Предлагаемое изобретение относится к нейропсихофизиологии и смежным с ней областям знаний и практики, поскольку в нем рассматривают фундаментальные основы дифференциальной психофизиологии.
Прототип - способ определения типов высшей нервной деятельности (ВНД) центральной нервной системы (ЦНС), общих для животных и людей, предложен русским физиологом И.П.Павловым [1] и сотрудниками его лаборатории [1, с.: 485-501].
Исследуя ВНД животных (собак) с помощью метода условных рефлексов, Павлов обнаружил три пары количественных свойств нервных процессов [1, 2, 3, 4, 5]:
1 - силу и слабость, которые установил по работоспособности клеток нервной ткани, их способности длительно поддерживать превышение процесса возбуждения над торможением;
2 - уравновешенность и неуравновешенность, что определил по количественному соотношению процессов возбуждения и торможения;
3 - подвижность и инертность, что охарактеризовал по скорости переключения нервной системы с возбужденного состояния на тормозное, и наоборот.
В результате Павлов выделил четыре основных типа ВНД:
I - слабый;
II - сильный неуравновешенный;
III - сильный уравновешенный подвижный;
IV - сильный уравновешенный инертный.
Эти типы ВНД Павлов считал общими для животных и людей, представляя их как совокупное проявление генетических свойств их организмов, так и возникших в течение жизни благодаря рефлекторному влиянию первой сигнальной системы (1СС). Предлагая эту классификацию, Павлов говорил об ее приблизительности, о том, что существует множество смешанных типов ВНД, промежуточных (еще 92 типа).
Говоря о совокупном проявлении генетических свойств организмов в типах ВНД, в их фенотипах, Павлов не выделял особенной роли ни соматической, ни вегетативной функции, ни их нервных систем, хотя и говорил о большом значении соматической функции в развитии ВНД животных и людей.
Однако Павлов предложил использовать параллельно свою классификацию типов ВНД с классификацией темпераментов по Гиппократу, основанной на роли той или иной жидкости у разных организмов, которая, по существу, является вегетативной классификацией темпераментов:
- меланхолического - по названию черной желчи (мелан хоул);
- холерического - соответственно, по названию желтой желчи (хоул);
- сангвинического - по названию крови (сангва);
- флегматического - по древнегреческому названию лимфы (флегма).
Тем самым Павлов, фактически, подчеркнул роль вегетативной функции в происхождении типов ВНД, общих для животных и людей, как и постоянно подчеркивал генетическую роль в происхождении типов ВНД.
Однако ни до, ни после Павлова никто какие-либо гены или генетические факторы, напрямую отвечающие за развитие нервной ткани (НТ) головного мозга (ГМ), не обнаружил ни в органах соматической, ни в органах вегетативной функции организма, ни в отделах ГМ и фокусах взаимодействия сигналов по A.M.Иваницкому [3].
Поэтому вопрос о предполагаемой генетической базе развития НТ ГМ и связанными с ней общими типами ВНД еще открыт, и что «основная проблема дифференциальной психофизиологии - понимание механизмов, обеспечивающих индивидуально-типологических особенностей человека, включая особенности его характера, все еще остается во многом не изученной» [5, с.516].
Огромное влияние на развитие типов ВНД у людей, на их фенотипы оказывает развитие второй сигнальной системы (2СС), связанной с речью. «Слово сделало нас людьми», - писал Павлов. 2СС «охватывает все виды символизации» и «использует не только речевые знаки, но и самые различные средства, включая музыкальные звуки, рисунки, математические символы, художественные образы, а также производные от речи и тесно связанные с ней реакции человека, например мимико-жестикуляционные и эмоциональные голосовые реакции, обобщенные образы, возникающие на основе абстрактных понятий, и т.п.» [3].
Учитывая взаимное влияние 1СС и 2СС, а также реально замечаемое наследственное влияние на течение нервных процессов, Павлов выделил три специфических, «человеческих» типа ВНД, идентифицировав их в виде художественного, мыслительного и среднего типов, а также говорил о четвертом, редком, гениальном.
Как и между общими типами ВНД, так и между «человеческими», считал Павлов, тоже имеется множество промежуточных.
Найденные Павловым и его школой динамические свойства НТ ГМ позволяют увидеть не только процесс формирования ВНД у фенотипа, но и - на уровне генотипа животных и человека.
Основными недостатками существующих взглядов по формированию генетической основы общих типов ВНД являются:
1 - недооценка ведущей роли соматической функции в филогенезе и онтогенезе нервной ткани ГМ организма животного;
2 - отсутствие учета роли соотношения количественных показателей импульсов от разных типов двигательных единиц скелетной мышечной ткани в их афферентной и эфферентной импульсной связи с НТ ГМ, с развитием и свойствами этой ткани.
Цель данного изобретения - определение конкретных генетических факторов, влияющих на развитие свойств нервной ткани головного мозга и формирование типов ВНД.
Итак, поставленную цель достигают, учитывая явное различие в филогенезе и онтогенезе растительных и животных форм органического мира, которое состоит в том, что основная функция у растений - вегетативная, а функции организма животных делят на две группы: соматические и вегетативные.
Движение со множеством степеней свободы, лежащее в основе соматической функции, обусловило резкое усложнение строения животных форм, для координации функционирования органов которых и для взаимодействия с окружающей средой понадобилась физиологическая регуляция (ФР). У животных и человека рассматривают два механизма ФР взаимодействующих между собой органов [6]:
1 - гуморальный,
2 - нервный.
Гуморальная ФР для передачи информации использует жидкие среды организма. Сигналы передаются посредством химических веществ, гормонов, электролитов, биологически активных веществ и т.д. [6], которые относят к работе вегетативной функции организма.
Особенностями гуморальной и, в частности, гормональной ФР являются:
1 - отсутствие точного адресата - с током биологических жидкостей вещества могут доставляться к любым клеткам организма;
2 - скорость доставки информации небольшая - определяется скоростью тока биологических жидкостей (0,5-5 м·с-1);
3 - продолжительное время действия.
Нервная ФР для переработки и передачи информации опосредуется через ЦНС и периферическую нервную систему. Сигналы передаются с помощью нервных импульсов.
Деятельность ЦНС, включающей в себя головной и спинной мозг, осуществляющей общий контроль и управление соматической функцией, анатомически осуществляется за счет НТ.
Особенностями нервной ФР являются:
1 - наличие точного адресата - сигналы доставляются к строго определенным органам и тканям;
2 - большая скорость доставки информации - скорость передачи нервного импульса - до 120 м·с-1;
3 - кратковременность действия по сравнению с гуморальной ФР.
Сравнение всех этих особенностей гуморальной и нервной ФР уже говорит о более мобильной роли последней во взаимодействии органов внутри организма и, тем более, о возможностях ее адекватной реакции на внешние раздражители - с одной стороны, а также о ее преимущественном влиянии на развитие НТ ГМ и ее свойств, и, следовательно, на формирование типов ВНД - с другой.
