Эта заявка заявляет права в соответствии с 35 U.S.С. §119(е) на все преимущества заявок US 60/627,763, зарегистрированной 12 ноября 2004 года; US 60/642,886, зарегистрированной 11 января 2006 года; US 60/649,508, зарегистрированной 2 февраля 2005 года; US 60/662,468, зарегистрированной 15 марта 2005 года; US 60/669,311, зарегистрированной 6 апреля 2005 года; US 60/681,607, зарегистрированной 16 мая 2005 года, US 60/690,200, зарегистрированной 13 июня 2005 года; US 60/696,609, зарегистрированной 5 июля 2005 года; US 60/703,018, зарегистрированной 27 июля 2005 года и US 60/726,453, зарегистрированной 12 августа 2005 года, которые включены путем отсылки во всей своей полноте.
Область техники, к которой относится изобретение
Это изобретение имеет отношение к оптимизированным вариантам иммуноглобулинов IgG, к конструкторским способам их получения и к их применению, в особенности в терапевтических целях.
Уровень техники
Антитела представляют собой иммунологические белки, которые связывают специфический антиген. У большинства млекопитающих, включая людей и мышей, антитела построены из спаренных тяжелых и легких полипептидных цепей. Каждая цепь собирается из индивидуальных доменов иммуноглобулина (Ig), и, таким образом, для таких белков используют общий термин иммуноглобулин. Каждая цепь собирается из двух различных участков, относящихся к вариабельной и константной области. Вариабельные области легкой и тяжелой цепей демонстрируют значительное различие в последовательности между антителами и отвечают за связывание с антигеном-мишенью. Константные области демонстрируют меньшие различия в последовательности и отвечают за связывание с большим количеством природных белков для достижения значимых биохимических эффектов. У человека существуют пять различных классов антител, включающих IgA (которые включают подклассы IgA1 и IgA2), IgD, IgE, IgG (которые включают подклассы IgG1, IgG2, IgG3 и IgG4) и IgM. Характерной особенностью этих классов антител являются их константные области, хотя более тонкие различия могут существовать и в V-области. Фигура 1 демонстрирует антитело IgG1, используемое здесь в качестве примера, чтобы описать основные структурные черты иммуноглобулинов. Антитела IgG представляют собой тетрамерные белки, состоящие из двух тяжелых цепей и двух легких цепей. Тяжелая цепь IgG состоит из четырех иммуноглобулиновых доменов, присоединенных от N-конца к С-концу в порядке VH-СН1-СН2-СН3, относящихся к вариабельному домену тяжелой цепи, константному домену 1 тяжелой цепи, константному домену 2 тяжелой цепи и константному домену 3 тяжелой цепи соответственно (также называемых VH-Cγ1-Cγ2-Cγ3, относящихся к вариабельному домену тяжелой цепи, константному домену гамма-1, константному домену гамма-2 и константному домену гамма-3 соответственно). Легкая цепь IgG состоит из двух иммуноглобулиновых доменов, присоединенных от N-конца к С-концу в порядке VL-CL, относящихся к вариабельному домену легкой цепи и константному домену легкой цепи соответственно.
Вариабельная область антитела содержит антиген-связывающие детерминанты молекулы и, таким образом, определяет специфичность антитела в отношении антигена-мишени. Вариабельная область называется так потому, что она наиболее сильно различается по последовательности от других антител внутри одного и того же класса. Наибольшие различия в последовательности обнаруживаются в гипервариабельных участках (CDR). Всего существует 6 участков CDR, по три на каждую тяжелую и легкую цепи, обозначаемые как VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3. Вариабельный участок, находящийся вне участков CDR, относится к каркасному участку (FR). Среди различных антител наблюдают различия последовательности в участках FR, хотя и не такие существенные, как в участках CDR. В целом, это характерное строение антител обеспечивает стабильную основу (участок FR), на которой значительное разнообразие связывания антигенов (CDR) может использоваться иммунной системой для достижения специфичности для широкого множества антигенов. Для множества фрагментов вариабельных областей различных организмов доступно некоторое количество структур, определенных с высоким разрешением, некоторые из них находятся в свободном виде, а некоторые находятся в комплексе с антигенами. Последовательность и структурные особенности вариабельных областей антител хорошо охарактеризованы (Morea et al., 1997, Biophys Chem 68:9-16; Morea et al., 2000, Methods 20:267-279, работы включены путем отсылки во всей своей полноте), консервативные свойства антител позволили развить множество инженерных технологий для производства антител (Maynard et al., 2000, Annu Rev Biomed Eng 2:339-376, работа включена путем отсылки во всей своей полноте). Например, возможно пересадить CDR от одного антитела, например мышиного антитела, на каркасный участок другого антитела, например человеческого антитела. Этот процесс, называемый в этой области техники «гуманизацией», позволяет получить менее иммуногенные терапевтические антитела из антител, не имеющих отношения к человеческим. Фрагменты, включающие вариабельную область, могут существовать в отсутствие других областей антитела, такие фрагменты включают, например, фрагмент, связывающий антиген (Fab), включающий VH-Cγ1 и VH-CL; вариабельный фрагмент (Fv), включающий VH и VL; одноцепочечный вариабельный фрагмент (scFv), включающий VH и VL, связанные между собой в одну и ту же цепь; а также множество других фрагментов вариабельной области (Little et al., 2000, Immunol Today 21:364-370, работа включена путем отсылки во всей своей полноте).
Участок антитела Fc взаимодействует с различными рецепторами Fc и лигандами, которые сообщают ему множество важных функциональных возможностей, так называемые эффекторные функции. В иммуноглобулинах IgG Fc-участок, как показано на фигурах 1 и 2, включает иммуноглобулиновый домен Сγ2 и Сγ3 и N-концевой шарнир, ведущий в Сγ2. Важное семейство рецепторов Fc класса IgG включает гамма-рецепторы Fc (FcγRs). Эти рецепторы осуществляют связь между антителами и клеточными механизмами иммунной системы (Raghavan et al., 1996, Annu Rev Cell Dev Biol 12:181-220; Ravetch et al., 2001, Annu Rev Immunol 19:275-290, все работы включены путем отсылки во всей своей полноте). У людей это семейство белков включает FcγRI (CD64), включающий изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), включающий изоформы FcγRIIa (включающий аллотипы Н131 и R131), FcγRIIb (включающий FcγRIIb-1 и FcγRIIb-2) и FcγRIIc; и FcγRIII (CD16), включающий изоформы FcγRIIIa (включающий аллотипы V158 и F158) и FcγRIIIb (включающий аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2) (Jefferis et al., 2002, Immunol Lett 82:57-65, работа включена путем отсылки во всей своей полноте). Эти рецепторы обычно имеют внеклеточный домен, который опосредует связывание с Fc, мембранно-связанный участок и внутриклеточный домен, который может опосредовать некоторые сигнальные события внутри клетки. Эти рецепторы экспрессируются в различных иммунных клетках, включающих моноциты, макрофаги, нейтрофилы, дендритные клетки, эозинофилы, тучные клетки, тромбоциты, В-клетки, большие зернистые лимфоциты, клетки Лангерганса, натуральные киллеры (NK) и γγ Т-клетки. Образование комплекса Fc/FcγR направляет эти эффекторные клетки к местам связывания антигенов, что обычно приводит к проведению сигнала в клетках и важным последующим иммунным ответам, таким как высвобождение воспалительных медиаторов, активация В-клеток, эндоцитоз, фагоцитоз и цитотоксическая атака. Способность опосредовать цитотоксическую и фаготоксическую эффекторную функцию является потенциальным механизмом, с помощью которого антитела разрушают клетки-мишени. Опосредованная клетками реакция, в которой неспецифические цитотоксические клетки, экспрессирующие FcγRs, узнают антитело, связанное на поверхности клетки-мишени, и затем вызывают лизис клетки-мишени, называется антитело-зависимой клеточно-опосредованной цитотоксичностью (ADCC) (Raghavan et al., 1996, Annu Rev Cell Dev Biol 12:181-220; Gheitie et al., 2000, Annu Rev Immunol 18:739-766; Ravetch et al., 2001, Annu Rev Immunol 19:275-290, все работы включены путем отсылки во всей своей полноте). Опосредованную клетками реакцию, в которой неспецифические цитотоксические клетки, экспрессирущие FcγRs, узнают антитело, связанное на поверхности клетки-мишени, и затем вызывают фагоцитоз клетки-мишени, называют антитело-зависимым клеточно-опосредованным фагоцитозом клеток-мишеней (ADCP). Было определено некоторое количество структур внеклеточных доменов FcγRs человека, включающих FcγRsIIa (код доступа в базе данных pdb 1H9V, включен путем отсылки во всей своей полноте) (Sonderman et al., 2001, J Mol Biol 309:737-749, работа включена путем отсылки во всей своей полноте) (код доступа в базе данных pdb 1FCG, включен путем отсылки во всей своей полноте) (Maxwell et al., 1999, Nat Struc Biol 6:437-442, работа включена путем отсылки во всей своей полноте), FcγRsIIb (код доступа в базе данных pdb 2FCB, включен путем отсылки во всей своей полноте) (Sondermann et al., 1999, Embo J 18:1095-1103, работа включена путем отсылки во всей своей полноте); и FcγRsIIIb (код доступа в базу данных pdb 1E4J, включен путем отсылки во всей своей полноте) (Sondermann et al., 2000, Nature 406:267-273, работа включена путем отсылки во всей своей полноте). Все FcγRs связывают один и тот же Fc-участок в N-концевом участке домена Сγ2 и предшествующего шарнирного участка, показанных на фигуре 1. Это взаимодействие хорошо охарактеризовано структурно (Sondermann et al., 2001, J Mol Biol 309:737-749, работа включена путем отсылки во всей своей полноте), и было определено несколько структур Fc человека, связывающихся с внеклеточным доменом FcγRsIIIb человека (код доступа в базе данных pdb 1E4K, включен путем отсылки во всей своей полноте) (Sondermann et al., 2000, Nature 406:267-273, работа включена путем отсылки во всей своей полноте) (код доступа в базе данных pdb 1IIS и 1IIX, включен путем отсылки во всей своей полноте) (Radaev et al., 2001, J Biol Chem 276:16469-16477, работа включена путем отсылки во всей своей полноте), а также структура комплекса Fc/FcєRIα человеческого IgE (код доступа в базе данных pdb 1F6A, включен путем отсылки во всей своей полноте) (Garman et al., 2000, Nature 406:259-266, работа включена путем отсылки во всей своей полноте).
Различные подклассы IgG обладают различным сродством к FcγRs, обычно IgG1 и IgG3 связываются с рецепторами значительно лучше, чем IgG2 и IgG4 (Jefferis et al., 2002, Immunol Lett. 82:57-65, работа включена путем отсылки во всей своей полноте). Все FcγRs связываются с одним и тем же Fc-участком IgG, но с различным сродством: FcγRI, связывающий с самым высоким сродством, имеет Kd для IgG1, равную 10-8М, тогда как рецепторы с низким сродством, FcγRII и FcγRIII, связываются при 10-6М и 10-5М соответственно. Внеклеточные домены FcγRIIIa и FcγRIIIb идентичны на 96%, однако FcγRIIIb не имеет внутриклеточного сигнального домена. Кроме того, поскольку FcγRI, FcγRIIa/с и FcγRIIIa являются положительными регуляторами активации, запускаемой иммунным комплексом, они характеризуются наличием внутриклеточного домена, в состав которого входит иммунорецепторный тирозин-зависимый активационный мотив (ITAM), FcγRIIb имеет в своем составе иммунорецепторный тирозин-зависимый ингибиторный мотив (ITIM) и поэтому является ингибитором. Таким образом, первые рецепторы относятся к активирующим рецепторам, a FcγRIIb относится к ингибирующим рецепторам. Рецепторы также различаются по характеру и уровням экспрессии на различных иммунных клетках. Еще один уровень сложности состоит в существовании полиморфизма FcγR в протеоме человека. Особенно важным полиморфизмом с клиническим значением являются формы V158/F158 FcγRIIIa. IgG1 человека связывается с более высоким сродством с аллотипом V158 по сравнению с аллотипом F158. Показано, что различие в сродстве и предположительно влияние этого различия на ADCC и/или ADCP являются существенным показателем эффективности антитела к CD20 ритуксимаба (Rituxan®, BiogenIdec). Пациенты с аллотипом V158 благоприятно реагируют на лечение ритуксимабом, однако пациенты с аллотипом F158, обладающим более низкой аффиностью, слабо отвечают на такое лечение (Cartron et al., 2002, Blood 99:754-758, работа включена путем отсылки во всей своей полноте). Приблизительно 10-20% людей являются гомозиготами V158/V158, 45% являются гетерозиготами V158/F158 и 35-45% людей являются гомозиготами F158/F158 (Lehmbecher et al., 1999, Blood 94:4220-4232; Cartron et al., 2002, Blood 99:754-758, работы включены путем отсылки во всей своей полноте). Таким образом, 80-90% людей являются слабо реагирующими, так как у них есть, по меньшей мере, одна аллель F158 FcγRIIIa.
Перекрывающийся, но индивидуальный участок на Fc, показанный на фигуре 1, служит контактной поверхностью для белка комплемента C1q. Тем же самым способом, при котором связывание Fc/FcγR опосредует ADCC, связывание Fc/C1q опосредует зависимую от комплемента цитотоксичность (CDC). Чтобы сформировать комплекс С1, C1q образует комплекс с сериновыми протеазами С1r и C1s. C1q способен к связыванию с шестью антителами, хотя для активации комплементного каскада достаточно связывания с двумя IgG. Подобно взаимодействию Fc с FcγRs, различные подклассы IgG обладают различным сродством к C1q, обычно IgG1 и IgG3 лучше связываются с FcγRs, чем IgG2 и IgG4 (Jefferis et al., 2002, Immunol Lett. 82:57-65, работа включена путем отсылки во всей своей полноте.
В IgG участок на Fc между доменами Сγ2 и Сγ3 (фигура 1) опосредует взаимодействие с неонатальным рецептором FcRn, связывание которого приводит к рециркуляции эндоцитозного антитела от эндосомы обратно в кровяное русло (Raghavan et al., 1996, Annu Rev Cell Dev Biol 12:181-220; Ghetie et al., 2000, Annu Rev Immunol 18:739-766, обе работы включены путем отсылки во всей своей полноте). Этот процесс, сопряженный с устранением почечной фильтрации из-за большого габарита полноразмерной молекулы, приводит к тому, что время полужизни подходящих антител в сыворотке крови составляет диапазон от одной до трех недель. Связывание Fc с FcRn также играет ключевую роль в транспорте антител. Участок связывания для FcRn на Fc также является участком, в котором связываются бактериальные протеины А и G. Прочное связывание с этими белками обычно используют в качестве способа очистки антител при применении протеин А- или протеин G- аффинной хроматографии во время очистки белков. Таким образом, точное воспроизведение этого участка на Fc очень существенно как для клинических свойств антител, так и для их очистки. Имеющиеся в распоряжении структуры крысиного комплекса Fc/FcRn (Burmeister et al., 1994, Nature, 372:379-383; Martin et al., 2001, Mol Cell 7:867-877, обе работы включены путем отсылки во всей своей полноте) и комплексов с протеинами А и G (Deisenhofer et al., 1981, Biochemistry 20:2361-2370; Sauer-Eriksson et al., 1995, Structure 3:265-278; Tashiro et al., 1995, Curr Opin Struct Biol 5:471-481, все работы включены путем отсылки во всей своей полноте) обеспечивают понимание того, как происходит взаимодействие Fc с этими белками. Рецептор FcRn также отвечает за перенос IgG в желудочно-кишечный тракт новорожденных и в просвет эпителия кишечника у взрослых (Ghetie and Ward, Annu. Rev. Immunol., 2000, 18:739-766; Yoshida et al., Immunity, 2004, 20(6):769-783, обе работы включены путем отсылки во всей своей полноте).
Исследования, проведенные на доменах Fcγ крысы и человека, продемонстрировали важность некоторых остатков Fc в связывании FcRn. Последовательности в Fc-участках у крыс и человека идентичны приблизительно на 65% (остатки 237-443 в нумерации по Kabat et al.). См. фигуры 3, 4 и 5, показывающие результаты выравнивания Fc, тяжелой цепи FcRn и легкой цепи FcRn (бета-2-микроглобулин) крысы/человека. На основе существующей структуры комплекса Fc/FcRn крысы была построена модель комплекса Fc/FcRn человека (Martin et al., 2001, Mol. Cell 7:867-877, работа включена путем отсылки во всей своей полноте). Последовательности у крысы и человека содержат некоторые общие для них остатки, которые критичны для связывания FcRn, например Н310 и Н435 (Medesan et al., 1997, J. Immunol. 158(5):221-7; Shields et al., 2001, J. Biol. Chem. 276(9):6591-6604, обе работы включены путем отсылки во всей своей полноте). Во многих положениях, однако, белки у крысы и человека имеют разные аминокислоты, обеспечивая для остатков в последовательности у человека другое окружение и возможно другие особенности по сравнению с последовательностью у крысы. Эта вариабельность ограничивает способность переносить характеристики одного гомолога на другой гомолог.
В мышином Fcγ случайная мутация и селекция с использованием фагового дисплея в участках Т252, Т254 и Т256 приводят к тройному мутанту T252L/T254S/T256F, у которого сродство к FcRn повышается в 3,5 раза и в 1,5 раза увеличивается время полужизни (Ghetie et al., 1997, Nat. Biotech. 15(7):637-640, работа включена путем отсылки во всей своей полноте).
Кристаллические структуры комплекса Fc/FcRn крысы идентифицировали остатки Fc, важные для связывания FcRn (Burmeister et al., Nature, 372:379-383 (1994); Martin et al. Molecular Cell. 7:867-877 (2001), обе работы включены путем отсылки во всей своей полноте). Первоначальная структура комплекса Fc/FcRn была получена в 1994 году с разрешением 6 Е (таблица 2а, Burmeister et al., Nature, 372:379-383 (1994), работа включена путем отсылки во всей своей полноте). Структура с более высоким разрешением, выполненная в 2001 году Martin et al., показала более детальный вид расположения боковых цепей (Martin et al. Molecular Cell. 7:867-877 (2001), работа включена путем отсылки во всей своей полноте). Эта кристаллическая структура Fc крысы, связанного с FcRn крысы, была разрешена с использованием димера Fc с одним мономером, содержащим мутации Т252G/I253G/Т254G/Н310Е/Н433Е/Н435Е, у которого нарушено связывание с FcRn, и одним мономером, содержащим мономер Fc дикого типа.
Мутационные исследования Fcγ человека были выполнены на некоторых остатках, которые важны для связывания с FcRn и которые, как было показано, имеют увеличенное время полужизни в сыворотке крови. Hinton et al. в Fcγ человека мутировали три остатка по отдельности от 19 общих аминокислот. Hinton et al. обнаружили, что некоторые точечные мутации двойного мутанта повышали сродство при связывании FcRn (Hinton et al., 2004, J. Biol. Chem. 279(8):6213-6216, работа включена путем отсылки во всей своей полноте). Две мутации повышали время полужизни у обезьян. Shields et al. мутировали остатки, почти что исключительно на Ala и изучали их связывание с FcRn и FcγR's (Shields et al., 2001, J Biol Chem 276:6591-6604, работа включена путем отсылки во всей своей полноте).
Чтобы отобрать Fc-мутанты, которые связывают FcRn с повышенным сродством, Dall'Acqua et al. использовали фаговый дисплей (Dall'Acqua et al., 2002, J. Immunol. 169:5171-5180, работа включена путем отсылки во всей своей полноте). Отобранные последовательности ДНК были двойными и тройными мутантами. В ссылке представлены белки, кодируемые многими отобранными последовательностями, и обнаружены некоторые белки, которые связываются с FcRn более прочно, чем Fc дикого типа.
Для введения антител и белков, слитых с Fc, в качестве терапевтических средств необходимы инъекции с предписанной частотой, зависящей от характеристик выведения и времени полужизни белков. Более длительное время полужизни in vivo делает возможным более редкие инъекции или сниженные дозировки, что, несомненно, выгодно. Хотя мутации, сделанные ранее в Fc-домене, привели к получению некоторых белков с повышенным сродством при связывании с FcRn и увеличенными временами полужизни in vivo, эти мутации не выяснили оптимальные мутации и увеличенное время полужизни in vivo.
Ключевой чертой Fc-участка является консервативное N-гликозилирование, показанное на фигуре 1, которое обнаруживают на остатке N297. Этот углевод или олигосахарид, как его иногда называют, играет особую структурную и функциональную роль в антителе, что является одной из принципиальных причин, по которой антитела должны продуцироваться с применением экспрессионных систем млекопитающих (Umana et al., 1999, Nat Biotechnol 17:176-180; Davies et al., 2001, Biotechnol Bioeng 74:288-294; Mimura et al., 2001, J Biol Chem 276:45539-45547; Radaev et al., 2001, J Biol Chem 276:16478-16483; Shields et al., 2001, J Biol Chem 276:6591-6604; Shields et al., 2002, J Biol Chem 277:26773-26740; Simmons et al., 2002, J Immunol Methods 263:133-147). Однако влияние углевода незначительно, если имеется любой специфический контакт с FcγR (Radaev et al., 2001, J Biol Chem 276:16469-16477; Radaev et al., 2001, J Biol Chem 276:16469-16477 и Krapp et al., 2003, J. Mol. Biol. 325:979-989, работы включены путем отсылки во всей своей полноте).
Антитела были разработаны для терапевтического применения. Наиболее типичные публикации, имеющие отношение к такому лечению включают Chamow et al., 1996, Trends Biotechnol 14:52-60; Ashkenazi et al., 1997, Curr Opin Immunol 9:195-200; Cragg et al., 1999, Curr. Opin. Immunol. 11:541-547; Glennie et al., 2000, Immunol. Today 21:403-410; McLaughlin et al., 1998, J. Clin. Oncol. 16:2855-2833 и Cobleigh et. al., 1999, J. Clin. Oncol. 17:2639-2648, все работы включены путем отсылки во всей своей полноте. В настоящее время для противораковой терапии любое небольшое улучшение в коэффициенте смертности определяется как успех. Определенные IgG-варианты, раскрытые здесь, по меньшей мере, частично, увеличивают эффективность антител в ограничении дальнейшего роста или разрушения раковых клеток-мишеней.
Противоопухолевая эффективность антител реализуется через повышение их способности опосредовать цитотоксические эффекторные функции, такие как ADCC, ADCP и CDC. Примеры включают работы Clynes et al., 1998, Proc Natl Acad Sci USA 95:652-656; Clynes et al., 2000, Nat Med 6:443-446 и Cartron et al., 2002, Blood 99:754-758, все работы включены путем отсылки во всей своей полноте.
IgG1 человека - антитело, наиболее часто применяемое в терапевтических целях, и большая часть конструкторских исследований была выполнена в этом контексте. Различные изотипы класса IgG, включающие IgG1, IgG2, IgG3 и IgG4, однако, имеют уникальные физические, биологические и клинические свойства. В этой области техники существует необходимость разрабатывать такие варианты, чтобы улучшить связывание с FcRn и/или увеличить время полужизни in vivo по сравнению с природными полипептидами IgG. Настоящая заявка отвечает всем этим и другим требованиям.
Раскрытие изобретения
Настоящее изобретение раскрывает создание новых вариантов Fc-доменов, включая те, которые обнаружены в антителах, слитых Fc и иммуноадгезинах, которые обладают повышенным связыванием с рецептором FcRn и повышенным удержанием в сыворотке крови in vivo. Дополнительный аспект изобретения - повышение связывания с FcRn по сравнению с диким типом, в особенности при низких значениях рН, около 6,0 для того, чтобы обеспечить связывание Fc/FcRn в эндосомах. Дополнительный аспект настоящего изобретения - предпочтительное связывание сконструированных вариантов при рН около 6 по сравнению с их связыванием при рН около 7,4 для того, чтобы облегчить высвобождение Fc в кровь после клеточного рециклирования.
Еще один аспект настоящего изобретения имеет отношение к конструированию вариантов Fc со сниженным связыванием с FcRn и сниженным временем полужизни in vivo. Такие белки, включающие мутации для снижения сродства к FcRn и/или времени полужизни in vivo, полезны для многих вариантов терапии и диагностики, включая доставку и мониторинг радиоактивных лекарственных средств, где в идеальном случае время полужизни радиоактивной метки приблизительно равно времени полужизни in vivo ее белкового конъюгата.
Еще один аспект настоящего изобретения имеет отношение к изменению связывания Fc-домена с FcR's, например с FcgRI, FcgRIIa, FcgRIIb, FcgRIIIa у людей. Эти рецепторы отвечают за индукцию различных эффекторных функций антител. Поэтому еще один аспект изобретения имеет отношение к изменению эффекторных функций Fc-домена, таких как зависимая антитело-зависимая клеточно-опосредованная цитотоксичность (ADCC), зависимая от комплемента цитотоксичность (CDC) и антитело-зависимый клеточно-опосредованный фагоцитоз (ADCP).
Еще один аспект настоящего изобретения имеет отношение к Fc-вариантам, которые обладают и измененным связыванием с FcRn, и измененным связыванием с Fcg, чтобы повлиять и на время полужизни in vivo, и на эффекторные функции белка, включающего Fc. Например, эти варианты могут иметь увеличенное время полужизни in vivo, а также улучшенную ADCC. Варианты, например, могут иметь увеличенное время полужизни in vivo и сниженную CDC.
Еще в одном аспекте изобретение обеспечивает рекомбинантные нуклеиновые кислоты, кодирующие вариантные Fc-белки, экспрессионные векторы и клетки хозяина.
В дополнительном аспекте изобретение обеспечивает способы продуцирования белка, включающего Fc, включающие культивирование клеток-хозяев изобретения в условиях, подходящих для экспрессии белка.
В еще одном аспекте изобретение обеспечивает фармацевтические композиции, включающие вариантный Fc-белок изобретения и фармацевтический носитель.
Еще в одном аспекте изобретение обеспечивает способы лечения нарушений, включающие введение пациенту белка, включающего вариантный Fc изобретения.
В дополнительном аспекте изобретение предоставляет Fc-участок варианта в исходном Fc-полипептиде, включающий, по меньшей мере, одну модификацию в Fc-участке указанного исходного полипептида, где указанный вариантный белок демонстрирует измененное связывание с FcRn по сравнению с исходным полипептидом, и где указанный Fc-вариант включает, по меньшей мере, одну модификацию, выбираемую из группы, состоящей из:

где нумерация соответствует индексу EU и символ «^» означает вставку после указанного положения, а символ «#» означает делецию в этом указанном положении.
Еще в одном аспекте изобретение обеспечивает Fc-варианты, включающие, по меньшей мере, одну модификацию, выбираемую из группы, состоящей из:


В дополнительном аспекте изобретение обеспечивает Fc-варианты, включающие, по меньшей мере, одну модификацию, выбираемую из группы, состоящей из:

Краткое описание чертежей
Фигура 1: Структура и функция антител. Показана модель полноразмерного человеческого антитела IgG1, смоделированного с использованием структуры гуманизированного фрагмента Fab с кодом доступа 1СЕ1 в базе данных pdb (James et al., 1999, J Mol Biol 289:293-301, работа включена путем отсылки во всей своей полноте) и структуры Fc-фрагмента человеческого IgG1 с кодом доступа 1DN2 в базе данных pdb (DeLano et al., 2000, Science 287:1279-1283, работа включена путем отсылки во всей своей полноте). Подвижный шарнир, который связывает участки Fab и Fc, не показан. IgG1 представляет собой гомодимер, состоящий из гетеродимеров, составленных из двух легких цепей и двух тяжелых цепей. Домены Ig, которые составляют антитело, отмечены и включают VL и CL в легкой цепи и VH, С-гамма-1 (Cγ1), С-гамма-2 (Сγ2) и С-гамма-3 (Сγ3) для тяжелой цепи. Fc-участок отмечен. Места связывания важных белков отмечены и включают место связывания антигена в вариабельной области и места связывания FcγRs, FcRn, C1q и протеинов А и G в участке Fc.
Фигура 2: Последовательности IgG человека, применяемые в настоящем изобретении, с нумерацией в соответствии с индексом EU базы данных Kabat et al.
Фигура 3: Пример последовательностей IgG человека и грызунов, применяемых в настоящем изобретении, с нумерацией в соответствии с индексом EU базы данных Kabat et al.
Фигура 4: Пример последовательностей тяжелых цепей FcRn человека и грызунов, применяемых в настоящем изобретении.
Фигура 5: Пример последовательностей бета-2-микроглобулина человека и грызунов, применяемых в настоящем изобретении.
Фигура 6: Модель комплекса Fc/FcRn человека, созданная на основании структур крысы (Burmeister et al., 1994, Nature, 372:379-383; Martin et al., 2001, Mol Cell 7:867-877, обе работы включены путем отсылки во всей своей полноте). Некоторые остатки гистидина показаны в виде заполняющих пространство атомов в цепях FcRn (светлосерые) и в Fc-полипептиде (темно-серые).
Фигура 7: Иллюстрация некоторых концепций, применяемых при конструировании вариантов, включающих вставки и делеции.
Фигура 8: Варианты настоящего изобретения.
Фигура 9: Варианты настоящего изобретения.
Фигура 10: Варианты настоящего изобретения.
Фигура 11: Схема вектора pcDNA3.1 Zeo+, который можно применять в конструкциях Fc-вариантов.
Фигура 12: Данные по конкуренции связывания Fc дикого типа и Fc-вариантов настоящего изобретения с FcRn. Ha каждой панели Fc-варианты настоящего изобретения показаны в виде левой кривой (красной или темно-серой), а трастузумаб дикого типа показан в виде правой кривой (синей или светло-серой).
Фигура 13: Обобщенные результаты FcRn-связывающих свойств Fc-вариантов. Колонки справа налево показывают модификации связывания FcRn, используемый иммуноглобулин, другие модификации, относительное сродство к FcRn, измеренное с помощью конкурентного анализа AlphaScreen™, по сравнению с диким типом (значение медианы), число проведенных анализов и идентификационный номер белка. Относительное сродство FcRn, которое выше, чем 1,0, показывает улучшение связывания по сравнению с диким типом.
Фигура 14: Данные по связыванию FcRn Fc-вариантами настоящего изобретения. Fc-варианты настоящего изобретения представляют собой алемтузумаб и трастузумаб. Показано кратное усиление связывания по сравнению с диким типом.
Фигура 15: Обобщенные эксперименты, проведенные методом поверхностного плазменного резонанса с Fc-вариантами, имеющими улучшенное связывание с FcRn. Гистограмма показывает кратность увеличения сродства при связывании с FcRn каждого варианта по отношению к Fc-домену дикого типа.
Фигура 16: Эксперименты, проведенные методом поверхностного плазменного резонанса с антителами дикого типа и вариантами настоящего изобретения. Показанные кривые представляют собой ассоциацию и диссоциацию Fc-вариантного антитела с FcRn при рН 6,0.
Фигура 17: Анализы связывания Fc-вариантов настоящего изобретения с FcRn. Показаны результаты анализа прямого связывания методом AlphaScreen™ при рН 6,0 (а и b) и при рН 7,0 (с).
Фигура 18: Анализы связывания Fc-вариантов настоящего изобретения с FcRn. Приведены единицы поверхностного плазмонного резонанса, полученные при связывании Fc-варианта с прикрепленным на поверхности FcRn.
Осуществление изобретения
Настоящее изобретение раскрывает создание новых вариантов Fc-доменов, включая те, которые обнаружены в антителах, слитых Fc и иммуноадгезинах, которые обладают усиленным связыванием с FcRn-рецептором. Как указано здесь, связывание с FcRn приводит к их более длительному удержанию в сыворотке крови in vivo.
Чтобы увеличить удержание Fc-белков in vivo, увеличение сродства при связывании должно быть приблизительно при рН 6,0 без сопутствующего увеличения сродства приблизительно при рН 7,4. Хотя это еще выясняется, полагают, что Fc-участки имеют более длительное время полужизни in vivo, так как связывание с FcRn при рН 6 в эндосоме приводит к захвату Fc (Ghetie and Ward, 1997, Immunol. Today, 18(12):592-598, работа включена путем отсылки во всей своей полноте). Эндосомальный компартмент затем вновь возвращает Fc на клеточную поверхность. После того, как компартмент открывается во внеклеточное пространство, более высокие значения рН около 7,4 индуцируют высвобождение Fc назад в кровяное русло. Dall' Acqua et al. показали, что Fc-мутанты с усиленным связыванием FcRn при рН 6 и 7,4 в действительности имеют сниженные концентрации в сыворотке крови и то же самое время полужизни, что и Fc дикого типа (Dall' Acqua et al., 2002, J. Immunol., 169:5171-5180, работа включена путем отсылки во всей своей полноте). Считают, что повышенное сродство Fc к FcRn при рН 7,4 не позволяет Fc высвобождаться обратно в кровь. Таким образом, мутации Fc, которые будут повышать время полужизни Fc in vivo, будут в идеальном случае увеличивать связывание FcRn при низких значениях рН, и в то же время все еще обеспечивать высвобождение Fc при более высоких значениях рН. Аминокислота гистидин изменяет свой заряд в диапазоне рН от 6,0 до 7,4. Следовательно, не удивительно обнаружить остатки His в важных положениях в комплексе Fc/FcRn (фигура 6).
