Область техники
Изобретение относится к биотехнологии, а именно к литической протеазе AlpB бактерии Lysobacter sp. XL1, а также к кодирующему этот фермент фрагменту ДНК бактерии Lysobacter sp. XL1 и способу получения фермента.
Клонирование фрагмента ДНК бактерии Lysobacter sp. XL1, кодирующего литическую протеазу AlpB, в составе экспрессионных векторов и введение таких рекомбинантных ДНК в клетки, например, бактерий, дрожжей, грибов, насекомых или млекопитающих, позволяет проводить наработку, выделение, очистку рекомбинантной литической протеазы AlpB, которая может быть использована в медицине, ветеринарии, растениеводстве, пищевой и косметической промышленности, а также в научно-исследовательских целях.
Уровень техники
На сегодняшний день известно несколько бактериолитических препаратов, основанных на внеклеточных бактериолитических ферментах. Например, ахромопептидаза из бактерии Achromobacter lyticus (TBL-1; Wako Pure Chemical Industries, Osaka), лизостафин из бактерии Staphylococcus simulans (Fluka, Sigma, Merck) и лизоамидаза из бактерии Lysobacter sp. XL1. Бактериолитические препараты отличаются как по составу входящих в них ферментов, так и по субстратной специфичности последних.
В состав препарата ахромопептидаза входит три литических фермента, продуцируемых бактерией Achromobacter lyticus M497-1 (Kessler F. «β-Lytic endopeptidases». In: Barett AJ (ed) Methods in Enzymol, 1995, Acad. Press, NY, p.740-756. Li S.L., Norioka S., Sakiyama F. «Molecular cloning and nucleotide sequence of the beta-lytic protease gene from Achromobacter lyticus». Journal of Bacteriology, 1990, V.172, p.6506. Li S.L., Norioka S., Sakiyama F. «Purification, staphylolytic activity, and cleavage sites of α-lytic protease from Achromobacter lyticus». Journal of Biochemistry, 1997, V.122, p.772-778. Li S.L., Norioka S., Sakiyama F. «Bacteriolytic activity and specificity of Achromobacter α-lytic protease». Journal of Biochemistry, 1998, V.124, p.332-339. Li S.L., Norioka S., Sakiyama F. «Purification, characterization, and primary structure of a novel cell wall hydrolytic amidase, Cwha, from Achromobacter lyticus». Journal of Biochemistry, 2000, V.127, p.1033-1039). Два из этих ферментов по первичной последовательности практически идентичны α- и β-литическим эндопептидазам бактерии Lysobacter enzymogenes.
Лизостафин представляет собой ферментный препарат, разрушающий клеточные стенки бактерий рода Staphylococcus. Препарат содержит три ферментных компонента, каждый из которых действует на разные участки пептидогликана. Наиболее активным компонентом лизостафина является Gly-Gly-эндопептидаза (Recsei Р.А., Gruss A.D., Novick R.P. «Cloning, sequence, and expression of the lysostaphin gene from Staphylococcus simulans». Proc. Natl. Acad. Sci USA, 1987, V.84, p.1127-1131).
Набор ферментов бактериолитического комплекса лизоамидаза отличается от такового ахромопептидазы и лизостафина. В ее состав входят: эндопептидазы Л1, Л4 и Л5, амидаза Л2 и мурамидаза Л3. Лизоамидаза обладает большим набором бактериолитических ферментов. Вследствие этого лизоамидаза разрушает более широкий спектр микроорганизмов, в частности бактерии родов Streptococcus, Staphylococcus, Micrococcus, Peptostreptococcus, Corinebacterium, Streptomyces, Bacillus, Arthrobacter и некоторые другие бактерии (Kulaev I.S., Stepnaya O.A., Tsfasman I.M., Chermenskaya T.S., Akimenko V.K., Ledova L.A., Zubritskaya L.G. «Bacteriolytic complex, method for production said complex and strain for earring out said method». 2006, Patent US 7150985. Severin A.I., Ilchenko V.Ya., Kulaev I.S. «Hydrolysis by Lisoamidase of Staphylococcus aureus 209 P cell wall peptidoglycan». Biochemistry (Russian), 1990, V.55, p.2078-2089. Severin Valiakhmetov A.Ya., Ilchenko V.Ya., Plotnikova Z.S., Kulaev I.S. «Hydrolysis of peptidoglycan from Micrococcus luteus BKM B-1314 cell walls by Lysoamidase». Biochemistry (Russian), 1990, V.55, p.986-992. Severin A.I., Tomasz A. «Selective lysis of cultures and cell walls of penicillin-resistant but not penicillin-susceptible Streptococcus pneumoniae strains by a murein hydrolase complex». Journal of Bacteriology, 1995, V.177, p.3316-3319), дрожжи Sacharomyces, Candida, Pseudozyma, препятствует прорастанию грибных и бактериальных спор (Ryzanova L.P., Stepnaya О.A., Suzina N.Е., Kulaev I.S. «Antifungal action of the lytic enzyme complex from Lysobacter sp. XL1». Process Biochemistry, 2005, V.40, p.557-564. Kulaev I.S., Stepnaya O.A., Tsfasman I.M., Chermenskaya T.S., Akimenko V.K., Ledova L.A., Zubritskaya L.G. «Bacteriolytic complex, method for production said complex and strain for earring out said method». 2006, Patent US 7,150,985 B2. Begunova Е.A., Chaika I.A., Stepnaya O.A., Safieva D.O., Budashov I.A, Kurochkin I.N., Varfolomeev S.D., Kulaev I.S. «Study of the effect of Lysoamidase on the spore form of bacteria of the genus Bacillus». Docl Biochem Biophys, 2006, V.408, p.152-154).
