Предмет изобретения
Настоящее изобретение относится к новым ферментам, которые переносят сиаловые кислоты от донорной молекулы (например, олигосахаридов, полисиаловых кислот, гликозилированных белков, гликозилированных пептидов, гликозилированных липидов (например, ганглиозидов) и других гликозилированных низкомолекулярных и высокомолекулярных молекул) на акцепторную молекулу (например, олиго- и полисахариды, гликозилированные белки, гликозилированные пептиды, гликозилированные липиды и другие гликозилированные низкомолекулярные и высокомолекулярные молекулы) (транс-сиалидазам). Эти ферменты были выделены из одноклеточного организма Trypanosoma congolense.
Кроме того, изобретение относится к функциональным эквивалентам этих ферментов; кодирующим эти ферменты и их функциональные эквиваленты последовательностям нуклеиновых кислот; экспрессионным конструкциям и векторам, которые содержат эти последовательности; рекомбинантным микроорганизмам, которые несут кодирующую последовательность нуклеиновой кислоты согласно изобретению; способам рекомбинантного получения ферментов по изобретению; способам выделения ферментов по изобретению из Trypanosoma congolense; способам ферментативного сиалилирования акцепторных молекул с применением ферментов по изобретению; эффекторам транс-сиалидаз по изобретению; применению последовательностей нуклеиновых кислот, ферментов, эффекторов или продуктов сиалилирования по изобретению для производства вакцин, медикаментов, пищевых продуктов или пищевых добавок; а также к самим полученным продуктам по изобретению.
Уровень техники
Транс-сиалидазы могут переносить сиаловые кислоты, предпочтительно альфа-2,3-связанные сиаловые кислоты, от донорной молекулы на акцепторную молекулу, при этом снова образуются альфа-2,3-гликозидные связи, предпочтительно на β-концевом галактозном остатке.
Под термином сиаловые кислоты понимают все N- и О-производные нейраминовой кислоты (Blix et al., 1957). Нейраминовая кислота (5-амино-3,5-дидезокси-D-глицеро-D-галакто-нонуло-пираносон-кислота) является аминосахаром со скелетом из девяти атомов углерода, который благодаря карбоксильной группе при втором С-атоме приобретает очень кислый рК-показатель 2,2 и поэтому является отрицательно заряженным при физиологических условиях. Незамещенная форма является очень нестабильной и не встречается в природе в свободной форме (Schauer, 1982). Правда, теперь стали известны более 40 природных производных нейраминовой кислоты (Schauer und Kamerling, 1997). Двумя наиболее часто встречающимися в природе сиаловыми кислотами являются N-ацетилнейраминовая кислота (Neu5Ac), предшественник всех гликозидно связанных сиаловых кислот (Schauer, 1991), и N-гликолилнейраминовая кислота (Neu5Gc), которая возникает в результате гидроксилирования метильной группы N-ацетильного остатка СМР-Neu5Ac (Shaw und Schauer, 1988). Гидроксильные группы этих обеих сиаловых кислот могут быть заменены ацетильными, лактильными, сульфатными и фосфатными остатками в различной комбинации, что приводит к большому структурному разнообразию сиаловых кислот (Schauer, 1991; Schauer und Kamerling, 1997).
Наибольшая часть природно встречающихся сиаловых кислот существует в связанном виде в виде компонента олигосахаридов, полисахаридов и особенно гликоконъюгатов (Schauer, 1982). Но уже известны также полисиаловые кислоты из трансгенного микробного производства. Сиалилированные гликоконъюгаты встречаются прежде всего в наружной мембране клеток, но являются также важными компонентами сыворотки и мукозной слизи (Traving und Schauer, 1998). Сиаловые кислоты защищают гликопротеины и клетки от атаки протеазами и другими ферментами и, следовательно, от расщепления (Reuter et al., 1988). Содержащие сиаловые кислоты слизистые оболочки желудочно-кишечного тракта образуют не только защиту от пищеварительных ферментов, но также защищают лежащие ниже ткани от проникновения патогенных бактерий (Keim und Schauer, 1997).
Очень важную функцию выполняют сиаловые кислоты при молекулярных и клеточных процессах распознавания. При этом они маскируют рецепторы и таким образом препятствуют взаимодействиям между рецепторами и лигандами (Schauer, 1985; Kelm und Schauer, 1997). Так, например, сиаловые кислоты защищают гликопротеины сыворотки и эритроциты от расщепления и фагоцитоза, маскируя лежащие под ними галактозные остатки. При отщеплении концевых сиаловых кислот субтерминальные галактозные остатки могут связываться лектинами на гепатоцитах или фагоцитах, и происходит эндоцитоз сывороточных белков или эритроцитов. Следующим примером является защита гомологичных тканей, а также многих высокосиалилированных опухолей от узнавания при помощи иммунной системы (Pilatte et al., 1993). В случае потери защищающего слоя сиаловых кислот могут произойти аутоиммунные реакции.
Сиаловые кислоты служат также в качестве сайтов узнавания для гомологичных клеток и гормонов и, следовательно, играют важную роль при клеточных взаимодействиях (Kelm und Schauer, 1997). При воспалениях, например, эндотелиальные клетки экспрессируют на их поверхности селектины, которые узнают определенные сиалилированные структуры (например, сиалил-Льюис Х) на лейкоцитах, так что они связываются с эндотелиальными клетками и могут проникать в ткань (Lasky, 1995). Кроме того, действие транс-сиалидаз влияет на активацию Т-клеток гуморальной иммунной защиты (Gao et al., 2001). Сиалоадгезины (Siglecs), такие как миелин-ассоциированный гликопротеин (MAG), также связываются с высокой специфичностью с сиалилированными гликанами (Kelm et al., 1996; Crocker et al., 1998). Миелин-ассоциированный гликопротеин участвует в нервной системе, в частности, в миелинизации и в регуляции роста аксонов. Таким образом, неудивительно, что недавно было установлено, что транс-сиалидазы посредством переноса сиаловых кислот участвуют в дифференцировке нервных клеток и глиальных клеток (Chuenkova et al., 2001). CD-22 является следующим связывающим сиаловую кислоту рецептором, который встречается на лимфоцитах и делает возможным «диалог» Т- и В-лимфоцитов. Семейство Siglecs состоит в среднем из более чем 10 молекулярно-биологически охарактеризованных предшественников.
Однако сиаловые кислоты важны не только при гомологичных процессах распознавания, но представляют собой также рецепторы для некоторых бактерий, вирусов и токсинов. Так, например, происходит связывание столбнячного токсина с ганглиозидами нервных синапсов через сиаловые кислоты (Schauer et al., 1995). Специфическая в отношении сиаловых кислот адгезия через микробные лектины (Sharon und Lis, 1997) часто является решающей стадией при инфекционных заболеваниях, например, при вызываемом некоторыми штаммами E. coli менингите новорожденных или при инфекциях слизистой оболочки желудка Helicobacter pylori. Прежде всего возбудители гриппа А и В прикрепляются через сиаловую кислоту к клеткам, которые должны инфицироваться (Schauer, 2000).
Модификации сиаловых кислот, в частности, О-ацетилирование, имеют большое значение при регуляции молекулярного и клеточного распознавания (Schauer, 1991). Так, вирусы гриппа С связываются специфически с 9-О-ацетилированными сиаловыми кислотами на эпителии бронхов (Herrler et al., 1985), тогда как О-ацетилирование препятствует связыванию вирусов гриппа А и В (Higa et al., 1985). Но прежде всего О-ацетилирование сиаловых кислот является очень важным для морфогенеза и развития различных тканей (Varki et al., 1991). В случае нейроэктодермальных опухолей оно повышается (Hubl et al., 2000; Fahr und Schauer, 2001), а при раке толстой кишки понижается (Cornfield et al., 1999). Сиаловые кислоты являются незаменимыми модуляторами биологического поведения опухолей (Schauer, 2000).
Описание фигур
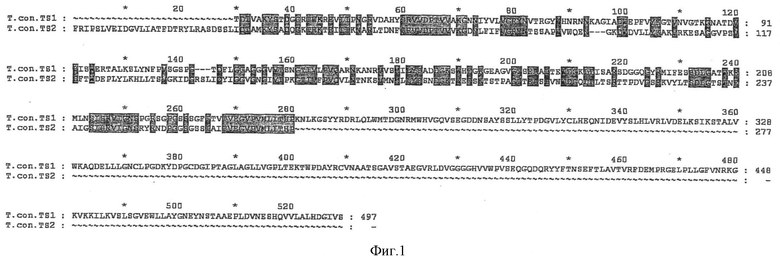
Фиг.1 показывает сравнение аминокислотных последовательностей транс-сиалидаз TS1 и TS2 по изобретению. Идентичные аминокислоты в обеих последовательностях отмчены в виде темных участков. Соответствие (идентичность) обеих частичных последовательностей составляет только приблизительно 50%.
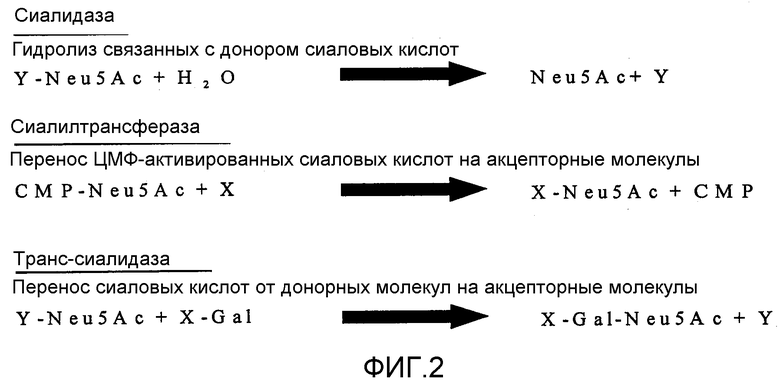
Фиг.2 показывает различные виды реакций сиалидазы, сиалилтрансфераз и транс-сиалидаз.
