Перекрестная ссылка на родственные заявки
Настоящее изобретение является национальной фазой международной заявки РСТ/JP06/0309606, поданной 2 мая 2006, по которой испрашивается приоритет предварительной патентной заявки США No. 60/679466, поданной 9 мая 2005, предварительной патентной заявки США No. 60/738434, поданной 21 ноября 2005, и предварительной патентной заявки США No. 60/748919, поданной 8 декабря 2005, каждая из которых приведена в описании в качестве ссылки в полном объеме.
Перечень последовательностей
В приложении к описанию также представлен перечень последовательностей, направленный посредством EFS 31 октября 2007 согласно 37 C.R.F. § I.52(e)(5); текстовой файл перечня последовательностей обозначен 077375,0499SEQLISTING.txt имеет размер 48642 байта и создан 3 октября 2007. Перечень последовательностей, поданный в электронной версии, не ограничивает рамки описания и не содержит новых объектов.
Область техники, к которой относится изобретение
Настоящее изобретение главным образом относится к иммунотерапии при лечении заболевания человека и уменьшению ее побочных эффектов. Более конкретно, настоящее изобретение относится к применению анти-PD-1-антител и к применению комбинированной иммунотерапии, включая комбинацию анти-CTLA-4 и анти-PD1-антител, для лечения рака и/или для уменьшения частоты или тяжести побочных эффектов, связанных с лечением такими отдельно используемыми антителами.
Уровень техники
Белок программируемой смерти 1 (PD-1) является ингибиторным членом семейства рецепторов CD28, которое включает в себя также CD28, CTLA-4, ICOS и BTLA. PD-1 экспрессируется активированными В-клетками, Т-клетками и миелоидными клетками (Agata et al., supra; Okazaki et al. (2002) Curr. Opin. Immunol. 14: 391779-82; Bennet et al. (2003) J Immunol 170:711-8). Первоначальные члены данного семейства, CD28 и ICOS, были обнаружены по функциональным действиям на увеличение пролиферации Т-клеток после добавления моноклональных антител (Hutloff et al. (1999) Nature 397:263-266; Hansen et al. (1980) Immunogenics 10:247-260). PD-1 был обнаружен скринингом на дифференциальную экспрессию в апоптотических клетках (Ishida et al. (1992) EMBO J 11:3887-95). Другие члены данного семейства, CTLA-4 и BTLA, были обнаружены скринингом на дифференциальную экспрессию в цитотоксических Т-лимфоцитах и TH1-клетках, соответственно. CD28, ICOS и CTLA-4, все, имеют неспаренный остаток цистеина, дающий возможность гомодимеризации. В противоположность этому, предполагается, что PD-1 существует в виде мономера, не имея неспаренного остатка цистеина, характерного для других членов семейства CD28.
Ген PD-1 является трансмембранным белком типа I 55 кДа, который является частью суперсемейства генов Ig (Agata et al. (1996) Int Immunol 8:765-72). PD-1 содержит мембранопроксимальный иммунорецепторный ингибирующий мотив на основе тирозина (ITIM) и мембранодистальный мотив переключения на основе тирозина (ITSM) (Thomas, M.L. (1995) J Exp Med 181:1953-6; Vivier, E и Daeron, M (1997) Immunol Today 18:286-91). PD-1, хотя и является структурно сходным с CTLA-4, лишен мотива MYPPPY, который является критическим для связывания B7-1 и B7-2. Были идентифицированы два лиганда для PD-1, PD-L1 и PD-L2, которые, как было показано, отрицательно регулируют активацию Т-клеток после связывания с PD-1 (Freeman et al. (2000) J Exp Med 192:1027-34; Latchman et al. (2001) Nat Immunol 2:261-8; Carter et al. (2002) Eur J Immunol 32:634-43). Как PD-L1, так и PD-L2 являются гомологами B7, которые связываются с PD-1, но не связываются с другими членами семейства CD28. Один лиганд для PD-1, PD-L1, является изобилующим в различных типах рака человека (Dong et al. (2002) Nat. Med. 8:787-9). Взаимодействие между PD-1 и PD-L1 приводит к уменьшению инфильтрирующих опухоль лимфоцитов, уменьшению опосредованной рецептором Т-клеток пролиферации и ускользанию от иммунологического надзора раковых клеток (Dong et al. (2003) J. Mol. Med. 81:281-7; Blank et al. (2005) Cancer Immunol. Immunother. 54:307-314; Konishi et al. (2004) Clin. Cancer Res. 10:5094-100). Иммуносупрессия может быть обращена ингибированием локального взаимодействия PD-1 с PD-L1, и это действие является аддитивным при блокировании взаимодействия PD-1 с PD-L2 (Iwai et al. (2002) Proc. Nat'l. Acad. Sci. USA 99:12293-7; Brown et al. (2003) J. Immunol. 170:1257-66).
PD-1 является ингибирующим членом семейства CD28, экспрессируемым на активированных B-клетках, T-клетках и миелоидных клетках (Agata et al., supra; Okazaki et al. (2002) Curr Opin Immunol 14: 391779-82; Bennett et al. (2003) J Immunol 170:711-8). PD-1-недостаточные животные развивают различные аутоиммунные фенотипы, включая аутоиммунную кардиопатию и подобный волчанке синдром с артритом и нефритом (Nishimura et al. (1999) Immunity 11:141-51; Nishimura et al. (2001) Science 291:319-22). Кроме того, было обнаружено, что PD-1 играет роль в аутоиммунном энцефаломиелите, системной красной волчанке, болезни трансплантат против хозяина (GVHD), диабете типа I и ревматоидном артрите (Salama et al. (2003) J Exp Med 198:71-78; Prokunina and Alarcon-Riquelme (2004) Hum Mol Genet 13:R143; Nielsen et al. (2004) Lupus 13:510). Было показано, что в линии мышиных опухолевых В-клеток ITSM PD-1 является необходимым для блокирования BCR-опосредованного вхождения Ca2+ и фосфорилирования тирозина, находящихся ниже по ходу процесса эффекторных молекул (Okazaki et al. (2001) PNAS 98:13866-71).
Таким образом, желательными являются агенты, которые распознают PD-1, и способы применения таких агентов.
Описание изобретения
Настоящее изобретение обеспечивает выделенные моноклональные антитела, в частности, моноклональные антитела человека, которые связываются с PD-1 и которые проявляют многочисленные желаемые свойства. Такие свойства включают, например, высокую аффинность связывания с PD-1 человека, но отсутствие существенной перекрестной реактивности с CD28, CTLA-4 или ICOS человека. Кроме того, было показано, что антитела настоящего изобретения модулируют иммунные реакции. Таким образом, другой аспект настоящего изобретения относится к способам модуляции иммунных реакций с использованием анти-PD-1-антител. В частности, настоящее изобретение обеспечивает способ ингибирования роста опухолевых клеток in vivo с использованием анти-PD-1-антител.
В одном аспекте настоящее изобретение относится к выделенному моноклональному антителу или его антигенсвязывающей части, где данное антитело проявляет по меньшей мере одно из следующих свойств:
(а) связывается с PD-1 человека с KD 1×10-7М или менее;
(b) по существу не связывается с CD28, CTLA-4 или ICOS человека;
(с) увеличивает пролиферацию Т-клеток в анализе реакции лимфоцитов в смешанной культуре (MLR);
(d) увеличивает продуцирование интерферона-гамма в анализе MLR;
(е) увеличивает секрецию интерлейкина-2 (IL-2) в анализе MLR;
(f) связывается с PD-1 человека и PD-1 собакоподобной обезьяны;
(g) ингибирует связывание PD-L1 и/или PD-L2 с PD-1;
(h) стимулирует антигенспецифические ответные реакции памяти;
(i) стимулирует ответные реакции антител;
(j) ингибирует рост опухолевых клеток in vivo.
Предпочтительно, данное антитело является антителом человека, хотя в альтернативном варианте осуществления данное антитело может быть мышиным антителом, химерным антителом или гуманизированным антителом.
В более предпочтительных вариантах осуществления данное антитело связывается с PD-1 человека с KD 5×10-8М или менее, связывается с PD-1 человека с KD 1×10-8М или менее, связывается с PD-1 человека с KD 5×10-9М или менее, или связывается с PD-1 человека с KD 1×10-8М - KD 1×10-10М.
В другом варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее:
(а) вариабельную область тяжелой цепи человека, которая содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 2, 3, 4, 5, 6 и 7; и
(b) вариабельную область легкой цепи человека, которая содержит аминокислотную последовательность, состоящую из SEQ ID NO:8, 9, 10, 11, 12, 13 и 14;
В различных вариантах осуществления ссылочное антитело содержит:
(а) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:1; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:8;
или ссылочное антитело содержит:
(а) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:2; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:9;
или ссылочное антитело содержит:
(а) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:3; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:10;
или ссылочное антитело содержит:
(а) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:4; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:11;
или ссылочное антитело содержит:
(а) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:5; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:12;
или ссылочное антитело содержит:
(а) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:6; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:13;
или ссылочное антитело содержит:
(а) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:7; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность SEQ ID NO:14.
В другом аспекте настоящее изобретение относится к выделенному моноклональному антителу или его антигенсвязывающей части, содержащему вариабельную область тяжелой цепи, которая является продуктом или получена из гена VH 3-33, где данное антитело специфически связывает PD-1. Кроме того, настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область тяжелой цепи, которая является продуктом или получена из гена VH 4-39, где данное антитело специфически связывает PD-1. Кроме того, настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область тяжелой цепи, которая является продуктом или получена из гена VK L6, где данное антитело специфически связывает PD-1. Кроме того, настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область легкой цепи, которая является продуктом или получена из гена VK L15, где данное антитело специфически связывает PD-1.
В предпочтительном варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее:
(а) вариабельную область тяжелой цепи гена VH 3-33 человека; и
(b) вариабельную область легкой цепи гена VK L6 человека;
где данное антитело специфически связывается с PD-1.
В другом предпочтительном варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее:
(а) вариабельную область тяжелой цепи гена VH 4-39 человека; и
(b) вариабельную область легкой цепи гена VK L15 человека;
где данное антитело специфически связывается с PD-1.
В другом аспекте настоящее изобретение обеспечивает выделенное моноклональное антитело человека, или его антигенсвязывающую часть, содержащее:
вариабельную область тяжелой цепи, которая содержит последовательности CDR1, CDR2 и CDR3; и
вариабельную область легкой цепи, которая содержит последовательности CDR1, CDR2 и CDR3, где
(а) последовательность CDR3 вариабельной области тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:29, 30, 31, 32, 33, 34 и 35, и их консервативных модификаций;
(b) последовательность CDR3 вариабельной области легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:50, 51, 52, 53, 54, 55 и 56, и их консервативных модификаций; и
(с) данное антитело специфически связывается с PD-1 человека.
Предпочтительно, последовательность CDR2 вариабельной области тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:22, 23, 24, 25, 26, 27 и 28, и их консервативных модификаций; и последовательность CDR2 вариабельной области легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:43, 44, 45, 46, 47 и 49, и их консервативных модификаций. Предпочтительно, последовательность CDR1 вариабельной области тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:15, 16, 17, 18, 19, 20 и 21, и их консервативных модификаций; и последовательность CDR1 вариабельной области легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:36, 37, 38, 39, 40, 41 и 42, и их консервативных модификаций.
Еще в одном аспекте настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(а) вариабельная область тяжелой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:1, 2, 3, 4, 5, 6 и 7;
(b) вариабельная область легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:8, 9, 10, 11, 12, 13 и 14;
(с) данное антитело связывается с PD-1 человека с KD 1×10-7М или менее; и
(d) данное антитело по существу не связывается с CD28, CTLA-4 или ICOS человека.
В предпочтительном варианте осуществления данные антитела дополнительно обладают по меньшей мере одним из следующих свойств:
(а) данное антитело увеличивает пролиферацию Т-клеток в анализе MLR;
(b) данное антитело увеличивает продуцирование интерферона-гамма в анализе MLR или
(с) данное антитело увеличивает секрецию IL-2 в анализе MLR.
Дополнительно или альтернативно, данное антитело может обладать одним или несколькими другими признаками, перечисленными выше.
В предпочтительном варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее:
(а) CDR1 вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:15, 16, 17, 18, 19, 20 и 21;
(b) CDR2 вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:22, 23, 24, 25, 26, 27 и 28;
(с) CDR3 вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:29, 30, 31, 32, 33, 34 и 35;
(d) CDR1 вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:36, 37, 38, 39, 40, 41 и 42;
(е) CDR2 вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:43, 44, 45, 46, 47, 48 и 49; и
(f) CDR3 вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:50, 51, 52, 53, 54, 55 и 56;
где данное антитело специфически связывает PD-1.
Предпочтительная комбинация содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:15;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:22;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:29;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:36;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:43, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:50.
Другая предпочтительная комбинация содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:16;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:23;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:30;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:37;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:44, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:51.
Другая предпочтительная комбинация содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:17;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:24;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:31;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:38;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:45, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:52.
Другая предпочтительная комбинация содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:18;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:25;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:32;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:39;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:46, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:53.
Другая предпочтительная комбинация содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:19;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:26;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:33;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:40;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:47, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:54.
Другая предпочтительная комбинация содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:20;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:27;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:34;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:41;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:48, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:55.
Другая предпочтительная комбинация содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:21;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:28;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:35;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:42;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:49, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:56.
Другие предпочтительные антитела настоящего изобретения, или их антигенсвязывающие части, содержат:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 2, 3, 4, 5, 6 и 7, и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:8, 9, 10, 11, 12, 13 и 14;
где данное антитело специфически связывает PD-1.
Предпочтительная комбинация содержит:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:1, и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8.
Другая предпочтительная комбинация содержит:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:2, и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:9.
Другая предпочтительная комбинация содержит:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:3, и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:10.
Другая предпочтительная комбинация содержит:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:4, и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:11.
Другая предпочтительная комбинация содержит:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:5, и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:12.
Другая предпочтительная комбинация содержит:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:6, и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:13.
Другая предпочтительная комбинация содержит:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:7, и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:14.
Антитела настоящего изобретения могут быть, например, полноразмерными антителами, например, изотипа IgG1 или IgG4. Альтернативно, данные антитела могут быть фрагментами антител, такими как Fab или F(ab')2 фрагменты, или одноцепочечными антителами.
Настоящее изобретение обеспечивает также иммуноконъюгат, содержащий антитело настоящего изобретения или его антигенсвязывающую часть, связанные с терапевтическим агентом, таким как цитотоксин или радиоактивный изотоп. Настоящее изобретение обеспечивает также биспецифическую молекулу, содержащую антитело или его антигенсвязывающую часть настоящего изобретения, связанные со второй функциональной частью, имеющей другую специфичность связывания, чем указанное антитело или его антигенсвязывающая часть.
Обеспечены также композиции, содержащие антитело или его антигенсвязывающую часть, или иммуноконъюгат или биспецифическую молекулу настоящего изобретения и фармацевтически приемлемый носитель.
Молекулы нуклеиновых кислот, кодирующие данные антитела или их антигенсвязывающие части настоящего изобретения, также включены в настоящее изобретение, а также экспрессирующие векторы, содержащие такие нуклеиновые кислоты, и клетки-хозяева, содержащие такие экспрессирующие векторы. Кроме того, настоящее изобретение обеспечивает трансгенную мышь, содержащую трансгены тяжелой и легкой цепей иммуноглобулина человека, причем указанная мышь экспрессирует антитело настоящего изобретения, а также гибридомы, полученные из такой мыши, причем указанная гибридома продуцирует антитело настоящего изобретения.
Еще в одном аспекте настоящее изобретение обеспечивает способ модуляции иммунной реакции у субъекта, предусматривающий введение указанному субъекту антитела или его антигенсвязывающей части настоящего изобретения таким образом, что иммунная реакция у данного субъекта модулируется. Предпочтительно, антитело настоящего изобретения усиливает, стимулирует или увеличивает иммунную реакцию у данного субъекта.
В следующем аспекте настоящее изобретение обеспечивает способ ингибирования роста опухолевых клеток у субъекта, предусматривающий введение субъекту терапевтически эффективного количества анти-PD-1-антитела или его антигенсвязывающей части. Антитела настоящего изобретения предпочтительны для применения в данном способе, хотя вместо них могут быть использованы другие анти-PD-1-антитела (самостоятельно или в комбинации с анти-PD-1-антителом настоящего изобретения). Например, в способе ингибирования роста опухоли может быть использовано химерное, гуманизированное или полностью анти-PD-1-антитело человека.
В следующем аспекте настоящее изобретение обеспечивает способ лечения инфекционного заболевания у субъекта, предусматривающий введение субъекту терапевтически эффективного количества анти-PD-1-антитела или его антигенсвязывающей части. Антитела настоящего изобретения предпочтительны для применения в данном способе, хотя вместо них могут быть использованы другие анти-PD-1-антитела (самостоятельно или в комбинации с анти-PD-1-антителом настоящего изобретения). Например, в способе лечения инфекционного заболевания может быть использовано химерное, гуманизированное или полностью анти-PD-1-антитело человека.
Кроме того, настоящее изобретение обеспечивает способ усиления иммунной реакции на антиген у субъекта, предусматривающий введение данному субъекту: (i) антигена и (ii) анти-PD-1-антитела или его антигенсвязывающей части таким образом, что иммунная реакция на данный антиген у субъекта усиливается. Таким антигеном может быть, например, опухолевый антиген, вирусный антиген, бактериальный антиген или антиген из патогена. Антитела настоящего изобретения предпочтительны для применения в данном способе, хотя вместо них могут быть использованы другие анти-PD-1-антитела (самостоятельно или в комбинации с анти-PD-1-антителом настоящего изобретения). Например, в способе усиления иммунной реакции на антиген у субъекта может быть использовано химерное, гуманизированное или полностью анти-PD-1-антитело человека.
Настоящее изобретение обеспечивает также способы получения анти-PD-1-антител «второй генерации» на основе последовательностей анти-PD-1-антител настоящего изобретения. Например, настоящее изобретение обеспечивает способ получения анти-PD-1-антитела, предусматривающий:
(а) обеспечение: (i) последовательности вариабельной области тяжелой цепи антитела, содержащей последовательность CDR1, которая выбрана из группы, состоящей из SEQ ID NO:15, 16, 17, 18, 19, 20 и 21, и/или последовательность CDR2, которая выбрана из группы, состоящей из SEQ ID NO:22, 23, 24, 25, 26, 27 и 28; и/или последовательность CDR3, которая выбрана из группы, состоящей из SEQ ID NO:29, 30, 31, 32, 33, 34 и 35; или (ii) последовательности вариабельной области легкой цепи антитела, содержащей последовательность CDR1, которая выбрана из группы, состоящей из SEQ ID NO:36, 37, 38, 39, 40, 41 и 42, и/или последовательность CDR2, которая выбрана из группы, состоящей из SEQ ID NO:43, 44, 45, 46, 47, 48 и 49, и/или последовательность CDR3, которая выбрана из группы, состоящей из SEQ ID NO:50, 51, 52, 53, 54, 55 и 56;
(b) изменение по меньшей мере одного аминокислотного остатка по меньшей мере в одной последовательности вариабельной области антитела, причем указанная последовательность выбрана из последовательности вариабельной области тяжелой цепи антитела и последовательности вариабельной области легкой цепи антитела, для создания по меньшей мере одной измененной последовательности антитела; и
(с) экспрессию измененной последовательности антитела в виде белка.
Другие признаки и преимущества настоящего изобретения будут очевидны из следующего подробного описания и примеров, которые не должны рассматриваться как ограничивающие. Содержание всех цитируемых ссылок, вводов GenBank, патентов и опубликованных заявок на патент специально включено в данное описание посредством ссылки.
Краткое описание чертежей
На фиг.1А показана нуклеотидная последовательность (SEQ ID NO:57) и аминокислотная последовательность (SEQ ID NO:1) вариабельной области тяжелой цепи моноклонального антитела человека 17D8. Области CDR1 (SEQ ID NO:15), CDR2 (SEQ ID NO:22) и CDR3 (SEQ ID NO:29) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V, D и J.
На фиг.1В показана нуклеотидная последовательность (SEQ ID NO:64) и аминокислотная последовательность (SEQ ID NO:8) вариабельной области легкой цепи моноклонального антитела человека 17D8. Области CDR1 (SEQ ID NO:36), CDR2 (SEQ ID NO:43) и CDR3 (SEQ ID NO:50) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.2А показана нуклеотидная последовательность (SEQ ID NO:58) и аминокислотная последовательность (SEQ ID NO:2) вариабельной области тяжелой цепи моноклонального антитела человека 2D3. Области CDR1 (SEQ ID NO:16), CDR2 (SEQ ID NO:23) и CDR3 (SEQ ID NO:30) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.2В показана нуклеотидная последовательность (SEQ ID NO:65) и аминокислотная последовательность (SEQ ID NO:9) вариабельной области легкой цепи моноклонального антитела человека 2D3. Области CDR1 (SEQ ID NO:37), CDR2 (SEQ ID NO:44) и CDR3 (SEQ ID NO:51) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.3А показана нуклеотидная последовательность (SEQ ID NO:59) и аминокислотная последовательность (SEQ ID NO:3) вариабельной области тяжелой цепи моноклонального антитела человека 4Н1. Области CDR1 (SEQ ID NO:17), CDR2 (SEQ ID NO:24) и CDR3 (SEQ ID NO:31) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.3В показана нуклеотидная последовательность (SEQ ID NO:66) и аминокислотная последовательность (SEQ ID NO:10) вариабельной области легкой цепи моноклонального антитела человека 4Н1. Области CDR1 (SEQ ID NO:38), CDR2 (SEQ ID NO:45) и CDR3 (SEQ ID NO:52) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.4А показана нуклеотидная последовательность (SEQ ID NO:60) и аминокислотная последовательность (SEQ ID NO:4) вариабельной области тяжелой цепи моноклонального антитела человека 5С4. Области CDR1 (SEQ ID NO:18), CDR2 (SEQ ID NO:25) и CDR3 (SEQ ID NO:32) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.4В показана нуклеотидная последовательность (SEQ ID NO:67) и аминокислотная последовательность (SEQ ID NO:11) вариабельной области легкой цепи моноклонального антитела человека 5С4. Области CDR1 (SEQ ID NO:39), CDR2 (SEQ ID NO:46) и CDR3 (SEQ ID NO:53) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.5А показана нуклеотидная последовательность (SEQ ID NO:61) и аминокислотная последовательность (SEQ ID NO:5) вариабельной области тяжелой цепи моноклонального антитела человека 4А11. Области CDR1 (SEQ ID NO:19), CDR2 (SEQ ID NO:26) и CDR3 (SEQ ID NO:33) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.5В показана нуклеотидная последовательность (SEQ ID NO:68) и аминокислотная последовательность (SEQ ID NO:12) вариабельной области легкой цепи моноклонального антитела человека 4А11. Области CDR1 (SEQ ID NO:40), CDR2 (SEQ ID NO:47) и CDR3 (SEQ ID NO:54) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.6А показана нуклеотидная последовательность (SEQ ID NO:62) и аминокислотная последовательность (SEQ ID NO:6) вариабельной области тяжелой цепи моноклонального антитела человека 7D3. Области CDR1 (SEQ ID NO:20), CDR2 (SEQ ID NO:27) и CDR3 (SEQ ID NO:34) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.6В показана нуклеотидная последовательность (SEQ ID NO:69) и аминокислотная последовательность (SEQ ID NO:13) вариабельной области легкой цепи моноклонального антитела человека 7D3. Области CDR1 (SEQ ID NO:41), CDR2 (SEQ ID NO:48) и CDR3 (SEQ ID NO:55) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.7А показана нуклеотидная последовательность (SEQ ID NO:63) и аминокислотная последовательность (SEQ ID NO:7) вариабельной области тяжелой цепи моноклонального антитела человека 5F4. Области CDR1 (SEQ ID NO:21), CDR2 (SEQ ID NO:28) и CDR3 (SEQ ID NO:35) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.7В показана нуклеотидная последовательность (SEQ ID NO:70) и аминокислотная последовательность (SEQ ID NO:14) вариабельной области легкой цепи моноклонального антитела человека 5F4. Области CDR1 (SEQ ID NO:42), CDR2 (SEQ ID NO:49) и CDR3 (SEQ ID NO:56) показаны прерывистой линией и указаны деривации (источники) зародышевой линии V и J.
На фиг.8 показано сопоставление аминокислотной последовательности вариабельной области тяжелой цепи 17D8, 2D3, 4H1, 5C4 и 7D3 с аминокислотной последовательностью VH 3-33 зародышевой линии человека (SEQ ID NO:71).
На фиг.9 показано сопоставление аминокислотной последовательности вариабельной области легкой цепи 17D8, 2D3 и 7D3 с аминокислотной последовательностью Vk L6 зародышевой линии человека (SEQ ID NO:73).
На фиг.10 показано сопоставление аминокислотной последовательности вариабельной области легкой цепи 4Н1 и 5С4 с аминокислотной последовательностью Vk L6 зародышевой линии человека (SEQ ID NO:73).
На фиг.11 показано сопоставление аминокислотной последовательности вариабельной области тяжелой цепи 4А11 и 5F4 с аминокислотной последовательностью VH 4-39 зародышевой линии человека (SEQ ID NO:72).
На фиг.12 показано сопоставление аминокислотной последовательности вариабельной области легкой цепи 4А11 и 5F4 с аминокислотной последовательностью Vk L15 зародышевой линии человека (SEQ ID NO:74).
На фиг.13А-13В показаны результаты экспериментов с проточной цитометрией, демонстрирующие, что моноклональные антитела человека 5С4 и 4Н1, направленные против PD-1 человека, связываются с клеточной поверхностью клеток СНО, трансфицированных полноразмерным PD-1 человека. На фиг.13А показан график проточной цитометрии для 5С4. На фиг.13В показан график проточной цитометрии для 4Н1. Тонкая линия представляет связывание с клетками СНО, и жирная линия представляет связывание с СНО-hPD-1-клетками.
На фиг.14 показывает график, демонстрирующий, что моноклональные антитела 17D8, 2D3, 4H1, 5C34 и 4А11, направленные против PD-1 человека, специфически связываются с PD-1, но не с другими членами семейства CD28.
На фиг.15А-15С показаны результаты экспериментов с проточной цитометрией, демонстрирующие, что моноклональные антитела человека 5С4 и 4Н1, направленные против PD-1 человека, связываются с PD-1 на клеточной поверхности. На фиг.15А показано связывание с активированными Т-клетками человека. На фиг.15В показано связывание с Т-клетками собакоподобной обезьяны. На фиг.15С показано связывание с трансфицированными клетками СНО, экспрессирующими PD-1.
На фиг.16А-16С показаны результаты экспериментов, демонстрирующие, что моноклональные антитела человека против PD-1 человека стимулируют пролиферацию Т-клеток, секрецию IFN-гамма и секрецию IL-2 в анализе реакции лимфоцитов в смешенной культуре. Фиг.16А является диаграммой в виде столбцов, показывающей зависимую от концентрации пролиферацию Т-клеток; фиг.16В является диаграммой в виде столбцов, показывающей зависимую от концентрации секрецию IFN-гамма; фиг.16С является диаграммой в виде столбцов, показывающей зависимую от концентрации секрецию IL-2.
На фиг.17А-17В показаны результаты экспериментов с проточной цитометрией, демонстрирующие, что моноклональные антитела человека против PD-1 человека блокируют связывание PD-L1 и PD-L2 с трансфицированными клетками СНО, экспрессирующими PD-1. Фиг.17А является графиком, показывающим ингибирование связывания PD-L1; фиг.17В является графиком, показывающим ингибирование связывания PD-L2.
На фиг.18 показывает результаты экспериментов с проточной цитометрией, демонстрирующие, что моноклональные антитела человека против PD-1 человека не стимулируют апоптоз Т-клеток.
На фиг.19 показывает результаты экспериментов, демонстрирующие, что анти-PD-1-HuMab имеют зависимое от концентрации действие на секрецию IFN-гамма PBMC из CMV-положительных доноров, когда PBMC стимулировали лизатом CMV и анти-PD-1-антителом.
На фиг.20 показывает результаты экспериментов по росту опухоли на системе мышиной модели, демонстрирующие, что лечение опухоли мыши in vivo анти-PD-1-антителами ингибирует рост опухоли.
На фиг.21А-21D показан объем опухоли во времени у отдельных мышей, которым имплантировали опухолевые клетки ободочной кишки МС38 (PD-1-) и в тот же день подвергали одной из следующих терапий: (A) IgG мыши (контроль), (B) анти-CTLA-4-антитело, (C) анти-PD-1-антитело и (D) анти-CTLA-4-антитело и анти-PD-1-антитело. Мышей подвергали последующей обработке антителами в дни 3, 6 и 10, как описано в примере 13, и объем опухоли подвергали мониторингу на протяжении 60 дней.
На фиг.22 показан средний объем опухоли мышей, показанный на фиг.21
На фиг.23 показана медиана объема опухоли мышей, показанного на фиг.21.
На фиг.24A-24D показан объем опухоли во времени у отдельных мышей, которым были имплантированы опухолевые клетки MC38 ободочной кишки (PD-L1-) и спустя одну неделю были подвергнуты одной из следующих терапий: (A) IgG мыши (контроль), (B) анти-CTLA-4-антитело, (C) анти-PD-1-антитело и (D) анти-CTLA-4-антитело и анти-PD-1-антитело. Объем опухоли в первый день обработки составлял приблизительно 315 мм3. Мышей подвергали последующей обработке антителами в дни 3, 6 и 10, как описано в примере 14.
На фиг.25 показан средний объем опухоли мышей, показанный на фиг.24.
На фиг.26 показана медиана объема опухоли мышей, показанного на фиг.24.
На фиг.27 показан средний объем опухоли во времени у отдельных мышей, которым были имплантированы опухолевые клетки MC38 ободочной кишки (PD-L1-) (день -7) и затем были подвергнуты в дни 0, 3, 6 и 10 после имплантации (как описано в примере 15) одной из следующих терапий: (A) IgG мыши в качестве контроля (20 мг/кг, X20) (B) анти-PD-1-антитело (10 мг/кг) и IgG мыши (10 мг/кг) (P10X10), (C) анти-CTLA-4-антитело (10 мг/кг) и IgG мыши (10 мг/кг) (C10X10), (D) анти-CTLA-4-антитело и анти-PD-1-антитело (10 мг/кг каждого) (C10P10), (E) анти-CTLA-4-антитело и анти-PD-1-антитело (3 мг/кг каждого) (C3P3), и (F) анти-CTLA-4-антитело и анти-PD-1-антитело (1 мг/кг каждого) (C1P1). Две группы мышей были подвергнуты обработке каждым антителом последовательно следующим образом: (G) анти-CTLA-4-антитело (10 мг/кг, день 0), анти-CTLA-4-антитело (10 мг/кг, день 3), анти-PD-1-антитело (10 мг/кг, день 6) и анти-PD-1-антитело (10 мг/кг, день 10) (C10C10P10P10); и (H) анти-PD-1-антитело (10 мг/кг, день 0), анти-PD-1-антитело (10 мг/кг, день 3), анти-CTLA-4-антитело (10 мг/кг, день 6) и анти-CTLA-4-антитело (10 мг/кг, день 10) (10 мг/кг, день 10) (P10P10C10C10).
На фиг.28 показан средний объем опухоли мышей, показанный на фиг.27.
На фиг.29 показана медиана объема опухоли мышей, показанного на фиг.27.
На фиг.30A-30F показан объем опухоли во времени у отдельных мышей, которым были имплантированы клетки фибросаркомы SA1/N (PD-L1-) и спустя один день были подвергнуты одной из следующих терапий: (A) PBS (забуференный фосфатом физиологический раствор) (контроль-носитель), (B) IgG мыши (контроль-антитело, 10 мг/кг), (C) анти-PD-1-антитело (10 мг/кг), (D) анти-CTLA-4-антитело (10 мг/кг), (E) анти-CTLA-4-антитело (0,2 мг/кг) и (F) анти-PD-1-антитело (10 мг/кг) и анти-CTLA-4-антитело (0,2 мг/кг). Мышей подвергали последующей обработке антителами в дни 4, 7 и 11, как описано в примере 16, и объем опухоли подвергали мониторингу на протяжении 41 дня.
