ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к ферменту сериновой протеазе, в частности к ферменту сериновой протеазе грибов, подходящей для различного использования, в частности в моющих средствах. Изобретение относится к молекуле нуклеиновой кислоты, кодирующей указанный фермент, рекомбинантному вектору, клетке-хозяину, продуцирующей указанный фермент, ферментной композиции, содержащей указанный фермент, а также процессу изготовления такой композиции. Данное изобретение также относится к различным применениям указанного фермента и композиций, содержащих указанный фермент.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Микробные протеазы относятся к наиболее важным гидролитическим ферментам и находят применение в различных отраслях промышленности, таких как производство моющих средств, пищевая промышленность, кожная промышленность, фармацевтическая промышленность, диагностика, переработка отходов и извлечение серебра. На долю внеклеточных протеаз микробного происхождения приходится основная часть продаж ферментов промышленного назначения во всем мире (Cherry and Fidantsef, 2003). Приблизительно 90% существующих на рынке протеаз представляют собой ферменты, обладающие моющими свойствами (Gupta et al., 2002). В настоящее время существующие на рынке детергентные (моющие) препараты имеют в своем составе щелочные сериновые протеазы естественного происхождения (ЕС 3.4.21) из субтилизинового семейства, или субтилизинов (ЕС 3.4.21.62), вырабатываемые представителями рода Bacillus, или рекомбинантные препараты этих протеаз (Maurer, 2004).
Примерами существующих на рынке протеаз являются субтилизин Carlsberg (Алькалаза®), субтилизин 309 (Савиназа®), субтилизин 147 (Эспераза®), Канназа®, Эверлаза®, Овозим®, а также протеаза для мытья в холодной воде Поларзим® (Novozymes A/S, DK), Пурафект®, Пурафект® Окс, Пурафект® Прайм и Пропераза® (Genencor Int., Inc., USA), а также протеазы серий BLAP S и X (Henkel, DE).
Несколько щелочных сериновых протеаз и генов, кодирующих эти ферменты, были также выделены из эукариотических организмов, включая дрожжи и нитчатые грибы. Патент США 3,652,399 и ЕР 519229 (Takeda Chemical Industries, Ltd., JP) описывают щелочную протеазу, вырабатываемую родом Fusarium (бесполая стадия, телеоморфа) или Gibberella, (половая стадия, анаморфа), в частности видами Fusarium sp. S-19-5 (АТСС 20192, IFO 8884), F. oxysporum f. sp. lini (IFO 5880) или G. saubinetti (АТСС 20193, IFO6608), подходящую для изготовления моющих средств и других чистящих композиций. WO1994025583 (NovoNordisk A/S, DK) описывает активный фермент трипсиноподобную протеазу, получаемую из видов Fusarium, в частности, из штамма F. oxysporum (DSM 2672), и последовательность ДНК, кодирующую его. Аминокислотная и нуклеотидная последовательности сериновых протеаз, получаемых из F. equiseti и F. acuminatum были описаны в WO 2010125174 и WO 2010125175, соответственно (АВ Enzymes Oy, FI). Также были описаны щелочные протеазы, получаемые из грибов видов Tritirachium и Conidiobolus (обзор Anwar and Saleemuddin 1998).
Главной проблемой при применении протеаз в жидких моющих средствах является их нестабильность. В жидких моющих средствах ферменты находятся в непосредственном контакте с водой и хаотропными агентами, такими как анионные сурфактанты и комплексообразующе агенты, что может привести к необратимой денатурации. Протеазы разрушают белки, включая другие ферменты, входящие в состав моющих средств, и самих себя. Автопротеолиз усиливается в присутствии сурфактантов и при нагревании. Таким образом, стабильность жидких моющих средств, содержащих протеазы, представляет основное препятствие для совершенствования продукции (Maurer, 2010).
Для улучшения стабильности сериновых протеаз, применяемых в промышленности, используют различные способы. WO 92/03529 (NovoNordisk A/S, DK), US 2009/096916 (Genencor Int. Inc., US) и WO 2007/145963 (Procter & Gamble Co., US) описывают применение обратимого ингибитора протеаз пептидного или белкового происхождения. Жидкие детергентные композиции, содержащие протеазы, часто имеют в своем составе такие ингибиторы протеаз, как борная кислота с добавлением или без добавления полиолов для ингибирования активности протеаз. Одним из примеров таких ингибитров является 4-формил-фенил-бороновая кислота (4-FPBA), описанная в US 2010/0120649 (Novozymes A/S, DK). Стабильность протеаз также была улучшена с применением комбинации галоидных солей с полиолами (WO 02/08398, Genencor Int. Inc., US). EP 0352244A2 (NovoNordisk A/S, DK) предлагает улучшение стабильности ферментов, полученных из Bacillus, с помощью амфотерных соединений, таких как сурфактанты.
Основываясь на данных кристаллографии и анализа сходства последовательностей гомологичных белков, можно создавать варианты с улучшенной стабильностью и/или улучшенным качеством работы. С помощью сайт-специфического и/или случайного мутагенеза были разработаны варианты сериновых протеаз естественного происхождения, обладающие улучшенной каталитической эффективностью и/или улучшенной стабильностью по отношению к температуре, окисляющим агентам и различным условиям стирки, а также улучшенной стабильностью в жидких моющих средствах при хранении.
Термомиколин ЕС 3.4.21.65, выделенный как внеклеточная щелочная эндопептидаза, вырабатывается термофильным грибом Malbranchea pulcella var. sulfurea. Термомиколин представляет собой одноцепочечный белок, состоящий из 325 аминокислотных остатков. Он имеет активный центр с последовательностью Leu-Ser-(Gly)-Thr-Ser*-Met-, типичной для представителей семейства субтилизинов. Термомиколин имеет одну дисульфидную связь, что является редкостью. Термомиколин является не настолько термостабильным, как внеклеточные сериновые протеазы термофильных бактерий, но гораздо более стабильным, чем большинство протеиназ грибов (Gaucher and Stevenson, 2004). Согласно Ong and Gaucher (1975), термическая инактивация термомиколина происходит при 73°С в присутствии 10 мМ Ca2+. Термомиколин гидролизует казеин в широком диапазоне рН. Оптимальное значение рН для гидролиза казеина составляет приблизительно 8,5.
Abu-Shady et al. (2001) описывают свойства протеазы Malbranchea sulfurea, представляющей собой местный изолят из образцов почв, собранных на скотобойнях в Египте, выращенный в культуре для получения протеазного фермента. В этой публикации описана относительная активность протеазы М. sulphurea в присутствии некоторых моющих средств в низких концентрациях (0,7%) при 30-90°С при инкубации в течение 15-60 мин, т.е. при условиях, напоминающих условия стирки. Однако, в ней не указывается каких-либо сведений о способности удалять загрязнения или стабильности этой протеазы в самом моющем средстве при хранении, являющихся необходимыми характеристиками для возможного применения протеазы в составе моющих средств. В публикации также описаны температурный и рН-профили частично очищенной протеазы. Температурой оптимум протеазы составляет 50°С, а оптимум рН - 9,0.
Несмотря на тот факт, что было опубликовано множество публикаций, обзоров и статей, описывающих сериновые протеазы грибов, полученные из различных микроорганизмов, например, низкотемпературные щелочные протеазы таких микроорганизмов, как актиномицеты (Nocardiopsis dassonvillei) и грибы (Paecilomyces marquandii), например, ЕР 0290567 и ЕР 0290569 (Novo Nordisk A/S, DK), по-прежнему существует большая потребность в новых протеазах, подходящих и являющихся эффективными для модификации, разложения и удаления белкового материала различных видов загрязнений, в частности, при низких или средних температурах, и обладающих стабильностью в присутствии моющих средств, обладающих различными свойствами. В связи с наличием у сериновых протеаз автокаталитических свойств, стабильность в процессе хранения также имеет большое значение.
Кроме того, желательно, чтобы сериновые протеазы было можно получать в большом количестве, а процесс их дальнейшей обработки путем простого отделения от ферментативного бульона и мицелия был экономически эффективным.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Целью настоящего изобретения является получение сериновой протеазы грибного происхождения, активной в широком диапазоне рН и особенно хорошо функционирующей при низких и средних температурах. Сериновые протеазы для применения в моющих средствах должны обладать стабильностью в присутствии моющих средств и обладать совместимостью с моющими средствами. В частности, целью изобретения является получение сериновой протеазы, способной удалять белковые субстанции, включая загрязнения при стирке белья и мытье посуды, при более низких температурах по сравнению с существующими на рынке ферментными препаратами, за счет чего можно будет, например, осуществлять чистку более деликатных материалов и сохранять энергию. Следующей целью изобретения является получение молекулы нуклеиновой кислоты, кодирующей указанный фермент, рекомбинантного вектора, клетки-хозяина, продуцирующей указанный фермент, композиции, содержащей указанный фермент, процесса изготовления такой композиции, а также применений указанного фермента и композиций, содержащих указанный фермент.
Сериновая протеаза грибного происхождения согласно изобретению может быть получена в высокопродуктивных грибах-хозяевах, а процесс ее дальнейшей обработки, например, отделение от ферментативного бульона и мицелия, легко выполним.
Настоящее изобретение относится к ферменту сериновой протеазе грибов, обладающей активностью сериновой протеазы и содержащей аминокислотную последовательность, по меньшей мере на 66% идентичную аминокислотной последовательности, представленной в SEQ ID No:18. Предпочтительно сериновую протеазу получают из Malbranchea cinnamomea (Lib.) Oorschot de Hoog.
В контексте настоящего изобретения "получена из" обозначает не только сериновую протеазу, которая была выработана или могла быть выработана штаммом указанного организма, но также сериновую протеазу, кодируемую последовательностью ДНК, выделенной из такого штамма, и выработанную организмом-хозяином, трансформированным указанной последовательностью ДНК. Наконец, термин обозначает сериновую протеазу, кодируемую последовательностью ДНК синтетического происхождения и/или происходящей из кДНК, и обладающую отличительными признаками указанной сериновой протеазы.
Предпочтительно, изобретение относится к ферменту сериновой протеазе грибов, обладающей активностью сериновой протеазы и содержащей аминокислотную последовательность по меньшей мере на 66% идентичную аминокислотной последовательности зрелой протеазы Malbranchea ALKO4122, представленной в SEQ ID No:18, или аминокислотной последовательности зрелой протеазы Malbranchea ALKO4122, представленной в SEQ ID No:18.
Фермент сериновую протеазу грибов по изобретению получают из Malbranchea, предпочтительно из Malbranchea cinnamomea (Lib.) Oorschot de Hoog (синоним Malbranchea pulchella var. sulfurea (Miehe) Cooney & R. Emers). Согласно наиболее предпочтительному воплощению фермент сериновую протеазу по изобретению получают из штамма Malbranchea ALKO4122, депонированного под номером CBS 128533, или из штамма Malbranchea ALKO4178, депонированного под номером CBS 128564. Фермент протеаза Malbranchea ALKO4178 по существу идентична ферменту протеазе штамма Malbranchea ALKO4122.
Фермент сериновая протеаза грибов имеет молекулярный вес от 20 до 35 кДа. Температурный оптимум фермента находится в диапазоне от 30°С до 80°С при рН 8,5, предпочтительно составляет приблизительно 70°С. Оптимум рН фермента находится в диапазоне по меньшей мере от рН 6 до рН 10 при 50°С, предпочтительно составляет рН 10. Температурные и рН характеристики определяли при времени реакции 30 мин и использовании казеина в качестве субстрата.
Сериновая протеаза грибов по изобретению способна разлагать или удалять белковые загрязнения в присутствии моющих средств при температуре от 0°С до 90°С, предпочтительно при температуре от 5°С до 60°С, наиболее предпочтительно при температуре от 10°С до 40°С.
Фермент сериновая протеаза грибов по изобретению кодируется последовательностью выделенного полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, имеющим последовательность, включенную в плазмиду pALK3092, содержащую нуклеотидную последовательность SEQ ID No:11, депонированную в Е.coli RF8758 под регистрационным номером DSM 24426, или последовательность SEQ ID No:17, кодирующую зрелую протеазу ALKO4122. Альтернативно, фермент сериновая протеаза грибов по изобретению кодируется последовательностью выделенного полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, имеющим последовательность, включенную в плазмиду pALK3093, содержащую нуклеотидную последовательность SEQ ID No:12, депонированную в Е.coli RF8759 под регистрационным номером DSM 24427.
Указанный фермент кодируется последовательностью выделенного полинуклеотида, которая кодирует полипептид, содержащий аминокислотную последовательность зрелой протеазы Malbranchea ALKO4122, согласно SEQ ID No:18, или аминокислотную последовательность, по меньшей мере на 66% идентичную аминокислотной последовательности зрелой протеазы Malbranchea ALKO4122, согласно SEQ ID No:18. Предпочтительно, указанный фермент кодируется молекулой выделенной нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID No:17.
Полноразмерный фермент сериновая протеаза грибов по изобретению кодируется полинуклеотидной последовательностью, включенной в pALK3094, депонированной в Escherichia coli RF8791 под регистрационным номером DSM 24410.
Фермент сериновая протеаза грибов продуцируется при рекомбинантной экспрессии вектора, содержащего молекулу нуклеиновой кислоты, кодирующую сериновую протеазу грибов по изобретению, функционально связанную с регуляторными последовательностями, способными направлять экспрессию фермента сериновой протеазы в подходящем хозяине. Подходящие хозяева включают гетерологичных хозяев, предпочтительно микроорганизмов родов Trichoderma, Aspergillus, Fusarium, Humicola, Chrysosporium, Neurospora, Rhizopus, Penicillium, Myceliophthora и Mortiriella.
Предпочтительно, указанный фермент продуцируется представителями Trichoderma или Aspergillus, наиболее предпочтительно, Т. reesei.
Настоящее изобретение также относится к молекуле выделенной нуклеиновой кислоты, содержащей полинуклеотидную последовательность, кодирующую фермент сериновую протеазу, выбранной из группы, состоящей из:
(а) молекулы нуклеиновой кислоты, кодирующей полипептид, обладающий активностью сериновой протеазы и содержащий аминокислотную последовательность, представленную в SEQ ID No:18;
(б) молекулы нуклеиновой кислоты, кодирующей полипептид, обладающий активностью сериновой протеазы и по меньшей мере на 66% идентичный аминокислотной последовательности по SEQ ID No:18;
(в) молекулы нуклеиновой кислоты, содержащей кодирующую последовательность нуклеотидной последовательности, представленной в SEQ ID No:17;
(г) молекулы нуклеиновой кислоты, содержащей кодирующую последовательность полинуклеотидной последовательности, содержащейся в DSM 24410;
(д) молекулы нуклеиновой кислоты, кодирующая последовательность которой отличается от кодирующей последовательности любой молекулы нуклеиновой кислоты от (в) до (г) вследствие вырожденности генетического кода; и
(е) молекулы нуклеиновой кислоты, гибридизующейся в жестких условиях с молекулой нуклеиновой кислоты, содержащейся в DSM 24426 или представленной в SEQ ID No:17, кодирующей полипептид, имеющий активность сериновой протеазы и аминокислотную последовательность по меньшей мере на 66% идентичную аминокислотной последовательности, представленной в SEQ ID No:18.
Изобретение также относится к рекомбинантному вектору экспрессии, содержащему нуклеотидную последовательность по изобретению, функционально связанную с регуляторными последовательностями, способными направлять экспрессию гена указанной сериновой протеазы в подходящем хозяине. Подходящие хозяева включают гетерологичных хозяев, предпочтительно микроорганизмов родов Trichoderma, Aspergillus, Fusarium, Humicola, Chrysosporium, Neurospora, Rhizopus, Penicillium, Myceliophthora и Mortiriella. Предпочтительно, указанный фермент вырабатывается представителями Trichoderma или Aspergillus, наиболее предпочтительно Т. reesei.
Изобретение также касается клетки-хозяина, содержащей рекомбинантный вектор экспрессии, согласно описанию выше. Предпочтительно, клетка-хозяин представляет собой микроорганизм, такой как нитчатый гриб. Предпочтительные хозяева относятся к родам Trichoderma, Aspergillus, Fusarium, Humicola, Chrysosporium, Neurospora, Rhizopus, Penicillium, Myceliophthora и Mortiriella. Более предпочтительно хозяин является представителем Trichoderma или Aspergillus, наиболее предпочтительно - нитчатым грибом Т. reesei.
Настоящее изобретение относится к процессу получения полипептида, обладающего активностью сериновой протеазы, указанный процесс включает этапы культивирования клетки-хозяина по изобретению и выделения полипептида. Также в рамках изобретения находится полипептид, обладающий активностью сериновой протеазы, кодируемый последовательностью нуклеиновой кислоты по изобретению, который можно получить в ходе процесса, описанного выше.
Изобретение относится к процессу изготовления ферментного препарата, включающему этапы культивирования клетки-хозяина по изобретению и либо выделения полипептида из клеток, либо отделения клеток от культуральной среды и получения супернатанта. В рамках изобретения также находится ферментный препарат, получаемый в ходе процесса, описанного выше.
Изобретение относится к ферментному препарату, который содержит фермент сериновую протеазу по изобретению.
Изобретение также относится к композиции, содержащей фермент сериновую протеазу по изобретению.
Ферментный препарат или композиция (например, моющее средство), содержащие фермент протеазу по изобретению, могут также содержать другие ферменты, выбранные из группы, состоящей из протеаз (отличных от протеазы по изобретению), амилаз, целлюлаз, липаз, ксиланаз, маннаназ, кутиназ, пектиназ и оксидаз с добавлением или без добавления медиатора, а также подходящего дополнительного компонента, выбранного из группы, состоящей из стабилизаторов, буферов, сурфактантов, отбеливающих агентов, медиаторов, антикоррозийных агентов, структурообразователей, препятствующих повторному осаждению агентов, оптических отбеливателей, красителей, пигментов, отдушек, щелочей, абразивов, консервантов и т.д.
Кондиционированная культуральная среда хозяина-продуцента может использоваться как таковая или из нее могут быть удалены клетки-хозяева и/или она может быть сконцентрирована, отфильтрована или разделена на фракции. Кроме того, она может быть обезвожена. Ферментный препарат и композиция, содержащие фермент сериновую протеазу по изобретению, могут находиться в форме жидкости, порошка, гранулята или таблеток. Фермент в составе препарата или композиции может находиться в иммобилизованной форме.
Кроме того, в рамках изобретения находится применение фермента сериновой протеазы или ферментного препарата по изобретению в составе моющих средств, для обработки волокон, для обработки белкового материала, такого как шерсть, волосы, кожа, шелк, для обработки пищевых продуктов или кормов или для любого другого использования, включающего модификацию, разложение или удаление белкового материала. В частности, фермент или ферментный препарат можно применять в качестве моющей добавки в жидких моющих средствах, порошкообразных моющих средствах и таблетированных моющих средствах.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фигуре 1 (1А и 1Б) показана нуклеотидная последовательность гена протеазы Malbranchea ALKO4122, и части последовательностей его промотора (50 нуклеотидов до ATG) и терминатора (60 нуклеотидов после стоп-кодона), а также предсказанная аминокислотная последовательность кодируемой протеазы. Предполагаемый сигнальный пептид, проанализированный в программе SignalP V3.0, обозначен строчными буквами и подчеркнут. Пропоследовательность и предсказанные аминокислоты пропоследовательности указаны строчными буквами. Нуклеотидная и пептидная последовательности зрелого пептида указаны заглавными буквами. Последовательности трех предполагаемых интронов указаны строчными буквами, курсивом и выделены пунктирной линией под нуклеотидной последовательностью. Стоп-кодон указан звездочкой под последовательностью. Расположение праймеров DET27 (5'-смысловой праймер, s) и DET28 (3'-антисмысловой праймер, as), использовавшихся для клонирования гена протеазы Malbranchea ALKO4178 при помощи ПЦР, подчеркнуто двойной линией.
На Фигуре 1А показана нуклеотидная последовательность гена протеазы Malbranchea ALKO4122 и часть его промотора. Последовательность гена протеазы включает нуклеотиды от 51 до 960, участок последовательности, кодирующий аминокислотную последовательность протеазы от Met 1 до Val 279 и первый кодон Ala 280.
На Фигуре 1Б показана нуклеотидная последовательность гена протеазы Malbranchea ALKO4122 и часть его терминатора. Последовательность гена протеазы включает нуклеотиды от 961 до 1486 (включая стоп-кодон ТАА), участок последовательности, кодирующий аминокислотную последовательность протеазы от Ala 280 (два последних кодона) до Arg 401.
На Фигуре 2 схематично изображена карта плазмиды pALK2777, на основе которой была сконструирована экспрессионная кассета pALK3097. Ген протеазы Malbranchea лигировали между последовательностями промотора cbh1 (cel7A) (сшивание полностью совпадающих концов в сайте SacII) и терминатора (в сайте BamHI в линкере) с плазмидой pALK2777, расщепленной SacII - BamHI. Более подробное описание приведено в Примере 2. Плазмида pALK2777 включает синтетический маркерный ген amdS для скрининга трансформантов. pcbh1, промотор cbh1; tcbh1, терминатор cbh1; s_amdS, синтетический маркерный ген amdS (кДНК); линкер, линкерная последовательность, включающая, например, сайт рестрикции BamHI.
На Фигуре 3 схематично изображена кассета pALK3097, выделенная из каркаса вектора путем расщепления NotI и использованная для экспрессии гена протеазы Malbranchea ALKO4122 в Trichoderma reesei. pcbh1, промотор cbh1; tcbh1, терминатор cbh1; s_amdS, синтетический маркерный ген amdS (кДНК); линкер, линкерная последовательность.
На Фигуре 4 показаны результаты исследования частично очищенного рекомбинантного белка при денатурирующем электрофорезе в 12% полиакриламидном геле. Дорожка 1: образец частично очищенной протеазы Malbranchea. Дорожка 2: макер молекулярного веса (Page Ruler Unstained Protein Ladder, Fermentas).
На Фигуре 5 (5A и 5Б) показана относительная активность фермента при различных значениях температуры и рН.
На Фигуре 5A показан температурный профиль рекомбинантной протеазы Malbranchea и Савиназы® 16L, исследованный при рН 8,5 с использованием казеина в качестве субстрата и времени реакции 30 мин. Точки на графике представляют собой усредненные значения трех отдельных измерений.
На Фигуре 5Б показано влияние рН на активность рекомбинантной протеазы Malbrachea и Савиназы® 16L. В качестве буфера использовали 40 мМ буфер Бриттона-Робинсона, в качестве субстрата использовали казеин, время реакции составляло 30 мин, реакцию проводили при температуре 50°С. Точки на графике представляют собой усредненные значения трех отдельных измерений.