Фиг.3 показывает сравнение аминокислотной последовательности сиалидазы из Trypanosoma rangeli (T. r. S), транс-сиалидазы Trypanosoma cruzi (T. cr. TS) и транс-сиалидазы Trypanosoma brucei brucei (T. b. br. TS) с частичными последовательностями обеих транс-сиалидаз из Trypanosoma congolense (T. con. TS1 и T. con. TS2) по изобретению. Аминокислоты, которые во всех последовательностях являются идентичными, изображены белыми на темно-сером фоне. Аминокислоты, которые идентичны по меньшей мере в 4 из 5 последовательностей, напечатаны черным шрифтом на темно-сером фоне, тогда как аминокислоты, которые являются идентичными по меньшей мере в 3 из 5 последовательностей, представлены светло-серым цветом.
Сущность изобретения
Задачей изобретения была разработка нового средства, при помощи которого можно влиять на регулируемые сиаловой кислотой биологические или патобиологические процессы.
Вышеуказанную задачу удалось неожиданным образом решить с помощью новых ферментов с транс-сиалидазной активностью и их кодирующих последовательностей из Trypanosoma congolense.
Первый предмет изобретения относится к полинуклеотидам, которые кодируют белки с транс-сиалидазной активностью и могут быть выделены из Trypanosoma congolense, причем эти белки предпочтительно катализируют перенос сиаловой кислоты от донорной молекулы на акцепторную молекулу.
Предпочтительные полинуклеотиды включают в себя по меньшей мере одну последовательность нуклеиновой кислоты в соответствии с SEQ ID NO:1 или 3 или представляют их фрагменты, которые включают в себя по меньшей мере 15 связанных нуклеотидных остатков. Предметом изобретения являются также комплементарные им полинуклеотиды и фрагменты; и нуклеотидные последовательности, произведенные из этих полинуклеотидов вследствие вырожденности генетического кода.
Следующий предмет изобретения относится к олигонуклеотидам, которые гибридизуются с полинуклеотидом по изобретению, в частности, при строгих условиях.
Предметом изобретения являются также полинуклеотиды, которые гибридизуются с олигонуклеотидом в соответствии с приведенным выше определением, в частности, при строгих условиях и кодируют генный продукт из микроорганизмов рода Trypanosoma.
Предметом изобретения являются также полипептиды, которые кодируются полинуклеотидом, который включает в себя последовательность нуклеиновой кислоты в соответствии с приведенным выше определением; или которые обнаруживают аминокислотную последовательность, которая включает в себя по меньшей мере 10 связанных аминокислот в соответствии с SEQ ID NO:2 или 4; а также их функциональные эквиваленты, которые обладают транс-сиалидазной активностью.
Предметом изобретения являются, в частности, транс-сиалидазы или их функциональные эквиваленты с транс-сиалидазной активностью, отличающиеся следующими частичными аминокислотными последовательностями:
TDTVAKYSTDGGRTWKREVIIPNGR (положения 1-25 согласно SEQ ID NO:2)
FRIPSLVEIDGVLIATFDTRYLRASDSSLI (положения 1-30 согласно SEQ ID NO:4)
Предпочтительная транс-сиалидаза 1 (TS1) отличается по меньшей мере одной из следующих характеристик:
Другая предпочтительная транс-сиалидаза 2 (TS2) отличается по меньшей мере одной из следующих характеристик:
Вышеописанные полинуклеотиды и полипептиды по изобретению, в частности кодирующие последовательности нуклеиновых кислот и аминокислотные последовательности, происходят из организма Trypanosoma congolense. Однако они доступны также с использованием синтетических, в частности химических, биохимических, ферментативных, генотехнологических и трансгенных способов.
Кроме того, предметом изобретения являются функциональные эквиваленты транс-сиалидаз данного изобретения.
Кроме того, предметом изобретения являются экспрессионные кассеты, включающие в себя последовательность нуклеиновой кислоты в соответствии с вышеописанным определением в функциональной связи по меньшей мере с одной регуляторной последовательностью нуклеиновой кислоты. Кроме того, изобретение включает в себя также рекомбинантные векторы, содержащие по меньшей мере одну из этих экспрессионных кассет.
Кроме того, предметом изобретения являются прокариотические и эукариотические хозяева, трансформированные по меньшей мере одним вектором в соответствии с приведенным выше определением.
Кроме того, изобретение относится к применению экспрессионной кассеты, вектора или хозяина в соответствии с приведенным выше определением для рекомбинантного получения белка с транс-сиалидазной активностью.
Предметом изобретения является также способ ферментативного сиалилирования акцепторной молекулы, отличающийся тем, что акцепторную молекулу инкубируют с содержащим остатки сиаловой кислоты донором в присутствии транс-сиалидазы в соответствии с вышеописанным определением и выделяют сиалилированный акцептор.
Подобные способы отличаются по меньшей мере одним дополнительным свойством из следующих свойств:
а) донор выбран из связанных с олигонуклеотидами, полисахаридами, полисиаловыми кислотами, гликопротеинами и гликолипидами сиаловых кислот, таких как, в частности, лактоферрины, гликозилированные белки молока и казеины и их фрагменты;
b) акцептор выбран из содержащих β-галактозу полимеров, таких как β-галактоолигосахариды, лактит, лактобионовая кислота, метил-β-лактозид, ацетиллактозамины, галактопиранозиды, транс-галактоолигосахариды, полигалактоза и другие гликоконъюгаты со связанной на конце β(1-3)- или β(1-4)-галактозой; или галактозы.
Следующий аспект изобретения относится к применению транс-сиалидазы по изобретению, кодирующей ее последовательности нуклеиновой кислоты или полученного продукта сиалилирования по изобретению для получения лекарственного средства, пищевого продукта, пищевой добавки или пищевого компонента для предупреждения или лечения контролируемых сиаловой кислотой паразитарных, бактериальных или вирусных инфекций; для лечения опухолевых заболеваний; для лечения заболеваний, которые связаны с нарушением развития ткани; для лечения заболеваний иммунной системы; для лечения аутоиммунных реакций; для лечения заболеваний с нарушенной коммуникацией клеток; и/или для лечения воспалений.
В частности, предметом изобретения является применение транс-сиалидазы по изобретению в соответствии с вышеуказанным определением для развития вакцины против трипаносомоза или для развития ферментных ингибиторов для лечения или предупреждения вызываемых трипаносомами инфекций.
Кроме того, изобретение относится к применению транс-сиалидазы, кодирующей ее последовательности нуклеиновой кислоты или полученных в соответствии с изобретением сиалилированных продуктов для получения лекарственного средства, пищевой добавки или пищевого продукта для защиты гомологичных клеток или тканей или гликопротеинов от ферментативного воздействия.
Кроме того, предметом изобретения является применение транс-сиалидазы, кодирующей ее последовательности нуклеиновой кислоты или полученных в соответствии с изобретением сиалилированных продуктов для получения лекарственного средства, пищевой добавки или пищевого продукта для воздействия на развитие и/или морфогенез тканей организма.
Кроме того, изобретение относится к эффекторам транс-сиалидазной активности, выбранным из
а) полипептидов-лигандов, которые взаимодействуют с транс-сиалидазой в соответствии с вышеуказанным определением;
b) низкомолекулярных эффекторов, которые модулируют биологическую активность транс-сиалидазы в соответствии с вышеуказанным определением; и
с) антисмысловых последовательностей нуклеиновых кислот относительно последовательности нуклеиновой кислоты в соответствии с вышеуказанным определением.
Кроме того, изобретение относится к применению такого эффектора для получения лекарственного средства, пищевой добавки или пищевого продукта для лечения или профилактики ассоциированных с транс-сиалидазной активностью заболеваний.
Кроме того, предметом изобретения является способ выделения фермента с транс-сиалидазной активностью, в котором
а) Trypanosoma congolense культивируют в среде и
b) желаемый продукт выделяют ионообменной хроматографией с использованием градиента соли, в случае необходимости с последующими изоэлектрическим фокусированием, гель-фильтрацией, аффинной хроматографией и/или осаждением белка.
Наконец, изобретение относится к фармацевтическому или генотерапевтическому средству, содержащему в фармацевтическом или генотерапевтическом носителе по меньшей мере один эффектор в соответствии с вышеуказанным определением.
Подробное описание изобретения
i) Значение изобретения
Значение изобретения возникает из возможного в результате осуществления изобретения влияния на контролируемые сиаловой кислотой механизмы паразитарной, бактериальной и вирусной инфекции, влияния на клеточную коммуникацию и на иммунную систему и изменения механизмов регуляции и развития тканей человека и животных, а также опухолей. Это достигается нацеленным переносом сиаловых кислот на биологически релевантные гликоструктуры (гликаны, производные гликанов и гликоконъюгаты) с использованием описанных здесь транс-сиалидаз.
Из переноса сиаловых кислот на выбранные структуры-носители возникают, например, продукты для изменения воспалительных реакций, изменения клеточных взаимодействий в теле человека и животного, защита гомологичных тканей от атак собственной иммунной системы (аутоиммунных реакций), «перерождение» раковых клеток в организме пациента, посредством чего гомологичная иммунная система снова борется с ними (терапия рака и предупреждение рака), подавление проникновения патогенных бактерий в организм человека и животного, предупреждение и подавление вирусных инфекций слизистой оболочки желудка посредством Helicobacter pylori, подавление вызываемого бактериями и вирусами менингита новорожденных, превентивное и терапевтическое влияние на рецепторы эукариотических и прокариотических патогенных организмов, бактерий, вирусов и токсинов во избежание развертывания их действия в организме человека и животного, ингибирование связывания холерного токсина со слизистыми оболочками пищеварительного тракта человека и животных, развитие вакцин против трипаносомоза, развитие ферментных ингибиторов для подавления (терапии) инфекций Trypanosoma, влияние на молекулярные и клеточные процессы распознавания в организме человека и животного, защита гликопротеинов и клеток от атаки протеазами и другими ферментами, среди прочего также для защиты от расщепления молекул ферментами пищеварительного тракта человека и животных, влияние на развитие тканей и влияние на морфогенез тканей в организме.