На фиг.31 показан средний объем опухоли мышей, показанный на фиг.29.
На фиг.32 показана медиана объема опухоли мышей, показанного на фиг.29.
На фиг.33А-33J показан объем опухоли во времени у отдельных мышей, которым были имплантированы клетки фибросаркомы SA1/N (PD-L1-) и затем были подвергнуты в дни 7, 10, 13 и 17 после имплантации (как описано на фиг.17) одной из следующих терапий: (A) PBS (контроль-носитель), (B) IgG мыши (контроль-антитело, 10 мг/кг), (C) анти-CTLA-4-антитело (0,25 мг/кг), (D) анти-CTLA-4-антитело (05 мг/кг), (E) анти-CTLA-4-антитело (5 мг/кг), (F) анти-PD-1-антитело (3 мг/кг), (G) анти-PD-1-антитело (10 мг/кг), (H) анти-PD-1-антитело (10 мг/кг) и анти-CTLA-4-антитело (0,25 мг/кг), (I) анти-PD-1-антитело (10 мг/кг) и анти-CTLA-4-антитело (0,5 мг/кг), и (F) анти-PD-1-антитело (3 мг/кг) и анти-CTLA-4-антитело (0,5 мг/кг). Объем опухоли в первый день обработки составлял приблизительно 110 мм3.
На фиг.34 показан средний объем опухоли в мышах, показанных на фиг.33.
На фиг.35 показана медиана объема опухоли мышей, показанного на фиг.33.
На фиг.36A и 36B показан объем опухоли во времени у отдельных мышей, которым были имплантированы клетки фибросаркомы SA1/N (PD-L1-) и затем были подвергнуты в дни 10, 13, 16 и 19 после имплантации (как описано в примере 17) одной из следующих терапий: (A) IgG мыши (контроль-антитело, 10 мг/кг) или (B) анти-PD-1-антитело (10 мг/кг) и анти-CTLA-4-антитело (1 мг/кг). Объем опухоли в первый день обработки составлял приблизительно 250 мм3.
На фиг.37 показан средний объем опухоли в мышах, показанных на фиг.36.
На фиг.38 показана медиана объема опухоли мышей, показанного на фиг.36.
На фиг.39 показан средний процент и медиана процента ингибирования опухоли, рассчитанные из объемов опухолей, показанных на фиг.33 и 36.
На фиг.40A-40D показан объем опухоли у мышей BALB/c, которым были имплантированы подкожно клетки почечной аденокарциномы RENCA (PD-L1+) (Murphy and Hrushesky (1973) J. Nat'l. Cancer Res. 50:1013-1025) (день -12) и затем были подвергнуты внутрибрюшинно в дни 0, 3, 6 и 9 после имплантации одной из следующих терапий: (A) IgG мыши (контроль-антитело, 20 мг/кг), (B) анти-PD-1-антитело (10 мг/кг), (C) анти-CTLA-4-антитело (10 мг/кг) и (D) анти-PD-1-антитело (10 мг/кг) в комбинации с анти-CTLA-4-антителом (10 мг/кг). Объем опухоли в первый день обработки составлял приблизительно 115 мм3.
На фиг.41 показано, что связывание мышиного слитого белка PD-L2-Fc с мышиным PD-1 (mPD-1) блокируется анти-mPD-1-антителом 4H2 зависимым от дозы образом. Связывание детектируют измерением флуоресценции FITC-меченым ослиным антителом против IgG крысы при помощи ELISA. Чем больше MFI (средняя интенсивность флуоресценции), тем больше связывание.
На фиг.42 показаны кривые связывания анти-mPD-1-антител с иммобилизованным слитым белком mPD-1-Fc, полученные при помощи ELISA.
На фиг.43 показана кривая связывания крысиного анти-mPD-1-антитела 4H2.B3 с mPD-1-экспрессирующими CHO-клетками. Связывание детектировали ослиными антителами против крысиного IgG, FITC-конъюгированными и измеренными при помощи FACS (MFI).
На фиг.44 показана кривая связывания слитого белка mPD-L1-hFc с mPD-1-экспрессирующими СНО-клетками в присутствии увеличивающихся концентраций анти-mPD-1-антитела 4H2.B3. Связывание детектировали козьими антителами против IgG человека IgG, FITC-конъюгированными и измеренными при помощи FACS (MFI).
На фиг.45 показаны кривые связывания крысиного анти-mPD-1-антитела 4H2.B3 с mPD-1-экспрессирующими СНО-клетками в сравнении с химерным крысиным:мышиным анти-mPD-1-антителом 4H2.
На фиг.46 показаны кривые связывания слитого белка mPD-L1-hFc с mPD-1-экспрессирующими СНО-клетками в присутствии увеличивающихся концентраций либо крысиного анти-mPD-1-антитела 4H2.B3, либо химерного крысиного:мышиного анти-mPD-1-антитела 4H2.
На фиг.47 показан средний объем опухоли не содержащих опухоли мышей, предварительно обработанных анти-PD-1-антителом и повторно имплантированных клетками фибросаркомы SA1/N (PD-L1-). Показан также средний объем опухоли необработанных мышей (контроль, предварительно неимплантированные или необработанные), имплантированных клетками фибросаркомы SA1/N.
На фиг.48 показывает объем опухоли во времени у отдельных мышей, которые выживали без опухоли после имплантации опухолевых клеток MC38 ободочной кишки (PD-L1-) и обработки анти-PD1-антителом или комбинацией анти-PD1-антитела с анти-CTLA-4-антителом), повторно имплантированных в 10 раз большим количеством опухолевых клеток MC38 ободочной кишки, чем при начальной обработке. Показан также средний объем опухоли необработанных мышей (контроль, предварительно неимплантированные или необработанные), имплантированных опухолевыми клетками МС38 ободочной кишки.
На фиг.49 показан средний объем опухоли мышей, показанный на фиг.48.
На фиг.50 показан средний объем опухоли во времени у отдельных мышей, которым были имплантированы опухолевые клетки CT26 ободочной кишки.
На фиг.51A-B показаны результаты экспериментов, демонстрирующих, что моноклональные антитела человека против PD-1 человека стимулируют пролиферацию Т-клеток и секрецию IFN-гамма в культурах, содержащих регуляторные Т-клетки. Фиг.51A является диаграммой в виде столбцов, показывающей зависимую от концентрации пролиферацию Т-клеток с использованием HuMAb 5C4; Фиг.51B является диаграммой в виде столбцов, показывающей зависимую от концентрации секрецию IFN-гамма с использованием HuMAb 5C4.
На фиг.52A-B показаны результаты экспериментов, демонстрирующие, что моноклональные антитела человека против PD-1 человека стимулируют пролиферацию Т-клеток и секрецию IFN-гамма в культурах, содержащих активированные Т-клетки. Фиг.52A является диаграммой в виде столбцов, показывающей зависимую от концентрации пролиферацию Т-клеток с использованием HuMAb 5C4; Фиг.52B является диаграммой в виде столбцов, показывающей зависимую от концентрации секрецию IFN-гамма с использованием HuMAb 5C4.
На фиг.53 показаны результаты анализа антителозависимой клеточноопосредованной цитотоксичности (ADCC), демонстрирующие, что моноклональные анти-PD-1-антитела человека убивают активированные Т-клетки человека в ADCC зависимым от концентрации образом относительно Fc-области анти-PD-1-антитела.
На фиг.54 показаны результаты анализа комплементзависимой цитотоксичности (CDC), демонстрирующие, что моноклональные анти-PD-1-антитела человека не убивают активированные Т-клетки человека в CDC зависимым от концентрации образом.
Наилучший способ осуществления изобретения
В одном аспекте настоящее изобретение относится к выделенным моноклональным антителам, в частности, моноклональным антителам человека, которые специфически связываются с PD-1. В некоторых вариантах осуществления антитела настоящего изобретения проявляют одно или несколько желаемых функциональных свойств, таких как высокая аффинность связывания с PD-1, отсутствие перекрестной реактивности c другими членами семейства CD28, способность стимулировать пролиферацию Т-клеток, секрецию IFN-гамма и/или IL-2 в реакциях смешанных лимфоцитов, способность ингибировать связывание одного или нескольких лигандов PD-1 (например, PD-L1 и/или PD-L2), способность перекрестно реагировать с PD-1 собакоподобной обезьяны, способность стимулировать антигенспецифические реакции памяти, способность стимулировать ответные реакции антител и/или способность ингибировать рост опухолевых клеток in vivo. Дополнительно или альтернативно, антитела настоящего изобретения получены из конкретных последовательностей зародышевой линии тяжелой и легкой цепи и/или содержат конкретные структурные признаки, такие как CDR-области, содержащие конкретные аминокислотные последовательности. В другом аспекте настоящее изобретение относится к комбинированному применению моноклональных антител, которые специфически связываются с PD-1, и моноклональных антител, которые специфически связываются с CTLA-4.
Настоящее изобретение обеспечивает, например, выделенные антитела, способы получения таких антител, иммуноконъюгатов и биспецифических молекул, содержащих такие антитела, и фармацевтические композиции, содержащие такие антитела, иммуноконъюгаты или биспецифические молекулы настоящего изобретения.
В другом аспекте настоящее изобретение относится к способам ингибирования роста опухолевых клеток у субъекта с использованием анти-PD-1-антител. Как показано в данном описании, анти-PD-1-антитела способны ингибировать рост опухолевых клеток in vivo. Настоящее изобретение относится также к способам применения данных антител для модификации иммунной реакции, а также для лечения заболеваний, таких как рак или инфекционное заболевание, или для стимуляции защитной аутоиммунной реакции, или для стимуляции антигенспецифических иммунных реакций (например, совместным введением анти-PD-1-антитела с представляющим интерес антигеном).
Для более легкого понимания настоящего изобретения сначала будут определены некоторые термины. Дополнительные определения объяснены в подробном описании.
Термины “программируемая гибель 1,” “программируемая гибель клеток 1,” “белок PD-1,” “PD-1,” PD1,” “PDCD1,” “hPD-1” и “hPD-I” используются взаимозаменяемо и включают варианты, изоформы, видовые гомологи PD-1 человека и аналоги, имеющие по меньшей мере один общий эпитоп с PD-1. Полная последовательность PD-1 может быть найдена под номером доступа GenBank Accession No. U64863.
Термины “цитотоксический T-лимфоцит-ассоциированный антиген-4,” “CTLA-4,” “CTLA4,” “антиген CTLA-4” и “CD152” (см., например, Murata, Am. J. Pathol. (1999) 155:453-460) используются взаимозаменяемо и включают варианты, изоформы, видовые гомологи CTLA-4 человека и аналоги, имеющие по меньшей мере один общий эпитоп с CTLA-4 (см., например, Balzano (1992) Int. J. Cancer Suppl. 7:28-32). Полная последовательность CTLA-4 может быть найдена под номером доступа GenBank Accession No. L15006.
Термин “иммунная реакция" относится к действию, например, лимфоцитов, антигенпрезентирующих клеток, фагоцитарных клеток, гранулоцитов и растворимых макромолекул, продуцируемых вышеуказанными клетками или печенью (включая антитела, цитокины и комплемент), которое приводит к селективному повреждению, деструкции или элиминации из организма человека внедряющихся патогенов, клеток или тканей, инфицированных патогенами, раковых клеток или, в случаях аутоиммунитета или патологического воспаления, здоровых клеток или тканей человека.
“Путь трансдукции сигнала” относится к биохимической связи между различными молекулами трансдукции сигналов, которые играют роль в передаче сигнала от одной части клетки к другой части клетки. Как использовано в данном описании, фраза “рецептор клеточной поверхности" включает, например, молекулы и комплексы молекул, способные получать сигнал и передавать такой сигнал через плазматическую мембрану клетки. Примером «рецептора клеточной поверхности» настоящего изобретения является рецептор PD-1.
Термин “антитело”, как использовано в данном описании, включает целые антитела и любой антигенсвязывающий фрагмент (т.е. “антигенсвязывающую часть”) или его отдельные цепи. Термин “антитело" относится к гликопротеину, содержащему по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, взаимосвязанные дисульфидными связями, или его антигенсвязывающей части. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно называемую в данном описании как VH) и константную область тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно называемую в данном описании как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые определяющими комплементарность областями (CDR), разбросанные между области, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат домен связывания, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями хозяина или факторами, включая различные клетки иммунной системы (например, эффекторными клетками), и первый компонент (Clq) классической системы комплемента.
Термин “антигенсвязывающая часть" антитела (или просто “часть антитела"), как использовано в данном описании, относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, PD-1). Было показано, что антигенсвязывающая функция антитела может выполняться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, включенных в термин “антигенсвязывающая часть" антитела включают (i) Fab-фрагмент, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2-фрагмент, двухвалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH в едином плече антитела, (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH; и (vi) выделенная определяющая комплементарность область (CDR). Кроме того, две области Fv-фрагмента, VL и VH, кодируются разными генами, они могут быть соединены при помощи рекомбинантных способов с использованием синтетического линкера, который дает возможность получать их в виде единой белковой цепи, в которой области VL и VH спариваются с образованием одновалентных молекул (известных как одноцепочечный Fv (scFv); см., например, Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Предполагается, что такие одноцепочечные молекулы также включены в термин “антигенсвязывающая часть" антитела. Такие фрагменты антител получают с использованием общепринятых способов, известных специалистам в данной области, и эти фрагменты подвергают скринингу таким же образом, как и интактные антитела.
“Выделенное антитело", как использовано в данном описании, означает антитело, которое по существу не содержит других антител, имеющих отличающиеся антигенные специфичности (например, выделенное антитело, которое специфически связывает PD-1, является по существу свободным от антител, которые специфически связывают антигены, отличные от PD-1). Однако выделенное антитело, которое связывает PD-1, может иметь перекрестную реактивность с другими антигенами, такими как молекулы PD-1 из других видов. Кроме того, выделенное антитело может быть по существу свободным от другого клеточного материала и/или химикалиев.
Термины «моноклональное антитело» или «композиция моноклонального антитела» относятся в данном описании к препарату молекул антител единого молекулярного состава. Композиция моноклонального антитела проявляет единственную специфичность связывания и аффинность в отношении конкретного эпитопа.
Термин «антитело человека» включает в данном описании антитела, имеющие вариабельные области, в которых как каркасные, так и CDR-области получены из последовательностей иммуноглобулина зародышевой линии человека. Кроме того, если данное антитело содержит константную область, то константная область также получена из последовательностей иммуноглобулина зародышевой линии человека. Антитела человека настоящего изобретения могут включать в себя аминокислотные остатки, не кодируемые последовательностями иммуноглобулина зародышевой линии человека (например, мутации, введенные случайным или сайт-специфическим мутагенезом in vitro, или соматическая мутация in vivo). Однако термин «антитело человека», как использовано в данном описании, не включает антитела, в которых CDR-последовательности, полученные из зародышевой линии другого вида млекопитающего, такого как мышь, были привиты на каркасные последовательности человека.
Термин «моноклональное антитело человека» означает антитела, проявляющие единственную специфичность связывания, которые имеют вариабельные области, в которых как каркасные, так и CDR-области получены из последовательностей иммуноглобулина зародышевой линии человека. В одном варианте осуществления моноклональные антитела человека получают с использованием гибридомы, которая включает в себя В-клетку, полученную из трансгенного животного (не человека), например, трансгенной мыши, имеющего геном, содержащий трансген тяжелой цепи и трансген легкой цепи человека, слитые с иммортализованной клеткой.
Термин «рекомбинантное антитело человека», как использовано в данном описании, включает все антитела человека, которые были получены, экспрессированы, созданы или выделены рекомбинантными способами, например, (а) антитела, выделенные из животного (например, мыши), которое является трансгенным или трансхромосомным в отношении генов иммуноглобулина человека, или полученной из них гибридомы (описанной дополнительно ниже), (b) антитела, выделенные из клетки-хозяина, трансформированной для экспрессии антитела человека, например, из трансфектомы, (с) антитела, выделенные из библиотеки рекомбинантных, комбинаторных антител человека, и (d) антитела, полученные, экспрессированные, созданные или выделенные любым другим способом, включающим сплайсинг последовательностей генов иммуноглобулинов человека в другие ДНК-последовательности. Такие рекомбинантные антитела человека имеют вариабельные области, в которых каркасные и CDR-области получены из последовательностей иммуноглобулина зародышевой линии человека. Однако в некоторых вариантах осуществления такие рекомбинантные антитела человека могут быть подвергнуты in vitro мутагенезу (или, когда используют животное, трансгенное в отношении последовательностей Ig, соматическому мутагенезу), и, следовательно, аминокислотные последовательности областей VH и VL рекомбинантных антител являются последовательностями, которые, хотя и получены из последовательностей VH и VL зародышевой линии человека или являются родственными указанным последовательностям, могут не существовать в природе в репертуаре зародышевой линии антител человека in vivo.
Как использовано в данном описании, «изотип» относится к классу антитела (например, IgM или IgG1), который кодируется генами константной области тяжелой цепи.
Фразы «антитело, распознающие антиген» и «антитело, специфическое в отношении антигена» являются в данном описании взаимозаменяемыми с термином «антитело, которое специфически связывается с антигеном».
Термин «производные антитела человека» относится к любой модифицированной форме антитела человека, например, конъюгату антитела и другого агента или антитела.
Термин «гуманизированное антитело» относится к антителам, в которых последовательности CDR, полученные из зародышевой линии другого вида млекопитающего, такого как мышь, были привиты на каркасные последовательности человека. В таких каркасных последовательностях человека могут быть произведены дополнительные модификации каркасной области.
Термин «химерное антитело» относится к антителам, в которых последовательности вариабельной области получены из одного вида, и последовательности константной области получены из другого вида, таким как антитело, в котором последовательности вариабельной области получены из мышиного антитела, и последовательности константной области получены из антитела человека.
Как использовано в данном описании, фраза антитело, которое «специфически связывается с PD-1 человека» относится к антителу, которое связывается с PD-1 человека с KD 1×10-7М или менее, более предпочтительно 5×10-8М или менее, более предпочтительно 1×10-8М или менее, более предпочтительно 5×10-9М или менее.
Термин “Kассоц” или “Ka”, как использовано в данном описании, относится к скорости ассоциации конкретного взаимодействия антитело-антиген, тогда как термин “Kдис” или “Kд” относится к скорости диссоциации конкретного взаимодействия антитело-антиген. Термин «KD» в данном описании относится к константе диссоциации, которую получают из отношения Kд к Ка (т.е. Кд/Ка), и ее выражают в виде молярной концентрации (М). Величины KD для антител могут быть определены с использованием способов, хорошо установленных в данной области. Предпочтительным способом определения KD антитела является способ с использованием резонанса поверхностных плазмонов, предпочтительно с использованием биосенсорной системы, такой как система Biacore®.
Как использовано в данном описании, термин «высокая аффинность» в отношении IgG-антитела относится к антителу, имеющему KD 10-8М, более предпочтительно 10-9 или менее и, даже более предпочтительно, 10-10М или менее в отношении антигена-мишени. Однако «высокая аффинность» связывания может варьироваться для других изотипов антигена. Например, «высокая аффинность» связывания для изотипа IgM относится к антителу, имеющему KD 10-7М или менее, более предпочтительно 10-8М или менее, даже более предпочтительно 10-9М или менее.
Термин «лечение» или «терапия» относится к введению активного агента с целью излечения, лечения, уменьшения симптомов, облегчения, изменения, вылечивания, купирования, улучшения состояния (например, заболевания), симптомов указанного состояния или воздействия на состояние (или заболевание), симптомы указанного состояния или для предупреждения или задержки проявления симптомов, осложнений, биохимических показателей заболевания или иным образом остановки или ингибирования дальнейшего развития указанного заболевания, симптомов или нарушения статистически значимым образом.
«Побочные эффекты» (АЕ) означает в данном описании любой неблагоприятный и обычно неожиданный, даже нежелательный, признак (включая аномальное лабораторное открытие), симптом или заболевание, ассоциированное с применением медицинского лечения. Например, неблагоприятное событие может быть ассоциировано с активацией иммунной системы или экспансии клеток иммунной системы (например, Т-клеток) в ответ на лечение. Медицинское лечение может иметь одно или несколько ассоциированных побочных эффектов (АЕ), и каждый АЕ может иметь один и тот же или различный уровень тяжести. Ссылка на способы, способные «изменять неблагоприятные события» означает схему лечения, которая уменьшает частоту и/или тяжесть одного или нескольких АЕ, ассоциированных с применением другой схемы лечении.
Как использовано в данном описании, «гиперпролиферативное заболевание» означает состояния, в которых рост клеток увеличивается выше нормальных уровней. Например, гиперпролиферативные заболевания или нарушения включают злокачественные заболевания (например, рак пищевода, рак ободочной кишки, билиарный рак) и незлокачественные заболевания (например, атеросклероз, доброкачественную гиперплазию, доброкачественную гиперплазию предстательной железы).
«Субтерапевтическая доза» означает дозу терапевтического соединения при введении отдельно для лечения гиперпролиферативного заболевания (например, рака). Например, субтерапевтической дозой CTLA-4-антитела является единственная доза данного антитела при менее чем приблизительно 3 мг/кг, т.е. известная доза анти-CTLA-4-антитела.
Применение альтернативы (например, «или»), как следует понимать, означает одну, обе или любую комбинацию указанных альтернатив. Неопределенные артикли «а» или “an” должны пониматься как «один или несколько» из любых указанных или перечисленных компонентов.
Как использовано в данном описании, «приблизительно» или «состоящий по существу из» означает «в пределах приемлемого диапазона ошибок» для конкретной величины, определенной специалистом в данной области, которая будет зависеть отчасти от того, как данная величина измерена или определена, т.е. от ограничений конкретной системы измерения. Например, «приблизительно» или «состоящий по существу из» может означать «в пределах 1 стандартного отклонения или более чем 1 стандартного отклонения», согласно практике в данной области. Альтернативно, «приблизительно» или «состоящий по существу из» может означать диапазон до 20%. Кроме того, в частности, в отношении биологических систем или процессов, эти термины могут означать «вплоть до значения или до 5-кратного значения величины». При использовании конкретных величин в данном описании и в формуле изобретения, если нет других указаний, значение «приблизительно» или «состоящий по существу из», как предполагается, должно быть в пределах приемлемого диапазона ошибок для этой конкретной величины.
Должно быть понятно, что в данном описании любой диапазон концентраций, диапазон процентов, диапазон отношений или диапазон целых чисел включает в себя величину любого целого числа в пределах указанного диапазона и, необязательно, его частей (например, одну десятую и одну сотую целого числа), если нет других указаний.
Как использовано в данном описании, термин «субъект» включает любого человека или любого животного, не являющегося человеком. Термин «животное-не человек» включает всех позвоночных, например, млекопитающих и не-млекопитающих, таких как приматы (не человек), овцы, собаки, лошади, коровы, куры, земноводные, рептилии и т.д. За исключением случаев, когда даются другие указания, термины «пациент» или «субъект» используются взаимозаменяемо.
Анти-PD-1-антитела
Антитела настоящего изобретения характеризуются конкретными функциональными признаками или свойствами указанных антител. Например, данные антитела связываются с PD-1 (например, связываются с PD-1 человека и могут перекрестно реагировать с PD-1 из другого вида, такого как собакоподобная обезьяна). Предпочтительно, антитело настоящего изобретения связывается с PD-1 с высокой аффинностью, например, с KD 1×10-7М или менее. Анти-PD-1-антитело настоящего изобретения предпочтительно проявляют одну или несколько из следующих характеристик:
(а) связывается с PD-1 человека с KD 1×10-7М или менее;
(b) по существу не связывается с CD28, CTLA-4 или ICOS человека;
(с) увеличивает пролиферацию Т-клеток в анализе реакции лимфоцитов в смешанной культуре (MLR);
(d) увеличивает продуцирование интерферона-гамма в анализе MLR;
(f) связывается с PD-1 человека и PD-1 собакоподобной обезьяны;
(g) ингибирует связывание PD-L1 и/или PD-L2 с PD-1;
(h) стимулирует антигенспецифические ответные реакции памяти;
(i) стимулирует ответные реакции антител;
(j) ингибирует рост опухолевых клеток in vivo.
Предпочтительно, данное антитело связывается с PD-1 человека с KD 5×10-8М или менее, связывается с PD-1 человека с KD 1×10-8М или менее, связывается с PD-1 человека с KD 5×10-9М или менее, или связывается с PD-1 человека с KD 1×10-8М-KD 1×10-10М.
Антитело настоящего изобретения может проявлять любую комбинацию вышеперечисленных признаков, например, двух, трех, четырех, пяти или более вышеперечисленных признаков.
Стандартные анализы для оценки связывающей способности антител против PD-1 известны в данной области, включая, например, ELISA, Вестерн-блоттинг и RIA. Кинетика связывания (например, аффинность связывания) антител может оцениваться также стандартными анализами, известными в данной области, такими как Biacore. Подходящие анализы для оценки любой из вышеописанных характеристик описаны подробно в примерах.
Моноклональные антитела 17D8, 2D3, 4H1, 5C4, 4А11, 7D3 и 5F4
Предпочтительные антитела настоящего изобретения являются моноклональными антителами человека 17D8, 2D3, 4H1, 5C4, 4А11, 7D3 и 5F4, выделенными и структурно охарактеризованными, как описано в примерах 1 и 2. Аминокислотные последовательности VH 17D8, 2D3, 4H1, 5C4, 4А11, 7D3 и 5F4 показаны в SEQ ID NO:1, 2, 3, 4, 5, 6 и 7, соответственно. Аминокислотные последовательности VL 17D8, 2D3, 4H1, 5C4, 4А11, 7D3 и 5F4 показаны в SEQ ID NO:8, 9, 10, 11, 12, 13 и 14, соответственно.
При условии, что каждое из указанных антител может связываться с PD-1, последовательности VH и VL могут быть «смешаны и спарены» для создания других анти-PD-1-связывающих молекул настоящего изобретения. PD-1-связывание таких «смешанных и спаренных» антител может быть испытано с использованием анализов связывания, описанных выше и в примерах (например, ELISA). Предпочтительно, при смешивании и спаривании цепей VH и VL последовательность VH из конкретного VH/VL-спаривания заменяется структурно сходной последовательностью VH. Подобным образом, последовательность VL из конкретного VH/VL-спаривания заменяется структурно сходной последовательностью VL.
Таким образом, в одном аспекте настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее:
(а) вариабельную область тяжелой цепи человека, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 2, 3, 4, 5, 6 и 7; и
(b) вариабельную область легкой цепи человека, содержащую аминокислотную последовательность, состоящую из SEQ ID NO:8, 9, 10, 11, 12, 13 и 14;
где данное антитело специфически связывает PD-1, предпочтительно PD-1 человека.
Предпочтительные комбинации тяжелых и легких цепей включают:
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:1; и (b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8; или
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:2; и (b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:9; или
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:3; и (b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:10; или
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:4; и (b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:11; или
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:5; и (b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:12; или
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:6; и (b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:13; или
(а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:7; и (b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:14.
В другом аспекте настоящее изобретение обеспечивает антитела, которые содержат CDR1, CDR2 и CDR3 тяжелой и легкой цепи 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4 или их комбинации. Аминокислотные последовательности CDR1 VH 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4 показаны в SEQ ID NO:15, 16, 17, 18, 19, 20 и 21, соответственно. Аминокислотные последовательности CDR2 VH 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4 показаны в SEQ ID NO:22, 23, 24, 25, 26, 27 и 28, соответственно. Аминокислотные последовательности CDR3 VH 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4 показаны в SEQ ID NO:29, 30, 31, 32, 33, 34 и 35, соответственно. Аминокислотные последовательности CDR1 Vκ 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4 показаны в SEQ ID NO:36, 37, 38, 39, 40, 41 и 42, соответственно. Аминокислотные последовательности CDR2 Vκ 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4 показаны в SEQ ID NO:43, 44, 45, 46, 47, 48 и 49, соответственно. Аминокислотные последовательности CDR3 Vκ 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4 показаны SEQ ID NO:50, 51, 52, 53, 54, 55 и 56, соответственно. CDR-области показаны прерывистой линией с использованием системы Кабата (Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242).
При условии, что каждое из указанных антител может связываться с PD-1 и что специфичность антигенсвязывания обеспечивается первично областями CDR1, CDR2 и CDR3, последовательности CDR1, CDR2 и CDR3 VH и последовательности CDR1, CDR2 и CDR3 Vκ могут быть “смешаны и спарены” (т.е. CDR из разных антител могут быть смешаны и спарены, хотя каждое антитело должно содержать CDR1, CDR2 и CDR3 и VH CDR1, CDR2 и CDR3 Vκ) для создания других анти-PD-1-связывающих молекул настоящего изобретения. PD-1-связывание таких “смешанных и спаренных” антител может быть испытано с использованием анализов связывания, описанных выше и в примерах (например, анализ ELISA, Biacore). Предпочтительно, при смешивании и спаривании последовательностей CDR VH, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VH заменяют структурно сходной (сходными) последовательностью (последовательностями) CDR. Подобным образом, при смешивании и спаривании последовательностей CDR Vκ, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности Vκ заменяют структурно сходной (сходными) последовательностью (последовательностями) CDR. Специалисту в данной области будет очевидно, что новые последовательности VH и VL могут быть созданы заменой одной или нескольких последовательностей CDR-областей VH и/или VL структурно сходными последовательностями из последовательностей CDR, описанных в данном описании для моноклональных антител 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4.
Таким образом, в другом аспекте настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее:
(а) CDR1 вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:15, 16, 17, 18, 19, 20 и 21;
(b) CDR2 вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:22, 23, 24, 25, 26, 27 и 28;
(с) CDR3 вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:29, 30, 31, 32, 33, 34 и 35;
(d) CDR1 вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:36, 37, 38, 39, 40, 41 и 42;
(е) CDR2 вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:43, 44, 45, 46, 47, 48 и 49; и
(f) CDR3 вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:50, 51, 52, 53, 54, 55 и 56:
где данное антитело специфически связывает PD-1, предпочтительно PD-1 человека.
В предпочтительном варианте осуществления данное антитело содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:15;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:22;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:29;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:36;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:43; и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:50.
В другом предпочтительном варианте осуществления данное антитело содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:16;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:23;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:30;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:37;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:44, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:51.
В другом предпочтительном варианте осуществления данное антитело содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:17;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:24;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:31;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:38;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:45, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:52.
В другом предпочтительном варианте осуществления данное антитело содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:18;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:25;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:32;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:39;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:46, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:53.
В другом предпочтительном варианте осуществления данное антитело содержит:
(а) CDR1 вариабельной области тяжелой цепи, содержащей SEQ ID NO:19;
(b) CDR2 вариабельной области тяжелой цепи, содержащей SEQ ID NO:26;
(с) CDR3 вариабельной области тяжелой цепи, содержащей SEQ ID NO:33;
(d) CDR1 вариабельной области легкой цепи, содержащей SEQ ID NO:40;
(е) CDR2 вариабельной области легкой цепи, содержащей SEQ ID NO:47, и
(f) CDR3 вариабельной области легкой цепи, содержащей SEQ ID NO:54.
Антитела, имеющие конкретные последовательности зародышевой линии
В некоторых вариантах осуществления антитело настоящего изобретения содержит вариабельную область тяжелой цепи из гена тяжелой цепи иммуноглобулина конкретной зародышевой линии и/или вариабельную область легкой цепи из гена легкой цепи иммуноглобулина конкретной зародышевой линии.
Например, в предпочтительном варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область тяжелой цепи, которая является продуктом или получена из гена VH 3-33, где данное антитело специфически связывает PD-1, предпочтительно PD-1 человека. В другом предпочтительном варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область тяжелой цепи, которая является продуктом или получена из гена VH 4-39, где данное антитело специфически связывает PD-1, предпочтительно PD-1 человека. Еще в одном предпочтительном варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область легкой цепи, которая является продуктом или получена из гена VK L6, где данное антитело специфически связывает PD-1, предпочтительно PD-1 человека. Еще в одном предпочтительном варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область легкой цепи, которая является продуктом или получена из гена VK L15, где данное антитело специфически связывает PD-1, предпочтительно PD-1 человека. Еще в одном предпочтительном варианте осуществления настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, где данное антитело:
(а) содержит вариабельную область тяжелой цепи, которая является продуктом гена VH 3-33 или гена 4-39 человека (причем указанный кодирует аминокислотную последовательность, представленную в SEQ ID NO:71 или 73, соответственно);
(b) содержит вариабельную область легкой цепи гена VK L6 или гена L15 человека (причем указанный кодирует аминокислотную последовательность, представленную в SEQ ID NO:72 или 74, соответственно), и
(с) специфически связывается с PD-1.