Отсюда поэтому следует, что классификация темпераментов по Гиппократу, основанная на роли жидких веществ в организме, - несостоятельна, так как пункты 1, 2 и 3 особенностей гуморальной ФР не соответствуют ни безусловной, ни условной рефлекторной деятельности холерического и сангвинического (по крайней мере) темпераментов, а также соответствующим им II-III типам ВНД по Павлову.
Известно, что вегетативная функция непосредственно не управляется сознанием, и эфферентно-афферентная нервноимпульсная связь вегетативных органов с ГМ идет непроизвольно. Соответственно этому и вегетативные компоненты двигательных реакций внутри этих органов, осуществляемые гладкими мышцами, происходят относительно медленно и ритмично. Этим их значительно отличают от параметров движений поперечнополосатых скелетных мышц органов соматической функции, способных сокращаться произвольно, с большой амплитудой и силой, с высокой скоростью и частотой, что в большей мере во многих случаях соответствует запросу сознания и адекватной реакции на воздействие окружающей среды.
На основании вышеизложенного устанавливают превосходящее, доминантное влияние, во-первых, соматической функции, ее нервноимпульской связи с НТ ГМ по сравнению с подобной связью вегетативной функции, а во-вторых, выделяют роль скелетных поперечнополосатых мышц в развитии свойств НТ ГМ и, следовательно, в формировании типов ВНД.
Этот вывод подтверждает и тот факт, что согласно анатомии человека относительная мышечная масса новорожденного составляет около 23% [7, с.243], а уже масса мышц мужчин составляет около 40 - 45%, а у женщин - 30% от массы тела [7, 8, 9], что в разы превышает массу вышеупоминавшихся жидкостей; а среди этих мышц масса поперечнополосатых на порядок больше массы гладких мышц.
Такой скачок возрастания относительной величины мышечной массы в организме говорит об ее доминантности и в развитии, поскольку относительный рост других органов становится меньше (сумма всех относительных величин равна 100%). Эта доминантность безусловно поддержана соответствующим возрастанием относительной роли афферентно-эфферентной нервноимпульсной связи между мышечной тканью и НТ ГМ.
Таким образом, в отличие от взглядов Павлова, устанавливают конкретную функцию - соматическую, превалирующую во влиянии на развитие НТ ГМ, и устанавливают конкретную структуру - скелетные поперечнополосатые мышцы, сокращающиеся произвольно и обуславливающие доминантную нервноимпульсную связь, сильное и мобильное воздействие на НТ ГМ, на ее развитие.
При этом выделяют тот факт, что не только пара конкурирующих процессов - возбуждение и торможение - влияет на формирование типов ВНД и других свойств НТ ГМ, но и значительные отличия в параметрах импульсов от двух видов двигательных единиц (д.е.) поперечнополосатых мышц как на афферентном пути к НТ ГМ, так и отличия между импульсами к ним от НТ ГМ на эфферентном пути.
«По скорости сокращения мышечных волокон двигательные единицы делят на медленные (время одиночного сокращения около 120 мсек) и быстрые (около 60 мсек). Они различаются и по типу обмена веществ. В медленных, содержащих большое количество миоглобина и запас кислорода, происходят в основном окислительные процессы; в быстрых - анаэробные (безкислородные) процессы. От количественного соотношения в мышце медленных и быстрых двигательных единиц зависят ее скоростные свойства» [4], а также сила и предрасположенность к работе на выносливость в том или ином режиме деятельности. Медленные мышечные волокна содержат много митохондрий, которые в основном отсутствуют или их значительно меньше у быстрых мышечных волокон, что напрямую связывают с видами биоэнергетики д.е. мышц.
Различия указанных биоэнергетических процессов, обеспечивающих ресинтез аденозинтрифосфорной кислоты (АТФ), являющейся непосредственным источником энергии сокращения всех типов мышечных волокон, вызывают разный состав жидких веществ в организме для разложения на белки, жиры и углеводы продуктов питания, (что и было подмечено Гиппократом), и что опосредованно подтверждает рассматриваемую ведущую роль в формировании типов ВНД мышечной системы соматической функции, так как вегетативная система количественно в основном обеспечивает ее функционирование. А в зависимости от этого зависят и возбудимость клеток НТ ГМ, лабильность процессов возбуждения и торможения в них, состав крови и скорость ресинтеза АТФ в НТ ГМ.
Ниже в абстрактном виде (математической форме) записи показывают отличия трактовки процессов формирования типов ВНД по Павлову от трактовки процессов в данном предлагаемом изобретении.
Первую пару свойств по Павлову, указанную выше, записывают неравенством:

где Jв и Jт - соответственно суммарный афферентный импульс процесса возбуждения НТ и суммарный импульс процесса торможения, что при Jт>0 дает:

Вторую пару свойств записывают в виде нескольких выражений:

при тех же обозначениях и условии.
Третью пару свойств записывают в виде производных по времени:

Сравнение выражений (1а) и (2) говорит о том, что второе содержит и первое, дублирует его.
Ниже не только не отрицают само наличие свойств, обнаруженных Павловым в постнатальном периоде онтогенеза, но и, учитывая их, дают им и обоснования происхождения уже и в пренатальном периоде.
Указанные выше свойства типов ВНД И.П.Павловым исследованы на динамике условных рефлексов (У.Р.), «сшибки» У.Р. возбуждения и У.Р. торможения, перенапряжения раздражительного процесса [1], а в данном изобретении такие же свойства типов ВНД рассматривают на базе конкуренции в первую очередь безусловных рефлексов организма - различных по величине афферентных импульсов от д.е. мышц.
Так, пару свойств - уравновешенность и неуравновешенность импульсов, конкурирующих даже до рождения плода, воздействующих на НТ ГМ и формирующих типы ВНД, трактуют как соотношение афферентных импульсов от быстрых Jвб и от медленных Jвм двигательных единиц скелетных мышц:

где индекс «в» означает, что он относится к импульсам возбуждения НТ ГМ соответственно от быстрых - индекс «б» или от медленных - индекс «м» двигательных единиц скелетных мышц.
Пару свойств - подвижность и инертность - записывают с учетом этой же трактовки индексов в виде:

Эти записи (4) и (5) с учетом процессов торможения импульсов каждого вида д.е. дают уже так:

и 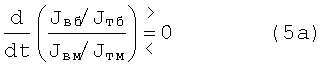
где индекс «т» означает, что он относится к импульсам торможения соответственно быстрых - индекс «б» или медленных - индекс «м» двигательных единиц.