Транс-сиалидазы по изобретению характеризуются следующими ДНК- и аминокислотными последовательностями, а также другими ДНК-последовательностями-гомологами, например, с более чем 60-процентным соответствием (идентичностью) относительно этих частичных последовательностей.
ii) Данные последовательностей относительно предпочтительных транс-сиалидаз
(1) Информации для последовательности фермента TS1:
Признаки ДНК частичной последовательности TS1:
Длина: 1491 п.н.
Тип: нуклеиновая кислота
Форма тяжа: двойной тяж
Происхождение: Trypanosoma congolense
ДНК-последовательность фермента TS1 (SEQ ID NO:1):

Аминокислотная последовательность фермента TS1 (SEQ ID NO:2):

(2) Информации для последовательности фермента TS2:
Признаки ДНК частичной последовательности TS1:
Длина: 831 п.н.
Тип: нуклеиновая кислота
Форма тяжа: двойной тяж
Происхождение: Trypanosoma congolense
ДНК-последовательность фермента TS2 (SEQ ID NO:3):

Аминокислотная последовательность фермента TS2 (SEQ ID NO:4):

Частичные последовательности аминокислот фермента TS1 и фермента TS2 имеют соответствие (идентичность) только приблизительно 50%. Поэтому эти частичные последовательности характеризуются однозначно как два различных вещества (см. фиг.1).
(iii) Описание свойств новых обнаруженных ферментов TS1 и TS2
а) Физико-химические свойства этих веществ
Основные данные обеих транс-сиалидаз TS1 и TS2
4 мМ Cu2+, Zn2+, Fe2+, Co2+: небольшое влияние
10 мМ N-ацетил-2,3-дидегидро-2-дезоксинейраминовая кислота: небольшое влияние
b) Биологические свойства этих веществ
Оба заявленных здесь вещества являются двумя ферментами, которые переносят сиаловые кислоты от донорной молекулы на акцепторную молекулу.
В качестве доноров в случае обоих ферментов, хорошими донорами для сиаловых кислот, которые могут переноситься этими ферментами, являются гликаны, например, олигосахариды, полисахариды, полисиаловые кислоты, гликопротеины и гликолипиды, связанные сиаловые кислоты. Среди гликопротеинов находятся, в частности, лактоферрины (из человека, коровы, козы, овцы, лошади, верблюда и других животных), гликозилированные белки молока (из человека, коровы, козы, овцы, лошади, верблюда и других животных), а также другие гликозилированные белки человеческого, животного и растительного происхождения, а также их части, такие как, например, гликомакропептид из казеинов этих животных. В качестве доноров могут использоваться также ганглиозиды.
Обе транс-сиалидазы имеют хорошую акцепторную специфичность в отношении галактоолигосахарида, в частности, в отношении бета-галактоолигосахарида, такого как, например, Vivinal GOS фирмы Borculo Domo Ingredients (BDI) и Oligomate 55 фирмы Yakult. Кроме того, в качестве акцепторов могут действовать лактит, лактобионовая кислота, метил-β-лактозид, ацетиллактозамины, галактопиранозиды, транс-галактоолигосахариды, полигалактозы и другие гликоконъюгаты с концевой β(1,3)- или β(1-4)-связанной галактозой. Метилирование галактозных остатков приводит к уменьшению акцепторной функции. Метилирование глюкозных остатков (например, у лактозы) имеет незначительное влияние на акцепторную функцию. Моносахарид галактоза также служит в качестве акцептора, хотя и с меньшей специфичностью.
Фермент TS1 обнаруживает вдвое более эффективный перенос сиаловых кислот на соответствующие акцепторы, чем фермент TS2. Субстраты могут быть сводобными, т.е. растворимыми, или также связанными с клеточной мембраной.
Известен также перенос альфа-2,3-связанных концевых сиаловых кислот на бета-1,4-связанные концевые галактозные остатки транс-сиалидазами Trypanosoma cruzi (Schenkman et al., 1991; Vandekerckhove et al., 1992; Scudder et al., 1993) и Trypanosoma brucei (Engstler et al., 1992, 1993, 1995). Однако TS1 и TS2 отличаются на основе различных ДНК- и аминокислотных последовательностей от этих уже известных ферментов. Таким образом, TS1 и TS2 характеризуются как однозначно новые вещества (транс-сиалидазы). Для дополнительного разграничения см. следующий параграф.
iv) Разграничение между настоящим изобретением и другими транс-сиалидазами, сиалидазами и сиалилтрансферазами
Фермент «транс-сиалидаза» был впервые описан в американском виде трипаносомы Trypanosoma cruzi (Schenkman et al., 1991). Немного позднее удалось обнаружить этот фермент также в американских видах Trypanosoma brucei gambiense, Trypanosoma brucei rhodesiense и Trypanosoma brucei brucei (Engstler et al., 1993, Pontes de Carvalho et al., 1993, Engstler et al., 1995). Кроме того, транс-сиалидазу детектировали в видах Endothypanum (паразитах, которые поражают ленивца) (Medina-Acosta et al., 1994), в Corinebacterium diphtheriae (Mattos-Guaraldi et al., 1998) и в плазме человека (Tertov et al., 2001). Уже задолго до обнаружения транс-сиалидаз были известны так называемые сиалидазы. Они являются гликогидралазами, которые переносят сиаловые кислоты от донорной молекулы исключительно на воду, эти сиаловые кислоты образуются также при гидролизе олигосахаридов и гликоконъюгатов.
Кроме того, определенные ферменты могут переносить активированные цитидинмонофосфатом (СМР) сиаловые кислоты на другие сахарные остатки, в основном на галактозу и N-ацетилгалактозамин. Эти ферменты называют сиалилтрансферазами (см. фиг.2).
Заявленные здесь транс-сиалидазы переносят сиаловые кислоты не исключительно от донорной молекулы на воду, как это делают просто сиалидазы. Однако, если подходящий акцептор отсутствует, заявленные здесь транс-сиалидазы гидролизуют сиаловые кислоты так же, как и простые сиалидазы. Заявленные здесь транс-сиалидазы не нуждаются также в активированных сиаловых кислотах для их реакции переноса, как ранее упомянутые сиалилтрансферазы. Транс-сиалидазы имеют также более широкую донорную и акцепторную специфичность, чем сиалилтрансферазы, и поэтому они являются особенно многосторонне применимыми. Таким образом, заявленные здесь транс-сиалидазы являются более предпочтительными для промышленного применения, чем просто сиалидазы и сиалилтрансферазы.
До сих пор известны только ДНК- и аминокислотные последовательности транс-сиалидаз Trypanosoma cruzi и Trypanosoma brucei brucei, а также ДНК- и аминокислотная последовательность простой сиалидазы из Trypanosoma rangeli. Заявленный здесь фермент TS1 имеет соответствие (идентичность) относительно соответствующей аминокислотной последовательности транс-сиалидазы из Trypanosoma brucei brucei ниже 60% и соответствие ниже 50% относительно соответствующей частичной последовательности Trypanosoma cruzi. Заявленный здесь фермент TS2 имеет соответствие (идентичность) относительно соответствующей аминокислотной последовательности транс-сиалидазы из Trypanosoma brucei brucei ниже 50% и соответствие ниже 50% относительно соответствующей частичной последовательности Trypanosoma cruzi (см. фиг.3). Далее, известно, что соответствие аминокислот между транс-сиалидазами трипаносом и известными сиалидазами и транс-сиалидазами бактерий и вирусов составляет только 20%-30% (Chuenkova et al., 1999, Montagna et al., 2002).
В случае описанных здесь ферментов речь идет, следовательно, о новых охарактеризованных веществах (ферментах), соответствие (идентичность) которых относительно соответствующих ДНК- и аминокислотных последовательностей других известных ферментов со сходной функцией составляет менее 60%.
v) Дополнительные пояснения в отношении настоящего изобретения
а) Полипептиды и функциональные эквиваленты
«Полипептиды» в контексте изобретения включают в себя характерные частичные фрагменты аминокислотных последовательностей по изобретению, а также аминокислотные последовательности ферментов по изобретению и их функциональные эквиваленты.
Таким образом, в соответствии с изобретением “функциональные эквиваленты” или “гомологи” включают в себя также конкретно раскрытые новые полипептиды или ферменты.
“Функциональные эквиваленты” или аналоги конкретно раскрытых полипептидов являются в пределах изобретения отличающимися от них полипептидами, которые, кроме того, обладают желаемой биологической активностью в соответствии с вышеуказанным определением (например, субстратной специфичностью).
Под “функциональными эквивалентами” в соответствии с изобретением понимают, в частности, мутанты, которые по меньшей мере в одном из вышеупомянутых положений последовательности обнаруживают другую аминокислоту, чем конкретно названная аминокислота, но, несмотря на это, обладают указанной здесь биологической активностью. “Функциональные эквиваленты” включают в себя, следовательно, мутанты, полученные посредством одного или нескольких добавлений, одной или нескольких замен, делеций и/или инверсий аминокислот, причем вышеупомянутые изменения могут обнаруживаться в любом положении последовательности, пока они приводят к мутанту с профилем свойств в соответствии с изобретением. Функциональная эквивалентность имеется, в частности, также в том случае, если картина реактивности между мутантом и неизмененным полипептидом качественно согласуется, т.е., например, превращаются одинаковые субстраты с различной скоростью.
«Функциональными эквивалентами» в вышеупомянутом смысле являются также предшественники описанных полипептидов, а также функциональные производные и соли этих полипептидов. Под термином «соли» понимают как соли карбоксильных групп, так и кислотно-аддитивные соли аминогрупп белковых молекул по изобретению. Соли карбоксильных групп могут быть получены известным per se способом и включают в себя неорганические соли, такие как, например, соли натрия, кальция, аммония, железа и цинка, а также соли с органическими основаниями, такими как, например, амины, например, триэтаноламин, аргинин, лизин, пиперидин и т.п. Кислотно-аддитивные соли, такие как, например, соли с минеральными кислотами, такими как соляная кислота или серная кислота, и соли с органическими кислотами, такими как уксусная кислота и щавелевая кислота, также являются предметом изобретения.
