ОБЛАСТЬ ТЕХНИКИ
Данное изобретение относится к носителю лекарства-антигена, дающего возможность трансмукозального и трансдермального введения, и более конкретно, к способу индуцирования мукозального иммунитета, который вызывает избирательное и эффективное, особенно селективное, продуцирование антиген-специфичных секреторных иммуноглобулинов А, мукозальной вакцине, предотвращению или лечению аллергии и системе доставки лекарства, отличающейся использованием носителя для желаемого антигена или лекарства.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Известно, что традиционные инактивированные вакцины, токсоиды и т.п. обладают следующими недостатками.
(1) Отсутствие защиты против инфекции по естественным путям проникновения инфекции.
Естественным путем проникновения инфекции, в частности бактерий, вирусов и т.п., является проникновение, например, через слизистую оболочку носовой полости, трахеи, кишечного тракта и т.п., в то время как путь введения вакцины является подкожным, внутримышечным и т.п., что отличается от вышеописанного пути. Желательно получить защиту от инфекции с помощью иммунизации тем путем, который соответствует действительным условиям проникновения природной инфекции, в частности защиту от инфекции в слизистой оболочке в результате введения вакцины через слизистую оболочку.
(2) Низкий мукозальный иммунитет.
У субъектов иммунизации вакцинами главным образом продуцируется в крови иммуноглобулин G (который далее будет обозначен просто как «IgG» или «IgG антитело»), и индуцируется гуморальный иммунный ответ.При этом секреторный иммуноглобулин А (который далее будет обозначен просто как «IgA» или «IgA антитело»), ответственный за мукозальный иммунитет, почти не продуцируется, и следовательно, развитие мукозального иммунитета не ожидается. Далее, необходимость и эффективность IgA антитела являются следующими: IgA антитело играет очень важную роль в клиническом иммунитете, отвечая за мукозальный иммунитет, то есть за защиту от инфекции в слизистой оболочке, которая является входом для инфекции в органы дыхания, такие как носовая полость и трахея, с капелями и воздухом, и в кишечный тракт через рот. Более того, IgА антитело обладает перекрестным иммунитетом, то есть активностью перекрестной нейтрализации и широким спектром защиты от инфекции как таковой, а также дает защиту против инфекции в отношении другого антигена, в то время как IgG антитело высокоспецифично для антигена, но имеет узкий спектр по защите от инфекции, что приводит к тому, что оно практически неэффективно при защите против инфекции в отношении другого антигена.
(3) Необходимость дополнительной вакцинации и дополнительных расходов.
Так как IgG антитело только после одной инъекции при первой иммунизации продуцируется в незначительном количестве и определенных эффектов ожидать не приходится, требуется повышать уровень IgG антитела в крови дополнительной иммунизацией, так называемой бустерной (повторной) иммунизацией (ревакцинацией), один или несколько раз, на основании условий позднего сохранения памяти IgG антител. Следовательно, требуются повторные затраты и усилия, и кроме того, часто получается, что шанс ревакцинации не эффективен для маленьких детей, особенно малышей двух лет и меньше, для которых вероятно уклониться от шанса ревакцинации, хотя и признаны эффекты для пожилых, взрослых и школьников, которые пользуются этим шансом.
Другими словами, традиционная инактивированная вакцина, токсоид и т.п. индуцирует в основном продуцирование IgG антитела в крови субъекта иммунизации вакциной и вызывает действия и эффекты по увеличению гуморального иммунитета и признана в этом качестве эффективной. Однако она имеет низкую продуктивность по индуцированию продуцирования IgА антитела или мукозального иммунитета, демонстрируя ограничение адекватной функции и эффективности по защите от естественной инфекции. В этих обстоятельствах до сих пор с различных сторон было предпринято много попыток по устранению этих недостатков традиционных вакцин. Например, предпринимались попытки улучшения вакцинного антигена в качественном или количественном отношении, экспериментального производства живых вакцин взамен инактивированных вакцин, разработки новых путей иммунизации, мукозальных вакцин и т.п., поиск адъювантов, которые повышают гуморальный иммунитет и обеспечивают его поддержание, разработки мукозальных иммуноадъювантов и т.п. Однако не удалось создать мукозальную вакцину, которая была бы безопасна и эффективна.
Далее будет пояснено, как разрабатываются мукозальные вакцины.
(1) Увеличение количества вакцинного антигена.
Была предпринята попытка увеличения количества вакцинного антигена, который вводят подкожно или внутримышечно, или увеличения количества IgG и IgA антител, которые секретируются в слизистую оболочку. Например, испытывался способ, при котором добавляют нейраминидазу белковой вирусной оболочки и смешивают с традиционной инактивированной противогриппозной вакциной для увеличения количества продуцируемых антител, или способ, при котором добавляют MF59 и смешивают как адъювант, и т.п. Однако обнаружилось, что эти способы имеют такие недостатки, как возникновение боли и сильная побочная реакция и т.п.
(2) Вакцины назального типа введения.
Был испытан способ, при котором жидкий сплит-антиген вводился непосредственно через нос (назально) для защиты от инфекции с помощью IgA антитела, что считается наиболее эффективным, но был отмечен факт, что количество продуцированного IgA явилось небольшим. Чтобы повысить способность продуцирования IgA антитела, была предпринята попытка, при которой термолабильный живой токсин Escherichia coli или токсин холеры добавили и смешали как адъювант со сплит-антигеном для повышения мукозального иммунного ответа, то есть способности продуцирования IgA антитела. Однако в условиях, когда безопасность токсина как адъюванта не была доказана, указанные испытания остановили, и способ не был внедрен в практику.
(3) Живая вакцина, использующая холодоаттенуированный штамм, который может быть введен в носовую полость.
В практику был внедрен способ, при котором использовался холодоаттенуированный штамм вируса гриппа, чья оптимальная температура роста составляет 25°С и который не растет при 39°С, но при этом не может быть отброшен риск возврата токсичности, так как механизм аттенуации родительского холодоаттенуированного штамма не ясен. Кроме того, активным ингредиентом вакцины является живой вирус, он имеет высокую силу инвазии в клетки и прекрасен для инициализации иммунитета, но часто встречаются умеренные симптомы гриппа, то есть обнаружены недостатки, и эта вакцина не может быть использована для людей, которые имеют высокий риск ухудшения состояния при заражении гриппом, пожилых людей и т.п.
(4) Другие вакцины.
Разработки векторной вакцины, у которой в качестве вирусного вектора выступает вирус коровьей оспы, аттенуированной живой вакцины методом обратной генетики, ДНК-вакцины, которая использует в качестве активного ингредиента ДНК или кДНК, продвигаются на лабораторном уровне, но не внедрены в практику.
Далее будет пояснено, как разрабатывается иммуноадъювант.
(1) Иммуноадъювант.
Иммуноадъювант - это общее наименование вещества, которое имеет активность регулятора, например обеспечивает усиление или ингибирование иммунного ответа. Эти вещества делятся в основном на два типа: вещество, которое относится к лекарственной форме, предназначенной для пролонгирования действия, удерживания и т.п. антигена в субъекте иммунизации, и вещество, которое помогает повышению, ингибированию и т.п. иммунного ответа. Из них первый, то есть адъювант для лекарственных форм, уже внедрен в практику, например вакцина или токсоид с использованием фосфата алюминия, квасцов и т.п. Однако пока не известно, чтобы в практику был внедрен второй из них, то есть адъювант, который помогает повышению/усилению иммунного ответа. Например, известны живая бактерия БЦЖ, полученная из бактерии, БЦЖ из клеточных оболочек микобактерий, эндотоксин, глюкан и т.п., синтезированный MDP, левамизол, Poly I-Poly С, бестатин и т.п., и интерфероны, такие как цитокины, ФНО, CSF и т.п., но считается, что необходима гарантия безопасности и эффективности для их практического применения из-за неудовлетворительной эффективности и т.п., адъювантных болезней, таких как артрит, хронический артритный ревматизм, гипер-γ-глобулинемия, анемия и т.п. Кроме того, известна методика (патентный документ 1) применения легочного сурфактанта/белка, полученного из высших животных, в качестве адъюванта, чтобы усилить индукцию гуморального иммунитета, но также не известно, чтобы она была внедрена в практику.
(2) Разработка мукозального иммуноадъюванта
Были разработаны различные адъюванты, например олигомер токсина В коклюша (патентный документ 2), токсин холеры (патентный документ 3), субъединица термолабильного энтеротоксина В Escherichia coli (LTB) (патентный документ 4), частицы крахмала (патентный документ 5), цепочный белок токсина В холеры (СТВ, от cholera toxin В) (патентный документ 6), субъединица В веротоксина 1 (патентный документ 7), олигонуклеотид (патентный документ 8), интерлейкин 12 (непатентный документ 1) и т.п. Однако до сих пор они не были внедрены в практику.
Как описано выше, широко признана необходимость замены традиционных вакцин, вводимых подкожно, внутримышечно и т.п., мукозальными вакцинами, которые индуцируют продуцирование IgA антитела в слизистой оболочке, которая является естественным путем проникновения вирусов. В частности, во всем мире ожидается, что в 21 веке будет разработана и внедрена в практику вакцина следующего поколения, так называемая мукозальная вакцина, которая индуцирует продуцирование IgA антитела и местный иммунитет или мукозальный иммунитет, однако такой результат еще не достигнут. Считается, что причиной этого является тот факт, что еще не определен и не создан безопасный и эффективный адъювант, который придает вакцине функцию индуцирования продуцирования IgA антитела и местного иммунитета или мукозального иммунитета.
Патентный документ 1: JP-T-2002-521460
Патентный документ 2: JP-A-3-135923
Патентный документ 3: JP-T-10-500102
Патентный документ 4: JP-T-2001-523729
Патентный документ 5: JP-T-2002-50452
Патентный документ 6: JP-A-2003-116385
Патентный документ 7: JP-A-2003-50452
Патентный документ 8: WO 00/20039
Непатентный документ 1: pp.4780-4788, vol. 71, 2003, Infection and Immunity.
Непатентный документ 2: pp.2-11, vol. 10, 2004, Journal of neonatal Nursing.
Непатентный документ 3: pp.9-14, vol. 74 (suppl. 1), 1998, Biology of the Neonate.
Непатентный документ 4: pp.452-458, vol. 24, 2001, American Journal of Respiratory Cell and Molecular Biology.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Задачи, на решение которых направлено изобретение, являются следующими: а именно, придать функцию индуцирования продуцирования IgA антитела и местного иммунитета или мукозального иммунитета; разработка безопасного и эффективного метода для этой цели; переход от традиционных вакцин для гуморального иммунитета к безопасным и эффективным вакцинам для мукозального иммунитета; предотвращение и лечение аллергии; создание системы трансмукозальной/трансдермальной доставки лекарства (для выражения «система доставки лекарства» далее будет использована аббревиатура «СДЛ») для введения и транспортировки лекарства через слизистую оболочку или кожу.
Данные задачи решаются тем, что предложен носитель лекарства-антигена, дающий возможность трансмукозального и трансдермального введения, а также способ индуцирования мукозального иммунитета, который дает предпочтительное и эффективное, в частности селективное, продуцирование антиген-специфичного секреторного иммуноглобулина А, а также мукозальная вакцина, агент для предотвращения или лечения аллергии, трансмукозальная или трансдермальная СДЛ, которая отличается использованием носителя для желаемого антигена или лекарства.
Применение или повсеместное использование этого носителя лекарства-антигена, предложенного согласно данному изобретению, приведет к распространению применений мукозальной вакцины против различных инфекций, агента для предотвращения или лечения аллергии, и трансмукозальной или трансдермальной СДЛ. Мукозальная вакцина, которая является иммунным средством, соответствующим действительным условиям природной инфекции, оказывает необыкновенно отличные эффекты по защите от инфекции в сравнении с традиционными вакцинами. Кроме того, IgА слизистой оболочки носовой полости, индуцируемый носителем лекарства-антигена, ведет к инактивации аллергена, что делает возможной пониженную сенсибилизацию. Далее, применение вышеупомянутой СДЛ для различных лекарств усиливает и содействует профилактическим и/или терапевтическим эффектам лекарств через трансмукозальное и трансдермальное введение. В результате данное изобретение в значительной степени улучшает лечение, здоровье и гигиену всего человечества, и также является хорошей обнадеживающей новостью для тех, кто имеет отношение к лечению, здоровью и гигиене в мире. Кроме того, оно в большой степени обеспечивает средство придания функции и действия, которое делает возможным трансмукозальное и трансдермальное введение, которое проще в сравнении с инъекцией, особенно для биологических препаратов, содержащих традиционные вакцины и вакцины будущего, токсоид и т.п., и различные лекарства.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1 представляет эффекты ингибирования инфекции вируса гриппа в (в) носовой полости и (г) альвеолах, в (а) носовой полости и (б) альвеолах при различных назальных введениях противогриппозной вакцины, и введении подкожной инъекцией противогриппозной вакцины.