На Фиг.6 (6А, 6Б, 6В и 6Г) показана способность рекомбинантной протеазы Malbranchea ALKO4122 удалять загрязнения при окрашивании кровью/молоком/чернилами (Арт.117, хлопок + полиэфир, серийный номер 11-08, новая партия, ЕМРА) при 10-50°С, рН прибл. 8, за 60 мин, в присутствии коммерческого жидкого моющего средства в концентрации 5 г/л. Для сравнения использовали коммерческие препараты протеаз Савиназа® 16L и Савиназа® Ультра 16L.
На Фиг.6А показана способность удалять загрязнения при 10°С.
На Фиг.6Б показана способность удалять загрязнения при 20°С.
На Фиг.6В показана способность удалять загрязнения при 30°С.
На Фиг.6Г показана способность удалять загрязнения при 50°С.
На Фиг.7 (7А, 7Б, 7В и 7Г) показана способность рекомбинантной протеазы Malbranchea ALKO4122 удалять загрязнения при окрашивании кровью/молоком/чернилами (Арт.117, хлопок + полиэфир, серийный номер 10-07, старая партия, ЕМРА) при 10-50°С, рН прибл. 8, за 60 мин, в присутствии коммерческого жидкого моющего средства в концентрации 5 г/л. Для сравнения использовали Савиназу® Ультра 16L.
Фиг.7А показана способность удалять загрязнения при 10°С.
Фиг.7Б показана способность удалять загрязнения при 20°С.
Фиг.7В показана способность удалять загрязнения при 30°С.
Фиг.7Г показана способность удалять загрязнения при 50°С.
На Фиг.8 (8А, 8Б, 8В и 8Г) показана способность рекомбинантной протеазы Malbranchea ALKO4122 удалять загрязнения при окрашивании кровью/молоком/чернилами (Арт.117, хлопок+полиэфир, серийный номер 11-08, новая партия, ЕМРА) при 10-50°С, рН прибл. 8, за 60 мин, в присутствии коммерческого жидкого моющего средства в концентрации 5 г/л. Для сравнения использовали Савиназу® 16L и Савиназу® Ультра 16L.
На Фиг.8А показана способность удалять загрязнения при 10°С (количество фермента рассчитывали по белку).
На Фиг.8Б показана способность удалять загрязнения при 20°С (количество фермента рассчитывали по белку).
На Фиг.8В показана способность удалять загрязнения при 30°С (количество фермента рассчитывали по белку).
На Фиг.8Г показана способность удалять загрязнения при 50°С (количество фермента рассчитывали по белку).
На Фигуре 9 (9А, 9Б, 9В, 9Г, 9Д, 9Е, 9Ж и 9З) показано качество работы рекомбинантной протеазы Malbranchea ALKO4122 в отношении различных загрязнений при испытании с применением лаундерометра в присутствии коммерческого жидкого моющего средства в концентрации 5 г/л при 30 и 60°С, за 60 мин, рН прибл. 8. Для сравнения использовали коммерческий препарат Савиназа® Ультра 16L.
На Фигуре 9А показано качество работы в отношении загрязнений от травы на хлопке, Арт.164 (серийный номер 23-03, ЕМРА) при 30°С.
На Фигуре 9Б показано качество работы в отношении загрязнений от травы на хлопке, Арт.164 (серийный номер 23-03, ЕМРА) при 60°С.
На Фигуре 9 В показано качество работы в отношении загрязнений от крови/молока/чернил на хлопке, Арт.116 (серийный номер 18-16, ЕМРА) при 30°C.
На Фигуре 9Г показано качество работы в отношении загрязнений от крови/молока/чернил на хлопке, Арт.116 (серийный номер 18-16, ЕМРА) при 60°С.
На Фигуре 9Д показано качество работы в отношении загрязнений от крови/молока/чернил на хлопке с полиэфиром, Арт.117 (серийный номер 11-08, новая партия, ЕМРА) при 30°С.
На Фигуре 9Е показано качество работы в отношении загрязнений от крови/молока/чернил на хлопке с полиэфиром, Арт.117 (серийный номер 11-08, новая партия, ЕМРА) при 60°С.
На Фигуре 9Ж показано качество работы в отношении загрязнений от крови/молока/чернил на хлопке с полиэфиром, Арт.117 (серийный номер 10-07, старая партия, ЕМРА) при 30°С.
На Фигуре 9З показано качество работы в отношении загрязнений от крови/молока/чернил на хлопке с полиэфиром, Арт.117 (серийный номер 10-07, старая партия, ЕМРА) при 60°С.
На Фигуре 10 (10А и 10Б) показано качество работы протеазы в присутствии порошкообразного моющего средства.
На Фигуре 10А показано качество работы рекомбинантной протеазы Malbranchea ALKO4122 в отношении загрязнений от крови/молока/чернил на хлопке с полиэфиром, Арт.117 (серийный номер 11-08, ЕМРА) в присутствии 5 г/л обычного коммерческого порошкообразного моющего средства (описано в Примере 8) при 50°С, за 60 мин, рН прибл. 10,5. Для сравнения использовали Савиназу® Ультра 16L.
На Фигуре 10Б показано качество работы рекомбинантной протеазы Malbranchea ALKO4122 в отношении загрязнений от крови/молока/чернил на хлопке с полиэфиром, Арт.117 (серийный номер 11-08, ЕМРА) в присутствии 5 г/л порошкообразного моющего средства, Арт.601, ЕМРА, 50°С, за 60 мин, рН прибл. 10. Для сравнения использовали Савиназу® Ультра 16L.
На Фигуре 11 (11А и 11Б) показана стабильность протеазы при хранении при 37°С.
На Фигуре 11А показана стабильность рекомбинантной протеазы Malbranchea ALKO4122 при хранении при 37°С, при использовании в качестве консерванта/стабилизатора Proxel LV или пропиленгликоля (PG).
На Фигуре 11Б показана стабильность рекомбинантной протеазы Fe_RF6318 (WO2010125174A1) при хранении при 37°С, при использовании в качестве консерванта/стабилизатора Proxel LV или пропиленгликоля.
На Фигуре 12 (12А и 12Б) показана стабильность протеазы в моющем средстве.
На Фигуре 12А показана стабильность рекомбинантной протеазы Malbranchea ALKO4122 protease в стандартном моющем средстве Ecolabel при 37°С, рН прибл. 7. Для сравнения использовали коммерческий препарат Савиназа® Ультра 16L и рекомбинантную протеазу Fe_RF6318 (WO2010125174A1). Количество ферментного препарата в моющем средстве составляло 4% (вес/вес).
На Фиг.12Б показана стабильность протеазы Malbranchea ALKO4122 в коммерческом жидком моющем средстве при 37°С (рН прибл. 8). Для сравнения использовали коммерческий препарат Савиназа® Ультра 16L и рекомбинантную протеазу Fe_RF6318 (WO2010125174A1). Количество ферментного препарата в моющем средстве составляло 4% (вес/вес).
Перечень последовательностей
SEQ ID NO:1 Последовательность смыслового праймера DET1, использованного в ПЦР для синтеза зонда протеазы Malbranchea.
SEQ ID NO:2 Последовательность смыслового праймера DET2, использованного в ПЦР для синтеза зонда протеазы Malbranchea.
SEQ ID NO:3 Последовательность антисмыслового праймера DET3, использованного в ПЦР для синтеза зонда протеазы Malbranchea.
SEQ ID NO:4 Последовательность антисмыслового праймера DET4, использованного в ПЦР для синтеза зонда протеазы Malbranchea.
SEQ ID NO:5 Последовательность смыслового праймера DET5, использованного в ПЦР для синтеза зонда протеазы Malbranchea.
SEQ ID NO:6 Консенсусная пептидная последовательность, использованная в дизайне смыслового праймера DET1 для ПЦР.
SEQ ID NO:7 Консенсусная пептидная последовательность, использованная в дизайне смыслового праймера DET2 для ПЦР.
SEQ ID NO:8 Консенсусная пептидная последовательность, использованная в дизайне антисмыслового праймера DET3 для ПЦР.
SEQ ID NO:9 Консенсусная пептидная последовательность, использованная в дизайне антисмыслового праймера DET4 для ПЦР.
SEQ ID NO:10 Консенсусная пептидная последовательность, использованная в дизайне смыслового праймера DET5 для ПЦР.
SEQ ID NO:11 Последовательность фрагмента, полученного в ПЦР с использованием в ходе реакции DET5 и DET4 и геномной ДНК Malbranchea ALKO4122 в качестве матрицы. Данный фрагмент содержит часть гена протеазы Malbranchea и вставлен в плазмиду pALK3092.
SEQ ID NO:12 Последовательность фрагмента, полученного в ПЦР с использованием в ходе реакции DET5 и DET4 и геномной ДНК Malbranchea ALKO4178 в качестве матрицы. Данный фрагмент содержит часть гена протеазы Malbranchea и вставлен в плазмиду pALK3093.
SEQ ID NO:13 Нуклеотидная последовательность, кодирующая полную аминокислотную последовательность протеазы Malbranchea ALKO4122. Полноразмерный ген включен в плазмиду pALK3094. Последовательность гена протеазы, клонированная из Malbranchea ALKO4178 с помощью ПЦР, была идентична данной последовательности.
SEQ ID NO:14 Полная аминокислотная последовательность протеазы Malbranchea ALKO4122, включающая аминокислоты полноразмерной протеазы от Met1 до Arg 401.
SEQ ID NO:15 Нуклеотидная последовательность, кодирующая аминокислотную последовательность протеазы Malbranchea ALKO4122 в форме профермента.
SEQ ID NO:16 Аминокислотная последовательность протеазы Malbranchea ALKO4122 в форме профермента, включающая аминокислоты полноразмерной протеазы от Gly 21 до Arg 401.
SEQ ID NO:17 Нуклеотидная последовательность, кодирующая аминокислотную последовательность зрелой формы протеазы Malbranchea ALKO4122.
SEQ ID NO:18 Аминокислотная последовательность зрелой формы протеазы Malbranchea ALKO4122, включающая аминокислоты полноразмерного фермента от Ala 121 до Arg 401.
SEQ ID NO:19 Последовательность смыслового праймера DET27, использованного в ПЦР для клонирования гена протеазы Malbranchea ALKO4178.
SEQ ID NO:20 Последовательность антисмыслового праймера DET28, использованного для клонирования гена протеазы Malbranchea ALKO4178.
SEQ ID NO:21 Последовательность смыслового праймера DET17, использованного для конструирования экспрессионной кассеты в pALK3097.
SEQ ID NO:22 Последовательность антисмыслового праймера DET18, использованного для конструирования экспрессионной кассеты в pALK3097.
Депонирование
Malbranchea ALKO4122 депонировали в Центральном Бюро Культур Грибов (Centraalbureau Voor Schimmelcultures, Uppsalalaan 8, 3508 AD, Utrecht, the Netherlands), 20 декабря 2010 г. под регистрационным номером CBS 128533.
Malbranchea ALKO4178 депонировали в Центральном Бюро Культур Грибов (Centraalbureau Voor Schimmelcultures, Uppsalalaan 8, 3508 AD, Utrecht, the Netherlands), 5 января 2011 г. под регистрационным номером CBS 128564.
Штамм Е.coli RF8791, содержащий плазмиду pALK3094, депонировали в Коллекции Микроорганизмов и Клеточных Культур Германии (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ), Inhoffenstrasse 7 В, D-38124 Braunschweig, Germany) 20 декабря 2010 под регистрационным номером DSM 24410.
Штамм E.coli RF8758, содержащий плазмиду pALK3092, депонировали в Коллекции Микроорганизмов и Клеточных Культур Германии (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ), Inhoffenstrasse 7 В, D-38124 Braunschweig, Germany) 3 января 2010 г. под регистрационным номером DSM 24426.
Штамм Е.coli RF8759, содержащий плазмиду pALK3093, депонировали в Коллекции Микроорганизмов и Клеточных Культур Германии (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSMZ), Inhoffenstrasse 7 В, D-38124 Braunschweig, Germany) 3 января 2010 г. под регистрационным номером DSM 24427.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В настоящем изобретении предложен фермент сериновая протеаза грибов. Данная протеаза является активной в широком диапазоне рН и имеет широкий температурный оптимум при мытье, в частности демонстрирует хорошее качество работы в низком температурном диапазоне, а также при средних и высоких температурах. Фермент идеально подходит для применения в составе моющих средств, выдерживает стандартные детергентные композиции и является эффективным в растворах моющих средств при низких концентрациях фермента. В частности, протеаза является активной при температурах 0°С - 90°С, предпочтительно в диапазоне от 5°С до 60°С, более предпочтительно от 10 до 40°С. Протеаза по изобретению также обладает высокой стабильностью в жидких детергентных композициях. Таким образом, в настоящем изобретении предложена новая сериновая протеаза для применения в составе моющих средств и для другого использования, в частности, в жидких составах. Сериновая протеаза грибов может быть получена в высокопродуктивных хозяевах-грибах, а процесс ее дальнейшей обработки, например, отделение от ферментативного бульона и мицелия легко осуществим.
В частности, в настоящем изобретении предложен фермент сериновая протеаза, обладающая активностью сериновой протеазы и содержащая аминокислотную последовательность, по меньшей мере на 66% идентичную аминокислотной последовательности, представленной в SEQ ID No:18. Предпочтительно, в настоящем изобретении предложен фермент сериновая протеаза грибов, обладающая активностью сериновой протеазы и содержащая аминокислотную последовательность, представленную в SEQ ID No:18. Предпочтительно, сериновая протеаза получена из Malbranchea cinnamomea (Lib.) Oorschot de Hoog.
Под "сериновой протеазой" или "сериновой эндопептидазой" или "сериновой эндопротеиназой" в данном изобретении понимают фермент, относящийся к классу ЕС 3.4.21 по Номенклатуре международного общества биохимии и молекулярной биологии. Протеазы можно классифицировать на основании групповой специфичности ингибиторов. Многообразная группа ингибиторов сериновых протеаз включает синтетические химические ингибиторы и естественные белковые ингибиторы. Таким образом, активность сериновых протеаз можно определять в тестах, основанных на расщеплении специфического субстрата, или в тестах с использованием любого белок-содержащего субстрата в подходящих условиях, в присутствии или в отсутствие специфического ингибитора сериновых протеаз.
Под термином "активность сериновой протеазы" в данном изобретении понимают гидролитическую активность в отношении белок-содержащего субстрата, например, казеина, гемоглобина и БСА. Способы исследования протеолитической активности хорошо известны в литературе и приводятся, например, Gupta et al. (2002).
Сериновые протеазы синетзируются в виде неактивных предшественников, или зимогенов в форме препроферментов, которые активируются при удалении сигнальной последовательности (секреторного сигнального пептида или препептида) и пропоследовательности (пропептида) с образованием активной зрелой формы фермента (Chen and Inouye, 2008). Этот процесс активации включает действие протеаз и может происходить при ограниченном аутопротеолитическом или автокаталитическом процессинге сериновой протеазы, например, в ходе посттрансляционных этапов синтеза или в кондиционированной культуральной среде или в ходе хранения культуральной среды или ферментного препарата. Активация профермента может также происходить при добавлении в культуральную среду во время или после культивирования организма-хозяина протеолитического фермента, способного конвертировать неактивный профермент в активный зрелый фермент. Укорочение фермента также может происходить, например, при усечении гена, кодирующего полипептид, до проведения трансформации хозяина-продуцента. "Препро-форма" сериновой протеазы в данном изобретении означает фермент, содержащий пре- и пропептиды. "Про-форма" означает фермент, который содержит пропептид, но не имеет препептида (сигнальную последовательность).
Термин "зрелый" означает форму фермента сериновой протеазы, которая после удаления сигнальной последовательности (препептида) и пропептида содержит аминокислоты, необходимые для осуществления ферментативной или каталитической активности. У нитчатых грибов она представляет собой нативную форму, секретируемую в культуральную среду. Первая аминокислота зрелой последовательности может быть идентифицирована при N-концевом секвенировании секретируемой протеазы. В случае, когда биохимические данные получить невозможно, координаты N-конца можно ориентировочно определить путем выравнивания аминокислотной последовательности с аминокислотной(ыми) последовательностью(ями) зрелого(ых) гомологичного(ых) белка(ов). Выравнивание можно выполнять, например, с помощью программы ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/).
Самой большой группой имеющихся на рынке сериновых протеаз являются "щелочные сериновые протеазы", название которых означает, что ферменты активны и стабильны при рН от 9 до 11 или даже при рН от 10 до 12,5 (Shimogaki et al., 1991), а их изоэлектрическая точка находится около рН 9. Определение оптимального для каталитической активности рН можно выполнять в подходящем буферном растворе при различных значениях рН, измеряя активность в отношении белкового субстрата. Как правило, протеазы, обладающие моющим действием, лучше всего функционируют, когда значения рН моющего раствора, в котором они работают, приблизительно совпадают со значениями изоэлектрической точки фермента. Величину изоэлектрической точки можно определить при изоэлектрическом фокусировании с помощью гелей с фиксированным градиентом рН на основе полиакриламида, крахмала или агарозы или приближенно рассчитать значение изоэлектрической точки по аминокислотной последовательности, например, с помощью функции pl/MW сервера ExPASy (http://expasy.org/tools/pi_tool.html: Gasteiger et al., 2003).
Молекулярная масса зрелых щелочных сериновых протеаз находится в диапазоне от 15 до 35 кДа, как правило, приблизительно между 25 и 30 кДа (Rao et al. 1998). Молекулярную массу сериновой протеазы можно определить с помощью масс-спектрометрии или при помощи денатурирующего электрофореза в полиакриламидном геле согласно Laemmli (1970). Молекулярную массу также можно предсказать по аминокислотной последовательности фермента.
Температурный оптимум у большинства сериновых протеаз составляет приблизительно 60°С (Rao et al., 1998). Температурный оптимум сериновой протеазы можно определить в подходящем буферном растворе при различной температуре с использованием казеина в качестве субстрата, как описано в Примере 3, или с использованием других субстратов и буферных систем, описанных в литературе (Gupta et al., 2002).
Зрелая рекомбинантная сериновая протеаза Malbranchea согласно изобретению имеет молекулярный вес приблизительно 29 кДа, при использовании казеина в качестве субстрата оптимальная температура составляет приблизительно 70°С при рН 8,5 и времени реакции 30 мин, фермент, проявляет активность при щелочных значениях рН, например, рН 10 при 50°С и времени реакции 30 мин с использованием казеина в качестве субстрата. Рекомбинантная сериновая протеаза Malbranchea демонстрирует хорошее качество работы в присутствии моющих средств с различными свойствами в широком температурном диапазоне, т.е. при низких, средних и даже при высоких значениях температур. Рекомбинантная сериновая протеаза Malbranchea в частности может применяться при температуре 60°С или ниже, в зависимости от условий мытья и наличия в моющих средствах вспомогательных ингредиентов и добавок.
Для улучшения качества работы сериновой протеазы Malbranchea при различном промышленном использовании, например, в моющих средствах, желательно улучшить свойства нативного фермента. Эти свойства включают, например, стабильность при хранении, стабильность в присутствии или отсутствии моющего средства, стабильность в отношении рН, устойчивость к окислению или устойчивость к отбеливающим агентам, а также специфичность к субстрату. Аутопротеолитическая активность фермента влияет на стабильность при хранении и должна быть как можно более низкой. Очевидно, что, например, в композициях для стирки белья и мытья посуды моющая способность модифицированной протеазы не должна ухудшаться по сравнению с родительским ферментом или предшественником фермента протеазы. Другими словами, желательно, чтобы варианты фермента обладали аналогичной или даже улучшенной моющей способностью и способностью удалять загрязнения по сравнению с родительской сериновой протеазой.
Полученные ферменты протеаз, в частности, сериновые протеазы можно очищать при помощи стандартных способов обработки ферментов, таких как высаливание, ультрафильтрация, ионообменная хроматография, аффинная хроматография, гель-фильтрация и хроматография гидрофобного взаимодействия. Очистку можно контролировать путем определения белка, исследования активности фермента и денатурирующего электрофореза в полиакриламидном геле. Можно определять активность фермента и стабильность очищенного фермента при различных значениях температуры и рН, а также молекулярную массу и изоэлектрическую точку.
Очистка рекомбинантной сериновой протеазы по настоящему изобретению показана в Примере 4. После центрифугирования и фильтрации культуральный супернатант наносили на обессоливающую колонку HiPrep 26/10 (GE Healthcare), уравновешенную 20 мМ MES, рН 5,3. После гель-фильтрации образец наносили на колонку S Sepharose HP объемом 1 мл (GE Healthcare), уравновешенную 20 мМ MES, рН 5,3. Для элюирования белков использовали увеличивающийся градиент NaCI (0,5 М). Фракции, содержащие протеазу, объединяли и концентрировали с помощью центрифужных фильтров Amicon Ultra-4 10,000 СО производства MILLIPORE. Затем образец очищали с помощью гель-фильтрации на колонке Superdex 75, уравновешенной 20 мМ MES, 150 мМ NaCl, рН 5,3. Фракции, содержащие протеазу, объединяли. Полученный образец анализировали при помощи денатурирующего электрофореза в полиакриламидном геле, Фиг.4. Очевидно, что фермент по данному изобретению можно выделять с помощью других известных способов очистки, которые можно использовать вместо описанных здесь способов или в дополнение к ним. Рекомбинантную сериновую протеазу очищали, как описано в Примере 4, и использовали для исследования температурного и рН профиля, как описано в Примере 5.
Протеазная активность, как правило, основана на деградации растворимых субстратов. При использовании моющих средств протеазы должны использовать субстраты, которые по меньшей мере частично нерастворимы. Следовательно, важным свойством протеазы, обладающей моющим действием, является способность адсорбироваться на таких нерастворимых фрагментах и гидролизовать их.
Фермент сериновая протеаза по изобретению может быть получена из любых организмов, включая бактерии, архебактерии, грибы, дрожжи и даже высшие эукариоты, такие как растения. Предпочтительно, указанный фермент происходит из грибов, включая нитчатые грибы и дрожжи, например, родов, выбранных из группы, включающей Malbranchea. Щелочные протеазы грибов обладают преимуществом по сравнению с бактериальными протеазами в связи с простотой их последующей обработки для получения фермента или ферментной композиции, не содержащей микроорганизмов. Мицелий может быть легко удален при помощи фильтрации перед очисткой фермента.
Слабо выраженный запах продуктов ферментации грибов по настоящему изобретению является преимуществом по сравнению с продуктами, произведенными Bacillus, которые обычно обладают неприятным запахом. Следовательно, в готовой композиции потребуется меньше отдушек для маскирования запаха, и это делает продукт подходящим для использования, когда применение отдушек является нежелательным.
