразумевает ссылку на специальные утверждающие положения, одобренные Конгрессом в 35 U.S.C. § 112, шестой параграф. В частности, термин «средства для создания двухцепочечных разрывов ДНК» относится к молекулярной структуре, которая способна расщеплять обе цепи двухцепочечной молекулы ДНК. Такие структуры включают полипептидные домены, содержащиеся во многих известных белках нуклеазах, например, домен нуклеазы FokI, каталитический домен, выбранный из группы, состоящей из белков MmeI, колицина-E7 (CEA7_ECOLX), колицина-E9, APFL, EndA, Endo I (END1_EC0LI), Endo G человека (NUCG_HUMAN), Endo G быка (NUCG_BOVIN), R.HinPII, I-BasI, I-BmoI, I-HmuI, I-TevI, I-TevII, I-TevIII, I-TwoI, R.MspI, R.MvaI, NucA, NucM, Vvn, Vvn_CLS, стафилококковой нуклеазы (NUC_STAAU), стафилококковой нуклеазы (NUC_STAHY), микрококковой нуклеазы (NUC_SHIFL), эндонуклеазы yncB, эндодезоксирибонуклеазы I (ENRN_BPT7), метназы, Nb.BsrDI, BsrDI A, Nt.BspD6I (большой субъединицы R.BspD6I), ss.BspD6I (малой субъединицы R.BspD6I), R.PIeI, MlyI, AlwI, MvaI269I, BsrI, BsmI, Nb.BtsCI, Nt.BtsCI, R1.BtsI, R2.BtsI, субъединицы 1 BbvCI, субъединицы 2 BbvCI, альфа-субъединицы Bpu10I, бета-субъединицы Bpu10I, BmrI, BfiI, I-CreI, hExoI (EX01JHUMAN), ExoI дрожжей (EXO1_YEAST), ExoI E.coli, TREX2 человека, TREX1 мыши, TREX1 человека, TREX1 быка, TREX1 крысы, DNA2 человека, DNA2 дрожжей(DNA2_YEAST).
Средства для репарации двухцепочечных разрывов ДНК: используемый в настоящем документе термин «средства для репарации двухцепочечных разрывов ДНК» также подразумевает ссылку на специальные утверждающие положения, одобренные Конгрессом в 35 U.S.C. § 112, шестой параграф. В частности, термин «средства для репарации двухцепочечных разрывов ДНК» относится к молекулярной структуре, которая способна облегчать/катализировать соединение концов двухцепочечных молекул ДНК, например, соединяя концы, образованные в результате расщепления одной двухцепочечной молекулы ДНК, или соединяя один конец, образованный в результате расщепления одной двухцепочечной молекулы ДНК, с концом экзогенной двухцепочечной молекулы ДНК. Такие структуры включают полипептидные домены, содержащиеся во многих известных белках лигазах, например, рекомбиназе Cre. В некоторых примерах одна и та же молекулярная структура может служить как средством создания двухцепочечных разрывов ДНК, так и средством репарации двухцепочечных разрывов ДНК, при этом одна и та же структура способствует как расщеплению, так и репарации двухцепочечной молекулы ДНК (например, рекомбиназа Hin).
Индукция сайт-специфических двухцепочечных разрывов в геноме индуцирует путь репарации ДНК в растительной клетке-хозяине, который устраняет двухцепочечные разрывы путем управляемой гомологией репарации (HDR) или путем негомологичного соединения концов (NHEJ). В научной литературе сообщается, что в растениях точное интегрирование гена или донорской ДНК в природные геномные или в предварительно сконструированные участки включает использование вводимого донорского конструкта(ов) ДНК, содержащего различные количества последовательности, гомологичной последовательностям, фланкирующим направленный двухцепочечный разрыв. Интегрирование таких доноров в определенный локус-мишень предположительно осуществляется по пути HDR. Исключительный расчет на подход HDR для генного таргетинга в растениях может иметь ограничения в связи с сообщениями, что путь репарации HDR не является доминантным путем репарации ДНК в сравнении с NHEJ. В опубликованной научной литературе, описывающей использование в растениях специфичных для мишени разрывов ДНК (при помощи ZFN, TALeNs или сконструированных мегануклеаз и так далее), путь NHEJ был описан в качестве способа внесения специфических точечных мутаций (вставок или делеций) в геном. В настоящем документе авторы изобретения сообщают, что при сайт-специфических двухцепочечных разрывах (индуцированных ZFN, TALeNs и так далее) в присутствии различных донорских конструкций ДНК с областями гомологии, составляющими от 0 до <10 п.н., может происходить специфическая вставка в направленный разрыв посредством репарационного пути NHEJ в растениях. Множество различных донорских конструкций ДНК с гомологией от нулевой до небольшой (1-10 п.н.) в диапазонах от линейной до кольцевой, от одноцепочечной до двухцепочечной можно направлять на специфические участки при помощи пути NHEJ. Основанное на NHEJ направление донорской ДНК в растительный геном может базироваться на «захвате липких концов», когда направленный двухцепочечный разрыв в геноме образован при помощи FokI (или других эндонуклеазных доменов II типа) и соответствующие липкие концы находятся на подходящих для NHEJ донорских конструкциях ДНК. Донорскую ДНК с липкими концами можно доставлять непосредственно в клетку в виде линейной донорской ДНК с заранее определенными выступающими фрагментами. Альтернативный подход заключается в образовании липких концов донорской ДНК in vivo путем совместной доставки направленной на геном хозяина ZFN и кольцевой донорской молекулы ДНК, содержащей по меньшей мере один сайт узнавания ZFN, который идентичен сайту узнавания мишени. Экспрессия по меньшей мере одной ZFN приводит к разрывам в геномной ДНК хозяина (природной или предварительно сконструированной) и кольцевой донорской ДНК, с образованием липких концов, которые соединяются при помощи репарационного пути NHEJ хозяина.
Можно иметь один или более сайтов разрезания ZFN на донорской молекуле (один сайт разрезания ZFN для линеаризации целой донорской молекулы, 2 одинаковых сайта ZFN для высвобождения меньшего по размеру донорского фрагмента ДНК или 2 разных сайта ZFN для высвобождения фрагмента из донора и соответствующего фрагмента из геномной ДНК хозяина (замена ДНК).
Таким образом, донорский полинуклеотид может представлять собой ДНК или РНК, одноцепочечную и/или двухцепочечную и может быть введен в клетку в линейной или кольцевой форме. Смотри, например, патентные публикации США №№20100047805 и 20110207221. Некоторые варианты осуществления настоящего изобретения могут также включать линейную экзогенную (донорскую) нуклеиновую кислоту(ты), композиции, содержащие такие нуклеиновые кислоты, и способы получения и использования таких линейных донорских молекул. В некоторых вариантах осуществления линейная донорская молекула стабильно сохраняется в клетке, в которую она введена. В других вариантах осуществления линейная донорская молекула модифицирована, чтобы противостоять экзонуклеолитическому расщеплению, например, путем помещения одной или более фосфоротиоатных фосфодиэфирных связей между одной или более парами оснований на концах донорской молекулы. Линейная экзогенная нуклеиновая кислота может также включать одноцепочечную специфическую ДНК.
III. Функциональные локусы FAD2
Локусы, обозначенные FAD2 (десатураза жирных кислот 2), включены в QTLs, вовлеченные в наследование комплексного мультигенного признака содержания жирных кислот в растениях. FAD2 кодирует фермент, отвечающий за десатурацию олеиновой кислоты (18:1) до линолевой кислоты (C18:2). Tanhuanpaa et al. (1998) Mol. Breed. 4: 543-50, Schierholt et al. (2001) Crop Sci. 41: 1444-9.
В пути биосинтеза масел в растениях десатуразы жирных кислот (FADs) играют ключевую роль в биосинтезе растительных липидов, и их активность в значительной степени влияет на состав жирных кислот. FADs в изобилии присутствуют в растениях, и анализ экспрессии свидетельствует о том, что мРНК FAD продуцируются в большом избытке. Кроме того, гены FAD экспрессируются в тканях и клетках различных типов, а также субклеточных компартментах, включая пластиды и эндоплазматический ретикулум.
Состав жирных кислот растений и характеристики масел, полученных из них во многих применениях, определяются относительными концентрациями основных компонентов жирных кислот: олеиновой, линолевой и линоленовой (C18:3). Концентрации этих жирных кислот преимущественно регулируются функционированием ферментов FAD2 и FAD3. Олеиновая кислота в растениях преобразуется в линолевую кислоту и линоленовую кислоту в соответствии со схемой:
Гены FAD2 были идентифицированы в основных видах растений и водорослей, включая, но не ограничиваясь ими, кукурузу, сою, хлопок, Arabidopsis, пшеницу, кормовые травы, рис, подсолнечник и Brassica, и изменение экспрессии FAD2 приводит к изменению профилей жирных кислот в этих организмах. Кроме того, растения, содержащие модифицированные гены FAD2, были введены в коммерческий оборот, и было показано, что разрушение гена FAD2 может приводить к улучшению питательных и функциональных свойств масла, производимого растением-хозяином, без ухудшения агрономических показателей растения-хозяина. Например, сорта канолы и подсолнечника, которые были введены в коммерческий оборот под торговой маркой Nexera® (Dow AgroSciences, LLC), характеризуются большим содержанием олеиновой кислоты, меньшим содержанием линолевой кислоты и меньшим содержанием линоленовой кислоты (и меньшим содержанием насыщенных жирных кислот) по сравнению с профилями канолы и подсолнечника дикого типа.
Как описано в Chi X., et al., ((2011) Genome-wide analysis of fatty acid desaturases in soybean (Glycine max), Plant Molecular Biology Reports 29, 769-783), включенной в настоящий документ посредством ссылки, известные функциональные генные копии FAD2 в сое были филогенетически отнесены к девяти подсемействам с аналогами в Arabidopsis: FAB2, FAD2, FAD3, FAD5, FAD6, FAD7, FAD8, SLD1 и DES1. Было обнаружено двадцать девять генов десатуразы, распределенных на по меньшей мере 15 из 20 хромосом сои. Структуры генов и составы мотивов в значительной степени консервативны среди подсемейств. На основании анализа данных микрочипов, большинство генов десатураз демонстрируют специфические временные и пространственные паттерны экспрессии в различных тканях и стадиях развития.
Локусы FAD2 можно модифицировать и/или разрушать в растении без отрицательного влияния на ценность растения, и во многих случаях фактически увеличивая его ценность, включая изменение экспрессии FAD2, изменение содержания/соотношения масел и/или интегрирование и экспрессию желаемых трансгенов. Кроме того, в связи с повсеместной распространенностью локусов FAD в растениях локусы FAD2 можно модифицировать и/или разрушать, по меньшей мере для некоторых целей, без нанесения ущерба во многих видах, включая, например, и без ограничения: канолу, сою, кукурузу, пшеницу, кормовые травы, Brassica sp., рис, томаты, ячмень, овес, сорго, хлопок и подсолнечник, а также грибы и водоросли. Варианты осуществления изобретения включают локусы FAD2 и их применение в качестве функциональных локусов для интегрирования экзогенных нуклеиновых кислот. В примерах локус FAD2 проявляет по меньшей мере одну из нескольких характерных особенностей, которые являются желательными в контексте его применения в качестве функционального локуса, включая, например, и без ограничения: то, что имеет место примерно постоянный уровень экспрессии на протяжении жизненного цикла организма-хозяина, и, что удивительно, вставка донорской ДНК в локус FAD2 не вызывает проблем с качеством и хорошим физическим состоянием хозяина.
В некоторых вариантах осуществления настоящего изобретения по меньшей мере один локус FAD2 (например, локус FAD2 2.3 и локус FAD2 2.6) используют в качестве сайта-мишени для сайт-специфического интегрирования экзогенной нуклеиновой кислоты (например, нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую интересующий полипептид). В конкретных вариантах осуществления интегрирование экзогенной нуклеиновой кислоты приводит к модификации локуса. Например, интегрирование экзогенной нуклеиновой кислоты может модифицировать локус таким образом, что приводит к разрушению (то есть, инактивации) гена FAD2.
В некоторых вариантах осуществления локус FAD2 может содержать нуклеотидную последовательность, которая может специфически гибридизоваться с комплементом нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 14-20. Например, локус FAD2 может содержать нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 14-20. В некоторых вариантах осуществления локус FAD2 может содержать нуклеотидную последовательность, которая в значительной степени идентична нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 14-20. Например, в некоторых вариантах осуществления локус FAD2 является гомологом FAD2 (например, ортологом или паралогом), содержащим нуклеотидную последовательность, которая по меньшей мере примерно на 85% идентична нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 14-20. Гомолог FAD2 может содержать нуклеотидную последовательность, которая является, например, и без ограничения: по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98%, по меньшей мере примерно на 99%, по меньшей мере примерно на 99,5%, 99,6%, 99,7%, 99,8% и/или по меньшей мере примерно на 99,9% идентичной нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 14-20. Такой гомолог FAD2 можно легко идентифицировать и выделять из любого полного или частичного генома различных организмов, который легко доступен для специалистов в данной области.
IV. Направленное интегрирование нуклеиновой кислоты в локус FAD2
Сайт-специфическое интегрирование экзогенной нуклеиновой кислоты в локус FAD2 можно осуществлять любым методом, известным специалистам в данной области. В некоторых вариантах осуществления интегрирование экзогенной нуклеиновой кислоты в локус FAD2 включает создание контакта клетки (например, выделенной клетки или клетки в ткани или организме) с молекулой нуклеиновой кислоты, содержащей экзогенную нуклеиновую кислоту. В примерах такая молекула нуклеиновой кислоты может содержать нуклеотидные последовательности, фланкирующие экзогенную нуклеиновую кислоту, которые способствуют гомологичной рекомбинации между молекулой нуклеиновой кислоты и по меньшей мере одним локусом FAD2. В конкретных примерах нуклеотидные последовательности, фланкирующие экзогенную нуклеиновую кислоту, которые способствуют гомологичной рекомбинации, могут быть комплементарны эндогенным нуклеотидам локуса FAD2. В конкретных примерах нуклеотидные последовательности, фланкирующие экзогенную нуклеиновую кислоту, которые способствуют гомологичной рекомбинации, могут быть комплементарны ранее интегрированным экзогенным нуклеотидам. В некоторых вариантах осуществления несколько экзогенных нуклеиновых кислот могут быть интегрированы в один локус FAD2, например, как при пакетировании генов.
Интегрирование нуклеиновой кислоты в локус FAD2 может облегчаться (например, катализироваться) в некоторых вариантах осуществления эндогенным клеточным аппаратом клетки-хозяина, таким как, например, и без ограничения, эндогенная ДНК и эндогенные ферменты рекомбиназы. В некоторых вариантах осуществления интегрирование нуклеиновой кислоты в локус FAD2 может облегчаться за счет одного или более факторов (например, полипептидов), доставленных в клетку-хозяина. Например, полипептиды нуклеазы(нуклеаз), рекомбиназы(рекомбиназ) и/или лигазы могут быть доставлены (либо независимо, либо в виде части химерного полипептида) путем создания контакта полипептидов с клеткой-хозяином или путем экспрессии полипептидов в клетке-хозяине. Соответственно, в некоторых примерах нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую по меньшей мере один полипептид нуклеазы, рекомбиназы и/или лигазы, можно вводить в клетку-хозяина либо одновременно, либо последовательно с нуклеиновой кислотой, предназначенной для сайт-специфического интегрирования в локус FAD2, при этом по меньшей мере один полипептид нуклеазы, рекомбиназы и/или лигазы экспрессируется с нуклеотидной последовательности в клетке-хозяине.
A. ДНК-связывающие полипептиды
В некоторых вариантах осуществления сайт-специфическое интегрирование можно осуществлять, используя факторы, способные узнавать и связываться с конкретными нуклеотидными последовательностями, например, в геноме организма-хозяина. Например, многие белки содержат полипептидные домены, способные узнавать и связываться с ДНК сайт-специфическим образом. Последовательность ДНК, которая узнается ДНК-связывающим полипептидом, можно назвать последовательностью-«мишенью». Полипептидные домены, способные узнавать и связываться с ДНК сайт-специфическим образом, как правило, свернуты правильно и действуют независимо, связывая ДНК сайт-специфическим образом, даже будучи экспрессированы в составе полипептида, отличного от белка, из которого домен был исходно выделен. Аналогично, последовательности-мишени для узнавания и связывания ДНК-связывающими полипептидами, как правило, способны быть узнанными и связанными такими полипептидами, даже если они находятся в крупных структурах ДНК (например, хромосоме), особенно если сайт, в котором расположена последовательность-мишень, является сайтом, который, как известно, доступен для растворимых клеточных белков (например, ген).
Хотя ДНК-связывающие полипептиды, идентифицированные в белках, которые существуют в природе, как правило, связываются с дискретной нуклеотидной последовательностью или мотивом (например, консенсусной последовательностью узнавания), существуют методы, известные в данной области, для модификации многих таких ДНК-связывающих полипептидов, чтобы они узнавали другую нуклеотидную последовательность или мотив. ДНК-связывающие полипептиды включают, например, и без ограничения: ДНК-связывающие домены «цинковые пальцы», лейциновые «молнии», ДНК-связывающие домены UPA, GAL4, TAL, LexA, репрессор Tet, LacR и рецептор стероидных гормонов.
В некоторых примерах ДНК-связывающий полипептид представляет собой «цинковый палец». Отдельные мотивы «цинковых пальцев» можно сконструировать для направления на цель и специфического связывания с любым из широкого диапазона сайтов ДНК. Канонические Cys2His2 (а также неканонические Cys3His) полипептиды «цинковые пальцы» связывают ДНК за счет вставки α-спирали в большую бороздку двойной спирали ДНК-мишени. Узнавание ДНК «цинковыми пальцами» является модульным; каждый «палец» контактирует преимущественно с тремя последовательными парами оснований в мишени и несколько ключевых остатков в полипептиде опосредуют узнавание. Путем включения нескольких ДНК-связывающих доменов «цинковых пальцев» в направленную эндонуклеазу специфичность ДНК-связывания направленной эндонуклеазы можно дополнительно повышать (и таким образом, специфичность любых генных регуляторных эффектов, привнесенных за счет этого, также можно повышать). Смотри, например, Urnov et al. (2005) Nature 435: 646-51. Таким образом, один или более ДНК-связывающих полипептидов «цинковых пальцев» можно конструировать и использовать таким образом, чтобы направленная эндонуклеаза, введенная в клетку-хозяина, взаимодействовала с последовательностью ДНК, уникальной в геноме клетки-хозяина.
