ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение раскрывает новые гены, кодирующие устойчивость к гербицидам, которые применимы в биологии растений, при селекции сельскохозяйственных культур и для культур растительных клеток.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
N-фосфонометилглицин, обычно называемый глифосатом, является важным агрономическим химикатом. Глифосат ингибирует фермент, который превращает фосфоенолпировиноградную кислоту (РЕР) и 3-фосфошикимовую кислоту (S3P) в 5-енолпирувил-3-фосфошикимовую кислоту. Ингибирование этого фермента (5-енолпирувилшикимат-3-фосфатсинтаза; обозначаемая в настоящем описании как "EPSPS"), уничтожает растительные клетки посредством отключения пути метаболизма шикимата, подавляя таким образом биосинтез ароматических кислот.
Поскольку гербициды класса глифосата подавляют биосинтез ароматических аминокислот, они не только уничтожают растительные клетки, но и являются токсичными для бактериальных клеток. Глифосат подавляет много бактериальных синтаз EPSP, и таким образом является токсичным для этих бактерий. Однако определенные бактериальные синтазы EPSP имеют высокую устойчивость к глифосату.
Растительные клетки, устойчивые к токсичности глифосата, могут быть получены посредством трансформирования растительных клеток для экспрессии глифосат-устойчивых бактериальных EPSP-синтаз. В частности, бактериальный ген из штамма CP4 Agrobacterium tumefaciens был использован для придания растительным клеткам устойчивости к гербицидам после экспрессии в растениях. Мутированная EPSP-синтаза из штамма CT7 Salmonella typhimurium придает устойчивость к глифосату бактериальным клеткам и придает устойчивость к глифосату растительным клеткам (Патенты США №№ 4535060; 4769061 и 5094945). Однако существует потребность в других генах устойчивости к гербицидам.
Кинетическая активность EPSPS может быть оценена посредством измерения высвобождения фосфата. Высвобождение фосфата определяется с использованием сопряженного анализа для флуоресцентного обнаружения фосфата на основе образования N-ацетил-3,7-дигидроксифеноксацина (Amplex® Red), как известно в области техники (Vazquez et al. (2003) Analytical Biochemistry 320: 292-298). Опубликованные условия анализа могут привести к насыщению анализа в экспериментах, где фосфат высвобождается очень быстро. Для измерения кинетической активности EPSPS необходимы дополнительные способы.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Представлены композиции и способы придания устойчивости или толерантности к гербициду бактериям, растениям, растительным клеткам, тканям и семенам. Композиции включают молекулы нуклеиновых кислот, кодирующие полипептиды устойчивости или толерантности к гербициду, векторы, содержащие такие молекулы нуклеиновых кислот, и клетки-хозяева, содержащие векторы. Композиции также включают антитела к полипептидам устойчивости или толерантности к гербициду. Как отмечено, нуклеотидные последовательности по изобретению могут быть использованы в конструкциях ДНК или экспрессионных кассетах для трансформации и экспрессии в организмах, включающих микроорганизмы и растения. Композиции также содержат трансформированные бактерии, растения, растительные клетки, ткани и семена. Кроме того, представлены способы получения полипептидов, кодирумых синтетическими нуклеотидами по изобретению.
Представлены изолированные молекулы нуклеиновых кислот и варианты таковых, кодирующие полипептиды устойчивости или толерантности к гербициду. Дополнительно, охвачены последовательности аминокислоты и варианты таковых, кодируемые полинуклеотидами, которые придают устойчивость или толерантность к гербициду. Настоящее изобретение представляет молекулы изолированных нуклеиновых кислот, содержащие нуклеотидную последовательность, представленную в SEQ ID NO: 1, 3 или 5, нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную в SEQ ID NO: 2, 4 или 6, нуклеотидную последовательность устойчивости к гербициду, депонированную в бактерии-хозяине в виде номеров доступа NRRL B-30888 или NRRL B-30949, так же, как и варианты и фрагменты таковых. Нуклеотидные последовательности, которые комплементарны нуклеотидным последовательностям по изобретению, или те, которые гибридизуются с последовательностями по изобретению, также охвачены.
Также представлены способы измерения ферментативной кинетической активности с применением флуорогенных субстратов.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фиг.1 показано выравнивание GRG23 ORF1 аминокислотной последовательности (SEQ ID NO:2) и GRG51 (SEQ ID NO:6) с Bacillus clausii (SEQ ID NO:7), Rubrobacer xylanophilus (SEQ ID NO:8), Escherichia coli (SEQ ID SEQ NO: 11), штаммом CP4 Agrobacterium sp. (SEQ ID NO:10) и Zea mays (SEQ ID NO:9).
На Фиг.2 показан график рассеяния ферментативной активности GRG23 (ось Y) как функции концентрации РЕР (ось X) при концентрациях глифосата 0, 3, 5 и 10 мМ.
На Фиг.3 показан график рассеяния Кm(app.) (ось Y) как функции концентрации глифосата (ось X). Точка пересечения оси X представляет Ki для глифосата.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящие изобретения теперь будут описаны более подробно со ссылкой на приложенные чертежи, в которых представлены несколько, но не все, варианты осуществления изобретений. Действительно, эти изобретения могут быть воплощены во многих различных формах и не должны быть рассмотрены как ограниченные вариантами осуществления, представленными здесь; скорее эти способы осуществления представлены так, чтобы это раскрытие удовлетворило применимые законные требования. Подобные номера на всем протяжении относятся к подобным элементам.
Множество модификаций и других вариантов осуществления изобретений, представленных здесь, будут понятны специалисту в области техники, к которой эти изобретения принадлежат, имеющих преимущество идеи изобретения, представленного в предшествующих описаниях и сопутствующих чертежах. Поэтому следует понимать, что изобретения не должны быть ограничены конкретными раскрытыми вариантами осуществления, и что модификации и другие варианты осуществления предназначены для включения в объем притязаний. Хотя здесь используются определенные термины, они применены только в типичном и описательном смысле, а не в целях ограничения.
Настоящее изобретение относится к композициям и способам регуляции в организмах устойчивости к гербициду, особенно у растений или растительных клеток. Способы включают трансформацию организмов нуклеотидными последовательностями, кодирующими ген устойчивости к глифосату по изобретению. Нуклеотидные последовательности по изобретению пригодны для получения растений, которые проявляют повышенную толерантность к гербицидному глифосату. Таким образом, представлены трансформированные бактерии, растения, растительные клетки, растительные ткани и семена. Композиции включают нуклеиновые кислоты и белки, имеющие отношение к толерантности к гербициду в микроорганизмах и растениях так же, как и трансформированные бактерии, растения, растительные ткани и семена. Раскрыты нуклеотидные последовательности гена устойчивости к глифосату (grg23 и grg51) и аминокислотные последовательности белков, кодируемых ими. Последовательности находят применение в конструировании экспрессионных векторов для последующей трансформации интересующих растений, в качестве зондов для выделения других генов устойчивости к глифосату, в качестве маркеров селекции, и т.п. Таким образом, под "геном устойчивости к глифосату по изобретению" подразумевается нуклеотидная последовательность, приведенная в SEQ ID NO:1 или 3, и варианты и фрагменты таковых (SEQ ID NO:5, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30 и 32), которые кодируют полипептид устойчивости или толерантности к глифосату. Аналогично, "полипептид устойчивости к глифосату по изобретению" представляет собой полипептид, обладающий аминокислотной последовательностью, приведенной в SEQ ID NO:2 или 4, и вариантами и фрагментами таковых (SEQ ID NO:6, 15, 17, 19, 21, 23, 25, 27, 29, 31 и 33), которые придают клетке-хозяину устойчивость или толерантность к глифосату.
Плазмиды, содержащие нуклеотидные последовательности устойчивости к гербициду по изобретению, депонированы в постоянной коллекции Коллекции Культур Службы сельскохозяйственных исследований, Северной Региональной Научно-исследовательской лаборатории (NRRL) 18 ноября 2005 под номером доступа NRRL B-30888 (grg23), и 26 июня 2006 под номером доступа NRRL B-30949 (grg51). Это депонирование будет поддерживаться в соответствии с Будапештским Договором о Международном Признании Депонирования Микроорганизмов для целей Патентной Процедуры. Это депонирование было сделано для удобства специалистов в области техники, и не допускается, что депонирование является необходимым по 35 своду законов США §112.
Под "глифосатом" подразумевается любая гербицидная форма N-фосфонометилглицина (включая любую соль такового) и другие формы, из которых получается анион глифосата in planta. "Белок устойчивости к гербициду" или белок, являющийся продуктом экспрессии "молекулы нуклеиновых кислот, кодирующей устойчивость к гербициду", включает белки, которые придают клетке способность быть толерантной к более высокой концентрации гербицида, чем клетки, которые не экспрессируют белок, или переносить определенную концентрацию гербицида в течение более длительного времени, чем клетки, которые не экспрессируют белок. "Белок устойчивости к глифосату" включает белок, который придает клетке способность оставаться толерантной к более высокой концентрации глифосата, чем клетки, которые не экспрессируют белок, или быть толерантной к определенной концентрации глифосата в течение более длинного промежутка времени, чем клетки, которые не экспрессируют белок. Под выражением "быть толерантным" или "толерантностью" подразумевается либо выживание, либо выполнение важнейших клеточных функций, таких как синтез белка и дыхание, таким способом, который с трудом отличим от необработанных клеток.
Изолированные молекулы нуклеиновых кислот и варианты и фрагменты таковых
Один аспект изобретения относится к изолированным молекулам нуклеиновых кислот, содержащим нуклеотидные последовательности, кодирующие белки и полипептиды устойчивости к гербициду или биологически активные части таковых так же, как и молекулы нуклеиновых кислот, подходящие для применения в качестве гибридизационных зондов для идентификации нуклеиновых кислот, кодирующих устойчивость к гербициду. Как используется в настоящем описании, термин "молекула нуклеиновой кислоты" предназначен включать молекулы ДНК (например, кДНК или геномной ДНК) и молекулы РНК (например, мРНК) и аналоги ДНК или РНК, полученные с использованием нуклеотидных аналогов. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной.
Нуклеотидные последовательности, кодирующие белки по настоящему изобретению, включают последовательности, приведенные в SEQ ID NO: 1, 3 и 5, нуклеотидную последовательность устойчивости к гербициду, введенную в бактериального хозяина под номерами доступа NRRL B-30888 и NRRL B-30949, и варианты, фрагменты, и комплементарные таковым. Под термином "комплементарный" подразумевается нуклеотидная последовательность, которая достаточно комплементарна данной нуклеотидной последовательности таким образом, что она может гибридизироваться с данной нуклеотидной последовательностью, образуя, таким образом, устойчивый дуплекс. Соответствующая аминокислотная последовательность для белка устойчивости к гербициду, кодируемая этими нуклеотидными последовательностями, приведена в SEQ ID NO:2, 4 или 6. Изобретение также охватывает молекулы нуклеиновых кислот, содержащие нуклеотидные последовательности, кодирующие неполноразмерные белки устойчивости к гербициду, и комплементарные таковым.
"Изолированная" или "очищенная" молекула нуклеиновой кислоты, или белка, или биологически активная часть таковой, является по существу свободной от другого клеточного материала или культуральной среды, если производится посредством рекомбинантных методик, или является по существу свободной от химических предшественников или других химических реагентов, если синтезируется химически. Предпочтительно, "изолированная" нуклеиновая кислота свободна от последовательностей (предпочтительно последовательностей, кодирующих белок), которые в природе примыкают к нуклеиновой кислоте (то есть, последовательности, расположенные на 5' и 3' концах нуклеиновой кислоты) в геномной ДНК организма, из которого получена нуклеиновая кислота. В целях изобретения термин "изолированная" применительно к молекулам нуклеиновых кислот исключает изолированные хромосомы. Например, в различных вариантах осуществления изолированная молекула нуклеиновой кислоты, кодирующая устойчивость к глифосату, может содержать меньше, чем приблизительно 5 т.п.н., 4 т.п.н., 3 т.п.н., 2 т.п.н., 1 т.п.н., 0,5 т.п.н или 0,1 т.п.н. нуклеотидной последовательности, которая в природе примыкает к молекуле нуклеиновой кислоты в геномной ДНК клетки, из которой получена нуклеиновая кислота. Белок устойчивости к гербициду, который по существу не содержит клеточный материал, включает получение белка, имеющего меньше, чем приблизительно 30%, 20%, 10% или 5% (сухой вес) белка устойчивости к не гербициду (также обозначаемый в настоящем описании как "загрязняющий белок").
Молекулы нуклеиновых кислот, которые являются фрагментами этих нуклеотидных последовательностей, кодирующих устойчивость к гербициду, также входят в объем настоящего изобретения. Под "фрагментом" понимается часть нуклеотидной последовательности, кодирующей белок устойчивости к гербициду. Фрагмент нуклеотидной последовательности может кодировать биологически активную часть белка устойчивости к гербициду, или это может быть фрагмент, который может быть использован в качестве гибридизационного зонда или ПЦР-праймера с использованием способов, раскрытых ниже. Молекулы нуклеиновых кислот, которые являются фрагментами нуклеотидной последовательности устойчивости к гербициду, содержат по меньшей мере приблизительно 15, 20, 50, 75, 100, 200, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050, 1100, 1150, 1200, 1250, 1300, 1350, 1400, 1450, 1500, 1550, 1600, 1650, 1700, 1750, 1800, 1850, 1900, 1950 последовательных нуклеотидов, или до числа нуклеотидов, присутствующих в полноразмерной нуклеотидной последовательности, кодирующей устойчивость к гербициду, раскрытой в настоящем описании (например, 1892 нуклеотидов для SEQ ID NO: 1, 1259 нуклеотидов для SEQ ID NO:3, и 1242 нуклеотидов для SEQ ID NO:5). Под "последовательными" нуклеотидами подразумеваются нуклеотидные остатки, которые являются непосредственно прилегающими друг к другу.
Фрагменты нуклеотидных последовательностей по настоящему изобретению главным образом будут кодировать фрагменты белка, которые сохраняют биологическую активность полноразмерного белка устойчивости к глифосату; то есть активность устойчивости к гербициду. Под выражением "сохраняет активность устойчивости к гербициду" подразумевается, что фрагмент будет иметь по меньшей мере приблизительно 30%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 70% или по меньшей мере приблизительно 80% активности устойчивости к гербициду полноразмерных белков устойчивости к глифосату, которые раскрыты в настоящем описании как SEQ ID NO:2, 4 или 6. Способы измерения активности устойчивости к гербициду известны в области техники. См., например, Патенты США №№ 4535060 и 5188642, каждый из которых включен в настоящее описание полностью посредством ссылки.
Фрагмент нуклеотидной последовательности, кодирующей устойчивость к гербициду, которая кодирует биологически активную часть белка по изобретению, будет кодировать по меньшей мере приблизительно 15, 25, 30, 50, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400 последовательных аминокислот, или вплоть до общего числа аминокислот в полноразмерном белке устойчивости к гербициду по настоящему изобретению (например, 436 аминокислот для SEQ ID NO:2, 413 аминокислот для SEQ ID NO: 4 и 413 аминокислот для SEQ ID NO:6).
Белки устойчивости к гербициду по настоящему изобретению кодируются нуклеотидной последовательностью, достаточно идентичной нуклеотидной последовательности SEQ ID NO:1, 3 или 5. Под термином "достаточно идентичная" подразумевают аминокислотную или нуклеотидную последовательность, которая имеет по меньшей мере приблизительно 60% или 65% идентичность последовательности, приблизительно 70% или 75% идентичность последовательности, приблизительно 80% или 85% идентичность последовательности, приблизительно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичность последовательности при сопоставлении с последовательностью сравнения с использованием одной из программ выравнивания, описанных в настоящем описании, с использованием стандартных параметров. Квалифицированный специалист в этой области техники признает, что эти значения могут быть подходящим образом приспособлены для определения соответствующих идентичностей белков, кодируемых двумя нуклеотидными последовательностями посредством принятия во внимание вырожденности кодонов, сходства аминокислот, расположения рамки считывания, и т.п.