Настоящее изобретение относится к сериновой протеазе грибов, которая демонстрирует хорошее качество работы в присутствии моющих средств с различными свойствами, в широком диапазоне температур, включая низкие и средние, от 0°С до 90°С, предпочтительно при температурах, варьирующих от 5 до 60°С, и наиболее предпочтительно при температурах, варьирующих от 10 до 40°С.
В настоящем изобретении хорошее качество работы в присутствии моющего средства означает, что фермент, в данном случае рекомбинантная сериновая протеаза грибов по изобретению, работает в более низком диапазоне температур по сравнению со многими существующими на рынке субтилизинами. Другими словами, хорошее качество работы означает, что фермент способен разлагать или удалять белковые загрязнения или субстанции в диапазоне температур от низких до средних, в частности в более низком диапазоне температур по сравнению с существующими на рынке продуктами субтилизина, например с существующим на рынке продуктом фермента субтилизин Савиназой® или Савиназой® Ультра 16L (Novozymes A/S, DK).
Сериновая протеаза грибов по изобретению может применяться в частности при температуре 60°С или ниже, в зависимости от условий мытья и наличия в моющих средствах вспомогательных ингредиентов и добавок. Фермент также работает при 50°С или ниже, при 40°С или ниже, при 30°С или ниже, при 20°С или ниже и при 10°С или ниже. Наиболее удивительным является то, что термофильный фермент, у которого температурный оптимум составляет приблизительно 70°С, является эффективным и может применяться при температурах ниже 40°С, даже при температурах ниже 30°С.
В присутствии моющего средства сериновая протеаза грибов по изобретению работает при температурах, описанных выше, и в частности, указанная сериновая протеаза грибов демонстрирует хорошее качество работы в присутствии моющего средства при 40°С или ниже. Способность сериновой протеазы грибов из Malbranchea удалять загрязнения в различных экспериментальных условиях, в отношении различных загрязнений, измеряемая как ΔL*, значительно лучше, чем у существующих на рынке продуктов Савиназа® и Савиназа® Ультра 16L (Novozymes A/S, DK). Результаты представлены в Примерах 6-8 и на Фигурах 6-10.
Согласно предпочтительному воплощению изобретения рекомбинантный фермент сериновая протеаза грибов представляет собой полипептид, обладающий активностью сериновой протеазы и содержащий аминокислотную последовательность зрелой протеазы Malbranchea ALKO4122, согласно SEQ ID No:18, или аминокислотную последовательность по меньшей мере на 66% идентичную аминокислотной последовательности зрелой протеазы Malbranchea ALKO4122, согласно SEQ ID No:18. Предпочтительные ферменты идентичны по меньшей мере на 66%, предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, еще более предпочтительно по меньшей мере на 80%. Еще более предпочтительно аминокислотные последовательности идентичны по меньшей мере на 85% или по меньшей мере на 90% или 95%, более предпочтительно по меньшей мере на 98%, наиболее предпочтительно по меньшей мере на 99% аминокислотной последовательности по SEQ ID No:18. Идентичность ферментов можно выявить при сравнении соответствующих участков зрелых последовательностей.
Сериновая протеаза по настоящему изобретению может быть получена из Malbranchea, предпочтительно из Malbranchea cinnamomea (Lib.) Oorschot de Hoog (синоним Malbranchea pulchella var. sulfurea (Miehe) Cooney & R. Emers.), которая является представителем ЕС3.4.21. Согласно наиболее предпочтительному воплощению, фермент сериновая протеаза по изобретению может быть получена из штамма Malbranchea ALKO4122, депонированного под регистрационным номером CBS 128533, или из штамма Malbranchea ALKO4178, депонированного под регистрационным номером CBS 128564. Протеаза Malbranchea ALKO4178 по существу идентична протеазе штамма Malbranchea ALKO4122.
Под термином "идентичность" здесь понимают идентичность двух аминокислотных последовательностей, сравниваемых друг с другом в пределах соответствующего участка последовательности с приблизительно одинаковым количеством аминокислот. Например, можно определять идентичность полноразмерной или зрелой последовательности двух аминокислотных последовательностей. Аминокислотные последовательности двух сравниваемых молекул могут различаться в одной или двух позициях, что, тем не менее, не нарушает биологическую функцию или структуру молекул. Такие вариации могут возникать естественным образом в различных организмах-хозяевах или вследствие мутаций аминокислотной последовательности или они могут быть получены в результате направленного мутагенеза. Вариации могут быть результатом делеций, замен, инсерций, вставок в одной или более позициях аминокислотной последовательности или их сочетания. Идентичность последовательностей определяется при выравнивании с помощью программы ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/) с параметрами, заданными по умолчанию (матрица сравнения аминокислот: Gonnet, введение разрыва: 10, удлинение разрыва: 0,20, расстояние между разрывами: 5).
Одно из предпочтительных воплощений изобретения представляет собой фермент сериновую протеазу грибов, имеющую активность сериновой протеазы и аминокислотную последовательность протеазы Malbranchea ALKO4122 согласно SEQ ID No:18. Зрелый фермент не имеет сигнальной последовательности, или препептида, а также пропоследовательности, или пропептида. Зрелая сериновая протеаза по изобретению содержит аминокислоты от Ala121 до Arg401 полноразмерной протеазы, описанной в SEQ ID No:14. Таким образом, в рамках изобретения также находится полноразмерный фермент протеаза Malbranchea ALKO4122, имеющая последовательность SEQ ID No:14, включающую сигнальную последовательность (препептид) и пропептид, а также профермент, не имеющий сигнальной последовательности (препептида), таким образом, имеющий последовательность SEQ ID No:16.
Настоящее изобретение относится к ферменту сериновой протеазе грибов, зрелая форма которого имеет молекулярную массу или молекулярный вес от 20 до 35 кДа, предпочтительно от 25 до 33 кДа, более предпочтительно от 28 до 30 кДа. Наиболее предпочтительной является предсказанная молекулярная масса, составляющая 29 кДа для зрелого полипептида, полученная с помощью функции Compute pl/MW сервера ExPASy (Gasteiger et al., 2003).
Фермент по изобретению эффективно разлагает белковые вещества в широком диапазоне температур. Сериновая протеаза грибов имеет температурный оптимум в диапазоне от 30°С до 80°С (по меньшей мере приблизительно 10% от максимальной активности), предпочтительно от 40°С до 80°С (по меньшей мере приблизительно 20% от максимальной активности) и более предпочтительно между 50°С и 80°С (по меньшей мере приблизительно 40% от максимальной активности), наиболее предпочтительно от 60°С до 80°С (по меньшей мере приблизительно 65% от максимальной активности), максимальная активность определяется при 70°С, при измерении при рН 8,5 и времени реакции 30 мин с использованием казеина в качестве субстрата, как описано в Примере 5.
Фермент имеет оптимум рН в диапазоне от рН 6 по меньшей мере до рН 10 при 50°С, времени реакции 30 мин и использовании казеина в качестве субстрата, как описано в Примере 5. В частности, оптимум рН находится между рН 6 и рН 10 (по меньшей мере приблизительно 60% от максимальной активности), более предпочтительно между рН 9 и рН 10 (по меньшей мере приблизительно 70% от максимальной активности) и наиболее предпочтительно приблизительно составляет рН 10.
Сериновая протеаза, соответственно, сериновая протеаза грибов по изобретению, "демонстрирует хорошее качество работы в присутствии моющего средства", т.е. способна разлагать или удалять белковые загрязнения или субстанции в присутствии моющего средства в диапазоне низких температур, в частности, в более низком температурном диапазоне по сравнению с существующими на рынке продуктами субтилизина, например, с существующими на рынке ферментативными продуктами Савиназа® или Савиназа® Ультра 16L (Novozymes A/S, DK). В присутствии моющего средства фермент по изобретению хорошо работает между 5°С и 60°С, предпочтительно при 50°С или ниже. Фермент также работает при температурах 40°С или ниже, или 30°С или ниже.
Согласно предпочтительному воплощению изобретения фермент сериновая протеаза грибов кодируется последовательностью выделенного полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом или зондом, последовательность которого включена в плазмиду pALK3092, содержащую нуклеотидную последовательность SEQ ID No:11 в Е.coli RF8758, депонированную в Коллекции Микроорганизмов и Клеточных Культур Германии (Deutsche Sammlung von Mikroorganismen und Zellkulturen, DSMZ) под регистрационным номером DSM 24426.
Аналогично, фермент сериновая протеаза грибов (полученная из Malbranchea ALKO4178) по изобретению кодируется последовательностью выделенного полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, последовательность которого включена в плазмиду pALK3093, содержащим нуклеотидную последовательность SEQ ID No:12, депонированную в Е.coli RF8759 под регистрационным номером DSM 24427.
Кроме того, фермент сериновая протеаза по изобретению кодируется последовательностью выделенного полинуклеотида, который гибридизуется в жестких условиях с полинуклеотидом, последовательность которого включена в плазмиду pALK3094, содержащую нуклеотидную последовательность SEQ ID No:17, депонированную в Е.coli RF8791 под регистрационным номером DSM 24410.
В настоящем изобретении ген протеазы Malbranchea был выделен при помощи зонда, полученного с помощью ПЦР, с применением гибридизации в жестких условиях, согласно описанию в Примере 1г. Для выделения кДНК или геномной ДНК организма-хозяина можно применять стандартные методы молекулярной биологии, например, методы, описанные в руководствах по молекулярной биологии, таких как Sambrook and Russell, 2001.
Гибридизацию с зондом ДНК, например, таким который представлен в SEQ ID NO:11, SEQ ID NO:12 или SEQ ID NO:17, состоящим из более чем 100-200 нуклеотидов, обычно осуществляют в условиях "высокой жесткости", т.е. гибридизацию проводят при температуре, которая на 20-25°С ниже рассчитанной температуры плавления (Tm) совершенного гибрида, при этом Tm рассчитывают согласно Bolton and McCarthy (1962). Обычно прегибридизацию и гибридизацию проводят по меньшей мере при 65°С в 6× SSC (или 6× SSPE), 5× растворе Денхардта, 0,5% (вес/об) SDS, 100 мкг/мл денатурированной фрагментированной ДНК из молок лососевых. Добавление 50% формамида позволяет снизить температуру гибридизации до 42°С. Отмывки проводят при низкой концентрации соли, например, в 2× SSC-0,5% SDS (вес/об) в течение 15 мин при комнатной температуре, затем в 2× SSC-0,1% SDS (вес/об) при комнатной температуре и, наконец, в 0,1× SSC-0,1% SDS (вес/об) по меньшей мере при 65°С, или при условиях, описанных в Примере 1г.
Согласно одному предпочтительному воплощению фермент сериновая протеаза грибов по изобретению кодируется молекулой выделенной нуклеиновой кислоты, которая кодирует полипептид, имеющий аминокислотную последовательность, описанную в SEQ ID No:18, или полипептид, по меньшей мере на 66% идентичный аминокислотной последовательности SEQ ID No:18. Предпочтительные ферменты идентичны по меньшей мере на 66%, предпочтительно по меньшей мере на 70%, более предпочтительно по меньшей мере на 75%, еще более предпочтительно по меньшей мере на 80%. Еще более предпочтительно аминокислотные последовательности по меньшей мере на 85% или по меньшей мере на 90% или 95%, более предпочтительно по меньшей мере на 98%, наиболее предпочтительно на 99% идентичны аминокислотной последовательности по SEQ ID No:18. Идентичность ферментов выявляют при сравнении соответствующих участков зрелых последовательностей.
Таким образом, в рамках изобретения находится последовательность полипептида, который кодируется молекулой нуклеиновой кислоты, кодирующей аминокислотную последовательность полноразмерной сериновой протеазы по изобретению, включающей препептид (сигнальную последовательность) и пропептид в дополнение к зрелой форме фермента, данная аминокислотная последовательность описана в SEQ ID No:14.
Также, в рамках изобретения находится последовательность полипептида, который кодируется молекулой нуклеиновой кислоты, кодирующей фермент сериновую протеазу по изобретению в форме пропептида, включающую пропептид в дополнение к зрелой форме фермента, данная аминокислотная последовательность описана в SEQ ID No:16.
Одно из предпочтительных воплощений изобретения представляет собой фермент сериновую протеазу грибов, кодируемую молекулой выделенной нуклеиновой кислоты, которая содержит нуклеотидную последовательность, кодирующую зрелую форму сериновой протеазы Malbranchea ALKO4122, имеющую последовательность SEQ ID No:18.
Согласно одному предпочтительному воплощению фермент сериновая протеаза грибов по изобретению кодируется молекулой выделенной нуклеиновой кислоты, содержащей нуклеотидную последовательность по SEQ ID No:17, кодирующую зрелую форму фермента Malbranchea ALKO4122 (SEQ ID No:18).
Таким образом, в рамках изобретения находится полипептид, кодируемый молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность SEQ ID No-,13, содержащую "кодирующую последовательность" фермента. Выражение "кодирующая последовательность" означает нуклеотидную последовательность, которая начинается у кодона, инициирующего трансляцию (ATG), и заканчивается у кодона, терминирующего трансляцию (ТАА, TAG или TGA). Транслированный полноразмерный полипептид, как правило, начинается с метионина и может содержать интронные участки.
Также в рамках изобретения находится фермент сериновая протеаза грибов, кодируемая молекулой нуклеиновой кислоты, содержащей нуклеотидную последовательность SEQ ID NO:15, которая кодирует профермент Malbranchea ALKO4122.
Согласно другому предпочтительному воплощению изобретения сериновая протеаза грибов кодируется полинуклеотидной последовательностью, включенной в плазмиду pALK3094, содержащую нуклеотидную последовательность SEQ ID No:13 в Е.coli RF8791, депонированную под регистрационным номером DSM 24410.
Одно воплощение изобретения представляет собой фермент сериновую протеазу, полученную с помощью рекомбинантного вектора экспрессии, содержащего молекулу нуклеиновой кислоты, которая кодирует фермент сериновую протеазу грибов, согласно описанию выше, функционально связанную с регуляторными последовательностями, способными направлять экспрессию указанного фермента сериновой протеазы в подходящем хозяине. Конструкция указанного рекомбинантного вектора экспрессии и применение указанного вектора описано более подробно в Примере 2.
Подходящие хозяева для продукции фермента сериновой протеазы грибов являются гомологичными или гетерологичными хозяевами, такими как хозяева-микроорганизмы, включая бактерии, дрожжи и грибы. Нитчатые грибы, такие как Trichoderma, Aspergillus, Fusarium, Humicola, Chrysosporium Neurospora, Rhizopus, Penicillium, Myceliophthora и Mortiriella являются предпочтительными хозяевами-продуцентами вследствие простоты их последующей обработки и выделения ферментного продукта. Подходящие хозяева включают такие виды, как Т. reesei, A. niger, A oryzae, A. sojae, A. awamori или виды штаммов A. japonicus, F. venenatum или F. oxysporum, H. insolens или Н. lanuginosa, N. crassa и С. lucknowense, некоторые из которых перечислены в качестве организмов-хозяев, продуцирующих ферменты, например, среди существующих на рынке ферментов в перечне Ассоциации производителей ферментных продуктов (AMFEP) 2009 (http://www.amfep.org/list.html). Более предпочтительно, фермент продуцирует хозяин, являющийся нитчатым грибом рода Trichoderma или Aspergillus, таким как Т. reesei или A. niger, A. oryzae или A. awamori. Согласно наиболее предпочтительному воплощению изобретения фермент сериновая протеаза грибов вырабатывается Т. reesei.
Настоящее изобретение также относится к молекуле выделенной нуклеиновой кислоты, содержащей полинуклеотидную последовательность, кодирующую фермент сериновую протеазу, выбранной из группы, состоящей из:
(а) молекулы нуклеиновой кислоты, кодирующей полипептид, обладающий активностью сериновой протеазы и содержащий аминокислотную последовательность, представленную в SEQ ID No:18;
(б) молекулы нуклеиновой кислоты, кодирующей полипептид, обладающий активностью сериновой протеазы и по меньшей мере на 66% идентичный аминокислотной последовательности no SEQ ID No:18;
(в) молекулы нуклеиновой кислоты, содержащей кодирующую последовательность нуклеотидной последовательности, представленной в SEQ ID No:17;
(г) молекулы нуклеиновой кислоты, содержащей кодирующую последовательность полинуклеотидной последовательности, содержащейся в DSM 24410;
(д) молекулы нуклеиновой кислоты, кодирующая последовательность которой отличается от кодирующей последовательности любой молекулы нуклеиновой кислоты от (в) до (г) вследствие вырожденности генетического кода; и
(е) молекулы нуклеиновой кислоты, гибридизующейся в жестких условиях с молекулой нуклеиновой кислоты, содержащейся в DSM 24426, или SEQ ID No:17, кодирующей полипептид, имеющий активность сериновой протеазы и аминокислотную последовательность, по меньшей мере на 66% идентичную аминокислотной последовательности, представленной в SEQ ID No:18.
Молекула нуклеиновой кислоты по изобретению может представлять собой РНК или ДНК, где ДНК может состоять из геномной ДНК или кДНК.
Для выделения и ферментативной обработки полинуклеотидной последовательности, кодирующей сериновую протеазу грибов по изобретению, можно использовать стандартные методы молекулярной биологии, включая выделение геномной и плазмидной ДНК, расщепление ДНК для получения фрагментов ДНК, секвенирования, трансформации E.coli и т.д. Фундаментальные методы описаны в основных руководствах по молекулярной биологии, например, Sambrook and Russell, 2001.
Выделение гена протеазы Malbranchea, кодирующего полипептид Malbranchea ALKO4122, описано в Примере 1. Вкратце, для выделения гена протеазы Malbranchea ALKO4122 использовали фрагмент ПЦР, полученный с использованием в ПЦР вырожденных олигонуклеотидных праймеров (SEQ ID No:5 и SEQ ID No:4). Фрагмент генома, включающий ген протеазы, лигировали с вектором pBluescript II KS+. Полноразмерный ген протеазы Malbranchea включали в плазмиду pALK3094, депонированную в Е.coli в коллекции культур DSMZ под регистрационным номером DSM 24410. Предсказанная аминокислотная последовательность сериновой протеазы была выведена по последовательности ДНК.
Нуклеотидная последовательность протеазы Malbranchea ALKO4122 (SEQ ID No:13), части последовательностей ее промотора и терминатора, а также выведенная аминокислотная последовательность (SEQ ID No:14) представлены на Фигурах 1А-Б. Размер гена составляет 1436 пн (включая стоп-кодон). Были найдены три предполагаемых интрона размером 72, 87 и 71 пн. Предсказанная последовательность белка состоит из 401 аминокислоты, включая предсказанную сигнальную последовательность из 20 аминокислот (SignalP V3.0; Nielsen et al., 1997 и Nielsen and Krogh, 1998) и предсказанный пропептид от Gly21 до Asp120. Предсказанная молекулярная масса зрелого полипептида составляет 28,5 кДа, а предсказанная изоэлектрическая точка составляет 6,15. Эти расчеты были сделаны с помощью функции Compute pl/MW сервера ExPASy (Gasteiger et al., 2003). Выведенная аминокислотная последовательность содержит три возможных сайта N-гликозилирования (Asn134, Asn172 и Asn277), но согласно данным CBS Server NetNGlyc V1.0, вероятными сайтами являются только два, Asn134 и Asn277. Поиск гомологии с опубликованными последовательностями протеаз осуществляли с помощью версии 2.2.25 программы BLASTP, NCBI (National Center for Biotechnology Information) (Altschul et al., 1990). Показатели идентичности последовательности зрелой протеазы Malbranchea с соответствующими участками большинства гомологичных последовательностей были получены при выравнивании с помощью программы ClustalW2 (матрица: Gonnet, введение разрыва: 10, удлинение разрыва: 0,20, расстояние между разрывами: 5; доступна на сайте www.ebi.ac.uk/Tools/msa/clustalw2/). Результаты приведены в Таблице 2.
Наиболее высокие значения показателей идентичности, полученные для зрелой протеазы Malbranchea ALKO4122 по настоящему изобретению (SEQ ID NO:18) при поиске в программе BLASTP, составили: 66% для предполагаемой субтилизин-подобной протеазы Coccidioides posadasii (EER24932.1) и гипотетического белка Coccidioides immitis CIMG_09197 (XP_001239485.1), 65% для гипотетического белка Uncinocarpus reesii UREG_05170 (EEP80328.1), 64% для гипотетического белка Coccidioides immitis CIMB_01394 (XP_001247623.1), предполагаемой субтилизин-подобной протеазы Coccidioides posadasii (EER23662.1), гипотетического белка Uncinocarpus reesii (EEP81307.1) и щелочной протеиназы Arthroderma otae (EEQ28657.1). Максимальная идентичность с запатентованными последовательностями составила 55% для SEQ ID NO:2 из US 5962765 (МЕ30270.1; протеаза Metarhizium anisopliae) и SEQ ID:15 из WO 8807581 (AAA54276.1; протеаза Tritirachium album). Последовательность зрелой протеазы Malbranchea ALKO4122 (SEQ ID NO:18) выравнивали со зрелыми последовательностями указанных выше гомологичных последовательностей при помощи программы ClustalW2. Значения показателя идентичности (оцениваемые в %), полученные при выравнивании в ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/). составляли 63% - 65%.
Таким образом, в рамках изобретения находится последовательность выделенного полинуклеотида или молекулы выделенной нуклеиновой кислоты, которая кодирует фермент сериновую протеазу грибов или полипептид, содержащий аминокислотную последовательность зрелой формы фермента Malbranchea ALKO4122, описанную в SEQ ID No:18, 15, т.е. аминокислоты от Ala121 до Arg401 полноразмерной сериновой протеазы по SEQ ID No:14.
Молекула нуклеиновой кислоты предпочтительно представляет собой молекулу, содержащую кодирующую последовательность, представленную в SEQ ID No:17, которая кодирует зрелую форму фермента сериновой протеазы грибов по данному изобретению.
Молекула выделенной нуклеиновой кислоты может представлять собой молекулу, содержащую кодирующую последовательность полинуклеотидной последовательности, содержащейся в DSM 24410, DSM 24426 или DSM 24427. DSM 24426 несет нуклеотидную последовательность ПЦР фрагмента (SEQ ID No:11), использованного при клонировании полноразмерного гена протеазы Malbranchea ALKO4122. DSM 24427 несет нуклеотидную последовательность ПЦР фрагмента (SEQ ID No:12), полученного из Malbranchea ALKO4178. DSM 24410 несет нуклеотидную последовательность гена полноразмерной протеазы Malbranchea ALKO4122 (SEQ ID No:13).