С помощью записи (4а) находят фактор, отвечающий за генетическую базу формирования типов ВНД. Для этого детализируют отдельные члены выражения (4а) таким образом:
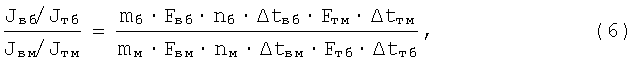
при этом 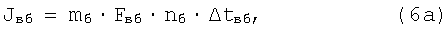
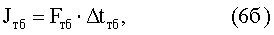
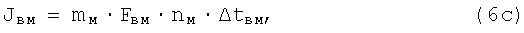
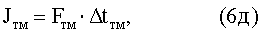
где mб, mм - соответственно число быстрых и число медленных д.е. конкретной мышцы (КМ);
Fвб, Fвм - сила одиночного афферентного импульса возбуждения соответственно от быстрых и медленных д.е. КМ;
nб, nм - число одиночных сокращений соответственно быстрых и медленных д.е., образующих одиночное сокращение КМ;
Δtвб, Δtвм - длительность импульса одиночного сокращения соответственно быстрых и медленных д.е. КМ;
Fтб, Fтм - сила импульса торможения афферентного импульса соответственно быстрых и медленных д.е. при одиночном сокращении КМ;
Δtтб, Δtтм - длительность импульса торможения афферентного импульса соответственно быстрых и медленных д.е. при одиночном сокращении КМ.
Правая часть выражения (6) состоит из экстенсивных и интенсивных параметров. Отношение экстенсивных параметров mб и mм между собой, как установлено в физиологии, для каждой скелетной мышцы - есть величина постоянная, не изменяемая со временем в течение жизни людей, что следует из наблюдений за близнецами и неблизнецами [10, 11, 12], и является генетическим фактором Фi - параметром каждой i-ой мышцы и у каждого человека индивидуальным, постоянным. Поэтому записывают для каждой скелетной мышцы выражение (6) в виде:
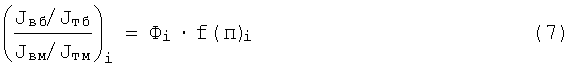
где f(п)i - функция всех переменных величин i-ой скелетной мышцы, входящих в правую часть выражения (6), за исключением mб и mм, отношение которых дает фактор i-ой мышцы Фi:

Скелетных мышц у человека порядка 500-600 [8, 9], поэтому значения Фi располагают в фиксированных точках на абсциссе - фиг.1 (см. чертеж). Фиг.1 является условным спектром значений
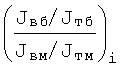
отложенных по ординате над осью абсцисс, или спектром значений
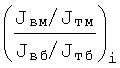
отложенных по ординате под осью абсцисс.
Цифрами вдоль оси абсцисс над этими спектрами показаны значения lg(Фi).
Этими спектрами условно показывают композицию соотношений афферентных импульсов от быстрых и медленных д.е. различных скелетных мышц всего организма, модулированных соответствующими им импульсами безусловного, запредельного, охранительного торможения, предохраняющими НТ ГМ от разрушительного воздействия очень сильных и/или очень продолжительных афферентных импульсов тех или иных д.е.
Как считают в физиологии [4, 7], так условно показывают и на фиг.2 (см. чертеж), только лишь небольшая часть скелетных мышц состоит почти целиком из быстрых или из медленных д.е. Цифрами вдоль оси абсцисс на фиг.2 показаны значения lg(Фi), а по оси ординат отложены относительные величины
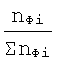
где nФi - число мышц, имеющих значение Фi;
ΣnФi - суммарное число мышц со всеми значениями Фi.
Эта (фиг.2) кривая распределения относительного количества мышц по соотношению быстрых и медленных д.е. в мышцах организма индивидуальна для каждого организма и является асимметричной кривой с одним или несколькими максимумами.
По сдвигу расположения среднестатистической величины Фср - всех таких значений Фi в ту или другую сторону от значения lg Фi,равного 0, на фиг.1 и фиг.2, с учетом парциальных количеств медленных мышечных волокон (ММВ) и быстрых (БМВ) - отношениям чисел ММВ или БМВ в конкретной мышце к общему числу всех аналогичных волокон скелетных мышц в организме - судят о типе ВНД. Это расположение Фср априори лежит внутри всего диапазона значений Фi - фиг.1 и ему соответствует какая-то конкретная мышца или группа мышц с конкретной величиной Фi.
Поскольку известно [5, с.30], что в скелетных мышцах соотношение красных (т.е. ММВ) и белых (т.е. БМВ) волокон индивидуально, «определяется генетически и фактически с возрастом не меняется», то и в сумме всех скелетных мышц в организме оно постоянно, генетически индивидуально в онтогенезе. Учитывая, что число тех или иных д.е. в мышце определяется отношением суммарного числа соответствующих волокон в ней к числу таких же волокон в аналогичной д.е., в итоге находят суммарный генетический фактор Ф всего организма - отношение всего числа БМВ ко всему числу ММВ в организме, который является генетической основой общего типа ВНД:

где NБМВ - общее число БМВ в организме,
NММВ - общее число ММВ в организме.
Так, по положению фактора Ф относительно единицы: Ф много больше, равно или около 1, или много меньше 1, судят о генетической основе типа ВНД конкретного организма. Значению Ф, равному 1, соответствует равное, 50% содержание в организме чисел БМВ и чисел ММВ. Большое увеличение содержания БМВ, т.е. Ф много больше 1, ведет к превосходству (по терминологии Гиппократа) холерических признаков, а большое увеличение содержания ММВ, т.е. Ф много меньше 1, ведет к превосходству флегматических. Сангвинический темперамент соответствует значениям Ф вблизи 1. При этом в данном изобретении в одном случае говорят о возрастании одаренности организма БМВ, а в другом случае - о возрастании одаренности организма ММВ.
Учитывая многочисленность возможных величин Фi у индивидов, находят большое генетическое разнообразие факторов Ф у сообществ видов животных и у сообщества людей, чем уже объясняют множество переходных типов ВНД, получаемых ими по наследству.
Однако БМВ по морфологическим, биохимическим и физиологическим характеристикам существенно неоднородны. Различают [5, 12, 13] БМВ типа IIB - белые быстрые мышечные волокна с низким содержанием миоглобина и митохондрий, и гликолитическим ресинтезом АТФ от БМВ типа IIA - со значительно большим, чем в БМВ типа IIВ, содержанием миоглобина и митохондрий, и окислительно-гликолитическими механизмами ресинтеза АТФ.
Считают, что БМВ типа IIА занимают промежуточное положение между ММВ, которые называют [5] также волокнами типа I, и БМВ типа IIB как по морфологическим и биоэнергетическим характеристикам, так и по параметрам идущих от них афферентных импульсов. Из-за окислительно-гликолитической биоэнергетики и морфологических особенностей двигательные единицы из БМВ типа IIA выполняют быстрые, энергичные сокращения с пониженной утомляемостью по сравнению с д.е. из БМВ типа IIB и их параметры Δtвб, входящие в соответствующие выражения, аналогичные (6) и (6а), находят у БМВ типа IIA значительно большими, чем у БМВ типа IIB, при одинаковых произвольных и непроизвольных режимах физической активности, а поэтому и значения Δtтб для их торможения тоже находят большими.