Примерами антител, имеющих VH и Vκ VH 3-33 и Vκ L6, соответственно, являются 17D8, 2D3, 4H1, 5C4 и 7D3. Примерами антител, имеющих VH и Vκ VH 4-39 и Vκ L15, соответственно, являются 4А11 и 5F4.
Как использовано в данном описании, антитело человека содержит вариабельные области тяжелой и легкой цепей, которые являются «продуктом» или получены из» последовательности конкретной зародышевой линии, если вариабельные области антитела получены из системы, которая использует гены иммуноглобулина зародышевой линии человека. Такие системы включают в себя иммунизацию трансгенной мыши, несущей гены иммуноглобулина человека, представляющим интерес антигеном или скрининг библиотеки генов иммуноглобулина человека, с фаговым дисплеем, с представляющим интерес антигеном. Антитело человека, которое является «продуктом или получено из» последовательности иммуноглобулина зародышевой линии человека, может быть идентифицировано, как таковое, сравнением аминокислотной последовательности антитела человека с аминокислотными последовательностями иммуноглобулинов зародышевой линии человека и отбором последовательности иммуноглобулина зародышевой линии человека, которая является наиболее близкой по последовательности (т.е. с большим % идентичности) с последовательностью антитела человека. Антитело человека, которое является «продуктом или получено из» последовательности иммуноглобулина зародышевой линии человека, могут содержать аминокислотные различия в сравнении с последовательностью зародышевой линии, вследствие, например, встречающихся в природе соматических мутаций или преднамеренного введения сайтнаправленной мутации. Однако выбранное антитело человека обычно является по меньшей мере на 90% идентичным по аминокислотной последовательности с аминокислотной последовательностью, кодируемой геном иммуноглобулина зародышевой линии человека, и содержит аминокислотные остатки, которые идентифицируют антитело человека, как являющееся антителом человека, при сравнении с аминокислотными последовательностями иммуноглобулина зародышевой линии другого вида (например, последовательностями зародышевой линии мыши). В некоторых случаях, антитело человека может быть по меньшей мере на 95% или даже по меньшей мере на 96%, 97%, 98% или 99% идентичным по аминокислотной последовательности с аминокислотной последовательностью, кодируемой геном иммуноглобулина данной зародышевой линии. Конкретно, антитело человека, полученное из последовательности конкретной зародышевой линии человека, будет обнаруживать не более чем 10 аминокислотных отличий от аминокислотной последовательности, кодируемой геном зародышевой линии иммуноглобулина. В этом случае антитело человека может обнаруживать не более чем 5, или даже не более чем 4, 3, 2 или 1 аминокислотное отличие от аминокислотной последовательности, кодируемой геном иммуноглобулина этой зародышевой линии.
Гомологичные антитела
Еще в одном варианте осуществления антитело настоящего изобретения содержит вариабельные области тяжелой и легкой цепей, содержащие аминокислотные последовательности, которые гомологичны аминокислотным последовательностям предпочтительных антител, описанных в данном описании, и где данные антитела сохраняют желаемые функциональные свойства анти-PD-1-антител настоящего изобретения.
Например, настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(а) вариабельная область тяжелой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:1, 2, 3, 4, 5, 6 и 7;
(b) вариабельная область легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:8, 9, 10, 11, 12, 13 и 14, и
данное антитело проявляет одно или несколько из следующих свойств:
(с) данное антитело связывается с PD-1 человека с KD 1×10-7М или менее; и
(d) данное антитело по существу не связывается с CD28, CTLA-4 или ICOS человека;
(е) данное антитело увеличивает пролиферацию Т-клеток в анализе MLR;
(f) данное антитело увеличивает продуцирование интерферона-гамма в анализе MLR;
(g) данное антитело увеличивает секрецию IL-2 в анализе MLR;
(h) данное антитело связывается с PD-1 человека и PD-1 собакоподобной обезьяны;
(i) данное антитело ингибирует связывание PD-L1 и/или PD-L2 с PD-1;
(j) данное антитело стимулирует антигенспецифические ответные реакции памяти;
(k) данное антитело стимулирует ответные реакции антител;
(l) данное антитело ингибирует рост опухолевых клеток in vivo.
В других вариантах осуществления аминокислотные последовательности VH и/или VL могут быть на 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологичны последовательностям, представленным выше. Антитело, имеющее области VH и VL, обладающие высокой гомологией (т.е. 80% или более) в отношении областей VH и VL представленных выше последовательностей, могут быть получены мутагенезом (например, сайтнаправленным или ПЦР-опосредованным мутагенезом) молекул нуклеиновых кислот, кодирующих SEQ ID NO:57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69 и 70, с последующим испытанием кодируемого измененного антитела на сохраненную функцию (т.е. на функции, представленные в (c)-(l) выше) с использованием описанных в данном описании функциональных анализов.
Как использовано в данном описании, процентная гомология между двумя аминокислотными последовательностями эквивалентна проценту идентичности между двумя последовательностями. Процент идентичности между двумя последовательностями является функцией числа идентичных положений, являющихся общими для данных последовательностей (т.е. % гомология=число идентичных положений/общее число положений×100), с учетом числа гэпов и длины каждого гэпа, которые должны быть введены для оптимального сопоставления указанных двух последовательностей. Это сравнение последовательностей и определение процента идентичности между двумя последовательностями может быть выполнено с использованием математического алгоритма, как описано в неограничивающих примерах ниже.
Процент идентичности между двумя аминокислотными последовательностями может быть определен с использованием алгоритма E. Meyers and W. Miller (Comput. Appl. Biosci., 4:11-17 (1988)), который был включен в программу ALIGN (версия 2.0), использующую таблицу массы остатков PAM120, штраф за длину гэпа 12 и штраф за открывание гэпа 4. Кроме того, процент идентичности между двумя аминокислотными последовательностями может быть определен с использованием алгоритма Needleman and Wunsch (J. Mol. Biol. 48:444-453 (1970)), который был включен в программу GAP в пакете программ GCG (доступном в ), использующую либо матрицу Blossum 62, либо матрицу PAM250 и массу гэпа 16, 14, 12, 10, 8, 6 или 4 и массу длины 1, 2, 3, 4, 5 или 6.
Дополнительно или альтернативно, белковые последовательности настоящего изобретения могут быть дополнительно использованы в качестве “запрашиваемой последовательности" для проведения поиска в публичных базах данных, например, для идентификации родственных последовательностей. Такие поиски могут быть проведены с использованием программы XBLAST (версия 2.0) Altschul, et al. (1990) J. Mol. Biol. 215:403-10. Поиски белков BLAST могут быть проведены с программой XBLAST, оценка=50, длина слова=3, для получения аминокислотных последовательностей, гомологичных молекулам антител настоящего изобретения. Для получения имеющих гэпы сопоставлений для целей сравнения может быть использована программа GAPPED BLAST, описанная в Altschul et al., (1997) Nucleic Acids Res. 25(17):3389-3402. При использовании программ BLAST и Gapped BLAST, могут быть использованы параметры по умолчанию соответствующих программ (например, XBLAST и NBLAST). (См. www.ncbi.nlm.nih.gov).
Антитела с консервативными модификациями
В некоторых вариантах осуществления антитело настоящего изобретения содержит вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, где одна или несколько из последовательностей CDR содержат указанные аминокислотные последовательности, выбранные на основе описанных в данном описании предпочтительных антител (например, 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4) или их консервативные модификации, и где указанные антитела сохраняют желаемые функциональные свойства анти-PD-1-антител настоящего изобретения. Таким образом, настоящее изобретение обеспечивает выделенное моноклональное антитело или его антигенсвязывающую часть, содержащее вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, где:
(а) последовательность CDR3 вариабельной области тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:29, 30, 31, 32, 33, 34 и 35, и их консервативных модификаций;
(b) последовательность CDR3 вариабельной области легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:50, 51, 52, 53, 54, 55 и 56, и их консервативных модификаций; и
где данное антитело проявляет одно или несколько из следующих свойств:
(с) данное антитело связывается с PD-1 человека с KD 1×10-7М или менее; и
(d) данное антитело по существу не связывается с CD28, CTLA-4 или ICOS человека;
(е) данное антитело увеличивает пролиферацию Т-клеток в анализе MLR;
(f) данное антитело увеличивает продуцирование интерферона-гамма в анализе MLR;
(g) данное антитело увеличивает секрецию IL-2 в анализе MLR;
(h) данное антитело связывается с PD-1 человека и PD-1 собакоподобной обезьяны;
(i) данное антитело ингибирует связывание PD-L1 и/или PD-L2 с PD-1;
(j) данное антитело стимулирует антигенспецифические ответные реакции памяти;
(k) данное антитело стимулирует ответные реакции антител;
(l) данное антитело ингибирует рост опухолевых клеток in vivo.
В предпочтительном варианте осуществления последовательность CDR2 вариабельной области тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:22, 23, 24, 25, 26, 27 и 28, и их консервативных модификаций; и последовательность CDR2 вариабельной области легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:43, 44, 45, 46, 47, 48 и 49, и их консервативных модификаций. В другом предпочтительном варианте осуществления последовательность CDR1 вариабельной области тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:15, 16, 17, 18, 19, 20 и 21, и их консервативных модификаций; и последовательность CDR1 вариабельной области легкой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:36, 37, 38, 39, 40, 41 и 42, и их консервативных модификаций.
Как использовано в данном описании, термин «консервативные модификации последовательностей» означает аминокислотные модификации, которые не влияют значимо на характеристики связывания или не изменяют характеристики связывания антитела, содержащего указанную аминокислотную последовательность. Такие консервативные модификации включают в себя аминокислотные замены, добавления и делеции. Модификации могут быть введены в антитело настоящего изобретения стандартными способами, известными в данной области, такими как сайтнаправленный мутагенез и ПЦР-опосредованный мутагенез. Консервативные аминокислотные замены являются заменами, в которых аминокислотный остаток заменен аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, были определены в данной области. Такие семейства включают в себя аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или несколько аминокислотных остатков в областях CDR антитела настоящего изобретения могут быть заменены другими аминокислотными остатками из того же самого семейства боковых цепей, и такое измененное антитело может быть тестировано на сохраненную функцию (т.е. функции, представленные в (с)-(l) выше), с использованием описанных в данном описании функциональных анализов.
Антитела, которые связываются с таким же эпитопом, что и анти-PD-1-антитела настоящего изобретения
В другом варианте осуществления настоящее изобретение обеспечивает антитела, которые связываются с таким же эпитопом на PD-1 человека, что и любое из моноклональных антител PD-1 настоящего изобретения (т.е. антитела, которые способны перекрестно конкурировать за связывание с PD-1 с любым из моноклональных антител настоящего изобретения). В предпочтительных вариантах осуществления ссылочным антителом для исследований перекрестной конкуренции может быть моноклональное антитело 17D8 (имеющее последовательности VH и VL, показанные в SEQ ID NO:1 и 8, соответственно) или моноклональное антитело 2D3 (имеющее последовательности VH и VL, показанные в SEQ ID NO:2 и 9, соответственно), или моноклональное антитело 4H1 (имеющее последовательности VH и VL, показанные в SEQ ID NO:3 и 10, соответственно), или моноклональное антитело 5C4 (имеющее последовательности VH и VL, показанные в SEQ ID NO:4 и 11, соответственно), или моноклональное антитело 4A11 (имеющее последовательности VH и VL, показанные в SEQ ID NO:5 и 12, или моноклональное антитело 7D3 (имеющее последовательности VH и VL, показанные в SEQ ID NO:6 и 13, или моноклональное антитело 5F4 (имеющее последовательности VH и VL, показанные в SEQ ID NO:7 и 14, соответственно). Такие перекрестно конкурирующие антитела могут быть идентифицированы на основе их способности перекрестно конкурировать с 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4 в стандартных анализах связывания PD-1. Например, анализ BIAcore, анализы ELISA или проточная цитометрия могут быть использованы для демонстрации перекрестной конкуренции с антителами настоящего изобретения. Способность тестируемого антитела ингибировать связывание, например, 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4, с PD-1 человека демонстрирует, что указанное тестируемое антитело может конкурировать с 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4 за связывание с PD-1 человека и, следовательно, связывается с таким же эпитопом на PD-1 человека, что и 17D8, 2D3, 4H1, 5С4 или 4А11. В предпочтительном варианте осуществления антителом, которое связывается с таким же эпитопом на PD-1 человека, что и 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4, является моноклональное антитело человека. Такие моноклональные антитела человека могут быть получены и выделены, как описано в примерах.
Сконструированные и модифицированные антитела
Кроме того, антитело настоящего изобретения может быть получено с использованием антитела, имеющего одну или несколько последовательностей VH и/или VL, описанных в данном описании, в качестве исходного материала для конструирования модифицированного антитела, которое может иметь измененные свойства в сравнении с исходным антителом. Антитело может быть сконструировано модификацией одного или нескольких остатков в одной или обеих вариабельных областях (т.е. VH и/или VL), например, в одной или нескольких областях CDR и/или в одной или нескольких каркасных областях. Дополнительно или альтернативно, антитело может быть сконструировано модификацией остатков в константной области (константных областях), например, для изменения эффекторной функции (эффекторных функций) данного антитела.
Одним типом конструирования вариабельной области, который может выполняться, является прививка CDR. Антитела взаимодействуют с антигенами-мишенями преимущественно через аминокислотные остатки, которые расположены в шести определяющих комплементарность областях (CDR) тяжелой и легкой цепи. По этой причине аминокислотные последовательности в CDR являются более разнообразными между отдельными индивидуумами, чем последовательности вне CDR. Поскольку последовательности CDR ответствены за большинство взаимодействий антитело-антиген, можно экспрессировать рекомбинантные антитела, которые имитируют свойства специфических встречающихся в природе антител, конструированием экспрессирующих векторов, которые включают в себя последовательности CDR из специфического встречающегося в природе антитела, привитые на каркасные последовательности из другого антитела с другими свойствами (см., например, Riechmann, L. et al. (1998) Nature 332:323-327; Jones, P. et al. (1986) Nature 321:522-525; Queen, C. et al. (1989) Proc. Natl. Acad. см. U.S.A. 86:10029-10033; патент США № 5225539, Winter, и патенты США №№ 5530101; 5585089; 5693762 и 6180370, Queen et al.).
Таким образом, другой вариант настоящего изобретения относится к выделенному моноклональному антителу или его антигенсвязывающей части, содержащему последовательности CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:15, 16, 17, 18, 19, 20 и 21, SEQ ID NO:22, 23, 24, 25, 26, 27 и 28 и SEQ ID NO:29, 30, 31, 32, 33, 34 и 35, соответственно, и последовательности CDR1, CDR2 и CDR3 вариабельной области легкой цепи, содержащей аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:36, 37, 38, 39, 40, 41 и 42, SEQ ID NO:43, 44, 45, 46, 47, 48 и 49, и SEQ ID NO:50, 51, 52, 53, 54, 55 и 56, соответственно. Таким образом, такие последовательности содержат последовательности CDR VH и VL моноклональных антител 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4, но могут содержать разные каркасные последовательности из указанных антител.
Такие каркасные последовательности могут быть получены из публичных баз данных или опубликованных ссылок, которые включают в себя последовательности генов антител зародышевой линии. Например, ДНК-последовательности зародышевой линии для генов вариабельной области тяжелой и легкой цепи человека могут быть найдены в базе данных последовательностей зародышевой линии человека "VBase" (доступной в Интернете в ), а также в Kabat, E A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242; Tomlinson, I.M., et al. (1992) "The Repertoire of Human Germline VH Sequences Reveals about Fifty Groups of VH Segments with Different Hypervariable Loops" J. Mol. Biol. 227:776-798; и Cox, J. P. L. et al. (1994) "A Directory of Human Germ-line VH Segments Reveals a Strong Bias in their Usage" Eur. J. Immunol. 24:827-836; содержание которых специально включено в данное описание посредством ссылки. В качестве другого примера, ДНК-последовательности зародышевой линии для генов вариабельной области тяжелой и легкой цепи могут быть найдены в базе данных GenBank. Например, следующие последовательности зародышевой линии тяжелой цепи, обнаруженные у мыши HCo7 HuMAb, доступны в сопутствующих номерах доступа GenBank: 1-69 (NG_0010109, NT_024637 и BC070333), 3-33 (NG_0010109 и NT_024637) и 3-7 (NG_0010109 и NT_024637). В качестве другого примера, следующие последовательности зародышевой линии тяжелой цепи, обнаруженные у мыши HCo12 HuMAb, доступны в сопутствующих номерах доступа GenBank: 1-69 (NG_0010109, NT_024637 и BC070333), 5-51 (NG_0010109 и NT_024637), 4-34 (NG_0010109 и NT_024637), 3-30.3 (AJ556644) и 3-23 (AJ406678).
Предпочтительными каркасными последовательностями для применения в антителах настоящего изобретения, являются последовательности, которые структурно сходны с каркасными последовательностями, используемыми в выбранных антителах настоящего изобретения, например, сходные с каркасными последовательностями VH 3-33 (SEQ ID NO:71) и/или каркасными последовательностями VH 4-39 (SEQ ID NO:73), и/или каркасными последовательностями VK L6 (SEQ ID NO:72), и/или каркасными последовательностями VK L15 (SEQ ID NO:74), используемыми предпочтительными моноклональными антителами настоящего изобретения. Последовательности CDR1, CDR2 и CDR3 VH и последовательности CDR1, CDR2 и CDR3 VK, могут быть привиты на каркасные области, которые имеют последовательность, идентичную последовательности, обнаруженной в гене иммуноглобулина зародышевой линии, из которой получена указанная последовательность, или такие последовательности CDR могут быть привиты на каркасные области, которые содержат одну или несколько мутаций в сравнении с последовательностями зародышевой линии. Например, было обнаружено, что в некоторых случаях предпочтительно мутировать остатки в каркасных областях для сохранения или усиления антигенсвязывающей способности данного антитела (см., например, патенты США №№ 5530101; 5585089; 5693762 и 6180370, Queen et al).
Другим типом модификации вариабельной области является мутирование аминокислотных остатков в областях CDR1, CDR2 и/или CDR3 VH и/или Vκ, посредством чего улучшая одно или несколько свойств связывания (например, аффинности) представляющего интерес антитела. Сайтнаправленный мутагенез или ПЦР-опосредованный мутагенез может быть выполнен для введения мутации (мутаций), и действие на связывание антител или другое представляющее интерес функциональное свойство может быть оценено в анализах in vitro или in vivo, описанных в данном описании и представленных в примерах. Предпочтительно, вводят консервативные модификации (обсуждаемые выше). Такими мутациями могут быть замены, добавления или делеции аминокислот, но предпочтительными являются замены. Кроме того, обычно изменяют не более чем один, два, три, четыре или пять остатков в области CDR.
Таким образом, в другом варианте осуществления настоящее изобретение обеспечивает выделенные моноклональные анти-PD-1-антитела, или их антигенсвязывающие части, содержащие вариабельную область тяжелой цепи, содержащую: (a) CDR1-область VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:15, 16, 17, 18, 19, 20 и 21, или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений, в сравнении с SEQ ID NO:15, 16, 17, 18, 19, 20 и 21; (b) CDR2-область VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:22, 23, 24, 25, 26, 27 и 28, или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений, в сравнении с SEQ ID NO:22, 23, 24, 25, 26, 27 и 28; (с) CDR3-область VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:29, 30, 31, 32, 33, 34 и 35, или аминокислотную последовательность, содержащую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений, в сравнении с SEQ ID NO:29, 30, 31, 32, 33, 34 и 35; (d) CDR1-область VK, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:36, 37, 38, 39, 40, 41 и 42, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений, в сравнении с SEQ ID NO:36, 37, 38, 39, 40, 41 и 42; (e) CDR2-область VK, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:43, 44, 45, 46, 47, 48 и 49, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений, в сравнении с SEQ ID NO:43, 44, 45, 46, 47, 48 и 49; и (f) CDR3-область VK, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:50, 51, 52, 53, 54, 55 и 56, или аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавлений, в сравнении с SEQ ID NO: 50, 51, 52, 53, 54, 55 и 56.
Сконструированные антитела настоящего изобретения включают в себя антитела, в которых модификации были произведены в каркасных остатках в VH и/или Vκ, например, для улучшения свойств данного антитела. Обычно такие каркасные модификации производят для уменьшения иммуногенности антитела. Например, одним подходом является «обратное мутирование» одного или нескольких каркасных остатков в соответствующую последовательность зародышевой линии. Более конкретно, антитело, которое было подвергнуто соматической мутации, может содержать каркасные остатки, которые отличаются от последовательности зародышевой линии, из которой получено данное антитело. Такие остатки могут быть идентифицированы сравнением каркасных последовательностей антитела с последовательностями зародышевой линии, из которой получено данное антитело.
Например, в таблице 1 ниже показано количество аминокислотных изменений в каркасных областях анти-PD-1-антител 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4, которые отличаются от последовательностей тяжелой цепи исходной зародышевой линии. Для возврата одного или нескольких аминокислотных остатков в указанных последовательностях каркасной области в их конфигурацию в зародышевой линии, соматические мутации могут быть «обратно-мутированы» в последовательность зародышевой линии, например, сайтнаправленным мутагенезом или ПЦР-опосредованным мутагенезом.
Аминокислотные изменения могут иметь место в каркасных областях анти-PD-1-антител, которые отличаются от исходной последовательности легкой цепи зародышевой линии. Например, для 17D8, аминокислотным остатком №47 (в FR2) VK является изолейцин, тогда как остатком в соответствующей последовательности VK L6 зародышевой линии является лейцин. Для возврата последовательностей каркасной области в их конфигурацию зародышевой линии, соматические мутации могут быть «обратно-мутированы» в последовательность зародышевой линии, например, сайтнаправленным мутагенезом или ПЦР-опосредованным мутагенезом (например, остаток №47 (остаток №13 FR2) VK 17D8 может быть «обратно-мутирован» из изолейцина в лейцин).
В качестве другого примера, для 4А11, аминокислотным остатком №20 (в FR1) VK является серин, тогда как остатком в соответствующей последовательности VK L15 зародышевой линии является треонин. Для возврата последовательностей каркасной области в их конфигурацию зародышевой линии, соматические мутации могут быть «обратно-мутированы» в последовательность зародышевой линии, например, остаток №20 VK 4А11 может быть «обратно-мутирован» из серина в треонин). Предполагается, что такие «обратно-мутированные» антитела также включены в настоящее изобретение.
Сопоставление VH-областей для 17D8, 2D3, 4H1, 5C4 и 7D3 с последовательностью VH 3-33 исходной зародышевой линии показано на фиг.8. Сопоставление VH-областей для 4А11 и 5F4 с последовательностью VH 4-39 исходной зародышевой линии показано на фиг.11.
Другой тип модификации каркаса включает мутацию одного или нескольких остатков в каркасной области или даже в одном или нескольких CDR-областей, для удаления эпитопов Т-клеток для уменьшения таким образом потенциальной иммуногенности антитела. Такой подход называют также «деиммунизацией», и он описаны более подробно в публикации патента США № 20030153043, Carr et al.
В качестве дополнения или альтернативы модификациям, произведенным в каркасных или CDR-областях, антитела настоящего изобретения могут быть сконструированы таким образом, что они включают в себя модификации в Fc-области, обычно для изменения одного или нескольких функциональных свойств антитела, таких как период полусуществования в сыворотке, фиксация комплемента, связывание рецептора Fc и/или антиген-независимая клеточноопосредованная цитотоксичность. Кроме того, антитело настоящего изобретения может быть также химически модифицировано (например, к антителу могут быть присоединены одна или несколько химических групп) или может быть модифицировано для изменения его гликозилирования, опять для изменения одного или нескольких функциональных свойств данного антитела. Каждый из указанных вариантов описан более подробно ниже. Нумерация остатков в Fc-области является нумерацией EU-индекса Кабата.
В одном варианте осуществления шарнирная область СН1 модифицирована таким образом, что количество остатков цистеина в шарнирной области изменено, например, увеличенным или уменьшенным. Такой подход описан дополнительно в патенте США № 5677425, Bodmer et al. Количество остатков цистеина в шарнирной области СН1 изменяют, например, для облегчения сборки легких и тяжелых цепей или для увеличения или уменьшения стабильности антитела.
В другом варианте осуществления шарнирную область Fc антитела мутируют для уменьшения биологического периода полусуществования антитела. Более конкретно, одну или несколько аминокислотных мутаций вводят в области контакта СН2-СН3-доменов Fc-шарнирного фрагмента таким образом, что данное антитело имеет нарушенное связывание стафилококкового белка А (SpA) в сравнении со связыванием Fc-шарнирного домена с SpA. Такой подход описан более подробно в патенте США № 6165745, Ward et al.
В другом варианте осуществления антитело модифицируют для увеличения биологического периода полусуществования антитела. Например, могут быть введены одна или несколько мутаций: T252L, T254S, T256F, как описано в патенте США № 6277375, Ward. Альтернативно, для увеличения биологического периода полусуществования, антитело может быть изменено в СН1 или СН2 области таким образом, что оно содержит эпитоп связывания рецептора реутилизации, образованного из двух петель СН2-домена Fc-области IgG, как описано в патентах США №№ 5869046 и 6121022, Presta et al.
Еще в одном варианте осуществления Fc-область изменяют заменой по меньшей мере одного аминокислотного остатка другим аминокислотным остатком для изменения эффекторной функции (эффекторных функций) данного антитела. Например, одна или несколько аминокислот, выбранных из аминокислотных остатков 234, 235, 236, 237, 297, 318, 320 и 322, могут быть заменены другим аминокислотным остатком таким образом, что антитело имеет измененную аффинность в отношении эффекторного лиганда, но сохраняет антигенсвязывающую способность исходного антитела. Эффекторным лигандом, в отношении которого аффинность изменяется, может быть, например, рецептор Fc или С1-компонент комплемента. Такой подход описан более подробно в патентах США №№ 5624821 и 5648260, Winter.
В другом примере, одна или несколько аминокислот, выбранных из аминокислотных остатков 329, 331 и 322, могут быть заменены другим аминокислотным остатком таким образом, что антитело имеет измененное связывание C1q и/или уменьшенную или устраненную комплементзависимую цитотоксичность (CDC). Такой подход описан более подробно в патенте США № 6194551, Idusogie et al.
В другом примере, один или несколько аминокислотных остатков в положениях аминокислот 231 и 239 изменяют, чтобы таким образом изменить способность антитела фиксировать комплемент. Такой подход описан более подробно в публикации РСТ WO 94/29351, Bodmer et al.
Еще в одном примере Fc-область модифицируют для увеличения способности антитела опосредовать антителозависимую клеточную цитотоксичность (ADCC) и/или для увеличения аффинности антитела в отношении рецептора Fcγ модификацией одной или нескольких аминокислот в следующих положениях:: 238, 239, 248, 249, 252, 254, 255, 256, 258, 265, 267, 268, 269, 270, 272, 276, 278, 280, 283, 285, 286, 289, 290, 292, 293, 294, 295, 296, 298, 301, 303, 305, 307, 309, 312, 315, 320, 322, 324, 326, 327, 329, 330, 331, 333, 334, 335, 337, 338, 340, 360, 373, 376, 378, 382, 388, 389, 398, 414, 416, 419, 430, 434, 435, 437, 438 или 439. Такой подход описан дополнительно в публикации PCT WO 00/42072, Presta. Кроме того, сайты связывания на IgG1 человека для FcγRI, FcγRII, FcγRIII и FcγRn были картированы и описаны варианты с улучшенным связыванием (см. Shields, R.L. et al. (2001) J. Biol. Chem. 276:6591-6604). Было показано, что конкретные мутации в положениях 256, 290, 298, 333, 334 и 339 улучшают связывание с FcγRIII. Дополнительно, было показано, что следующие мутанты улучшают связывание FcγRIII: T256A/S298A, S298A/E333A, S298A/K224A и S298A/E333A/K334A.
Еще в одном варианте осуществления модифицируют гликозилирование антитела. Например, может быть получено агликозилированное антитело (т.е. антитело лишено гликозилирования). Гликозилирование может быть изменено, например, для увеличения аффинности антитела в отношении антигена. Такие углеводные модификации могут быть выполнены, например, изменением одного или нескольких сайтов гликозилирования в последовательности антитела. Например, могут быть произведены одна или несколько аминокислотных замен, которые приводят к элиминации одного или нескольких сайтов гликозилирования каркаса вариабельной области, чтобы таким образом элиминировать гликозилирование в данном сайте. Такое агликозилироание может увеличивать аффинность антитела в отношении антигена. Такой подход описан более подробно в патентах США №№ 5714350 и 6350861, Co et al.
Дополнительно или альтернативно, может быть получено антитело, которое имеет измененный тип гликозилирования, например, гипофукозилированное антитело, имеющее уменьшенные количества остатков фукозила, или антитело, имеющее увеличенные разделяющие пополам структуры GlcNac. Было продемонстрировано, что такие измененные картины гликозилирования увеличивают ADCC-способность антител. Такие углеводные модификации могут быть выполнены, например, экспрессией антитела в клетке-хозяине с измененным аппаратом гликозилирования. Клетки с измененным аппаратом гликозилирования были описаны в данной области и могут быть использованы в качестве клеток-хозяев для экспрессии в них рекомбинантных антител настоящего изобретения, чтобы таким образом получить антитела с измененным гликозилированием. Например, линии клеток Ms704, Ms705 и Ms709 лишены гена фукозилтрансферазы, FUT8 (альфа(1,6)фукозилтрансферазы) таким образом, что антитела, экспрессируемые в клеточных линиях Ms704, Ms705 и Ms709, лишены фукозы на их углеводах. FUT8-/--клеточные линии Ms704, Ms705 и Ms709 были созданы нацеленным разрушением гена FUT8 в клетках CHO/DG44 с использованием двух вытесняющих векторов (см. публикацию патента США No. 20040110704, Yamane et al. и Yamane-Ohnuki et al. (2004) Biotechnol Bioeng 87:614-22). В качестве другого примера, в ЕР 1176195 Hanai et al. описывают клеточную линию с функционально разрушенным геном FUT8, который кодирует фукозилтрансферазу таким образом, что антитела, экспрессируемые в такой клеточной линии, проявляют гипофукозилирование или элиминацию альфа-1,6-связь-родственного фермента. Hanai et al. описывают также клеточные линии, которые имеют низкую ферментативную активность в отношении присоединения фукозы к N-ацетилглюкозамину, который связывается с Fc-областью антитела, или не имеет ферментативной активности, например, клеточную линию миеломы крысы YB2/0 (ATCC CRL 1662). В публикации PCT WO 03/035835, Presta описывает вариант клеточной линии СНО, клетки Lec13, с уменьшенной способностью присоединять фукозу к Asn(297)-связанным углеводам, что также приводит к гипофукозилированию антител, экспрессируемых в клетках-хозяевах (см. также Shields, R.L. et al. (2002) J. Biol. Chem. 277:26733-26740). В публикации PCT WO 99/54342, Umana et al. описывают клеточные линии, сконструированные для экспрессии гликопротеин-модифицирующих гликозилтрансфераз (например, бета(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII)) таким образом, что антитела, экспрессируемые в таких сконструированных клеточных линиях, проявляют увеличенное разделение на две половины структур GlcNac, что приводит к увеличенной активности ADCC в указанных антителах (см. также Umana et al. (1999) Nat. Biotech. 17:176-180). Альтернативно, остатки фукозы антитела могут быть отщеплены с использованием фермента фукозидазы. Например, фукозидаза альфа-L-фукозидаза удаляет фукозильные остатки из антител (Tarentino, A.L. et al. (1975) Biochem. 14:5516-23).