Чтобы определить процент идентичности двух последовательностей аминокислот или двух нуклеиновых кислот, последовательности выравнивают с целью оптимального сравнения. Процент идентичности между двумя последовательностями является функцией числа идентичных положений, которые являются общими для последовательностей (т.е. процент идентичности = число идентичных положений/общее количество положений (например, перекрывающиеся положения) × 100). В одном варианте осуществления две последовательности имеют одинаковую длину. Процент идентичности между двумя последовательностями может быть определен с использованием методик, подобных описанным ниже, с или без допущения пропусков. При вычислении процента идентичности, как правило, подсчитывают точные совпадения.
Определение идентичности процента между двумя последовательностями может быть достигнуто с использованием математического алгоритма. Неограничивающим примером математического алгоритма, используемого для сравнения двух последовательностей, является алгоритм Karlin and Altschul (1990) Proc. Natl Acad. Sci. USA 87:2264-2268, измененный как в Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-5877. Такой алгоритм включен в BLASTN и программы BLASTX от Altschul et al. (1990) J. Mol. Biol. 215:403-410. BLAST нуклеотидные поиски могут быть выполнены программой BLASTN, число очков = 100, длина слова = 12, для получения нуклеотидных последовательностей, гомологичных GDC-подобным молекулам нуклеиновых кислот по изобретению. BLAST поиски белка могут быть выполнены программой BLASTX, число очков = 50, длина слова = 3, для получения аминокислотных последовательностей, гомологичных молекулам белка устойчивости к гербициду по изобретению. Чтобы получить, с целью сравнения, выравнивания с разрывами, может быть использован Gapped BLAST, как описано в Altschul et al. (1997) Nucleic Acid Res. 25:3389-3402. В качестве альтернативы для выполнения повторного поиска, который обнаруживает отдаленные сходства между молекулами, может быть применен PSI-Blast. См. Altschul et al. (1997) выше. При использовании BLAST, Gapped BLAST и PSI-BLAST программ можно использовать параметры по умолчанию соответствующих программ (например, BLASTX и BLASTN). См. www.ncbi.nlm.nih.gov. Другим неограничивающим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм ClustalW (Higgins et al. (1994) Nucleic Acid Res. 22:4673-4680). ClustalW сравнивает последовательности и выравнивает аминокислотные или ДНК-последовательности полностью, и таким образом может предоставить данные о консервативности последовательности всей аминокислотной последовательности. Алгоритм ClustalW используется в нескольких коммерчески доступных пакетах программ для анализа ДНК/аминокислот, таких как модуль ALIGNX Набор Программы Vector NTI (Корпорация Invitrogen, Carlsbad, CA). После выравнивания аминокислотных последовательностей ClustalW может быть определен процент аминокислотной идентичности. Неограничивающим примером программы, пригодной для анализа выравниваний ClustalW, является GeneDocТМ. GenedocТМ (Karl Nicholas) позволяет оценивать аминокислотное сходство (или ДНК) и идентичность между несколькими белками. Другим неограничивающим примером математического алгоритма, используемого для сравнения последовательностей, является алгоритм Myers и Miller (1988) CABIOS 4:11-17. Такой алгоритм включен в программу ALIGN (версия 2.0), которая является частью пакета программ для выравнивания последовательности GCG (доступный от Accelrys, Inc., Сан-Диего, Калифорния). Используя программу ALIGN для сравнения аминокислотных последовательностей, могут быть применены таблица веса остатков PAM120, штраф длины разрыва 12 и штраф разрыва 4.
Если не указано иначе, Версия 10 GAP, которая использует алгоритм Needleman и Wunsch (1970) J. Mol. Biol. 48 (3):443-453, будет использована для определения идентичности последовательности или сходства с использованием следующих параметров: % идентичности и % сходства для нуклеотидной последовательности с использованием GAP Weight 50 и Length Weight 3 и nwsgapdna.cmp матрицы очков; % идентичности или % сходства для аминокислотной последовательности с использованием GAP Weight и Length Weight 2, и программа начисления очков BLOSUM62. Также могут быть использованы эквивалентные программы. Под "эквивалентной программой" подразумевают любую программу сравнения последовательности, которая для любых двух рассматриваемых последовательностей, производит выравнивание, имеющее идентичные совпадения нуклеотидных остатков и идентичный процент идентичности последовательностей по сравнению с соответствующим выравниванием, произведенным посредством GAP Версии 10.
Изобретение также охватывает молекулы вариантов нуклеиновой кислоты. "Варианты" нуклеотидных последовательностей, кодирующих устойчивость к гербициду, включают те последовательности, которые кодируют белок устойчивости к гербициду, раскрытый в настоящем описании, но который консервативно отличается из-за вырожденности генетического кода, так же, как и те, которые достаточно идентичны, как обсуждено выше (например, SEQ ID NO:5, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30 и 32 являются вариантами SEQ ID NO:1). Природные аллельные варианты могут быть идентифицированы с использованием известных методов молекулярной биологии, таких как полимеразная цепная реакция (PCR) и методы гибридизации, как изложено в общих чертах ниже. Вариантные нуклеотидные последовательности также включают синтетически полученные нуклеотидные последовательности, которые были получены, например, с использованием сайт-направленного мутагенеза, но которые все еще кодируют белки устойчивости к гербициду, раскрытые в настоящем изобретении, как обсуждено ниже.
Вариантные белки, охваченные настоящим изобретением, являются биологически активными, если они сохраняют желательную биологическую активность природного белка, то есть активность устойчивости к гербициду. Под "сохраняет активность устойчивости к гербициду" понимают, что вариант будет иметь по меньшей мере приблизительно 30%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно 70% или по меньшей мере приблизительно 80% активности устойчивости к гербициду природного белка. Способы измерения активности устойчивости к гербициду хорошо известны в области техники. См., например, Патенты США №№ 4535060 и 5188642, каждый из которых включен в настоящее изобретение полностью посредством ссылки.
Квалифицированный специалист в дальнейшем оценит, что в нуклеотидные последовательности по изобретению могут быть введены изменения посредством мутации, таким образом приводящей к изменениям в аминокислотной последовательности кодируемых белков устойчивости к гербициду, без изменения биологической активности белков. Таким образом, изолированные вариантные молекулы нуклеиновых кислот могут быть созданы посредством введения одной или нескольких нуклеотидных замен, добавлений или делеций в соответствующую нуклеотидную последовательность, раскрытую в настоящем описании, таким образом, что в кодируемый белок будут введены одна или несколько аминокислотных замен, добавлений или делеций. Мутации могут быть введены посредством стандартных методик, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Такие вариантные нуклеотидные последовательности также входят в объем настоящего изобретения.
Например, консервативные аминокислотные замены могут быть сделаны в одном или нескольких предсказанных несущественных аминокислотных остатках. "Несущественным" аминокислотным остатком является остаток, который может быть изменен в последовательности белка дикого типа устойчивости к гербициду без изменений биологической активности, тогда как "существенный" аминокислотный остаток необходим для биологической активности. "Консервативная аминокислотная замена" является такой, в которой аминокислотный остаток замещен на аминокислотный остаток, имеющий сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, были определены в области техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолецин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Аминокислотные замены могут быть сделаны в неконсервативных областях, которые сохраняют функцию. Главным образом, такие замены не были бы сделаны для консервативных аминокислотных остатков или для аминокислотных остатков, находящихся в пределах консервативного мотива, где такие остатки являются существенными для активности белка. Однако квалифицированный специалист в этой области техники понял бы, что функциональные варианты могут иметь малые консервативные или неконсервативные изменения в консервативных остатках.
Lys-22, Arg-124, Asp-313, Arg-344, Arg-386 и Lys-411 являются консервативными остатками EPSP-синтазы из E. coli (Schönbrunn et al. (2001) Proc. Natl. Acad. Sci. USA 98:1376-1380). Консервативные остатки, важные для EPSP-синтазной активности, также включают Arg-100, Asp-242 и Asp-384 (Selvapandiyan et al. (1995) FEBS Letters 374:253-256). Arg-27 связывается с S3P (Shuttleworth et al. (1999) Biochemistry 38:296-302).
В качестве альтернативы вариантные нуклеотидные последовательности могут быть получены посредством введения мутаций случайным образом вдоль всей или части кодирующей последовательности, например, посредством мутагенеза насыщения, и полученные мутанты могут быть проверены на способность обеспечивать активность устойчивости к гербициду для идентификации мутантов, которые сохраняют активность. После мутагенеза кодируемый белок может быть экспрессирован рекомбинантным способом, и активность белка может быть определена с использованием стандартных методов тестирования.
Используя способы, такие как ПЦР, гибридизация и т.п., могут быть идентифицированы соответствующие последовательности устойчивости к гербициду, такие последовательности, которые имеют существенную идентичность с последовательностями по изобретению. См., например, Sambrook и Russell (2001) Molecular Cloning: Laboratory Manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY) и Innis, et al. (1990) PCR PRotocols: A Guide to Methods and Applications (Academic Press, St.-Louis, MO).
В способе гибридизации вся или часть нуклеотидной последовательности устойчивости к гербициду может быть использована для скринирования кДНК или геномных библиотек. Способы создания таких кДНК и геномных библиотек являются общеизвестными в области техники и раскрыты в Sambrook and Russell (2001) выше. Так называемые гибридизационные зонды могут быть фрагментами геномной ДНК, фрагментами кДНК, фрагментами РНК или другими олигонуклеотидами, и могут быть помечены обнаруживаемой группой, такой как 32P или любым другим обнаруживаемым маркером, таким как другие радиоизотопы, флуоресцентное соединение, фермент или кофактор фермента. Зонды для гибридизации могут быть получены посредством мечения синтетических олигонуклеотидов, основываясь на известной(ых) нуклеотидной(ых) последовательности(ях), кодирующей(их) устойчивость к гербициду, раскрытой(ых) в настоящем описании. Дополнительно могут быть использованы вырожденные праймеры, сконструированные на основе консервативных нуклеотидов или аминокислотных остатков в нуклеотидной последовательности или кодируемой аминокислотной последовательности. Зонд, как правило, содержит область нуклеотидной последовательности, которая гибридизуется в жестких условиях с по меньшей мере приблизительно 12, по меньшей мере приблизительно 25, по меньшей мере приблизительно 50, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400, 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600 или 1800 последовательными нуклеотидами нуклеотидной последовательности(ей), кодирующей устойчивость к гербициду по изобретению, или фрагмента или варианта таковых. Способы для получения зондов для гибридизации являются общеизвестными в области техники и раскрыты в Sambrook and Russell (2001) выше, и Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY), каждый из которых включен в настоящее описание посредством ссылки.
Например, полная последовательность устойчивости к гербициду, раскрытая в настоящем описании, или одна или более частей таковой, могут быть использованы в качестве зонда, способного специфически гибридизоваться с соответствующим последовательностями устойчивости к гербициду и с матричными РНК. Для достижения специфической гибридизации при различных условиях, такие зонды включают последовательности, которые являются уникальными и представляют собой по меньшей мере приблизительно 10 нуклеотидов в длину и по меньшей мере приблизительно 20 нуклеотидов в длину. Такие зонды могут быть использованы для амплификации соответствующих последовательностей устойчивости к гербициду из выбранного организма посредством ПЦР. Эта методика может быть использована для выделения дополнительных кодирующих последовательностей из желательного организма или в качестве диагностического теста для определения присутствия кодирующих последовательностей в организме. Методы гибридизации включают гибридизационное скринирование высеянных на чашки библиотек ДНК (или бляшек, или колоний; см., например, Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY).
Гибридизация таких последовательностей может быть выполнена в жестких условиях. Под "жесткими условиями" или "жесткими условиями гибридизации" подразумевают условия, в которых зонд гибридизуется со своей последовательностью-мишенью в детектируемо большей степени, чем с другими последовательностями (например, по меньшей мере с 2-кратным превышением фона). Жесткие условия являются зависимыми от последовательности и будут различны при различных обстоятельствах. Посредством управления жесткостью гибридизации и/или условиями промывки последовательности-мишени, которые на 100% комплементарны зонду, могут быть идентифицированы (homologous probing). В качестве альтернативы, жесткие условия могут быть отрегулированы для позволения некоторого несоответствия в последовательностях так, чтобы были обнаружены более низкие степени сходства (heterologous probing). Обычно зонд составляет по длине меньше, чем приблизительно 1000 нуклеотидов, или меньше, чем приблизительно 500 нуклеотидов в длину.
Как правило, жесткими условиями будут такие, при которых концентрация соли меньше, чем приблизительно 1,5 М ион Na, как правило, от приблизительно 0,01 до 1,0 М концентрации иона Na (или других солей) при pH от 7,0 до 8,3 и температуре по меньшей мере приблизительно 30°C для коротких зондов (например, от 10 до 50 нуклеотидов) и по меньшей мере приблизительно 60°C для длинных зондов (например, больше, чем 50 нуклеотидов). Жесткие условия могут также быть достигнуты добавлением агентов для дестабилизации, таких как формамид. Иллюстративные условия низкой жесткости включают гибридизацию в буферном растворе 30%-35%-ного формамида, 1 М NaCl, 1%-ной SDS (натрия додецилсульфат) при 37°C, и промывку в 1X-2X SSC (20X SSC = 3,0 М NaCl/0,3 М тринатрия цитрат) при 50-55°C. Иллюстративные условия умеренной жесткости включают гибридизацию в 40%-45%-ном формамиде, 1,0 М NaCl, 1%-ном SDS при 37°C, и промывку в 0,5X-1X SSC при 55-60°C. Иллюстративные условия высокой жесткости включают гибридизацию в 50%-ном формамиде, 1 М NaCl, 1%-ном SDS при 37°C, и промывку в 0,1X SSC при 60-65°C. Необязательно, буферы для промывки могут содержать от приблизительно 0.1% до приблизительно 1% SDS. Продолжительность гибридизации составляет обычно менее чем приблизительно 24 часа, обычно от приблизительно 4 до приблизительно 12 часов.
Специфичность, как правило, является функцией постгибридизационных промывок, критическими факторами являются ионная сила и температура раствора для заключительной промывки. Для ДНК-ДНК гибридов, Тm может быть приближена к уравнению Meinkoth and Wahl (1984) Anal. Biochem. 138:267-284: Tm = 81,5°C + 16,6 (log M), + 0,41 (%GC) - 0,61 (% форм.) - 500/L; где М является молярностью одновалентных катионов, %GC - процент нуклеотидов гуанозина и цитозина в ДНК, % форм. - процент формамида в растворе гибридизации, и L - длина гибрида в парах азотистых оснований. Тm является температурой (при определенной ионной силе и pH), при которой 50% комплементарной последовательности-мишени гибридизуется с идеально соответствующей пробой. Тm уменьшается приблизительно на 1°C с каждым 1% несоответствия; таким образом, Тm, гибридизация и/или условия промывки могут быть отрегулированы для гибридизации с последовательностями желательной идентичности. Например, если требуется найти последовательности с ≥ 90%-ной идентичностью, Tm может быть уменьшена на 10°C. Обычно жесткие условия выбираются, чтобы быть примерно на 5°C ниже, чем температура точки плавления (Тm) для специфической последовательности и комплементарной ей при определенной ионной силе и pH. Однако при очень жестких условиях гибридизация и/или промывка могут проводиться при 1, 2, 3 или 4°C ниже, чем температура точки плавления (Тm); при умеренно жестких условиях гибридизация и/или промывка могут проводиться при 6, 7, 8, 9, или 10°C ниже, чем температура точки плавления (Тm); в условиях низкой жесткости гибридизация и/или промывка могут проводиться при 11, 12, 13, 14, 15 или 20°C ниже, чем температура точки плавления (Тm). Применяя уравнение, гибридизацию и составы промывки и желательную Тm, среднему специалисту понятно, что разнообразие в жесткости гибридизации и/или растворов промывки по существу описано. Если желательная степень несоответствия приводит к Тm меньше, чем 45°C (водный раствор) или 32°C (раствор формамида), предпочтительно увеличить концентрацию SSC так, чтобы можно было использовать более высокую температуру. Обширное руководство по гибридизации нуклеиновых кислот найдено в Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular Biology - Hybridization with Nucleic Acid Probes, Part I, Chapter 2 (Elsevier, New York); и Ausubel et al., eds. (1995) Current Protocols in Molecular Biology, Chapter 2 (Greene Publishing and Wiley-Interscience, New York). См. Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY).