Эндопептидаза Л5 является минорным компонентом бактериолитического препарата лизоамидаза. Данный фермент гидролизует пептидные и амидные связи.
Субстратами для него являются белки, пептиды, клеточные стенки микроорганизмов (дрожжей, бактерий). В бактериальных пептидогликанах эндопептидаза Л5 гидролизует пептидные связи пептидной части пептидогликана. Эндопептидаза Л5 способна разрушать живые клетки Micrococcus roseus, Micrococcus luteus, Candida utilis, Corinebacterium flavum, Alcaligenes faecalis, а также термически обработанные клетки Staphylococcus aureus. Bacillus subtilis, Erwinia carotovora, Sacharomyces cerevisiae, Listeria monocytogenes, Pseudomonas aeruginosa, P. putida и т.д.
Таким образом, эндопептидаза Л5 не только в комбинации с другими ферментами, но и сама по себе может использоваться для подавления соответствующей патогенной микрофлоры.
Однако содержание этого фермента в культуральной жидкости бактерии Lysobacter sp. XL1 невысоко и составляет 0,2 мг/л. Низкий уровень продукции делает получение эндопептидазы Л5 из культуральной жидкости природного штамма-продуцента дорогим и неэффективным.
Существенным недостатком ахромопептидазы, лизостафина и лизоамидазы является возможное непостоянство состава этих препаратов из-за использования нестандартизованного сырья для выращивания бактерий-продуцентов: Achromobacter lyticus, Staphylococcus simulans и Lysobacter sp. XL1 соответственно. Эта проблема может быть решена за счет получения высокоочищенных препаратов отдельных ферментов, чего можно достичь за счет клонирования генов этих ферментов и создания штаммов-суперпродуцентов.
В научной литературе представлены публикации, описывающие последовательности генов литических эндопептидаз.
В 1988 году была определена первичная нуклеотидная последовательность гена α-литической протеазы бактерии Lysobacter enzymogenes (Silen J.L., McGrath C.N., Smith K.R., Agard D.A. «Molecular analysis of the gene encoding alpha-lytic protease: evidence for a preproenzyme». Gene, 1988, V.69, p.237-244). Фермент синтезируется в виде предшественника, который включает последовательность сигнального пептида (33 а.о.), пропептид (166 а.о.) и последовательность зрелого фермента (198 а.о.).
В 1990 году клонирован ген β-литической эндопептидазы бактерии Achromobacter lyticus - blp (Li S.L., Norioka S., Sakiyama F. «Molecular cloning and nucleotide sequence of the β-lytic protease gene from Achromobacter lyticus». J. Bacteriol., 1990, V.172, p.6506-6511).
В дополнение к вышеперечисленным были клонированы несколько генов, кодирующих эндопептидазы, для которых не была показана литическая активность.
В 2002 году был клонирован ген lepA, кодирующий лизин-специфическую эндопептидазу (LepA) бактерии Lysobacter sp.IB-9374. (Chohnan S., Nonaka J., Teramoto K., Taniguchi K., Kameda Y., Tamura H., Kurusu Y., Norioka S., Masaki Т., Sakiyama F. «Lysobacter strain with high lysyl endopeptidase production». FEMS Microbiol. Lett., 2002, V.213, p.13-20).
В 2004 году из ДНК бактерии Lysobacter sp.IB-9374 был клонирован ген lepB, кодирующий вторую лизин-специфическую эндопептидазу LepB (Chohnan S., Shiraki K., Yokota К., Ohshima М., Kuroiwa N., Ahmed K., Masaki Т., Sakiyama F. «A Second Lysine-Specific Serine Protease from Lysobacter sp. Strain IB-9374». Journal of Bacteriology, 2004, V.186, p.5093-5100). Ген lepB располагается рядом с геном lepA. Аминокислотные последовательности зрелых эндопептидаз LepA и LepB идентичны на 72%. Однако эндопептидаза LepB менее активна и более чувствительна к температуре и денатурирующим агентам, чем LepA.
Ген apI, кодирующий лизин-специфичную протеазу I бактерии Achromobacter lyticus М497-1, был клонирован в 1989 году (Ohara Т., Makino K., Shinagawa H., Nakata A., Norioka S., Sakiyama F. «Cloning, nucleotide sequence and expression of Achromobacter protease gene». J. Biol. Chem., 1989, V.264, Issue 34, p.20625-20631).
Гены, кодирующие ферменты бактериолитического комплекса лизоамидаза бактерии Lysobacter sp. XL1, не были клонированы ранее.
Сущность изобретения
Одним из объектов изобретения является литическая протеаза AlpB бактерии Lysobacter sp. XL1, имеющая последовательность аминокислот, приведенную в SEQ ID №1.