В связи с постоянством генетических соотношений сумм чисел тех и других БМВ с суммой чисел ММВ в индивидуальном организме и различиям по параметрам Δtвб и Δtтб афферентных импульсов от этих БМВ находят различное их воздействие на НТ ГМ и, соответственно, на формирование ВНД животных и человека даже еще в утробном состоянии, в пренатальный период развития их организмов.
Ввиду вышеизложенного фактор Ф рассматривают состоящим из двух слагаемых:

где


где NБВМIIA - суммарное число БМВ типа IIA в организме,
NБМВIIB - суммарное число БМВ типа IIB в организме,
NММВ - суммарное число ММВ в организме.
Считают также [4, 5], что «в естественных условиях большинство сокращений скелетных мышц возникают в ответ на импульсную стимуляцию частотного характера и являются тетаническими». Соответственно этому в данном случае устанавливают, что различия в интенсивных количественных параметрах импульсов от д.е. из БМВ типа IIA, БМВ типа IIB и ММВ резко возрастают при переходе к рассмотрению тетанических сокращений.
Согласно [4, 5] «тетаническое сокращение характерно как для одиночного мышечного волокна, так и в целом для всей мышцы …. Важно подчеркнуть, что все виды тетанических сокращений по своей амплитуде превышают амплитуду одиночного сокращения». Доказывают также (Е.К.Жуков), что при тетаническом сокращении более сильно и эффективно работает сократительный аппарат мышечных волокон (меньше затраты энергии на ее потери), а «кроме того, способность мышцы к генерации тетанического сокращения существенно расширяет возможности ЦНС для управления сократительной активностью скелетных мышц» [5].
Поскольку возникновение тетанического сокращения мышц зависит от амплитуды и величин периодов сокращения и расслабления составляющих их д.е. и входящих в них мышечных волокон, устанавливают, что тетанус мышцы скорее происходит за счет более раннего возникновения тетануса от д.е. из БМВ типа IIA, чем при равных количествах от д.е. из других мышечных волокон при достаточно интенсивной физической нагрузке рассматриваемой мышцы.
При этом, соответственно, различают и количественные параметры импульсов торможения афферентных импульсов от всех этих д.е., что с учетом запаздывания возникновения этих тормозных импульсов и наличия, следовательно, проскока доли неослабленных аффертных импульсов к НТ ГМ от этих д.е. вызывает более значительные различия в воздействии на НТ ГМ афферентных импульсов от д.е. всех типов при тетанических сокращениях, чем при нететанических, одиночных сокращениях скелетных мышц.
С учетом тетанических сокращений мышц при вышерассмотренной трактовке фактора Ф находят, что классификации четырех общих типов ВНД по Павлову соответствуют лишь части общей композиции из факторов Ф, ФIIA и ФIIB.
Из выражений (9б) и (9в) видят, что

В многочисленности вариантов совокупностей экстенсивных и интенсивных параметров согласно выражениям (4а, 5а) и (6) видят многообразие способов развития НТ ГМ и возможные способы воздействия на общие типы ВНД с помощью опосредованного влияния режимов физической активности на те или иные группы мышц и организм в целом.
При этом устанавливают, что влияние 1СС на типы ВНД, на фенотипы состоит и в том, что эти источники возбуждения, увеличивая общее число источников возбуждения, и, следовательно, импульсов возбуждения НТ в ГМ, обуславливающих ее развитие, снижают относительную роль влияния базового, генетического фактора Ф на НТ в ГМ, не уменьшая его абсолютной постоянной роли. Находят также, что аналогично, но в значительно большей мере оказывает воздействие и 2СС на НТ в ГМ.
Роль показанного фактора Ф организма уже фактически сравнительно давно известна и ее уже используют в спорте для ранней специализации детей в разных видах спорта, особенно циклических, а также в спортивной физиологии для объяснения или планирования роста результатов [10-14]. А сам фактор практически возможно определить по экспресс-анализу с помощью биопсии.
Так, например, с помощью биопсии возможно определение абсолютных и относительных количеств всех типов волокон, как в отдельных мышцах различных органов, так и в мышцах всего организма. Однако известно [5, 12], что данные биопсии разных мышц различны. Но результаты биопсии некоторых отдельных мышц, например широкой мышцы бедра, в спортивной медицине весьма удачно экстраполируют на весь организм с целью определения относительного состава волокон во всем организме. Так, в статье «Бег» [11, с.55] читаем: «при исследовании мышц выдающихся бегунов методом биопсии (при помощи тонкой иглы кусочек мышцы вынимается и анализируется под микроскопом) оказалось, что у спринтеров 80-85% быстрых и 20-15% медленных волокон, а у марафонцев наоборот - 85-90% медленных и только 15-10% быстрых. Если спортсмен обладает преимущественно медленными волокнами и мечтает стать классным спринтером, то все его усилия будут обречены на провал.
В некоторых странах берут биопсию у 12-летних подростков и сразу определяют «специальность» будущего бегуна. Итак, установлено соответствие типа мышц классу беговой дистанции». Кроме биопсии для той же цели применяют и динамометрию мышц [12, с.88]. Последний метод - безболезненный и, как считает В.Н.Селуянов, более точен.
В подтверждение того, что не обязательно анализировать все мышцы организма, чтобы судить с достаточной точностью для практических целей об их относительном составе в нем, ярко говорит тест, применяемый в спортивном плавании. В работе [14, с.23-24] говорится: «В настоящее время точно установлено, что пловцы, склонные к достижению высоких результатов в спринте и имеющие высокий процент быстрых мышечных волокон, показывают высокий результат в прыжке вверх с места. Учитывая результаты такого прыжка, можно с достаточной точностью выявить пловцов-спринтеров, пловцов на средние и длинные дистанции (рис.1)». В подписи под этим рис.1 читаем: «зависимость между высотой прыжка вверх с места и предрасположенностью пловца к достижениям на различных дистанциях (J. Counsilman, 1980, переработано)».
Этот тест весьма примечателен еще тем, что говорит о существовании не только теоретически возможной корреляции относительного состава волокон отдельной группы мышц ног или отдельной мышцы с относительным составом волокон мышц всего организма, но и о практической подобной корреляции, так как в нециклическом движении - прыжке вверх с места - работают только толчковые мышцы ног, а в плавании работает уже большинство мышц всего организма: и ног, и рук, и туловища. Это говорит и о том, что вышерассмотренные биопсия и динамометрия отдельных мышц могут служить для целей экспресс-анализа относительного состава волокон мышц всего организма, что и используют успешно как показатель предрасположенности к той или иной специализации, особенно в циклических видах спорта.