* представляет уровень значимости от группы вакцинно-независимых (без носителя лекарства-антигена или адъюванта) введений по Т-тесту (р<0,01). (Пример 1)
Фиг.2 представляет количества IgА и IgG, представляющие собой противогриппозные антитела в растворе смыва из носовой полости после (а) назального введения и (б) подкожной инъекции противогриппозной вакцины. Белый столбец представляет количество IgA, и черный столбец представляет количество IgG, соответственно. (Пример 2)
Фиг.3 представляет влияние адъюванта на продуцирование специфичных противогриппозных антител, IgA и IgG, в растворе смыва из легких после (а) назального введения и (б) подкожной инъекции противогриппозной вакцины. (Пример 3)
Фиг.4 представляет влияние PSF-2 и СТВ на продуцирование специфичных противогриппозных антител, IgA и IgG, в крови после (а) назального введения и (б) подкожной инъекции противогриппозной вакцины. (Пример 4)
Фиг.5 представляет влияние PSF-2 и СТВ на уровень секреции TGF-β1 в слизистой оболочке (а) носовой полости и (б) альвеол после назального введения противогриппозной вакцины. (Пример 6)
Фиг.6 представляет влияние PSF-2 и СТВ на продуцирование специфичных противогриппозных антител в (а) носовой полости, (б) альвеолах и (в) крови после назального введения противогриппозной вакцины. (Пример 7)
Фиг.7 представляет влияние PSF-2 и СТВ на продуцирование различных цитокинов, секретируемых из носовой полости, альвеол и лимфоцитов селезенки после назального введения противогриппозной вакцины. (Пример 8)
Фиг.8 представляет влияние PSF-3 на продуцирование специфичных противогриппозных антител в (а) носовой полости, (б) альвеолах и (в) крови после назального введения противогриппозной вакцины. (Пример 9)
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Воплощения данного изобретения будут пояснены ниже.
1. Пояснение терминов и ингредиентов носителя лекарства-антигена
(1) Носитель лекарства антигена
Носитель (носитель лекарства антигена, далее обозначаемы как «носитель ЛА» или «НЛА») представляет собой комплекс липидов и белков, выполненный таким образом, чтобы сделать возможным трансмукозальное и трансдермальное введение антигена, лекарства и т.п. Носитель ЛА содержит компоненты (а)-(в), представленные ниже.
(а) Белок В легочного сурфактанта или его фрагмент (включая не только естественные фрагменты, полученные протеазой, но также искусственные фрагменты, полученные путем генетической инженерии или пептидным синтезом, или варианты фрагментов из-за замены и/или делеции по меньшей мере одной аминокислоты, входящей в состав такого фрагмента).
(б) Белок С легочного сурфактанта или его фрагмент (включая не только естественные фрагменты, полученные протеазой, но также искусственные фрагменты, полученные путем генетической инженерии или пептидным синтезом, или варианты фрагментов из-за замены и/или делеции по меньшей мере одной аминокислоты, входящей в состав такого фрагмента).
(в) Липид, такой как фосфолипид или жирная кислота. Форма и структура представляют собой мембрану (липидная мембрана в форме листа или рулона), сохраняющая полипептидные цепи в форме шипов или выступов, то есть в такой форме, где концы гидрофобных областей множественных полипептидных цепей погружены в липидную мембрану, чтобы быть внедренными в виде шипов в отличие от традиционной липидной везикулы (липосомы).
При желании антиген, лекарство и т.п. подвергаются сосуществованию, контакту, захвату, адгезии или связыванию (райдингу) с носителем лекарства-антигена согласно данному изобретению, что делает возможным трансмукозальное введение и трансдермальное введение такого антигена, лекарства и т.п. Другими словами вышеуказанный носитель является для него средством переноса, делающим возможным трансмукозальное введение и трансдермальное введение такого антигена, лекарства и т.п.
Кроме того, ингредиенты носителя лекарства-антигена, белки, полипептиды, пептиды или липиды, использованные в приготовлении/производстве носителя, то есть белки легочного сурфактанта В и С, их фрагменты, варианты фрагментов в силу замещения и/или делеции по меньшей мере одной аминокислоты, входящей в состав белка, из которого получен фрагмент, и т.п., и липид, такой как фосфолипид, жирная кислота, и т.п., будут пояснены в деталях далее.
(2) Легочный сурфактант
Легочный сурфактант был внедрен в практику с середины 1990-х для лечения респираторного дистресс-синдрома (RDS, от respiratory distress syndrome), и в настоящее время на рынке представлены и широко применяются различные препараты, полученные из человека, крупного рогатого скота, свиней и т.п. (непатентный документ 2). Кроме того, выпускаются и продаются препараты синтетических пептидов, содержащие активный домен, имеющий отношение к лечению RDS, и находится в развитии разработка конструкции или синтез аналогов SP-B и SP-C (непатентный документ 3). Состав и структура легочного сурфактанта являются следующими: комплекс, содержащий около 90% липида (67,3% фосфатидилхолина, 19,3% фосфатидилглицерина, 3,2% фосфатидилсерина, другие свободные жирные кислоты и т.п.), и около 10% белка (сурфактантные белки А, В, С и D, которые далее обозначены как «SP-A», «SP-B», «SP-C» и «SP-D», соответственно). Молекулярные массы составляют 28-36 кДа для SP-A, 15 кДа для SP-B, 3,5 кДа для SP-C и 43 кДа для SP-D. SP-A и SP-D являются гидрофильными (водорастворимым) и лектинеподобными (ассоциированными с мембраной). SP-B и SP-C являются гидрофобными (липидорастворимыми) и связываемыми с липидами, и обладают способностью быть погруженными в фосфолипидную мембрану и активностью сурфактанта. Гены белков легочных сурфактантов, полученных из человека, крупного рогатого скота, свиней и т.п., известны, например полноразмерная нуклеотидная последовательность гена человеческого SP-B имеет коллекционный номер J02761, а человеческого SP-C (и SP-C1) - J03890 в GenBank/NCBI (http//www.ncbi.nlm.nih.gov./). Кодирующая область (CDR) человеческих SP-B и SP-C, полученных из NCBI, и кодируемая ими аминокислотная последовательность описаны ниже.
SEQ ID No. 1: последовательность нуклеотидов CDR ДНК гена человеческого SP-B;
SEQ ID No. 2: полноразмерная аминокислотная последовательность человеческого SP-B, выведенная из SEQ ID No. 1;
SEQ ID No. 3: последовательность нуклеотидов CDR ДНК гена человеческого SP-C;
SEQ ID No. 4: полноразмерная аминокислотная последовательность человеческого SP-C, выведенная из SEQ ID No. 3;
SEQ ID No. 5: последовательность нуклеотидов CDR SP-C1, локализованная в ДНК гена человеческого SP-C; и
SEQ ID No. 6: полноразмерная аминокислотная последовательность человеческого SP-C1, выведенная из SEQ ID No. 5.
(3) Белки или пептиды, используемые в этом изобретении
В приготовлении/производстве носителя лекарства-антигена по настоящему изобретению могут быть использованы комбинация SP-B и SP-C и комбинация SP-B и SP-C1, полученные из млекопитающих, таких как человек, крупный рогатый скот, свиньи, киты, дельфины и т.п., или рыб, таких как тунец, акула, скаты, желтохвост и т.п. Например, может быть применена комбинация SP-B и SP-C и комбинация SP-B и SP-C1, представляющих собой белки человеческого происхождения, содержащие полноразмерную аминокислотную последовательность, описанную в SEQ ID No. 2, 4 и 6, соответственно. Далее, например, может быть применена гидрофобная (липидорастворимая) область SP-B и SP-C на основании величины гидрофобности Кайта-Дулитла (Kyte-Doolittle) и фрагмент, содержащий эту область, вариант фрагмента в силу замены и/или делеции по меньшей мере одной аминокислоты, входящей в состав пептида, из которого получен этот фрагмент. Например, может быть применен природный пептид или пептид, полученный генетической инженерией или химическим синтезом, содержащий аминокислотную последовательность, описанную в SEQ ID No. 7-20, длинноцепный пептид из них, содержащий такой пептид, и вариант или синтетический аналог с заменой и/или делецией по меньшей мере одной аминокислоты, входящей в состав такого пептида. Номер аминокислоты представлен порядковым номером, так что Met на N-конце каждой последовательности имеет номер 1, и далее идет нумерация в направлении С-конца (направление слева направо в описываемых последовательностях).
SEQ ID No. 7: аминокислотная последовательность с 214 до 225 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 8: Аминокислотная последовательность с 257 до 266 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 9: Аминокислотная последовательность с 29 до 58 в последовательностях SEQ ID No. 4 и 6 (SP-C фрагмент);
SEQ ID No. 10: Аминокислотная последовательность с 1 до 20 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 11: Аминокислотная последовательность с 102 до 110 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 12: Аминокислотная последовательность с 119 до 127 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 13: Аминокислотная последовательность с 136 до 142 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 14: Аминокислотная последовательность с 171 до 185 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 15: Аминокислотная последовательность с 201 до 279 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 16: Аминокислотная последовательность с 253 до 278 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 17: Аминокислотная последовательность с 300 до 307 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 18: Аминокислотная последовательность с 317 до 330 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 19: Аминокислотная последовательность с 344 до 351 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 20: Аминокислотная последовательность с 358 до 381 в последовательности SEQ ID No. 2 (SP-B фрагмент);
SEQ ID No. 21: Аминокислотная последовательность с 24 до 58 в последовательностях SEQ ID No. 4 и 6 (SP-C фрагмент).
Согласно данному изобретению может быть применена комбинация по меньшей мере одного типа, выбранного из группы, состоящей из белков SP-B, содержащих аминокислотные последовательности, описанные в SEQ ID No. 2, 7, 8 и 10-20, и их фрагментов, и по меньшей мере одного типа, выбранного из группы, состоящей из белков SP-C (и SP-C1), содержащих аминокислотные последовательности, описанные в SEQ ID No. 4, 6, 9 и 21, и их фрагментов.
(4) Липиды, использованные в данном изобретении
В качестве фосфолипидов предпочтительно использованы фосфолдипиды легочного сурфактанта, например фосфатидилхолин (лецитин), дипальмитоилфосфатидилхолин, фосфатидилсерин, и т.п. Кроме того, могут быть использованы дипальмитоилглицерофосфохолин, диацилглицерофосфоглицерин, фосфатидилглицерин (кардиолипин), дилауроилфосфатидилглицерин, димиристоилфосфатидилглицерин, дипальмитоилфосфатидилглицерин, дистеароилфосфатидилглицерин, фосфатидилинозитол, фосфатидилэтаноламин, фосфатидная кислота, сфингомиелин и т.п. Кроме того, в качестве жирной кислоты может быть использована лауриновая кислота, миристиновая кислота, пальмитиновая кислота, стеариновая кислота, пальмитолеиновая кислота, олеиновая кислота и т.п. Кроме того, могут быть использованы липиды, полученные из водных животных, таких как кит, тунец, дельфин и т.п., которые имеют активную легочную экспансию.
(5) Препараты легочного сурфактанта, находящиеся на рынке для лечения RDS
Согласно данному изобретению в качестве носителя ЛА могут быть использованы препараты легочного сурфактанта, имеющиеся на рынке, например Surfacten (товарный знак), Infasurf, Curosurf, Humansurf, Exosurf, Alveofact и т.п., которые были одобрены соответствующими органами в смысле их безопасности и эффективности в качестве лекарства для лечения RDS и содержат гидрофобные или липидорастворимые SP-B и SP-C и фосфолдипид. Кроме SP-B и SP-C находящиеся на рынке препараты, содержащие гидрофильные или водорастворимые SP-А и SP-D, могут быть применены после экстрагирования водорастворимых белков SP-A и SP-D 1-бутанолом и т.п. и их удаления до предела обнаружения или менее. Кроме того, применение сухого препарата является более желательным, чем жидкого препарата, в смысле доводки концентрации для использования в препарате носителя ЛА.
2. Новые данные, являющиеся основой данного изобретения
При таком сильном уровне техники данное изобретение совершено в результате тонких наблюдений и анализа авторами проб и ошибок, совершаемых на протяжении около 10 лет, глубоких знаний и опыта, а также новых идей, и оно основано на следующих неожиданно обнаруженных фактах.
(1) Традиционный адъювант усиливает антиген-представляющие явления, вызывая воспаление, и изначально имеется четыре типа активных белковых ингредиента, SP-A, SP-B, SP-C и SP-D, представляющих собой сурфактанты, секретируемые слизистой оболочкой легких или кишечного тракта. Было обнаружено, что когда вирусный антиген подвергается сосуществованию, контакту, захвату или адсорбции комплексом из комбинации SP-B и SP-C, исключая SP-A и SP-D, и фосфолипида, или комплексом из комбинации синтетических пептидов обоих фрагментов SP-B и SP-C, содержащих их липидорастворимую область (активную область), и липидной мембраны (вышеупомянутый носитель ЛА), антиген-представляющие клетки в слизистой оболочке носовой полости активируются, не вызывая воспаления, вирусные антигены внедряются в эти клетки с хорошей эффективностью, а также продуцирование противовирусных IgA слизистой оболочкой индуцируется эффективно и предпочтительно, и даже селективно, не вызывая индуцирования продуцирования IgG в слизистой оболочке или крови.
(2) Кроме того, было обнаружено, что путем добавления и смешивания комплекса из комбинации SP-B и SP-C и фосфолипида, или комплекса из комбинации синтетических пептидов обоих фрагментов SP-B и SP-C, содержащих их липидорастворимую область (активную область), и липидной мембраны (вышеупомянутый носитель ЛА), со сплит-антигеном вируса гриппа, который традиционно используется в качестве безопасного инактивированного вакцинного антигена, достигается селективное индуцирование секреторного продуцирования IgA с активацией антиген-представляющих клеток в достаточной степени усиленного и дополненного, которое является слабым при независимом сплит-антигене по сравнению с живой вакциной, с сохранением при этом высокой безопасности сплит-антигена.
3. Как были сделаны вышеуказанные неожиданные наблюдения
(1) Авторы данного изобретения проводили обширные исследования, чтобы объяснить механизм развития гриппа, а также способы лечения и предотвращения гриппа. При этом было выявлено, что легочный сурфактант адсорбирует и инактивирует триптазу Клара из протеаз, осуществляющих в дыхательных путях процессинг гемагглютинина (НА), который обнаруживает активность внедрения в мембрану вируса и инфекционную способность, путем рестрикционного расщепления НА (мембранного белка вируса гриппа), что приводит к ингибированию цикла роста этого вируса.