Предпочтительно, белок с «цинковыми пальцами» не является природным, он сконструирован для связывания с выбранным сайтом-мишенью. Смотри, например, Beerli et al. (2002) Nature Biotechnol. 20: 135-141, Pabo et al. (2001) Ann. Rev. Biochem. 70: 313-340, Isalan et al. (2001) Nature Biotechnol. 19: 656-660, Segal et al. (2001) Curr. Opin. Biotechnol. 12: 632-637, Choo et al. (2000) Curr. Opin. Struct. Biol. 10: 411-416, патенты США №№6453242, 6534261, 6599692, 6503717, 6689558, 7030215, 6794136, 7067317, 7262054, 7070934, 7361635, 7253273 и патентные публикации США №№2005/0064474, 2007/0218528, 2005/0267061, полное содержание которых включено в настоящий документ посредством ссылки.
Сконструированный связывающий домен «цинковый палец» может иметь новую специфичность связывания в сравнении с природным белком с «цинковыми пальцами». Способы конструирования включают, но не ограничиваются ими, рациональное проектирование и различные виды отбора. Рациональное проектирование включает, например, использование баз данных, содержащих триплетные (или квадруплетные) нуклеотидные последовательности и отдельные аминокислотные последовательности «цинковых пальцев», при этом каждая триплетная или квадруплетная нуклеотидная последовательность связана с одной или более аминокислотными последовательностями «цинковых пальцев», которые связывают конкретную триплетную или квадруплетную последовательность. Смотри, например, патенты США 6453242 и 6534261 того же заявителя, полное содержание которых включено в настоящий документ посредством ссылки.
Иллюстративные методы отбора, включая системы фагового дисплея и двойных гибридов, описаны в патентах США 5789538, 5925523, 6007988, 6013453, 6410248, 6140466, 6200759 и 6242568, а также в WO 98/37186, WO 98/53057, WO 00/27878, WO 01/88197 и GB 2338237. Кроме того, повышение специфичности связывания для связывающих доменов «цинковых пальцев» описано, например, в WO 02/077227 того же заявителя.
Кроме того, как раскрыто в данных и других литературных источниках, домены «цинковые пальцы» и/или белки с несколькими «цинковыми пальцами» могут быть связаны вместе при помощи любых подходящих последовательностей линкеров, включая, например, линкеры из 5 или более аминокислот в длину. Смотри также патенты США №№6479626, 6903185 и 7153949 для иллюстративных последовательностей линкеров, имеющих 6 или более аминокислот в длину. Белки, описанные в настоящем документе, могут включать любое сочетание подходящих линкеров между отдельными «цинковыми пальцами» белка.
Выбор сайтов-мишеней, ZFPs и способы разработки и конструирования слитых белков (и полинуклеотидов, кодирующих их) известны специалистам в данной области и описаны подробно в патентах США №№61400815, 789538, 6453242, 6534261, 5925523, 6007988, 6013453, 6200759, WO 95/19431, WO 96/06166, WO 98/53057, WO 98/54311, WO 00/27878, WO 01/60970, WO 01/88197, WO 02/099084, WO 98/53058, WO 98/53059, WO 98/53060, WO 02/016536 и WO 03/016496.
Кроме того, как раскрыто в данных и других литературных источниках, домены «цинковые пальцы» и/или белки с несколькими «цинковыми пальцами» могут быть связаны вместе при помощи любых подходящих последовательностей линкеров, включая, например, линкеры из 5 или более аминокислот в длину. Смотри также патенты США №№6479626, 6903185 и 7153949 для иллюстративных последовательностей линкеров, имеющих 6 или более аминокислот в длину. Белки, описанные в настоящем документе, могут включать любое сочетание подходящих линкеров между отдельными «цинковыми пальцами» белка.
В некоторых примерах ДНК-связывающий полипептид представляет собой ДНК-связывающий домен из GAL4. GAL4 является модульным трансактиватором в Saccharomyces cerevisiae, однако он также действует как трансактиватор и во многих других организмах. Смотри, например, Sadowski et al. (1988) Nature 335: 563-4. В данной регуляторной системе экспрессия генов, кодирующих ферменты метаболического пути галактозы в S. cerevisiae, строго регулируется доступным источником углерода. Johnston (1987) Microbiol. Rev. 51: 458-76. Транскрипционный контроль этих метаболических ферментов опосредован взаимодействием между белком положительной регуляции, GAL4, и симметричной последовательностью ДНК из 17 п.н., с которой GAL4 специфически связывается (UAS).
Природный GAL4 состоит из 881 аминокислотного остатка и имеет молекулярную массу 99 кДа. GAL4 содержит функционально автономные домены, совокупная активность которых определяет активность GAL4 in vivo. Ma and Ptashne (1987) Cell 48: 847-53), Brent and Ptashne (1985) Cell 43(3 Pt 2): 729-36. N-концевые 65 аминокислот GAL4 составляют ДНК-связывающий домен GAL4. Keegan et al. (1986) Science 231: 699-704, Johnston (1987) Nature 328: 353-5. Для специфичного для последовательности связывания необходимо наличие двухвалентного катиона, координируемого 6 остатками Cys, присутствующими в ДНК-связывающем домене. Скоординированный катион-содержащий домен взаимодействует с, и узнает консервативный триплет CCG на каждом конце 17-п.н. UAS посредством прямых контактов с большой бороздкой спирали ДНК. Marmorstein et al. (1992) Nature 356: 408-14. ДНК-связывающая функция белка помещает C-концевые активирующие транскрипцию домены в непосредственной близости от промотора таким образом, что активирующие домены могут управлять транскрипцией.
Дополнительные ДНК-связывающие полипептиды, которые можно использовать в некоторых вариантах осуществления, включают, например, и без ограничения, связывающую последовательность из AVRBS3-индуцируемого гена, консенсусную связывающую последовательность из AVRBS3-индуцируемого гена или синтетическую связывающую последовательность, сконструированную на их основе (например, ДНК-связывающий домен UPA), TAL, LexA (смотри, например, Brent & Ptashne (1985), выше), LacR (смотри, например, Labow et al. (1990) Mol. Cell. Biol. 10: 3343-56, Baim et al. (1991) Proc. Natl. Acad. Sci. USA 88(12): 5072-6), рецептор стероидных гормонов (Ellliston et al. (1990) J. Biol. Chem. 265: 11517-121), репрессор Tet (патент США 6271341) и мутантный репрессор Tet, который связывается с последовательностью оператора tet в присутствии, но не в отсутствие, тетрациклина (Tc), ДНК-связывающий домен NF-κB, а также компоненты регуляторной системы, описанной в Wang et al. (1994) Proc. Natl. Acad. Sci. USA 91(17): 8180-4, где используют слитый продукт из GAL4, рецептора гормона и VP16.
В некоторых вариантах осуществления ДНК-связывающий домен одной или более из нуклеаз, используемых в способах и композициях, описанных в настоящем документе, содержит природный или сконструированный (неприродный) ДНК-связывающий домен TAL-эффектора. Смотри, например, патентную публикацию США №20110301073, полное содержание которой включено в настоящий документ посредством ссылки. Патогенные для растений бактерии рода Xanthomonas, как известно, вызывают множество заболеваний важных сельскохозяйственных культур. Патогенность Xanthomonas зависит от консервативной системы секреции III типа (T3S), которая инъецирует более 25 различных эффекторных белков в растительную клетку. Среди этих инъецированных белков находятся подобные активаторам транскрипции (TAL) эффекторы, которые имитируют активаторы транскрипции растений и управляют транскриптомом растения (смотри Kay et al. (2007) Science 318: 648-651). Эти белки содержат ДНК-связывающий домен и домен активации транскрипции. Одним из наиболее хорошо охарактеризованных TAL-эффекторов является AvrBs3 из Xanthomonas campestgris pv. Vesicatoria (смотри Bonas et al. (1989) Mol Gen Genet 218: 127-136 и WO 2010079430). TAL-эффекторы содержат централизованный домен тандемных повторов, при этом каждый повтор содержит примерно 34 аминокислоты, которые являются ключевыми для специфичности ДНК-связывания данных белков. Кроме того, они содержат последовательность с ядерной локализацией и кислый домен активации транскрипции (для обзора смотри Schornack S, et al. (2006) J Plant Physiol 163(3): 256-272). Кроме того, в фитопатогенных бактериях Ralstonia solanacearum обнаружены два гена, обозначенные brg11 и hpx17, которые гомологичны семейству AvrBs3 из Xanthomonas, в R. solanacearum биоваре 1 штамме GMI1000 и в биоваре 4 штамме RS1000 (смотри Heuer et al. (2007) Appl and Envir Micro 73(13): 4379-4384). Эти гены на 98,9% идентичны друг другу по нуклеотидной последовательности, но отличаются делецией 1,575 п.н. в домене повтора hpx17. Однако оба генных продукта имеют менее 40% идентичности последовательностей с белками семейства AvrBs3 из Xanthomonas. Смотри, например, патенты США №№8420782 и 8440431 и патентную публикацию США №20110301073.
В других вариантах осуществления нуклеаза содержит систему CRISPR/Cas. Локус CRISPR (кластерные короткие палиндромные повторы, разделенные регулярными промежутками), который кодирует РНК компоненты системы, и локус cas (CRISPR-ассоциированный), который кодирует белки (Jansen et al., 2002. Mol. Microbiol. 43: 1565-1575, Makarova et al., 2002. Nucleic Acids Res. 30: 482-496, Makarova et al., 2006. Biol. Direct 1: 7, Haft et al., 2005. PLoS Comput. Biol. 1: e60), составляют генные последовательности нуклеазной системы CRISPR/Cas. Локусы CRISPR в микробных хозяевах содержат комбинацию CRISPR-ассоциированных (Cas) генов, а также некодирующие элементы РНК, способные программировать специфичность CRISPR-опосредованного расщепления нуклеиновых кислот.
CRISPR II типа является одной из наиболее хорошо охарактеризованных систем и осуществляет направленный двухцепочечный разрыв ДНК в четыре последовательных этапа. Во-первых, две некодирующие РНК, пре-crRNA комплекс и tracrRNA, транскрибируются с локуса CRISPR. Во-вторых, tracrRNA гибридизируется с областями повторов в пре-crRNA и опосредует процессинг пре-crRNA в зрелые crRNA, содержащие отдельные спейсерные последовательности. В-третьих, комплекс зрелые crRNA:tracrRNA направляет Cas9 на ДНК-мишень за счет спаривания оснований по принципу Уотсона-Крика между спейсером на crRNA и протоспейсером на ДНК-мишени рядом с прилегающим к протоспейсеру мотивом (PAM), дополнительное требование для узнавания мишени. В конечном итоге, Cas9 опосредует расщепление ДНК-мишени, с образованием двухцепочечного разрыва внутри протоспейсера. Активность системы CRISPR/Cas включает три этапа: (i) вставку чужеродных последовательностей ДНК в комплекс CRISPR для предотвращения будущих атак в процессе, называемом «адаптацией», (ii) экспрессию соответствующих белков, а также экспрессию и процессинг комплекса, с последующей (iii) РНК-опосредуемой интерференцией с чужеродной нуклеиновой кислотой. Таким образом, в бактериальной клетке несколько так называемых белков «Cas» связаны с естественной функцией системы CRISPR/Cas и принимают участие в таких функциях, как вставка чужеродной ДНК и так далее.
В некоторых вариантах осуществления белок Cas может быть «функциональным производным» природного белка Cas. «Функциональное производное» полипептида с природной последовательностью представляет собой соединение, имеющее качественное биологическое свойство, общее с полипептидом с природной последовательностью. «Функциональные производные» включают, но не ограничиваются ими, фрагменты полипептида с природной последовательностью и производные полипептида с природной последовательностью и их фрагменты при условии, что они имеют биологическую активность, общую с соответствующим полипептидом с природной последовательностью. Биологическая активность, рассматриваемая в настоящем документе, представляет собой способность функционального производного гидролизовать ДНК-субстрат до фрагментов. Термин «производное» охватывает как вариантные аминокислотные последовательности полипептида, ковалентные модификации, так и его слитые белки. Подходящие производные полипептида Cas или его фрагмента включают, но не ограничиваются ими, мутанты, слитые продукты, ковалентные модификации белка Cas или его фрагмента. Белок Cas, который включает белок Cas или его фрагмент, а также производные белка Cas или его фрагмента, можно получать из клеток или синтезировать химически или получать с использованием сочетания этих двух способов. Клетка может быть клеткой, которая естественным образом продуцирует белок Cas, или клеткой, которая естественным образом продуцирует белок Cas и была модифицирована генно-инженерными методами для продуцирования эндогенного белка Cas на более высоком уровне экспрессии или для продуцирования белка Cas с экзогенной введенной нуклеиновой кислоты, при этом нуклеиновая кислота кодирует Cas, который является таким же или отличающимся от эндогенного Cas. В некоторых случаях клетка не продуцирует белок Cas естественным образом и ее модифицируют генно-инженерными методами для продуцирования белка Cas.
В конкретных вариантах осуществления ДНК-связывающий полипептид специфически узнает и связывается с нуклеотидной последовательностью-мишенью, содержащейся в геномной нуклеиновой кислоте организма-хозяина. Любое количество отдельных вариантов нуклеотидной последовательности-мишени можно найти в геноме хозяина в некоторых примерах. Нуклеотидная последовательность-мишень может быть редкой в геноме организма (например, менее чем примерно 10, примерно 9, примерно 8, примерно 7, примерно 6, примерно 5, примерно 4, примерно 3, примерно 2 или примерно 1 копии(й) последовательности-мишени может существовать в геноме). Например, нуклеотидная последовательность-мишень может быть расположена в уникальном сайте в геноме организма. Нуклеотидные последовательности-мишени могут быть, например, и без ограничения, случайным образом распределены по геному относительно друг друга, расположены в разных группах сцепления в геноме, расположены в одной и той же группе сцепления, расположены на разных хромосомах, расположены на одной и той же хромосоме, расположены в геноме на сайтах, которые экспрессируются в аналогичных условиях в организме (например, под контролем одних и тех же, или в значительной степени функционально идентичных регуляторных факторов), и расположены близко друг к другу в геноме (например, последовательности-мишени могут находиться в составе нуклеиновых кислот, интегрированных в виде конкатемеров в геномных локусах).
B. Направленные эндонуклеазы
В конкретных вариантах осуществления ДНК-связывающий полипептид, который специфически узнает и связывается с нуклеотидной последовательностью-мишенью, может находиться в составе химерного полипептида так, чтобы придавать способность к специфическому связыванию с последовательностью-мишенью химерному полипептиду. В примерах такой химерный полипептид может содержать, например, и без ограничения, полипептиды нуклеазы, рекомбиназы и/или лигазы, такие как полипептиды, описанные выше. Химерные полипептиды, содержащие ДНК-связывающий полипептид и полипептид нуклеазы, рекомбиназы и/или лигазы, могут также содержать другие функциональные полипептидные мотивы и/или домены, такие как, например, и без ограничения: спейсерная последовательность, расположенная между функциональными полипептидами в химерном белке, лидерный пептид, пептид, который направляет слитый белок в органеллу (например, ядро), полипептиды, которые расщепляются клеточным ферментом, пептидные маркеры (например, Myc, His и так далее) и другие аминокислотные последовательности, которые не препятствуют функции химерного полипептида.
Функциональные полипептиды (например, ДНК-связывающие полипептиды и полипептиды нуклеаз) в химерном полипептиде могут быть функционально связаны. В некоторых вариантах осуществления функциональные полипептиды химерного полипептида могут быть функционально связаны за счет их экспрессии с одного полинуклеотида, кодирующего по меньшей мере функциональные полипептиды, лигированные друг с другом в рамке считывания, так, что образуется химерный ген, кодирующий химерный белок. В альтернативных вариантах осуществления функциональные полипептиды химерного полипептида могут быть функционально связаны с помощью других средств, например, за счет сшивки независимо экспрессированных полипептидов.
В некоторых вариантах осуществления ДНК-связывающий полипептид, который специфически узнает и связывается с нуклеотидной последовательностью-мишенью, может находиться в составе природного выделенного белка (или его мутанта), при этом природный выделенный белок или его мутант также содержит полипептид нуклеазы (и может также содержать полипептид рекомбиназы и/или лигазы). Примеры таких выделенных белков включают TALENs, рекомбиназы (например, рекомбиназу Cre, Hin, Tre и FLP), РНК-направляемые CRISPR-Cas9 и мегануклеазы.
Используемый в настоящем документе термин «направленная эндонуклеаза» относится к природным или сконструированным выделенным белкам и их мутантам, содержащим ДНК-связывающий полипептид и полипептид нуклеазы, а также к химерным полипептидам, содержащим ДНК-связывающий полипептид и нуклеазу. Любую направленную эндонуклеазу, содержащую ДНК-связывающий полипептид, который специфически узнает и связывается с нуклеотидной последовательностью-мишенью, находящейся в локусе FAD2 (например, либо вследствие того, что последовательность-мишень находится в составе природной последовательности в локусе, либо вследствие того, что последовательность-мишень была введена в локус, например, путем рекомбинации), можно использовать в некоторых вариантах осуществления.
Некоторые примеры химерных полипептидов, которые могут быть полезны в конкретных вариантах осуществления изобретения, включают, без ограничения, сочетания следующих полипептидов: ДНК-связывающие полипептиды «цинковые пальцы», полипептид нуклеазы FokI, домены TALE, лейциновые «молнии», ДНК-связывающие мотивы фактора транскрипции и узнающие и/или расщепляющие ДНК домены, выделенные, например, и без ограничения, из TALEN, рекомбиназы (например, рекомбиназ Cre, Hin, RecA, Tre и FLP), РНК-направляемых CRISPR-Cas9, мегануклеазы и других, известных специалистам в данной области. Конкретные примеры включают химерный белок, содержащий сайт-специфический ДНК-связывающий полипептид и полипептид нуклеазы. Химерные полипептиды можно конструировать способами, известными специалистам в данной области, для изменения последовательности узнавания ДНК-связывающего полипептида, находящегося в составе химерного полипептида, с тем, чтобы направлять химерный полипептид на конкретную интересующую нуклеотидную последовательность.
В некоторых вариантах осуществления химерный полипептид содержит ДНК-связывающий домен (например, «цинковый палец», домен TAL-эффектора и так далее) и домен нуклеазы (расщепляющий). Расщепляющий домен может быть гетерологичным для ДНК-связывающего домена, например, ДНК-связывающий домен «цинковый палец» и расщепляющий домен из нуклеазы или ДНК-связывающий домен TALEN и расщепляющий домен, или ДНК-связывающий домен мегануклеазы и расщепляющий домен из другой нуклеазы. Гетерологичные расщепляющие домены можно получать из любой эндонуклеазы или экзонуклеазы. Иллюстративные эндонуклеазы, из которых расщепляющий домен может быть получен, включают, но не ограничиваются ими, эндонуклеазы рестрикции и хоминг-эндонуклеазы. Смотри, например, 2002-2003 Catalogue, New England Biolabs, Beverly, MA, и Belfort et al. (1997) Nucleic Acids Res. 25: 3379-3388. Известны дополнительные ферменты, которые расщепляют ДНК (например, нуклеаза S1, нуклеаза золотистой фасоли, панкреатическая ДНКаза I, микрококковая нуклеаза, дрожжевая HO эндонуклеаза, смотри также Linn et al. (eds.) Nucleases, Cold Spring Harbor Laboratory Press, 1993). Один или более из этих ферментов (или их функциональных фрагментов) можно использовать в качестве источника расщепляющих доменов и расщепляющих полудоменов.