Молекула нуклеиновой кислоты по изобретению также может быть аналогом нуклеотидной последовательности, охарактеризованной выше. "Вырожденность" означает, что аналоги нуклеотидной последовательности, различаются по одному нуклеотиду или кодону или более, но кодируют рекомбинантную протеазу по изобретению.
Молекула нуклеиновой кислоты может также представлять собой молекулу нуклеиновой кислоты, гибридизующуюся в жестких условиях с ПЦР зондом, содержащимся в плазмидах pALK3092 или pALK3093, депонированных в E.coli под регистрационными номерами DSM 24426 и DSM 24427, соответственно, или с последовательностью ДНК SEQ ID NO:17, кодирующей зрелый полипептид, имеющий активность и аминокислотную последовательность сериновой протеазы. Гибридизуемая ДНК может происходить из грибов, принадлежащих к видам Malbranchea или она может происходить из других видов грибов.
Таким образом, в рамках изобретения находится молекула выделенной нуклеиновой кислоты, содержащая нуклеотидную последовательность, представленную в SEQ ID No:17, и ее аналоги.
Настоящее изобретение также относится к рекомбинантному вектору экспрессии или рекомбинантной экспрессионной конструкции, которые можно использовать для умножения или экспрессии последовательности нуклеиновой кислоты, кодирующей выбранную сериновую протеазу в подходящем эукариотическом или прокариотическом хозяине. Рекомбинантный экспрессирующий вектор содержит последовательности ДНК или нуклеиновой кислоты, которые способствуют или направляют экспрессию и секрецию сериновой протеазы, кодируемой последовательностью, в подходящем хозяине, например, промоторы, энхансеры, терминаторы (включая сигналы, терминирующие транскрипцию и трансляцию), а также сигнальные последовательности, функционально связанные с полинуклеотидной последовательностью, кодирующей указанную сериновую протеазу. Экспрессионный вектор может также содержать маркерные гены для селекции трансформированных штаммов или селективный маркер может быть введен в хозяина в составе другой векторной конструкции в ходе ко-трансформации. Указанные регуляторные последовательности могут быть гомологичными или гетерологичными в отношении организма-продуцента или они могут происходить из организма, из которого был выделен ген, кодирующий сериновую протеазу.
Примерами промоторов для экспрессии сериновой протеазы по изобретению в хозяевах, представляющих собой нитчатые грибы, являются промоторы TAKA-амилазы A. oryzae, щелочной протеазы ALP и триозофосфат-изомеразы, липазы Rhizopus miehei, глюкоамилазы Aspergillus niger или A. awamori (glaA), трипсиноподобной протеазы Fusarium oxysporum, промотор целлобиогидролазы 1 Chrysosporium lucknowense, целлобиогидролазы I Trichoderma reesei (Cel7A) и т.д.
Например, для обеспечения экспрессии в дрожжах, могут использоваться промоторы энолазы S. cerevisiae (ENO-1), галактокиназы (GAL1), алкоголь-дегидрогеназы (ADH2) и 3-фосфоглицерат-киназы.
Примерами последовательностей промоторов для направления транскрипции сериновой протеазы по изобретению в хозяевах-бактериях, являются промотор lac оперона Escherichia coli, промотор dagA агаразы Streptomyces coelicolor, промотор гена альфа-амилазы В. licheniformis (amyL), промотор гена мальтогенной амилазы В. stearothermophilus (amyM), промоторы генов xylA и xylB В. sublitis и т.д.
Подходящие терминаторы включают терминаторы генов, перечисленных выше, или любые охарактеризованные терминирующие последовательности.
Подходящие маркеры трансформации или селективные маркеры включают маркеры, устраняющие дефект у хозяина, например, гены dal В. subtilis или B. licheniformis или amdS и niaD Aspergillus. Отбор может также основываться на использовании маркеров, придающих устойчивость к антибиотикам, например, устойчивость к ампициллину, канамицину, хлорамфениколу, тетрациклину, флеомицину или гигромицину.
Предпочтительной является внеклеточная экспрессия сериновой протеазы по изобретению. Таким образом, рекомбинантный вектор содержит последовательности, способствующие секреции выбранным хозяином. Сигнальная последовательность сериновой протеазы по изобретению или препоследовательность или препептид могут быть включены в рекомбинантный экспрессионный вектор или природная сигнальная последовательность может быть заменена на другую сигнальную последовательность, способную облегчать экспрессию выбранным хозяином. Таким образом, выбранная сигнальная последовательность может быть гомологичной или гетерологичной для экспрессирующего хозяина. Природный пропептид может также быть заменен на другой пропептид. Пропептид может быть гомологичным или гетерологичным для экспрессирующего хозяина.
Примерами подходящих сигнальных последовательностей являются последовательности грибов или дрожжей, например, сигнальные последовательности генов с хорошей экспрессией. Такие сигнальные последовательности хорошо известны из литературных источников.
Рекомбинантный вектор может также содержать последовательности, способствующие интеграции вектора в хромосомную ДНК хозяина для достижения стабильной экспрессии и/или облегчения направленной доставки в определенное положение генома хозяина.
Протеаза Malbranchea ALKO4122 по изобретению с собственной сигнальной последовательностью была экспрессирована с промотора Т. reesei cbh1 (cel7A), как описано в Примере 2. Экспрессионная конструкция, использованная для трансформации хозяина Т. reesei, также включала терминатор cbh1 и синтетический маркер amdS для отбора трансформантов из нетрансформированных клеток.
Настоящее изобретение также относится к клеткам-хозяевам, содержащим рекомбинантный вектор экспрессии, согласно описанию выше. Подходящие хозяева для продукции фермента сериновой протеазы грибов представляют собой гомологичные или гетерологичные хозяева, такие как хозяева-микроогранизмы, включая бактерии, дрожжи и грибы. Также возможны продуцирующие системы в клетках растений или млекопитающих.
Предпочтительными хозяевами-продуцентами являются нитчатые грибы, такие как Trichoderma, Aspergillus, Fusarium, Humicola, Chrysosporium, Neurospora, Rhizopus, Penicillium, Myceliophthora и Mortiriella, вследствие простоты их последующей обработки и выделения ферментного продукта. Подходящими хозяевами, представляющими собой систему экспрессии и продукции, являются, например, продуцирующая система, разработанная для хозяев-нитчатых грибов Trichoderma reesei (EP 244234) или продуцирующие системы Aspergillus, такие как А. oryzae или A. niger (WO 9708325, US 5,843,745, US 5,770,418), A. awamori, A. sojae и штаммы типа A. japonicus, или продуцирующая система, разработанная для Fusarium, такая как F. oxysporum (Malardier et al., 1989) или F. venenatum, и для Neurospora crassa, Rhizopus miehei, Mortiriella alpinis, H. lanuginosa или H. insolens или для Chrysosporium lucknowense (US 6,573,086). Подходящими продуцирующими системами, разработанными для дрожжей, являются системы, разработанные для Saccharomyces, Schizosaccharomyces или Pichia pastoris. Подходящими продуцирующими системами, разработанными для бактерий, являются продуцирующие системы, разработанные для Bacillus, например, для В. subtilis, В. licheniformis, В. amyloliquefaciens, для Е.coli или для актиномицетов Streptomyces. Предпочтительно, сериновая протеаза по изобретению вырабатывается хозяевами, представляющими собой нитчатые грибы родов Trichoderma или Aspergillus, такими как Т. reesei, или A. niger, A oryzae, A. sojae, A. awamori или штаммы типа А. japonicus. Согласно наиболее предпочтительному воплощению изобретения фермент сериновая протеаза грибов вырабатывается Т. reesei.
Настоящее изобретение также относится к процессу получения полипептида, обладающего активностью сериновой протеазы, указанный процесс включает этапы культивирования естественной или рекомбинантной клетки-хозяина, несущей рекомбинантный вектор для экспрессии сериновой протеазы по изобретению, в подходящих условиях и, возможно, выделение указанного фермента. Среда для продукции может представлять собой среду, подходящую для выращивания организма-хозяина и содержащую индукторы для эффективной экспрессии. Подходящие среды хорошо известны из литературы.
Изобретение относится к полипептиду, обладающему активностью сериновой протеазы, указанный полипептид кодируется молекулой нуклеиновой кислоты по изобретению и может быть получен при помощи описанного выше способа.
Изобретение также относится к процессу получения ферментного препарата, содержащего полипептид, который обладает активностью сериновой протеазы, указанный процесс включает этапы культивирования клетки-хозяина, несущей экспрессирующий вектор по изобретению, и либо выделения полипептида из клеток, либо отделения клеток от культуральной среды и получения супернатанта, обладающего активностью сериновой протеазы.
Настоящее изобретение также относится к ферментному препарату, который содержит фермент сериновую протеазу, охарактеризованную выше. Ферментный препарат или композиция обладают активностью сериновой протеазы и могут быть получены в ходе процесса по изобретению.
Изобретение охватывает ферментный препарат, а также композицию, содержащие сериновую протеазу по изобретению.
Ферментный препарат или композиция (например, моющее средство), содержащие фермент протеазу по изобретению, могут также содержать другие ферменты, выбранные из группы, состоящей из протеаз (отличных от протеазы по изобретению), амилаз, липаз, целлюлаз, кутиназ, пектиназ, маннаназ, ксиланаз и оксидаз, таких как лакказа или пероксидаза, с добавлением или без добавления медиатора. Предполагается, что эти ферменты улучшают результаты работы сериновых протеаз по изобретению, например, за счет удаления углеводов и масел или жиров, присутствующих в обрабатываемом материале. Указанные ферменты могут иметь природное происхождение или быть рекомбинантными ферментами, полученными в штаммах-хозяевах, или могут быть добавлены в культуральный супернатант после завершения процесса продукции.
Указанный ферментный препарат или композиция могут также содержать один или более подходящих дополнительных компонентов, выбранных из группы, состоящей из сурфактантов, или поверхностно-активных агентов, буферов, антикоррозийных агентов, стабилизаторов, отбеливающих агентов, медиаторов, структурообразователей, щелочей, абразивов и консервантов, оптических отбеливателей, препятствующих повторному осаждению агентов, красителей, пигментов, отдушек, и т.д.
Сурфактанты целесообразно использовать для эмульгирования жиров и увлажнения поверхностей. Сурфактант может быть неионным, включая полуполярные и/или анионные и/или катионные и/или цвиттер-ионные.
Буферы можно добавлять к ферментному препарату или композиции для изменения рН или влияния на качество работы или стабильность других ингредиентов.
Подходящие стабилизаторы включают полиолы, такие как пропиленгликоль или глицерол, сахара или сахароспирты, молочную кислоту, борную кислоту или производные борной кислоты, пептиды и т.д.
Отбеливающий агент применяют для окисления и деградации органических соединений. Примерами подходящих химических отбеливающих систем являются источники H2O2, такие как перборат или перкарбонат с добавлением или без добавления образующих надкислоту активаторов отбеливания, таких как тетраацетилэтилендиамин, или, как вариант, пероксикислоты, например, амидного, имидного или сульфонного типа. Химические окислители можно частично или полностью заменить на окислительные ферменты, такие как лакказы или пероксидазы. Многие лакказы не эффективны в отсутствие медиаторов.
Структурообразователи или комплексообразующие агенты включают такие вещества, как цеолит, дифосфат, трифосфат, карбонат, цитрат и т.д. Ферментный препарат может также содержать один или более полимеров, таких как карбоксиметилцеллюлоза, полиэтиленгликоль, поливиниловый спирт, поливинилпирролидон, и т.д. Также можно добавлять смягчители, щелочи, консерванты для предупреждения порчи других ингредиентов, абразивы и вещества, изменяющие ценообразование и вязкость.
Согласно одному предпочтительному воплощению изобретения указанный ферментный препарат или композиция, содержащие указанный фермент, находятся в форме жидкости, порошка, гранулята или таблеток. Согласно предпочтительному воплощению изобретения указанная композиция представляет собой моющее средство в форме жидкости, порошка, гранулята или таблеток. Кроме того, фермент в составе препарата или композиции может быть в форме иммобилизованного фермента.
Сериновая протеаза по изобретению может, как и другие протеазы, в частности, щелочные протеазы, применяться в производстве моющих средств, белков, в пивоваренной, мясной, фотографической, кожной, молочной и фармацевтической промышленности (Kalisz, 1988; Rao et al., 1998). Например, ее можно применять в качестве альтернативы химическим соединениям для превращения волокнистых белковых отходов (например, рогов, перьев, ногтей и волос) в полезную биомассу, белковый концентрат или аминокислоты (Anwar and Saleemuddin, 1998). Применение сериновой протеазы по настоящему изобретению может, как в случае других ферментов, оказаться успешным при улучшении качества кож и сокращении загрязнений окружающей среды и сбережения энергии, и, как в случае щелочных протеаз, оно может оказаться полезным при синтезе пептидов и разделении смесей D, L-аминокислот. Примером использования сериновых протеаз в фармацевтической промышленности является использование субтилизина в сочетании с антибиотиками широкого спектра для лечения ожогов и ран, таким образом, сериновая протеаза грибов по настоящему изобретению может также найти подобное применение и может так же, как щелочные протеазы применяться для удаления крови с хирургических инструментов и очистки контактных линз или зубных протезов. Подобно щелочной протеазе Conidiobolus coronatus, сериновая протеаза грибов по настоящему изобретению может применяться для замены трипсина при культивировании клеток животных. Протеазы по изобретению могут также применяться для очистки мембран и разрушения биопленок. В хлебопечении протеазы могут применяться, например, для разрушения клейковинного каркаса, а в других областях пищевой промышленности для гидролиза пищевых белков, например, белков молока. Они могут также применяться, например, для обработки дрожжей, переработке животного сырья (экстрагировании большего количества белка из костей животных), создании новых ароматизаторов, удаления горечи, изменения эмульгирующих свойств, образования биологически активных пептидов и снижения аллергенности белков. Субстраты включают белки животного, растительного и микробного происхождения.
Промышленность, производящая моющие средства, в частности, промышленность, производящая моющие средства для стирки, стала одним из основных потребителей протеаз, проявляющих активность в широком диапазоне рН (Anwar and Saleemuddin, 1998). Идеальная протеаза, обладающая моющим действием, должна обладать широкой субстратной специфичностью, чтобы способствовать удалению разнообразных загрязнений от пищевых продуктов, травы, крови и других выделений организма. Она должна быть активной при рН и ионной силе раствора моющего средства, температуре и рН стирки, обладать устойчивостью при механических манипуляциях, а также в присутствии хелатирующих и окисляющих агентов, добавляемых в моющие средства. В связи с необходимостью энергосбережения в настоящее время желательно применять протеазу при более низких температурах.
Настоящее изобретение также относится к применению фермента сериновой протеазы или ферментного препарата в моющих средствах, для обработки текстильных волокон, для обработки белковых материалов, таких как шерсть, волосы, шелк, кожа, для обработки пищевых продуктов или кормов, или для любых других применений, включающих модификацию, разложение или удаление белкового материала.
Одно из предпочтительных воплощений изобретения, таким образом, представляет собой применение фермента сериновой протеазы, согласно описанию выше, в качестве моющей добавки, подходящей для моющих средств, используемых при стирке, и композиций для мытья посуды, включая композиции для автоматизированной мойки посуды.
Выражение "моющее средство" используется для обозначения вещества или субстанции, предназначенных для содействия чистке или имеющих чистящие свойства. Термин "моющее действие" обозначает наличие или степень выраженности чистящих свойств. Степень выраженности чистящих свойств можно исследовать с помощью различных белковых или белок-содержащих субстратах или загрязнениях или смесях загрязнений, связанных с твердым водонерастворимым носителем, таким как текстильные волокна или стекло. Типичные белковые субстанции включают кровь, молоко, чернила, яйца, траву и соусы. Для проведения испытаний на рынке существует большое разнообразие загрязнителей, содержащих белок. Функция фермента, обладающего моющим действием, состоит в разложении и удалении белок-содержащих загрязнений. Результаты испытаний зависят от вида загрязнения, композиции моющего средства, а также природы и состояния текстиля, используемого при испытании моющей способности (Maurer, 2004).
Термин "низкая температура" в контексте настоящей заявки означает температуру в диапазоне от 10°С до 30°С, которая согласно Экспериментам не является оптимальной для качества работы многих из существующих в настоящее время ферментных препаратов, в частности, ферментных моющих препаратов. Под термином "средняя температура" понимают температурный диапазон от 30°С до 60°С.
Термин "подходящий для применения в диапазоне низких или средних температур" включает промышленное использование, при котором желательно, чтобы фермент эффективно работал в диапазоне низких или средних температур (от 10°С до 60°С). Такое использование включает применение в пищевой, кормовой, кожной промышленности, фармацевтике, диагностике, переработке отходов и извлечении серебра. В данном документе считается, что такое использование исключает применение фермента сериновой протеазы по изобретению в качестве средства биологической борьбы для биологической защиты от патогенных грибов растений и нематод.
В настоящем изобретении термин "стабильность моющего средства" означает, что фермент или вариант фермента по существу сохраняет свою активность в растворе моющего средства во время хранения и/или мытья. Таким образом, он является эффективным при разложении или удалении белковых загрязнений или субстанций в присутствии моющего средства, такого как стандартное моющее средство Ecolabel light duty (wfk Testgewebe GmbH) или коммерческое жидкое моющее средство, описанное в Таблице 3 Примера 6. Стабильность можно оценивать, определяя остаточную активность, например, через несколько дней инкубации (при 37°С) в моющем средстве. Остаточную протеазную активность можно определять согласно способу, описанному в Примере 3, или любому другому способу, описанному в литературе (Gupta et al. 2002).
Термин "эффективное количество" сериновой протеазы означает количество фермента протеазы, необходимое для достижения ферментативной активности в определенной детергентной композиции. Предпочтительно, детергентная композиция по изобретению содержит приблизительно от 0,0001% до приблизительно 10% по весу варианта протеазы по изобретению в детергентной композиции, более предпочтительно от 0,001% до приблизительно 1%, еще более предпочтительно от 0,001% до приблизительно 0,5%.
Обычно моющую способность протеазы определяют как "эффективность удаления загрязнений" или "степень удаления загрязнений" или "выраженность чистящих свойств", означающие видимое и измеряемое увеличение осветления или изменения окраски загрязненного материала, например, искусственно загрязненных кусков материи или испытуемой ткани. Величину осветления или изменения окраски можно измерить, например, измеряя окраску как коэффициент отражения при помощи спектрофотометра с использованием координат цветового пространства L*a*b*, согласно описанию в Примерах 6-8. Обесцвечивание или удаление белкового загрязнения, указывающее на качество работы протеазы (эффективность удаления загрязнений) рассчитывают, например, как ΔL*, что означает величину осветления L* ткани, обработанной ферментом, за вычетом величины осветления L* ткани, обработанной моющим раствором без фермента (холостая проба или контроль). Присутствие моющего средства может улучшить качество работы фермента при удалении загрязнений. Сериновая протеаза по настоящему изобретению разрушает различные виды белковых загрязнений в щелочных условиях в присутствии моющих средств различного состава (Примеры 6-8).
Как показано в Примере 6, сериновая протеаза по изобретению удаляет загрязнения от крови/молока/чернил при 10-50°С и особенно при 30°С или ниже в присутствии коммерческого жидкого моющего средства значительно лучше, чем коммерческие препараты протеаз Савиназа® 16L и Савиназа® Ультра 16L (Фигуры 6 и 7). Ферментные препараты дозировали по единицам активности. Аналогичный эффект наблюдался, когда дозировку рассчитывали по количеству добавленного белка (Фигура 8). Удивительным является тот факт, что в условиях эксперимента при использовании казеина в качестве субстрата протеаза Malbranchea ALKO4122 проявляет оптимальную способность удалять загрязнения в очень широком диапазоне температур, в частности при низких температурах, таких как 10-30°С, несмотря на высокие значения температурного оптимума (прибл. 70°С, Фиг.5А). Савиназа®, обладающая схожим температурным профилем в условиях эксперимента (Фиг.5А), несомненно уступает по качеству работы при низких температурах стирки. Результаты этих испытаний указывают, что протеаза по изобретению демонстрирует превосходное качество работы в присутствии жидких моющих средств в широком диапазоне температур и даже при очень низких температурах стирки.
Кроме различных загрязнений от крови/молока/чернил протеаза Malbranchea ALKO4122 эффективно удаляет такие загрязнения как загрязнения от травы и какао при испытании в составе жидкого моющего средства при 30 и 60°С. Обработку проводили на лаундерометре ATLAS LP-2, дозировку фермента рассчитывали по активности. Результаты (Фигуры 9А - 3) показывают, что протеаза Malbranchea эффективна в отношении нескольких загрязнений как при температуре 30, так и 60°С и проявляет лучшую способность удалять загрязнения по сравнению с Савиназой® Ультра 16L. Malbranchea ALKO4122 также была эффективна в отношении загрязнений от арахисового масла/молока и яичного желтка.
Качество работы протеазы Malbranchea ALKO4122 также исследовали с применением обычного коммерческого порошкообразного моющего средства, содержащего отбеливающие агенты и оптические отбеливатели и с применением стандартного моющего средства ЕСЕ 77, не содержащего оптических отбеливателей и отбеливающих агентов, при 50°С при рН 10,5 или 10, соответственно, согласно описанию в Примере 8. Исследовали способность фермента удалять загрязнения от крови/молока/чернил с материала полиэфир-хлопок. Как показано на Фигурах 10А и Б, протеаза по изобретению подходит также для порошкообразных моющих средств в сильно щелочных условиях и проявляет эффект, сравнимый с Савиназой® Ультра 16L, при дозировании ферментного препарата в единицах активности.
Кроме моющей способности, фермент по настоящему изобретению в достаточной степени сохраняет свою активность при хранении, даже при хранении в жидких моющих средствах, как показано в Примерах 9 и 10. Протеаза Malbranchea ALKO4122 обладает превосходной стабильностью при хранении при 37°С по сравнению, например, с протеазой Fusarium Fe_RF6318 (WO2010125174A1) (Фигуры 11А и Б). На Фигурах 12А и 12Б показано, что протеаза Malbranchea обладает особенно высокой стабильностью в стандартном моющем средстве Ecolabel light duty (wfk Testgewebe GmbH) и коммерческом жидком моющем средстве (Таблица 3) при сравнении с протеазой Fusarium и обладает схожей стабильностью с существующей на рынке бактериальной протеазой Савиназа® 16L в моющем средстве Ecolabel. Хорошая стабильность протеазы согласно изобретению при повышенных температурах делает ее наиболее подходящей для использования в составе жидких моющих средств в странах с теплым климатом.