Другой модификацией антител, которая обсуждается данным изобретением, является пэгилирование. Антитело может быть пэгилировано, например, для увеличения биологического (например, в сыворотке) периода полусуществования антитела. Для пэгилирования антитела данное антитело или его фрагмент, обычно реагирует с полиэтиленгликолем (ПЭГ), таким как реакционноспособное сложноэфирное или альдегидное производное ПЭГ, в условиях, в которых одна или несколько групп ПЭГ становятся присоединенными к антителу или фрагменту антитела. Предпочтительно, пэгилирование проводят путем реакции ацилирования или реакции алкилирования с реакционноспособной молекулой ПЭГ (или аналогичным реакционноспособным водорастворимым полимером). Как использовано в данном описании, предполагается, что термин “полиэтиленгликоль” включает любую из форм ПЭГ, которые были использованы для дериватизации других белков, такую как моно(C1-C10)алкокси- или арилоксиполиэтиленгликоль или малеимид полиэтиленгликоля. В некоторых вариантах осуществления антителом, которое должно быть пэгилировано, является агликозилированное антитело. Способы пэгилирования белков известны в данной области и могут быть применены к антителам настоящего изобретения. См., например, EP 0154316, Nishimura et al. и EP 0401384, Ishikawa et al.
Способы конструирования антител
Как обсуждалось выше, анти-PD-1-антитела, имеющие описываемые последовательности VH и VK, могут быть использованы для создания новых анти-PD-1-антител модификацией последовательностей VH и/или VK или константной области (константных областей), присоединенных к ним. Таким образом, в другом аспекте настоящего изобретения структурные признаки анти-PD-1-антитела настоящего изобретения, например, 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4, используют для создания структурно родственных анти-PD-1-антител, которые сохраняют по меньшей мере одно функциональное свойство антител настоящего изобретения, например, связывание с PD-1 человека. Например, один или несколько CDR-областей 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4, или их мутации, могут быть комбинированы с известными каркасными областями и/или другими CDR для создания дополнительных, рекомбинантно сконструированных анти-PD-1-антител настоящего изобретения, как обсуждалось выше. Другие типы модификаций включают модификации, обсуждаемые в предыдущем разделе. Исходным материалом для способа конструирования является одна или несколько последовательностей VH и/или VK, обеспеченные настоящим изобретением, или один или несколько их CDR-областей. Для создания сконструированного антитела необязательно действительно получать (т.е. экспрессировать в виде белка) антитело, имеющее одну или несколько последовательностей VH и/или VK, обеспеченных настоящим изобретением, или один или несколько их CDR-областей. Предпочтительно, используют информацию, содержащуюся в последовательности (последовательностях), в качестве исходного материала для создания последовательности (последовательностей) «второй генерации», полученных из начальной последовательности (начальных последовательностей), и затем последовательность (последовательности) «второй генерации» получают и экспрессируют в виде белка.
Таким образом, в другом варианте осуществления настоящее изобретение обеспечивает способ получения анти-PD-1-антитела, предусматривающий:
(a) обеспечение: (i) последовательности вариабельной области тяжелой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO:15, 16, 17, 18, 19, 20 и 21, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NO:22, 23, 24, 25, 26, 27 и 28, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NO:29, 30, 31, 32, 33, 34 и 35; и/или (ii) последовательности вариабельной области легкой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO:36, 37, 38, 39, 40, 41 и 42, последовательность CDR2, выбранную из группы, состоящей из SEQ ID NO:43, 44, 45, 46, 47, 48 и 49, и/или CDR3, выбранную из группы, состоящей из SEQ ID NO:50, 51, 52, 53, 54, 55 и 56;
(b) изменение по меньшей мере одного аминокислотного остатка в последовательности вариабельной области тяжелой цепи антитела и/или последовательности вариабельной области легкой цепи антитела для создания по меньшей мере одной измененной последовательности антитела; и
(c) экспрессию измененной последовательности антитела в виде белка.
Для получения и экспрессии измененной последовательности антитела могут быть использованы стандартные способы молекулярной биологии.
Предпочтительно, антитело, кодируемое измененной последовательностью антитела (измененными последовательностями антитела) является антитело, которое сохраняет одно, несколько или все из функциональных свойств описанных в данном описании анти-PD-1-антител, причем указанные функциональные свойства включают, но не ограничиваются ими:
(а) данное антитело связывается с PD-1 человека с KD 1×10-7М или менее; и
(b) данное антитело по существу не связывается с CD28, CTLA-4 или ICOS человека;
(c) данное антитело увеличивает пролиферацию Т-клеток в анализе MLR;
(d) данное антитело увеличивает продуцирование интерферона-гамма в анализе MLR;
(e) данное антитело увеличивает секрецию IL-2 в анализе MLR;
(f) данное антитело связывается с PD-1 человека и PD-1 собакоподобной обезьяны;
(g) данное антитело ингибирует связывание PD-L1 и/или PD-L2 с PD-1;
(h) данное антитело стимулирует антигенспецифические ответные реакции памяти;
(i) данное антитело стимулирует ответные реакции антител;
(j) данное антитело ингибирует рост опухолевых клеток in vivo.
Функциональные свойства измененных антител могут оцениваться с использованием стандартных способов, доступных в данной области и/или описанных в данном описании, таких как представленные в примерах (например, проточная цитометрия, анализы связывания).
В некоторых вариантах способов конструирования антител настоящего изобретения мутации могут вводиться случайным образом или селективно вдоль всей или части кодирующей последовательности анти-PD-1-антитела, и полученные анти-PD-1-антитела могут быть подвергнуты скринингу на связывающую активность и/или другие функциональные свойства, как описано в данном описании. Мутационные способы были описаны в данной области. Например, в публикации РСТ WO 02/092780, Short описывает способы создания и скрининга мутаций антител с использованием насыщающего мутагенеза, сборки синтетическим лигированием или их комбинации. Альтернативно, в публикации РСТ WO 03/074679, Lazar et al. описывают компьютерные способы скрининга для оптимизации физико-химических свойств антител.
Молекулы нуклеиновых кислот, кодирующие антитела настоящего изобретения
Другой аспект настоящего изобретения относится к молекулам нуклеиновых кислот, которые кодируют антитела настоящего изобретения. Такие нуклеиновые кислоты могут присутствовать в цельных клетках, в лизате клеток или в частично очищенной или по существу очищенной форме. Нуклеиновая кислота является «выделенной» или «по существу очищенной» при очистке от других клеточных компонентов или других загрязнений, например, других клеточных нуклеиновых кислот или белков, стандартными способами, включающими обработку щелочью/ДСН, CsCl-бэндинг, колоночную хроматографию, электрофорез в агарозном геле и другие способы, хорошо известные в данной области. См., F. Ausubel, et al., ed. (1987) Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York. Нуклеиновая кислота настоящего изобретения может быть, например, ДНК или РНК, и может содержать или нет интронные последовательности. В предпочтительном варианте осуществления указанной нуклеиновой кислотой является молекула кДНК.
Нуклеиновые кислоты настоящего изобретения могут быть получены с использованием стандартных способов молекулярной биологии. Что касается антител, экспрессируемых гибридомами (например, гибридомами, полученными из трансгенных мышей, несущих гены иммуноглобулина человека, как описано дополнительно ниже), кДНК, кодирующие легкую и тяжелую цепи антитела, созданные гибридомой, могут быть получены стандартными способами ПЦР-амплификации или клонирования кДНК. Что касается антител, полученных из библиотеки генов иммуноглобулинов (например, использующей способы фагового дисплея), нуклеиновая кислота, кодирующая данное антитело, может быть извлечена из указанной библиотеки.
Предпочтительными молекулами нуклеиновых кислот настоящего изобретения являются молекулы нуклеиновых кислот, кодирующие последовательности VH и VL моноклональных антител 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 или 5F4. ДНК-последовательности, кодирующие последовательности VH 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4, показаны в SEQ ID NO:57, 58, 59, 60, 61, 62 и 63, соответственно. ДНК-последовательности, кодирующие последовательности VL 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4 показаны в SEQ ID NO:64, 65, 66, 67, 68, 69 и 70, соответственно.
После получения ДНК-фрагментов, кодирующих сегменты VH и VL, ДНК-фрагменты могут быть подвергнуты дополнительной манипуляции при помощи стандартных способов рекомбинантных ДНК, например, для превращения генов вариабельной области в гены полноразмерной цепи антитела, в гены Fab-фрагмента или в ген scFv. В указанных манипуляциях кодирующий фрагмент VH или VL ДНК функционально связан с другим ДНК-фрагментом, кодирующим другой белок, таким как константная область антитела или гибкий линкер. Термин «функционально связан» в данном описании означает, что два ДНК-фрагмента соединены таким образом, что аминокислотные последовательности, кодируемые такими двумя ДНК-фрагментами, остаются в рамке считывания.
Выделенная ДНК, кодирующая VH-область, может быть превращена в полноразмерный ген тяжелой цепи функциональным связыванием VH-кодирующей ДНК с другой молекулой ДНК, кодирующей константные области тяжелой цепи (CH1, CH2 и CH3). Последовательности генов константной области тяжелой цепи человека известны в данной области (см., например, Kabat, E. A., el al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242), и ДНК-фрагменты, включающие в себя указанные области, могут быть получены стандартной ПЦР-амплификацией. Константная область тяжелой цепи может быть константной областью IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно является константной областью IgG1 или IgG4. Что касается гена Fab-фрагмента, VH-кодирующая ДНК может быть функционально связана с другой молекулой ДНК, кодирующей только константную область тяжелой цепи СН1.
Выделенная ДНК, кодирующая VL-область, может быть превращена в полноразмерный ген легкой цепи (а также ген легкой цепи Fab) функциональным связыванием VL-кодирующей ДНК с другой молекулой ДНК, кодирующей константную область легкой цепи, CL. Последовательности генов константной области легкой цепи человека известны в данной области (см., например, Kabat, E.A., et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242), и ДНК-фрагменты, включающие в себя указанные области, могут быть получены стандартной ПЦР-амплификацией. Константная область легкой цепи может быть константной областью каппа или лямбда, но наиболее предпочтительно она является константной областью каппа.
Для создания гена scFv, VH и VL-кодирующие ДНК-фрагменты функционально связывают с другим фрагментом, кодирующим гибкий линкер, например, кодирующим аминокислотную последовательность (Gly4-Ser)3 таким образом, что последовательности VH и VL могут быть экспрессированы в виде непрерывного одноцепочечного белка с областями VH и VL, соединенными гибким линкером (см., например, Bird et al. (1988) Science 242:423-426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883; McCafferty et al., (1990) Nature 348:552-554).
Получение моноклональных антител настоящего изобретения
Моноклональные антитела (mAb) настоящего изобретения могут быть получены различными способами, включая общепринятую методику моноклональных антител, например, способ гибридизации соматических клеток Kohler and Milstein (1975) Nature 256: 495. Хотя предпочтительными являются методы гибридизации соматических клеток, в принципе могут быть использованы другие способы получения моноклонального антитела, например, вирусное или онкогенное превращение В-лимфоцитов.
Предпочтительной системой животного для получения гибридом является мышиная система. Получение гибридомы в мыши является хорошо установленной методикой. Протоколы и способы выделения иммунизированных спленоцитов для слияния известны в данной области. Партнеры слияния (например, мышиные миеломные клетки) и методы слияния также известны.
Химерные или гуманизированные антитела настоящего изобретения могут быть получены на основе последовательности мышиного моноклонального антитела, полученного, как описано выше. ДНК, кодирующая иммуноглобулины тяжелой и легкой цепи, может быть получена из представляющей интерес мышиной гибридомы и сконструированы таким образом, что они содержат последовательности не-мышиного иммуноглобулина (например, иммуноглобулина человека), с использованием стандартных способов молекулярной биологии. Например, для создания химерного антитела мышиные вариабельные области могут быть связаны с константными областями человека с использованием способов, известных в данной области (см., например, патент США № 4816567, Cabilly et al.). Для создания гуманизированного антитела мышиные CDR-области могут быть встроены в каркасную область человека с использованием способов, известных в данной области (см., например, патент США № 5225539, Winter, и патенты США №№ 5530101; 5585089; 5693762 и 6180370, Queen et al.).
В предпочтительном варианте осуществления антитела настоящего изобретения являются моноклональными антителами человека. Такие моноклональные антитела человека, направленные против PD-1, могут быть генерированы с использованием трансгенных или трансхромосомных мышей, несущих части иммунной системы человека, что более предпочтительно системам мыши. Такие трансгенные и трансхромосомные мыши включают мышей, называемых в данном описании мышами HuMAb и мышами КМ™, соответственно, и в целом называемых «мышами с Ig человека».
Мышь HuMAb® (Medarex, Inc.) содержит минилокусы гена иммуноглобулина человека, которые кодируют неупорядоченные последовательности тяжелой (µ и γ) и легкой цепи κ иммуноглобулина человека, вместе с нацеленными мутациями, которые инактивируют эндогенные локусы цепи µ и κ (см., например, Lonberg, et al. (1994) Nature 368(6474): 856-859). Таким образом, данные мыши обнаруживают уменьшенную экспрессию IgM мыши или κ, и в ответ на иммунизацию, введенные трансгены тяжелой и легкой цепей человека подвергаются переключению класса и соматической мутации с генерированием моноклонального IgGκ с высокой аффинностью (Lonberg, N. et al. (1994), supra; в обзоре Lonberg, N. (1994) Handbook of Experimental Pharmacology 113:49-101; Lonberg, N. and Huszar, D. (1995) Intern. Rev. Immunol. 13: 65-93, и Harding, F. and Lonberg, N. (1995) Ann. N.Y. Acad. Sci. 764:536-546). Получение и использование мышей HuMAb и геномные модификации, которые несут такие мыши, дополнительно описаны у Taylor, L. et al. (1992) в Nucleic Acids Research 20:6287-6295; Chen, J. et al. (1993) International Immunology 5: 647-656; Tuaillon et al. (1993) Proc. Natl. Acad. Sci. USA 90:3720-3724; Choi et al. (1993) Nature Genetics 4:117-123; Chen, J. et al. (1993) EMBO J. 12: 821-830; Tuaillon et al. (1994) J. Immunol. 152:2912-2920; Taylor, L. et al. (1994) International Immunology 6: 579-591; and Fishwild, D. et al. (1996) Nature Biotechnology 14: 845-851, содержание каждой из которых специально включено в данное описание посредством ссылки во всей их полноте. См. дополнительно, патенты США №№ 5545806; 5569825; 5625126; 5633425; 5789650; 5877397; 5661016; 5814318; 5874299 и 5770429; все на имя Lonberg и Kay; патент США № 5545807, Surani et al.; публикации PCT №№ WO 92/03918, WO 93/12227, WO 94/25585, WO 97/13852, WO 98/24884 и WO 99/45962, все на имя Lonberg и Kay; и публикацию PCT № WO 01/14424, Korman et al.
В другом варианте осуществления антитела человека настоящего изобретения могут быть индуцированы с использованием мыши, которая несет последовательности иммуноглобулина человека на трансгенах и трансхромосомах, такой как мышь, которая несет трансген тяжелой цепи человека и трансхромосому легкой цепи человека. Такие мыши, называемые “мышами KMTM”, подробно описаны в публикации PCT WO 02/43478, Ishida et al.
Дополнительно в данной области доступны альтернативные системы животных, экспрессирующие гены иммуноглобулина человека, и они могут быть использованы для индукции анти-PD-1-антител настоящего изобретения. Например, может быть использована альтернативная трансгенная система, называемая ксеномышью (Abgenix, Inc.); такие мыши описаны, например, в патентах США №№ 5939598; 6075181; 6114598; 6150584 и 6162963, Kucherlapati et al.
Кроме того, альтернативные трансхромосомные системы животных, экспрессирующие гены иммуноглобулина человека, доступны в данной области и могут быть использованы для индукции анти-PD-1-антител настоящего изобретения. Например, могут быть использованы мыши, несущие как трансхромосому тяжелой цепи человека, так и трансхромосому легкой цепи человека, называемые “TC-мышами”; такие мыши описаны Tomizuka et al. (2000) в Proc. Natl. Acad. Sci. USA 97:722-727. Кроме того, в данной области были описаны коровы, несущие трансхромосомы тяжелой и легкой цепей человека (Kuroiwa et al. (2002) Nature Biotechnology 20:889-894), и они могут быть использованы для индукции анти-PD-1-антител настоящего изобретения.
Моноклональные антитела человека настоящего изобретения могут быть также получены с использованием способов фагового дисплея для скрининга библиотек генов иммуноглобулинов человека. Такие способы фагового дисплея для выделения антител человека являются установленными в данной области. См., например, патенты США №№ 5223409; 5403484 и 5571698, Ladner et al.; патенты США №№ 5427908 и 5580717, Dower et al.; патенты США №№ 5969108 и 6172197, McCafferty et al.; и патенты США №№ 5885793; 6521404; 6544731; 6555313; 6582915 и 6593081, Griffiths et al.
Моноклональные антитела настоящего изобретения могут быть также получены с использованием мышей SCID, в которых иммунные клетки человека были реконструированы таким образом, что после иммунизации может быть генерирована реакция антител человека. Такие мыши описаны, например, в патентах США №№ 5476996 и 5698767, Wilson et al.
Иммунизация мышей с Ig человека
При использовании мышей с Ig человека для индуцирования антител человека настоящего изобретения, такие мыши могут быть иммунизированы очищенным или обогащенным препаратом антигена PD-1 и/или рекомбинантного PD-1 или слитого белка PD-1, как описано Lonberg, N. et al. (1994) Nature 368(6474): 856-859; Fishwild, D. et al. (1996) Nature Biotechnology 14: 845-851; и в публикациях PCT WO 98/24884 и WO 01/14424. Предпочтительно, мыши должны быть 6-16-недельными после первой инфузии. Например, очищенный или рекомбинантный препарат (5-50 мкг) антигена PD-1 может быть использован для внутрибрюшинной иммунизации мышей с Ig человека.
Подробные методики генерирования полных моноклональных антител к PD-1 человека описаны в примере 1 ниже. Кумулятивный опыт работы с различными антигенами показал, что трансгенные мыши отвечают при начальной иммунизации внутрибрюшинно (IP) антигеном в полном адъюванте Фрейнда с последующими иммунизациями IP каждую вторую неделю (всего до 6 недель) антигеном в неполном адъюванте Фрейнда. Однако обнаружено, что адъюванты, отличные от адъювантов Фрейнда, также являются эффективными. Кроме того, обнаружено, что цельные клетки в отсутствие адъюванта являются высокоиммуногенными. Иммунная реакция может быть подвергнута мониторингу в ходе протокола иммунизации, причем пробы плазмы получают ретроорбитальными кровопусканиями. Плазма может быть подвергнута скринингу при помощи ELISA (как описано ниже), и мыши с достаточными титрами анти-PD-1-иммуноглобулина человека могут быть использованы для слияний. Мыши могут быть подвергнуты внутривенному бустингу антигеном за 3 дня перед умерщвлением и удалением селезенки. Ожидается, что может быть необходимым проведение 2-3 слияний для каждой иммунизации. Обычно иммунизируют от 6 мышей до 24 мышей для каждого антигена. Обычно используют штаммы HCo7 и HCo12. Кроме того, как трансген HCo7, так и трансген HCo12 могут быть введены вместе в одну мышь, имеющую два разных трансгена тяжелой цепи человека (HCo7/HCo12). Альтернативно или дополнительно, может быть использован штамм KM-мышиTM, как описано в примере 1.
Генерирование гибридом, продуцирующих моноклональные антитела человека настоящего изобретения
Для генерирования гибридом, продуцирующих моноклональные антитела человека настоящего изобретения, спленоциты и/или клетки лимфатических узлов из иммунизированных мышей могут быть выделены и слиты с подходящей иммортализованной клеточной линией, такой как линия мышиных миеломных клеток. Полученные гибридомы могут быть подвергнуты скринингу на продуцирование антигенспецифических антител. Например, суспензии отдельных клеток лимфоцитов селезенки из иммунизированных мышей могут быть слиты до одной шестой количества несекретирующих P3X63-Ag8.653 миеломных клеток мыши (ATCC, CRL 1580) с 50% ПЭГ. Альтернативно, суспензии отдельных клеток лимфоцитов селезенки из иммунизированных мышей могут быть слиты при помощи способа электрослияния, основанного на использовании электрического поля, с использованием прибора с большой камерой для слияния клеток Cyto Pulse large chamber cell fusion electroporator (Cyto Pulse Sciences, Inc., Glen Burnie, MD). Клетки высевают приблизительно 2×105 клеток в плоскодонном микротитрационном планшете с последующим двухнедельным инкубированием в селективной среде, содержащей 20% фетальную сыворотку (Clone Serum), 18% кондиционированную среду "653", 5% ориген (IGEN), 4 мМ L-глутамин, 1 мМ пируват натрия, 5 мМ HEPES, 0,055 мМ 2-меркаптоэтанол, 50 единиц/мл пенициллина, 50 мг/мл стрептомицина, 50 мг/мл гентамицина и 1X HAT (Sigma; HAT добавляют спустя 24 часа после слияния). Спустя приблизительно две недели, клетки могут быть культивированы в среде, в которой НАТ заменена НТ. Затем отдельные лунки могут быть подвергнуты скринингу при помощи ELISA на моноклональные IgM и IgG антитела человека. После экстенсивного роста гибридом за средой можно наблюдать обычно спустя 10-14 дней. Секретирующие антитело гибридомы могут быть повторно высеяны, снова подвергнуты скринингу и, если они все еще являются положительными в отношении IgG человека, моноклональные антитела могут быть субклонированы по меньшей мере два раза с использованием лимитирующего разведения. Стабильные субклоны могут быть затем культивированы in vitro для генерирования небольших количеств антитела в среде для культуры ткани для характеристики.
Для очистки моноклональных антител человека отобранные гибридомы могут выращиваться во вращающихся колбах на два литра для очистки моноклональных антител. Супернатанты могут быть отфильтрованы и концентрированы перед аффинной хроматографией с использованием белка А-сефарозы (Pharmacia, Piscataway, N.J.). Элюированный IgG может контролироваться гель-электрофорезом и высокоэффективной жидкостной хроматографией для гарантии чистоты. Буферный раствор может быть заменен PBS и концентрация может быть определена по OD280 с использованием коэффициента экстинкции 1,43. Моноклональные антитела могут быть разделены на аликвоты и могут храниться при -80°С.
Генерирование трансфектом, продуцирующих моноклональные антитела настоящего изобретения
Антитела настоящего изобретения могут быть также получены в трансфектоме клетки-хозяина, например, с использованием комбинирования способов рекомбинантных ДНК и способов трансфекции генов, как хорошо известно в данной области (например, Morrison, S (1985) Science 229:1202).
Например, для экспрессии указанных антител или фрагментов антител, ДНК, кодирующие частичные или полноразмерные легкие и тяжелые цепи, могут быть получены стандартными способами молекулярной биологии (например, ПЦР-амплификацией или клонированием кДНК с использованием гибридомы, которая экспрессирует представляющее интерес антитело), и полученные ДНК могут быть встроены в экспрессирующие векторы таким образом, что гены функционально связаны с регуляторными последовательностями транскрипции и трансляции. В данном описании термин «функционально связанные» означает, что ген антитела лигирован в вектор таким образом, что регуляторные последовательности транскрипции и трансляции в векторе выполняют их предполагаемую функцию регуляции транскрипции и трансляции гена данного антитела. Экспрессирующий вектор и регуляторные последовательности экспрессии выбраны таким образом, что они являются совместимыми с используемой экспрессионной клеткой-хозяином. Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть встроены в отдельные векторы или, более конкретно, оба гена встроены в один и тот же экспрессирующий вектор. Гены антител встраивают в экспрессирующий вектор стандартными способами (например, лигированием комплементарных сайтов рестрикции на фрагменте гена антитела и векторе, или лигированием тупых концов, если сайты рестрикции отсутствуют). Вариабельные области легкой и тяжелой цепей антител, описанных выше, могут быть использованы для создания полноразмерных генов антител любого изотипа антитела встраиванием их в экспрессирующие векторы, уже кодирующие константные области тяжелой цепи и константные области легкой цепи желаемого изотипа таким образом, что VH-сегмент функционально связывается с СН-сегментом (сегментами) в данном векторе, и VK-сегмент функционально связывается с CL-сегментом в этом векторе. Дополнительно или альтернативно, рекомбинантный экспрессионный вектор может кодировать сигнальный пептид, который облегчает секрецию цепи антитела из клетки-хозяина. Ген цепи антитела может быть клонирован в вектор таким образом, что сигнальный пептид связывается в рамке считывания с амино-концом гена цепи антитела. Сигнальный пептид может быть сигнальным пептидом иммуноглобулина или гетерологичным сигнальным пептидом (т.е. сигнальным пептидом из белка, не являющегося иммуноглобулином).
В дополнение к генам цепей антитела, рекомбинантные экспрессирующие векторы настоящего изобретения несут регуляторные последовательности, которые регулируют экспрессию генов цепей антитела в клетке-хозяине. Термин «регуляторная последовательность» включает промоторы, энхансеры и другие элементы регуляции экспрессии (например, сигналы полиаденилирования), которые регулируют транскрипцию и трансляцию генов цепей антител. Такие регуляторные последовательности описаны, например, в Goeddel (Gene Expression Technology. Methods in Enzymology 185, Academic Press, San Diego, CA (1990)). Специалистам в данной области будет понятно, что конструкция экспрессирующего вектора, включая выбор регуляторных последовательностей, может зависеть от таких факторов, как выбор трансформируемого хозяина, уровень экспрессии желаемого белка и т.д. Предпочтительные регуляторные последовательности для экспрессии клетки-хозяина млекопитающего включают в себя вирусные элементы, которые определяют высокие уровни экспрессии белка в клетках млекопитающего, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV), вируса обезьян 40 (SV40), аденовируса (например, основной поздний промотор аденовируса (AdMLP) и полиомы. Альтернативно, могут быть использованы невирусные регуляторные последовательности, такие как промотор убиквитина или промотор β-глобина. Кроме того, могут быть использованы регуляторные элементы, составленные из последовательностей из разных источников, такие как промоторная система SRα, которая содержит последовательности из раннего промотора SV40, и длинный концевой повтор вируса типа 1 Т-клеточного лейкоза человека (Takebe, Y. et al. (1988) Mol. Cell. Biol. 8:466-472).
В дополнение к генам цепей антитела и регуляторным последовательностям, рекомбинантные экспрессирующие векторы настоящего изобретения могут нести дополнительные последовательности, например, последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, сайты инициации репликации), и гены селектируемых маркеров. Ген селектируемого маркера облегчает отбор клеток-хозяев, в которые был введен вектор (см. например, патенты США №№ U.S. Pat. 4399216, 4634665 и 5179017, все на имя Axel et al.). Например, обычно ген селектируемого маркера придает устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клетке-хозяину, в которую был введен данный вектор. Предпочтительные гены селектируемых маркеров включают в себя ген дигидрофолатредуктазы (DHFR) (для применения в dhfr-клетках-хозяевах с отбором/амплификацией в присутствии метотрексата) и ген neo (для отбора в присутствии G418).
Для экспрессии легкой и тяжелой цепей экспрессирующий вектор (экспрессирующие векторы), кодирующие тяжелую и легкую цепи, трансфицируют в клетку-хозяина стандартными способами. Имеется в виду, что различные формы термина «трансфекция» включают большое разнообразие способов, обычно используемых для введения экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например, электропорацию, осаждение фосфатом кальция, трансфекцию с использованием DEAE-декстрана и т.п. Хотя теоретически можно экспрессировать антитела настоящего изобретения в любых прокариотических или эукариотических клетках-хозяевах, наиболее предпочтительной является экспрессия антител в эукариотических клетках и, наиболее предпочтительно, в клетках-хозяевах млекопитающих, так как такие эукариотические клетки и, в частности, клетки млекопитающих, с большей вероятностью, чем прокариотические клетки, будут собираться и секретировать правильно уложенное и иммунологически активное антитело. Сообщалось, что прокариотическая экспрессия генов антител является неэффективной для получения высоких выходов активного антитела (Boss, M.A. and Wood, C. R. (1985) Immunology Today 6:12-13).
Предпочтительные клетки-хозяева млекопитающих для экспрессии рекомбинантных антител настоящего изобретения включают в себя клетки яичника китайского хомячка (клетки CHO) (включая dhfr-CHO-клетки, описанные Urlaub and Chasin, (1980) Proc. Natl. Acad. Sci. USA 77:4216-4220, используемые с DHFR-селектируемым маркером, например, как описано R.J. Kaufman and P.A. Sharp (1982) в Mol. Biol. 159:601-621), клетки миеломы NSO, клетки COS и клетки SP2. В частности, для применения с миеломными клетками NSO, другой предпочтительной системой экспрессии является система экспрессии гена GS, описанная в WO 87/04462, WO 89/01036 и EP 338841. При введении рекомбинантных экспрессирующих векторов, кодирующих гены антител, в клетки-хозяина млекопитающих, антитела получают культивированием клеток-хозяев в течение времени, достаточного для достижения прохождения экспрессии антитела в клетках-хозяевах или, более предпочтительно, секреции антитела в культуральную среду, в которой выращиваются клетки-хозяева. Антитела могут быть извлечены из культуральной среды с использованием стандартных методов очистки белка.
Характеристика связывания антител с антигеном
Антитела настоящего изобретения могут быть испытаны в отношении связывания с PD-1, например, при помощи стандартного ELISA. Кратко, микротитрационные планшеты покрывают очищенным PD-1 при 0,25 мкг/мл в PBS и затем блокируют 5% бычьим сывороточным альбумином в PBS. Разведения антитела (например, разведения плазмы из иммунизированных PD-1 мышей) добавляют в каждую лунку и инкубируют в течение 1-2 часов при 37°С. Планшеты промывают смесью PBS/Твин и затем инкубируют со вторым реагентом (например, для антител человека, поликлональными козьими антителами, специфическими в отношении IgG-Fc человека), конъюгированным со щелочной фосфатазой, в течение 1 часа при 37°С. После промывания планшеты проявляют с субстратом pNPP (1 мг/мл) и анализируют при OD 405-650. Предпочтительно, мыши, которые развивают наивысшие титры, будут использоваться для слияний.
Анализ ELISA, описанный выше, может быть также использован для скрининга на гибридомы, которые обнаруживают положительную реактивность с иммуногеном PD-1. Гибридомы, которые связываются с высокой авидностью с PD-1, субклонируют и дополнительно характеризуют. Один клон из каждой гибридомы, который сохраняет реактивность исходных клеток (согласно ELISA), может быть выбран для создания банка клеток из 5-10 флаконов, хранящегося при -140°С, и для очистки антитела.
Для очистки анти-PD-1-антител отобранные гибридомы могут быть выращены во вращающихся колбах на два литра для очистки моноклональных антител. Супернатанты могут быть отфильтрованы и концентрированы перед аффинной хроматографией с использованием белка А-сефарозы (Pharmacia, Piscataway, N.J.). Элюированный IgG может контролироваться гель-электрофорезом и высокоэффективной жидкостной хроматографией для гарантии чистоты. Буферный раствор может быть заменен на PBS и концентрация может быть определена по OD280 с использованием коэффициента экстинкции 1,43. Моноклональные антитела могут быть разделены на аликвоты и могут храниться при -80°С.
Для определения, связываются ли отобранные моноклональные анти-PD-1-антитела с уникальными эпитопами, каждое антитело может быть биотинилировано с использованием коммерчески доступных реагентов (Pierce, Rockford, IL). Конкурентные исследования с использованием немеченых моноклональных антител и биотинилированных моноклональных антител могут быть проведены с использованием покрытых PD-1 планшетов ELISA, как описано выше. Связывание биотинилированного mAb может быть детектировано с использованием зонда, содержащего стрептавидин-щелочную фосфатазу.
Для определения изотипа очищенных антител ELISA могут быть проведены с использованием реагентов, специфических в отношении антител конкретного изотипа. Например, для определения изотипа моноклонального антитела человека, лунки микротитрационного планшета могут быть покрыты 1 мкг/мл антитела против иммуноглобулина человека и оставлены при 4°С в течение ночи. После блокирования 1% BSA планшеты подвергают взаимодействию с 1 мкг/мл или менее тестируемых моноклональных антител или очищенными контролями изотипа, при температуре окружающей среды в течение одного или двух часов. Затем лунки могут быть подвергнуты взаимодействию со специфическими либо в отношении IgG1 человека, либо в отношении IgM человека, конъюгированными со щелочной фосфатазой зондами. Планшеты визуализируют и анализируют, как описано выше.