Обозначение «литическая протеаза AlpB бактерии Lysobacter sp. XL1» соответствует общепринятой номенклатуре обозначения генов и кодируемых ими белков и используется в данном изобретении для обозначения фермента, называемого ранее «эндопептидаза Л5 бактерии Lysobacter sp. XL1». Данный фермент гидролизует пептидные и амидные связи. Субстратами для него являются белки, пептиды, клеточные стенки микроорганизмов (дрожжей, бактерий). В бактериальных пептидогликанах Л5 гидролизует пептидные связи пептидной части пептидогликана.
Эндопептидаза Л5 способна разрушать живые клетки Micrococcus roseus, Micrococcus luteus, Candida utilis, Corinebacterium flavum, Alcaligenes faecalis, а также термически обработанные клетки Staphylococcus aureus, Bacillus subtilis, Erwinia carotovora, Sacharomyces cerevisiae, Listeria monocytogenes, Pseudomonas aeruginosa, P. putida и т.д.
Литическая протеаза AlpB имеет длину 399 а.о. и синтезируется в виде препрофермента (Фиг.1), который состоит из сигнального пептида длиной 28 а.о. (с 1 по 28 а.о., SEQ ID №1), прочасти длиной 166 а.о. (с 29 по 194 а.о., SEQ ID №1) и зрелого фермента длиной 205 а.о. (с 195 по 399 а.о., SEQ ID №1). Собственно зрелый фермент и обуславливает специфическую активность литической протеазы AlpB.
Другим объектом изобретения является фрагмент ДНК бактерии Lysobacter sp. XL1, представленный в SEQ ID №2, который содержит ген alpB, кодирующий литическую протеазу AlpB бактерии Lysobacter sp. XL1; варианты последовательности ДНК, представленной в SEQ ID №2, которые кодируют литическую протеазу AlpB бактерии Lysobacter sp. XL1. Данный фрагмент ДНК либо его варианты может быть использован для продукции литической протеазы AlpB как in vivo, так и in vitro.
Еще одним объектом изобретения является способ получения литической протеазы AlpB из неактивного денатурированного препарата профермента методом ренатурации и протеазной активации in vitro. Способ получения литической протеазы AlpB включает клонирование в бактерии Escherichia coli участка гена alpB бактерии Lysobacter sp. XL1, последовательность которого представлена в SEQ ID №2, кодирующего профермент литической протеазы AlpB с нарушенным сигнальным пептидом или без него, продукцию этого профермента в клетках бактерии Escherichia coli, наработку его в неактивной денатурированной форме в виде телец включения, ренатурацию и активацию литической протеазы AlpB in vitro.
Способ получения литической протеазы AlpB подразумевает клонирование в экспрессионном векторе в клетках бактерии Escherichia coli участка гена alpB бактерии Lysobacter sp. XL1. А именно, клонируют фрагмент гена alpB с 85 по 1200 н.о. в соответствии с последовательностью, представленной в SEQ ID №2. Данный участок гена alpB бактерии Lysobacter sp. XL1 кодирует профермент литической протеазы AlpB без сигнального пептида.
В полученной в результате клонирования рекомбинантной плазмиде фрагмент гена alpB помещается под контроль эффективных сигналов транскрипции и трансляции. Клонирование может быть проведено таким образом, что при экспрессии клонированного участка гена alpB в клетках бактерии Escherichia coli будет синтезироваться профермент, слитый на N- и/или С-концах с дополнительными аминокислотными последовательностями, предназначенными для повышения эффективности экспрессии, выделения и очистки продуцированного профермента литической протеазы AlpB.
Затем полученную рекомбинантную плазмиду вводят в штамм бактерии Escherichia coli, в котором проводится экспрессия клонированного гена. Профермент AlpB, не содержащий функциональный сигнальный пептид, накапливается в штамме-продуценте бактерии Escherichia coli в неактивной денатурированной форме в виде телец включения. Тельца включения растворяют в буфере, содержащем хаотропный агент. Получение активной формы литической протеазы AlpB проводят методом ренатурации in vitro и активации ренатурированого препарата при помощи протеиназной обработки.
Описание изобретения
Эндопептидаза Л5 бактерии Lysobacter sp. XL1 совместно с рядом других литических ферментов секретируется штаммом-хозяином в культуральную жидкость.
Решение проблемы повышения уровня продукции эндопептидазы Л5 может быть получено за счет создания искусственного штамма-продуцента этого фермента, что возможно на основании информации о последовательности ДНК бактерии Lysobacter sp. XL1, содержащей ген эндопептидазы Л5.
Выполнение работы проводили с использованием стандартных методик генной инженерии, описанных в книге (Sambrook J., Fritsch E.F., Maniatis T. "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press, 1989).
Для получения данного изобретения авторами была сконструирована библиотека генов бактерии Lysobacter sp. XL1 в клетках бактерии Escherichia coli. В составе полученной библиотеки генов был идентифицирован клон, содержащий фрагмент ДНК бактерии Lysobacter sp. XL1 с геном эндопептидазы Л5.
Для идентификации специфического клона было проведено следующее исследование. В процессе клонирования гена alpA литической протеазы AlpA была определена нуклеотидная последовательность фрагмента геномной ДНК бактерии Lysobacter sp. XL1. В данном фрагменте ДНК на расстоянии 601 п.н. от 3'-конца гена alpA был обнаружен ген ранее неизвестной литической эндопептидазы, который был обозначен alpB.
