По настоящей заявке испрашивается приоритет предварительной заявки США №60/581747, поданной 21 июня 2004 г.; содержание которой включено в настоящую заявку в качестве ссылки в полном объеме.
УРОВЕНЬ ТЕХНИКИ
Интерфероны (IFN) типа I (IFN-α, IFN-β, IFN-ω, IFN-τ) представляют собой семейство структурно родственных цитокинов, обладающих противовирусными, противоопухолевыми и иммуномодуляторными эффектами (Hardy et al. (2001) Blood, 97: 473; Cutrone and Langer (2001) J.Biol.Chem. 276: 17140). Локус IFNα человека включает два подсемейства. Первое подсемейство состоит из 14 неаллельных генов и 4 псевдогенов, имеющих гомологию не менее 80%. Второе подсемейство αII или омега (ω), содержит 5 псевдогенов и 1 функциональный ген, который на 70% гомологичен генам IFNα (Weissman and Weber (1986) Prog.Nucl.Acid Res. Mol. Biol., 33: 251-300). Подтипы IFNα обладают различными видами специфической активности, но имеют одинаковый биологический спектр (Streuli et al. (1981) Proc.Natl.Acad.Sci USA, 78:2848) и один и тот же клеточный рецептор (Agnet M. et. al. in “Interferon 5” Ed.I. Gresser p. 1-22, Academic Press, London 1983).
Интерферон β (IFNβ) закодирован одним геном, который имеет примерно 50% гомологию с генами IFNα.
Гамма-интерферон, который вырабатывается активированными лимфоцитами, не обладает какой-либо гомологией с интерферонами альфа/бета и не взаимодействует с их рецептором.
Все интерфероны человека типа I связываются с рецептором на клеточной поверхности (IFN альфа рецептором, IFNAR), состоящим из двух трансмембранных белков, т.е. IFNAR-1 и IFNAR-2 (Uze et. al. (1990) Cell, 60:225; Novick et al. (1994) Cell, 77:391). IFNAR-1 важен для высокоаффинного связывания и дифференциальной специфичности комплекса IFNAR (Cutrone, 2001, см.выше). Хотя функциональные различия каждого из подтипов IFN типа I не были установлены, предполагают, что каждый из них может осуществлять различные взаимодействия с компонентами рецептора IFNAR, ведущие к появлению потенциально различных сигналов (Cook et al. (1996) J.Biol.Chem. 271: 13448). В частности, исследования с использованием мутантных форм IFNAR-1 и IFNAR-2 позволили предположить, что сигналы альфа- и бета-интерферонов по-разному передаются рецептором за счет различного взаимодействия с соответствующими цепями (Lewerenz et al. (1998) J.Mol.Biol. 282: 585).
Первоначально функциональные исследования интерферонов типа I были сконцентрированы на природной защите против вирусных инфекций (Haller et al. (1981) J.Exp.Med., 154:199; Lindenmann et al. (1981) Methods Enzymol. 78: 181). Однако в более поздних исследованиях интерфероны типа I рассматривают как мощные иммунорегуляторные цитокины при адаптивном иммунном ответе. Конкретно, было показано, что интерфероны типа I облегчают дифференцировку «необученных» T-клеток в направлении пути Th1 (Brinkmann et al. (1993) J.Exp.Med, 178: 1655), усиливают выработку антител (Finkelman et al. (1991) J.Exp.Med. 174:1179) и поддерживают функциональную активность и сохранение памяти T-клеток (Santini et al. (2000) J.Exp.Med. 191:1777; Tough et al. (1996) Science, 272:1947).
В современных работах многочисленных исследовательских групп предполагается, что IFN-α может улучшать созревание или активацию дендритных клеток (DC) (Santini, et al. (2000) J.Exp.Med. 191:1777; Luft et al. (1998) J.Immunol. 161:1947; Luft et al. (2002) Int.Immunol. 14:367; Radvanyi et al. (1999) Scand.J.Immunol. 50:499). Кроме того, увеличение экспрессии интерферонов типа I было описано при многочисленных аутоиммунных заболеваниях (Foulis et al. (1987) Lancet 2: 1423; Hooks et al. (1982) Arthritis Rheum. 25:396; Hertzog et al. (1988) Clin. Immunol.Immunopathol. 48:192; Hopkins and Meager (1988) Clin.Exp.Immunol. 73:88; Arvin and Miller (1984) Arthritis Rheum. 27:582). Большинство изученных примеров этих заболеваний представляют собой инсулин-зависимый сахарный диабет (IDDM) (Foulis (1987), см. выше) и системную красную волчанку (SLE) (Hooks (1982), см. выше), которые связаны с повышенными уровнями IFN-α, а также ревматоидный артрит (RA) (Hertzog (1988), Hopkins and Meager (1988), Arvin and Miller (1984), см. выше), при котором более важную роль может играть IFN-β.
Кроме того, сообщалось, что введение интерферона α обостряет основное заболевание у пациентов с псориазом и рассеянным склерозом и вызывает SLE-подобный синдром у пациентов, у которых ранее отсутствовали аутоиммунные заболевания. Также было показано, что интерферон α вызывает гломерулонефрит у нормальных мышей и ускоряет начало спонтанного аутоиммунного заболевания мышей NZB/W. Кроме того, было показано, что в некоторых случаях IFN-α терапия ведет к нежелательным побочным эффектам, в том числе к повышению температуры и неврологическим расстройствам. Следовательно, имеются патологические состояния, при которых ингибирование активности интерферонов типа I может оказать благоприятный эффект у пациента, и существует потребность в средствах, эффективных для ингибирования активности интерферонов типа I.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделенным моноклональным антителам человека, которые связываются с IFNAR-1 и ингибируют биологическую активность интерферона типа I, предпочтительно нескольких интерферонов типа I. Кроме того, указанные антитела не связываются с тем же эпитопом, что и анти-IFNAR-1 антитело мыши, т.е. 64G12.
В одном из аспектов настоящее изобретение относится к выделенному антителу человека или его антиген-связывающему участку, причем антитело специфично связывается с IFNAR-1 и проявляет одно или несколько из следующих свойств:
a) связывается с IFNAR-1 с KD, равной 1·10-7 М, или с более высокой аффинностью;
b) ингибирует биологическую активность нескольких интерферонов типа I;
c) ингибирует активность IFN-α 2b в анализе пролиферации клеток Daudi;
d) ингибирует активность IFN омега в анализе пролиферации клеток Daudi;
e) ингибирует секрецию IP-10 моноядерными клетками периферической крови, вызванную IFN-α 2b;
f) ингибирует секрецию IP-10 моноядерными клетками периферической крови, вызванную IFN омега;
g) ингибирует развитие дендритных клеток, опосредованное плазмой системной красной волчанки; и
h) связывается с другим эпитопом, чем моноклональное антитело 64G12 мыши (ECACC, депозитарный № 92022605).
Предпочтительные антитела по настоящему изобретению специфично связываются c рецептором 1 интерферона альфа человека, причем связывание осуществляется с KD 1·10-8 М или большей аффинностью, или с KD 1·10-9 М или большей аффинностью, или с KD 5·10-10 М или большей аффинностью, или с KD 2·10-10 М или большей аффинностью.
В одном из аспектов настоящее изобретение относится к выделенному моноклональному антителу или его антиген-связывающему участку, включающим вариабельную область тяжелой цепи, которая является продуктом или производным генов VH 4-34 или 5-51 человека, причем указанное антитело специфично связывается с рецептором 1 интерферона альфа человека. В другом аспекте изобретение относится к выделенному моноклональному антителу или его антиген-связывающей части, включающим вариабельную область легкой цепи, которая является продуктом или производным генов VK L18 или A27 человека, причем указанное антитело специфично связывается с рецептором 1 интерферона альфа человека. В еще одном аспекте изобретение относится к выделенному моноклональному антителу человека или его антиген-связывающему участку, включающим:
(a) вариабельную область тяжелой цепи, которая является продуктом или производным генов VH 4-34 или 5-51 человека; и
(b) вариабельную область легкой цепи, которая является продуктом или производным генов VK L18 или A27 человека;
где антитело специфично связывается с рецептором 1 интерферона альфа человека.
В предпочтительных вариантах осуществления антитело включает вариабельную область тяжелой цепи гена VH 4-34 человека и вариабельную область легкой цепи гена VK L18 человека или же антитело включает вариабельную область тяжелой цепи гена VH 5-51 человека и вариабельную область легкой цепи гена VK A27 человека.
В другом аспекте изобретение относится к выделенному моноклональному антителу человека или его антиген-связывающему участку, включающим:
вариабельную область тяжелой цепи человека, включающую последовательности CDR1, CDR2 и CDR3; и вариабельную область легкой цепи человека, включающую последовательности CDR1, CDR2 и CDR3, причем:
(a) последовательность CDR3 вариабельной области тяжелой цепи человека включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 9, 10, 11 и 12 и их консервативных модификаций;
(b) последовательность CDR3 вариабельной области легкой цепи человека включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 21, 22, 23 и 24 и их консервативных модификаций;
(c) антитело специфично связывается с рецептором 1 интерферона альфа человека со сродством к связыванию не менее 1·10-8 М или большим; и
(d) антитело ингибирует биологическую активность хотя бы одного интерферона типа I.
Предпочтительно последовательность CDR2 вариабельной области тяжелой цепи человека включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 5, 6, 7 и 8 и их консервативных модификаций; и последовательность CDR2 вариабельной области легкой цепи человека включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 17, 18, 19 и 20 и их консервативных модификаций. Предпочтительно последовательность CDR1 вариабельной области тяжелой цепи человека включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 1, 2, 3 и 4 и их консервативных модификаций; и последовательность CDR1 вариабельной области легкой цепи человека включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 13, 14, 15 и 16 и их консервативных модификаций.
В другом аспекте изобретение относится к выделенному моноклональному антителу человека или его антиген-связывающему участку, включающим вариабельную область тяжелой цепи человека и вариабельную область легкой цепи человека, причем:
(a) вариабельная область тяжелой цепи человека включает аминокислотную последовательность, которая не менее чем на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 26, 27 и 28;
(b) вариабельная область легкой цепи человека включает аминокислотную последовательность, которая не менее чем на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 30, 31 и 32;
(c) антитело специфично связывается с рецептором 1 интерферона альфа человека со сродством к связыванию не менее 1·10-8 М или большим; и
(d) антитело ингибирует биологическую активность хотя бы одного интерферона типа I.
Предпочтительные антитела по настоящему изобретению включают выделенные моноклональные антитела человека или их антиген-связывающие участки, включающие:
(a) вариабельную область тяжелой цепи CDR1 человека, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, 2, 3 и 4;
(b) вариабельную область тяжелой цепи CDR2 человека, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, 6, 7 и 8;
(c) вариабельную область тяжелой цепи CDR3 человека, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, 10, 11 и 12;
(d) вариабельную область легкой цепи CDR1 человека, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13, 14, 15 и 16;
(e) вариабельную область легкой цепи CDR2 человека, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 17, 18, 19 и 20; и
(f) вариабельную область легкой цепи CDR3 человека, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, 22, 23 и 24;
причем антитело специфично связывается с рецептором 1 интерферона альфа человека со сродством к связыванию не менее 1·10-8 М или большим.
Предпочтительные комбинации областей CDR включают следующие:
(a) вариабельную область тяжелой цепи CDR1 человека, включающую SEQ ID NO:1;
(b) вариабельную область тяжелой цепи CDR2 человека, включающую SEQ ID NO:5;
(c) вариабельную область тяжелой цепи CDR3 человека, включающую SEQ ID NO:9;
(d) вариабельную область легкой цепи CDR1 человека, включающую SEQ ID NO:13;
(e) вариабельную область легкой цепи CDR2 человека, включающую SEQ ID NO:17; и
(f) вариабельную область легкой цепи CDR3 человека, включающую SEQ ID NO:21.
(a) вариабельную область тяжелой цепи CDR1 человека, включающую SEQ ID NO:2;
(b) вариабельную область тяжелой цепи CDR2 человека, включающую SEQ ID NO:6;
(c) вариабельную область тяжелой цепи CDR3 человека, включающую SEQ ID NO:10;
(d) вариабельную область легкой цепи CDR1 человека, включающую SEQ ID NO:14;
(e) вариабельную область легкой цепи CDR2 человека, включающую SEQ ID NO:18; и
(f) вариабельную область легкой цепи CDR3 человека, включающую SEQ ID NO:22.
(a) вариабельную область тяжелой цепи CDR1 человека, включающую SEQ ID NO:3;
(b) вариабельную область тяжелой цепи CDR2 человека, включающую SEQ ID NO:7;
(c) вариабельную область тяжелой цепи CDR3 человека, включающую SEQ ID NO:11;
(d) вариабельную область легкой цепи CDR1 человека, включающую SEQ ID NO:15;
(e) вариабельную область легкой цепи CDR2 человека, включающую SEQ ID NO:19; и
(f) вариабельную область легкой цепи CDR3 человека, включающую SEQ ID NO:23.
(a) вариабельную область тяжелой цепи CDR1 человека, включающую SEQ ID NO:4;
(b) вариабельную область тяжелой цепи CDR2 человека, включающую SEQ ID NO:8;
(c) вариабельную область тяжелой цепи CDR3 человека, включающую SEQ ID NO:12;
(d) вариабельную область легкой цепи CDR1 человека, включающую SEQ ID NO:16;
(e) вариабельную область легкой цепи CDR2 человека, включающую SEQ ID NO:20; и
(f) вариабельную область легкой цепи CDR3 человека, включающую SEQ ID NO:24.
Другие предпочтительные антитела по настоящему изобретению включают выделенные моноклональные антитела человека или их антиген-связывающие участки, включающие:
(a) вариабельную область тяжелой цепи человека, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 25, 26, 27 и 28; и
(b) вариабельную область легкой цепи человека, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 29, 30, 31 и 32;
причем антитело специфично связывается с рецептором 1 интерферона альфа человека со сродством к связыванию не менее 1·10-8 М или большим.
Предпочтительные комбинации тяжелой и легкой цепей включают следующие:
(a) вариабельную область тяжелой цепи человека, включающую аминокислотную последовательность SEQ ID NO:25; и
(b) вариабельную область легкой цепи человека, включающую аминокислотную последовательность SEQ ID NO:29.
(a) вариабельную область тяжелой цепи человека, включающую аминокислотную последовательность SEQ ID NO:26; и
(b) вариабельную область легкой цепи человека, включающую аминокислотную последовательность SEQ ID NO:30.
(a) вариабельную область тяжелой цепи человека, включающую аминокислотную последовательность SEQ ID NO:27; и
(b) вариабельную область легкой цепи человека, включающую аминокислотную последовательность SEQ ID NO:31.
(a) вариабельную область тяжелой цепи человека, включающую аминокислотную последовательность SEQ ID NO:28; и
(b) вариабельную область легкой цепи человека, включающую аминокислотную последовательность SEQ ID NO:32.
Другой аспект изобретения относится к антителам, которые конкурируют за связывание с IFNAR-1 с контрольными антителами, разработанными в настоящем изобретении. Соответственно в другом варианте осуществления изобретение относится к:
выделенному моноклональному антителу или его антиген-связывающему участку, причем антитело принимает участие в перекрестной конкуренции с контрольным антителом за связывание с рецептором 1 интерферона альфа человека, где контрольное антитело выбрано из группы, состоящей из:
(a) антитела, включающего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:25; и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:29;
(b) антитела, включающего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:26; и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:30;
(c) антитела, включающего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 27; и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:31; и
(d) антитела, включающего вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:28; и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:32.
В некоторых вариантах осуществления изобретение относится к антителу человека или его антиген-связывающему участку, причем антитело не связывается с тем же эпитопом, что и моноклональное антитело 64G12 мыши (ECACC, депозитарный № 92022605) (т.е. не участвует в перекрестной конкуренции с ним).
Антитела по настоящему изобретению могут принадлежать к любому изотипу. Предпочтительные антитела относятся к изотипам IgG1, IgG3 или IgG4. Антитела по настоящему изобретению могут быть полноразмерными антителами, включающими вариабельные и константные области, или же они могут являться их антиген-связывающими фрагментами, как например антителами с одной цепью или фрагментами Fab или Fab'2.
Кроме этого изобретение относится к иммуноконъюгату, включающему антитело по настоящему изобретению или его антиген-связывающий участок, связанные с терапевтическим средством, таким как цитотоксин или радиоактивный изотоп. Изобретение также относится к биспецифичной молекуле, включающей антитело или его антиген-связывающий участок по настоящему изобретению, связанные со вторым функциональным фрагментом, имеющим специфичность связывания, отличную от указанного антитела или его антиген-связывающего участка.
Композиции, содержащие антитело или его антиген-связывающий участок, или иммуноконъюгат, или биспецифичную молекулу по настоящему изобретению и, кроме того, фармацевтически приемлемый носитель, также относятся к настоящему изобретению.
Молекулы нуклеиновых кислот, кодирующие антитела или их антиген-связывающие участки по настоящему изобретению, также входят в объем изобретения, также как и векторы экспрессии, включающие такие нуклеиновые кислоты, и клетки-хозяева, содержащие такие векторы экспрессии. Кроме того, изобретение относится к трансгенным мышам, имеющим трансгены тяжелой и легкой цепей иммуноглобулина человека, причем мыши экспрессируют антитела по настоящему изобретению, а также к гибридомам, полученным из таких мышей, причем гибридомы вырабатывают антитела по настоящему изобретению.
Помимо этого изобретение относится к способам получения «второго поколения» антител анти-IFNAR-1, основанным на последовательностях антител анти-IFNAR-1, разработанных в настоящем изобретении. Например, изобретение относится к способу получения антитела анти-IFNAR-1, включающему:
(a) получение: (i) последовательности вариабельной области тяжелой цепи антитела, включающей последовательность CDR1, которая выбрана из группы, состоящей из SEQ ID NO: 1, 2, 3 и 4, последовательность CDR2, которая выбрана из группы, состоящей из SEQ ID NO: 5, 6, 7 и 8; и последовательность CDR3, которая выбрана из группы, состоящей из SEQ ID NO: 9, 10, 11 и 12; или (ii) последовательности вариабельной области легкой цепи антитела, включающей последовательность CDR1, которая выбрана из группы, состоящей из SEQ ID NO: 13, 14, 15 и 16, последовательность CDR2, которая выбрана из группы, состоящей из SEQ ID NO: 17, 18, 19 и 20, а также последовательность CDR3, которая выбрана из группы, состоящей из SEQ ID NO: 21, 22, 23 и 24;
(b) изменение хотя бы одного аминокислотного остатка в хотя бы одной вариабельной области последовательности антитела, причем упомянутая последовательность выбрана из последовательности вариабельной области тяжелой цепи антитела и последовательности вариабельной области легкой цепи антитела, для создания хотя бы одной измененной последовательности антитела; и
(c) экспрессию измененной последовательности антитела в виде белка.
Кроме этого настоящее изобретение относится к способу ингибирования биологического действия интерферона типа I на экспрессируемый клетками рецептор 1 интерферона альфа, включающий взаимодействие клетки с антителом по настоящему изобретению, с тем, чтобы ингибировать биологическую активность интерферона типа I. Изобретение также относится к способу лечения заболеваний или расстройств у индивидуума, опосредованных интерфероном типа I, в случае необходимости такого лечения, включающему введение индивидууму антитела или его антиген-связывающего участка по настоящему изобретению, с тем, чтобы подвергнуть лечению заболевание, опосредованное интерфероном типа I. Заболевание, опосредованное интерфероном типа I, может, например, представлять собой заболевание, опосредованное интерфероном альфа.
Примеры заболеваний или расстройств, для которых может быть эффективно применение способов по настоящему изобретению, включают системную красную волчанку, инсулин-зависимый сахарный диабет, воспалительные заболевания кишечника, рассеянный склероз, псориаз, аутоиммунный тиреоидит, ревматоидный артрит, гломерулонефрит, ВИЧ-инфекцию, СПИД, отторжение трансплантата и реакцию «трансплантат против хозяина».
Другие отличительные особенности и преимущества настоящего изобретения станут ясны из следующего подробного описания и примеров, которые не должны истолковываться как ограничивающие. Содержание всех ссылок, позиций в генобанке, патентов и опубликованных заявок на патенты, цитируемых в тексте настоящей заявки, непосредственно включено в настоящую заявку с помощью ссылки.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1A показана нуклеотидная последовательность (SEQ ID NO:33) и аминокислотная последовательность (SEQ ID NO:25) вариабельной области тяжелой цепи моноклонального антитела 3F11 человека. Выделены области CDR1(SEQ ID NO:1), CDR2(SEQ ID NO:5) и CDR3(SEQ ID NO:9).
На фиг.1B показана нуклеотидная последовательность (SEQ ID NO:37) и аминокислотная последовательность (SEQ ID NO:29) вариабельной области легкой цепи моноклонального антитела 3F11 человека. Выделены области CDR1(SEQ ID NO:13), CDR2(SEQ ID NO:17) и CDR3(SEQ ID NO:21).
На фиг.2A показана нуклеотидная последовательность (SEQ ID NO:34) и аминокислотная последовательность (SEQ ID NO:26) вариабельной области тяжелой цепи моноклонального антитела 4G5 человека. Выделены области CDR1(SEQ ID NO:2), CDR2(SEQ ID NO:6) и CDR3(SEQ ID NO:10).
На фиг.2B показана нуклеотидная последовательность (SEQ ID NO:38) и аминокислотная последовательность (SEQ ID NO:30) вариабельной области легкой цепи моноклонального антитела 4G5 человека. Выделены области CDR1(SEQ ID NO:14), CDR2(SEQ ID NO:18) и CDR3(SEQ ID NO:22).
На фиг.3A показана нуклеотидная последовательность (SEQ ID NO:35) и аминокислотная последовательность (SEQ ID NO:27) вариабельной области тяжелой цепи моноклонального антитела 11E2 человека. Выделены области CDR1(SEQ ID NO:3), CDR2(SEQ ID NO:7) и CDR3(SEQ ID NO:11).
На фиг.3B показана нуклеотидная последовательность (SEQ ID NO:39) и аминокислотная последовательность (SEQ ID NO:31) вариабельной области легкой цепи моноклонального антитела 11E2 человека. Выделены области CDR1(SEQ ID NO:15), CDR2(SEQ ID NO:19) и CDR3(SEQ ID NO:23).
На фиг.4A показана нуклеотидная последовательность (SEQ ID NO:36) и аминокислотная последовательность (SEQ ID NO:28) вариабельной области тяжелой цепи моноклонального антитела 9D4 человека. Выделены области CDR1(SEQ ID NO:4), CDR2(SEQ ID NO:8) и CDR3(SEQ ID NO:12).
На фиг.4B показана нуклеотидная последовательность (SEQ ID NO:40) и аминокислотная последовательность (SEQ ID NO:32) вариабельной области легкой цепи моноклонального антитела 9D4 человека. Выделены области CDR1(SEQ ID NO:16), CDR2(SEQ ID NO:20) и CDR3(SEQ ID NO:24).
На фиг.5 показано совмещение аминокислотной последовательности вариабельной области тяжелой цепи 3F11 с аминокислотной последовательностью зародышевой линии VH 4-34 человека (SEQ ID NO:41).
На фиг.6 показано совмещение аминокислотной последовательности вариабельной области тяжелой цепи 4G5 с аминокислотной последовательностью зародышевой линии VH 4-34 человека (SEQ ID NO:41).
На фиг.7 показано совмещение аминокислотной последовательности вариабельной области тяжелой цепи 11E2 и 9D4 с аминокислотной последовательностью зародышевой линии VH 5-51 человека (SEQ ID NO:42).
На фиг.8 показано совмещение аминокислотной последовательности вариабельной области легкой цепи 3F11 с аминокислотной последовательностью зародышевой линии VK L18 человека (SEQ ID NO:43).
На фиг.9 показано совмещение аминокислотной последовательности вариабельной области легкой цепи 4G5 с аминокислотной последовательностью зародышевой линии VK L18 человека (SEQ ID NO:43).
На фиг.10 показано совмещение аминокислотной последовательности вариабельной области легкой цепи 11E2 и 9D4 с аминокислотной последовательностью зародышевой линии VK A27 человека (SEQ ID NO:44).
Фиг.11 представляет собой график, показывающий результаты экспериментов, демонстрирующих, что моноклональное антитело человека, а именно 3F11, направленное против IFNAR-1 человека, не конкурирует с моноклональным антителом 64G12 мыши за связывание с IFNAR-1.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделенным моноклональным антителам, которые связываются с рецептором 1 интерферона альфа (IFNAR-1) и которые способны блокировать действие интерферонов типа I. В настоящем изобретении разработаны выделенные антитела, способы получения этих антител, иммуноконъюгаты и биспецифичные молекулы, включающие такие антитела, а также фармацевтические композиции, содержащие указанные антитела, иммуноконъюгаты или биспецифичные молекулы по настоящему изобретению. Кроме этого настоящее изобретение относится к способам применения этих антител для ингибирования связывания интерферона типа I с IFNAR-1 на клетках, экспрессирующих IFNAR-1, например, при лечении иммунно-опосредованных расстройств у индивидуума, включая аутоиммунные расстройства, отторжение трансплантатов и реакцию "трансплантат против хозяина" (GVHD).
Для более легкого понимания настоящего изобретения в начале определяются некоторые термины. Дополнительные определения приведены по ходу подробного описания изобретения.
Термины «рецептор 1 интерферона альфа», «IFNAR-1» и «антиген IFNAR-1» являются взаимозаменяемыми и включают варианты, изоформы, разновидности гомологов IFNAR-1 человека, а также его аналоги, имеющие хотя бы один общий эпитоп с IFNAR-1. Соответственно, антитела человека по настоящему изобретению в некоторых случаях могут вступать в перекрестное взаимодействие с IFNAR-1 других биологических видов, кроме человека, или другими белками, которые структурно родственны с IFNAR-1 человека (например, гомологами IFNAR-1 человека). В других случаях антитела могут быть полностью специфичны для IFNAR-1 человека и могут не проявлять признаков способности к перекрестным взаимодействиям других типов.
Полная последовательность кДНК IFNAR-1 человека имеет инвентарный номер генобанка NM_000629.
Термин «интерферон типа I» в настоящем описании предназначен для указания на членов семейства молекул интерферонов типа I, которые являются лигандами для IFNAR-1 (т.е. членов семейства молекул интерферонов типа I, которые способны связываться с IFNAR-1). Примерами лигандов, относящихся к интерферонам типа I, являются интерферон альфа 1, 2a, 2b, 4, 5, 6, 7, 8, 10, 14, 16, 17, 21, интерферон бета и интерферон омега.