Аналогично, расщепляющий полудомен можно получать из любой нуклеазы или ее части, как указано выше, которым требуется димеризация для расщепляющей активности. Как правило, для расщепления необходимы два слитых белка, если слитые белки содержат расщепляющие полудомены. Альтернативно, можно использовать один белок, содержащий два расщепляющих полудомена. Два расщепляющих полудомена можно получать из одной и той же эндонуклеазы (или ее функциональных фрагментов) или каждый расщепляющий полудомен можно получать из отдельной эндонуклеазы (или ее функциональных фрагментов). Кроме того, сайты-мишени для двух слитых белков предпочтительно расположены по отношению друг к другу таким образом, что связывание двух слитых белков с их соответствующими сайтами-мишенями приводит к размещению расщепляющих полудоменов в такой пространственной ориентации по отношению друг к другу, которая позволяет расщепляющим полудоменам образовывать функциональный расщепляющий домен, например, в результате димеризации. Таким образом, в некоторых вариантах осуществления ближние края сайтов-мишеней разделены 5-8 нуклеотидами или 15-18 нуклеотидами. Однако любое целое число нуклеотидов или пар нуклеотидов может находиться между двумя сайтами-мишенями (например, от 2 до 50 пар нуклеотидов или более). Как правило, сайт расщепления лежит между сайтами-мишенями.
Эндонуклеазы рестрикции (ферменты рестрикции) присутствуют во многих видах и способны к специфичному для последовательности связыванию с ДНК (в сайте узнавания) и расщеплению ДНК в сайте связывания или рядом с ним, например, таким образом, что одна или более экзогенных последовательностей (доноры/трансгены) интегрируются в сайтах связывания (мишенях) или рядом с ними. Некоторые ферменты рестрикции (например, типа IIS) расщепляют ДНК в сайтах, удаленных от сайта узнавания, и имеют разделяемые связывающие и расщепляющие домены. Например, фермент FokI типа IIS катализирует двухцепочечное расщепление ДНК через 9 нуклеотидов от его сайта узнавания на одной цепи и через 13 нуклеотидов от его сайта узнавания на другой. Смотри, например, патенты США 5356802, 5436150 и 5487994, а также Li et al. (1992) Proc. Natl. Acad. Sci. USA 89: 4275-4279, Li et al. (1993) Proc. Natl. Acad. Sci. USA 90: 2764-2768, Kim et al. (1994a) Proc. Natl. Acad. Sci. USA 91: 883-887, Kim et al. (1994b) J. Biol. Chem. 269: 31978-31982. Таким образом, в одном варианте осуществления слитые белки содержат расщепляющий домен (или расщепляющий полудомен) из по меньшей мере одного фермента рестрикции типа IIS и один или более связывающих доменов «цинковых пальцев», которые могут быть или не быть сконструированными.
Иллюстративным ферментом рестрикции типа IIS, расщепляющий домен которого отделим от связывающего домена, является FokI. Этот конкретный фермент активен в форме димера. Bitinaite et al. (1998) Proc. Natl. Acad. Sci. USA 95: 10570-10575. Соответственно, для целей настоящего изобретения часть фермента FokI, используемую в описанных слитых белках, считают расщепляющим полудоменом. Таким образом, для направленного двухцепочечного расщепления и/или направленной замены клеточных последовательностей с использованием слитых белков «цинковый палец»-FokI, два слитых белка, каждый из которых содержит расщепляющий полудомен FokI, можно использовать для восстановления каталитически активного расщепляющего домена. Альтернативно, также можно использовать одну полипептидную молекулу, содержащую ДНК-связывающий домен и два расщепляющих полудомена FokI.
Расщепляющий домен или расщепляющий полудомен может представлять собой любую часть белка, сохраняющую расщепляющую активность, или сохраняющую способность к мультимеризации (например, димеризации), с образованием функционального расщепляющего домена.
Иллюстративные ферменты рестрикции типа IIS описаны в патентной публикации США №20070134796, полное содержание которой включено в настоящий документ посредством ссылки. Дополнительные ферменты рестрикции также содержат разделяемые связывающие и расщепляющие домены, и они предусмотрены по настоящему изобретению. Смотри, например, Roberts et al. (2003) Nucleic Acids Res. 31: 418-420.
В некоторых вариантах осуществления расщепляющий домен содержит один или более сконструированных расщепляющих полудоменов (также называемых димеризующимися мутантными доменами), которые сводят к минимуму или предотвращают гомодимеризацию, как описано, например, в патентных публикациях США №№20050064474, 20060188987 и 20080131962, полное содержание всех из которых включено в настоящий документ посредством ссылки. Аминокислотные остатки в положениях 446, 447, 479, 483, 484, 486, 487, 490, 491, 496, 498, 499, 500, 531, 534, 537 и 538 в FokI все являются мишенями, влияющими на димеризацию расщепляющих полудоменов FokI.
Иллюстративные сконструированные расщепляющие полудомены FokI, которые образуют облигатные гетеродимеры, включают пару, в которой первый расщепляющий полудомен содержит мутации аминокислотных остатков в положениях 490 и 538 в FokI и второй расщепляющий полудомен содержит мутации аминокислотных остатков 486 и 499.
Таким образом, в одном варианте осуществления мутация в положении 490 заменяет Glu (E) на Lys (K), мутация в положении 538 заменяет Iso (I) на Lys (K), мутация в положении 486 заменяет Gln (Q) на Glu (E) и мутация в положении 499 заменяет Iso (I) на Lys (K). Конкретно, сконструированные расщепляющие полудомены, описанные в настоящем документе, были получены в результате мутаций в положениях 490 (E→K) и 538 (I→K) в одном расщепляющем полудомене, с получением сконструированного расщепляющего полудомена, обозначенного «E490K:I538K», и в результате мутаций в положениях 486 (Q→E) и 499 (I→L) в другом расщепляющем полудомене, с получением сконструированного расщепляющего полудомена, обозначенного «Q486E:I499L». Сконструированные расщепляющие полудомены, описанные в настоящем документе, являются облигатными мутантными гетеродимерами, для которых аберрантное расщепление сведено к минимуму или устранено. Смотри, например, патентную публикацию США №2008/0131962, содержание которой включено в настоящий документ посредством ссылки в полном объеме для всех целей.
В некоторых вариантах осуществления сконструированный расщепляющий полудомен содержит мутации в положениях 486, 499 и 496 (нумерация соответствует FokI дикого типа), например, мутации, которые заменяют остаток Gln (Q) дикого типа в положении 486 на остаток Glu (E), остаток Iso (I) дикого типа в положении 499 на остаток Leu (L) и остаток Asn (N) дикого типа в положении 496 на остаток Asp (D) или Glu (E) (в так называемых доменах «ELD» и «ELE», соответственно). В других вариантах осуществления сконструированный расщепляющий полудомен содержит мутации в положениях 490, 538 и 537 (нумерация соответствует FokI дикого типа), например, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 на остаток Lys (K), остаток Iso (I) дикого типа в положении 538 на остаток Lys (K) и остаток His (H) дикого типа в положении 537 на остаток Lys (K) или остаток Arg (R) (в так называемых доменах «KKK» и «KKR», соответственно). В других вариантах осуществления сконструированный расщепляющий полудомен содержит мутации в положениях 490 и 537 (нумерация соответствует FokI дикого типа), например, мутации, которые заменяют остаток Glu (E) дикого типа в положении 490 на остаток Lys (K) и остаток His (H) дикого типа в положении 537 на остаток Lys (K) или остаток Arg (R) (в так называемых доменах «KIK» и «KIR», соответственно). (Смотри патентную публикацию США №20110201055). Сконструированные расщепляющие полудомены, описанные в настоящем документе, можно получать с использованием любого подходящего метода, например, путем сайт-направленного мутагенеза расщепляющих полудоменов (FokI) дикого типа, как описано в патентных публикациях США №№20050064474, 20080131962 и 20110201055.
Альтернативно, нуклеазы могут быть собраны in vivo в сайте-мишени нуклеиновой кислоты с использованием так называемой технологии «разделенных ферментов» (смотри, например, патентную публикацию США №20090068164). Компоненты таких разделенных ферментов могут экспрессироваться либо на отдельных экспрессионных конструктах, либо могут быть связаны в одной открытой рамке считывания, в которой отдельные компоненты разделены, например, последовательностью саморасщепляющегося пептида 2A или IRES. Компоненты могут представлять собой отдельные связывающие домены «цинковые пальцы» или домены связывающего нуклеиновую кислоту домена мегануклеазы.
C. Нуклеазы с «цинковыми пальцами»
В конкретных вариантах осуществления химерный полипептид представляет собой специально разработанную нуклеазу с «цинковыми пальцами» (ZFN), которая может быть спроектирована для доставки специфического для сайта-мишени двухцепочечного разрыва ДНК, в который может быть интегрирована экзогенная нуклеиновая кислота или донорская ДНК (смотри патентную публикацию США 20100257638 того же заявителя, включенную в настоящий документ посредством ссылки). ZFNs представляют собой химерные полипептиды, содержащие неспецифический расщепляющий домен из эндонуклеазы рестрикции (например, FokI) и полипептид ДНК-связывающего домена «цинкового пальца». Смотри, например, Huang et al. (1996) J. Protein Chem. 15: 481-9, Kim et al. (1997a) Proc. Natl. Acad. Sci. USA 94: 3616-20, Kim et al. (1996) Proc. Natl. Acad. Sci. USA 93: 1156-60, Kim et al. (1994) Proc Natl. Acad. Sci. USA 91: 883-7, Kim et al. (1997b) Proc. Natl. Acad. Sci. USA 94: 12875-9, Kim et al. (1997c) Gene 203: 43-9, Kim et al. (1998) Biol. Chem. 379: 489-95, Nahon and Raveh (1998) Nucleic Acids Res. 26: 1233-9, Smith et al. (1999) Nucleic Acids Res. 27: 674-81. В некоторых вариантах осуществления ZFNs содержат неканонические ДНК-связывающие домены «цинковые пальцы» (смотри патентную публикацию США 20080182332 того же заявителя, включенную в настоящий документ посредством ссылки). Эндонуклеаза рестрикции FokI должна димеризоваться через домен нуклеазы, чтобы расщеплять ДНК и вносить двухцепочный разрыв. Следовательно, ZFNs, содержащим домен нуклеазы из такой эндонуклеазы, также требуется димеризация домена нуклеазы, чтобы расщеплять ДНК-мишень. Mani et al. (2005) Biochem. Biophys. Res. Commun. 334: 1191-7, Smith et al. (2000) Nucleic Acids Res. 28: 3361-9. Димеризации ZFN могут способствовать два смежных, противоположно ориентированных ДНК-связывающих сайта. Там же.
Гибкость и специфичность системы ZFN обеспечивает уровень контроля, прежде недостижимый при использовании известных стратегий опосредованного рекомбиназой корректирования генов. В качестве одного из примеров, ZFNs можно легко сконструировать, например, для узнавания специфических последовательностей нуклеиновой кислоты. Wu et al. (2007) Cell. Mol. Life Sci. 64: 2933-44 (смотри патентные публикации США 20090205083, 20110189775, 20110167521 и 20100199389, полное содержание которых включено в настоящий документ посредством ссылки). Рандомизация кодонов для узнающих остатков «цинковых пальцев» позволяет отбирать новые «пальцы», которые имеют высокую аффинность для произвольно выбранных последовательностей ДНК. Кроме того, «цинковые пальцы» являются природными ДНК-связывающими молекулами, и было показано, что сконструированные «цинковые пальцы» действуют на предназначенные для них мишени в живых клетках. Таким образом, нуклеазы на основе «цинковых пальцев» могут быть направлены на специфические, но произвольно выбранные сайты узнавания.
В конкретных примерах способ сайт-специфического интегрирования экзогенной нуклеиновой кислоты в по меньшей мере один функциональный локус FAD2 хозяина включает введение в клетку-хозяина ZFN, при этом ZFN узнает и связывается с нуклеотидной последовательностью-мишенью, причем нуклеотидная последовательность-мишень находится в по меньшей мере одном локусе FAD2 хозяина. В некоторых примерах нуклеотидная последовательность-мишень не находится в геноме хозяина в каком-либо ином положении, чем по меньшей мере один локус FAD2. Например, ДНК-связывающий полипептид из ZFN можно сконструировать для узнавания и связывания с нуклеотидной последовательностью-мишенью, идентифицированной в по меньшей мере одном локусе FAD2 (например, путем секвенирования локуса FAD2). Способ сайт-специфического интегрирования экзогенной нуклеиновой кислоты в по меньшей мере один функциональный локус FAD2 хозяина, который включает введение в клетку-хозяина ZFN, может также включать введение в клетку экзогенной нуклеиновой кислоты, при этом рекомбинация экзогенной нуклеиновой кислоты в нуклеиновую кислоту хозяина, содержащую по меньшей мере один локус FAD2, облегчается сайт-специфическим узнаванием и связыванием ZFN с последовательностью-мишенью (и последующим расщеплением нуклеиновой кислоты, содержащей локус FAD2).
V. Экзогенные нуклеиновые кислоты для интегрирования в локус FAD2
Варианты осуществления изобретения могут включать одну или более нуклеиновых кислот, выбранных из группы, состоящей из: экзогенной нуклеиновой кислоты для сайт-специфического интегрирования в по меньшей мере один локус FAD2, например, и без ограничения, PTU, ELP, ETIP или ORF; нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую направленную эндонуклеазу; и вектор, содержащий по меньшей мере либо одну, либо обе из вышеуказанных. Таким образом, конкретные нуклеиновые кислоты для использования в некоторых вариантах осуществления включают нуклеотидные последовательности, кодирующие полипептид, структурные нуклеотидные последовательности и/или сайты узнавания и связывания ДНК-связывающего полипептида.
A. Экзогенные молекулы нуклеиновой кислоты для сайт-специфического интегрирования
Как отмечалось выше, вставку экзогенной последовательности (также называемой «донорской последовательностью» или «донором», или «трансгеном») осуществляют, например, для экспрессии полипептида, корректировки мутантного гена или для повышения экспрессии гена дикого типа. Очевидно, что донорская последовательность, как правило, не идентична геномной последовательности, в которую ее помещают. Донорская последовательность может содержать негомологичную последовательность, фланкированную двумя областями гомологии для осуществления эффективной HDR в интересующий участок. Кроме того, донорские последовательности могут содержать молекулу вектора, содержащую последовательности, которые не гомологичны интересующей области в клеточном хроматине. Донорская молекула может содержать несколько прерывистых областей гомологии с клеточным хроматином. Например, для направленной вставки последовательностей, обычно не присутствующих в интересующей области, указанные последовательности могут присутствовать в донорской молекуле нуклеиновой кислоты и быть фланкированы областями гомологии с последовательностью в интересующей области.
Донорский полинуклеотид может представлять собой ДНК или РНК, быть одноцепочечным или двухцепочечным и может быть введен в клетку в линейной или кольцевой форме. Смотри, например, патентные публикации США №№20100047805, 20110281361, 20110207221 и патентную заявку США №13/889162. При введении в линейной форме концы донорской последовательности могут быть защищены (например, от экзонуклеолитической деградации) методами, известными специалистам в данной области. Например, один или более дидезоксинуклеотидных остатков добавляют к 3'-концу линейной молекулы и/или самокомплементарные олигонуклеотиды лигируют на одном или обоих концах. Смотри, например, Chang et al. (1987) Proc. Natl. Acad. Sci. USA 84: 4959-4963, Nehls et al. (1996) Science 272: 886-889. Дополнительные методы защиты экзогенных полинуклеотидов от деградации включают, но не ограничиваются ими, добавление концевой аминогруппы(групп) и использование модифицированных межнуклеотидных связей, таких как, например, фосфоротиоаты, фосфорамидаты, а также остатков O-метилрибозы или дезоксирибозы.
Полинуклеотид можно вводить в клетку в виде части молекулы вектора, имеющего дополнительные последовательности, такие как, например, точки начала репликации, промоторы и гены, кодирующие устойчивость к антибиотику. Кроме того, донорские полинуклеотиды можно вводить в виде «голой» нуклеиновой кислоты, в виде нуклеиновой кислоты в комплексе с таким средством, как липосома или полоксамер, или можно доставлять с помощью вирусов (например, аденовируса, AAV, герпесвируса, ретровируса, лентивируса и дефектного по интегразе лентивируса (IDLV)).
Донор, как правило, интегрируется таким образом, что его экспрессия управляется эндогенным промотором в сайте интеграции, а именно, промотором, который управляет экспрессией эндогенного гена, в который донор интегрирован (например, FAD2). Однако очевидно, что донор может содержать промотор и/или энхансер, например, конститутивный промотор или индуцируемый, или тканеспецифичный промотор.
Кроме того, хотя для экспрессии в этом нет необходимости, экзогенные последовательности могут также включать регуляторные последовательности транскрипции или трансляции, например, промоторы, энхансеры, инсуляторы, внутренний сайт связывания рибосом, последовательности, кодирующие пептиды 2A и/или сигналы полиаденилирования.
Экзогенные нуклеиновые кислоты, которые могут быть интегрированы сайт-специфическим образом в по меньшей мере один локус FAD2 для модификации локуса FAD2, в вариантах осуществления включают, например, и без ограничения, нуклеиновые кислоты, содержащие нуклеотидную последовательность, кодирующую интересующий полипептид; нуклеиновые кислоты, содержащие нужный с агрономической точки зрения ген; нуклеиновые кислоты, содержащие нуклеотидную последовательность, кодирующую молекулу РНКи, или нуклеиновые кислоты, которые разрушают ген FAD2.
В некоторых вариантах осуществления экзогенная нуклеиновая кислота интегрируется в локус FAD2 для модификации локуса FAD2, при этом нуклеиновая кислота содержит нужный с агрономической точки зрения ген или нуклеотидную последовательность, кодирующую интересующий полипептид, таким образом, что нужный с агрономической точки зрения ген или нуклеотидная последовательность экспрессируется в организме хозяина с локуса FAD2. В некоторых примерах интересующий полипептид (например, чужеродный белок) экспрессируется с нуклеотидной последовательности, кодирующей интересующий полипептид, в коммерческих количествах. В таких примерах интересующий полипептид можно экстрагировать из клетки, ткани или биомассы хозяина. В некоторых вариантах осуществления хозяином является растение, и растительный материал, поставляемый для коммерческого производства интересующего полипептида, может представлять собой растение, часть растения, ткань растения или клетку растения. В некоторых примерах часть растения может быть семенем растения. Экстракцию белка из растительной биомассы можно осуществлять известными методами, которые описаны, например, в Heney and Orr (1981) Anal. Biochem. 114: 92-6.