Ранее из культуральной жидкости бактерии Lysobacter sp. XL1 была очищена и определена последовательность аминокислот N-конца эндопептидазы Л5.
Затем установленная нуклеотидная последовательность гена alpB была транслирована in silico и проведен ее компьютерный анализ. В результате анализа было показано соответствие аминокислотных последовательностей N-конца эндопептидазы Л5 и фрагмента аминокислотной последовательности белка, предсказанного из нуклеотидной последовательности гена alpB бактерии Lysobacter sp. XL1 (Фиг.2).
Анализ базы данных последовательностей ДНК и белков GenBank (Benson D.A., Karsch-Mizrachi I., Lipman D.J., Ostell J., Sayers E.W. «GenBank». Nucleic Acids Res. 2009, V.37 (Database issue), D26-31. http://www.ncbi.nlm.nih.gov/Genbank/index.html) показал, что идентифицированная нами эндопептидаза Л5 является уникальным ферментом, так как идентичных белков обнаружено не было.
Таким образом, в клонированном фрагменте ДНК бактерии Lysobacter sp. XL1 был идентифицирован ген эндопептидазы Л5, который в соответствии с общепринятой номенклатурой был обозначен alpB, и кодируемый этим геном фермент был обозначен как «литическая протеаза AlpB». Нуклеотидная последовательность гена alpB представлена в SEQ ID №2. Аминокислотная последовательность литической протеазы AlpB представлена в SEQ ID №1.
Литическая протеаза AlpB имеет длину 399 а.о. и синтезируется в виде препрофермента (Фиг.1), который состоит из сигнального пептида длиной 28 а.о. (с 1 по 28 а.о., SEQ ID №1), прочасти длиной 166 а.о. (с 29 по 194 а.о., SEQ ID №1) и зрелого фермента длиной 205 а.о. (с 195 по 399 а.о., SEQ ID №1). Последний обуславливает специфическую активность литической протеазы AlpB.
Наличие сигнального пептида и сайт его отщепления были предсказаны при помощи программы «SignalP 3.0» (http://www.cbs.dtu.dk/services/SignalP/) (Nielsen H., Engelbrecht J., Brunak S., Gunnar von Heijne. «Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites». Protein Engineering, 1997, V.10, p.1-6). Граница прочасти и зрелой части литической протеазы AlpB была установлена исходя из полученных нами данных об N-концевой последовательности зрелого фермента, выделенного из культуральной жидкости бактерии Lysobacter sp. XL1.
Установленные нами данные о нуклеотидной последовательности гена alpB, a также данные об аминокислотной последовательности кодируемого им белка позволяют проводить конструирование штаммов-продуцентов литической протеазы AlpB бактерии Lysobacter sp. XL1.
Клонирование фрагмента ДНК бактерии Lysobacter sp. XL1, кодирующего литическую протеазу AlpB, в составе экспрессионных векторов и введение таких рекомбинантных ДНК в соответствующие штаммы-хозяева, например клетки бактерий, дрожжей, грибов, насекомых или млекопитающих, позволяет проводить наработку, выделение и очистку рекомбинантного литической протеазы AlpB и ее последующее использование для лизиса патогенных микроорганизмов.
Перечень фигур
В SEQ ID №1 приведена аминокислотная последовательность литической протеазы AlpB бактерии Lysobacter sp. XL1.
В SEQ ID №2 приведена нуклеотидная последовательность гена alpB бактерии Lysobacter sp. XL1.
На Фиг.1 представлена структура препрофермента AlpB бактерии Lysobacter sp. XL1.
На Фиг.2 представлено выравнивание фрагментов предсказанной аминокислотной последовательности литической протеазы AlpB бактерии Lysobacter sp. XL1 и экспериментально установленной аминокислотной последовательности N-конца зрелой формы эндопептидазы Л5 бактерии Lysobacter sp. XL1.
На Фиг.3 представлена экспрессия гена alpB бактерии Lysobacter sp. XL1 в клетках бактерии Escherichia coli, которая проявляется в виде зон лизиса вокруг клеток бактерии-продуцента литической протеазы AlpB.
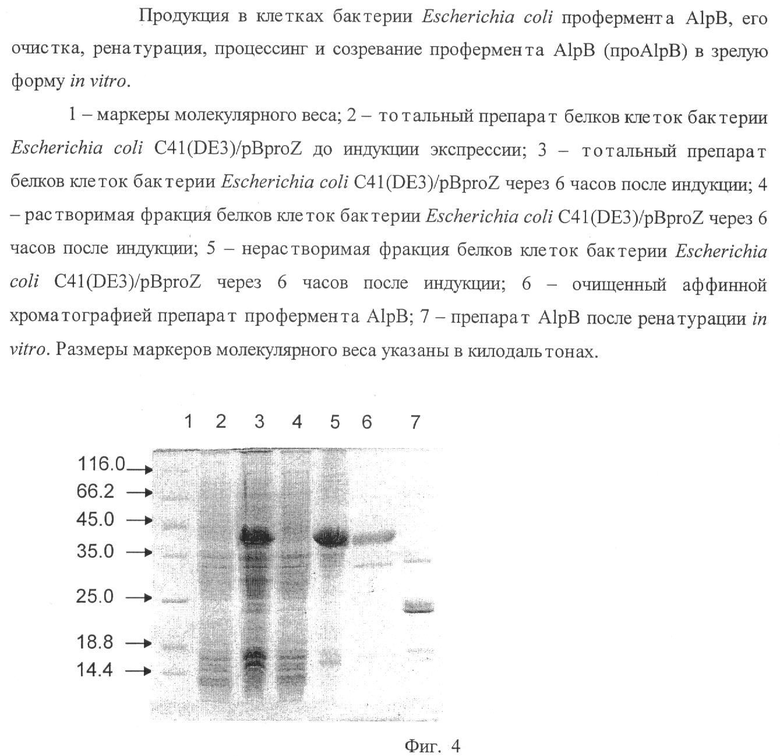
На Фиг.4 представлена продукция в клетках бактерии Escherichia coli профермента AlpB, его очистка, ренатурация, процессинг и созревание профермента AlpB (про AlpB) в зрелую форму in vitro.