Термин «иммунный ответ» относится к действию, например, лимфоцитов, антигенпредставляющих клеток, фагоцитов, гранулоцитов и растворимых макромолекул, вырабатываемых указанными клетками или печенью (включая антитела, цитокины и комплемент), которое приводит к селективному повреждению, или разрушению, или удалению из организма человека проникших в него патогенов, клеток или тканей, инфицированных патогенами, раковых клеток или, в случаях аутоиммунитета или патологического воспаления, нормальных клеток или тканей.
Термин «путь трансдукции сигнала» относится к биохимической связи между различными молекулами, преобразующими сигнал, которые играют роль в передаче сигнала из одной части клетки в другую ее часть. В настоящем описании фраза «рецептор клеточной поверхности» включает в себя, например, молекулы и комплексы молекул, способные получать сигнал и передавать этот сигнал через цитоплазматическую оболочку клетки. Примером «рецептора клеточной поверхности» в настоящем изобретении является рецептор IFNAR-1.
Термин «антитело» в рамках настоящего описания включает целые антитела и любые их антиген-связывающие фрагменты (т.е. «антиген-связывающие участки») или отдельные цепи. Термин «антитело» относится к гликопротеинам, содержащим как минимум две тяжелых цепи (H) и две легких цепи (L), связанных дисульфидными связями, или к их антиген-связывающим участкам. В каждую тяжелую цепь включена вариабельная область тяжелой цепи (сокращенно обозначаемая в тексте VH) и константная область тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов CH1, CH2 и CH3. В каждую из легких цепей включена вариабельная область легкой цепи (сокращенно обозначаемая в тексте VL) и константная область легкой цепи. Константная область легкой цепи состоит из одного домена, обозначаемого CL. Области VH и VL далее могут быть подразделены на области гипервариабельности, называемые областями, определяющими комплементарность (CDR), которые чередуются с более консервативными фрагментами, именуемыми каркасными областями (FR). Каждая из цепей VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелых и легких цепей содержат связывающие домены, которые взаимодействуют с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с хозяйскими тканями или факторами, включая различные клетки иммунной системы (например, клетки-эффекторы) и первый компонент (Clq) классической системы комплемента.
Термин «антиген-связывающий участок» антитела (или просто «участок антитела») в настоящем описании относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфично связываться с антигеном (например, IFNAR-1). Было показано, что антиген-связывающая функция антитела может осуществляться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, охваченных термином «антиген-связывающий участок» антитела, включают (i) фрагмент Fab, т.е. моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) фрагмент F(ab')2, т.е. бивалентный фрагмент, включающий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fd, состоящий из доменов VH и CH1; (iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела; (v) фрагмент dAb (Ward et al. (1989) Nature, 341:544-546), который состоит из домена VH; и (vi) выделенную область, определяющую комплементарность (CDR). Кроме того, хотя два домена фрагмента Fv, т.е. VL и VH, закодированы отдельными генами, они могут быть объединены синтетическим линкером с использованием рекомбинантных способов, причем линкер дает возможность получить их в виде единой белковой цепи, в которой пара областей VL и VH образует моновалентные молекулы (известные как одноцепочечные Fv (scFv); см., например, Bird et al. (1988) Science, 242: 423-426; и Huston et al. (1988) Proc.Natl.Acad.Sci. USA, 85: 5879-5883). Имеется в виду, что такие одноцепочечные антитела также охвачены термином «антиген-связывающий участок» антитела. Эти фрагменты антител получают с использованием стандартных методик, известных специалистам в данной области техники, и фрагменты подвергают отбору по их полезности тем же способом, что и интактные антитела.
Термин «выделенное антитело» в настоящем изобретении предназначен для указания на антитело, которое в основном свободно от других антител, обладающих другими антигенными специфичностями (например, выделенное антитело, которое специфично связывается с IFNAR-1, в основном свободно от антител, которые специфично связываются с другими антигенами, отличающимися от IFNAR-1). Однако выделенное антитело, которое специфично связывается с IFNAR-1, может обладать перекрестной реакционной способностью в отношении других антигенов, таких как молекулы IFNAR-1 других видов. Кроме того, выделенное антитело может быть в основном свободно от других клеточных веществ и/или химических соединений.
Термины «моноклональное антитело» или «композиция на основе моноклонального антитела» в настоящем описании относятся к препарату на основе молекул антитела одного молекулярного состава. Композиция на основе моноклонального антитела проявляет одну специфичность связывания и сродство к конкретному эпитопу.
Предполагается, что в настоящем описании термин «антитело человека» включает антитела, имеющие вариабельные области, в которых как каркасные, так и CDR области получены из иммуноглобулиновых последовательностей зародышевой линии клеток человека. Кроме того, если антитело содержит константную область, константная область также получена из иммуноглобулиновых последовательностей зародышевой линии клеток человека. Антитела человека по настоящему изобретению могут включать аминокислотные остатки, которые не закодированы иммуноглобулиновыми последовательностями зародышевой линии клеток человека (например, мутации, внесенные случайным или сайт-специфичным мутагенезом in vitro или соматической мутацией in vivo). Однако в настоящем описании не предполагается, что термин «антитело человека» включает антитела с последовательностями CDR, полученными из зародышевых линий клеток других видов млекопитающих, как например мышей, которые привиты к каркасным последовательностям человека.
Термин «моноклональное антитело человека» относится к антителам, демонстрирующим одну специфичность связывания, которые имеют вариабельные области, в которых как каркасные, так и CDR области получены из иммуноглобулиновых последовательностей зародышевой линии клеток человека. В одном из вариантов осуществления моноклональные антитела человека вырабатываются гибридомой, которая включает B-клетки, полученные из трансгенных животных, не относящихся к человеческому роду, например трансгенных мышей, имеющих геном, который включает трансген тяжелой цепи человека и трансген легкой цепи человека, слитый с иммортализованной клеткой.
Термин «рекомбинантное антитело человека» в настоящем описании включает все антитела человека, которые образуются, экспрессируются, создаются или изолируются средствами рекомбинантных технологий, как например (a) антитела, изолированные из животных (например, мышей), которые являются трансгенными или трансхромосомными, для получения из них генов иммуноглобулина человека или гибридомы (описанного ниже по тексту), (b) антитела, выделенные из клеток-хозяев, преобразованных для экспрессии антител человека, например, из трансфектомы, (c) антитела, выделенные из рекомбинантных, комбинаторных библиотек антител человека, и (d) антитела, полученные, экспрессированные, созданные или выделенные любым другим способом, который включает сплайсинг последовательностей гена иммуноглобулина человека с другими последовательностями ДНК. Такие рекомбинантные антитела человека имеют вариабельные области, в которых каркасные и CDR области получены из иммуноглобулиновых последовательностей зародышевой линии клеток человека. Однако в некоторых вариантах осуществления такие рекомбинантные антитела человека могут быть подвергнуты in vitro мутагенезу (или, если используется животное, являющееся трансгенным вследствие наличия последовательностей Ig человека, in vivo соматическому мутагенезу) и, таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител являются последовательностями, которые, будучи получены из и являясь родственными последовательностям VH и VL зародышевых линий клеток человека, могут отсутствовать в природе в ассортименте антител зародышевой линии человека in vivo.
В настоящем описании термин «изотип» относится к классу антител (например, IgM или IgGI), которые закодированы генами константной области тяжелой цепи.
Термин «специфичное связывание» относится в настоящем описании к связыванию антитела с заданным антигеном. Как правило, антитело связывается с константой диссоциации (KD), равной 10-7 М или меньшей, причем связывание с заданным антигеном происходит с KD, которая, по крайней мере, в два раза меньше KD для связывания этого антитела с неспецифичным антигеном (например, BSA, казеином), который отличается от заданного или близкородственного антигена. Фразы «антитело, распознающее антиген» и «антитело, специфичное для антигена» в настоящем описании являются взаимозаменяемыми с термином «антитело, которое специфично связывается с антигеном».
Термин «Kасс» или «Ka» в настоящем изобретении предназначен для указания на скорость ассоциации при конкретном взаимодействии антитело-антиген, тогда как термин «Kдис» или «Kd» в настоящем описании предназначен для указания на скорость диссоциации при конкретном взаимодействии антитело-антиген. Термин «KD» в настоящем описании предназначен для указания на константу диссоциации, которая получается из отношения Kd к Ka (т.е. Kd/Ka) и выражается в единицах молярной концентрации (M). Значения KD антител могут быть определены с применением методик, хорошо отработанных в технике. Предпочтительный способ определения KD антитела состоит в использовании поверхностного плазмонного резонанса, предпочтительно с применением биосенсорной системы, такой как система Biacore®.
В настоящем описании термин «высокое сродство» к IgG антителу относится к антителу, имеющему KD 10-8 М или менее, более предпочтительно 10-9 М или менее и еще более предпочтительно 10-10 М или менее. Однако связывание с «высоким сродством» может изменяться для антител других изотипов. Например, связывание с «высоким сродством» для изотипа IgM относится к антителу, имеющему KD 10-7 М или менее, более предпочтительно 10-8 М или менее.
В настоящем описании термин «субъект» включает людей и животных, не относящихся к человеческому роду. Термин «животное, не относящееся к человеческому роду» включает всех позвоночных, например млекопитающих и не млекопитающих, таких как приматы, овцы, собаки, кошки, лошади, коровы, куры, земноводные, пресмыкающиеся и т.д.
В следующих далее подразделах различные аспекты настоящего изобретения описаны с дополнительными подробностями.
Антитела анти-IFNAR-1
Антитела по настоящему изобретению характеризуются определенными функциональными особенностями или свойствами антител. Например, эти антитела специфично связываются с IFNAR-1, предпочтительно IFNAR-1 человека. Кроме этого антитела могут вступать в перекрестное взаимодействие с IFNAR-1 одного или нескольких приматов, кроме человека, как например обезьян cynomolgus и/или макак резус. Предпочтительно антитело по настоящему изобретению связывается с IFNAR-1 с высоким сродством, например, с KD, равной 10-7 M или менее, более предпочтительно с KD, равной 10-8 M или менее, или 10-9 M или менее, или даже 5·10-10 M или менее, или 2·10-10 M или менее.
Кроме того, антитела по настоящему изобретению способны ингибировать биологическую активность интерферонов типа I. Эти антитела ингибируют биологическую активность хотя бы одного интерферона типа I и предпочтительно ингибируют биологическую активность большого количества интерферонов типа I (т.е. как минимум двух, более предпочтительно как минимум трех, или как минимум четырех, или как минимум пяти, или как минимум шести, или как минимум семи, или как минимум восьми, или как минимум девяти, или как минимум десяти, или как минимум 11, или как минимум 12, или как минимум 13, или как минимум 14, или как минимум 15 различных подтипов интерферонов типа I). В предпочтительном варианте осуществления антитело ингибирует биологическую активность следующих интерферонов типа I: α1, α2a, α2b, α4, α5, α6, α7, α8, α10, α14, α16, α17, α21, бета и омега. В других предпочтительных вариантах осуществления антитело ингибирует активность лимфобластоидного интерферона и/или лейкоцитарного интерферона.
Способность антитела ингибировать биологическую активность интерферонов типа I может быть проверена в одном или нескольких анализах, известных в технике. Неограничивающие примеры включают ингибирование пролиферации клеток Daudi, опосредованное ингибированием интерферона типа I, ингибирование вызванной интерфероном типа I экспрессии IP-10 моноядерными клетками перифиерической крови (PBMC), ингибирование развития дендритных клеток, опосредованное плазмой системной красной волчанки (SLE), а также ингибирование противовирусной активности интерферона типа I. Антитело «ингибирует биологическую активность интерферонов типа I», если оно эту ингибирует активность как минимум на 20%, более предпочтительно как минимум на 30%, еще более предпочтительно как минимум на 40%, как минимум на 50%, как минимум на 60%, как минимум на 70%, как минимум на 80% или как минимум на 90%, по сравнению с неспецифичным контрольным антителом.
В предпочтительных вариантах осуществления антитело ингибирует активность IFN-α 2b в анализе, основанном на пролиферации клеток Daudi, ингибирует активность IFN омега в анализе, основанном на пролиферации клеток Daudi, ингибирует секрецию IP-10 клетками PBMC, вызванную IFN-α 2b или IFN омега, и/или ингибирует развитие дендритных клеток, опосредованное плазмой SLE.
В другом предпочтительном варианте осуществления антитело не вступает в перекрестную конкуренцию (т.е. связывается с другим эпитопом) с анти-IFNAR-1 антителом 64G12 мыши (депонировано как ECACC, депозитарный №92022605).
В примерах более подробно описаны анализы для оценки функциональной активности анти-IFNAR антител. Предпочтительные антитела по настоящему изобретению проявляют хотя бы одно, более предпочтительно два, три, четыре, пять или более, из следующих свойств:
a) специфично связываются с IFNAR-1 (предпочтительно IFNAR-1 человека);
b) связываются с IFNAR-1 с высоким сродством, так что KD составляет 1·10-8 М или более высоким сродством;
c) ингибируют биологическую активность нескольких интерферонов типа I;
d) ингибируют активность IFN-α 2b в анализе пролиферации клеток Daudi;
e) ингибируют активность IFN омега в анализе пролиферации клеток Daudi;
f) ингибируют секрецию IP-10 моноядерными клетками периферической крови, вызванную IFN-α 2b;
g) ингибируют секрецию IP-10 моноядерными клетками периферической крови, вызванную IFN омега;
h) ингибируют развитие дендритных клеток, опосредованное плазмой системной красной волчанки; и
i) связываются с другим эпитопом (например, не участвует в перекрестной конкуренции), чем моноклональное антитело 64G12 мыши (ECACC, депозитарный № 92022605).
Антитело по настоящему изобретению может проявлять любые комбинации описанных выше функциональных особенностей и/или функциональных особенностей, описанных в примерах.
Моноклональные антитела 3F11, 4G5, 11E2 и 9D4
Предпочтительными антителами по настоящему изобретению являются моноклональные антитела человека 3F11, 4G5, 11E2 и 9D4, выделенные и структурно охарактеризованные, как описано в примерах. Аминокислотные последовательности VH 3F11, 4G5, 11E2 и 9D4 показаны в SEQ ID NO: 25, 26, 27 и 28 соответственно. Аминокислотные последовательности VL 3F11, 4G5, 11E2 и 9D4 показаны в SEQ ID NO: 29, 30, 31 и 32 соответственно.
С учетом того, что каждое из этих антител может связываться с IFNAR-1, последовательности VH и VL могут быть «смешаны и подобраны в пары» для создания других анти-IFNAR-1 связывающих молекул по настоящему изобретению. Связывание таких «смешанных и подобранных в пары» антител с IFNAR-1 может быть исследовано с использованием описанных в настоящей заявке анализов связывания (например, ELISA) и/или с использованием функциональных анализов ингибирования интерферона типа I, описанных в примерах. Предпочтительно, если цепи VH и VL смешаны и подобраны в пары, последовательность VH из конкретной пары VH/VL заменена подобной по структуре последовательностью VH. Аналогично, предпочтительно последовательность VL из конкретной пары VH/VL заменена подобной по структуре последовательностью VL. Например, последовательности VH и VL антител 3F11 и 4G5 особенно подходят для смешивания и подбора в пару, поскольку эти антитела используют последовательности VH и VL, полученные из одних и тех же последовательностей зародышевых линий (VH 4-34 и VK L18), и, таким образом, они проявляют структурное подобие. В дополнение, последовательности VH и VL 11E2 и 9D4 особенно подходят для смешивания и подбора в пару, т.к. эти антитела используют последовательности VH и VL, полученные из одних и тех же последовательностей зародышевых линий (VH 5-51 и VK A27), и, таким образом, они проявляют структурное подобие.
Соответственно в одном из аспектов настоящее изобретение относится к выделенному моноклональному антителу или его антиген-связывающему участку, включающим:
(a) вариабельную область тяжелой цепи, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 25, 26, 27 и 28; и
(b) вариабельную область легкой цепи, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 29, 30, 31 и 32;
где антитело специфично связывается IFNAR-1.
Предпочтительные комбинации тяжелой и легкой цепей включают:
(a) вариабельную область тяжелой цепи, включающую аминокислотную последовательность SEQ ID NO: 25; и (b) вариабельную область легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 29; или
(a) вариабельную область тяжелой цепи, включающую аминокислотную последовательность SEQ ID NO: 26; и (b) вариабельную область легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 30; или
(a) вариабельную область тяжелой цепи, включающую аминокислотную последовательность SEQ ID NO: 27; и (b) вариабельную область легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 31; или
(a) вариабельную область тяжелой цепи, включающую аминокислотную последовательность SEQ ID NO: 28; и (b) вариабельную область легкой цепи, включающую аминокислотную последовательность SEQ ID NO: 32.
В другом аспекте настоящее изобретение относится к антителам, которые включают тяжелые и легкие цепи CDR1, CDR2 и CDR3 антител 3F11, 4G5, 11E2 и 9D4 или их комбинации. Аминокислотные последовательности VH CDR1 3F11, 4G5, 11E2 и 9D4 показаны в SEQ ID NO: 1, 2, 3 и 4. Аминокислотные последовательности VH CDR2 3F11, 4G5, 11E2 и 9D4 показаны в SEQ ID NO: 5, 6, 7 и 8. Аминокислотные последовательности VH CDR3 3F11, 4G5, 11E2 и 9D4 показаны в SEQ ID NO: 9, 10, 11 и 12. Аминокислотные последовательности VK CDR1 3F11, 4G5, 11E2 и 9D4 показаны в SEQ ID NO: 13, 14, 15 и 16. Аминокислотные последовательности VK CDR2 3F11, 4G5, 11E2 и 9D4 показаны в SEQ ID NO: 17, 18, 19 и 20. Аминокислотные последовательности VK CDR3 3F11, 4G5, 11E2 и 9D4 показаны в SEQ ID NO: 21, 22, 23 и 24. Области CDR выделены с использованием системы Kabat (Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S.Department of Health and Human Services, NIH Publication No 91-3242).
С учетом того, что каждое из этих антител может связываться с IFNAR-1, и того, что антиген-связывающая специфичность обеспечивается в первую очередь областями CDR1, 2 и 3, последовательности VH CDR1, 2 и 3 и VK CDR1, 2 и 3 могут быть «смешаны и подобраны в пары» (т.е. CDR различных антител могут быть смешаны и подобраны в пары, хотя каждое антитело должно содержать VH CDR1, 2 и 3 и VK CDR1, 2 и 3) для создания других анти-IFNAR-1 связывающих молекул по настоящему изобретению. Связывание таких «смешанных и подобранных в пары» антител с IFNAR-1 может быть исследовано с использованием анализов связывания, описанных выше и в примерах (например, ELISA). Предпочтительно, если последовательности VH CDR смешаны и подобраны в пары, последовательности CDR1, CDR2 и/или CDR3 из конкретной последовательности VH заменены подобной по структуре последовательностью (последовательностями) CDR. Аналогично, если последовательности VK CDR смешаны и подобраны в пары, последовательности CDR1, CDR2 и/или CDR3 из конкретной последовательности VK предпочтительно заменены подобной по структуре последовательностью (последовательностями) CDR. Например, последовательности VH CDR1 антител 3F11 и 4G5 обладают некоторым структурным сходством и, следовательно, подходят для смешивания и подбора в пару. В качестве другого примера, последовательности VH CDR1 антител 11E2 и 9D4 обладают некоторым структурным сходством и, следовательно, подходят для смешивания и подбора в пару. В качестве еще одного примера, последовательности VK CDR1 антител 3F11 и 4G5 обладают некоторым структурным сходством. В качестве еще одного примера, последовательности VH CDR1 антител 11E2 и 9D4 обладают некоторым структурным сходством. Рядовой специалист в данной области техники легко поймет, что новые последовательности VH и VL могут быть созданы замещением одной или нескольких последовательностей областей CDR VH и/или VL, структурно сходными последовательностями из последовательностей CDR, раскрытых в настоящем описании для моноклональных антител 3F11, 4G5, 11E2 и 9D4.
Соответственно в другом аспекте изобретение относится к выделенному моноклональному антителу или его антиген-связывающему участку, включающим:
(a) вариабельную область тяжелой цепи CDR1, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, 2, 3 и 4;
(b) вариабельную область тяжелой цепи CDR2, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, 6, 7 и 8;
(c) вариабельную область тяжелой цепи CDR3, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, 10, 11 и 12;
(d) вариабельную область легкой цепи CDR1, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13, 14, 15 и 16;
(e) вариабельную область легкой цепи CDR2, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 17, 18, 19 и 20; и
(f) вариабельную область легкой цепи CDR3, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, 22, 23 и 24;
причем антитело специфично связывается с IFNAR-1.
В предпочтительных вариантах осуществления антитело включает:
(a) вариабельную область тяжелой цепи CDR1, включающую SEQ ID NO:1;
(b) вариабельную область тяжелой цепи CDR2, включающую SEQ ID NO:5;
(c) вариабельную область тяжелой цепи CDR3, включающую SEQ ID NO:9;
(d) вариабельную область легкой цепи CDR1, включающую SEQ ID NO:13;
(e) вариабельную область легкой цепи CDR2, включающую SEQ ID NO:17; и
(f) вариабельную область легкой цепи CDR3, включающую SEQ ID NO:21.
В другом предпочтительном варианте осуществления антитело включает:
(a) вариабельную область тяжелой цепи CDR1, включающую SEQ ID NO:2;
(b) вариабельную область тяжелой цепи CDR2, включающую SEQ ID NO:6;
(c) вариабельную область тяжелой цепи CDR3, включающую SEQ ID NO:10;
(d) вариабельную область легкой цепи CDR1, включающую SEQ ID NO:14;
(e) вариабельную область легкой цепи CDR2, включающую SEQ ID NO:18; и
(f) вариабельную область легкой цепи CDR3, включающую SEQ ID NO:22.
В другом предпочтительном варианте осуществления антитело включает:
(a) вариабельную область тяжелой цепи CDR1, включающую SEQ ID NO:3;
(b) вариабельную область тяжелой цепи CDR2, включающую SEQ ID NO:7;
(c) вариабельную область тяжелой цепи CDR3, включающую SEQ ID NO:11;
(d) вариабельную область легкой цепи CDR1, включающую SEQ ID NO:15;
(e) вариабельную область легкой цепи CDR2, включающую SEQ ID NO:19; и
(f) вариабельную область легкой цепи CDR3, включающую SEQ ID NO:23.
В еще одном предпочтительном варианте осуществления антитело включает:
(a) вариабельную область тяжелой цепи CDR1, включающую SEQ ID NO:4;
(b) вариабельную область тяжелой цепи CDR2, включающую SEQ ID NO:8;
(c) вариабельную область тяжелой цепи CDR3, включающую SEQ ID NO:12;
(d) вариабельную область легкой цепи CDR1, включающую SEQ ID NO:16;
(e) вариабельную область легкой цепи CDR2, включающую SEQ ID NO:20; и
(f) вариабельную область легкой цепи CDR3, включающую SEQ ID NO:24.
Антитела, которые связываются с тем же эпитопом, что и 3F11, 4G5, 11E2 и 9D4
В другом варианте осуществления изобретение относится к антителам, которые связываются с тем же эпитопом на IFNAR-1 человека, что и моноклональные антитела 3F11, 4G5, 11E2 или 9D4 (имеющие последовательности VH, которые показаны в SEQ ID NO: 25, 26, 27 и 28 соответственно, и последовательности VL, которые показаны в SEQ ID NO: 29, 30, 31 и 32 соответственно). Такие антитела могут быть идентифицированы на основе их способности вступать в перекрестную конкуренцию с 3F11, 4G5, 11E2 или 9D4 в стандартных анализах связывания с IFNAR-1. Способность исследуемого антитела ингибировать связывание 3F11, 4G5, 11E2 или 9D4 с IFNAR-1 человека демонстрирует, что исследуемое антитело может конкурировать с 3F11, 4G5, 11E2 или 9D4 за связывание с IFNAR-1 человека и, таким образом, связываться с тем же эпитопом IFNAR-1 человека, что и антитела 3F11, 4G5, 11E2 или 9D4. В предпочтительном варианте осуществления антитело, которое связывается с тем же эпитопом IFNAR-1 человека, что и 3F11, 4G5, 11E2 или 9D4, представляет собой моноклональное антитело человека. Такие моноклональные антитела человека могут быть получены и выделены, как описано в примерах.
В другом предпочтительном варианте осуществления антитело связывается с другим эпитопом (т.е. не участвует в перекрестной конкуренции), чем моноклональное антитело 64G12 мыши (ECACC, депозитарный № 92022605).
Антитела, имеющие конкретные последовательности зародышевых линий
В некоторых вариантах осуществления антитело по настоящему изобретению включает вариабельную область тяжелой цепи из гена тяжелой цепи иммуноглобулина конкретной зародышевой линии и/или вариабельную область легкой цепи из гена легкой цепи иммуноглобулина конкретной зародышевой линии.
Например, в предпочтительном варианте осуществления изобретение относится к выделенному анти-IFNAR-1 моноклональному антителу или его антиген-связывающему участку, причем антитело:
(a) включает вариабельную область тяжелой цепи из генов VH 4-34 или 5-51 человека;
(b) включает вариабельную область легкой цепи из генов VK L18 или A27 человека; и
(c) антитело специфично связывается с IFNAR-1.
Примеры антител, имеющих VH и VK из VH 4-34 и VK L18 соответственно, включают 3F11 и 4G5. Примеры антител, имеющих VH и VK из VH 5-51 и VK A27 соответственно, включают 11E2 и 9D4.