Изолированные белки и варианты и фрагменты таковых
Белки устойчивости к гербициду также охвачены объемом настоящего изобретения. Под "белком устойчивости к гербициду" или "белком толерантности к гербициду" подразумевают белок, имеющий аминокислотную последовательность, представленную в SEQ ID NO:2, 4 или 6. Фрагменты, биологически активные части и варианты таковых также представлены и могут быть использованы для осуществления на практике способов по настоящему изобретению.
"Фрагменты" или "биологически активные части" включают полипептидные фрагменты, содержащие часть аминокислотной последовательности, кодирующей белок устойчивости к гербициду, как приведено в SEQ ID N0:2, 4, или 6, и которые сохраняют активность устойчивости к гербициду. Биологически активная часть белка устойчивости к гербициду может быть полипептидом, который, например, имеет 10, 25, 50, 100 или больше аминокислот в длину. Такие биологически активные части могут быть получены посредством рекомбинантных методик и оценены на активность устойчивости к гербициду. Способы измерения активности устойчивости к гербициду хорошо известны в области техники. См., например, Патенты США №№ 4535060 и 5188642, каждый из которых включен в настоящее описание полностью посредством ссылки. Как используется в настоящем описании, фрагмент содержит по меньшей мере 8 последовательных аминокислот SEQ ID NO: 2, 4 или 6. Однако изобретение охватывает другие фрагменты, такие как, например, любой фрагмент в белке, больший, чем приблизительно 10, 20, 30, 50, 100, 150, 200, 250, 300, 350 или 400 аминокислот.
Под "вариантами" подразумеваются белки или полипептиды, имеющие аминокислотную последовательность, которая по меньшей мере приблизительно на 60%, 65%, по меньшей мере приблизительно на 70%, 75%, по меньшей мере приблизительно на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, идентична аминокислотой последовательности SEQ ID NO:2, 4, или 6 (например, SEQ ID NO:6, 15, 17, 19, 21, 23, 25, 27, 29, 31 и 33 являются вариантами SEQ ID NO:2). Варианты также включают полипептиды, кодируемые молекулой нуклеиновой кислоты, которая гибридизуется с молекулой нуклеиновой кислоты SEQ ID NO:1, 3 или 5, или комплементарна таковым, в жестких условиях. Варианты включают полипептиды, которые отличаются по аминокислотной последовательности вследствие мутагенеза. Вариантные белки, охваченные настоящим изобретением, являются биологически активными, так что они продолжают обладать желательной биологической активностью исходного белка, то есть сохраняют активность устойчивости к гербициду. Способы измерения активности устойчивости к гербициду хорошо известны в области техники. См., например, Патенты США №№ 4535060 и 5188642, каждый из которых включен в настоящее описание полностью посредством ссылки.
Бактериальные гены, такие как ген grg23 или grg51 по настоящему изобретению, весьма часто обладают множественными инициирующими кодонами метионина вблизи начала открытой рамки считывания. Часто инициирование трансляции в одном или нескольких из этих старт-кодонов приведет к образованию функционального белка. Эти старт-кодоны могут включать ATG-кодоны. Однако бактерии, такие как Bacillus sp., также узнают кодон GTG как старт-кодон, и белки, которые начинают транслироваться в кодонах GTG, содержат метионин в первой аминокислоте. Кроме того, это не часто определяется a priori, какие из этих кодонов используются в природе в бактерии. Таким образом, понятно, что использование одного из дополнительных кодонов метионина может привести к образованию вариантов grg23 или grg51, которые обладают устойчивостью к гербициду. Эти белки устойчивости к гербициду входят в объем настоящего изобретения и могут быть использованы в способах по настоящему изобретению.
Также охвачены антитела к полипептидам по настоящему изобретению, или к вариантам или фрагментам таковых. Способы получения антител хорошо известны в области техники (см., например, Harlow and Lane (1988) Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory, Cold Spring Harbor, NY); Патент США № 4 196 265).
Измененные или улучшенные варианты
Признано, что последовательность ДНК grg23 или grg51 может быть изменена посредством различных способов, и что эти изменения могут привести к ДНК последовательностям, кодирующим белки с аминокислотными последовательностями, отличными от тех, которые кодируются grg23 или grg51. Этот белок может быть изменен различными путями, включая аминокислотные замены, делеции, усечения и инсерции. Способы таких манипуляций являются общеизвестными в области техники. Например, варианты аминокислотной последовательности белка GRG23 или GRG51 могут быть получены посредством мутаций в ДНК. Это также может быть достигнуто посредством одной из нескольких форм мутагенеза и/или посредством направленной эволюции. В некоторых аспектах изменения, кодируемые в аминокислотной последовательности, не будут по существу затрагивать функцию белка. Такие варианты будут обладать желательной активностью устойчивости к гербициду. Однако понятно, что способность GRG23 или GRG51 придавать устойчивость к гербициду может быть улучшена посредством использования таких методик в композициях по настоящему изобретению. Например, GRG23 или GRG51 могут экспрессироваться в клетках-хозяевах, которые проявляют высокие уровни ошибочного включения во время репликации ДНК, таких как XL-1 Red (Stratagene, La Jolla, СА). После воспроизведения в таких штаммах, ДНК grg23 или grg51 может быть изолирована (например, посредством получения плазмидной ДНК или посредством ПЦР-амплификации и клонирования получающегося ПЦР-фрагмента в вектор) и культивироваться в немутагенных штаммах. Клоны, содержащие мутации в grg23 или grg51, могут быть идентифицированы посредством измерения улучшенной устойчивости к гербициду, такому как глифосат, например, посредством выращивания клеток на увеличивающихся концентрациях глифосата и тестирования для поиска клонов, которые обладают толерантностью к увеличивающимся концентрациям глифосата.
В качестве альтернативы, могут быть сделаны изменения с последовательностью белка многих белков с амино- или карбоксиконца, по существу не затрагивая активность. Эти изменения могут включать инсерции, делеции или изменения, вводимые посредством современных молекулярные способов, таких как ПЦР, включая ПЦР-амплификацию, которая изменяет или удлиняет последовательность, кодирующую белок, посредством включения последовательностей, кодирующих аминокислоты в олигонуклеотидах, используемых при ПЦР-амплификации. В качестве альтернативы добавленные последовательности белка могут включать полные последовательности, кодирующие белок, такие как те, которые обычно применяются в области техники для получения гибридных белков. Такие гибридные белки часто применяются для (1) увеличения экспрессии интересующего белка; (2) введения связывающего домена, ферментативной активности или эпитопа для облегчения либо очищения белка, либо обнаружения белка или других экспериментальных применений, известных в области техники; или (3) адресной секреции или трансляции белка в субклеточную органеллу, такую как периплазматическое пространство грамотрицательных бактерий или эндоплазматическая сеть эукариотических клеток, последняя из которых часто приводит к гликозилированию белка.
Вариантные нуклеотидные и аминокислотные последовательности по настоящему изобретению также охватывают последовательности, полученные посредством мутагенных и рекомбинантных процедур, таких как перетасовка ДНК. С помощью такой процедуры одна или несколько различных кодирующих областей белков устойчивости к гербициду может быть использована для создания нового белка устойчивости к гербициду, обладающего желательными свойствами. Таким образом, библиотеки рекомбинантных полинуклеотидов формируются из числа родственных полинуклеотидных последовательностей, включающих области последовательности, которые обладают существенной идентичностью последовательности и могут быть гомологично рекомбинированы in vitro или in vivo. Например, применяя этот подход, фрагменты последовательности, кодирующей интересующий домен, могут быть перетасованы между геном устойчивости к гербициду по изобретению и другими известными генами устойчивости к гербициду для получения нового гена, кодирующего белок с улучшенным интересующим свойством, таким как увеличенная активность устойчивости к глифосату. Стратегии для такой перетасовки ДНК известны в области техники. См., например, Stemmer (1994) Proc. Natl. Acad. Sci. USA 91:10747-10751; Stemmer (1994) Nature 370:389-391; Crameri et al. (1997) Nature Biotech. 15:436-438; Moore et al. (1997) J. Mol. Biol. 272:336-347; Zhang et al. (1997) Proc. Natl. Acad. Sci. USA 94:4504-4509; Crameri et al. (1998) Nature 391 :288-291; и Патенты США №№ 5605793 и 5837458.
Трансформация бактериальных или растительных клеток
Представленное в настоящем описании является новыми изолированными генами, которые придают устойчивость к гербициду. Также представленное является аминокислотными последовательностями белков GRG23 и GRG51. Белок, образующийся при трансляции этого гена, позволяет клеткам функционировать в присутствии концентраций гербицида, которые иначе были бы токсичны для клеток, включая растительные клетки и бактериальные клетки.
В одном аспекте изобретения ген grg23 или grg51 применим в качестве маркерного гена для оценки трансформации бактериальных или растительных клеток.
Посредством конструирования grg23 или grg51 для (1) экспрессии с бактериальным промотором, стимулирующим транскрипцию в тестируемом организме, (2) правильной трансляции для образования неизмененного пептида GRG23 или GRG51, и (3) перемещения клеток в другую токсичную концентрацию гербицида, могут быть идентифицированы клетки, которые были трансформированы ДНК посредством их устойчивости к гербициду. Под "промотором" подразумевают последовательность нуклеиновой кислоты, которая функционирует, чтобы направить транскрипцию расположенной далее по направлению кодирующей последовательности. Промотор, вместе с другими последовательностями нуклеиновой кислоты, которые регулируют транскрипцию и трансляцию (также называемые "регуляторные последовательности"), необходим для экспрессии интересующей последовательности ДНК.
Трансформация бактериальных клеток осуществляется посредством одной из нескольких методик, известных в области техники, включая, но не ограничиваясь электропорацией или химической трансформацией (См., например, Ausubel (ed.) (1994) Current Protocols in Molecular Biology (John Wiley and Sons, Inc., Indianapolis, IN)). Маркеры, несущие устойчивость к токсичным веществам, применяются для идентификации трансформированных клеток (захвативших и экспрессирующих тестируемую ДНК) от нетрансформированных клеток (тех, которые не содержат или не экспрессируют тестируемую ДНК). В одном аспекте изобретения, ген grg23 или grg51 применяется как маркерный ген для оценки трансформации бактериальных или растительных клеток.
Трансформация растительных клеток может осуществляться подобным способом. Под "растением" подразумевают целые растения, органы растения (например, листья, стебли, корни, и т.д.), семена, растительные клетки, побеги, эмбрионы и потомство такового. Растительные клетки могут быть дифференцироваными или недифференцироваными (например, каллюс, суспензионные культуры клеток, протопласты, клетки листьев, клетки корня, клетки флоэмы, пыльца). "Трансгенные растения" или "трансформированные растения" или "стабильно трансформированные" растения, клетки или ткани относятся к растениям, которые вобрали или интегрировали последовательности экзогенной нуклеиновой кислоты или фрагменты ДНК в растительную клетку. Под "стабильной трансформацией" подразумевают, что нуклеотидная конструкция, введенная в растение, интегрирует в геном растения и способна к тому, чтобы быть унаследованной потомством такового.
Ген grg23 или grg51 по изобретению может быть модифицирован для получения или усиления экспрессии в растительных клетках. Последовательности устойчивости к гербициду по изобретению могут быть предоставлены в экспрессионных кассетах для экспрессии в интересующем растении. "Кассета для экспрессии в растении" включает конструкции ДНК, которые способны приводить к экспрессии белка с открытой рамки считывания в растительной клетке. Кассета будет включать в 5'-3' направлении транскрипции, область инициации транскрипции (то есть промотор), функционально связанную с последовательностью ДНК по изобретению, и область терминации транскрипции и трансляции (то есть область терминации), функционирующие в растениях. Кассета может дополнительно содержать по меньшей мере один дополнительный ген для котрансформации в организм, такой как ген маркера селекции. В качестве альтернативы дополнительный ген(ы) может быть предоставлен в нескольких кассетах для экспрессии. Такая кассета для экспрессии предоставлена с множеством рестриктных сайтов для инсерции последовательности устойчивости к гербициду под транскрипционной регуляцией регуляторных областей.
Промотор может быть нативным, или аналогичным, или инородным, или гетерологичным, растению-хозяину и/или к последовательности ДНК по изобретению. Дополнительно, промотор может быть встречающейся в природе последовательностью или, в противоположность, синтетической последовательностью. Если промотор является "нативным" или "гомологичным" растению-хозяину, это означает, что промотор происходит из нативного растения, в которое введен промотор. Если промотор является "инородным" или "гетерологичным" последовательности ДНК по изобретению, это означает, что промотор не происходит или не является встречающимся в природе промотором, функционально связанным с последовательностью ДНК по изобретению. "Гетерологичный" обычно относится к последовательностям нуклеиновой кислоты, которые не являются эндогенными для клетки или частью нативного генома, в котором они находятся, и были добавлены к клетке посредством инфекции, трансфекции, микроинъекции, электропорации, микропроекции или т.п. Под "функционально связанный" подразумевают функциональную связь между промотором и второй последовательностью, если последовательность промотора инициирует и опосредует транскрипцию с последовательности ДНК, соответствующей второй последовательности. Обычно “функционально связанный” означает, что связанные последовательности нуклеиновой кислоты являются последовательными и при необходимости присоединения двух кодирующих белок последовательных областей последовательны в одной и той же самой рамке считывания.
Часто такие конструкции будут также содержать 5' и 3' нетранслируемые области. Такие конструкции могут содержать "сигнальную последовательность" или "лидерную последовательность" для облегчения котрансляции или посттрансляционного транспорта интересующего пептида в определенные внутриклеточные структуры, такие как хлоропласт (или другой пластид), эндоплазматическая сеть или аппарат Гольджи, или быть секретированным. Например, ген может в результате конструирования содержать сигнальный пептид для облегчения передачи пептида в эндоплазматическую сеть. Под "сигнальной последовательностью" подразумевают последовательность, которая известна или, как считают, приводит к котрансляционному или посттрансляционному транспорту пептида через клеточную мембрану. У эукариот этот транспорт обычно вовлекает секрецию в аппарат Гольджи, с некоторым гликозилированием. Под "лидерной последовательностью" понимают любую последовательность, которая при трансляции приводит к аминокислотной последовательности, достаточной для запуска котрансляционного транспорта пептидной цепи в субклеточную органеллу. Таким образом, указанное выше включает транспорт, направленный лидерной последовательностью, и/или гликозилирование посредством прохода в эндоплазматическую сеть, проходов к вакуолям, пластидам, включая хлоропласты, митохондрии и т.п. Кассета для экспрессии в растении может также быть составлена таким образом, чтобы содержать интрон, процессинг мРНК которого необходим для экспрессии.
Под "3' нетранслируемой областью" подразумевают нуклеотидную последовательность, расположенную по ходу транскрипции кодирующей последовательности. Последовательности сигнала полиаденилирования и другие последовательности, кодирующие регуляторные сигналы, способные воздействовать на добавление полиадениловокислых хвостов к 3-концу предшественника мРНК, являются 3' нетранслируемыми областями. Под "5' нетранслируемой областью" понимают нуклеотидную последовательность, расположенную против хода трансткрипции кодирующей последовательности.
Другие расположенные против хода транскрипции или по ходу транскрипции нетранслируемые элементы включают энхансеры. Энхансерами являются нуклеотидные последовательности, которые приводят к увеличению экспрессию промоторной области. Энхансеры известны в области техники и включают, но не ограничены, область энхансера SV40 и энхансерный элемент 35S.