(2) В результате длительных исследований было обнаружено, что в дополнение к описанному выше действию легочный сурфактант селективно активирует мукозальные антиген-представляющие клетки, и таким образом активирует проявление иммунитета против вирусного антигена, приводя к индукции секреторных IgA, но не приводя к индукции IgG. Далее, было показано, что SP-B и SP-C являются такими же важными, как и липидный ингредиент, в качестве активных ингредиентов, усиливающих мукозальный иммунитет, в легочном сурфактанте. Была уточнена активная область этих белковых ингредиентов, и оценена эффективность усиления мукозального иммунитета.
(3) Далее, исследования были продолжены в отношении дающего биологическую защиту вещества в слизистой оболочке дыхательных путей и защиты против вирусной инфекции как описано выше, и было доказано, что легочный сурфактант, секретируемый в живом организме, вовлечен в селективное индуцирование продуцирования IgA в качестве мукозального иммуноадъюванта из живого организма.
(4) Были продолжены исследования в отношении того факта, что (а) легочный сурфактант, который изначально является физиологически активным веществом в живом организме, обладает свойством адсорбирования специфических биовеществ (Kido H., et al. FEBS Lett. Pulmonary surfactant is a potential endogenous inhibitor of proteolytic activation of Sendai virus and influenza A virus, 322 (29), 115-119, 1992), (б) легочный сурфактант секретируется из альвеолярных клеток типа II или клеток Клара и затем селективно включается в макрофаги (Akira Suwabe, J. Jpn. Med. Soc. Biol. Interface; Surfactant metabolism disorder in alveolar proteinemia, 33, 10-13, 2002), и (в) легочный сурфактант включается и метаболизируется в аналогичных клетках, например антиген-представляющих клетках (дендритных клетках).
В результате было обнаружено, что только SP-B и SP-C среди белковых ингредиентов, и липидные ингредиенты легочного сурфактанта действуют в качестве «носителя ЛА» мукозальной вакцины, которая селективно индуцирует продуцирование IgA, и что область активного ингредиента SP-B или активного домена индукции мукозального иммунитета представляет собой пептид, содержащий следующие аминокислотные последовательности:
SP-B 214-225: Leu Ile Lys Arg Ile Gin Ala Met Ile Pro Lys Gly (SEQ ID No. 7); и
SP-B 257-266: Leu Leu Asp Thr Leu Leu Gly Arg Met Leu (SEQ ID No. 8).
Кроме того, было обнаружено, что область активного ингредиента SP-C или активного домена индуцирования мукозального иммунитета представляет собой пептид, содержащий следующую аминокислотную последовательность:
SP-C 29-58: Cys Pro Val His Leu Lys Arg Leu Leu Ile Val Val Val Val Val Val Leu Ile Val Val Val Ile Val Gly Ala Leu Leu Met Gly Leu (SEQ ID No. 9).
(5) Кроме того, было обнаружено в качестве механизма селективного индуцирования IgA, что активные ингредиенты легочного сурфактанта эффективно проводят представление антигена на Т-лимфоцитах, индуцируя увеличение экспрессии МНС класса II, CD40 и В7-2 антиген-представляющих дендритных клеток и, кроме того, способствуют переключению класса на lgA-продуцирующие В-лимфоциты, индуцируя цитокины TGF-β1 в местной слизистой оболочке.
4. Задачи данного изобретения, полностью основанные на наблюдениях и элементах, описанных выше, являются следующими:
(1) Первой задачей является создание способа мукозального иммунитета. Путем обеспечения «носителя ЛА» и его использования данное изобретение достигает селективного индуцирования продуцирования антиген-специфичного IgA, который является активным веществом мукозального иммунитета, а также создает основу для индукции безопасного и эффективного (без побочных эффектов) мукозального иммунитета и способ для этого.
(2) Второй задачей является улучшение качеств, относящихся к безопасности, эффективности и однородности носителя ЛА, путем применения синтетического пептида. Данное изобретение предлагает комплекс (носитель ЛА), который приготовлен с синтетическим пептидом активного домена SP-B индуктора мукозального иммунитета (содержащего каждые аминокислотные последовательности SP-B 214-225 и SP-B 257-266, описанные выше), синтетическим пептидом активного домена SP-C индуктора мукозального иммунитета (содержащего каждую аминокислотную последовательность SP-C 29-58, описанную выше), его синтетическим аналогом, длинноцепным синтетическим пептидом, содержащим эти аминокислотные последовательности в качестве своей части, и т.п. и липидным ингредиентом легочного сурфактанта, и улучшает качество носителя ЛА.
(3) Третьей задачей данного изобретения является превращение традиционной подкожной вакцины в вакцину для мукозального введения. Данное изобретение использует носитель ЛА в инактивированной вакцине вируса, инфицирующего дыхательные пути, например инактивированной вакцине гриппа, SARS, кори, краснухи, свинки и т.п., и инактивированной вакцине вируса, инфицирующего кишечный тракт, например инактивированной вакцине ротавируса, полиовируса и т.п., и превращает вакцину для их подкожного введения в мукозальную вакцину
(4) Четвертой задачей данного изобретения является обеспечить способ использования носителя ЛА в инактивированной вакцине против вирусной инфекции через слизистую оболочку кроме дыхательных путей и кишечного тракта, например инактивированной вакцины против СПИДа, гепатита В, гепатита С и т.п.
(5) Пятой задачей является обеспечить способ использования носителя ЛА в ДНК вакцине, живой вакцине, предотвращение и лечение аллергии и т.п.
(6) Шестой задачей является обеспечить способ использования носителя ЛА при трансдермальном введении (аппликация, наклеивание и т.п.) в качестве пути иммунитета, который может индуцировать IgA не в слизистой оболочке.
(7) Седьмой задачей является открыть путь для применения и использования носителя ЛА в земледелии, рыбоводстве и т.п., а не только как СДЛ или фармацевтических средств.
Носитель ЛА, предлагаемый согласно данному изобретению, имеет проявления и действия, отличные от адъювантов, которые традиционно использовались в иммунологии, как показано ниже. А именно, традиционные адъюванты вводили обычно подкожно или внутримышечно, что вызывало местную воспалительную реакцию и привлекало антиген-представляющие клетки или В- и Т-лимфоциты, и они содержат чужеродное вещество, сообщающее им способность быть активным ингредиентом. Кроме того, они использовались в комбинации с минеральными маслами или солями металлов, что вызывает пролонгированное действие и удержание антигена с поддержанием воспалительной реакции в течение длительного периода времени. Кроме того, известные в качестве традиционных вакцин/адъювантов, таких как описано выше, чужеродное вещество, такое как живой термолабильный токсин Escherichia coli и токсин холеры, имеют риск вызвать опасное действие или побочные эффекты. Напротив, носитель ЛА по данному изобретению не вызывает местной воспалительной реакции. Кроме того, носитель ЛА получен из биологических ингредиентов и более того, он ограничен активным ингредиентом легочного сурфактанта или его активным доменом и использует пептид с низким молекулярным весом, содержащий такой домен или область домена, для получения эффективной мукозальной вакцины. Согласно этому носитель ЛА является весьма безопасным и неинвазивным.
5. Согласно данному изобретению предложено, соответственно, следующее (1)-(5).
(1) Носитель лекарства-антигена, представляющий собой комплекс, содержащий по меньшей мере один фрагмент, выбранный из белка В легочного сурфактанта или множественных фрагментов, полученных из этого белка В, по меньшей мере один фрагмент, выбранный из белка С легочного сурфактанта или множественных фрагментов, полученных из этого белка С, и по меньшей мере один липид.
Более конкретно, носитель ЛА представляет собой комплекс, содержащий по меньшей мере в сумме три вещества, которые выбраны по меньшей мере в виде одного типа каждой из следующих групп: Группы I (группа белка В легочного сурфактанта, и природные и синтетические полипептиды, полученные или происходящие из этого белка В), Группы II (группа белка С легочного сурфактанта, и природные и синтетические полипептиды, полученные или происходящие из этого белка С) и Группы III (группа липидов, таких как фосфолипид, жирная кислота и т.п.). [Группа I] Белок В легочного сурфактанта и полипептид, содержащий следующую аминокислотную последовательность, описанную в SEQ ID No. 2 (аминокислоты нумеруются по порядку от Met на N-конце каждой последовательности, которая считается первой аминокислотой, к С-концу): 1-381 (SEQ ID No. 2), 214-225 (SEQ ID No. 7), 257-266 (SEQ ID No. 8), 1-20 (SEQ ID No. 10), 102-110 (SEQ ID No. 11), 119-127 (SEQ ID No. 12), 136-142 (SEQ ID No. 13), 171-185 (SEQ ID No. 14), 201-279 (SEQ ID No. 15), 253-278 (SEQ ID No. 16), 300-307 (SEQ ID No. 17), 317-330 (SEQ ID No. 18), 344-351 (SEQ ID No. 19), 358-381 (SEQ ID No. 20), полипептид, содержащий по меньшей мере одну последовательность из вышеуказанных аминокислотных последовательностей в качестве активного домена, полипептид, полученный в результате замещения и/или делеции по меньшей мере одной аминокислоты в вышеуказанных аминокислотных последовательностях, его синтетический аналог, его основа, модифицированная сахаридом или сахаридной цепью и т.п.
[Группа II] Белок С легочного сурфактанта и полипептид, содержащий следующую аминокислотную последовательность, описанную в SEQ ID No. 4 (аминокислоты нумеруются по порядку от Met на N-конце каждой последовательности, который считается первой аминокислотой, к С-концу): 1-197 (SEQ ID No. 4), 29-58 (SEQ ID No. 9), 24-58 (SEQ ID No. 21), полипептид, содержащий аминокислотную последовательность с номерами 1-191 последовательности SEQ ID No. 6, полипептид, содержащий по меньшей мере одну последовательность из вышеуказанных аминокислотных последовательностей в качестве активного домена, полипептид, полученный в результате замещения и/или делеции по меньшей мере одной аминокислоты в вышеуказанных аминокислотных последовательностях, его синтетический аналог, его основа, модифицированная сахаридом или сахаридной цепью и т.п.
[Группа III] Липид, такой как фосфолипид, подобный фосфатидилхолину, дипальмитоилфосфатидилхолину, фосфатидилсерину, дипальмитоилглицерофосфохолину, диацилглицерофосфоглицеролу, фосфатидилглицерину, фрсфатидилинозитолу, фосфатидилэтаноламину, фосфатидной кислоте и т.п., жирная кислота, подобная лауриновой кислоте, миристиновой кислоте, пальмитиновой кислоте, стеариновой кислоте, олеиновой кислоте, и т.п., и т.п.
Кроме того, носитель лекарства-антигена предпочтительно имеет форму и структуру, где вышеуказанная Группа III представляет собой липидную мембрану в форме листа или рулона, и множественные цепи каждой из вышеуказанных Группы I и Группы II внедрены в форме шипов (выступов) с концами гидрофобных областей, вдающимися в липидную мембрану.
(2) Мукозальная вакцина, которая характеризуется индуцированием мукозального иммунитета, который получается путем подвергания носителя лекарства-антигена вышеуказанного (1) сосуществованию, контакту, захвату или адсорбции с антигеном.
(3) Агент для предотвращения и лечения аллергии, который характеризуется индуцированием мукозального иммунитета, полученный путем подвергания носителя лекарства-антигена вышеуказанного (1) сосуществованию, контакту захвату или адсорбции с аллергеном. Действие и эффективность имеет место за счет дезактивации и снижения сенситизации к аллергену, такому как кедровая пыльца, приносимая воздухом и всасываемая, клещ и т.п., мукозальными IgA в носовой полости или носовой части глотки.
(4) Трансмукозальная и/или транодермальная СДЛ, которая получена подверганием носителя ЛА вышеуказанного (1) сосуществованию, контакту, захвату или адсорбции с лекарством в эффективном количестве.
(5) Способ индуцирования мукозального иммунитета, отличающийся тем, что водят мукозальную вакцину, которая получена подверганием носителя ЛА вышеуказанного (1) сосуществованию, контакту, захвату или адсорбции с антигеном, в нос или верхние дыхательные пути.
Индуцирование мукозального иммунитета по изобретению в вышеуказанных (2), (3) и (5) предпочтительно характеризуется усилением продуцирования IgA антитела в местной слизистой оболочке, и далее усилением продуцирования TGF-β1 и цитокинов Th2-типа в местной слизистой оболочке.
6. Далее здесь будут пояснены воплощения данного изобретения.
(1) Состав носителя ЛА
Ниже следуют массовые % сухого вещества трех вышеуказанных групп: Группы I (группа белка В легочного сурфактанта и природных и синтетических полипептидов, полученных или происходящих из этого белка В), Группы II (группа белка С легочного сурфактанта и природных и синтетических полипептидов, полученных или происходящих из этого белка С) и Группы III (группа липидов, таких как фосфолипид, жирная кислота и т.п.): от около 0,1 до около 6,0 мас.% Группы I, от около 0,1 до около 6,0 мас.% Группы II и от около 88 до около 99,8 мас.% Группы III. При приготовлении носителя лекарства-антигена композиция доводится до: Группа I% + Группа II% + Группа III% = 100% по массе.
(2) Приготовление носителя ЛА
Способы приготовления являются следующими, например: взвешивают соответственно 2 мг Группы I, 2 мг Группы II и 96 мг Группы III (Группа I% + Группа II% + Группа III% = 100% по массе) и суспендируют до однородности в 5 мл изотонического раствора, например физиологического раствора или фосфатно-солевого раствора (PBS). Полученную суспензию используют в виде раствора носителя лекарства-антигена (100 мг/5 мл). Носитель готовят для каждого использования. Для получения суспензии может быть использован ультразвук, гомогенизатор, миксер, шейкер и т.п. Ультразвук при избыточном воздействии вызывает изменение в жидкости (увеличение вязкости), и, таким образом, предпочтительно использование миксера, например камерного миксера (например миксера Вортекс (Vortex, товарный знак)).