Возможность осуществления предлагаемых изобретений подтверждается приведенными ниже примерами, но не ограничивается ими.
Пример 1. Клонирование гена alpB в экспрессионный вектор рЕТ-29а(+) и экспрессия его в клетках бактерии Escherichia coli штамма BL21(DE3)/pLysE.
1. Фрагмент ДНК бактерии Lysobacter sp. XL1, который соответствует с 1 по 1200 н.о. последовательности, представленной в SEQ ID №2, амплифицируют при помощи ПЦР с использованием олигонуклеотидных праймеров ALPII-up и ALPII-lo. Праймеры ALPII-up и ALPII-lo имеют следующие последовательности нуклеотидов:
5'-TTTTTACATATGTCCGTATCGAAGTCGAATCT-3' (ALPII-up);
5'-AAATTACTCGAGTCAGCTCGTGACCAGGGC-3' (ALPII-lo).
Для проведения последующей процедуры клонирования в последовательность праймера ALPII-up вводят сайт эндонуклеазы рестрикции NdeI, а в последовательность праймера ALPII-lo сайт эндонуклеазы рестрикции XhoI.
ПЦР-смесь объемом 25 мкл содержит 1х буфер для ДНК-полимеразы Pwo, 3 мМ MgSO4, 200 мкМ каждого дезоксирибонуклеозидтрифосфата, 4% диметилсульфоксида, 0,5 мкМ каждого праймера, 20 нг геномной ДНК бактерии Lysobacter sp. XL1 и 1 единицу активности ДНК-полимеразы Pwo. ПЦР проводят на приборе «GeneAmp PCR System 2400» (Perken Elmer, США) при следующем режиме: 94°С - 3 мин, затем 3 цикла: 94°С - 30 с, 60°С - 30 с, 72°С - 70 с, затем 27 циклов: 94°С - 30 с, 66°С - 30 с, 72°С - 70 с, затем достройка в течение 3 мин.
2. Полученный ПЦР-фрагмент и плазмиду рЕТ-29а(+) (Novagen, США) гидролизуют эндонуклеазами рестрикции NdeI и XhoI и лигируют между собой при помощи ДНК-лигазы фага Т4.
3. Лигазной смесью трансформируют клетки штамма бактерии Escherichia coli DH5α (Cohen SN, Chang AC, and Hsu L. Nonchromosomal antibiotic resistance in bacteria: genetic transformation of Escherichia coli by R-factor DNA, Proc Natl Acad Sci USA 1972 Aug; 69(8) 2110-4) и проводят скрининг устойчивых к канамицину клонов на наличие рекомбинантной плазмиды.
4. Проводят выделение рекомбинантных плазмид, их рестрикционный анализ и секвенируют в составе этих плазмид фрагмент ДНК, соответствующий последовательности гена alpB бактерии Lysobacter sp. XL1. Анализируют данные секвенирования и отбирают рекомбинантную плазмиду, в которой последовательность гена alpB соответствует последовательности, представленной в SEQ ID №2. В результате получают рекомбинантную плазмиду, обозначенную, например, pALPII-29a, которая имеет размер 6441 п.н. и содержит ген alpB бактерии Lysobacter sp. XL1 под контролем гибридного промотора T7 lac.
5. Клетки штамма бактерии Escherichia coli BL21(DE3)/pLysE трансформируют плазмидой pALPII-29a или рЕТ-29а(+) и проводят селекцию трансформантов на агаризованной среде LB (бакто-триптон - 10 г/л, бакто-дрожжевой экстракт - 5 г/л, NaCl - 10 г/л), содержащей канамицин в концентрации 25 мкг/мл и хлорамфеникол в концентрации 30 мкг/мл.
6. Клетки бактерии Escherichia coli BL21(DE3)/pLysE, содержащие плазмиду pALPII-29a или рЕТ-29а(+), перекалывают на индикаторные чашки с агаризованной средой Дэвиса (декстроза - 1 г/л, калий фосфорнокислый однозамещенный - 2 г/л, калий фосфорнокислый двузамещенный - 7 г/л, цитрат натрия - 0,5 г/л, сульфат магния - 0,1 г/л, сульфат аммония - 1,0 г/л, агар - 15 г/л), содержащей канамицин в концентрации 25 мкг/мл, хлорамфеникол в концентрации 30 мкг/мл, 0,1 мМ негидролизуемого индуктора изопропил-β-D-тиогалактопиранозида и 2 мг/мл клеточных стенок автоклавированного золотистого стафилококка Staphylococcus aureus 209-P. Чашки инкубируют при 28°С в течение 1,5-2 суток. Об экспрессии гена alpB судят по появлению зон лизиса вокруг колоний (Фиг.3).