“Функциональные производные” полипептидов по изобретению могут быть также получены на функциональных боковых группах аминокислот или на их N- или С-концевых сторонах с использованием известных способов. Подобные производные включают в себя, например, алифатические эфиры групп карбоновых кислот, амиды групп карбоновых кислот, полученные превращением с аммиаком или с первичным или вторичным амином; N-ацилпроизводные свободных аминогрупп, полученные превращением с ацильными группами; или О-ацилпроизводные свободных гидроксигрупп, полученные превращением с ацильными группами.
«Функциональные эквиваленты» включают в себя, конечно, также полипептиды, которые доступны из других организмов, а также природно встречающиеся варианты. Например, посредством сравнения последовательностей могут быть установлены участки гомологичных районов последовательностей и в приложении к конкретным задачам изобретения могут быть получены эквивалентные ферменты.
«Функциональные эквиваленты» включают в себя также фрагменты, предпочтительно отдельные домены или мотивы последовательности, полипептидов данного изобретения, которые, например, обнаруживают желаемую биологическую функцию.
«Функциональными эквивалентами» являются, кроме того, слитые белки, которые обнаруживают одну из вышеупомянутых полипептидных последовательностей или произведенные из нее функциональные эквиваленты и по меньшей мере одну дополнительную функционально от нее отличающуюся, гетерологичную последовательность в функциональном N- или С-концевом связывании (т.е. без взаимного значимого функционального ухудшения частей слитого белка). Неограничительными примерами подобных гетерологичных последовательностей являются, например, сигнальные пептиды, ферменты, иммуноглобулины, поверхностные антигены, рецепторы или лиганды рецепторов.
“Функциональными эквивалентами” транс-сиалидаз по изобретению являются, в частности, ферменты, аминокислотные последовательности или аминокислотые частичные последовательности которых обнаруживают относительно соответствующей аминокислотной последовательности или аминокислотной частичной последовательности согласно SEQ ID NO:2 или 4 идентичность последовательности (гомологию последовательности) по меньшей мере 60%, в частности, по меньшей мере 70%, например, 75%, 80%, 85%, 90%, 95%, 98% или 99%, рассчитанную в соответствии с алгоритмом Pearson und Lipman, Proc. Natl. Acad. Sci. (USA) 85(8), 1988, 2444-2448.
В случае возможного гликозилирования белка эквиваленты согласно изобретению включают в себя белки вышеупомянутого типа в дегликозилированной или гликозилированной форме, а также полученные изменением картины гликозилирования измененные формы.
Гомологи белков и полипептидов по изобретению могут быть получены при помощи мутагенеза, например, посредством точковой мутации, удлинения или укорочения белка. Понятие “гомолог” относится в данном контексте также к вариантной форме белка, которая действует в качестве агониста или антагониста активности белка.
Гомологи белков по изобретению могут быть идентифицированы посредством скрининга комбинаторных банков мутантов, например, укороченных мутантов. Например, может быть создан мозаичный банк белков-вариантов при помощи комбинаторного мутагенеза на уровне нуклеиновых кислот, например, посредством ферментативного лигирования смеси синтетических олигонуклеотидов. Имеется множество способов, которые могут быть использованы для получения банков потенциальных гомологов из одной вырожденной олигонуклеотидной последовательности. Химический синтез вырожденной последовательности гена может выполняться в автоматизированном ДНК-синтезаторе, и синтетический ген может быть затем лигирован в подходящий экспрессирующий вектор. Применение вырожденного набора генов позволяет обеспечение всех последовательностей в одной смеси, которые кодируют желаемый набор потенциальных белковых последовательностей. Способы синтеза вырожденных олигонуклеотидов известны специалисту в данной области (например, Narang, S.A. (1982) Tetrahedron 39:3; Itakura et al. (1984) Annu. Rev. Biochem, 53:323; Itakura et al. (1984) Science 198:1056; Ike et al. (1983) Nucleic Acids Res. 11:477).
b) Полинуклеотиды
«Полинуклеотиды» в контексте изобретения включают в себя характерные частичные фрагменты последовательностей нуклеиновых кислот по изобретению, которые кодируют частичные аминокислотные последовательности ферментов по изобретению, а также последовательности нуклеиновых кислот, которые кодируют ферменты и их функциональные эквиваленты. Предпочтительно, полинуклеотиды включают в себя более приблизительно 20, в частности, более приблизительно 30, например, более приблизительно 45 или более приблизительно 60 остатков нуклеиновых кислот.
«Олигонуклеотиды» включают в себя, в частности, последовательность менее чем приблизительно 60, предпочтительно, менее чем приблизительно 45, в частности, менее чем приблизительно 30, или менее чем приблизительно 20 остатков нуклеиновых кислот.
Все упомянутые здесь «последовательности нуклеиновых кислот» могут быть получены известным per se образом при помощи химического синтеза из нуклеотидных элементарных звеньев, как, например, посредством конденсации фрагментов отдельных перекрывающихся, комплементарных элементарных звеньев нуклеиновых кислот двойной спирали. Химический синтез олигонуклеотидов может происходить, например, известным образом, в соответствии с фосфороамидитным способом (Voet, Voet, 2. Aufgabe, Wiley Press New York, Seiten 896-897). Присоединение синтетических олигонуклеотидов и заполнение гэпов с использованием фрагмента Кленова ДНК-полимеразы и реакций лигирования, а также общие способы клонирования описаны в Sambrook et al. (1989), Molecular Cloning: A Laboratоry Manual, Cold Spring Harbor Laboratory Press.
Предметом изобретения являются также последовательности нуклеиновых кислот (одноцепочечные и двухцепочечные ДНК- и РНК-последовательности, такие как, например, кДНК и мРНК), кодирующие один из вышеупомянутых полипептидов и их функциональных эквивалентов, которые, например, доступны также с использованием искусственных аналогов нуклеотидов.
Изобретение относится как к выделенным молекулам нуклеиновых кислот, которые кодируют полипептиды или белки согласно изобретению или их биологически активные сегменты, так и к фрагментам нуклеиновых кислот, которые, например, могут использоваться для применения в качестве гибридизационных зондов или праймеров для идентификации или амплификации кодирующих нуклеиновых кислот по изобретению.
Кроме того, молекулы нуклеиновых кислот по изобретению могут содержать нетранслируемые последовательности 3'- и/или 5'-концов кодирующей области гена.
«Выделенную» молекулу нуклеиновой кислоты отделяют от других молекул нуклеиновых кислот, которые присутствуют в природном источнике нуклеиновой кислоты, и она может, кроме того, быть по существу свободной от другого клеточного материала или от культуральной среды, если ее получают рекомбинантными способами, или может не содержать химических предшественников или других химикалиев, если ее синтезируют химически.
Молекула нуклеиновой кислоты по изобретению может быть выделена при помощи молекулярно-биологических стандартных способов и обеспеченной настоящим изобретением информации последовательности. Например, кДНК может быть выделена из подходящего кДНК-банка с использованием одной из конкретно раскрытых полных последовательностей или их сегмента в качестве гибридизационного зонда и стандартных способов гибридизации (как, например, описано в Sambrook, J., Fritsch, E.F. and Maniatis, T. Molecular Cloning: A Laboratоry Manual, Second Edition, Cold Spring Harbor Press Laboratory, Cold Spring Harbor, NY, 1989). Кроме того, молекула нуклеиновой кислоты, включающая в себя раскрытые последовательности или их сегмент, может быть выделена с использованием олигонуклеотидных праймеров, которые были изготовлены на основе этой последовательности. Амплифицированная таким образом нуклеиновая кислота может быть клонирована в подходящий вектор и охарактеризована при помощи анализа последовательности (секвенирования) ДНК. Олигонуклеотиды по изобретению могут быть получены, кроме того, стандартными способами синтеза, например, с использованием автоматизированного ДНК-синтезатора.
Изобретение включает в себя также молекулы нуклеиновых кислот, «комплементарные» конкретным описанным нуклеотидным последовательностям или их сегменту.
Нуклеотидные последовательности по изобретению делают возможным изготовление зондов и праймеров, которые применимы для идентификации и/или клонирования гомологичных последовательностей в других типах клеток и организмах. Такие зонды или праймеры включают в себя обычно район нуклеотидной последовательности, который при строгих условиях гибридизуется по меньшей мере с приблизительно 12, предпочтительно по меньшей мере с приблизительно 25, например, приблизительно 40, 50 или 75 следующими друг за другом нуклеотидами смысловой цепи последовательности нуклеиновой кислоты по изобретению или соответствующей антисмысловой цепи.
Дополнительные последовательности нуклеиновых кислот по изобретению произведены из SEQ ID NO:1 и 3 и отличаются от них добавлением, заменой, инсерцией или делецией одного или нескольких нуклеотидов, но кодируют, кроме того, полипептиды с желаемым профилем свойств.
В соответствии с изобретением включены также такие последовательности нуклеиновых кислот, которые включают в себя так называемые молчащие мутации или изменены в соответствии с использованием кодонов специального организма-источника или организма-хозяина, в сравнении с конкретной названной последовательностью, а также природно встречающиеся варианты, такие как, например, их сплайсинговые варианты или аллельные варианты. Предметом изобретения являются также полученные посредством консервативных нуклеотидных замен последовательности (т.е. рассматриваемую аминокислоту заменяют аминокислотой со сходными зарядом, размером, полярностью и/или растворимостью).
Предметом изобретения являются также молекулы, произведенные посредством полиморфизмов последовательностей конкретных раскрытых нуклеиновых кислот. Эти генетические полиморфизмы могут существовать между индивидуумами внутри популяции на основе природной изменчивости. Эти природные вариации вызывают обычно дисперсию 1-5% в нуклеотидной последовательности гена.
Кроме того, изобретение включает в себя также последовательности нуклеиновых кислот, которые гибридизуются с вышеупомянутыми кодирующими последовательностями или являются комплементарными им. Эти полинуклеотиды могут быть найдены при просмотре геномных банков или кДНК-банков и в случае необходимости размножены с подходящими праймерами при помощи ПЦР и после этого, например, выделены с подходящими зондами. Следующую возможность представляет трансформация подходящих микроорганизмов полинуклеотидами или векторами по изобретению, размножение этих микроорганизмов и, следовательно, полинуклеотидов, и их последующее выделение. Кроме того, полинуклеотиды по изобретению могут также синтезироваться химическими путями.