Область терминации может быть нативной по отношению к области инициации транскрипции, может быть нативной по отношению к последовательности устойчивости к гербициду по настоящему изобретению, или может быть получена из другого источника. Удобные области терминации доступны из Ti-плазмиды A. tumefaciens, такие как области терминации октопинсинтазы и нопалинсинтазы. См. также Guerineau et al. (1991) Mol. Gen. Genet. 262:141-144; Proudfoot (1991) Cell 64:671-674; Sanfacon et al. (1991) Genes Dev. 5:141-149; Mogen et al. (1990) Plant Cell 2:1261-1272; Munroe et al. (1990) Gene 91:151-158; Ballas et al. (1989) Nucleic Acids Res. 17:7891-7903; и Joshi et al. (1987) Nucleic Acid Res. 25 15:9627-9639.
По необходимости, ген(ы) может быть оптимизирован для увеличения экспрессии в трансформированной клетке-хозяине. Таким образом, гены могут быть синтезированы, применяя кодоны, предпочтительные для клетки-хозяина, для улучшенной экспрессии, или могут быть синтезированы, применяя кодоны, предпочтительные для хозяина, используя повторы. Обычно GC содержание гена будет увеличено. См., например, Campbell and Gowri (1990) Plant Physiol. 92:1-11 для обсуждения кодонов, предпочтительных для хозяина. Способы синтезирования генов, предпочтительных для хозяина, являются широко известными в области техники. См., например, Патенты США №№ 6320100; 6075185; 5380831 и 5436391, Опубликованные заявки США №№ 20040005600 и 20010003849, и Murray et al. (1989) Nucleic Acids Res. 17:477-498, включенные в настоящее описание посредством ссылки.
В одном варианте осуществления интересующие нуклеиновые кислоты нацелены для экспрессии в хлоропласт. В данной способе, если интересующая нуклеиновая кислота непосредственно не вставлена в хлоропласт, кассета для экспрессии будет дополнительно содержать нуклеиновую кислоту, кодирующую транзитный пептид для направления интересующего генетического продукта в хлоропласты. Такие транзитные пептиды известны в области техники. См., например, Von Heijne et al. (1991) Plant Mol. Biol. Rep. 9:104-126; Clark et al. (1989) J. Biol. Chem. 264:17544-17550; Della-Cioppa et al. (1987) Plant Physiol. 84:965-968; Romer et al. (1993) Biochem. Biophys. Res. Commun. 196:1414-1421; и Shah et al. (1986) Science 233:478-481.
Интересующие нуклеиновые кислоты, которые будет нацелены в хлоропласт, могут быть оптимизированы для экспрессии в хлоропласте, принимая во внимание различия в использовании кодонов между ядром растения и этой органеллой. Таким образом, интересующие нуклеиновые кислоты могут быть синтезированы, применяя кодоны, предпочтительные для хлоропласта. См., например, Патент США № 5380831, включенный в настоящее описание посредством ссылки.
Как правило, данная "кассета для экспрессии в растении" будет вставлена в "вектор для трансформации растения". Под "вектором для трансформации" подразумевают молекулу ДНК, которая необходима для эффективной трансформации клетки. Такая молекула может состоять из одной или нескольких кассет для экспрессии, и может быть организована в более чем одну "векторную" молекулу ДНК. Например, двойные векторы являются векторами для трансформации растения, которые используют два непоследовательных ДНК-вектора для кодирования всех необходимых цис- и транс-действующих функций для трансформации растительных клеток (Hellens and Mullineaux (2000) Trends in Plant Science 5:446-451). "Вектор" относится к конструкции нуклеиновой кислоты, сконструированной для передачи между различными клетками-хозяевами. "Вектор для экспрессии" относится к вектору, который способен внедряться, интегрировать и экспрессировать гетерологичные последовательности ДНК или фрагменты в чужеродной клетке.
Этот вектор для трансформации растения может включать один или несколько ДНК-векторов, необходимых для достижения трансформации растения. Например, обычной практикой в области техники является использование векторов для трансформации растения, которые состоят из более чем одного последовательного ДНК-сегмента. Эти векторы часто упоминаются в области техники как "двойные векторы". Двойные векторы так же, как и векторы со вспомогательными плазмидами, чаще всего применяются для Agrobacterium-опосредованной трансформации, когда необходимы весьма большие размер и сложность сегментов ДНК для достижения эффективной трансформации, и выгодно разделить функции между отдельными молекулами ДНК. Двойные векторы, как правило, содержат плазмидный вектор, который содержит цис-действующие последовательности, необходимые для T-ДНК перемещения (как, например, левую границу и правую границу), маркер селекции, который сконструирован так, чтобы быть способным к экспрессии в растительной клетке, и интересующий ген (ген, сконструированный так, чтобы быть способным к экспрессии в растительной клетке, для которой желательно образование трансгенных растений). Также на этом плазмидном векторе присутствуют последовательности, необходимые для репликации в бактерии. Цис-действующие последовательности устроены таким образом, чтобы обеспечивать эффективную передачу в растительные клетки и экспрессию в них. Например, ген маркера селекции и интересующий ген расположены между левой и правой границами. Часто второй плазмидный вектор содержит транс-действующие факторы, которые опосредуют Т-ДНК передачу от Agrobacterium к растительным клеткам. Эта плазмида часто содержит факторы вирулентности (Vir-гены), которые позволяют инфицировать растительные клетки посредством Agrobacterium, и перенос ДНК посредством разрезания в последовательностях границы и vir-опосредованный перенос ДНК, как это понимают в области техники (Hellens and Mullineaux (2000) Trends in Plant Science, 5:446-451). Несколько типов штаммов Agrobacterium (например, LBA4404, GV3101, EHA101, EHA105, и т.д.), могут быть использованы для трансформации растения. Второй плазмидный вектор не является необходимым для трансформации растений посредством других способов, таких как микропроецирование, микроинъекция, электропорация, полиэтиленгликоль, и т.д.
Трансформация растения
Способы по изобретению затрагивают введение нуклеотидной конструкции в растение. Под "введением" подразумевают предоставление растению нуклеотидной конструкции таким образом, что конструкция получает доступ внутрь клетки растения. Способы по изобретению не требуют использования конкретного способа введения растению нуклеотидной конструкции, если нуклеотидная конструкция получает доступ по меньшей мере внутрь одной клетки растения. Способы введения нуклеотидных конструкций в растения известны в области техники, включая, но не ограничиваясь способами стабильной трансформации, способами временной трансформации и вирус-опосредованными способами.
В общем, способы трансформации растений затрагивают перенос гетерологичной ДНК в растительные клетки-мишени (например, незрелые или зрелые эмбрионы, суспензионные культуры, недифференцированный каллюс, протопласты, и т.д.), после чего посредством применения максимального порогового уровня подходящей селекции (в зависимости от гена маркера селекции и в этом случае "глифосата") восстанавливают трансформированные растительные клетки из группы нетрансформированной клеточной массы. Эксплантаты, как правило, переносятся свежему доступу той же самой среды и культивируются обычным способом. Впоследствии трансформированные клетки дифференцируются в побеги после перемещения в среду для регенерации, дополненную максимальным пороговым уровнем вещества селекции (например, "глифосат"). Побеги затем переносят в селективную среду для выращивания растений для восстановления корневых побегов или саженцев. Трансгенный саженец далее вырастает в зрелые растения и производит плодородные семена (например, Hiei et al. (1994) The Plant Journal 6:271-282; Ishida et al. (1996) Nature Biotechnology 14:745-750). Эксплантаты, как правило, переносят свежими в ту же самую среду и культивируют обычным способом. Общее описание методов и способов получения трансгенных растений найдено в Ayres and Park (1994) Critical Reviews in Plant Science 13:219-239 и Bommineni and Jauhar (1997) Maydica 42:107-120. Поскольку трансформированный материал содержит много клеток; и трансформированные и нетрансформированные клетки присутствуют в любой части обработанного каллюса-мишени или ткани, или группы клеток. Способность уничтожать нетрансформированные клетки и позволять трансформированным клеткам размножаться дает культуры трансформированных растений. Часто способность удалить нетрансформированные клетки является ограничением для быстрого восстановления трансформированных растительных клеток и успешному образованию трансгенных растений. Для подтверждения присутствия интегрированного гетерологичного интересующего гена в геноме трансгенного растения могут быть применены молекулярные и биохимические способы.
Получение трансгенных растений может быть выполнено посредством одного из нескольких способов, включая, но не ограничиваясь введением гетерологичной ДНК посредством Agrobacterium в растительные клетки (Agrobacterium-опосредованная трансформация), бомбардировка растительных клеток гетерологичной чужеродной ДНК, прикрепленной к частицам, и различными другими направленными способами без частиц (например, Hiei et al. (1994) The Plant Journal 6:271-282; Ishida et al. (1996) Nature Biotechnology 14:745-750; Ayres and Park (1994) Critical Reviews in Plant Science 13:219-239; Bommineni and Jauhar (1997) Maydica 42:107-120), для переноса ДНК.
Способы трансформации хлоропластов хорошо известны в области техники. См., например, Svab et al. (1990) Proc. Natl. Acad. Sci. USA 87:8526-8530; Svab and Maliga (1993) Proc. Natl. Acad. Sci. USA 90:913-917; Svab and Maliga (1993) EMBO J. 12:601-606. Способ основан на доставке с помощью “ружья” ДНК-частиц, содержащих маркер селекции, адресующей ДНК к геному пластид посредством гомологичной рекомбинации. Помимо этого, трансформация пластид может быть достигнута посредством трансактивации молчащего трансгена в пластиде посредством ткане-предпочтительной экспрессии РНК полимеразы, кодируемой ядре и направленной в пластиды. Такая система была упомянута в McBride et al. (1994) Proc. Natl. Acad. Sci. USA 90:5873-5877.
Клетки, которые были трансформированы, могут быть выращены в растения в соответствии с общепринятыми способами. См., например, McCormick et al. (1986) Plant Cell Reports 5:81-84. Эти растения затем могут быть выращены и опылены либо тем же самым трансформированным штаммом, либо отличными штаммами, и полученный гибрид имеет конститутивную экспрессию желательной идентифицированной фенотипической особенности. Два или несколько поколений могут быть выращены для уверенности в том, что экспрессия желательной фенотипической особенности стабильно поддерживается и наследуется, а затем собирают семена для подтверждения, что экспрессия желательной фенотипической особенности была достигнута. Таким образом, настоящее изобретение предоставляет трансформированные семена (также называемые "трансгенные семена"), обладающие нуклеотидной конструкцией по изобретению, например, кассетой для экспрессии по изобретению, стабильно включенной в их геном.
Измерение EPSPS-активности
В одном варианте осуществления настоящего изобретения EPSPS-фермент устойчивости к глифосату обладает Кm для фосфоенолпирувата (РЕР) между приблизительно 1 и приблизительно 15O мкМ, включая приблизительно 2 мкМ, приблизительно 3, 4, 5, 6, 7, 8, 9, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130 или приблизительно 140 мкМ, и Ki (глифосат)/Кm (РЕР) между приблизительно 500 и приблизительно 1000, приблизительно 550, приблизительно 600, 650, 700, 750, 800, 850, 900, 950 или до приблизительно 1000. Как используется в настоящем описании, Km и Ki измерены в условиях, в которых фермент подчиняется кинетике Михаэлиса-Ментена, рН приблизительно 7. В одной неограничивающей методике измерения используется фермент в очищенном виде в хлориде калия и буфере HEPES при рН 7 при комнатной температуре и используется концентрация глифосата от 0 до 10 мМ.
Кинетическая активность EPSPS может быть измерена, например, посредством измерения высвобождения фосфата, который образуется во время катализа субстрата EPSPS (например, РЕР и S3P) с получением последующего продукта реакции (например, 5-енолпирувил-3-фосфошикимовой кислоты), используя флуоресцентный тест, описанный Vazquez et al. (2003) Anal. Biochem. 320 (2):292-298. Этот тест основан на окислении нефлуоресцентного соединения N-ацетил-3,7-дигидроксифеноксацина (Amplex® Red, Invitrogen, Carlsbad, CA) во флуоресцентное соединение резоруфина посредством перекиси водорода (Zhou и Panchuk-Voloshina (1997) Anal. Biochem. 253:169-174). Реакция основывается на использовании фосфата пуриннуклеозидфосфорилазы (PNP), ксантиноксидазы (XOD) и пероксидазы хрена (HRP). Высвобождение фосфата связано с уровнем флюоресценции, которая образуется при преобразовании Amplex® Red в резоруфин. Флюоресценцию можно измерить, например, используя флуориметровый фильтр, спектрофотометр для считывания планшетов, спектрофлуориметр, спектрофотометр или подобные, применяя способы, хорошо известные в области техники. Флюоресценция, образовавшаяся посредством реакции, может быть обнаружена, применяя флюориметрическую установку с возбуждением в диапазоне от приблизительно 530 до приблизительно 560 нм и эмиссией приблизительно 590 нм. Поглощение можно измерять (например, применяя спектрофотометр или спектрофотометр для считывания планшетов) приблизительно при 565 нм.
В одном варианте осуществления настоящее изобретение охватывает изменение ранее указанных условий теста для расширения динамического диапазона теста для приспособления к более широкому диапазону концентраций субстрата. Изменение включает концентрацию XOD по меньшей мере 1 Е/мл, приблизительно 1 до приблизительно 1,25 Е/мл, от приблизительно 1,25 до приблизительно 1,5 Е/мл, от приблизительно 1,5 до приблизительно 2 Е/мл, или больше чем 2 Е/мл; концентрацию PNP более чем 0,1 Е/мл, от приблизительно 0,1 до приблизительно 0,5 Е/мл, от приблизительно 0,5 до приблизительно 1 Е/мл, от приблизительно 1 до приблизительно 1,5 Е/мл, от приблизительно 1,5 до приблизительно 2 Е/мл, или более чем 2 Е/мл; и концентрацию Amplex® Red более чем 100 мкМ, от приблизительно 100 до приблизительно 200 мкМ, от приблизительно 200 до приблизительно 300 мкМ, от приблизительно 300 до приблизительно 400 мкМ, от приблизительно 400 до приблизительно 500 мкМ, от приблизительно 500 до приблизительно 600 мкМ, от приблизительно 700 до приблизительно 800 мкМ, от приблизительно 800 до приблизительно 900 мкМ, от приблизительно 900 до приблизительно 1000 мкМ, или более чем приблизительно 1000 мкМ. Эта модификация может быть применена к тестам, измеряющим кинетическую активность любого фермента, в котором фосфат высвобождается во время реакции, катализируемой посредством фермента.
Растения
Настоящее изобретение может быть использовано для трансформации любых разновидностей растений, включая, но не ограничиваясь однодольными и двудольными. Примеры интересующих растений включают, но не ограничены зерновыми (кукуруза), сорго, пшеницей, подсолнечником, томатом, крестоцветными, перцами, картофелем, хлопчатником, рисом, соей, сахарной свеклой, сахарным тростником, табаком, ячменем и масличными культурами, Brassica sp., люцерной, рожью, просом, сафлором, арахисом, бататом, кассавой, кофе, кокосовым орехом, ананасом, цитрусовыми деревьями, какао, чаем, бананом, авокадо, инжиром, гуавой, манго, маслинами, папайей, орехом кешью, макадамией, миндалем, овсом, овощами, декоративными растениями и хвойными.
Овощи включают, но не ограничены, томатами, салатом, зелеными бобами, лимскими бобами, горохом, и членами рода Curcumis, такими как огурец, мускусная дыня и дыня мускуса. Декоративные растения включают, но не ограничены азалией, гортензией, гибискусом, розами, тюльпанами, нарциссами, петуниями, гвоздиками, молочаем и хризантемой. Предпочтительно, растениями по настоящему изобретению являются хлебные злаки (например, кукуруза, сорго, пшеница, подсолнечник, помидор, крестоцветные, перцы, картофель, хлопчатник, рис, соя, сахарная свекла, сахарный тростник, табак, ячмень, масличные культуры, и т.д.).