Пример 2. Получение экспрессионной конструкции, содержащей фрагмент гена alpB, кодирующий профермент литической протеазы AlpB бактерии Lysobacter sp. XL1 без сигнального пептида, и экспрессия данного фрагмента гена alpB в клетках бактерии Escherichia coli.
1. Фрагмент ДНК бактерии Lysobacter sp. XL1, который соответствует с 85 по 1200 н.о. последовательности, представленной в SEQ ID №2, амплифицируют при помощи ПЦР с использованием праймеров BproZ-up и ALPII-lo. Праймеры BproZ-up и ALPII-lo имеют следующие последовательности нуклеотидов:
5'-TTTTTCATATGACCGACGGTCTCAACCCGA-3' (BproZ-up);
5'-AAATTACTCGAGTCAGCTCGTGACCAGGGC-3' (ALPII-lo).
Для проведения последующей процедуры клонирования в последовательность праймера BproZ-up вводят сайт эндонуклеазы рестрикции NdeI, а в последовательность праймера ALPII-lo - сайт эндонуклеазы рестрикции XhoI.
ПЦР-смесь объемом 25 мкл содержит 1х буфер для ДНК-полимеразы Pwo, 3 мМ MgSO4, 200 мкМ каждого дезоксирибонуклеозидтрифосфата, 4% диметилсульфоксида, 0,5 мкМ каждого праймера, 0,1 нг плазмидной ДНК pALPII-29a и 1 единицу активности ДНК-полимеразы Pwo. ПЦР проводят на приборе «GeneAmp PCR System 2400» (Perken Elmer, США) при следующем режиме: 94°С - 3 мин, затем 4 цикла: 94°С - 30 с, 60,5°С - 30 с, 72°С - 70 с, затем 26 циклов: 94°С - 30 с, 70°С - 30 с, 72°С - 70 с, затем достройка в течение 3 мин.
2. Полученный ПЦР-фрагмент и плазмиду pET-15b (Novagen, США) гидролизуют эндонуклеазами рестрикции NdeI и XhoI и лигируют между собой при помощи ДНК-лигазы фага Т4.
3. Лигазной смесью трансформируют клетки бактерии Escherichia coli штамма DH5α (по примеру 1) и проводят скрининг устойчивых к ампициллину клонов на наличие рекомбинантной плазмиды.
4. Проводят выделение рекомбинантных плазмид, их рестрикционный анализ и секвенируют в составе этих плазмид фрагмент ДНК, соответствующий последовательности гена alpB бактерии Lysobacter sp. XL1. Анализируют данные секвенирования и отбирают рекомбинантную плазмиду, в которой последовательность фрагмента гена alpB соответствует последовательности, представленной в SEQ ID №2. В результате получают рекомбинантную плазмиду, обозначенную как pBproZ. Данная плазмида имеет размер 6824 п.н., содержит фрагмент гена alpB бактерии Lysobacter sp. XL1 под контролем гибридного промотора T7lac и кодирует профермент литической протеазы AlpB бактерии Lysobacter sp. XL1 без сигнального пептида, слитый на N-конце с последовательностью из 6-ти остатков гистидина.
5. Клетки бактерии Escherichia coli штамма C41(DE3) трансформируют плазмидой pBproZ или рЕТ-15b (по примеру 1) и проводят селекцию трансформантов на агаризованной среде LB, содержащей ампициллин в концентрации 100 мкг/мл.
6. 100 мл среды LB, содержащей ампициллин в концентрации 100 мкг/мл, засевают инокулятом клеток бактерии Escherichia coli C41(DE3)/pBproZ, полученным смывом с чашек, и инкубируют на роторной качалке при 37°С и 150 об/мин до достижения культурой оптической плотности 0,7 при длине волны 590 нм. Экспрессию индуцируют добавлением изопропил-β-D-тиогалактопиранозида до концентрации 1 мМ. Индуцированную культуру инкубируют на роторной качалке 37°С и 150 об/мин в течение 6-ти часов.
7. Осаждают биомассу центрифугированием при 4°С и 4000×g в течение 20 минут.
8. Осажденную биомассу ресуспендируют в 10 мл буфера, содержащего 50 мМ Трис-HCl рН 8.0, 100 мМ NaCl и 5 мМ 2-меркаптоэтанола, после чего разрушают при помощи ультразвуковой дезинтеграции на ледяной бане. Полученный лизат центрифугируют 20 минут при 10000×g и температуре 4°С.
9. Клеточные фракции анализируют электрофорезом в 12% полиакриламидном геле в денатурирующих условиях на наличие пробелка AlpB. В данных условиях пробелок AlpB накапливается в нерастворимой фракции клеток в тельцах включения (Фиг.4).
Пример 3. Выделение, очистка и ренатурация литической протеазы AlpB из телец включения, образуемых при продукции профермента AlpB в клетках бактерии Escherichia coli.