Под свойством “гибридизоваться” с полинуклеотидом имеют в виду способность поли- или олигонуклеотида связываться при строгих условиях с почти комплементарной последовательностью, в то время как при этих условиях не образуются неспецифические связи между некомплементарными партнерами. Для этого эти последовательности должны быть комплементарными до 70-100%, предпочтительно до 90-100%. Свойство комплементарных последовательностей специфически связываться друг с другом используют, например, в Нозерн- или Саузерн-блоттинге или при связывании праймера в ПЦР или ОТ-ПЦР. Для этого обычно используют олигонуклеотиды с длиной от 30 пар оснований.
Под “строгими условиями” имеют в виду условия, если, например, после Саузерн- или Нозерн-блоттинга ДНК- или РНК-фрагменты на мембранах гибридизуются с зондом при специфических условиях, т.е. при температуре 60-70°С (38-42°С при 50% гибридизационных растворах, которые содержат 50% формамид). Кроме того, эти условия являются специфическими или строгими, если сразу же после гибридизации проводят также специфически стадии промывок для элюции неспецифически гибридизованных ДНК- или РНК-зондов. При специфических стадиях промывок речь идет обычно о двухкратной промывке при 20-25°С в течение 5-10 минут 2 × SSC-буфером, который содержит 0,1% ДСН (додецилсульфат натрия), и последущих двухкратных промывках буфером более низкой ионной силы (например, 0,1 × SSC с 0,1% ДМН) при более высокой температуре (например, 64°С) [20 × SSC:3 М NaCl, 0,3 М Na-цитрат, рН 7,0]. При этом связанными друг с другом остаются только комплементарные в высокой степени нуклеиновые кислоты. Установление строгих условий известно специалисту в данной области и описано, например, в Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, N.Y. (1989), 6.3.1-6.3.6.
Следующий аспект изобретения относится к «антисмысловым» нуклеиновым кислотам. Они включают в себя последовательность нуклеиновой кислоты, которая комплементарна последовательности кодирующей «смысловой» нуклеиновой кислоты. Антисмысловая нуклеиновая кислота может быть комплементарна всей кодирующей цепи или только ее сегменту. В следующем варианте осуществления антисмысловая молекула нуклеиновой кислоты является антисмысловой относительно некодирующей области кодирующей цепи последовательности нуклеиновой кислоты. Термин «некодирующая область» относится к сегментам последовательности, называемым 5'- и 3'-нетранслируемыми областями.
Антисмысловой олигонуклеотид может иметь длину, например, приблизительно 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 нуклеотидов. Антисмысловые нуклеиновые кислоты данного изобретения могут конструироваться при помощи химического синтеза и ферментативных реакций лигирования с использованием известных в данной области способов. Антисмысловая нуклеиновая кислота может химически синтезироваться, причем используются природно встречающиеся нуклеотиды или различные модифицированные нуклеотиды, которые образованы таким образом, что они повышают биологическую стабильность молекул или повышают физическую стабильность дуплекса, который возникает между антисмысловой и смысловой нуклеиновой кислотой. Например, могут использоваться фосфоротиоатные производные и акридинзамещенные нуклеотиды. Примерами модифицированных нуклеотидов, которые могут использоваться для получения антисмысловой нуклеиновой кислоты, являются, например, 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-иодурацил, гипоксантин, ксантин, 4-ацетилцитозин и т.п.
Антисмысловые молекулы нуклеиновых кислот по изобретению обычно вводят в клетку или производят in situ, так что они могут гибридизоваться с клеточной мРНК и/или кодирующей ДНК или связываться с ними, так что экспрессия белка ингибируется, например, ингибированием транскрипции и/или трансляции.
Термины “экспрессировать” или “усиление” или “сверхэкспрессия” описывают в контексте настоящего изобретения образование или повышение внутриклеточной активности одного или нескольких ферментов в микроорганизме, которые кодируются соответствующей ДНК. Для этого можно, например, ввести в организм ген, заменить присутствующий ген другим геном, повысить копийность гена или генов, использовать сильный промотор или использовать ген, который кодирует соответствующий фермент с высокой активностью, и можно в случае необходимости комбинировать эти меры.
Термины “ослабить” и “уменьшить” описывают в контексте настоящего изобретения ослабление или уменьшение внутриклеточной активности одного или нескольких ферментов в микроорганизме, которые кодируются соответствующей ДНК. Для этого можно, например, делетировать ген в организме, заменить присутствующий ген другим геном, понизить копийность транскрипта гена или генов, использовать слабый промотор или использовать ген, который кодирует соответствующий фермент с более низкой активностью, и можно в случае необходимости комбинировать эти меры.
с) Экспрессионные конструкции и векторы:
Кроме того, предметом изобретения являются экспрессионные конструкции, содержащие под генетическим контролем регуляторных последовательностей нуклеиновых кислот последовательность нуклеиновой кислоты, кодирующую полипептид по изобретению; а также векторы, включающие в себя по меньшей мере одну из этих экспрессионных конструкций. Предпочтительно, такие конструкции по изобретению включают в себя 5'-против хода транскрипции (слева) от соответствующей кодирующей последовательности промотор и 3'-по ходу транскрипции (справа) терминирующую последовательность, а также в случае необходимости дополнительные регуляторные элементы, а именно, в каждом случае функционально связанные с кодирующей последовательностью.
Под “функциональным связыванием” понимают последовательное расположение промотора, кодирующей последовательности, терминатора и в случае необходимости дополнительных регуляторных элементов таким образом, что каждый из регуляторных элементов может выполнять надлежащим образом его функцию при экспрессии кодирующей последовательности. Примерами функционально связанных последовательностей являются нацеливающие последовательности, а также энхансеры, сигналы полиаденилирования и т.п. Дополнительные регуляторные элементы включают в себя селектируемые маркеры, сигналы амплификации, сайты инициации репликации (ориджин) и т.п. Подходящие регуляторные последовательности описаны, например, в Goeddel, Gene Expression Technology: Methods in Enzymology 195, Academic Press, San Diego, CA (1990).
Дополнительно к искусственным регуляторным последовательностям может присутствовать еще природная регуляторная последовательность перед собственно структурным геном. Посредством генетической модификации эта природная регуляция может быть в случае необходимости выключена, и экспрессия генов может повышаться или понижаться. Но генная конструкция может быть также построена более просто, т.е. не инсертируются дополнительные регуляторные сигналы перед структурным геном и не удаляется природный промотор с его регуляцией. Вместо этого природную регуляторную последовательность мутируют таким образом, что регуляция более не происходит и экспрессия гена повышается или уменьшается. Последовательности нуклеиновых кислот могут содержаться в данной конструкции гена в виде одной или в виде нескольких копий.
Примерами применимых промоторов являются: cos-, tac-, trp-, tet-, trp-tet-, lpp-, lac-, lpp-lac-, laclq-, T7-, T5-, T3-, gal-, trc-, ara-, SP6- лямбда-PR- или лямбда-PL-промотор, которые находят применения предпочтительно в грамотрицательных бактериях; а также грамположительные промоторы amy и SPO2, промоторы дрожжей ADC1, MFальфа, AC, P-60, CYC1, GAPDH или промоторы растений CaMV/35S, SSU, OCS, lib4, usp, STLS1, B33, not или промотор убиквитина или фазеолина. Особенно предпочтительным является применение индуцируемых промоторов, таких как, например, индуцируемые светом и особенно индуцируемые температурой промоторы, такие как PrPl-промотор. В принципе могут использоваться все природные промоторы с их регуляторными последовательностями. Кроме того, могут также предпочтительно использоваться синтетические промоторы.
Вышеупомянутые регуляторные последовательности должны делать возможной нацеленную экспрессию последовательностей нуклеиновых кислот и экспрессию белков. Это может означать, например, в зависимости от организма-хозяина, что ген экспрессируется или сверхэкспрессируется только после индукции, или что он экспрессируется и/или сверхэкспрессируется сразу же.
Регуляторные последовательности или факторы могут при этом предпочтительно положительно влиять на экспрессию и благодаря этому повышать или понижать ее. Так, может происходить усиление регуляторных элементов предпочтительно на уровне транскрипции, с использованием сильных сигналов транскрипции, таких как промоторы и/или энхансеры. Наряду с этим, возможно также усиление трансляции улучшением, например, стабильности мРНК.
Получение экспрессионной кассеты выполняют посредством слияния подходящего промотора с подходящей кодирующей последовательностью нуклеиновой кислоты, а также терминирующим сигналом или сигналом полиаденилирования. Для этого используют общепринятые способы рекомбинации и клонирования, такие как, например, описанные в Т. Maniatis, E.F., Fritsch und J. Sambrook, Molecular Cloning: A Laboratоry Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor (1989), а также в T.J. Silhavy, M.L.Berman und L.W. Enquist, Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1984) и в Ausubel F.M. et al., Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley Interscience (1987).
Рекомбинантную конструкцию нуклеиновой кислоты или генную конструкцию инсертируют для экспрессии в подходящем организме-хозяине в хозяин-специфический вектор, который делает возможной оптимальную экспрессию этого гена в хозяине.
«Векторы» хорошо известны специалисту в данной области и могут быть взяты, например, из “Cloning Vectors” (Pouwels P.H. et al., Hrsg, Elsevier, Amsterdam-New York-Oxford, 1985). Под векторами следует понимать, кроме плазмид, также все другие известные специалисту векторы, такие как, например, фаги, вирусы, такие как SV40, CMV, бакуловирус и аденовирус, транспозоны, IS-элементы (мобильные нуклеотидные последовательности, встречающиеся в геномах бактериальных популяций), фагмиды, космиды и линейные или кольцевые ДНК. Эти векторы могут реплицироваться автономно в организме-хозяине или хромосомно.