Для спецификации может быть принято 96 мг липида Группы III, например смеси 71 мг фосфатидилхолина, 21 мг фосфатидилглицерина и 4 мг фосфатидилсерина и т.п. (общее количество липидов 96 мг). Кроме того, в случае использования препаратов легочных сурфактантов, имеющихся в продаже для лечения RDS, о которых точно известно, что они содержат SP-B и SP-C, но не SP-A и SP-D, суспензию, приготовленную в соответствии с инструкцией по использованию, можно применять как таковую или как раствор носителя лекарства-антигена.
(3) Приготовление мукозальной вакцины
Мукозальную вакцину готовят путем добавления и смешивания раствора носителя лекарства-антигена с неразведенным раствором вакцины, так что отношение сухого веса количества носителя лекарства-антигена (Н) к количеству антигена в вакцине (А) составляет от около 0,2 до около 5. Например, если содержание антигена 1 мкг/мл в 1000 мл неразведенного раствора вакцины, и весовое соотношение принято как А/Н=1, добавление и смешивание количеств раствора носителя лекарства-антигена (100 мг/5 мл), который был приготовлен в вышеупомянутом (2), составляет 50 мкл. Для однородного смешивания может быть использован гомогенизатор, миксер, шейкер, мешалка и т.п.
(4) Приготовление трансмукозальной и трансдермальной СДЛ
Трансмукозальная или трансдермальная СДЛ может быть приготовлена с использованием лекарства вместо антигена таким же образом, как в вышеуказанном (3).
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее устройство и эффекты данного изобретения будут объяснены в деталях на иллюстративных примерах. Однако данное изобретение не ограничено только этими примерами, объяснениями и описанием.
Исходные материалы, способы проведения экспериментов и т.п., использованные в этих примерах, раскрыты ниже.
(1) Легочный сурфактант
Легочный сурфактант, используемый в качестве «носителя лекарства-антигена» (носитель ЛА), представляет собой препарат из бычьего легкого, полученный способом Howgood, et al. (PSF-1) (Howgood S, et al.,: Effects of a surfactant-associated protein and calcium ions on the structure and surface activity of lung surfactant lipids. Biochemistry, 24, 184-190, 1985), или в существенной степени препарат, полученный экстрагированием первичного препарата 1-бутанолом для удаления водорастворимых белковых ингредиентов, SP-A и SP-D, или уменьшения их до предела обнаружения или менее (PSF-2) (Haagasman HP, et al.,: The major lung surfactant protein, SP28-36, is a calcium-dependent, carbohydrate binding protein, J. Biol. Chem. 262, 13977-13880, 1987), и далее любой синтетический пептид из активных областей липидорастворимых белков, SP-B или SP-C, который содержит 40% по весу или более от целого фосфолипида, содержащего в основном фосфатидилхолин и дипальмитоилфосфатидилхолин, и далее содержит 10-20% фосфатидилглицерина и 2-5% фосфатидилсерина, которые сходны с липидом легочного сурфактанта, или препарат, содержащий 0-3,5% обоих пептидов в комбинации. Среди этих препаратов были исследованы препараты, содержащие оба пептида в комбинации активных областей SP-B и SP-C (PSF-3), образец, содержащий синтетический пептид активной области SP-B (PSF-4) и образец, содержащий синтетический пептид активной области SP-C (PSF-5). В качестве альтернативы также используются известные образцы, соответствующие PSF-1 или PSF-2, например Survanta, Infasurf, Curosurf, Humansurf, Exosurf, Alveofact и т.п. (товарные знаки).
(2) Животное
6-недельные самки BALB/c мыши и 10-недельные морские свинки Hartley были приобретены в Japan SLC, Inc. (Shizuoka, Japan) и использованы. Все эксперименты на животных были проведены в клетке для инфицированных животных (уровень Р2) центра экспериментов на животных медицинского факультета Университета Токушима (University of Tokushima) и осуществлены в соответствии с руководством комитета по экспериментам на животных медицинского факультета Университета Токушима.
(3) Приготовление противогриппозной вакцины сплит-типа
Приготовление противогриппозной вакцины сплит-типа было осуществлено по следующей методике, использующей суспензию, полученную из выращенной яйцеклетки, в которую был введен вирус гриппа штамма A Aichi/68/2/H3N2 (1×108 бляшкообразующих единиц [PFU, от plaque forming unit]) (предоставленного профессором Masanobu Ouchi из класса микробиологии медицинской школы Кавасаки (Kawasaki)), β-пропиолактон (Wako Pure Chemical Industries, Ltd., Osaka, Japan) был добавлен к суспензии вируса, которая была диализирована в 0,004 М PBS (TakaraBio, Inc., Shiga, Tokyo, Japan) в течение ночи до 0,05% содержания жидкости и 8 нМ конечной концентрации и инкубирована в течение 18 часов на ледяной бане. Затем она была инкубирована в течение 1,5 часов при 37°С для гидролиза β-пропиолактона. Затем был добавлен Tween 20 (Wako Pure Chemical Industries, Ltd.) до 0,1% конечной концентрации и далее добавлен диэтиловый эфир (Wako Pure Chemical Industries, Ltd.) в том же количестве, что и Tween, и смесь перемешана со встряхиванием в течение двух часов при 4°С. Этот раствор центрифугировали на 2000 об/мин в течение 5 минут, чтобы собрать водный слой. Далее осуществили удаление диэтилового эфира из водного слоя с помощью Automatic Environmental SpeedVac System (SAVANT INSTRUMENTS, INC., New York, USA). Затем отфильтровали на 0,45 мкм фильтре (MILLIPORE, Massachusetts, USA) для получения инактивированной противогриппозной вакцины сплит-типа.
(4) Иммунизация
К противогриппозной вакцине сплит-типа, полученной способом, описанным выше, были добавлены при перемешивании вышеупомянутый легочный сурфактант (PSF-1), легочный сурфактант, экстрагированный 1-бутанолом (PSF-2), синтетические пептиды активных областей SP-B и SP-C и липиды легочного сурфактанта (PSF-3, -4 и -5), известный препарат легочного сурфактанта или субъединица В токсина холеры (СТВ, SIGMA, Missouri, USA) в качестве иммуноадъюванта. Легочный сурфактант или соответствующий препарат, описанный выше, суспендировали в PBS in situ в концентрации, требуемой для введения вакцины, и обрабатывали ультразвуком в течение 5 мин при комнатной температуре до получения однородной суспензии. Добавляли 0,1 мкг противогриппозной вакцины сплит-типа на 0,1 мкг сухого веса легочного сурфактанта или соответствующего препарата, перемешивали на мешалке вортекс (vortex) и оставляли на 1 час при комнатной температуре до использования. СТВ добавляли in situ аналогичным образом и смешивали 0,1 мкг на 0,1 мкг противогриппозной вакцины сплит-типа (Watanabe I, et al.: Characterization of protective immune responses induced by nasal influenza vaccine containing mutant cholera toxin as a safe Adjuvant (CT112K). Vaccine 2002; 20: 3443-55).
Для назального введения вакцины вышеуказанный отрегулированный продукт разбавляли в PBS до 0,1 мкг/1 мкл раствора фосфатно-солевого буфера (PBS, от phosphate buffered saline) и вводили в обе стороны по 1 мкл, соответственно, одному животному, то есть всего 2 мкл было введено через нос в обе стороны носовой полости мыши, анестезированной эфиром. Для традиционной подкожной инъекции, которую проводили для сравнения, противогриппозную вакцину сплит-типа разбавляли в PBS до получения раствора 0,1 мкг/50 мкл и вводили в гиподерму шеи мыши. В контрольной группе PBS был введен в том же количестве, что и вакцинный раствор. Через 4 недели проводили вторую иммунизацию тем же образом, что и первую иммунизацию, а через две недели после второй иммунизации готовили растворы смывов из носовой полости и альвеол и сыворотку мыши, которые использовались для измерения вирус-специфичных IgA и IgG и исследования вирусной инфекции.
(5) Приготовление растворов смывов из носовой полости и альвеол и сыворотки мыши
Мышь, которой была введена вакцина, была вскрыта разрезанием брюшной полости и грудной клетки под анестезией фенобарбиталом, трахея также вскрыта разрезанием, в легкое был введен клиновидный венозный катетер Atom 3 Fr (Atom medical Corporation, Tokyo, Japan), был влит 1 мл физиологического раствора, и раствор был собран. Это повторяли трижды, и собранный раствор в общем объеме 3 мл был использован в качестве раствора смыва из альвеол. После сбора раствора смыва из альвеол венозный катетер atom был помещен через открытую трахею в направлении носовой полости, и был влит 1 мл физиологического раствора, а раствор, выходящий из носа, собран. Этот раствор был использован в качестве раствора смыва из носа. Далее забор крови осуществлялся из сердца, и сыворотку готовили центрифугированием на 5000 об/мин в течение 10 минут.
(6) Количественное определение белка
Содержание белка в растворах смывов из носа и легких и в сыворотке измерялось с помощью набора ВСА Protein Assay Reagent Kit (PIERCE, Illinois, USA) (Smith PK., et al.,: Measurement of protein using bicinchoninic acid. Anal. Biochem., 150, 76-85, 1985). Оптическую плотность на 562 нм измеряли с помощью SPECTRAmax PLUS 384 (Molecular Devices Corporation, California, USA).
(7) Исследование вирусной инфекции и оценка величины инфекции.
Для инфицирования был использован штамм вируса гриппа, штамм А Aichi/68/2/H3N2, который являлся тем же, что и штамм, использованный при приготовлении противогриппозной вакцины сплит-типа. Через две недели после завершения второй иммунизации мышей анестезировали эфиром, и суспензию, полученную из яйцеклетки, в которой рос вирус гриппа, закапывали для инфицирования в обе стороны носовой полости в количестве 7×104 PFU/3 мкл. Через три недели после инфекции растворы смыва из носовой полости и альвеол готовили способом, описанным выше, и использовали для оценки величины вирусной инфекции. Оценку величины вирусной инфекции осуществляли с использованием клеток А 549 (предоставленных профессором Masanobu Ouchi из класса микробиологии медицинской школы Kawasaki). 549 клеток инкубировали в условиях 5% фетальной бычьей сыворотки/DMEM (Gibco, New York, USA). 549 клеток субкультивировали до 100% конфлуентности в 6 планшетах для инкубации (Greiner Deutsche Stuttgart) и заменяли на бессывороточную среду через 24 часа. Растворы смыва из носовой полости и легких мышей, инфицированных гриппом, раскапывали по 500 мкл, соответственно, в каждую лунку, и инкубировали в СО2 инкубаторе при 37°С в течение 12-16 часов. К этому раствору добавили 1% PBS раствор эритроцитов, которые получили из морской свинки, и смесь оставляли на 5 мин при комнатной температуре. Смесь промывали 1 мМ Са2+/Mg2+ PBS, и оценку величины вирусной инфекции проводили подсчетом клеток, агглютинирующих эритроциты, как клеток, инфицированных вирусом (Tashiro M., Homma М.: Pneumotropism of Sendai virus in relation to protease-mediated activation in mouse lungs. Infect. Immun. 39, 879-888, 1983).
(8) Очистка специфичных противогриппозных антител IgA и IgG
Очистка специфичных противогриппозных антител IgA и IgG осуществлялась как описано ниже для использования в качестве стандарта при количественном определении в анализе ELISA. Фракции IgG выделяли из растворов смыва из легких мышей, которым была введена вакцина и которые были инфицированы вирусом, с помощью аффинной хроматографии на колонке с 4 В сефарозой и рекомбинантным экспрессируемым Белком G Е.coli (ZYMED LABORTORIES INC, San Francisco, USA). Козьи IgG против мышиного IgA (SIGMA) были связаны на колонке с BrCN-активированной 4 В сефарозой (Amersham Bioscience, New Jersey, USA), и с их использованием фракции IgA были очищены аффинной хроматографией от проходящей фракции Белка G. Для очистки вирус-специфичных антител из этих IgG и IgA фракций инактивированная противогриппозная вакцина сплит-типа, использованная для иммунизации, была связана на колонке с BrCN-активированной сефарозой, и с ее использованием антиген-аффинной хроматографией из IgG и IgA фракций были очищены противогриппозные специфичные IgA и IgG, соответственно. Связывание белка гриппа сплит-типа в качестве лиганда с колонкой было осуществлено с помощью реакции связывания в 0,1 М NaHCO3/0,5 М NaCl буферном растворе (рН 8,5) с удалением несвязанного лиганда буферным раствором 0,1 М уксусная кислота/0,5 М NaCl (рН 8.5) и нейтрализацией с помощью PBS (рН 7,5). После реакции аффинного связывания в PBS (рН 7,5) и удаления несвязанных антител, каждая из аффинных хроматографий была подвергнута элюированию специфичных антител буферным раствором глицин-НСl (рН 2,8). Элюированные фракции были немедленно нейтрализованы с помощью буферного раствора 0,5 М Tris HCI (рН 9,0) и диализированы в MilliQ воду и лиофилизированы, чтобы использовать растворенными в PBS in situ.