Аналогично, нужные с агрономической точки зрения гены могут быть экспрессированы в трансформированных клетках растений, растениях и/или их потомстве. Например, растение может быть генетически модифицировано с помощью способов по конкретным вариантам осуществления для экспрессии различных фенотипов интересующих с агрономической точки зрения генов с по меньшей мере одного локуса FAD2.
В некоторых вариантах осуществления нуклеиновые кислоты, содержащие нужный с агрономической точки зрения ген или нуклеотидную последовательность, кодирующую интересующий полипептид, могут включать, например, и без ограничения: ген, придающий устойчивость к вредителям или заболеваниям (смотри, например, Jones et al. (1994) Science 266: 789 (клонирование гена томатов Cf-9 для устойчивости к Cladosporium fulvum), Martin et al. (1993) Science 262: 1432, Mindrinos et al. (1994) Cell 78: 1089 (ген RSP2 для устойчивости к Pseudomonas syringae), международную патентную публикацию PCT № WO 96/30517 (устойчивость к соевой цистообразующей нематоде), международную патентную публикацию PCT № WO 93/19181); ген, кодирующий белок Bacillus thuringiensis, его производное или синтетический полипептид, смоделированный на его основе (смотри, например, Geiser et al. (1986) Gene 48: 109 (клонирование и нуклеотидная последовательность гена Bt δ-эндотоксина, кроме того, молекулы ДНК, кодирующие гены δ-эндотоксина можно приобретать в Американской коллекции типовых культур (Manassas, VA), например, под регистрационными №№ ATCC 40098, 67136, 31995 и 31998)); ген, кодирующий лектин (смотри, например, Van Damme et al. (1994) Plant Molec. Biol. 24: 25 (нуклеотидные последовательности некоторых генов манноза-связывающего лектина Clivia miniata)); ген, кодирующий витамин-связывающий белок, например, авидин (смотри международную патентную публикацию PCT № US 93/06487 (использование авидина и гомологов авидина в качестве ларвицидов против насекомых-вредителей)); ген, кодирующий ингибитор фермента, например, протеазы, ингибитор протеазы или ингибитор амилазы (смотри, например, Abe et al. (1987) J. Biol. Chem. 262: 16793 (нуклеотидная последовательность ингибитора цистеиновой протеазы риса), Huub et al. (1993) Plant Molec. Biol. 21: 985 (нуклеотидная последовательность кДНК, кодирующей ингибитор протеиназы I табака), Sumitani et al. (1993) Biosci. Biotech. Biochem. 57: 1243 (нуклеотидная последовательность ингибитора альфа-амилазы Streptomyces nitrosporeus) и патент США 5494813); ген, кодирующий специфичный для насекомых гормон или феромон, например, экдистероидный или ювенильный гормон, его вариант, миметик на его основе, или его антагонист или агонист (смотри, например, Hammock et al. (1990) Nature 344: 458 (бакуловирусная экспрессия клонированной эстеразы ювенильного гормона, инактиватора ювенильного гормона)); ген, кодирующий специфичный для насекомых пептид или нейропептид, который при экспрессии нарушает физиологические процессы подвергшегося воздействию вредителя (смотри, например, Regan (1994) J. Biol. Chem. 269: 9 (экспрессионное клонирование приводит к получению ДНК, кодирующей рецептор диуретического гормона насекомых), Pratt et al. (1989) Biochem. Biophys. Res. Comm. 163: 1243 (аллостатин в Diploptera puntata) и патент США 5266317 (гены, кодирующие специфичные для насекомых паралитические нейротоксины)); ген, кодирующий специфичный для насекомых яд, вырабатываемый в природе змеями, осами или другими организмами (смотри, например, Pang et al. (1992) Gene 116: 165 (гетерологичная экспрессия в растениях гена, кодирующего токсичный для насекомых пептид скорпиона)); ген, кодирующий фермент, ответственный за гипераккумуляцию монотерпена, сесквитерпена, стероида, гидроксамовой кислоты, фенилпропаноидного производного или другой молекулы с инсектицидной активностью; ген, кодирующий фермент, вовлеченный в модификацию, включая посттрансляционную модификацию, биологически активной молекулы, например, гликолитического фермента, протеолитического фермента, липолитического фермента, нуклеазы, циклазы, трансаминазы, эстеразы, гидролазы, фосфатазы, киназы, фосфорилазы, полимеразы, эластазы, хитиназы или глюканазы, либо природный, либо синтетический (смотри, например, международную патентную публикацию PCT № WO 93/02197 (нуклеотидная последовательность гена каллазы), кроме того, молекулы ДНК, содержащие кодирующие хитиназу последовательности, могут быть получены, например, из ATCC под регистрационными №№39637 и 67152, Kramer et al. (1993) Insect Biochem. Molec. Biol. 23: 691 (нуклеотидная последовательность кДНК, кодирующей хитиназу табачного бражника) и Kawalleck et al. (1993) Plant Molec. Biol. 21: 673 (нуклеотидная последовательность гена ubi4-2 полиубиквитина петрушки)); ген, кодирующий молекулу, стимулирующую сигнальную трансдукцию (смотри, например, Botella et al. (1994) Plant Molec. Biol. 24: 757 (нуклеотидные последовательности кДНК-клонов кальмодулина золотистой фасоли) и Griess et al. (1994) Plant Physiol. 104: 1467 (нуклеотидная последовательность кДНК-клона кальмодулина кукурузы)); ген, кодирующий пептид с гидрофобным моментом (смотри, например, международную патентную публикацию PCT № WO 95/16776 (пептидные производные тахиплезина, ингибирующие грибковые патогены растений) и международную патентную публикацию PCT № WO 95/18855 (синтетические противомикробные пептиды, придающие устойчивость к заболеваниям)); ген, кодирующий мембранную пермеазу, образователь каналов или блокатор каналов (смотри, например, Jaynes et al. (1993) Plant Sci 89: 43 (гетерологичная экспрессия аналога литического пептида цекропина-β для придания трансгенным растениям табака устойчивости к Pseudomonas solanacearum)); ген, кодирующий вирусный инвазивный белок или полученный из него комплексный токсин (смотри, например, Beachy et al. (1990) Ann. rev. Phytopathol. 28: 451); ген, кодирующий специфичное для насекомых антитело или полученный из него иммунотоксин (смотри, например, Taylor et al., Abstract #497, Seventh Int'l Symposium on Molecular Plant-Microbe Interactions (Edinburgh, Scotland) (1994) (ферментативная инактивация в трансгенных растениях табака за счет выработки одноцепочечных фрагментов антитела)); ген, кодирующий специфичное для вируса антитело (смотри, например, Tavladoraki et al. (1993) Nature 366: 469 (трансгенные растения, экспрессирующие гены рекомбинантного антитела, защищены от атаки вирусов)); ген, кодирующий останавливающий развитие белок, продуцируемый в природе патогенами или паразитами (смотри, например, Lamb et al. (1992) Bio/Technology 10: 1436 (грибковые эндо-α-1,4-D-полигалактуроназы способствуют грибковой колонизации и высвобождению питательных веществ для растений путем солюбилизации клеточной стенки растений гомо-α-1,4-D-галактуроназой), Toubart et al. (1992) Plant J. 2: 367 (клонирование и характеризация гена, кодирующего ингибирующий эндополигалактуроназу белок фасоли)); ген, кодирующий останавливающий развитие белок, продуцируемый в природе растениями (смотри, например, Logemann et al. (1992) Bio/Technology 10: 305 (трансгенные растения, экспрессирующие ген, инактивирующий рибосомы ячменя, обладают повышенной устойчивостью к грибковым заболеваниям)).
В некоторых вариантах осуществления нуклеиновые кислоты, содержащие нужный с агрономической точки зрения ген или нуклеотидную последовательность, кодирующую интересующий полипептид, могут также и/или альтернативно включать, например, и без ограничения: гены, придающие устойчивость к гербицидам, таким как гербицид, который ингибирует точку роста или меристему, например, имидазолинон или сульфонилмочевина (иллюстративные гены в этой категории кодируют мутантные ферменты ALS и AHAS, описанные, например, в Lee et al. (1988) EMBO J. 7: 1241, и Miki et al. (1990) Theor. Appl. Genet. 80: 449, соответственно); устойчивость к глифосату, придаваемую, например, мутантными генами 5-енолпирувилшикимат-3-фосфатсинтазы (EPSPs) (за счет введения рекомбинантных нуклеиновых кислот и/или различных форм in vivo мутагенеза природных генов EPSPs (включая, но не ограничиваясь ими, CP4, DMMG и DGT-28); гены aroA и гены глифосат-ацетилтрансферазы (GAT), соответственно); другим фосфоновым соединениям, таким как глюфосинат (гены фосфинотрицин-ацетилтрансферазы (PAT) из видов Streptomyces, включая Streptomyces hygroscopicus и Streptomyces viridichromogenes), и пиридинокси- или феноксипропионовым кислотам и циклогексонам (гены, кодирующие ингибитор АССазы). Смотри, например, патенты США 4940835 и 6248876 (нуклеотидные последовательности форм EPSPs, которые способны придавать растениям устойчивость к глифосату). Молекулу ДНК, кодирующую мутантный ген aroA, можно получать в ATCC под регистрационным № 39256. Смотри также патент США №4769061 (нуклеотидная последовательность мутантного гена aroA). В европейской патентной заявке №0333033 и патенте США №4975374 раскрыты нуклеотидные последовательности генов глутаминсинтетазы, которые могут придавать устойчивость к таким гербицидам, как L-фосфинотрицин. Нуклеотидные последовательности иллюстративных генов PAT приведены в европейской патентной заявке № 0 242 246, и DeGreef et al. (1989) Bio/Technology 7: 61 (получение трансгенных растений, экспрессирующих химерные гены bar, кодирующие активность PAT). Примеры генов, придающих устойчивость к феноксипропионовым кислотам и циклогексонам, таким как сетоксидим и галоксифоп, включают гены Acc1-S1, Acc1-S2 и Acc1-S3, описанные в Marshall et al. (1992) Theor. Appl. Genet. 83: 435. Гены GAT, способные придавать устойчивость к глифосату, описаны, например, в WO 2005012515. Гены, придающие устойчивость к 2,4-D, пиридинокси- и феноксипропионовой кислоте, ауксиновым гербицидам, описаны, например, в WO 2005107437 и WO 2007053482.
Нуклеиновые кислоты, содержащие нужный с агрономической точки зрения ген или нуклеотидную последовательность, кодирующую интересующий полипептид, могут также включать, например, и без ограничения: ген, придающий устойчивость к такому гербициду, который ингибирует фотосинтез, например триазину (гены psbA и gs+) или бензонитрилу (ген нитрилазы). Смотри, например, Przibila et al. (1991) Plant Cell 3: 169 (трансформация Chlamydomonas плазмидами, кодирующими мутантные гены psbA). Нуклеотидные последовательности генов нитрилазы раскрыты в патенте США 4810648, и молекулы ДНК, содержащие эти гены, доступны под регистрационными №№ ATCC 53435, 67441 и 67442. Смотри также Hayes et al. (1992) Biochem. J. 285: 173 (клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу).
В некоторых вариантах осуществления нуклеиновые кислоты, содержащие нужный с агрономической точки зрения ген или нуклеотидную последовательность, кодирующую интересующий полипептид, могут также и/или альтернативно включать гены, которые придают или вносят вклад в добавляющий ценность признак, например, и без ограничения: модифицированный метаболизм жирных кислот, например, путем трансформации растения антисмысловым геном стеарил-ACP-десатуразы для увеличения содержания стеариновой кислоты в растении (смотри, например, Knultzon et al. (1992) Proc. Natl. Acad. Sci. U.S.A. 89: 2624); сниженное содержание фитатов, например, введение кодирующего фитазу гена может увеличивать разрушение фитатов, добавляя больше свободного фосфата для трансформированного растения (смотри, например, Van Hartingsveldt et al. (1993) Gene 127: 87 (нуклеотидная последовательность гена фитазы Aspergillus niger), можно вводить ген для снижения содержания фитатов в кукурузе, например, это можно осуществлять путем клонирования и затем повторного введения ДНК, ассоциированной с одним аллелем, которая может приводить к созданию мутантов кукурузы, характеризующихся низкими уровнями фитиновой кислоты (смотри Raboy et al. (1990) Maydica 35: 383)); и модифицированный углеводный состав, получаемый, например, путем трансформации растений геном, кодирующим фермент, который изменяет характер ветвления крахмала (смотри, например, Shiroza et al. (1988) J. Bacteol. 170: 810 (нуклеотидная последовательность мутантного гена фруктозилтрансферазы Streptococcus), Steinmetz et al. (1985) Mol. Gen. Genet. 20: 220 (ген левансахаразы), Pen et al. (1992) Bio/Technology 10: 292 (α-амилаза), Elliot et al. (1993) Plant Molec. Biol. 21: 515 (нуклеотидные последовательности генов инвертазы томатов), Sogaard et al. (1993) J. Biol. Chem. 268: 22480 (ген α-амилазы ячменя) и Fisher et al. (1993) Plant Physiol. 102: 1045 (фермент ветвления крахмала II эндосперма кукурузы)).
В некоторых вариантах осуществления экзогенная нуклеиновая кислота интегрируется в локус FAD2 для модификации локуса FAD2, при этом нуклеиновая кислота содержит PTU или ELP, так что, например, облегчается последующее сайт-специфическое интегрирование второй экзогенной нуклеиновой кислоты в сайт PTU или ELP. Смотри также патентную заявку США №13/889162.
Для опосредованного направленными эндонуклеазами интегрирования интересующей молекулы нуклеиновой кислоты в геном растения путем направленного интегрирования необходима доставка направленных эндонуклеаз или кодирующих направленные эндонуклеазы молекул нуклеиновой кислоты, с последующей экспрессией функционального белка направленной эндонуклеазы в хозяине. Предпочтительно, экзогенная нуклеиновая кислота также присутствует в клетке-хозяине в то самое время, когда в ней происходит доставка или экспрессия направленной эндонуклеазы, так что функциональный белок направленной эндонуклеазы индуцирует двухцепочечные разрывы в сайте(ах)-мишени в по меньшей мере одном локусе FAD2, которые затем восстанавливаются, например, путем управляемого гомологией интегрирования экзогенной нуклеиновой кислоты в локус. Специалистам в данной области понятно, что экспрессию функционального белка направленной эндонуклеазы можно осуществлять несколькими методами, включая, но не ограничиваясь ими, трансгенез кодирующего направленную эндонуклеазу конструкта и временную экспрессию кодирующего направленную эндонуклеазу конструкта. В обоих этих случаях экспрессию функционального белка направленной эндонуклеазы и доставку экзогенной нуклеиновой кислоты в клетку-хозяина можно осуществлять одновременно, чтобы стимулировать направленное интегрирование в локус FAD2.
Особым преимуществом, получаемым в вариантах осуществления с использованием ZFNs в качестве направленных эндонуклеаз, является то, что необходимость в димеризации расщепляющих доменов химерных нуклеаз с «цинковыми пальцами» приводит к высокому уровню специфичности узнавания и, следовательно, расщепления последовательности. Поскольку каждый набор из трех «пальцев» связывает девять последовательных пар оснований, для двух химерных нуклеаз обязательно требуется 18-п.н. мишень, если каждый домен «цинковый палец» имеет идеальную специфичность. Любая конкретная последовательность такой длины, по прогнозам, будет уникальной в составе одного генома (предположительно примерно 109 п.н.). Bibikova et al. (2001) Mol. Cell. Biol. 21(1): 289-97, Wu et al. (2007), выше. Кроме того, дополнительные «пальцы» могут обеспечивать повышенную специфичность, Beerli et al. (1998) Proc. Natl. Acad. Sci. USA 95: 14628-33, Kim and Pabo (1998) Proc. Natl. Acad. Sci. USA 95: 2812-7, Liu et al. (1997) Proc. Natl. Acad. Sci. USA 94: 5525-30, так что количество «цинковых пальцев» в каждом ДНК-связывающем домене можно увеличивать для обеспечения еще большей специфичности. Например, специфичность можно дополнительно увеличивать путем использования пары ZFNs с 4, 5, 6 или более «пальцами», которые узнают 24-п.н. последовательность. Urnov et al. (2005) Nature 435: 646-51. Таким образом, ZFNs можно использовать так, что последовательность узнавания для введения в геном растения-хозяина будет уникальной в составе генома.
B. Молекулы нуклеиновой кислоты, содержащие нуклеотидную последовательность, кодирующую направленную эндонуклеазу
В некоторых вариантах осуществления нуклеотидную последовательность, кодирующую направленную эндонуклеазу, можно конструировать путем манипулирования (например, лигирования) природными нуклеотидными последовательностями, кодирующими полипептиды, находящиеся в составе направленной эндонуклеазы. Например, нуклеотидную последовательность гена, кодирующего белок, содержащий ДНК-связывающий полипептид, можно внимательно изучать для идентификации нуклеотидной последовательности гена, которая соответствует ДНК-связывающему полипептиду, и эту нуклеотидную последовательность можно использовать в качестве элемента нуклеотидной последовательности, кодирующей направленную эндонуклеазу, содержащую ДНК-связывающий полипептид. Альтернативно, аминокислотную последовательность направленной эндонуклеазы можно использовать для выведения нуклеотидной последовательности, кодирующей направленную эндонуклеазу, например, с учетом вырожденности генетического кода.
В иллюстративных молекулах нуклеиновой кислоты, содержащих нуклеотидную последовательность, кодирующую направленную эндонуклеазу, последний кодон первой полинуклеотидной последовательности, кодирующей полипептид нуклеазы, и первый кодон второй полинуклеотидной последовательности, кодирующей ДНК-связывающий полипептид, можно разделять любым количеством триплетов нуклеотидов, например, без кодирования интрона или сигнала остановки «стоп». Аналогично, последний кодон нуклеотидной последовательности, кодирующей первую полинуклеотидную последовательность, кодирующую ДНК-связывающий полипептид, и первый кодон второй полинуклеотидной последовательности, кодирующей полипептид нуклеазы, можно разделять любым количеством триплетов нуклеотидов. В этих и других вариантах осуществления последний кодон (то есть, наиболее 3'-расположенный в нуклеотидной последовательности) первой полинуклеотидной последовательности, кодирующей полипептид нуклеазы, и вторая полинуклеотидная последовательность, кодирующая ДНК-связывающий полипептид, могут быть слиты в фазовом регистре с первым кодоном дополнительной полинуклеотидной кодирующей последовательности, непосредственно примыкающей к ним, или отделенной от них не более чем короткой пептидной последовательностью, например, такой как закодированная синтетическим нуклеотидным линкером (например, нуклеотидным линкером, который мог быть использован для получения слитого продукта). Примеры таких дополнительных полинуклеотидных последовательностей включают, например, и без ограничения, маркеры, направленные пептиды и сайты ферментативного расщепления. Аналогично, первый кодон, наиболее 5'-расположенный (в нуклеотидной последовательности), первой и второй полинуклеотидных последовательностей могут быть слиты в фазовом регистре с последним кодоном дополнительной полинуклеотидной кодирующей последовательности, непосредственно примыкающей к ним, или отделенной от них не более чем короткой пептидной последовательностью.