Это изобретение является особенно подходящим для любого члена однодольного семейства растений, включая, но не ограничиваясь кукурузой, рисом, ячменем, овсом, пшеницей, сорго, рожью, сахарным тростником, ананасом, бататом, луком, бананом, кокосовым орехом и финиками.
Оценка трансформации растения
После введения гетерологичной чужеродной ДНК в растительные клетки трансформация или интеграция гетерологичного гена в геном растения подтверждается посредством различных способов, таких как анализ нуклеиновых кислот, белков и метаболитов, связанных с встроившимся геном.
ПЦР-анализ является быстрым методом для скрининга трансформированных клеток, ткани или побегов на наличие введенного гена на более ранней стадии перед пересаживанием в почву (Sambrook and Russell (2001) Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY)). ПЦР выполняется с использованием олигонуклеотидных праймеров, специфичных для интересующего гена или для вектора Agrobacterium в качестве фона, и т.д.
Трансформация растения может быть подтверждена посредством Саузерн-блот анализа геномной ДНК (Sambrook and Russell (2001) выше). В общем, общая ДНК экстрагируется из трансформанта, расщепляется соответствующими рестрицирующими ферментами, фракционируется в агарозном геле и переносится на нейлоновую мембрану или нитроцеллюлозу. Мембрана или "блот" могут далее быть исследованы, например, фрагментом ДНК-мишени, меченным радиоактивным изотопом 32P, для подтверждения интеграции введенного гена в геном растения согласно стандартным методикам (Sambrook и Russell, 2001, выше).
При Нозерн-анализе из определенных тканей трансформанта выделяют РНК, фракционируют в агарозном формальдегидном геле и переносят на нейлоновый фильтр согласно стандартным методикам, которые обычно используются в области техники (Sambrook and Russell (2001) выше). Посредством гибридизации фильтра с радиоактивным зондом, полученным из GDC посредством способов, известных в области техники, проверяют экспрессию РНК, кодируемой grg23 или grg51 (Sambrook and Russell (2001) выше).
Для определения наличия белка, кодируемого геном устойчивости к гербициду посредством стандартных методик, на трансгенных растениях могут быть выполнены вестерн-блоттинг и биохимические тесты и т.п. (Sambrook and Russell (2001) выше), применяя антитела, которые связываются с одним или несколькими эпитопами, присутствующими в белке устойчивости к гербициду.
Следующие примеры предлагаются в качестве иллюстрации, а не в качестве ограничения.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Пример 1: Получение ATX21308
ATX 21308 был выделен посредством высеивания образцов почвы на Обогащенную Минимальную Питательную среду 3 (EMM3), содержащую фосфаты и 50 мМ глифосата. Поскольку EMM3 не содержит никаких ароматических аминокислот, штамм должен быть устойчивым к глифосату, чтобы расти на этой питательной среде.
Приблизительно один грамм почвы был растворен приблизительно в 10 мл воды и перемешан на вортексе в течение 5 секунд. 100 мкл этой суспензии добавлены к 1 мл EMM3 с фосфатом, но без глифосата. EMM3 содержит (на литр, рН 7,0): 10 г сахарозы, 1 г NH4Cl, 0,2 г MgSO4 · 7H2O, 0,01 г FeSO4 · 7H2O, 0,007 г MnSO4 · H2O и 10 мл раствора фосфата, содержащего (на литр, рН 7,0) 210 г Na2HPO4 и 90 г NaH2PO4. Культуру взбалтывают в крутящемся барабане для тканевых культур при 21°C в течение ночи и затем высевают на агар EMM3, содержащий 50 мМ глифосата. После трех дней изолят высеивают на агар Luria Bertani (LB) для подтверждения единственности морфологии. После шести дней единичная колонию переносят на агар EMM3, содержащий 50 мМ глифосата. Изолят выращивают в течение ночи на чашках с 50 мМ глифосатом. Один специфический штамм, обозначенный как ATX21308, был выбран в силу его способности к росту в присутствии высоких концентраций глифосата. Этот штамм проверен на способность к росту в присутствии глифосата в жидкой культуре и способен к выращиванию при приблизительно 300 мМ глифосата при отработанных условиях.
Пример 2. Подготовка и скринирование космидных библиотек
Полная ДНК была извлечена из культуры ATX21308 с использованием способов, широко известных в области техники. ДНК была частично расщеплена ферментом рестрикции Sau3A1 и лигирована с векторным фрагментом SuperCos (Stratagene) согласно указаниям изготовителя. Продукты лигирования были упакованы в фаговые частицы с использованием упаковочного экстракта GigaPack XL (Stratagene), трансфицированы в клетки E. coli и высеяны на агаре LB, содержащем 50 мкг/мл канамицина для отбора колоний, содержащих космиды.
Индивидуальные колонии были переколоты в 384-луночные планшеты, содержащие бульон LB и 50 мкг/мл канамицина, и выращивались до насыщения. Клетки этих культур были разбавлены 1:10, затем прикреплены на агаровые чашки M63, содержащие 50 мкг/мл канамицина и либо 0 мМ, 10 мМ, 20 мМ или 50 мМ глифосата. [Агаровая среда M63 100 мМ KH2PO4, 15 мМ (NH4)2SO4, 50 мкМ CaCl2, 1 мкM FeSO4, 50 мкM MgCl2, 55 мМ глюкозы, 25 мг/литр L-пролина, 10 мг/л тиамина HCl, достаточно NaOH для доведения рН до 7,0, и 15 г/л агара]. Трансформанты с более быстрым ростом при более высоких концентрациях глифосата, были изолированы и расщеплены рестрикцирующим ферментом EcoRI для идентификации космид со сходным рисунком рестрикции. Были идентифицированы несколько клонов, которые ростут в присутствии глифосата и имеют сходные EcoRI рисунки рестрикции. Один из этих космидных клонов, pAX1924, был выбран для дальнейших экспериментов.
Пример 3. Идентификация grg23 в космиде pAX1924
Для идентификации гена(ов), ответственного за устойчивость к глифосату, продемонстрированную космидой pAX1924, ДНК из этого клона мутагенизировали подвижными элементами. В этом способе индентифицировали клоны, которые подверглись инсерциям транспозона и потеряли способность придавать устойчивость к глифосату. Местоположение инсерций транспозона устанавливает открытую рамку считывания, ответственную за фенотип устойчивости к глифосату.
Космида pAX1924 in vitro была подвергнута мутагенезу транспозоном с использованием EZ::TN Insertion Kit (Epicentre, Madison, WI) согласно протоколу изготовителя. По этому способу фрагмент транспозона рандомизированно вставляется в космидную ДНК и таким образом рандомизированно разрушает функцию генов в космиде. Этот транспозон содержит ген, кодирующий устойчивости к триметоприму, таким образом, клоны с инсерцией транспозона могут быть выбраны посредством способности к росту в присутствии этого антибиотика. Местоположения инсерций транспозона могут быть определены посредством составления генетической карты фрагмента рестрикции или посредством секвенирование с праймеров, которые отжигаются на транспозоне. Клоны pAX1924 со вставками транспозона высевали на питательную среду M63, содержащую глифосат. Было идентифицировано множество транспозон-содержащих клонов, которые утеряли способность к росту в присутствии глифосата, указывая, что транспозон разрушил ген, ответственный за устойчивость.
Последовательность ДНК определяли для области pАХ1924, содержащей инсерции транспозона, применяя способы секвенирования, хорошо известные в области техники. Используя эту информацию о последовательности, синтезировали ДНК-праймеры и использовали для определения последовательности ДНК pAX1924 области, вмещающей инсерции транспозона. Анализ полученных последовательностей ДНК показывает, что эта область содержит единственный ген. Этот ген обозначен в настоящем описании как grg23. Анализ grg23 показывает, что он способен к образованию двух возможных белков в бактериальных клетках из-за наличия потенциальных дополнительных участков старта трансляции. Первый ORF (ORF1) начинается со старт-кодона GTG в положениях 109-111 из SEQ ID NO:1, и заканчивается стоп-кодом TAG у нуклеотидов 1417-1419 из SEQ ID NO:1. Второй ORF (ORF2) начинается со старт-кодона ATG у нуклеотидов 178-180 из SEQ ID NO:1 и заканчивается стоп-кодоном TAG у нуклеотидов 1417-1419 из SEQ ID NO:1. Трансляция ORF1 приводит к аминокислотной последовательности, приведенной в SEQ ID NO:2. Трансляция ORF2 приводит к аминокислотной последовательности, приведенной в SEQ ID NO:4.
Анализ области ДНК, окружающей grg23, предполагает, что ORF2 предшествует участку связывания рибосомы, тогда как не очевидно, что участок связывания рибосомы предшествует началу трансляции ORF1. Кроме того, выравнивание обеих открытых рамок считывания с репрезентативными EPSPS-ферментами показывает, что немногие EPSPS-ферменты содержат данное N-концевое удлинение, кодируемое в пределах ORF1. Таким образом, функциональный ORF, кодируемый grg23 в бактериях, является ORF2. Поэтому, как используется в настоящем описании, GRG23 относится к тому, что кодируется ORF2 (нуклеотиды 178-1419 SEQ ID NO: 1). Тем не менее, известно в области техники, что EPSPS-ферменты являются весьма толерантными к дополнительным аминокислотам на их N-конце. Поэтому экспрессия ORF1 (нуклеотиды 109-1419 из SEQ ID NO:1) должна также приводить к EPSPS, который обладает устойчивостью к глифосату.
Для проверки способности ORF2 функционировать в качестве EPSPS и обладать устойчивостью к глифосату в клетке, эта открытая рамка считывания может быть субклонирована и экспрессирована в E. coli.
Пример 4. Субклонирование grg23 в векторы для экспрессии в E. coli.
Ген, кодирующий GRG23 ORF2 (начинающийся с ATG (положения 178-180 из SEQ ID NO: 1), экспрессирующий белок из 413 аминокислот), был субклонирован в pUC18 и pRSF1b, применяя ту же самую стратегию клонирования, изложенную в общих чертах выше. Был синтезирован ПЦР-праймер [5' CAGGGATCCGGCATGGAAACTGATCGACTAGTG 3'], который добавляет сайт BamHI, за которым следует GGC (5'-GGATCCGGC-3') непосредственно с 5' от старт-участка. Был синтезирован 2-й праймер [5'ATTGGCGCGCCCTAGCCGGGGAGCGTAAG 3'], который добавлял сайт AscI непосредственно с 3' от стоп-последовательности (5'-GGCGCGCC-3'). Область, кодирующую grg23, амплифицировали посредством ПЦР, применяя PFUULTRA™ ДНК-полимеразу (Stratagene). После ПЦР-амплификации grg23, применяя данные праймеры и расщепление рестриктазами BamHI/AscI, ПЦР-продукт был лигирован в pUC19 (ращепленный BamHI и AscI) и pRSF1b (расщепленный BamHI и AscI), и были получены колонии, содержащие вставку. Клон pUC18-grg23 (обозначенный в настоящем описании как pAX1927) был подтвержден посредством разрезания рестриктазами и посредством секвенирования ДНК.
Подобным образом вектор для экспрессии pAX1909 был расщеплен BamHI и AscI, и вектор, содержащий фрагмент, был выделен из геля посредством способов, широко известных в области техники. pAX1909 является производным PRSF-1b (Novagen, San-Diego, CA), измененный так, чтобы содержать участок BamHI непосредственно с 3' области, кодирующей обогащенный гистидином "His-Tag". Таким образом, белки, клонированные в pAX1909, являются гибридами в одной рамке, которые содержат дополнительные аминокислоты MAHHHHHHGSG. Векторы, такие как pAX1909, как правило, разработаны для экспрессии и очистки белков, и эти способы широко известны в области техники.
Расщепленный ПЦР-продукт, полученный выше, был лигирован в расщепленный вектор pAX1909, и были получены колонии, содержащие вставку. Клон pAX1909-grg23 (обозначенный в настоящем описании как pAX1926) был подтвержден посредством расщепления рестриктазами и посредством секвенирования ДНК. Свойство конструкции pAX1926 таково, что предсказанный продукт трансляции GRG23 содержит аминоконцевое удлинение, состоящее из MAHHHHHH. Это N-концевое удлинение включает 'гистидиновую метку' или метку 'six-His', которая применима для облегчения очистки белка GRG23, что также широко известно в области техники.
Плазмида pAX1926, содержащая grg23 ORF2, была депонирована в Коллекции культур службы сельскохозяйственных исследований (NRRL) 18 ноября 2005, и ей присвоен номер доступа NRRL B-30888.
Пример 5. grs23 придает устойчивость к высоким уровням глифосата
Конструкцию pUC18-Grg23 (pAX1927) трансформировали в E. coli штамм DH5α и высевали на чашки с агаром LB, дополненные карбенициллином (0,1 мг/мл). Были выбраны две колонии, ресуспендированы в стерилизованной воде и высеяны на чашки M63, содержащие или 0 мМ, 25 мМ, 50 мМ или 100 мМ глифосата. Изопропил-B-D-тиогалактопиранозид (IPTG; 0,1 мМ), был также добавлен на чашки. В качестве контроля, клетки, содержащие только один вектор pUC18, были трансформированы и высеяны на чашки с глифосатом. После 2 дней роста эти чашки были проверены на рост (Таблица 1).
Этот результат подтверждает, что экспрессия grg23 с образованием GRG23-ORF2 придает устойчивость к глифосату в E. coli к по меньшей мере 100 мМ. Дополнительно, рост E. сoli, содержащих pAX1927, более выражен в присутствии глифосата, чем в отсутствие глифосата.
Пример 6. Гомология GRG23 с другими белками
Выведенная аминокислотная последовательность GRG23 имеет гомологию с EPSPS-ферментами, указывая, что grg23 кодирует EPSPS.
Изучение выведенной аминокислотной последовательности GRG-ORF2 (SEQ ID NO:4) показывает, что в ней не содержится четырех доменов, типичных для II Класса EPSPS-ферментов. Таким образом, она является новой, не принадлежащей к классу II, EPSPS устойчивости к глифосату.
Поиск публично доступных белковых баз данных, такой как SWISSPROT, показывает, что у GRG23 есть аминокислотное сходство с широким классом EPSPS-ферментов. Однако ни один белок во всех базах данных не имеет более чем 50%-ной гомологии с аминокислотной последовательностью GRG23. Репрезентативное выравнивание GRG23 с другими EPSPS-ферментами показано на Фигуре 1.
Пример 7. Очистка GRG23
Конструкцию pRSF1b-grg23 (pAX1926) экспрессировали в E. coli после индукции IPTG и очищали в одну стадию, применяя кобальтовую хроматографическую колонку, как известно в области техники. После элюции с колонки, очищенный GRG23 был диализован против 50 мМ HEPES/100 мМ KCl, рН 7,0. Белок был более чем на 95% чистый по оценке посредством PAGE. Количество GRG23 было определено количественно, применяя способ Брэдфорда, как это известно в области техники (Bradford (1976) Anal. Biochem. 72:248-254).