То есть, этот тест подтверждает показанную выше возможность теоретической корреляции среднестатистической величины Фср скелетных мышц всего организма с Фi одной мышцы или группы мышц.
С другой стороны, о предрасположенности организма к определенной специализации судят и по темпераменту, о чем говорят специалисты разных видов спорта. Например, весьма красноречива цитата из статьи [11, с.59-60]: «Небольшое отступление относительно психической организации бегунов на различные дистанции. Сегодня уже точно установлено, что заметные успехи возможны лишь при соответствии психики спортсмена той дистанции, на которой он специализируется. Возможно, это проливает свет на некоторые «пусковые» механизмы того или иного двигателя, объясняет казалось бы невозможные результаты. Спринтеры всегда взрывные, импульсивные, возбудимые, непоседливые, зачастую агрессивные. Любят спорить, эмоционально неустойчивы. Иногда эмоциональный взрыв приводит к мощному всплеску энергии, и тогда за считанные секунды приходит победа или рекорд. Стайеры степенные и неторопливые, рассудительные и невозмутимые. Всегда уравновешенные. Без этих качеств трудно было бы ежедневно осиливать тяжелую и монотонную работу. Разговаривая с бегуном или наблюдая за ним, можно определить склонность его таланта. Итальянец Меннеа или кубинец Леонард в общении быстры, их мимика подвижна, жестикуляция активна. Их мнения категоричны, недовольны - взрываются, рады - готовы плясать. Наши Моисеев или Антипов обдумывают каждое слово, доброжелательны, в их присутствии чувствуешь какое-то внутреннее спокойствие, любое дело совершают спокойно, без суеты. Пропажа шиповок накануне старта - серьезная неприятность для бегуна, фактически это его единственный инвентарь. Спринтера подобная история начисто выбьет из колеи, стайер перенесет ее с добродушной усмешкой. В этом плане исключений совсем мало».
Беспристрастные, обширные, глубокие, полезные, многочисленные и статистически достоверные наблюдения делают и два крупных специалиста в другом виде спорта - В.Н.Платонов и С.М.Вайцеховский [14]:
«Личностные и психические качества пловца служат не только критериями оценки перспективности, но и дополнительными критериями предрасположенности к специализации на коротких, средних и длинных дистанциях. Необходимо учитывать, что у спортсменов, специализирующихся на коротких дистанциях, обычно наблюдается слабый (реактивный) тип нервной системы. При этом основными признаками являются категоричность в суждении, высокая возбудимость, быстрота смены настроения, легкость приспособления к новым условиям жизни и тренировки, быстрота перехода от сна к бодрствованию, быстрота усвоения навыков, низкая концентрация внимания и необходимость применения специальных приемов для его активации.
Специализирующихся на длинных дистанциях отличает спокойное, устойчивое настроение, трудолюбие и дисциплинированность, плохая приспособляемость к новым условиям. У них отмечаются пониженная чувствительность к слабым раздражителям, высокая концентрация внимания, слабая переключаемость, малая эмоциональность, объективная оценка своих возможностей, настойчивость и упорство».
Аналогичные предпосылки к специализации видят и в других циклических видах спорта: конькобежном, лыжном, гребном, велосипедном.
Но теперь обратим внимание и на тот факт, что, с одной стороны, на базе соотношения типов волокон в организме основывают энергетические предпосылки к специализации индивида в спорте, что, как уже говорилось выше, часто и давно уже используют на практике, учитывая соотношение типов волокон в организме, а с другой стороны, как было только что показано в цитатах, к этой специализации ведут темперамент и черты характера, связанные с типом ВНД индивида. То есть, налицо имеем компоненты факторного анализа [15]: «Множество попарных корреляций между фактами является материалом для факторного анализа. За статистически значимыми коррелирующими друг с другом фактами лежит один и тот же фактор».
О необходимости применения факторного анализа в подобных случаях говорят и в работе [16, с.152]: «выделены свойства общие и парциальные, которые функционируют в разных структурах головного мозга, по-разному сказываются в поведении и предполагают различающиеся измерения. Последние по природе носят статистический характер и требуют для интерпретации использования корреляционного, дисперсионного или факторного анализа результатов».
То есть, в данном случае в основе двух попарных корреляций в предрасположенности к специализации лежит один и тот же фактор - коррелируют между собой относительные составы мышечных волокон соматической функции в организмах - генетически заданные факторы индивидов и их общие типы ВНД, темпераменты индивидов, тоже считающиеся генетически заданными (И.П.Павлов и др.) и наиболее ярко проявляющиеся в экстремальных условиях спорта.
Математически это проявление видят и в том, что при нахождении производной по выражению (5а) (или аналогичной формулы с учетом условных рефлексов для постнатального периода) постоянная величина - генетический фактор Ф - выходит за знак дифференциала.
Таким образом, именно метод факторного анализа позволяет дать объективное, никем конкретно не высказываемое и, следовательно, беспристрастное, независимое, но аргументированное многочисленными фактами заключение, фактически уже существующее в природе и подтверждающее заключение, которое делают в изобретении и не через «предрасположенность», а напрямую, математическими символами показывают наглядный вывод связи между морфофункциональными и нейрофизиологическими параметрами у людей и у животных.
Вышеприведенные цифровые данные из ссылочных работ по содержанию мышечных волокон у разных индивидов показывают широкий диапазон рассматриваемых относительных величин у людей.
Так у прирожденных спринтеров максимальное значение содержания БМВ во всем организме достигает 90% от общего числа волокон, а остальные 10% составляют ММВ. Следовательно, соотношение этих чисел равно 9 (а при 91% и 9% соответственно, такое соотношение было бы уже больше 10).
У прирожденных марафонцев цифры противоположные: БМВ - около 10%, ММВ - 90% и их соотношение равно 1/9 (а при 9% БМВ и 91% ММВ их соотношение было бы уже меньше 0,1).
У средневиков находят относительное содержание БМВ и ММВ от общего количества этих волокон приблизительно равным - около 50%, то есть их соотношение составляет около 1.
Таким образом, эти цифры показывают, что диапазон величин фактора Ф у разных людей изменяется почти от 10 до 0,1, то есть практически на два порядка.
Находят, что с точки зрения различий в генетической основе общих типов ВНД величины Ф более трех уже относят к величинам много больше единицы, а величины Ф менее одной трети - к величинам много меньшим единицы.