Согласно предпочтительному воплощению изобретения сериновая протеаза грибов по изобретению может найти применение в жидких моющих средствах и порошкообразных моющих средствах, как показано в Примерах 6-10. Фермент или ферментный препарат по изобретению могут быть изготовлены для применения при ручной и машинной стирке или могут быть изготовлены для применения при чистке твердых поверхностей в домашних условиях или предпочтительно при ручной или автоматизированной мойке посуды.
В заключение можно отметить, что предложен новый фермент, термостабильная сериновая протеаза грибов, происходящая из термофильных микроорганизмов, которая особенно хорошо функционирует при низких температурах и является совместимой и стабильной в жидких детергентных композициях, вследствие чего требуется меньше стабилизаторов и других дополнительных компонентов.
Для иллюстрации изобретения приведены следующие примеры, относящиеся к некоторым воплощениям изобретения, однако изобретение не исчерпывается этими примерами.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Пример 1. Синтез зондов для клонирования генов протеазы Malbranchea и клонирования гена протеазы штаммов Malbranchea ALKO4122 и ALKO4178
(а) Выделение ДНК и использованные методы молекулярной биологии
Для выделения и ферментативной обработки ДНК использовали стандартные методы молекулярной биологии (например, выделение плазмидной ДНК, расщепление ДНК для получения фрагментов ДНК) при трансформации Е.coli, секвенировании и т.д. Были использованы стандартные методы, либо в соответствии с описанием производителей ферментов, реагентов и наборов, либо по описанию в стандартных пособиях по молекулярной биологии, например, Sambrook and Russell (2001). Выделение геномной ДНК из нитчатых грибов производили в соответствии с подробным описанием Raeder and Broda (1985).
(б) Праймеры для создания зонда
Зонды для клонирования генов, кодирующих протеазы Malbranchea, синтезировали при помощи ПЦР. Вырожденные смысловой и антисмысловой олигонуклеотиды сконструировали, руководствуясь консенсусными аминокислотными последовательностями протеаз грибов (например, Fusarium equiseti RF6318, Fusarium acuminatum RF7182, Т. reesei Prb1 по WO2010125174, WO2010125175 и WO2011003968, AB Enzymes Oy, соответственно). Дополнительный 5'-праймер (DET5) синтезировали в соответствии с выбранным участком опубликованной N-концевой последовательности Термомиколина Malbranchea cinnamomea (всего 34 аминокислоты; регистрационный номер Swiss-Prot P13858.1). Для создания праймера DET5 использовали аминокислоты 20-28 вышеупомянутой последовательности. Последовательности праймеров и пептидные последовательности, использованные для их создания, приведены в Таблице 1 (SEQ ID NOs:1-5 и SEQ ID NOs:6-10, соответственно). Наименования олигонуклеотидов, SEQ ID NO, длина, вырожденность и нуклеотидная последовательность также приведены в таблице наряду с пептидными последовательностями, использованными при дизайне праймеров, и их SEQ ID NOs:6-10.
(в) ПЦР реакции и выбор зондов для «локирования полноразмерных генов протеазы
Препараты геномной ДНК, выделенной из Malbranchea ALKO4122 и ALKO4178, использовали в качестве матриц в реакциях ПЦР. Malbranchea ALKO4122 была идентифицирована службой идентификации Центрального Бюро Культур Грибов CBS (Utrecht, The Netherlands) как Malbranchea cinnamomea (Lib.) Oorschot de Hoog (синоним Malbranchea pulchella var. sulfurea (Miehe) Cooney & R. Emers.) в январе 2011. Malbranchea ALKO4178 депонировали в коллекции культур Roal Oy как Malbranchea sulfurea, но дальнейшей идентификации этого изолята не проводилось ни CBS, ни другим институтом, специализирующимся на идентификации нитчатых грибов.
Для выделения геномной ДНК мицелий Malbranchea ALKO4122 и ALKO4178 выращивали путем культивирования штаммов при 37°С в течение 4 дней при 250 об/мин в 50 мл глюкозо-дрожжевой среды (25 г глюкозы, 27,5 г дрожжевого экстракта, рН 6,5). Мицелий собирали, выделяли геномную ДНК и использовали в качестве матрицы для синтеза зонда. Реакционные смеси для ПЦР содержали буфер Phusion® GC (Finnzymes/Fisher Scientific, Finland), 0,2 мМ дНТФ, 50 пмоль каждого праймера, DMSO (1/10 объема), 1 ед полимеразы Phusion® (Finnzymes/Fisher Scientific) и приблизительно 0,5-1 мкг геномной ДНК на 50 мкл объема реакционной смеси. Условия для проведения ПЦР реакции были следующими: 1 мин первоначальной денатурации при 98°С, за которой следовало 28 циклов по 10 сек при 98°С, отжиг при 45, 47, 52 и 57°С в течение 30 сек, элонгация при 72°С в течение 30 сек и конечная элонгация при 72°С в течение 5 мин. Комбинация праймеров DET5 (SEQ ID NO:5) и DET4 (SEQ ID NO:4) позволила получить при трех наиболее низких температурах отжига (см. выше) ДНК-продукт, имеющий размер приблизительно 0,8 кб. Этот размер соответствовал ожидаемому размеру гена протеазы, согласно расчетам, базирующимся на опубликованных последовательностях протеаз грибов. При использовании в качестве матрицы геномной ДНК ALKO4122 или ALKO4178 был получен продукт схожего размера. ДНК-продукты выделяли и очищали от компонентов реакционной смеси ПЦР и лигировали с вектором pCR®4 Blunt-TOPO согласно инструкциям производителя (Invitrogen, USA). Полученные плазмиды были обозначены pALK3092 (с ПЦР продуктом ALKO4122 в качестве вставки) и pALK3093 (с ПЦР продуктом ALKO4178 в качестве вставки). Вставки секвенировали из pALK3092 (SEQ ID NO:11) и pALK3093 (SEQ ID NO:12). Обе вставки имели длину 791 пн и отличались между собой только на один нуклеотид в синтезированном участке (не принимая во внимание последовательности праймеров). Нуклеотид 323 во вставке pALK3092 представлял собой цитозин, а во вставке pALK3093 он представлял собой тимин. Штаммы E.coli RF8758 и RF8759, содержащие плазмиды pALK3092 и pALK3093, соответственно, были депонированы в коллекции DSMZ под регистрационными номерами DSM 24426 и DSM 24427, соответственно.
Поиск в программе BLASTP с использованием полученных последовательностей выполняли с помощью версии программы 2.2.23 на сайте NCBI (National Center for Biotechnology Information) с параметрами, заданными по умолчанию (Altschul et al., 1990). Последовательности кодируемых аминокислот демонстрировали гомологию, например, с гипотетическими и консервативными гипотетическими белками Uncinocarpus reesii (XP_002584481.1 и ХР_002583205.1, соответственно) и с предполагаемым субтилизин-подобным белком Coccidioides posadasii (EER24932.1), а также с гипотетическим белком Coccidioides immitis (ХР_001239485.1) с показателем идентичности 59-65%. Таким образом, результаты указывают, что фрагменты ДНК, полученные в ходе реакций ПЦР, представляли собой части генов, кодирующих протеазы, и таким образом, могли использоваться в качестве зондов для скрининга полноразмерного(ых) гена(ов) протеазы gene(s) Malbranchea. Однако, кодируемая аминокислотная последовательность, следующая за последовательностью, кодируемой праймером DET5, не была идентична опубликованной последовательности термомиколина Malbranchea (аминокислоты 29-34, регистрационный номер Р13858).
(г) Клонирование полноразмерного гена протеазы Malbranchea ALKO4122
Геномную ДНК Malbranchea ALKO4122 и ALKO4178 расщепляли с помощью нескольких рестрикционных ферментов для анализа методом Саузерн-блоттинга. Продукты расщепления разделяли на геле и выполняли Саузерн-перенос и гибридизацию. Зонд для Саузерн-блоттинга был помечен в ходе ПЦР с использованием прямого и обратного праймеров М13 и матрицы pALK3092 (Пример 1в). Последовательность зонда включала SEQ ID NO:11 (Пример 1в). Мечение зонда с помощью раствора для мечения PCR DIG (дигоксигенин) и гибридизацию выполняли согласно инструкциям производителя (Roche, Germany). Гибридизацию проводили в течение ночи при 68°С. После гибридизации фильтры отмывали следующим образом: 2 отмывки по 5 мин при комнатной температуре с помощью 2×SSC-0,1% SDS, с последующими 2 отмывками по 15 мин при 68°C с помощью 0,1×SSC-0,1% SDS.
Среди продуктов расщепления геномной ДНК Malbranchea ALKO4122 и ALKO4178 было обнаружено несколько гибридизующихся фрагментов. Для большинства продуктов расщепления из обоих геномов были получены идентичные результаты. Гибридизующийся фрагмент размером приблизительно 2,7 кб был получен при расщеплении геномной ДНК рестриктазой XbaI. По результатам анализа фрагмент, полученный с помощью XbaI, содержал полноразмерный ген протеазы (по результатам двойного расщепления геномной ДНК по сайтам рестрикции XbaI и PstI, содержащимся в последовательности зонда, и расчетах размера, при которых руководствовались опубликованными последовательностями протеаз грибов). Фрагменты ДНК выделяли из продукта расщепления геномной ДНК ALKO4122 с помощью XbaI, имеющего размер гибридизующегося фрагмента (приблизительно 2,7 кб). Выделение осуществляли из агарозного геля. Выделенные фрагменты геномной ДНК клонировали в вектор pBluescript II KS+ (Stratagene, USA), расщепленный XbaI. Лигированную смесь использовали для трансформации клеток Escherichia coli XL10-Gold (Stratagene), которые затем высевали на чашки со средой LB (Луриа-Бертани), содержащей 50 мкг/мл ампициллина. Проводили скрининг положительных колоний E.coli с помощью гибридизации колоний с использованием в качестве зонда вставки pALK3092, помеченного при помощи ПЦР. Гибридизацию проводили так же, как было описано для продуктов расщепления геномной ДНК. Всего на чашках было выбрано 20 клонов, 8 из которых по результатам расщепления рестриктазой XbaI и гибридизации с использованием Саузерн-блоттинга (которую проводили в соответствии с описанием гибридизации геномной ДНК) содержали вставки, имеющие ожидаемый размер и гибридизующиеся с зондом pALK3092. Вставку, полученную при расщеплении XbaI (2961 пн) секвенировали и подтвердили, что она содержала ген полноразмерной протеазы Malbranchea ALKO4122 (SEQ ID NO:13). Плазмида, содержащая указанный выше фрагмент геномной ДНК, полученный при расщеплении XbaI, была обозначена pALK3094. Штамм E.coli RF8791, содержащий плазмиду pALK3094, депонировали в коллекции DSMZ под регистрационным номером DSM 24410.
(д) Характеризация гена, кодирующего протеазу Malbranchea ALKO4122, и предсказанная аминокислотная последовательность протеазы
Последовательность гена (SEQ ID NO:13), части последовательностей его промотора и терминатора, а также выведенная аминокислотная последовательность кодируемой протеазы (SEQ ID NO:14) показаны на Фигуре 1. Длина гена составляет 1436 пн (включая стоп-кодон). Было обнаружено три предполагаемых интрона размером 72, 87 и 71 пн. Интроны имели консенсусные 5' и 3' концевые последовательности, а также внутренние консенсусные последовательности, соответствующие тем, что были обнаружены у нескольких интронов грибов (Gurr et al., 1987). Выведенная последовательность белка (SEQ ID NO:14) состояла из 401 аминокислоты, включая предсказанную сигнальную последовательность из 20 аминокислот (SignalP V3.0; Nielsen et al., 1997 and Nielsen and Krogh, 1998) и предсказанную пропоследовательность из 100 аминокислот. Координаты N-конца зрелой протеазы, Ala 121, определили при сравнении с другими последовательностями протеаз грибов. Предсказанная молекулярная масса зрелой протеазы составляла 28530 Да, а предсказанная величина изоэлектрической точки составляла 6,15. Эти расчеты были сделаны с помощью функции Compute pl/MW сервера ExPASy (Gasteiger et al., 2003). Выведенная зрелая аминокислотная последовательность содержала три возможных сайта N-гликозилирования в положениях Asn134, Asn172 и Asn277 зрелой последовательности (аминокислоты в положениях 254, 292 и 397 на Фигуре 1), но по данным CBS Server NetNGlyc V1.0 возможные сайты находились только в положениях Asn134 и Asn277.
N-концевая последовательность протеазы Malbranchea ALKO4122 только частично соответствовала ранее опубликованной N-концевой последовательности термомиколина М. cinnamonea (P13858): только первые 28 аминокислот в этих двух последовательностях были идентичны.
Последовательность зонда гена протеазы Malbranchea ALKO4122 в pALK3092 и геномная последовательность гена протеазы ALKO4122 различались на один нуклеотид (в соответствующем участке). При этом геномная последовательность гена протеазы ALKO4122 была идентична последовательности зонда протеазы ALKO4178 в pALK3093 (в соответствующем участке). Таким образом, различие по одному нуклеотиду во вставке в pALK3092 (Пример 1в) вероятнее всего было следствием мутации в ходе реакции ПЦР.
(е) Клонирование гена протеазы Malbranchea ALKO4178
Праймеры DET27 (SEQ ID NO:19) и DET28 (SEQ ID NO:20) были сконструированы для синтеза полноразмерного гена протеазы Malbranchea ALKO4178 с помощью ПЦР. DET 27 представлял собой смысловой праймер, комплементарный области промотора (нуклеотиды от -44 до -24 относительно АТС), а DET28 представлял собой антисмысловой праймер, комплементарный области терминатора (нуклеотиды от 55 до 36 относительно стоп-кодона) гена протеазы Malbranchea ALKO4122 (Фиг.1). Реакционные смеси для ПЦР содержали буфер 1XPhusion® HF (Finnzymes/Fisher Scientific, Finland), 0,2 мМ дНТФ, 75 пмоль каждого праймера, 2 ед полимеразы Phusion® (Finnzymes/Fisher Scientific) и приблизительно 1,5 мкг геномной ДНК на 200 мкл объема реакционной смеси. Условия для проведения реакции ПЦР были следующими: 30 сек первоначальной денатурации при 98°С, за которой следовало 24 цикла по 10 сек при 98°С, отжиг при 60 и 65°С в течение 30 сек, элонгация при 72°С в течение 60 сек и конечная элонгация при 72°С в течение 7 мин. В ходе реакций были получены фрагменты ожидаемой длины (~1,5 кб). Были выделены фрагменты, полученные в ходе двух отдельных реакций ПЦР, которые лигировали в векторы pCR4®Blunt-TOPO®. Вставки секвенировали из отдельных клонов. Последовательности вставок у обоих клонов были идентичны друг с другом и с геном протеазы Malbranchea ALKO4122, участками его промотора и терминатора (Фиг.1). Плазмида, содержащая ген протеазы Malbranchea ALKO4178 (фрагмент получен с помощью ПЦР), была обозначена pALK3147. Клон Е.coli, содержащий эту плазмиду, был депонирован в коллекции культур Roal Oy как RF9332.
(ж) Исследование гомологии, идентичности и выравнивания
Аминокислотную последовательность зрелой протеазы Malbranchea ALKO4122 (SEQ ID NO:18) использовали для поиска гомологичных последовательностей протеаз в общедоступных источниках. Поиск проводили как в базе повторяющихся последовательностей белков, так и в базе запатентованных последовательностей белков GenBank. При поиске использовали версию 2.2.25 программы BLASTP NCBI (National Center for Biotechnology Information) с настройками, заданными по умолчанию (Altschul et al., 1990). Максимальная идентичность, полученная при анализе всех повторяющихся последовательностей, составила 66% для предполагаемой субтилизин-подобной протеазы Coccidioides posadasii (EER24932.1) и гипотетического белка Coccidioides immitis CIMG_09197 (ХР_001239485.1), 65% для гипотетического белка Uncinocarpus reesii UREG_05170 (ЕЕР80328.1), 64% для гипотетического белка Coccidioides immitis CIMB_01394 (ХР_001247623.1), предполагаемой субтилизин-подобной протеазы Coccidioides posadasii (EER23662.1), гипотетического белка Uncinocarpus reesii (EEP81307.1) и щелочной протеиназы Arthroderma otae (EEQ28657.1). EER24932.1 и ХР_001239485.1 различались между собой только по трем аминокислотам, а ХР_001247623.1 и EER23662.1 различались между собой по одной аминокислоте. Последовательность протеазы Malbranchea ALKO4122 выравнивали с упомянутыми выше гомологичными последовательностями при помощи программы ClustalW2. Для выравнивания использовали предполагаемые зрелые последовательности каждой протеазы. Зрелые последовательности каждой протеазы сравнивали друг с другом. Значения показателя идентичности (оцениваемые в %), полученные при выравнивании с помощью программы ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/), приведены в Таблице 2. Зрелые аминокислотные последовательности, не включающие сигнальных пептидов и пропептидов, выравнивали с использованием параметров, заданных по умолчанию (матрица сравнения аминокислот: Gonnet, введение разрыва: 10, удлинение разрыва: 0,20, расстояние между разрывами: 5). SEQ ID NO:18 представляет собой зрелую аминокислотную последовательность протеазы ALKO4122. Полученные значения показателя идентичности (оцениваемые в %) составляли 63% - 65%.
Максимальная идентичность последовательностей в базе запатентованных последовательностей составила 55% для SEQ ID NO:2 из US 5962765 (ААЕ30270.1; протеаза Metarhizium anisopliae) и SEQ ID:15 из WO 8807581 (AAA54276.1; протеаза Tritirachium album).
Пример 2. Продукция рекомбинантной протеазы Malbranchea ALKO4122 в Trichoderma reesei
(а) Подготовка вектора для продукции и продуцирующих штаммов
Экспрессионная плазмида pALK3097 была сконструирована для продукции рекомбинантной протеазы Malbranchea ALKO4122 в Trichoderma reesei. Ген с собственной сигнальной последовательностью был присоединен к промотору Т. reesei cbh1 (cel7A) при помощи ПЦР благодаря полной комплементарности. Праймеры, использованные в реакции ПЦР, были обозначены DET17 (5'-праймер; SEQ ID NO:21) и DET18 (3'-праймер; SEQ ID NO:22). Праймер DET17 содержал часть промотора cbh1, от сайта рестрикции SacII (в положении -16 от ATG) до положения -1 и начало гена протеазы Malbranchea (26 нуклеотидов, включая стартовый кодон ATG), а также 3 дополнительных нуклеотида на 5'-конце. DET18 содержал 26 нуклеотидов конца гена протеазы Malbranchea (включая STOP кодон) и линкер, содержащий сайт рестрикции BamHI для сшивания гена по его 3'-концу с линкером pALK2777 (терминатор cbh1; см. ниже). Нативный терминатор гена протеазы не был включен в конструкцию.
Ген протеазы был вырезан из ПЦР-фрагмента путем расщепления SacII - BamHI и сшит с каркасом экспрессионного вектора pALK2777, расщепленного теми же ферментами (Фиг.2). Плазмида pALK2777 содержала промотор cbh1 (до сайта SacII), линкер, содержащий, например, сайт BamHI, терминатор cbh1 и синтетический ген amdS для скрининга трансформантов. Синтетический ген amdS в pALK2777 содержал терминатор, последовательность которого была укорочена (до сайта XbaI) по сравнению с нативным геном amdS (Kelly and Hynes, 1985). Кроме того, были удалены интроны нативного гена amdS, а выбранные рестрикционные сайты промотора amdS и гена были модифицированы для облегчения конструкции и выделения экспрессионных кассет. Однако, аминокислотная последовательность, кодируемая синтетическим геном amdS, была идентична последовательности, кодируемой геном amdS дикого типа.
Линейная экспрессионная кассета размером 7,2 кб (представлена на Фигуре 3) была выделена из каркаса вектора при расщеплении Nott и была использована для трансформации протопластов Trichoderma reesei. Использованный штамм-хозяин Т. reesei не продуцирует ни одной из четырех основных целлюлаз Т. reesei (CBHI, CBHII, EGI, EGII). Трансформации проводили согласно Penttila et al. (1987) с модификациями, описанными Karhunen et al. (1993). Трансформанты выделяли на чашках с селекционными средами в виде отдельных конидий с последующим высеванием их на PD агар для споруляции.
(б) Продукция протеазы во встряхиваемых колбах и лабораторном биореакторе
Со скошенного PD агара трансформантов высевали во встряхиваемые колбы, содержащие 50 мл многокомпонентной среды на основе лактозы, индуцирующей синтез целлюлазы (Joutsjoki et al., 1993), забуференной 5% KH2PO4 с рН 6,0. Продукцию протеазы трансформантами исследовали в супернатантах культур после выращивания трансформантов в течение 5 дней при 30°С, 250 об/мин. При денатурирующем электрофорезе в полиакриламидном геле кондиционированного культурального супернатанта была обнаружена выраженная полоса белка размером приблизительно 30 кДа, соответствующая ожидаемой массе рекомбинантной протеазы. В образцах определяли протеазную активность с использованием казеина в качестве субстрата, как описано в Примере 3. В культуральных супернатантах определялась активность, которая была несомненно выше, чем у хозяев. Интеграцию экспрессионной кассеты в геном грибов у избранных трансформантов подтверждали при помощи Саузерн-блоттинга, в котором использовали несколько продуктов расщепления геномной ДНК, а в качестве зонда использовали экспрессионную кассету pALK3097.
Трансформанты Т. reesei, проявлявшие наибольшую протеазную активность при культивировании во встряхиваемых колбах, были отобраны для культивирования в лабораторных биореакторах. При культивировании использовали многокомпонентную среду, индуцирующую синтез целлюлазы. Кондиционированную культуральную среду, полученную при культивировании, или концентрированные образцы использовали в качестве стартового материала для очистки и дальнейшей биохимической характеризации рекомбинантной протеазы Malbranchea ALKO4122 (Примеры 3-5), а также в испытаниях, представленных в Примерах 6-10.
Пример 3. Исследование протеазной активности
Протеазную активность определяли с использованием казеина в качестве субстрата. Скорость деградации казеина под действием протеазы определяли, регистрируя высвобождение кислоторастворимых фрагментов пептидов, как функцию времени. Количественное определение кислоторастворимых пептидов проводили спектрофотометрически. Результаты выражали на 1 мкг тирозина в мл (или грамм) за мин.
Вначале готовили растворы всех необходимых реагентов в деионизированной воде, Milli-Q или аналогах следующим образом.
(STW) Искусственная водопроводная вода:
Готовили следующие матричные растворы:
(A) 5,8 г CaCl2 × 2 H2O/200 мл H2O
(B) 2,8 г MgCl2 × 6 H2O/200 мл Н2О
(C) 4,2 г NaHCO3/200 мл H2O
10 мл этих растворов добавляли в заданном порядке к 300 мл H2O при перемешивании, затем доводили объем до 1 литра H2O. Полученный раствор был назван искусственной водопроводной водой.