Пример 8 Кинетические тесты на активность GRG23
Образцы очищенных белков анализировали на активность EPSPS, используя кинетический тест, включающий инкубацию PEP (Sigma, St. Louis, MO) и S3P в буфере, содержащем хлорид калия и HEPES при рН 7.0. Высвобождение фосфата детектировали, используя двойной тест для флуоресцентной детекции фосфата на основе образования Amplex Red, как это известно в области техники (Vazquez et al., (2003) Anal. Biochem. 320:292-298). Опубликованные условия анализа могут привести к насыщению анализируемого в экспериментах, в которых фосфат высвобождается очень быстро. Это насыщение несколько ограничивает динамический диапазон анализируемого, и требует определенного диапазона концентраций фермента. Было определено, что кинетическое ограничение флуоресцентного теста для фосфата очевидно из-за комбинации факторов, включая ограничение инозина и PNP. В настоящем изобретении условия теста были разработаны таким образом, что привели к значительному улучшению динамического диапазона и позволяют применять более широкий диапазон концентраций фермента и субстрата. Условия теста, которые были значительно изменены, включают концентрации пуриннуклеозидфосфорилазы (PNP), ксантиноксидаза (XOD), AMPLEX® Red и инозина, для каждого из которых были увеличены концентрации в тесте для приспособления к более высокой скорости оборота фосфата. Этот тест был применен для измерения активности EPSPS в 96-луночном формате со следующими улучшениями:
Улучшенный флуоресцентный тест
Ферментативные тесты проводились в 96-луночных планшетах в общем объеме 50 мкл. Реакции проводились при комнатной температуре при рН 7,0. Все компоненты теста, кроме РЕР, EPSPS и S3P, были объединены в Master Mix и расфасованы по отдельным пробиркам в 96-луночный планшет, применяя многоканальную пипетку. В каждую лунку затем были добавлены подходящие концентрации РЕР. Свежие разведения EPSPS были приготовлены и добавлены в соответствующие лунки. Каждый тест начинали посредством добавления S3P.
Данные по скорости были нанесены на диаграмму, и кинетические параметры Кm и Kcat были определены посредством применения Уравнения Михаэлиса-Ментена, применяя программу для обработки нелинейной кривой (KALEID AGRAPH®, Synergy Sofware). Значения Ki были определены посредством измерения Кm (app) при множественных концентрациях глифосата, и была составлена диаграмма Кm (арр) как функция концентрации ингибитора.
Влияние глифосата на Кm(арр) GRG23
Посредством построения графика Кm(арр) как функции концентрации глифосата может быть получено линейное представление устойчивости GRG23 к глифосату. Точка пересечения полученной линии с осью Х является -Ki. Построение этой линии по данным из Таблицы 3 приводит к следующим значениям:
Кинетические значения для GRG23
GRG23 очень устойчив к глифосату, с Ki более чем 9 мМ, и отношением Ki/Km более чем 800.
Пример 9: Выделение ATX21308
Для штамма ATX 21313 приблизительно один грамм почвы суспендировали в 10 мл воды, и были использованы 100 мкл инокуляции 1 мл культуры в минеральную питательную среду солей А (MSMA) без глифосата. MSMA содержит (на 1 литр, рН 7,0) 1 г NH4Cl, 10 г сахарозы, 0,2 г MgSO4 · 7H2O, 0,01 г FeSO4 · 7H2O, 0,007 г MnSO4 · H2O, дополненная фосфатами. После ночной инкубации культура была высеяна на твердую среду, содержащую MSMA и 50 мМ глифосата, инкубирована в течение нескольких дней, и инокулирована на агаровые чашки Luria Bertani для подтверждения единственного типа колоний. Рост в присутствии 50 мМ глифосата был рекомендован после повторного выращивания на MSMA, 50 мМ агаровых чашках. Этот способ выделения привел к штамму ATX21313, который был способен хорошо расти в таких условиях.
Пример 10. Клонирование устойчивых к глифосату EPSP-синтаз
Геномную ДНК была извлечена из штамма ATX21313, и полученную ДНК частично была расщеплена ферментом рестрикции Sau3A1 для получения фрагментов ДНК с размером приблизительно 5 тысяч пар нуклеотидов. Эти молекулы ДНК были отбраны по размеру на агарозном геле, выделены и лигированы в "плечи" LAMBDA ZAP® вектора, который предварительно расщепляли BamHI. Лигированные "плечи" далее были упаковыны в фаговые частицы, и были определены титры бактериофага, как известно в области техники. Полученные библиотеки амплифицировали посредством способов, известных в области техники, для получения библиотечного титра между 3×107 и 3×108 БОЕ/мл. Для каждой независимой библиотеки, E. coli (XL1 Blue MRF') была затем котрансфицирована бактериофагом из амплифицированной библиотеки так же, как и М13 фагом-помощником для обеспечения массового вырезания библиотеки в виде инфекционной, кольцевой оцДНК, как известно в области техники (Short et al. (1988) Nucleic Acids Research 16:7583-7600). После центрифугирования коинфицированных клеток супернатант, содержащий бактериофаг, нагревали до 65-70°C в течение 15-20 минут для инактивации остаточных частиц фага лямбды. Разведения полученной оцДНК плазмидной библиотеки трансфицировали в свежую культуру компетентных E. coli клеток XL1 Blue MRF', а также клеток XL-Blue MRF' (ΔaroA) (XL1 Blue MRF'). Полученные трасфицированные клетки высевали на чашки M63, содержащие канамицин, 0,1 мкМ IPTG и или 0 мМ, 20 мМ или 50 мМ глифосата. Этот способ скринирования позволяет идентифицировать клоны, содержащие толерантные к глифосату EPSP-синтазы, так же, как и клоны, несущие толерантность к глифосату. Колонии, растущие на 20 мM или 50 мМ глифосата в штамме ΔaroA или XL-Blue MRF', были переколоты, и их плазмиды проанализированы посредством рестриктного разрезания для идентификации плазмид с общим рисунком рестрикции. Отдельные плазмиды секвенировали посредством способов, известных в области техники, с предпочтением, отданным плазмидам, которые проявляли устойчивость к 50 мМ глифосату.
Используя этот подход, который иногда модифицировали для каждой библиотеки, как известно и ценится в области техники, были опознаны клоны библиотеки, содержащие гены EPSP-синтазы.
Последовательности областей полученных клонов были идентифицированы как области EPSP-синтазы.
Пример 11. ДНК и белковая последовательности EPSP-синтаз
Последовательность ДНК EPSP-синтазы, устойчивой к глифосату, была определена для pAX1967 посредством способов, известных в области техники. Последовательность ДНК grg51 представлена в настоящем описании как SEQ ID NO:5. Предсказанный продукт трансляции grg51 (GRG51) представлен в настоящем описании как SEQ ID NO:6. GRG51 демонстрирует 97% аминокислотную идентичность с GRG23 (SEQ ID NO:2).
Плазмида pAX1926, содержащая grg23 ORF2, была депонирована в Коллекции культур службы сельскохозяйственных исследований (NRRL) 18 ноября 2005, и ей присвоен номер доступа NRRL B-30949.
Таблица 5 суммирует гомологию GRG23 и GRG51 с другими EPSP-синтазными ферментами.
Аминокислотная идентичность GRG23-ORF1 и GRG51 с репрезентативными EPSPS-ферментам
Пример 12. Клонирование новых устойчивых к глифосату EPSP-синтаз в E. coli вектор для экспрессии
Ген grg51, содержащийся в pAX1967, был субклонирован в E. coli вектор для экспpессии рRSF1b (Invitrogen). Полученные клоны были подтверждены посредством ДНК секвенирования, и применены для индукции экспрессии grg51 в E. coli. Экспрессированный His-теговый белок был затем очищен, как известно в области техники.
Пример 13. Устойчивость к глифосату EPSP-синтаз
Клетки, содержащие pAX1967, были высеяны на M63 + чашки, содержащие антибиотик и или 0 мМ или 20 мМ глифосата. Рост был оценен после двух дней выращивания при 37°C. Наблюдали, что GRG51 придавал клеткам E. coli устойчивость к 20 мМ глифосату (Таблица 6).
Глифосатный скрининг
Пример 14. Конструирование syngrg23 и экспрессия
Последовательность нового гена, кодирующая белок GRG23 (SEQ ID NO:2; Заявка на Патент США № 60/741166, зарегистрированная 1 декабря 2005), была сконструирована и синтезирована. Эта последовательность представлена как SEQ ID NO: 12. Эта открытая рамка считывания, сконструированная в настоящем описании как "syngrg23", была клонирована в вектор для экспрессии pRSF1b (Invitrogen) посредством способов, известных в области техники.
Ген Syngrg23, кодирующий GRG23, был клонирован в вектор pUC19 для создания pAX748. Для амплификации syngrg23 с pAX748 с использованием MUTAZYME® II системы (Stratagene) для введения случайных мутаций в syngrg23, кодирующий область, были применены ПЦР-праймеры, которые фланкировали syngrg23 в этом векторе. Матрица была разбавлена 1:50 в подверженной ошибкам реакции ПЦР, и амплификация выполнялась в течение 30 циклов. Полученный ПЦР-продукт был расщеплен ферментами рестрикции BamHI и SgsI, очищен от геля и лигирован в вектор pRSF1b для создания библиотеки мутированных syngrg23.
Библиотеки мутированных syngrg23 были трансформированы в E. coli штамм BL21*DE3 star (Invitrogen). После трансформации отдельные колонии были высеяны на среду 1x M63, содержащую 150 мМ глифосата для отбора клонов, которые сохранили ферментативную активность и устойчивость роста.
Пример 15. Скринирование на устойчивость к глифосату на чашках
Библиотечные лигирования были трансформированы в компетентные клетки E.coli BL21*DE3 (Invitrogen). Трансформации были выполнены согласно инструкциям изготовителя со следующими модификациями. После инкубации в течение 1 часа при 37°C в среде SOC, клетки были осаждены центрифугированием (5 минут, 1000 × g, 4°C). Клетки промывали 1 мл M63+, снова центрифугировали, и удаляли супернатант. Клетки промывали во второй раз 1 мл M63+ и повторно ресуспендировали в 200 мкл M63+.
Для отбора мутантных ферментов GRG23, придающих устойчивость к глифосату в E. coli, клетки были высеяны на агаровые чашки со средой M63+, содержащие 150 мМ глифосата, 0,05 мМ IPTG (изопропил-бета-D-тиогалактопиранозид) и 50 мкг/мл канамицина. Среда M63+ 100 мМ KH2PO4, 15 мМ (NH4)2SO4, 50 мкМ CaCl2, 1 мкM FeSO4, 50 мкM MgCl2, 55 мМ глюкозы, 25 мг/литр L-пролина, 10 мг/л тиамина HCl, достаточно NaOH для доведения рН до 7,0, и 15 г/л агара. Чашки инкубировали в течение 36 часов при 37°C.
Отдельные колонии были переколоты, высеяны в 384-луночные планшеты. Таким образом, были созданы два 384-луночных планшета. Третий планшет с 384 клонами был собран из колоний, которые росли на чашках без глифосата.
Пример 16. Изоляция и анализ вариантов GRG23, устойчивых к глифосату
Клетки BL21*DE3, трансформированные мутагенизированными syngrg23 и/или вариантами grg23, были идентифицированы посредством роста на чашках с глифосатом. Экстракты мутагенизированных syngrg23 и вариантов grgr23 были приготовлены и протестированы на улучшение ферментативной активности. Колонии, идентифицированные на чашках с глифосатом, были прикреплены в 96-луночные блоки, содержащие среду LB, и были выращены до O.D. приблизительно 0,6. Затем был добавлен IPTG (0,5 мм), и блоки инкубировали в течение ночи при 20°C для индукции экспрессии белка. Экстракты белка были приготовлены из осадков клеток, применяя реактив для культур РОР (Novagen) и Lysonase (Novagen), и ферментативная активность в неочищенных лизатах была измерена после нагревания экстрактов в течение 30 минут при 37°C. Экстракты с активностью большей, чем два стандартных отклонения выше среднего из ряда экстрактов, содержащих соответствующий контрольный белок (например, GRG23), были выбраны для дальнейшего анализа.
Клоны, показывающие повышенную активность после инкубации в качестве неочищенных экстрактов, были выращены в 250 мл культуры LB, и экспрессия белка была вызвана IPTG. После индукции мутантный белок GRG23 был очищен из каждой культуры посредством аффинной хроматографии с использованием кобальтовой смолы (Novagen). Далее очищенные белки были проверены на ферментативную активность после нагревания в течение 0, 2, 4 и приблизительно 16 часов при 37°C.
Пример 17. Улучшенные варианты GRG23
Из библиотеки ДНК мутагенизированного syngrg23 было идентифицировано несколько клонов с улучшенной активностью. Были определены последовательности ДНК клонов, соответствующих этим экстрактам. В Таблице 7 показаны аминокислотные замены, идентифицированные в шести вариантах GRG23, которые сохраняли устойчивость к глифосату: grg23(L3P1.B20) (SEQ ID NO:26), кодирующий аминокислотную последовательность GRG23 (L3P1.B20) (SEQ ID NO:27); grg23(L3P1.B3) (SEQ ID NO:28) кодирующий аминокислотную последовательность GRG23(L3P1.B3) (SEQ ID NO:29); grg23(L3P1.F18) (SEQ ID NO:30), кодирующий аминокислотную последовательность GRG23(L3P1.F18) (SEQ ID NO:31); и grg23(L3P1.O23) SEQ ID NO:31 SEQ, кодирующий аминокислотную последовательность GRG23(L3P1.O23) SEQ ID NO:32).
Мутации, идентифицированные в устойчивых к глифосату вариантах GRG23
Клоны были выращены в 250 мл культуры LB, и была вызвана экспрессия белка, и белок был выделен, как описано выше. Очищенные белки затем были протестированы на ферментативную активность после нагревания в течение 0, 2 и 4 и приблизительно 16 часов при 37°C. Оказалось, один из клонов, обозначенный "M5", обладал увеличенным соотношением его ферментативной активности после продолжительной инкубации при 37°C (Таблица 8). Была определена последовательность ДНК данного клона, и ген определяется в настоящем описании как grg23 (ace1) (SEQ ID NO: 14). Белок, экспрессированный с grg23 (ace1), обозначен GRG23 (ACE1) (SEQ ID NO: 15).
Время полужизни GRG23 (ACE1) в сравнении с GRG23 при повышенной температуре
GRG23 (ACEl) содержит 2 аминокислотные замены в сравнении с белком GRG23 дикого типа: A49→T и S276→T. Вектор pRSF1b, который содержит этот ген, обозначен pAX3801. На фигуре 1 представлена относительная устойчивость GRG23 (ACEl) в сравнении с GRG23 при повышенных температурах.
Пример 18. Определение EPSPS-активности вариантов GRG-23
Экстракты, содержащие вариантные белки GRG23, были протестированы на синтазную активность EPSP, как описано в Заявке на Патент США № 60/741166, зарегистрированной 1 декабря 2005, включенной в настоящее описание полностью посредством ссылка. Тестирования были проведены в суммарном объеме 50 мкл, содержащем 0,5 мМ шикимат-3-фосфата, 200 мкМ фосфоенолпирувата (РЕР), 1 Е/мл ксантиноксидазы, 2 Е/мл нуклеозидфосфорилазы, 2,25 мМ инозина, 1 Е/мл пероксидазы хрена, 0-2 мМ глифосата, 50 мМ HEPES/KOH pH 7,0, 100 мМ KCl и AMPLEX® Red (Invitrogen) согласно инструкциям изготовителя. Экстракты инкубировали со всеми компонентами теста, кроме шикимат-3-фосфата, в течение 5 минут при комнатной температуре, и тестирование было начато посредством добавления шикимат-3-фосфата. EPSP-синтазная активность была измерена с использованием флуоресцентный спектрометр Spectramax Gemini XPS (Molecular Dynamics, возбуждение: 555 нм; эмиссия: 590 нм).
Следуя полному определению, кинетические параметры были выполнены на очищенном белке, как описано ранее (Заявка на Патент США № 60/741166, зарегистрированная 1 декабря 2005), доводя количество белка, определенное посредством метода Брэдфорда, как известно в области техники. Для любой одной концентрации глифосата EPSP-синтазная активность была измерена как функция широкого диапазона концентраций РЕР. Данные укладывались в уравнение Михаэлиса-Ментена с использованием программного обеспечения KALEIDAGRAPH® (Synergy Software), для определения Кm (кажущаяся Кm) EPSP-синтазы при той концентрации глифосата. Кажущиеся значения Km были определены при не менее чем 4 концентрациях глифосата, и Ki EPSPS для глифосата были вычислены по графику кажущихся Кm по сравнению с концентрацией глифосата, используя уравнение (мл*x/(m2+x); m1=1; m2=1), как известно в области техники.