Последовательность, разделяющая полинуклеотидные последовательности, кодирующие функциональные полипептиды в направленной эндонуклеазе (например, ДНК-связывающий полипептид и полипептид нуклеазы), могут, например, состоять из любой последовательности так, чтобы кодируемая аминокислотная последовательность не изменяла в значительной степени трансляцию направленной эндонуклеазы. В связи с автономным характером известных полипептидов нуклеазы и известных ДНК-связывающих полипептидов, в примерах промежуточные последовательности не будут препятствовать соответствующим функциям данных структур.
C. Векторы и экспрессионные конструкты
В некоторых вариантах осуществления по меньшей мере одну молекулу(ы) нуклеиновой кислоты, содержащую по меньшей мере одну экзогенную полинуклеотидную последовательность, кодирующую интересующий полипептид и/или направленную эндонуклеазу, можно вводить в клетку, ткань или организм для экспрессии в них. Например, молекулу нуклеиновой кислоты, содержащую полинуклеотидную последовательность, кодирующую направленную эндонуклеазу, которая специфически узнает нуклеотидную последовательность, содержащуюся в по меньшей мере одном локусе FAD2, можно вводить в клетку для экспрессии направленной эндонуклеазы, и молекулу нуклеиновой кислоты, содержащую полинуклеотидную последовательность, кодирующую интересующий полипептид, можно вводить в клетку так, что полинуклеотидная последовательность, кодирующая интересующий полипептид, интегрируется в по меньшей мере один локус FAD2, например, путем гомологичной рекомбинации после внесения двухцепочечного разрыва в локусе экспрессированной направленной эндонуклеазой, и интересующий полипептид экспрессируется с интегрированной полинуклеотидной последовательности.
В некоторых вариантах осуществления молекула нуклеиновой кислоты, такая как одна из вышеперечисленных, может, например, представлять собой векторную систему, включая, например, и без ограничения, линейную плазмиду или замкнутую кольцевую плазмиду. В конкретных примерах вектор может представлять собой экспрессионный вектор. Нуклеотидные последовательности по конкретным вариантам осуществления могут, например, быть интегрированы в вектор так, что нуклеотидная последовательность является функционально связанной с одной или более регуляторными последовательностями. Множество векторов доступны для данной цели, и выбор конкретного вектора может зависеть, например, от размера нуклеиновой кислоты, которую предстоит вставлять в вектор, конкретной клетки-хозяина, которую предстоит трансформировать вектором, и/или количества любого кодируемого полипептида, экспрессия которого желательна. Вектор, как правило, содержит различные компоненты, характер которых зависит от функции вектора (например, амплификация ДНК или экспрессия ДНК) и конкретной клетки(ок)-хозяина, с которой вектор совместим.
В некоторых вариантах осуществления регуляторная последовательность, функционально связанная с одной или более кодирующими последовательностями, может представлять собой последовательность промотора, который функционирует в клетке-хозяине, такой как бактериальная клетка, клетка водоросли, грибная клетка или растительная клетка, при этом молекула нуклеиновой кислоты будет амплифицироваться или экспрессироваться. Некоторые варианты осуществления могут включать трансформирующий вектор растений, который содержит нуклеотидную последовательность, содержащую по меньшей мере одну регуляторную последовательность, функционально связанную с одной или более нуклеотидными последовательностями, кодирующими интересующий полипептид или направленную эндонуклеазу, при этом одна или более нуклеотидные последовательности могут экспрессироваться под контролем регуляторной последовательности(ей) в клетке, ткани или организме растения для продуцирования интересующего полипептида или направленной эндонуклеазы.
Промоторы, подходящие для использования в молекулах нуклеиновой кислоты по некоторым вариантам осуществления, включают те, которые являются индуцируемыми, тканеспецифичными, вирусными, синтетическими или конститутивными, все из которых хорошо известны в данной области. Неограничивающие примеры промоторов, которые могут быть полезны в вариантах осуществления изобретения, приведены в: патентах США №№6437217 (промотор RS81 кукурузы), 5641876 (промотор актина риса), 6426446 (промотор RS324 кукурузы), 6429362 (промотор PR-1 кукурузы), 6232526 (промотор A3 кукурузы), 6177611 (конститутивные промоторы кукурузы), 5322938, 5352605, 5359142 и 5530196 (промотор 35S), 6433252 (промотор олеозина L3 кукурузы), 6429357 (промотор актина 2 риса и интрон актина 2 риса), 6294714 (индуцируемые светом промоторы), 6140078 (индуцируемые солью промоторы), 6252138 (индуцируемые патогенами промоторы), 6175060 (индуцируемые дефицитом фосфора промоторы), 6388170 (двунаправленные промоторы), 6635806 (промотор гамма-коиксина), 5447858 (промотор соевого белка теплового шока) и в патентной заявке США с серийным №09/757089 (промотор альдолазы хлоропластов кукурузы).
Дополнительные иллюстративные промоторы включают промотор нопалинсинтазы (NOS) (Ebert et al. (1987) Proc. Natl. Acad. Sci. USA 84(16): 5745-9), промотор октопинсинтазы (OCS) (который находится на индуцирующих опухоли плазмидах Agrobacterium tumefaciens), колимовирусные промоторы, такие как промотор 19S вируса мозаики цветной капусты (CaMV) (Lawton et al. (1987) Plant Mol. Biol. 9: 315-24), промотор 35S CaMV (Odell et al. (1985) Nature 313: 810-2, промотор 35S вируса мозаики норичника (Walker et al. (1987) Proc. Natl. Acad. Sci. USA 84(19): 6624-8), промотор сахароза синтазы (Yang and Russell (1990) Proc. Natl. Acad. Sci. USA 87: 4144-8), промотор комплекса гена R (Chandler et al. (1989) Plant Cell 1: 1175-83), промотор гена хлорофилл a/b-связывающего белка, 35S CaMV (патенты США №№5322938, 5352605, 5359142 и 5530196), 35S FMV (патенты США №№6051753 и 5378619), промотор PC1SV (патент США № 5850019), промотор SCP1 (патент США №6677503) и промоторы AGRtu.nos (регистрационный № GenBank V00087, Depicker et al. (1982) J. Mol. Appl. Genet. 1: 561-73, Bevan et al. (1983) Nature 304: 184-7).
В конкретных вариантах осуществления молекулы нуклеиновой кислоты могут содержать тканеспецифичный промотор. Тканеспецифичный промотор представляет собой нуклеотидную последовательность, которая направляет транскрипцию функционально связанной нуклеотидной последовательности на высоком уровне в ткани, для которой промотор является специфичным, по сравнению с другими тканями организма. Примеры тканеспецифичных промоторов включают, без ограничения: специфичные для тапетума промоторы, специфичные для пыльника промоторы, специфичные для пыльцы промоторы (смотри, например, патент США №7141424 и международную патентную публикацию PCT № WO 99/042587), специфичные для семяпочки промоторы, (смотри, например, патентную заявку США №2001/047525 A1), специфичные для плодов промоторы (смотри, например, патенты США №№4943674 и 5753475) и специфичные для семян промоторы (смотри, например, патенты США №№5420034 и 5608152). В некоторых вариантах осуществления можно использовать специфичный для стадии развития промотор (например, промотор, активный на более поздней стадии развития).
Дополнительные регуляторные последовательности, которые в некоторых вариантах осуществления могут быть функционально связаны с молекулой нуклеиновой кислоты, включают 5' UTRs, расположенные между последовательностью промотора и кодирующей последовательностью, которые действуют как лидерные последовательности трансляции. Лидерная последовательность трансляции присутствует в полностью процессированной мРНК и она может влиять на процессинг первичного транскрипта и/или стабильность РНК. Примеры лидерных последовательностей трансляции включают лидерные последовательности белков теплового шока кукурузы и петунии (патент США №5362865), лидерные последовательности белков оболочки вирусов растений, лидерные последовательности rubisco растений и другие. Смотри, например, Turner and Foster (1995) Molecular Biotech. 3(3): 225-36. Неограничивающие примеры 5' UTRs включают: GmHsp (патент США №5659122), PhDnaK (патент США № 5362865), AtAnt1, TEV (Carrington and Freed (1990) J. Virol. 64: 1590-7) и AGRtunos (регистрационный № GenBank V00087, а также Bevan et al. (1983), выше).
Дополнительные регуляторные последовательности, которые в некоторых вариантах осуществления могут быть функционально связаны с молекулой нуклеиновой кислоты, также включают 3'-нетранслируемые последовательности, 3'-области терминации транскрипции или области полиаденилирования. Они представляют собой генетические элементы, расположенные ниже по ходу транскрипции от нуклеотидной последовательности, и включают полинуклеотиды, образующие сигнал полиаденилирования и/или другие регуляторные сигналы, способные влиять на транскрипцию или процессинг мРНК. Сигнал полиаденилирования в растениях действует, вызывая присоединение полиаденилат нуклеотидов к 3'-концу мРНК-предшественника. Последовательность полиаденилирования можно получать из различных генов растений, или из генов T-ДНК. Неограничивающим примером 3'-области терминации транскрипции является 3'-область нопалинсинтазы (nos 3'; Fraley et al. (1983) Proc. Natl. Acad. Sci. USA 80: 4803-7). Пример использования различных 3'-нетранслируемых областей приведен в Ingelbrecht et al. (1989) Plant Cell 1: 671-80. Неограничивающие примеры сигналов полиаденилирования включают сигнал полиаденилирования из гена RbcS2 Pisum sativum (Ps.RbcS2-E9; Coruzzi et al. (1984) EMBO J. 3: 1671-9) и AGRtu.nos (регистрационный № GenBank E01312).
Дополнительная информация о регуляторных последовательностях, которые могут быть полезны в конкретных вариантах осуществления, приведена, например, в Goeddel (1990) «Gene Expression Technology», Methods Enzymol. 185, Academic Press, San Diego, CA.
Рекомбинантная молекула нуклеиновой кислоты или вектор могут содержать селективный маркер, который придает селектируемый фенотип трансформированной клетке, такой как клетка растения. Селективные маркеры также можно использовать для отбора клеток или организмов, которые содержат молекулу нуклеиновой кислоты, содержащую селективный маркер. Маркер может кодировать устойчивость к биоциду, устойчивость к антибиотику (например, канамицину, генетицину (G418), блеомицину и гигромицину) или устойчивость к гербициду (например, глифосату). Примеры селективных маркеров включают, но не ограничиваются ими: ген neo, который придает устойчивость к канамицину и может быть отобран с использованием, например, канамицина и G418; ген bar, который придает устойчивость к биалафосу; мутантный ген EPSP-синтазы, который придает устойчивость к глифосату; ген нитрилазы, который придает устойчивость к бромоксинилу; мутантный ген ацетолактатсинтазы (ALS), который придает устойчивость к имидазолинону или сульфонилмочевине, и ген устойчивости к метотрексату DHFR. Доступно множество селективных маркеров, которые придают устойчивость к химическим веществам, включая, например, и без ограничения, ампициллин, блеомицин, хлорамфеникол, гентамицин, гигромицин, канамицин, линкомицин, метотрексат, фосфинотрицин, пуромицин, спектиномицин, рифампицин, стрептомицин и тетрациклин. Примеры таких селективных маркеров приведены, например, в патентах США 5550318, 5633435, 5780708 и 6118047.
Молекула нуклеиновой кислоты или вектор могут также, или альтернативно, включать скринируемый маркер. Скринируемые маркеры можно использовать для мониторинга экспрессии. Иллюстративные скринируемые маркеры включают ген β-глюкуронидазы или uidA (GUS), кодирующий фермент, для которого известны различные хромогенные субстраты (Jefferson et al. (1987) Plant Mol. Biol. Rep. 5: 387-405); ген R-локуса, кодирующий продукт, который регулирует выработку пигментов антоцианинов (красного цвета) в тканях растений (Dellaporta et al. (1988) «Molecular cloning of the maize R-nj allele by transposon tagging with Ac», в: 18th Stadler Genetics Symposium, P. Gustafson and R. Appels, eds., Plenum, NY (pp. 263-82); ген β-лактамазы (Sutcliffe et al. (1978) Proc. Natl. Acad. Sci. USA 75: 3737-41); ген, кодирующий фермент, для которого известны различные хромогенные субстраты (например, PADAC, хромогенный цефалоспорин); ген люциферазы (Ow et al. (1986) Science 234: 856-9); ген xylE, кодирующий катехол диоксигеназу, которая преобразует хромогенные катехолы (Zukowski et al. (1983) Gene 46(2-3): 247-55); ген амилазы (Ikatu et al. (1990) Bio/Technol. 8: 241-2); ген тирозиназы, кодирующий фермент, способный окислять тирозин до ДОФА и допахинона, который, в свою очередь, конденсируется в меланин (Katz et al. (1983) J. Gen. Microbiol. 129: 2703-14), и ген α-галактозидазы.
Все из нуклеотидных последовательностей, которые кодируют, например, конкретный интересующий полипептид или конкретную направленную эндонуклеазу, могут легко узнать специалисты в данной области. Вырожденность генетического кода обеспечивает конечное число кодирующих последовательностей для конкретной аминокислотной последовательности. Выбор конкретной последовательности для кодирования полипептида по вариантам осуществления изобретения остается на усмотрение практикующего специалиста. Для разных вариантов применения могут быть желательны разные кодирующие последовательности.
В некоторых вариантах осуществления может быть желательной модификация нуклеотидов нуклеиновой кислоты, например, для повышения экспрессии полинуклеотидной последовательности, содержащейся в нуклеиновой кислоте конкретного хозяина. Генетический код является избыточным с 64 возможными кодонами, однако большинство организмов предпочтительно используют подгруппу этих кодонов. Кодоны, которые используются каким-либо видом наиболее часто, называют оптимальными кодонами, а те, которые используются не очень часто, классифицируют как редкие или малоиспользуемые кодоны. Zhang et al. (1991) Gene 105: 61-72. Кодоны могут быть заменены, чтобы отражать предпочтительное использование кодонов конкретным хозяином, в процессе того, что иногда называют «оптимизацией кодонов». Оптимизированные кодирующие последовательности, содержащие кодоны, предпочитаемые конкретным прокариотическим или эукариотическим хозяином, можно получать, например, для повышения скорости трансляции или для получения рекомбинантных РНК-транскриптов, имеющих нужные свойства (например, большее время полужизни по сравнению с транскриптами, полученными с неоптимизированной последовательности).
В вариантах осуществления изобретения нуклеиновые кислоты можно вводить в клетку-хозяина любым методом, известным специалистам в данной области, в том числе, например, и без ограничения: путем трансформации протопластов (смотри, например, патент США 5508184), путем опосредованного обезвоживанием/ингибированием поглощения ДНК (смотри, например, Potrykus et al. (1985) Mol. Gen. Genet. 199: 183-8), путем электропорации (смотри, например, патент США 5384253), путем перемешивания с волокнами карбида кремния (смотри, например, патенты США 5302523 и 5464765), путем опосредованной Agrobacterium трансформации (смотри, например, патенты США 5563055, 5591616, 5693512, 5824877, 5981840 и 6384301) и путем ускорения покрытых ДНК частиц (смотри, например, патенты США 5015580, 5550318, 5538880, 6160208, 6399861 и 6403865). Благодаря применению таких методик, как эти, можно стабильно трансформировать клетки практически любого вида. В некоторых вариантах осуществления трансформирующая ДНК интегрируется в геном клетки-хозяина. В случае многоклеточных видов трансгенные клетки могут быть регенерированы в трансгенный организм. Любую из этих методик можно использовать для получения трансгенного растения, например, содержащего одну или более нуклеотидных последовательностей по изобретению в геноме трансгенного растения.
Наиболее широко используемый метод введения экспрессионного вектора в растения основан на природной системе трансформации Agrobacterium. A. tumefaciens и A. rhizogenes являются патогенными для растений почвенными бактериями, которые генетически трансформируют растительные клетки. Ti и Ri-плазмиды A. tumefaciens и A. rhizogenes, соответственно, несут гены, ответственные за генетическую трансформацию растения. Ti-плазмиды (индуцирующие опухоли) содержат крупный сегмент, известный как T-ДНК, который передается трансформированным растениям. Другой сегмент Ti-плазмиды, область vir, отвечает за перенос T-ДНК. Область T-ДНК ограничена слева и справа границами, каждая из которых состоит из концевых повторяющихся нуклеотидных последовательностей. В некоторых модифицированных бинарных векторах индуцирующие опухоли гены были делетированы, и функции области vir используют для переноса чужеродной ДНК, ограниченной пограничными последовательностями T-ДНК. T-область может также содержать, например, селективный маркер для эффективного отбора трансгенных растений и клеток, а также сайт множественного клонирования для вставки последовательностей, предназначенных для переноса, таких как нуклеиновая кислота, кодирующая слитый белок по изобретению.
Таким образом, в некоторых вариантах осуществления трансформирующий вектор растений получен из Ti-плазмиды A. tumefaciens (смотри, например, патенты США №№4536475, 4693977, 4886937 и 5501967, и европейский патент EP 0122791) или Ri-плазмиды A. rhizogenes. Дополнительные трансформирующие векторы растений включают, например, и без ограничения, те, которые описаны в Herrera-Estrella et al. (1983) Nature 303: 209-13, Bevan et al. (1983), выше, Klee et al. (1985) Bio/Technol. 3: 637-42 и в европейском патенте EP 0120516, а также те, которые получены из любых вышеперечисленных. Другие бактерии, такие как Sinorhizobium, Rhizobium и Mesorhizobium, естественным образом взаимодействующие с растениями, можно модифицировать для осуществления переноса генов в целый ряд разнообразных растений. Эти связанные с растениями симбиотические бактерии можно сделать компетентными для переноса генов, получая как нейтрализованную Ti-плазмиду, так и подходящий бинарный вектор.
После введения экзогенной ДНК в клетки-реципиенты, трансформированные клетки, как правило, идентифицируют для дальнейшего культивирования и регенерации растений. Для улучшения способности идентифицировать трансформированные клетки можно использовать ген селективного или скринируемого маркера, как описано выше, с вектором, используемым для получения трансформанта. В случае использования селективного маркера, трансформированные клетки идентифицируют в популяции потенциально трансформированных клеток, подвергая клетки воздействию селективного средства или средств. В случае использования скринируемого маркера, можно проводить скрининг клеток на признак желаемого маркерного гена.