Анти-PD-1-IgG человека могут быть дополнительно испытаны на реактивность с антигеном PD-1 при помощи Вестерн-блоттинга. Кратко, PD-1 может быть получен и подвергнут электрофорезу в додецилсульфатполиакриламидном геле. После электрофореза разделенные антигены переносят на нитроцеллюлозные мембраны, блокируют 10% фетальной телячьей сывороткой и зондируют тестируемыми моноклональными антителами. Связывание IgG человека может быть детектировано с использованием комплекса антитело против IgG человека - щелочная фосфатаза и обнаружено с помощью таблеток субстрата BCIP/NBT (Sigma Chem. Co., St. Louis, Mo.).
Иммуноконъюгаты
В другом аспекте настоящее изобретение описывает анти-PD-1-антитело или его фрагмент, конъюгированные с терапевтической частью молекулы, такой как цитотоксин, лекарственное средство (например, иммуносупрессор) или радиотоксин. Такие конъюгаты называют «иммуноконъюгатами». Иммуноконъюгаты, которые включают один или несколько цитотоксинов, называют «иммунотоксинами». Цитотоксин или цитотоксический агент включает в себя любой агент, который является вредным для клеток (например, убивает клетки). Примеры включают в себя таксол, цитохалазин В, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин, и их аналоги или гомологи. Терапевтические агенты включают также, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, цитарабин, 5-фторурацил-декарбазин), алкилирующие агенты (например, мехлорэтамин, тиоэпахлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин С и цис-дихлордиаминплатина (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (АМС)) и антимитотические агенты (например, винкристин и винбластин).
Другие предпочтительные примеры терапевтических цитотоксинов, которые могут быть конъюгированы с антителом настоящего изобретения, включают дуокармицины, калихеамицины, майтансины и ауристатины, и их производные. Один пример конъюгата калихеамицин-антитело является коммерчески доступным (Mylotarg™; Wyeth-Ayerst).
Цитотоксины могут быть конъюгированы с антителами настоящего изобретения с использованием линкерной технологии, доступной в данной области. Примеры типов линкеров, которые используют для конъюгирования цитотоксина с антителом, включают в себя, но не ограничиваются ими, гидразоны, простые тиоэфиры, сложные эфиры, дисульфиды и пептидсодержащие линкеры. Может быть выбран линкер, который, например, является чувствительным к расщеплению при низком рН в лизосомном компартменте или чувствительным к расщеплению протеазами, например протеазами, предпочтительно экспрессируемыми в опухолевой ткани, такими как катепсины (например, катепсины В, С, D).
В отношении дополнительного обсуждения типов цитотоксинов, линкеров и способов конъюгации терапевтических агентов с антителами см. также Saito, G. et al. (2003) Adv. Drug Deliv. Rev. 55:199-215; Trail, P.A. et al. (2003) Cancer Immunol. Immunother. 52:328-337; Payne, G. (2003) Cancer Cell 3:207-212; Allen, T.M. (2002) Nat. Rev. Cancer 2:750-763; Pastan, I. and Kreitman, R.J. (2002) Curr. Opin. Investig. Drugs 3:1089-1091; Senter, P.D. and Springer, C.J. (2001) Adv. Drug Deliv. Rev. 53:247-264.
Антитела настоящего изобретения могут быть также конъюгированы с радиоактивным изотопом для генерирования цитотоксических радиофармацевтических веществ, также называемых радиоиммуноконъюгатами. Примеры радиоактивных изотопов, которые могут быть конъюгированы с антителами для диагностического или терапевтического применения, включают, но не ограничиваются ими, иод131, индий111, иттрий90 и лютеций177. Способы получения радиоиммуноконъюгатов являются установленными в данной области. Примеры радиоиммуноконъюгатов являются коммерчески доступными, включая Zevalin™ (IDEC Pharmaceuticals) и Bexxar™ (Corixa Pharmaceuticals), и сходные способы могут быть использованы для получения радиоиммуноконъюгатов с использованием антител настоящего изобретения.
Конъюгаты антител настоящего изобретения могут быть использованы для модификации конкретной биологической ответные реакции, и лекарственная часть молекулы не должна быть ограничена классическими химическими терапевтическими агентами. Например, лекарственной частью молекулы может быть белок или полипептид, обладающий желаемой биологической активностью. Такие белки могут включать в себя, например, ферментативно активный токсин или его активный фрагмент, такой как абрин, рицин А, экзотоксин Pseudomonas или дифтерийный токсин; такой белок, как фактор некроза опухолей или интерферон-γ; или модификаторы биологической реакции, такие как, например, лимфокины, интерлейкин-1 (“IL-1”), интерлейкин-2 ("IL-2"), интерлейкин-6 (“IL-6"), колониестимулирующий фактор гранулоцитов-макрофагов (“GM-CSF"), колониестимулирующий фактор гранулоцитов (“G-CSF") или другие факторы роста.
Способы конъюгирования такой терапевтической части молекулы с антителами хорошо известны, см., например, Arnon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Antibodies For Drug Delivery", in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", in Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera et al. (eds.), pp. 475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", in Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), и Thorpe et al., "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-58 (1982).
Биспецифические молекулы
В другом аспекте настоящее изобретение описывает биспецифические молекулы, содержащие анти-PD-1-антитело или его фрагмент настоящего изобретения. Антитело настоящего изобретения или его антигенсвязывающие части могут быть дериватизованы или связаны с другой функциональной молекулой, например, другим пептидом или белком (например, другим антителом или лигандом для рецептора) с получением биспецифической молекулы, которая связывается по меньшей мере с двумя различными сайтами связывания или молекулами-мишенями. Антитело настоящего изобретения может быть фактически дериватизировано или связано с более чем одной, функциональной молекулой, с получением полиспецифических молекул, которые связываются с более чем двумя различными сайтами связывания и/или молекулами-мишенями; предполагается, что такие полиспецифические молекулы включены в термин «биспецифическая молекула» данного описания. Для создания биспецифической молекулы настоящего изобретения антитело настоящего изобретения может быть функционально связано (например, химическим связыванием, генетическим слиянием, нековалентной ассоциацией или другим образом) с одной или несколькими другими связывающими молекулами, такими как другое антитело, фрагмент антитела, пептид или связывающий миметик таким образом, что образуется биспецифическая молекула.
Таким образом, настоящее изобретение включает биспецифические молекулы, содержащие по меньшей мере одну первую специфичность связывания в отношении PD-1 и вторую специфичность связывания в отношении второго эпитопа-мишени. В конкретном варианте осуществления настоящего изобретения вторым эпитопом-мишенью является рецептор Fc, например, FcγRI человека (CD64) или рецептор Fcα человека (CD89). Таким образом, настоящее изобретение включает биспецифические молекулы, способные связываться как с FcγR или FcαR экспрессирующими эффекторными клетками (например, моноцитами, макрофагами или полиморфонуклеарными клетками (PMN), так и с клетками-мишенями, экспрессирующими PD-1. Такие биспецифические молекулы нацеливают экспрессирующие PD-1 клетки на эффекторную клетку и запускают опосредованные рецептором Fc активности эффекторных клеток, такие как фагоцитоз экспрессирующих PD-1 клеток, антителозависимую клеточноопосредованную цитотоксичность (ADCC), высвобождение цитокинов или генерирование супероксидного аниона.
В одном варианте осуществления настоящего изобретения, в котором биспецифическая молекула является полиспецифической, данная молекула может дополнительно включать третью специфичность связывания и специфичность анти-PD-1-связывания. В одном варианте осуществления третьей специфичностью связывания является часть фактора анти-усиления (EF), например, молекула, которая связывается с поверхностным белком, участвующим в цитотоксической активности, и усиливает, таким образом, иммунную реакцию против клетки-мишени. «Частью фактора анти-усиления (EF)» может быть антитело, функциональный фрагмент антитела или лиганд, который связывается с конкретной молекулой, например, антигеном или рецептором, и приводит, таким образом, к усилению эффекта связывающих детерминант для рецептора Fc или клеточного антигена-мишени. «Часть фактора анти-усиления (EF)» может связываться с рецептором Fc или клеточным антигеном-мишенью. Альтернативно, «часть фактора анти-усиления» может связываться с частицей, которая отличается от частицы, с которой связываются первая и вторая специфичности связывания. Например, «часть фактора анти-усиления» может связывать цитотоксическую Т-клетку (например, через CD2, CD3, CD8, CD28, CD4, CD40, ICAM-1 или другую иммунную клетку в увеличенной реакции против клетки-мишени).
В одном варианте осуществления биспецифические молекулы настоящего изобретения содержат в качестве специфичности связывания по меньшей мере одно антитело или фрагмент антитела, включая, например, Fab, Fab', F(ab')2, Fv или одноцепочечный Fv. Данное антитело может быть димером легкой цепи или тяжелой цепи или любого минимального фрагмента антитела, такого как Fv или любая одноцепочечная конструкция, как описано Ladner et al. в патенте США No. 4946778, содержание которого включено в данное описание посредством ссылки.
В одном варианте осуществления специфичность связывания для рецептора Fcγ обеспечивается моноклональным антителом, связывание которого не блокируется иммуноглобулином G человека (IgG). Как использовано в данном описании, термин “рецептор IgG" относится к любому из восьми генов γ-цепи, локализованным на хромосоме 1. Такие гены кодируют в целом двенадцать трансмембранных или растворимых изоформ рецептора, которые сгруппированы в три класса рецепторов Fcγ: FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16). В одном предпочтительном варианте осуществления рецептором FcγR является FcγRI человека с высокой аффинностью. FcγRI человека является молекулой 72 кДа, которая обнаруживает высокую аффинность в отношении мономерного IgG (108-109М-1).
Получение и характеристика некоторых предпочтительных моноклональных анти-Fcγ-антител описаны Fanger et al. в публикации PCT WO 88/00052 и в патенте США № 4954617, описания которых включены в данное описание посредством ссылки во всей своей полноте. Данные антитела связываются с эпитопом FcγRI, FcγRII или FcγRIII в сайте, который отличается от сайта связывания Fcγ данного рецептора, и, следовательно, их связывание по существу не блокируется физиологическими уровнями IgG. Специфическими анти-FcγRI-антителами, применимыми в данном изобретении, являются mAb 22, mAb 32, mAb 44, mAb 62 и mAb 197. Гибридома, продуцирующая mAb 32, доступна из Американской Коллекции Типовых Культур, ATCC Accession No. (номер доступа) HB9469. В других вариантах осуществления анти-рецептор Fcγ-антителом является гуманизированная форма моноклонального антитела 22 (H22). Получение и характеристика антитела Н22 описаны Graziano, R.F. et al. (1995) в J. Immunol 155 (10): 4996-5002 и публикации PCT WO 94/10332. Продуцирующая антитело H22 клеточная линия была депонирована в Американской Коллекции Типовых Культур под номером HA022CL1 и имеет номер доступа CRL 11177.
В других предпочтительных вариантах осуществления специфичность связывания в отношении рецептора Fc обеспечена антителом, которое связывается с рецептором IgA человека, например, рецептором Fc-альфа (FcαRI (CD89)), связывание которого предпочтительно не блокируется иммуноглобулином А человека (IgA). Термин “рецептор IgA" включает генный продукт одного α-гена (FcαRI), расположенного на хромосоме 19. Известно, что указанный ген кодирует несколько альтернативно сплайсированных трансмембранных изоформ 55-110 кДа. FcαRI (CD89) конститутивно экспрессируется на популяциях моноцитов/макрофагов, эозинофильных и нейтрофильных гранулоцитов, но не на популяциях неэффекторных клеток. FcαRI имеет среднюю аффинность (≈5×107M-1) в отношении как IgA1, так и IgA2, которая увеличивается после воздействия цитокинов, таких как G-CSF или GM-CSF (Morton, H.C. et al. (1996) Critical Reviews in Immunology 16:423-440). Были описаны четыре FcαRI-специфических моноклональных антитела, идентифицированных как A3, A59, A62 и A77, которые связывают FcαRI вне домена связывания лиганда IgA (Monteiro, R.C. et al. (1992) J. Immunol. 148:1764).
FcαRI и FcγRI являются запускающими рецепторами для применения в биспецифических молекулах настоящего изобретения, так как они (1) экспрессируются первично на иммунных эффекторных клетках, например, моноцитах, PMN, макрофагах и дендритных клетках; (2) экспрессируются при высоких уровнях (например, 5000-100000 на клетку); (3) являются медиаторами цитотоксических активностей (например, ADCC, фагоцитоза); (4) опосредуют усиленную презентацию антигенов, включая аутоантигены, которые являются их мишенями.
Хотя предпочтительными являются моноклональные антитела человека, другими антителами, которые могут использоваться в биспецифических антителах, являются мышиные, химерные и гуманизированные моноклональные антитела.
Биспецифические молекулы настоящего изобретения могут быть получены конъюгацией составляющих специфичностей связывания, например, анти-FcR- и анти-PD-1-специфичностей связывания, с использованием известных в данной области способов. Например, каждая специфичность связывания биспецифической молекулы может быть получена отдельно, и затем они могут быть конъюгированы друг с другом. Когда специфичностями связывания являются белки или пептиды, разнообразные связывающие или сшивающие агенты могут быть использованы для ковалентной конъюгации. Примеры сшивающих агентов включают белок A, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 5,5'-дитиобис(2-нитробензойную кислоту) (DTNB), o-фенилендималеимид (oPDM), N-сукцинимидил-3-(2-пиридилтио)пропионат (SPDP) и сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC) (см., например, Karpovsky et al. (1984) J. Exp. Med. 160:1686; Liu, MA et al. (1985) Proc. Natl. Acad. Sci. USA 82:8648). Другие способы включают способы, описанные в Paulus (1985) Behring Ins. Mitt. No. 78, 118-132; Brennan et al. (1985) Science 229:81-83) и Glennie et al. (1987) J. Immunol. 139: 2367-2375). Предпочтительными конъюгирующими агентами являются SATA и сульфо-SMCC, оба доступные от Pierce Chemical Co. (Rockford, IL).
Когда специфичностями связывания являются антитела, они могут быть конъюгированы через образование сульфгидрильной связи шарнирных областей С-концов двух тяжелых цепей. В особенно предпочтительном варианте осуществления шарнирная область модифицирована таким образом, что она содержит нечетное число сульфгидрильных остатков, предпочтительно один сульфгидрильный остаток, перед конъюгацией.
Альтернативно, обе специфичности связывания могут быть кодированы в одном и том же векторе и экспрессированы и собраны в одной и той же клетке-хозяине. Данный способ особенно полезен, когда биспецифической молекулой является mAb×mAb, mAb×Fab, Fab×F(ab')2 или лиганд×Fab-слитый белок. Биспецифической молекулой настоящего изобретения может быть одноцепочечная молекула, содержащая одно одноцепочечное антитело и детерминанту связывания, или одноцепочечная биспецифическая молекула, содержащая две детерминанты связывания. Биспецифические молекулы могут содержать по меньшей мере две одноцепочечные молекулы. Способы получении биспецифических молекул описаны, например, в патенте США № 5260203; патенте США № 5455030; патенте США № 4881175; патенте США № 5132405; патенте США № 5091513; патенте США № 5476786; патенте США № 5013653; патенте США № 5258498 и патенте США № 5482858.
Связывание биспецифических молекул с их специфическими мишенями может быть подтверждено, например, твердофазным иммуноферментным анализом (ELISA), радиоиммуноанализом (RIA), FACS-анализом, биоанализом (например, ингибированием роста) или Вестерн-блот-анализом. Каждый из указанных анализов обычно детектирует присутствие представляющих конкретный интерес комплексов белок-антитело с использованием меченого реагента (например, антитела), специфического в отношении представляющего интерес комплекса. Например, комплексы FcR-антитело могут быть детектированы с использованием, например, ферментсвязанного антитела или фрагмента антитела, которые распознают комплексы антитело-Fcγ и специфически связываются с ними. Альтернативно, комплексы могут быть детектированы с использованием различных других иммуноанализов. Например, антитело может быть радиоактивно меченым и может быть использовано в радиоиммуноанализе (RIA) (см., например, Weintraub, B., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986, которая включена в данное описание посредством ссылки). Радиоактивный изотоп может быть детектирован такими средствами, как использование γ-счетчика или сцинтилляционного счетчика, или авторадиографией.
Фармацевтические композиции
В другом аспекте настоящее изобретение обеспечивает композицию, например, фармацевтическую композицию, содержащую одно моноклональное антитело или комбинацию моноклональных антител, или их антигенсвязывающую часть (антигенсвязывающие части), настоящего изобретения, приготовленные вместе с фармацевтически приемлемым носителем. Такие композиции могут содержать одно антитело или комбинацию (например, два или более различных) антител, или иммуноконъюгаты, или биспецифические молекулы настоящего изобретения. Например, фармацевтическая композиция настоящего изобретения может содержать комбинацию антител (или иммуноконъюгаты, или биспецифические антитела), которые связываются с разными эпитопами на антигене-мишени или которые имеют комплементарные (дополняющие) активности.
Фармацевтические композиции настоящего изобретения также могут быть введены в комбинированной терапии, т.е. комбинированные с другими агентами. Например, комбинированная терапия может включать анти-PD-1-антитело настоящего изобретения, комбинированное по меньшей мере с одним другим противовоспалительным или иммуносупрессорным агентом. Примеры терапевтических агентов, которые могут быть использованы в комбинированной терапии, описаны более подробно ниже в разделе о применении антител настоящего изобретения.
Как использовано в данном описании, «фармацевтически приемлемый носитель» включает любые и все растворители, задерживающие абсорбцию агенты и т.п., которые являются физиологически совместимыми. Предпочтительно, носитель является подходящим для внутривенного, внутримышечного, подкожного, парентерального, спинномозгового или эпидермального введения (например, инъекцией или инфузией). В зависимости от способа введения, активное соединение, т.е. антитело, иммуноконъюгат или биспецифическая молекула, могут быть покрыты материалом для защиты данного соединения от действия кислот и других природных условий, которые могут инактивировать данное соединение.
Фармацевтические соединения настоящего изобретения могут включать в себя одну или несколько фармацевтически приемлемых солей. «Фармацевтически приемлемая соль» означает соль, которая сохраняет желаемую биологическую активность исходного соединения и не вызывает никаких нежелательных токсикологических эффектов (см., например, Berge, S.M., et al. (1977) J. Pharm. Sci. 66:1-19). Примеры таких солей включают кислотно-аддитивные соли и основно-аддитивные соли. Кислотно-аддитивные соли включают соли, образованные из нетоксичных неорганических кислот, таких как хлористоводородная, азотная, фосфорная, серная, бромистоводородная, иодистоводородная, фосфористая и т.п., а также из нетоксичных органических кислот, таких как алифатические моно- и дикарбоновые кислоты, фенилзамещенные алкановые кислоты, гидроксиалкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты и т.п. Основно-аддитивные соли включают соли, образованные из щелочных и щелочноземельных металлов, таких как натрий, калий, магний, кальций и т.п., а также из нетоксичных органических аминов, таких как N,N'-дибензилэтилендиамин, N-метилглюкамин, хлорпрокаин, холин, диэтаноламин, этилендиамин, прокаин и т.п.
Фармацевтические композиции настоящего изобретения могут содержать также фармацевтически приемлемый антиоксидант. Примеры фармацевтически приемлемых антиоксидантов включают: (1) водорастворимые антиоксиданты, такие как аскорбиновая кислота, гидрохлорид цистеина, бисульфат натрия, метабисульфит натрия, сульфат натрия и т.п.; (2) растворимые в масле антиоксиданты, такие как аскорбилпальмитат, бутилированный гидроксианизол (ВНА), бутилированный гидрокситолуол (ВНТ), лецитин, пропилгаллат, альфа-токоферол и т.п.; и (3) образующие хелаты металлов агенты, такие как лимонная кислота, этилендиаминтетрауксусная кислота (EDTA), сорбит, винная кислота, фосфорная кислота и т.п.
Примеры подходящих водных и неводных носителей, которые могут быть использованы в фармацевтических композициях настоящего изобретения, включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.) и их подходящие смеси, растительные масла, такие как оливковое масло, и инъецируемые органические эфиры, такие как этилолеат. Подходящая текучесть может поддерживаться, например, применением материалов покрытий, таких как лецитин, поддержанием требуемого размера частиц в случае дисперсий и применением поверхностно-активных веществ.
Композиции также могут содержать адъюванты, такие как консерванты, увлажняющие агенты, эмульгирующие агенты и диспергирующие агенты. Предотвращение присутствия микроорганизмов может гарантироваться процедурами стерилизации, supra, и включением различных антибактериальных и противогрибковых агентов, например, парабена, хлорбутанола, фенолсорбиновой кислоты и т.п. Может быть желательным включение в композиции изотонических агентов, таких как сахара, хлорид натрия и т.п.,. Кроме того, пролонгированная абсорбция инъектируемой фармацевтической формы может осуществлена включением агентов, которые замедляют абсорбцию, таких как моностеарат алюминия и желатин.
Фармацевтически приемлемые носители включают в себя стерильные растворы или дисперсии и стерильные порошки для немедленного приготовления стерильных инъекционных растворов или дисперсии. Применение таких сред и агентов для фармацевтически активных веществ известно в данной области. За исключением случаев, когда какие-либо общепринятые среда или агент являются несовместимыми с активным соединением, рассматривается его применение в фармацевтической композиции настоящего изобретения. В композиции могут быть также включены дополнительные активные соединения.
Терапевтические композиции обычно должны быть стерильными и стабильными в условиях приготовления и хранения. Композиция может быть приготовлена в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, пригодной для высокой концентрации лекарственного средства. Носителем может быть растворитель или дисперсионная среда, содержащая, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Необходимая текучесть может поддерживаться, например, с использованием покрытия, такого как лецитин, поддержанием требуемого размера частиц в случае дисперсии и с использованием поверхностно-активных веществ. Во многих случаях, будет предпочтительно включать в композицию изотонические агенты, например, сахара, полиспирты, такие как маннит, сорбит, или хлорид натрия. Пролонгированная абсорбция инъектируемых композиций может быть осуществлена включением агентов, которые замедляют абсорбцию, таких как моностеаратные соли и желатин.
Стерильные инъекционные растворы могут быть приготовлены включением активного соединения в требуемом количестве в подходящем растворителе с одним ингредиентом или несколькими ингредиентами, перечисленными выше, при необходимости, с последующей микрофильтрацией. Обычно, дисперсии готовят включением активного соединения в стерильный наполнитель, который содержит дисперсионную среду в качестве основы и требуемые другие ингредиенты из перечисленных выше ингредиентов. В случае стерильных порошков для приготовления стерильных инъекционных растворов, предпочтительными способами приготовления являются сушка в вакууме и сушка вымораживанием (лиофилизация), которые дают порошок активного ингредиента с любым дополнительным желаемым ингредиентом из его предварительно стерильно-отфильтрованного раствора.
Количество активного ингредиента, которое может быть объединено с материалом-носителем для приготовления отдельной дозированной формы, будет варьироваться в зависимости от подлежащего лечению субъекта и конкретного способа введения. Количество активного ингредиента, которое может быть объединено с материалом-носителем для приготовления отдельной дозированной формы будет обычно количеством композиции, которое производит терапевтическое действие. Обычно, в расчете на сто процентов, это количество будет находиться в диапазоне приблизительно от 0,01 процента до приблизительно девяносто девяти процентов активного ингредиента, предпочтительно приблизительно от 0,1 процента до приблизительно 70 процентов, наиболее предпочтительно приблизительно от 1 процента до приблизительно 30 процентов активного ингредиента в комбинации с фармацевтически приемлемым носителем.
Схемы введения доз приспосабливают для обеспечения оптимальной желаемой ответной реакции (например, терапевтической реакции). Например, может вводиться единственная ударная доза (болюс), несколько разделенных доз на протяжении времени или доза может быть пропорционально уменьшена или увеличена в зависимости от случаев острой необходимости терапевтической ситуации. Парентеральные композиции особенно предпочтительно готовить в единичных дозированных формах для легкости введения и однородности дозы. «Единичная дозированная форма» в данном описании означает физически дискретные единицы, подходящие в качестве разовой дозы для субъектов, которые должны подвергаться лечению; каждая единица дозы содержит заданное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта, вместе с требуемым фармацевтическим носителем. Спецификация в отношении единичных дозированных форм настоящего изобретения диктуется (а) уникальными характеристиками активного соединения и конкретным терапевтическим эффектом, который должен быть достигнут и зависит от них, и (b) ограничениями, присущими в области компаундирования такого активного соединения, для воздействия на чувствительность у индивидуумов.
Для введения антитела доза находится в диапазоне приблизительно 0,0001-100 мг/кг и более часто 0,01-5 мг/кг массы тела хозяина. Например, дозы могут составлять 0,3 мг/кг массы тела, 5 мг/кг массы тела или 10 мг/кг массы тела или находиться в диапазоне 1-10 мг/кг. Примерная схема лечения включает введение один раз в неделю, один раз каждые две недели, один раз каждые три недели, один раз каждые четыре недели, один раз в месяц, один раз каждые 3 месяца или один раз каждые 3-6 месяцев. Предпочтительные схемы введения доз для анти-PD-1-антитела настоящего изобретения включают 1 мг/кг массы тела или 3 мг/кг массы тела внутривенным введением, причем данное антитело вводят с использованием одной из следующих схем введения доз: (i) каждые четыре недели шесть доз, затем каждые три месяца; (ii) каждые три недели; (iii) 3 мг/кг массы тела один раз и затем 1 мг/кг массы тела каждые три недели.
В некоторых способах, два или более моноклональных антител с разными специфичностями связывания вводят одновременно, и в этом случае доза каждого антитела находится в указанных диапазонах. Антитело обычно вводят в виде множественных введений. Интервалы между отдельными дозами могут быть, например, недельными, месячными, каждые три месяца или один раз в год. Интервалы могут быть также нерегулярными, в зависимости от измерения уровней в крови антитела к антигену в конкретном пациенте. В некоторых способах дозу корректируют для достижения концентрации антитела в плазме приблизительно 1-1000 мкг/мл и в некоторых способах приблизительно 25-300 мкг/мл.
Альтернативно, антитело может вводиться в виде формы непрерывного высвобождения, и в этом случае требуется менее частое введение. Доза и частота введения варьируются в зависимости от периода полусуществования антитела у пациента. Обычно, антитела человека обнаруживают самый длинный период полусуществования, за ними следуют гуманизированные антитела, химерные антитела и антитела не человека. Доза и частота введения могут варьироваться в зависимости от того, является ли лечение профилактическим или терапевтическим. В профилактических применениях, вводят относительно низкую дозу при относительно нечастых интервалах на протяжении продолжительного периода времени. Некоторые пациенты продолжают получать лечение в течение всей их оставшейся жизни. В терапевтических применениях, иногда требуется относительно высокая доза при относительно коротких интервалах, пока прогрессирование заболевания не уменьшается или не терминируется, и, предпочтительно, пока пациент не обнаружит частичное или полное уменьшение симптомов заболевания. После этого пациент может получать дозы в соответствии с профилактической схемой введения.
Фактические уровни доз активных ингредиентов в фармацевтических композициях настоящего изобретения могут варьироваться таким образом, чтобы получить количество активного ингредиента, которое является эффективным для достижения желаемой терапевтической реакции для конкретного пациента, конкретной композиции и конкретного способа введения, без проявления токсичности в отношении данного пациента. Выбранный уровень дозы будет зависеть от различных фармакокинетических факторов, включающих активность конкретных используемых композиций настоящего изобретения или их сложного эфира, соли или амида, способ введения, время введения, скорость выведения конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, используемые в комбинации с применяемыми конкретными композициями, возраст, пол, массу, состояние, общее здоровье и предыдущую историю болезни пациента, и подобные факторы, хорошо известные в данных областях медицины.
«Терапевтически эффективная доза» анти-PD-1-антитела настоящего изобретения предпочтительно приводит к уменьшению тяжести симптомов заболевания, увеличению частоты и продолжительности бессимптомных периодов или предупреждению ухудшения или потери трудоспособности вследствие данной болезни. Например, для лечения опухолей, «терапевтически эффективная доза» предпочтительно ингибирует рост клеток или рост опухоли по меньшей мере приблизительно на 20%, более предпочтительно по меньшей мере приблизительно на 40%, даже более предпочтительно по меньшей мере приблизительно на 60% и еще более предпочтительно по меньшей мере приблизительно на 80% относительно не подвергнутых лечению субъектов. Способность соединения ингибировать рост опухоли может оцениваться на системе модели животного, предсказывающей эффективность в опухолях человека. Альтернативно, такое свойство композиции может быть оценено испытанием способности данного соединения к ингибированию, и такие анализы ингибирования in vitro известны квалифицированному практику. Терапевтически эффективное количество терапевтического соединения может уменьшать размер опухоли или иным образом ослаблять симптомы у субъекта. Специалист в данной области сможет определить такие количества на основе таких факторов, как размер субъекта, тяжесть симптомов у субъекта и конкретная выбранная композиция или выбранный способ введения.
В другом аспекте настоящее изобретение обеспечивает фармацевтический набор компонентов, содержащий анти-PD-1-антитело и анти-CTLA-4-антитело, описанные в данном описании. Набор может дополнительно содержать инструкции для применения в лечении гиперпролиферативного заболевания (такого как рак, как описано здесь). В другом варианте осуществления анти-PD-1 и анти-CTLA-4-антитела могут быть совместно упакованы в единичной дозированной форме.
В некоторых вариантах осуществления два или несколько моноклональных антител с различными специфичностями связывания (например, анти-PD-1-антитело и анти-CTLA-4-антитело) вводят одновременно, и в этом случае доза каждого вводимого антитела находится в указанных диапазонах. Антитело может вводиться в виде единственной дозы или, более часто, может вводиться в виде множественных введений. Интервалы между отдельными единственными дозами могут составлять один раз в неделю, один раз в месяц, каждые три месяца или один раз в год. Интервалы могут быть также нерегулярными, в соответствии с измерениями уровней в крови антитела к антигену-мишени у пациента. В некоторых способах доза корректируется таким образом, чтобы получать концентрацию антитела в плазме приблизительно 1-1000 мкг/мл, и в некоторых способах приблизительно 25-300 мкг/мл.
Композиция настоящего изобретения может вводиться посредством одного или нескольких способов введения с использованием одного или нескольких различных способов, известных в данной области. Как будет понятно специалисту в данной области, способ и/или схема введения будут варьироваться в зависимости от желаемых результатов. Предпочтительные способы введения антител настоящего изобретения включают внутривенный, внутримышечный, внутрикожный, внутрибрюшинный, подкожный, спинномозговой или другие парентеральные способы введения, например, инъекцией или инфузией. Фраза «парентеральное введение» означает в данном описании способы введения, другие, чем энтеральное и местное введение, обычно посредством инъекции, и включает, но без ограничения, внутривенную, внутримышечную, внутриартериальную, внутриоболочечную, внутрикапсулярную, внутриглазную, внутрисердечную, внутрикожную, внутрибрюшинную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, субкапсулярную, субарахноидальную, вводимую в спинномозговой канал, эпидуральную и надчревную инъекцию и инфузию.
Альтернативно, антитело настоящего изобретения может вводиться не-парентеральным способом, таким как местный, эпидермальный способ или способ введения через слизистую оболочку, например, интраназально, перорально, вагинально, ректально, сублингвально или местно.
Активные соединения могут быть приготовлены с носителями, которые будут защищать данное соединение против быстрого высвобождения, например формы регулируемого высвобождения, включая имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Могут быть использованы биодеградируемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Многие способы приготовления таких форм запатентованы или обычно известны специалистам в данной области. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Терапевтические композиции могут вводиться медицинскими устройствами, известными в данной области. Например, в предпочтительном варианте осуществления терапевтическая композиция настоящего изобретения может водиться безигольным устройством для подкожных инъекций, таким как устройства, описанные в патентах США №№ 5399163; 5383851; 5312335; 5064413; 4941880; 4790824 или 4596556. Примеры хорошо известных имплантатов и модулей, применимых в данном изобретении, включают: патент США № 4487603, в котором описан имплантируемый микроинфузионный насос для введения лекарственного средства при регулируемой скорости; патент США № 4486194, в котором описано терапевтическое устройство для введения лекарственного средства через кожу; патент США № 4447233, в котором описан насос для инфузии лекарственного средства для доставки лекарственного средства при точной скорости инфузии; патент США № 4447224, в котором описано имплантируемое устройство для инфузии с вариабельным током для непрерывной доставки лекарственного средства; патент США № 4439196, в котором описана система осмотической доставки лекарственного средства, имеющую многокамерные компартменты; и патент США № 4475196, в котором описана система осмотической доставки лекарственного средства. Приведенные патенты включены в данное описание посредством ссылки. Многие другие подобные имплантаты, системы доставки и модули известны специалистам в данной области.