В качестве примеров экспрессионных векторов могут быть названы:
Обычные слитые экспрессионные векторы, такие как pGEX (Pharmacia Biotech Inc; Smith, D.B. und Johnson, K.S. (1988) Gene 67:31-40), pMAL (New England Biolabs, Beverly, MA) и pRIT 5 (Pharmacia, Piscataway, NJ), у которых глутатион-S-трансфераза, мальтозу Е-связывающий белок или белок А слиты с рекомбинантным целевым белком.
Экспрессионные векторы, не являющиеся слитыми белками, такие как pTrc (Amann et al., (1989) Gene 69:301-315) и pET 11d (Studier et al. Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Kalifornien (1990) 60-89).
Дрожжевой экспрессионный вектор для экспрессии в дрожжах S. cerevisiae, такой как pYepSec1 (Baldari et al., (1987) Embo J. 6:229-234), pMFa (Kurjan und Herskowitz (1982) Cell 30:933-943), pJRY88 (Schultz et al. (1987) Gene 54:113-123), а также pYES2 (Invitrogen Corporation, San Diego, CA). Векторы и способы конструирования векторов, которые пригодны для применения в других грибах, таких как нитевидные грибы, включают в себя векторы и способы, которые подробно описаны в van den Hondel, C.A.M.J.J. & Punt, P.J. (1991) “Gene transfer systems and vector development for filamentous fungi, in: Applied Molecular Genetics of Fungi, J.F. Peberdy et al., Hrsg., S. 1-28, Cambridge University Press: Cambridge.
Бакуловирусные векторы, которые пригодны для экспрессии белков в культивируемых клетках насекомых (или Sf9-клетках), включают в себя pAc-серию (Smith et al., (1983) Mol. Cell Biol., 3:2156-2165) и pVL-серию (Lucklow und Summers (1989) Virology 170:31-39).
Экспрессионные векторы растений, такие, какие подробно описаны в: Becker, D., Kemper, E., Schell, J. und Masterson, R. (1992) “New plant binary vectors with selectable markers located proximal to the left border” Plant Mol. Biol. 20:1195-1197; и Bevan, M.W. (1984) “Binary Agrobacterium vectors for plant transformation”, Nucl. Acids Res. 12:8711-8721.
Экспрессионные векторы млекопитающих, такие как pCDM8 (Seed, B. (1987) Nature 329:840) и pMT2PC (Kaufman et al. (1987) EMBO J. 6:187-195).
Дополнительные подходящие системы экспрессии для прокариотических и эукариотических клеток описаны в главах 16 и 17 Sambrook, J., Fritsch, E.F. and Maniatis, T. Molecular Cloning: A Laboratоry Manual, Second Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
d) Рекомбинантные микроорганизмы:
При помощи векторов по изобретению могут быть получены рекомбинантные микроорганизмы, которые, например, трансформированы по меньшей мере одним вектором по изобретению и могут быть использованы для получения полипептидов по изобретению. Предпочтительно, вышеописанные рекомбинантные конструкции по изобретению вводят в подходящую систему-хозяин и экспрессируют. При этом используют предпочтительно известные специалисту общепринятые способы клонирования и трансфекции, такие как, например, копреципитация, слияние протопластов, электропорация, ретровирусная трансфекция и т.п., чтобы заставить вышеупомянутые нуклеиновые кислоты экспрессироваться в соответствующей системе экспрессии. Подходящие системы описаны, например, в Current Protocols in Molecular Biology, F. Ausubel et al., Hrsg, Wiley Interscience, New York 1997 или Sambrook et al. Molecular Cloning: A Laboratоry Manual, Second Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
В соответствии с изобретением могут быть получены также гомологично рекомбинированные микроорганизмы. Для этого получают вектор, который содержит по меньшей мере один сегмент гена по изобретению или кодирующей последовательности, в который были введены в случае необходимости делеция, добавление или замена одной аминокислоты для изменения последовательности по изобретению, например, функционального разрушения (“нокаут”-вектор). Введенная последовательность может быть, например, также гомологом из родственного микроорганизма или быть произведена из источника, являющегося млекопитающим, дрожжами или насекомым. Альтернативно, используемый для гомологичной рекомбинации вектор может быть построен таким образом, что эндогенный ген мутирован при гомологичной рекомбинации или по-другому изменен, однако все еще кодирует функциональный белок (например, лежащая слева (против хода транскрипции) регуляторная область может быть изменена таким образом, что посредством этого изменяется экспрессия эндогенного белка). Этот измененный сегмент гена по изобретению находится в векторе гомологичной рекомбинации. Конструирование подходящих векторов для гомологичной рекомбинации описано, например, в Thomas, K.R. und Capecchi, M.R. (1987) Cell 51:503.
В качестве организмов-хозяев пригодны в принципе все организмы, которые делают возможной экспрессию нуклеиновых кислот по изобретению, их аллельных вариантов, их функциональных эквивалентов или производных. Под организмами-хозяевами понимают, например, бактерии, грибы, дрожжи, растительные клетки или клетки животных. Предпочтительными организмами являются бактерии, такие как бактерии родов Escherichia, например, Escherichia coli, Streptomyces, Bacillus или Pseudomonas, эукариотические микроорганизмы, такие как Saccharomyces cerevisiae, Aspergillus, высшие эукариотические клетки из животных или растений, например, клетки Sf0 или СНО.
Отбор успешно трансформированных организмов может происходить с использованием маркерных генов, которые также содержатся в векторе или в экспрессионной кассете. Примерами таких маркерных генов являются гены устойчивости к антибиотикам и гены ферментов, катализирующих цветную реакцию, которая влияет на окраску трансформированных клеток. Они могут также отбираться при помощи автоматизированного сортинга клеток. Успешно трансформированные вектором микроорганизмы, которые несут соответствующий ген устойчивости к антибиотикам (например, к G418 или гигромицину), могут отбираться также с использованием соответствующих содержащих антибиотики сред или питательных сред. Маркерные белки, которые презентированы на клеточной поверхности, могут быть использованы для отбора с использованием аффинной хроматографии.
Комбинация из организмов-хозяев и подходящих для этих организмов векторов, таких как плазмиды, вирусы или фаги, например, плазмиды с системой РНК-полимераза/промотор, фаги 8 или 7 или другие умеренные фаги или транспозоны и/или дополнительные предпочтительные регуляторные последовательности, образует систему экспрессии. Например, под термином “система экспрессии” следует понимать комбинацию из клеток млекопитающего, таких как клетки СНО, и векторов, таких как вектор pcDNA3neo, которые пригодны для клеток млекопитающих.
Наиболее желательно, генный продукт может экспрессироваться также в трансгенных организмах, таких как трансгенные животные, в частности, мыши, овцы, или трансгенных растениях.
е) Рекомбинантное получение полипептидов:
Кроме того, предметом изобретения являются способы рекомбинантного получения полипептида по изобретению или его функциональных, биологически активных фрагментов, в которых продуцирующий полипептиды микроорганизм культивируют, в случае необходимости индуцируют экспрессию полипептидов и выделяют их из культуры. Полипептиды могут таким образом продуцироваться также и в крупномасштабном производстве, если это желательно.
Рекомбинантные микроорганизмы могут культивироваться и ферментироваться согласно известным способам. Бактерии могут, например, размножаться в ТВ- или LB-среде и при температуре 20-40°С и показателе рН 6-9. В частности, подходящие условия культивирования описаны, например, в Т. Maniatis, E.F., Fritsch und J. Sambrook, Molecular Cloning: A Laboratоry Manual, Second Edition, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989).
Затем эти клетки, в том случае, если полипептиды секретируются в культуральную среду, растворяют и продукт получают согласно известным способам выделения белков из лизата. Эти клетки могут быть растворены на выбор посредством высокочастотного ультразвука, посредством высокого давления, например, во French-прессе, посредством осмолиза, посредством обработки детергентами, литическими ферментами или органическими растворителями, с использованием гомогенизаторов или комбинацией нескольких из названных способов.
Очистка полипептидов может достигаться известными, хроматографическими способами, такими как хроматография с молекулярными ситами (гель-фильтрация), такими как хроматография с использованием Q-сефарозы, ионообменная хроматография и гидрофобная хроматография, а также другими обычными способами, такими как ультрафильтрация, кристаллизация, высаливание, диализ и нативный гель-электрофорез. Подходящие способы описаны, например, в Cooper, F.G., Biochemische Arbeitsmethoden, Verlag Walter de Gruyter, Berlin, New York или в Scopes, R., Protein Purification, Springer Verlag, New York, Heidelberg, Berlin. То же самое относится к полученным нерекомбинантно полипептидам.
Особенно предпочтительно использовать для выделения рекомбинантных белков векторные системы или олигонуклеотиды, которые удлиняют кДНК на определенные нуклеотидные последовательности и тем самым кодируют удлиненные полипептиды или слитые белки, которые, например, служат для более простой очистки. Подобными подходящими модификациями являются, например, действующие в качестве якоря так называемые «метки», такие как, например, известная как гекса-гистидиновый якорь модификация или эпитопы, которые могут узнаваться в качестве антигенов антителами (описанные, например, в Harlow, E. and Lane, D., 1988, Antibodies: A Laboratory Manual, Cold Spring Harbor (N.Y.) Press). Эти якоря могут использоваться для прикрепления белков к твердому носителю, такому как полимерный матрикс, который, например, может быть помещен в хроматографическую колонку, или к микротитрационному планшету или иному носителю.
Одновременно эти якоря могут использоваться также для узнавания белков. Кроме того, для узнавания белков могут также применяться обычные маркеры, такие как флуоресцентные красители, ферментные маркеры, которые после реакции с субстратом образуют детектируемый продукт реакции, или радиоактивные маркеры, по отдельности или в комбинации с якорями для дериватизации белков.
f) Очистка желаемых продуктов силилирования из культуры
Получение желаемого продукта из микроорганизма или из культурального супернатанта может выполняться с использованием различных, известных в данной области способов. Если желаемый продукт не секретируется из клеток, клетки могут быть собраны из культуры медленным центрифугированием, клетки могут быть лизированы при помощи стандартных способов, таких как обработка механическим усилием или обработка ультразвуком. Остатки клеток удаляют центрифугированием и фракцию супернатанта, которая содержит растворимые белки, получают для дополнительной очистки желаемого соединения. Если продукт клеток секретируется, клетки удаляют из культуры медленным центрифугированием и фракцию супернатанта сохраняют для дальнейшей очистки.