Клетки, которые выживают при воздействии селективного средства, или клетки, которые были признаны положительными при скрининге, можно культивировать в среде, которая способствует регенерации растений. В некоторых вариантах осуществления любую подходящую для культивирования растительных тканей среду (например, среды MS и N6) можно модифицировать, включая дополнительные вещества, такие как факторы роста. Ткань можно поддерживать на основной среде с регуляторами роста до тех пор, пока не образуется достаточное количество ткани для начала работы по регенерации растений, или после неоднократных раундов отбора вручную до тех пор, пока морфология тканей не станет подходящей для регенерации (например, по меньшей мере 2 недели), затем переносить в среду, способствующую образованию побегов. Культуры периодически пересаживают до тех пор, пока побеги не разовьются в достаточной степени. После образования побегов их пересаживают в среду, способствующую образованию корней. После того, как корни сформируются в достаточной степени, растения можно пересаживать в почву для дальнейшего роста и созревания.
Для подтверждения присутствия интересующей молекулы нуклеиновой кислоты (например, нуклеотидной последовательности, кодирующей полипептид, содержащий по меньшей мере один слитый белок по изобретению) в регенерирующем растении можно проводить различные анализы. Такие анализы включают, например: молекулярно-биологические анализы, такие как саузерн- и нозерн-блоттинг, ПЦР и секвенирование нуклеиновой кислоты; биохимические анализы, такие как обнаружение присутствия белкового продукта, например, иммунологическими методами (ELISA и/или вестерн-блоттинг), или ферментативной функции; анализ частей растения, например, анализ листьев или корней, и анализ фенотипа целого регенерированного растения.
Интеграционные события можно анализировать, например, ПЦР амплификацией с использованием, например, олигонуклеотидных праймеров, которые специфичны для интересующей нуклеотидной последовательности. Следует понимать, что генотипирование методом ПЦР включает, но не ограничивается ими, амплификацию полимеразной цепной реакцией (ПЦР) геномной ДНК, полученной из выделенной ткани растения-хозяина, предположительно содержащего интересующую молекулу нуклеиновой кислоты, интегрированную в геном, с последующим стандартным клонированием и анализом последовательности продуктов ПЦР-амплификации. Методы генотипирования с помощью ПЦР подробно описаны (смотри, например, Rios, G. et al. (2002) Plant J. 32: 243-53) и могут быть применены к геномной ДНК, полученной из любого вида растения или типа ткани, включая клеточные культуры.
Трансгенное растение, полученное с использованием методов Agrobacterium-зависимой трансформации, как правило, содержит от одной до нескольких копий рекомбинантной ДНК. Одну рекомбинантную последовательность ДНК называют «трансгенным событием» или «интеграционным событием». Такие трансгенные растения являются гетерозиготными в отношении вставленной последовательности ДНК. В некоторых вариантах осуществления трансгенное растение, гомозиготное в отношении трансгена, можно получать путем полового скрещивания (самоопыления) независимого сегреганта трансгенного растения, содержащего одну экзогенную последовательность гена, с самим собой, например, растения F0, с получением семян F1. Одна четверть полученных семян F1 будет гомозиготной в отношении трансгена. При проращивании семян F1 образуются растения, которые можно тестировать на гетерозиготность, как правило, с использованием анализа SNP или анализа термоамплификации, который позволяет различать между гетерозиготами и гомозиготами (то есть, анализ зиготности).
В дополнение к непосредственной трансформации растения или клетки растения молекулой нуклеиновой кислоты в некоторых вариантах осуществления, трансгенные растения можно получать в конкретных вариантах осуществления путем скрещивания первого растения, имеющего по меньшей мере одно трансгенное событие, со вторым растением, лишенным такого события. Например, нуклеиновую кислоту, содержащую по меньшей мере один модифицированный локус FAD2, в который экзогенная нуклеиновая кислота была интегрирована сайт-специфическим образом, можно вводить в растение первой линии, которая поддается трансформации, для получения трансгенного растения, которое можно скрещивать с растением второй линии для интрогрессии по меньшей мере одного модифицированного локуса FAD2 (и, следовательно, экзогенной нуклеиновой кислоты) в растение второй линии.
Для подтверждения присутствия интересующей молекулы нуклеиновой кислоты в регенерирующих растениях можно проводить различные анализы. Такие анализы включают, например: молекулярно-биологические анализы, такие как саузерн- и нозерн-блоттинг, и ПЦР; биохимические анализы, такие как обнаружение присутствия белкового продукта, например, иммунологическими методами (ELISA и/или вестерн-блоттинг), или ферментативной функции; анализ частей растения, например, анализ листьев или корней, и анализ фенотипа целого регенерированного растения.
Можно проводить скрининг направленных интеграционных событий, например, методом ПЦР-амплификации с использованием, например, олигонуклеотидных праймеров, специфичных для интересующей молекулы нуклеиновой кислоты. Следует понимать, что генотипирование методом ПЦР включает, но не ограничивается ими, амплификацию полимеразной цепной реакцией (ПЦР) геномной ДНК, полученной из выделенной каллюсной ткани растения-хозяина, предположительно содержащего интересующую молекулу нуклеиновой кислоты, интегрированную в геном, с последующим стандартным клонированием и анализом последовательности продуктов ПЦР-амплификации. Методы генотипирования с помощью ПЦР подробно описаны (например, Rios, G. et al. (2002) Plant J. 32: 243-53) и могут быть применены к геномной ДНК, полученной из любого вида растения или типа ткани, включая клеточные культуры. Сочетания олигонуклеотидных праймеров, которые связываются как с последовательностью-мишенью, так и с введенной последовательностью, можно использовать последовательно или комплексно в реакциях ПЦР-амплификации. Можно создавать олигонуклеотидные праймеры, сконструированные для отжига с сайтом-мишенью, введенными последовательностями нуклеиновой кислоты и/или их сочетаниями. Таким образом, стратегии генотипирования методом ПЦР могут включать (но не ограничиваются ими) амплификацию специфических последовательностей в геноме растения, амплификацию нескольких специфических последовательностей в геноме растения, амплификацию неспецифических последовательностей в геноме растения, или их сочетание. Специалист в данной области может разработать дополнительные сочетания праймеров и реакции амплификации для анализа генома. Например, можно разрабатывать набор прямых и обратных олигонуклеотидных праймеров для отжига с нуклеотидной последовательностью(ми), специфичных для мишени за пределами введенной последовательности нуклеиновой кислоты.
Можно разрабатывать прямые и обратные олигонуклеотидные праймеры для отжига конкретно с введенной интересующей молекулой нуклеиновой кислоты, например, в последовательности, соответствующей кодирующей области в составе интересующей молекулы нуклеиновой кислоты, или других частях интересующей молекулы нуклеиновой кислоты. Эти праймеры можно использовать в сочетании с праймерами, описанными выше. Олигонуклеотидные праймеры могут быть синтезированы в соответствии с желаемой последовательностью и являются коммерчески доступными (например, от компании Integrated DNA Technologies, Inc., Coralville, IA). Вслед за амплификацией можно проводить клонирование и секвенирование или прямой анализ последовательности продуктов амплификации. Специалисту в данной области могут быть известны альтернативные методы анализа продуктов амплификации, полученных в процессе генотипирования методом ПЦР. В одном варианте осуществления в ПЦР-амплификации используют олигонуклеотидные праймеры, специфичные для гена-мишени.
VI. Трансгенные растения и растительные материалы, содержащие нуклеиновую кислоту, интегрированную в функциональный локус FAD2
В некоторых вариантах осуществления предложено трансгенное растение, которое содержит растительную клетку, содержащую по меньшей мере один модифицированный (например, разрушенный и/или содержащий направленно интегрированную экзогенную последовательность) локус FAD2 (например, локус FAD2 2.3 и/или локус FAD2 2.6 сои). В конкретных вариантах осуществления такое растение можно получать путем трансформации ткани растения или клетки растения и регенерации целого растения. В других вариантах осуществления такое растение можно получать путем введения экзогенной нуклеиновой кислоты в по меньшей мере один локус FAD2 сайт-специфическим образом или путем интрогрессии модифицированного локуса FAD2 в зародышевую плазму. Также предложены растительные материалы, содержащие такую растительную клетку. Такой растительный материал можно получать из растения, содержащего растительную клетку.
Трансгенное растение или растительный материал, содержащие клетку растения, содержащую по меньшей мере один модифицированный локус FAD2, в некоторых вариантах осуществления могут проявлять одну или более из следующих характеристик: экспрессию направленной эндонуклеазы в клетке растения, экспрессию интересующего полипептида в клетке растения (или в ее пластиде), экспрессию направленной эндонуклеазы в ядре клетки растения, локализацию направленной эндонуклеазы в клетке растения, интеграцию в локус FAD2 в геноме клетки растения, интеграцию нуклеотидной последовательности, кодирующей интересующий полипептид или нужный с агрономической точки зрения ген, в локус FAD2 в геноме клетки растения и/или присутствие РНК-транскрипта, соответствующего кодирующей последовательности, интегрированной в локус FAD2 в геноме клетки растения. Такое растение может дополнительно иметь один или более из желаемых признаков, включая, например, и без ограничения, те, которые возникают в результате экспрессии эндогенной или трансгенной нуклеотидной последовательности, экспрессия которой регулируется интересующим полипептидом или нужным с агрономической точки зрения геном, интегрированным в локус FAD2 в геноме клетки растения; устойчивость к насекомым, другим вредителям и болезнетворным агентам; устойчивость к гербицидам; повышенную стабильность, урожайность или долговечность при хранении; толерантность к условиям окружающей среды; производство фармацевтических продуктов; производство промышленных продуктов и улучшенные питательные свойства.
Трансгенное растение по изобретению может представлять собой любое растение, способное быть трансформированным нуклеиновой кислотой, которая впоследствии интегрируется в по меньшей мере один локус FAD2 при использовании способов, описанных в настоящем документе. Соответственно, растение может быть двудольным или однодольным. Неограничивающие примеры двудольных растений, которые можно использовать в настоящих способах, включают Arabidopsis, люцерну, бобы, брокколи, капусту, канолу, морковь, цветную капусту, сельдерей, китайскую капусту, хлопок, огурцы, баклажаны, салат, дыню, горох, перец, арахис, картофель, тыкву, редис, рапс, шпинат, сою, сквош, сахарную свеклу, подсолнечник, табак, томаты и арбуз. Неограничивающие примеры однодольных растений, которые можно использовать в настоящих способах, включают кукурузу, ячмень, лук, рис, сорго, пшеницу, рожь, просо, сахарный тростник, овес, тритикале, просо прутьевидное и газонные травы. Трансгенные растения по изобретению можно использовать или культивировать любым способом.
Некоторые варианты осуществления также относятся к товарам, производимым из трансгенных растений по изобретению. Товары включают, например, и без ограничения: продукты питания, муку, масла или дробленые или цельные зерна, или семена растения, содержащего одну или более нуклеотидных последовательностей, интегрированных в по меньшей мере один локус FAD2. Обнаружение одной или более таких нуклеотидных последовательностей в одном или более товарах или товарных продуктах свидетельствует де-факто, что товар или товарный продукт был, по меньшей мере частично, произведен из трансгенного растения, полученного в соответствии с одним из вариантов осуществления изобретения. В некоторых вариантах осуществления трансгенное растение или семя, содержащее растительную клетку, содержащую по меньшей мере один модифицированный локус FAD2, может содержать по меньшей мере одно другое трансгенное событие в своем геноме, включая, без ограничения: трансгенное событие, с которого транскрибируется молекула РНКи; ген, кодирующий инсектицидный белок (например, инсектицидный белок Bacillus thuringiensis); ген устойчивости к гербициду (например, ген, обеспечивающий устойчивость к глифосату) и ген, вносящий вклад в создание желаемого фенотипа трансгенного растения (например, повышенной урожайности, измененного метаболизма жирных кислот или восстановления цитоплазматической мужской стерильности).
Трансгенное растение, содержащее растительную клетку, содержащую по меньшей мере один модифицированный локус FAD2, может иметь один или более желаемых признаков. Такие признаки могут включать, например: устойчивость к насекомым, другим вредителям и болезнетворным агентам, устойчивость к гербицидам, повышенную стабильность, урожайность или долговечность при хранении, толерантность к условиям окружающей среды, производство фармацевтических продуктов, производство промышленных продуктов и улучшенные питательные свойства. Желаемые признаки могут быть приданы одной или более молекулами нуклеиновой кислоты, интегрированными путем направленной рекомбинации в локус FAD2, которые экспрессируются в растении, проявляющем желаемые признаки. Таким образом, в некоторых вариантах осуществления желаемый признак может быть связан с присутствием трансгена(ов) в растении, который был введен в геном растения в сайт по меньшей мере одного модифицированного локуса FAD2. В дополнительном варианте осуществления желаемый признак можно получать при помощи общепринятой селекции, при этом признак может быть придан одной или более молекулами нуклеиновой кислоты, интегрированными путем направленной рекомбинации в по меньшей мере один модифицированный локус FAD2.
Трансгенные растения по изобретению можно использовать или культивировать любым способом, при этом желательно наличие по меньшей мере одного модифицированного локуса FAD2. Соответственно, растение может быть спроектировано так, чтобы, среди прочего, иметь один или более желаемых признаков, будучи трансформированным молекулами нуклеиновой кислоты, которые впоследствии интегрируются сайт-специфическим образом в по меньшей мере один локус FAD2 по изобретению, и быть высажено и культивировано любым методом, известным специалистам в данной области.
VII. Селекция с помощью маркеров трансгенных растений, содержащих нуклеиновую кислоту, интегрированную в функциональный локус FAD2
Предложены молекулярные маркеры, которые связаны (например, тесно связаны) с fad2 в Glycine max. Например, идентифицированы сегменты ДНК, содержащие последовательности, определяющие признак HO (fad2). Эти сегменты расположены вокруг и между маркерами, которые связаны (например, тесно связаны) с мутантными аллелями в геномной группе сцепления. Таким образом, также предложены молекулы нуклеиновой кислоты, содержащие мутантный ген FAD2, имеющий инактивирующую мутацию. Идентифицированные сегменты и их маркеры включены в объект настоящего изобретения, частично за счет их положения в группах сцепления в геноме Glycine max.
Все литературные источники, включая публикации, патенты и патентные заявки, цитированные в настоящем документе, включены в настоящий документ посредством ссылки в такой степени, чтобы они не противоречили четко сформулированным деталям данного изобретения, и так же включены в такой степени, как если было бы индивидуально и специально указано, что каждый литературный источник включен посредством ссылки и изложен в полном объеме в настоящем документе. Литература, обсуждаемая в настоящем документе, приведена исключительно с целью ее раскрытия до даты подачи настоящей заявки. Ничто в настоящем документе не должно быть истолковано как признание того, что авторы изобретения не имеют права датировать задним числом такое раскрытие на основании предшествующего изобретения. Следующие примеры приведены для иллюстрации некоторых конкретных признаков и/или вариантов осуществления. Примеры не должны быть истолкованы как ограничивающие изобретение приведенными в них конкретными признаками или вариантами осуществления.
ПРИМЕРЫ
Пример 1: Секвенирование последовательностей-мишеней FAD2 из пяти культиваров сои
Реакции секвенирования
Геномную ДНК выделяли из тканей сои. Геномную ДНК выделяли и очищали из суспензии лиофилизированных эмбриогенных клеток в случае культиваров X5 и Westag и из молодых листьев в случае культиваров Jack, Williams 82 и Maverick. Геномную ДНК экстрагировали с использованием набора DNeasy Plant Mini Kit™ (Qiagen; Carlsbad, CA) в соответствии с протоколами производителя.
Гены FAD2 2.3 и 2.6
Последовательности геномной ДНК FAD2 2.3 и FAD2 2.6 амплифицировали методом ПЦР с использованием праймеров MA49 (SEQ ID NO: 1 caagggttccaaacacaaagcc) и MA51 (SEQ ID NO: 2 catcaatacttgttcctgtacc) или MA50 (SEQ ID NO: 3 gaagaagcctctctcaagggttc) и MA51. Последовательность геномной ДНК получали для фрагмента из оснований примерно с 40 по 1140 в 1140-п.н. гене. Условия реакции ПЦР были следующими: 1 мин при 98°C для начальной денатурации, затем 35 циклов по 30 сек при 98°C, 15 сек при 60°C, 3 мин при 72°C и окончательное удлинение в течение 5 мин при 72°C.
Полученные ПЦР-ампликоны суспендировали в TE буфере (1 мкг в TE буфере) и разрезали на фрагменты размером примерно 300 п.н. при помощи генератора ультразвука Covaris E220 System™ (Covaris; Woburn, MA) с использованием следующих настроек: пиковая мощность падающей волны 140, относительная длительность 10%, 200 циклов на импульс и время обработки 430 секунд. Библиотеки секвенирования спаренных концов Illumina™ (San Diego, CA) получали с использованием набора PrepX DNA library kit™ (IntegenX; Pleasanton, CA) на Apollo 324TM Automation System™ (IntegenX), следуя протоколу, рекомендованному производителем. Вкратце, разрезанные фрагменты ДНК с 5' или 3' липкими концами были преобразованы в 5' фосфорилированную ДНК с тупыми концами. Один удлиняющий остаток аденина (A) добавляли к 3'-концу фрагментов ДНК с восстановленными концами, с последующим лигированием с индексными парноконцевыми (PE) адаптерами Illumina™. Наконец, лигированные с адаптерами продукты библиотеки извлекали из робота и обогащали с помощью ПЦР, используя реагенты Illumina TruSeq PCR™ в условиях термоциклирования 30 сек при 98°C для начальной денатурации, затем 10 циклов по 10 сек при 98°C, 30 сек при 60°C, 30 сек при 72°C и окончательное удлинение в течение 5 мин при 72°C. Обогащенные библиотеки нормировали до концентрации 2 нМ и объединяли. Затем объединенные библиотеки денатурировали гидроксидом натрия и разбавляли до концентрации 6 пМ в буфере гибридизация для нанесения на Miseq flow cell™ (Illumina; San Diego, CA). Прогон 2×150 циклов с 6 циклами индексных последовательностей проводили на приборе Miseq™ в соответствии с рекомендованным протоколом компании Illumina.
Проводили тримминг полученных в реакциях секвенирования в приборе Illumina Miseq™ парных чтений для TruSeq adapter sequences™ (Illumina). После тримминга чтения были картированы на референсный геном сои культивара Williams 82 с использованием программы Burrows Wheeler Aligner (BWA) (Li H. and Durbin R. (2009) Fast и accurate short read alignment with Burrows-Wheeler Transform. Bioinformatics, 25: 1754-60).
Каждый культивар сои обрабатывали как отдельный образец и, таким образом, чтения при секвенировании каждого культивара картировали отдельно. Области на геноме, где глубина картированных чтений секвенирования была выше нуля, были изучены. Поскольку это были чтения секвенирования с ампликонов, ожидалось, что только специфические области на геномах будут иметь картированные на них чтения. Чтения секвенирования картировались на соевые хромосомы 10 и 20 у различных образцов. Для каждого образца получали консенсусную последовательность с использованием компьютерной программы Mpileup. Эти результаты свидетельствовали о том, что чтения секвенирования картировались на два паралогичных предполагаемых гена FAD. Полученные чтения последовательностей выравнивали для идентификации SNPs/вариаций между паралогичными предполагаемыми генными последовательностями, полученными для культиваров X5, Westag, Jack, Williams 82 и Maverick.