В настоящем описании антитело человека включает вариабельные области тяжелой или легкой цепей «из», или «полученные из», или «являющиеся продуктом» определенных последовательностей зародышевых линий, если вариабельные области антитела получены из системы, которая использует гены иммуноглобулина зародышевых линий человека. Такие системы включают иммунизирование трансгенной мыши, имеющей гены иммуноглобулина человека, представляющим интерес антигеном или скрининг библиотеки генов иммуноглобулина человека в фаг-дисплейной системе с представляющим интерес антигеном. Антитело человека, в отношении которого употребляются термины «из», или «полученное из», или «являющееся продуктом» иммуноглобулиновой последовательности зародышевой линии человека, может быть идентифицировано само по себе, путем сравнения аминокислотной последовательности антитела человека с аминокислотными последовательностями иммуноглобулинов зародышевой линии человека и выбора иммуноглобулиновой последовательности зародышевой линии человека, которая наиболее точно соответствует (т.е. наибольший % соответствия) последовательности антитела человека. Антитело человека, в отношении которого употребляются термины «из», или «полученное из», или «являющееся продуктом» иммуноглобулиновой последовательности конкретной зародышевой линии человека, может содержать отличия в аминокислотах по сравнению с последовательностью зародышевой линии, благодаря, например, природным соматическим мутациям или намеренному введению сайт-направленной мутации. Однако выбранное антитело человека по аминокислотной последовательности, как правило, хотя бы на 90% идентично аминокислотной последовательности, закодированной геном иммуноглобулина зародышевой линии человека, и содержит аминокислотные остатки, которые определяют человеческую природу антитела человека при сравнении с иммуноглобулиновыми аминокислотными последовательностями зародышевых линий других видов (например, последовательностями зародышевых линий мыши). В некоторых случаях антитело человека может быть, по крайней мере, на 95% или даже, по крайней мере, на 96%, 97%, 98% или 99% идентично по аминокислотной последовательности, аминокислотной последовательности, закодированной геном иммуноглобулина зародышевой линии. Как правило, в антителе человека, полученном из конкретной последовательности зародышевой линии человека, не будет выявляться более 10 различий в остатках аминокислот по сравнению с аминокислотной последовательностью, закодированной геном иммуноглобулина зародышевой линии человека. В некоторых случаях в антителе человека может выявляться не более 5 или даже не более 4, 3, 2 или 1 различия в остатках аминокислот по сравнению с аминокислотной последовательностью, закодированной геном иммуноглобулина зародышевой линии.
Гомологичные антитела
В еще одном варианте осуществления антитело по настоящему изобретению включает вариабельные области тяжелой и легкой цепей, содержащие аминокислотные последовательности, которые гомологичны аминокислотным последовательностям описанных в настоящей заявке предпочтительных антител, причем антитела сохраняют желаемые функциональные свойства анти-IFNAR-1 антител по настоящему изобретению. Например, изобретение относится к выделенному моноклональному антителу или его антиген-связывающему участку, включающему вариабельную область тяжелой цепи и вариабельную область легкой цепи, в которых:
(a) вариабельная область тяжелой цепи включает аминокислотную последовательность, которая как минимум на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 25, 26, 27 и 28;
(b) вариабельная область легкой цепи включает аминокислотную последовательность, которая как минимум на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 29, 30, 31 и 32;
(c) антитело специфично связывается с IFNAR-1 и проявляет как минимум одно из функциональных свойств, описанных в настоящей заявке, предпочтительно несколько из описанных в заявке свойств.
В других вариантах осуществления аминокислотные последовательности VH и/или VL могут быть на 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологичны приведенным выше последовательностям. Антитело, в котором области VH и VL имеют высокую (т.е. 80% или более) степень гомологии областям VH и VL приведенных выше последовательностей, может быть получено мутагенезом (например, сайт-направленным или PCR-опосредованным мутагенезом) молекул нуклеиновых кислот, кодирующих SEQ ID NO: 33, 34, 35, 36, 37, 38, 39 или 40 с последующим тестированием изменившегося закодированного антитела на сохранение его функций (т.е. функций, изложенных выше в пп. (c), (d) и (e)) с применением описанных в настоящей заявке функциональных анализов.
В настоящем описании процент гомологии между двумя аминокислотными последовательностями эквивалентен проценту идентичности между этими двумя последовательностями. Процент идентичности между двумя последовательностями является функцией числа идентичных позиций, которые присутствуют в двух последовательностях одновременно (т.е. % гомологии = число идентичных позиций/общее число позиций ×100), с учетом числа разрывов и длины каждого разрыва, которые следует ввести для оптимального совмещения этих двух последовательностей. Сравнение последовательностей и определение процента идентичности между двумя последовательностями может выполняться с использованием математического алгоритма, как описано в приведенном ниже неограничивающем примере.
Процент идентичности между двумя аминокислотными последовательностями может быть определен с применением алгоритма E.Meyers и W.Miller (Comput. Appl.Biosci., 4:11-17 (1988)), который был включен в программу ALIGN (версия 2.0) при использовании таблицы весовых остатков PAM120, параметра “gap length penalty”, равного 12, и параметра “gap penalty”, равного 4. Кроме этого процент идентичности между двумя аминокислотными последовательностями может быть определен с применением алгоритма Needleman и Wunsch (J.Mol.Biol. 48:444-453 (1970)), который был включен в программу GAP программного пакета GCG (доступен на сайте http://www.gcg.com), с использованием либо матрицы Blossum 62, либо матрицы PAM250, а также параметра “gap weight” 16, 14, 12, 10, 8, 6 или 4 и “length weight” 1, 2, 3, 4, 5 или 6.
В качестве дополнения или альтернативы, белковые последовательности по настоящему изобретению могут дополнительно использоваться в качестве «элемента базы данных» для осуществления поиска по общедоступным базам данных с целью, например, идентификации родственных последовательностей. Такие поиски могут выполняться с применением программы XBLAST (версия 2.0) Altschul, et al. (1990) J.Mol.Biol. 215:403-10. Для получения аминокислотных последовательностей, гомологичных молекулам антител по настоящему изобретению, можно выполнить поиски белков BLAST с помощью программы XBLAST, при значениях параметров “score”=50, “wordlength”=3. Для получения пропущенных совмещений с целями сравнения, может быть использована программа Gapped BLAST, как описано Altschul, et al. (1997) Nucleic Acid Res. 25(17):3389-3402. При использовании программ BLAST и Gapped BLAST могут применяться параметры соответствующей программы по умолчанию (например, XBLAST и NBLAST). См. http://www.ncbi.nlm.nih.gov.
Антитела с консервативными модификациями
В некоторых вариантах осуществления антитело по настоящему изобретению включает вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, где одна или несколько из этих последовательностей CDR включают определенные аминокислотные последовательности, основанные на предпочтительных антителах, описанных в настоящей заявке (например, 3F11, 4G5, 11E2 и 9D4), или их консервативные модификации, и где антитела сохраняют желаемые функциональные свойства анти-IFNAR-1 антител по настоящему изобретению. Соответственно изобретение относится к выделенному моноклональному антителу или его антиген-связывающему участку, включающему вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, причем:
(a) последовательность CDR3 вариабельной области тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 9, 10, 11 и 12 и их консервативных модификаций;
(b) последовательность CDR3 вариабельной области легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 21, 22, 23 и 24 и их консервативных модификаций; и
(c) антитело специфично связывается с IFNAR-1 и проявляет как минимум одно из функциональных свойств, описанных в настоящей заявке, предпочтительно несколько из описанных в заявке свойств.
В другом варианте осуществления последовательность CDR2 вариабельной области тяжелой цепи включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 5, 6, 7 и 8 и их консервативных модификаций; и последовательность CDR2 вариабельной области легкой цепи включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 17, 18, 19 и 20 и их консервативных модификаций. В еще одном варианте осуществления последовательность CDR1 вариабельной области тяжелой цепи включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 1, 2, 3 и 4 и их консервативных модификаций; и последовательность CDR1 вариабельной области легкой цепи включает аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 13, 14, 15 и 16 и их консервативных модификаций.
В настоящей заявке термин «консервативные модификации последовательностей» предназначен для указания на модификации аминокислотных последовательностей, которые не оказывают существенного влияния или существенно не изменяют характеристики связывания антитела, содержащего аминокислотную последовательность. Такие консервативные модификации включают замещения, добавления или делеции аминокислот. Модификации в антитело по настоящему изобретению могут вноситься с помощью стандартных методик, известных в технике, таких как сайт-направленный мутагенез и PCR-опосредованный мутагенез. Консервативными замещениями аминокислот являются такие замещения, при которых аминокислотный остаток замещается аминокислотным остатком, имеющим сходную боковую цепь. В технике были определены семейства аминокислотных остатков, имеющих сходные боковые цепи. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), боковыми цепями с бета-разветвлением (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или несколько аминокислотных остатков в областях CDR антитела по настоящему изобретению могут быть заменены другими аминокислотными остатками с боковой цепью из того же семейства, и измененное антитело может быть протестировано с точки зрения сохранения функций (т.е. функций, изложенных выше в пп. (c), (d) и (e)) с применением описанных в настоящей заявке функциональных анализов.
Сконструированные и модифицированные антитела
Помимо описанного выше, антитело по настоящему изобретению может быть получено с использованием антитела, имеющего одну или несколько последовательностей VH и/или VL, раскрытых в настоящей заявке, в качестве исходного вещества для конструирования модифицированного антитела, которое может иметь свойства, отличающиеся от исходного антитела. Антитело может быть сконструировано с помощью модификации одного или большего числа остатков в одной или обеих вариабельных областях (т.е. VH и/или VL), например в одной или нескольких областях CDR и/или в одной или нескольких каркасных областях. В качестве дополнения или альтернативы, антитело может быть сконструировано за счет модификации остатков в константной области (областях), например, для изменения эффекторной функции (функций) антитела.
Одним из возможных типов конструирования вариабельной области является пересадка CDR. Антитела преимущественно взаимодействуют с целевыми антигенами через аминокислотные остатки, которые расположены в легкой и тяжелой цепях в шести определяющих комплементарность областях (CDR). Поэтому у различных антител аминокислотные последовательности в CDR различаются в большей степени, чем за пределами CDR. Поскольку последовательности CDR отвечают за большинство взаимодействий антитело-антиген, имеется возможность экспрессии рекомбинантных антител, которые имитируют свойства конкретных природных антител, путем конструирования векторов экспрессии, которые включают последовательности CDR из конкретного природного антитела, пересаженные на каркасные последовательности другого антитела с отличающимися свойствами (см., например, Riechmann, L. et al. (1998) Nature, 332:323-327; Jones, P. et al. (1986) Nature, 321: 522-525; Oueen, C. et al (1989) Proc.Natl.Acad.See.USA 86:10029-10033; Патент США №5 225 539, выданный Winter, и патенты США №№ 5530101; 5585089; 5693762 и 6180370, выданные Queen и соавторам).
Соответственно другой вариант осуществления настоящего изобретения имеет отношение к выделенному моноклональному антителу или его антиген-связывающему участку, включающему: вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, включающие аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 1, 2, 3 и 4, SEQ ID NO: 5, 6, 7 и 8 и SEQ ID NO: 9, 10, 11 и 12 соответственно, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, включающие аминокислотные последовательности, выбранные из группы, состоящей из SEQ ID NO: 13, 14, 15 и 16, SEQ ID NO: 17, 18, 19 и 20 и SEQ ID NO: 21, 22, 23 и 24 соответственно. Таким образом, эти антитела содержат последовательности CDR VH и VL моноклональных антител 3F11, 4G5, 11E2 или 9D4, и, кроме того, они могут содержать различные каркасные последовательности из этих антител.
Такие каркасные последовательности могут быть получены из общедоступных баз данных ДНК или опубликованной справочной информации, которая включает последовательности генов антител зародышевых линий. Например, последовательности ДНК зародышевых линий для генов вариабельных областей тяжелой и легкой цепей человека могут быть найдены в базе данных “VBase” последовательностей зародышевых линий человека (доступной в Интернете по адресу www.mrc-cpe.cam.ac.uk/vbase), а также в Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No 91-3242; Tomlinson, I.M., et al. (1992) “The Repertoire of Human Germline VH Sequences Reveals about Fifty Groups of VH Segments with Different Hypervariable Loops”. J. Mol. Biol., 227:776-798; и Cox, J. P.L. et al. (1994) “A Directory of Human Germ-line VH Segments Reveals a Strong Bias in their Usage”. Eur. J. Immunol., 24:827-836; причем содержание каждого из указанных источников непосредственно включено в настоящую заявку при помощи ссылки.
Предпочтительными каркасными последовательностями для применения в антителах по настоящему изобретению являются те последовательности, которые структурно близки к каркасным последовательностям, имеющимся в выбранных антителах по настоящему изобретению, например близки к каркасным последовательностям VH 4-34 и VL L18, имеющимся в моноклональных антителах 3F11 и 4G5, или к каркасным последовательностям VH 5-51 и VL A27, имеющимся в моноклональных антителах 11E2 и 9D4. Последовательности VH CDR1, 2 и 3 SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 и 12, а также последовательности VL CDR1, 2 и 3 SEQ ID NO: 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23 и 24 могут быть пересажены в каркасные области, которые имеют те же последовательности, которые были обнаружены в гене иммуноглобулина зародышевой линии, из которого образуется каркасная последовательность, или последовательности CDR могут быть пересажены в каркасные области, которые содержат одну или большее число мутаций по сравнению с последовательностями зародышевых линий. Например, было обнаружено, что в некоторых случаях выгодно подвергнуть мутации остатки в каркасных областях для сохранения или улучшения антиген-связывающей способности антитела (см., например, патенты США №№ 5530101; 5585089; 5693762 и 6180370, выданные Queen и соавторам).
Другой тип модификации вариабельной области заключается в мутации аминокислотных остатков в областях CDR1, CDR2 и/или CDR3 VH и/или VL, чтобы таким образом улучшить одно или несколько связывающих свойств (например, сродство) представляющего интерес антитела. Для введения мутации (мутаций) могут осуществляться сайт-направленный мутагенез или PCR-опосредованный мутагенез, причем влияние на связывание антитела или другие интересующие функциональные свойства может быть оценено в анализах in vitro или in vivo, как описано в настоящей заявке и показано в примерах. Предпочтительно (как обсуждалось выше) вводятся консервативные модификации. Мутации могут представлять собой замещение, добавление и делецию аминокислот, но предпочтительно они являются замещениями. Кроме того, как правило, в области CDR изменяют не более пяти остатков.
Соответственно в другом варианте осуществления в изобретении разработаны выделенные анти-IFNAR-1 моноклональные антитела или их антиген-связывающие участки, включающие вариабельные области тяжелых цепей, содержащие: (a) область VH CDR1, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1, 2, 3 и 4, или аминокислотную последовательность, в которой имеются одно, два, три, четыре или пять замещений, делеций или добавлений аминокислот по сравнению с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1, 2, 3 и 4; (b) область VH CDR2, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, 6, 7 и 8, или аминокислотную последовательность, в которой имеются одно, два, три, четыре или пять замещений, делеций или добавлений аминокислот по сравнению с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 5, 6, 7 и 8; (c) область VH CDR3, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 9, 10, 11 и 12, или аминокислотную последовательность, в которой имеются одно, два, три, четыре или пять замещений, делеций или добавлений аминокислот по сравнению с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 9, 10, 11 и 12; (d) область VL CDR1, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 13, 14, 15 и 16, или аминокислотную последовательность, в которой имеются одно, два, три, четыре или пять замещений, делеций или добавлений аминокислот по сравнению с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 13, 14, 15 и 16; (e) область VL CDR2, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 17, 18, 19 и 20, или аминокислотную последовательность, в которой имеются одно, два, три, четыре или пять замещений, делеций или добавлений аминокислот по сравнению с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 17, 18, 19 и 20; и (f) область VL CDR3, включающую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21, 22, 23 и 24, или аминокислотную последовательность, в которой имеются одно, два, три, четыре или пять замещений, делеций или добавлений аминокислот по сравнению с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 17, 18, 19 и 20.
Сконструированные антитела по настоящему изобретению включают такие антитела, в которых были осуществлены модификации в каркасных остатках в VH и/или VL, например, для улучшения свойств антитела. Как правило, такие каркасные модификации осуществляются для уменьшения иммуногенности антитела. Например, один из подходов заключается в «обратной мутации» одного или нескольких каркасных остатков к соответствующей последовательности зародышевой линии. Более конкретно, антитело, которое подверглось соматической мутации, может содержать в каркасной области остатки, отличающиеся от последовательности зародышевой линии, из которой получено антитело. Такие остатки могут быть идентифицированы путем сравнения каркасных последовательностей антитела с последовательностями зародышевых линий, из которых получено антитело. Например, для 3F11 аминокислотный остаток №43 (в FR2) VH представляет собой треонин, тогда как этот же остаток в соответствующей последовательности зародышевой линии VH 4-34 представляет собой аланин (см. фиг.5). Для возвращения последовательности этой каркасной области к ее зародышевой конфигурации, соматические мутации могут подвергаться «обратной мутации» к последовательности зародышевой линии, с помощью, например, сайт-направленного мутагенеза или PCR-опосредованного мутагенеза (например, остаток 43 VH 3F11 может быть превращен из треонина в аланин путем «обратной мутации»). В качестве другого примера, аминокислотный остаток №81 VH антитела 4G5 (в FR3) представляет собой аспарагин, тогда как этот же остаток в соответствующей последовательности зародышевой линии VH 4-34 представляет собой лизин (см. фиг.6). Для возвращения этих последовательностей каркасной области к их зародышевой конфигурации, соматические мутации могут подвергаться «обратной мутации» из аспарагина в лизин. В качестве еще одного примера, для антител 11E2 и 9D4 аминокислотный остаток №28 (в FR1) VH представляет собой изолейцин, тогда как этот же остаток в соответствующей последовательности зародышевой линии VH 5-51 представляет собой серин (см. фиг.7). Для возвращения этих последовательностей каркасной области к их зародышевой конфигурации, соматические мутации могут подвергаться «обратной мутации» из изолейцина в серин. Предполагается, что такие подвергшиеся «обратной мутации» антитела также включены в объем настоящего изобретения.
Другой тип каркасных модификаций включает один или несколько мутирующих остатков в каркасной области или даже в одной или нескольких областях CDR, для устранения T-клеточных эпитопов, чтобы тем самым уменьшить потенциальную иммуногенность антитела. Этот подход также называют «деиммунизацией» и он более подробно описан в патентной публикации США №20030153043 Carr и соавторами.
В качестве дополнения или альтернативы модификациям, произведенным в каркасных или CDR областях, антитела по настоящему изобретению могут быть сконструированы так, чтобы включать модификации в области Fc, как правило, для изменения одного или нескольких функциональных свойств антитела, как например времени полужизни в сыворотке, фиксации комплемента, связывания рецептора Fc и/или антиген-зависимой клеточной цитотоксичности. Кроме того, антитело по настоящему изобретению может быть химически модифицировано (например, к антителу может быть присоединен один или несколько химических фрагментов) или подвергнуться модификации для изменения его гликозилирования, т.е. вновь для изменения одного или нескольких функциональных свойств антитела. Каждый из этих вариантов осуществления более подробно описан ниже по тексту. Нумерация остатков в области Fc соответствует EU индексу Kabat.
В одном из вариантов осуществления шарнирную область CH1 изменяют таким образом, что в этой области изменяется количество остатков цистеина, например увеличивается или уменьшается. Этот подход дополнительно описан в патенте США № 5677425 Bodmer и соавторами. Число цистеиновых остатков в шарнирной области CH1 изменяют, например, для облегчения сборки легкой и тяжелой цепей или для увеличения или уменьшения стабильности антитела.
В другом варианте осуществления шарнирную область Fc антитела подвергают мутации для уменьшения биологического времени полужизни антитела. Более конкретно, одну или несколько мутаций аминокислот вводят в пограничную область доменов CH2-CH3 шарнирного фрагмента Fc, так что антитело приобретает ухудшенное связывание белка A Staphylococcyl (SpA), по сравнению со связыванием природного Fc-шарнирного домена SpA. Этот подход более подробно описан в патенте США № 6165745 Ward и соавторами.
В другом варианте осуществления антитело модифицируют для увеличения времени его биологической полужизни. Возможны различные подходы. Например, могут быть введены одна или несколько из следующих мутаций: T252L, T254S, T256F, как описано в патенте США №6277375, выданном Ward. С другой стороны, для увеличения времени биологической полужизни антитело может быть подвергнуто изменениям в областях CH1 или CL для включения эпитопа, связывающего восстанавливающий рецептор, взятого из двух петель домена CH2 области Fc IgG, как описано в патентах США №№ 5869046 и 6121022 Presta и соавторами.
В еще одном варианте осуществления область Fc изменяют путем замены хотя бы одного аминокислотного остатка другим аминокислотным остатком для изменения эффекторной функции (функций) антитела. Например, одна или несколько аминокислот, выбранных из аминокислотных остатков 234, 235, 236, 237, 297, 318, 320 и 322, могут быть заменены другими аминокислотными остатками, такими, что антитело приобретает измененное сродство к эффекторному лиганду, но сохраняет антиген-связывающую способность исходного антитела. Эффекторным лигандом, к которому изменяется сродство, может быть, например, рецептор Fc, компонента комплемента C1. Этот подход более подробно описан в патентах США №№ 5624821 и 5648260 Winter и соавторами.
В другом примере одна или несколько аминокислот, выбранных из аминокислотных остатков 329, 331 и 322, могут быть заменены другим аминокислотным остатком, так что изменяется связывание антитела с C1q и/или снижается или ликвидируется комплемент-зависимая цитотоксичность (CDC). Этот подход более подробно описан в патенте США № 6194551 Idusogie и соавторами.
В еще одном примере изменяют один или несколько аминокислотных остатков в положениях 231 и 239, изменяя тем самым способность антитела останавливать комплемент. Этот подход дополнительно описан в публикации PCT WO 94/29351 Bodmer и соавторами.
В еще одном примере область Fc модифицируют для увеличения способности антитела опосредовать зависимую от него клеточную цитотоксичность (ADCC) и/или увеличивать сродство антитела к рецептору Fcγ, за счет модификации одного или нескольких аминокислотных остатков в следующих положениях: 238, 239, 248, 249, 252, 254, 255, 256, 258, 265, 267, 268, 269, 270, 272, 276, 278, 280, 283, 285, 286, 289, 290, 292, 293, 294, 295, 296, 298, 301, 303, 305, 307, 309, 312, 315, 320, 322, 324, 326, 327, 329, 330, 331, 333, 334, 335, 337, 338, 340, 360, 373, 376, 378, 382, 388, 389, 398, 414, 416, 419, 430, 434, 435, 437, 438 или 439. Этот подход дополнительно описан в публикации PCT WO 00/42072 Presta. Кроме того, было осуществлено картирование сайтов связывания IgG1 человека для FcγR1, FcγRII, FcγRIII и FcRn, и были описаны варианты с улучшенным связыванием (cм. Shields, R.L. et al. (2001) J.Biol.Chem. 276:6591-6604). Было показано, что определенные мутации в положениях 256, 290, 298, 333, 334 и 339 улучшают связывание с FcγRIII. Кроме этого было показано, что следующие комбинированные продукты мутаций улучшают связывание FcγRIII: T256A/S298A, S298A/E333A, S298A/K224A и S298A/E333A/K334A.
В еще одном варианте осуществления модификации подвергается гликозилирование антитела. Например, может быть получено дегликозилированное антитело (т.е. антитело, не имеющее гликозилирования). Гликозилирование может быть подвергнуто изменениям, например, для увеличения сродства антитела к антигену. Такие модификации углеводного состава могут осуществляться, например, изменением одного или нескольких сайтов гликозилирования в последовательности антитела. Например, может быть проведено замещение одного или нескольких аминокислотных остатков, которое приводит к уничтожению одного или нескольких сайтов гликозилирования каркаса вариабельных областей, чтобы тем самым устранить гликозилирование по этому сайту. Такое дегликозилирование может увеличивать сродство антитела к антигену. Подобный подход описан с дополнительными подробностями в патентах США №№ 5714350 и 6350861 Co и соавторами.
В качестве дополнения или альтернативы, может быть получено такое антитело, которое имеет измененный тип гликозилирования, как например гипофукозилированное антитело, имеющее уменьшенное количество фукозильных остатков, или антитело, имеющее увеличенное количество биссекторных структур GlcNac. Было показано, что такие измененные модели гликозилирования увеличивают ADCC способность антител. Указанные углеводные модификации могут осуществляться, например, путем экспрессии антитела в клетке-хозяине с измененным комплексом механизмов гликозилирования. Клетки с измененным комплексом механизмов гликозилирования описаны в технике и могут применяться в качестве клеток-хозяев, в которых осуществляется экспрессия рекомбинантных антител по настоящему изобретению, для получения тем самым антитела с измененным гликозилированием. Например, в EP 1176195 Hanai и соавторами описана клеточная линия с функционально нарушенным геном FUT8, который кодирует фукозилтрансферазу, так что антитела, экспрессируемые в такой клеточной линии, выявляют гипофукозилирование. В публикации PCT WO 03/035835 Presta описывает вариант клеточной линии CHO, а именно клетки Lec 13, с пониженной способностью присоединять фукозу к Asn(297)-связанным углеводам, также приводящую к гипофукозилированию антител, экспрессируемых в этой клетке-хозяине (см. также Shields, R.L. et al. (2002) J.Biol.Chem. 277:26733-26740). В публикации PCT WO 99/54342 Umana и соавторы описывают клеточные линии, созданные для экспрессии гликопротеин-модифицирующих гликозилтрансфераз (например, бета(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII)), так что антитела, экспрессируемые в разработанных клеточных линиях, выявляют повышенное количество биссекторных структур GlcNac, что приводит к возросшей ADCC активности антител (см. также Umana et al. (1999) Nat. Biotech. 17:176-180).
Другой модификацией антител по настоящей заявке, которая рассматривается в настоящем изобретении, является пэгилирование. Антитело может быть пэгилировано, например, для увеличения его биологического (например, в сыворотке) времени полужизни. Для осуществления пэгилирования антитела это антитело или его фрагмент, как правило, вводят во взаимодействие с полиэтиленгликолем (ПЭГ), как например реакционноспособным сложным эфиром или альдегидным производным ПЭГ, в условиях, при которых одна или несколько групп ПЭГ присоединяются к антителу или его фрагменту. Предпочтительно пэгилирование выполняют с помощью реакции ацилирования или реакции алкилирования с реакционноспособной молекулой ПЭГ (или аналогичного реакционноспособного водорастворимого полимера). В настоящем описании термин «полиэтиленгликоль» служит для обозначения любых форм ПЭГ, которые применялись для получения производных других белков, таких как моно(C1-C10)алкокси или арилокси полиэтиленгликолей или полиэтиленгликольмалеимид. В некоторых вариантах осуществления антитело, которое предполагается пэгилировать, представляет собой дегликозилированное антитело. Способы пэгилирования белков известны в технике, и они могут быть применены к антителам по настоящему изобретению. См., например, EP 0154316 Nishimura et al. и EP 0401384 Ishikawa et al.