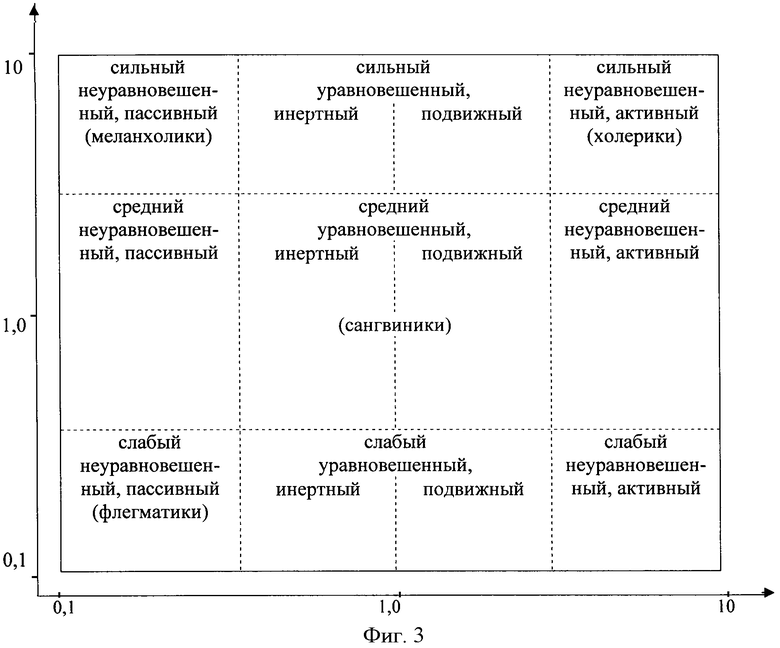
В связи с тем, что при каждом Ф приблизительно в подобном диапазоне величин различают и соотношение ФIIA/ФIIB у разных индивидов, то общую композицию для различных Ф и их разновидностей представляют в виде совокупности точек на плоскости - фиг.3 (см. чертеж), координаты которых и по вертикали - Ф и по горизонтали - ФIIA/ФIIB лежат в пределах величин от больше 0,1 до меньше 10. В эту композицию входят 12 существенно различающихся групп типов ВНД:
если Ф больше 3 и отношение ФIIA к ФIIB больше 3, определяют сильный неуравновешенный активный тип ВНД;
если Ф больше 3, а отношение ФIIA к ФIIB равно от 1 до 3, определяют сильный уравновешенный подвижный тип ВНД;
если Ф больше 3, а отношение ФIIA к ФIIB равно от 0,3 до 1, определяют сильный уравновешенный инертный тип ВНД;
если Ф больше 3, а отношение ФIIA к ФIIB меньше 0,3, определяют сильный неуравновешенный пассивный тип ВНД;
если Ф от 0,3 до 3, а отношение ФIIA к ФIIB больше 3, определяют средний неуравновешенный активный тип ВНД;
если Ф от 0,3 до 3, а отношение ФIIA к ФIIB от 1 до 3, определяют средний уравновешенный подвижный тип ВНД;
если Ф от 0,3 до 3, а отношение ФIIA к ФIIB от 1 до 0,3, определяют средний уравновешенный инертный тип ВНД;
если Ф от 0,3 до 3, а отношение ФIIA к ФIIB меньше 0,3, определяют средний неуравновешенный пассивный тип ВНД;
если Ф меньше 0,3, а отношение ФIIA к ФIIB больше 3, определяют слабый неуравновешенный активный тип ВНД;
если Ф меньше 0,3, а отношение ФIIA к ФIIB от 1 до 3, определяют слабый уравновешенный подвижный тип ВНД;
если Ф меньше 0,3, а отношение ФIIA к ФIIB от 0,3 до 1, определяют слабый уравновешенный инертный тип ВНД;
если Ф и отношение ФIIA к ФIIB меньше 0,3, определяют слабый неуравновешенный пассивный тип ВНД.
Знанием общего соотношения чисел БМВ и ММВ в организме заранее устанавливают тип ВНД и сопутствующий ему темперамент человека и используют их для активного раннего воздействия на развитие и сохранение способностей человека, определяемые физиологическим строением его органов 1СС, путем подбора соответствующего режима физической активности, влияющей на развитие мышц данного органа и на НТ ГМ в целом. А также, устанавливая предлагаемым способом тип ВНД, целенаправленно физическими упражнениями изменяют соответствующую физическую активность, понижая или повышая ее, чем одновременно, совместно с 2СС, влияют на корректировку темперамента в желаемом направлении, на концентрацию внимания, усидчивости, обучаемости и т.д., то есть тем самым и обратно - на развитие и восприятие форм 2СС и когнитивное обучение.
Устанавливают также, что подобно влиянию афферентных импульсов на развитие и свойства НТ ГМ непосредственно в моторной зоне коры больших полушарий, влияют и индуцируемые ее же участками эфферентные импульсы с различными параметрами к соответствующим д.е., поскольку при произвольном режиме индуцирования этих импульсов она (НТ) вырабатывает и такие, на образование которых нервными клетками тратится энергия, значительно превышающая адаптационный уровень ее на конкретные подобные, но менее интенсивные действия предыдущего периода. Взаимозависимая эфферентно-афферентная импульсная связь между НТ двигательной зоны ГМ и мышечной тканью органов соматической функции влияет, как показал А.А.Ухтомский, и на другие зоны ГМ путем развития множественных связей с ними и образованием структур мозгового звена функциональных органов, развивающихся прижизненно при освоении новых внешних и внутренних действий и реакций на 1СС и 2СС.
Таким образом, согласно вышеизложенному, генетическую основу общего типа ВНД организма человека и животного определяют по композиции морфо-функционального параметра Ф - генетического соотношения общего числа БМВ к общему числу ММВ всех его скелетных поперечнополосатых мышц соматической функции (выражение (9)), а различные соотношения всех чисел БМВ типа IIA и всех чисел БМВ типа IIB между собой (выражение (10)) при конкретном факторе Ф составляют разновидности этого фактора у различных людей и животных.
С помощью подбора режима физической активности для тех или других мышечных волокон в органах соматической функции усиливают или ослабляют влияние на развитие активности нервной ткани головного мозга, чем управляют развитием предрасположенностей к способностям, самими способностями и их сохранением.
В установленной генетической многовариантности общих типов ВНД и многообразии влияния интенсивных параметров на свойства НТ ГМ со стороны соматической функции видят фундаментальность филогенеза, пошедшего по этому пути развития органического мира разумного, и почву для восприятия и адаптации к информации 1СС, а также почву для развития и восприятия многообразия форм 2СС и когнитивности обучения в онтогенезе.
«Тип поведения и темперамент связаны не однозначно. Темперамент задает динамику психики и поведения, но не определяет их содержание и направление. Да и сама динамика может быть при необходимости скрыта или усилена. От одинакового темперамента можно прийти к различным характерам, и наоборот. Темперамент врожден, поскольку обусловлен свойствами нервной системы человека. Значит индивид не может изменить своего темперамента, но вполне в состоянии как-то подправлять его внешние проявления. Но это не является изменением или воспитанием темперамента. Это формирование, развитие, социализация, воспитание характера и личности в целом» - из работы [16, с.154].