Выравнивание последовательностей выполняли при помощи программы AlignX® из компьютерной программы Vector NTI Advance 11.0 (Life Technologies, Carlsbad, CA) и они приведены на фигуре 1 и фигуре 2. В программе AlignX® используется модифицированный алгоритм Clustal W для выравнивания нескольких последовательностей, или белковых, или нуклеотидных последовательностей, для сравнения подобия и для аннотаций. Как показано на фигуре 1 и фигуре 2, анализ выделенных последовательностей показывает, что соответствующие последовательности FAD2 2.3 и FAD2 2.6 имеют высокие уровни сходства последовательностей.
Ген FAD2 2.3 соответствует основаниям 49417070-49418219 на хромосоме 10 референсной геномной последовательности Williams 82 (SEQ ID NO: 4). Последовательности генов из пяти культиваров (Williams 82, Jack, Maverick, X5 и Westag) были идентичными по основаниям 41-1140 (покрытие, полученное с использованными праймерами). Ген FAD2 2.6 соответствует основаниям 34178330-34179475 на хромосоме 20 референсной геномной последовательности Williams 82 (SEQ ID NO: 9). Различия последовательностей были идентифицированы в последовательности X5 относительно референсной последовательности Williams 82 в положениях 233 (C>T), 352 (A>G), 633 (C>T), 645 (T>C), 658 (T>C), 894 (A>G). В культиваре Maverick были такие же изменения оснований, что и в X5, в положениях 352 и 894.
Пример 2: Разработка связывающих доменов «цинковых пальцев», специфичных для генов FAD2
Белки с «цинковыми пальцами», направленные против последовательностей ДНК, кодирующих различные функциональные последовательности локуса гена FAD2, были разработаны, как описано ранее. Смотри, например, Urnov et al. (2005) Nature 435: 646-651. Иллюстративная последовательность-мишень и спирали узнавания приведены в таблице 1 (сайты-мишени) и таблице 2 (конструкции областей спирали узнавания). Направляющие сайты нуклеазы с «цинковыми пальцами» (ZFN) были разработаны для связывания сайтов-мишеней FAD2. Конструкции с «цинковыми пальцами» для FAD2 были вставлены в экспрессионные векторы с «цинковыми пальцами», кодирующие белок, имеющий по меньшей мере один «палец» со структурой CCHC. Смотри патентную публикацию США №2008/0182332. В частности, последний «палец» в каждом белке имел каркас CCHC для спирали узнавания. Неканонические кодирующие «цинковые пальцы» последовательности были слиты с доменом нуклеазы фермента рестрикции FokI типа IIS (аминокислоты 384-579 последовательности, приведенной в Wah et al., (1998) Proc. Natl. Acad. Sci. USA 95: 10564-10569) через линкер ZC из четырех аминокислот и сигнал ядерной локализации opaque-2, полученный из Zea mays, с образованием нуклеаз с «цинковыми пальцами» (ZFNs) для FAD2. Как домены FokI дикого типа, так и домены eHF-FokI (смотри патентную публикацию США №20110201055) были использованы для конструирования ZFNs 1-3, при том, что только домены eHF-FokI были использованы для ZFNs 4-7.
Активность ZFNs, разработанных для FAD2 2.3 и 2.6, проверяли в анализе DLSSA (смотри публикацию патентной заявки США №20110301073) для идентификации ZFNs с наивысшей активностью. Оценивали расщепление ZFNs соответствующих последовательностей FAD2 2.3 и 2.6, клонированных в клетки млекопитающих (фигура 3). Активность сравнивали с высокоактивной референсной ZFN (8266:8196); указана базовая активность.
Сайты-мишени FAD2 для «цинковых пальцев»
Конструкции «цинковых пальцев» для FAD2 мономеров ZFN для сайта связывания ZFN
Пример 3: Конструкты нуклеаз с «цинковыми пальцами» и доноров
Конструкты ZFN
Были разработаны и получены плазмидные векторы, содержащие экспрессионные конструкты ZFN для иллюстративных нуклеаз с «цинковыми пальцами», которые были идентифицированы с помощью анализа, описанного в примере 2.
Каждая кодирующая «цинковые пальцы» последовательность была слита с последовательностью, кодирующей сигнал ядерной локализации opaque-2 (Maddaloni et al. (1989) Nuc. Acids Res. 17(18): 7532), которая была помещена выше нуклеазы с «цинковыми пальцами». Затем слитая последовательность сигнал ядерной локализации opaque-2::нуклеаза с «цинковыми пальцами» была спарена с комплементарной слитой последовательностью сигнал ядерной локализации opaque-2::нуклеаза с «цинковыми пальцами». Как таковой, каждый конструкт состоял из одной открытой рамки считывания, состоящей из двух слитых последовательностей сигнал ядерной локализации opaque-2::нуклеаза с «цинковыми пальцами», разделенных последовательностью 2A из вируса Thosea asigna (Mattion et al. (1996) J. Virol. 70: 8124-8127). Экспрессия слитых белков управлялась относительно сильным конститутивным промотором, таким как промотор из вируса мозаики жилок маниоки (CsVMV), и они были фланкированы 3' нетранслируемой областью ORF23 Agrobacterium tumefaciens (AtuORF23 3'UTR).
Векторы были собраны с использованием In-Fusion™ Advantage Technology (Clontech, Mountain View, CA). Эндонуклеазы рестрикции приобретали у компании New England BioLabs (NEB; Ipswich, MA) и ДНК-лигазу T4 (Invitrogen) использовали для лигирования ДНК. Плазмиды получали с использованием набора NucleoSpin® Plasmid Kit (Macherey-Nagel Inc., Bethlehem, PA) или Plasmid Midi Kit (Qiagen), следуя инструкциям поставщиков. Фрагменты ДНК выделяли, используя набор QIAquick Gel Extraction Kit™ (Qiagen) после электрофореза в агарозном геле с трис-ацетатным буфером. Сначала проводили скрининг колоний всех собранных плазмид путем рестрикционного расщепления минипрепаратов ДНК. Плазмидная ДНК отобранных клонов была секвенирована коммерческим поставщиков услуг секвенирования (Eurofins MWG Operon, Huntsville, AL). Данные по последовательностям были собраны и проанализированы с использованием программы Sequencher™ (Gene Codes Corp., Ann Arbor, MI).
Перед доставкой в протопласты Glycine max плазмидную ДНК получали из культур E. coli с использованием наборов Pure Yield Plasmid Maxiprep System® (Promega Corporation, Madison, WI) или Plasmid Maxi Kit® (Qiagen, Valencia, CA), следуя инструкциям поставщиков.
Полученные одиннадцать плазмидных конструктов: pDAB115603 (содержащий ZFN37354 и ZFN37355 конструкт с eHF-FokI), pDAB115600 (содержащий ZFN37354 и ZFN37355 конструкт с FokI дикого типа), pDAB115605 (содержащий ZFN37370 и ZFN37371 конструкт с eHF-FokI), pDAB115601 (содержащий ZFN37370 и ZFN37371 конструкт с FokI дикого типа), pDAB115606 (содержащий ZFN37374 и ZFN37375 конструкт с eHF-FokI), pDAB115604 (содержащий ZFN37366 и ZFN37367 конструкт с eHF-FokI), pDAB115607 (содержащий ZFN37384 и ZFN37385 конструкт с eHF-FokI), pDAB115602 (содержащий ZFN37384 и ZFN37385 конструкт с FokI дикого типа), pDAB115609 (содержащий ZFN37398 и ZFN37399 конструкт с eHF-FokI) и pDAB115608 (содержащий ZFN37392 и ZFN37393 конструкт с eHF-FokI) подтверждали расщеплением ферментами рестрикции и секвенированием ДНК.
Конструкты доноров
Векторы доноров для FAD2 конструировали путем объединения синтезированных de novo линейных фрагментов в многокопийном плазмидном векторе. pDAB115620 (фигура 4) и pDAB115622 (фигура 5) использовали для интегрирования доноров в локусы FAD2 генома сои. Оба вектора доноров были синтезированы для содержания доменов связывания нуклеазы с «цинковыми пальцами». pDAB115620 («донор 1») содержит домен связывания ZFN 37354:37355, домен связывания ZFN 37366:37367, домен связывания ZFN 37370:37371 и домен связывания ZFN 37374:37375. pDAB115622 («донор 2») содержит домен связывания ZFN 37384:37385, домен связывания ZFN 37392:37393 и домен связывания ZFN 37398:37399. Домены связывания ZFN узнаются соответствующей экспрессированной нуклеазой с «цинковыми пальцами» и расщепляются в процессе совместной трансформации клеток растения вектором донора и вектором нуклеазы с «цинковыми пальцами».
Пример 4: Трансформация протопластов сои
Был разработан метод трансформации протопластов сои (например, Glycine max культивар Maverick). Протопласты выделяли из суспензионной культуры Maverick, полученной из каллюса, полученного из листовых эксплантатов. Ниже приведена соответствующая методика.
Культивирование
Суспензии клеток сои субкультивировали каждые 7 дней путем разведения 1:5 в свежей среде LS (Linsmaier and Skoog, 1965), содержащей 3% (масс/об) сахарозы, 0,5 мг/л 2,4-D и 7 г бактоагара, pH 5,7. Все эксперименты проводили, начиная с 7 дня после субкультивирования, используя протокол, приведенный ниже.
Выделение протопластов
Тридцать миллилитров суспензионной культуры Maverick через 7 дней после субкультивирования переносили в 50-мл коническую центрифужную пробирку и центрифугировали при 200 g в течение 3 минут, получая примерно 10-мл объем осажденных клеток (SCV) на пробирку. Супернатант удаляли, не затрагивая осадок клеток. Двадцать миллилитров раствора ферментов (0,3% пектолиазы (320952, MP Biomedicals), 3% целлюлазы («Onozuka» R10™, Yakult Pharmaceuticals, Japan)) в растворе MMG (4 мМ MES, 0,6 M маннит, 15 мМ MgCl2, pH 6,0) добавляли на каждые 4 SCV суспензионных клеток и пробирки оборачивали парафильмом™. Пробирки помещали на платформу шейкера на ночь (примерно 16-18 часов). На следующее утро аликвоту обработанных клеток рассматривали под микроскопом, чтобы убедиться, что расщепление клеточных стенок было достаточным.
Очистка протопластов
Растворы, содержащие клетки/ферменты, медленно фильтровали через 100-мкм клеточное сито. Клеточное сито промывали 10 мл среды W5+ (1,82 мМ MES, 192 мМ NaCl, 154 мМ CaCl2, 4,7 мМ KCl, pH 6,0). Этап фильтрования повторяли с использованием 70-мкм сита. Конечный объем доводили до 40 мл добавлением 10 мл среды W5+. Клетки перемешивали, переворачивая пробирку. Протопласты медленно наслаивали на 8 мл плотного раствора сахарозы (500 мМ сахароза, 1 мМ CaCl2, 5 мМ MES-KOH, pH 6,0), добавляя плотный раствор на дно 50-мл конической центрифужной пробирки, содержащей клетки. Пробирки центрифугировали при 350 g в течение 15 минут в бакет-роторе. Наконечник 5-мл пипетки использовали, чтобы медленно удалить содержащую протопласты полосу (примерно 7-8 мл). Затем протопласты переносили в 50-мл коническую пробирку и добавляли 25 мл промывочной среды W5+. Пробирки медленно переворачивали и центрифугировали в течение 10 минут при 200 g. Супернатант удаляли, добавляли 10 мл раствора MMG и пробирку медленно переворачивали для ресуспендирования протопластов. Плотность протопластов определяли при помощи гемоцитометра или проточного цитометра. Как правило, из 4 PCV суспензии клеток получали примерно 2 миллиона протопластов.
Трансформация протопластов с помощью ПЭГ
Концентрацию протопластов доводили до 1,6 миллионов/мл, используя MMG. Аликвоты по 300 мкл протопластов (примерно 500000 протопластов) переносили в 2-мл стерильные пробирки. Суспензию протопластов регулярно перемешивали при перенесении протопластов в пробирки. Плазмидную ДНК добавляли к аликвотам протопластов в соответствии с дизайном эксперимента. Штатив с пробирками, содержащими протопласты, медленно переворачивали 3 раза, каждый раз по 1 минуте, для смешивания ДНК и протопластов. Протопласты инкубировали в течение 5 минут при комнатной температуре. Триста микролитров раствора полиэтиленгликоля (ПЭГ 4000) (40% этиленгликоля (81240-Sigma Aldrich), 0,3 M маннит, 0,4 M CaCl2) добавляли к протопластам и штатив с пробирками переворачивали для перемешивания в течение 1 мин и инкубировали в течение 5 мин, осторожно переворачивая дважды за время инкубации. Один миллилитр W5+ медленно добавляли в пробирки и штатив с пробирками переворачивали 15-20 раз. Затем пробирки центрифугировали при 350 g в течение 5 мин и супернатант удаляли, не затрагивая осадок. Один миллилитр среды WI (4 мМ MES, 0,6 M маннит, 20 мМ KCl, pH 6,0) добавляли в каждую пробирку, и штатив осторожно переворачивали для ресуспендирования осадков. Штатив накрывали алюминиевой фольгой и укладывали набок, оставляя для инкубации на ночь при 23°C.
Измерение частоты трансформации и сбор протопластов
Количественное определение протопластов и эффективности трансформации осуществляли с использованием проточного цитометра Quanta Flow Cytometer™ (Beckman-Coulter Inc). Примерно через 16-18 часов после трансформации отбирали по 100 мкл из каждой параллельной пробы, помещали в лунки 96-луночного планшета и разбавляли 1:1 раствором WI. Параллельные пробы ресуспендировали 3 раза и 100 мкл использовали для количественного определения методом проточной цитометрии. Перед использованием образцов для анализа их центрифугировали при 200 g в течение 5 мин, супернатанты удаляли и образцы быстро замораживали в жидком азоте. Затем образцы помещали в морозильную камеру при -80°C до обработки для молекулярного анализа.
Пример 5: Расщепление нуклеазой с «цинковыми пальцами» и интегрирование доноров
Сконструированные ZFNs вводили трансформацией в протопласты сои, используя описанную выше методику трансформации. Эффективность расщепления в отношении локуса FAD2 оценивали для различных ZFNs при помощи анализа на разрушение локуса, описанного в предварительной патентной заявке США №61/736856. Кроме того, опосредованное нуклеазой с «цинковыми пальцами» интегрирование донорской последовательности в локусы FAD2 оценивали в анализе «внутренней-внешней» ПЦР и полученные ПЦР-ампликоны секвенировали для характеризации интегрирования доноров в геном сои.
Эксперименты включали группы обработки с содержанием только вектора донора, только вектора ZFN или сочетания векторов ZFN и донора (таблица 3). Кроме того, эксперименты включали в качестве отрицательного контроля группы обработки с нетрансформированными клетками или клетками, трансформированными контрольным вектором, pDAB7221 (фигура 6), содержащим экспрессионную кассету GFP, управляемую промотором CsVMV и фланкированную 3'-UTR AtuORF24 в многокопийной плазмиде. Трансформированные образцы собирали примерно через 18-24 часов после трансфекции. Экспериментальные данные показали высокую активность плазмиды ZFN, pDAB115601, и эта плазмида ZFN была использована в качестве положительного контроля во всех последующих экспериментах.
Как указано в таблице 3, в экспериментах по трансформации было задействовано в общей сложности 80 мкг ДНК, при этом в случае необходимости добавляли плазмиду pDAB7221 для доведения общего количества ДНК до 80 мкг. Отношение вектора донора к ZFN-экспрессирующей плазмиде составляло примерно 10:1. Для каждого эксперимента или вида обработки использовали по шесть экспериментальных параллельных проб, которые обрабатывали и анализировали независимо. Оценку ZFNs проводили в двух сериях экспериментов, при этом плазмиду ZFN, pDAB115601, использовали во всех конечных экспериментах в качестве положительного контроля.
Дизайн эксперимента. Плазмиды ZFN оценивали в двух наборах (F2 ZFNs 1-3 и F2 ZFNs 4-7). Векторы доноров, подходящие для протестированных плазмид ZFN, использовали в экспериментах по направлению на мишень. Для каждого вида обработки использовали по шесть параллельных проб
Анализ направления на мишень
Образцы ДНК из экспериментов по направлению на мишень анализировали с использованием анализа разрушения локуса для обнаружения модификаций в сайтах расщепления ZFN в FAD2 или оценки направления на мишень с помощью NHEJ. Был разработан анализ кПЦР для количественного определения интактных сайтов связывания ZFN в мишенях FAD2. Опосредованная ZFN вставка донора или расщепление с последующей репарацией NHEJ приводит к утрате сайта связывания ZFN и последующему уменьшению обнаруживаемого сигнала кПЦР. ZFNs, обладающие значительной расщепляющей активностью, приводят к образованию ампликонов с пониженным сигналом по сравнению с тем, что происходит при обработке только донором. Праймеры и зонды, используемые в анализе разрушения локуса, приведены в таблице 4, и их относительные положения на локусах FAD2 показаны на фигуре 7.
Результаты сравнивали с сигналом, полученным от интактных локусов FAD2 в необработанных клетках сои. Обработка протопластов FAD2 2.3 ZFN2_WT ZFN (оба эксперимента) и FAD2 2.6 ZFNs ZFN4_HF (один эксперимент), а также F2 ZFN5_HF (оба эксперимента) в присутствии соответствующих векторов доноров приводила к статистически значимому снижению сигнала по сравнению с тем, который получен в случае интактной последовательности (только донор).
Праймеры и зонды для ПЦР в анализе разрушения локуса
Локус-специфическая «внутренняя-внешняя» ПЦР
Для подтверждения направленной вставки донора ДНК из всех вариантов обработки подвергали анализу локус-специфической «внутренней-внешней» ПЦР. В экспериментах был разработан вектор донора для содержания сайтов связывания для всех ZFNs, которые были протестированы для направленного интегрирования в локус FAD2. Совместная доставка ZFN и донора в клетки сои приводит к расщеплению сайта связывания ZFN в мишени и в векторе донора и последующему интегрированию донора в расщепленный локус FAD2 по механизму негомологичного соединения концов. Концы хромосомного сайта FAD2 и линеаризованного вектора донора, образованные в результате расщепления ZFN, подвергаются процессингу перед интегрированием в локус FAD2, и это может приводить к несовершенным продуктам соединения концов. Подтверждение направленного интегрирования в мишень выполняли на основе стратегии «внутренней-внешней» ПЦР, когда «внешний» праймер узнает последовательность в природном геномном локусе и «внутренний» праймер связывается с последовательностью внутри ДНК донора. Анализ «внутренней-внешней» ПЦР проводили как с 5'-, так и с 3'-конца от мест соединения вставки.