0,3 М раствор Tris в искусственной водопроводной воде:
36,3 г Trizma base (SIGMA Т-1503) растворяли в искусственной водопроводной воде и доводили объем до 1 литра.
Раствор казеина:
6 г казеина (Hammarsten usd. 12840) добавляли к 350 мл искусственной водопроводной воды и растворяли при перемешивании на магнитной мешалке в течение 10 мин. Добавляли 50 мл раствора Tris и раствор вновь перемешивали в течение 10 мин. Затем раствор нагревали до 70°С. После этого давали температуре снизиться до 50°С и доводили рН до 8,5 при помощи 0,1М NaOH. Перемешивание продолжали до достижения комнатной температуры. Объем раствора доводили до 500 мл искусственной водопроводной водой. Раствор субстрата хранили максимально в течение 3 суток в холодильнике (или в замороженном виде).
110 мМ раствор трихлоруксусной кислоты (раствор для остановки реакции):
18 г ТХУ (Merck 807) растворяли в Н20 и доводили объем до 1 литра.
0,5 М Na2CO:
53 г Na2CO3 растворяли в H2O и доводили объем до 1 литра.
Раствор Фолина:
25 мл 2N фенолового реагента Фолина-Чокальтеу (SIGMA, F 9252) доводили до 100 мл с помощью H2O.
Буфер для разведения образцов:
Образец разводили в 50 мМ Tris-HCl буфере, рН 8,5.
При наиболее подходящем разведении оптическая плотность после проведения реакции должна составлять 0,4-0,8.
Исследование:
В начале исследования 2,5 мл раствора субстрата нагревали в пробирках в течение 5 мин при 50°С. После этого добавляли 0,5 мл разбавленного раствора фермента, перемешивали на вортексе и проводили реакцию при 50°С строго в течение 30 мин. Холостую пробу готовили аналогичным образом, но раствор для остановки реакции (110 мМ ТХУ) добавляли в пробирки до внесения образца. После проведения реакции в пробирки (кроме холостой пробы) добавляли по 2,5 мл останавливающего раствора, содержимое перемешивали и оставляли на 30 минут при комнатной температуре. Пробирки центрифугировали при 4000 об/мин в течение 10 минут (Hettich Rotanta 460). Смешивали 1 мл прозрачного супернатанта с 2,5 мл 0,5 М Na2CO3 и 0,5 мл разведенного реагента Фолина. По истечении 10 мин (развитие окраски) измеряли оптическую плотность смеси (окраску) при 660 нм против холостой пробы.
В каждом измерении параллельно анализировали по меньшей мере два образца.
Пример 4. Очистка рекомбинантной протеазы
Клетки и твердые частицы удаляли из кондиционированной культуральной среды, полученной при ферментации (Пример 2), путем центрифугирования в течение 30 мин при 50000 g при +4°С (Sorvall RC6 plus). Для очистки протеазы использовали 8 мл супернатанта. Все этапы очистки проводили в холодной комнате. После центрифугирования образцы фильтровали через фильтр с порами 0,44 мкм (MILLEX HV Millipore) до нанесения на обессоливающую колонку HiPrep 26/10 (from GE Healthcare), уравновешенную 20 мМ MES, рН 5,3. После гель-фильтрации образцы наносили на колонку S Sepharose HP объемом 1 мл (from GE Healthcare), уравновешенную 20 мМ MES, рН 5,3. Для элюирования белков использовали увеличивающийся градиент NaCl (0,5 М). Фракции, содержащие протеазу, объединяли и концентрировали с помощью центрифужных фильтров Amicon Ultra-4 10,000 СО производства MILLIPORE. Затем образец очищали с помощью гель-фильтрации на колонке Superdex 75, уравновешенной 20 мМ MES, 150 мМ NaCl, pH 5,3. Фракции, содержащие протеазу, объединяли и использовали для дальнейшего исследования профилей pH и температуры. Полученный образец анализировали при помощи денатурирующего электрофореза в полиакриламидном геле, Фиг.4.
Пример 5. Характеризация рекомбинантной протеазы
Температурный профиль
Температурный профиль рекомбинантной протеазы Malbranchea и Савиназы® 16L был получен в ходе исследования, описанного в Примере 3. Результаты приведены на Фиг.5А. Температурный оптимум протеазы составлял приблизительно 70°С.
рН-профиль
рН-профиль протеазы Malbranchea и Савиназы® 16L определяли при 50°C с использованием казеина в качестве субстрата, как описано в Примере 3, за исключением того, что образцы ферментов разводили, а казеин растворяли в 40 мМ буфере Бриттона-Робинсона, реакцию проводили при pH 6-10, время реакции составляло 30 мин, а для остановки ферментативной реакции использовали 0,11 М раствор ТХУ, содержавший 0,22 М ацетат натрия и 0,33 М уксусную кислоту. Результаты приведены на Фиг.5Б. Рекомбинантная протеаза демонстрировала относительную активность, превышающую 50%, при pH от 6 до 10, при этом наивысшая активность наблюдалась при pH приблизительно 10.
Пример 6. Способность протеазы Malbranchea ALKO4122 удалять загрязнения в присутствии жидкого моющего средства при различных температурах
У протеазы Malbranchea ALKO4122, полученной в Trichoderma согласно описанию в Примере 2 (б), исследовали способность удалять загрязнения от стандартного красителя кровь/молоко/чернила при 10, 20, 30 и 50°С в присутствии коммерческого жидкого моющего средства (Таблица 3) при концентрации 5 г/л. В качестве тестового материала использовали искусственно загрязненный образец ткани с нанесенным стандартным красителем, Арт.117 (кровь/молоко/чернила, полиэфир+хлопок, серийный номер No 11-08 или 10-07, ЕМРА Testmaterialen AG, Switzerland). Для сравнения использовали коммерческие препараты протеазы Савиназа® 16L и Савиназа® Ультра 16L (содержащий ингибитор протеаз, 4-FBPA), а также выполняли обработку в отсутствие фермента (контроль). Дозировка каждого ферментного препарата составляла 0-8 или 0-16 единиц активности (мкг тирозина/мин) на мл моющего раствора. Активность измеряли, как описано в Примере 3.
5 г коммерческого жидкого моющего средства растворяли в 1 литре водопроводной воды (dH ≤4), хорошо размешивали на магнитной мешалке и нагревали до температуры стирки. Величина рН моющего раствора составляла прибл. 8. Загрязненную ткань вначале разрезали на лоскуты размером 1,5 см × 1,5 см и закругляли углы кусочков ткани. Затем кусочки помещали в лунки планшетов для микротитрования (Nunc 150200). В каждую лунку диаметром 2 см поверх ткани добавляли по 1,5 мл моющего раствора, содержащего моющее средство и раствор фермента в воде (прибл. 50 мкл). Планшеты с образцами инкубировали в шейкер-инкубаторе Infers Ecotron при 10, 20, 30 и 50°С в течение 60 мин при 130 об/мин. После этого лоскутки аккуратно прополаскивали в проточной воде (прибл. при температуре стирки) и высушивали в течение ночи на воздухе при комнатной температуре на решетке, защищая от солнечного света.
Способность удалять загрязнения оценивали, измеряя окраску как коэффициент отражения при помощи спектрофотометра Minolta CM 2500, используя координаты цветового пространства L*a*b* (illuminant D65/20). После обработки определяли окраску с обеих сторон лоскутков. Каждое значение представляло собой средний результат, полученный по меньшей мере для 2 параллельно обрабатываемых образцов ткани при измерении окраски с обеих сторон ткани. Обесцвечивание загрязнений от крови/молока/чернил, указывающее на качество работы протеазы (эффективность удаления загрязнений) рассчитывали как ΔL* (дельта L*), что означало величину осветления L* ткани, обработанной ферментом, за вычетом величины осветления L* ткани, обработанной моющим раствором без фермента (холостая проба или контроль).
Результаты, приведенные на Фигурах 6А-6Г, указывают, что протеаза Malbranchea ALKO4122 значительно лучше влияет на загрязнения от крови/молока/чернил (Арт.117, серийный номер 11-08) при всех исследованных температурах по сравнению с имеющимися на рынке препаратами Савиназы®. Было отмечено, что загрязнения от крови/молока/чернил предыдущей партии (серийный номер 10-07) было гораздо труднее удалить без фермента (только с помощью моющего средства), и следовательно, вклад фермента (ΔL*) был существенно выше по сравнению с испытаниями, проводимыми на новой партии 11-08. Препарат Malbranchea был еще более эффективным в отношении этого трудноудаляемого застарелого загрязнения по сравнению с Савиназой® (Фиг.7А-7Г). Кроме того, если дозировку рассчитывали по количеству добавляемого белка (Фигуры 8А-8Г), эффективность удаления загрязнений была наиболее высокой у протеазы Malbranchea sulfurea ALKO4122. Количество белка в ферментных препаратах определяли при помощи набора для определения концентрации белка Bio-Rad protein assay (Bio-Rad Laboratories, Hercules, CA), с использованием бычьего гамма-глобулина (Bio-Rad) в качестве стандарта.
Неожиданным оказался тот факт, что протеаза Malbranchea ALKO4122 проявляла прекрасную способность удалять загрязнения в очень широком температурном диапазоне, в частности при низких температурах, например 10-30°С, несмотря на высокие значения ее температурного оптимума в условиях эксперимента при использовании казеина в качестве субстрата (прибл. 70°С, Фиг.5А). Савиназа®, обладавшая схожим температурным профилем в условиях эксперимента (Фиг.5А), несомненно уступала по качеству работы при низких температурах стирки. Результаты этих испытаний указывают, что протеаза Malbranchea ALKO4122 демонстрирует превосходное качество работы в присутствии жидких моющих средств в широком диапазоне температур и даже при очень низких температурах стирки.
Пример 7. Испытания протеазы Malbranchea ALKO4122 в присутствии жидкого моющего средства с использованием различных загрязнений, проводимые на лаундерометре
Исследовали способность протеазы Malbranchea ALKO4122, полученной в Trichoderma согласно описанию в Примере 2 (б), удалять различные загрязнения в присутствии коммерческого жидкого моющего средства при 30 и 60°С в сравнении с коммерческим препаратом протеазы Савиназа® Ультра 16 L. Использовали следующие искусственно загрязненные образцы тканей производства ЕМРА: кровь/молоко/чернила (Арт.117, серийный номер 11-08 и 10-07, полиэфир+хлопок), кровь/молоко/чернила (Арт.116, серийный номер 18-16, хлопок), трава (Арт.164, серийный номер 23-03, хлопок) и какао (Арт112, серийный номер 31-06, хлопок). Ткани разрезали на лоскутки размером 6 см × 6 см и обшивали концы зигзагообразным швом.
Обработку для удаления загрязнений проводили на лаундерометре LP-2 следующим образом. Вначале прогревали лаундерометр до 30 или 60°С. Затем в 1,2-литровые контейнеры, содержащие 250 мл прогретого моющего раствора и загрязненные лоскутки, добавляли фермент по 0, 2, 5, 10 и 20 (40) единиц активности на мл моющего раствора. Активность (мкг тирозина/мин) измеряли согласно описанию в Примере 3. Моющий раствор содержал 5 г коммерческого жидкого моющего средства (Таблица 3) на литр водопроводной воды (dH и 4) и имел рН прибл. 8. Лаундерометр запускали на 60 мин при 30°С со скоростью вращения 42 об/мин. После этого лоскутки аккуратно прополаскивали в проточной воде (прибл. 20°С) и высушивали в течение ночи на воздухе при комнатной температуре на решетке, защищая от солнечного света.
Окраску лоскутков после обработки определяли при помощи спектрофотометра Minolta CM 2500 с использованием координат цветового пространства L*a*b* и рассчитывали способность удалять загрязнения как ΔL*, согласно описанию в Примере 6. После обработки определяли окраску с обеих сторон лоскутков. Каждое значение представляло собой среднее по меньшей мере из 12 измерений для каждого лоскутка. При измерении избегали участков, где в результате сминания ткани при обработке образовались заломы (в хлопковых образцах Арт.116 и Арт.112).
Результаты, приведенные на Фигурах 9А-93, указывают, что протеаза Malbranchea sulfurea ALKO4122 оказывает лучшее воздействие на загрязнения от травы (Арт.164, серийный номер 23-03) и различные загрязнения кровь/молоко/чернила (Арт.117, серийный номер 11-08 и 10-07, Арт.116, серийный номер 18-16), как при 30, так и при 60°С по сравнению с коммерческим препаратом протеазы Савиназой® Ультра 16L. Дозировка 10-20 единиц Савиназы® Ультра 16L на моющий раствор соответствовала дозировке приблизительно 0,4-0,7% ферментного препарата на вес моющего средства, что находится в стандартном диапазоне, в котором используются ферменты, обладающие моющим действием.
Кроме того, результаты, полученные с загрязнением от какао, были лучше у Malbranchea по сравнению с Савиназой® Ультра 16L (данные не приведены). В дополнительных испытаниях на лаундерометре (данные не приведены) было обнаружено, что протеаза Malbranchea эффективна при 30°С также в отношении следующих загрязнений (Центр для тестирования материалов, Center for Testmaterials BV, The Netherlands): арахисовое масло, пигменты, высокобелковое молоко (С-10), желток яйца, пигмент, состаренный (CS-38), экстракт травы (CS-08). Во всех испытаниях использовали одно и то же коммерческое жидкое моющее средство. Результаты этих испытаний указывают, что протеаза Malbranchea эффективна в отношении нескольких загрязнений в широком диапазоне температур 30-60°С.
Пример 8. Способность протеазы Malbranchea ALKO4122 удалять загрязнения в присутствии порошкообразных моющих средств
Способность протеазы Malbranchea ALKO4122, полученной в Trichoderma удалять стандартные загрязнения кровь/молоко/чернила в присутствии моющего средства (5 г/л) исследовали при 50°С в ходе испытания, аналогичного описанному в Примере 7, за исключением того, что использовали обычное коммерческое порошкообразное моющее средство, содержащее отбеливающие агенты, оптические отбеливатели и фосфаты/фосфонаты, а также стандартное моющее средство ЕСЕ 77, не содержащее оптического отбеливателя и отбеливающих агентов (Арт.601, ЕМРА). Окраску лоскутков после обработки определяли при помощи спектрофотометра Minolta CM 2500 с использованием координат цветового пространства L*a*b* и рассчитывали способность удалять загрязнения ΔL*, как описано в Примере 6.
Результаты (Фигуры 10А и 10Б) показывают, что протеаза Malbranchea ALKO4122 может также применяться вместе с порошкообразными моющими средствами в сильно щелочных условиях (рН прибл. 10-10,5).
Пример 9. Стабильность протеазы Malbranchea ALKO4122 при хранении
Исследовали протеазу Malbranchea ALKO4122, полученную в Trichoderma, при хранении при 37°C с использованием следующих составов: 1) 0,2% (вес/вес) Proxel LV (консервант, Arch Biosides, U.K), рН доводили до 6, 2) 20% пропиленгликоль, рН доводили до 6, 3) 50% пропиленгликоль, рН доводили до 5,5. Образцы упаковывали в пробирки Sarstedt's (13 мл) и инкубировали при 37°С. Определяли активность (согласно описанию в Примере 3) через определенные интервалы и сравнивали с результатами, полученными ранее для протеазы Fusarium Fe_RF6318 (WO2010125174A1) в аналогичных условиях.
На Фиг.11 показано, что протеаза Malbranchea ALKO4122 демонстрирует превосходную стабильность при хранении при повышенных температурах (37°С) по сравнению с протеазой Fusarium дикого типа Fe_RF6318. При использовании пропиленгликоля в качестве стабилизатора остаточная активность протеазы Fusarium составляла около 20% через 7 дней инкубации, тогда как протеаза Malbranchea не теряла активности за это время.
Пример 10.Стабильность протеазы Malbranchea ALKO4122 в жидких моющих средствах
Исследовали стабильность протеазы Malbranchea ALKO4122, полученной в Trichoderma, в коммерческом жидком моющем средстве при 37°С (Таблица 3) и стандартном моющем средстве Ecolabel light duty (Ch. Nr. 196-391, wfk Testgewebe GmbH). В качестве референтных препаратов использовали Савиназу® 16L и протеазу Fusarium Fe_RF6318 (WO2010125174A1).
Использовали следующие тестовые системы: 0,4 г ферментного препарата и 9,6 г раствора моющего средства тщательно перемешивали в пробирках Sarstedt (13 мл). Перед смешиванием моющего средства с другими компонентами в него добавляли 0,2% Proxel LV в качестве консерванта. Значение рН образцов, приготовленных в коммерческом жидком моющем средстве составляло прибл. 8. Значения рН образцов, приготовленных в стандартном моющем средстве Ecolabel составляло прибл. 7,2. Пробирки инкубировали при 37°С и определяли протеазную активность через определенные интервалы согласно способу, описанному в Примере 3.
На Фиг.12 показано, что Malbranchea ALKO4122 демонстрирует очень хорошую стабильность при хранении при повышенных температурах (37°С) в жидких моющих средствах, при сравнении с протеазой Fusarium дикого типа Fe_RF6318 (WO2010125174A1). Протеаза Malbranchea обладала схожей стабильностью с Савиназой® 16L в референтном моющем средстве Ecolabel.
При комнатной температуре наблюдалась превосходная стабильность (данные не приведены).
Ссылки
Abu-Shady MR, EI-Gindy AA, Saad RR, Ibrahim ZM. 2001. Production, Partial Purification and Some Properties of Thermostabile Alkaline Protease from Malbranchea sulfurea and its Compatibility with Commercial Detergents. Afr. J. Mycol. And Biotech. 9 (3) (17-26).
Altschul SF, W Gish, W Miller, EW Myers and DJ Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410.
AMFEP, 2009. Association of Manufacturers and Formulators of Enzyme products, List of enzymes at http://www.amfep.org/list.html (updated October 2009).
Anwar, A and M Saleemuddin. 1998. Alkaline proteases: A review. Bioresource Technology 64:175-183.
Bolton, ET and BJ McCarthy. 1962. A general method for the isolation of RNA complementary to DNA. Proc. Nat. Acad. Sci. USA 48:1390-1397.
Chen, Y-J, and M Inouye, 2008. The intramolecular chaperone-mediated protein folding. Curr. Opin. Struct. Biol. 18:765-770.
Cherry, J. R., and Fidantsef, A. L. 2003. Directed evolution of industrial enzymes: an update. Curr. Opin. Biotechnol. 14:438-443.
Gasteiger, E, A Gattiker, С Hoogland, I Ivanyi, RD Appel and A Bairoch. 2003.
Gaucher G M, Stevenson K J 2004. Thermomycolin. Handbook of Proteolytic Enzymes 2nd Ed.: 1834-1835
ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 31:3784-3788.
Gupta, R, QK Beg, S Khan and В Chauhan. 2002. An overview on fermentation, downstream processing and properties of microbial alkaline protease. Appl. Microbiol. Biotechnol. 60:381-395.
Gurr, SJ, Uncles, SE, and Kinghorn JR. 1987. The structure and organization of nuclear genes in filamentous fungi. pp 93-139. In (JR Kinghorn, ed.) Gene Structure in Eukaryotic Microbes. IRL Press, Oxford.
Joutsjoki, VV, TK Torkkeli, and KMH Nevalainen. 1993. Transformation of Trichoderma reesei with the Hormoconis resinae glucoamylase P (gamP) gene: production of a heterologous glucoamylase by Trichoderma reesei. Curr. Genet. 24:223-228.
Kalisz, HM. 1988. Microbial proteinases. Adv. Biochem. Eng. Biotechnol. 36:1-65.
Karhunen T, A Mantyla, KMH Nevalainen, and PL Suominen. 1993. High frequency one-step gene replacement in Trichoderma reesei. I. Endoglucanase I overproduction. Mol. Gen. Genet. 241:515-522.
Kelly and Hynes 1985. Transformation of Aspergillus niger by the amdS gene of Aspergillus nidulans. The EMBO Journal 4(2):475-479.
Laemmli, UK. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680-685.
Malardier L, MJ Daboussi, J Julien, F Roussel, С Scazzocchio and Y Brygoo. 1989. Cloning of the nitrate reductase gene (niaD) of Aspergillus nidulans and its use for transformation of Fusarium oxysporum. Gene 78:147-156.
Maurer, K-H. 2004. Detergent proteases. Curr. Opin. Biotechnol. 15:330-334.
Maurer, K-H, 2010. Enzymes, Detergent, pp.1-17. In (MC Flickinger ed.) Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology, John Wiley & Sons, Inc.
Nielsen H, J Engelbrecht, S Brunak and G von Heijne. 1997. Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Engineering 10:1-6.
Nielsen H and A Krogh. 1998. Prediction of signal peptides and signal anchors by a hidden Markov model. In: Proceedings of the Sixth International Conference on Intelligent Systems for Molecular Biology (ISMB 6), AAAI Press, Menlo Park, California, pp.122-130.
Ong PH and Gaucher GM, 1975. Production, purification and characterization of thermomycolase, the extracellular serine protease of the thermophilic fungus Malbranchea pulchella var. sulfurea. Can. J. Microbiol. 22:165-175.
Penttila M, H Nevalainen, M Ratto, E Salminen, and J Knowles. 1987. A versatile transformation system for the cellulolytic filamentous fungus Trichoderma reesei. Gene 61:155-164.
Raeder U and P Broda. 1985. Rapid preparation of DNA from filamentous fungi. Lett. Appl. Microbiol. 1:17-20.
Rao, MB, AM Tanksale, MS Ghatge and W Deshpande. 1998. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev. 62:597-635.
Sambrook J and DW Russell. 2001. Molecular cloning, a laboratory manual. Cold Spring Harbor Laboratory, New York, US.