(9) Количественное определение противогриппозных антител
Содержание противогриппозных IgA и IgG в растворах смывов из носовой полости и альвеол и в сыворотке было количественно определено в анализе ELISA. Анализ ELISA был осуществлен согласно способу для набора Mouse ELISA Quantitization фирмы BETHYL LABORATORIES (Texas, USA). В каждую лунку 96-луночного иммунопланшета Nunc (Nalgen Nunc International, New York, USA), был добавлен 1 мкг вакцины и 100 мкл 1 мкг/мл PBS раствора бычьего сывороточного альбумина (BSA, SIGMA, Missouri, USA) и реакцию по затвердеванию слоя осуществляли в течение ночи при 4°С. Затем каждую лунку промывали трижды промывным раствором (50 мМ Tris, 0,14 М NaCl, 0,05% Tween 20, pH 8,0) для удаления раствора вакцины. В каждую лунку добавляли 50 мМ Tris-HCl буферного раствора, содержащего 0,15 М NaCl и 1% BSA, и реакцию блокировали при комнатной температуре в течение 1 часа. Каждую лунку трижды промывали промывным раствором и затем добавляли 100 мкл растворов смывов из носовой полости и альвеол и сыворотку, разбавленные подходящим количеством буферного раствора для связывания препарата (50 mM Tris, 0,15 М NaCl, 1% BSA, 0,05% Tween 20, pH 8,0), и смесь оставляли для реагирования при комнатной температуре на 2 часа. Реакцию окрашивания проводили с ТМВ Microwell Peroxidase Substrate System (Kirkegaard & Perry Laboratories, Inc. Maryland, USA), используя в качестве вторичных антител конъюгированные с пероксидазой хрена (HRP) козьи антитела против мышиного IgA или IgG (BETHYL LABORATORIES INC.). Реакцию останавливали добавлением 100 мкл 2М H2SO4 (Wako Pure Chemical Industries, Ltd.) в каждую лунку и измеряли оптическую плотность на 450 нм с помощью SPECTRAmax PLUS 384. 10 нг противогриппозных IgA и IgG, очищенных из указанного выше раствора смыва из легких, обрабатывали таким же образом как стандарт для количественного определения и использовали полученную оптическую плотность.
(10) Приготовление дендритных клеток и проточная цитометрия
Приготовление дендритных клеток осуществляли из носа, легкого и селезенки мыши из каждой группы (4 в одной группе) через 2 дня после первой иммунизации по способу Gonzalez-Juarrero М (Gonzalez-Juarrero М, Orme IM.: Characterization of murine lung dendritic cells infected with Mycobacterium tuberculosis. Infect Immun. 2001; 69: 1127-33).
Приготовление дендритных клеток из носа и обработку их коллагеназой осуществляли согласно методу Асанума (Asanuma H, et al.: Characterization of mouse nasal lymphocytes isolated by enzymatic extraction with collagenase. J. Immunol. Methods 1995; 187: 41-51). Дендритные клетки, приготовленные из каждой ткани, промывали 1 мМ EDTA/PBS и добавляли на каждые 106 клеток, где каждая 1 мкг/мл FIТС-конъюгированных Anti-IА/IЕ (МНС класс II) и РЕ-конъюгированных Anti CD40 или F.IТС-конъюгированных Anti-CD80 (B7-1) и РЕ-конъюгированных Anti CD86 (В7-2) (BD Bioscience, New Jersey, USA), и оставляли для взаимодействия на 30 мин на льду в виде 50 мкл 1 мМ суспензии EDTA/PBS. Несвязанные антитела удаляли, осуществляя дважды промывку с помощью 1 мл 1 мМ EDTA/PBS, и ресуспендировали клетки в 1 мл 1 мМ EDTA/PBS. С использованием этого осуществляли определение фактора модификации на поверхности клеток с помощью BD FACS Callibur (BD Bioscience).
(11) Количественное определение TGF-β1
Количество секретированного TGF-β1 в растворах смыва из носовой полости и альвеол определяли количественно в анализе ELISA. Анализ ELISA проводили с использованием набора TGF-β1 ELISA (BIOSOURCE INTERNATIONAL, California, USA), согласно инструкциям, приложенным к набору.
(12) Количественное определение различных цитокинов
Количество секретированных цитокинов, соответственно, в растворах смывов из носовой полости и альвеол и из лимфоцитов селезенки (интерлейкины: ИЛ-4, ИЛ-5, ИЛ-6 и ИЛ-13), количественно определяли с помощью коммерческого набора ELISA.
(13) Приготовление PSF-3, содержащих синтетические пептиды SP-B и SP-C и фосфолипид
Каждый пептид SP-B 253-278 (SEQ ID No. 16) и SP-C 24-58 (SEQ ID No. 21) был химически синтезирован известным способом. Эти пептиды добавляли к фосфолипидной мембране (дипальмитоилфосфатидилхолин (75), фосфатидилглицерин (25) и пальмитиновая кислота (10)) для получения плоской фосфолипидной мембраны и готовили носитель ЛА PSF-3.
ПРИМЕР 1
Сравнение назальной противогриппозной вакцины и противогриппозной вакцины для подкожных инъекций в отношении ингибирующего действия на рост вируса
В качестве назальной вакцины было введено 0,1 мкг противогриппозной вакцины сплит-типа в обе ноздри мыши BALB/c по 1 мкл, соответственно как раствор PBS одного или с 0,1 мкг экстрагированного 1-бутанолом (исключая SP-A и SP-D) сурфактанта (ниже PSF-2) в качестве «носителя ЛА», или 0,1 мкг СТВ. В качестве вакцины для подкожной инъекции в гиподерму шеи BALB/c мыши была введена вакцина в том же количестве, что и назальная вакцина, в виде в общем 50 мкл раствора PBS, одного или с добавлением PSF-2 в качестве носителя ЛА или СТВ в качестве адъюванта. Через 4 недели была осуществлена вторая иммунизация тем же образом, что и первая иммунизация. Контрольной группе было введено то же количество PBS, соответственно. Через 2 недели после второй иммунизации осуществлена назальная инфекция вирусом гриппа в количестве 6,6×104 PFU в виде 3 мкл раствора PBS. Через 3 дня после инфекции мышь была убита, были приготовлены растворы смыва из носовой полости и альвеол, и с их использованием была проведена оценка величины вирусной инфекции (m=15-20; cpeднee ± SE; * уровень значимости по Т-тесту был p<0,01 в группе введения вакцины).
Как показано на Фиг.1 (а) и (б), в случае назального введения противогриппозной вакцины, рост вируса гриппа в растворах смывов из носовой полости и альвеол был ингибирован даже при независимом введении вакцины, но PSF-2 или СТВ усиливал эффекты значительно и рост вируса гриппа был почти совершенно подавлен, убеждая в эффектах вакцины.
Хотя и не показано на фиг.1 (а) и (б), но в случае использования PSF-1 или PSF-3 вместо PSF-2, были получены схожие эффекты. Схожие явления имели место для PSF-4 и -5, но эффекты были ослаблены.
Хотя и не показано на фиг.1, даже в случае, когда PSF-2 или СТВ были введены назально независимо, соответственно при первой иммунизации и второй иммунизации, ни в одном случае не были обнаружены эффекты подавления роста вируса, следовательно, определили, что эффекты PSF-2 или СТВ являются эффектами усиления по отношению к эффектам вакцины.
С другой стороны в случае подкожной инъекции противогриппозной вакцины, как показано на (в) и (г) на Фиг.1, титр (PFU) вируса гриппа в растворах смывов из носовой полости и альвеол был значительно уменьшен даже самостоятельной вакциной, так что была обнаружена эффективность вакцины, но эффективности усиления PSF-2 или СТВ не были видны. Другими словами, в случае подкожного введения, эффективности усиления иммунитета PSF-2 или СТВ не были обнаружены или были обнаружены, но очень слабые. Хотя и не показано на фиг.1, в этом эксперименте даже в случае, когда PSF-2 или СТВ введены подкожно самостоятельно, соответственно при первой иммунизации и второй иммунизации, эффективности подавления роста вируса не были обнаружены ни в одном случае. Хотя и не показано на фиг.1, в случае применения PSF-1, PSF-3, PSF-4 и PSF-5 вместо PSF-2 не были обнаружены эффективности усиления действия вакцины.
ПРИМЕР 2
Влияние PSF-2 или СТВ на продуцирование специфичных противогриппозных антител (IgA и IgG) в растворе смыва из носовой полости после (а) назального введения и (б) введения подкожной инъекцией противогриппозной вакцины
Тем же образом, что описан для Фиг.1, в качестве назальной вакцины было введено 0,1 мкг противогриппозной вакцины сплит-типа в обе ноздри мыши BALB/c в 1 мкл, соответственно, то есть всего 2 мкл, в виде раствора PBS только или с 0,1 мкг PSF-2 в качестве «носителя ЛА», или 0,1 мкг СТВ в качестве адъюванта. В качестве подкожной вакцины в гиподерму шеи BALB/c мыши была введена вакцина в том же количестве, что и назальная вакцина, PSF-2 или СТВ в виде 50 мкл раствора PBS. Через 4 недели была осуществлена вторая иммунизация тем же образом, что и первая иммунизация. Контрольной группе было введено то же количество PBS, соответственно. Через 2 недели после второй иммунизации осуществлена назальная инфекция вирусом гриппа в количестве 6,6×104 PFU в виде 3 мкл раствора PBS. Через 3 дня после инфекции мышь была убита, был приготовлен раствор смыва из носовой полости, и с его использованием была проведена оценка величины вирусной инфекции (n=15-20; cpeднee ± SE; * уровень значимости по Т-тесту был p<0,01 в группе введения вакцины).
Результаты показаны на Фиг.2 (а) и (б). При назальном введении противогриппозной вакцины сплит-типа специфичные противогриппозные антитела IgA селективно продуцировались в носовой полости, и таким образом в растворе смыва их число возросло, но PSF-2 или СТВ увеличили количество специфичных IgA в равной степени значительно. С другой стороны, даже в случае подкожной инъекции, было обнаружено возрастание количества специфичных IgA в растворе смыва из носовой полости от самостоятельной противогриппозной вакцины сплит-типа, однако степень его была ниже, чем для назального введения. Кроме того, в случае подкожной инъекции вакцины, эффективность усиления иммунитета PSF-2 или СТВ не была обнаружена ни в одном случае при продуцировании IgA или IgG. Хотя и не показано на фиг.2, но в случае использования PSF-1 или PSF-3 вместо PSF-2, были получены сходные эффективности. Схожие явления наблюдались для PSF-4 и -5, но эффективности были ослаблены.
ПРИМЕР 3
Влияние PSF-2 или СТВ на продуцирование специфичных противогриппозных антител (IgA и IgG) в растворе смыва из легких после (а) назального введения и (б) введения подкожной инъекцией противогриппозной вакцины
В качестве назальной вакцины было введено 0,1 мкг противогриппозной вакцины сплит-типа в обе ноздри мыши BALB/c по 1 мкл соответственно, то есть всего 2 мкл, в виде PBS раствора одного или с 0,1 мкг PSF-2 в качестве «носителя ЛА», или 0,1 мкг СТВ в качестве адъюванта. В качестве вакцины для подкожной инъекции в гиподерму шеи BALB/c мыши была введена вакцина в том же количестве, что и назальная вакцина, PSF-2 или СТВ в виде 50 мкл раствора PBS. Через 4 недели была осуществлена вторая иммунизация тем же образом, что и первая иммунизация. Контрольной группе было введено то же количество PBS, соответственно. Через 2 недели после второй иммунизации осуществлена назальная инфекция вирусом гриппа в количестве 6,6×104 PFU в виде 3 мкл раствора PBS. Через 3 дня после инфекции мышь была убита, были приготовлены растворы смыва из легких, и с их использованием оценивали влияние носителя лекарства-антигена на продуцирование специфичных противогриппозных антител (IgA и IgG) (n=15-20; cpeднee ± SE; * уровень значимости по Т-тесту был р<0,01 в группе введения вакцины).
Как показано на Фиг.3 (а) и (б), эффективности способствования PSF-2 или СТВ продуцированию специфичных противогриппозных антител были значительными, аналогично случаю назального введения вакцины, и между ними не было большой разницы. Эти эффективности усиления иммунитета были специфичны для IgA, но не были обнаружены для IgG. В случае подкожной инъекции возрастали IgA в растворе смыва из легких, но эффективности усиления иммунитета PSF-2 или СТВ не были обнаружены аналогично случаю растворов смыва из носовой полости. Хотя и не показано на фиг.3, в этом эксперименте даже в случае, когда PSF-2 или СТВ были введены назально или подкожно самостоятельно, соответственно при первой иммунизации и второй иммунизации, ни в одном случае не было обнаружено возрастание вирус-специфичных антител IgA или IgG в растворах смыва из легких, и следовательно определили, что эффективности PSF-2 или СТВ являются эффективностями усиления по отношению к эффективностям вакцины. Хотя и не показано на фиг.3, но в случае использования PSF-1 или PSF-3 вместо PSF-2, были получены сходные эффективности. Схожие явления наблюдались для PSF-4 и -5, но эффективности были ослаблены.
ПРИМЕР 4
Влияние PSF-2 или СТВ на продуцирование специфичных противогриппозных антител (IgA и IgG) в крови после (а) назального введения и (б) подкожной инъекции противогриппозной вакцины
В качестве назальной вакцины было введено 0,1 мкг противогриппозной вакцины сплит-типа в обе ноздри мыши BALB/c по 1 мкл соответственно, то есть всего 2 мкл, в виде PBS раствора одного или с 0,1 мкг PSF-2 в качестве «носителя ЛА», или 0,1 мкг СТВ в качестве адъюванта. В качестве подкожной вакцины в гиподерму шеи BALB/c мыши была введена вакцина в том же количестве, что и назальная вакцина, PSF-2 или СТВ в виде 50 мкл раствора PBS. Через 4 недели была осуществлена вторая иммунизация тем же образом, что и первая иммунизация. Контрольной группе было введено то же количество PBS, соответственно. Через 2 недели после второй иммунизации мышь была убита, был взят образец крови из сердца, из него была получена сыворотка и с ее использованием подсчитано количество экспрессированных противогриппозных антител (n=15-20; cpeднee ± SE).