Способы конструирования антител
Таким образом, в другом аспекте настоящего изобретения структурные особенности анти-IFNAR-1 антител по настоящему изобретению, например 3F11, 4G5, 11E2 и 9D4, используются для создания структурно родственных анти-IFNAR-1 антител, которые сохраняют как минимум одно функциональное свойство антител по настоящему изобретению, как например связывание с IFNAR-1. Например, одна или несколько областей CDR 3F11, 4G5, 11E2 или 9D4 или продуктов их мутаций могут быть рекомбинантно объединены с известными каркасными областями и/или другими CDR для создания дополнительных рекомбинантно-сконструированных анти-IFNAR-1 антител по настоящему изобретению, как обсуждалось выше. Другие типы модификаций включают модификации, описанные в предыдущем разделе. Исходным материалом для способа конструирования являются одна или несколько последовательностей из числа VH и/или VL, разработанных в настоящем изобретении, или одна или несколько из областей CDR этих последовательностей. Для создания сконструированного антитела нет необходимости действительно получать (например, экспрессировать в виде белка) антитело, имеющее одну или несколько из числа последовательностей VH и/или VL, разработанных в настоящем изобретении, или одну или несколько из их областей CDR. Скорее, информация, содержащаяся в последовательности (последовательностях), используется в качестве исходного материала для создания «второго поколения» последовательности (последовательностей), полученных из исходной последовательности (последовательностей), и затем получают «второе поколение» последовательности (последовательностей) и экспрессируют в виде белка.
Соответственно в еще одном варианте осуществления изобретение относится к способу получения анти-IFNAR-1 антитела, включающему:
(a) получение: (i) последовательности вариабельной области тяжелой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO: 1, 2, 3 и 4, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NO: 5, 6, 7 и 8, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NO: 9, 10, 11 и 12; и (ii) последовательности вариабельной области легкой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO: 13, 14, 15 и 16, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NO: 17, 18, 19 и 20, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NO: 21, 22, 23 и 24;
(b) изменение хотя бы одного аминокислотного остатка в первой последовательности антитела и/или второй последовательности антитела для создания хотя бы одной измененной последовательности антитела; и
(c) получение измененной последовательности антитела; и
(d) экспрессию измененной последовательности антитела в виде белка.
Для получения и экспрессии измененной последовательности антитела могут применяться стандартные методики молекулярной биологии.
Предпочтительно антитело, закодированное измененной последовательностью (последовательностями), представляет такое антитело, которое сохраняет одно, некоторые или все функциональные свойства описанных в настоящей заявке анти-IFNAR-1 антител, причем указанные функциональные свойства включают, не ограничиваясь перечисленным:
(i) связывание с IFNAR-1;
(ii) ингибирование связывания интерферонов типа I с IFNAR-1;
(iii) связывание с живыми клетками, экспрессирующими IFNAR-1 человека;
(iv) связывание с IFNAR-1 человека с KD, равной 10-8 M или менее (например, 10-9 М или 10-10 М или менее);
(v) связывание с уникальным эпитопом на IFNAR-1 (для исключения возможности того, что моноклональные антитела с комплементарными видами активности при использовании их в комбинации принимали бы участие в конкуренции за связывание с одним и тем же эпитопом).
Функциональные свойства измененных антител можно оценить при использовании стандартных анализов, известных в технике и/или описанных в настоящей заявке. Например, способность антитела связываться с IFNAR-1 может быть определена с помощью стандартных анализов связывания, таких, которые описаны в примерах (например, ELISA).
В некоторых вариантах осуществления способов конструирования антител по настоящему изобретению мутации могут вводиться случайно или селективно во всю последовательность, кодирующую анти-IFNAR-1 антитело, или в ее часть (например, последовательность, кодирующую 3F11, 4G5, 11E2 или 9D4), и полученные модифицированные анти-IFNAR-1 антитела могут подвергаться скринингу по признаку активности связывания и/или других функциональных свойств, как описано в настоящей заявке. Мутационные методики были описаны в технике. Например, в опубликованной согласно PCT WO 02/092780 Short описывает способы создания и скрининга мутаций антител с применением насыщающего мутагенеза, сборки синтетическим сшиванием или их комбинации. С другой стороны, в публикации PCT WO 03/074679 Lazar и соавторами описаны способы, использующие вычислительные методики скрининга для оптимизации физиохимических свойств антител.
Молекулы нуклеиновых кислот, кодирующие антитела по настоящему изобретению
Другой аспект настоящего изобретения имеет отношение к молекулам нуклеиновых кислот, которые кодируют антитела по настоящему изобретению. Нуклеиновые кислоты могут присутствовать в целых клетках, или клеточном лизате, или в частично очищенной, или в основном чистой форме. Нуклеиновая кислота является «выделеннной» или «приведенной в практически чистое состояние», когда она очищена от других клеточных компонентов или других загрязняющих примесей, например других клеточных нуклеиновых кислот или белков, с помощью стандартных методик, в т.ч. обработки щелочью/SDS, расслоения в CsCl, колоночной хроматографии, электрофореза агарозного геля и других, хорошо известных в технике. См. F.Ausubel, et al., ed. (1987) Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York. Нуклеиновые кислоты по настоящему изобретению могут представлять собой, например, ДНК или РНК и могут содержать или не содержать интронные последовательности. В предпочтительном варианте осуществления нуклеиновая кислота представляет собой молекулу кДНК.
Нуклеиновые кислоты по настоящему изобретению могут быть получены при использовании стандартных методик молекулярной биологии. Для антител, экспрессируемых гибридомами (например, гибридомами, полученными из трансгенной мыши, несущей гены иммуноглобулина человека, как описано далее по тексту), созданные гибридомой кДНК, кодирующие легкие и тяжелые цепи антитела, могут быть получены стандартными методиками амплификации PCR или клонирования кДНК. Для антител, полученных из библиотеки генов иммуноглобулина (например, с применением методики фагового дисплея), нуклеиновая кислота, кодирующая антитело, может быть восстановлена из библиотеки.
Предпочтительными молекулами нуклеиновых кислот по настоящему изобретению являются молекулы, кодирующие последовательности VH и VL моноклональных антител 3F11, 4G5, 11E2 и 9D4. Последовательности ДНК, кодирующие последовательности VH и VL 3F11, показаны на SEQ ID NO: 33 и 37 соответственно. Последовательности ДНК, кодирующие последовательности VH и VL 4G5, показаны на SEQ ID NO: 34 и 38 соответственно. Последовательности ДНК, кодирующие последовательности VH и VL 11E2, показаны на SEQ ID NO: 35 и 39 соответственно. Последовательности ДНК, кодирующие последовательности VH и VL 9D4, показаны на SEQ ID NO: 36 и 40 соответственно.
Если фрагменты ДНК, кодирующие сегменты VH и VL, уже когда-либо были получены, эти фрагменты ДНК в дальнейшем можно обрабатывать с помощью стандартных методик работы с рекомбинантными ДНК, например превращать гены вариабельной области в гены полноразмерной цепи антитела, гены фрагмента Fab или ген scFv. В ходе этих превращений фрагмент ДНК, кодирующий VL или VH, оказывается операционно-связанным с другим фрагментом ДНК, кодирующим другую белковую последовательность, такую как константная область или гибкий линкер антитела. Термин «операционно-связанный» в данном контексте используется для обозначения того, что два фрагмента ДНК объединены таким образом, что аминокислотные последовательности, кодируемые этими двумя фрагментами ДНК, сохраняются в рамке считывания.
Выделенную ДНК, кодирующую область VH, можно превратить в полноразмерный ген тяжелой цепи с помощью операционного связывания кодирующей VH ДНК с другой молекулой ДНК, кодирующей константные области тяжелой цепи (CH1, CH2 и CH3). Последовательности генов константных областей тяжелых цепей человека известны в технике (см., например, Kabat, E.A., et al. (1991) Sequenced of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No 91-3242), и фрагменты ДНК, соответствующие этим областям, могут быть получены стандартной PCR-амплификацией. Константные области тяжелой цепи могут быть константными областями IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно являются константными областями IgG1 или IgG4. Для гена фрагмента Fab тяжелой цепи, кодирующая VH ДНК может быть операционно связана с другой молекулой ДНК, кодирующей только константную область CH1 тяжелой цепи.
Выделенная ДНК, кодирующая область VL, может быть превращена в полноразмерный ген легкой цепи (а также в ген Fab легкой цепи) с помощью операционного связывания VL-кодирующей молекулы ДНК с другой молекулой ДНК, кодирующей константную область легкой цепи, т.е. CL. Последовательности генов константных областей легких цепей человека известны в технике (см., например, Kabat, E.A., et al. (1991) Sequenced of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No 91-3242), и фрагменты ДНК, соответствующие этим областям, могут быть получены стандартной PCR-амплификацией. Константные области легких цепей могут быть константными областями каппа или лямбда, но наиболее предпочтительно константной областью каппа.
Для создания гена scFv, VH и VL-кодирующие фрагменты ДНК операционно связывают с другим фрагментом, кодирующим гибкий линкер, например кодирующим аминокислотную последовательность (Gly4-Ser)3, так что последовательности VH и VL могут быть экспрессированы в виде непрерывного одноцепочечного белка, с областями VH и VL, связанными гибким линкером (см., например, Bird et al., (1988) Science, 242:423-426; Huston et al. (1988) Proc.Natl.Acad.Sci USA, 85:5879-5883; McCafferty et al., (1990) Nature, 348:552-554).
Получение моноклональных антител по настоящему изобретению
Моноклональные антитела (mAb) по настоящему изобретению могут быть получены разнообразными способами, включая традиционную совокупность методик работы с моноклональными антителами, например стандартную методику гибридизации соматических клеток, разработанную Kohler и Milstein (1975) Nature, 256:495. Хотя предпочтительными являются методики гибридизации соматических клеток, в принципе, могут применяться другие способы получения моноклональных антител, например вирусная или онкогенная трансформация B-лимфоцитов.
Предпочтительной животной системой для получения гибридом является мышиная система. Методика получения гибридом в мышах является очень хорошо разработанной. В технике известны протоколы иммунизации и методики выделения иммунизированных спленоцитов для слияния. Также известны клетки-партнеры при слиянии (например, клетки миеломы мыши) и методики слияния.
Химерные или гуманизированные антитела по настоящему изобретению могут быть получены на основе последовательностей моноклональных антител мыши, полученных, как описано выше. ДНК, кодирующая тяжелые и легкие цепи иммуноглобулинов, может быть получена с применением стандартных методик молекулярной биологии из соответствующей гибридомы мыши и сконструирована таким образом, что она содержит не мышиные (например, человеческие) иммуноглобулиновые последовательности. Например, для создания химерного антитела вариабельные области мыши могут быть связаны с константными областями человека с применением известных в технике способов (см., например, патент США № 4816567, выданный Cabilly и соавторам). Для создания гуманизированного антитела области CDR мыши могут быть внедрены в каркас человека с применением способов, известных в технике (см., например, патент США № 5225539, выданный Winter, и патенты США №№ 5530101; 5585089; 5693762 и 6180370, выданные Queen и соавторам).
В предпочтительном варианте осуществления антитела по настоящему изобретению являются моноклональными антителами человека. Такие моноклональные антитела человека, направленные против IFNAR-1, могут быть получены с применением трансгенных или трансхромосомных мышей, имеющих части человеческой, а не иммунной системы мыши. Эти трансгенные и трансхромосомные мыши включают мышей, которые в настоящей заявке называются мышами HuMAb и мышами KM соответственно, и, вместе взятые, именуются «мышами с Ig человека».
Мышь HuMAb® (Medarex, Inc.) содержит минилокусы иммуноглобулинового гена человека, которые кодируют неперегруппированные последовательности тяжелой (µ и γ) и κ легкой цепей иммуноглобулина человека, совместно с нацеленными мутациями, которые инактивируют эндогенные локусы µ и κ цепей (см., например, Lonberg, et al. (1994) Nature, 368 (6474):856-859). Соответственно эта мышь демонстрирует уменьшенную экспрессию мышиного IgM или κ, и в ответ на иммунизацию, введенные трансгены тяжелой и легкой цепей человека испытывают переключение классов и соматическую мутацию, генерируя моноклональный IgGκ человека с высоким сродством (Lonberg, N. et al. (1994), см. выше; обзор в Lonberg, N. (1994) Handbook of Experimental Pharmacology, 113: 49-101; Lonberg, N. and Huszar, D. (1995) Intern.Rev. Immunol. 13:65-93, а также Harding, F. and Lonberg, N. (1995) Ann.N.Y.Acad.Sci. 764:536-546). Получение и использование мышей HuMAb и модификации генома, осуществляемые такой мышью, более подробно описаны в Taylor, L. et al. (1992) Nucleic Acids Research, 20:6287-6295; Chen, J. et al. (1993) International Immunology, 5:647-656; Tuaillon et al. (1993) Proc. Natl. Acad. Sci. USA, 90:3720-3724: Choi et al. (1993) Nature Genetics 4:117-123; Chen, J. et al. (1993) EMBO J. 12:821-830; Tuaillon et al. (1994) J. Immunol, 152: 2912-2920; Taylor, L. et al. (1994) International Immunology, 6:579-591; и Fishwild, D. et al. (1996) Nature Biotecnology, 14:845-851, причем содержание всех перечисленных работ непосредственно включено в настоящую заявку во всей полноте с помощью ссылки. Дополнительно см. патенты США №№ 5545806; 5569825; 5625126; 5633425; 5789650; 5877397; 5661016; 5814318; 5874299 и 5770429; все выданы Lonberg и Kay; патент США №5545807, выданный Surani и соавторам; публикации PCT №№ WO92/03918, WO93/12227, WO94/25585, WO97/13852, WO98/24884 и WO99/45962 все Lonberg и Kay; а также публикацию PCT № WO 01/14424 Korman и соавторов.
В другом варианте осуществления антитела человека по настоящему изобретению могут быть получены с применением мышей, которые являются носителями иммуноглобулиновых последовательностей человека на трансгенах или трансхромосомах, как например мышей, являющихся носителями трансгена тяжелой цепи человека и трансхромосомы легкой цепи человека. Такие мыши, называемые в настоящем описании «мыши KM», подробно описаны в публикации PCT № WO 02/43478 Ishida и соавторами.
В технике известны и другие альтернативные трансгенные животные системы, экспрессирующие гены иммуноглобулина человека, причем эти системы могут применяться для получения анти-IFNAR-1 антител по настоящему изобретению. Например, может применяться альтернативная трансгенная система, называемая Xenomouse (Abgenix, Inc.); такие мыши описаны, например, в патентах США №№ 5939598; 6075181; 6114598; 6150584 и 6162963, выданных Kucherlapati и соавторам.
Кроме того, в технике имеются альтернативные трансхромосомные животные системы, экспрессирующие иммуноглобулиновые гены человека, и их можно применять для выработки анти-IFNAR-1 антител по настоящему изобретению. Например, могут применяться мыши, являющиеся носителями как трансхромосомы тяжелой цепи человека, так и трансхромосомы легкой цепи человека, называемые «TC мышами»; такие мыши описаны в Tomizuka et al. (2000) Proc.Natl.Acad.Sci. USA, 97:722-727. Кроме того, в технике были описаны коровы, являющиеся носителями трансхромосом тяжелой и легкой цепей человека (Kuroiwa et al. (2002) Nature Biotechnology, 20:889-894), причем их можно применять для выработки анти-IFNAR-1 антител по настоящему изобретению.
Моноклональные антитела человека по настоящему изобретению также могут быть получены с применением методик фагового дисплея для скрининга библиотек иммуноглобулиновых генов человека. В технике разработаны такие методики фагового дисплея для выделенных антител человека. См., например, патенты США №№ 5223409; 5403484; и 5571698, выданные Ladner и соавторам; патенты США №№ 5427908 и 5580717, выданные Dower и соавторам; патенты США №№ 5969108 и 6172197, выданные McCafferty и соавторам; и патенты США №№ 5885793; 6521404; 6544731; 6555313; 6582915 и 6593081, выданные Griffiths и соавторам.
Моноклональные антитела человека по настоящему изобретению также могут быть получены с применением мышей SCID, в которых воспроизведены иммунные клетки человека, так что при иммунизации генерируется иммунный ответ с выработкой антител человека. Такие мыши описаны, например, в патентах США №№ 5476996 и 5698767, выданных Wilson и соавторам.
Иммунизация мышей с Ig человека
Если для выработки антител человека по настоящему изобретению используют мышей с Ig человека, такие мыши могут быть иммунизированы очищенными или обогащенными препаратами антигена IFNAR-1 и/или клетками, экспрессирующими IFNAR-1, как описано в Lonberg, N. et al. (1994) Nature, 368 (6474):856-859; Fishwild, D. et al (1996) Nature Biotechnology, 14: 845-851; и публикациях PCT WO 98/24884 и WO 01/14424. Предпочтительно при первой инфузии мыши должны иметь возраст 6-16 недель. Например, для иммунизации мышей с Ig человека может быть применено интраперитонеальное введение очищенного или обогащенного препарата (5-50 мкг) антигена IFNAR-1. В случае если иммунизация с применением очищенного или обогащенного препарата антигена IFNAR-1 не приводит к образованию антител, мышь также может быть иммунизирована клетками, экспрессирующими IFNAR-1, например линией T-клеток человека, с целью стимулирования иммунных ответов.
В примере 1 далее по тексту подробно описана методика генерирования полностью моноклональных антител человека IFNAR-1. Обобщенный опыт, относящийся к различным антигенам, показал, что трансгенные мыши дают иммунную реакцию, при первоначальной интраперитонеальной (IP) иммунизации антигеном в полном адъюванте Фрейнда, с последующими еженедельными иммунизациями (общим числом до 6) антигеном в неполном адъюванте Фрейнда. Однако было найдено, что другие адъюванты, кроме адъюванта Фрейнда, также являются эффективными. Кроме этого было найдено, что целые клетки в отсутствие адъюванта обладают высокой иммуногенностью. За иммунным ответом можно наблюдать по ходу выполнения протокола иммунизации с помощью образцов плазмы, получаемых при ретроорбитальных кровотечениях. Эта плазма может быть подвергнута скринингу с помощью ELISA (как описано ниже), и мыши с достаточными титрами анти-IFNAR-1 иммуноглобулина человека могут быть использованы для слияний. Воздействие на мышь может быть усилено внутривенным введением антигена за 3 дня до умерщвления и удаления селезенки. Ожидается, что для каждой иммунизации может быть необходимо выполнить 2-3 слияния. Для каждого антигена, как правило, иммунизируют от 6 до 24 мышей. Обычно используют обе линии HCo7 и HCo12. Кроме этого оба трансгена HCo7 и HCo12 могут быть совместно воспроизведены в одной мыши, имеющей два различных трансгена тяжелой цепи человека (HCo7/HCo12).
Генерация гибридом, вырабатывающих моноклональные антитела человека по настоящему изобретению
Для генерации гибридом, вырабатывающих моноклональные антитела человека по настоящему изобретению, спленоциты или клетки лимфатических узлов иммунизированной мыши могут быть выделены и слиты с подходящими клетками иммортализованной линии, как например линии миеломы мыши. Полученные гибридомы могут быть подвергнуты скринингу по критерию выработки антиген-специфичных антител. Например, суспензии одиночных клеток лимфоцитов селезенки, полученных из иммунизированных мышей, могут быть слиты в количественном соотношении один к шести P3X63-Ag8.653 с клетками несекретирующей миеломы мыши (ATCC, CRL 1580) c 50% ПЭГ. Клетки размещают на планшетах в количестве примерно 2·105 на плоскодонный микротитровальный планшет, с последующим двухнедельным инкубированием в селективной среде, содержащей 20% фетальной сыворотки, 18% кондиционированной среды «653», 5% Origen (IGEN), 4 мМ L-глутамина, 1 мМ пирувата натрия, 5 мМ HEPES, 0,055 мМ 2-меркаптоэтанола, 50 ед/мл пенициллина, 50 мг/мл стрептомицина, 50 мг/мл гентамицина и 1X HAT (Sigma; HAT добавляют 24 часа после слияния). По прошествии приблизительно двух недель клетки могут быть культивированы в среде, в которой HAT заменен на HT. Затем индивидуальные лунки можно исследовать способом ELISA на моноклональные антитела IgM и IgG человека. Если имеет место экстенсивный рост гибридомы, среду обычно можно исследовать через 10-14 дней. Гибридомы, секретирующие антитела, могут быть повторно размещены на планшетах, вновь подвергнуты скринингу, и в случае повторного положительного результата в отношении IgG человека, моноклональные антитела могут быть субклонированы, по крайней мере, дважды с помощью серийных разведений. Затем стабильные субклоны могут быть культивированы in vitro с целью генерации небольших количеств антитела в тканевой культурной среде для идентификации.
Для очистки моноклональных антител человека отобранные гибридомы могут быть выращены в двухлитровых роллерных колбах для очистки моноклональных антител. Перед осуществлением аффинной хроматографии с протеин А-сефарозой (Pharmacia, Piscataway, N.J.) супернатанты можно подвергнуть фильтрованию и концентрированию. Для гарантии чистоты элюированный IgG может быть проверен гель-электрофорезом и высокоэффективной жидкостной хроматографией. Буферный раствор можно заменить на PBS и концентрацию можно определить по OD280 (оптической плотности) с использованием коэффициента экстинкции 1,43. Полученные моноклональные антитела можно разделить на аликвоты и хранить при -80°C.
Генерация трансфектом, вырабатывающих моноклональные антитела человека по настоящему изобретению
Антитела человека по настоящему изобретению также могут быть получены в клетке-хозяине, называемой трансфектомой, с применением, например, хорошо известных в технике методик на основе комбинантной или рекомбинантной ДНК, а также способов трансфекции генов (например, Morrison, S. (1985) Science, 229:1202).
Например, для того чтобы экспрессировать антитело или его фрагмент, с помощью стандартных методик молекулярной биологии (например, PCR-амплификации или клонирования кДНК с использованием гибридомы, которая экспрессирует представляющее интерес антитело) могут быть получены молекулы ДНК, кодирующие полноразмерные легкие и тяжелые цепи или их части, и эти молекулы ДНК могут быть включены в векторы экспрессии, так чтобы гены были операционно связаны с транскрипционными и трансляционными управляющими последовательностями. В данном контексте термин «операционно-связанные» предназначен для обозначения того, что ген антитела встроен в вектор таким образом, что транскрипционные и трансляционные управляющие последовательности в векторе исполняют предназначенные им функции управления транскрипцией и трансляцией генов антитела. Вектор экспрессии и управляющие экспрессией последовательности выбирают так, чтобы они были совместимы с применяемой для экспрессии клеткой-хозяином. Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть включены в отдельные векторы, или чаще оба гена включают в один и тот же вектор экспрессии. Гены антитела включают в вектор экспрессии стандартными способами (например, сшиванием комплементарных сайтов рестрикции на фрагменте гена антитела и вектора или сшиванием «тупых концов», если сайты рестрикции отсутствуют). Вариабельные области тяжелых и легких цепей антител, описанных в настоящем изобретении, могут быть использованы для создания полноразмерных генов любых изотипов антител путем включения их в векторы экспрессии, уже кодирующие константные области тяжелой цепи и легкой цепи желаемого изотипа, так что сегмент VH операционно связан в векторе с сегментом (сегментами) CH и сегмент VL операционно связан в векторе с сегментом CL. В качестве дополнения или альтернативы рекомбинантный вектор экспрессии может кодировать сигнальный пептид, который облегчает секрецию цепи антитела из клетки-хозяина. Ген цепи антитела может быть клонирован в вектор так, чтобы сигнальный пептид был связан в рамке с аминоконцом гена цепи антитела. Сигнальный пептид может быть сигнальным пептидом иммуноглобулина или гетерологичным сигнальным пептидом (т.е. сигнальным пептидом, отличным от иммуноглобулина белка).
В дополнение к генам цепи антитела рекомбинантные векторы экспрессии по настоящему изобретению несут регуляторные последовательности, которые управляют экспрессией генов цепи антитела в клетке-хозяине. Предполагается, что термин «регуляторная последовательность» включает промоторные, энхансерные и другие элементы управления экспрессией (например, сигналы полиаденилирования), которые регулируют транскрипцию или трансляцию генов цепи антитела. Такие регуляторные последовательности описаны, например, у Goeddel (Gene Expression Technology. Methods in Enzymology 185, Academic Press, San Diego, CA (1990)). Специалист в данной области техники примет во внимание, что создание вектора экспрессии, включая выбор регуляторных последовательностей, может зависеть от таких факторов, как выбор клетки-хозяина, которую предстоит трансформировать, желательного уровня экспрессии белка и т.д. Предпочтительные регуляторные последовательности для экспрессии в клетках-хозяевах млекопитающих включают вирусные элементы, которые приводят к высоким уровням экспрессии белков в клетках млекопитающих, как например промоторы и/или энхансеры, полученные из цитомегаловируса (CMV), вируса обезьяны 40 (SV40), аденовируса (например, большой поздний промотор аденовируса (AdMLP)) и полиомы. С другой стороны, могут применяться регуляторные последовательности невирусной природы, такие как промотор убиквитина или промотор β-глобина. Кроме этого могут применяться регуляторные элементы, созданные на основе последовательностей из других источников, таких как система промоторов SRα, которая содержит последовательности из раннего промотора SV40, и длинный концевой повтор вируса T-клеточной лейкемии человека типа I (Takebe, Y. et al. (1988) Mol.Cell. Biol. 8:466-472).
Кроме генов цепи антитела и регуляторных последовательностей, рекомбинантные векторы экспрессии по настоящему изобретению могут включать дополнительные последовательности, как например последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, сайты инициации репликации) и гены селектируемого маркера. Ген селектируемого маркера облегчает выбор клеток-хозяев, в которых был введен данный вектор (см., например, патенты США №№ 4399216, 4634665 и 5179017, все Axel и соавторов). Например, как правило, ген селектируемого маркера придает клеткам-хозяевам, в которые был введен вектор, устойчивость к лекарствам, таким как G418, гигромицин или метотрексат. Предпочтительные гены селектируемого маркера включают ген дигидрофолатредуктазы (DHFR) (для применения в dhfr-клетках-хозяевах с селекцией/амплификацией метотрексата) и неоген (для селекции G418).