Shimogaki, H, K Takenchi, T. Nishino, M. Ohdera, T. Kudo, K. Ohba, MV Iwama and M Irie. 1991. Purification and properties of a novel surface active agent and alkaline-resistant protease from Bcillus sp. Y. Agric. Biol. Chem. 55:2251-2258.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВАЯ ПРОТЕАЗА ГРИБОВ И ЕЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2566549C2 |
| ПРИМЕНЕНИЕ ВАРИАНТОВ ПРОТЕАЗЫ | 2010 |
|
RU2639534C2 |
| ПРОТЕАЗА Streptomyces | 2008 |
|
RU2486244C2 |
| Способы и композиции, содержащие варианты сериновой протеазы | 2012 |
|
RU2663114C2 |
| ВАРИАНТЫ СУБТИЛАЗ | 2010 |
|
RU2651525C2 |
| СПОСОБЫ УЛУЧШЕНИЯ ЭФФЕКТИВНОСТИ БЕЛКОВ | 2008 |
|
RU2569106C2 |
| МОЮЩАЯ КОМПОЗИЦИЯ | 2015 |
|
RU2737535C2 |
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2423525C2 |
| CПОСОБ МЫТЬЯ ПОСУДЫ | 2007 |
|
RU2509152C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЭНДОПЕПТИДАЗНОЙ АКТИВНОСТЬЮ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2011 |
|
RU2583293C2 |

















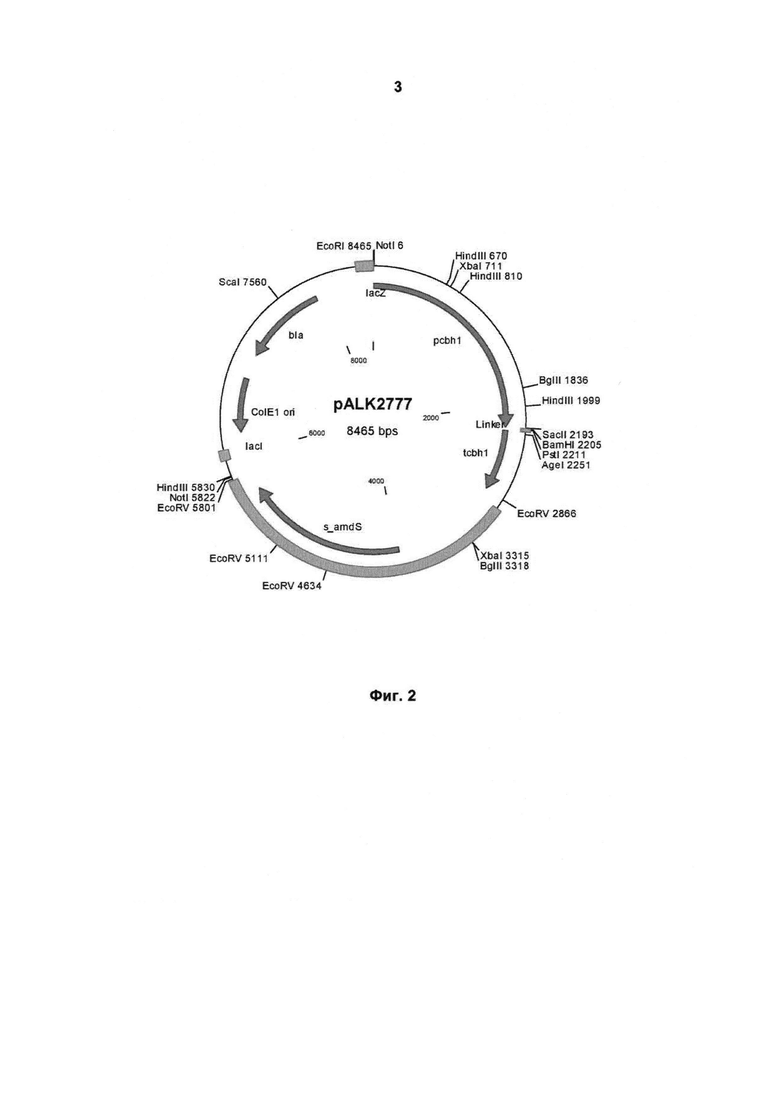
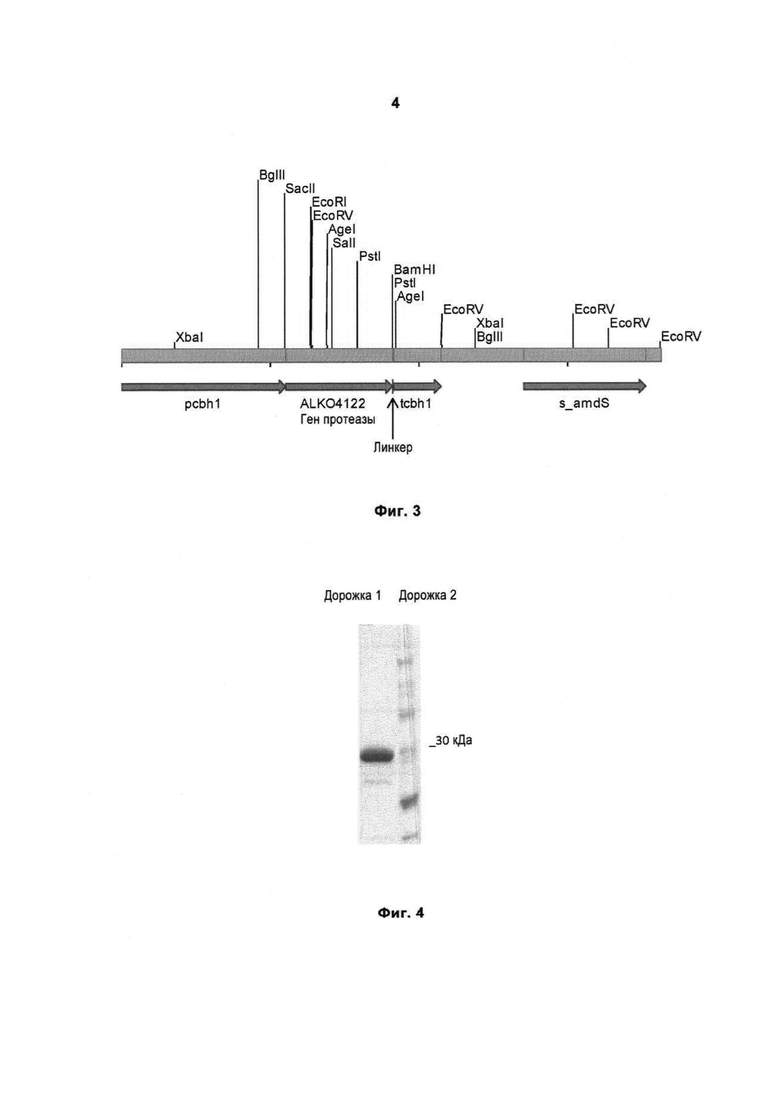
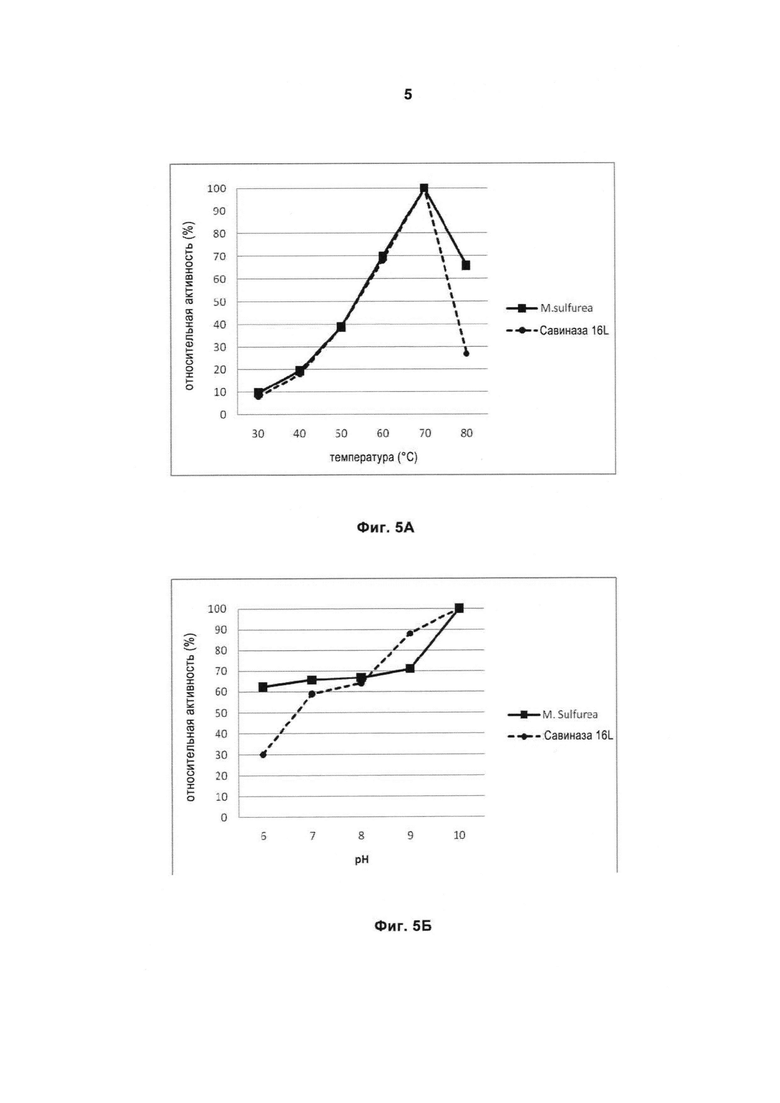
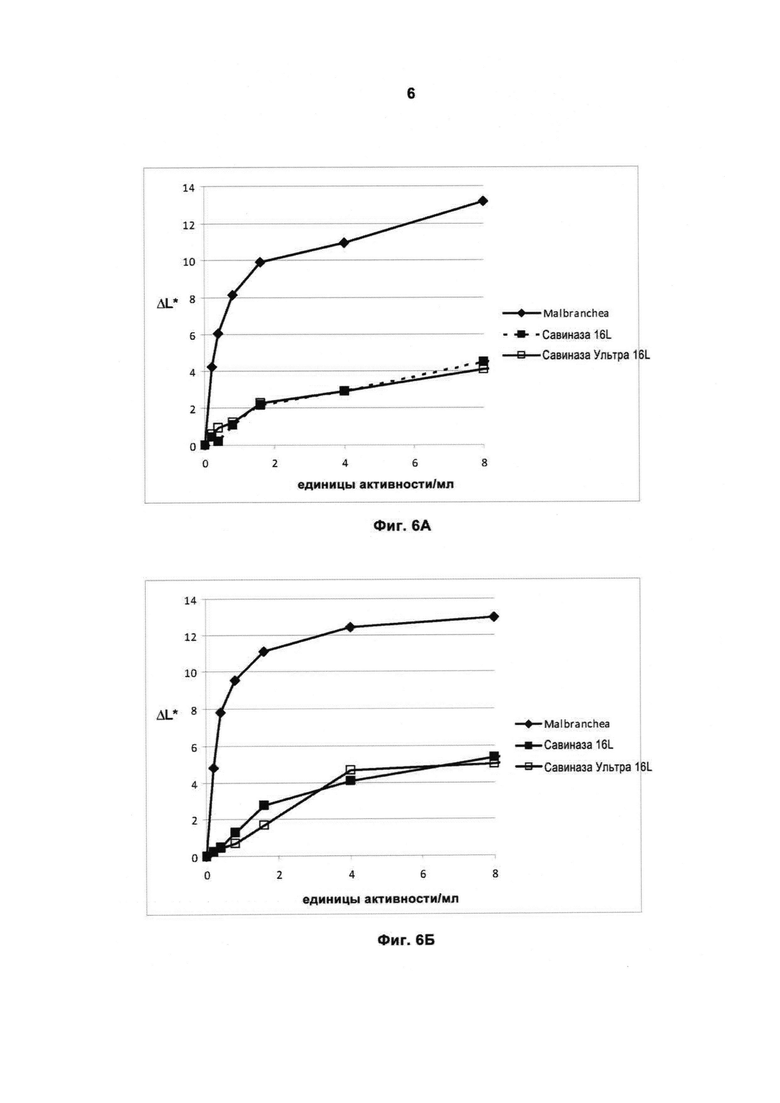
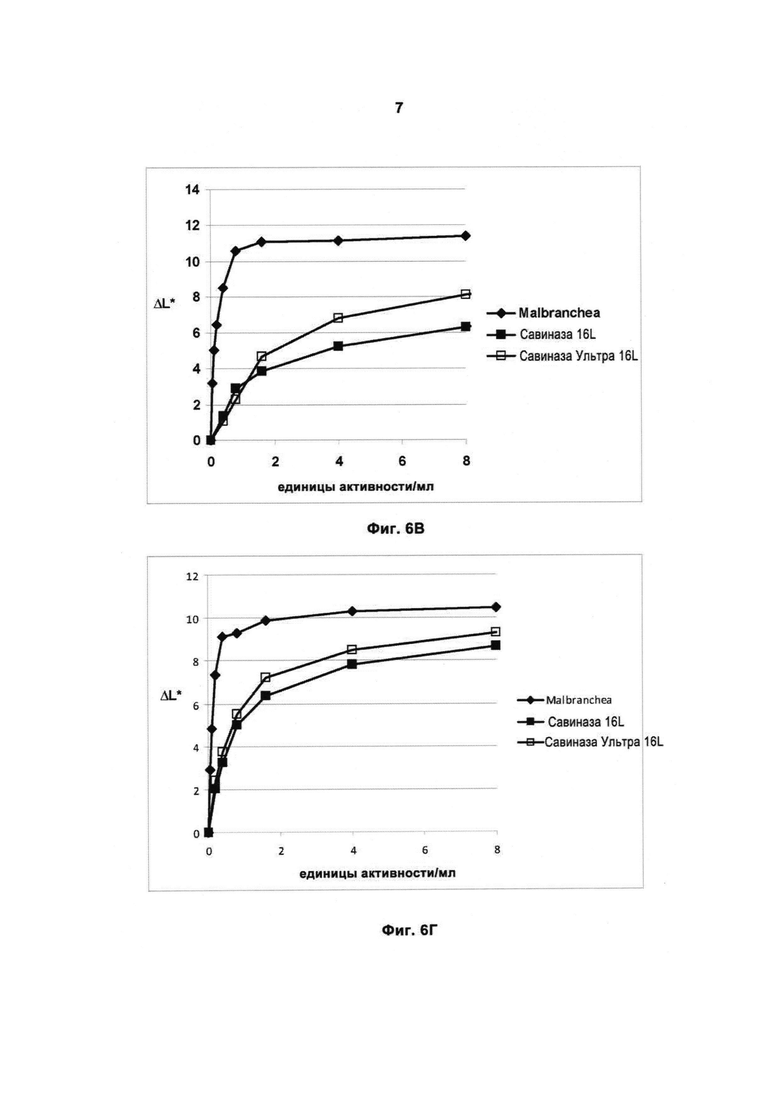

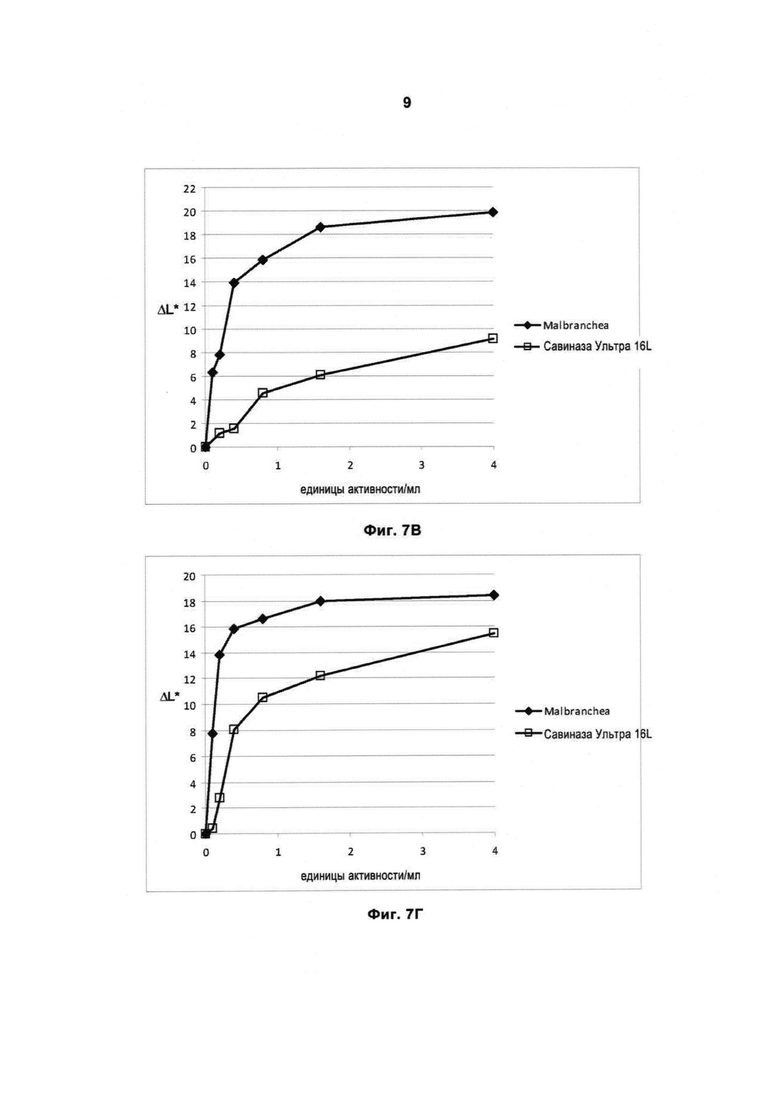

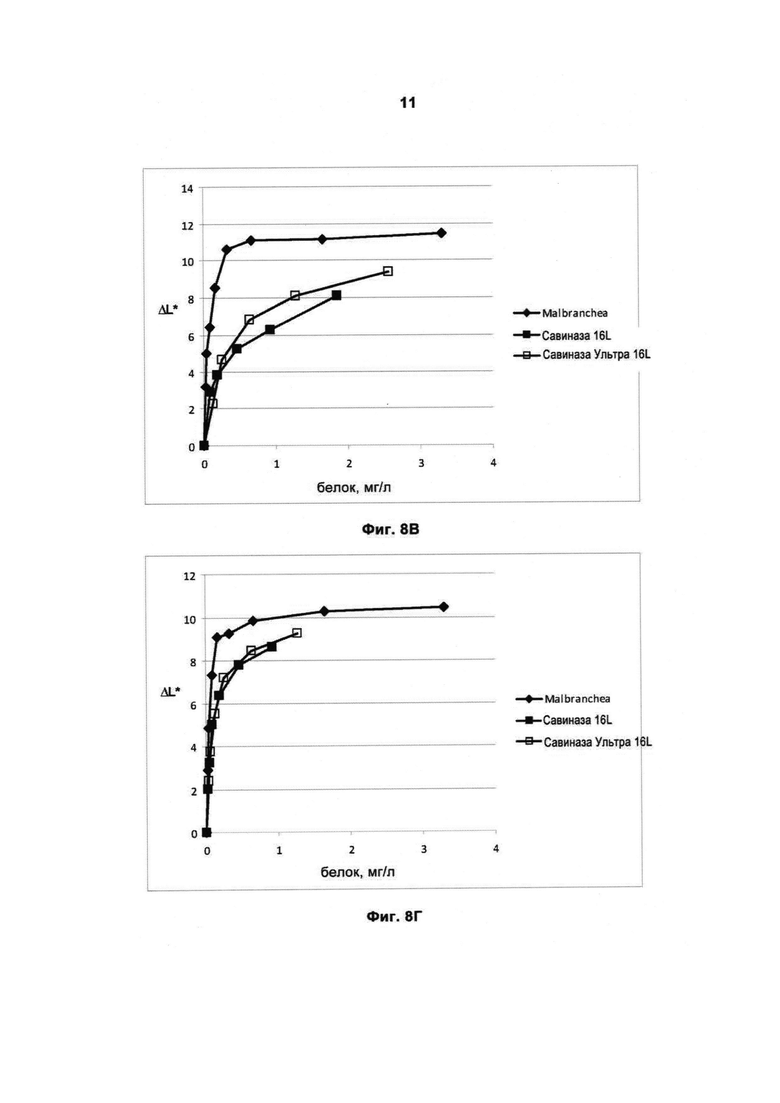
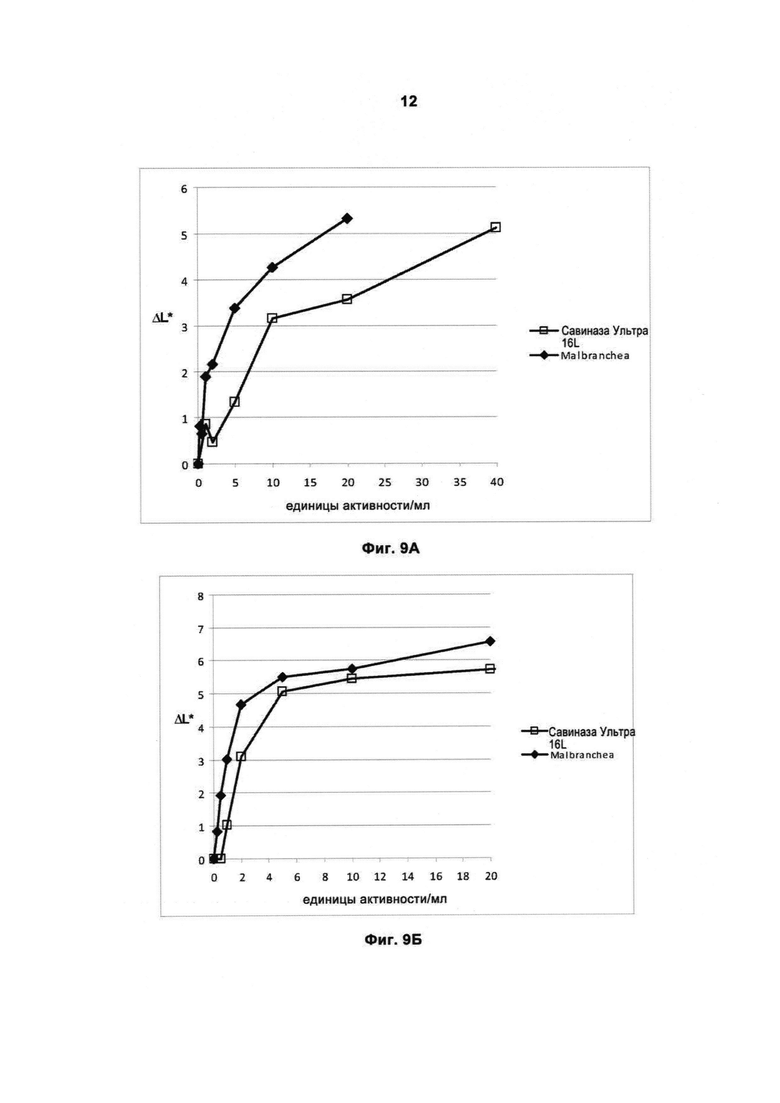
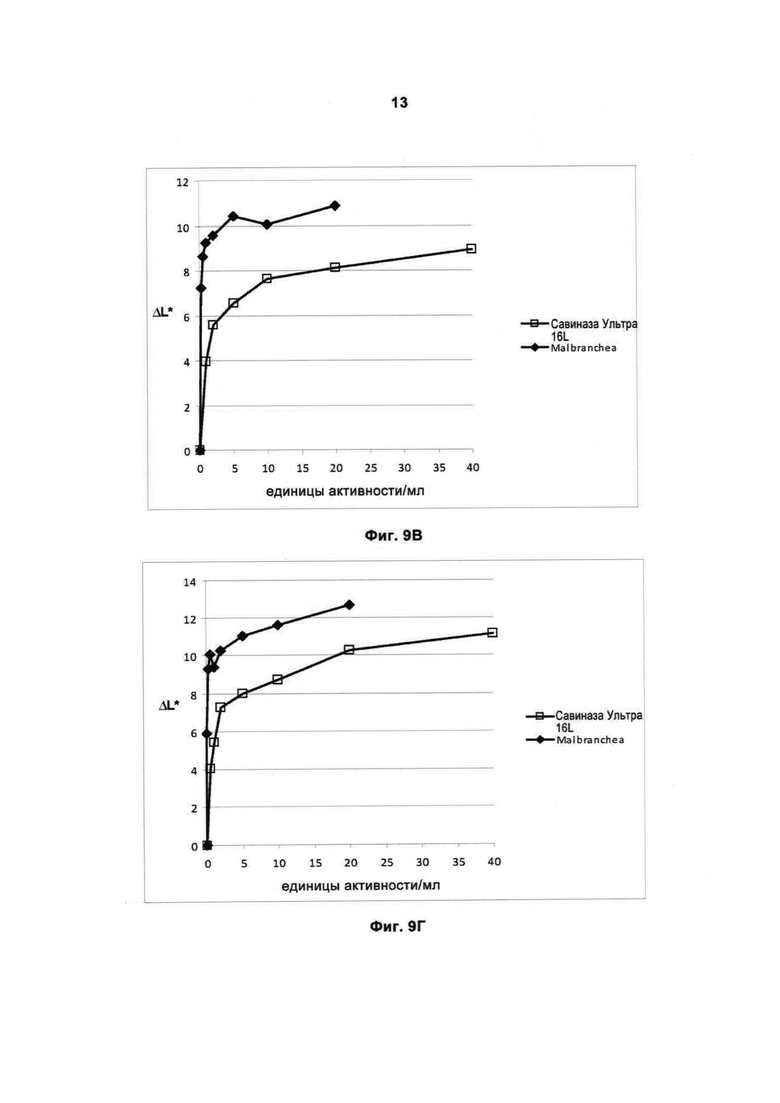

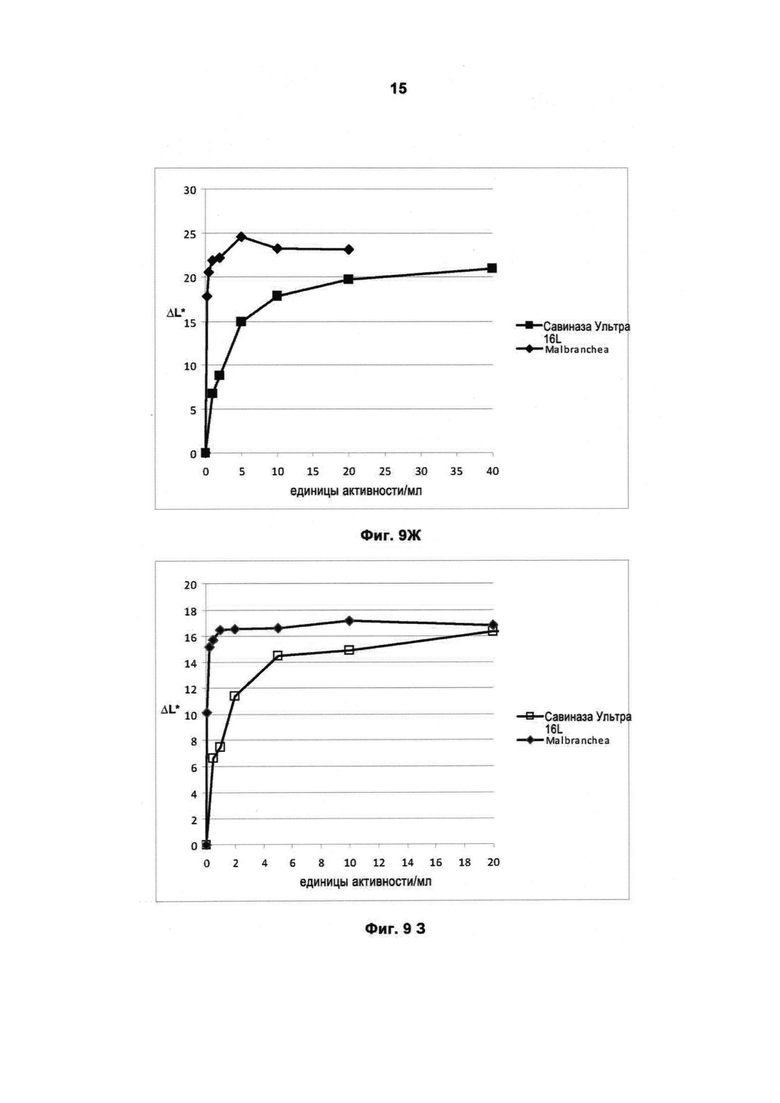
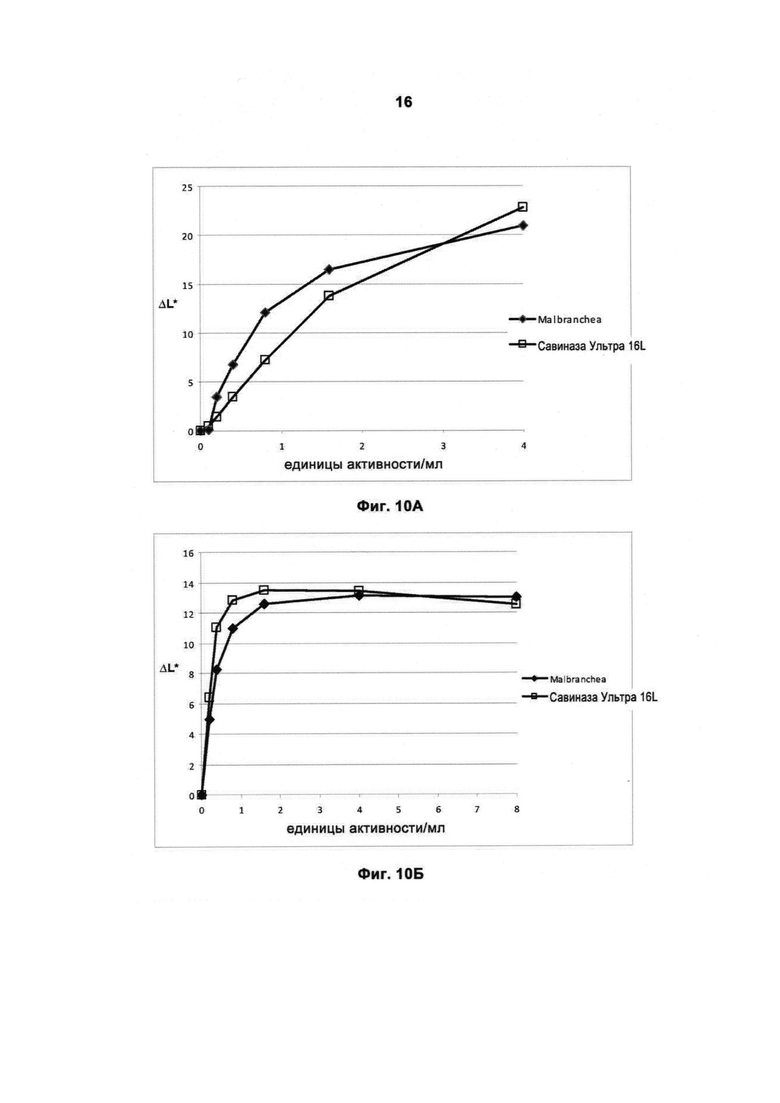
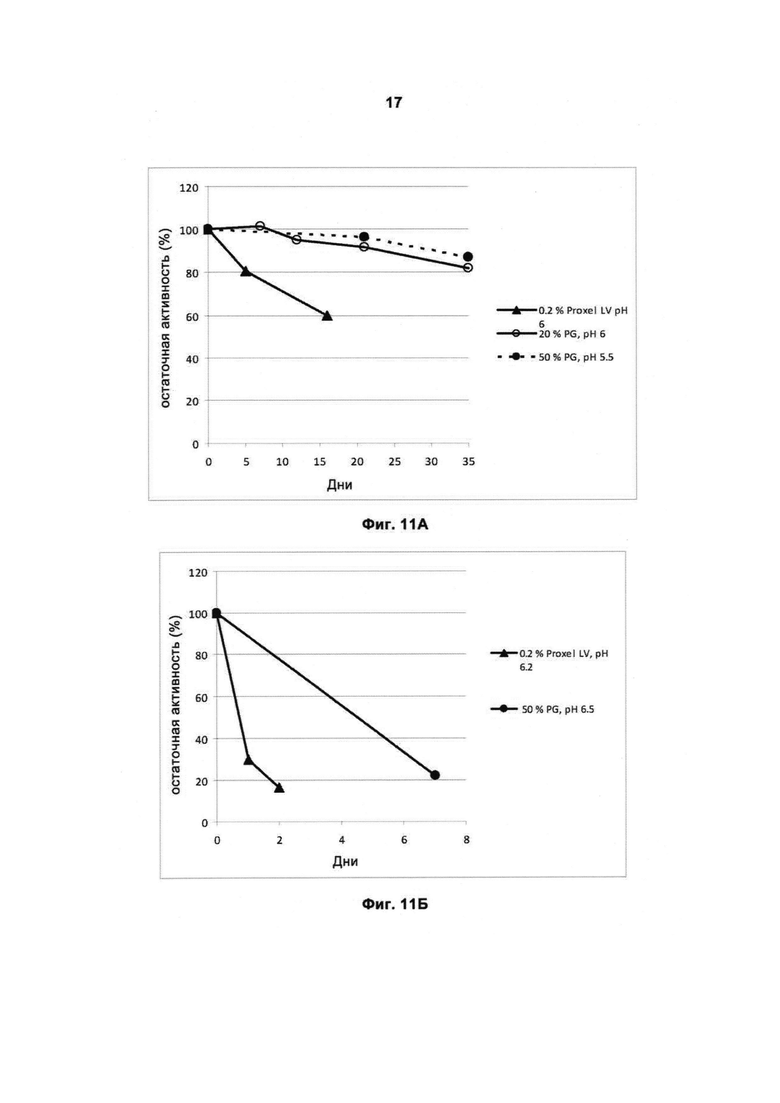
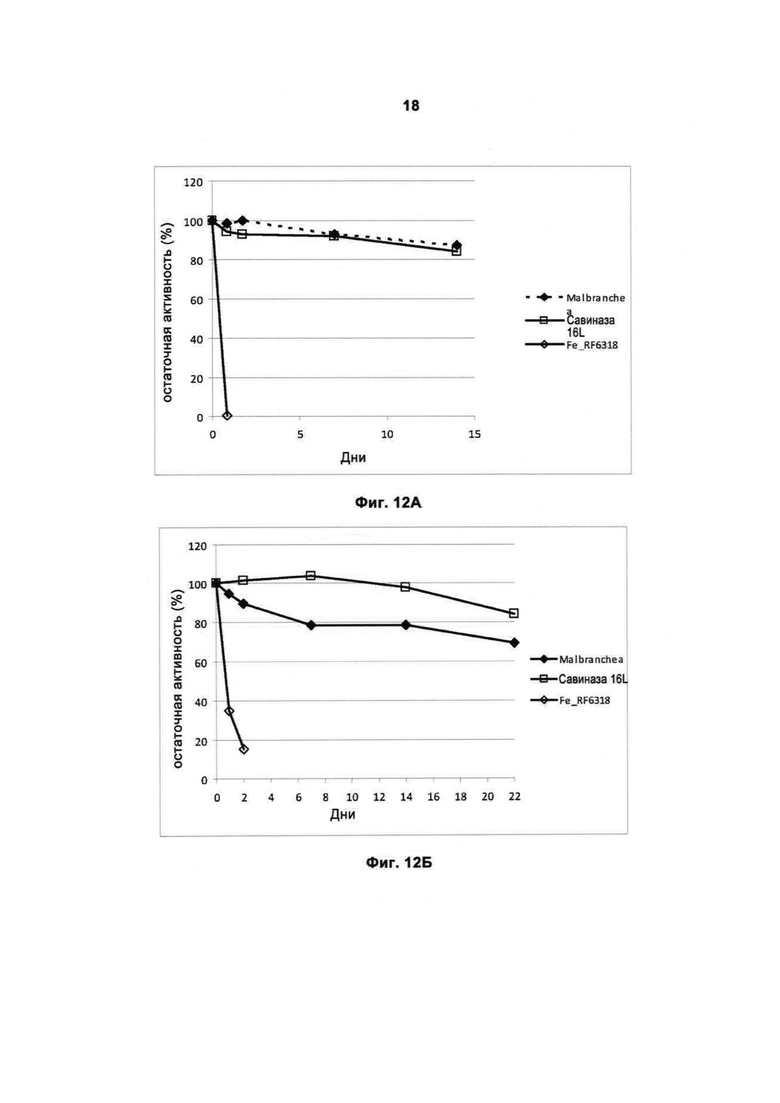
Группа изобретений относится к области биотехнологии. Представлена молекула выделенной нуклеиновой кислоты, кодирующая фермент сериновую протеазу. Представлен способ продукции полипептида, обладающего активностью сериновой протеазы. Представлен рекомбинантный фермент, обладающий активностью сериновой протеазы и содержащий аминокислотную последовательность, кодируемую молекулой вышеуказанной нуклеиновой кислоты. Представлен ферментный препарат, обладающий активностью сериновой протеазы, который содержит вышеуказанный фермент. Представлена детергентная композиция, содержащая вышеуказанный рекомбинантный фермент или вышеуказанный ферментный препарат. Группа изобретений может быть использована для обработки волокон, белкового материала, пищевых продуктов или любого другого использования, включающего модификацию, разложение или удаление белковой субстанции, в том числе и в составе моющих средств. 9 н. и 17 з.п. ф-лы, 33 ил., 3 табл., 10 пр.
1. Молекула выделенной нуклеиновой кислоты, кодирующая фермент сериновую протеазу, где молекула нуклеиновой кислоты выбрана из группы, состоящей из:
(а) молекулы нуклеиновой кислоты, кодирующей полипептид, обладающий активностью сериновой протеазы и содержащий аминокислотную последовательность, представленную в SEQ ID NO: 18;
(б) молекулы нуклеиновой кислоты, кодирующей полипептид, обладающий активностью сериновой протеазы и по меньшей мере на 85% идентичный аминокислотной последовательности SEQ ID NO: 18;
(в) молекулы нуклеиновой кислоты, содержащей кодирующую последовательность нуклеотидной последовательности, представленной в SEQ ID NO: 17;
(г) молекулы нуклеиновой кислоты, содержащей кодирующую последовательность полинуклеотидной последовательности, представленной в SEQ ID NO: 17, содержащейся в штамме, депонированном в DSMZ под номером DSM 24410;
(д) молекулы нуклеиновой кислоты, кодирующая последовательность которой отличается от кодирующей последовательности любой молекулы нуклеиновой кислоты от (в) до (г) вследствие вырожденности генетического кода; и
(е) молекулы нуклеиновой кислоты, содержащей полинуклеотидную последовательность, которая гибридизуется в жестких условиях с молекулой нуклеиновой кислоты по SEQ ID NO: 17.
2. Молекула выделенной нуклеиновой кислоты по п.1, характеризуемая тем, что указанная молекула нуклеиновой кислоты, содержащая полинуклеотидную последовательность, гибридизуется в жестких условиях с полинуклеотидной последовательностью, содержащейся в плазмиде pALK3092, содержащей нуклеотидную последовательность SEQ ID No: 11 в Е. coli RF8758, депонированной под регистрационным номером DSM 24426, или она гибридизуется в жестких условиях с полинуклеотидной последовательностью, содержащейся в плазмиде pALK3093 содержащей нуклеотидную последовательность SEQ ID No: 12, депонированной в Е. coli RF8759 под регистрационным номером DSM 24427.
3. Молекула выделенной нуклеиновой кислоты по п.1 или 2, где молекула выделенной нуклеиновой кислоты содержит полинуклеотидную последовательность, которая гибридизуется в жестких условиях с молекулой нуклеиновой кислоты по SEQ ID NO: 17.
4. Рекомбинантный вектор экспрессии, содержащий нуклеотидную последовательность по любому из пп.1-3, функционально связанную с регуляторными последовательностями, способными направлять экспрессию указанного фермента сериновой протеазы в подходящем хозяине.
5. Клетка-хозяин для продукции сериновой протеазы, кодируемой молекулой нуклеиновой кислоты по п.1, содержащая рекомбинантный вектор экспрессии по п.4.