Как показано на Фиг.4 (а) и (б) для количества продуцирования противогриппозных антител IgA (белая полоса) и IgG (черная полоса), обнаружено небольшое возрастание IgA или IgG при назальном введении вакцины, но увеличения IgA или IgG из-за PSF-2 или СТВ не обнаружено, то есть эффективности усиления иммунитета не обнаружено. С другой стороны, в случае подкожной инъекции обнаружено значительное увеличение IgG и отчетливое увеличение IgA для противогриппозной вакцины сплит-типа, но также в этом случае не было увеличения продуцирования антител из-за PSF-2 или СТВ, то есть эффективности усиления иммунитета не были обнаружены. В частности, в случае подкожной инъекции, IgG возрастал специфично в крови. В случае использования PSF-1 вместо PSF-2, были получены практически аналогичные эффекты (не показано).
Кроме того, даже в случае, когда PSF-2 или СТВ были введены назально или подкожно самостоятельно, соответственно в первой и во второй иммунизации, не было обнаружено возрастания вирус-специфичных IgA или IgG в крови, и таким образом эффективности PSF-2 или СТВ были определены как эффективности усиления действия вакцины (не показано).
ПРИМЕР 5
Влияние PSF-2 или СТВ на антиген-представляющую способность дендритных клеток носа, легкого или селезенки после (а) назального введения и (б) подкожной инъекции противогриппозной вакцины
В качестве назальной вакцины было введено 0,1 мкг противогриппозной вакцины сплит-типа в обе ноздри мыши BALB/c в общем объеме 2 мкл в виде 1 мкл PBS раствора одного или с 0,1 мкг PSF-2, или 0,1 мкг СТВ. В качестве подкожной вакцины в гиподерму шеи BALB/c мыши была введена противогриппозная вакцина сплит-типа в том же количестве, что и назальная вакцина, PSF-2 или СТВ в виде 50 мкл раствора PBS. Через 2 дня мышь была убита, были приготовлены образцы дендритных клеток из носа, легкого или селезенки, и уровни экспрессии МНС класса II, CD40, В7-1 (CD80) и В7-2 (CD80) на поверхности клеток были измерены проточной цитометрией.
В результате было обнаружено возрастание экспрессии молекул, имеющих отношение к представлению антигена, CD40 и В7-2, на поверхности мембраны дендритных клеток (антиген-представляющих клеток) в носу, куда вакцину вводили с СТВ, и эффектности адъюванта были обнаружены на молекулярном уровне. В случае PSF-2, возрастание экспрессии молекул МНС II было также обнаружено в дополнение к CD40 и В7-2 в дендритных клетках носа, и было также показано, что по меньшей мере три молекулы дендритных клеток вовлечены в эффекты усиления иммунитета.
Однако не было обнаружено отчетливого изменения в CD40, В7-2 или МНС II молекулах дендритных клеток легкого и селезенки. В случае использования PSF-1 или PSF-3 вместо PSF-2, были получены сходные эффективности. Схожие явления наблюдались для PSF-4 и -5, но эффективности были ослаблены.
ПРИМЕР 6
Влияние PSF-2 или СТВ на уровень секреции TGF-β1 в слизистой оболочке (а) носовой полости и (б) альвеол при назальном введении противогриппозной вакцины
В качестве назальной вакцины было введено 0,1 мкг противогриппозной вакцины сплит-типа в обе ноздри мыши BALB/c в общем объеме 2 мкл в виде 1 мкл PBS раствора одного или с 0,1 мкг PSF-2, или 0,1 мкг СТВ. В качестве подкожной вакцины в гиподерму шеи BALB/c мыши была введена вакцина в том же количестве, что и назальная вакцина, PSF-2 или СТВ в виде 50 мкл раствора PBS. Через 4 недели была осуществлена вторая иммунизация тем же образом, что и первая иммунизация. Контрольной группе было введено то же количество PBS, соответственно. Через 2 недели после второй иммунизации мышь была убита, был приготовлен раствор смыва из носовой полости и с его использованием было осуществлено количественное определение секреции TGF-β1 (n=15-20; cpeднee ± SE; * уровень значимости по Т-тесту был p<0,01 в группе введения вакцины).
Было известно, что для дифференциации В-клеток в IgA-продуцирующие клетки (переключение класса) важна местная концентрация TGF-(31, присутствующая в продуцирующих клетках (Stavnezer, J.: Regulation of antibody production and class switching by TGF-beta. J. Immunol. 155(4), 1647-1651, 1995). И здесь была исследована концентрация TGF-β1 в местной слизистой оболочке (а) носовой полости или (б) альвеол.
Концентрация TGF-β1 в местной слизистой оболочке носовой полости или альвеол, куда была введена противогриппозная вакцина сплит-типа, значительно возрастала в присутствии PSF-2 или СТВ. Степень возрастания была такова, что существенной разницы между PSF-2 и СТВ не было. Результаты показаны на Фиг.5 (а) и (б). Было обнаружено, что PSF-2, полученное из живого организма, увеличивало концентрацию TGF-β1, которая является стимулом активирования дифференциации В клеток, секретирующих IgA, также как СТВ, представляющий собой экзогенный токсин. Хотя и не показано на фиг.5, но в случае использования PSF-1 или PSF-3 вместо PSF-2, были получены схожие эффекты. Схожие явления имели место для PSF-4 и -5, но эффекты были ослаблены. Кроме того, даже в случае, когда PSF-2 или СТВ были введены назально или подкожно самостоятельно, соответственно при первой иммунизации и второй иммунизации, не было обнаружено увеличение концентрации TGF-β1, следовательно, определили, что эффекты PSF-2 или СТВ являются эффектами усиления по отношению к эффектам вакцины (не показано).
ПРИМЕР 7
Влияние PSF-2 или СТВ на продуцирование специфичных противогриппозных антител (IgA и IgG) в (а) носовой полости, (б) альвеолах и (в) крови после назального введения противогриппозной вакцины
В качестве назальной вакцины было введено 0,2 мкг противогриппозной вакцины сплит-типа в обе ноздри мыши BALB/c по 1 мкл соответственно, то есть всего 2 мкл, в виде PBS раствора одного или с 0,2 мкг PSF-2 в качестве «носителя ЛА», или 0,2 мкг СТВ в качестве адъюванта. Через 4 недели была осуществлена вторая иммунизация тем же образом, что и первая иммунизация. Контрольной группе было введено то же количество PBS, соответственно. Через 2 недели после второй иммунизации мышь была убита в каждой группе, были приготовлены растворы смыва из носовой полости и альвеол, препарат сыворотки крови из сердца, и с их использованием проводили количественную оценку экспрессированного количества противогриппозных антител (IgA и IgG) (n=15-20; cpeднee ± SE; * уровень значимости по Т-тесту был p<0,01 относительно самостоятельного введения вакцины).
Как показано на Фиг.6 (а) и (б), в растворах смыва из носовой полости и альвеол, противогриппозные антитела IgA (синие кружки) показали значительное повышение в случае, когда были введены PSF-2 и вакцина. Однако как показано на Фиг.6 (в), возрастания IgG (красные кружки) обнаружено не было.
С другой стороны, в случае, когда были введены СТВ и вакцина, IgA и IgG возросли в растворах смыва из носовой полости и альвеол (Фиг.6 (а) и (б)), и также значительно в крови (Фиг.6 (в)).
Как описано выше, в случае СТВ, он реагировал с антигеном, введенным назально, и вызывал системный иммунный ответ, а также вызывал развитие местного иммунитета, как обычно сообщалось. С другой стороны, в случае PSF-2, он вызывал только развитие местного иммунитета.
Кроме того, J. Freek van Iwaarden, et al. (Непатентный документ 4) сообщали, что если искусственно удалить макрофаги из легкого, SP-B и липиды могут вызвать системную иммунную реакцию, но в случае когда макрофаги не удалены, они не индуцируют иммунитет. Кроме того, в этом документе количество SP-B и липидов, необходимое, чтобы индуцировать системную иммунную реакцию, составляет 250-300 мкл, что существенно отличается от вводимого количества PSF-2 в вышеуказанном примере (0,2 мкл). При этом местный мукозальный иммунитет там вообще не упоминается.
ПРИМЕР 8
Влияние PSF-2 или СТВ на различные цитокины, секретируемые из носа, легких или лимфоцитов селезенки, при назальном введении противогриппозной вакцины
В качестве назальной вакцины в верхние дыхательные пути мыши BALB/c было введено 0,2 мкг противогриппозной вакцины сплит-типа в общем объеме 2 мкл в виде 1 мкл раствора PBS только или с 0,2 мкг PSF-2, или 0,2 мкг СТВ. Через 2 недели после второй иммунизации мышь убивали и проводили количественную оценку секретированного количества TGF-β1 и цитокинов (ИЛ-4, ИЛ-5, ИЛ-6 и ИЛ-13) из носа, легких или лимфоцитов селезенки (n=15-20; среднее ± SЕ; +++, р=0,06; ++, Р=0,05; +, Р=0,01 по сравнению с введением только вакцины).
Как показано на Фиг.7 (а) - (д), было обнаружено, что TGF-β1, ИЛ-5 и ИЛ-6 значительно увеличиваются в секрете, в то время как ИЛ-4 and ИЛ-13 значительно возрастают в местной слизистой оболочке (носа, легких) после введения вакцины совместно с PSF-2. С другой стороны, не наблюдалось значительного возрастания какого-либо из цитокинов в селезенке.
На этих результатах было показано, что PSF-2 повышает цитокины Тh2-типа, которые способствуют дифференциации и индукции продуцирования В клетками IgA.
ПРИМЕР 9
Влияние PSF-3 на продуцирование специфичных противогриппозных антител (IgA и IgG) в (а) носовой полости, (б) альвеолах и (в) крови при назальном введении противогриппозной вакцины
В качестве назальной вакцины в обе ноздри мыши BALB/c было введено 0,2 мкг противогриппозной вакцины сплит-типа по 1 мкл, соответственно, то есть всего 2 мкл, в виде раствора PBS только или с 0,2 мкг PSF-3 (смесь того же количества фрагмента SP-B, то есть 253-278 пептидов, описанных в SEQ ID No. 16, и фрагмента SP-C, то есть 24-58 пептидов, описанных в SEQ ID No. 21) в качестве "носителя ЛА", или 0,2 мкг липидного ингредиента. Через 4 недели провели вторую иммунизацию таким же образом, как и первую. Контрольной группе ввели то же количество PBS соответственно. Через две недели после второй иммунизации убили мышь в каждой группе и приготовили растворы смыва из носовой полости и альвеол и препарат сыворотки крови из сердца, и с их использованием осуществили количественное определение экспрессированного количества противогриппозных антител (n=15-30; среднее ± SЕ; +, р<0,01 по отношению к введению только вакцины).
Как показано на Фиг.8 (а) и (б), в растворах смыва из носовой полости и альвеол, противогриппозные антитела IgA (синие кружки) показали значительное увеличение в случае, когда были введены PSF-3 и вакцина, но значительного увеличения IgG (красные кружки) не наблюдалось. С другой стороны, в сыворотке (в), ни IgA (синие кружки), ни IgG (красные кружки) не возросли значительно.
Промышленная применимость
«Носитель ЛА» согласно данному изобретению обладает функцией транспорта всех веществ, таких как антиген, лекарство, питательное вещество и т.п. из слизистой оболочки носа, трахеи, кишечника и т.п. или кожи в клетки, а также индуцирует предпочтительное и селективное продуцирование IgA, что делает возможным мукозальную вакцину, предотвращение и лечение аллергии, трансмукозальную и трансдермальную СДЛ, и трансмукозальное и трансдермальное введение полезного вещества, такого как лекарство, питательное вещество и т.п.. Более того, он уже одобрен для клинического применения и признан безопасным в ряде стран.
Таким образом, применение и использование «носителя ЛА» можно ожидать в очень широком круге отраслей промышленности, таких как лекарства/фармацевтические средства в биологических препаратах, СДЛ и т.п., промышленности пищи и напитков в функциональных пищевых продуктах, диетических продуктах, сельском хозяйстве и сельскохозяйственных химикатах, при выращивании и культивировании сельскохозяйственных продуктов, мероприятиях по защите от заболеваний, уничтожению насекомых и т.п., разведении рыб в вакцинах против заболеваний рыб и их введении и т.п., восстановлении или сохранении окружающей среды, против муравьев, против насекомых и т.п.
