Находят, что и независимость, и объективность фактов, на основании которых осуществлено подтверждение - вывод факторного анализа, состоят в следующем.
а. Возможные мнения, заключения, основанные на сенсорном восприятии кого-либо кем-либо, не всегда объективны, поскольку субъективны по своей сути.
Это - мнения фенотипа, имеющего некий генотип с безусловными рефлексами, который оброс «камуфляжем» из условных рефлексов от воздействия 1СС и 2СС, сформировавшимся на базе этого генотипа и его безусловных рефлексов в онтогенезе.
К тому же, это - мнения о фенотипе, тоже имеющем свой генотип и свой «камуфляж» из своих условных рефлексов.
Освободиться от обоих «камуфляжей» одновременно наблюдателю и наблюдаемому - весьма непростая задача, поскольку неизвестна генетическая основа, лежащая за каждым из этих «камуфляжей».
В решении этого вопроса помогают длительность наблюдения за индивидом в различных жизненных ситуациях, особенно в экстремальных, стрессовых условиях, поскольку, как считают в психологии, «камуфляж» индивида в экстремальных, стрессовых условиях в значительное мере «спадает», а черты темперамента больше коррелируют с его типом ВНД [5, с.502]: «Удельное значение врожденного и приобретенного в фенотипе может меняться в зависимости от условий. В необычных, экстремальных условиях на первый план в поведении выступают преимущественно врожденные механизмы ВНД».
Фактически об этом же говорят и в работе [16, с.154]: «Б.М.Теплов отмечал, что в обыденных обстоятельствах, в привычных для личности условиях деятельности нет обязательных проявлений темперамента. Нормальный человек ведет себя не так, как велит нервная система, а как он считает принятым в данных обстоятельствах. Темперамент проявляется лишь в экстремальных условиях, да и не только он…».
В основе появления экстремальных, стрессовых условий лежит одновременное сильное воздействие на организм по крайней мере двух взаимопротивоположных рефлексов, особенно безусловных, или перенапряжение раздражительного процесса. Такие стрессовые условия часто создавали в экспериментах с собаками в лаборатории И.П.Павлова, когда проводили «сшибку» раздражительного и тормозного процессов, то есть, когда раздражительный условный рефлекс переделывался в тормозной условный рефлекс, или наоборот. В этих вынужденных, принудительных условиях порою более ярко проявлялись динамические параметры типов ВНД животных. Однако очень часто эти «сшибки» приводили и к стойкой патологии нервной деятельности животного, от чего затем было трудно его вылечить полностью [1].
Естественно, что такие эксперименты с людьми небезопасны. Однако есть вид добровольной деятельности у людей, в которой регулярно, длительное время человек под влиянием разнообразных сильных мотиваций подвергает себя воздействию экстремальных условий. Это - спорт, особенно - «большой» спорт, при развивающих нагрузках на организм, а также на соревнованиях.
При этом сталкиваются два процесса: с одной стороны - раздражительные условный и безусловный рефлексы, основанные на внешних и внутренних мотивациях (экономические выгоды, самоутверждение, лидерство и т.д.), ведущие к повышению физических нагрузок на организм, а с другой стороны - оборонительный, тормозной безусловный рефлекс самосохранения гомеостаза организма, защищающий имеющуюся на каждый конкретный момент адаптированную базу течения энергетических процессов в организме.
Именно противоборство этих процессов в течение длительного времени подготовки к соревнованиям позволяет тренерам и медикам наблюдать более чистое, более свободное от «камуфляжа» проявление темперамента, динамики типа ВНД спортсмена и связанных с ними черт его характера, что было выше показано, и что явилось второй попарной корреляцией для факторного анализа.
б. По поводу экстраполяции Фi - данных биопсии по конкретной мышце на фактор Ф всего организма.
В спортивной практике широко распространена биопсия широкой мышцы бедра, что оправдывают успешной спортивной ориентацией. Но возможно, что есть еще более «точные» мышцы для определения фактора Ф или группа мышц.
Литература
1. Павлов И.П., Двадцатилетний опыт объективного изучения высшей нервной деятельности (поведения) животных, Медгиз, 1951, М., с.: 366-369, 394-397, 432-444, 448-462, 463-466.
2. Немов Р.С., Психология, М., 2008, с.384-390.
3. Данилова Н.Н., Психофизиология, М., 2000, с.257, 279-280.
4. Под ред. Васильевой В.В., Физиология человека, М., ФиС, 1984, с.53-59, 83, 91-94.
5. Циркин В.И., Трухина С.И., Физиологические основы психической деятельности и поведения человека. Медицинская книга, М., 2001, с.29-33, 350, 500-516.
6. Леванова Н.Д., Фирсова С.С., Нормальная физиология, Ростов на Дону, Феникс, 2007.
7. Дубровский В.И., Спортивная физиология, М., 2005, с.243.
8. Самусев Р.П., Липченко В.Я., Атлас анатомии человека, М., 2002.
9. Боянович Ю.В., Балакирев Н.П., Атлас анатомии человека. Ростов на Дону, Феникс, 2005.
10. Торлина Т., Природу побеждает тот, кто ей повинуется, М., Наука и жизнь, №6, 1980, с.25.
11. Залесский М., Леонидов Л., Бег, М., Наука и жизнь, №6, 1980, с.60.
12. Селуянов В.Н., Знание слепо без интуиции, Л.С., №21, с.88-98, М., 2002.
13. Селуянов В.Н., Интуиция слепа без знания, Л.С., №23, с.67-77, М., 2002.
14. Платонов В.Н., Вайцеховский С.М., Тренировка пловцов высокого класса, М., ФиС, 1985, с.23-26.
15. Большая психологическая энциклопедия, Эксмо, М., с.479.