Кинетика GRG23 (ACE1) в сравнении с GRG23
Пример 19. Идентификация grg23 (ace2).
GRG23(ACE1) содержит две аминокислотные замены по сравнению с GRG23. Чтобы определить, могли ли дополнительные замены в этих положениях далее улучшить активность, была получена ДНК библиотека, которая привела к клонам, экспрессирующим белки, которые были преимущественно мутированы в обоих положения 49 и 276 GRG23. Клоны, несущие устойчивость к глифосату, были выбраны посредством роста на чашках с глифосатом, и выращены и протестированы на кинетические свойства, как было описано.
Удивительно, один клон, обозначенный в настоящем описании grg23(ace2) (SEQ ID NO: 16), кодирующий GRG23(ACE2) белок (SEQ ID NO: 17), был идентифицирован как обладающий улучшенной термостабильностью. Последовательность ДНК grg23(ace2) показала, что GRG23 (ACE2) содержит единственную аминокислотную замену (остаток 276 GRG23 на аргинин).
Пример 20. Сравнение GRG23 и GRG51, и мутагенез различных остатков
Для оценки пермутаций аминокислотных последовательностей, вероятных из сравнения аминокислотных последовательностей GRG23 и GRG51, были получены две библиотеки. Первая библиотека вводила разнообразие из аминокислотной последовательности GRG51 в кодирующую область grg23(ace2). Вторая библиотека вводила разнообразие из аминокислотной последовательности GRG23 (ACE2) в кодирующий область grg51.
Клоны полученных библиотек были протестированы на (1) способность придавать клетке устойчивость к глифосату, и (2) активность после продолжительной инкубации при 37°C. В общей сложности десять клонов были секвенированы и проанализированы более подробно. Один конкретный клон, определяемый в настоящем описании как grg51.4 (SEQ ID NO: 18), кодирующий белок GRG51.4 (SEQ ID NO: 19), содержит несколько аминокислотных замен и относительно GRG23(ACE2) и относительно GRG51. Аминокислотные замены, присутствующие в GRG51.4 относительно GRG23(ACE2), были впоследствии введены в ген grg23(ace2) для получения grg23(ace3) (SEQ ID NO:20), который кодирует белок GRG23(ACE3) (SEQ ID NO:21). GRG23(ACE3) проявляет наивысшую активность и термостабильность в сравнении с GRG23 и GRG23(ACE2).
GRG23(ace1) был мутагенизирован, и клоны были протестированы для идентификации клонов, экспрессирующих варианты с улучшенной термостабильностью и/или активностью. Один клон, grg23(L5P2.J2) (SEQ ID NO:22), кодирующий GRG23(L5P2.J2) (SEQ ID NO:23), был идентифицирова на основании его улучшенных кинетических свойств. GRG23(L5P2.J2), содержит три аминокислотные замены относительно GRG23(ACE1), как показано в следующей Таблице 10.
Аминокислотные замены в GRG23 (L5P2.J2)
Для создания клонов, которые содержат каждый идентифицированную аминокислотную замену GRG23(L5P2.J2) в кодирующей области grg23(ace3), был применен мутагенез олигонуклеотидами. Клон был идентифицирован как клон, кодирующий белок, имеющий улучшенные кинетические свойства по сравнению с GRG23(ACE3), и обозначен как grg23(ace4) (SEQ ID NO:24). Белок, кодируемый grg23(ace4), обозначен как GRG23(ACE4) (SEQ ID NO: 25), содержит единственную аминокислотную замену относительно GRG23(ACE3) (Валин 101 на фенилаланин). На основе этих результатов был выполнен отдельный мутагенез олигонуклеотидами, чтобы проанализировать кинетику каждых возможных аминокислотных замен в положении 101. Ни одна из аминокислотных замен не привела к дополнительному улучшению кинетических свойств по сравнению с GRG23 (ACE4).
Кинетика улучшенных вариантов
Пример 21. Конструирование grg23 или grg51 для трансформации растения
Открытая рамка считывания (ORF) grg23 или grg51 была амплифицирована посредством ПЦР с полноразмерной кДНК матрицы. Рестриктные сайты HindIII были добавлены к каждому концу ORF во время ПЦР. Дополнительно, нуклеотидная последовательность ACC была добавлена сразу с 5' от старт-кодона гена для увеличения эффективности трансляции (Kozak (1987) Nucleic Acid Research 15:8125-8148; Joshi (1987) Nucleic Acid Research 15:6643-6653). Продукт ПЦР был клонирован и секвенирован, применяя способы, известные в области техники, для подтверждения того, что никакие мутации не введены во время ПЦР.
Плазмида, содержащая grg23 или grg51 продукта ПЦР, была расщеплена HindII, и фрагмент, содержащий интактную ORF, был изолирован. Этот фрагмент был клонирован в рестриктные сайты HindIII плазмиды pAX200, вектор для экспрессии в растениях, содержащий промотор актина риса (McElroy etal. (1991) Molec. Gen. Genet. 231:150-160) и терминатор PinII (An et al. (1989) The Plant Cell 1:115-122). Фрагмент промотор-ген-терминатор из этой промежуточной плазмиды субклонировали в плазмиду pSB11 (Japan Tobacco, Inc) для образования итоговой плазмиды, например, pSB11GRG23. pSB11GRG23 была организована таким образом, что 3,91 т.п.н. фрагмент ДНК, содержащий конструкцию промотор-grg23-терминатор может быть расщеплен посредством двойного расщепления KpnI и PmeI и использован для трансформации растений посредством аэрозольной лучевой инъекции. Структура pSB11GRG23 была проверена посредством рестрикционного разрезания и электрофореза в геле и посредством секвенирования через различные связки для клонирования.
Плазмида была перенесена в Agrobacterium tumefaciens, штамм LBA4404, который также несет плазмиду pSB1 (Japan Tobacco, Inc), используя процедуры трехродительского скрещивания, хорошо известные в области техники, и высевая в среду, содержащую спектиномицин. Плазмида pSB11GRG23 несет устойчивость к сектиномицину, но является плазмидой узкого круга хозяев и не может размножаться в Agrobacterium. Колонии, устойчивые к спектиномицину, появляются, когда pSВ11GRG23 встраивается в плазмиду широкого круга хозяев pSB1 посредством гомологичной рекомбинации. Продукт коинтеграта pSB1 и pSB11GRG23 проверен посредством гибридизации по Саузерну. Штамм Agrobacterium, несущий коинтеграт, используется для трансформации кукурузы посредством способа PureIntro (Japan Tobacco).
Пример 22. Трансформация grs23 или grg51 в растительные клетки
Початки кукурузы собирали спустя 8-12 дней после опыления. Из початков выделяли эмбрионы, и данные эмбрионы, размером 0,8-1,5 мм, применяли для трансформации. Эмбрионы высеивали на чашки стороной шейного щитка наверх в подходящей среде для инкубации, такой как среда DN62A5S (3,98 г/л Солей N6; 1 мл/л (1000x базового раствора) Витаминов N6; 800 мг/л L-аспарагина; 100 мг/л Myo-инозитола 1,4 г/л L-пролина; 100 мг/л казаминовых кислот; 50 г/л сахарозы; 1 мл/л (1 мг/мл базового раствора) 2,4-D). Однако среды и соли, отличные от DN62A5S, являются подходящими и общеизвестны в области техники. Эмбрионы инкубировала в течение ночи при 25°C в темноте.
Полученные эксплантаты переносили в квадратные лунки (30-40 на чашке), переносили в осмотические среды в течение 30-45 минут, затем переносили на лучевую чашку (см., например, Публикацию РСТ № WO/0138514 и Патент США № 5240842).
ДНК-конструкции, сконструированные для экспрессии GRG23 в растительных клетках, переносили в ткани растения с использованием катализатора луча аэрозоля, применяя по существу условия, как описано в Публикации РСТ № WO/0138514. После облучения эмбрионы инкубировали в течение 30 минут в осмотических средах и помещали в инкубационные среды на ночь при 25°C в темноте. Для того чтобы избежать чрезмерного повреждения облученных эксплантатов, их инкубировали в течение по меньшей мере 24 часов до перемещения в восстанавливающую среду. Эмбрионы затем распределяли в среду для восстановления в течение 5 дней при 25°C в темноте, затем переносили в селекционную среду. Эксплантаты культивировали в селективной среде вплоть до восьми недель, в зависимости от природы и особенностей конкретно применяемой селекции. После периода селекции полученный каллюс переносили на среду для созревания эмбрионов до тех пор, пока не наблюдалось образование зрелых соматических эмбрионов. Полученные зрелые соматические эмбрионы далее помещали под слабое освещение, и запускали процесс регенерации посредством способов, известных в области техники. Полученным побегам позволяли укорениться в среде для выращивания растений и полученные растения переносили в горшки питомника и размножали как трансгенные растения.
Материалы
Доводят pH раствора до pH 5,8 1N KOH/1N KCl, добавляют Gelrite (Sigma) до 3 г/л и автоклавируют. После охлаждения до 50°C доводят до 2 мл/л 5 м/мл базовым раствором нитрата серебра (Phytotechnology Labs). Рецепт рассчитан приблизительно на 20 чашек.
Пример 23. Трансформация grs23 или grg51 в растительные клетки кукурузы посредством Agrobacterium-опосредованной трансформации.
Початки собирали спустя 8-12 дней после опыления. Из початков выделяли эмбрионы, и эти эмбрионы, размером 0,8-1,5 мм, применяли для трансформации. Эмбрионы высеивали на чашки стороной шейного щитка наверх в подходящей среде для инкубации в течение ночи при 25°C в темноте. Однако по существу не обязательно культивировать эмбрионы в течение ночи. Эмбрионы, которые контактируют со штаммом Agrobacterium, содержащим соответствующие векторы для плазмиды Ti-опосредованной передачи в течение 5-10 минут, затем высевают на чашки со средой для совместного культивирования в течение 3 дней (при 25°C в темноте). После совместного культивирования эксплантаты переносили в среду для периода восстановления в течение пяти дней (в 25°C в темноте). Эксплантаты инкубировали на селективной среде вплоть до восьми недель в зависимости от природы и особенностей конкретного способа селекции.
После периода селекции, полученный каллюс переносили в среду для созревания эмбриона до тех пор, пока не наблюдается образование зрелых соматических эмбрионов. Полученные зрелые соматические эмбрионы далее помещали под слабое освещение и запускали процесс регенерации посредством способов, известных в области техники. Полученным побегам позволяли укорениться в среде для выращивания растений, и полученные растения переносили в горшки питомника и размножали как трансгенные растения.
Все публикации и заявки на патенты, упомянутые в описании, указывают на уровень опыта специалистов в области техники, к которой принадлежит настоящее изобретение. Все публикации и заявки на патенты включены в настоящее описание посредством ссылки в той степени, как если бы конкретно и отдельно указано, что каждая отдельная публикация или заявка на патент включены посредством ссылки.
Хотя предшествующее изобретение было описано подробно посредством иллюстрации и примера для лучшего понимания, очевидно, что определенные изменения и модификации могут быть осуществлены в рамках приложенной формулы изобретения.






