Для экспрессии легкой и тяжелой цепей вектор (векторы) экспрессии, кодирующие легкие и тяжелые цепи, трансфицируют в клетку-хозяина с помощью стандартных методик. Предполагается, что различные формы термина «трансфекция» охватывают широкий круг методик, обычно применяемых для введения экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например электропорация, осаждение с фосфатом кальция, трансфекция с DEAE-декстраном и т.п. Хотя теоретически возможно экспрессировать антитела по настоящему изобретению либо в прокариотических, либо в эукариотических клетках-хозяевах, экспрессия антител в эукариотических клетках, и лучше всего в клетках-хозяевах млекопитающих является наиболее предпочтительной, т.к. такие эукариотические клетки, и в частности клетки млекопитающих, с большей вероятностью, чем прокариотические клетки, способны собирать и секретировать антитела с надлежащей пространственной структурой и иммунологической активностью. Экспрессия генов антител в прокариотических клетках, как сообщалось, является неэффективной для получения высоких выходов активных антител (Boss, M.A. and Wood, C.R. (1985) Immunology Today, 6:12-13).
Предпочтительные клетки-хозяева млекопитающих для экспрессии рекомбинантных антител по настоящему изобретению включают клетки яичника китайского хомяка (клетки CHO) (включая dhfr-CHO-клетки, описанные в Urlaub and Chasin, (1980) Proc.Natl.Acad.Sci. USA, 77:4216-4220, используемые вместе с DHFR селектируемым маркером, например, как описано в R.J.Kaufman and P.A.Sharp (1982) Mol.Biol. 159:601-621), клетки миеломы NSO, клетки COS и клетки SP2. В частности, другой предпочтительной системой экспрессии для использования с клетками миеломы NSO является система экспрессии гена GS, раскрытая в WO 87/04462, WO 89/01036 и EP 338841. Когда рекомбинантные векторы экспрессии, кодирующие гены антитела, вводят в клетки-хозяева млекопитающих, выработка антител происходит при культивировании клеток-хозяев в течение периода времени, достаточного для осуществления экспрессии антител в клетках-хозяевах, более предпочтительно для секреции антител в культурную среду, в которой выращиваются клетки-хозяева. Антитела могут быть выделены из культурной среды с применением стандартных способов очистки белков.
Определение характеристик связывания антитела с антигеном
Антитела по настоящему изобретению могут быть исследованы на связывание с IFNAR-1, например, с помощью стандартного ELISA. Вкратце, микротитровальные планшеты покрывают очищенным IFNAR-1 в концентрации 0,25 мкг/мл в PBS и затем блокируют 5% альбумином бычьей сыворотки в PBS. Антитело в различных разбавлениях (например, разбавления плазмы IFNAR-1-иммунизированной мыши) добавляют в каждую лунку и инкубируют 1-2 часа при 37°C. Планшеты промывают смесью PBS/Tween и затем инкубируют с вторичным реагентом (например, в случае антител человека, с козьим античеловеческим IgG Fc-специфичным поликлональным реагентом), конъюгированным со щелочной фосфатазой, в течение 1 часа при 37°C. После промывания планшеты обрабатывают субстратом pNPP (1 мг/мл) и анализируют OD при 405-650. Предпочтительно для слияния будет использоваться мышь, продемонстрировавшая наиболее высокие титры.
Как указано выше, анализ ELISA также может использоваться для отбора гибридом, которые показывают положительную реактивность в отношении иммуногена IFNAR-1. Гибридомы, которые связываются с IFNAR-1 с высокой авидностью, подвергают субклонированию и далее определяют характеристики. Один клон из каждой гибридомы, который сохраняет реактивность исходных клеток (согласно ELISA), может быть выбран для создания 5-10 флаконов с банками клеток, хранящихся при -140°C, и для очистки антител.
Для того чтобы очистить анти-IFNAR-1 антитела, отобранные гибридомы могут быть выращены в двухлитровых роллерных колбах для очистки моноклональных антител. Супернатанты могут быть подвергнуты фильтрованию и концентрированию перед проведением аффинной хроматографии с протеин A-сефарозой (Pharmacia, Piscataway, NJ). Элюированный иммуноглобулин для гарантии чистоты может быть проверен с помощью гель-электрофореза и высокоэффективной жидкостной хроматографии. Буферный раствор может быть заменен на PBS, и концентрация может быть определена по OD280 с использованием коэффициента экстинкции 1,43. Моноклональные антитела могут быть разделены на аликвоты и сохранены при -80°C.
Для того чтобы определить, связываются ли отобранные анти-IFNAR-1 моноклональные антитела с единственными эпитопами, каждое антитело может быть биотинилировано с применением коммерчески доступных реагентов (Pierce, Rockford, IL). Исследование конкурентного связывания с использованием немеченых моноклональных антител и биотинилированых моноклональных антител может быть выполнено с применением планшетов ELISA, покрытых IFNAR-1, как описано выше. Связывание биотинилированных mAb может быть обнаружено с помощью зонда стрептококк-авидин-щелочная фосфатаза.
Для определения изотипа очищенных антител может быть осуществлен ELISA-анализ изотипа с применением реагентов, специфичных для антител конкретного изотипа. Например, для определения изотипа моноклонального антитела человека лунки микротитровальных планшетов могут быть покрыты в течение ночи при 4°C 1 мкг/мл античеловеческого иммуноглобулина. После блокирования 1% BSA содержимое планшетов вводят во взаимодействие с 1 мкг/мл или менее тестовых моноклональных антител или очищенных контрольных образцов изотипа, при комнатной температуре в течение одного или двух часов. Затем содержимое лунок может быть введено во взаимодействие либо с IgG1 человека, либо с конъюгированными зондами IgM-специфичной щелочной фосфатазы человека. Планшеты обрабатывают и анализируют, как описано выше.
Для демонстрации связывания моноклональных антител с живыми клетками, экспрессирующими IFNAR-1, может быть использована проточная цитометрия. Вкратце, линии клеток, экспрессирующие IFNAR-1 (выращенные в стандартных условиях роста), смешивают с моноклональными антителами в различных концентрациях в PBS, содержащем 0,1% BSA и 10% фетальной сыворотки телят, после чего инкубируют при 37°C в течение 1 часа. После промывания клетки вводят во взаимодействие с меченным флуоресцеином античеловеческим IgG антителом в тех же условиях, в которых осуществлялось первичное окрашивание антител. Образцы можно анализировать на приборе FACScan, используя свойства света и боковое рассеяние при возбуждении флуоресценции отдельных клеток. Альтернативный анализ с применением флуоресцентной микроскопии может быть выполнен либо дополнительно к проточной цитометрии, либо вместо нее. Клетки могут быть окрашены в полном соответствии с приведенным выше описанием и исследованы флуоресцентной микроскопией. Этот метод делает возможным визуализацию отдельных клеток, но может обладать пониженной чувствительностью в зависимости от плотности антигена.
Далее, анти-IFNAR-1 человека IgG могут быть испытаны на реакционную способность в отношении антигена IFNAR-1 с помощью вестерн-блоттинга. Вкратце, клеточные экстракты, полученные из клеток, экспрессирующих IFNAR-1, могут быть получены и подвергнуты электрофорезу в геле додецилсульфат натрия - полиакриламид. После электрофореза разделенные антигены переносят на мембраны из нитроцеллюлозы, блокируют 10% фетальной телячьей сывороткой и проводят испытание с тестируемыми моноклональными антителами. Связывание IgG человека может быть обнаружено при использовании античеловеческого IgG - щелочной фосфатазы и проявлено с помощью субстратных таблеток BCIP/NBT (Sigma Chem.Co., St. Louis, MO).
Иммуноконъюгаты
В другом аспекте настоящее изобретение включает анти-IFNAR-1 антитело или его фрагмент, конъюгированные с терапевтическим фрагментом, таким как цитотоксин, лекарственное средство (например, иммунодепрессант) или радиотоксин. Такие конъюгаты в настоящей заявке именуются «иммуноконъюгатами». Иммуноконъюгаты, которые включают один или несколько цитотоксинов, называются «иммунотоксинами». Цитотоксин или цитотоксичное средство включают любое средство, которое приносит вред клеткам (например, убивает). Примеры включают таксол, цитохалазин B, грамицидин D, этидиум бромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин, а также их аналоги и гомологи. Терапевтические средства также включают, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил, декарбазин), алкилирующие средства (например, мехлорэтамин, тиоепа хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин C и цис-дихлордиамин платина (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин, (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)), а также антимитотические средства (например, винкристин и винбластин).
Другие предпочтительные примеры терапевтических цитотоксинов, которые могут быть конъюгированы с антителами по настоящему изобретению, включают дуокармицины, калихеамицины, майтанзины и ауристатины, а также их производные. Примером конъюгата антитела с калихеамицином является коммерчески доступный препарат (MylotargTM; Wyeth-Ayerst).
Цитотоксины могут быть конъюгированы с антителами по настоящему изобретению с применением имеющейся в технике линкерной технологии. Примеры типов линкеров, которые применялись для конъюгации цитотоксина к антителу, включают, не ограничиваясь перечисленными, гидразоны, тиоэфиры, сложные эфиры, дисульфиды и пептидсодержащие линкеры. Может быть выбран линкер, который, например, чувствителен к расщеплению в лизосомальном компартменте при низких значениях pH или чувствителен к расщеплению под действием протеаз, как например протеаз, преимущественно экспрессируемых в опухолевой ткани, таких как катепсины (например, катепсины B, C, D).
Для ознакомления с более подробным обсуждением типов цитотоксинов, линкеров и способов конъюгации терапевтических средств и антител см. также Saito, G. et al. (2003) Adv. Drug Deliv. Rev. 55:199-215; Trail, P.A. et al. (2003) Cancer Immunol. Immunother. 52:328-337; Payne, G. (2003) Cancer Cell. 3:207-212; Allen, T.M. (2002) Nat. Rev. Cancer 2:750-763; Pastan, I. and Kreitman, R.J. (2002) Curr. Opin. Investig. Drugs. 3:1089-1091; Senter, P.D. and Springer, C.J. (2001) Adv. Drug. Deliv. Rev. 53:247-264.
Антитела по настоящему изобретению также могут быть конъюгированы с радиоактивными изотопами дл получения цитотоксичных радиоактивных фармацевтических препаратов, также именуемых радиоиммуноконъюгатами. Примеры радиоактивных изотопов, которые могут быть конъюгированы с антителами для диагностического и терапевтического применения, включают, не ограничиваясь перечисленными, йод131, индий111, иттрий90 и лютеций177. Способы получения радиоиммуноконъюгатов разработаны в технике. Примеры коммерчески доступных радиоиммуноконъюгатов включают ZevalinTM (IDEC Pharmaceuticals) и BexxarTM (Corixa Pharmaceuticals), причем аналогичные способы могут применяться для получения радиоиммуноконъюгатов, включающих антитела по настоящему изобретению.
Конъюгаты антител по настоящему изобретению могут применяться для модификации данной биологической реакции, причем не следует считать, что фрагменты лекарственного препарата ограничены кругом классических химических лекарственных средств. Например, лекарственный фрагмент может представлять собой белок или полипептид, обладающий желаемой биологической активностью. Такие белки могут включать, например, ферментно-активные токсины или их активные фрагменты, такие как абрин, рицин A, псевдомонадный экзотоксин или дифтерийный токсин; белок, такой как фактор некроза опухолей или интерферон-γ; или модификаторы биологической реакции, такие как, например, лимфокины, интерлейкин-1 (“IL-1”), интерлейкин-2 (“IL-2”), интерлейкин-6 (“IL-6”), гранулоцит-макрофаг колониестимулирующий фактор (“GM-CSF”), гранулоцит колониестимулирующий фактор (“G-CSF”) или другие факторы роста.
Хорошо известны методики конъюгации таких терапевтических фрагментов с антителами, см., например, Amon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Antibodies For Drug Delivery", in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe. "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", in Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera el al. (eds.), pp. 475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", in Monoclonal Antibodies For Cancer Detection And Therapy. Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), and Thorpe et al. "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-58(1982).
Биспецифичные молекулы
В другом аспекте настоящее изобретение относится к биспецифичным молекулам, включающим анти-IFNAR-1 антитело по настоящему изобретению или его фрагмент. Антитело по настоящему изобретению или его антиген-связывающий участок может быть превращен в производное или связан с другой функциональной молекулой, например другим пептидом или белком (например, другим антителом или лигандом рецептора), для получения биспецифичной молекулы, которая связывается как минимум с двумя различными сайтами связывания или молекулами-мишенями. Фактически антитело по настоящему изобретению может быть превращено в производное или связано с более чем одной из других функциональных молекул для получения мультиспецифичных молекул, которые связываются более чем с двумя различными сайтами связывания и/или молекулами-мишенями; причем имеется в виду, что в настоящем описании такие мультиспецифичные молекулы также охвачены термином «биспецифичная молекула». Для создания биспецифичной молекулы по настоящему изобретению антитело по настоящему изобретению может быть функционально связано (например, путем химического сочетания, генетического слияния, нековалентной ассоциации или другим способом) с одной или несколькими другими связывающимися молекулами, такими как другое антитело, фрагмент антитела, пептид или связывающийся миметик, так что образуется биспецифичная молекула.
Соответственно настоящее изобретение включает биспецифичные молекулы, содержащие как минимум один первый специфично связывающийся фрагмент для IFNAR-1 и второй специфично связывающийся фрагмент для другого целевого эпитопа. В конкретном варианте осуществления настоящего изобретения второй целевой эпитоп представляет собой Fc рецептор, например FcγRI человека (CD64) или Fcα рецептор человека (CD89). Следовательно, изобретение включает биспецифичные молекулы, способные как к связыванию с эффекторными клетками, экспрессирующими FcγR, FcαR или FcεR (например, моноцитами, макрофагами или полиморфоядерными клетками (PMN)), так и с клетками-мишенями, экспрессирующими IFNAR-1. Эти биспецифичные молекулы нацеливают IFNAR-1 экспрессирующие клетки на эффекторные клетки и инициируют различные виды активности эффекторных клеток, опосредованные рецептором Fc, такие как фагоцитоз IFNAR-1 экспрессирующих клеток, зависимая от антител клеточно-опосредованная цитотоксичность (ADCC), высвобождение цитокинов или генерация супероксидного аниона.
В варианте осуществления настоящего изобретения, в котором биспецифичная молекула является мультиспецифичной, эта молекула может дополнительно включать третий специфично связывающийся фрагмент, помимо специфично связывающегося анти-Fc фрагмента и специфично связывающегося анти-IFNAR-1 фрагмента. В одном из вариантов осуществления третий специфично связывающийся фрагмент представляет собой участок антифактора усиления (EF), например молекулу, которая связывается с поверхностным белком, вовлеченным в цитотоксическую активность, и тем самым усиливает иммунную реакцию против клетки-мишени. «Участок антифактора усиления» может представлять собой антитело, функциональный фрагмент антитела или лиганд, который связывается с данной молекулой, например антигеном или рецептором, и тем самым приводит к усилению воздействия связывающих детерминант на рецептор Fc или антиген клетки-мишени. «Участок антифактора усиления» может связывать Fc рецептор или антиген клетки-мишени. С другой стороны, участок антифактора усиления может связываться с объектом, который отличается от объекта, с которым связываются первый и второй специфично связывающиеся фрагменты. Например, участок антифактора усиления может связывать цитотоксичные T-клетки (например, через CD2, CD3, CD8, CD28, CD4, CD40, ICAM-1 или другие иммунные клетки, что приводит к усилению иммунной реакции против клетки-мишени).
В одном из вариантов осуществления биспецифичные молекулы по настоящему изобретению включают в качестве специфично связывающегося фрагмента как минимум одно антитело или его фрагмент, включающий, например, Fab, Fab', F(ab')2, Fv или одноцепочечный Fv. Антитело может быть также димером легкой цепи или тяжелой цепи или любым их минимальным фрагментом, таким как фрагмент Fv или одноцепочечная конструкция, как описано Ladner и соавторами в патенте США № 4946778, содержание которого непосредственно включено в настоящую заявку с помощью ссылки.
В одном из вариантов осуществления специфичность связывания с рецептором Fcγ обеспечивается моноклональным антителом, связывание которого не блокировано иммуноглобулином G человека (IgG). В настоящем описании термин «рецептор IgG» относится к любому из восьми генов γ-цепи, находящихся на хромосоме 1. Эти гены кодируют в общей сложности двенадцать изоформ трансмембранных или растворимых рецепторов, которые сгруппированы в три класса рецепторов Fcγ: FcγRI (CD64), FcγRII(CD32) и FcγRIII(CD16). В одном из предпочтительных вариантов осуществления рецептор Fcγ представляет собой высокоафинный FcγRI человека. FcγRI представляет собой молекулу массой 72 кДа, которая проявляет высокое сродство к мономерному IgG (108-109 M-1).
Получение и свойства некоторых предпочтительных анти-Fcγ моноклональных антител описаны Fanger и соавторами в публикации PCT WO 88/00052 и патенте США № 4954617, содержание которого полностью включено в настоящую заявку с помощью ссылки. Эти антитела связываются с эпитопом FcγRI, FcγRII или FcγRIII по сайту, который отличается от сайта связывания Fcγ рецептора, и, таким образом, их связывание не блокируется в существенной степени физиологическими уровнями IgG. Специфичными анти-FcγRI антителами, применимыми в настоящем изобретении, являются mAb 22, mAb 32, mAb 44, mAb 62 и mAb197. Гибридома, вырабатывающая mAb 32, доступна из американской коллекции типовых культур (ATCC), инвентарный № HB9469. В других вариантах осуществления анти-Fcγ рецептор антитело представляет собой гуманизированную форму моноклонального антитела 22 (H22). Получение и свойства антитела H22 описаны в Graziano, R.F. et al. (1995) J.Immunol, 155 (10):4996-5002 и публикации PCT WO 94/10332. Линия клеток, вырабатывающих антитело H22, была депонирована в американской коллекции типовых культур с обозначением HA022CL1 и имеет инвентарный №CRL 11177.
В других предпочтительных вариантах осуществления специфичность связывания с рецептором Fc обеспечена антителом, которое связывается с рецептором IgA человека, например Fc-альфа рецептором (FcαRI(CD89)), связывание которого предпочтительно не блокируется иммуноглобулином A человека (IgA). Имеется в виду, что термин «рецептор IgA» включает продукт α-гена (FcαRI), расположенного на хромосоме 19. Известно, что этот ген кодирует несколько альтернативно сплайсированных трансмембранных изоформ, массой от 55 до 110 кДа. FcαRI(CD89) конститутивно экспрессируется на моноцитах/макрофагах, эозинофильных и нейтрофильных гранулоцитах, но не на популяциях неэффекторных клеток. FcαRI обладает средним сродством (≈5·107 M-1) к обеим изоформам IgA1 и IgA2, которое увеличивается при действии цитокинов, таких как G-CSF или GM-CSF (Morton, H.C. et al. (1996) Critical Reviews in Immunology, 16:423-440). Были описаны четыре FcαRI-специфичных моноклональных антитела, идентифицированных как A3, A59, A62 и A77, которые связываются с FcαRI за пределами домена, связывающего лиганд IgA (Monteiro, R.C. et al. (1992) J.Immunol. 148:1764).
FcαRI и FcγRI представляют собой предпочтительные триггерные рецепторы для применения в биспецифичных молекулах по настоящему изобретению, поскольку они (1) в основном экспрессируются на иммунных эффекторных клетках, например моноцитах, PMN, макрофагах и дендритных клетках; (2) экспрессируются в больших количествах (например, 5000-100000 на клетку); (3) являются медиаторами активности цитокинов (например, ADCC, фагоцитоза); (4) опосредуют усиление презентации антигенов, в т.ч. нацеленных на них собственных антигенов.
Хотя предпочтительными являются моноклональные антитела человека, другими антителами, которые могут применяться в биспецифичных молекулах по настоящему изобретению, являются мышиные, химерные и гуманизированные моноклональные антитела.
Биспецифичные молекулы по настоящему изобретению могут быть получены конъюгацией составляющих специфично связывающихся фрагментов, например анти-FcR и анти-IFNAR-1 фрагментов, с применением известных в технике способов. Например, каждый из специфично связывающихся фрагментов биспецифичной молекулы может быть получен отдельно и затем конъюгирован с другим фрагментом. Если специфично связывающиеся фрагменты являются белками или пептидами, может применяться большое количество связывающих или поперечно-сшивающих реагентов для ковалентной конъюгации. Примеры поперечно-сшивающих реагентов включают белок A, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 5,5'-дитиобис(2-нитробензойную кислоту) (DTNB), о-фенилендималеимид (oPDM), N-сукцинимидил-3-(2-пиридилтио)пропионат (SPDP) и сульфосукцинимидил 4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC) (см., например, Karpovsky et al. (1984) J.Exp.Med. 160:1686; Liu, MA et al. (1985) Proc. Natl. Acad. Sci. USA, 82:8648). Другие методики включают способы, описанные в следующих публикациях Paulus (1985) Behring Ins.Mitt. No.78, 118-132; Brennan et al. (1985) Science, 229:81-83 и Glennie et al. (1987) J.Immunol. 139:2367-2375. Предпочтительными реагентами для конъюгации являются SATA и сульфо-SMCC, причем оба этих реагента могут быть приобретены у Pierce Chemical Co. (Rockford, IL).
Если специфично связывающимися фрагментами являются антитела, они могут быть конъюгированы через сульфгидрильное связывание C-концов шарнирных областей двух тяжелых цепей. В особенно предпочтительном варианте осуществления шарнирную область модифицируют так, чтобы она до конъюгации содержала нечетное число сульфгидрильных остатков, предпочтительно один.
С другой стороны, оба специфично связывающихся фрагмента могут быть закодированы в одном и том же векторе и экспрессированы и собраны в одной и той же клетке-хозяине. Этот метод особенно применим, если биспецифичная молекула представляет собой mAb · mAb, mAb · Fab, Fab · F(ab')2 или лиганд · Fab слитый белок. Биспецифичная молекула по настоящему изобретению может быть одноцепочечной молекулой, включающей одно одноцепочечное антитело и детерминанту связывания, или одноцепочечной биспецифичной молекулой, включающей две детерминанты связывания. Биспецифичные молекулы могут включать как минимум две одноцепочечные молекулы. Способы получения биспецифичных молекул описаны, например, в патенте США № 5260203; патенте США № 5455030; патенте США № 4881175; патенте США № 5132405; патенте США № 5091513; патенте США № 5476786; патенте США № 5013653; патенте США № 5258498; и патенте США № 5482858.
Связывание биспецифичных молекул с их конкретными мишенями может быть подтверждено, например, твердофазным иммуноферментным анализом (ELISA), радиоиммунным анализом (RIA), FACS анализом, биологическим анализом (например, ингибированием роста) или вестерн-блоттингом. Каждый из перечисленных способов анализа, как правило, обнаруживает наличие интересующих комплексов белок-антитело за счет применения меченого реагента (например, антитела), специфичного в отношении интересующего комплекса. Например, комплексы FcR-антитело могут быть обнаружены с применением, например, антитела или его фрагмента, связанного с ферментом, которые распознают и специфично связываются с комплексами антитело-FcR. С другой стороны, эти комплексы могут быть обнаружены с применением любого из многих других способов иммунного анализа. Например, антитело может быть радиоактивно меченым и применяться в радиоиммунном анализе (RIA) (см., например, Weintraub, B., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986, причем данная публикация включена в настоящую заявку с помощью ссылки). Радиоактивный изотоп может быть обнаружен такими средствами, как использование γ-счетчика или сцинтилляционного счетчика или ауторадиографией.
Фармацевтические композиции
В другом аспекте настоящее изобретение относится к композициям, например фармацевтическим композициям, содержащим одно моноклональное тело или его антиген-связывающий участок (участки) или комбинацию таких тел или участков по настоящему изобретению, в сочетании с фармацевтически приемлемым носителем. Такие композиции могут включать одно антитело или их комбинацию (например, двух или нескольких различных антител), или иммуноконъюгаты, или биспецифичные молекулы по настоящему изобретению. Например, фармацевтическая композиция по настоящему изобретению может включать комбинацию антител (или иммуноконъюгатов, или биспецифичных антител), которые связываются с различными эпитопами на целевом антигене или которые имеют дополнительные виды активности.
Фармацевтические композиции по настоящему изобретению также могут назначаться в комбинированной терапии, т.е. в сочетании с другими средствами. Например, комбинированная терапия может включать анти-IFNAR-1 антитело по настоящему изобретению в сочетании как минимум с одним иммунодепрессивным средством.
В настоящем описании термин «фармацевтически приемлемый носитель» включает все без исключения растворители, дисперсионные среды, покрытия, антибактериальные и антигрибковые средства, изотонические и замедляющие абсорбцию средства и т.п., которые являются физиологически совместимыми. Предпочтительно носитель подходит для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, с помощью инъекции или инфузии). В зависимости от пути введения, действующее соединение, т.е. антитело, иммуноконъюгат или биспецифичная молекула, может быть покрыто соответствующим веществом для защиты соединения от действия кислот и других естественных условий, которые могут инактивировать действующее соединение.
Фармацевтические соединения по настоящему изобретению могут включать одну или несколько фармацевтически приемлемых солей. Термин «фармацевтически приемлемая соль» относится к соли, которая сохраняет желаемую биологическую активность исходного соединения и не дает каких бы то ни было нежелательных токсикологических эффектов (см., например, Berge, S.M., et al. (1977) J.Pharm. Sci. 66:1-19). Примеры таких солей включают кислотно-аддитивные и основно-аддитивные соли. Кислотно-аддитивные соли включают соли, полученные из нетоксичных неорганических кислот, таких как хлористоводородная, азотная, фосфорная, серная, бромистоводородная, йодистоводородная, фосфорная и т.п., а также из нетоксичных органических кислот, таких как алифатические моно- и дикарбоновые кислоты, фенилзамещенные алкановые кислоты, гидроксиалкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты и т.п. Основно-аддитивные соли включают соли, полученные из щелочных и щелочноземельных металлов, таких как натрий, калий, магний, кальций и т.п., а также из нетоксичных органических аминов, таких как N,N'-дибензилэтилендиамин, N-метилглюкамин, хлорпрокаин, холин, диэтаноламин, этилендиамин, прокаин и т.п.
Кроме этого фармацевтическая композиция по настоящему изобретению также может включать фармацевтически приемлемый антиоксидант. Примеры фармацевтически приемлемых антиоксидантов включают: (1) водорастворимые антиоксиданты, такие как аскорбиновая кислота, гидрохлорид цистеина, бисульфат натрия, метабисульфит натрия, сульфит натрия и т.п.; (2) маслорастворимые антиоксиданты, такие как аскорбил пальмитат, бутилированный гидроксианизол (BHA), бутилированный гидрокситолуол (BHT), лецитин, пропилгаллат, альфа-токоферол и т.п.; и (3) хелатирующие металл реагенты, такие как лимонная кислота, этилендиаминтетрауксусная кислота (ЭДТУ), сорбит, винная кислота, фосфорная кислота и т.п.