Фракция супернатанта из обоих способов очистки может быть подвергнута хроматографии с подходящей смолой, при которой желаемая молекула с более высокой селективностью, чем примеси, либо удерживается хроматографической смолой, либо проходит через нее. Эти стадии хроматографии могут в случае необходимости повторяться, причем используют те же самые или другие хроматографические смолы. Специалисту известны выбор подходящей хроматографической смолы и ее эффективное применение для определенной, подлежащей очистке молекулы. Очищенный продукт может концентрироваться фильтрованием или ультрафильтрацией и храниться при температуре, при которой стабильность этого продукта является максимальной.
Из уровня техники известны многие способы очистки. Эти способы очистки описаны, например, в Bailey, J.E. & Ollis, D.F. Biochemical Engineering Fundamentals, McGraw-Hill: New York (1986).
Идентичность и чистота выделенных соединений может быть определена способами существующего уровня техники. Они включают в себя высокоэффективную жидкостную хроматографию (ВЖХ), спектроскопический способ, способ с изменением окраски, тонкослойную хроматографию, NIRS, ферментативный тест или микробиологические тесты. Эти способы анализа рассматриваются, например, в: Patek et al. (1994) Appl. Environ. Microbiol. 60:133-140; Malakchova et al. (1996) Biotekhnologiya 11 27-32; и Schmidt et al. (1998) Bioprocess Engineer. 19:67-70. Ullman's Encyclopedia of Industrial Chemistry (1996) Bd. F27, VCH: Weinheim, S. 89-90, S. 521-540, S. 540-547, S. 559-566, 575-581 и S. 581-587; Michal, G. (1999) Biochemical Pathways: An Atlas of Biochemistry and Molecular Biology, John Wiley and Sons; Fallon, A. et al. (1987) Applications of HPLC in Biochemistry in: Laboratory Techniques in Biochemistry and Molecular Biology, Bd. 17.
Следующие неограничивающие примеры описывают конкретные варианты осуществления изобретения.
Примеры получения, очистки и применения заявленных здесь транс-сиалидаз
Общая часть:
Проведенные в рамках настоящего изобретения стадии клонирования, такие как, например, расщепления рестриктазами, электрофорез в агарозном геле, очистка ДНК-фрагментов, перенос нуклеиновых кислот на нитроцеллюлозу и найлоновые мембраны, связывание ДНК-фрагментов, трансформация клеток, выращивание бактерий, размножение фагов и анализ последовательностей (секвенирование) рекомбинантных ДНК проводили, как описано Sambrook et al. (1989) в указанном выше месте.
Пример 1: Выделение фермента из культур Trypanosoma congolense
Проциклические формы Trypanosoma congolense (депонированные в Schweizer Tropeninstitut Basel (STIB) в виде штамма № 249) могут выращиваться при 27°С без СО2 в среде SM/SDM 79, которая содержит 10% фетальную телячью сыворотку и гемин. Спустя три-четыре дня количество клеток вырастает с 1×106 до 7×106/мл и культуральный супернатант отделяют центрифугированием, фильтруют и концентрируют ультрафильтрацией. В полученном таким образом культуральном супернатанте могут быть установлены 84% активности фермента, в то время как еще 16% активности фермента может детектироваться в виде связанной с клеточным остатком активности. Концентрированные культуральные супернатанты используют непосредственно в качестве концентрата фермента для реакций транс-сиалидазы. Желаемые сиалилированные молекулы выделяют после реакции из культурального супернатанта.
Пример 2: Очистка фермента
Для выделения чистых ферментов концентрированный культуральный супернатант наносят на ионообменную колонку (Q-сефарозу). Колонку элюируют после промывания градиентами соли. TS2 элюируется концентрацией соли до 0,2 М, TS1 градиентами соли менее 0,2 М. После элюции оба фермента разделяют с использованием изоэлектрического фокусирования, гель-фильтрации (Сефадекс G150 SF), аффинной хроматографии или осаждения белка до кажущейся гомогенности.
Пример 3: Определение активности фермента
Для определения активности переноса транс-сиалидазы 25 мкл раствора фермента в 50 мМ Бис-Трис-буфере, рН 7,0, инкубируют вместе с 1 мМ Neu5Ac-α(2-3)лактозой в качестве донора и 0,5 мМ 4-метилумбеллиферилгалактозидом в качестве акцептора в конечном объеме 50 мкл при 37°С в течение 2 часов. Инкубирование останавливают добавлением 1 мл ледяной воды. После этого реакционную смесь наносят на предварительно заполненные 0,3 мл Q-сефарозы FF (ацетатной формы) и предварительно уравновешенные водой колонки. После вымывания акцептора водой и выбрасывания свободного объема (200 мкл 1н HCl) элюируют продукт 1н HCl (700 мкл). После кислотного гидролиза продукта при 95°С в течение 45 минут и охлаждения на льду пробу 250-290 мкл нейтрализуют 2н NaOH и 300 мкл 1 М глицин/NaOH-буфера рН 10,0. Измеряют флуоресценцию высвобождающегося метилумбеллиферона в черных 96-луночных планшетах (Microfluor, Dynex, U.S.A) при длине волны возбуждения 365 нм и длине волны испускания 450 нм. Активность фермента соответствует интенсивности измеренной флуоресценции и может считываться по построенной предварительно калибровочной кривой (Способ Engstler et al. 1992).
Пример 4: Получение трансгенных продукционных организмов (бактерий, дрожжей, грибов, растений) в отношении этого фермента
Описанные здесь впервые частичные ДНК-последовательности TS1 и TS2 позволяют с использованием рутинно применяемых стандартных способов установить полноразмерные ДНК-последовательности этих ферментов (в частности, потому, что представленные здесь ДНК-последовательности не содержат некодирующих интронов). Соответствующими стандартными способами являются, например, способ «полимеразной цепной реакции» (ПЦР-способ), «блоттинг по Саузерну» включающий в себя способы для геномных ДНК или кДНК и мРНК, которые могут проводиться с использованием коммерчески доступных наборов, например, из фирм Invitrogen или Clontech. Эти способы известны специалисту в данной области и описаны, inter alia, в: Ausubel et al.: Current Protocols in Molecular Biology, Edition 1989 und 2001. Полноразмерную ДНК соответствующих ферментов или функциональные частичные последовательности из нее вводят в желаемые организмы-продуценты при помощи стандартных способов трансформации (Ausubel et al.). Эти трансгенные организмы продуцируют TS1 и TS2, и они могут быть выделены из трансгенных организмов и/или их культуральных супернатантов. В качестве организмов-реципиентов ДНК, которые кодируют TS1 и TS2, используют прокариотические бактерии, эукариотические микроорганизмы, дрожжи и другие грибы, культуры эукариотических клеток, водоросли, растения, семена, животных, части животных, ткани, гибридомы, трансгенные организмы и генобиологические, генотерапевтические и трансгенные рекомбинанты, а также полученные из них организмы, органы, ткани и клетки. Заявленные здесь ферменты могут быть выделены из соответствующих целых трансгенных организмов или их частей, из их культуральных супернатантов, из органов, тканей, клеток, биологических жидкостей, экссудатов, яиц, крови, лимфы, молока, растений, водорослей и семян, а также из их частей.
Пример 5: Пример реакции ферментов
6 кг гликомакропептида (GMP) растворяют вместе с 1 кг галактоолигосахарида с длиной цепи 6-10 сахарных остатков в доступном в продаже 50 мМ БисТрис-буфере рН 7,0 (например, фирмы Merck, Darmstadt). Раствор смешивают с 1 литром содержащего транс-сиалидазу культурального супернатанта Trypanosoma congolense и культивируют при 37°С в течение 3 часов. После этого периода времени транс-сиалидаза переносит сиаловые кислоты от GMP на галактоолигосахарид. Сиалилированные продукты могут быть при помощи обычных хроматографических способов (Ausubel et al.) или технологий фильтрования отделены и очищены и предоставлены в чистом состоянии для приготовления композиций продукта.
Гликомакропептид (GMP) является побочным продуктом приготовления сыра из коровьего молока. После осаждения казеина для приготовления сыра он может быть выделен из оставшейся молочной сыворотки при помощи технологии фильтрования.
Галактоолигосахариды получают превращением лактозы с использованием получаемого из фирм фермента бета-галактозидазы. При этом превращении бета-галактозидаза расщепляет, с одной стороны, лактозу до ее мономерного сахара. С другой стороны, при этом расщеплении возникают в побочной реакции также более длиннноцепочечные галактоолигосахариды, которые могут быть отделены и затем предоставлены в качестве акцепторов для транс-сиалидазной реакции.
Пример 6: Использование ферментов
Оба изолированных фермента могут использоваться, например, для сиалилирования содержащих бета-галактозу полимеров (таких как аравийская камедь и т.д.) и, в частности, для полилактозаминов и галактанов, а также для галактоолигосахаридов (GOS), в частности, для бета-галактоолигосахаридов, таких как, например, Vivinal GOS фирмы Borculo Domo Ingredients (BDI) и Oligomate 55 фирмы Yakult.
Эти полимерные галактозо-сахара и новообразованные галактоолигосахариды (получение, как описано в примере 5) могут сиалилироваться при помощи заявленных в данном патенте транс-сиалидаз. В качестве доноров для сиаловых кислот могут использоваться все вышеупомянутые доноры и, в частности, гликомакропептид из казеинов (человека, коровы, козы, овцы, лошади, верблюда и других животных). Сиалилированные сахарные структуры обнаруживают увеличенное сходство с кислыми сахарами, которые могут быть также найдены в теле человека и иметь там многосторонние функции.
Литература
Blix, F. G., Gottschalk, A. und Klenk, E. (1957). Proposed nomenclature in the field of neuraminic and sialic acids. Nature 179: 1088.