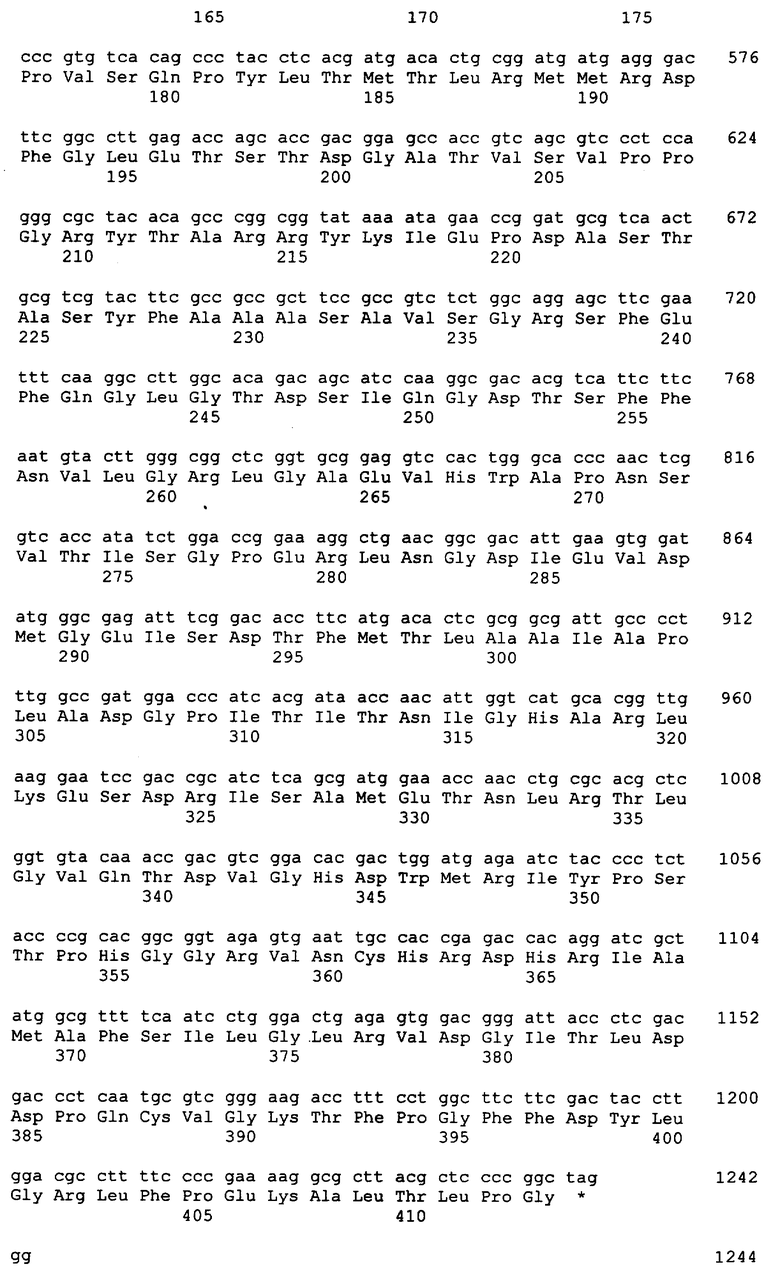







| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ КЛАСС ГЕНОВ УСТОЙЧИВОСТИ К ГЛИФОСАТУ | 2013 |
|
RU2634411C2 |
| ГЕН АХМ1554, КОДИРУЮЩИЙ ДЕЛЬТА-ЭНДОТОКСИН, И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2016 |
|
RU2755282C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТА ИЗ BRASSICA | 2013 |
|
RU2636035C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2632648C2 |
| НОВЫЙ КЛАСС ГЕНОВ УСТОЙЧИВОСТИ К ГЛИФОСАТУ | 2013 |
|
RU2721122C2 |
| ТРАНЗИТНЫЙ ПЕПТИД ХЛОРОПЛАСТОВ | 2013 |
|
RU2636036C2 |
| СИНТЕТИЧЕСКИЕ ПРОИСХОДЯЩИЕ ИЗ BRASSICA ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2626535C2 |
| ИНСЕКТИЦИДНЫЕ ПОЛИПЕПТИДЫ С ШИРОКИМ СПЕКТРОМ АКТИВНОСТИ И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2750459C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТОВ | 2013 |
|
RU2720973C2 |
| Белок устойчивости к гербицидам, кодирующий его ген и их применение | 2016 |
|
RU2692553C2 |
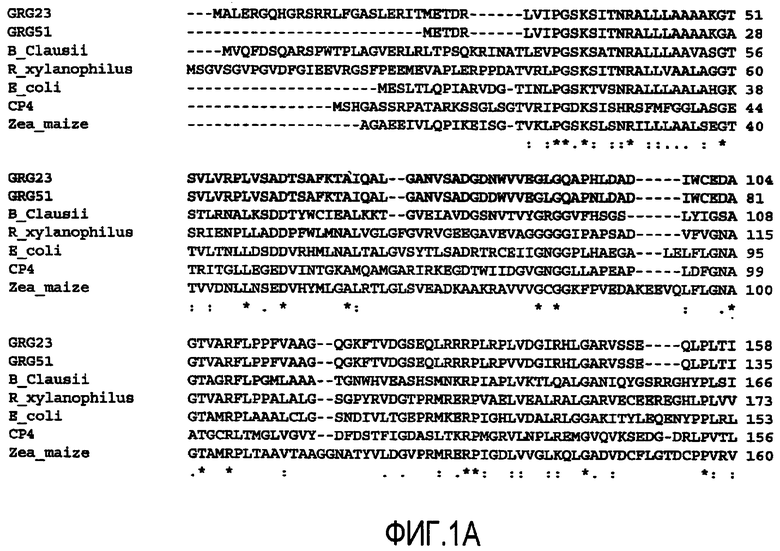
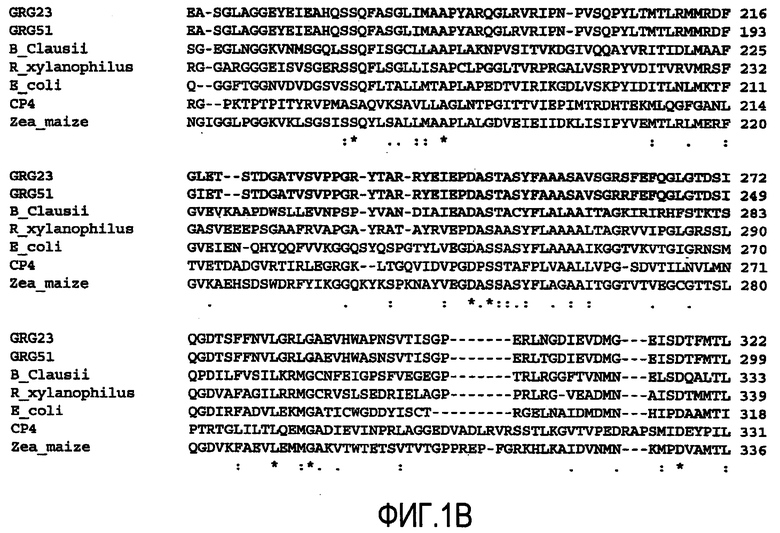

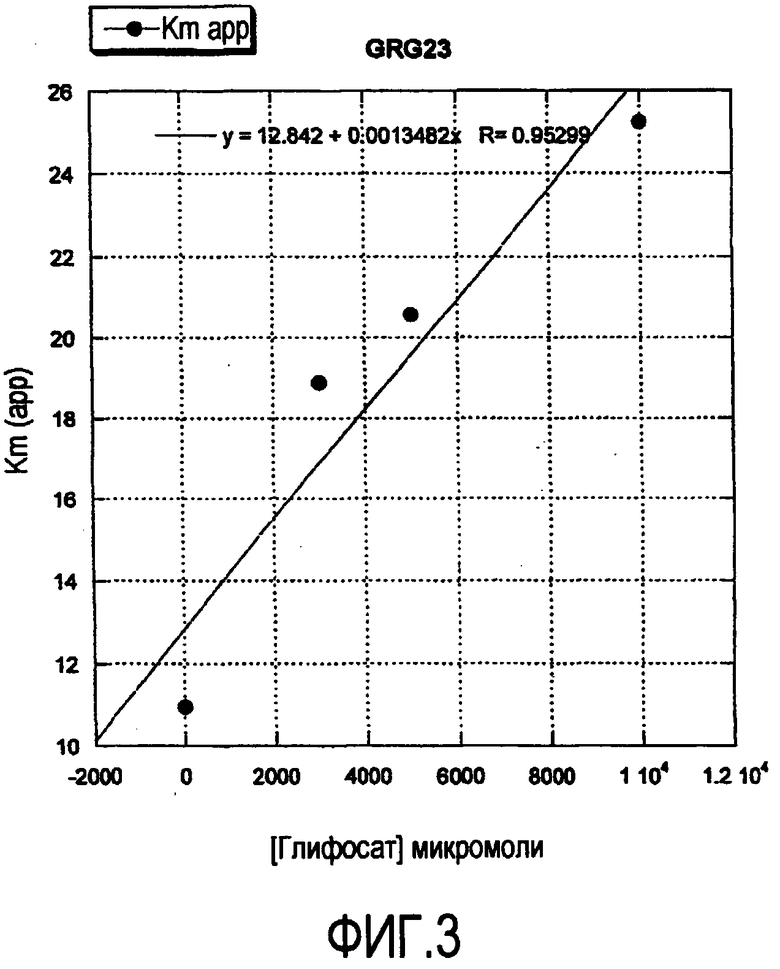
Нуклеотидные последовательности, кодирующие полипептиды устойчивости к глифосату, используют в конструкциях ДНК или экспрессионных кассетах для трансформации или экспрессии в организмах, включая микроорганизмы и растения. Трансформированные организмы приобретают устойчивость к гербициду. 9 н. и 8 з.п. ф-лы, 5 ил., 11 табл.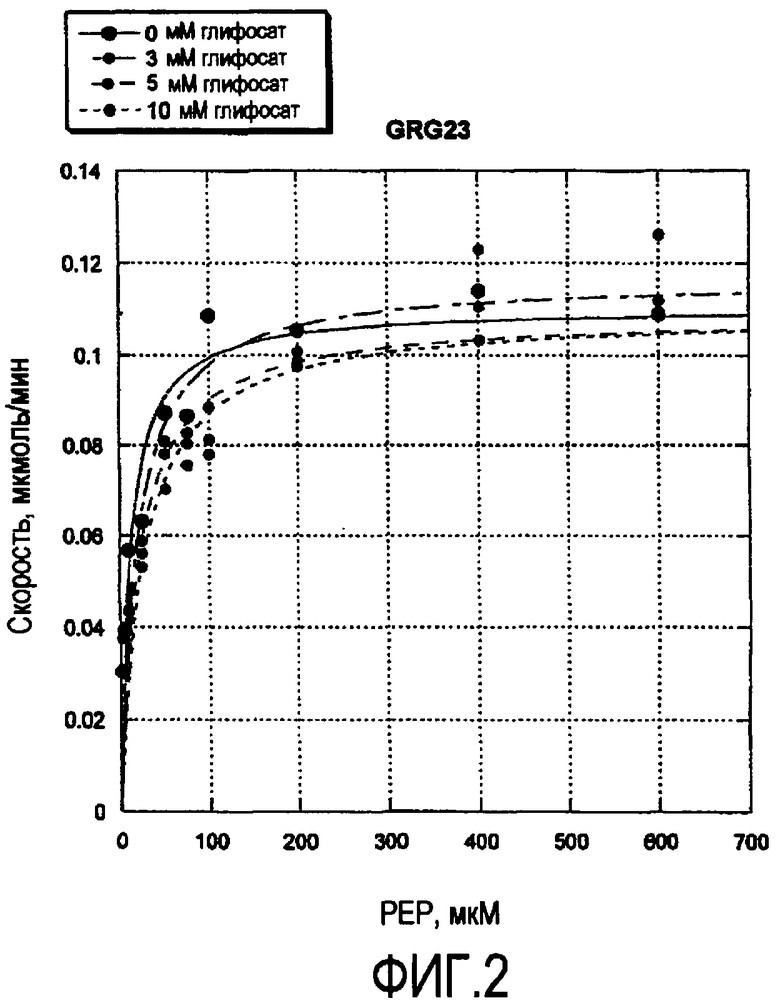
1. Изолированная молекула нуклеиновой кислоты, кодирующая полипептид, обладающий активностью устойчивости к гербициду, выбранная из группы, состоящей из:
a) молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:1, 3 или 5 или комплементарную таковой;
b) молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, имеющую по меньшей мере 80%-ную идентичность последовательности с нуклеотидной последовательностью SEQ ID NO:1, 3 или 5 или комплементарной таковой;
c) нуклеотидной последовательности устойчивости к гербициду вставки ДНК плазмиды, депонированной под Номером Доступа NRRL В-30888 или В-30949 или комплементарной таковой;
d) молекулы нуклеиновой кислоты, которая кодирует полипептид, содержащий аминокислотную последовательность SEQ ID NO:2, 4 или 6;
и
e) молекулы нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую полипептид, обладающий по меньшей мере 80%-ной идентичностью аминокислотной последовательности с аминокислотной последовательностью SEQ ID NO:2, 4 или 6.
2. Изолированная молекула нуклеиновой кислоты по п.1, где указанная нуклеотидная последовательность является синтетической последовательностью, которая была сконструирована для экспрессии в растении.
3. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п.1.
4. Вектор экспрессии по п.3, дополнительно содержащий молекулу нуклеиновой кислоты, кодирующую гетерологичный полипептид.
5. Клетка-хозяин, обладающая активностью устойчивости к гербициду, где указанная клетка-хозяин содержит вектор экспрессии по п.3.
6. Клетка-хозяин по п.5, которая является бактериальной клеткой-хозяином.
7. Клетка-хозяин по п.5, которая является растительной клеткой.
8. Трансгенное растение, обладающее активностью устойчивости к гербициду, где указанное растение содержит клетку-хозяина по п.7.
9. Растение по п.8, где указанное растение выбрано из группы, состоящей из кукурузы, сорго обыкновенного, пшеницы, подсолнечника, томата, крестоцветных, перцев, картофеля, хлопка, риса, сои, сахарной свеклы, сахарного тростника, табака, ячменя и масличных культур.
10. Трансформированное семя растения по п.8, где указанное семя содержит молекулу нуклеиновой кислоты, кодирующую полипептид, обладающий активностью устойчивости к гербициду.
11. Изолированный полипептид устойчивости к гербициду, выбранный из группы, состоящей из:
a) полипептида, содержащего аминокислотную последовательность SEQ ID NO:2, 4 или 6;
b) полипептида, кодируемого нуклеотидной последовательностью SEQ ID NO:1,3 или 5;
c) полипептида, содержащего аминокислотную последовательность, имеющую по меньшей мере 80%-ную идентичность последовательности с аминокислотной последовательностью SEQ ID NO:2, 4 или 6, где указанный полипептид имеет активность устойчивости к гербициду;
d) полипептида, который кодируется нуклеотидной последовательностью, которая по меньшей мере на 80% идентична нуклеотидной последовательности SEQ ID NO:1, 3 или 5, где указанный полипептид имеет активность устойчивости к гербициду; и
e) полипептида, который кодируется нуклеотидной последовательностью устойчивости к гербициду вставки ДНК плазмиды, депонированной под Номером Доступа NRRL В-30888 или NRRL В-30949.
12. Полипептид по п.11, дополнительно содержащий гетерологичную аминокислотную последовательность.
13. Способ получения полипептида с активностью устойчивости к гербициду, включающий культивирование клетки-хозяина по п.5 в условиях, в которых экспрессируется молекула нуклеиновой кислоты, кодирующая полипептид, где указанный полипептид выбран из группы, состоящей из:
a) полипептида, обладающего активностью устойчивости к гербициду, содержащего аминокислотную последовательность SEQ ID NO:2, 4 или 6;
b) полипептида, обладающего активностью устойчивости к гербициду, кодируемого последовательностью нуклеиновой кислоты SEQ ID NO:1, 3 или 5;
c) полипептида, содержащего аминокислотную последовательность, имеющую по меньшей мере 80%-ную идентичность последовательности с полипептидом с аминокислотной последовательностью SEQ ID NO:2, 4 или 6, где указанный полипептид имеет активность устойчивости к гербициду;
d) полипептида, кодируемого молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность, имеющую по меньшей мере 80%-ную идентичность последовательности с последовательностью нуклеиновой кислоты SEQ ID NO:1, 3 или 5, где указанный полипептид имеет активность устойчивости к гербициду; и
е) полипептида, обладающего активностью устойчивости к гербициду, который кодируется нуклеотидной последовательностью устойчивости к гербициду вставки ДНК плазмиды, депонированной под Номером Доступа NRRL В-30888 или NRRL В-30949.
14. Способ придания растениям устойчивости к гербициду, включающий трансформацию указанного растения ДНК-конструкцией, содержащей промотор, который управляет экспрессией в растительной клетке, функционально связанный с нуклеотидной последовательностью, по меньшей мере на 80% идентичной нуклеотидной последовательности SEQ ID NO:1, 3 или 5, где указанная нуклеотидная последовательность кодирует полипептид, обладающий активностью устойчивости к гербициду; и регенерацию трансформированного растения.
15. Способ по п.14, где указанным гербицидом является глифосат.
16. Растение, обладающее активностью устойчивости к гербициду, имеющее стабильно встроенную в его геном ДНК-конструкцию, содержащую нуклеотидную последовательность, которая кодирует белок, имеющий активность устойчивости к гербициду, где указанная нуклеотидная последовательность выбрана из группы, состоящей из:
a) нуклеотидной последовательности SEQ ID NO:1, 3 или 5, где указанная нуклеотидная последовательность кодирует полипептид, обладающий активностью устойчивости к гербициду;
b) нуклеотидной последовательности, имеющей по меньшей мере 80%-ную идентичность последовательности с нуклеотидной последовательностью SEQ ID NO:1, 3 или 5, где указанная нуклеотидная последовательность кодирует полипептид, обладающий активностью устойчивости к гербициду;
c) нуклеотидной последовательности, кодирующей полипептид, обладающий активностью устойчивости к гербициду, содержащий аминокислотную последовательность SEQ ID NO:2, 4 или 6;
d) нуклеотидной последовательности, кодирующей полипептид, имеющий по меньшей мере 80%-ную идентичность аминокислотной последовательности с аминокислотной последовательностью SEQ ID NO:2, 4 или 6, где указанный полипептид обладает активностью устойчивости к гербициду; и
e) нуклеотидной последовательности устойчивости к гербициду вставки ДНК плазмиды, депонированной под Номером Доступа NRRL В-30888 или NRRL В-30949;
где указанная нуклеотидная последовательность является функционально связанной с промотором, который управляет экспрессией кодирующей последовательности в растительной клетке.
17. Растение по п.16, где указанным растением является растительная клетка.
| ВЫДЕЛЕННЫЙ ПОЛИНУКЛЕОТИД, СПОСОБНЫЙ ПРИДАВАТЬ РАСТЕНИЮ УСТОЙЧИВОСТЬ ИЛИ ТОЛЕРАНТНОСТЬ К ГЛИФОСАТНОМУ ГЕРБИЦИДУ, ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ, ТОЛЕРАНТНЫХ ИЛИ УСТОЙЧИВЫХ К ГЛИФОСАТНОМУ ГЕРБИЦИДУ, СПОСОБ РЕГЕНЕРАЦИИ ТРАНСФОРМИРОВАННОГО РАСТЕНИЯ И СПОСОБ СЕЛЕКТИВНОЙ БОРЬБЫ С СОРНЯКАМИ | 2000 |
|
RU2235778C2 |
| ИЗОЛИРОВАННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК (ВАРИАНТЫ), РЕКОМБИНАНТНАЯ ДВУНИТЕВАЯ МОЛЕКУЛА ДНК И СПОСОБ ПОЛУЧЕНИЯ ГЕНЕТИЧЕСКИ ТРАНСФОРМИРОВАННЫХ РАСТЕНИЙ | 1991 |
|
RU2146290C1 |
| APPL | |||
| AND ENVIRON | |||
| MICROBIOLOGY, 1988; 54(11): 2860-2870 | |||
| APPL | |||
| AND ENVIRON | |||
| MICROBIOLOGY, 1988; 54(5): 1293-1296 | |||
| JOURNAL OF BIOLOGICAL CHEMISTRY, 1991; 266(33): 22364-9. | |||