В некоторых вариантах осуществления моноклональные антитела человека настоящего изобретения могут быть приготовлены для гарантии правильного распределения in vivo. Например, гематоэнцефалический барьер (ВВВ) исключает многие высокогидрофильные соединения. Для гарантии того, что терапевтические соединения настоящего изобретения пересекают ВВВ (если желательно), они могут быть приготовлены, например, в липосомах. В отношении способов приготовления липосом, см., например, патенты США №№ 4522811; 5374548 и 5399331. Липосомы могут содержать одну или несколько групп, которые селективно транспортируются в специфические клетки или органы и, следовательно, усиливают нацеленную доставку лекарственного средства (см., например, V.V. Ranade (1989) J. Clin. Pharmacol. 29:685). Примеры нацеливающих частей молекул включают фолат и биотин (см., например, патент США 5416016, Low et al.); маннозиды (Umezawa et al., (1988) Biochem. Biophys. Res. Commun. 153:1038); антитела (P.G. Bloeman et al. (1995) FEBS Lett. 357:140; M. Owais et al. (1995) Antimicrob. Agents Chemother. 39:180); рецептор белка А поверхностно-активного вещества (Briscoe et al. (1995) Am. J. Physiol. 1233:134); p120 (Schreier et al. (1994) J. Biol. Chem. 269:9090); см. также K. Keinanen; M.L. Laukkanen (1994) FEBS Lett. 346:123; J.J. Killion; I.J. Fidler (1994) Immunomethods 4:273.
Применения и способы настоящего изобретения
Антитела, композиции антител и способы настоящего изобретения имеют многочисленные применения in vitro и in vivo, например, детектирование PD-1 или усиление иммунной реакции блокадой PD-1. В предпочтительном варианте осуществления антитела настоящего изобретения являются антителами человека. Например, молекулы могут быть введены в клетки в культуре, in vitro и in vivo, или субъектам-людям, например, in vivo для усиления иммунитета в различных ситуациях. Таким образом, в одном аспекте настоящее изобретение обеспечивают способ модификации иммунного ответа у субъекта, предусматривающий введение указанному субъекту антитела или его антигенсвязывающей части настоящего изобретения таким образом, что иммунная реакция у субъекта модифицируется. Предпочтительно, указанная реакция усиливается, стимулируется или положительно регулируется.
Как использовано в данном описании, термин “субъект" включает человека и животных, не являющихся человеком. Термин животное (не человек) включает в себя всех позвоночных, например, млекопитающих и не-млекопитающих, таких как приматы (не человек), овцы, собаки, кошки, коровы, лошади, куры, земноводные и пресмыкающиеся, хотя млекопитающие являются предпочтительными, например, приматы (не человек), овцы, собаки, кошки, коровы и лошади. Предпочтительные субъекты включают пациентов-людей, нуждающихся в усилении иммунного ответа. Данные способы предпочтительно являются подходящими для лечения пациентов-людей, имеющих нарушение, которое может подвергаться лечению увеличением опосредованной Т-клетками иммунной реакции. В предпочтительном варианте осуществления данные способы предпочтительно пригодны для лечения клеток рака in vivo. Для достижения антигенспецифического усиления иммунитета анти-PD-1-антитела могут вводиться вместе с представляющим интерес антигеном. При введении антител к PD-1 вместе с другим агентом, указанные два агента могут вводиться в любом порядке или одновременно.
Настоящее изобретение обеспечивает дополнительно способы детектирования присутствия антигена PD-1 человека в пробе или измерения количества антигена PD-1 человека, предусматривающие контактирование пробы и контрольной пробы с моноклональным антителом человека или его антигенсвязывающей частью, которое специфически связывается с PD-1 человека, в условиях, которые делают возможным образование комплекса между антителом или его частью и PD-1 человека. Затем образование комплекса детектируют, причем различие образования комплекса данной пробой в сравнении с контрольной пробой является показателем присутствия антигена PD-1 человека в пробе.
Вследствие специфического связывания антител настоящего изобретения с PD-1, в сравнении с CD28, ICOS и CTLA-4, антитела настоящего изобретения могут быть использованы для специфического детектирования экспрессии PD-1 на поверхности клеток и, кроме того, могут быть использованы для очистки PD-1 при помощи иммуноаффинной очистки.
Рак
Блокада PD-1 антителами может усиливать иммунную реакцию пациента на раковые клетки. Лиганд для PD-1, PD-L1, не экспрессируется в здоровых клетках человека, но является изобилующим в различных типах рака человека (Dong et al. (2002) Nat Med 8:787-9). Взаимодействие между PD-1 и PD-L1 приводит к уменьшению инфильтрирующих опухоль лимфоцитов, уменьшению опосредованной рецептором Т-клеток пролиферации и ускользанию раковых клеток от иммунологического надзора (Dong et al. (2003) J Mol Med 81:281-7; Blank et al. (2005) Cancer Immunol. Immunother. 54:307-314; Konishi et al. (2004) Clin. Cancer Res. 10:5094-100). Иммунная супрессия может быть изменена ингибированием локального взаимодействия PD-1 с PD-L1, и это действие является аддитивным при блокировании также взаимодействия PD-1 с PD-L2 (Iwai et al. (2002) PNAS 99:12293-7; Brown et al. (2003) J. Immunol. 170:1257-66). Хотя предыдущие исследования показали, что пролиферация Т-клеток может быть восстановлена ингибированием взаимодействия PD-1 с PD-L1, не было сообщений о прямом действии на рост раковой опухоли in vivo блокированием взаимодействия PD-1/PD-L1. В одном аспекте настоящее изобретение относится к лечению субъекта in vivo с использованием анти-PD-1-антитела таким образом, что рост раковых тканей ингибируется. Анти-PD-1-антитело может быть использовано одно для ингибирования роста раковых опухолей. Альтернативно, анти-PD-1-антитело может быть использовано вместе с другими иммуногенными агентами, стандартными способами лечения рака или другими антителами, как описано ниже.
Таким образом, в одном варианте осуществления настоящее изобретение обеспечивает способ ингибирования роста опухолевых клеток у субъекта, предусматривающий введение указанному субъекту терапевтически эффективного количества анти-PD-1-антитела или его антигенсвязывающей части. Предпочтительно, антителом является анти-PD-1-антитело человека (например, любое из анти-PD-1-антител человека, описанных в данном описании). Дополнительно или альтернативно, антителом может быть химерное или гуманизированное анти-PD-1-антитело.
Предпочтительными типами рака, рост которых может быть ингибирован с использованием антител настоящего изобретения, включают типы рака, отвечающие на иммунотерапию. Неограничивающие примеры предпочтительных типов рака включают меланому (например, метастатическую злокачественную меланому), рак почки (например, гипернефрому), рак предстательной железы (например, не отвечающую на гормоны аденокарциному предстательной железы), рак молочной железы, рак ободочной кишки и рак легкого (например, немелкоклеточный рак легкого). Кроме того, настоящее изобретение включает трудноизлечимые или рецидивирующие злокачественные опухоли, рост которых может быть ингибирован антителами настоящего изобретения.
Примеры других типов рака, которые могут подвергаться лечению способами настоящего изобретения, включают костный рак, рак поджелудочной железы, рак кожи, рак головы и шеи, кожную или внутриглазную злокачественную меланому, рак матки, рак яичника, ректальный рак, рак анальной области, рак желудка, рак яичек, рак матки, рак фаллопиевых труб, рак эндометрия, рак шейки матки, рак вагины, рак вульвы, болезнь Ходжкина, не-Ходжкинскую лимфому, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркому мягкой ткани, рак мочеиспускательного канала, рак пениса, хронические или острые лейкозы, включая острый миелоидный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, солидные опухоли детского возраста, лимфоцитарную лимфому, рак мочевого пузыря, рак почки или мочеточника, рак почечных лоханок, неоплазму центральной нервной системы (ЦНС), лимфому первичной ЦНС, ангиогенез опухоли, опухоль спинного мозга, глиому ствола мозга, аденому гипофиза, саркому Капоши, эпидермоидный рак, плоскоклеточный рак, Т-клеточную лимфому, индуцированные условиями окружающей среды раковые заболевания, включая раковые заболевания, индуцированные асбестом, и комбинации указанных типов рака. Настоящее изобретение также полезно для лечения метастазирующих типов рака, которые экспрессируют PD-L2 (Iwai et al. (2005) Int. Immunol. 17:133-144).
Необязательно, антитела к PD-1 могут быть объединены с иммуногенным агентом, таким как раковые клетки, очищенные опухолевые антигены (включая рекомбинантные белки, пептиды и углеводные молекулы), клетки и клетки, трансфицированные генами, кодирующими иммуностимулирующие цитокины (He et al (2004) J. Immunol. 173:4919-28). Неограничивающие примеры опухолевых вакцин, которые могут быть использованы, включают пептиды антигенов меланомы, например, пептиды gp100, антигены MAGE, Trp-2, MART1 и/или тирозиназу, или опухолевые клетки, трансфицированные для экспрессии цитокина GM-CSF (обсуждаемые дополнительно ниже).
Было показано, что у людей некоторые опухоли являются иммуногенными, например, меланомы. Считается, что при увеличении порога активации Т-клеток посредством блокады PD-1 можно ожидать активации опухолевых реакций в хозяине.
Блокада PD-1 является, по-видимому, наиболее эффективной при комбинировании с протоколом вакцинации. Были разработаны многие экспериментальные стратегии вакцинации против опухолей (см. Rosenberg, S., 2000, Development of Cancer Vaccines, ASCO Educational Book Spring: 60-62; Logothetis, C., 2000, ASCO Educational Book Spring: 300-302; Khayat, D. 2000, ASCO Educational Book Spring: 414-428; Foon, K. 2000, ASCO Educational Book Spring: 730-738; см. также Restifo, N. and Sznol, M., Cancer Vaccines, Ch. 61, pp. 3023-3043 in DeVita, V. et al. (eds.), 1997, Cancer: Principles and Practice of Oncology. Fifth Edition). В одной из таких стратегий вакцину готовят с использованием аутологичных или аллогенных опухолевых клеток. Было показано, что клеточные вакцины являются наиболее эффективными при трансдукции опухолевых клеток для экспрессии GM-CSF. Было показано, что GM-CSF является сильным активатором презентации антигена для опухолевой вакцинации (Dranoff et al. (1993) Proc. Natl. Acad. Sci U.S.A. 90: 3539-43).
Исследование экспрессии генов и широкомасштабных распределений экспрессии в различных опухолях привело к определению так называемых опухолеспецифических антигенов (Rosenberg, SA (1999) Immunity 10: 281-7). Во многих случаях, опухолеспецифические антигены являются антигенами дифференцировки, экспрессируемыми в опухолях и в клетке, из которой возникла данная опухоль, например, антигенами меланоцитов gp100, антигенами MAGE и Trp-2. Более важно, было показано, что многие из указанных антигенов являются мишенями опухолеспецифических Т-клеток, обнаруженных в данном хозяине. Блокада PD-1 может быть использована вместе с набором рекомбинантных белков и/или пептидов, экспрессируемых в опухоли, для генерирования иммунной реакции на данные белки. Белки в норме рассматриваются иммунной системой как аутоантигены и, следовательно, являются толерантными к ней. Опухолевый антиген может также включать в себя белок теломеразу, который необходим для синтеза теломеров хромосом и который экспрессируется в более чем 85% типов рака человека, и только в ограниченном количестве соматических тканей (Kim, N et al. (1994) Science 266: 2011-2013). (Соматические ткани могут быть защищены от иммунной атаки различными способами). Опухолевым антигеном могут быть также «неоантигены», экспрессируемые в раковых клетках вследствие соматических мутаций, которые изменяют последовательность белка или создают слитые белки между двумя неродственными последовательностями (например, bcr-abl в хромосоме Philadelphia) или идиотип из В-клеточных опухолей.
Другие опухолевые вакцины могут включать в себя белки из вирусов, участвующие в раковых заболеваниях человека, таких как папилломавирусы человека (HPV), вирусы гепатита (HBV и HCV) и герпесвирус саркомы Капоши (KHSV). Другой формой опухолеспецифического антигена, который может быть использован вместе с блокадой PD-1, является белки теплового шока (HSP), выделенные из самой ткани опухоли. Белки теплового шока содержат фрагменты белков из опухолевых клеток, и такие HSP являются высокоэффективными в доставке антигенпрезентирующих клеток для индукции противоопухолевого иммунитета (Suot, R & Srivastava, P (1995) Science 269:1585-1588; Tamura, Y. et al. (1997) Science 278:117-120).
Дендритные клетки (DC) являются сильными антигенпрезентирующими клетками, которые могут быть использованы для прайминга антигенспецифических реакций. DC могут продуцироваться ex vivo и загружаться разными белковыми и пептидными антигенами, а также экстрактами опухолевых клеток (Nestle, F. et al. (1998) Nature Medicine 4: 328-332). DC могут также переноситься генетическими средствами для экспрессии таких опухолевых антигенов. DC были также слиты непосредственно с опухолевыми клетками для цели иммунизации (Kugler, A. et al. (2000) Nature Medicine 6:332-336). В качестве способа вакцинации, иммунизация DC может быть эффективно объединена с блокадой PD-1 для активации более сильных противоопухолевых реакций.
Блокада PD-1 может также комбинироваться со стандартными раковыми терапиями. В этих случаях, можно уменьшать дозу вводимого химиотерапевтического реагента (Mokyr, M. et al. (1998) Cancer Research 58: 5301-5304). Примером такой комбинации является анти-PD-1-антитело в комбинации с интерлейкином-2 (IL-2) для лечения меланомы. Научным обоснованием такого комбинированного применения блокады PD-1 и химиотерапии является то, что гибель клеток, которая является следствием цитотоксического действия большинства химиотерапевтических соединений, должна приводить к увеличенным уровням опухолевого антигена в пути презентации антигена. Другими комбинированными терапиями, которые могут приводить к синергизму с блокадой PD-1 посредством гибели клеток, являются облучение, хирургия и выключение эндокринной функции. Каждый из таких протоколов создает источник опухолевого антигена в хозяине. Ингибиторы ангиогенеза могут также комбинироваться с блокадой PD-1. Ингибирование ангиогенеза приводит к гибели опухолевых клеток, которые могут подавать опухолевый антиген в пути презентации антигенов хозяина.
Блокирующие PD-1 антитела могут быть также использованы в комбинации с биспецифическими антителами, которые нацеливают экспрессирующие Fc-альфа- или рецептор Fc-гамма эффекторные клетки на опухолевые клетки (см., например, патенты США №№ 5922845 и 5837243). Биспецифические антитела могут быть использованы для нацеливания на два отдельных антигена. Например, биспецифические антитела против рецептора Fc/против опухолевого антигена (например, Her-2/neu) использовали для нацеливания макрофагов на участки опухоли. Такое нацеливание может более эффективно активировать опухолеспецифические реакции. Т-клеточная ветвь данных реакций может увеличиваться с использованием блокады PD-1. Альтернативно, антиген может доставляться непосредственно к DC с использованием биспецифических антител, которые связываются с опухолевым антигеном и специфическим в отношении дендритных клеток маркером поверхности клеток.
Опухоли избегают иммунологического надзора хозяина большим разнообразием механизмов. Многие из таких механизмов могут быть преодолены инактивацией белков, которые экспрессируются опухолями и которые являются иммуносупрессивными. Они включают, среди прочего, TGF-бета (Kehrl, J. et al. (1986) J. Exp. Med. 163: 1037-1050), IL-10 (Howard, M. & O'Garra, A. (1992) Immunology Today 13: 198-200) и Fas-лиганд (Hahne, M. et al. (1996) Science 274: 1363-1365). Антитела к каждому из указанных агентов могут быть использованы в комбинации с анти-PD-1-антителом для противодействия эффектам иммуносупрессивного агента и содействия иммунным реакциям хозяина против опухоли.
Другие антитела, которые могут быть использованы для активации иммунологической откликаемости хозяина, могут быть использованы в комбинации с анти-PD-1-антителом. Они включают в себя молекулы на поверхности дендритных клеток, которые активируют функцию DC и презентацию антигена. Анти-CD40-антитела способны эффективно заменять активность Т-клеток-хелперов (Ridge, J. et al. (1998) Nature 393: 474-478) и могут быть использованы вместе с анти-PD-1-антителами (Ito, N. et al. (2000) Immunobiology 201 (5) 527-40). Активация антител к костимулирующим Т-клетки молекулам, таким как CTLA-4 (например, патент США № 5811097), OX-40 (Weinberg, A. et al. (2000) Immunol 164: 2160-2169), 4-1BB (Melero, I. et al. (1997) Nature Medicine 3: 682-685 (1997), and ICOS (Hutloff, A. et al. (1999) Nature 397: 262-266) может также обеспечивать увеличенные уровни активации Т-клеток.
Пересадку (трансплантацию) костного мозга используют в настоящее время для лечения различных опухолей гемопоэтического происхождения. Хотя реакция трансплантат против хозяина является следствием такой терапии, можно получить терапевтическую пользу из реакций трансплантат против хозяина. Блокада PD-1 может быть использована для эффективности пересаженных опухолеспецифических Т-клеток донора.
Имеются также несколько экспериментальных протоколов терапии, которые включают активацию ex vivo и размножение антигенспецифических Т-клеток и адоптивный (со стойким приживлением) перенос указанных клеток реципиентам для получения антигенспецифических Т-клеток против опухоли (Greenberg, R. & Riddell, S. (1999) Science 285: 546-51). Такие способы могут быть также использованы для активации Т-клеточных реакций на инфекционные агенты, такие как CMV. Можно ожидать, что активация ex vivo в присутствии анти-PD-1-антител будет увеличивать встречаемость и активность адоптивно перенесенных Т-клеток.
Инфекционные заболевания
Другие способы настоящего изобретения используют для лечения пациентов, которые были подвергнуты действию конкретных токсинов или патогенов. Таким образом, другой аспект настоящего изобретения обеспечивает способ лечения инфекционного заболевания у субъекта, предусматривающий введение указанному субъекту анти-PD-1-антитела или его антигенсвязывающей части таким образом, что субъект подвергается лечению в отношении указанного инфекционного заболевания. Предпочтительно, антителом является анти-PD-1-антитело человека (например, любое из описанных в данном описании анти-PD-1-антител человека). Дополнительно или альтернативно, данное антитело может быть химерным или гуманизированным антителом.
Подобно ее использованию в отношении опухолей, как обсуждалось выше, опосредованная антителом блокада PD-1 может быть использована самостоятельно или в качестве дополнения в комбинации с вакцинами, для стимуляции иммунной реакции на патогенны, токсины и аутоантигены. Примеры патогенов, для которых может использоваться, в частности, такой терапевтический подход, включают патогены, для которых в настоящее время нет эффективной вакцины, или патогены, для которых общепринятые вакцины являются менее чем полностью эффективными. Они включают, но не ограничиваются ими, ВИЧ, вирусы гепатита (A, B и C), вирусы гриппа, герпесвирусы, возбудителей гиардиоза, возбудителей малярии, лейшманиоза, Staphylococcus aureus, Pseudomonas aeruginosa. Блокада PD-1 особенно полезна против установленных инфекций. Новые эпитопы распознаются как чужеродные в момент введения анти-PD-1-антител, провоцируя таким образом сильную Т-клеточную реакцию, которая не заглушается негативными сигналами через PD-1.
Некоторые примеры патогенных вирусов, вызывающих инфекции, которые могут подвергаться лечению способами настоящего изобретения, включают ВИЧ, вирус гепатита (А, В или С), герпесвирус (например, VZV, HSV-1, HAV-6, HSV-II и CMV, вирус Эпштейна-Барра), аденовирус, вирус гриппа, флавивирусы, эховирус, риновирус, вирус Коксаки, коронавирус, респираторно-синцитиальный вирус, вирус эпидемического паротита, ротавирус, вирус кори, вирус коревой краснухи, парвовирус, вирус коровьей оспы, HTLV-вирус, вирус Денге, папилломавирус, вирус контагиозного моллюска, полиовирус, вирус бешенства, JC-вирус (вирус полиомы человека, прогрессирующей многоочаговой лейкоэнцефалопатии) и вирус арбовирусного энцефалита.
Некоторые примеры патогенных бактерий, вызывающих инфекции, которые могут подвергаться лечению способами настоящего изобретения включают хламидии, бактерии рода Rickettsia, микобактерии, стафилококки, стрептококки, пневмококки, менингококки и гонококки, Klebsiella, Proteus, Serratia, Pseudomonas, Legionella, дифтерийные бактерии, Salmonella, бациллы, возбудитель холеры, возбудитель столбняка, возбудитель ботулизма, возбудитель сибирской язвы, возбудитель чумы, возбудитель лептоспироза и бактерии, вызывающие болезнь Лайма.
Некоторые примеры патогенных грибов, вызывающих инфекции, которые могут подвергаться лечению способами настоящего изобретения, включают Candida (albicans, krusei, glabrata, tropicalis и т.д.), Cryptococcus neoformans, Aspergillus (fumigatus, niger и т.д.), род Mucorales (mucor, absidia, rhizophus), Sporothrix schenkii, Blastomyces dermatitidis, Paracoccidioides brasiliensis, Coccidioides immitis и Histoplasma capsulatum.
Некоторые примеры патогенных паразитов, вызывающих инфекции, которые могут подвергаться лечению способами настоящего изобретения, включают Entamoeba histolytica, Balantidium coli, Naegleriafowleri, Acanthamoeba sp., Giardia lambia, Cryptosporidium sp., Pneumocystis carinii, Plasmodium vivax, Babesia microti, Trypanosoma brucei, Trypanosoma cruzi, Leishmania donovani, Toxoplasma gondi и Nippostrongylus brasiliensis.
Во всех вышеуказанных способах блокада PD-1 может комбинироваться с другими формами иммунотерапии, такими как лечение цитокинами (например, интерферонами, GM-CSF, G-CSF, IL-2), или терапия с использованием биспецифических антител, которая обеспечивает усиленную презентацию опухолевых антигенов (см., например, Holliger (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak (1994) Structure 2:1121-1123).
Аутоиммунные реакции
Анти-PD-1-антитела могут провоцировать и размножать аутоиммунные реакции. Действительно, индукции противоопухолевых реакций с использованием опухолевых клеток и пептидных вакцин выявила, что многие противоопухолевые реакции включают в себя реактивности против собственных антигенов (депигментацию, наблюдаемую в модифицированной анти-CTLA-4+GM-CSF меланоме В16 в Elsas et al. supra; депигментацию в вакцинированных Trp-2 мышах (Overwijk, W. et al. (1999) Proc. Natl. Acad. Sci. U.S.A. 96: 2982-2987); аутоиммунный простатит, индуцированный противоопухолевыми вакцинами TRAMP (Hurwitz, A. (2000) supra), вакцинацией пептидным антигеном меланомы и витилиго, наблюдаемые в клинических исследованиях на человеке (Rosenberg, SA and White, DE (1996) J. Immunother Emphasis Tumor Immunol 19 (1): 81-4).
Таким образом, можно рассматривать применение блокады против PD-1 вместе с различными аутологичными белками для создания протоколов вакцинации для эффективного генерирования иммунных реакций против аутологичных белков для лечения заболевания. Например, болезнь Альцгеймера включает в себя нежелательное накапливание Аβ-пептида в амилоидных отложениях в головном мозге; реакции антител против амилоида способны очищать амилоидные отложения (Schenk et al., (1999) Nature 400: 173-177).
Другие аутологичные белки могут также использоваться в качестве мишеней, например, IgE, для лечения аллергии и астмы, и TNFα для ревматоидного артрита. Наконец, реакции антител на различные гормоны могут быть индуцированы с использованием анти-PD-1-антитела. Реакции нейтрализующих антител на репродуктивные гормоны могут быть использованы для контрацепции. Может быть также использована реакция нейтрализующих антител на гормоны и другие растворимые факторы, которые необходимы для роста конкретных опухолей и могут также рассматриваться как возможные мишени вакцинации.
Способы, аналогичные способам, описанным выше для использования анти-PD-1-антитела, могут быть использованы для индукции терапевтических аутоиммунных реакций для лечения пациентов, имеющих нежелательное накапливание других аутоантигенов, таких как амилоидные отложения, включая Аβ в болезни Альцгеймера, цитокинов, таких как TNFα, и IgE.
Вакцины
Анти-PD-1-антитела могут быть использованы для стимуляции антигенспецифических иммунных реакций совместным введением анти-PD-1-антитела с представляющим интерес антигеном (например, вакциной). Таким образом, в другом аспекте настоящее изобретение обеспечивает способ усиления иммунной реакции на антиген у субъекта, предусматривающий введение указанному субъекту: (i) антигена; и (ii) анти-PD-1-антитела или его антигенсвязывающей части таким образом, что иммунная реакция на данный антиген у субъекта усиливается. Предпочтительно, антителом является анти-PD-1-антитело человека (например, любое из описанных в данном описании анти-PD-1-антител человека). Дополнительно или альтернативно, таким антителом может быть химерное антитело или гуманизированное антитело. Антигеном может быть, например, опухолевый антиген, вирусный антиген, бактериальный антиген или антиген из патогена. Неограничивающие примеры таких антигенов включают антигены, обсуждаемые в разделах выше, такие как опухолевые антигены (или противоопухолевые вакцины), обсуждаемые выше, или антигены из вирусов, бактерий или других патогенов, описанные в данном описании.
Подходящие пути введения композиций антител (например, моноклональных антител человека, полиспецифических и биспецифических молекул или иммуноконъюгатов) настоящего изобретения in vivo и in vitro хорошо известны в данной области и могут быть выбраны специалистами в данной области. Например, композиции антител могут вводиться инъекцией (например, внутривенной или подкожной). Подходящие дозы используемых молекул будут зависеть от возраста и массы субъекта и концентрации и/или формы композиции антител.
Как описано выше, анти-PD-1-антитела человека настоящего изобретения могут вводиться совместно с одним или несколькими другими терапевтическими агентами, например, цитотоксическим агентом, радиотоксическим агентом или иммуносупрессивным агентом. Антитело может быть связано с данным агентом (в виде иммунокомплекса) или может быть введено отдельно от данного агента. В последнем случае (в случае отдельного введения) антитело может вводиться до, после или совместно с указанным агентом или может вводиться совместно с другими известными терапиями, например, противораковой терапией, например, облучением. Такие терапевтические агенты включают, среди прочего, противоопухолевые агенты, такие как доксорубицин (адриамицин), цисплатин, блеомицина сульфат, кармустин, хлорамбуцил, декарбазин и циклофосфамидгидроксимочевина, которые, сами по себе, являются эффективными только при уровнях, которые являются токсичными или субтоксичными для пациента. Цисплатин вводят внутривенно в виде 100 мг на дозу один раз каждые четыре недели, и адриамицин вводят внутривенно в виде дозы 60-75 мг/мл один раз каждые 21 день. Совместное введение анти-PD-1-антител человека или их антигенсвязывающих частей настоящего изобретения с химиотерапевтическими агентами обеспечивает два противораковых агента, которые действуют посредством различных механизмов, которые оказывают цитотоксическое действие в отношении опухолевых клеток человека. Такое совместное введение может решить проблемы, связанные с развитием устойчивости к лекарственным средствам или изменением антигенности опухолевых клеток, которые будут делать их нереактивными с данным антителом.
В объем настоящего изобретения входят также наборы, содержащие композиции антител настоящего изобретения (например, антител человека, биспецифических или полиспецифических молекул или иммуноконъюгатов) и инструкции для применения. Кроме того, набор содержит по меньшей мере один дополнительный реагент или один или несколько дополнительных антител человека настоящего изобретения (например, антитело человека, имеющее дополняющую активность, которое связывается с эпитопом в антигене PD-1, отличающимся от эпитопа первого антитела человека). Наборы обычно содержат этикетку, указывающую предполагаемое применение содержимого данного набора. Термин «этикетка» включает любое описание или записанный материал, поставляемый с набором, или материал, который иным образом прилагается к набору.
Комбинированная терапия
Настоящее изобретение основано, частично, на следующих экспериментальных данных. Модели опухоли мыши (рака ободочной кишки МС38 и фибросаркомы SA1/N) использовали для испытания действия in vivo лечения опухоли комбинированием иммуностимулирующих терапевтических антител - анти-CTLA-4 и анти-PD-1-антител. Иммунотерапевтическую комбинацию предоставляли либо одновременно с имплантатом опухолевых клеток (примеры 14 и 17), либо после того, как опухолевые клетки были имплантированы, в течение времени, достаточного для того, чтобы они стали развившейся (установленной) опухолью (примеры 15, 16 и 18). Независимо от хронометрирования лечения антителами, было обнаружено, что лечение одним анти-CTLA-4-антителом и одним анти-PD-1-антителом (химерным антителом, в котором крысиное антитело против мышиного PD-1 было модифицировано Fc-областью мышиного иммуноглобулина, см. пример 1) имело умеренное действие на уменьшение роста опухоли на модели опухоли МС38 (см., например, фиг.21, 24 и 27). Одно анти-CTLA-4-антитело было вполне эффективным на модели опухоли SA1/N (см. фиг.30D), которая требовала более низкой дозы анти-CTLA-4-антитела для комбинированных исследований на данной модели. Тем не менее, комбинированное лечение анти-CTLA-4-антителом и анти-PD-1-антителом показала неожиданное, значительно более высокое действие на уменьшение роста опухолей в сравнении с лечением каждым антителом по отдельности (см., например, фиг.21D, 24D, 30F и 33H-J). Кроме того, результаты примеров 14, 16 и 18 показывают, что комбинированное лечение анти-CTLA-4-антителом и анти-PD-1-антителом имело значительное синергическое действие на рост опухолей даже при субоптимальных терапевтических дозах в сравнении с лечением любым из антител по отдельности (т.е. комбинированная терапия была неожиданно более эффективной при субтерапевтических дозах, чем любая монотерапия). Не желая быть связанными теорией, авторы считают возможным, что увеличением порога активации Т-клеток блокадой PD-1 и CTLA-4 можно активировать противоопухолевые реакции у хозяина.
В одном варианте осуществления настоящее изобретение обеспечивает способ лечения гиперпролиферативного заболевания, предусматривающий введение анти-PD-1-антитела и анти-CTLA-4-антитела субъекту. В дополнительных вариантах осуществления анти-PD-1-антитело вводят в субтерапевтической дозе, анти-CTLA-4-антитело вводят в субтерапевтической дозе или оба вводят в субтерапевтической дозе. В другом варианте осуществления настоящее изобретение обеспечивает способ изменения побочного эффекта, связанного с лечением гиперпролиферативного заболевания иммуностимулирующим агентом, предусматривающий введение анти-PD-1-антитела и субтерапевтической дозы анти-CTLA-4-антитела субъекту. В некоторых вариантах осуществления субъектом является человек. В некоторых вариантах осуществления анти-CTLA-4-антителом является моноклональное антитело последовательности человека 10D1, и анти-PD-1-антителом является моноклональное антитело последовательности человека, такое как 17D8, 2D3, 4H1, 5C4 и 4А11. Моноклональные антитела с последовательностью человека 17D8, 2D3, 4H1, 5C4 и 4А11 были выделены и структурно охарактеризованы, как описано в предварительном патенте США № 60/679466.
Моноклональные антитела анти-CTLA-4-антитело и анти-PD-1-антитело (mAb) и антитела с последовательностью человека настоящего изобретения могут быть получены различными способами, включая общепринятую методику получения моноклональных антител, например, вирусное или онкогенное превращение В-лимфоцитов. Одной системой животного для получения гибридом является мышиная система. Продуцирование гибридом у мыши является хорошо установленной методикой. Протоколы и способы выделения иммунизированных спленоцитов для слияния известны в данной области. Партнеры слияния (например, мышиные миеломные клетки) и методы слияния также известны (см., например, Harlow and Lane (1988) “Antibodies: A Laboratory Manual,” Cold Spring Harbor Laboratory Press, Cold Spring Harbor New York).