| название | год | авторы | номер документа |
|---|---|---|---|
| МУКОЗАЛЬНАЯ ВАКЦИНА | 2011 |
|
RU2570394C2 |
| ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ДЛЯ ВВЕДЕНИЯ ЧЕРЕЗ СЛИЗИСТУЮ И СПОСОБ ПРЕДОТВРАЩЕНИЯ ГРИППА | 2004 |
|
RU2390351C2 |
| ИНДУКТОР СЕКРЕТОРНЫХ АНТИТЕЛ IgA И IgG | 2006 |
|
RU2440138C2 |
| ИММУНОГЕННЫЙ ПЕПТИД ПРОТИВ СТРЕПТОКОККОВ ГРУППЫ А | 2018 |
|
RU2775621C2 |
| МУКОЗАЛЬНАЯ ВАКЦИНА ДЛЯ ИММУНОТЕРАПИИ ЗАБОЛЕВАНИЙ, ОБУСЛОВЛЕННЫХ ВИРУСАМИ ПАПИЛЛОМЫ ЧЕЛОВЕКА, И СПОСОБ ЛЕЧЕНИЯ С ЕЕ ИСПОЛЬЗОВАНИЕМ (ВАРИАНТЫ) | 2008 |
|
RU2377305C1 |
| Фармацевтическая композиция на основе PLGA для индукции эффективного мукозального иммунного ответа | 2018 |
|
RU2742580C2 |
| ИНТРАНАЗАЛЬНАЯ ВАКЦИНА, ИНДУЦИРУЮЩАЯ КЛЕТОЧНО-ОПОСРЕДОВАННЫЙ ИММУНИТЕТ | 2019 |
|
RU2796973C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ АТТЕНУИРОВАННЫЕ ШТАММЫ Streptococcus pneumoniae, И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772131C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ ГЛИКОПРОТЕИНА G ВИРУСОВ HENDRA И NIPAH | 2012 |
|
RU2681529C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ГРИБКОВЫХ И БАКТЕРИАЛЬНЫХ ПАТОГЕНОВ | 2014 |
|
RU2717306C2 |
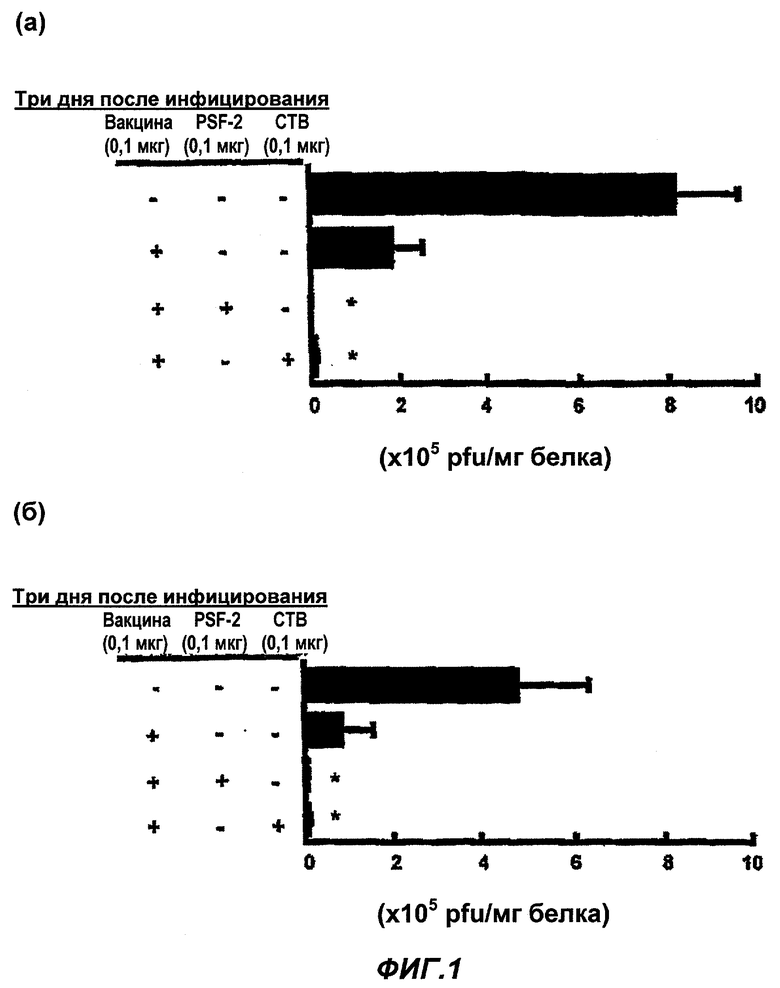
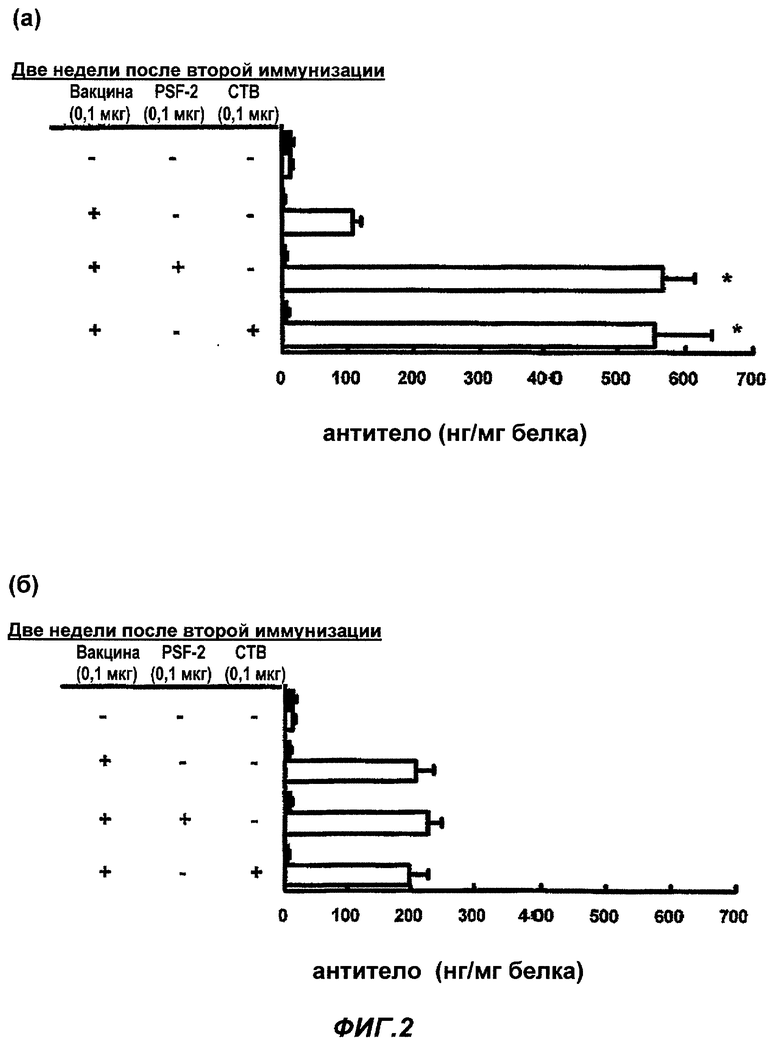
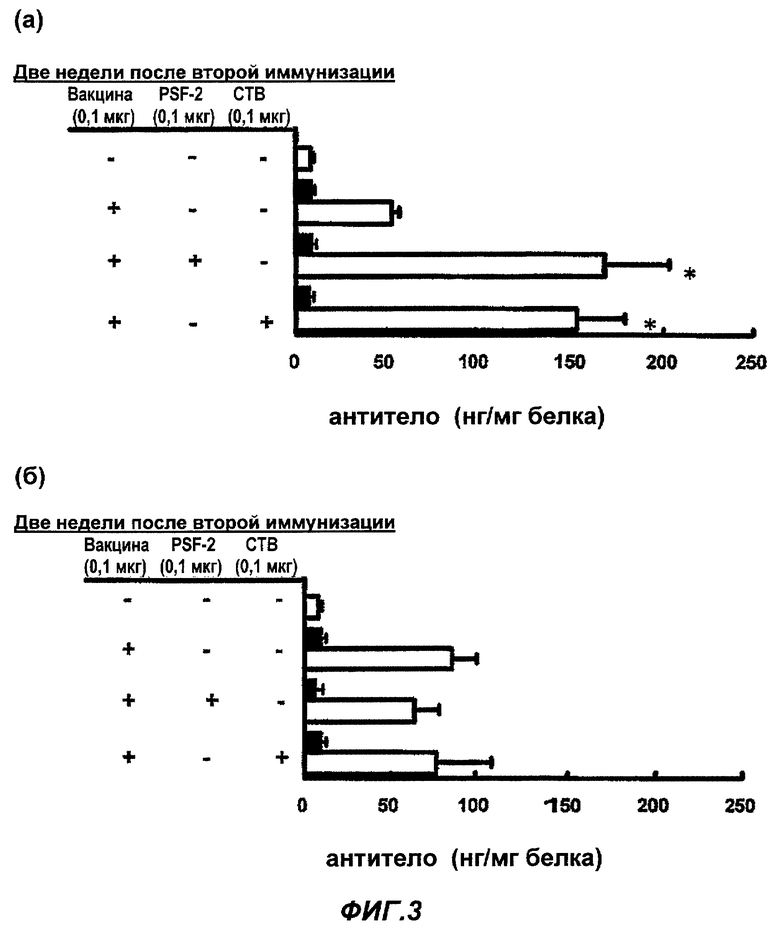
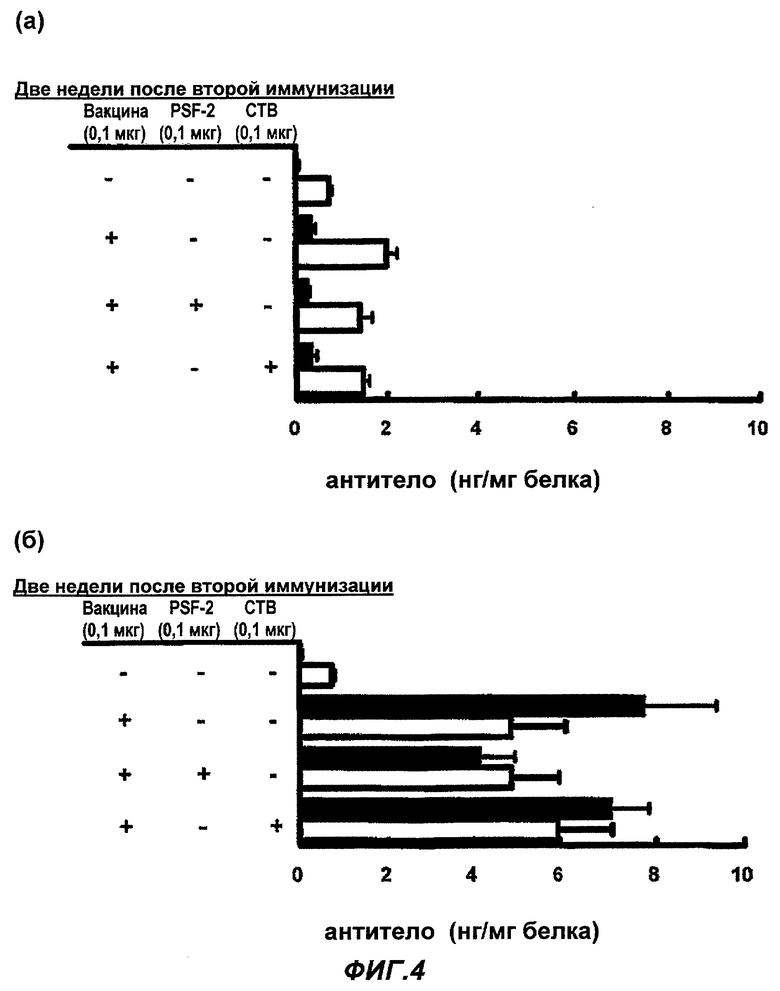
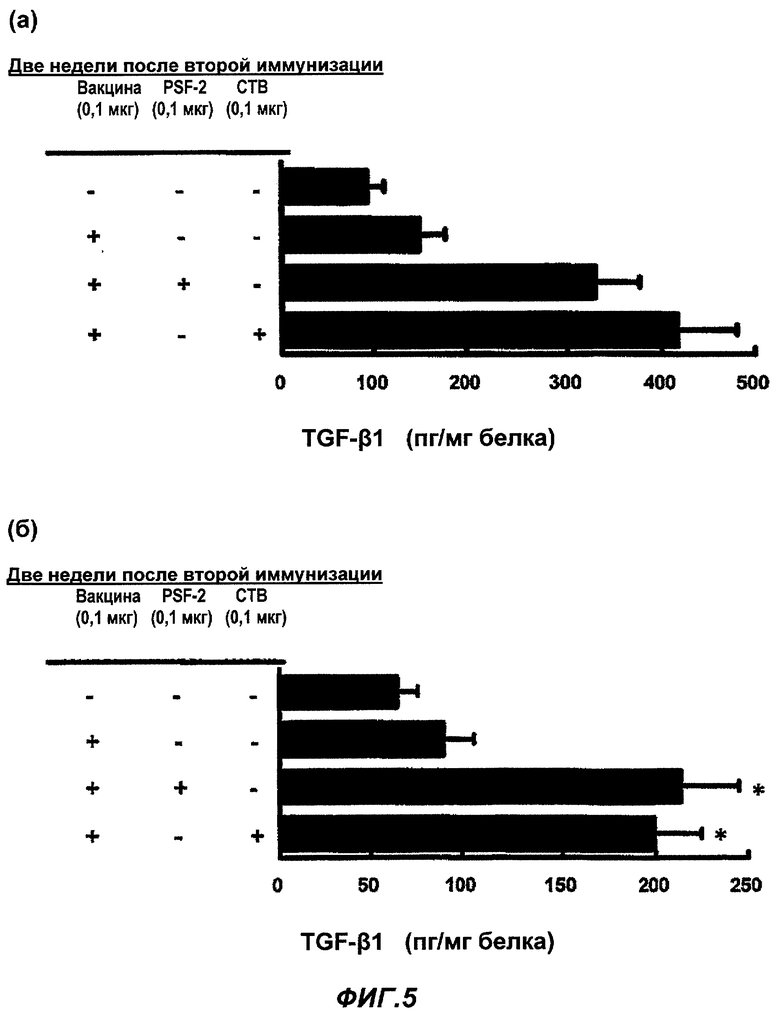
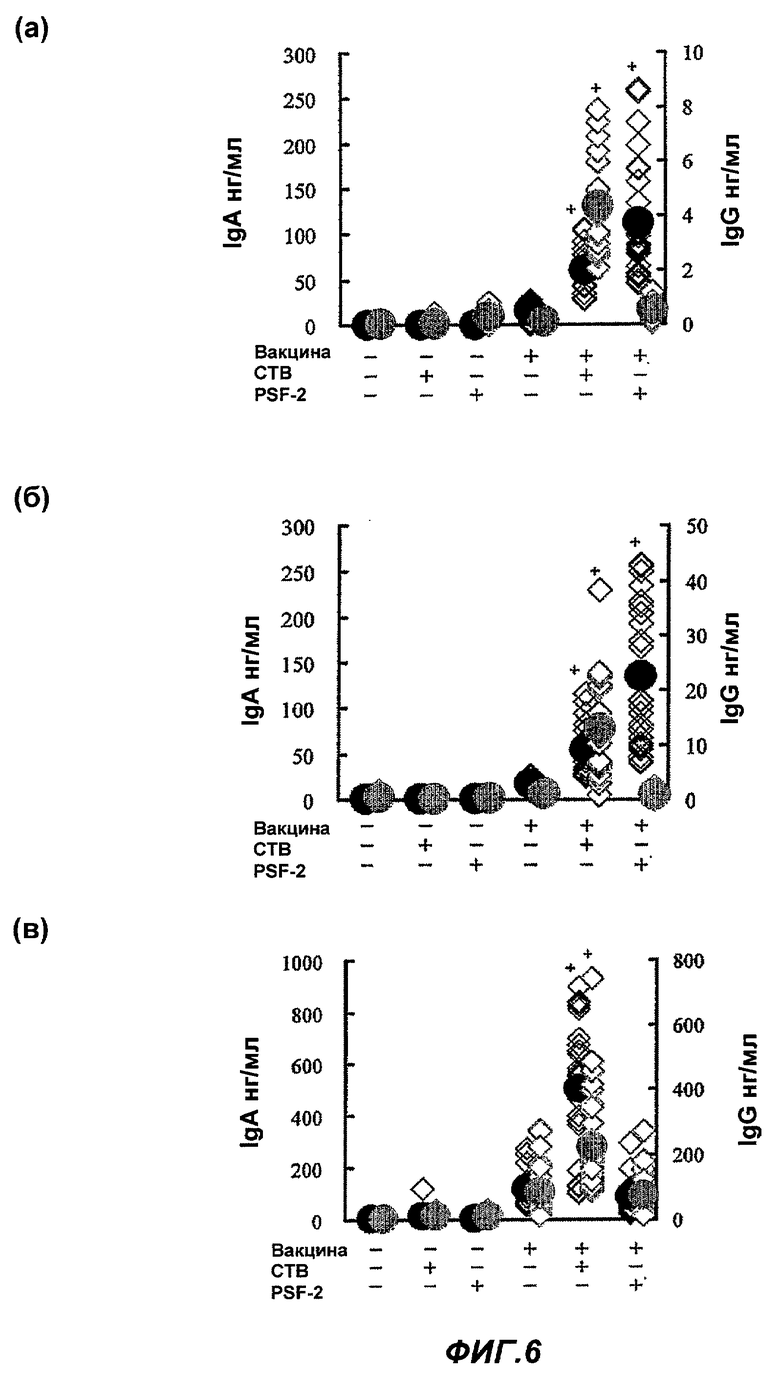
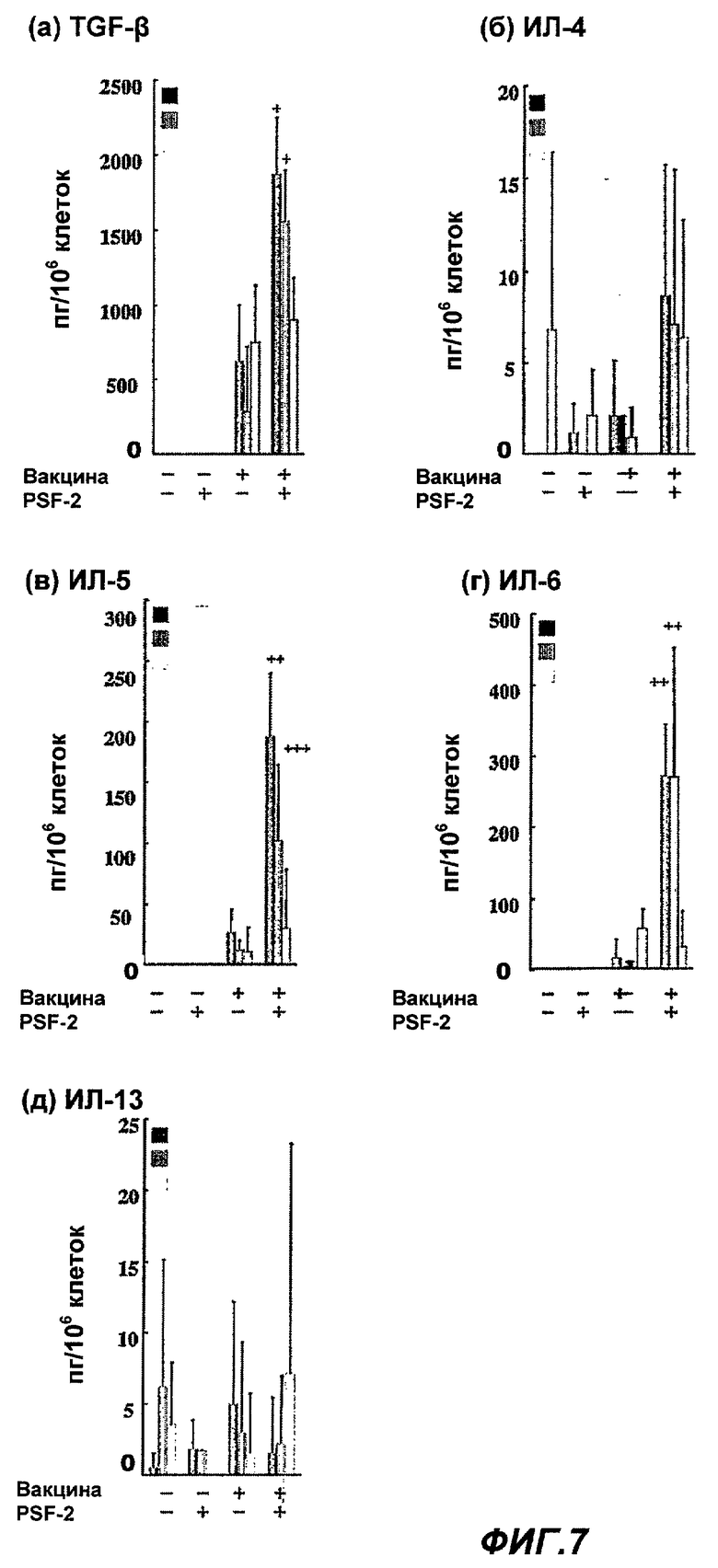
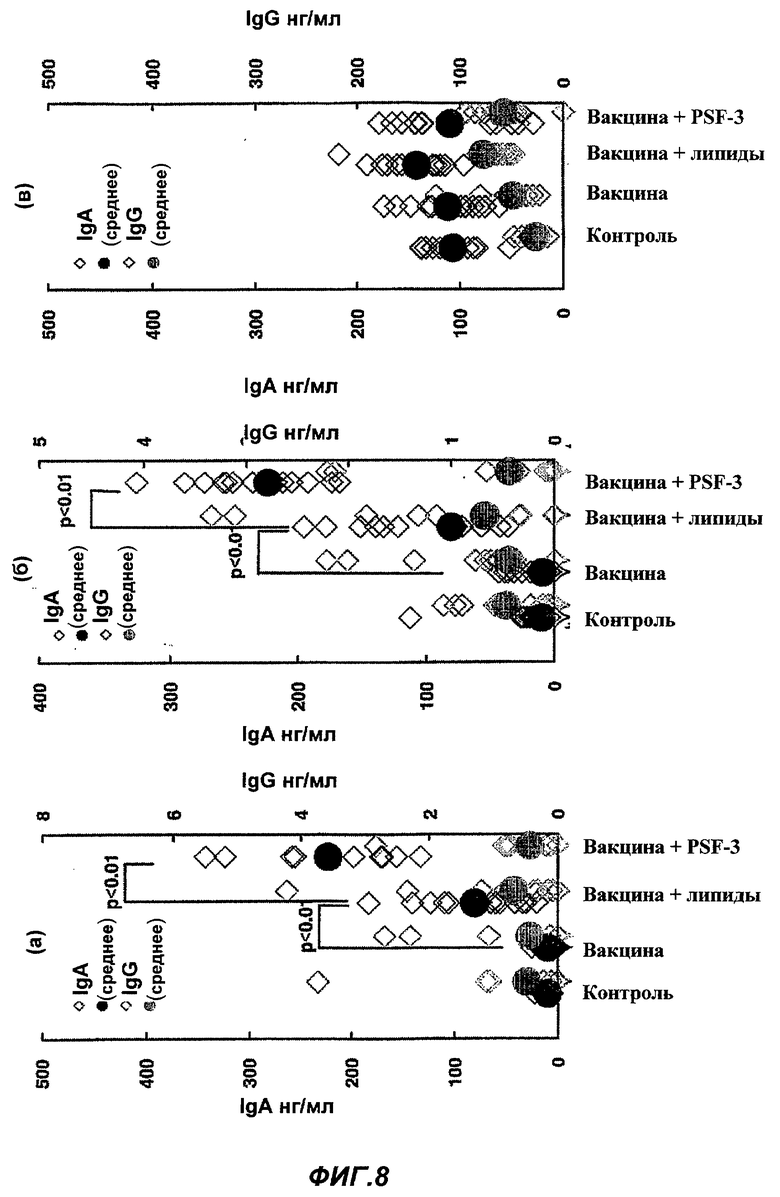
Изобретение относится к биотехнологии. Описана мукозальная вакцина, содержащая носитель лекарства-антигена, представляющий собой комплекс, содержащий белок В легочного сурфактанта или по меньшей мере один его фрагмент, белок С легочного сурфактанта или по меньшей мере один его фрагмент, и по меньшей мере один липид, и антиген, причем указанная мукозальная вакцина индуцирует терапевтически значимый мукозальный иммунитет. Изобретение позволяет индуцировать терапевтически значимый мукозальный иммунитет. 2 н. и 13 з.п. ф-лы, 8 ил.
1. Мукозальная вакцина, содержащая
(i) носитель лекарства-антигена, представляющий собой комплекс, содержащий белок В легочного сурфактанта или по меньшей мере один его фрагмент, белок С легочного сурфактанта или по меньшей мере один его фрагмент и по меньшей мере один липид, и
(ii) антиген,
причем указанная мукозальная вакцина индуцирует терапевтически значимый мукозальный иммунитет.
2. Мукозальная вакцина по п.1, где фрагмент белка В представляет собой пептид, содержащий аминокислотную последовательность SEQ ID №2, 7, 8, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20.
3. Мукозальная вакцина по п.1, где фрагмент белка С представляет собой пептид, содержащий аминокислотную последовательность SEQ ID №4, 9, 21 или 6.