16. Под редакцией Сосновского Б.А., Психология, учебник, М., Юрайт, 2005, с.151-154.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ ГЕНЕТИЧЕСКОЙ ОСНОВЫ ОБЩИХ ТИПОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ | 2008 |
|
RU2386399C9 |
| СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ И РЕАБИЛИТАЦИИ ЛОКАЛЬНОЙ ИМПУЛЬСНОЙ ЭЛЕКТРОСТИМУЛЯЦИЕЙ | 1999 |
|
RU2160127C1 |
| Способ остеопатической коррекции соматических дисфункций организма человека | 2024 |
|
RU2834332C1 |
| СПОСОБ ДИСТАНЦИОННОЙ МУЛЬТИВОЛНОВОЙ ЭЛЕКТРОМАГНИТНОЙ РАДИОНЕЙРОИНЖЕНЕРИИ ГОЛОВНОГО МОЗГА ЧЕЛОВЕКА | 2015 |
|
RU2621547C2 |
| Способ стимулирования ангиогенеза, лечения нейродегенеративных заболеваний и регенерации скелетной мышцы с помощью плазмидных конструкций с генами ангиогенных и нейротрофических факторов | 2020 |
|
RU2805082C2 |
| СПОСОБ ДИАГНОСТИКИ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ БИОМЕМБРАН ПРИ МАГНИТНЫХ АППЛИКАЦИЯХ ПО "ЭФФЕКТУ МОЛЧАНИЯ" В ДУГЕ СОМАТИЧЕСКОГО РЕФЛЕКСА | 1996 |
|
RU2144311C1 |
| СПОСОБ ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКИ ПСИХОСОМАТИЧЕСКИХ И СОМАТИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2672886C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ РАБОЧИХ КАЧЕСТВ СЛУЖЕБНЫХ СОБАК | 2016 |
|
RU2621382C1 |
| СПОСОБ ОЗДОРОВЛЕНИЯ ОРГАНИЗМА | 2003 |
|
RU2228771C1 |
| СПОСОБ ЛЕЧЕНИЯ НЕЙРОСЕНСОРНОЙ ТУГОУХОСТИ ЭЛЕКТРОСТИМУЛЯЦИЕЙ | 2000 |
|
RU2166967C1 |
Изобретение относится к области медицины, а именно к нейропсихофизиологии. Находят сумму чисел медленных мышечных волокон (NММВ), сумму быстрых мышечных волокон (NБМВ), сумму быстрых мышечных волокон типа IIА (NБМВIIA) и сумму быстрых мышечных волокон типа IIВ - (NБМВIIВ). Рассчитывают генетически суммарный фактор организма - Ф по формуле: Ф=NБМВ/NММВ, а также суммарный фактор БМВ типа IIА в организме - ФIIА и суммарный фактор БМВ типа IIВ - ФIIВ по формулам: ФIIA=NБМВIIA/NММВ, ФIIB=NБМВIIB/NММВ. Если Ф больше 3 и отношение ФIIА к ФIIВ больше 3, определяют сильный неуравновешенный активный тип ВНД; если Ф больше 3, а отношение ФIIА к ФIIВ равно от 1 до 3, определяют сильный уравновешенный подвижный тип ВНД; если Ф больше 3, а отношение ФIIА к ФIIВ равно от 0,3 до 1, определяют сильный уравновешенный инертный тип ВНД; если Ф больше 3, а отношение ФIIА к ФIIB меньше 0,3, определяют сильный неуравновешенный пассивный тип ВНД; если Ф от 0,3 до 3, а отношение ФIIА к ФIIВ больше 3, определяют средний неуравновешенный активный тип ВНД; если Ф от 0,3 до 3, а отношение ФIIА к ФIIВ от 1 до 3, определяют средний уравновешенный подвижный тип ВНД; если Ф от 0,3 до 3, а отношение ФIIА к ФIIВ от 1 до 0,3, определяют средний уравновешенный инертный тип ВНД; если Ф от 0,3 до 3, а отношение ФIIА к ФIIВ меньше 0,3, определяют средний неуравновешенный пассивный тип ВНД; если Ф меньше 0,3, а отношение ФIIA к ФIIB больше 3, определяют слабый неуравновешенный активный тип ВНД; если Ф меньше 0,3, а отношение ФIIА к ФIIВ от 1 до 3, определяют слабый уравновешенный подвижный тип ВНД; если Ф меньше 0,3, а отношение ФIIA к ФIIB от 0,3 до 1, определяют слабый уравновешенный инертный тип ВНД; если Ф и отношение ФIIА к ФIIВ меньше 0,3, определяют слабый неуравновешенный пассивный тип ВНД. Способ расширяет арсенал средств определения типов. 3 рис.
Способ определения общих типов высшей нервной деятельности - ВНД людей и животных, отличающийся тем, что находят сумму чисел медленных мышечных волокон - NММВ, сумму быстрых мышечных волокон - NБMB, сумму быстрых мышечных волокон типа IIA-NБMBIIA и сумму быстрых мышечных волокон типа IIB-NБMBIIB, рассчитывают генетически суммарный фактор организма - Ф по формуле:
Ф=NБМВ/NММВ,
а также суммарный фактор БМВ типа IIА в организме - ФIIA и суммарный фактор БМВ типа IIВ-ФIIB по формулам: ФIIA=NБМВIIA/NММВ, ФIIB=NБМВIIB/NММВ и,
если Ф больше 3 и отношение - ФIIА к ФIIВ больше 3, определяют сильный неуравновешенный активный тип ВНД;
если Ф больше 3, а отношение ФIIА к ФIIВ равно от 1 до 3, определяют сильный уравновешенный подвижный тип ВНД;
если Ф больше 3, а отношение ФIIА к ФIIВ равно от 0,3 до 1, определяют сильный уравновешенный инертный тип ВНД;
если Ф больше 3, а отношение ФIIА к ФIIB меньше 0,3, определяют сильный неуравновешенный пассивный тип ВНД;
если Ф от 0,3 до 3, а отношение ФIIА к ФIIВ больше 3, определяют средний неуравновешенный активный тип ВНД;
если Ф от 0,3 до 3, а отношение ФIIА к ФIIВ от 1 до 3, определяют средний уравновешенный подвижный тип ВНД;
если Ф от 0,3 до 3, а отношение ФIIА к ФIIВ от 1 до 0,3, определяют средний уравновешенный инертный тип ВНД;
если Ф от 0,3 до 3, а отношение ФIIА к ФIIB меньше 0,3, определяют средний неуравновешенный пассивный тип ВНД;
если Ф меньше 0,3, а отношение ФIIА к ФIIВ больше 3, определяют слабый неуравновешенный активный тип ВНД;
если Ф меньше 0,3, а отношение ФIIА к ФIIВ от 1 до 3, определяют слабый уравновешенный подвижный тип ВНД;
если Ф меньше 0,3, а отношение ФIIА к ФIIВ от 0,3 до 1, определяют слабый уравновешенный инертный тип ВНД;
если Ф и отношение ФIIА к ФIIВ меньше 0,3, определяют слабый неуравновешенный пассивный тип ВНД.
| ПАВЛОВ И.П | |||
| Двадцатилетний опыт объективного изучения высшей нервной деятельности животных | |||
| - М.: Медгиз, 1951, с.366-369, 394-397, 432-444, 448-462, 463-466 | |||
| СПОСОБ ОПРЕДЕЛЕНИЯ ПСИХОЛОГИЧЕСКОГО ТИПА ПО АНУАШВИЛИ | 2001 |
|
RU2201131C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ТИПОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ У БЫКОВ-ПРОИЗВОДИТЕЛЕЙ | 2005 |
|
RU2292133C1 |
| WO 2008016693 А1, 07.02.2008 | |||
| СУЛУЯНОВ В.Н | |||
| Сердце не машина | |||
| Опубликовано в Интернете по адресу | |||