Анти-CTLA-4-антитела настоящего изобретения могут связываться с эпитопом на CTLA-4 человека таким образом, что они ингибируют взаимодействие CTLA-4 с противорецептором В7 человека. Поскольку взаимодействие CTLA-4 человека с В7 человека трансдуцирует сигнал, приводящий к инактивации Т-клеток, несущих рецептор CTLA-4 человека, антагонизм такого взаимодействия эффективно индуцирует, увеличивает или пролонгирует активацию Т-клеток, несущих рецептор CTLA-4 человека, пролонгируя или увеличивая тем самым иммунную реакцию. Анти-CTLA-4-антитела описаны в патентах США №№ 5811097; 5855887; 6051227; в публикациях заявок PCT №№ WO 01/14424 и WO 00/37504; и в публикации патента США № 2002/0039581. Каждая из приведенных ссылок специально включена в данное описание посредством ссылки, с целью описания анти-CTLA-4-антител. Примером клинического анти-CTLA-4-антитела является моноклональное антитело человека 10D1, описанное в WO 01/14424 и Заявке на патент США № 09/644668. Антитело 10D1 вводили в единственной дозе и во множественных дозах, самостоятельно или в комбинации с вакциной, химиотерапией и интерлейкином-2 более чем 500 пациентам с диагнозом метастатической меланомы, рака предстательной железы, лимфомы, почечноклеточного рака, рака молочной железы, рака яичника и ВИЧ. Другие анти-CTLA-4-антитела, включенные способами настоящего изобретения, включают, например, антитела, описанные в WO 98/42752; WO 00/37504; патенте США № 6207156; Hurwitz et al. (1998) Proc. Natl. Acad. Sci. USA 95(17):10067-10071; Camacho et al. (2004) J. Clin. Oncology 22(145): Abstract No. 2505 (antibody CP-675206) и Mokyr et al. (1998) Cancer Res. 58:5301-5304. В некоторых вариантах осуществления способы настоящего изобретения включают в себя применение анти-CTLA-4-антитела, которое является антителом с последовательностью человека, предпочтительно моноклональным антителом, и в другом варианте осуществления оно является моноклональным антителом 10D1.
В некоторых вариантах осуществления анти-CTLA-4-антитело связывается с CTLA-4 человека с KD 5×10-8M или менее, связывается с CTLA-4 человека с KD 1×10-8M или менее, связывается с CTLA-4 человека с KD 5×10-9M или менее или связывается с CTLA-4 человека с KD 1×10-8M-1×10-10M или менее.
Комбинация антител полезна для усиления иммунной реакции против гиперпролиферативного заболевания блокадой PD-1 и CTLA-4. В предпочтительном варианте осуществления антитела настоящего изобретения являются антителами человека. Например, молекулы могут вводиться в клетки в культуре, in vitro или ex vivo, или субъектам-людям, например, in vivo, для усиления иммунитета в различных ситуациях. Таким образом, в одном аспекте настоящее изобретение обеспечивает способ модификации иммунной реакции у субъекта, предусматривающий введение указанному субъекту комбинации антител или комбинации их антигенсвязывающих частей, настоящего изобретения таким образом, что иммунная реакция у данного субъекта модифицируется. Предпочтительно, данная реакция усиливается, стимулируется или положительно регулируется. В другом варианте осуществления данное открытие обеспечивает способ изменения неблагоприятных (побочных) событий, связанных с лечением гиперпролиферативного заболевания иммуностимулирующим терапевтическим агентом, предусматривающий введение анти-PD-1-антитела и субтерапевтической дозы анти-CTLA-4-антитела субъекту.
Блокада PD-1 и CTLA-4 антителами может усиливать иммунную реакцию на раковые клетки у пациента. Типы рака, рост которых может быть ингибирован антителами настоящего изобретения, включают типы рака, обычно отвечающие на иммунотерапию. Репрезентативные примеры рака для лечения комбинированной терапией настоящего изобретения включают меланому (например, метастатическую злокачественную меланому), рак почки, рак предстательной железы, рак молочной железы, рак ободочной кишки и рак легкого. Примеры других типов рака, которые могут подвергаться лечению с использованием способов настоящего изобретения, включают костный рак, рак поджелудочной железы, рак кожи, рак головы и шеи, кожную или внутриглазную злокачественную меланому, рак матки, рак яичника, ректальный рак, рак анальной области, рак желудка, рак яичек, рак матки, рак фаллопиевых труб, рак эндометрия, рак шейки матки, рак вагины, рак вульвы, болезнь Ходжкина, не-Ходжкинскую лимфому, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечника, саркому мягкой ткани, рак мочеиспускательного канала, рак пениса, хронические или острые лейкозы, включая острый миелоидный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, солидные опухоли детского возраста, лимфоцитарную лимфому, рак мочевого пузыря, рак почки или мочеточника, рак почечных лоханок, неоплазму центральной нервной системы (ЦНС), лимфому первичной ЦНС, ангиогенез опухоли, опухоль спинного мозга, глиому ствола мозга, аденому гипофиза, саркому Капоши, эпидермоидный рак, плоскоклеточный рак, Т-клеточную лимфому, индуцированные условиями окружающей среды раковые заболевания, включая раковые заболевания, индуцированные асбестом, и комбинации указанных типов рака. Настоящее изобретение полезно также для лечения метастазирующих типов рака.
В некоторых вариантах осуществления комбинация терапевтических антител, обсуждаемая в данном описании, может вводиться одновременно в виде единой композиции в фармацевтически приемлемом носителе или одновременно в виде отдельных композиций, в которых каждое антитело находится в фармацевтическом носителе. В другом варианте осуществления комбинация терапевтических антител может вводиться последовательно. Например, анти-CTLA-4-антитело и анти-PD-1-антитело могут вводиться последовательно таким образом, что анти-CTLA-4-антитело вводят первым, и анти-PD-1-антитело вводят вторым, или анти-PD-1-антитело вводят первым, а анти-CTLA-4-антител вводят вторым. Кроме того, если вводят последовательно более чем одну дозу комбинированной терапии, порядок последовательного введения может быть обратным или может сохраняться один и тот же порядок введения в каждой временной точке введения, последовательные введения могут комбинироваться с совместными введениями или любой их комбинацией. Например, первое введение комбинации анти-CTLA-4-антитела и анти-PD-1-антитела может быть совместным, второе введение может быть последовательным с анти-CTLA-4-антителом первым и анти-PD-1-антителом вторым, и третье введение может быть последовательным с анти-PD-1-антителом первым и анти-CTLA-4-антител вторым, и т.д. Другая репрезентативная схема введения доз может включать первое введение, которое является последовательным с анти-PD-1-антителом первым и анти-CTLA-4-антителом вторым, и следующие введения могут быть одновременными.
Необязательно, комбинация анти-PD-1-антител и анти-CTLA-4-антител может дополнительно комбинироваться с иммуногенным агентом, таким как раковые клетки, очищенные опухолевые антигены (включая рекомбинантные белки, пептиды и молекулы углеводов), клетки и клетки, трансфицированные генами, кодирующими иммуностимулирующие цитокины (He et al. (2004) J. Immunol. 173:4919-28). Неограничивающие примеры опухолевых вакцин, которые могут быть использованы, включают в себя пептиды антигенов меланомы, например, пептиды gp100, антигены MAGE, Trp-2, MART1 и/или тирозиназу, или опухолевые клетки, трансфицированные для экспрессии цитокина GM-CSF (обсуждаемые дополнительно ниже).
Комбинированная блокада PD-1 и CTLA-4 может быть дополнительно комбинирована с протоколом вакцинации. Были разработаны многие экспериментальные стратегии для вакцинации против опухолей (см. Rosenberg, S. (2000) Development of Cancer Vaccines, ASCO Educational Book Spring: 60-62; Logothetis, C., 2000, ASCO Educational Book Spring: 300-302; Khayat, D. (2000) ASCO Educational Book Spring: 414-428; Foon, K. (2000) ASCO Educational Book Spring: 730-738; см. также Restifo and Sznol, Cancer Vaccines, Ch. 61, pp. 3023-3043 in DeVita et al. (eds.), 1997, Cancer: Principles and Practice of Oncology. Fifth Edition). В одной из таких стратегий вакцину готовят с использованием аутологичных или аллогенных опухолевых клеток. Было показано, что клеточные вакцины являются наиболее эффективными при трансдукции опухолевых клеток для экспрессии GM-CSF. Было показано, что GM-CSF является сильным активатором презентации антигена для опухолевой вакцинации (Dranoff et al. (1993) Proc. Natl. Acad. Sci U.S.A. 90: 3539-43).
Исследование экспрессии генов и широкомасштабных распределений экспрессии в различных опухолях привело к определению так называемых опухолеспецифических антигенов (Rosenberg (1999) Immunity 10: 281-7). Во многих случаях, опухолеспецифические антигены являются антигенами дифференцировки, экспрессируемыми в опухолях и в клетке, из которой возникла данная опухоль, например, антигенами меланоцитов gp100, антигенами MAGE и Trp-2. Более важно, было показано, что многие из указанных антигенов являются мишенями опухолеспецифических Т-клеток, обнаруженных у данного хозяина. В некоторых вариантах осуществления комбинированная блокада PD-1 и CTLA-4, использующая описанные выше композиции антител, может быть использована вместе с набором рекомбинантных белков и/или пептидов, экспрессируемых в опухоли, для генерирования иммунной реакции на указанные белки. Белки в норме рассматриваются иммунной системой как аутоантигены и, следовательно, они являются толерантными к ней. Опухолевый антиген может также включать в себя белок теломеразу, который необходим для синтеза теломеров хромосом и который экспрессируется в более чем 85% типов рака человека и только в ограниченном количестве соматических тканей (Kim, N et al. (1994) Science 266: 2011-2013). (Соматические ткани могут быть защищены от иммунной атаки различными способами). Опухолевым антигеном могут быть также «неоантигены», экспрессируемые в раковых клетках вследствие соматических мутаций, которые изменяют последовательность белка или создают слитые белки между двумя неродственными последовательностями (например, bcr-abl в хромосоме Philadelphia) или идиотип из В-клеточных опухолей.
Другие опухолевые вакцины могут включать белки из вирусов, участвующие в раковых заболеваниях человека, таких как папилломавирусы человека (HPV), вирусы гепатита (HBV и HCV) и вирус саркомы Капоши (KHSV). Другой формой опухолеспецифического антигена, который может быть использован вместе с блокадой PD-1, является белки теплового шока (HSP), выделенные из самой ткани опухоли. Белки теплового шока содержат фрагменты белков из опухолевых клеток, и такие HSP являются высокоэффективными в доставке антигенпрезентирующих клеток для индукции противоопухолевого иммунитета (Suot, R & Srivastava, P (1995) Science 269:1585-1588; Tamura, Y. et al. (1997) Science 278:117-120).
Дендритные клетки (DC) являются сильными антигенпрезентирующими клетками, которые могут быть использованы для прайминга антигенспецифических реакций. DC могут продуцироваться ex vivo и загружаться разными белковыми и пептидными антигенами, а также экстрактами опухолевых клеток (Nestle et al. (1998) Nature Medicine 4: 328-332). DC могут также переноситься генетическими средствами для экспрессии опухолевых антигенов. DC были также слиты непосредственно с опухолевыми клетками для целей иммунизации (Kugler et al. (2000) Nature Medicine 6:332-336). В качестве способа вакцинации, иммунизация DC может быть дополнительно эффективно объединена с блокадой PD-1 и CTLA-4 для активации более сильных противоопухолевых реакций.
Комбинированная блокада PD-1 и CTLA-4 может также дополнительно комбинироваться со стандартными раковыми терапиями. Например, комбинированная блокада PD-1 и CTLA-4 может эффективно комбинироваться с химиотерапевтическими схемами лечения. В этих случаях, как наблюдали с комбинацией анти-PD-1-антител и анти-CTLA-4-антител, можно уменьшать дозу вводимого химиотерапевтического реагента (Mokyr et al. (1998) Cancer Research 58: 5301-5304). Примером такой комбинации является комбинация анти-PD-1-антител и анти-CTLA-4-антител, дополнительно в комбинации с интерлейкином-2 (IL-2) для лечения меланомы. Научным обоснованием так комбинированного применения блокады PD-1 и CTLA-4 с химиотерапией является то, что гибель клеток, которая является следствием цитотоксического действия большинства химиотерапевтических соединений, должна приводить к увеличенным уровням опухолевого антигена в пути презентации антигена. Другими комбинированными терапиями, которые могут приводить к синергизму с комбинированной блокадой PD-1 и CTLA-4 посредством смерти клеток, являются облучение, хирургия и выключение эндокринной функции. Каждый из указанных протоколов создает источник опухолевого антигена у хозяина. Ингибиторы ангиогенеза могут также комбинироваться с комбинированной блокадой PD-1 и CTLA-4. Ингибирование ангиогенеза приводит к гибели опухолевых клеток, которые могут подавать опухолевый антиген в пути презентации антигенов хозяина.
Комбинация блокирующих PD-1 и CTLA-4 антител может быть также использована в комбинации с биспецифическими антителами, которые нацеливают экспрессирующие рецептор Fcα или Fcγ эффекторные клетки на опухолевые клетки (см., например, патенты США №№ 5922845 и 5837243). Биспецифические антитела могут быть использованы для нацеливания на два отдельных антигена. Например, биспецифические антитела против рецептора Fc/против опухолевого антигена (например, Her-2/neu) использовали для нацеливания макрофагов на участки опухоли. Такое нацеливание может более эффективно активировать опухолеспецифические реакции. Т-клеточная ветвь указанных реакций может увеличиваться с использованием комбинированной блокады PD-1 и CTLA-4. Альтернативно, антиген может доставляться непосредственно к DC с использованием биспецифических антител, которые связываются с опухолевым антигеном и специфическим в отношении дендритных клеток маркером поверхности клеток.
В другом примере комбинация анти-PD-1-антител и анти-CTLA-4-антител может быть использована вместе с противоопухолевыми антителами, такими как Rituxan® (ритуксимаб), Herceptin® (трастуцумаб), Bexxar® (тозитумомаб), Zevalin® (тбритумомаб), Campath® (алемтуцумаб), Lymphocide® (эпртуцумаб), Avastin® (бевацицумаб) и Tarceva® (эрлотиниб), и т.п. В качестве примера и без связи с теорией, лечение противораковым антителом и противораковым антителом, конъюгированным с токсином, может приводить к гибели раковых клеток (например, опухолевых клеток), которые могут потенцировать иммунную реакцию, опосредованную CTLA-4 или PD-1. В примере варианта осуществления лечение гиперпролиферативного заболевания (например, раковой опухоли) может включать противораковое антитело в комбинации с анти-PD-1 и анти-CTLA-4-антителами, одновременно или последовательно, или любой их комбинации, которые могут потенцировать противоопухолевые иммунные реакции хозяина.
Опухоли избегают иммунологического надзора хозяина большим разнообразием механизмов. Многие из таких механизмов могут быть преодолены инактивацией белков, которые экспрессируются опухолями и которые являются иммуносупрессивными. Они включают, среди прочего, TGF-бета (Kehrl, J. et al. (1986) J. Exp. Med. 163: 1037-1050), IL-10 (Howard, M. & O'Garra, A. (1992) Immunology Today 13: 198-200) и Fas-лиганд (Hahne, M. et al. (1996) Science 274: 1363-1365). В другом примере, антитела к каждому из указанных агентов могут быть комбинированы с комбинацией анти-PD-1-антитела и анти-CTLA-4-антитела для противодействия эффектам иммуносупрессивных агентов и содействия иммунным реакциям хозяина против опухоли.
Другие антитела, которые могут быть использованы для активации иммунологической откликаемости хозяина, могут быть дополнительно использованы в комбинации с комбинацией анти-PD-1-антитела и анти-CTLA-4-антитела. Они включают в себя молекулы на поверхности дендритных клеток, которые активируют функцию DC и презентацию антигена. Анти-CD40-антитела способны эффективно заменять активность Т-клеток-хелперов (Ridge, J. et al. (1998) Nature 393: 474-478) и могут быть использованы вместе с комбинацией анти-PD-1-антитела и анти-CTLA-4-антитела (Ito, N. et al. (2000) Immunobiology 201 (5) 527-40). Активация антител к костимулирующим Т-клетки молекулам, таким как OX-40 (Weinberg, A. et al. (2000) Immunol 164: 2160-2169), 4-1BB (Melero, I. et al. (1997) Nature Medicine 3: 682-685 (1997) и ICOS (Hutloff, A. et al. (1999) Nature 397: 262-266), может также обеспечивать увеличенные уровни активации Т-клеток.
Пересадку (трансплантацию) костного мозга используют в настоящее время для лечения различных опухолей гемопоэтического происхождения. Хотя реакция трансплантат против хозяина является следствием такой терапии, можно получить терапевтическую пользу из реакций трансплантат против хозяина. Комбинированная блокада PD-1 и CTLA-4 может быть использована для эффективности пересаженных опухолеспецифических Т-клеток донора.
Имеются также несколько экспериментальных протоколов терапии, которые включают активацию ex vivo и размножение антигенспецифических Т-клеток и адоптивный (со стойким приживлением) перенос указанных клеток реципиентам для получения антигенспецифических Т-клеток против опухоли (Greenberg, R. & Riddell, S. (1999) Science 285: 546-51). Такие способы могут быть также использованы для активации Т-клеточных реакций на инфекционные агенты, такие как CMV. Можно ожидать, что активация ex vivo в присутствии анти-PD-1-антител будет увеличивать встречаемость и активность адоптивно перенесенных Т-клеток.
Как описано в данном описании, органы могут проявлять иммуноассоциированные побочные эффекты после терапии иммуностимулирующими терапевтическими антителами, такие как эффекты, возникающие в желудочно-кишечном тракте (диарея и колит) и на коже (сыпь и зуд), после лечения анти-CTLA-4-антителом. Например, не связанные с ободочной кишкой иммуноассоциированные желудочно-кишечные неблагоприятные эффекты наблюдали также в пищеводе (эзофагит), двенадцатиперстной кишке (дуоденит) и подвздошной кишке (илеит) после лечения анти-CTLA-4-антителом.
В некоторых вариантах осуществления настоящее изобретение обеспечивает способ изменения побочных эффектов, связанных с лечением гиперпролиферативного заболевания иммуностимулирующим агентом, предусматривающий введение анти-PD-1-антитела и субтерапевтической дозы анти-CTLA-4-антител субъекту. Например, способы настоящего изобретения обеспечивают способы уменьшения встречаемости индуцированных иммуностимулирующим антителом колита или диареи введением неабсорбируемого стероида пациенту. Поскольку любой пациент, который будет получать иммуностимулирующее терапевтическое антитело, имеет риск развития колита или диареи, индуцированных таким антителом, вся популяция пациентов подходит для терапии в соответствии со способами настоящего изобретения. Хотя стероиды вводили для лечения воспалительного заболевания пищеварительного тракта (IBD) и предотвращения обострений IBD, их не использовали для предотвращения (уменьшения встречаемости) IBD у пациентов, которые не имели диагноза IBD. Существенные побочные действия, связанные со стероидами, даже неабсорбируемыми стероидами, расхолаживали врачей при рассмотрении профилактического применения.
В следующих вариантах осуществления комбинированная блокада PD-1 и CTLA-4 (т.е. иммуностимулирующих терапевтических антител анти-PD-1-антитела и анти-CTLA-4-антитела) может дополнительно комбинироваться с применением любого неабсорбируемого стероида. Как использовано в данном описании, «неабсорбируемый стероид» означает глюкокортикоид, который проявляет экстенсивный метаболизм первого прохождения таким образом, что после метаболизма в печени биодоступность стероида является низкой, т.е. приблизительно 20%. В одном варианте осуществления настоящего изобретения, неабсорбируемым стероидом является будесонид. Будесонид является локально действующим глюкокортикостероидом, который экстенсивно метаболизируется, прежде всего печенью, после пероральнрого введения. ENTOCORT EC® (Astra-Zeneca) является зависимой от pH и времени пероральной формой будесонида, разработанной для оптимизации доставки лекарственного средства в подвздошную кишку и через ободочную кишку. ENTOCORT EC® одобрен в США для лечения мягкой-умеренной болезни Крона, включающей подвздошную и/или восходящую ободочную кишку. Обычная пероральная доза ENTOCORT EC® для лечения болезни Крона составляет 6-9 мг/день. ENTOCORT EC® высвобождается в кишечнике перед абсорбцией и сохраняется в слизистой оболочке кишечника. После прохождения его через ткань слизистой оболочки кишечника, ENTOCORT EC® экстенсивно метаболизируется системой цитохрома Р450 в печени до метаболитов с незначительной глюкокортикоидной активностью. Таким образом, биодоступность является низкой (приблизительно 10%). Такая низкая биодоступность будесонида приводит к улучшенному терапевтическому индексу в сравнении с другими глюкокортикоидами с менее экстенсивным метаболизмом первого прохождения. Будесонид приводит к меньшим побочным эффектам, включая меньшую гипоталамическую-гипофизарную супрессию, чем у системно действующих кортикостероидов. Однако продолжительное введение ENTOCORT EC® может привести к системным эффектам глюкокортикоидов, таким как супрессия гиперкортицизма и надпочечников. См. PDR 58th ed. 2004; 608-610.
В других вариантах осуществления комбинированная блокада PD-1 и CTLA-4 (т.е. иммуностимулирующих терапевтических анти-PD-1 и анти-CTLA-4-антител) вместе с неабсорбируемым стероидом может быть дополнительно комбинирована с салицилатом. Салицилаты включают агенты 5-ASA, такие как, например: сульфасалазин (AZULFIDINE®, Pharmacia & UpJohn); олсалазин (DIPENTUM®, Pharmacia & UpJohn); балсалазин (COLAZAL®, Salix Pharmaceuticals, Inc.) и месаламин (ASACOL®, Procter & Gamble Pharmaceuticals; PENTASA®, Shire US; CANASA®, Axcan Scandipharm, Inc.; ROWASA®, Solvay).
В соответствии со способами настоящего изобретения, салицилат, вводимый в комбинации с анти-PD-1 и анти-CTLA-4-антителами и неабсорбируемым стероидом, может предусматривать любое перекрывающееся или последовательное введение салицилата и неабсорбируемого стероида с целью уменьшения заболеваемости колитом, индуцируемой иммуностимулирующими антителами. Таким образом, например, способы уменьшения заболеваемости колитом, индуцируемой иммуностимулирующими антителами по данному изобретению, предусматривают введение салицилата и неабсорбируемого стероида одновременно или последовательно (например, салицилат вводят спустя 6 часов после введения неабсорбируемого стероида) или любой их комбинации. Кроме того, в соответствии с данным изобретением, салицилат и неабсорбируемый стероид могут вводиться одним и тем же способом (например, оба вводят перорально) или различными способами (например, салицилат вводят перорально, и неабсорбируемый стероид вводят ректально), которые могут отличаться от способа (способов), используемых для введения анти-PD-1 и анти-CTLA-4-антител.
Настоящее изобретение дополнительно иллюстрировано следующими примерами, которые не должны рассматриваться как дополнительное ограничение. Содержание всех фигур и всех цитируемых ссылок, патентов и опубликованных заявок на патент, специально включены в данное описание посредством ссылки.
ПРИМЕРЫ
Пример 1
Генерирование моноклональных антител человека против PD-1
Антиген
Протоколы иммунизации использовали в качестве антигена как (i) рекомбинантный слитый белок, содержащий внеклеточную часть PD-1, так и (ii) мембраносвязанный полноразмерный PD-1. Оба антигена генерировали рекомбинантными способами трансфекции в клеточной линии СНО.
Трансгенные мыши HuMab и KM™
Полные моноклональные антитела к PD-1 получали с использованием штамма НСо7 трансгенных мышей HuMab и штамма КМ трансгенных трансхромосомных мышей, каждый из которых экспрессирует гены антител человека. В каждом из указанных мышиных штаммов эндогенный ген легкой цепи каппа мыши был гомозиготно разрушен, как описано в Chen et al. (1993) EMBO J. 12:811-820 и эндогенный ген тяжелой цепи мыши был гомозиготно разрушен, как описано в примере 1 публикации PCT WO 01/09187. Каждый из указанных штаммов мыши несет трансген легкой цепи каппа человека, KCo5, как описано в Fishwild et al. (1996) Nature Biotechnology 14:845-851. Штамм HCo7 несет трансген тяжелой цепи человека HCo7, как описано в патентах США №№ 5545806; 5625825 и 5545807. Штамм KM содержит трансхромосому SC20, как описано в публикации PCT WO 02/43478.
Иммунизации HuMab и KM:
Для генерирования полных моноклональных антител человека к PD-1, мышей HuMab и мышей KMTM иммунизировали очищенным рекомбинантным слитым белком PD-1 и PD-1-трансфицированными клетками CHO в качестве антигена. Общие схемы иммунизации для мышей HuMab описаны Lonberg, N. et al (1994) в Nature 368(6474): 856-859; Fishwild, D. et al. (1996) Nature Biotechnology 14: 845-851 и публикации PCT WO 98/24884. Мыши имели возраст 6-16 недель после первой инфузии антигена. Рекомбинантный препарат (5-50 мкг) антигена PD-1-слитого белка и 5-10×106 клеток использовали для иммунизации мышей HuMab и мышей KMTM внутрибрюшинно, подкожно (SC) или посредством инъекции в подушечку лапы.
Трансгенных мышей иммунизировали дважды антигеном в полном адъюванте Фрейнда или адъюванте Риби IP, с последующими иммунизациями IP в течение 3-21 дня (в целом до 11 иммунизаций) антигеном в неполном адъюванте Фрейнда или адъюванте Риби. Иммунную реакцию подвергали мониторингу с использованием ретроорбитальных кровопусканий. Плазму подвергали скринингу при помощи ELISA (как описано ниже) и мышей с достаточными титрами анти-PD-1-иммуноглобулина человека использовали для слияний. Мышей повторно иммунизировали внутривенно антигеном 3 дня перед умерщвлением и извлечением селезенки. Обычно выполняли 10-35 слияний для каждого антигена. Каждым антигеном иммунизировали несколько дюжин мышей.
Отбор мышей HuMab или мышей KMTM, продуцирующих анти-PD-1-антитела:
Для отбора мышей HuMab или мышей KMTM, продуцирующих антитела, которые связывают PD-1, сыворотки из иммунизированных мышей испытывали при помощи ELISA, как описано Fishwild, D. et al. (1996). Кратко, микротитрационные планшеты покрывали очищенным рекомбинантным PD-1-слитым белком из трансфицированных клеток СНО при 1-2 мкг/мл в PBS, 100 мкл на лунку, инкубировали при 4°С в течение ночи и затем блокировали 200 мкл на лунку 5% фетальной телячьей сыворотки в смеси PBS/Твин (0,05%). Разведения сывороток из иммунизированных PD-1 мышей добавляли в каждую лунку и инкубировали в течение 1-2 часов при температуре окружающей среды. Планшеты промывали смесью PBS/Твин и затем инкубировали с поликлональным козьим антителом против IgG человека, конъюгированным с пероксидазой хрена (HRP), в течение 1 часа при комнатной температуре. После промывания, планшеты проявляли с субстратом ABTS (Sigma, A-1888, 0,22 мг/мл) и анализировали на спектрофотометре при OD 415-495. Мышей, которые развивали самые высокие титры анти-PD-1-антител, использовали для слияний. Слияния выполняли, как описано ниже, и супернатанты гибридом испытывали на анти-PD-1-активность при помощи ELISA.
Генерирование гибридом, продуцирующих моноклональные антитела к PD-1 человека:
Спленоциты мышей, выделенные из мышей HuMab или KM, сливали с мышиной линией миеломных клеток либо с использованием стандартных протоколов на основе ПЭГ, либо с использованием электрослияния на основе электрического поля при помощи электропоратора для слияния клеток с большой камерой (Cyto Pulse Sciences, Inc., Glen Burnie, MD). Затем полученные гибридомы подвергали скринингу на продуцирование антигенспецифических антител. Суспензии отдельных клеток спленоцитов из иммунизированных мышей сливали с одной четвертой количества не секретирующих SP2/0 мышиных миеломных клеток (АТСС, CRL 1581) c 50% ПЭГ (Sigma). Клетки высевали при приблизительно при 1×105 на лунку в плоскодонном микротитрационном планшете с последующей приблизительно двухнедельной инкубацией в селективной среде, содержащей 10% фетальную телячью сыворотку, 10% P388D1 (ATCC, CRL TIB-63) кондиционированную среду, 3-5% ориген (IGEN) в DMEM (Mediatech, CRL 10013, с высокой глюкозой, L-глутамином и пируватом натрия) плюс 5 мМ HEPES, 0,055 мМ 2-меркаптоэтанол, 50 мг/мл гентамицин и 1× HAT (Sigma, CRL P-7185). Спустя 1-2 недели, клетки культивировали в среде, в которой HAT была заменена HT. Затем отдельные клетки подвергали скринингу при помощи ELISA (как описано выше) на моноклональные анти-PD-1-IgG человека. После появления экстенсивного роста гибридом среду подвергали мониторингу обычно спустя 10-14 дней. Секретирующие антитела гибридомы повторно высевали, снова подвергали скринингу и, если они были все еще положительными в отношении IgG человека, моноклональные анти-PD-1-антитела субклонировали по меньшей мере два раза с использованием лимитирующего разведения. Затем стабильные субклоны культивировали in vitro для генерирования малых количеств антитела в среде для культуры ткани для последующей характеристики.
Для последующего анализа были отобраны гибридомные клоны 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4.