На Фигурах 8-10 и 17-19 представлены дополнительные данные по вариантам, раскрытым в первоначально поданном описании. На Фигурах 20, 27 и 30 приведены данные, полученные in vivo.
На Фигуре 20 представлено обобщение по результатам измерения аффинности связывания Fc-вариантов настоящего изобретения с FcRn человека, макаки и мыши, полученные методом поверхностного плазменного резонанса (SPR). Значения, превышающие единицу, указывают на увеличение связывания Fc-варианта настоящего изобретения с FcRn. Измерения проводились с использованием аппроксимации кривых, полученных методом SPR, в соответствии с моделью связывания по Лэнгмюру при стехиометрии связывания 1:1.
На Фигуре 27 представлены результаты, полученные in vivo. Показаны значения концентрации антител дикого типа (WT) и вариантов в сыворотке мышей с нокином человеческого FcRn. Использовались следующие антитела против VEGF: антитела дикого типа (WT) - открытые квадратики, V308F - закрытые квадратики, P257L - закрытые треугольники, и P257N - крестики.
На Фигуре 30 представлены данные по связыванию антител дикого типа и вариантов с FcRn на поверхности клеток 293Т.
Дополнительный аспект изобретения - повышение связывания с FcRn по сравнению с диким типом, в особенности при низких значениях рН, около 6,0, чтобы облегчить связывание Fc/FcRn в эндосоме. Также в заявке раскрыты Fc-варианты с измененным связыванием с FcRn и измененным связыванием с другим классом Fc-рецепторов, FcγR's, так как было показано, что различное связывание с FcγRs, в особенности повышенное связывание с FcγRIIIb и пониженное связывание с FcγRIIb, приводит к увеличению эффективности.
Определения
Чтобы заявку можно было бы более полно понять, ниже приведены некоторые определения. Разумеется, что такие определения включают грамматические эквиваленты.
Термин «ADCC» или «антитело-зависимая клеточно-опосредованная цитотоксичность», который используют здесь, означает опосредованную клетками реакцию, в которой неспецифические цитотоксические клетки, экспрессирующие FcγRs, распознают антитела, связанные на клетке-мишени, и, впоследствии, вызывают лизис клетки-мишени.
Термин «ADCP» или «антитело-зависимый клеточно-опосредованный фагоцитоз», который используют здесь, означает опосредованную клетками реакцию, в которой неспецифические цитотоксические клетки, экспрессирующие FcγRs, распознают антитела, связанные на клетке-мишени, и впоследствии вызывают фагоцитоз клетки-мишени.
Термин «модификация аминокислот» означает здесь замену, вставку и/или делецию в полипептидной последовательности.
Термины «замена аминокислот» или «замена» означают здесь замещение аминокислоты в определенном положении в исходной полипептидной последовательности на другую аминокислоту. Например, замена E272Y относится к вариантному полипептиду, в данном случае к Fc-варианту, в котором глутаминовую кислоту в положении 272 замещают на тирозин.
Термин «вставка аминокислоты» или «вставка» означает здесь добавление аминокислоты в определенное положение в исходной полипептидной цепи. Например, -233Е или ^233Е обозначает вставку глутаминовой кислоты после положения 233 и до положения 234. Кроме того, -233ADE или ^233ADE обозначает вставку AlaAspGlu после положения 233 и перед положением 234.
Термин «делеция аминокислоты» или «делеция», который используют здесь, означает удаление аминокислоты, в частности, в определенном положении исходной полипептидной последовательности. Например, Е233- или Е233# обозначают делецию глутаминовой кислоты в положении 233. Кроме того, EDA233- или EDA233# делецию удаление последовательности GluAspAla, которая начинается от положения 233.
Термин «вариантный белок», или «вариант белка», или «вариант», который здесь используют, означает белок, которая отличается от исходного белка, по меньшей мере, одной аминокислотной модификацией. Вариант белка может относиться к белку самому по себе, к композиции, которая включает белок, или к последовательности аминокислот, которая его кодирует. Предпочтительно, когда вариант белка имеет, по меньшей мере, одну аминокислотную модификацию по сравнению с исходным белком, например приблизительно от одной до приблизительно десяти аминокислотных модификаций и предпочтительно приблизительно от одной до приблизительно пяти аминокислотных модификаций по сравнению с исходным белком. Последовательность варианта белка здесь предпочтительно будет иметь, по меньшей мере, 80%-ную гомологию с последовательностью исходного белка, наиболее предпочтительно, по меньшей мере, примерно 90%-ную гомологию, более предпочтительно, по меньшей мере, примерно 95%-ную гомологию. Вариантный белок может относиться к вариантному белку самому по себе, к композиции, которая включает вариант белка, или к последовательности аминокислот, которая его кодирует. Соответственно термин «вариант антитела» или «вариантное антитело», который здесь используют, обозначает антитело, которое отличается от исходного антитела, по меньшей мере, одной аминокислотной модификацией, термин «IgG-вариант» или вариантный «IgG», используемый здесь, обозначает антитело, которое отличается от исходного IgG, по меньшей мере, одной аминокислотной модификацией, и термин «вариант иммуноглобулина» или «вариантный иммуноглобулин», который используют здесь, обозначает иммуноглобулиновую последовательность, которая отличается от исходной иммуноглобулиновой последовательности, по меньшей мере, одной аминокислотной модификацией. Варианты могут включать неприродные аминокислоты. Примеры включают US 6586207; WO 98/48032; WO 03/073238; US 2004-021988 A1; WO 05/35727A2; WO 05/74524A2; J.W.Chin et al., (2002) Journal of the AmericanChemical Society 124:9026-9027; J.W.Chin & P.G.Schultz, (2002), ChemBioChem 11:1135-1137; J.W.Chin et al., (2002), PICAS United States of America 99:11020-11024; и L.Wang & P.G.Schultz, (2002), Chem. 1-10, все работы включены путем отсылки во всей своей полноте).
Термин «белок» здесь означает, по меньшей мере, две ковалентно связанные аминокислоты, термин включает в себя белки, полипептиды, олигопептиды и пептиды. Пептидильная группа может включать встречающиеся в природе аминокислоты и пептидные связи или синтетические пептидоподобные структуры, то есть «аналоги», например пептоиды (см. Simon et al., PNAS USA 89(20):9367 (1992), работа включена путем отсылки во всей своей полноте). Аминокислоты могут быть или природными, или не встречающимися в природе, как это хорошо известно в этой области техники. Например, гомофенилаланин, цитруллин и норлейцин рассматриваются как аминокислоты, применяемые в целях настоящего изобретения, и могут применяться как D- и L- (R или S) конфигурации аминокислот. Варианты настоящего изобретения могут включать модификации, которые включают применение ненасыщенных аминокислот, в том числе и применение, например, технологий, развиваемых Schultz et al., включая, но не ограничиваясь методами, описанными Cropp&Schultz, 2004, Trends Genet. 20(12):625-30, Anderson et al., 2004, Proc. Natl. Acad. Sci. USA 101(2):7566-71, Zhang et al., 2003, 303(5656):371-3 и Chin et al., 2003, Science, 301(5635):964-7, все работы включены путем отсылки во всей своей полноте). Кроме того, полипептиды могут включать получение синтетических производных одной или нескольких боковых или основных цепей, гликозилирование, ПЭГилирование, циклическую перестановку, циклизацию, линкеры к другим молекулам, слияние с белками или белковыми доменами и присоединение белковых тагов или меток.
Термин «остаток», который здесь используют, означает положение в белке и ассоциированную с ним аминокислотную идентичность. Например, аспарагин-297 (также называемый Asn297, также называемый N297) является остатком антитела IgG1 человека.
Термин «Fab» или «Fab-участок», используемый здесь, означает полипептиды, которые включают VH-, CH1-, VL- и CL-домены иммуноглобулинов. Fab может иметь отношение к этому участку в изолированном виде или этот участок в контексте полноразмерного антитела или фрагмента антитела.
Термин «модификация подкласса IgG», используемый здесь, означает аминокислотную модификацию, которая превращает аминокислоту одного изотипа IgG в соответствующую аминокислоту в другом, выровненном по последовательности изотипе IgG. Например, так как IgG1 в EU-положении 296 включает тирозин, a IgG2 - фенилаланин, то замена F296Y в IgG2 означает модификацию подкласса IgG.
Термин «не встречающаяся в природе модификация», используемый здесь, означает аминокислотную модификацию, которая не является изотипической. Например, так как ни один из IgG не включает глутаминовую кислоту в положении 332, замена I332E в IgG1, IgG2, IgG3 или IgG4 рассматриваются как не встречающаяся в природе модификация.
Термины «аминокислота» или «идентичность аминокислоты» используют здесь для обозначения одной из 20 встречающихся в природе аминокислот или любых их неприродных аналогов, которые могут присутствовать в специфическом определенном положении.
Термин «эффекторная функция» здесь означает биохимическое явление, которое происходит в результате взаимодействия Fc-участка антитела с Fc-рецептором или лигандом. Эффекторные функции включают, но не ограничиваются ADCC, ADCP и CDC.
«Эффекторная клетка» здесь означает клетку иммунной системы, которая экспрессирует один или несколько Fc-рецепторов и является посредником для одной или нескольких эффекторных функций. Эффекторные клетки включают, но не ограничиваются только ими, моноциты, макрофаги, нейтрофилы, дендритные клетки, эозинофилы, тучные клетки, тромбоциты, В-клетки, большие гранулярные лимфоциты, клетки Лангерганса, натуральные клетки-киллеры (NK) и γδТ-клетки, которые могут происходить из любого организма, включая, но не ограничиваясь людьми, мышами, крысами, кроликами и обезьянами.
Термин «Fc-лиганд IgG», который здесь используют, означает молекулу, предпочтительно полипептид из любого организма, который связывает Fc-участок антитела IgG с формированием комплекса Fc/Fc-лиганд. Fc-лиганды включают, но не ограничиваются только ими, FcγRI, FcγRII, FcγRIII, FcγRn, C1q, С3, маннан-связывающий лектин, маннозный рецептор, стафилококковый белок А, стрептококковый белок G и вирусный FcγR. Fc-лиганды также включают гомологи Fc-рецептора (FcRH), которые относятся к семейству Fc-рецепторов, гомологичных FcγRs (Davis et al., 2002, Immunological Reviews 190:123-136, работа включена путем отсылки во всей своей полноте). Fc-лиганды могут включать до сих пор не открытые молекулы, которые связываются с Fc. Особыми Fc-лигандами IgG являются FcRn и Fc-гамма-рецепторы. Термин «Fc-лиганд», который здесь используют, означает молекулу, предпочтительно полипептид из любого организма, который связывает Fc-участок антитела с формированием комплекса Fc/Fc-лиганд.
Термины «Fc-гамма-рецептор» или «FcγR», которые здесь используют, означают любого представителя семейства белков, которые связывают Fc-фрагмент антитела IgG и которые кодируются геном FcγR. У людей это семейство включает, но не ограничивается только ими, FcγRI (CD64), включая изоформы FcγRIa, FcγRIb и FcγRIc; FcγRII (CD32), включая изоформы FcγRIIa (включая аллотипы Н131 и R131), FcγRIIb (включая FcγRIIb-1 и FcγRIIb-2) и FcγRIIc; и FcγRIII (CD16), включая изоформы FcγRIIIa (включая аллотипы V158 и F158), и FcγRIIIb (включая аллотипы FcγRIIIb-NA1 и FcγRIIIb-NA2) (Jefferis et al., 2002, Immunol Lett 82:57-65, работа включена путем отсылки во всей своей полноте), а также любые до сих пор не открытые человеческие FcγRs или изоформы или аллотипы FcγR. FcγR может происходить из любого организма, включающего людей, мышей, крыс, кроликов и обезьян, но не ограниченного ими. Мышиные FcγRs включают, но не ограничиваются перечисленными, FcγRI (CD64), FcγRII (CD32), FcγRIII (CD16) и FcγRIII-2 (CD 16-2), так же как и любые до сих пор не открытые мышиные FcγRs или изоформы или аллотипы FcγR.
Термин «FcRn» или «неонатальный Fc-рецептор», используемый здесь, означает белок, который связывает Fc-участок антитела IgG и кодируется, по меньшей мере, частично геном FcRn. FcRn может происходить из любого организма, включая людей, мышей, крыс, кроликов и обезьян, но не ограничивается только ими. Как известно в этой области техники, функциональный FcRn-белок включает два полипептида, часто называемых тяжелой цепью и легкой цепью. Легкая цепь - бета-2-микроглобулин, а тяжелая цепь кодируется геном FcRn. Если в тексте специально не оговорено, FcRn или FcRn-белок относятся к комплексу тяжелой цепи FcRn с бета-2-микроглобулином. Последовательности FcRn, представляющие особый интерес, в особенности последовательности человека, показаны на чертежах.
Термин «исходный полипептид», который используют в тексте, означает полипептид, который значительно модифицирован для создания варианта. Исходный полипептид может быть полипептидом, встречающимся в природе, или вариантной или сконструированной разновидностью встречающегося в природе полипептида. Исходный полипептид может относиться как к полипептиду самому по себе, к композиции, которая включает исходный полипептид, или к последовательности аминокислот, которая его кодирует. Соответственно, термин «исходный иммуноглобулин», который здесь используют, означает немодифицированный иммуноглобулиновый полипептид, который затем модифицируют для создания варианта, а термин «исходное антитело», который здесь используют, означает немодифицированное антитело, которое затем модифицируют для создания вариантного антитела. Следует отметить, что «исходное антитело» включает известные имеющиеся в продаже антитела, рекомбинантно-продуцируемые антитела, как отмечено ниже.
Термин «положение», который здесь используют, означает определенное место в последовательности белка. Положения могут быть пронумерованы последовательно или в соответствии с установленным форматом, например в соответствии с индексом EU в базе данных Kabat. Например, положение 297 - это положение в человеческом антителе IgG1.
Термин «антиген-мишень», который здесь используют, означает молекулу, которая специфически связывается с вариабельной областью определенного антитела. Антиген-мишень может быть белком, углеводом, липидом или другим химическим веществом.
Термин «клетка-мишень», который здесь используют, означает клетку, которая экспрессирует антиген-мишень.
Термин «вариабельная область», который здесь используют, означает часть иммуноглобулина, которая включает один или несколько Ig-доменов, в основном кодируемых любым из Vκ, Vλ- и/или VH- генов, которые содержат генетические локусы каппа, лямбда и локус тяжелой цепи иммуноглобулина соответственно.
Термин «дикий тип или WT», который здесь используют, означает аминокислотную последовательность или нуклеотидную последовательность, которые обнаружены в природе, включая аллельные вариации. WT-белок имеет аминокислотную последовательность или нуклеотидную последовательность, которые не являются преднамеренно модифицированными.
Настоящее изобретение направлено на антитела, которые демонстрируют модулированное связывание с FcRn (модуляция, включающая повышенное или пониженное связывание). Например, в некоторых случаях повышенное связывание приводит к клеточному рециклированию антитела и таким образом к увеличенному времени полужизни, например, в случае терапевтических антител. Альтернативно, требуется сниженное связывание с FcRn, например в случае диагностических антител или терапевтических антител, которые содержат радиоактивную метку. Кроме того, в настоящем изобретении находят свое применение антитела, демонстрирующие повышенное связывание с FcRn и измененное связывание с другими Fc-рецепторами, например с FcγRs.
Антитела
Настоящее изобретение направлено на антитела, которые включают аминокислотные модификации, которые модулируют связывание с FcRw. Особый интерес представляют антитела, которые минимально включают Fc-участок, или его функциональный вариант, которые проявляют увеличенное средство связывания с FcRn при пониженных рН и не проявляют в существенной мере измененного связывания при более высоких рН.
Традиционные структурные единицы антител обычно включают тетрамер. Каждый тетрамер обычно состоит из двух идентичных пар полипептидных цепей, каждая пара имеет одну «легкую» (обычно с молекулярной массой около 25 кДа) и одну «тяжелую» цепь (обычно с молекулярной массой около 50-70 кДа). У человека легкие цепи классифицируются как легкие цепи каппа и лямбда. Тяжелые цепи классифицируют как цепи мю, дельта, гамма, альфа или эпсилон и определяют такие изотипы антител, как IgM, IgD, IgG, IgA и IgE соответственно. Иммуноглобулин IgG имеет несколько подклассов, включающих, но неограниченных IgG1, IgG2, IgG3 и IgG4. Иммуноглобулин IgM имеет подклассы, включающие, но не ограниченные IgM1 и IgM2. Таким образом, термин «изотип», который здесь используют, означает любой из подклассов иммуноглобулинов с определенными химическими и антигенными характеристиками их константных областей. Известными изотипами иммуноглобулинов человека являются IgG1, IgG2, IgG3, IgG4, IgA1, IgA2, IgM1, IgM2, IgD и IgE.
N-концевая часть каждой цепи включает вариабельную область, состоящую приблизительно из 100-110 или более аминокислот, которая исходно отвечает за узнавание антигена. В вариабельной области в каждом V-домене тяжелой цепи и легкой цепи собраны по три петли для формирования антиген-связывающего участка. Каждая петля относится к гипервариабельному участку (в дальнейшем именуемому как «CDR»), в котором различие аминокислотной последовательности наиболее значительно.
Карбокси-концевая часть каждой цепи определяет константную область, исходно отвечающую за эффекторную функцию. Kabat et al. собрали множество первичных последовательностей вариабельных областей тяжелых цепей и легких цепей. На основании степени консервативности последовательностей они классифицировали индивидуальные первичные последовательности на CDR и каркасную часть и создали их список (см. SEQUENCES OF IMMUNOLOGICAL INTEREST, 5™ edition, NIH publication. No. 91-3242, Kabat et al., работа включена путем отсылки во всей своей полноте).
В IgG-подклассе иммуноглобулинов в тяжелой цепи существует несколько иммуноглобулиновых доменов. Термин «иммуноглобулиновый (Ig) домен» здесь означает участок иммуноглобулина, имеющий обособленную третичную структуру. В настоящем изобретении представляют интерес домены тяжелых цепей, включая константные тяжелые (СН) домены, и домены шарнирного участка. В контексте IgG-антител каждый из IgG-изотипов имеет по три СН-участка. Соответственно «СН»-домены в контексте IgG представляют собой: «СН1», относящийся к положениям 118-220 в соответствии с EU-индексом в базе данных Kabat. «CH2» относится к положениям 237-340 в соответствии с EU-индексом в базе данных Kabat и «СН3» относится к положениям 341-447 в соответствии с EU-индексом в базе данных Kabat.
Другой тип Ig-домена тяжелой цепи относится к шарнирной области. Термины «шарнир» или «шарнирный участок», или шарнирный участок антитела», используемые здесь, относятся к подвижному полипептиду, включающему аминокислоты между первым и вторым константными доменами антитела. Структурно СН1-домен IgG заканчивается в EU-положении 220, а СН2-домен IgG начинается у остатка в EU-положении 237. Таким образом, в IgG шарнир антитела определен здесь как домен, включающий положения от 221 (D221 в IgG1) до 236 (G236 в IgG1), где нумерация соответствует EU-индексу в базе данных Kabat. В некоторых воплощениях, например в контексте Fc-участка, включен нижний шарнир, обычно имеющий отношение к положениям от 226 или 230.
Особый интерес в настоящем изобретении представляют Fc-участки. Термины «Fc» или «Fc-участок» означают здесь полипептид, включающий константную область антитела за исключением первого константного участка иммуноглобулинового домена и в некоторых классах часть шарнира. Таким образом, Fc относится к двум последним константным участкам иммуноглобулиновых доменов IgA, IgD и IgG, и к последним трем константным участкам иммуноглобулиновых доменов IgE и IgM и к подвижному N-концевому шарниру этих доменов. Fc IgA и IgM могут включать цепь J. Как показано на фигуре 1 для IgG, Fc включает иммуноглобулиновые домены С-гамма-2 и С-гамма-3 (Cg2 и Cg3) и участок нижнего шарнира между С-гамма-1 (Cg1) и С-гамма-2 (Cg2). Несмотря на то, что поверхности раздела Fc-участка могут различаться, Fc-участок тяжелой цепи человеческого IgG обычно включает остатки С226 или Р230 на С-конце, где нумерация соответствует индексу EU базы данных Kabat. Fc может иметь отношение к этому участку в изолированном виде или к участку в контексте Fc-полипептида, как описано ниже. Термин «Fc-полипептид», используемый здесь, означает полипептид, который включает весь или часть Fc-участка. Fc-полипептиды включают антитела, слитые Fc-белки, изолированные Fc и Fc-фрагменты.
В некоторых воплощениях антитела являются полноразмерными. Под «полноразмерным антителом» здесь подразумевают структуру, которая создает природную биологическую форму антитела, включающую вариабельную и константную области, включающие одну или несколько модификаций, как здесь было отмечено.
Альтернативно, антитела могут представлять собой множество структур, включающих, но не ограниченных только ими, фрагменты антител, моноклональные антитела, биспецифические антитела, миниантитела, доменные антитела, синтетические антитела (иногда называемые здесь «миметиками антител»), химерные антитела, гуманизированные антитела, слитые антитела (иногда называемые «конъюгатами антител») и их фрагменты соответственно.
Фрагменты антител
В одном воплощении антитело представляет собой фрагмент антитела. Особый интерес представляют антитела, которые включают Fc-участки, слитые Fc и константную область тяжелой цепи (СН1-шарнир-СН2-СН3), также снова включающие слитые константные тяжелые участки.
Специфические фрагменты антител включают, но не ограничиваются только ими, (i) Fab-фрагмент, состоящий из VL-, VH-, CL- и СН1-доменов, (ii) Fd-фрагмент, состоящий из VH- СН1-доменов, (iii) Fv-фрагмент, состоящий из VL- и VH-доменов отдельного антитела; (iv) dAb-фрагмент (Ward et al., 1989, Nature 341:544-546, работа включена путем отсылки во всей своей полноте), который состоит из отдельного вариабельного участка, (v) изолированные участки CDR, (vi) F(аb')2-фрагменты, бивалентный фрагмент, включающий два связанных Fab-фрагмента, (vii) моноцепочечные молекулы Fv (scFv), где VH- и VL-домены связаны через пептидный линкер, что позволяет двум доменам ассоциировать с образованием антиген-связывающего участка (Bird et al., 1988, Science 242:423-426, Huston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-5883, работы включены путем отсылки во всей своей полноте), (viii) биспецифичный одноцепочечный Fv (WO 03/11161, включенный сюда путем отсылки) и (ix) димеры и тримеры малых фрагментов антител, мультивалентные или мультиспецифичные фрагменты, сконструированные путем слияния генов (Tomlinson et al., 2000, Methods Enzymol. 326:461-479; WO94/13804; Hollinger et al., 1993, Proc. Natl. Acad. Sci. USA 90:6444-6448, работы включены путем отсылки во всей своей полноте). Фрагменты антител могут быть модифицированы. Например, молекулы могут быть стабилизированы путем включения дисульфидных мостиков, связывающих VH- и VL-домены (Reiter et al., 1996, Nature. Biotech. 14:1239-1245, работа включена путем отсылки во всей своей полноте).
Химерные и гуманизированные антитела
В некоторых воплощениях компоненты остова могут быть смесью из различных видов. Собственно, если антитело является каким-либо антителом, такое антитело может быть химерным антителом и/или гуманизированным антителом. В основном, как «химерные антитела», так и «гуманизированные антитела» относятся к антителам, которые объединяют области от более чем одного вида. Например, «химерные антитела» обычно включают вариабельную область (области) от мыши (или, в некоторых случаях, от крысы) и константную область (области) от человека. «Гуманизированные антитела» обычно относятся к антителам, не являющимся человеческими, которые включают каркасные участки вариабельного домена с заменой на последовательности, обнаруженные в человеческих антителах. Обычно в гуманизированном антителе антитело целиком, за исключением CDR's, кодируется полинуклеотидом человеческого проихождения или идентично такому антителу, за исключением его CDR's. CDR's, некоторые или все, которые кодируются нуклеиновыми кислотами, происходящими из организмов, не являющихся человеком, пересаживают в бета-складчатый каркасный участок вариабельного участка человеческого антитела для создания антитела, специфичность которого определяется пересаженными CDR's. Создание таких антител описано, например, в WO 92/11018, Jones et al., 1986, Nature 321:522-525; Verhoeyen et al., 1988, Science, 239:1534-1536, работы включены путем отсылки во всей своей полноте. «Обратная мутация» выбранных аминокислотных остатков акцепторного каркасного участка на соответствующие донорные аминокислотные остатки часто необходима для восстановления аффинности, которая теряется в исходной конструкции при пересадке (US 5530101; US 5585089; US 5693761; US 5693762; US 6180370; US 5859205; US 5821337; US 6054297; US 6407213, все работы включены путем отсылки во всей своей полноте). Оптимально гуманизированное антитело также будет включать, по меньшей мере, часть константной области иммуноглобулина, обычно принадлежащей иммуноглобулину человека, и, следовательно, будет включать человеческий Fc-участок. Гуманизированные антитела также можно получить, используя мышей с генетически сконструированной иммунной системой. Roque et al., 2004, Biotechnol. Prog. 20:639-654, работа включена путем отсылки во всей своей полноте. В этой области техники хорошо известно множество методов гуманизации и восстановления исходной формы антител, не принадлежащих к человеку (см. Tsurushita & Vasquez, 2004, Humanization of Monoclonal Antibodies, Molecular Biology of В Cells, 533-545, Elsevier Science (USA), и ссылки, процитированные там, все работы включены путем отсылки во всей своей полноте). Методы гуманизации включают, но не ограничиваются методами, описанными в работах Jones et al., 1986, Nature 321:522-525; Riechmann et al., 1988; Nature 332:323-329; Verhoeyen et al., 1988, science, 239:1534-1536; Queen et al., 1989, Proc. Natl. Acad. Sci., USA 86:10029-33; He et al., 1998, J. Immunol. 160:1029-1035; Carter et al., 1992, Proc. Natl. Acad. Sci. USA 89:4285-9, Presta et al., 1997, Cancer Res. 57(20):4593-9; Gorman et al., 1991, Proc. Natl. Acad. Sci. USA 88:4181-4185; O'Connor et al., 1998, Protein Eng 11:321-8, все работы включены путем отсылки во всей своей полноте. Гуманизация или другие методы снижения иммуногенности вариабельных областей антител, не являющихся человеческими, могут включать методы восстановления поверхности, как описано, например, в работе Roguska et al., 1994, Proc. Natl. Acad. Sci. USA 91:969-973, включенной путем отсылки во всей своей полноте. В одном из воплощений исходное антитело является афинно созревшим, как это известно в этой области техники. Для гуманизации и аффинного созревания можно применять структурные методы, как, например, описано в US 11/004,590. Для гуманизации и аффинного созревания вариабельных областей антител можно применять селекционные методы, которые включают, но не ограничиваются методами, описанными в работах Wu et al., 1999, J. Mol. Biol. 294:151-162; Baca et al., 1997, J. Biol. Chem. 272(16):10678-10684; Rosok et al., 1996, J. Biol. Chem. 271(37): 22611-22618; Rader et al., 1998, Proc. Natl. Acad. Sci. USA 95: 8910-8915; Krauss et al., 2003, Protein Engineering 16(10):753-759, включенных путем отсылки во всей своей полноте. Другие методы гуманизации могут включать пересадку только частей CDR, эти методы включают, но не ограничиваются методами, описанными в работах US 091810,502; Tan et al., 2002, J. Immunol. 169:1119-1125; De Pascalis et al., 2002, J. Immunol. 169:3076-3084, включенных сюда путем отсылки во всей своей полноте.
Биспецифические антитела
В одном из воплощений антитела изобретения являются мультиспецифичным антителом и исключительно биспецифичным антителом, также называемыми иногда «диателами». Это антитела, которые связывают два (или более) различных антигенов. Диатела можно изготовить множеством способов, известных в этой области техники (Holliger and Winter, 1993, Current Opinion Biotechnol. 4:446-449, работа включена путем отсылки во всей своей полноте), например, получив химическим путем или из гибридных гибридом.
Миниантитела
В одном из воплощений антитело является миниантителом. Миниантитела представляют собой доведенные до минимума антителоподобные белки, включающие scFv, присоединенный к CH1-домену. Hu et al., 1996, Cancer Res. 56:30553061, работа включена путем отсылки во всей своей полноте. В некоторых случаях scFv может быть присоединен к Fc-участку и может включать некоторую часть или весь шарнирный участок.
Человеческие антитела
В одном из воплощений антитело является полностью человеческим антителом, по меньшей мере, с одной модификацией, как уже было указано. «Полностью человеческое антитело» или «абсолютно человеческое антитело» относится к человеческому антителу, имеющему последовательность антитела, происходящего из человеческой хромосомы с модификациями, указанными в тексте заявки.
Слитые антитела
В одном из воплощений антитела изобретения представляют собой слитые белки антител (иногда называемые здесь «конъюгатом антител»). Один тип слитых антител включает слитые Fc, в которых Fc-участок связан с партнером по слиянию. Термин «слитый Fc». который используют в тексте, означает белок, в котором один или несколько полипептидов эффективно присоединены к Fc-участку. Слитый Fc здесь является синонимом терминов «иммуноадгезин», «слитый Ig», «химера Ig» и «рецепторный глобулин» (иногда пишется через тире), использовавшихся ранее (Chamow et al., 1996, Trends Biotechnol 14:52-60; Ashkenazi et al., 1997 Curr Opin Immunol 9:195-200). Слитый Fc объединяет в себе Fc-участок иммуноглобулина с партнером по слиянию, который, в основном, может быть любым белком или небольшой молекулой. Фактически любой белок или небольшая молекула могут быть присоединены к Fc для создания слитого Fc. Белковые партнеры по слиянию не ограничиваются данными примерами, но могут включать вариабельную область антитела, связывающий участок рецептора-мишени, адгезионную молекулу, лиганд, фермент, цитокин, хемокин или какой-либо другой белок или белковый домен. Небольшая молекула в качестве партнеров по слиянию может включать любое терапевтическое средство, которое направляет слитый Fc к терапевтической мишени. Такими мишенями могут быть любые молекулы, предпочтительно внеклеточный рецептор, который вовлечен в заболевание. Таким образом, IgG-варианты могут быть присоединены к одному или нескольким партнерам по слиянию. В одном альтернативном воплощении IgG-вариант конъюгирован или эффективно присоединен к другому терапевтическому соединению. Терапевтическое соединение может быть цитотоксичным средством, химиотерапевтическим средством, токсином, радиоизотопом, цитокином или другим терапевтически активным средством. IgG может быть присоединен к одному из множества небелковых полимеров, например к полиэтиленгликолю, пропиленгликолю, полиалкиленам или сополимерам полиэтиленгликоля и пропиленгликоля.
Кроме слитых Fc, слитые антитела включают слияние константной области тяжелой цепи с одним или несколькими партнерами по слиянию (кроме того, включая вариабельную область любого антитела), в то время как другие слитые антитела представляют собой в значительной степени полные антитела или полноразмерные антитела с партнерами по слиянию. В одном из воплощений роль партнера по слиянию состоит в опосредовании связывания с мишенью и, таким образом, он является функциональным аналогом вариабельных областей антитела (и на самом деле может им быть). Фактически любой белок или небольшая молекула могут быть присоединены к Fc для создания слитого Fc (или слитого антитела). Белковые партнеры по слиянию не ограничиваются данными примерами, но могут включать связывающий участок рецептора-мишени, адгезионную молекулу, лиганд, фермент, цитокин, хемокин или какой-либо другой белок или белковый домен. Небольшая молекула в качестве партнеров по слиянию может включать любое терапевтическое средство, которое направляет слитый Fc к терапевтической мишени. Такими мишенями могут быть любые молекулы, предпочтительно внеклеточный рецептор, который вовлечен в заболевание.
Партнер по слиянию может быть белковым и небелковым; последний обычно создается с использованием функциональных групп, расположенных на антителе и партнере по слиянию. Например, в этой области техники известны линкеры; например, хорошо известны гомо- или гетеро-бифункциональные линкеры (см. каталог 1994 Pierce Chemical Company технический раздел, относящийся к поперечно-сшивающим агентам, стр.155-200, включенный сюда путем отсылки).
Подходящие конъюгаты включают, но не ограничиваются только ими, метки, как описано ниже, лекарственные средства и цитотоксические средства, включающие, но не ограниченные цитотоксическими лекарственными средствами (например, химиотерапевтическими средствами) или токсинами или активными фрагментами таких токсинов. Подходящие токсины и их соответствующие фрагменты включают дифтерийную цепь А, цепь А экзотоксина, цепь А рицина, цепь А абрина, курцин, кротин, феномицин, эномицин и т.п. Цитотоксические средства также включают радиоактивные химические вещества, сделанные путем слияния радиоизотопов с антителами или путем связывания радионуклидов с хелатирующими агентами, которые были ковалентно присоединены к антителу. Дополнительные воплощения применяют калихеамицин, ауристатины, гелданамицин, майтанзин и дуокармицины и аналоги (последние описаны в US 2003/0050331 А1, включенным путем отсылки во всей своей полноте).