Chuenkova M., Pereira M., (1999). Taylor G. trans-sialidase of Trypanosoma cruzi: Location of galactose-binding site(S). BIOCHEM BIOPHYS RES COMMUN; 262: 549-556.
Chuenkova M.V., Pereira M.A. (2001). The T. cruzi trans-sialidase induces PC12 cell differentiation via MAPK/ERK pathway. Neuroreport 12: 3715-3718.
Corfield A.P., Myerscough N., Warren B.F., Durdey P., Paraskeva C. and: Schauer R. (1999). Reduction of sialic acid O-acetylation in human colon mucins in the adenoma-carcinoma sequence, Glycoconjugate J. 16: 307-317.
Crocker, P. R., Clark, E. A., Filbin, M., Gordon, S., Jones, Y., Kehrl, J. H., Kelm, S., Le Douarin, N., Powell, L., Roder, J., Schnaar, R. L., Sgroi, D. C., Stamenkovic, K., Schauer, R., Schachner, M., van den Berg, T. K., van der Merwe, P. A., Watt, S. M. und Varki, A. (1998). Siglecs: a family of sialic-acid binding lectins. Glycobiology 8, v.
Engstler, M., Reuter, G. und Schauer, R. (1992). Purification and characterization of a novel sialidase found in procyclic culture forms of Trypanosoma brucei. Mol. Biochem. Parasitol. 54: 21-30.
Engstler M., Reuter G., Schauer R. (1993). The developmentally regulated trans-sialidase from Trypanosoma brucei sialylates the procyclic acidic repetitive protein. Mol Biochem Parasitol; 61: 1-13.
Engstler M., Schauer R., Brun R. (1995). Distribution of developmentally regulated trans-sialidases in the Kinetoplastida and characterization of a shed trans- sialidase activity from procyclic Trypanosoma congolense. Acta Trop; 59: 117-129.
Fahr C. and Schauer R. (2001). Detection of Sialic Acids and Gangliosides with Special Reference to 9-O-Acetylated Species in Basaliomas and Normal Human Skin, J. Invest. Dermatol. 116: 254-260.
Gao W, Pereira MA. (2001). Trypanosoma cruzi trans-sialidase potentiates T cell activation through antigen-presenting cells: role of IL-6 and Bruton's tyrosine kinase. Eur J Immunol 31: 1503-1512.
Herrler, G., Rott, R., Klenk, H. D., Muller, H. P., Shukla, A. K. und Schauer, R. (1985). The receptor-destroying enzyme of influenza C virus is neuraminate-O-acetylesterase. EMBO J. 4, 1503-1506.
Higa, H. H., Rogers, G. N. und Paulson, J. C. (1985). Influenza virus hemagglutinins differentiate between receptor determinants bearing N-acetyl-, N-glycolyl-, and N,O-diacetyl-neuraminic acids. Virology 144: 279-282.
Hubl U., Ishida H., Kiso M., Hasegawa A., and Schauer R. (2000). Studies on the Specificity and Sensitivity of the Influenza C Virus Binding Assay for O-Acetylated Sialic Acids and Its Application to Human Melanomas, J. Biochem. 127: 1021-1031.
Kelm, S. und Schauer, R. (1997). Sialic acids in molecular and cellular interactions. Int. Rev.Cytol. 175: 137-240.
Kelm, S., Schauer, R. und Crocker, P. R. (1996). The Sialoadhesins - a family of sialic acid-dependent cellular recognition molecules within the immunoglobulin superfamily. Glycoconj. J. 13: 913-926.
Lasky, L. A. (1995). Selectin-carbohydrate interactions and the initiation of the inflamma-tory response. Ann. Rev. Biochem. 64, 113-139.
Mattos-Guaraldi, A.L, Formiga, L.C.D. and Andrade, A.F.B. (1998). FEMS Microbiology Letters, 168: 167-172.
Medina-Acosta E., Paul S., Tomlinson S., Pontes-de-Carvalho L.C. (1994). Combined occurrence of trypanosomal sialidase/trans-sialidase activities and leishmanial metalloproteinase gene homologues in Endotrypanum sp. Mol Biochem Parasitol; 64: 273-282.
Montagna G, Cremona ML, Paris G, Amaya MF, Buschiazzo A, Alzari PM, Frasch AC (2002). The trans-sialidase from the african trypanosome Trypanosoma brucei. Eur J Biochem; 269: 2941-2950.
Pilatte, Y., Bignon, J. und Lambré, C. R. (1993). Sialic acids as important molecules in the regulation of the immune system: pathophysiological implications of sialidases in immunity. Glycobiology 3: 201-218.
Pontes de Carvalho L.C., Tomlinson S., Vandekerckhove F., Bienen E.J., Clarkson A.B., Jiang M.S., Hart G.W., Nussenzweig V. (1993). Characterization of a novel trans-sialidase of Trypanosoma brucei procyclic trypomastigotes and identification of procyclin as the main sialic acid acceptor. J Exp Med; 177: 465-474.
Reuter, G. und Schauer, R. (1988). Nomenclature of sialic acids. Glycoconj. J. 5: 133-135.
Reuter, G., Kelm, S. und Schauer, R. (1988). Chemistry and biology of cell surface glyco-conjugates. Acta Histochem. Suppl. 36: 51-79.
Schauer R. (2000), Achievements and challenges of sialic acid research, Glycoconjugate J. 17: 485-499.
Schauer, R. (1982). Chemistry, metabolism, and biological functions of sialic acids. Adv. Carbohydr. Chem. Biochem. 40: 131-234.
Schauer, R. (1985). Sialic acids and their role as biological masks. Trends Biochem. Sci. 10, 357-360.
Schauer, R. (1991). Biosynthesis and function of N- and O-substituted sialic acids. Glyco-biology 1: 449-452.
Schauer, R. und Kamerling, J. P. (1997). Chemistry, biochemistry and biology of sialic acids. In Glycoproteins II. J. Montreuil, J. F. G. Vliegenthart, und H. Schachter, eds. (Amsterdam: Elsevier), pp. 243-402.
Schauer, R., Kelm, S., Reuter, G., Roggentin, P. und Shaw, L. (1995). Biochemistry and role of sialic acids. In Biology of sialic acids. A. Rosenberg, ed. (New York: Plenum Press), pp. 7-67.
Schenkman, S., Jiang, M. S., Hart, G. W. und Nussenzweig, V. (1991). A novel cell surface trans-sialidase of Trypanosoma cruzi generates a stage-specific epitope required for invasion of mammalian cells. Cell 65; 1117-1125.
Sharon, N. und Lis, H. (1997). Microbial lectins and their glycoprotein receptors. In Glyco-proteins II. J. Montreuil, J. F. G. Vliegenthart und H. Schachter, eds. (Amsterdam: Elsevier), pp. 475-506.
Shaw, L. und Schauer, R. (1988). The biosynthesis of N-glycoloylneuraminic acid occurs by hydroxylation of the CMP-glycoside of N-acetylneuraminic acid. Biol. Chem. Hoppe-Seyler 369: 477-486.
Tertov V.V., Kaplun V.V., Sobenin I.A., Boytsova, E.Y., Bovin N.V. & Orekhov A.N. (2001). Human plasma trans-sialidase causes atherogenic modification of low density lipoprotein. Atherosclerosis 159: 103-115.
Traving, C. und Schauer, R. (1998). Structure, function and metabolism of sialic acids. CMLS, Cell. Mol. Life Sci. 54: 1330-1349.
Varki, A., Hooshmand, F., Diaz, S., Varki, N. M. und Hedrick, S. M. (1991). Developmental abnormalities in transgenic mice expressing a sialic acid-specific 9-O-acetylesterase. Cell 65; 65-74.








Изобретение относится к биотехнологии и касается фермента транс-сиалидазы. Этот фермент был выделен из одноклеточного организма Trypanosoma congolense. Транс-сиалидаза характеризуется одной из следующих аминокислотных последовательностей: SEQ ID NO:2, SEQ ID NO:4 или последовательностью, обладающей по меньшей мере 75% идентичностью с одной из вышеуказанных последовательностей. Изобретение позволяет расширить арсенал ферментов с транс-сиалидазной активностью. 5 з.п. ф-лы, 3 ил., 1 табл.
1. Транссиалидаза, которая может быть получена из Trypanosoma congolense, характеризующаяся одной из следующих аминокислотных последовательностей: SEQ ID NO: 2, SEQ ID NO: 4, или последовательностью, обладающей по меньшей мере 75%-ной идентичностью с одной из вышеуказанных последовательностей.
2. Транссиалидаза 1 (TS1) по п.1, отличающаяся по меньшей мере одной из следующих характеристик:
3. Транссиалидаза 2 (TS2) по п.1, отличающаяся по меньшей мере одной из следующих характеристик:
4. Транссиалидаза по одному из пп.1-3, происходящая из организма Trypanosoma congolense.
5. Транссиалидаза по одному из пп.1-3, полученная с использованием синтетических, в частности химических, биохимических, ферментативных, генотехнологических и трансгенных способов.
6. Транссиалидаза по п.2 или 3, аминокислотная последовательность или аминокислотная частичная последовательность которой обнаруживает относительно соответствующей аминокислотной последовательности или аминокислотной частичной последовательности согласно SEQ ID NO: 2 или 4 идентичность последовательности по меньшей мере 75%, рассчитанную в соответствии с алгоритмом Pearson und Lipraan, Proc. Natl. Acad. Sci. (USA) 85(8), 1988, 2444-2448; и содержит одну или несколько делеций, добавлений, замен или инверсий отдельных или нескольких аминокислотных остатков; причем остается сохраненной способность к катализу переноса сиаловых кислот от донора на акцептор.
| PONTES DE CARVALHO LC | |||
| Characterization of a novel trans-sialidase of Trypanosoma brucei procyclic trypomastigotes and identification of procyclin as the main sialic acid acceptor | |||
| J Exp Med | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| MONTAGNA G, The trans-sialidase from the african trypanosome Trypanosoma brucei | |||
| Eur J Biochem | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| WO | |||