6. Клетка-хозяин по п.5, характеризуемая тем, что указанный хозяин представляет собой хозяин-микроорганизм.
7. Клетка-хозяин по п.5 или 6, характеризуемая тем, что указанный хозяин представляет собой нитчатый гриб.
8. Клетка-хозяин по любому из пп. 5, 6, характеризуемая тем, что указанный хозяин относится к родам Trichoderma, Aspergillus, Fusarium, Humicola, Chrysosporium, Neurospora, Rhizopus, Penicillium, Myceliophthora и Mortiriella.
9. Клетка-хозяин по п. 8, характеризуемая тем, что указанный хозяин представляет собой Trichoderma или Aspergillus.
10. Клетка-хозяин по п.9, характеризуемая тем, что указанный хозяин представляет собой Т. reesei.
11. Способ продукции полипептида, обладающего активностью сериновой протеазы, указанный процесс включает этапы культивирования клетки-хозяина согласно любому из пп.5-10 и выделения полипептида.
12. Способ по п.11, где молекула выделенной нуклеиновой кислоты включается в геном клетки-хозяина и проводится культивирование в многокомпонентной среде, индуцирующей синтез целлюлазы.
13. Рекомбинантный фермент сериновая протеаза, характеризуемый тем, что указанный фермент обладает активностью сериновой протеазы и содержит аминокислотную последовательность, кодируемую молекулой выделенной нуклеиновой кислоты по любому из пп.1-3.
14. Рекомбинантный фермент сериновая протеаза по п.13, характеризуемый тем, что указанный фермент содержит аминокислотную последовательность, представленную в SEQ ID NO: 18.
15. Рекомбинантный фермент сериновая протеаза по любому из пп. 13, 14, характеризуемая тем, что указанный фермент получен из депонированного штамма CBS 128533 Malbranchea ALKO4122 или из депонированного штамма CBS 128564 Malbranchea ALKO4178.
16. Рекомбинантный фермент сериновая протеаза по любому из пп. 13, 14, характеризуемый тем, что зрелая форма указанного фермента имеет молекулярную массу от 20 до 35 кДа, предпочтительно от 25 до 33 кДа, наиболее предпочтительно от 28 до 30 кДа.
17. Рекомбинантный фермент сериновая протеаза по любому из пп. 13, 14, характеризуемый тем, что указанный фермент имеет температурный оптимум при pH 8,5, времени реакции 30 мин и использовании казеина в качестве субстрата, составляющий от 50°C до 80°C, предпочтительно 70°C.
18. Рекомбинантная сериновая протеаза по любому из пп. 13, 14, характеризуемая тем, что указанный фермент имеет pH оптимум при температуре 50°C, времени реакции 30 мин и использовании казеина в качестве субстрата, равный pH 10.
19. Ферментный препарат, обладающий активностью сериновой протеазы, который содержит рекомбинантный фермент сериновую протеазу по любому из пп.13-18.
20. Ферментный препарат по п.19, характеризуемый тем, что указанный препарат содержит другие ферменты, выбранные из группы, состоящей из протеаз, амилаз, целлюлаз, липаз, ксиланаз, маннаназ, кутиназ, пектиназ и оксидаз, с добавлением или без добавления медиатора.
21. Ферментный препарат по любому из пп. 19, 20, характеризуемый тем, что указанный препарат содержит один или более дополнительных компонентов, выбранных из группы, состоящей из стабилизаторов, буферов, сурфактантов, структурообразователей, отбеливающих агентов, медиаторов, антикоррозийных агентов, препятствующих повторному осаждению агентов, щелочей, абразивов, оптических отбеливателей, красителей, отдушек, пигментов и консервантов.
22. Ферментный препарат по любому из пп. 19, 20, характеризуемый тем, что указанный ферментный препарат находится в форме жидкости, порошка, гранулята или таблеток.
23. Применение рекомбинантного фермента сериновой протеазы по любому из пп.13-16 или ферментного препарата по любому из пп.19-22 в моющих средствах для обработки волокон, для обработки белковых материалов, предпочтительно выбранных из шерсти, волос, кожи и шелка, для обработки пищевых продуктов или кормов или для применений, включающих модификацию, разложение или удаление белковой субстанции.
24. Применение рекомбинантного фермента сериновой протеазы по любому из пп.13-16 или ферментного препарата по любому из пп.19-22 в качестве моющей добавки, предпочтительно в жидких моющих средствах, в порошкообразных моющих средствах и в таблетированных моющих средствах.
25. Детергентная композиция, характеризуемая тем, что указанная композиция содержит сурфактанты, рекомбинантный фермент сериновую протеазу по любому из пп.13-16 или ферментный препарат по любому из пп.19-22, а также дополнительные компоненты, выбранные из группы, состоящей из стабилизаторов, буферов, структурообразователей, отбеливающих агентов, медиаторов, антикоррозийных агентов, препятствующих повторному осаждению агентов, щелочей, абразивов, оптических отбеливателей, красителей, отдушек, пигментов и консервантов.
26. Детергентная композиция по п.25, характеризуемая тем, что указанная композиция содержит другие ферменты, выбранные из группы, состоящей из протеаз, амилаз, целлюлаз, липаз, ксиланаз, маннаназ, кутиназ, пектиназ и оксидаз, с добавлением или без добавления медиатора.
| WO2008048392 A1, 24.04.2008 | |||
| WO2010125174 A1, 04.11.2010 | |||
| ВАРИАНТЫ ПРОТЕАЗЫ, ЗАМЕЩЕННЫЕ В НЕСКОЛЬКИХ ПОЛОЖЕНИЯХ | 1998 |
|
RU2269572C2 |