4. Мукозальная вакцина по п.1, где по меньшей мере один липид выбран из липидной группы, состоящей из фосфатидилхолина, дипальмитоилфосфатидилхолина, фосфатидилсерина, дипальмитоилглицерофосфохолина, диацилглицерофосфоглицерола, фосфатидилглицерола, фосфатидилинозитола, фосфатидилэтаноламина, фосфатидной кислоты, лауриновой кислоты, миристиновой кислоты, пальмитиновой кислоты, стеариновой кислоты и олеиновой кислоты.
5. Мукозальная вакцина по п.4, где фрагмент белка В представляет собой пептид, содержащий аминокислотную последовательность SEQ ID №2, 7, 8, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20.
6. Мукозальная вакцина по п.4, где фрагмент белка С представляет собой пептид, содержащий аминокислотную последовательность SEQ ID №4, 9, 21 или 6.
7. Мукозальная вакцина по любому из пп.1-6, где указанный липид представляет собой мембрану в форме листа или в форме рулона и множественные цепи по меньшей мере одного фрагмента, выбранного из белка В легочного сурфактанта или множественных фрагментов белка В, и по меньшей мере одного фрагмента, выбранного из белка С легочного сурфактанта или множественных фрагментов белка С, внедрены в виде шипов с концами гидрофобных областей, вдающихся в липидную мембрану.
8. Мукозальная вакцина по любому из пп.1-7, где индуцирование мукозального иммунитета осуществляется стимуляцией продуцирования TGF-β1 и цитокинов типа Th2 в местной слизистой оболочке.
9. Мукозальная вакцина по любому из пп.1-7, где индуцирование мукозального иммунитета осуществляется стимуляцией продуцирования IgA антитела в местной слизистой оболочке.
10. Мукозальная вакцина по п.9, отличающаяся тем, что индуцирование мукозального иммунитета осуществляется стимуляцией продуцирования TGF-β1 и цитокинов типа Th2 в местной слизистой оболочке.
11. Мукозальная вакцина, как определено в любом из пп.1-10, для применения в терапии.
12. Применение мукозальной вакцины по любому из пп.1-7 для изготовления медикамента для индуцирования мукозального иммунитета, причем указанная мукозальная вакцина вводится в носовую полость или верхние дыхательные пути.
13. Применение по п.12, где индуцирование мукозального иммунитета осуществляется стимуляцией продуцирования TGF-β1 и цитокинов типа Th2 в местной слизистой оболочке.
14. Применение по п.12, где индуцирование мукозального иммунитета осуществляется стимуляцией продуцирования IgA антитела в местной слизистой оболочке.
15. Применение по п.14, где индуцирование мукозального иммунитета осуществляется стимуляцией продуцирования TGF-β1 и цитокинов типа Th2 в местной слизистой оболочке.
| JP 2003523348 Т, 05.08.2003 | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| J | |||
| Freek van Iwaarden et al | |||
| "Alveolar Macrophages, Surfactant Lipids and Surfactant Protein В Regulate the Induction of Immune Responses via the Airways", Am | |||
| J | |||
| Respir | |||
| Cell Mol.Biol., vol.24, p.452-458, 2001 | |||
| РЫБАЛЬСКИЙ Н.Г., СЕРОВА М.А., ИГНАТЬЕВА Г.А., СТАРЧЕУС А.П | |||
| Моноклональные антитела и гибридомы | |||
| - М.: ВАСХНИЛ, 1989. | |||