Ковалентные модификации антител
Ковалентные модификации антител включены в объем этого изобретения и, в основном, но не всегда, сделаны посттрансляционно. Например, несколько типов ковалентных модификаций антитела введены в молекулу путем реакции специфических аминокислотных остатков антитела с органическим дериватизирующим агентом, который способен реагировать с выбираемыми боковыми цепями или N- или С-концевыми остатками.
Остатки цистеина наиболее часто реагируют с α-галогенацетатами (и соответствующими аминами), такими как хлоруксусная кислота или хлорацетамид, с образованием карбоксиметильных или карбоксиамидометильных производных. Остатки цистеина можно также перевести в производные с помощью реакции с бромтрифторацетоном, α-бром-β-(5-имидазоил)пропионовой кислотой, хлорацетилфосфатом, N-алкилмалеимидами, 3-нитро-2-пиридилдисульфидом, метил-2-пиридилдисульфидом, пара-хлормеркурибензоатом, 2-хлормеркури-4-нитрофенолом или хлор-7-нитробензо-2-окса-1,3-диазолом и т.п.
Остатки гистидина превращают в производные с помощью реакции с диэтилпирокарбонатом при рН 5,5-7,0, так как это соединение относительно специфично по отношению к боковой цепи гистидина. Также полезен пара-бромфенацилбромид; реакцию предпочтительно проводят в 0,1 М какодилате натрия при рН 6,0.
Остатки лизина и концевые аминогруппы вводят в реакцию с ангидридами янтарной и других карбоновых кислот. Получение производных с помощью этих агентов оказывает эффект изменения заряда остатков лизина. Другие подходящие реагенты для получения производных остатков, содержащих альфа-аминогруппы, включают имидоэфиры, такие как метиловый эфир пиколинимидата; пиридоксальфосфат; пиридоксаль; хлорборгидрид; тринитробензосульфоновая кислота; O-метилизомочевина; 2,4-пентандион и реакция с глиоксилатом, катализируемая трансаминазой.
Остатки аргинина модифицируют с помощью реакции с одним или несколькими общепринятыми реагентами, среди них фенилглиоксаль, 2,3-бутандион, 1,2-циклогександион и нингидрин. Для превращения остатков аргинина в их производные необходимо, чтобы реакцию проводили в щелочных условиях из-за высокого значения рКа гуанидиновой функциональной группы. Кроме того, эти реагенты могут реагировать с лизиновыми группами, а также с эпсилон-аминогруппой аргинина.
Могут быть сделаны специфические модификации остатков тирозина, причем особый интерес представляет введение в остатки тирозина спектральных меток с помощью реакции с ароматическими соединениями диазония или тетранитрометаном. Чаще всего для образования O-ацетилтирозиновых производных и 3-нитро-производных применяют N-ацетилимидазол и тетранитрометан соответственно. Остатки тирозина иодируют, применяя 125I или 131I для получения меченых белков для применения в радиоиммунологических анализах, подходящим является метод с использованием хлорамина Т, описанный выше.
Боковые карбоксильные группы (аспартильная или глутамильная) селективно модифицируют с помощью реакции с карбодиимидами (R'-N=C=N-R'), где R и R' необязательно являются различными алкильными группами, такими как 1-циклогексил-3-(2-морфолинил-4-этил)карбодиимид или 1-этил-3-(4-азониа-4,4- диметилпентил)карбодиимид. Кроме того, остатки аспарагиновой и глутаминовой кислот превращают в остатки аспарагина и глутамина с помощью реакции с ионами аммония.
Дериватизация с помощью бифункциональных агентов полезна для сшивания антител с водонерастворимой поддерживающей матрицей или поверхностью для использования во многих методах, в дополнение к методам, описанным ниже. Обычно применяемые сшивающие агенты включают, например, 1,1-бис(диазоацетил)-2-фенилэтан, глутаровый альдегид, N-гидроксисукцинимидные эфиры, например эфиры с 4-азидосалициловой кислотой, гомофункциональные имидоэфиры, включающие дисукцинимидильные эфиры, такие как 3,3'-дитиобис(сукцинимидилпропионат) и бифункциональные малеимиды, такие как бис-N-малеимидо-1,8-октан. Дериватизирующие агенты, такие как метил-3-[(пара-азидофенил)дитио]пропиоимидат, позволяют получить фотоактивируемые интермедиаты, которые способны к образованию поперечных сшивок в присутствии света. Альтернативно, реакционноспособные водонерастворимые матрицы, такие как активированные цианогенбромидом углеводы и реакционноспособные субстраты, описанные в US Pat. Nos. 3,969,287; 3,691,016; 4,195,128; 4,247,642; 4,229,537 и 4,330,440, включенные сюда путем отсылки во всей своей полноте, применяют для иммобилизации белков.
Остатки глутамина и аспарагина часто деамидируют для получения соответствующих остатков глутаминовой и аспарагиновой кислот. Альтернативно, эти остатки можно деамидировать в мягких кислых условиях. Оба способа образования этих остатков входят в объем настоящего изобретения.
Другие модификации включают гидроксилирование пролина и лизина, фосфорилирование гидроксильных групп остатков серина и треонина, метилирование α-аминогрупп лизиновых, аргининовых и гистидиновых боковых цепей (Т.Е. Creighton, Proteins: Structure and Molecular Properties, W.H. Freeman&Co., San Francisco, pp.79-86 [1983], работа включена путем отсылки во всей своей полноте), ацетилирование N-концевого амина и амидирование любой С-концевой карбоксильной группы.
Гликозилирование
Другим типом ковалентной модификации является гликозилирование. В другом воплощении IgG-варианты, раскрытые здесь, могут быть модифицированы для включения одной или нескольких сконструированных гликоформ. Термин «сконструированные гликоформы», который здесь используют, означает смесь углеводов, которую ковалентно присоединяют к IgG, где указанные углеводы химически отличаются от углеводов исходного IgG. Сконструированные гликоформы могут быть полезными для многих целей, включающих, но не ограниченных увеличением или снижением эффекторной функции. Сконструированные гликоформы могут быть созданы с помощью множества способов, известных в этой области техники (Umaňa et al., 1999, Nat Biotechnol 17:176-180; Davies et al., 2001, Biotechnol Bioeng 74:288-294; Shields et al., 2002, J Biol Chem 277:26733-26740; Shinkawa et al., 2003, J Biol Chem 278:3466-3473; US 6,602,684; US 10/277,370; US 10/113,929; PCT WO 00/61739A1; PCT WO 01/29246A1; PCT WO 02/31140A1; PCT WO 02/30954A1, все источники включены путем отсылки во всей своей полноте (PotelligentTM technology [Biowa, Inc., Princeton, NJ]; GlycoMAb™ glycosylation engineering technology [GLYCART biotechnology AG, Zűrich, Switzerland]). Многие из этих методик основаны на регулировании уровня фукозилированных и/или разветвления олигосахаридов, которые ковалентно присоединены к Fc-участку, например, с помощью экспрессии IgG в различных организмах или клеточных линиях, сконструированных или сделанных иным способом (например, в клетках Lec-13 СНО или клетках крысиной гибридомы YB2/0), с помощью регулирования ферментов, вовлеченных в путь гликозилирования (например, FUT8 [α1,6-фукозилтрансферазы] и/или β1-4-N-ацетилглюкозаминилтрансферазы III [GnTIII]) или с помощью модификации углевода(ов) после экспрессии IgG. Сконструированная гликоформа обычно имеет отношение к различным углеводам или олигосахаридам; поэтому IgG-вариант, например антитело или слитый Fc, могут включать сконструированную гликоформу. Альтернативно, сконструированная гликоформа может иметь отношение к варианту IgG, который включает различный углевод или олигосахарид. Как известно в этой области техники, способы гликозилирования могут зависеть как от последовательности белка (например, от присутствия или отсутствия особенных аминокислотных остатков, пригодных для гликозилирования), так и от клетки-хозяина или организма, в котором продуцируют белок. Отдельные экспрессионные системы обсуждаются ниже.
Гликозилирование полипептидов обычно происходит либо через N-связь, либо через O-связь. N-гликозилирование имеет отношение к присоединению молекулы углевода к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где Х - любая аминокислота кроме пролина, являются последовательностями узнавания для энзиматического присоединения молекулы углевода к боковой цепи аспарагина. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. O-гликозилирование имеет отношение к присоединению одного из сахаров: N-ацетилгалактозамина, галактозы или ксилозы, к гидроксиаминокислоте, чаще всего серину или треонину, хотя также могут использоваться 5-гидроксипролин или 5-гидроксилизин.
Включение участков гликозилирования в антитело удобно проводить путем изменения аминокислотной последовательности, так чтобы она содержала одну или более трипептидных последовательностей из описанных выше (для связанных с атомом N сайтов гликозилирования). Изменение можно сделать с помощью присоединения или замещения одного или нескольких остатков серина или треонина для начала последовательности (для связанных с атомом О сайтов гликозилирования). Для удобства аминокислотную последовательность антитела предпочтительно модифицируют путем изменения на уровне ДНК, в особенности путем мутирования ДНК, кодирующей полипептид-мишень, в заранее выбранных основаниях, так чтобы образовались кодоны, которые будут транслироваться в желаемые аминокислоты.
Другими способами повышения количества молекул углеводов в антителе является химическое или энзиматическое присоединение гликозидов к белку. Эти процедуры полезны в тех методиках, которые не требуют продуцирование белка в клетке-хозяине, которая обладает способностью к гликозилированию по N- и O-связям. В зависимости от применяемого способа присоединения сахар(сахара) могут присоединяться к (а) аргинину и гистидину, (b) свободным карбоксильным группам, (с) свободным сульфгидрильным группам, таким как цистеин, (d) свободным гидроксильным группам, таким как серин, треонин или гидроксипролин, (е) ароматическим остаткам, таким как фенилаланин, тирозин или триптофан, или (f) амидной группе глутамина. Эти методы описаны в WO 87/05330 и в работе Aplin and Wriston, 1981, CRC Crit. Rev. Biochem., pp.259-306, включенных сюда путем отсылки во всей своей полноте.
Удаление углеводной части, присутствующей в исходном антителе, может быть проведено химически или энзиматически. Для химического дегликозилирования необходимо воздействие на белок соединения трифторметансульфоновой кислоты или эквивалентного соединения. Эта обработка приводит к отщеплению большинства или всех сахаров, за исключением связанных сахаров (N-ацетилглюкозамина или N-ацетилгалактозамина), в то время как уходящий белок остается интактным. Химическое дегликозилирование описано Hakimuddin et al., 1987, Arch. Biochem. Biophys. 259:52 и Edge et al., 1981, Anal. Biochem. 118:131, обе работы включены путем отсылки во всей своей полноте. Энзиматическое отщепление углеводной части полипептида может быть достигнуто при использовании множества эндо- и экзогликозидаз, как описано в работе Thotakura et al., 1987, Meth. Enzymol. 138:350, включенной путем отсылки во всей своей полноте. Гликозилирование в потенциальных участках гликозилирования можно предотвратить при использовании соединения туникамицина, как описано в работе Duskin et al., 1982, J. Biol. Chem. 257:3105, включенной сюда путем отсылки во всей своей полноте. Туникамицин блокирует образование белок-N-гликозидных связей.
Другой тип ковалентной модификации антитела включает присоединение антитела к различным небелковым полимерам, включающим, но не ограниченным различными полиолами, такими как полиэтиленгликоль, полипропиленгликоль или полиоксиалкилены, способом, описанным, например, в 2005-2006 PEG Catalog from Nektar Therapeutics (доступно на веб-сайте Nektar), US Patents 4,640,835; 4,496,689; 4,301,144; 4,670,417; 4,791,192 или 4,179,337, все они включены путем отсылки во всей своей полноте. Кроме того, как известно в этой области техники, в различных положениях внутри антитела можно сделать аминокислотные замены, чтобы облегчить присоединение полимеров, таких как ПЭГ. См., например, публикацию US Publication No. 2005/011037А1, включенную путем отсылки во всей своей полноте.
Меченые антитела
В некоторых воплощениях ковалентная модификация антител изобретения включает внедрение одной или нескольких меток. В некоторых случаях их рассматривают как слитые антитела. Термин «метящая группа» означает любую детектируемую метку. В некоторых воплощениях меченая группа присоединена к антителу через спейсерные ножки различной длины для снижения потенциального стерического затруднения. Различные методы получения меченых белков известны в этой области техники и могут быть использованы в осуществлении настоящего изобретения.
В основном, метки бывают трех классов, в зависимости от анализа, в которых эти метки обнаруживают: а) изотопные метки, которые могут быть радиоактивными или тяжелыми изотопами; b) магнитные метки (например, магнитные частицы); с) редокс-активные вещества; d) оптические краски; энзиматические группы (например, пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза); е) биотинилированные группы и f) заранее определенные полипептидные эпитопы, узнаваемые вторичными репортерами (например, пара последовательностей «лейциновой молнии», связывающие места для вторичных антител, металл-связывающие домены, эпитопные метки и т.д.). В некоторых воплощениях меченая группа присоединена к антителу через спейсерные ножки различной длины для снижения потенциального стерического несоответствия. Различные методы получения меченых белков известны в этой области техники и могут быть использованы в осуществлении настоящего изобретения.
Специфические метки включают оптические краски, включающие, но не ограниченные только ими, хромофоры, люминофоры и флуорофоры, последние во многих случаях являются специфичными. Флуорофоры могут быть или флуоресцентными «небольшими молекулами», или флуоресцентными метками белковой природы.
Термин «флуоресцентная метка» означает любую молекулу, которую можно детектировать, используя присущие ей флуоресцентные свойства. Подходящие флуоресцентные метки включают, но не ограничиваются только ими, флуоресцеин, родамин, тетраметилродамин, эозин, эритрозин, кумарин, метилкумарины, пирен, малахитовый зеленый, стильбен, Lucifer Yellow, Cascade BlueJ, Texas Red, IAEDANS, EDANS, BODIPY FL, LC Red 640, Су 5, Су 5.5, LC Red 705, Oregon green, краски Alexa-Fluor (Alexa Fluor 350, Alexa Fluor 430, Alexa Fluor 488, Alexa Fluor 546, Alexa Fluor 568, Alexa Fluor 594, Alexa Fluor 633, Alexa Fluor 660, Alexa Fluor 680), Cascade Blue, Cascade Yellow и R-фикоэритрин (РЕ) (Molecular Probes, Eugene, OR), FITC, родамин и Texas Red (Pierce, Rockford, IL), Cy5, Cy5.5, Cy7 (Amersham Life Science, Pittsburgh, PA). Подходящие оптические краски, включающие флуорофоры, описаны в книге Molecular Probes Handbook by Richard P. Haugland, включенной сюда путем отсылки во всей своей полноте.
Подходящие белковые флуоресцентные метки включают, но не ограничиваются только ими, зеленый флуоресцентный белок, включая GFP из видов Renilla, Ptilosarcus или Aequorea (Chalfie et al., 1994, Science 263:802-805), EGFP (Clontech Laboratories, Inc., код доступа в Genbank U55762), синий флуоресцентный белок (BFP, Quantum Biotechnologies, Inc. 1801 de Maisonneuve Blvd. West, 8th Floor, Montreal, Quebec, Canada H3H 1J9; Stauber, 1998, Biotechniques 24:462-471; Helm et al., 1996, Cur. Biol. 6:178-182), желтый белок с усиленной флуоресценцией (EYFP, Clontech Laboratories, Inc.), люциферазу (Ichiki et al., 1993, J. Immunol. 150:5408-5417), β-галактозидазу (Nolan et al., 1988, Proc. Natl. Acad. Sci. USA 85:2603-2607) и Renilla (WO 92/15673, WO 95/07463, WO 98/14605, WO 98/26277, WO 99/49019, US Patent Nos. 5292658, 5418155, 5683888, 5741668, 5777079, 5804387, 5874304, 5876995, 5925558). Все указанные выше источники в этом параграфе непосредственно включены путем отсылки.
IgG-варианты
В одном из воплощений изобретение обеспечивает вариантные IgG-белки. Как минимум IgG-варианты включают фрагмент антитела, включающий СН2-СН3-участок тяжелой цепи. Кроме того, подходящие IgG-варианты включают Fc-домены (например, включающие нижний шарнирный участок), а также IgG-варианты, включающие константную область тяжелой цепи (СН1-шарнир-СН2-СН3), также полезны в настоящем изобретении, все они могут быть присоединены к партнерам по слиянию.
IgG-вариант включает одну или несколько аминокислотных модификаций, относящихся к исходному IgG-полипептиду, в некоторых случаях относящихся к дикому типу IgG. IgG-вариант может обладать одним или несколькими оптимизированными свойствами. IgG-вариант отличается по аминокислотной последовательности от своего исходного IgG фактически, по меньшей мере, одной аминокислотной модификацией. Таким образом, IgG-варианты имеют, по меньшей мере, одну аминокислотную модификацию по сравнению с исходным белком. Альтернативно, IgG-варианты могут иметь больше, чем одну, например, приблизительно от одной до пятидесяти аминокислотных модификаций, предпочтительно приблизительно от одной до десяти аминокислотных модификаций и наиболее предпочтительно приблизительно от одной до приблизительно пяти аминокислотных модификаций по сравнению с исходным белком.
Таким образом, последовательности IgG-вариантов и исходного Fc-полипептида в значительной степени гомологичны. Например, вариантные последовательности IgG-вариантов здесь будут иметь приблизительно 80%-ную гомологию с последовательностью исходного IgG-варианта, предпочтительно, по меньшей мере, приблизительно 90%-ную гомологию и более предпочтительно, по меньшей мере, приблизительно 95%-ную гомологию. Модификации могут быть сделаны генетически с использованием молекулярной биологии, или могут быть сделаны энзиматически или химически.





Специалисты в этой области техники признают, что упомянутый выше список мишеней относится не только к специфическим белкам и биомолекулам, но и к биохимическому пути или путям, которые их включают. Например, ссылка на CTLA-4 в качестве антигена-мишени подразумевает, что лиганды и рецепторы, которые составляют Т-клеточный костимулирующий путь, включающий CTLA-4, В7-1, В7-2, CD28 и любые другие не открытые в настоящее время лиганды и рецепторы, которые связывают эти белки, также являются мишенями. Таким образом, термин «мишень», который здесь используют, относится не только к специфическим биомолекулам, но и к ряду белков, которые взаимодействуют с указанной мишенью и участниками биохимического пути, которому принадлежит указанная мишень. Специалисты в этой области техники в дальнейшем поймут, что любой из указанных выше антигенов-мишеней, лигандов или рецепторов, которые их связывают, или других участников соответствующего биохимического пути может быть эффективно присоединен к Fc-вариантам настоящего изобретения для образования слитого Fc. Так, например, слитый Fc, который нацелен на EGFR, может быть сконструирован эффективным присоединением Fc-варианта к EGF, TGF-b или любому другому лиганду, исследованному или не открытому в настоящее время, который связывается с EGFR. Соответственно Fc-вариант настоящего изобретения может быть эффективно присоединен к EGFR для образования слитого Fc, который связывает EGFR, TGF-b или любой другой лиганд, исследованный или не открытый в настоящее время, который связывается с EGFR. Таким образом, фактически любой полипептид, или лиганд, или рецептор, или некоторый другой белок или белковый домен, включающий, но не ограниченный указанными выше мишенями и белками, которые составляют соответствующие биохимические пути, может быть эффективно присоединен к Fc-вариантам настоящего изобретения для разработки слитого Fc.
Выбор подходящего антигена зависит от желаемого практического применения. Для противораковой терапии желательно иметь мишень, экспрессия которой будет ограничена в раковых клетках. Некоторые мишени, для которых доказано, что они особенно хорошо поддаются лечению антителами, являются мишенями с сигнальными функциями. Другие терапевтические антитела проявляют свои эффекты путем блокирования передачи сигналов рецептором путем ингибирования связывания рецептора и лиганда, узнаваемого этим рецептором. Другой механизм действия терапевтических антител состоит в том, что они вызывают супрессирующую регуляцию рецептора. Другие антитела не функционируют путем передачи сигнала через свой антиген-мишень. В некоторых случаях применяют антитела, направленные против агентов, вызывающих инфекционные заболевания.
В одном из воплощений Fc-варианты настоящего изобретения вводят в антитело вместе с цитокином. Альтернативно, Fc-варианты сливают или конъюгируют с цитокином. Термин «цитокин», используемый здесь, означает общий термин для белков, высвобождаемых одной клеточной популяцией, которые действуют на другие клетки как межклеточные медиаторы. Примерами таких цитокинов являются лимфокины, монокины и традиционные пептидные гормоны. Например, как описано в работе Penichet et al., 2001, J Immunol Methods 248:91-101, прямо включенной сюда путем отсылки, цитокины могут быть слиты с антителом для предоставления набора желаемых свойств. Примерами таких цитокинов являются лимфокины, монокины и обычные полипептидные гормоны. К цитокинам относятся гормон роста, например гормон роста человека, N-метионильный гормон роста человека и бычий гормон роста; паратгормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, например фоликулстимулирующий гормон (FSH), тироидстимулирующий гормон (TSH) и лютеинизирующий гормон (LH); печеночный фактор роста; фактор роста фибробластов; пролактин; плацентарный лактоген; фактор некроза опухолей-альфа и -бета; антимюллеров гормон; мышиный гонадотропин-ассоциированный пептид; ингибин; активин; эндотелиальный фактор роста сосудов; интегрин; тромбопоэтин (ТРО), факторы роста нервов, например NGF-beta; фактор роста тромбоцитов; трансформирующие факторы роста (TGFs), например TGF-альфа и TGF-бета; инсулиноподобный фактор роста I и II; эритропоэтин (ЕРО); остеоиндуктивные факторы; интерфероны, например интерферон-альфа, -бета и -гамма; колониестимулирующие факторы (CSFs), например макрофагальный-колониестимулирующий фактор (N-CSF), гранулоцитарный-макрофагальный-CSF (GM-CSF) и гранулоцитарный-CSF (G-CSF); интерлейкины (ILs), например IL-1, IL-1альфа, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12; IL-15, фактор некроза опухолей, например TNF-альфа или TNF-бета; С5а и другие полипептидные факторы, включающие LIF и лиганд Kit (KL). Используемый здесь термин «цитокин» включает белки из природных источников или из рекомбинантных клеточных культур и биологически активные эквиваленты цитокинов с природными последовательностями.
Цитокины и растворимые мишени, например представители суперсемейства TNF, являются предпочтительными мишенями при применении вариантов настоящего изобретения. Например, антитела к VEGF и антитела к CTLA-4, и антитела к TNF или их фрагменты, представляют собой в особенности хорошие антитела для применения Fc-вариантов с усиленным связыванием FcRn. Терапевтические средства, направленные на эти мишени, часто применяют при лечении аутоиммунных заболеваний и для этого требуются многократные инъекции в течение долгого периода времени. Поэтому особенно предпочтительны увеличенное время полужизни в сыворотке крови и менее частые воздействия, обеспечиваемые вариантами настоящего изобретения.
Для некоторых антител и слитых Fc, которые одобрены для применения, находятся на стадии клинических испытаний или разработки, могут быть полезны Fc-варианты настоящего изобретения. Эти антитела и слитые Fc называются здесь «клиническими продуктами и кандидатами». Таким образом, в предпочтительном воплощении Fc-полипептиды настоящего изобретения могут найти применение в ряду клинических продуктов и кандидатов. Например, для некоторых антител, мишенью которых является CD20, могут быть полезны Fc-полипептиды настоящего изобретения. Например, Fc-полипептиды настоящего изобретения могут найти применение в антителах, которые в значительной степени сходны с ритуксимабом (Rituxan®, IDEC/Genentech/Roche) (см. например, US 5,736,137), химерным антителом к CD20, одобренным для лечения неходжкинской лимфомы; HuMax-CD20, антителом к CD20, в настоящее время разрабатываемым компанией «Genmab», антителом к CD20, описанным в патенте US 5,500,362, АМЕ-133 (Applied Molecular Evolution), hA20 (Immunomedic, Inc.), HumaLYM (Intracel) и PRO70769 (PCT/US2003/040426 под названием «Immunoglobulin Variants and Uses Thereof»). Для некоторых антител, мишенью которых являются представители семейства рецепторов эпидермального фактора роста, включающих EGFR (ЕrbВ-1), Her2/neu (ErbB-2), Неr3 (ErbB-3), Her4 (Erb-4), могут быть полезны Fc-полипептиды настоящего изобретения. Например, Fc-полипептиды настоящего изобретения могут найти применение в антителе, которое в значительной степени сходно с трастузумабом (Herceptin®, Genentech) (см., например, US 5,677,171), гуманизированным антителом к Her2/neu, одобренном для лечения рака молочной железы; пертузумабом (rhuMab-2C4, Omnitarg™), в настоящее время разрабатываемым компанией «Genentech»; антителом к Нег2, описанным в US 4,753,894; цетуксимабом (Erbitux®, Imclone) (US 4,943,533; PCT WO 96/40210), химерным антителом к EGFR, проходящим клинические испытания для применения при множестве видов рака; ABX-EFG (US 6,235,883), в настоящее время разрабатываемым компанией «Abgenix/Immunex/Amgen»; HuMax-EGFr (US 10/172,317), в настоящее время разрабатываемым компанией «Genmab»; 425, EMD55900, EMD62000 и EMD72000 (Merck KGaA) (US 5558864; Murthy et al., 1987, Arch Biochem Biophys. 252(2):549-60; Rodeck et al., 1987, J Cell Biochem. 35(4):315-20; Kettleborough et al., 1991, Protein Eng. 4(7):773-83); ICR62 (Institute of Cancer Research) (PCT WO 95/20045; Modjtahedi et al., 1993, J Cell Biophys. 1993, 22(1-3):129-46; Modjtahedi et al., 1993, Br J Cancer. 1993, 67(2):247-53; Modjtahedi et al., 1996, Br J Cancer, 73(2):228-35; Modjtahedi et al., 2003, Int J Cancer, 105(2):273-80); TheraCIM hR3 (YM Biosciences, Canada and Centro de Immunologia Molecular, Cuba (US 5,891,996; US 6,506,883; Mateo et al., 1997, Immunotechnology, 3(1):71-81); mAb-806 (Ludwig Institute for Cancer Research, Memorial Sloan-Kettering) (Jungbluth et al., 2003, Proc Natl Acad Sci USA. 100(2):639-44); KSB-102 (KS Biomedix); MR1-1 (IVAX, National Cancer Institute) (PCT WO 0162931A2) и SC100 (Scancell) (PCT WO 01/88138). В другом предпочтительном воплощении Fc-полипептиды настоящего изобретения могут найти применение в алемтузумабе (Campath®, Millenium), гуманизированном моноклональном антителе, в настоящее время одобренном для лечения хронической В-клеточной лимфоцитарной лейкемии. Fc-полипептиды настоящего изобретения могут найти применение во множестве антител или слитых Fc, которые в значительной степени сходны с другими клиническими продуктами и кандидатами, включающими, но не ограниченными ими, муромонаб-CD3 (Orthoclone OKT3®), антитело к CD3, разработанное компанией «Ortho Biotech/Johnson & Johnson», ибритумомаб тиуксетан (Zevalin®), антитело к CD20, разработанное компанией «IDEC/Schering AG», гемтузумаб озогамицин (Mylotarg®), антитело к CD33 (белок р67), разработанное компанией «Celltech/Wyeth», алефасепт (Amevive®), антитело к LFA-3, слитое с Fc, разработанное компанией «Biogen», абциксимаб (ReoPro®), разработанный компанией «Centocor/Lilly», басиликсимаб (Simultec®), разработанный компанией «Novartis», паливизумаб (Sinagis®), разработанный компанией «MedImmune», инфликсимаб (Remicade®), антитело к TNF-альфа, разработанное компанией «Centocor», адалимумаб (Humira®), антитело к TNF-альфа, разработанное компанией «Abbott, Humicade™», антитело к TNF-альфа, разработанное компанией «Celltech», этанерцепт (Enbrel®), антитело к TNF-альфа, слитое с Fc, разработанное компанией «Immunex/Amgen», АВХ-CBL, антитело к CD 147, разрабатываемое компанией «Abgenix», ABX-IL8, антитело к IL8, разрабатываемое компанией «Abgenix», ABX-MA1, антитело к MUC18, разрабатываемое компанией «Abgenix», пемтумомаб (R1549, 90Y-muHMFG1), антитело к MUC1 в разработке компанией «Antisoma», Therex (R1550), антитело к MUC1, разрабатываемое компанией «Antisoma», AngioMab (AS1405), разрабатываемый компанией «Antisoma», HuBC-1, разрабатываемый компанией «Antisoma», Thioplatin (AS1407), разрабатываемый компанией «Antisoma», Antegren® (натализумаб), антитело к альфа-4-бета-1(VLA-4) и альфа-4-бета-7, разрабатываемое компанией «Biogen», VLA-1 mAb, антитело к интегрину VLA-1, разрабатываемое компанией «Biogen», LTBR mAb, антитело к рецептору лимфотоксина-бета (LTBR), разрабатываемое компанией «Biogen», CAT-152, антитело к TGFβ2, разрабатываемое компанией «Cambridge Antibody Technology», J695, антитело к IL-12, разрабатываемое компаниями «Cambridge Antibody Technology» и «Abbott», CAT-192, антитело к TGFβ1, разрабатываемое компаниями «Cambridge Antibody Technology» и «Genzyme», CAT-213, антитело к эотаксину-1, разрабатываемое компанией «Cambridge Antibody Technology», LymphoStat-B™ анти-Blys антитело, разрабатываемое компаниями «Cambridge Antibody Technology» и «Human Genome Sciences Inc.», TRAIL-R1mAb, антитело к TRAIL-R1, разрабатываемое компаниями «Cambridge Antibody Technology» и «Human Genome Sciences Inc.», Avastin™ (бевацизумаб, rhuMAb-VEGF), антитело к VEGF, разрабатываемое компанией «Genentech», антитело к семейству рецепторов HER, разрабатываемое компанией «Genentech», Anti-Tissue Factor (ATF), антитело к тканевому фактору, разрабатываемое компанией «Genentech», Xolair™ (омализумаб), антитело к иммуноглобулинам IgE, разрабатываемое компанией «Genentech», Raptiva™ (ефализумаб), антитело к CD-11, разрабатываемое компанией «Genentech» and «Xoma», антитело MLN-02 (ранее называвшееся LDP-02), разрабатываемое компаниями «Genentech» и «Millenium Pharmaceuticals», HuMax CD4, антитело к CD4, разрабатываемое компанией «Genmab», HuMax-IL15, антитело к IL15, разрабатываемое компаниями «Genmab» и «Amgen», HuMax-Inflam, разрабатываемый компаниями «Genmab» и «Medarex», HuMax-Cancer, антитело к гепараназе I, разрабатываемое компанией «Genmab» and «Medarex» and Oxford GcoSciences, препарат HuMax-Lymphoma, разрабатываемый компаниями «Genmab» и «Amgen», HuMax-TAC, разрабатываемый компанией «Genmab», IDEC-131 и антитело к CD40L, разрабатываемые компанией «IDEC Pharmaceuticals», IDEC-151 (кленоликсимаб), антитело к CD4, разрабатываемое компанией «IDEC Pharmaceuticals», IDEC-114, антитело к CD80, разрабатываемое компанией «IDEC Pharmaceuticals», IDEC-152, антитело к CD23, разрабатываемое компанией «IDEC Pharmaceuticals», антитела к фактору, ингибирующему миграцию макрофагов (MIF), разрабатываемые компанией «IDEC Pharmaceuticals», BEC2, антиидиотипическое антитело, разрабатываемое компанией «Imclone», IMC-1C11, антитело к KDR, разрабатываемое компанией «Imclone», DC101, антитело к flk-1, разрабатываемое компанией «Imclone», антитела к VE-кадгерину, разрабатываемые компанией «Imclone», CEA-Cide™ (лабетузумаб), антитело к карциноэмбриональному антигену (СЕА), разрабатываемое компанией «Immunomedics», LymphoCide™ (эпратузумаб), антитело к CD22, разрабатываемое компанией «Immunomedics», AFP-Cide, разрабатываемый компанией «Immunomedics», MyelomaCide, разрабатываемый компанией «Immunomedics», LkoCide, разрабатываемый компанией «Immunomedics», ProstaCide, разрабатываемый компанией «Immunomedics», MDX-010, антитело к CTLA4, разрабатываемое компанией «Medarex», MDX-060, антитело к CD30, разрабатываемое компанией «Medarex», MDX-070, разрабатываемое компанией «Medarex», MDX-018, разрабатываемое компанией «Medarex», Osidem™ (IDM-1) и антитело к Her2, разрабатываемые компаниями «Medarex» и «Immuno-Designed Moleculas», HuMax™-CD4, антитело к CD-4, разрабатываемое компанией «Medarex» и «Genmab», HuMax-IL15, антитело к IL15, разрабатываемое компанией «Medarex» и «Genmab», CNTO 148, анти-TNFα антитело, разрабатываемое компаниями «Medarex» и «Centocor/J&J», CNTO 1275, антитело к цитокину, разрабатываемое компанией «Centocor/J&J», MOR101 и MOR102, антитела к внутриклеточной адгезионной молекуле-1 (ICAM-1) (CD54), разрабатываемые компанией «MorphoSys», MOR201, антитело к рецептору 3 фактора роста фибробластов (FGFR-3), разрабатываемое компанией «MorphoSys», Nuvion® (висилизумаб), антитело к CD-3, разрабатываемое компанией «Protein Design Labs», HuZaF™, антитело к интерферону-гамма, разрабатываемое компанией «Protein Design Labs», антитело к интегрину α5β1, разрабатываемое компанией «Protein Design Labs», антитело к IL-12, разрабатываемое компанией «Protein Design Labs», ING-1, антитело к Ер-САМ, разрабатываемое компанией «Xoma» и MLN01, антитело к бета2-интегрину, разрабатываемое компанией «Xoma», все указанные выше источники в этом параграфе прямо включены путем отсылки.