Примеры подходящих водных и неводных носителей, которые могут применяться в фармацевтических композициях по настоящему изобретению, включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.), а также их подходящие смеси, растительные масла, такие как оливковое масло, и пригодные для инъекций органические сложные эфиры, такие как этилолеат. Надлежащая подвижность может поддерживаться, например, применением покрывающих материалов, таких как лецитин, в случае дисперсий сохранением требуемого размера частиц, а также применением ПАВ.
Описываемые композиции могут также содержать вспомогательные вещества, такие как консерванты, смачивающие средства, эмульгаторы и диспергирующие средства. Защита от присутствия микроорганизмов может быть гарантирована как процедурами стерилизации, см. выше, так и включением различных антибактериальных и антигрибковых средств, например парабена, хлорбутанола, фенола, сорбиновой кислоты и т.п. Также может быть желательно включить в состав композиций изотонические средства, такие как сахара, хлорид натрия и т.п. Кроме того, длительная абсорбция инъецируемых лекарственных форм может быть достигнута введением агентов, которые задерживают абсорбцию, таких как моностеарат алюминия и желатин.
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии, а также стерильные порошки для экстемпорального приготовления стерильных пригодных для инъекций растворов или суспензий. Применение таких сред и агентов для фармацевтически активных веществ известно в технике. В фармацевтических композициях по настоящему изобретению предполагается применение любой традиционной среды или агента, если они не являются несовместимыми с действующим соединением. Кроме этого в композиции могут быть включены дополнительные действующие вещества.
Терапевтические композиции, как правило, должны быть стерильными и стабильными в условиях производства и хранения. Композиция может принимать форму раствора, микроэмульсии, липосом или другой заданной структуры, подходящей для высокой концентрации лекарственного средства. Носитель может быть растворителем или дисперсионной средой, содержащей, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Надлежащая подвижность может поддерживаться, например, применением покрывающих материалов, таких как лецитин, в случае дисперсий сохранением требуемого размера частиц, а также применением ПАВ. Во многих случаях будет предпочтительно включить в состав композиции изотонические средства, например сахара, полиспирты, такие как манит и сорбит, или хлорид натрия. Пролонгированная абсорбция инъецируемых композиций может быть достигнута за счет введения в композиции агентов, которые задерживают абсорбцию, например моностеаратные соли и желатин.
Стерильные пригодные для инъекций растворы могут быть получены введением действующего соединения в требуемом количестве в подходящий растворитель вместе с одним из перечисленных выше ингредиентов или их комбинацией, если это необходимо, с последующей стерилизацией с помощью микрофильтрации. В основном дисперсии получают введением действующего соединения в стерильный носитель, который содержит основную дисперсионную среду и другие требуемые ингредиенты из числа перечисленных выше. В случае стерильных порошков для приготовления стерильных пригодных для инъекций растворов, предпочтительными способами получения являются высушивание в вакууме и сушка вымораживанием (лиофилизация), которые позволяют получить порошок действующего ингредиента и любого дополнительного желаемого ингредиента из его полученного предварительно стерильного фильтрованного раствора.
Количество действующего ингредиента, которое может быть объединено с веществом носителя для получения лекарственной формы разовой дозировки, будет изменяться в зависимости от подвергаемого лечению субъекта и конкретного способа введения. Количество действующего ингредиента, которое может быть объединено с веществом носителя для получения лекарственной формы разовой дозировки, будет, как правило, являться таким количеством композиции, которое приводит к терапевтическому эффекту. В основном, исходя из процентной шкалы, это количество будет находиться в пределах от примерно 0,01 процента до примерно 99 процентов действующего ингредиента, предпочтительно от примерно 0,1 процента до примерно 70 процентов, наиболее предпочтительно от примерно 1 процента до примерно 30 процентов действующего ингредиента в комбинации с фармацевтически приемлемым носителем.
Требования по дозировке регулируются для обеспечения оптимальной желаемой реакции (например, терапевтической реакции). Например, может быть осуществлено однократное болюсное введение, может быть введено несколько отдельных доз в определенный промежуток времени или же доза может быть пропорционально уменьшена или увеличена в зависимости от требований текущей ситуации в процессе лечения. Особенно предпочтительно получать парентеральные композиции в виде лекарственных форм разовой дозировки с целью облегчения введения и равномерности дозирования. В настоящем описании термин «лекарственные формы разовой дозировки» относится к физически дискретным единицам, пригодным в качестве доз для однократного введения субъектам, подвергающимся лечению; каждая единица содержит заранее установленное количество действующего соединения, рассчитанное на получение желаемого терапевтического эффекта, в сочетании с требуемым фармацевтическим носителем. Спецификация лекарственных форм разовой дозировки по настоящему изобретению обусловливается и непосредственно зависит от (a) конкретных характеристик действующего соединения и конкретного терапевтического результата, который должен быть достигнут, и (b) ограничений по чувствительности у отдельных пациентов, существующих в технике составления смесей, подобных применяемому для лечения действующему соединению.
При введении антитела дозировка изменяется в диапазоне от примерно 0,0001 до 100 мг/кг и чаще от 0,01 до 5 мг/кг массы тела пациента, принимающего препарат. Например, дозировки могут составлять 0,3 мг/кг массы тела, 1 мг/кг массы тела, 3 мг/кг массы тела, 5 мг/кг массы тела или 10 мг/кг массы тела или в диапазоне 1-10 мг/кг. Типовой режим лечения определяет введение один раз в неделю, один раз каждые две недели, один раз каждые три недели, один раз в месяц, один раз каждые три месяца, один раз каждые 3-6 месяцев. Предпочтительные режимы дозирования для анти-IFNAR-1 антитела по настоящему изобретению включают 1 мг/кг массы тела или 3 мг/кг массы тела при внутривенном введении, причем антитело вводят, используя один из следующих календарных планов: (i) каждые четыре недели для первых шести доз, затем каждые три месяца; (ii) каждые три недели; (iii) однократное введение 3 мг/кг массы тела, затем 1 мг/кг массы тела каждые три недели.
В некоторых способах два или несколько моноклональных антител с различными специфичностями связывания вводятся одновременно, и в этом случае дозировка каждого из вводимых антител находится в указанных диапазонах. Антитела обычно вводят по многим причинам. Промежутки между отдельными введениями могут быть, например, недельными, месячными, трехмесячными или годичными. Промежутки также могут быть неравными и определяться в соответствии с измерениями уровней антитела к целевому антигену в крови пациента. В некоторых способах дозировку регулируют для достижения концентрации антитела в плазме примерно 1-1000 мкг/мл и в отдельных способах примерно 25-300 мкг/мл.
С другой стороны, антитела могут вводиться в виде составов с продолжительным высвобождением, причем в этом случае требуется менее частое введение. Дозировка и частота введения меняются в зависимости от времени полужизни антитела в организме пациента. Как правило, наиболее продолжительное время жизни демонстрируют антитела человека, за ними следуют гуманизированные антитела, химерные антитела и антитела, не являющиеся антителами человека. Дозировка и частота введения могут меняться в зависимости от того, является ли лечение профилактическим или терапевтическим. При профилактическом применении вводят относительно низкие дозы через относительно продолжительные промежутки на протяжении длительного периода времени. Некоторые пациенты продолжают получать лечение в течение всей жизни. При терапевтическом применении иногда требуются относительно высокие дозы в относительно короткие промежутки времени до тех пор, пока развитие заболевания не замедлится или не прекратится, и предпочтительно до тех пор, пока пациент не продемонстрирует частичное или полное облегчение симптомов заболевания. После этого лекарственное средство может вводиться пациенту в профилактическом режиме.
Фактические уровни дозировки действующих ингредиентов в фармацевтических композициях по настоящему изобретению могут подвергаться изменениям, с тем чтобы получить количество действующего ингредиента, которое является эффективным для достижения желаемой терапевтической реакции у конкретного пациента, для данной композиции и способа введения, без проявлений токсичности для пациента. Выбранный уровень дозировки будет зависеть от широкого круга фармакокинетических факторов, включая активность конкретной применяемой композиции по настоящему изобретению или ее сложного эфира, соли или амида, пути введения, времени введения, скорости выведения конкретного применяемого соединения, продолжительности лечения, других лекарственных средств, соединений и/или веществ, применяемых в комбинации с конкретными применяемыми композициями, возраста, пола, массы, болезненного состояния, общего состояния здоровья и предыдущей истории болезни пациента, подвергаемого лечению, и подобных факторов, хорошо известных в медицине.
«Терапевтически эффективная дозировка» анти-IFNAR-1 антитела по настоящему изобретению предпочтительно приводит к уменьшению тяжести симптомов болезни, увеличению частоты и продолжительности бессимптомных периодов или предотвращению ухудшения здоровья или потери трудоспособности из-за развития заболевания. В случае, например, системной красной волчанки (SLE), терапевтически эффективная доза предпочтительно предотвращает дальнейшее ухудшение физических симптомов, связанных с SLE, таких как, например, боль, усталость и слабость. Терапевтически эффективная доза также предпочтительно предотвращает или замедляет наступление SLE, что, например, может быть желательно, если имеются первоначальные или предварительные признаки заболевания. Также сюда входит замедление хронического развития, связанное с SLE. Лабораторные исследования, применяемые в диагностике SLE, включают химические анализы, гематологию, серологию и радиологию. Соответственно, любой клинический или биохимический анализ, который контролирует что-либо из перечисленного выше, может применяться для определения того, является ли данная доза терапевтически эффективной для лечения SLE. Обычный специалист в данной области техники был бы способен установить такие количества, на основании таких факторов, как размер субъекта, тяжесть симптомов у субъекта, а также конкретная выбранная композиция или выбранный путь введения.
Композиции по настоящему изобретению могут вводиться одним или несколькими путями с применением одного или нескольких из числа разнообразных способов, известных в технике. Как поймет специалист в данной области техники, путь и/или форма введения будут изменяться в зависимости от желаемых результатов. Предпочтительные пути введения антител по настоящему изобретению включают внутривенный, внутримышечный, внутрикожный, интраперитонеальный, подкожный, спинальный или другие парентеральные пути введения, например, с помощью инъекции или инфузии. Фраза «парентеральное введение» в настоящем описании означает формы введения, отличные от кишечного и местного введения, как правило, с помощью инъекции или инфузии, т.е. не ограничиваясь перечисленным, внутривенные, внутримышечные, внутриартериальные, интратекальные, внутрисуставные, внутриглазничные, внутрисердечные, внутрикожные, интраперитонеальные, транстрахеальные, подкожные, субкутикулярные, внутрисуставные, субкапсулярные, субарахноидальные, интраспинальные, эпидуральные и внутригрудинные инъекции и инфузии.
С другой стороны, антитела по настоящему изобретению могут вводиться непарентеральными путями, такими как местный и эпидермальный пути введения или через слизистые оболочки, например интраназально, перорально, вагинально, ректально, сублингвально или топически.
Препараты действующих соединений могут быть получены с применением носителей, которые будут предохранять эти соединения от быстрого высвобождения, т.е. могут применяться составы с регулируемым высвобождением, в т.ч. имплантаты, трансдермальные пластыри и микрокапсулированные системы доставки. Могут применяться биоразрушаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Большое количество способов получения таких составов запатентовано или известно всем специалистам в данной области техники. См, например, Sustained and Controlled Release Drug Delivery Systems, J.R.Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Терапевтические композиции могут вводиться с помощью известных в технике медицинских приборов. Например, в предпочтительном варианте осуществления терапевтическую композицию по настоящему изобретению можно вводить с помощью гиподермического безыгольного инъектора, как например устройства, раскрытого в патентах США №№ 5399163; 5383851; 5312335; 5064413; 4941880; 4790824; или 4596556. Примеры хорошо известных имплантатов и модулей, применимых в настоящем изобретении, включают: патент США № 4487603, в котором раскрыт имплантируемый микроинфузионный насос для дозирования лекарственного средства с регулируемой скоростью; патент США № 4486194, в котором раскрыт терапевтический прибор для введения лекарственных средств через кожу; патент США № 4447233, в котором раскрыт насос для инфузии лекарственных препаратов предназначенный для доставки лекарственного средства при точной скорости инфузии; патент США № 4447224, в котором раскрыт имплантируемый инфузионный аппарат с изменяемым потоком для непрерывной доставки лекарств; патент США № 4439196, в котором раскрыта осмотическая система доставки лекарств, имеющая многокамерные отсеки; и патент США № 4475196, в котором раскрыта осмотическая система доставки лекарств. Эти патенты включены в настоящую заявку с помощью ссылки. Многие другие подобные имплантаты, системы доставки и модули известны специалистам в данной области техники.
В некоторых вариантах осуществления моноклональные антитела человека по настоящему изобретению могут быть включены в такие составы, чтобы гарантировать надлежащее распределение in vivo. Например, барьер кровь-мозг (BBB) исключает проникновение многих высокогидрофильных соединений. Для гарантии проникновения терапевтических соединений по настоящему изобретению через BBB (если это желательно), они могут быть включены в состав, например, на основе липосом. Для ознакомления со способами получения липосом см., например, патенты США 4522811; 5374548; и 5399331. Липосомы могут включать один или несколько фрагментов, которые селективно транспортируются в конкретные клетки или органы и, таким образом, улучшают целенаправленную доставку лекарств (см., например, V.V.Ranade (1989) J.Clin.Pharmacol. 29:685). Типовые нацеливающие фрагменты включают фолат или биотин (см., например, патент США 5416016, выданный Low и соавторам); маннозиты (Umezawa et al. (1988) Biochem.Biophys.Res.Commun. 153:1038); антитела (P.G.Bloeman et al. (1995) FEBS Lett. 357:140; M.Owais et al. (1995) Antimicrob.Agents Chemother. 39:180); поверхностно-активный рецептор белка A (Briscoe et al. (1995) Am.J.Physiol. 1233:134); p120 (Schreier et al. (1994) J.Biol.Chem. 269:9090); см. также K.Keinanen; M.L.Laukkanen (1994) FEBS Lett. 346:123; J.J.Killion:I.J.Fidler (1994) Immunomethods. 4:273.
Применения и способы по настоящему изобретению
Антитела (а также иммуноконъюгаты и биспецифичные молекулы) по настоящему изобретению имеют диагностические и терапевтические применения in vitro и in vivo. Например, эти молекулы могут вводиться в клетки в культуре, например, in vitro или ex vivo, или субъекту, например, in vivo, для лечения профилактики или диагностики большого количества расстройств. Предполагается, что термин «субъект» в настоящем описании включает человека и животных, не принадлежащих к человеческому роду. Животные включают всех позвоночных, например, млекопитающих и немлекопитающих, таких как не принадлежащие к человеческому роду приматы, овцы, собаки, кошки, коровы, лошади, куры, земноводные и пресмыкающиеся. Способы, разработанные в настоящем изобретении, особенно подходят для лечения пациентов из числа людей, имеющих расстройство, связанное с отклоняющейся от нормы или неадекватной экспрессией интерферона типа I (например, чрезмерной экспрессией).
Если антитела к IFNAR-1 вводят совместно с другим средством, оба эти препарата могут вводиться в любом порядке или одновременно. Например, анти-IFNAR-1 антитело по настоящему изобретению может применяться в сочетании с одним или несколькими из следующих средств: анти-IFNα антитело, антитело против рецептора IFNγ, растворимый рецептор IFNγ, анти-TNF антитело, антитело против рецептора TNF и/или растворимого рецептора TNF (см., например, патент США № 5888511). Кроме того, анти-IFNAR-1 антитело по настоящему изобретению может применяться в сочетании с антагонистом лиганда Flt3 (см., например, заявку на патент США №2002/0160974).
В одном из вариантов осуществления антитела (а также иммуноконъюгаты и биспецифичные молекулы) по настоящему изобретению могут применяться для определения уровней IFNAR-1 или уровней клеток, которые экспрессируют IFNAR-1. Это может быть достигнуто, например, введением во взаимодействие образца (как, например, образца in vitro) и контрольного образца с анти-IFNAR-1 антителом при условиях, которые делают возможным образование комплекса между антителом и IFNAR-1. Любые комплексы, образованные антителом и IFNAR-1, обнаруживают и сравнивают в исследуемом и контрольном образце. При использовании композиций по настоящему изобретению могут реализовываться, например, стандартные способы обнаружения, хорошо известные в технике, такие как ELISA и поточная цитометрия.
Соответственно в одном из аспектов изобретение дополнительно относится к способам обнаружения наличия IFNAR-1 (например, антигена IFNAR-1 человека) в образце или измерения количества IFNAR-1, включающим введение во взаимодействие образца и контрольного образца с антителом по настоящему изобретению или его антиген-связывающим участком, которые специфично связываются с IFNAR-1, при условиях, которые делают возможным образование комплекса между антителом или его фрагментом и IFNAR-1. Затем устанавливают образование комплекса, причем разница в комплексообразовании между образцом и контрольным образцом является показателем наличия IFNAR-1 в образце.
Также в объем настоящего изобретения входят комплекты, включающие композиции по настоящему изобретению (например, антитела, антитела человека, иммуноконъюгаты и биспецифичные молекулы) и инструкции по применению. Такой комплект может дополнительно содержать как минимум один дополнительный реагент или одно или несколько дополнительных антител по настоящему изобретению (например, антитело, обладающее комплементарной активностью, которое связывается с эпитопом на целевом антигене, отличном от первого антитела). Комплекты, как правило, включают этикетку с указаниями на предполагаемое применение содержимого комплекта. Термин «этикетка» включает любой письменный или записанный материал, нанесенный на комплект или поставляемый вместе с ним или каким-либо другим образом сопровождающий комплект.
IFNAR-1 является одним из клеточных рецепторов интерферонов типа I, и интерфероны типа I, как известно, являются иммунорегуляторными цитокинами, которые вовлечены в том числе в дифференцировку T-клеток, выработку и активность антител и сохранение памяти T-клеток. Кроме того, увеличенная экспрессия интерферонов типа I была отмечена при многих аутоиммунных заболеваниях, ВИЧ-инфекции, при отторжении трансплантатов и реакции «трансплантат против хозяина» (GVHD). Соответственно, анти-IFNAR-1 антитела (а также иммуноконъюгаты и биспецифичные молекулы) по настоящему изобретению, которые ингибируют функциональную активность интерферонов типа I, могут применяться при разнообразных клинических показаниях, включающих отклоняющуюся от нормы или нежелательную активность интерферонов типа I. Следовательно, настоящее изобретение относится к способу ингибирования заболеваний и расстройств, опосредованных интерфероном типа I, причем способ включает введение антитела или его антиген-связывающего участка по настоящему изобретению (или иммуноконъюгата, или биспецифичной молекулы по настоящему изобретению), с тем чтобы подвергнуть лечению заболевание или расстройство, опосредованное интерфероном типа I.
Конкретные примеры аутоиммунных состояний, при которых могут применяться антитела по настоящему изобретению, включают, не ограничиваясь перечисленным, следующие: системная красная волчанка (SLE), инсулин-зависимый сахарный диабет (IDDM), воспалительные заболевания кишечника (IBD) (в т.ч. болезнь Крона, язвенный колит и глютеиновая болезнь), рассеянный склероз (MS), псориаз, аутоиммунный тиреоидит, ревматоидный артрит (RA) и гломерулонефрит. Кроме того, композиции антител по настоящему изобретению могут применяться для ингибирования или предотвращения отторжения трансплантата или лечения реакции «трансплантат против хозяина» (GVHD) или при лечении ВИЧ-инфекции/СПИДа.
Высокие уровни IFNα наблюдались в сыворотке пациентов с системной красной волчанкой (SLE) (см., например, Kim et al. (1987) Clin.Exp.Immunol. 70:562-569). Кроме того, было показано, что введение IFNα, например, при лечении рака или вирусных инфекций вызывает SLE (Garcia-Porrua et al. (1998) Clin.Exp.Rheumatol. 16:107-108). Соответственно, в другом варианте осуществления анти-IFNAR-1 антитела по настоящему изобретению могут применяться при лечении SLE путем введения антитела субъекту при необходимости лечения. Антитело может применяться само по себе или в сочетании с другими средствами против SLE, такими как нестероидные противовоспалительные препараты (NSAID), анальгетики, кортикостероиды (например, преднизон, гидрокортизон), иммунодепрессанты (такие как циклофосфамид, азатиоприн и метотрексат), антималярийные препараты (такие как гидроксихлорохин) и биологические лекарственные средства, которые ингибируют выработку антител против dsДНК (двухцепочечной ДНК) (например, LJP 394).
IFNα также вовлечен в патологию диабета типа I. Например, сообщалось о наличии иммунореактивного IFNα в бета-клетках поджелудочной железы у пациентов с диабетом типа I (Foulis et al. (1987) Lancet. 2:1423-1427). Кроме этого было показано, что длительное применение IFNα в антивирусной терапии вызывает диабет типа I (Waguri et al. (1994) Diabetes Res.Clin.Pract. 23:33-36). Соответственно, в другом варианте осуществления анти-IFNAR-1 антитела по настоящему изобретению могут применяться в лечении диабета типа I путем введения антитела субъекту в случае необходимости лечения. Антитело может применяться само по себе или в сочетании с другими антидиабетическими средствами, такими как инсулин.
Было показано, что антитела к IFNAR являются эффективными в животных моделях воспалительных заболеваний кишечника (см. заявку на патент США 60/465155). Таким образом, анти-IFNAR-1 антитела по настоящему изобретению могут применяться в лечении воспалительных заболеваний кишечника (IBD), включая язвенный колит и болезнь Крона, путем введения антитела субъекту в случае необходимости лечения. Это антитело может применяться само по себе или в сочетании с другими анти-IBD средствами, как например лекарственными препаратами, содержащими мезаламин (включая сульфасалазин и другие средства, содержащие 5-аминосалициловую кислоту (5-ASA), такие как олсалазин и балсалазид), нестероидные противовоспалительные препараты (NSAID), анальгетики, кортикостероиды (например, преднизон, гидрокортизон), ингибиторы TNF (включая адилимумаб (Humira®), этанерсепт (Enbrel®) и инфликсимаб (Remicade®)), иммунодепрессанты (такие как 6-меркаптопурин, азатиоприн и циклоспорин A), а также антибиотики.
Также наблюдали, что лечение с использованием IFNα вызывает аутоиммунный тиреоидит (Monzani et al. (2004) Clin.Exp.Med. 3:199-210; Prummel and Laurberg (2003) Thyroid. 13:547-551). Соответственно в другом варианте осуществления анти-IFNAR антитела по настоящему изобретению могут применяться при лечении аутоиммунных заболеваний щитовидной железы, включая аутоиммунный первичный гипотиреоидизм, базедову болезнь, тиреоидит Хашимото и деструктивный тиреоидит с гипотиреоидизмом, путем введения антитела субъекту при необходимости лечения. Это антитело может применяться само по себе или в комбинации с другими средствами или способами лечения, как например антитиреоидными лекарственными препаратами, радиоактивным йодом и частичной тиреоидэктомией.
Повышенные уровни интерферонов типа I, в особенности IFNβ, наблюдались в сыворотке пациентов с RA (см., например, Hertzog et al. (1988) Clin.Immunol.Immunopath. 48:192). Таким образом, в одном из вариантов осуществления анти-IFNAR-1 антитела по настоящему изобретению могут применяться в лечении RA путем введения этих антител субъекту в случае необходимости такого лечения. Антитела могут применяться сами по себе или в комбинации с одним или несколькими другими анти-RA средствами, такими как нестероидные противовоспалительные препараты (NSAID), ингибиторы COX-2, анальгетики, кортикостероиды (например, преднизон, гидрокортизон), золото, иммунодепрессанты (например, метотрексат), средства для истощения популяции B-клеток (например, RituxanTM), агонисты B-клеток (например, LymphoStat-BTM) и анти-TNF-α средства (например, EMBRELTM, HUMIRA® и REMICADETM).
Сообщалось, что введение IFNα обостряет псориаз. Соответственно в другом варианте осуществления анти-IFNAR-1 антитело по настоящему изобретению может применяться в лечении псориаза и псориатического артрита путем введения антитела субъекту при необходимости такого лечения. Антитело может применяться само по себе или в сочетании с одним или несколькими другими видами лечения от псориаза, такими как светолечение, местная терапия (например, местное введение глюкокортикоидов) или системная терапия (например, метотрексат, синтетический ретиноид, циклоспорин), анти-TNFα средство (например, EMBRELTM, HUMIRA® и REMICADETM) и ингибитор T-клеток (например, RaptivaTM).
Кроме этого высокие уровни IFNα наблюдались в крови пациентов с ВИЧ-инфекцией, причем их наличие является прогнозирующим признаком развития СПИДа (DeStefano et al. (1982) J.Infec.Disease. 146:451; Vadhan-Raj et al. (1986) Cancer Res. 46:417). Таким образом, в другом варианте осуществления анти-IFNAR-1 антитело по настоящему изобретению применяют в лечении ВИЧ-инфекции или СПИДа путем введения антитела субъекту в случае необходимости лечения. Антитело может быть применено само по себе или в сочетании с другими средствами против ВИЧ, такими как нуклеозидные ингибиторы обратной транскриптазы, ненуклеозидные ингибиторы обратной транскриптазы, ингибиторы протеазы и ингибиторы слияния.
Было продемонстрировано, что антитела к IFNAR-1 являются эффективными в ингибировании отторжения аллотрансплантата и продлении долговечности аллотрансплантата (см., например, Tovey et al. (1996) J.Leukoc.Biol. 59:512-517; Benizri et al. (1998) J.Interferon Cytokine Res. 18:273-284). Соответственно, анти-IFNAR-1 антитела по настоящему изобретению также могут применяться у реципиентов трансплантатов для ингибирования отторжения аллотрансплантатов и/или продления долговечности аллотрансплантатов. Настоящее изобретение относится к способу ингибирования отторжения трансплантата путем введения анти-IFNAR-1 антитела по настоящему изобретению реципиенту трансплантата в случае необходимости лечения. Примеры трансплантатов тканей, которые могут быть подвергнуты лечению, включают, не ограничиваясь перечисленным, печень, легкие, почки, сердце, тонкую кишку и островные клетки поджелудочной железы, также сюда входит лечение реакции «трансплантат против хозяина» (GVHD). Антитело может применяться само по себе или в комбинации с другими средствами для ингибирования отторжения трансплантата, такими как иммунодепрессивные средства (например, циклоспорин, азатиоприн, метилпреднизолон, преднизолон, преднизон, микофенолята мофетил, сирилимус, рапамицин, такролимус), противоинфекционные средства (например, ацикловир, клотримазол, ганцикловир, нистатин, триметопримсульфаметоксазол), диуретики (например, буметанид, фуросемид, метолазон) и противоязвенные лекарственные средства (например, циметидин, фарнотидин, лансопразол, омепразол, ранитидин, сукралфат).