1. Трансформируют компетентные клетки бактерии Escherichia coli C41(DE3) плазмидой pBproZ (по примеру 1), несущей ген alpB, без последовательности, кодирующей сигнальный пептид. Трансформированные клетки высевают на чашки с агаризованной средой LB, содержащей ампициллин в концентрации 75 мкг/мл, и инкубируют ночь при 37°С. В результате трансформации получают клетки бактерии Escherichia coli C41(DE3)/pBproZ.
2. 100 мл среды LB, содержащей ампициллин в концентрации 100 мкг/мл засевают инокулятом клеток бактерии Escherichia coli C41(DE3)/pBproZ, полученным смывом с чашек, и инкубируют на роторной качалке при 37°С и 150 об/мин до достижения культурой оптической плотности 0,7 при длине волны 590 нм. Экспрессию индуцируют добавлением негидролизуемого индуктора изопропил-β-D-тиогалактопиранозида до 1 мМ. Индуцированную культуру инкубируют на роторной качалке 37°С и 150 об/мин в течение 6-ти часов.
3. Осаждают биомассу центрифугированием при 4°С и 4000×g в течение 20 минут.
4. Осажденную биомассу ресуспендируют в 10 мл буфера, содержащего 50 мМ Трис-HCl рН 8.0, 100 мМ NaCl и 5 мМ 2-меркаптоэтанола, после чего разрушают пятью сериями ультразвуковой дезинтеграции на ледяной бане. Полученный лизат центрифугируют 20 минут при 4°С на скорости 10000×g.
5. Дважды отмывают осажденные тельца включения 10 мл буфера, содержащего 50 мМ Трис-HCl рН 8.0, 100 мМ NaCl, 5 мМ 2-меркаптоэтанола и 0.1% Тритона Х-100. После каждой отмывки тельца включения осаждают центрифугированием при 10000×g и 4°С в течение 20 минут.
6. Растворяют тельца включения в 10 мл буфера, содержащего 8 М мочевины, 50 мМ натриево-фосфатного буфера рН 7,0 и 300 мМ NaCl. Для окончательного растворения инкубируют препараты в течение ночи при 4°С. Центрифугируют растворенные тельца включения 20 минут при 4°С при 1000×g.
7. Супернатант наносят на 5 мл сорбента Ni-NTA (Qiagen, Германия), собирают проскок, затем колонку промывают 50 мл буфера, содержащего 8 М мочевины, 50 мМ натриево-фосфатного буфера рН 7, 1 М NaCl и 20 мМ имидазола.
8. Элюируют белки с сорбента 15 мл буфера, содержащего 8 М мочевины, 50 мМ натриево-фосфатного буфера рН 7,0, 300 мМ NaCl и 200 мМ имидазола.
9. Анализируют эффективность очистки при помощи электрофореза в 12% полиакриламидном геле в денатурирующих условиях.
10. Повторяют стадии 7-9 до максимального извлечения белка из первоначального раствора.
11. Диализуют препарат при 4°С в течение ночи против 100 объемов буфера, содержащего 8 М мочевины, 10 мМ натриево-фосфатного буфера рН 8,0, 5 мМ 2-меркаптоэтанола и 1 мМ этилендиаминтетрауксусной кислоты.
12. Оценивают концентрацию полученного белкового препарата спектрофотометрически при длине волны 280 нм. Выход белка составляет ~67 мг при очистке из 100 мл культуры.
13. Разбавляют полученный препарат проAlpB буфером, содержащим 8 М мочевины, 10 мМ натриево-фосфатного буфера рН 8,0 и 5 мМ 2-меркаптоэтанола до концентрации 0,1 мг/мл.
14. Диализуют разбавленный до концентрации 0,1 мг/мл препарат проAlpB в течение ночи против 100 объемов раствора, содержащего 50 мМ Трис-HCl рН 8,5, 250 мМ NaCl, 0,4 М сахарозы, 1 мМ этилендиаминтетрауксусной кислоты и 1 мМ дитиотреитола.
15. Центрифугируют препарат после диализа при 1000×g и 4°С в течение 20 минут, осадок выбрасывают.
16. Активируют полученный зрелый белок AlpB добавлением трипсина в молярном соотношении 1:1000 с последующей инкубацией 15 минут при 37°С.
17. Анализируют эффективность ренатурации электрофорезом в 12% полиакриламидном геле в денатурирующих условиях (фиг.4).