Fc-полипептиды настоящего изобретения могут быть включены в указанные выше клинические кандидаты и продукты или в антитела и слитые Fc, которые в значительной степени сходны с ними. Fc-полипептиды настоящего изобретения могут быть включены в разновидности указанных выше клинических кандидатов и продуктов, которые представляют собой гуманизированные, аффинно-созревшие, сконструированные или модифицированные каким-либо способом разновидности.
В одном из воплощений Fc-полипептиды настоящего изобретения применяют для лечения аутоиммунных, воспалительных заболеваний или по показаниям при трансплантации. Антигены-мишени и клинические продукты и кандидаты, которые важны при таких заболеваниях, включают, но не ограничиваются ими, антитела к α4β7-интегрину, такие как LDP-02, антитела к бета-2-интегрину, такие как LDP-01, антитела к комплементу (С5), такие как 5G1.1, антитела к CD2, такие как BTI-322, MEDI-507, антитела к CD3, такие как ОKТ3, SMART anti-CD3, антитела к CD4, такие как IDEC-151, MDX-CD4, ОКТ4А, антитела к CD11а, антитела к CD14, такие как IC14, антитела к CD18, антитела к CD23, такие как IDEC 152, антитела к CD25, такие как зенапакс, антитела к CD40L, такие как 5с8, Antova, IDEC-131, антитела к CD64, такие как MDX-33, антитела к CD80, такие как IDEC-114, антитела к CD147, такие как ABX-CBL, антитела к Е-селектину, такие как CDP850, антитела к gpIIb/IIIа, такие как ReoPro/Abcixima, антитела к ICAM-3, такие как ICM3, антитела к ICE, такие как VX-740, антитела к FcR1, такие как MDX-33, антитела к IgE, такие как rhuMab-E25, антитела к IL-4, такие как SB-240683, антитела к IL-5, такие как SB-240563, SCH55700, антитела к IL-8, такие как ABX-IL8, антитела к интерферону-гамма, антитела к TNF (TNF, TNFa, TNFa, TNF-альфа), такие как CDP571, CDP870, D2E7, инфликсимаб, MAK-195F и антитела к VLA-4, такие как антегрен.
Fc-варианты настоящего изобретения с увеличенным связыванием с FcRn могут быть использованы в TNF-ингибиторных молекулах для обеспечения улучшенных свойств. Полезные TNF-ингибиторные молекулы включают любую молекулу, которая ингибирует действие TNF-альфа у млекопитающих. Подходящие примеры включают слитый Fc-белок Enbrel® (этанерцепт) и антитела Humira® (адалимумаб) и Remicade® (инфликсимаб). Моноклональные антитела (такие как ремикад и хумира), сконструированные с помощью Fc-вариантов настоящего изобретения, для улучшения связывания с FcRn, можно преобразовать в антитела с лучшей эффективостью путем увеличения времени полужизни.
В некоторых воплощениях применяют антитела против инфекционных заболеваний. Антитела против эукариотических клеток включают антитела, направленные на клетки дрожжей, включающие, но не ограниченные Saccharomyces cerevisiae, Hansenula polymorpha, Kluyveromyces fragilis и K.lactis, Pichia guillerimondii и Р.pastoris, Schizosaccharomyces pombe, Plasmodium falciparium и Yarrowia lipolytica.
Также полезны антитела к другим клеткам грибов, включающих антигены-мишени, ассоциированные со штаммами Candida, включающими Candida glabrata, Candida albicans, С.krusei, С.lusitaniae и С.maltosa, а также представители рода Aspergillus, Cryptococcus, Histoplasma, Coccidioides, Blastomyces и Penicillium и др.
Антитела, направленные против антигенов-мишеней, ассоциированных с простейшими, включают, но не ограничиваются антителами, ассоциированными с Trypanosoma, представителями рода Leishmania, включающими Leishmania donovanii, Plasmodium spp., Pneumocystis carinii, Cryptosporidium parvum, Giardia lamblia, Entamoeba histolytica и Cyclospora cayetanensis.
Полезны также антитела против антигенов прокариотов, включающие антитела против подходящих бактерий, таких как патогенные и непатогенные прокариоты, включающие, но не ограниченные только ими, Bacillus, включая Bacillus anthracis; Vibrio, например V. cholerae; Escherichia, например энтеротоксигенные Е.coli, Shigella, например S. dysenteriae; Salmonella, например S. typhi; Mycobacterium, например М. tuberculosis, M. leprae; Clostridium, например С. botulinum, С. tetani, С. difficile, С. perfringens; Corynebacterium, например С. diphtheriae; Streptococcus, S. pyogenes, S. pneumoniae; Staphylococcus, например P. aureus; Haemophilus, например H. influenzae; Neisseria, например N. meningitidis, N. gonorrhoeae; Yersinia, например Y. lamblia, Ypestis, Pseudomonas, например P. aeruginosa, P. putida; Chlamydia, например С.trachomatis; Bordetella, например В. pertussis; Treponema, например Т. palladium; В. anthracis, Y pestis, Brucella spp., F. tularensis, B. mallei, B. pseudomallei, C. botulinum, Salmonella spp., SEB V. cholerae toxin B, Е. coli O157:H7, Listeria spp., Trichosporon beigelii, Rhodotorula species, Hansenula anomala, Enterobacter sp., Klebsiella sp., Listeria sp., Mycoplasma sp. и т.п.
В некоторых аспектах антитела направлены против вирусных инфекций; эти вирусы включают, но не ограничиваются только ими, ортомиксовирусы (например, вирус гриппа), парамиксовирусы (например, респираторно-синцитиальный вирус, вирус свинки, вирус кори), аденовирусы, риновирусы, коронавирусы, реовирусы, тогавирусы (например, вирус краснухи), парвовирусы, поксвирусы (например, вирус натуральной оспы, вирус коровьей оспы), энтеровирусы (например, полиовирус, вирус Коксаки), вирусы гепатита (включая А, В и С), вирусы герпеса (например, вирус простого герпеса, вирус ветряной оспы, цитомегаловирус, вирус Эпштейна-Барр), ротавирусы, вирусы Норфолк, хантавирус, аренавирус, рабдовирус (например, вирус бешенства), ретровирусы (включая HIV, HTLV-I и II), паповавирусы (например, вирус папилломы), полиомавирусы и пикорнавирусы и т.п.
Свойства оптимизированных IgG-вариантов
Настоящее изобретение также обеспечивает IgG-варианты, которые оптимизированы под широкий спектр терапевтически важных свойств. IgG-вариант, сконструированный или спрогнозированный для демонстрации одного или нескольких оптимизированных свойств, называется здесь «оптимизированным IgG-вариантом». Наиболее предпочтительные свойства, которые могут быть оптимизированы, включают, но не ограничиваются увеличением или уменьшением сродства к FcRn и увеличение или уменьшение времени полужизни in vivo. Подходящие воплощения включают антитела, которые демонстрируют улучшенное сродство при связывании с FcRn при сниженных значениях рН, таких как значения рН, ассоциированные с эндосомами, например при рН 6,0, и в то же время не проявляют соответствующего улучшенного сродства при связывании при более высоких значениях рН, например при рН 7,4, что позволяет повысить поглощение в эндосомы при нормальных скоростях высвобождения. Сходным образом, эти антитела с модулированным связыванием FcRn необязательно обладают другими желаемыми свойствами, такими как модулированное связывание FcγR, такое как описано в US 11/174,287, 11/124,640, 10/822,231, 10/672,280, 10/379,392 и патентной заявке, озаглавленной «IgG Immunoglobulin variants with optimized effector function» и поданной 21 октября 2005 г. под номером ________. Таким образом, оптимизированные свойства также включают, но не ограничиваются только ими, повышение или снижение сродства к FcγR. В одном необязательном воплощении IgG-варианты оптимизированы для того, чтобы увеличить их сродство к активаторному FcγR человека, предпочтительно к FcγRIIIa в дополнение к FcRn-связывающему профилю. В еще одном необязательном альтернативном предпочтительном воплощении IgG-варианты оптимизированы для того, чтобы снизить их сродство к ингибиторному рецептору FcγRIIb человека. Значит, особые воплощения включают применение антител, которые демонстрируют повышенное связывание с FcRn и повышенное связывание с FcγRIIIa. В других воплощениях применяют антитела, которые демонстрируют повышенное связывание с FcRn и повышенное связывание с FcγRIIIa. Эти воплощения предусматривают обеспечение IgG-полипептидов с усиленными терапевтическими свойствами для людей, например, с усиленной эффекторной функцией и более высокой противораковой эффективностью. В альтернативном воплощении IgG-варианты оптимизированы таким образом, что у них повышено или понижено сродство к FcRn и повышено или понижено сродство к FcγR человека, включая, но не ограничиваясь FcγRI, FcγRIIa, FcγRIIb, FcγRIIc, FcγRIIIa и FcγRIIIb, включая их аллельные вариации. Эти воплощения предусматривают обеспечение IgG-полипептидов с усиленными терапевтическими свойствами для людей, например, с увеличенным временем полужизни в сыворотке крови и пониженной эффекторной функцией. В других воплощениях IgG-варианты обеспечивают повышенное сродство к FcRn и повышенное сродство к одному или нескольким FcγRs, и, кроме того, пониженное сродство к одному или нескольким FcγRs. Например, IgG-вариант может обладать улучшенным связыванием с FcRn и FcγRIIIa и, кроме того, пониженным связыванием с FcγRIIb. Альтернативно, IgG-вариант может обладать пониженным связыванием с FcRn и FcγR's. В другом воплощении IgG-вариант может обладать пониженным сродством к FcRn и повышенным сродством к FcγRIIb, и, кроме того, пониженным сродством к одному или нескольким активаторным FcγRs. В еще одном воплощении IgG-вариант может иметь увеличенное время полужизни в сыворотке крови и ослабленные эффекторные функции.
Предпочтительные воплощения включают оптимизацию связывания с человеческим FcRn и FcγR, однако в альтернативных воплощениях IgG-варианты обладают повышенным или пониженным сродством к FcRn и FcγRs из организмов, не являющихся людьми, включающих, но не ограниченных грызунами и приматами, не являющимися людьми. IgG-варианты, у которых оптимизировано связывание с FcRn, не относящегося к человеческому, могут найти применение в проведении исследований. Например, модели на мышах пригодны для исследования множества болезней и позволяют испытывать свойства, такие как эффективность, токсичность и фармакокинетика определенного кандидата в лекарственное средство. Как известно специалистам в этой области техники, раковые клетки могут быть привиты или введены в мышь для имитации рака людей, этот процесс известен как ксенотрансплантация. Испытание IgG-вариантов, которые включают IgG-варианты, оптимизированные для FcRn, может предоставить полезную информацию относительно характеристик клиренса белка, механизма его клиренса и т.п. IgG-варианты также могут быть оптимизированы для усиления функциональности и/или повышения растворимости в агликозилированной форме. Fc-лиганды включают, но не ограничиваются перечисленными, FcRn, FcγRs, C1q и протеины А и G, и могут быть из любого источника, включающего, но не ограниченного человеком, мышью, крысой, кроликом или обезьяной, но предпочтительным источником является человек. В альтернативном предпочтительном воплощении IgG-варианты оптимизируют, чтобы они стали более стабильными и/или более растворимыми, чем агликозилированная форма исходного IgG-варианта.
IgG-варианты могут включать модификации, которые модулируют взаимодействие с Fc-лигандами, которые не относятся к FcRn и FcγRs, которые включают, но не ограничиваются перечисленными, белки комплемента и гомологи Fc-рецепторов (FcRHs). FcRHs включают, но не ограничиваются перечисленными, FcRH1, FcRH2, FcRH3, FcRH4, FcRH5 и FcRH6 (Davis et al., 2002, Immunol. Reviews 190:123-136, работа включена путем отсылки во всей своей полноте).
Предпочтительно специфичность IgG-варианта к Fc-лиганду будет определять его терапевтическое применение. Применимость конкретного IgG-варианта для терапевтических целей будет зависеть от эпитопа или формы антигена-мишени и заболевания или симптома, которые подвергают лечению. Для большинства мишеней и показаний может быть предпочтительным усиленное связывание FcRn, так как усиленное связывание FcRn может приводить к увеличению времени полужизни в сыворотке крови. Увеличенное время полужизни в сыворотке крови позволяет использовать меньшую частоту введения доз или низкую дозировку лекарственных средств. Это может быть в особенности предпочтительно, если терапевтическое средство применяют в ответ на показания, для которых необходимо повторное введение средства. Для некоторых мишеней и показаний может быть предпочтительным пониженное сродство к FcRn. Это может быть особенно предпочтительным, если желательно иметь Fc-вариант с повышенным клиренсом или сниженным временем полужизни в сыворотке крови, например в Fc-полипептиде, применяемом в качестве визиографических средств или в виде радиоактивных терапевтических средств.
Можно применять IgG-варианты, которые включают IgG-варианты, обеспечивающие повышенное сродство к FcRn с усиленными активаторными FcγR и/или пониженное сродство к ингибиторным FcγR. Для некоторых мишеней и показаний, кроме того, может быть полезным применение IgG-вариантов, которые обеспечивают различную селективность для различных активаторных FcγR; например, в некоторых случаях может быть желательным усиленное связывание с FcγRIIa и FcγRIIIa, но не с FcγRI, тогда как в других случаях может быть предпочтительным повышенное связывание только с FcγRIIa. Для некоторых мишеней и показаний может быть предпочтительным применение IgG-вариантов, которые изменяют связывание FcRn и усиливают эффекторные функции, как опосредованные FcγR, так и эффекторные функции, опосредованные комплементом, тогда как в других случаях может быть полезным применять IgG-варианты, которые усиливают связывание FcRn или увеличивают время полужизни в сыворотке крови и усиливают эффекторные функции, как опосредованные FcγR, так и эффекторные функции, опосредованные комплементом. Для некоторых мишеней или раковых показаний может быть полезным снизить или полностью устранить одну или несколько эффекторных функций, например с помощью нокаута связывания C1q, одного или нескольких FcγR, FcRn, одного или нескольких других Fc-лигандов. Для других мишеней и показаний может быть предпочтительным применять IgG-варианты, которые обеспечивают повышенное связывание с ингибиторным FcγRIIb и, кроме того, обеспечивают связывание с активаторными FcγR на уровне WT, на пониженном уровне или не связываются с активаторными FcγR вовсе. Это может быть особенно полезным, например, если целью IgG-варианта является ингибирование воспаления или аутоиммунного заболевания или модуляция некоторым образом иммунной системы. Так как аутоиммунные заболевания в основном продолжаются в течение долгого периода времени и для лечения используют повторное введение доз, в особенности предпочтительно лечение этих заболеваний с помощью Fc-вариантов с увеличенным временем полужизни вследствие усиления FcRn.
Чтобы улучшить стабильность, растворимость, функционирование или клиническое применение IgG, можно провести модификацию. В предпочтительном воплощении IgG-варианты могут включать модификации для снижения иммуногенности у людей. В наиболее предпочтительном воплощении иммуногенность IgG-варианта снижена с помощью методов, описанных в патенте US 11/004,590, включенном путем отсылки во всей своей полноте. В альтернативных воплощениях IgG-варианты гуманизированы (Clark, 2000, Immunol Today 21:397-402, работа включена путем отсылки во всей своей полноте).
IgG-варианты могут включать модификации, которые снижают иммуногенность. Модификации, предназначенные для снижения иммуногенности, могут включать модификации, которые ухудшают связывание процессированных пептидов, происходящих из последовательности-предшественника, с белками главного комплекса гистосовместимости (МНС). Например, аминокислотные модификации можно сконструировать таким образом, чтобы в них не было бы вовсе или было бы минимальное количество иммунных эпитопов, предположительно связывающихся с высоким сродством с любыми превалирующими аллелями МНС. В этой области техники известно несколько методов идентификации в белковых последовательностях эпитопов, связывающихся с МНС, и они могут быть использованы для определения суммы баллов для эпитопов в варианте IgG. См., например, WO 98/52976; WO 02/079232; WO 00/13317; US 09/903,378; US 10/039,170; US 60/222,697; US 10/754,296; PCT WO 01/21823 и РСТ WO 02/00165; Mallios, 1999, Bioinformatics 15: 432-439; Mallios, 2001, Bioinformatics 17: 942-948; Sturniolo et al., 1999, Nature Biotech. 17: 555-561; WO 98/59244; WO 02/069232; WO 02/77187; Marshall et al., 1995, J. Immunol. 154: 5927-5933 и Hammer et al., 1994, J. Exp. Med. 180: 2353-2358, все работы включены путем отсылки во всей своей полноте. Для определения суммы баллов для связывания при взаимодействии конкретного пептида и МНС можно использовать информацию, основанную на последовательности (см., например, Mallios, 1999, Bioinformatics 15: 432-439; Mallios, 2001, Bioinformatics 17: p942-948; Sturniolo et. al., 1999, Nature Biotech. 17: 555-561, все работы включены путем отсылки во всей своей полноте).
Конструирование IgG-вариантов
Варианты настоящего изобретения могут быть сконструированы различными способами. Варианты, как описано здесь, могут быть вставками, делециями, заменами, другими модификациями или их комбинациями и другими изменениями. В частности, новейшим воплощением настоящего изобретения является конструирование вставок и делеций, которые как улучшают, так и ухудшают связывание Fc-полипептида с Fc-лигандом. Как раскрыто в этой заявке, могут быть сделаны вставки и делеции, которые повышают или понижают сродство Fc-полипептида к FcRn. Вставки и делеции могут быть сконструированы с помощью рационально продуманных подходов или с помощью подходов, которые включают применение случайных компонентов, таких как создание библиотек случайных или полуслучайных вариантов или скрининг. В альтернативном воплощении раскрыты замены, которые увеличивают или снижают сродство Fc-полипептида к FcRn.
Вставки и делеции
Вариантные Fc-полипептиды могут быть созданы путем замены исходной аминокислоты в каком-либо положении Fc-полипептида на вариантную аминокислоту. С помощью замены одной или нескольких аминокислот в Fc-полипептиде на вариантные аминокислоты в этих положениях изменяются боковые цепи. Наиболее полезные замены модифицируют свойства Fc путем изменения боковых цепей Fc. Замещенные боковые цепи могут взаимодействовать непосредственно или непрямым способом с партнером, связывающимся с Fc, ассоциированным с функцией или свойством Fc. По меньшей мере, одна замена меняет ковалентную структуру одной или нескольких боковых цепей исходного Fc-полипептида.
Альтернативно, могут быть созданы вариантные Fc-полипептиды, которые меняют ковалентную структуру основной цепи исходного Fc-полипептида. Атомы белка, входящие в состав основной цепи молекулы, включают пептидный атом азота, альфа-углеродный атом, карбонил или пептидный атом углерода и карбонильный кислород. Изменение ковалентной структуры основной цепи обеспечивает дополнительные способы изменения свойств Fc-полипептидов. Ковалентная структура основной цепи Fc может быть изменена путем добавления атомов в основную цепь молекулы, например путем вставки одной или нескольких аминокислот, или удалением атомов из основной цепи, например путем делеции одной или нескольких аминокислот. Ковалентную структуру основной цепи можно также модифицировать с помощью изменения индивидуальных атомов основной цепи на другие атомы (Deechongkit et al., J. Am. Che. Soc. 2004, 126(51):16762-71, работа включена путем отсылки во всей своей полноте). Как известно в этой области техники и проиллюстрировано здесь, вставки и делеции аминокислот в Fc-полипептидах могут быть сделаны с помощью вставок или делеций соответствующих нуклеотидов в ДНК, кодирующей Fc-полипептид. Альтернативно, как известно в этой области техники, вставки и делеции могут быть сделаны во время синтеза Fc-полипептидов.
Конструирование вставок и делеций аминокислот, которые изменяют взаимодействие Fc-полипептида с одним или несколькими партнерами по связыванию (например, с Fc-гаммаR's, FcRn, C1q) могут быть сделаны исходя из структуры комплекса Fc-полипептида и его партнера по связыванию. В менее предпочтительном воплощении конструирование можно провести исходя из структуры Fc-полипептида и информации о Fc-участке, вовлеченном в связывание с партнером по связыванию. Эту информацию можно получить с помощью экспериментов по мутагенезу, экспериментов с применением фагового дисплея, путем сравнения по гомологии, с помощью компьютерного моделирования и другими способами.
Предпочтительные положения в аминокислотной последовательности для вставок и делеции, которые влияют на взаимодействия при связывании Fc, но не оказывают влияния на общую структуру, стабильность, экспрессию или применение Fc-полипептида, находятся в участках взаимодействия Fc/партнер Fc по связыванию. Чтобы модифицировать связывание FcRn с Fc-полипептидом, предпочтительными местами расположения петель для вставок и делеций являются положения 244-257, 279-284, 307-317, 383-390 и 428-435 (нумерация в соответствии с EU-индексом в базе данных Kabat et al., Burmeister et al., 1994, Nature, 372:379-383; Martin et al., 2001, Mol Cell 7:867-877, все работы включены путем отсылки во всей своей полноте). Чтобы модифицировать связывание Fc-гамма-рецептора с Fc-полипептидом, предпочтительными местами расположения петель для вставок и делеций являются положения 229-239, 266-273, 294-299 и 324-331 (нумерация в соответствии с EU-индексом в базе данных Kabat et al., код pdb 1E4K.pdb Sondermann et al., 2000, Nature 406:267, все работы включены путем отсылки во всей своей полноте). Петли являются участками полипептида, не вовлеченными в структуру альфа-спиралей и бета-складок (van Holde, Johnson and Ho. Principles of Physical Biochemistry. Prentice Hall, New Jersey 1998, Chapter 1 pp2-67, работа включена путем отсылки во всей своей полноте). Положения петель являются предпочтительными, так как атомы основной цепи молекулы обычно более подвижны и менее вероятно вовлечены в образование водородных связей по сравнению с атомами основной цепи альфа-спиралей и бета-складок. Поэтому менее вероятно, что удлинение или укорачивание петель из-за вставки или делеции одной или нескольких аминокислот приведет к крупным, разрушительным изменениям в Fc-полипептиде, включая его стабильность, экспрессию и другие проблемы.
Вставки и делеции могут быть использованы для изменения длины полипептида. Например, в области петель изменение длины петли приведет к измененной подвижности и конформационной энтропии петли. Вставки в петлю будут в основном повышать конформационную энтропию петли, которую можно определить как константу Больцмана, умноженную на натуральный логарифм числа возможных конформаций (van Holde, Johnson and Ho. Principles of Physical Biochemistry. Prentice Hall, New Jersey 1998, Chapter 1 pp78, работа включена путем отсылки во всей своей полноте). При вставке в полипептид, по меньшей мере, одной аминокислоты общее количество конформаций, возможных в полипептиде, увеличивается. Эти дополнительные конформации могут быть полезными для образования благоприятных взаимодействий Fc/партнер Fc по связыванию, так как Fc-полипептид может использовать одну из дополнительных конформаций при связывании белка, связывающегося с Fc. В этом случае вставка может привести к более сильным взаимодействиям Fc/партнера Fc по связыванию. Если дополнительные конформации не используются на поверхности связывания, то вставка может привести к более слабым взаимодействиям Fc/партнера Fc по связыванию, так как дополнительные конформации должны конкурировать с конформацией, компетентной в отношении связывания. Сходным образом делеция полипептидного сегмента может также привести как к более прочным, так и менее прочным взаимодействиям Fc/партнера Fc по связыванию. Если делеция сегмента, которая уменьшает возможное число конформаций основной цепи молекулы, ликвидирует конформацию, компетентную в отношении связывания, то делеция может привести к более слабым взаимодействиям Fc/партнера Fc по связыванию. Если делеция сегмента не ликвидирует конформацию, компетентную в отношении связывания, то делеция может привести к более сильным взаимодействиям Fc/партнера Fc по связыванию, так как делеция может ликвидировать конформации, конкурирующие с конформацией, компетентной в отношении связывания.
Вставки и делеции могут быть использованы для изменения положений и ориентации аминокислот в Fc-полипептиде. Так как вставки и делеции могут вызывать изменения в ковалентной структуре основной цепи, они обязательно вызовут изменение в положениях атомов основной цепи. Фигура 7 сравнивает положения основной цепи в некоторых сегментах петли, обозначенных L1-L4, в трех различных основных цепях. Исходная основная структура содержит четыре сегмента петли, в то же время делеция в основной цепи приводит к потере сегмента L1, а вставка сегмента включает дополнительный сегмент, расположенный перед сегментом L1, например перед N-концом. Делеции и вставки вызывают масштабное изменение в структуре основной цепи вблизи места вставки или делеции. Из-за удаления сегмента, расположенного около N-концевой части петли, например сегмента L1, петля укорачивается, и оставшиеся сегменты смещают свое положение ближе к N-концу петли. Это оказывает эффект на передвижение сегмента L2 по направлению к прежнему расположению сегмента L1 и по направлению к N-концу петли. Это изменение положения сегмента L2 относительно сегмента L1 может усилить связывание комплекса Fc/партнера Fc по связыванию и является предпочтительным, если имеется предварительная информация, предполагающая, что аминокислота или аминокислоты, расположенные в L2, делают благоприятными взаимодействия с партнером по связыванию Fc при расположении в L1. Например, если L2 содержит аланин и тирозин и замена двух аминокислот в L1 на аланин и тирозин ранее привела к Fc-варианту с улучшенным связыванием, то делеция L1 может создать Fc-вариант с повышенным сродством к партнеру по связыванию с Fc.
Сходным образом, вставка полипептидного сегмента в Fc-полипептид на N-концевой части петли приведет к тому, что положение сегментов петли будет смещаться по направлению к С-концевой части петли. На фигуре 7 вставка одной или нескольких аминокислот перед сегментом L1, например на N-конце, изменяет конформацию основной цепи, включая перемещение сегмента L1 по направлению к С-концу петли. Такой тип вставки предпочтителен, если известно, что аминокислоты, расположенные в сегменте L1, благоприятствуют взаимодействию, когда они находятся в положении L2, так как вставка может приводить к более сильным взаимодействиям Fc/партнера Fc по связыванию. Если желательно более слабые взаимодействия Fc/партнера Fc по связыванию, то можно использовать вставку для смещения неблагоприятной аминокислоты в новое положение. Вставленный, удаленный и исходный сегменты (L1-L4 на фигуре 7) могут быть одной или несколькими аминокислотами в Fc-полипептиде.
Альтернативно, аналогично вставкам и делециям на N-концевом участке петель могут быть использованы вставки и делеции на С-концевом участке петель. Вставки на С-конце петель могут приводить к передвижению положений N-концевого участка вставки по направлению к N-концу петли. Делеции на С-конце петель могут приводить к передвижению положений N-концевого участка делеции по направлению к С-концу петли. Выбор при использовании вставки или делеции на N-концевом или С-концевом участке петли основан на аминокислотах, расположенных в петле, необходимости увеличения или уменьшения сродства в комплексе Fc/партнер Fc по связыванию и желаемого позиционного сдвига.
Вставки и делеции могут применяться в любом участке Fc-полипептида, включая участки петель, альфа-спиралей и бета-складок. Предпочтительные локализации вставок и делеции включают участки петель, которые не относятся к участкам альфа-спиралей и бета-складок. Петли предпочтительны потому, что они обычно воспринимают изменения в основной цепи лучше, чем альфа-спирали и бета-складки. Особенно предпочтительными локализациями вставок или делеций, которые приводят к более сильным белок/белковым взаимодействиям, являются N-концевой и С-концевой край петли. Если боковые цепи петли вовлечены во взаимодействия Fc/партнера Fc по связыванию, то менее вероятно, что вставки или делеция на краях приведут к очень вредным изменениям в связывающих взаимодействиях. Более вероятно, что делеции точно по центру петли ликвидируют важные остатки на поверхности взаимодействия Fc/партнера Fc по связыванию, а вставки точно по центру петли, более вероятно, создадут неблагоприятные взаимодействия на поверхности связывания Fc/партнера Fc по связыванию. Число удаляемых или вставляемых остатков может быть определено объемом желаемого изменения основной цепи, предпочтительно около 15 вставок или делеций или меньше, более предпочтительно около 10 вставок или делеций или меньше и наиболее предпочтительно около 5 вставок или делеций или меньше.
После того, как расположение и размер делеционного Fc-варианта были спроектированы, окончательно определяют полную последовательность полипептида, и полипептид можно создать с помощью способов, известных в этой области техники.
Инсерционные Fc-варианты, однако, имеют дополнительные стадии конструирования последовательности, по меньшей мере, из одной вставляемой аминокислоты. Вставки полярных остатков, включающих Ser, Thr, Asn, Gln, Ala, Gly, His, предпочтительны в положениях, которые, как ожидают, будут в Fc-полипептиде экспонированы. В особенности предпочтительны меньшие по размерам аминокислоты, включающие Ser, Thr и Ala, так как менее вероятно, что небольшой размер будет стерически препятствовать взаимодействиям Fc/партнера Fc по связыванию. Ser и Thr также обладают способностью к образованию водородных связей с атомами партнера Fc по связыванию.
Вставки также могут обладать дополнительной гибкостью, которую можно сконструировать в инсерционном полипептиде, чтобы создать благоприятные взаимодействия с партнером Fc по связыванию, как было бы необходимо в случае, если желательно более сильное связывание Fc/партнера Fc по связыванию. Длина вставки в основную цепь может быть определена путем моделирования вариантной основной цепи с помощью простой базовой последовательности, используемой для вставки. Например, можно сконструировать и смоделировать вставки полисерина, полиглицина или полиаланина различной длины. Моделирование можно проводить с помощью множества методов, включающих гомологичное моделирование, основанное на известных трехмерных структурах гомологов, включающих вставки, и с помощью компьютерного моделирования, включающего MODELLER (M.A. Marti-Renom et al., Annu. Rev. Biophys. Biomol. Struct. 29, 291-325, 2000) и ROSETTA (Kuhlman et al. (2003). Science 302, 1364-8), обе работы включены путем отсылки во всей своей полноте. Обычно вначале создают различные конформации основной цепи, а конечную структуру основной цепи можно определить после того, как будут установлены идентичности боковой цепи. Боковые цепи могут быть спроектированы с помощью алгоритмов PDA® (US 6,188,965; 6,269,312; 6,403,312; 6,801,861; 6,804,611; 6,792,356, 6,950,754 и US 09/782,004; 09/927,790; 10/101,499; 10/666,307; 10/666311; 10/218,102, все включены путем отсылки во всей своей полноте).
Вставки и делеции можно спроектировать в любых полипептидах помимо Fc-полипептидов с помощью методов, описанных здесь. Например, вставки или делеции в представителе суперсемейства TNF, APRIL, можно спроектировать, используя его трехмерную структуру (PDB код IXU1.pdb, Hymowitz, et al. (2005) J.Biol.Chem. 280:7218, включены путем отсылки во всей своей полноте). Вставки или делеции можно спроектировать для усиления связывания APRIL с его рецептором, TACI. Предпочтительными остатками петли в качестве участков для вставки или делеции являются остатки Ser118-Val124, Asp164-Phe167, Pro192-Ala198, Pro221-Lys226. Эти петли взаимодействуют с TACI в комплексе APRIL/TACI и опосредуют связывание.