Далее настоящее изобретение проиллюстрировано нижеследующими примерами, которые не должны истолковываться как дополнительные ограничения. Содержание всего иллюстративного материала и всех ссылок, патентов и опубликованных заявок на патенты, цитированных на протяжении данного описания, прямо включено в настоящую заявку с помощью ссылки.
Пример 1: Получение моноклональных антител человека против IFNAR-1
Антиген
Растворимый IFNAR-1, содержащий внеклеточный домен IFNAR-1, получали рекомбинантными способами и использовали в качестве антигена для иммунизации.
Трансгенные мыши HuMab
Полностью моноклональные антитела человека к IFNAR-1 получали с использованием линий HuMab трансгенных мышей HCo7, HCo12 и HCo7·HCo12, каждая из которых экспрессирует гены антител человека. В каждой из этих линий мышей, эндогенный ген каппа легкой цепи мыши подвергали гомозиготному разрушению, как описано в Chen et al. (1993) EMBO J. 12:811-820 и эндогенный ген тяжелой цепи мыши подвергали гомозиготному разрушению, как описано в примере 1 опубликованной согласно PCT WO 01/09187. Каждая из этих линий мышей несет трансген каппа легкой цепи KCo5 человека, как описано в Fishwild et al. (1996) Nature Biotechnology. 14:845-851. Линия HCo7 несет трансген тяжелой цепи HCo7 человека, как описано в патентах США №№ 5545806; 5625825 и 5545807. Линия HCo12 несет трансген тяжелой цепи HCo12 человека, как описано в примере 2 опубликованной согласно PCT WO 01/09187. Линия HCo7·HCo12 несет как трансген HCo7, так и трансген HCo12, причем она была получена совместным размножением особей двух предыдущих линий.
Иммунизация мышей HuMab
Для получения полностью моноклональных антител человека к IFNAR-1, мышей HuMab иммунизировали очищенным рекомбинантным IFNAR-1 в качестве антигена. Общие схемы иммунизации мышей HuMab описаны в Lonberg, N. et al (1994) Nature. 368 (6474): 856-859; Fishwild, D. et al. (1996) Nature Biotechnology. 14: 845-851 и опубликованной согласно PCT WO 98/24884. При первой инфузии антигена возраст мышей составлял 6-16 недель. Очищенный рекомбинантный препарат (5-50 мкг) растворимого антигена IFNAR-1 вводили для иммунизации мышей HuMab интраперитонеально, подкожно (Sc) или с помощью инъекции в подушечки лап.
Трансгенных мышей дважды иммунизировали антигеном в полном адъюванте Фрейнда или адъюванте Ribi, интраперитонеально (IP), подкожно (Sc) либо через подушечки лап (FP), после чего через 3-21 дней следовала IP, Sc или FP иммунизация (в общей сложности до 11 иммунизаций) антигеном в неполном адъюванте Фрейнда или Ribi. За иммунной реакцией наблюдали с помощью ретроорбитальных кровотечений. Осуществляли скрининг плазмы с помощью ELISA (как описано ниже) и мышей с достаточными титрами анти-IFNAR-1 иммуноглобулина человека использовали для слияний. За 2 или 3 дня до умерщвления и удаления селезенки иммунную реакцию мышей поддерживали внутривенным введением антигена. Как правило, для каждого антигена проводили 10-35 слияний. Для каждого антигена иммунизировали несколько дюжин мышей.
Отбор HuMab мышей, вырабатывающих анти-IFNAR-1 антитела
Для отбора HuMab мышей, вырабатывающих антитела, которые связываются с IFNAR-1, сыворотку из иммунизированных мышей исследовали способом ELISA, как описано D.Fishwild и соавторами (1996). Вкратце, микротитровальные планшеты покрывали очищенным рекомбинантным IFNAR-1 из E.coli в концентрации 1-2 мкг/мл в PBS, 50 мкл/лунку, инкубировали при 4°C в течение ночи, затем блокировали 200 мкл/лунку 5% сывороткой цыплят в PBS/Tween (0,05%). Плазму из IFNAR-1 иммунизированных мышей в различных разбавлениях добавляли в каждую лунку и инкубировали в течение 1-2 часов при комнатной температуре. Планшеты промывали PBS/Tween и затем инкубировали с козьим Fc поликлональным антителом против IgG человека, конъюгированным с пероксидазой хрена (HRP), в течение 1 часа при комнатной температуре. После промывания планшеты обрабатывали субстратом ABTS (Sigma, A-1888, 0,22 мг/мл) и анализировали спектрофотометром, определяя OD 415-495. Мышей, продемонстрировавших наивысшие титры анти-IFNAR-1 антител, использовали для слияний. Слияния проводили, как описано ниже, и супернатанты гибридом тестировали на анти-IFNAR-1 активность способом ELISA.
Получение гибридом, вырабатывающих моноклональные антитела человека к IFNAR-1
Слияние спленоцитов мыши, выделенных из HuMab мышей, с клеточной линией миеломы мыши в присутствии ПЭГ осуществляли на основе стандартных протоколов. Затем проводили скрининг полученных гибридом по критерию выработки антиген-специфичных антител. Осуществляли слияние суспензии отдельных клеток селезеночных лимфоцитов из иммунизированных мышей с одной четвертой частью по количеству SP2/0 несекретирующих клеток миеломы мыши (ATCC, CRL 1581) с 50% ПЭГ (Sigma). Клетки распределяли по плоскодонным микротитровальным планшетам в количестве приблизительно 1·105/лунка с последующей примерно двухнедельной инкубацией в селективной среде, содержащей 10% фетальной телячьей сыворотки, 10% кондиционированной среды P388D1 (ATCC, CRL TIB-63), 3-5% Origen (IGEN) в DMEM (Mediatech, CRL 10013 с высоким содержанием глюкозы, L-глутамина и пирувата натрия) c добавками 5 мМ HEPES, 0,055 мМ 2-меркаптоэтанола, 50 мг/мл гентамицина и 1·HAT (Sigma, SRL P-7185). Через 1-2 недели клетки культивировали в среде, в которой HAT заменяли HT. Затем лунки по отдельности подвергали скринингу способом ELISA (описан выше) на анти-IFNAR-1 человека моноклональные IgG антитела. Если имел место экстенсивный рост гибридомы, среду обычно исследовали через 10-14 дней. Гибридомы, секретировавшие антитела, повторно распределяли по планшетам, вновь подвергали скринингу и в случае повторного положительного результата на IgG человека анти-IFNAR-1 моноклональные антитела субклонировали как минимум дважды с помощью серийных разведений. Затем стабильные субклоны культивировали in vitro с целью генерации небольших количеств антитела в тканевой культурной среде для последующего получения характеристик.
Клоны гибридом 3F11, 4G5, 11E2 и 9D4 отбирали для дальнейшего анализа.
Пример 2: Получение структурных характеристик моноклональных антител человека 3F11, 4G5, 11E2 и 9D4
Последовательности кДНК, кодирующие вариабельные области тяжелой и легкой цепей моноклональных антител 3F11, 4G5, 11E2 и 9D4, получали из гибридом 3F11, 4G5, 11E2 и 9D4 соответственно, применяя стандартные PCR методики и секвенировали с использованием стандартных методик секвенирования ДНК.
Нуклеотидные и аминокислотные последовательности вариабельной области тяжелой цепи 3F11 показаны на фиг.1A и в SEQ ID NО: 33 и 25 соответственно.
Нуклеотидные и аминокислотные последовательности вариабельной области легкой цепи 3F11 показаны на фиг.1B и в SEQ ID NО: 37 и 29 соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 3F11 с известными иммуноглобулиновыми последовательностями тяжелой цепи зародышевых линий человека продемонстрировало, что в тяжелой цепи 3F11 использован VH сегмент зародышевой линии клеток человека VH 4-34, неустановленный D сегмент и JH сегмент зародышевой линии JH 6b человека. Совмещение VH последовательности 3F11 с последовательностью зародышевой линии VH 4-34 показано на фиг.5. Дальнейший анализ VH последовательности 3F11 с использованием системы определения CDR областей по Кабату привел к выявлению CDR1, CDR2 и CDR3 областей тяжелой цепи, как показано на фиг.1A и 5 и в SEQ ID NО: 1, 5 и 9 соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 3F11 с известными иммуноглобулиновыми последовательностями легкой цепи зародышевых линий человека продемонстрировало, что в легкой цепи 3F11 использован VL сегмент зародышевой линии клеток VK L18 человека и JK сегмент зародышевой линии JK 5 человека. Совмещение VL последовательности 3F11 с последовательностью зародышевой линии VK L18 показано на фиг.8. Дальнейший анализ VL последовательности 3F11 с использованием системы определения CDR областей по Кабату привел к выявлению CDR1, CDR2 и CDR3 областей легкой цепи, как показано на фиг.1B и 8 и в SEQ ID NО: 13, 17 и 21 соответственно.
Нуклеотидные и аминокислотные последовательности вариабельной области тяжелой цепи 4G5 показаны на фиг.2A и в SEQ ID NО:34 и 26 соответственно.
Нуклеотидные и аминокислотные последовательности вариабельной области легкой цепи 4G5 показаны на фиг.2B и в SEQ ID NО:38 и 30 соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 4G5 с известными иммуноглобулиновыми последовательностями тяжелой цепи зародышевых линий человека продемонстрировало, что в тяжелой цепи 4G5 использован VH сегмент зародышевой линии клеток VH 4-34 человека, неустановленный D сегмент и JH сегмент зародышевой линии JH 4b человека. Совмещение VH последовательности 4G5 с последовательностью зародышевой линии VH 4-34 показано на фиг.6. Дальнейший анализ VH последовательности 4G5 с использованием системы определения CDR областей по Кабату привел к выявлению CDR1, CDR2 и CDR3 областей тяжелой цепи, как показано на фиг.2A и 6 и в SEQ ID NО: 2, 6 и 10 соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 4G5 с известными иммуноглобулиновыми последовательностями легкой цепи зародышевых линий человека продемонстрировало, что в легкой цепи 4G5 использован VL сегмент зародышевой линии клеток VK L18 человека и JK сегмент зародышевой линии JK 2 человека. Совмещение VL последовательности 4G5 с последовательностью зародышевой линии VK L18 показано на фиг.9. Дальнейший анализ VL последовательности 4G5 с использованием системы определения CDR областей по Кабату привел к выявлению CDR1, CDR2 и CDR3 областей легкой цепи, как показано на фиг.2B и 9 и в SEQ ID NО: 14, 18 и 22 соответственно.
Нуклеотидные и аминокислотные последовательности вариабельной области тяжелой цепи 11E2 показаны на фиг.3A и в SEQ ID NО: 35 и 27 соответственно.
Нуклеотидные и аминокислотные последовательности вариабельной области легкой цепи 11E2 показаны на фиг.3B и в SEQ ID NО: 39 и 31 соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 11E2 с известными иммуноглобулиновыми последовательностями тяжелой цепи зародышевых линий человека продемонстрировало, что тяжелая цепь 11E2 является производным или обладает высокой степенью подобия VH сегменту зародышевой линии клеток VH 5-51 человека, неустановленному D сегменту и JH сегменту зародышевой линии JH 4b человека. Совмещение VH последовательности 11E2 с последовательностью зародышевой линии VH 5-51 показано на фиг.7. Дальнейший анализ VH последовательности 11E2 с использованием системы определения CDR областей по Кабату привел к выявлению CDR1, CDR2 и CDR3 областей тяжелой цепи, как показано на фиг.3A и 7 и в SEQ ID NО: 3, 7 и 11 соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 11E2 с известными иммуноглобулиновыми последовательностями легкой цепи зародышевых линий человека продемонстрировало, что в легкой цепи 11E2 использован VL сегмент зародышевой линии клеток VK A27 человека и JK сегмент зародышевой линии JK 5 человека. Совмещение VL последовательности 11E2 с последовательностью зародышевой линии VK A27 показано на фиг.10. Дальнейший анализ VL последовательности 11E2 с использованием системы определения CDR областей по Кабату привел к выявлению CDR1, CDR2 и CDR3 областей легкой цепи, как показано на фиг.3B и 10 и в SEQ ID NО:15, 19 и 23 соответственно.
Нуклеотидные и аминокислотные последовательности вариабельной области тяжелой цепи 9D4 показаны на фиг.4A и в SEQ ID NО:36 и 28 соответственно.
Нуклеотидные и аминокислотные последовательности вариабельной области легкой цепи 9D4 показаны на фиг.4B и в SEQ ID NО:40 и 32 соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 9D4 с известными иммуноглобулиновыми последовательностями тяжелой цепи зародышевых линий человека продемонстрировало, что тяжелая цепь 9D4 является производным или обладает высокой степенью подобия VH сегменту зародышевой линии клеток VH 5-51 человека, неустановленному D сегменту и JH сегменту зародышевой линии JH 4b человека. Совмещение VH последовательности 9D4 с последовательностью зародышевой линии VH 5-51 показано на фиг.7. Дальнейший анализ VH последовательности 9D4 с использованием системы определения CDR областей по Кабату привел к выявлению CDR1, CDR2 и CDR3 областей тяжелой цепи, как показано на фиг.4A и 7 и в SEQ ID NО:4, 8 и 12 соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 9D4 с известными иммуноглобулиновыми последовательностями легкой цепи зародышевых линий человека продемонстрировало, что в легкой цепи 9D4 использован VL сегмент зародышевой линии клеток VK A27 человека и JK сегмент зародышевой линии JK 5 человека. Совмещение VL последовательности 9D4 с последовательностью зародышевой линии VK A27 показано на фиг.10. Дальнейший анализ VL последовательности 9D4 с использованием системы определения CDR областей по Кабату привел к выявлению CDR1, CDR2 и CDR3 областей легкой цепи, как показано на фиг.3B и 10 и в SEQ ID NО:16, 20 и 24 соответственно.
Пример 3: Моноклональные анти-IFNAR-1 антитела человека ингибируют биологическую активность интерферона α2b
Клеточная линия Daudi, полученная из B-лимфобластной лимфомы Беркита человека, экспрессирует высокие уровни IFNAR-1, и рост этих клеток ингибируется интерферонами типа I. Для измерения функциональной блокирующей способности анти-IFNAR-1 антител человека проводили два различных анализа, а именно анализ клеточной пролиферации и репортерный анализ.
В первом анализе клетки Daudi культивировали с интерфероном α2b в присутствии или отсутствие антитела и измеряли пролиферацию по поглощению 3[H]-тимидина. Клетки Daudi (ATCC CCL-213) выращивали в RPMI, содержащей 10% FCS и 2 мМ бета-меркаптоэтанола (среда). Клетки центрифугировали и ресуспендировали при концентрации 1·106 клеток/мл в среде с добавкой 1% альбумина сыворотки человека (среда и HS). К каждой лунке 96-луночного планшета добавляли 100 мкл 200 Ед/мл интерферона α2b (Shering Corporation), содержащего соответствующую концентрацию антитела. В лунки добавляли 100 мкл клеток Daudi в среде и HS и затем планшеты инкубировали в течение 48 часов при 37°C. Планшеты метили 1 мкКюри 3[H]-тимидина и инкубировали в течение еще 24 часов. Удаляли содержимое лунок, собирая его на 96-луночном волокнистом фильтровальном планшете, и производили подсчет радиоактивности с применением сцинтилляционного счетчика TopCount (Packard). Строили график числа отсчетов за минуту как функцию концентрации антитела и анализировали данные с помощью нелинейной регрессии, т.е. сигмоидной зависимости доза-реакция (с переменной крутизной), используя программное обеспечение Prism (San Diego, CA).
Во втором анализе клетки U937 трансфицировали конструктом, в котором стимулируемый интерфероном чувствительный элемент связан с репортерным геном (ISRE-RG), и измеряли способность гуманизированных анти-IFNAR-1 антител блокировать индуцированную интерфероном экспрессию репортерного гена. Клетки выращивали в RPMI, содержащей 10% FCS и 2 мМ бетамеркаптоэтанол (среда). Клетки (1·106 клеток/мл) ресуспендировали в среде с добавкой 2% сыворотки человека. В 96-луночный планшет добавляли по 100 мкл клеток. Антитела последовательно разбавляли в среде, содержащей 200 Ед/мл интерферона α2b (Shering corporation), и в каждую лунку добавляли по 100 мкл. Планшеты инкубировали в течение ночи при 37°C. После этого инкубирования измеряли экспрессию репортерного гена с помощью поточной цитометрии. Строили график средней геометрической интенсивности флуоресценции как функции концентрации антитела и анализировали данные с помощью нелинейной регрессии, т.е. сигмоидной зависимости доза-реакция (с переменной крутизной), используя программное обеспечение Prism (San Diego, CA).
Используя две методики анализа, описанные выше, сравнивали эффективность моноклонального антитела 3F11, мышиного анти-IFNAR-1 антитела 64G12 (ECACC, депозитарный № 92022605) и гуманизированного анти-IFNAR-1 антитела D1 H3K1 (дополнительно описанного в заявке США, порядковый № 60/465058). Антитело 3F11 продемонстрировало в 5-10 раз большую эффективность, чем антитело мыши, и в 6-30 раз большую эффективность, чем гуманизированное антитело. Результаты суммированы ниже в таблице 1.
Блокирующая способность анти-IFNAR-1 антитела человека по отношению к IFN альфа 2b
Пример 4: Моноклональные анти-IFNAR-1 антитела человека ингибируют биологическую активность IFN омега
Исследовали способность анти-IFNAR-1 антитела человека ингибировать реакции IFN омега с использованием анализа пролиферации клеток Daudi, описанного выше в примере 3. В каждую лунку 96-луночного планшета добавляли 100 мкл 200 Ед/мл интерферона омега (PBL), содержащего антитело в соответствующих концентрациях. Антитела человека 3F11, 4G5, 11E2 и 9D4 оказывались в 4-18 раз более эффективными (как измерено по EC50), чем антитело 64G12 мыши. Результаты суммированы ниже в таблице 2.
Блокирующая способность моноклональных анти-IFNAR-1 антител человека по отношению к IFN омега
Пример 5: Моноклональные анти-IFNAR-1 антитела человека ингибируют биологическую активность IFN различных типов I
Как указано в примере 3, интерферон альфа ингибирует пролиферацию клеток Daudi (лимфома Беркита, ATCC #CCL-213) дозозависимым образом. Нейтрализующее антитело, которое блокирует связывание интерферона с его рецептором, будет восстанавливать пролиферацию. Применяя анализ пролиферации клеток, исследовали специфичность очищенных анти-IFN альфа антител человека, тестируя блокирование природного лимфобластоидного IFNα, природного лейкоцитарного интерферона, 13 подтипов рекомбинантного IFN альфа, IFN бета и IFN омега.
Клетки Daudi выращивали в культурной среде (RPMI 1640, с добавками 10% FCS, 1x2-ME, L-глутамина, пенициллина и стрептомицина) с добавкой и без добавки IFNα в 96-луночном плоскодонном планшете для культивирования клеток. Каждый из протестированных интерферонов типа I исследовали при EC50 и смешивали с 2-кратными последовательными разбавлениями анти-IFNAR-1 антитела 3F11, как правило, от 50 мкг/мл (312 нМ) до 381 пг/мл (2,4 пМ). Смесь антитело/IFN добавляли к клеткам в Daudi 96-луночном планшете с дном до конечной плотности 1·104 клеток Daudi/100 мкл/лунка, и инкубировали при 37°C, в атмосфере 5% CO2 в течение 72 часов. Пролиферацию анализировали добавлением MTS (Promega), 20 мкл/лунка, и измеряли OD при 490 нм с дальнейшей 3-часовой инкубацией. Количество жизнеспособных клеток было пропорционально зарегистрированному значению OD. Рассчитывали процент блокирования интерферона по отношению к пролиферации клеток Daudi в отсутствие IFN (=100% блокирования) и в присутствии только IFN (=0% блокирования). Результаты для антитела 3F11 рассчитывали в соответствии со степенью блокирования, получая профиль специфичности подтипа IFNα. Результаты продемонстрировали, что антитело 3F11 человека против рецептора 1 интерферона альфа ингибирует действие многочисленных подтипов интерферона альфа, включая IFNα 6, 2b, 2a, 1, 16, 10, 8, 5, 14, 17, 7, 4 и 21, а также лимфобластоидного IFN, IFN лейкоцитов и IFN омега. 3F11 является ингибитором IFN бета более низкого уровня, хотя наблюдалось более чем 50% ингибирование. Процент блокирования и среднее квадратичное отклонение для различных интерферонов показано ниже в таблице 3.
Ингибирование различных интерферонов типа I антителом 3F11
Пример 6: Ингибирование индуцированной IFN секреции IP-10 анти-IFNAR-1 антителами
Было показано, что добавление IFN альфа 2b к клеточной культурной среде индуцирует секрецию IP-10 нормальными моноядерными клетками периферической крови (PBMNC). Исследовали активность анти-IFNAR-1 антитела 3F11 человека в ингибировании индуцированной интерфероном секреции IP-10 нормальными культурами PBMNC с помощью анализа связывания способом ELISA.
Клетки PBMNC инкубировали в культурной среде (RPMI 1640+10% FBS+1% сыворотка человека) с лейкоцитарный IFN, IFN альфа 2b или IFNω в течение 24-48 часов.
Супернатанты собирали и анализировали на концентрацию IP-10/CXCL10, используя комплект для количественного сэндвич-анализа ELISA (Quantikine®, R&D Systems) в разбавлении 1:30 согласно рекомендациям производителя. Результаты анализа продемонстрировали, что моноклональные антитела человека 3F11 ингибируют индуцированную лейкоцитарным IFN, рекомбинантным IFNα 2b и рекомбинантным IFNω секрецию IP-10 нормальной культурой PBMNC. Эти результаты показаны в таблице 4.
Ингибирование IFN-индуцированной экспрессии IP-10 на нормальных PBMNC антителами
антитело
IP-10 (пг/мл)
IFN
IP-10 (пг/мл)
IP-10 (пг/мл)
IP-10 (пг/мл)
Пример 7: Анализ перекрестной конкуренции моноклональных анти-IFNAR-1 антител человека
Чтобы определить, связываются ли моноклональные антитела человека с тем же эпитопом, что и моноклональное антитело 64G12 мыши, проводили анализ перекрестной конкуренции с помощью ELISA для определения, конкурируют ли указанные антитела за один связывающий эпитоп.
96-луночные планшеты покрывали растворимым IFNAR-1 человека, полученным из клеток CHO, в концентрации 1 мкг/мл в свежеприготовленном DPBS в количестве 100 мкл/лунка (Mediatech). Моноклональные антитела человека 3F11, 4G5, 11E2 и 9D4 в концентрации 20 мкг/мл добавляли в ряд лунок 1 и последовательно разбавляли в соотношении 1:2 в лунках, относящихся к рядам от 1 до 12, с последующим инкубированием в течение 45 минут. Добавляли моноклональное антитело 64G12 мыши при концентрации EC75, равной 0,3 мкг/мл, в количестве 50 мкл на лунку и инкубировали планшеты в течение 30 минут. Планшеты 3 раза промывали c помощью автоматического устройства для промывки планшетов Elx 405 (BIO-TEK Instruments). Прошедшее аффинную очистку F(ab')2 козье антимышиное IgG (Fcγ специфичное) антитело с пероксидазой разбавляли в соотношении 1:3000 в PBS и добавляли в качестве конъюгата для детектирования (Jackson ImmunoResearch Laboratories, cat. 115-036-0710). После инкубирования в течение 1 часа планшеты 3 раза промывали c помощью автоматического устройства для промывки планшетов Elx 405. Раствор ABTS (800 мкл исходного раствора ABTS, 8 мкл 30% H2O2 и 100 мл цитратно-фосфатного буфера) в концентрации 27,8 мг/мл добавляли в каждую лунку и инкубировали в течение 20 минут. Планшеты считывали при 415 нм, используя 490 нм в качестве контрольной длины волны. Результаты приведены на фиг.11. Результаты показывают, что моноклональные анти-IFNAR-1 антитела человека, а именно 3F11, 4G5, 11E2 и 9D4, не конкурируют с 64G12 за связывание с IFNAR-1 и, таким образом, связываются с другим эпитопом на IFNAR-1, чем 64G12.
Пример 8: Ингибирование антителами развития дендритных клеток, опосредованного SLE плазмой
SLE плазма индуцирует развитие дендритных клеток из нормальных моноцитов человека. В этом примере очищенное моноклональное анти-IFNAR-1 антитело человека 3F11 испытывали на ингибирование развития дендритных клеток, которое оценивали по способности антител ингибировать индукцию маркеров клеточной поверхности CD38, MHC класса I и CD123 под действием SLE плазмы.
25 мл лейкоцитарной пленки разбавляли PBS в 4 раза. Образец разделяли по 4·50-мл коническим пробиркам и под них вводили в виде слоя 15 мл среды для фракционирования лимфоцитов (ICN Biomedicals). После 30-мин вращения при 500g лейкоцитарный слой, содержащий PBMC, удаляли и промывали PBS. Клетки ресуспендировали в культурной среде в количестве 4·106 клеток/мл. Моноциты изолировали инкубированием PBMC (2,0·107 клеток/5 мл/25 см3 колбу) в течение 1,5 часа при 37°C в культурной среде с последующим двукратным вымыванием неадгезивных клеток. После второго промывания клетки культивировали в среде, содержащей 1% сыворотку человека, инактивированную нагреванием. В колбы с культурами добавляли двадцатипятипроцентную плазму SLE пациентов плюс/минус нейтрализующие антитела и контрольный изотип (30 мкг/мл); смесь IFN альфа 2b (100 и 10 МЕ/мл) и 25% нормальной плазмы человека использовали в качестве положительного контроля индукции маркеров. Колбы инкубировали при 37°C в атмосфере 5% CO2 от трех до семи дней. Из каждой колбы собирали кондиционированную среду и выделяли суспендированные клетки центрифугированием при 1000 об/мин на роторе Sorvall RTH-750. Осажденные клетки сохраняли на льду и слой жидкости замораживали при -80°C для ELISA.