18. Анализируют бактериолитическую активность полученного препарата с использованием в качестве субстрата автоклавированных клеток Staphylococcus aureus 209-Р. Для этого к 2 мл суспензии клеток стафилококка в 0,01 М Tris-HCl, рН 8,0 (поглощение при 540 нм 0,5) добавляют ферментный препарат в объеме 100 мкл. Смесь инкубируют 5-30 мин при 37°С. Реакцию останавливают помещением пробирок в ледяную баню. Затем измеряют поглощение при длине волны 540 нм на спектрофотометре. За единицу бактериолитической активности (ЛЕ) принимают такое количество фермента, которое приводит к снижению поглощения бактериальной суспензии на 0,01 оптическую единицу за 1 мин. Полученные препараты зрелого активного белка AlpB в концентрации 0,04 мг/мл обладают удельной активностью 1520 ЛЕ/мл.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛИТИЧЕСКАЯ ПРОТЕАЗА AlpA БАКТЕРИИ LYSOBACTER SP. XL1, ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ЛИТИЧЕСКУЮ ПРОТЕАЗУ AlpA БАКТЕРИИ LYSOBACTER SP. XL1, И СПОСОБ ПОЛУЧЕНИЯ ЛИТИЧЕСКОЙ ПРОТЕАЗЫ AlpA БАКТЕРИИ LYSOBACTER SP. XL1 | 2009 |
|
RU2407782C2 |
| Применение Т5 промотора для экспрессии гена бета-литической протеазы Lysobacter capsici VKM B-2533 и штамм L. capsici blp P-продуцент данной протеазы | 2021 |
|
RU2787787C1 |
| ШТАММ БАКТЕРИИ Escerichia coli XL1-blue/pQS-G3, ПРОДУЦЕНТ ТЕРМОСТАБИЛЬНОЙ ЛИПАЗЫ БАКТЕРИИ Geobacillus stearothermophilus G3 | 2013 |
|
RU2540873C1 |
| ФЕРМЕНТ ПРОТЕАЗА И ЕГО ПРИМЕНЕНИЯ | 2012 |
|
RU2611043C2 |
| ГИДРОЛАЗА ПЕПТИДОГЛИКАНА, ЭКСПРЕССИОННАЯ ПЛАЗМИДА, СОДЕРЖАЩАЯ ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ГИДРОЛАЗУ ПЕПТИДОГЛИКАНА, БАКТЕРИЯ-ПРОДУЦЕНТ И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ГИДРОЛАЗЫ ПЕПТИДОГЛИКАНА | 2012 |
|
RU2547584C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pEstPc, КОДИРУЮЩАЯ ПОЛИПЕПТИД СО СВОЙСТВАМИ ЭСТЕРАЗЫ Psychrobacter cryohalolentis K5, И ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ ПОЛИПЕПТИДА СО СВОЙСТВАМИ ЭСТЕРАЗЫ Psychrobacter cryohalolentis K5 | 2011 |
|
RU2478708C1 |
| НОВАЯ ПРОТЕАЗА ГРИБОВ И ЕЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2566549C2 |
| Нуклеотидная последовательность, кодирующая фермент литиказу, и панель олигонуклеотидов для получения синтетической нуклеотидной последовательности гена литиказы | 2023 |
|
RU2826150C1 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2351653C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |





Изобретение относится к биотехнологии и представляет собой литическую протеазу AlpB бактерии Lysobacter sp. XL1. Изобретение относится также к фрагменту ДНК бактерии Lysobacter sp. XL1, кодирующему литическую протеазу AlpB, и способу получения литической протеазы AlpB in vitro. Для получения фермента проводят клонирование в клетках бактерии Escherichia coli участка фрагмента ДНК, содержащего фрагмент гена alpB бактерии Lysobacter sp. XL1, кодирующего профермент литической протеазы AlpB с нарушенным сигнальным пептидом или без него, продукцию этого профермента в клетках бактерии Escherichia coli, наработку его в неактивной денатурированной форме в виде телец включения, ренатурацию и активацию профермента литической протеазы AlpB in vitro. Изобретение позволяет расширить арсенал литических протеаз. 3 н.п. ф-лы, 4 ил.
1. Литическая протеаза AlpB бактерии Lysobacter sp. XL1, имеющая последовательность аминокислот, приведенную в SEQ ID №1.
2. Фрагмент ДНК бактерии Lysobacter sp. XL1, представленный в SEQ ID №2, который содержит ген alpB, кодирующий литическую протеазу AlpB бактерии Lysobacter sp. XL1 по п.1 или варианты фрагмента ДНК, представленного в SEQ ID №2, которые кодируют литическую протеазу AlpB бактерии Lysobacter sp. XL1 по п.1.
3. Способ получения литической протеазы AlpB по п.1, включающий клонирование в клетках бактерии Escherichia coli участка фрагмента ДНК по п.2, содержащего фрагмент гена alpB бактерии Lysobacter sp. XL1, кодирующего профермент литической протеазы AlpB с нарушенным сигнальным пептидом или без него, продукцию этого профермента в клетках бактерии Escherichia coli, наработку его в неактивной денатурированной форме в виде телец включения, ренатурацию и активацию профермента литической протеазы AlpB in vitro.
| CHOHNAN S, et | |||
| al., A second lysine-specific serine protease from Lysobacter sp | |||
| Способ и устройство для просушки движущегося бумажного, папочного и т.п. полотна в сушильной части бумагоделательной машины | 1926 |
|
SU9374A1 |
| J Bacteriol | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| EPSTEIN D.M | |||
| et | |||
| al., The alpha-lytic protease gene of Lysobacter enzymogenes | |||
| The nucleotide sequence predicts a large prepro-peptide with homology to pro-peptides of other chymotrypsin-like enzymes, J | |||
| Biol | |||
| Chem, 263(32), 1988, p.16586-16590 | |||
| БАКТЕРИОЛИТИЧЕСКИЙ КОМПЛЕКС, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ШТАММ ДЛЯ ОСУЩЕСТВЛЕНИЯ СПОСОБА | 2000 |
|
RU2193063C2 |