Полипептиды, включающие варианты
IgG-варианты могут быть основаны на последовательностях IgG человека, и, таким образом, последовательности IgG человека применяют как «основные» последовательности, с которыми сравнивают другие последовательности, включающие, но не ограниченные последовательностями из других организмов, например последовательностями грызунов и приматов. IgG-варианты могут также включать последовательности из других классов иммуноглобулинов, таких как IgA, IgE, IgD, IgM и т.п. Предполагают, что хотя IgG-варианты конструируют в контексте одного исходного IgG, варианты могут быть сконструированы и «перенесены» в контекст другого, второго исходного IgG. Это делают путем определения «эквивалентных» или «соответствующих» остатков и замен между первым и вторым IgG, которое обычно основано на последовательности или структурной гомологии между последовательностями IgGs. Для установления гомологии аминокислотную последовательность первого IgG, указанного здесь, прямо сравнивают с последовательностью второго IgG. После процедуры выравнивания последовательностей с помощью одной или нескольких программ установления гомологии, известных специалистам в этой области (например, с помощью остатков, консервативных у различных видов), допускающих при необходимости существование вставок и делеций, чтобы сохранить выравнивание (то есть избежать исключения консервативных остатков из-за случайных вставок и делеций), устанавливают аминокислотные остатки, эквивалентные определенным аминокислотам в первичной последовательности IgG-варианта. Выравнивание консервативных остатков предпочтительно должно сохранять 100% таких остатков. Однако выравнивание более чем 75% или не менее 50% консервативных остатков также достаточно для определения эквивалентных остатков. Эквивалентные остатки можно также установить с помощью определения гомологии между первым и вторым IgG на уровне третичной структуры для IgGs, для которых третичная структура уже установлена. В этом случае эквивалентными называют остатки, для которых атомные координаты двух или нескольких атомов основной цепи определенного аминокислотного остатка в исходном полипептиде или его предшественнике (N на N, СА на СА, С на С, О на О) при выравнивании находятся в пределах 0.13 нм и предпочтительно в пределах 0.1 нм. Выравнивание достигается после того, как модель наилучшего соответствия ориентировали и располагали таким образом, чтобы обеспечить максимальное перекрывание атомных координат, относящихся к атомам белка, которые не являются атомами водорода. Независимо от того, сколько определяют эквивалентных или соответствующих остатков, и независимо от особенности исходного IgG, на основании которого создают IgGs, речь идет о том, что известные IgG-варианты могут быть спроектированы в любой второй исходный IgG, который имеет значительную гомологию в последовательности и структуре с IgG-вариантом. Таким образом, например, если получают вариантное антитело, в котором исходное антитело является IgG1 человека, с помощью методов, описанных выше или других методов определения эквивалентных остатков, вариантное тело может быть спроектировано в другое исходное антитело IgG1, которое связывает другой антиген, в исходное антитело IgG2 человека, исходное антитело IgA человека, исходное мышиное антитело IgG2a или IgG2b и т.п. Кроме того, как описано выше, контекст исходного IgG-варианта не влияет на возможность перевода IgG-вариантов в другие исходные IgGs.
Обеспечиваются способы создания, продуцирования и скрининга IgG-вариантов. Это не означает, что описанные способы ограничены рамками какого-либо практического применения или механизма действия. Скорее, предоставляемые способы предназначены для иллюстрирования в общих чертах того, что один или несколько IgG-вариантов можно сконструировать, продуцировать и отобрать экспериментально, для того чтобы получить IgG-варианты с оптимизированной эффекторной функцией. Множество методов для конструирования, продуцирования и тестирования антител и исходных вариантов описано в US 10/754,296 и US 10/672,280, оба включены путем отсылки во всей своей полноте.
Для проектирования IgG-вариантов с оптимизированной эффекторной функцией можно применять множество методов конструирования белков. В одном воплощении можно применять основанный на известной структуре метод конструирования, в котором для управления заменами применяют имеющуюся в распоряжении информацию о структуре, где замены проектируют, основываясь на их энергетическом соответствии при компьютерных расчетах. См., например, US 10/754,296 и US 10/672,280 и ссылки, процитированные в них, все они включены путем отсылки во всей своей полноте.
Для управления заменами в установленных положениях может быть использовано выравнивание последовательностей. Специалистам в этой области техники будет понятно, что использование информации о последовательностях может сдерживать введение замен, которые потенциально вредны для структуры белка. Источники последовательностей могут сильно различаться и включают в себя одну или несколько известных баз данных, включающих, но не ограниченных перечисленными источниками, базу данных Kabat (Northwestern University); Johnson & Wu, 2001, Nucleic Acids Res 29:205-206; Johnson & Wu, 2000, Nucleic Acids Res 28:214-218), базу данных IMGT (IMGT, международную информационную систему ImMunoGeneTics information system®; Lefranc et al., 1999, Nucleic Acids Res 27:209-212; Ruiz et al., 2000 Nucleic Acids Res 28:219-221; Lefranc et al., 2001, Nucleic Acids Res 29:207-209; Lefranc et al., 2003, Nucleic Acids Res 31:307-310) и VBASE, все включены путем отсылки во всей своей полноте. Информацию о последовательностях антител можно получить, собрать и/или создать из выравниваний последовательностей зародышевой линии клеток или существующих в природе антител из любого организма, включающего, но не ограниченного только млекопитающими. Специалистам в этой области техники будет ясно, что использование последовательностей, которые являются человеческими или главным образом человеческими, могут, кроме того, приносить пользу из-за их меньшей иммуногенности при введении человеку. Другие базы данных, которые обычно представляют собой базы данных нуклеиновых кислот или белков, то есть не имеют отношения только к антителам, включают, но не ограничиваются SwissProt, GenBank, Entrez и EMBL Nucleotide Sequence Database. Выравниваемые последовательности могут включать последовательности VH, VL, СН и/или CL. Существует большое количество программ выравнивания, основанных на последовательностях, и способов, известных в этой области техники, и все они находят применение для создания выравниваний последовательностей.
Альтернативно, могут применяться методы случайного или полуслучайного мутагенеза для получения аминокислотных модификаций в желаемых положениях. В этих случаях положения выбирают случайным образом или аминокислотные замены производят с помощью упрощенных приемов. Например, все остатки могут быть мутированы в аланин, этот процесс называют аланиновым сканированием. Такие способы могут быть сопряжены с более сложными инженерными подходами, в которых для отбора более высоких уровней разновидностей последовательностей применяют селекционные методы. Как известно в этой области техники, существует многообразие селекционных технологий, которые могут применяться в таких подходах и которые включают, например, технологии дисплея, например, фаговый дисплей, рибосомальный дисплей, дисплей на поверхностности клеток и т.п., как описано ниже.
Методы получения и отбора IgG-вариантов хорошо известны в этой области техники. Основные методы молекулярной биологии антител, экспрессии, очистки и скрининга описаны в книге Antibody Engineering, edited by Duebel&Kontermann, Springer-Verlag, Heidelberg, 2001; и в работе Hayhurst&Georgiou, 2001, Curr Opin Chem Biol 5:683-689; Maynard&Georgiou, 2000, Annu Rev Biomed Eng 2:339-76. Также см. методы, описанные в US 10/754,296; US 10/672,280 и US 10/822,231 и 11/124,620, все включены путем отсылки во всей своей полноте.
Предпочтительные варианты настоящего изобретения включают те, которые представлены на фигуре 8. Альтернативно предпочтительные варианты настоящего изобретения включают те, которые представлены на фигуре 9. Кроме того, альтернативно предпочтительные варианты настоящего изобретения включают те, которые представлены на фигуре 10. Особенно предпочтительные варианты настоящего изобретения включают G358H и N434Y. Наиболее предпочтительные варианты настоящего изобретения включают 257С, 257М, 257L, 257N, 257Y, 279Q, 279Y, 308F и 308Y.
Изготовление IgG-вариантов
IgG-варианты могут быть изготовлены любыми методами, известными в этой области техники. В одном из воплощений для создания нуклеиновых кислот, которые кодируют последовательности какого-либо представителя и которые могут затем быть клонированы в клетки хозяина, экспрессированы и при необходимости проанализированы, применяют последовательности IgG-вариантов. Эти способы выполняют, применяя хорошо известные процедуры, и разнообразные методы, которые могут найти применение, описаны в книге Molecular Cloning - A Laboratory Manual, 3nd Ed. (Maniatis, Cold Spring Harbor Laboratory Press, New York, 2001) и Current Protocols in Molecular Biology (John Wiley&Sons), обе включены путем отсылки во всей своей полноте. Для экспрессии белка нуклеиновые кислоты, которые кодируют IgG-варианты, могут быть включены в экспрессионный вектор. Экспрессионные векторы обычно включают белок, эффективно соединенный, то есть находящийся в функциональных взаимоотношениях с управляющими или регулирующими последовательностями, селективными маркерами и партнерами по слиянию и/или дополнительными элементами. IgG-варианты можно создать путем культивирования клетки хозяина, трансформированной нуклеиновой кислотой, предпочтительно экспрессионным вектором, включающим нуклеиновую кислоту, кодирующую IgG-варианты, в условиях, подходящих для того, чтобы индуцировать или вызвать экспрессию белка. Может быть использовано огромное разнообразие подходящих клеток хозяина, включающих, но не ограниченных перечисленными, клетки млекопитающих, бактерии, клетки насекомых и дрожжей. Например, множество клеточных линий, которые могут найти применение в настоящем изобретении, описаны в каталоге клеточных линий АТСС, доступном в американской коллекции типовых культур (American Type Culture Collection) и включенном сюда путем отсылки во всей своей полноте. Методы введения экзогенной нуклеиновой кислоты в клетки хозяина хорошо известны в этой области техники и меняются в зависимости от того, какую клетку хозяина применяют.
В предпочтительном воплощении IgG-варианты очищают или выделяют после экспрессии. Белки можно выделить или очистить с помощью многочисленных способов, известных специалистам в этой области техники. Стандартные способы очистки включают хроматографические методы, электрофорез, иммунологические способы, осаждение, диализ и изоэлектрофокусирование. Как хорошо известно в этой области техники, разнообразные природные белки связывают антитела, например бактериальные протеины A, G и L, и эти белки могут найти применение при очистке. Очистку часто можно проводить при помощи определенного партнера по слиянию. Например, белки могут быть очищены при помощи глутатионовой смолы, если используется слитый GST; при помощи N+2-аффинной хроматографии, если используется His-tag, или при помощи иммобилизованного антитела против flag при применении flag-метки. Для ознакомления с подходящими способами очистки, см. книгу Antibody Purification: Principles and Practice, 3rd Ed., Scopes, Springer-Verlag, NY, 1994, включенную сюда путем отсылки во всей своей полноте.
IgG-варианты могут быть подвергнуты скринингу при помощи различных способов, включающих, но не ограниченных перечисленными, способы, в которых применяют анализы in vitro, анализы in vivo, клеточные анализы и селекционные методики. В процедурах скрининга могут применяться автоматические и высокопроизводительные технологии скрининга. В скрининге могут применяться партнер по слиянию или метка, например иммунная метка, изотопная метка или метка, представляющая собой небольшую молекулу, например флуоресцентный или колориметрический краситель.
В предпочтительном воплощении в анализе in vitro отбирают функциональные и/или биофизические свойства IgG-вариантов. В предпочтительном воплощении белок отбирают на основании его функциональных возможностей, например способности катализировать реакцию или сродства при связывании его мишени. Анализы связывания могут быть проведены с применением целого ряда способов, известных в этой области техники, включающих, но не ограниченных перечисленными, анализы, основанные на FRET (резонансный перенос энергии флуоресценции) и BRET (резонансный перенос энергии биолюминесценции), AlphaScreen™ (гомогенный анализ близости с применением усиленной люминесценции), сцинтилляционный анализ близости, ELISA (иммуноферментный анализ), SPR (поверхностный плазменный резонанс, также известный как BIACORE®), калориметрию изотермического титрования, дифференциальную сканирующую калориметрию, гель-электрофорез и хроматографию, включающую гельфильтрацию. Эти и другие способы могут использовать преимущества некоторых партнеров по слиянию или меток. В анализах могут быть использованы разнообразные способы детекции, включающие, но не ограниченные перечисленными, хромогенные, флуоресцентные, люминесцентные и изотопные метки. Биофизические свойства белков, например стабильность или растворимость, могут быть отобраны с помощью разнообразных способов, известных в этой области техники. Стабильность белка можно определить с помощью измерения термодинамического равновесия между свернутым и развернутым состояниями. Например, IgG-варианты могут быть развернуты с помощью химического денатурирующего соединения, нагревания или рН, и за этим переходом можно следить с помощью способов, включающих, но не ограниченных перечисленными, спектроскопию кругового дихроизма, флуоресцентную спектроскопию, абсорбционную спектроскопию, спектроскопию ЯМР, калориметрию и протеолиз. Как признано специалистами в этой области техники, кинетические параметры переходов свернутого и развернутого состояния также могут быть прослежены с помощью этих и других технологий. Растворимость и общая структурная целостность IgG-вариантов может быть охарактеризована качественно и количественно с помощью широкого спектра способов, известных в этой области техники. Способы, которые могут найти применение для характеристики биофизических свойств IgG-вариантов, включают гель-электрофорез, хроматографию, такую как эксклюзионную хроматографию и обратно-фазовую высокоэффективную жидкостную хроматографию, масс-спектрометрию, ультрафиолетовую абсорбционную спектроскопию, флуоресцентную спектроскопию, спектроскопию кругового дихроизма, калориметрию изотермического титрования, дифференциальную сканирующую калориметрию, аналитическое ультрацентрифугирование, динамическое светорассеяние, протеолиз и межмолекулярное сшивание, измерение мутности, анализ задержки на фильтре, иммунологические анализы, анализ связывания флуоресцентных красителей, анализ окрашивания белков, микроскопию и детекцию агрегатов с помощью анализа ELISA или другого анализа связывания. Возможен также структурный анализ с применением кристаллографических методов, использующих рентгеновские лучи, и спектроскопия ЯМР.
Как известно в этой области техники, группа способов скрининга позволяет выбирать полезных участников библиотеки. Указанные здесь способы относят к "способам селекции", и эти способы находят применение при скрининге IgG-вариантов. Если библиотеки белков подвергают скринингу с помощью способа селекции, то размножают, выделяют и/или отмечают только тех участников библиотеки, которые являются полезными, то есть которые отвечают определенным критериям селекции. Считают, что в связи с получением только наиболее подходящих вариантов такие способы позволяют осуществлять скрининг библиотек, которые более масштабны по сравнению с библиотеками, которые отбирают с помощью способов, анализирующих индивидуальную пригодность участников библиотеки. Селекцию можно осуществить любым способом, методикой или с помощью партнера по слиянию, который ковалентно или нековалентно соединяет фенотип белка с его генотипом, то есть функцию белка с нуклеиновой кислотой, которая его кодирует. Например, использование в качестве способа селекции метода фагового дисплея становится возможным при слиянии участников библиотеки с белком гена III. В этом случае селекцию или получение IgG-вариантов, которые соответствуют определенным критериям, например сродству при связывании с белковыми мишенями, также применяют для отбора или получения нуклеиновой кислоты, которая их кодирует. После выделения ген или гены, кодирующие IgG-варианты, могут быть затем амплифицированы. Этот процесс выделения и амплификации, называемый пэннингом, может быть повторен и позволяет обогатить библиотеку полезными вариантами IgG. В конце концов, секвенирование присоединенной нуклеиновой кислоты позволяет идентифицировать ген.
В этой области техники известно множество способов селекции, которые могут найти применение для скрининга белковых библиотек. Они включают, но не ограничиваются перечисленными способами, метод фагового дисплея (Phage display of peptides and proteins: a laboratory manual, Kay et al., 1996 Academic Press, San Diego, CA, 1996; Lowman et al., 1991, Biochemistry 30:10832-10838; Smith, 1985, Science 228:1315-1317) и его модификации, такие как метод селективного инфицирования фагом (Malmborg et al., 1997, J Mol Biol 273:544-551), метод селективно инфекционного фага (Krebber et al., 1997, J Mol Biol 268:619-630) и пэннинг отложенной инфективности (Benhar et al., 200, J Mol Biol 301:893-904), дисплей на клеточной поверхности (Wittrup, 2001, Curr Opin Biotechnol, 12:395-399), например дисплей на бактериях (Georgiou et al., 1997, Nat Biotechnol 15:29-34; Geourgiou et al., 1993, Trends Biotechnol 11:6-10; Lee et al., 2000, Nat Biotechnol 18:645-648; Jim et al., 1998, Nat Biotechnol 16:576-80), на дрожжах (Boder & Wittrup, 2000, Methods Enzymol 328:430-44; Boder & Wittrup, 1997, Nat Biotechnol 15:553-557) и на клетках млекопитающих (Whitehom et al., 1995, Biotechnology 13:1215-1219), a также технологии дисплея in vitro (Amstutz et al., 2001, Curr Opin Biotechnol 12:400-405), такие как дисплей на полисомах (Matheakis et al., 1994, Proc Natl Acad Sci USA 91:9022-9026), дисплей на рибосомах (Hanes et al., 1997 Proc Natl Acad Sci USA 94:4937-4942), дисплей на мРНК (Roberts & Szostak, 1997, Proc Natl Acad Sci USA 94:12297-12302; Nemoto et al., 1997, FEBS Lett 414:405-408) и система дисплея инактивации рибосом (Zhow et al., 2002, J Am Chem Soc 124:538-543). Все источники включены в этот параграф путем отсылки во всей своей полноте.
Другие способы селекции, которые могут найти применение, включают методы, которые не относятся к дисплею, например, это способы селекции in vivo, включающие, но не ограниченные перечисленными, экспрессию в периплазме и цитометрический скрининг (Chen et al., 2001, Nat Biotechnol 19:537-542, работа включена путем отсылки во всей своей полноте), комплементационный анализ белковых фрагментов (Johnsson & Varshavsky, 1994, Proc Natl Acad Sci USA 91:10340-10344; Pelletier et al., Proc Natl Acad Sci USA 95:12141-12146, все работы включены путем отсылки во всей своей полноте) и дрожжевая система скрининга двойных гибридов (Fields & Song, 1989, Nature 340:245-246, работа включена путем отсылки во всей своей полноте), применяемая в селективном режиме (Visintin et al., 1999, Proc Natl Acad Sci USA 96:11723-11728, работа включена путем отсылки во всей своей полноте). В альтернативном воплощении селекцию можно проводить с помощью партнера по слиянию, который связывается со специфической последовательностью экспрессионного вектора, таким образом, ковалентно или нековалентно связывая партнера по слиянию и ассоциированного с ним участника библиотеки Fc-вариантов с нуклеиновой кислотой, которая их кодирует. Например, патенты US 09/642,574; US 10/080,376; US 09/792,630; US 10/023,208; US 09/792,626; US 10/082,671; US 09/953,351; US 10/097,100; US 60/366,658; PCT WO 00/22906; PCT WO 01/49058; PCT WO 02/04852; PCT WO 02/04853; PCT WO 02/08023; PCT WO 01/28702; и PCT WO 02/07466, все включенные сюда путем отсылки во всей своей полноте, описывают такого партнера по слиянию и технику, которая может найти применение. В альтернативном воплощении селекция in vivo может происходить в том случае, если экспрессия белка наделяет клетки некоторыми преимуществами роста, воспроизведения или выживаемости.
Подгруппа способов селекции, относящихся к "направленной эволюции", представляет собой способы, которые включают соединение или размножение полезных последовательностей во время селекции, иногда с помощью включения новых мутаций. Как признается специалистами в этой области техники, способы направленной эволюции могут способствовать идентификации наиболее полезных последовательностей библиотеки и могут увеличивать разнообразие отбираемых последовательностей. В этой области техники известно множество способов направленной эволюции, которые могут найти применение для скрининга библиотек IgG-вариантов и которые включают, но не ограничиваются перечисленными, перестановку ДНК (PCT WO 00/42561 A3; PCT WO 01/70347 A3), перестановку экзона (US 6,365,377; Kolkman & Stemmer, 2001, Nat Biotechnol 19:423-428), перестановку семейства (Crameri et al., 1998, Nature 391:288-291; US 6,376,246), RACHITT™ (Coco et al., 2001, Nat Biotechnol 19:354-359; PCT WO 02/06469), STEP и случайный прайминг рекомбинации in vitro (Zhao et al., 1998, Nat Biotechnol 16:258-261; Shao et al., 1998, Nucleic Acids Res 26:681-683), сборку генов, зависимую от экзонуклеазы (US 6,352,842; US 6,361,974), метод Gene Site Saturation Mutagenesis™ (US 6,358,709), метод Gene Reassembly™ (US 6,358,709), SCRATCHY (Lutz et al., 2001, Proc Natl Acad Sci USA 98:11248-11253), способы фрагментации ДНК (Kikuchi et al., Gene 236:159-167), перестановку одноцепочечной ДНК (Kikuchi et al., 2000, Gene 243:133-137) и технологию конструирования белков направленной эволюции AMEsystem™ (прикладная молекулярная эволюция) (US 5,824,514; US 5,817,483; US 5,814,476; US 5,763,192; US 5,723,323). Все источники, процитированные в параграфе, включены путем отсылки во всей своей полноте.
В предпочтительном воплощении IgG-варианты подвергают скринингу, применяя один или несколько клеточных анализов или анализов in vivo. Для таких анализов обычно добавляют экзогенно очищенные или неочищенные белки, так чтобы клетки были подвергнуты воздействию определенных вариантов или пула вариантов, принадлежащих библиотеке. Эти анализы обычно, но не всегда, основаны на функции IgG, то есть на способности IgG связывать свой антиген-мишень и опосредовать некоторые биохимические события, например эффекторную функцию, ингибирование связывания лиганда/рецептора, апоптоз и т.п. Такие анализы часто включают мониторинг ответа клеток на IgG, например, такого как выживание клеток, клеточная смерть, изменение в клеточной морфологии или активация транскрипции, например клеточная экспрессия природного гена или репортерного гена. Например, в таких анализах можно измерять способность IgG-вариантов вызывать ADCC, ADCP или CDC. Для некоторых анализов кроме клеток-мишеней может быть необходимым добавление дополнительных клеток или компонентов, например сывороточного комплемента или эффекторных клеток, таких как моноциты периферической крови (РВМС), NK-клетки, макрофаги и т.п. Такие дополнительные клетки могут происходить из любого организма, но предпочтительно, чтобы они были клетками человека, мышей, крыс, кроликов и обезьян. Антитела могут вызвать апоптоз некоторых клеточных линий, экспрессирующих мишень, или они могут опосредовать атаку на клетки-мишени с помощью иммунных клеток, которые были добавлены для анализа. В этой области техники известны способы мониторинга клеточной смерти или жизнеспособности, и они включают применение красителей, иммунохимических, цитохимических и радиоактивных реагентов. Например, анализ с окрашиванием каспаз дает возможность измерять уровень апоптоза, а поглощение и высвобождение радиоактивных субстратов или флуоресцентных красителей, таких как Аламаровый синий, позволяет наблюдать за клеточным ростом или клеточной активацией. В предпочтительном воплощении применяют анализ цитотоксичности DELFIA®EuTDA-based cytotoxicity assay (Perkin Elmer, MA). Альтернативно, за мертвыми или поврежденными клетками-мишенями можно следить с помощью высвобождения одного или нескольких природных внутриклеточных белков, например лактатдегидрогеназы. Активация транскрипции также может служить способом анализа функции в клеточных измерениях. В этом случае за ответом можно следить с помощью природных генов или белков, которые могут регулироваться с помощью активации, например, можно измерить высвобождение определенных интерлейкинов, или, альтернативно, ответ может быть опосредован репортерной конструкцией. Клеточные анализы могут также включать измерение морфологических изменений клеток в ответ на присутствие белка. Типы клеток в таких анализах могут быть прокариотическим или эукариотическими, и можно применять разнообразные клеточные линии, которые известны в этой области техники. Альтернативно, отбор с использованием клеток проводят с помощью клеток, которые были трансформированы или трансфицированы с помощью нуклеиновой кислоты, кодирующей варианты. То есть IgG-варианты не добавляют экзогенно к клеткам. Например, в одном из воплощений в отборе, основанном на применении клеток, используют метод представления на клеточной поверхности. Можно применять партнера по слиянию, который делает возможным представление IgG-вариантов на поверхности клеток (Witrrup, 2001, Curr Opin Biotechnol, 12:395-399, работа включена путем отсылки во всей своей полноте).
В предпочтительном воплощении иммуногенность IgG-вариантов определяют экспериментально, применяя один или несколько клеточных анализов. Для экспериментального подтверждения эпитопов можно применять несколько методов. В предпочтительном воплощении для экспериментальной количественной оценки иммуногенности применяют активацию Т-клеток ex vivo. В этом методе антиген-презентирующие клетки и нативные Т-клетки из подходящих доноров один или несколько раз провоцируют пептидом или целым белком, представляющим интерес. Затем активацию Т-клеток детектируют, применяя некоторое количество методов, например с помощью мониторинга за продукцией цитокинов или измерения поглощения тимидина, меченного тритием. В наиболее предпочтительном воплощении следят за продукцией гамма-интерферона, используя анализ Elispot (Schmittel et al., 2000, J. Immunol. Meth. 24:17-24, работа включена путем отсылки во всей своей полноте).
Биологические свойства IgG-вариантов можно охарактеризовать в экспериментах на клетках, тканях или целом организме. Как известно в этой области техники, лекарственные препараты часто испытывают на животных, включающих, но не ограниченных перечисленными, мышей, крыс, кроликов, собак, кошек, свиней и обезьян, для того чтобы определить эффективность лекарственных средств для лечения болезней или в моделях заболеваний; или чтобы измерить фармакокинетику, токсичность и другие свойства лекарственных средств. Животные могут быть отнесены к моделям заболеваний. Терапевтические средства часто испытывают на мышах, включающих, но не ограниченных перечисленными, безволосых мышей, мышей SCID, ксенотрансплантантных мышей и трансгенных мышей (включая нокины и нокауты). Такой эксперимент может предоставить очень важные сведения для определения потенциала белка, применяемого в качестве терапевтического средства. Любой организм, желательно млекопитающее, может быть использован в испытаниях. Например, в связи с тем, что генетически обезьяны схожи с человеком, они могут быть подходящими терапевтическими моделями, и таким образом могут быть использованы для тестирования эффективности, токсичности, фармакокинетики или других свойств IgG. В конечном итоге для апробации их в качестве лекарственных средств требуются испытания на людях, и, конечно поэтому, такие эксперименты предусмотрены. Таким образом, IgG могут быть испытаны на людях для определения терапевтической эффективности, токсичности, иммуногенности, фармакокинетики и/или других клинических свойств.
Способы применения IgG-вариантов
IgG-варианты могут найти применение в широком диапазоне продуктов. В одном из воплощений IgG-вариант является терапевтическим, диагностическим или исследовательским реагентом, предпочтительно терапевтическим средством. IgG-вариант может найти применение в композиции антител, которые могут быть моноклональными или поликлональными. В предпочтительном воплощении IgG-варианты применяют для уничтожения клеток-мишеней, которые несут антиген-мишень, например раковых клеток. В альтернативном воплощении IgG-варианты применяют для того, чтобы блокировать антиген-мишень, быть антагонистами или агонистами антигена-мишени, например, чтобы быть антагонистами для цитокина или цитокинового рецептора. В альтернативном предпочтительном воплощении IgG-варианты применяют для того, чтобы блокировать антиген-мишень, быть антагонистами или агонистами антигена-мишени и чтобы уничтожать клетки-мишени, несущие антиген-мишень.
IgG-варианты могут применяться в различных терапевтических целях. В предпочтительном воплощении антитела, включающие IgG-вариант, вводят пациенту для лечения антитело-зависимого расстройства. Термин «пациент» в целях настоящего изобретения включает как людей, так и других животных, предпочтительно млекопитающих и наиболее предпочтительно людей. Термины «антитело-зависимое расстройство», или «антитело-чувствительное расстройство», или «состояние», или «заболевание» здесь означают расстройство, которое может быть улучшено введением фармацевтической композиции, включающей IgG-вариант. Антитело-зависимые расстройства включают, но не ограничиваются ими, аутоиммунные заболевания, иммунологические заболевания, инфекционные заболевания, воспалительные заболевания, неврологические заболевания и онкологические и неопластические заболевания, включающие рак. Термины «рак» и «раковый» имеют отношение или описывают физиологическое состояние у млекопитающих, которое обычно характеризуют неконтролируемым клеточным ростом. Примеры рака включают, но не ограничиваются ими, карциному, лимфому, бластому, саркому (включая липосаркому), нейроэндокринные опухоли, мезотелиому, шваному, менингиому, аденокарциному, меланому и лейкемию или лимфоидные злокачественные новообразования.
В одном воплощении IgG-вариант является единственным терапевтически активным средством, вводимым пациенту. Альтернативно, IgG-вариант вводят вместе с одним или несколькими терапевтическими средствами, включающими, но не ограниченными ими, цитотоксические средства, химиотерапевтические средства, цитокины, средства, ингибирующие рост, антигормональные средства, ингибиторы киназ, антиангиогенные средства, кардиопротекторы или другие терапевтические средства. IgG-варианты могут быть введены дополнительно к одному или нескольким другим терапевтическим режимам. Например, IgG-вариант может быть введен пациенту вместе с химиотерапией, радиотерапией или одновременно с химиотерапией и радиотерапией. В одном из воплощений IgG-вариант может быть введен в сочетании с одним или несколькими антителами, которые могут быть или не быть IgG-вариантом. В соответствии с другим воплощением для воздействия на раковые клетки ex vivo применяют IgG-вариант и один или несколько видов противораковой терапии. Полагают, что такое воздействие ех vivo может быть полезным при трансплантации костного мозга и, в особенности, при аутогенной трансплантации костного мозга. Конечно же, полагают, что IgG-варианты можно применять еще и в комбинации с другими терапевтическими методиками, такими как хирургия.
Для введения вместе вариантами IgG могут найти применение множество других терапевтических средств. В одном из воплощений IgG вводят вместе с антиангиогенным средством. Термин «антиангиогенное средство», который здесь используют, означает соединение, которое блокирует или в некоторой степени мешает развитию кровеносных сосудов. Антиангиогенное средство, например, может быть небольшой молекулой или белком, например антителом, слитым Fc или цитокином, которые связываются с фактором роста или рецептором фактора роста, вовлеченными в стимуляцию ангиогенеза. Предпочтительным антиангиогенным фактором здесь является антитело, которое связывается с эндотелиальным фактором роста сосудов (VEGF). В альтернативном воплощении IgG вводят вместе с терапевтическим средством, которое включает или усиливает адаптивный иммунный ответ, например, с антителом, мишенью которого является CTLA-4. В альтернативном воплощении IgG вводят вместе с ингибитором тирозинкиназы. Термин «ингибитор тирозинкиназы». который здесь используют, означает молекулу, которая до некоторой степени ингибирует тирозинкиназную активность тирозинкиназы. В альтернативном воплощении IgG-варианты вводят вместе с цитокином.
Предусмотрены фармацевтические композиции, в состав которых входят IgG-вариант и одно или несколько терапевтически активных средств. Композиции IgG-вариантов получают для хранения путем смешивания IgG, имеющего желаемую степень чистоты, с необязательными фармацевтически приемлемыми носителями, наполнителями или стабилизаторами (Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed., 1980, работа включена путем отсылки во всей своей полноте) в виде лиофилизированных композиций или водных растворов. Композиции, применяемые для введения in vivo, предпочтительно являются стерильными. Это легко сделать с помощью фильтрации через стерильные фильтрационные мембраны или другими способами. IgG-варианты и другие терапевтически активные средства, раскрытые здесь, можно применять в композиции в виде иммунолипосом и/или поместить в микрокапсулы.
Концентрация терапевтически активного IgG-варианта в композиции может варьировать приблизительно от 0,1 до 100% по массе. В предпочтительном воплощении концентрация IgG находится в диапазоне от 0,003 до 1,0 М. Для лечения пациента можно вводить терапевтически эффективную дозу IgG-варианта. Термин «терапевтически эффективная доза» здесь означает дозу, которая вызывает эффекты, для которых ее вводят. Точная доза будет зависеть от цели лечения и устанавливается специалистом с использованием известных методик. Дозировки могут составлять от 0,01 до 100 мг/кг веса тела или более, например 0,01, 0,1, 1,0, 10 или 50 мг/кг веса тела, предпочтительно от 1 до 10 мг/кг. Как известно в этой области техники, может понадобиться корректировка, учитывающая деградацию белка, соотношение системной доставки против локализованной доставки, и уровень синтеза новых протеаз, а также возраст, вес тела, общее состояние здоровья, пол, питание, время введения, взаимодействие лекарств и тяжесть состояния, и это будет выяснено специалистами в процессе рутинной экспериментальной работы.
Введение фармацевтической композиции, включающей IgG-вариант, предпочтительно в форме стерильного водного раствора, может быть осуществлено многими способами, включающими, но не ограниченными только ими, пероральный, подкожный, внутривенный, парентеральный, внутриносовой, внутриушной, внутриглазной, ректальный, вагинальный, чрескожный, поверхностный (например, при применении гелей, мазей, лосьонов, кремов и т.д.), внутрибрюшинный, внутримышечный, внутрилегочный (например, при применении метода ингаляции AERx®, коммерчески доступного от компании «Aradigm», или при применении системы доставки в легкие Inhance™, коммерчески доступной от компании «Nektar Terapeutics») способы введения и т.д. Терапевтические средства, описанные здесь, могут быть дополнительно введены вместе с другими терапевтическими средствами, то есть терапевтические средства, описанные здесь, могут быть введены совместно с помощью других видов терапии или вместе с терапевтическими средствами, которые включают, например, небольшие молекулы, другие биологические соединения, радиационную терапию, хирургию и т.п.