Адгезивные клетки выделяли из колбы промыванием PBS (2 мл) с последующим 15-минутным инкубированием в версене (ЭДТУ) (3 мл) в случае необходимости. В конце инкубирования с версеном колбу отскабливали и в заключение промывали PBS (2 мл). PBS и версен вместе со смытыми клетками объединяли с клетками, выделенными из собранной кодиционированной среды. Объединенную суспензию клеток центрифугировали при 1000 об/мин на роторе Sorvall RTH-750, полученный осадок ресуспендировали до 300 мкл в окрашивающем буфере (PBS + 0,1М ЭДТУ + 2% FBS +1% HS) и распределяли по 100 мкл/лунка в 96-луночный планшет с V-образным дном. Планшет подвергали импульсному центрифугированию при 2800 об/мин на роторе Sorvall RTH-750 и осажденные клетки ресуспендировали по 25 мкл/лунка в меченных флуорохромом антителах следующим образом: (1) мышиные анти-MHC I-FITC + мышиные анти-CD38-PE и (2) контрольный изотип мышиные IgG-FITC+мышиный IgG-PE. Защищенные от света планшеты инкубировали на льду в течение 45 минут. Клетки промывали три раза добавлением 200 мкл окрашивающего буфера с последующим импульсным центрифугированием и затем ресуспендировали в 200 мкл 2% параформальдегида в PBS. Окрашивание дендритных клеток анализировали поточной цитометрией с использованием Becton Dickinson FACScaliburTM. На графике переднее рассеяние - боковое рассеяние были начерчены окна для удаления загрязняющих клеток из анализа. Моноклональное анти-IFNAR-1 антитело человека ингибирует IFN альфа-зависимый процесс развития дендритных клеток, что продемонстрировано с помощью нормализованной экспрессии маркеров клеточной поверхности MHC класса I, CD38 и CD123 в присутствии 3F11. Результаты показаны ниже в таблице 5, где части таблицы (A) и (B) суммируют результаты для образцов двух представителей доноров SLE.
Ингибирование созревания дендритных клеток
(A)
Плазма донора №40* (13,3 МЕ/мл**)
Плазма донора №59* (75,3 МЕ/мл**)
Пример 9: Анализ связывания анти-IFNAR-1 антител человека с клетками Daudi или человеческими моноядерными клетками периферической крови по Scatchard
Моноядерные клетки периферической крови человека получали из свежей крови по стандартным протоколам, используя разделение на ступенчатом градиенте Ficol. Клетки Daudi получали из ATCC и выращивали в RPMI, содержащей 10% фетальную телячью сыворотку (FBS). Клетки дважды промывали RPMI, содержащей 10% FBS при 4 градусах, и количество клеток доводили до 4·107 клеток/мл в среде RPMI, содержащей 10% фетальной телячьей сыворотки (связывающий буфер). Планшеты Millipore (MAFB NOB) покрывали 1% обезжиренным сухим молоком в воде и хранили при 4°C в течение ночи. Планшеты промывали связывающим буфером и добавляли 25 мкл немеченого антитела (1000-кратный избыток) в связывающем буфере в контрольные лунки 96-луночного фильтровального планшета из стекловолокна Millipore (неспецифическое связывание NSB). Двадцать пять микролитров чистого буфера добавляли в контрольную лунку с максимальным связыванием (полное связывание). Добавляли двадцать пять микролитров 125I-анти-IFNAR-1 антитела в изменяющейся концентрации и 25 мкл клеток Daudi или моноядерных клеток периферической крови человека (4·107 клеток/мл) в связывающем буфере. Планшеты инкубировали в течение 2 часов на шейкере при 200 об/мин при 4°C. По завершении инкубации планшеты Millipore дважды промывали 0,2 мл холодного связывающего буфера. Фильтры удаляли и подсчитывали на гамма-счетчике. Определение равновесного связывания проводили с использованием параметров одного сайта связывания с помощью программного обеспечения Prism (San Diego, CA).
По результатам описанного выше анализа связывания по Scatchard, значения KD антитела для клеток Daudi и для моноядерных клеток периферической крови человека составляли приблизительно 0,2 нМ и 0,5 нМ соответственно.
Перечень последовательностей

















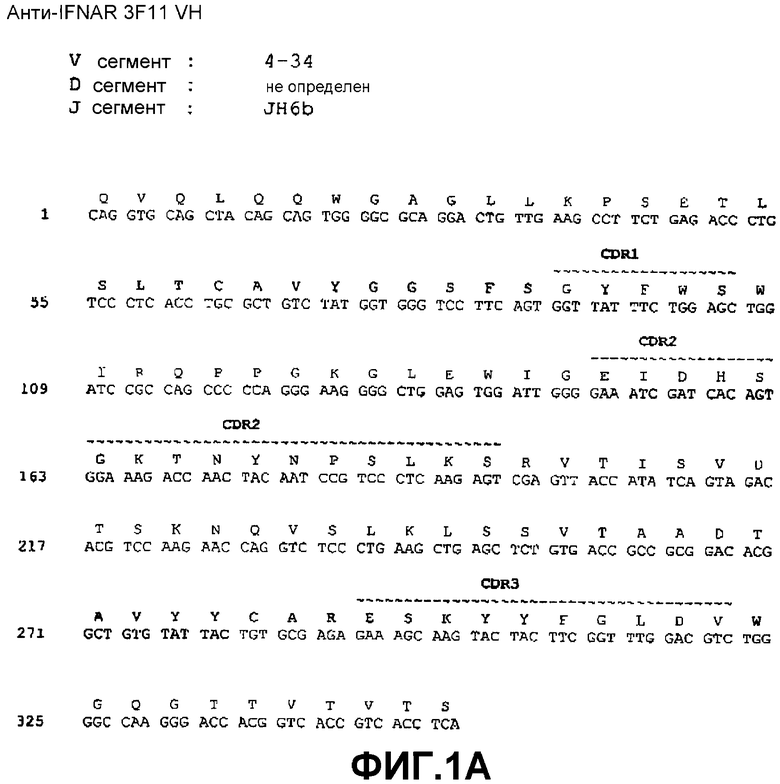




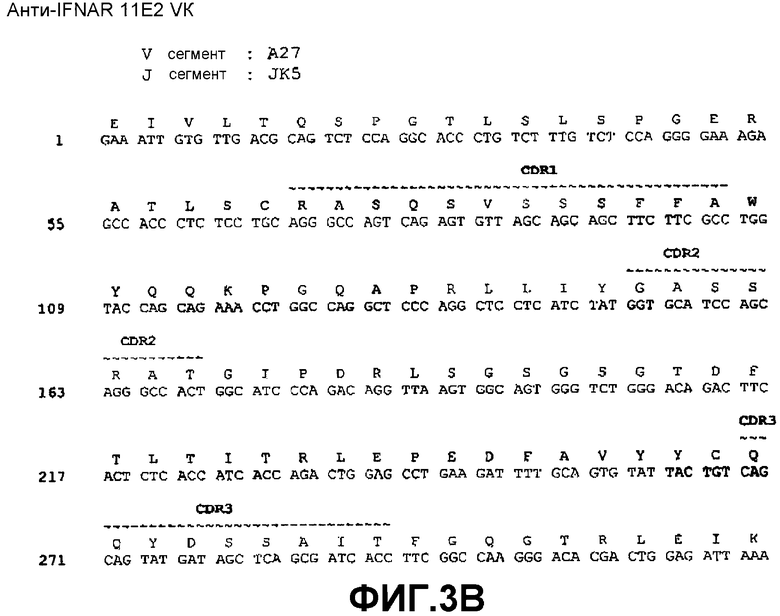


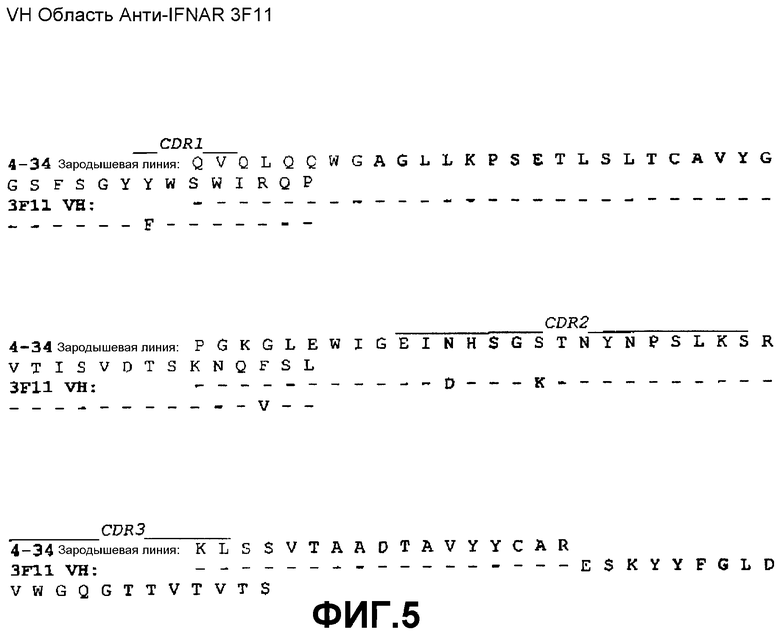
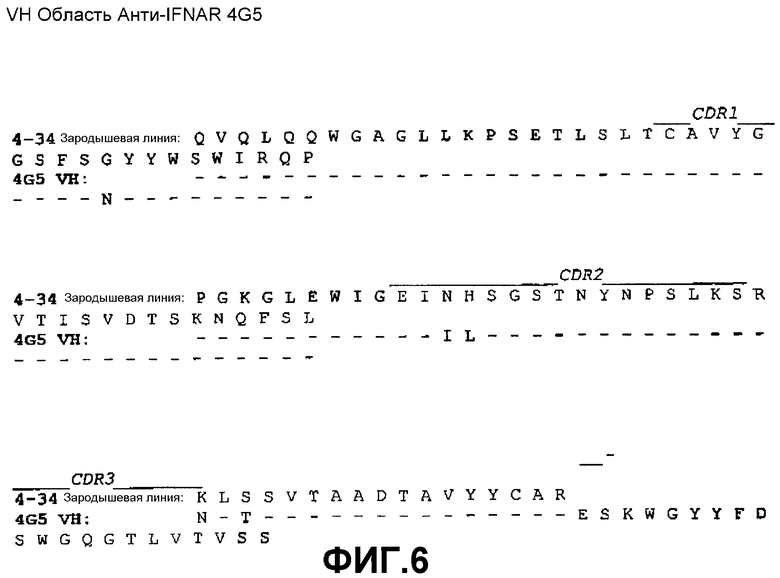
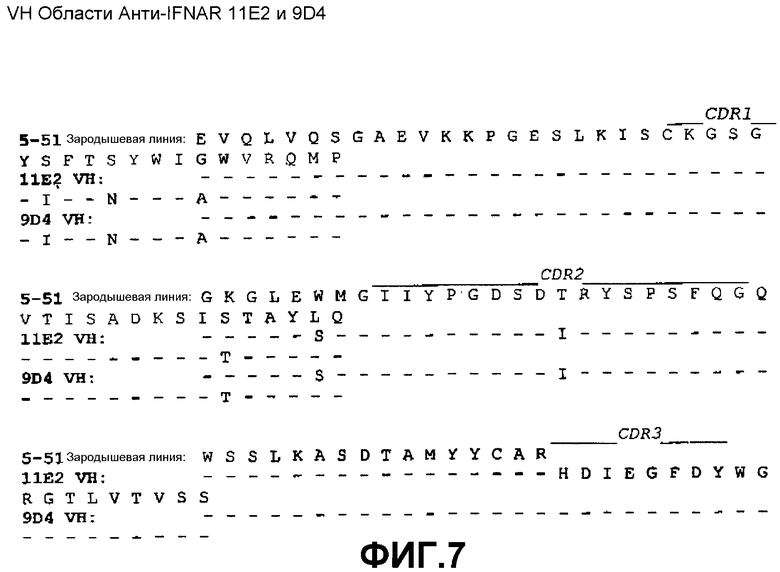
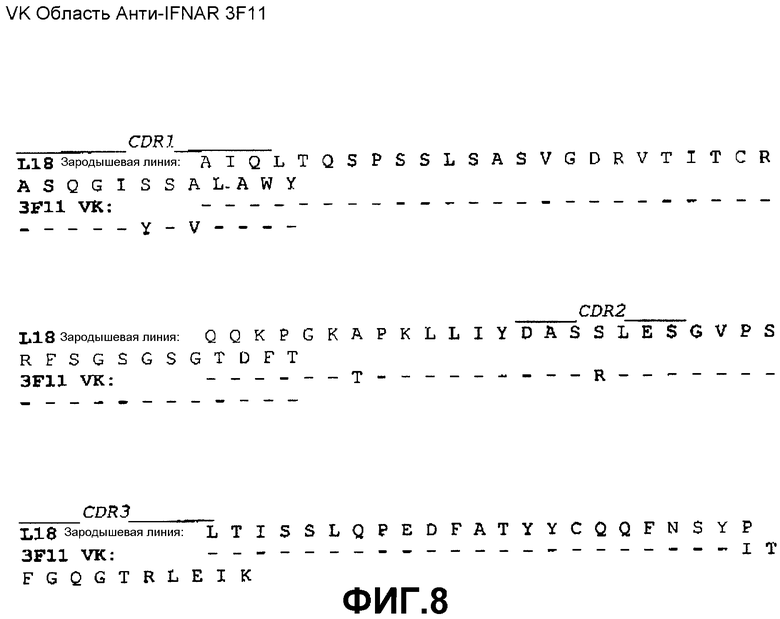
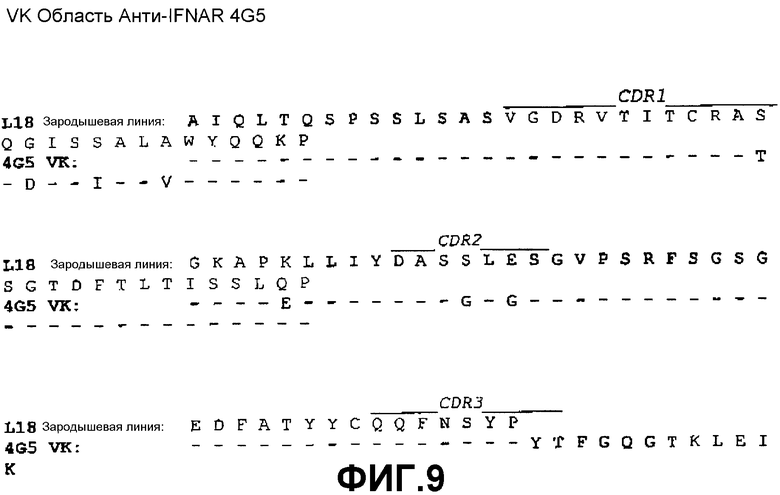
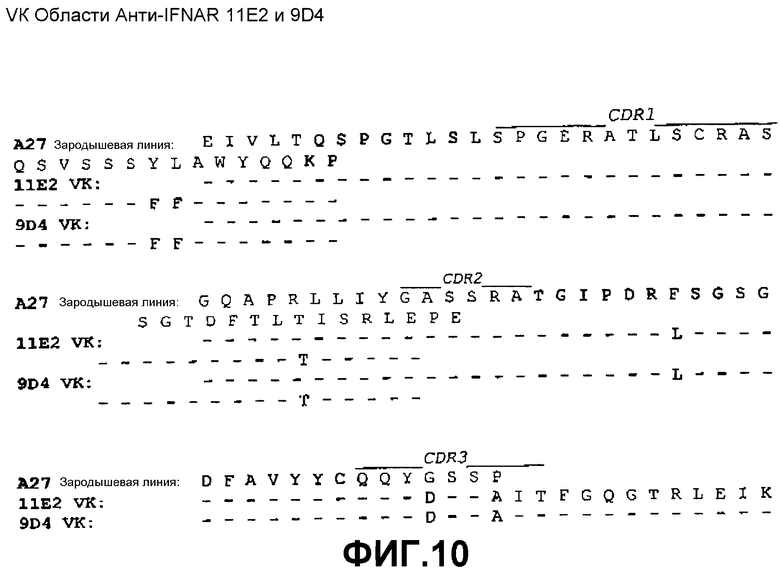
Настоящее изобретение относится к иммунологии и биотехнологии. Предложены варианты выделенного моноклонального антитела, специфичного к IFNAR-1. Описаны: биспецифичная молекула, иммуноконъюгат и композиции для лечения заболеваний и расстройств, опосредованных IFNAR-1 на основе моноклонального антитела. Раскрыты также способы ингибирования биологической активности интерферона типа I, способ лечения заболеваний или расстройств, опосредованных интерфероном типа I, с использованием антитела. Предложены: кодирующая антитело нуклеиновая кислота, вектор для экспрессии антитела, клетка, трансформированная вектором, а также способ получения антител и гибридома, продуцирующая антитело. Использование изобретения обеспечивает новые ингибирующие IFNAR-1 антитела, которые блокируют IFNAR-1 и связывают иной его эпитоп, по сравнению с известным антителом 64G12, что может найти дальнейшее применение в медицине. 15 н. и 14 з.п. ф-лы, 11 ил., 6 табл.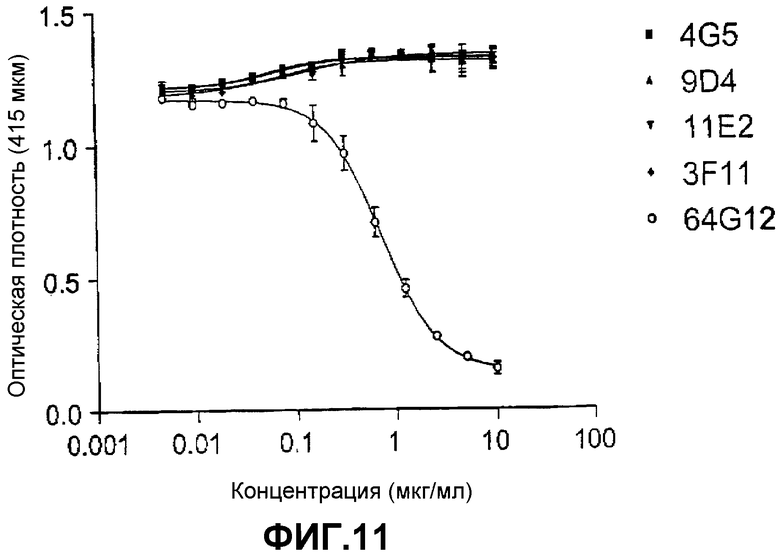
1. Выделенное моноклональное антитело человека, которое специфично связывается с рецептором 1 интерферона альфа человека (IFNAR-1), или его антигенсвязывающий участок, содержащее:
(a) вариабельную область тяжелой цепи CDR1 человека, содержащую SEQ ID NO: 4;
(b) вариабельную область тяжелой цепи CDR2 человека, содержащую SEQ ID NO: 8;
(c) вариабельную область тяжелой цепи CDR3 человека, содержащую SEQ ID NO: 12;
(d) вариабельную область легкой цепи CDR1 человека, содержащую SEQ ID NO: 16;
(e) вариабельную область легкой цепи CDR2 человека, содержащую SEQ ID NO: 20; и
(f) вариабельную область легкой цепи CDR3 человека, содержащую SEQ ID NO: 24.
2. Выделенное моноклональное антитело человека, которое специфично связывается с рецептором 1 интерферона альфа человека (IFNAR-1), или его антигенсвязывающий участок, содержащее:
(a) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность SEQ ID NO: 28; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность SEQ ID NO: 32.
3. Антитело человека по любому из пп.1 и 2 или 26-32, которое не связывается с тем же самым эпитопом, что и моноклональное антитело 64G12 мыши (депозитарный номер ЕСАСС №92022605).
4. Композиция для лечения заболеваний и расстройств, опосредованных интерфероном типа I, содержащая терапевтически эффективное количество антитела или его антигенсвязывающий участок по любому из пп.1 и 2 или 26-32, и фармацевтически приемлемый носитель.
5. Иммуноконъюгат для лечения заболеваний и расстройств, опосредованных интерфероном типа I, содержащий антитело или его антигенсвязывающий участок по любому из пп.1 и 2 или 26-32, связанный с терапевтическим средством.
6. Иммуноконъюгат по п.5, дополнительно содержащий фармацевтически приемлемый носитель.
7. Иммуноконъюгат по п.5, в котором терапевтическое средство является цитотоксином.
8. Иммуноконъюгат по п.7, дополнительно содержащий фармацевтически приемлемый носитель.
9. Иммуноконъюгат по п.5, в котором терапевтическое средство представляет собой радиоактивный изотоп.
10. Иммуноконъюгат по п.9, дополнительно содержащий фармацевтически приемлемый носитель.
11. Биспецифичная молекула для лечения заболеваний и расстройств, опосредованных интерфероном типа I, содержащая антитело или его антигенсвязывающий участок по любому из пп.1 и 2 или 26-32, связанная со вторым функциональным фрагментом, имеющим другую специфичность связывания по сравнению с указанным антителом или его антигенсвязывающим участком.
12. Композиция, содержащая терапевтически эффективное количество биспецифичной молекулы по п.11 и фармацевтически приемлемый носитель, для лечения заболеваний и расстройств, опосредованных интерфероном типа I.
13. Выделенная молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающий участок по любому из пп.1 и 2 или 26-32.
14. Вектор экспрессии, включающий молекулу нуклеиновой кислоты по п.13.
15. Клетка-хозяин, содержащая вектор экспрессии по п.14.
16. Антитело по любому из пп.1, 2, 3 или 27-33, полученное путем иммунизации рекомбинантным белком IFNAR-1 трансгенной мыши НСо7, НСо12 и НСо7×НСо12, несущее трансгены тяжелой и легкой цепей иммуноглобулина человека.
17. Гибридома, продуцирующая антитело по любому из пп.1, 2, 3 и 26-32, причем гибридома получена из трансгенной мыши НСо7, НСо12 и НСо7×НСо12.
18. Способ получения анти-IFNAR-1 антитела по любому из пп.1, 2 или 26-32, включающий:
(a) получение:
(i) последовательности вариабельной области тяжелой цепи антитела, содержащей последовательность CDR1 SEQ ID NO: 4, последовательность CDR2 SEQ ID NO: 8; и последовательность CDR3 SEQ ID NO: 12; или (ii) последовательности вариабельной области легкой цепи антитела, содержащей последовательность CDR1 SEQ ID NO: 16, последовательность CDR2 SEQ ID NO: 20, а также последовательность CDR3 SEQ ID NO: 24;
(b) изменение по меньшей мере одного аминокислотного остатка по меньшей мере одной вариабельной области последовательности антитела, причем упомянутая последовательность выбрана из последовательности вариабельной области тяжелой цепи антитела и последовательности вариабельной области легкой цепи антитела, для создания хотя бы одной измененной последовательности антитела; и
(с) экспрессию измененной последовательности антитела в виде белка, где антитело ингибирует биологическую активность IFN-β; ингибирует активность IFN-α2b в анализе пролиферации клеток Daudi; ингибирует активность IFN омега в анализе пролиферации клеток Daudi; ингибирует секрецию IP-10 мононуклеарными клетками периферической крови, индуцированную IFN-α2b; ингибирует секрецию IP-10 мононуклеарными клетками периферической крови, вызванную IFN омега; ингибирует развитие дендритных клеток, опосредованное плазмой пациента с системной красной волчанкой; и связывается с эпитопом, отличным от эпитопа связывания моноклонального антитела 64G12 мыши (депозитарный номер ЕСАСС №92022605).
19. Способ ингибирования биологической активности интерферона типа I на клетке, экспрессирующей рецептор 1 интерферона альфа, включающий взаимодействие клетки с антителом по любому из пп.1, 2 или 26-32, с тем чтобы ингибировать биологическую активность интерферона типа I.
20. Способ лечения заболеваний или расстройств, опосредованных интерфероном типа I, у индивидуума, включающий введение индивидууму антитела или его антигенсвязывающего участка, по любому из пп.1, 2 или 26-32, для лечения заболевания, опосредованного интерфероном типа I, у индивидуума.
21. Способ по п.20, где заболевание, опосредованное интерфероном типа I, представляет собой заболевание, опосредованное интерфероном альфа.
22. Способ по п.20, где заболевание или расстройство представляет собой системную красную волчанку.
23. Способ по п.20, где заболевание или расстройство выбрано из группы, состоящей из инсулинзависимого сахарного диабета, воспалительного заболевания кишечника, рассеянного склероза, псориаза, аутоиммунного тиреоидита, ревматоидного артрита и гломерулонефрита.
24. Способ по п.20, где заболевание или расстройство представляет собой ВИЧ-инфекцию или СПИД.
25. Способ по п.20, где заболевание или расстройство представляет собой отторжение трансплантата или реакцию «трансплантат против хозяина».
26. Выделенное моноклональное антитело человека, которое связывается с рецептором 1 интерферона альфа человека (IFNAR-1), или его антигенсвязывающий участок, содержащее вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, где:
(a) последовательность CDR3 вариабельной области тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 12 и консервативные модификации указанной последовательности; и
(b) последовательность CDR3 вариабельной области легкой цепи содержит аминокислотную последовательность SEQ ID NO: 24 и консервативные модификации указанной последовательности.
27. Антитело по п.26, где последовательность CDR2 вариабельной области тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 8 и консервативные модификации указанной последовательности;
последовательность CDR2 вариабельной области легкой цепи содержит аминокислотную последовательность SEQ ID NO: 20 и консервативные модификации указанной последовательности.
28. Антитело по п.27, где последовательность CDR1 вариабельной области тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 4 и консервативные модификации указанной последовательности; последовательность CDR1 вариабельной области легкой цепи содержит аминокислотную последовательность SEQ ID NO: 16 и консервативные модификации указанной последовательности.
29. Выделенное моноклональное антитело человека, которое связывается с рецептором 1 интерферона альфа человека (IFNAR-1), или его антигенсвязывающий участок, в котором:
(a) вариабельная область тяжелой цепи содержит аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 28; и
(b) вариабельная область легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 32.
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| GOLDMAN et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| J Interferon Cytokine Res | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| RU 94045826 A1, 10.11.1996. | |||