Все протестированные ZFNs продемонстрировали некоторые признаки направления на мишень и интегрирования донорского фрагмента в локус FAD2 сои по меньшей мере в одном эксперименте, что было определено по ПЦР-продукту в образцах донора и ZFN. Результаты направления на мишень с интегрированием донора при использовании следующих ZFNs: F2 ZFN2_WT, F2 ZFN2_HF и F2 ZFN4_HF были воспроизводимы, поскольку ПЦР-продукты образовывались в по меньшей мере 2 из 6 экспериментальных параллельных проб как на 5'-, так и на 3'-концах (таблица 5).
Результаты направления на мишень с помощью NHEJ в локусе FAD2 в протопластах сои. Число параллельных проб, положительных по результатам «внутренней-внешней» ПЦР в независимых экспериментах по направлению на мишень, приведено для экспериментов или вариантов обработки
Секвенирование продуктов «внутренней-внешней» ПЦР
Два из ампликонов (ожидаемого размера) из каждого из экспериментов по направлению на мишень с последующей «внутренней-внешней» ПЦР, проводимых с pDAB1115620 и F2 ZFN2_WT или pDAB1115620 и F2 ZFN2_HF, были клонированы в плазмиду. Полученную плазмиду секвенировали методом секвенирования по Сэнгеру. Последовательности выравнивали с референсной последовательностью, в которой одноцепочечные 4-п.н. концы, которые предположительно возникали в результате расщепления FokI, были продублированы для представления всех возможных комбинаций концов. Десять уникальных паттернов последовательностей были обнаружены для 23 полученных клонированных последовательностей (фигура 8). Во всех паттернах последовательностей сохранялась часть геномной референсной последовательности FAD2, расположенная между сайтами связывания ZFN (GAAATTTC), однако в паттернах последовательностей также имели место делеции относительно геномной референсной последовательности FAD2. Последовательности 4WT1 и 4WT4 имели делеции, которые продолжались в сайт связывания ZFN на 3'-конце последовательности GAAATTTC. Две последовательности, 1HF4 и 6HF4, имели вставки в одно основание. Наблюдаемые паттерны последовательностей ДНК продемонстрировали, что имело место направление донорской ДНК в локус FAD2 сои.
При том, что некоторые иллюстративные варианты осуществления были описаны в настоящем документе, специалистам в данной области будет понятно и очевидно, что множество добавлений, изъятий и модификаций иллюстративных вариантов осуществления можно использовать без отклонения от объема нижеследующей формулы изобретения. Кроме того, признаки из одного варианта осуществления можно комбинировать с признаками из другого варианта осуществления.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛОКУСЫ ФУНКЦИОНАЛЬНОСТИ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ К УЧАСТКУ-МИШЕНИ СВЯЗЫВАЮЩИЕ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656159C2 |
| ЛОКУСЫ FAD3 ДЛЯ ВЫПОЛНЕНИЯ ОПЕРАЦИЙ И СООТВЕТСТВУЮЩИЕ СВЯЗЫВАЮЩИЕСЯ СО СПЕЦИФИЧЕСКИМИ САЙТАМИ-МИШЕНЯМИ БЕЛКИ, СПОСОБНЫЕ К ВЫЗОВУ НАПРАВЛЕННЫХ РАЗРЫВОВ | 2013 |
|
RU2665811C2 |
| СКОНСТРУИРОВАННАЯ СПОСОБАМИ ИНЖЕНЕРИИ ПЛАТФОРМА ДЛЯ ВСТРАИВАНИЯ ТРАНСГЕНА (ETIP) ДЛЯ НАЦЕЛИВАНИЯ ГЕНОВ И СТЭКИНГА ПРИЗНАКОВ | 2013 |
|
RU2666916C2 |
| БЫСТРЫЙ НАПРАВЛЕННЫЙ АНАЛИЗ СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР ДЛЯ ОПРЕДЕЛЕНИЯ ДОНОРНОЙ ВСТАВКИ | 2014 |
|
RU2668819C2 |
| НАПРАВЛЕННАЯ МОДИФИКАЦИЯ МАЛАТДЕГИДРОГЕНАЗЫ | 2013 |
|
RU2658437C2 |
| ОБОГАЩЕНИЕ АКТИВИРУЕМОЙ ФЛУОРЕСЦЕНЦИЕЙ СОРТИРОВКИ КЛЕТОК (FACS) ДЛЯ СОЗДАНИЯ РАСТЕНИЙ | 2013 |
|
RU2679510C2 |
| УНИВЕРСАЛЬНАЯ ДОНОРНАЯ СИСТЕМА ДЛЯ НАПРАВЛЕННОГО ВОЗДЕЙСТВИЯ НА ГЕНЫ | 2014 |
|
RU2687369C2 |
| СПОСОБЫ ОБНАРУЖЕНИЯ ДНК ДЛЯ САЙТ-СПЕЦИФИЧЕСКОЙ НУКЛЕАЗНОЙ АКТИВНОСТИ | 2013 |
|
RU2678001C2 |
| ОПТИМАЛЬНЫЕ ЛОКУСЫ СОИ | 2014 |
|
RU2719150C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ОПОСРЕДОВАННОЙ НУКЛЕАЗОЙ НАПРАВЛЕННОЙ ИНТЕГРАЦИИ ТРАНСГЕНОВ | 2013 |
|
RU2650819C2 |




































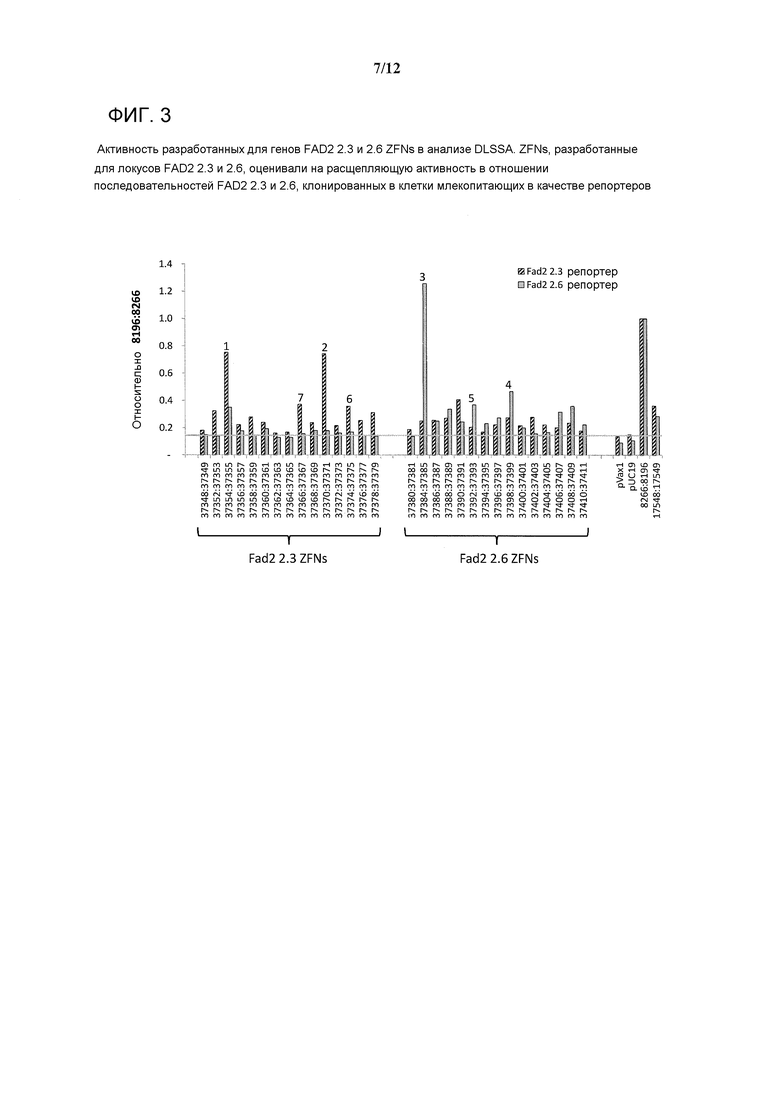
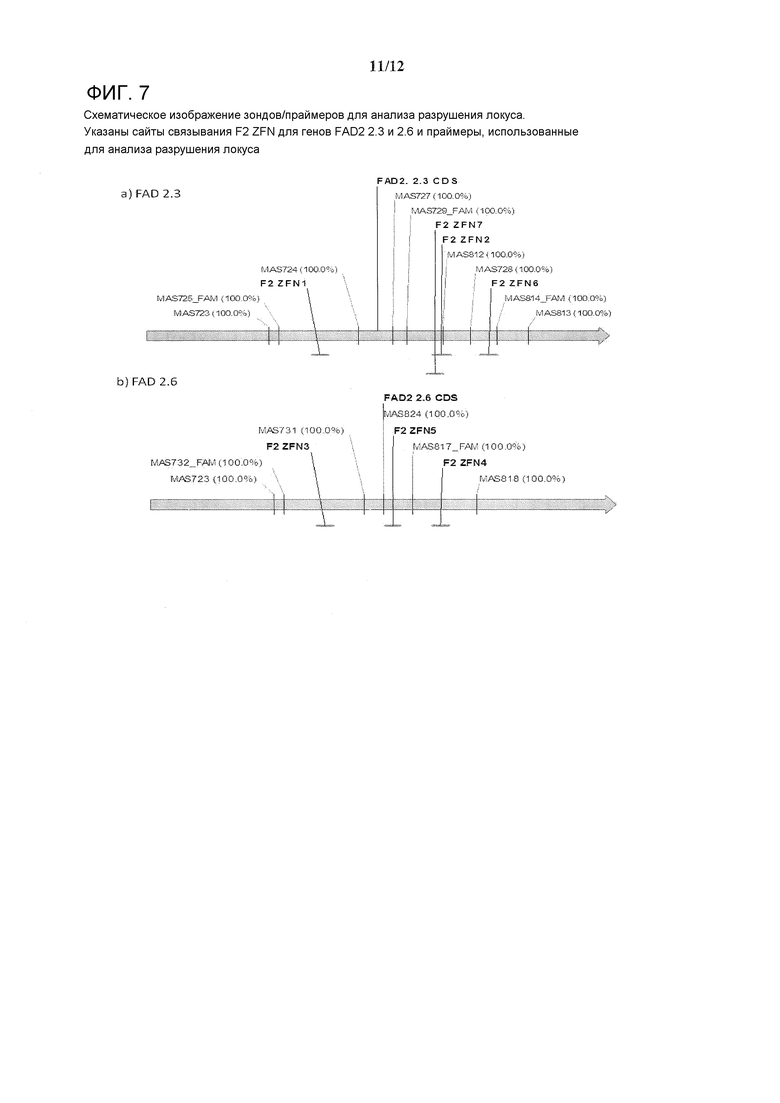
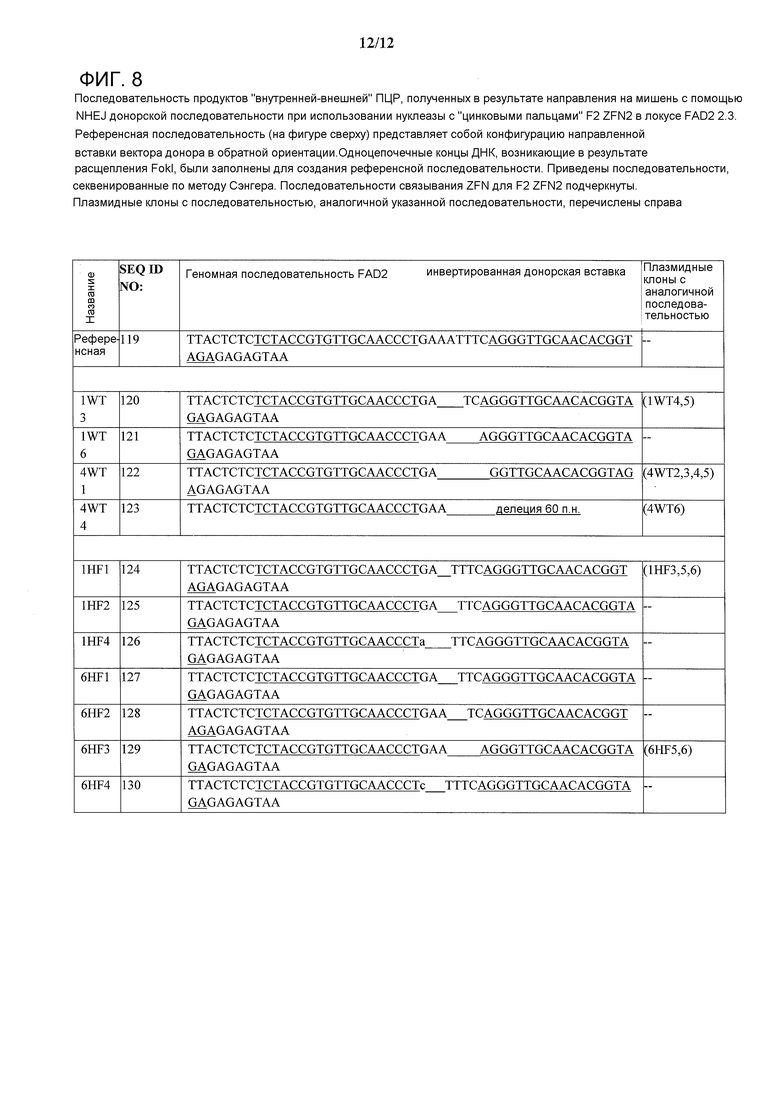
Изобретение относится к области биохимии, в частности к способу интегрирования интересующей последовательности нуклеиновой кислоты в ген FAD2 в клетку сои, включающему расщепление сайт-специфическим образом гена FAD2 клетки сои с использованием нуклеазы с «цинковыми пальцами». Также раскрыта сайт-специфическая нуклеаза с «цинковыми пальцами» для использования с целью модификации гена FAD2. Изобретение позволяет эффективно интегрировать интересующую последовательность нуклеиновой кислоты в клетку растения сои. 2 н. и 4 з.п. ф-лы, 8 ил., 5 табл., 5 пр.
1. Способ интегрирования интересующей последовательности нуклеиновой кислоты в ген FAD2 в клетке сои, включающий:
расщепление сайт-специфическим образом гена FAD2 клетки сои с использованием нуклеазы с «цинковыми пальцами», содержащей от трех до шести доменов «цинковых пальцев», где каждый домен «цинковый палец» содержит спиральную область узнавания,
где
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 24, 25, 26, 27, 28, 29, и расщепляет участок-мишень SEQ ID NO: 14 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 30, 31, 32, 33, 34, и расщепляет участок-мишень SEQ ID NO: 14 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 35, 33, 36, 37, 38, 39, и расщепляет участок-мишень SEQ ID NO: 15 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 40, 41, 24, 42, 33, 43, и расщепляет участок-мишень SEQ ID NO: 15 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 44, 45, 46, 47, 48, 49, и расщепляет участок-мишень SEQ ID NO: 16 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 50, 51, 52, 53, 54, 55, и расщепляет участок-мишень SEQ ID NO: 16 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 56, 57, 58, 59, 60, 61, и расщепляет участок-мишень SEQ ID NO: 17 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 62, 63, 64, 65, 36, 66, и расщепляет участок-мишень SEQ ID NO: 17 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 24, 25, 26, 27, 67, 68, и расщепляет участок-мишень SEQ ID NO: 18 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 30, 31, 32, 33, 69, и расщепляет участок-мишень SEQ ID NO: 18 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 56, 57, 58, 58, 36, 70, и расщепляет участок-мишень SEQ ID NO: 19 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 62, 63, 40, 71, 36, 66, и расщепляет участок-мишень SEQ ID NO: 19 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 72, 73, 30, 74, 75, и расщепляет участок-мишень SEQ ID NO: 20 или вблизи него; или
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 24, 76, 77, 78, 79, и расщепляет участок-мишень SEQ ID NO: 20 или вблизи него;
с созданием, тем самым, разрыва в гене FAD2, и
интегрирование в указанный разрыв интересующей последовательности нуклеиновой кислоты, присутствующей в клетке.
2. Способ по п.1, в котором ген FAD2 представляет собой ген FAD2 2.3, FAD2 2.6 или оба.
3. Способ по п.1 или 2, в котором расщепление сайт-специфическим образом является специфическим для некоторых, но не для всех копий FAD2 2.3 и FAD2 2.6.
4. Способ по любому из пп. 1-3, в котором расщепление сайт-специфическим образом является специфическим для некоторых, но не для всех копий FAD2 2.3, или некоторых, но не для всех копий FAD2 2.6.
5. Способ по любому из пп. 1-4, в котором интересующая последовательность нуклеиновой кислоты выбрана из группы, состоящей из последовательности, содержащей сайт-мишень связывания ДНК-связывающего домена, одного или более генов устойчивости к инсектицидам, одного или более генов толерантности к гербицидам, одного или более генов, определяющих эффективность использования азота, одного или более генов, определяющих эффективность использования воды, одного или более генов, определяющих пищевые свойства, одного или более ДНК-связывающих генов, одного или более генов селективных маркеров, а также их сочетаний.
6. Сайт-специфическая нуклеаза с «цинковыми пальцами» для использования с целью модификации гена FAD2, где указанная нуклеаза с «цинковыми пальцами» содержит от трех до шести доменов «цинковых пальцев», где каждый домен «цинковый палец» содержит спиральную область узнавания,
где
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 24, 25, 26, 27, 28, 29, и расщепляет участок-мишень SEQ ID NO: 14 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 30, 31, 32, 33, 34, и расщепляет участок-мишень SEQ ID NO: 14 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 35, 33, 36, 37, 38, 39, и расщепляет участок-мишень SEQ ID NO: 15 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 40, 41, 24, 42, 33, 43, и расщепляет участок-мишень SEQ ID NO: 15 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 44, 45, 46, 47, 48, 49, и расщепляет участок-мишень SEQ ID NO: 16 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 50, 51, 52, 53, 54, 55, и расщепляет участок-мишень SEQ ID NO: 16 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 56, 57, 58, 59, 60, 61, и расщепляет участок-мишень SEQ ID NO: 17 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 62, 63, 64, 65, 36, 66, и расщепляет участок-мишень SEQ ID NO: 17 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 24, 25, 26, 27, 67, 68, и расщепляет участок-мишень SEQ ID NO: 18 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 30, 31, 32, 33, 69, и расщепляет участок-мишень SEQ ID NO: 18 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 56, 57, 58, 58, 36, 70, и расщепляет участок-мишень SEQ ID NO: 19 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 62, 63, 40, 71, 36, 66, и расщепляет участок-мишень SEQ ID NO: 19 или вблизи него;
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 72, 73, 30, 74, 75, и расщепляет участок-мишень SEQ ID NO: 20 или вблизи него; или
указанная нуклеаза с «цинковыми пальцами» содержит распознающие спиральные области в следующем порядке, представленные в SEQ ID NO: 24, 76, 77, 78, 79, и расщепляет участок-мишень SEQ ID NO: 20 или вблизи него.