Примеры
Ниже представлены примеры, иллюстрирующие настоящее изобретение. Это не означает, что приведенные примеры ограничивают объем настоящего изобретения любыми частными способами применения или теориями действия. Во всех положениях, обсуждаемых в настоящем изобретении, нумерация соответствует индексу EU в базе данных Kabat (Kabat et al., 1991, Sequences of Proteins of Immunological Interest, 5th Ed., United States Public Health Service, National Intitules of Health, Bethesda, работа включена путем отсылки во всей своей полноте). Специалисты в этой области техники признают, что эта договоренность включает непоследовательную нумерацию в определенных участках последовательности иммуноглобулинов, что позволяет нормализованно ссылаться на консервативные положения в семействах иммуноглобулинов. Таким образом, положения любого предоставленного иммуноглобулина, определяемые с помощью индекса EU, не обязательно совпадают с его последовательной нумерацией.
Пример 1: Конструирование ДНК, экспрессия и очистка Fc-вариантов
Fc-варианты были сконструированы с использованием Fc-домена IgG1 человека и вариабельной области трастузумаба (Herceptin®, Genentech). Fc-полипептиды были частью алемтузумаба, трастузумаба или АС10. Алемтузумаб (Campath®, торговая марка фирмы «Millenium») представляет собой гуманизированное моноклональное антитело, одобренное в настоящее время для лечения хронической В-клеточной лимфоцитарной лейкемии (Hale et al., 1990. Tissue Antigens 35:118-127, работа включена путем отсылки во всей своей полноте). Трастузумаб (Herceptin®, торговая марка фирмы «Genentech») представляет собой антитело к Her2/neu для лечения метастатического рака молочной железы. АС10 представляет собой моноклональное антитело к CD30. Вариабельную область герцептина составляли, используя метод рекурсивной ПЦР. Эту вариабельную область затем клонировали вместе с IgG1 в вектор pcDNA3.1/Zeo(+) (Invitrogen), показанный на фигуре 11. Плазмиды размножали в клетках One Shot TOP10 Е. coli (Invitrogen) и очищали с помощью набора Hi-Speed Plasmid Maxi Kit (Qiagen). Плазмиды секвенировали, чтобы проверить на присутствие клонированных вставок.
Сайт-направленный мутагенез проводили с помощью метода Quikchange™ (Stratagene). Плазмиды, содержащие желаемые замены, вставки и делеции, размножали в клетках One Shot TOP10 Е. coli (Invitrogen) и очищали с помощью набора Hi-Speed Plasmid Maxi Kit (Qiagen). ДНК секвенировали для подтверждения точности последовательностей.
Плазмиды, содержащие ген тяжелых цепей (VH-Cγ1-Cγ2-Сγ3) (дикий тип или варианты), ко-трансфицировали с плазмидой, содержащей ген легких цепей (VL-Cκ), в клетки 293Т. Среду собирали через 5 дней после трансфекции и антитела из супернатанта очищали с помощью аффинной хроматографии на колонке с протеином A (Pierce). Концентрацию антител определяли с помощью анализа с бицинхониновой кислотой (ВСА) (Pierce).
Пример 2: Измерения сродства связывания
Связывание Fc-полипептидов с Fc-лигандами анализировали с помощью измерения поверхностного плазменного резонанса. Измерения поверхностного плазменного резонанса (SPR) проводили с помощью прибора BIAcore 3000 (BIAcore AB). Антитело дикого типа или вариантное антитело фиксировали с помощью иммобилизованного протеина L (Pierce Biotechnology, Rockford, IL) и измеряли связывание с рецепторным аналитом. Протеин L ковалентно связывали с сенсорным чипом СМ5 в концентрации 1 мкМ в 10 мМ ацетате натрия, рН 4,5 на сенсорном чипе СМ5, используя N-гидроксисукцинимид/N-этил-N'-(-3-диметиламино-пропил)карбодиимид (NHS/EDC) со скоростью потока 5 мкл/мин. Проточную кювету 1 каждого сенсорного чипа обрабатывали NHS/EDC в качестве отрицательного контроля связывания. Рабочим буфером был 0,01 М HEPES рН 7,4, 0,15М NaCl, 3 мМ ЭДТА, 0,005% масс/масс поверхностно-активное вещество Р20 (HBS-EP, Biacore, Upsala, Sweden), а буфером для регенерации чипа был 10 мМ глицин НС1 рН 1,5. 125 нМ антитело трастузумаб, дикого типа или вариант, связывали с протеин L-СМ5-чипом в HBS-EP при скорости 1 мкл/мин в течение 5 минут. Аналит FcRn-His-GST, FcRn, слитый с His-меткой и глутатион-S-трансферазой, в серийном разведении в диапазоне от 1 до 250 нМ, вводили инъекцией для ассоциации в течение 20 минут, для диссоциации в течение 10 минут в HBS-EP при скорости 10 мкл/мин. Ответ, измеряемый в резонансных единицах (RU) и отражающий близкое к стационарному связывание, обнаруживали через 1200 секунд после инъекции рецептора. Цикл, проведенный только с антителом и буфером, обеспечивал базовый уровень сигнала. Строили график зависимости RU от 1/log концентрации и аппроксимировали в сигмоидальную кривую доза-ответ с помощью метода нелинейной регрессии, используя программу GraphPad Prism.
Связывание Fc-полипептидов с Fc-лигандами также проводили с помощью анализа AlphaScreen™ (гомогенный анализ близости с применением усиленной люминесценции). AlphaScreen™ - нерадиоактивный люминесцентный анализ измерения близости. Возбуждение лазером донорных бусинок возбуждает кислород, который в случае близко расположенных акцепторных бусинок будет вызывать каскад хемилюминесцентных явлений, приводящих в конце концов к эмиссии флуоресценции при 520-620 нм. Принципиальным преимуществом анализа AlphaScreen™ является его чувствительность. Так как одна донорная бусинка выделяет до 60000 возбужденных молекул кислорода в секунду, амплификация сигнала становится чрезвычайно высокой, что позволяет детектировать величины аттомолярного уровня (10-18). Антитело дикого типа биотинилировали с помощью стандартных методов для присоединения к стрептавидиновым донорным бусинкам и меченый Fc-лиганд, например FcRn, присоединяли к акцепторным бусинкам, хелатирующим глутатион. AlphaScreen™ применяли в качестве анализа прямого связывания, в котором взаимодействие Fc/Fc-лиганд приводило к объединению донорных и акцепторных бусинок. Кроме того, AlphaScreen™ применяли в конкурентном анализе для скрининга сконструированных Fc-полипептидов. В отсутствие конкурирующих Fc-полипептидов антитело дикого типа и FcRn взаимодействовали и давали сигнал при 520-620 нм. Немеченые Fc-домены конкурировали с взаимодействием Fc дикого типа/FcRn, количественно уменьшая флуоресценцию, что дает возможность определить относительное сродство связывания.
Пример 3: FcRn-связывающие свойства Fc-вариантов.
Сродство связывания Fc IgG1 с FcRn измеряли с вариантными антителами, используя AlphaScreen™. Fc-полипептиды были частью алемтузумаба или трастузумаба. Алемтузумаб (Campath®, Ilex) представляет собой гуманизированное моноклональное антитело, одобренное в настоящее время для лечения хронической В-клеточной лимфоцитарной лейкемии (Hale et al., 1990. Tissue Antigens 35:118-127, работа включена путем отсылки во всей своей полноте). Трастузумаб (Herceptin®, «Genentech») представляет собой антитело к Her2/neu для лечения метастатического рака молочной железы.
Собирали данные конкурентного анализа AlphaScreen™ для измерения относительного связывания Fc-вариантов по сравнению с антителом дикого типа при рН 6,0. Примеры сигнала AlphaScreen™ как функции конкурентного антитела представлены на фигуре 12. Кривые для 12 вариантов: P257L, P257N, V279E, V279Q, V279Y, ^281S, E283F, V284E, L306Y, T307V, V308F и Q311V, показывают повышенное сродство, так как на графике в соответствующем окошке кривая для каждого варианта смещается влево от кривой для дикого типа. Данные конкурентного анализа AlphaScreen™ для Fc-вариантов настоящего изобретения суммированы на фигурах 13 и 14. Составлена таблица относительного связывания FcRn вариантов по сравнению с диким типом. Значения больше чем 1 демонстрируют улучшенное связывание Fc-варианта с FcRn по сравнению с диким типом. Например, вариант E283L и V284E демонстрируют связывание, которое в 9,5 и 26 раз сильнее, чем у дикого типа, соответственно. Измерения поверхностного плазменного резонанса многих вариантов также показывают повышенное связывание с FcRn, как показано на фигурах 15 и 16.
В положении 257 все варианты, у которых убрали иминокислоту пролин и заменили аминокислоту, у которой в основной цепи нет атома N, на ковалентную связь боковой цепи, обеспечивают большую подвижность основной цепи, которая предоставляет большую свободу Fc-домена для лучшего связывания FcRn. В частности, варианты с L и N в положении 257 обладают сильным связыванием FcRn при рН 6, что демонстрирует, что 4-атомная боковая цепь и гамма-разветвленный элемент боковой цепи помогает Fc-домену делать продуктивным, то есть более сильным, взаимодействие с FcRn. Положение 308 взаимодействует с положением 257. Оба эти положения, в свою очередь, взаимодействуют с Н310, который прямо вовлечен во взаимодействия Fc/FcRn (таблица 2, Burmeister et al., 1994, Nature, 372:379-383, работа включена путем отсылки во всей своей полноте). Сродство Fc-вариантов V308F и V308Y к FcRn в 2,9 раза и в 4,3 раза выше по сравнению с диким типом (фигура 13). Положения 279 и 385 взаимодействуют с FcRn, так как все варианты V279E, V279Q и V279Y, и G385H, и G385N сильнее связываются с FcRn. Все эти варианты имеют отношение к аминокислотам, которые способны к образованию водородных связей.
Fc-вариант N434Y обладает особенно сильным связыванием с FcRn при рН 6,0, как показано на фигуре 13. Единичный вариант N434Y обладает 16-кратным повышенным связыванием. Комбинации этих вариантов с другими модификациями приводят к еще более прочному связыванию. Например, P257L/N434Y, ^281S/N434Y и V308F/N434Y демонстрируют усиление связывания FcRn в 830,180 и 350 раз.
Пример 4: Варианты, включающие вставки и делеции
Были сконструированы вставки и делеции, которые изменяют силу взаимодействия Fc/FcRn и были измерены их связывающие свойства в отношении различных Fc-лигандов. Для усиления FcRn-связывающих свойств Fc-домена был спроектирован Fc-вариант с вставленным остатком Ser между 281 и 282 остатками, в соответствии с EU-индексом по Kabat et al. Этот вариант нывается ^281S, где символ «^» означает вставку сразу после указанного положения. Вставленной последовательности, которая может включать более одного остатка, присваивают следующий позиционный номер. Этот Fc-вариант был сконструирован в каппа-IgG1-антителе трастузумабе (Herceptin®, «Genentech») с помощью способов, раскрытых в этой заявке. Вставка на участке между остатками 281 и 282 сдвигает остатки петли Fc к С-концевой части остатка 281 по направлению к С-концевой части петли и изменяет расположение боковой цепи. Предполагают, что смещение в С-концевой части петли у Fc-вариантов, включающих замены в положениях 282, 283 и 284, было выгодным (смотри фигуру 14). Для перемещения положения FcRn-связывающей петли был также сконструирован другой вариант с делецией N286, называемый иногда N286#. Оба этих варианта демонстрируют усиленное сродство с FcRn при рН 6,0.
Результаты анализа AlphaScreen™ показывают связывание варианта ^281S и других вариантов с FcRn. Эти результаты AlphaScreen™ собирали в качестве прямого анализа связывания. Более высокие уровни хемилюминесцентного сигнала демонстрируют более прочное связывание. При повышении концентраций вариантов в анализе получают более мощные сигналы. Эти данные при рН 6,0, представленные на фигурах 17а и 17b, показывают повышенное сродство вариантов ^281S, P257L, P257L/^281S (комбинация вариантов с заменой и вставкой) и других вариантов по сравнению с Fc дикого типа. Также показана двойная замена, T250Q/M428L, которая, как было показано ранее, имеет увеличенное время полужизни в сыворотке крови у обезьян (Hinton et al., 2004, J. Biol. Chem. 279(8):6213-6216, работа включена путем отсылки во всей своей полноте). Вставка ^281S сама по себе усиливает связывание Fc/FcRn. Кроме того, ^281S повышает связывание P257L, если в варианте P257L/^281S объединяют две модификации, как показано в результатах для точки ~40 нМ. Результаты, приведенные на фигуре 17с, демонстрируют, что эти варианты не обладают повышенным связыванием FcRn при рН 7,0. Пониженное сродство при рН 7,0 желательно для увеличения времени полужизни in vivo, так как это дает возможность высвобождению Fc-полипептидов из FcRn во внеклеточное пространство, что является важной стадией в рециклировании Fc.
Эксперименты с поверхностным плазменным резонансом также демонстрируют улучшенное связывание ^281S с FcRn. Фигура 18 показывает единицы сигнала, возникающие при связывании различных Fc-вариантов с FcRn на поверхности чипов. После того, как вариант полностью связывается с чипом, записывают единицы сигнала, которые показаны на ординате. Вставка ^281S демонстрирует связывающие свойства, сравнимые с другими вариантами, показанными здесь, которые обладают повышенным сродством к FcRn по сравнению с диким типом (см., например, фигуры 13, 14 и 15).
Делеционный вариант N286#, включающий делецию N286, также демонстрирует повышенное сродство к FcRn по сравнению с диким типом. Этот вариант обладает 2-кратным увеличением сродства к FcRn, как показано на фигуре 13, также здесь представлены данные анализа AlphaScreen™ по исследованию конкуренции при рН 6,0. Для ингибирования связывания с Fc дикого типа, присоединенного к донорным бусинкам, с FcRn, присоединенным с акцепторными бусинками, использовали различные варианты. Для ингибирования связывания донорных и акцепторных бусинок через комплекс Fc/FcRn требовалось в два раза меньше свободного N286# по сравнению со свободным Fc дикого типа. Это показывает 2-кратное усиление связывания N286# по сравнению с диким типом.
Другие Fc-варианты, включающие вставки и делении, обладали пониженным сродством к FcRn. Инсерпионный вариант ^254N обладает значительно сниженным связыванием FcRn, как следовало ожидать из природы варианта и его расположения. В этом варианте вставка Asn размещена в середине FcRn-связывающей петли. Эта вставка обладает лишь 1,1% связывающего сродства по сравнению с диким типом (фигура 13).
Несмотря на то, что частные воплощения настоящего изобретения были описаны выше в целях иллюстрации, специалистам в этой области техники будет очевидно, что можно осуществлять множество модификаций деталей без отступления от изобретения, как оно описано в прилагаемой в конце текста формуле изобретения. Все процитированные здесь публикации включены путем отсылки во всей своей полноте.
| название | год | авторы | номер документа |
|---|---|---|---|
| Fc-ВАРИАНТЫ С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С FcRn | 2008 |
|
RU2517621C2 |
| ВАРИАНТЫ Fc С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ C FcRn | 2008 |
|
RU2529951C2 |
| ВАРИАНТЫ Fc С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С FcRn | 2021 |
|
RU2833597C2 |
| ВАРИАНТЫ Fc С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С FcRn | 2014 |
|
RU2700882C2 |
| FC ВАРИАНТЫ АНТИТЕЛА | 2012 |
|
RU2607014C2 |
| АФФИННАЯ ХРОМАТОГРАФИЯ С ПРИМЕНЕНИЕМ FC-РЕЦЕПТОРОВ | 2013 |
|
RU2624128C2 |
| ОПТИМИЗИРОВАННЫЕ Fc-ВАРИАНТЫ, ИМЕЮЩИЕ ИЗМЕНЕННОЕ СВЯЗЫВАНИЕ С FcγR, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2004 |
|
RU2337107C2 |
| ИНТРАТЕКАЛЬНОЕ ВВЕДЕНИЕ ВЕКТОРОВ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННЫХ ВИРУСОВ ДЛЯ ГЕННОЙ ТЕРАПИИ | 2016 |
|
RU2775138C2 |
| МОДИФИЦИРОВАННЫЕ АСИММЕТРИЧНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕ Fc-РЕЦЕПТОР, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2687043C2 |
| Химерный улучшенный рекомбинантный фактор VIII | 2015 |
|
RU2647769C2 |









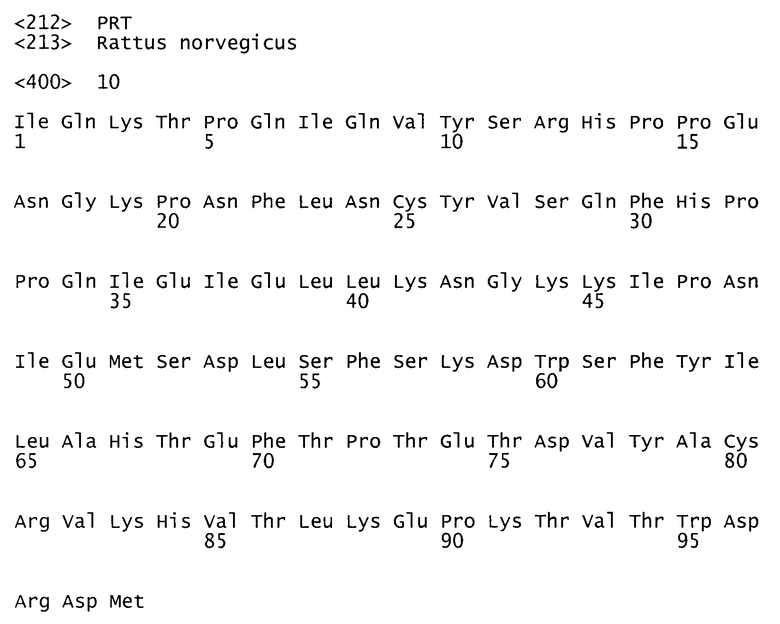
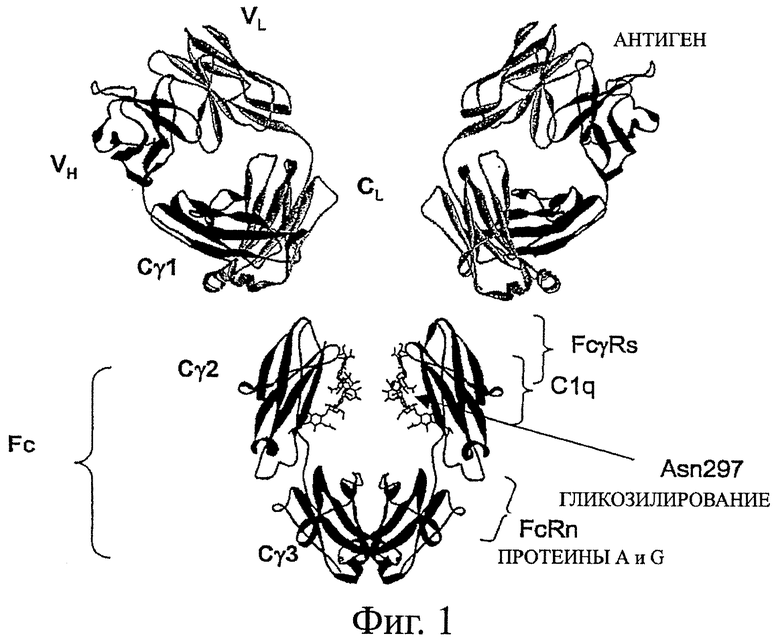
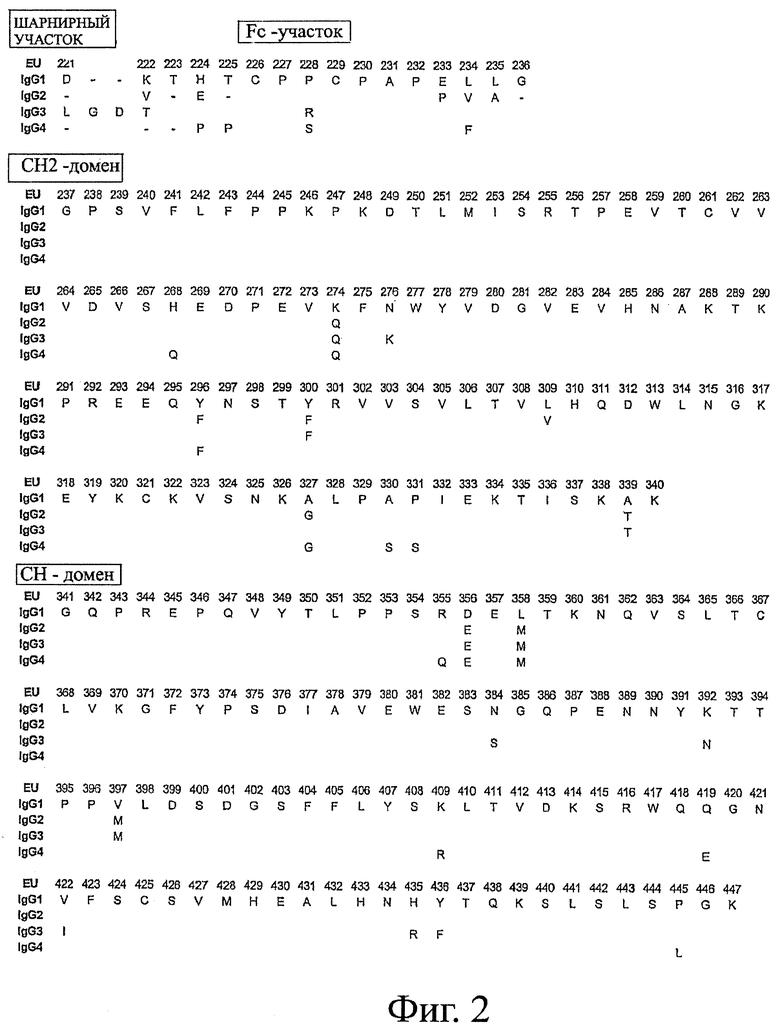


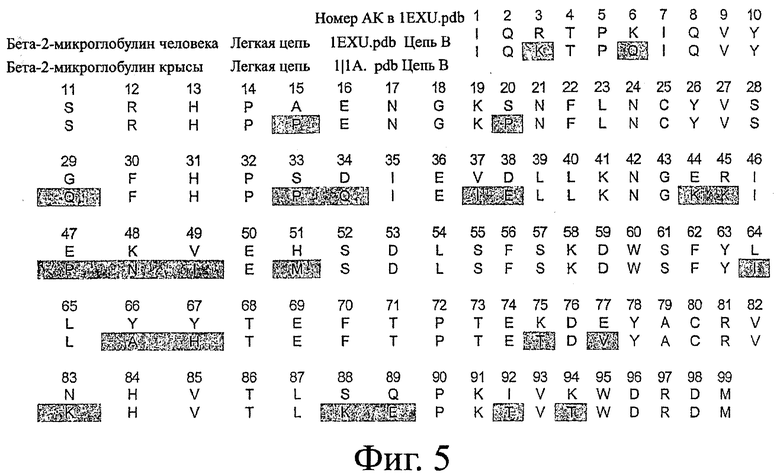
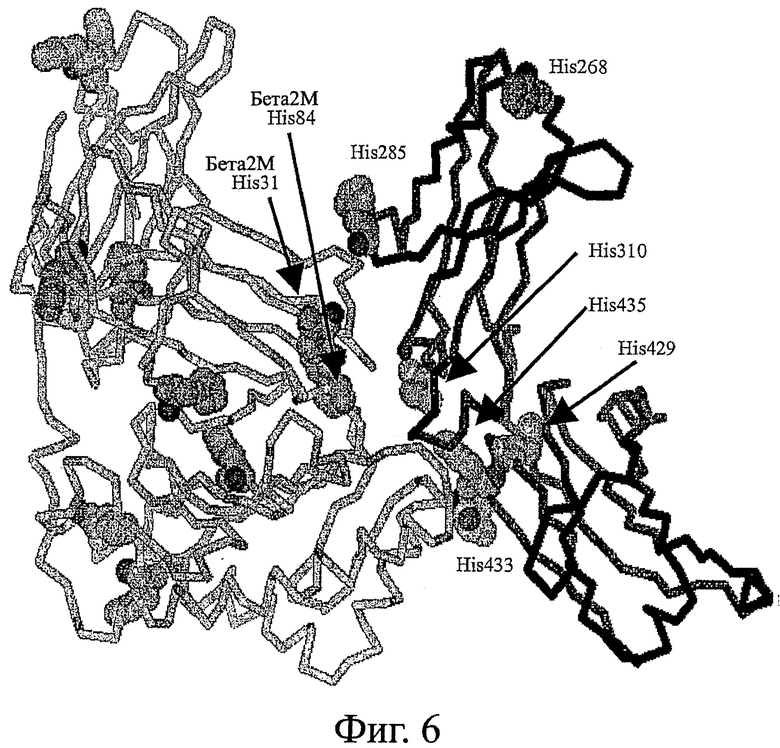
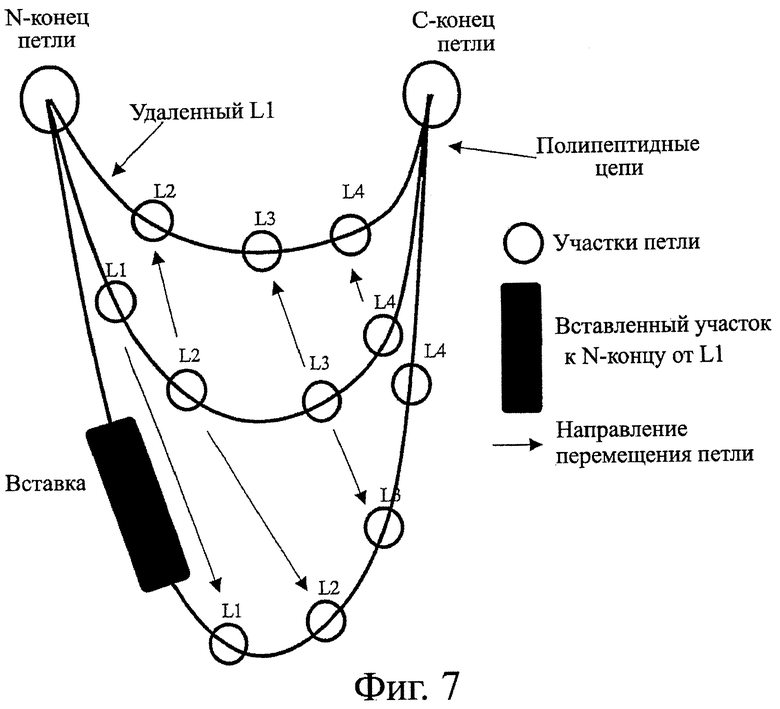
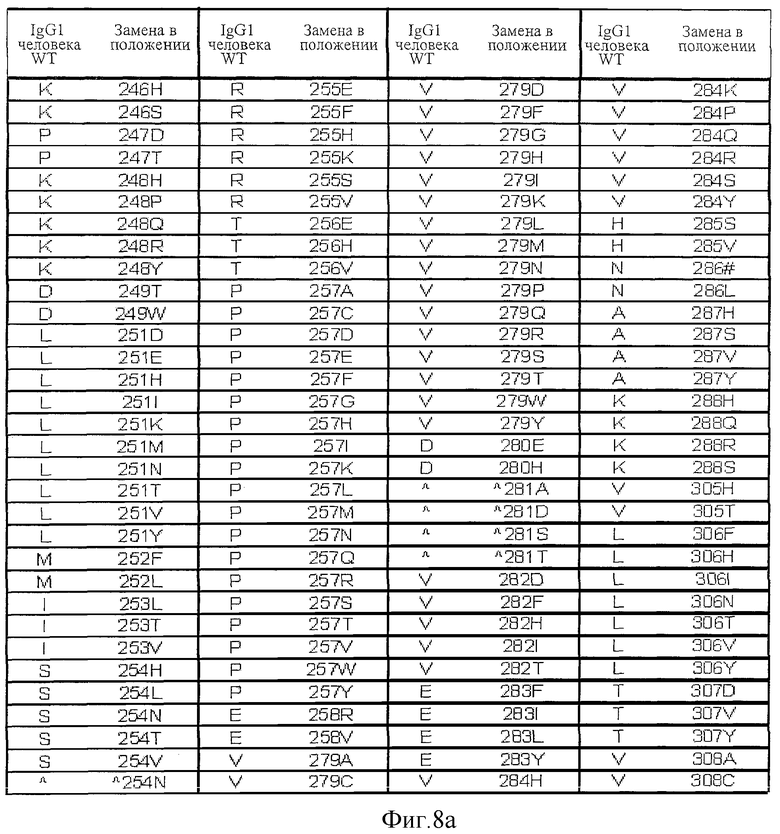
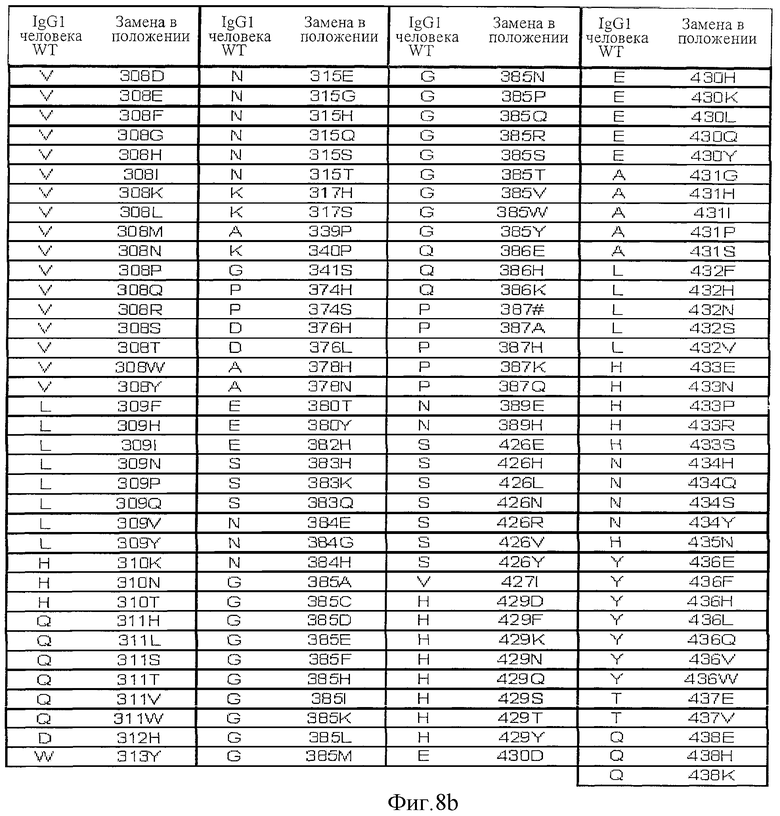
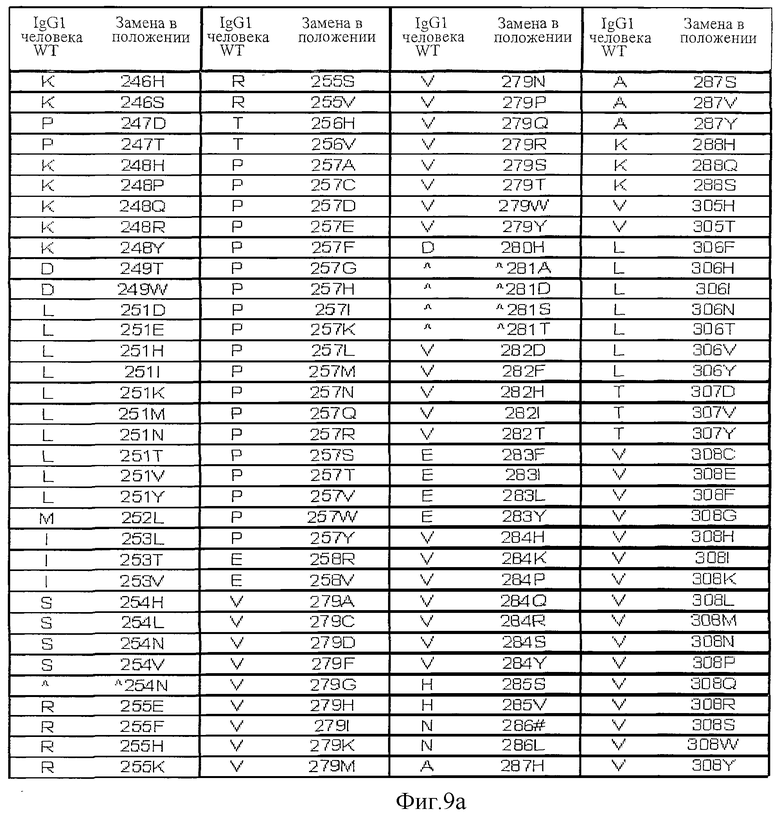

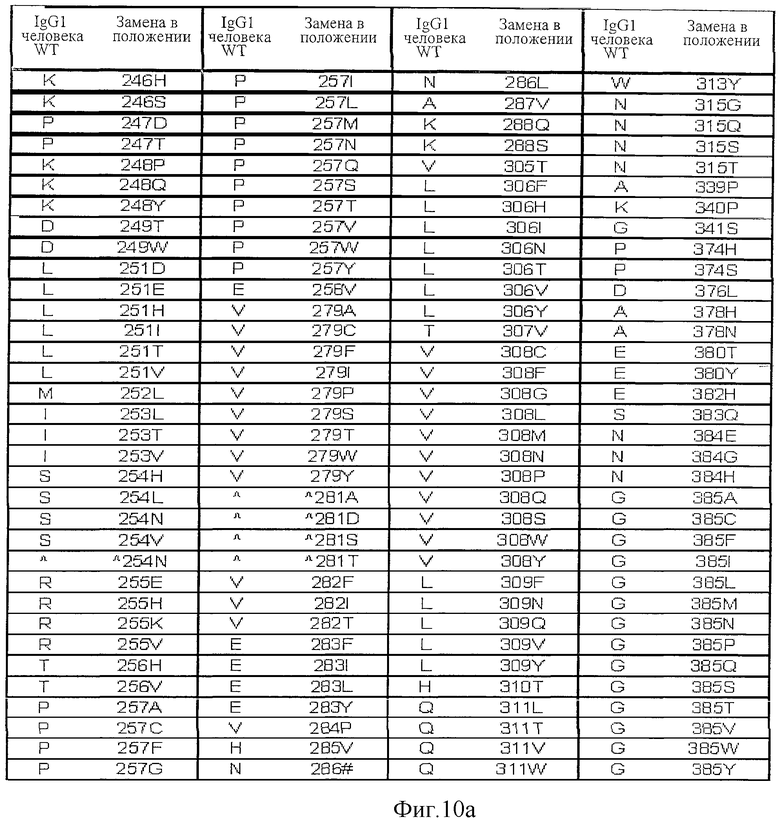
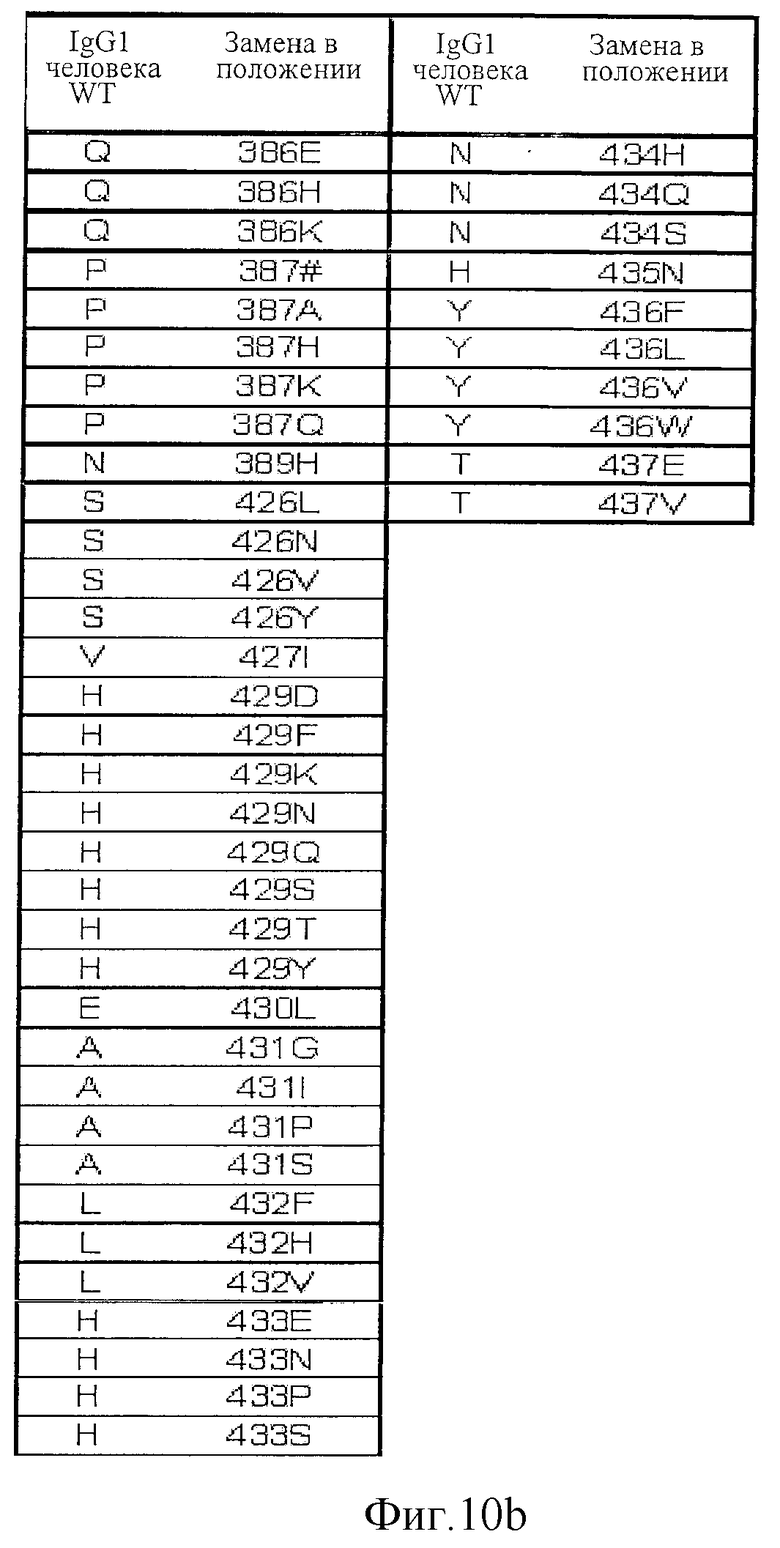
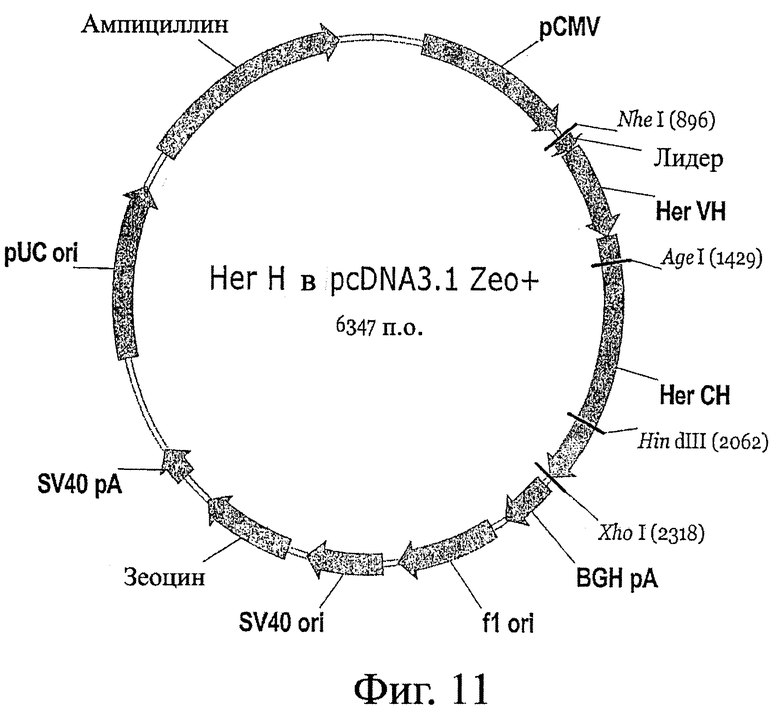
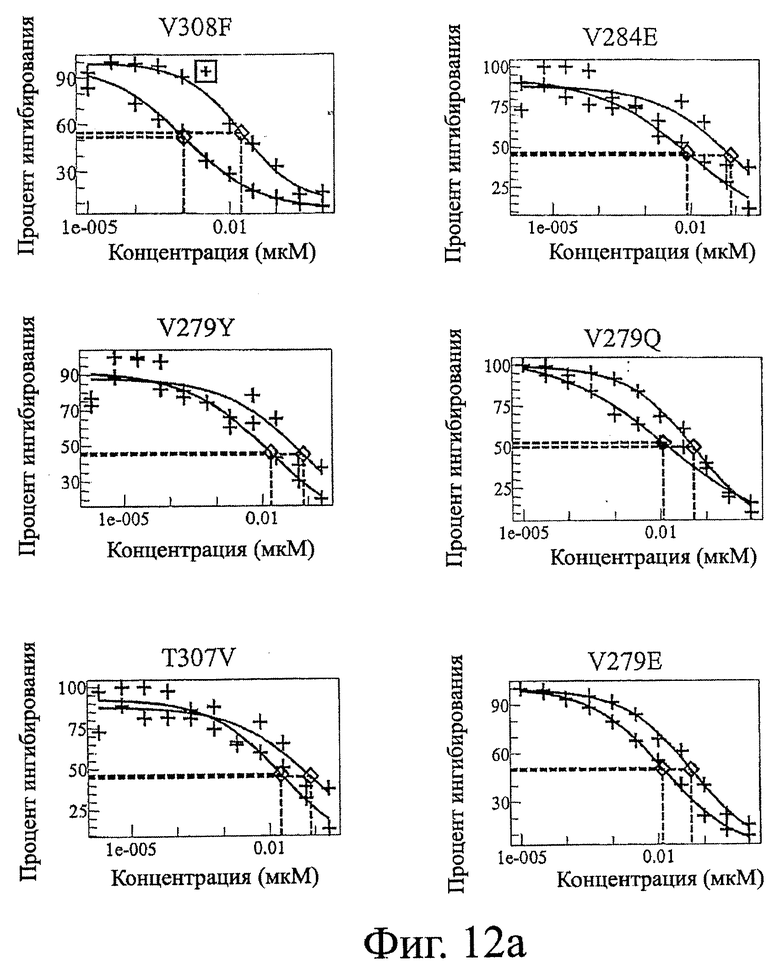
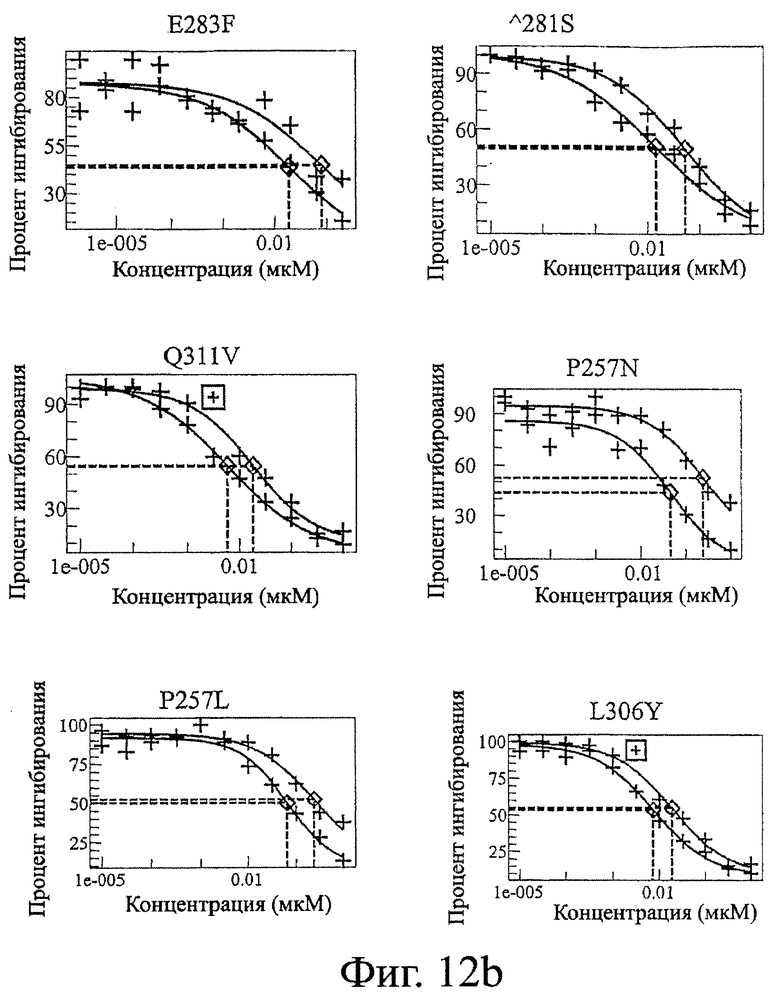
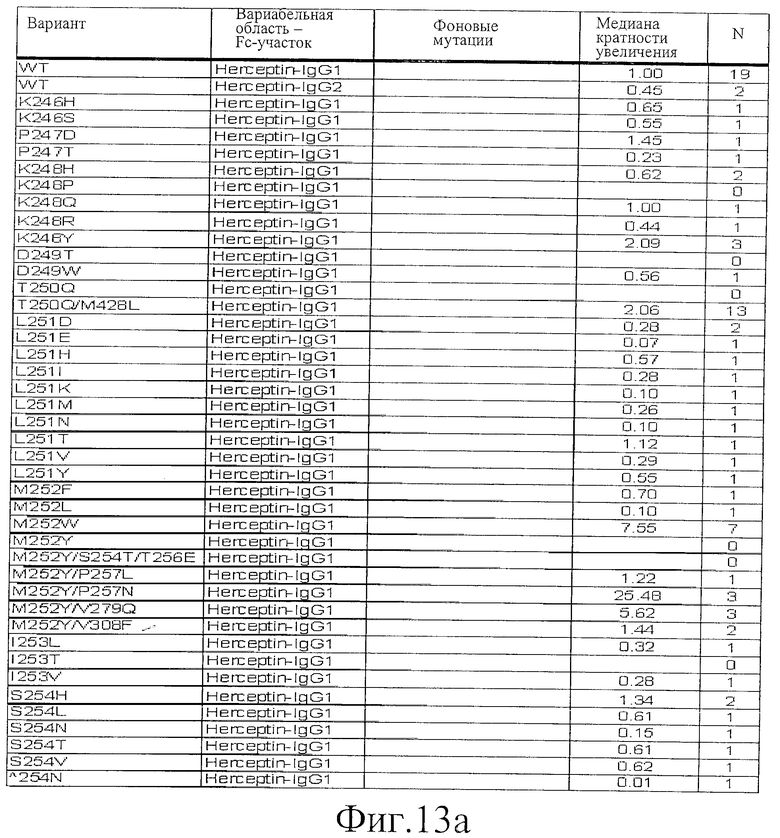
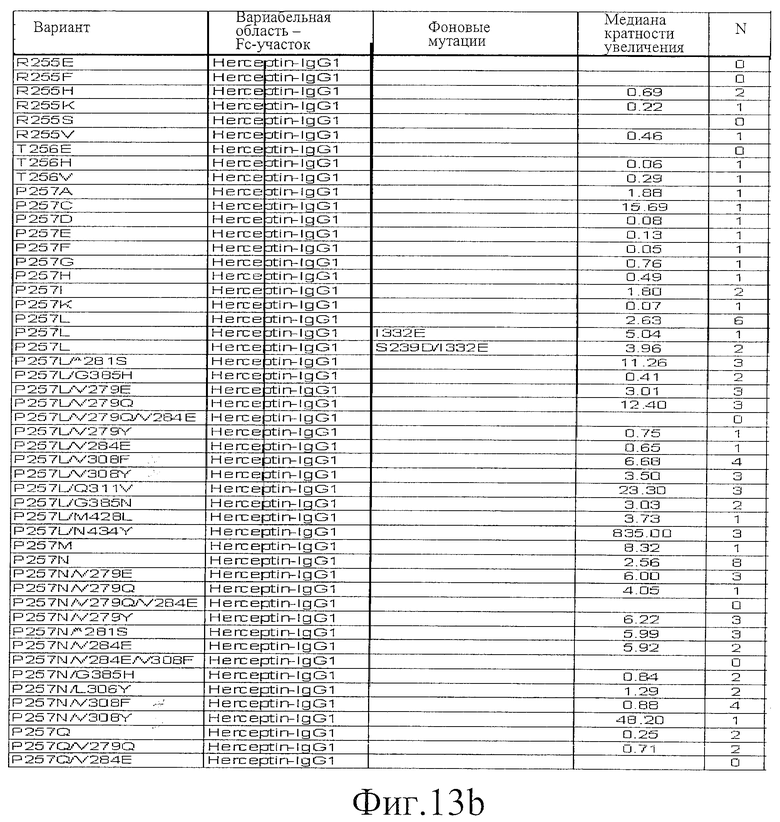
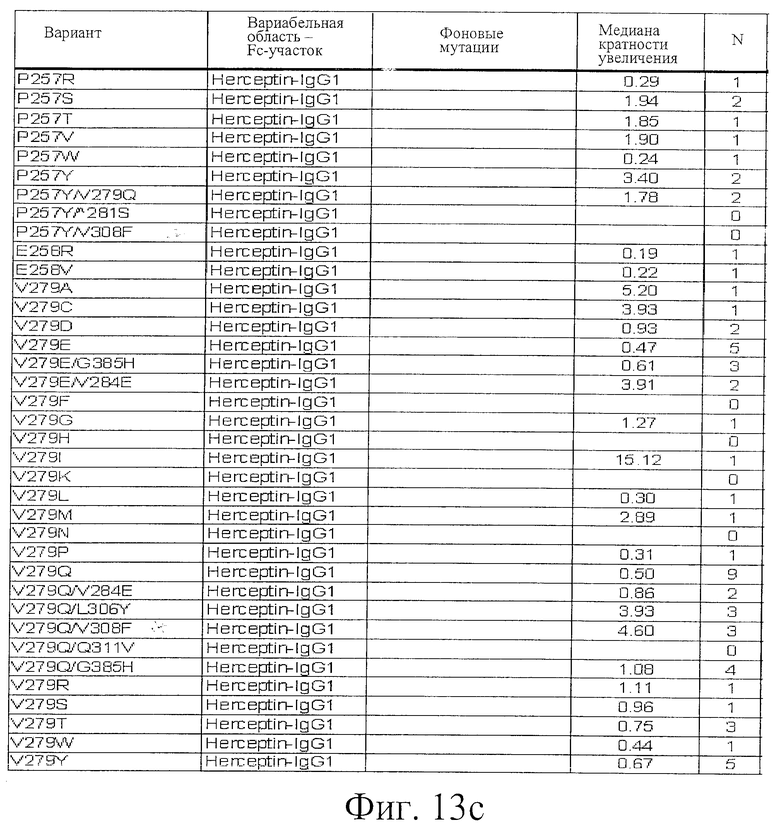
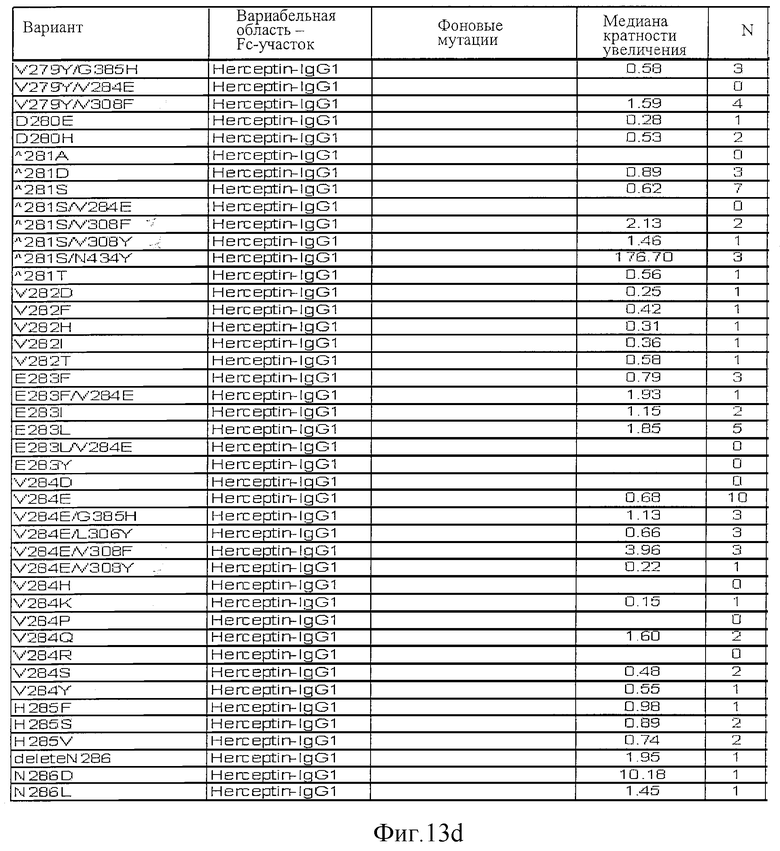
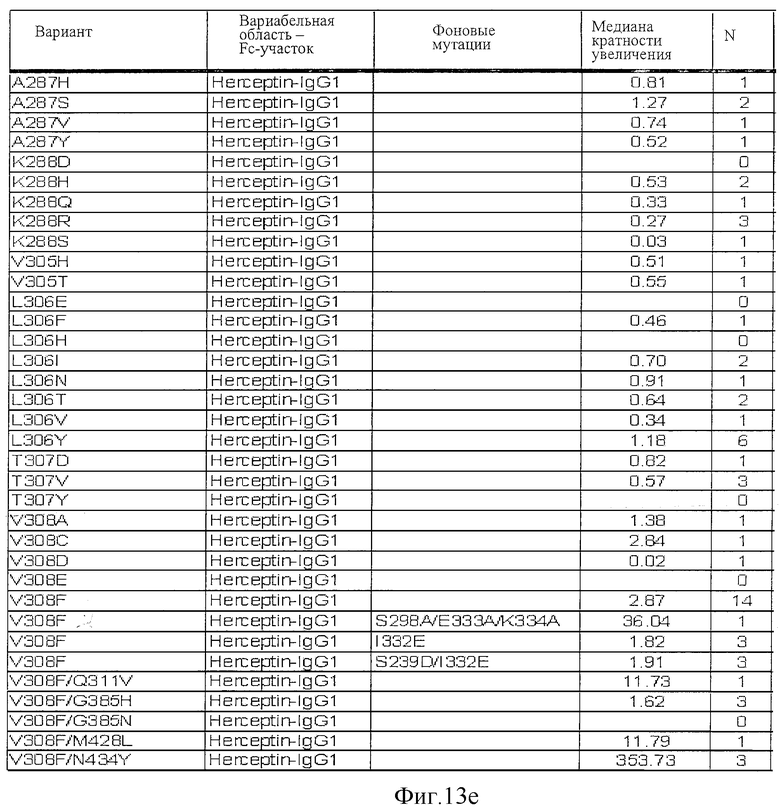
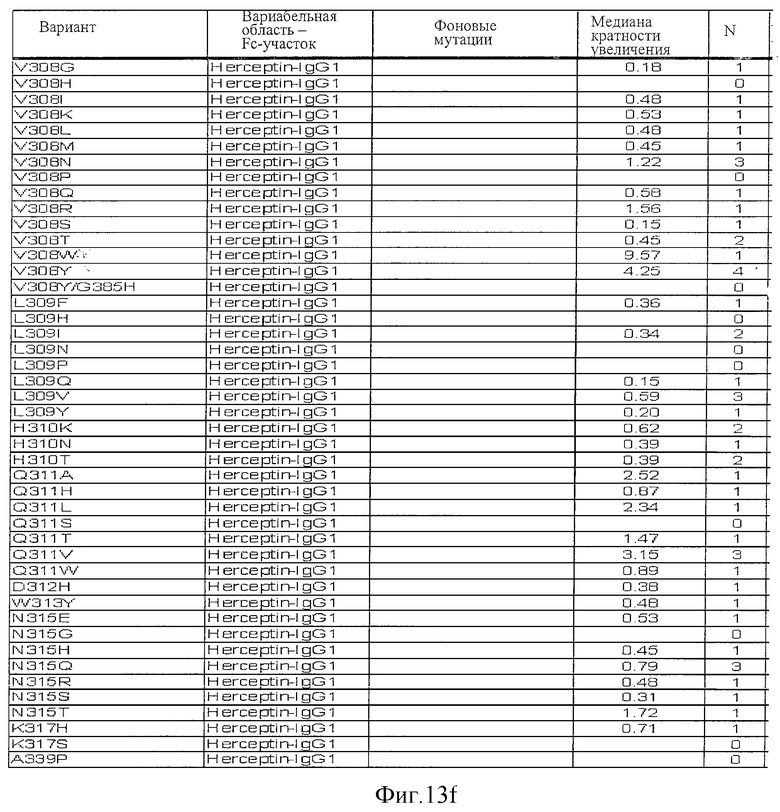
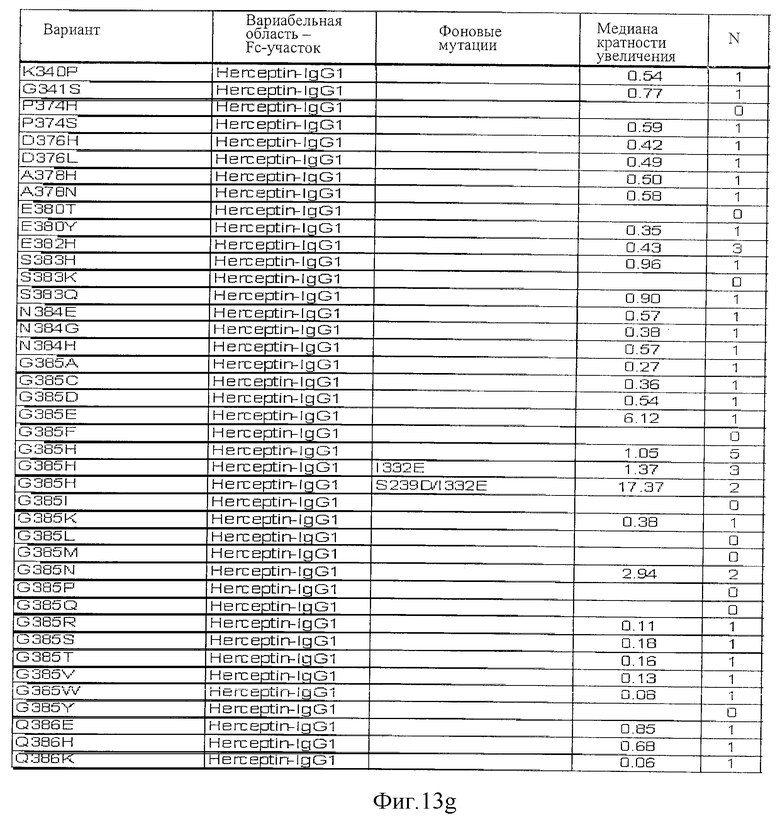
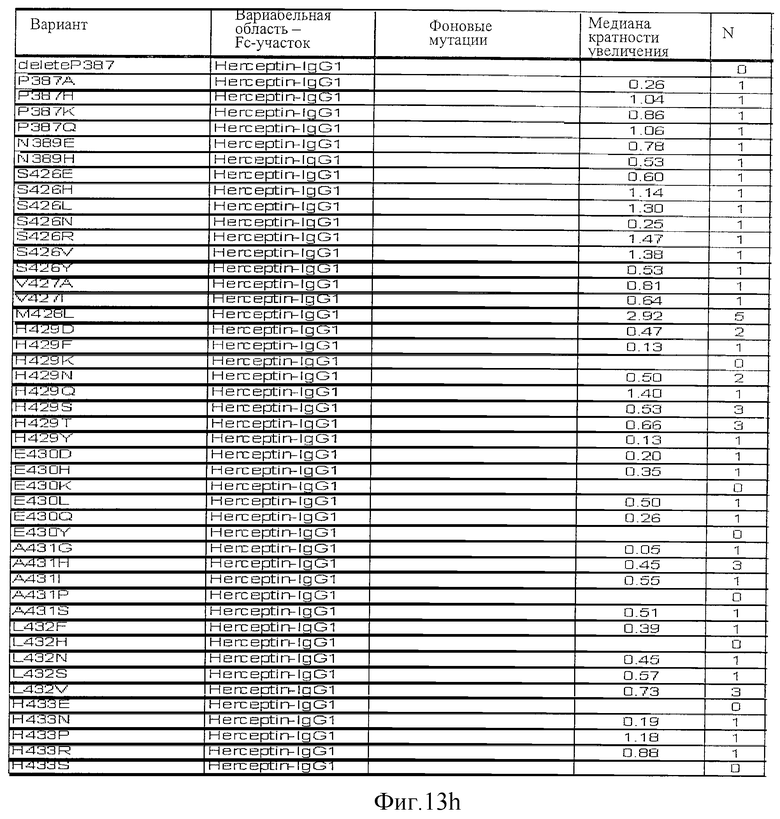
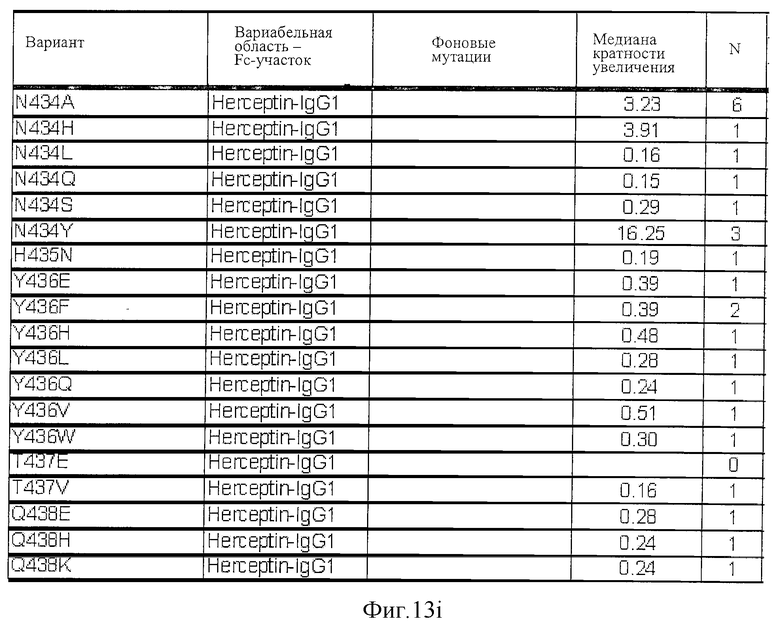
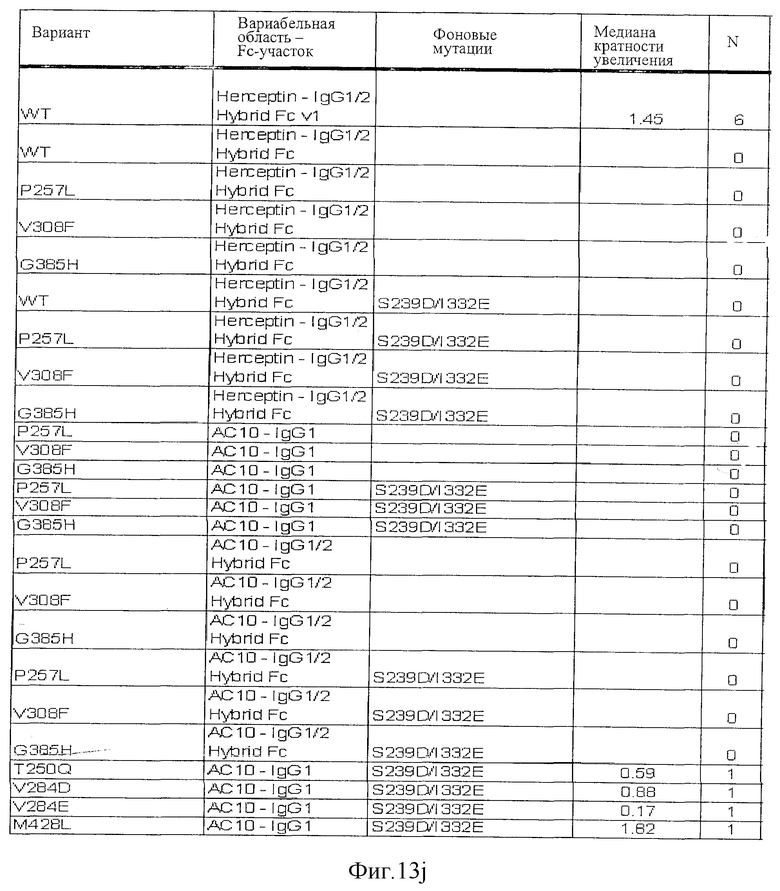
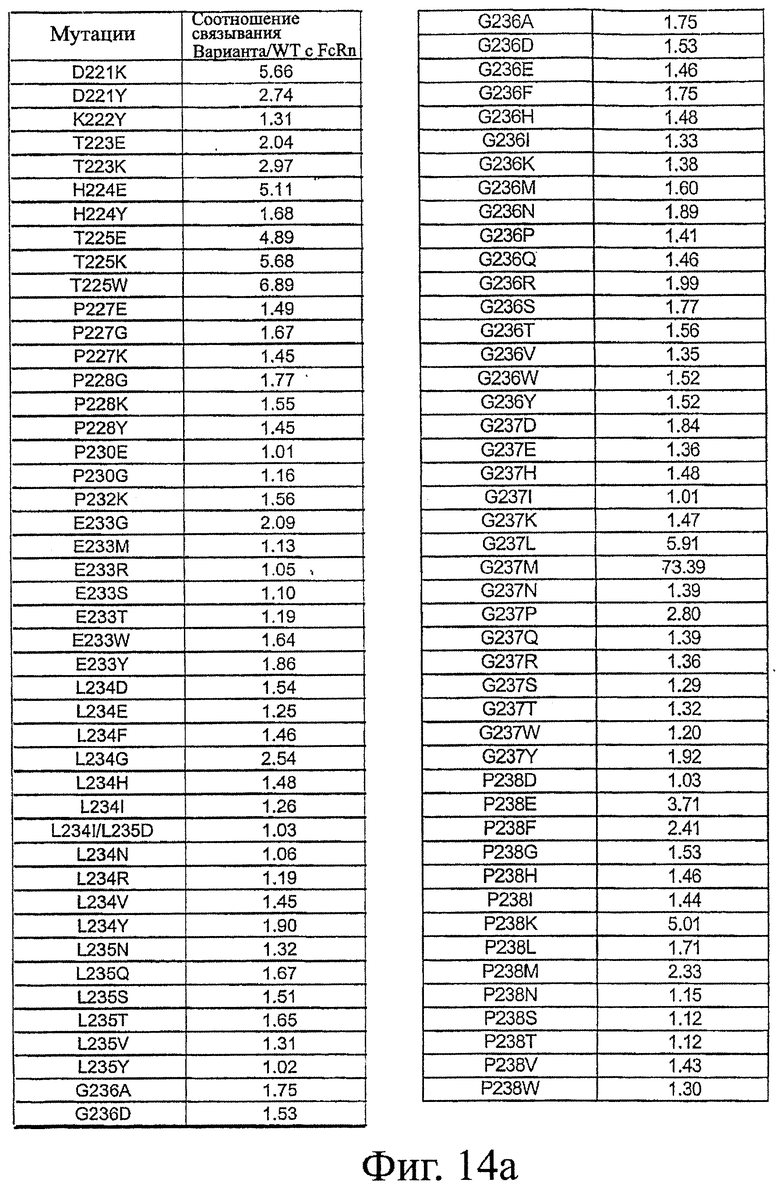
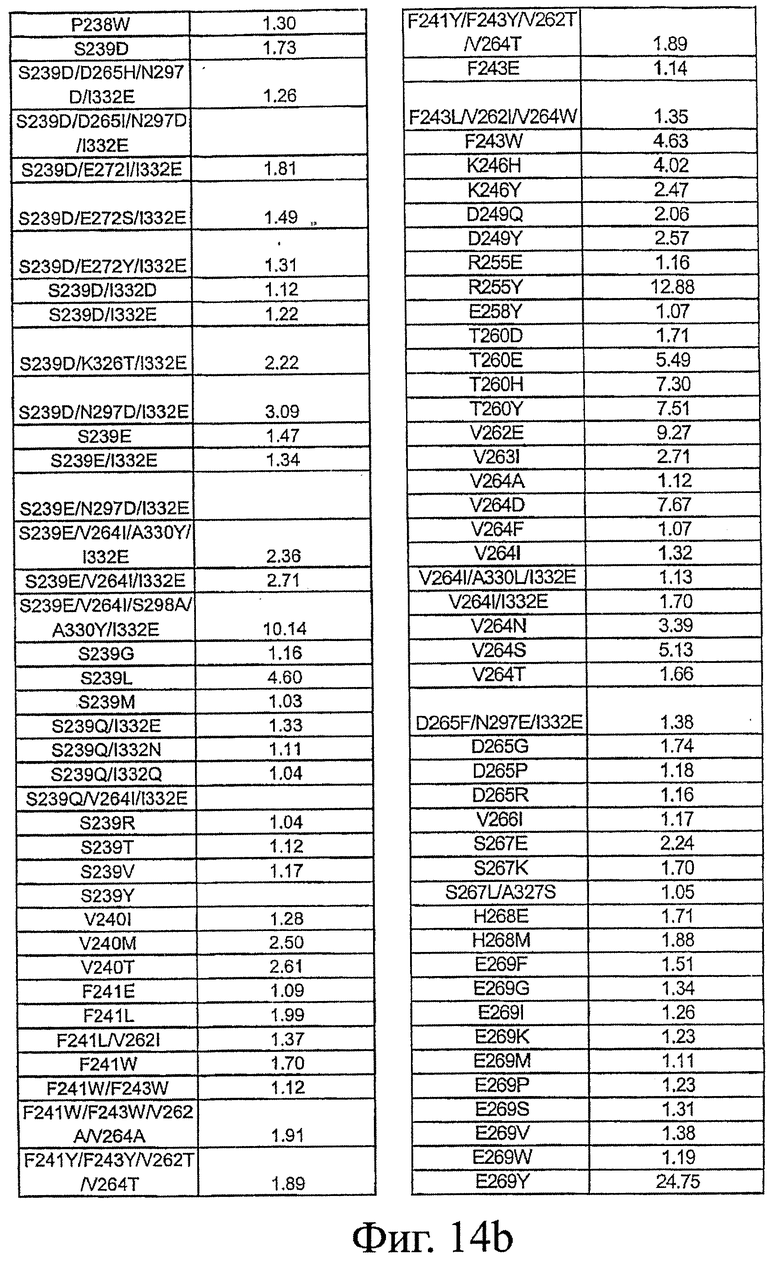
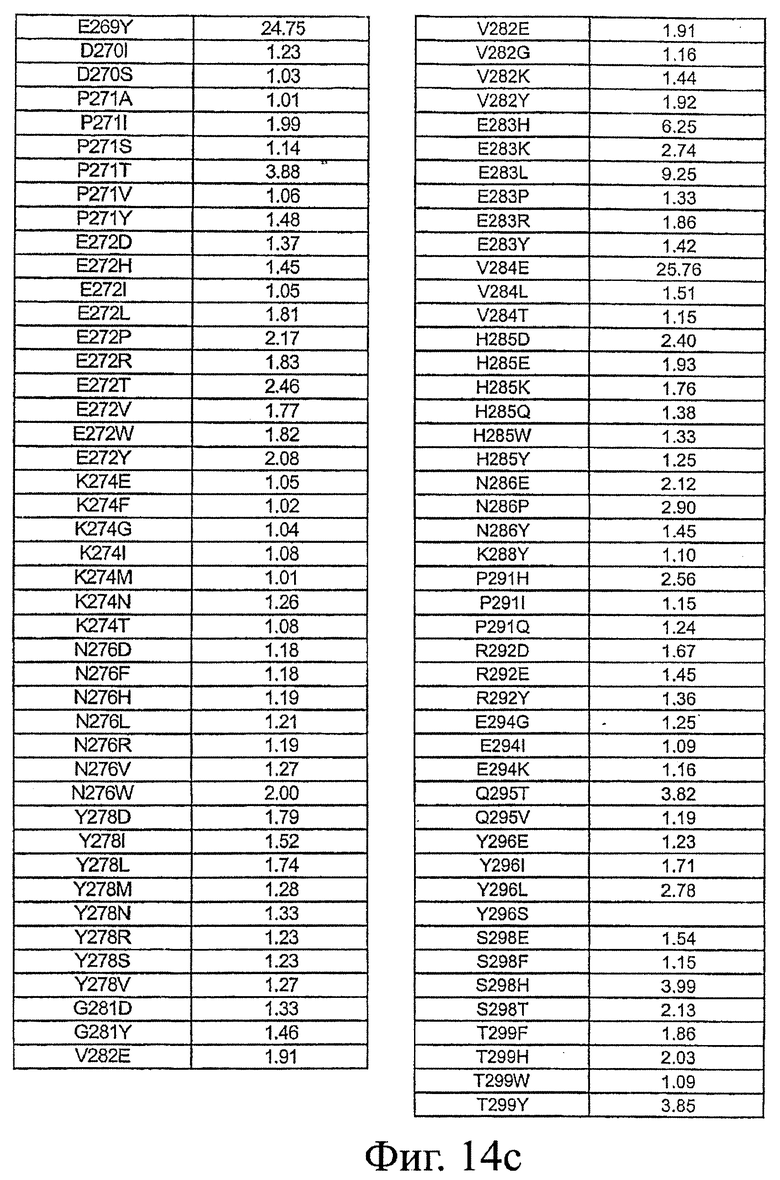
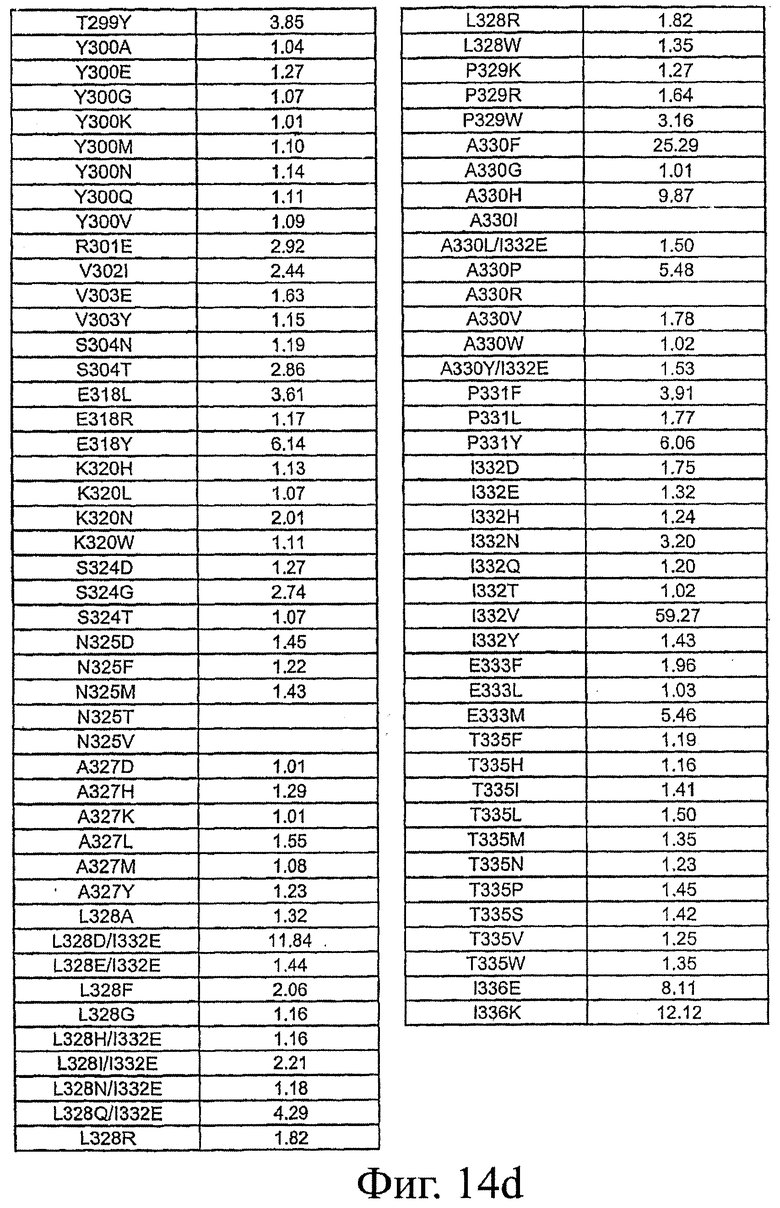
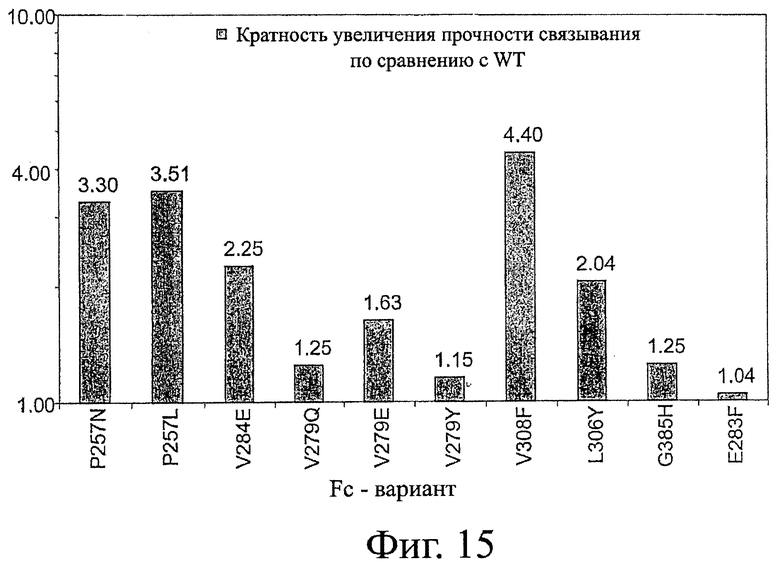
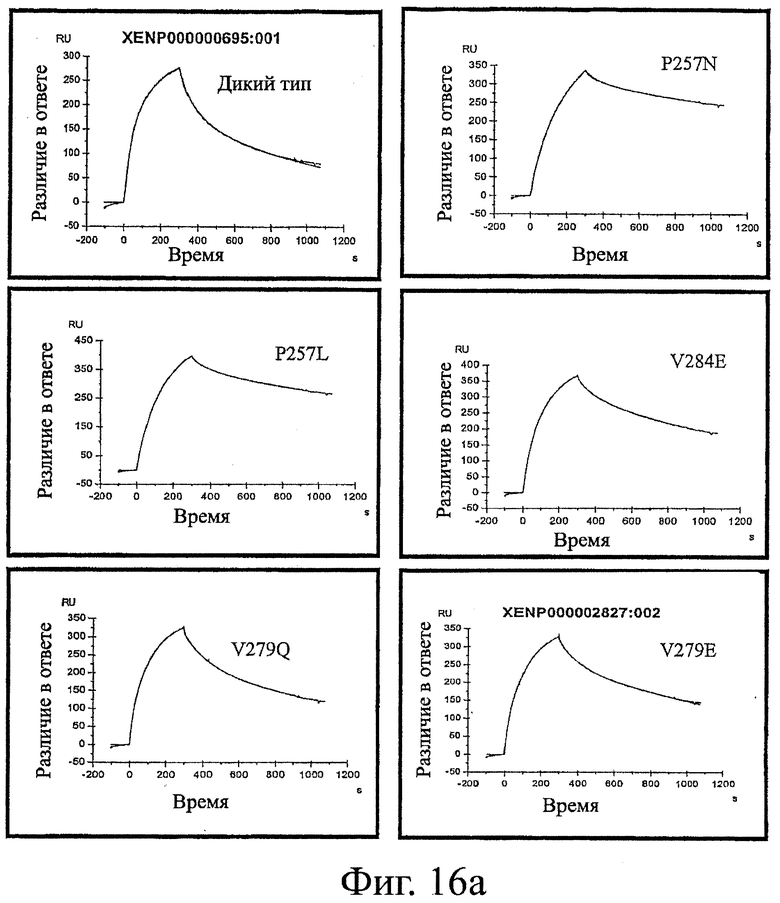
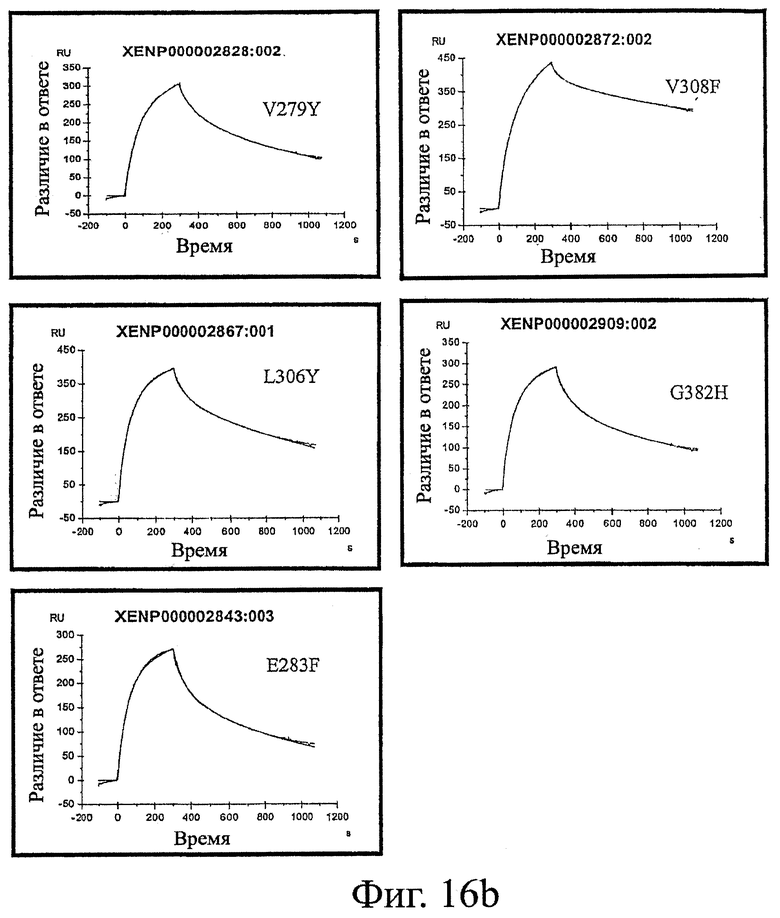
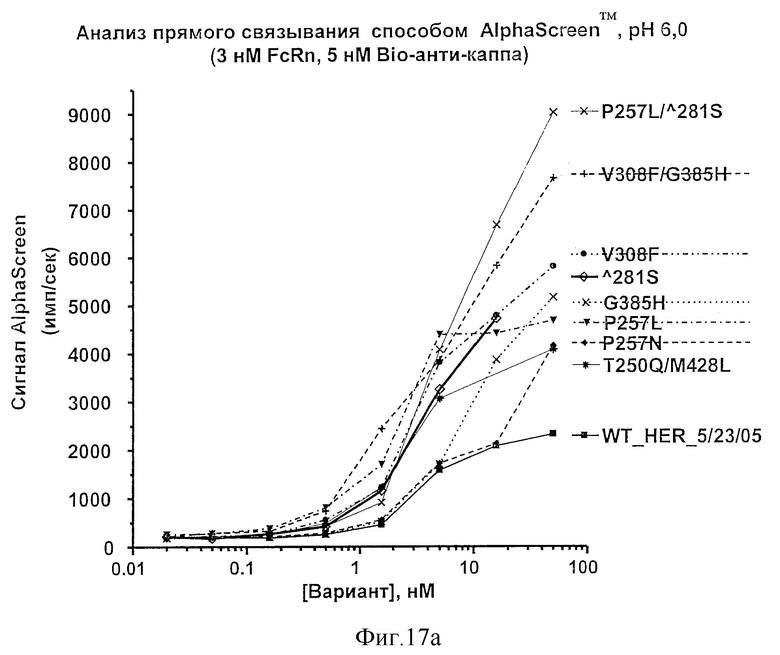

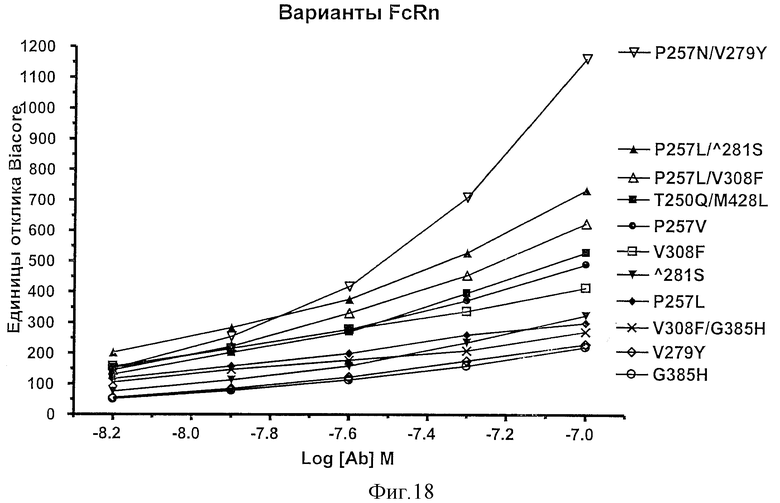
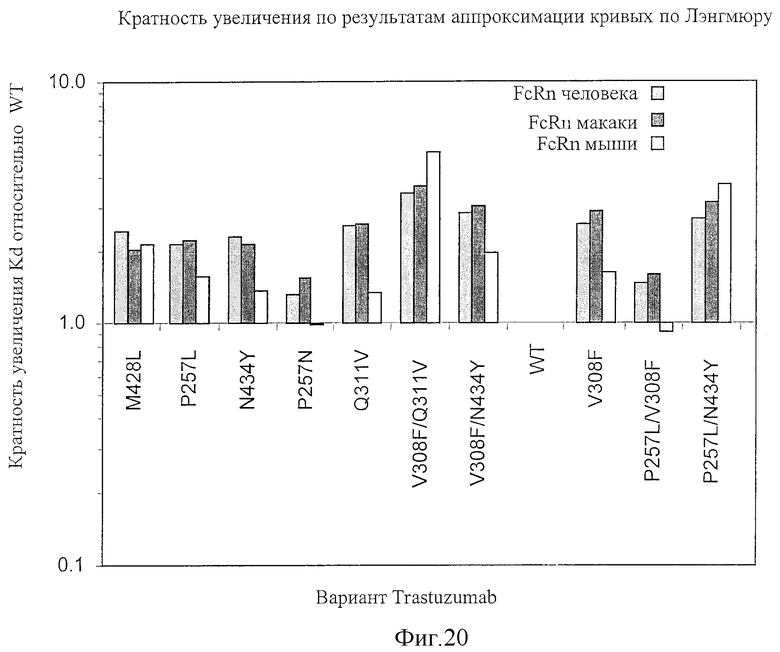
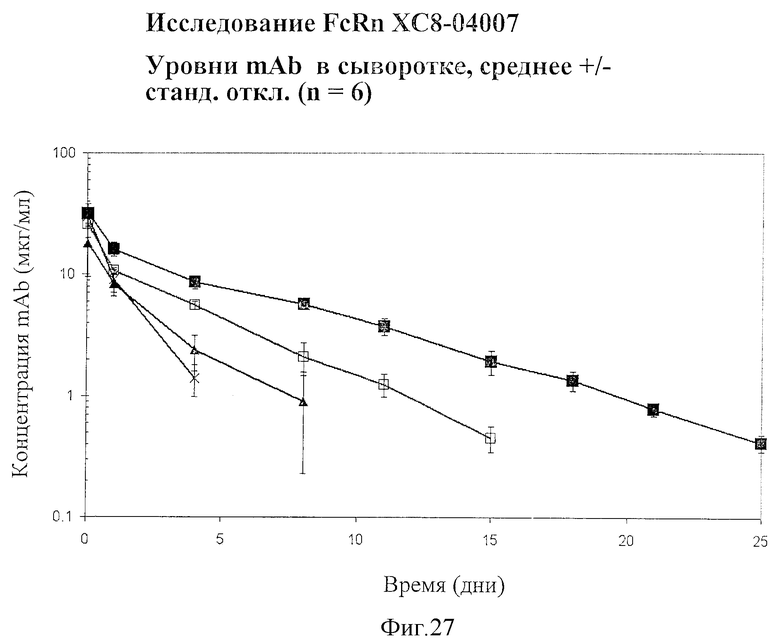
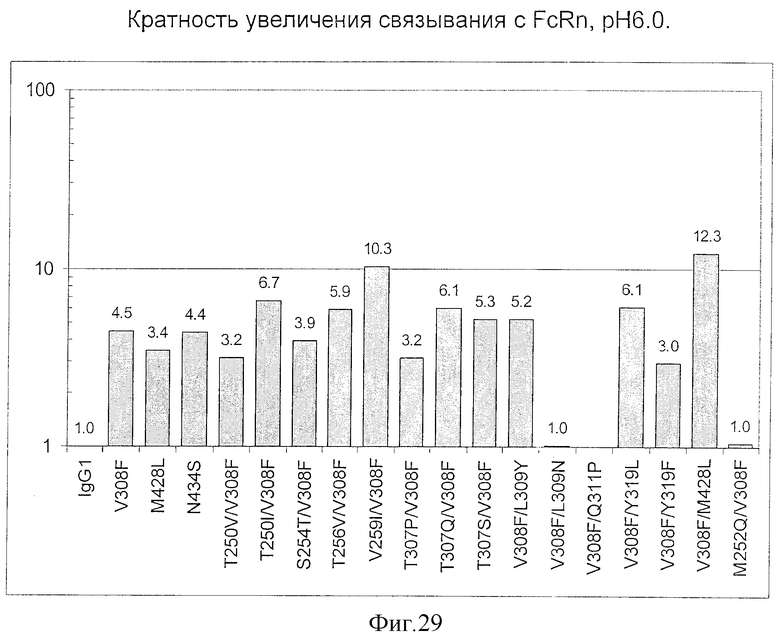
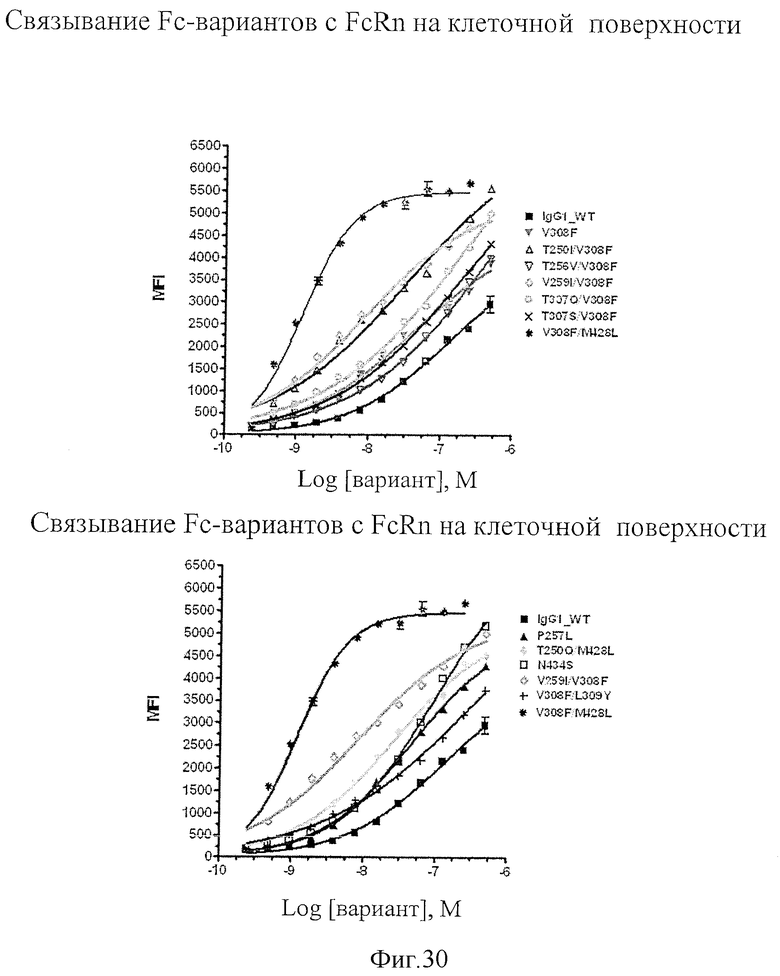


Изобретение относится к иммунологии и биотехнологии. Предложены варианты полипептидов с Fc-участком из IgG, которые обладают увеличенным связыванием FcRn за счет введения мутации 308С, 308F, 308W, 308Y или измененным связыванием при введении мутации 252Y/308F, 257L/308F, 257L/308Y, 257N/308Y, 279Q/308F, 279Y/308F, ^281S/308F, ^281S/308Y, 284E/308F, 298A/308F/333A/334A, 308F/332E, 308F/311V, 308F/G385H, 308F/428L, и 308F/434Y. Описано антитело с указанными мутациями в Fc-области, а также использование таких Fc-областей для получения слитого белка. Использование изобретения обеспечивает новые Fc-варианты с измененным связыванием FcRn, что может найти применение в медицине для получения различных конструкций, с пролонгированным временем удержания в сыворотке крови in vivo или, наоборот, в случае терапии с использованием радиоактивных лекарственных средств, с пониженным временем удержания в сыворотке крови in vivo. 4 н. и 10 з.п. ф-лы, 44 ил.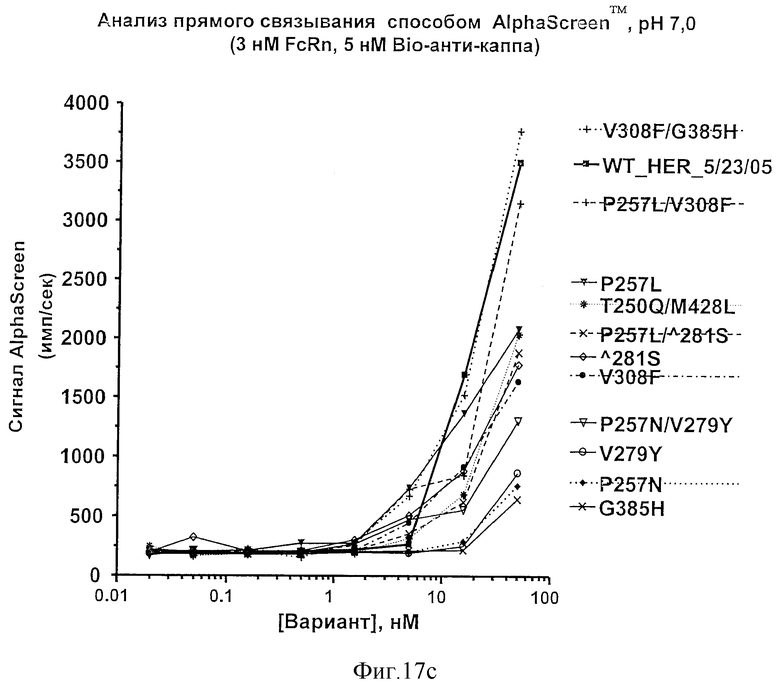
1. Полипептид, демонстрирующий измененное связывание с FcRn, включающий Fc-вариант исходного Fc-полипептида IgG, содержащий модификацию в Fc-участке, выбранную из группы, состоящей из 308С, 308F, 308W, 308Y, где указанный Fc-вариант демонстрирует увеличенное связывание с FcRn по сравнению с исходным Fc-полипептидом, где нумерация соответствует ЕU-индексу.
2. Полипептид по п.1, где указанный Fc-вариант содержит, по меньшей мере, одну замену, выбранную из группы, состоящей из: 308F, 308W и 308Y.
3. Полипептид по п.1 или 2, где указанный Fc-вариант содержит 308F.
4. Полипептид по любому из пп.1-3, где указанный Fc-вариант демонстрирует измененное связывание с FcγR по сравнению с указанным исходным Fc-полипептидом.
5. Полипептид по любому из пп.1-4, где указанный Fc-вариант демонстрирует повышенное связывание с FcγR.
6. Полипептид по любому из пп.1-4, где указанный Fc-вариант демонстрирует пониженное связывание с FcγR.
7. Полипептид по любому из пп.1-4, где указанный Fc-вариант демонстрирует измененное связывание с FcRn по сравнению с исходным Fc-полипептидом, и где указанный полипептид дополнительно характеризуется тем, что полипептид обладает специфичностью по отношению к молекуле-мишени, которую выбирают из группы, состоящей из цитокина, растворимого белкового фактора и белка, экспрессируемого на раковых клетках.
8. Полипептид, демонстрирующий измененное связывание с FcRn, включающий Fc-вариант исходного Fc-полипептида IgG, где указанный Fc-вариант демонстрирует увеличенное связывание с FcRn по сравнению с исходным Fc-полипептидом, где указанный Fc-вариант включает модификации в Fc-участке указанного исходного полипептида, выбранные из группы, состоящей из 252Y/308F, 257L/308F, 257L/308Y, 257N/308Y, 279Q/308F, 279Y/308F, ^281S/308F, ^281S/308Y, 284E/308F, 298A/308F/333A/334A, 308F/332E, 308F/311V, 308F/G385H, 308F/428L и 308F/434Y, где ^ означает инсерцию, где нумерация соответствует EU-индексу.
9. Полипептид по п.8, где указанный Fc-вариант демонстрирует измененное связывание с FcγR по сравнению с указанным исходным Fc-полипептидом.
10. Полипептид по любому из пп.8 и 9, где указанный Fc-вариант демонстрирует повышенное связывание с FcγR.
11. Полипептид по любому из пп.8 и 9, где указанный Fc-вариант демонстрирует пониженное связывание с FcγR.
12. Полипептид по любому из пп.8 и 9, где указанный Fc-вариант демонстрирует измененное связывание с FcRn по сравнению с исходным Fc-полипептидом, и где указанный полипептид дополнительно характеризуется тем, что полипептид обладает специфичностью по отношению к молекуле-мишени, которую выбирают из группы, состоящей из цитокина, растворимого белкового фактора и белка, экспрессируемого на раковых клетках.
13. Антитело, демонстрирующее измененное связывание с FcRn, содержащее Fc-вариант по любому из пп.1-12.
14. Применение Fc-варианта, охарактеризованного в любом из пп.1-13, для получения слитого с Fc белка, демонстрирующего увеличенное связывание с FcRn.
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| DALL'ACQUA et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ПРОИСХОДЯЩИЕ ОТ ИММУНОГЛОБУЛИНОВ, КОТОРЫЕ НЕ СТИМУЛИРУЮТ ЛИЗИС, ОПОСРЕДОВАННЫЙ КОМПЛЕМЕНТОМ | 1999 |
|
RU2226196C2 |