Пример 2
Структурная характеристика моноклональных антител человека 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4
кДНК-последовательности, кодирующие вариабельные области тяжелой и легкой цепей моноклональных антител 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4, получали из гибридом 17D8, 2D3, 4H1, 5C4, 4A11, 7D3 и 5F4, соответственно, с использованием стандартных ПЦР-способов и секвенировали с использованием стандартных способов секвенирования ДНК.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи 17D8 показаны на фиг.1А и в SEQ ID NO:57 и 1, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи 17D8 показаны на фиг.1В и в SEQ ID NO:64 и 8, соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 17D8 с известными последовательностями тяжелых цепей иммуноглобулина зародышевой линии человека продемонстрировало, что тяжелая цепь 17D8 использует VH-сегмент из сегмента VH 3-33 зародышевой линии человека, неопределенный сегмент D и сегмент JH из JH 4b зародышевой линии человека. Сопоставление последовательности VH 17D8 c последовательностью VH 3-33 зародышевой линии человека показано на фиг.8. Дальнейший анализ последовательности VH 17D8 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 тяжелой цепи, как показано на фиг.1A и 8 и в SEQ ID NO:15, 22 и 29, соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 17D8 с известными последовательностями легкой цепи зародышевой линии человека продемонстрировало, что легкая цепь 17D8 использует сегмент VL из Vκ L6 зародышевой линии человека и сегмент JK из JK 4 зародышевой линии человека. Сопоставление последовательности VL 17D8 с последовательностью Vκ L6 зародышевой линии человека показано на фиг.9. Дальнейший анализ последовательности VL 17D8 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 легкой цепи, как показано на фиг.1В и 9 и в SEQ ID NO:36, 43 и 50, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи 2D3 показаны на фиг.2А и в SEQ ID NO:58 и 2, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи 2D3 показаны на фиг.2В и в SEQ ID NO:65 и 9, соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 2D3 с известными последовательностями тяжелых цепей иммуноглобулина зародышевой линии человека продемонстрировало, что тяжелая цепь 2D3 использует VH-сегмент из сегмента VH 3-33 зародышевой линии человека, сегмент D из 7-27 зародышевой линии человека и сегмент JH из JH 4b зародышевой линии человека. Сопоставление последовательности VH 2D38 c последовательностью VH 3-33 зародышевой линии человека показано на фиг.8. Дальнейший анализ последовательности VH 17D8 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 тяжелой цепи, как показано на фиг.2A и 8 и в SEQ ID NO:16, 23 и 30, соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 2D38 с известными последовательностями легкой цепи зародышевой линии человека продемонстрировало, что легкая цепь 2D38 использует сегмент VL из Vκ L6 зародышевой линии человека и сегмент JK из JK 4 зародышевой линии человека. Сопоставление последовательности VL 2D3 с последовательностью Vκ L6 зародышевой линии человека показано на фиг.9. Дальнейший анализ последовательности VL 2D3 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 легкой цепи, как показано на фиг.2В и 9 и в SEQ ID NO:37, 44 и 51, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи 4Н1 показаны на фиг.3А и в SEQ ID NO:59 и 3, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи 4Н1 показаны на фиг.3В и в SEQ ID NO:66 и 10, соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 4Н1 с известными последовательностями тяжелых цепей иммуноглобулина зародышевой линии человека продемонстрировало, что тяжелая цепь 4Н1 использует VH-сегмент из сегмента VH 3-33 зародышевой линии человека, неопределенный сегмент D и сегмент JH из JH 4b зародышевой линии человека. Сопоставление последовательности VH 4Н1 c последовательностью VH 3-33 зародышевой линии человека показано на фиг.8. Дальнейший анализ последовательности VH 4Н1 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 тяжелой цепи, как показано на фиг.3А и 8 и в SEQ ID NO:17, 24 и 31, соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 4Н1 с известными последовательностями легкой цепи зародышевой линии человека продемонстрировало, что легкая цепь 4Н1 использует сегмент VL из Vκ L6 зародышевой линии человека и сегмент JK из JK 1 зародышевой линии человека. Сопоставление последовательности VL 4Н1 с последовательностью Vκ L6 зародышевой линии человека показано на фиг.10. Дальнейший анализ последовательности VL 4Н1 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 легкой цепи, как показано на фиг.3В и 10 и в SEQ ID NO:38, 45 и 52, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи 5С4 показаны на фиг.4А и в SEQ ID NO:60 и 4, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи 5С4 показаны на фиг.4В и в SEQ ID NO:67 и 11, соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 5С4 с известными последовательностями тяжелых цепей иммуноглобулина зародышевой линии человека продемонстрировало, что тяжелая цепь 5С4 использует сегмент VH из сегмента VH 3-33 зародышевой линии человека, неопределенный сегмент D и сегмент JH из JH 4b зародышевой линии человека. Сопоставление последовательности VH 5С4 c последовательностью VH 3-33 зародышевой линии человека показано на фиг.8. Дальнейший анализ последовательности VH 5С4 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 тяжелой цепи, как показано на фиг.4А и 8, и в SEQ ID NO:18, 25 и 32, соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 5С4 с известными последовательностями легкой цепи зародышевой линии человека продемонстрировало, что легкая цепь 5С4 использует сегмент VL из Vκ L6 зародышевой линии человека и сегмент JK из JK 1 зародышевой линии человека. Сопоставление последовательности VL 5С4 с последовательностью Vκ L6 зародышевой линии человека показано на фиг.10. Дальнейший анализ последовательности VL 5С4 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 легкой цепи, как показано на фиг.4В и 10 и в SEQ ID NO:39, 46 и 53, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи 4А11 показаны на фиг.5А и в SEQ ID NO:61 и 5, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи 4А11 показаны на фиг.5В и в SEQ ID NO:68 и 12, соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 4А11 с известными последовательностями тяжелых цепей иммуноглобулина зародышевой линии человека продемонстрировало, что тяжелая цепь 4А11 использует сегмент VH из сегмента VH 4-39 зародышевой линии человека, сегмент D из 3-9 зародышевой линии человека и сегмент JH из JH 4b зародышевой линии человека. Сопоставление последовательности VH 4А11 c последовательностью VH 4-39 зародышевой линии человека показано на фиг.11. Дальнейший анализ последовательности VH 4А11 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 тяжелой цепи, как показано на фиг.5А и 11 и в SEQ ID NO:19, 26 и 33, соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 4А11 с известными последовательностями легкой цепи зародышевой линии человека продемонстрировало, что легкая цепь 4А11 использует сегмент VL из Vκ L15 зародышевой линии человека и сегмент JK из JK 1 зародышевой линии человека. Сопоставление последовательности VL 4А11 с последовательностью Vκ L6 зародышевой линии человека показано на фиг.12. Дальнейший анализ последовательности VL 4А11 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 легкой цепи, как показано на фиг.5В и 12 и в SEQ ID NO:40, 47 и 54, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи 7D3 показаны на фиг.7А и в SEQ ID NO:62 и 6, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи 7D3 показаны на фиг.7В и в SEQ ID NO:69 и 13, соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 7D3 с известными последовательностями тяжелых цепей иммуноглобулина зародышевой линии человека продемонстрировало, что тяжелая цепь 7D3 использует сегмент VH из сегмента VH 3-33 зародышевой линии человека, сегмент D из 7-27 зародышевой линии человека и сегмент JH из JH 4b зародышевой линии человека. Сопоставление последовательности VH 7D3 c последовательностью VH 3-33 зародышевой линии человека показано на фиг.8. Дальнейший анализ последовательности VH 7D3 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 тяжелой цепи, как показано на фиг.6А и 8 и в SEQ ID NO:20, 27 и 34, соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 7D3 с известными последовательностями легкой цепи зародышевой линии человека продемонстрировало, что легкая цепь 4А11 использует сегмент VL из Vκ L6 зародышевой линии человека и сегмент JK из JK 4 зародышевой линии человека. Сопоставление последовательности VL 7D3 с последовательностью Vκ L6 зародышевой линии человека показано на фиг.9. Дальнейший анализ последовательности VL 7D3 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 легкой цепи, как показано на фиг.6В и 9 и в SEQ ID NO:41, 48 и 55, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи 5F4 показаны на фиг.7А и в SEQ ID NO:63 и 7, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи 5F4 показаны на фиг.7В и в SEQ ID NO:70 и 14, соответственно.
Сравнение иммуноглобулиновой последовательности тяжелой цепи 5F4 с известными последовательностями тяжелых цепей иммуноглобулина зародышевой линии человека продемонстрировало, что тяжелая цепь 5F4 использует VH-сегмент из сегмента VH 4-39 зародышевой линии человека, сегмент D из 3-9 зародышевой линии человека и сегмент JH из JH 4b зародышевой линии человека. Сопоставление последовательности VH 5F4 c последовательностью VH 4-39 зародышевой линии человека показано на фиг.11. Дальнейший анализ последовательности VH 5F4 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 тяжелой цепи, как показано на фиг.7А и 11, и в SEQ ID NO:21, 28 и 35, соответственно.
Сравнение иммуноглобулиновой последовательности легкой цепи 5F4 с известными последовательностями легкой цепи зародышевой линии человека продемонстрировало, что легкая цепь 5F4 использует сегмент VL из Vκ L15 зародышевой линии человека и сегмент JK из JK 1 зародышевой линии человека. Сопоставление последовательности VL 5F4 с последовательностью Vκ L6 зародышевой линии человека показано на фиг.12. Дальнейший анализ последовательности VL 5F4 с использованием системы Кабата определения CDR-областей привел к очерчиванию областей CDR1, CDR2 и CD3 легкой цепи, как показано на фиг.7В и 12 и в SEQ ID NO:42, 49 и 56, соответственно.
Пример 3
Характеристика специфичности связывания и кинетики связывания моноклональных анти-PD-1-антител человека
В данном примере аффинность связывания и кинетику связывания анти-PD-1-антител испытывали при помощи анализа Biacore. Специфичность связывания и перекрестную конкуренцию испытывали при помощи проточной цитометрии.
Аффинность и кинетика связывания
Анти-PD-1-антитела характеризовали в отношении аффинностей и кинетики связывания при помощи анализа Biacore (Biacore AB, Uppsala, Sweden). Очищенный рекомбинантный PD-1-слитый белок человека ковалентно связывали с чипом СМ5 (покрытым карбоксиметилдекстраном чипом) через первичные аминогруппы, с использованием стандартного химического способа связывания аминогрупп и набора, обеспеченного Biacore. Связывание измеряли с использованием протекания антител в буфере HBS EP (обеспечиваемом Biacore AB) при концентрации 267 нМ при скорости потока 50 мкл/мин. Кинетику ассоциации антиген-антитело прослеживали в течение 3 минут и кинетику диссоциации прослеживали в течение 7 минут. Кривые ассоциации и диссоциации строили относительно модели связывания 1:1 Лангмуира с использованием программного обеспечения BIAevaluation (Biacore AB). Для минимизации эффектов авидности в оценке констант связывания для построения кривых использовали только начальный сегмент данных, соответствующих фазам ассоциации и диссоциации. Величины KD, kon и koff, которые были определены, показаны в таблице 2.
Специфичность связывания согласно проточной цитометрии
Получали линии клеток яичника китайского хомячка (CHO), которые экспрессируют рекомбинантный PD-1 человека на клеточной поверхности, и использовали их для определения специфичности моноклональных анти-PD-1-антител человека при помощи проточной цитометрии. Клетки CHO трансфицировали экспрессионными плазмидами, содержащими полноразмерную кДНК, кодирующую трансмембранные формы PD-1. Связывание моноклональных анти-PD-1-антител человека 5С4 и 4Н1 оценивали инкубированием трансфицированных клеток с моноклональными анти-PD-1-антителами при концентрации 20 мкг/мл. Клетки промывали и связывание детектировали с использованием FITC-меченого антитела против IgG Ab человека. Анализы проточной цитометрии выполняли с использованием проточной цитометрии FACScan (Becton Dickinson, San Jose, CA). Результаты изображены на фиг. 13A (5C4) и 13B (4H1). Моноклональные анти-PD-1-антитела связывались с клетками СНО, трансфицированными PD-1, но не с клетками СНО, которые не были трансфицированы PD-1 человека. Приведенные данные демонстрируют специфичность моноклональных анти-PD-1-антител человека в отношении PD-1.
Специфичность связывания согласно ELISA в сравнении с другими членами семейства CD28
Сравнение связывания анти-PD-1-антител с членами семейства CD28 выполняли при помощи стандартного ELISA с использованием четырех различных членов семейства CD28 для испытания специфичности связывания в отношении PD-1.
Слитые белки членов семейства CD28, ICOS, CTLA-4 и CD28 (R&D Biosystems), испытывали на связывание с моноклональными анти-PD-1-антителами человека 17D8, 2D3, 4H1, 5C4 и 4A11. Выполняли стандартные процедуры ELISA. Моноклональные анти-PD-1-антитела человека добавляли при концентрации 20 мкг/мл. Козье (специфическое в отношении каппа-цепи) поликлональное антитело против IgG человека, конъюгированное с пероксидазой хрена (HRP), использовали в качестве второго антитела. Результаты показаны на фиг.14. Каждое из моноклональных анти-PD-1-антител человека связывалось с высокой специфичностью с PD-1, но не с другими членами семейства CD28.
Пример 4
Характеристика связывания анти-PD-1-антитела с PD-1, экспрессируемым на поверхности клеток человека и обезьяны
Анти-PD-1-антитела испытывали на связывание с клетками, экспрессирующими PD-1, на их клеточной поверхности, при помощи проточной цитометрии.
Активированные Т-клетки человека, мононуклеарные клетки периферической крови (PBMC) обезьяны и клетки СНО, трансфицированные PD-1, испытывали на связывание антител. Т-клетки человека и PBMC собакоподобной обезьяны активировали анти-PD-1-антителом для индукции экспрессии PD-1 на Т-клетках перед связыванием с моноклональным анти-PD-1-антителом человека. Связывание моноклональных анти-PD-1-антител человека 5С4 и 4Н1 оценивали инкубированием трансфицированных клеток либо с формами IgG1, либо с формами IgG4 моноклональных анти-PD-1-антител человека при различных концентрациях. Клетки промывали и связывание детектировали FITC-меченым антителом против IgG человека. Анализы проточной цитометрии выполняли с использованием проточной цитометрии FACScan (Becton Dickinson, San Jose, CA). Результаты показаны на фиг. 15А (активированные Т-клетки человека), 15В (PBMC собакоподобной обезьяны) и 15С (PD-1-трансфицированные клетки СНО). Моноклональные анти-PD-1-антитела 5С4 и 4Н1 связывались с активированными Т-клетками человека, активированными PBMC обезьяны и клетками СНО, трансфицированными PD-1 человека, как измерено по средней интенсивности флуоресценции (MFI) окрашивания. Приведенные данные демонстрируют, что анти-PD-1-HuMAb связываются с PD-1 поверхности клеток как человека, так и собакоподобной обезьяны.
Пример 5
Действие анти-PD-1-антител человека на пролиферацию клеток и продуцирование цитокинов в реакции смешанных лимфоцитов
Реакцию смешанных лимфоцитов использовали для демонстрации эффекта блокирования PD-1-пути в отношении лимфоцитарных эффекторных клеток. Т-клетки в данном анализе тестировали на пролиферацию, секрецию IFN-гамма и секрецию IL-2 в присутствии и в отсутствие антитела анти-PD-1-HuMAb.
Т-клетки очищали из PBMC с использованием колонки обогащения CD4+ T-клеток человека (R&D systems). Каждая культура содержала 105 очищенных Т-клеток и 104 аллогенных дендритных клеток в общем объеме 200 мкл. Моноклональное анти-PD-1-антитело 5C4, 4H1, 17D8, 2D3 или Fab-фрагмент 5C4 добавляли к каждой культуре при различных концентрациях антител. Отсутствие антитела или контрольное изотипически сходное антитело добавляли в качестве отрицательного контроля. Клетки культивировали в течение 5 дней при 37°С. После дня 5 из каждой культуры отбирали 100 мкл среды для измерения цитокинов. Уровни IFN-гамма и других цитокинов измеряли с использованием наборов OptEIA ELISA (BD Biosciences). Клетки метили 3H-тимидином, культивировали еще в течение 18 часов и анализировали на пролиферацию клеток. Результаты показаны на фиг.16А (пролиферация Т-клеток), 16В (секреция IFN-γ) и 16С (секреция IL-2). Моноклональные анти-PD-1-антитела стимулировали пролиферацию Т-клеток, секрецию IFN-гамма и секрецию IL-2 зависимым от концентрации образом. Fab-фрагмент 5С4 также стимулировал пролиферацию Т-клеток, секрецию IFN-гамма и секрецию IL-2 зависимым от концентрации образом. В противоположность этому, культуры, содержащие контрольное изотопически сходное антитело, не обнаруживали увеличения пролиферации Т-клеток, секреции IFN-гамма или IL-2.
Пример 6
Блокирование связывания лиганда с PD-1 анти-PD-1-антителами человека
Анти-PD-1-HuMAb испытывали на способность блокировать связывание лигандов PD-L1 и PD-L2 с PD-1, экспрессируемым на трансфицированных клетках СНО, с использованием анализа проточной цитометрии.
Экспрессирующие PD-1 клетки CHO суспендировали в FACS-буфере (PBS с 4% фетальной телячьей сывороткой). Различные концентрации анти-PD-1-HuMAb 5C4 и 4H1 добавляли к суспензии клеток и инкубировали при 4°С в течение 30 минут. Несвязанное антитело вымывали и в пробирки добавляли либо FITC-меченый PD-L1-слитый белок, либо FITC-меченый PD-L2-слитый белок и инкубировали при 4°С в течение 30 минут. Анализы проточной цитометрии выполняли с использованием проточного цитометра FACScan (Becton Dickinson, San Jose, CA). Результаты изображены на фиг.17А (блокирование PD-L1) и 17B (блокирование PD-L2). Моноклональные анти-PD-1-антитела 5C4 и 4H1 блокировали связывание PD-L1 и PD-L2 с клетками CHO, трансфицированными PD-1 человека, как измерено по средней интенсивности флуоресценции (MFI) окрашивания. Приведенные данные демонстрируют, что анти-PD-1-HuMAb блокируют связывание лиганда (как PD-L1, так и PD-L2) с PD-1 поверхности клеток.
Пример 7
Действие анти-PD-1-антител человека на высвобождение цитокинов в крови человека
Анти-PD-1-HuMAb смешивали со свежей цельной кровью человека для определения, только ли анти-PD-1-HuMAb стимулируют высвобождение определенных цитокинов из клеток крови человека.
500 мкл гепаринизированной свежей цельной крови человека добавляли в каждую лунку. В каждую лунку добавляли либо 10 мкг, либо 100 мкг анти-PD-1-HuMAb (4H1 или 5C4, последнее в виде изотипа IgG1 или изотипа IgG4). Некоторые лунки инкубировали с анти-CD3-антителом в качестве положительного контроля или с IgG1 или IgG4-антителом человека в качестве изотипически сходных отрицательных контролей. Клетки инкубировали при 37°C в течение либо 6, либо 24 часов. Клетки осаждали центрифугированием и плазму собирали для измерения цитокинов IFN-гамма, TNF-альфа, IL-2, IL-4, IL-6, IL-10 и IL-12 с использованием цитометрического анализа цитокинов на множестве гранул (BD Biosciences). Концентрация каждого цитокина (пг/мл) показана в таблицах 3а, с 6-часовой инкубацией, и 3b, с 24-часовой инкубацией, ниже. Полученные результаты показывают, что обработка только анти-PD-1-антителами 5C4 и 4H1 не стимулировала клетки крови человека для высвобождения ни одного из цитокинов IFN-гамма, TNF-альфа, IL-2, IL-4, IL-6, IL-10 и IL-12.
Пример 8
Действие анти-PD-1-антител на апоптоз Т-клеток
Действие анти-PD-1-антител на индукцию апоптоза Т-клеток измеряли с использованием тест окрашивания аннексином V.
Т-клетки культивировали в реакции смешанных лимфоцитов, как описано выше в примере 5. Анти-PD-1-антитело 5C4 добавляли в пробирку в концентрации 25 мкг/мл. Неспецифическое антитело использовали в качестве контроля. Аннексин V и пропидийиодид добавляли в соответствии со стандартным протоколом (BD Biosciences). Полученную смесь инкубировали в течение 15 минут в темноте при комнатной температуре и затем анализировали с использованием проточного цитометра FACScan (Becton Dickinson, San Jose, CA). Результаты показаны на фиг.18. Анти-PD-1-антитело 5C4 не действовало на апоптоз Т-клеток.
Пример 9
Действие анти-PD-1-антител на секрецию цитокина стимулированными вирусом клетками PBMC из вирус-положительного донора
В данном примере мононуклеарные клетки периферической крови (PBMC) из донора, положительного в отношении CMV, выделяли и подвергали действию лизата CMV в присутствии и в отсутствие анти-PD-1-антител для испытания действия указанных антител на секрецию цитокина, стимулированную антигеном.
2×105 PMBC человека из CMV-положительного донора культивировали в общем объеме 200 мкл и добавляли в каждую лунку вместе с лизатом CMV-инфицированных клеток. Анти-PD-1-HuMAb 5C4 добавляли в каждую лунку в различных концентрациях в течение 4 дней. После дня 4, 100 мкл среды отбирали из каждой культуры для измерения цитокина. Уровень IFN-гамма измеряли с использованием наборов OptEIA ELISA (BD Biosciences). Клетки метили 3H-тимидином, культивировали еще в течение 18 часов и анализировали на пролиферацию клеток. Пролиферацию клеток анализировали с использованием реагента Cell Titer-Glo reagent (Promega). Результаты показаны на фиг.19. Анти-PD-1-HuMab 5C4 увеличивало секрецию IFN-гамма зависимым от концентрации образом. Полученные результаты показывают, что анти-PD-1-HuMAb могут стимулировать высвобождение IFN-гамма в Т-клеточной ответные реакции памяти из клеток PBMC, стимулированных предварительно антигеном.
Пример 10
Действие анти-PD-1-антитела на реакцию вторичного антитела на антиген
Мышей иммунизировали и повторно иммунизировали TI-антигеном (DNP-Ficoll), а также обрабатывали крысиным анти-mPD-1-антителом или контрольным антителом для испытания действия анти-PD-1-антитела на титры антител.
Самок мышей C57BL6 делили на две группы с 6 мышами на одну группу. Одну группу обрабатывали контрольным крысиным IgG, и другую крысиным анти-mPD-1-антителом. Этих мышей иммунизировали 5 мкг DNP-Ficoll (T1-антигена) в 50 мкл полного адъюванта Фрейнда (CFA) i.p. в день 0. Любое контрольное крысиное IgG-антитело или крысиное анти-mPD-1-антитело (200 мкг/мышь) вводили i.p. в дни -1, 0 и 2. После четырех недель мышей повторно иммунизировали 5 мкг DNP-Ficoll в 50 мкл неполного адъюванта Фрейнда (IFA) i.p. в день 0. Крысиное анти-mPD-1-антитело или контрольное антитело (200 µg/мышь) вводили i.p. в дни 0 и 1. Титры антител измеряли стандартным анализом ELISA в день 7 после вторичной иммунизации. Полученные результаты показаны в таблице 4 ниже. У мышей, обработанных анти-mPD-1-антителом, как изотип IgM, так и изотип IgG3 показали наивысшее увеличение титра после иммунизации Т1-антигеном, в сравнении с мышами, обработанными контрольным антителом. Полученные результаты демонстрируют, что обработка анти-PD-1-антителом может увеличивать титры антитела в реакции на T1-антиген.
антитела
Пример 11
Лечение на модели опухоли in vivo с использованием анти-PD-1-антител
Мышей, имплантированных раковой опухолью, лечили in vivo анти-PD-1-антителами для испытания действия in vivo указанных антител на рост опухоли. В качестве положительного контроля использовали анти-CTLA-4-антитела, поскольку было показано, что такие антитела ингибируют рост опухоли in vivo.
В данном эксперименте используемым анти-PD-1-антителом было химерное крысиное анти-mPD-1-антитело, полученное с использованием хорошо известных лабораторных способов. Для генерирования крысиного анти-mPD-1-антитела, крыс иммунизировали мышиными клетками, трансфицированными для экспрессии рекомбинантного мышиного PD-1-слитого белка (R&D Systems Catalog No. 1021-PD), и моноклональные антитела подвергали скринингу на связывание с мышиным антигеном PD-1 при помощи анализа ELISA. Затем V-области крысиного анти-PD-1-антитела рекомбинантно связывали с константной областью мышиного IgG1 с использованием стандартных способов молекулярной биологии и подвергали повторному скринингу на связывание с мышиным PD-1 при помощи ELISA и FACS. Полученное химерное крысиное антитело против мышиного PD-1 (крысиное анти-mPD-1-антитело), используемое в данном случае, было названо 4Н2.
Для исследований на опухолях, самок мышей AJ в возрасте 6-8 недель (Harlan Laboratories) делили случайным образом по массе на 6 групп. Мышей имплантировали подкожно в правом боку 2×106 клетками фибросаркомы SA1/N, растворенными в 200 мкл DMEM в день 0. Мышей обрабатывали PBS-носителем или антителами при 10 мг/кг. Животным вводили дозу внутрибрюшинной инъекцией приблизительно 200 мкл PBS, содержащих антитело или носитель, в дни 1, 4, 8 и 11. Каждая группа, содержала 10 животных и все группы состояли из следующих групп: (i) группа носителя, (ii) контроль IgG мыши, (iii) контроль IgG хомячка, (iv) антитело хомячка против мышиного CTLA-4 и (v) химерное анти-PD-1-антитело 4Н2. Мышей подвергали мониторингу два раза в неделю на рост опухоли в течение приблизительно 6 недель. С использованием электронного штангенциркуля опухоли измеряли в трех измерениях (высота×ширина×длина) и рассчитывали объем опухоли. Мышей подвергали эвтаназии, когда опухоли достигали конечной точки (1500 мм3) или обнаруживали большую, чем 15%, потерю массы. Результаты показаны на фиг.20. Анти-PD-1-антитело продлевало среднее время до достижения объема опухоли конечной точки (1500 мм3) с ~25 дней в контрольных группах до ~40 дней. Таким образом, лечение анти-PD-1-антителом оказывало прямое ингибирующее действие на рост опухоли in vivo.
Пример 12
Генерирование химерного (крыса-мышь) анти-PD-1-антитела 4H2
Крысиное моноклональное антитело против мышиных анти-PD-1-антител (крысиное анти-mPD-1) получали из крыс, иммунизированных mPD-1-hFc-слитым белком с использованием стандартных способов получения гибридом (см. Kohler and Milstein (1975) Nature 256:495; и Harlow and Lane (1988) Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor New York). Восемь гибридом субклонировали и антитела выделяли и подвергали скринингу на их способность блокировать связывание мышиного PD-L2 (mPD-L2) с mPD-1. Были идентифицированы несколько анти-mPD-1-антител, способных блокировать связывание mPD-L2 с mPD-1 (см. например, активность 4H2, фиг.41), и определяли аффинность связывания нескольких из указанных антител с mPD-1-Fc-слитым белком при помощи ELISA (фиг.42).
Было дополнительно охарактеризовано антитело 4H2.B3, которое называют взаимозаменяемо “4H2.” Клетки CHO, экспрессирующие мышиный PD-1, конструировали и инкубировали с анти-mPD-1-антителом 4H2 при диапазоне концентраций от 200 мкг/мл до 0,012 мкг/мл для определения аффинности связывания 4H2 с PD-1. Связывание анти-mPD-1-антитела с экспрессирующими PD-1 клетками CHO детектировали инкубированием с ослиным антителом против крысиного IgG, конъюгированным с FITC, и измеряли при помощи FACS. Анти-mPD-1-антитело имело EC50 (50% эффективную концентрацию) приблизительно 0,38 мкг (фиг.43) и KD 4,7×10-9M. Для испытания ингибирования связывания PD-L1 с PD-1, выполняли тот же самый анализ, за исключением того, что клетки инкубировали также с 0,16 мкг mPD-L1-hFc-слитым белком, затем связывание PD-L1 с экспрессирующими PD-1 клетками CHO детектировали инкубированием с козьими антителами против IgG человека (Fc-специфическими), конъюгированными с FITC, и измерением сигнала связывания при помощи FACS (MFI, средней интенсивности флуоресценции). Анти-mPD-1-антитело имело EC50 приблизительно 0,72 мкг (фиг.44).
Для применении на мышиных моделях опухолей крысиное антитело анти-mPD-1 должно быть модифицировано таким образом, чтобы иммунная система мыши не могла нейтрализовать иммунотерапевтическое антитело (т.е. таким образом данное антитело будет иметь лучшую фармакокинетику), и чтобы избежать антителозависимой клеточной цитотоксичности (ADCC) уменьшением Fc-рецепторных взаимодействий (т.е. таким образом блокада анти-PD-1 может быть оценена как ухудшенная эффектами ADCC). Было определено, что исходное крысиное анти-mPD-1-антитело, 4H2, является крысиным изотипом IgG2. Таким образом, Fc-часть этого антитела 4Н2 была заменена Fc-частью из мышиного изотипа IgG1. С использованием описанного выше анализа было обнаружено, что аффинность связывания химерного крыса-мышь антитела 4Н2 в отношении mPD-1 является сравнимой с аффинностью связывания крысиного анти-mPD-1-антитела 4Н2.В3 (фиг.45). Подобным образом, ингибирование связывания PD-L1 с PD-1 было сравнимым для обоих антител (фиг.46). Таким образом, химерное крыса-мышь анти-mPD-1-антитело 4Н2 использовали для испытания терапевтической эффективности анти-PD-1-антитела в комбинации с анти-CTLA-4-антителом.
Пример 13
Эффективность in vivo комбинированной терапии (анти-CTLA-4 и анти-PD-1-антител) на установлении и росте опухоли
Раковые клетки ободочной кишки MC38 (PD-L1-) (доступные от доктора Dr. N. Restifo, National Cancer Institute, Bethesda, MD; or Jeffrey Schlom, National Institutes of Health, Bethesda, MD) имплантировали мышам C57BL/6 (2×106 клеток/мышь). В день 0 (т.е. в день, когда клетки МС38 имплантировали мышам), каждую из четырех групп из 10 мышей инъецировали внутрибрюшинно (IP) одним из следующих вариантов: (1) IgG мыши (контроль), (2) моноклональное анти-CTLA-4-антитело 9D9 (мышиное анти-mCTLA-4-антитело, полученное от J. Allison, Memorial Sloan-Kettering Cancer Center, New York, NY), (3) моноклональное анти-PD-1-антитело 4H2 (химерное антитело, в котором крысиное анти-mPD-1-антитело было модифицировано Fc-областью мыши, как описано в примере 6), или (4) анти-CTLA-4-антитело 9D9 и анти-PD-1-антитело 4H2. Затем инъекции антител вводили дополнительно в дни 3, 6 и 10. Лечение только одним антителом проводили при дозе 10 мг/кг, и комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела проводили при 5 мг/кг каждого антитела (т.е. 10 мг/кг антител в целом). С использованием электронного штангенциркуля опухоли измеряли в трех измерениях (высота×ширина×длина) и рассчитывали объем опухоли. Мышей подвергали эвтаназии, когда опухоли достигали указанной конечной точки опухоли. Полученные результаты показаны в таблице 5 и на фиг.21.
Процент не имеющих опухолей мышей после лечения анти-PD-1-антителами и/или анти-CTLA-4-антителами
Восемь мышей в группе IgG достигали конечной точки опухоли приблизительно в день 30, а две мыши (86066 и 87260) в группе IgG имели изъязвленные опухоли (фиг.21A). В группе только с анти-CTLA-4-антителом, семь мышей достигали конечной точки опухоли приблизительно в день 60, одна мышь имела изъязвленную опухоль (84952), одна мышь имела опухоль с объемом, меньшим, чем 1500 мм3 (85246), и одна мышь не имела опухоли (86057) (фиг.21B). В группе только с анти-PD-1-антителом, шесть мышей достигали конечной точки опухоли приблизительно в день 60, одна мышь имела изъязвленную опухоль (86055), и три мыши не содержали опухолей (84955, 85239 и 86750) (фиг.21C). В группе с комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела, четыре мыши достигали конечной точки опухоли приблизительно в день 40, и шесть мышей не содержали опухолей (84596, 85240, 86056, 86071, 86082 и 86761) (фиг.21D).
На фиг.22 показано, что средний объем опухоли, измеренный в день 21, составлял приблизительно 2955 мм3 для контрольной IgG-группы; приблизительно 655 мм3 для группы только с анти-CTLA-4-антителом, приблизительно 510 мм3 для группы только с анти-PD-1-антителом, и приблизительно 280 мм3 для группы с комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела. На фиг.23 показано, что медиана объема опухоли, измеренного в день 21 была приблизительно на 2715 мм3 для группы IgG; приблизительно 625 мм3 для группы только с анти-CTLA-4-антителом; приблизительно 525 мм3 для группы только с анти-PD-1-антителом; и приблизительно 10 мм3 для группы с комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела (и снижалась до 0 мм3 в день 32).
Данное исследование показывает, что на мышиной модели опухоли лечение одним анти-CTLA-4-антителом и лечение одним анти-PD-1-антителом имеет умеренное действие на рост опухоли, и что комбинированное лечение анти-CTLA-4-антителом и анти-PD-1-антителом имеет значительно большее действие на рост опухоли. Интересно отметить, что комбинированное лечение анти-CTLA-4-антителом и анти-PD-1-антителом имеет более существенное действие на рост опухоли при дозе 5 мг/кг каждого антитела в сравнении с действием каждого антитела по отдельности при введении каждого в более высокой дозе 10 мг/кг.
Пример 14
Эффективность in vivo комбинированной терапии (анти-CTLA-4-антитела и анти-PD-1-антитела) на рост установленной опухоли
Раковые клетки MC38 ободочной кишки (PD-L1-) имплантировали мышам C57BL/6 (2×106 клеток/мышь) в течение времени, достаточного (приблизительно 6-7 дней) для образования опухолей. В день 6 после имплантации (день -1) выполняли измерения опухоли и мышей делили случайным образом на основе среднего объема опухоли (приблизительно 250 мм3) на 11 групп для последующей терапии антителами. В день 0 (т.е. спустя одну неделю после имплантации клеток MC38) мышей инъецировали IP (1) IgG мыши (контроль), (2) моноклональным анти-CTLA-4-антителом 9D9, (3) моноклональным анти-PD-1-антителом 4H2 или (4) анти-CTLA-4-антителом 9D9 и моноклональным анти-PD-1-антителом 4H2 при концентрации 10 мг/кг мыши. Инъекции антител вводили также в дни 3, 6 и 10. Используемые композиции моноклональных антител имели низкие уровни эндотоксина и по существу не агрегировали. С использованием электронного штангенциркуля опухоли измеряли в трех измерениях (высота×ширина×длина) и рассчитывали объем опухоли. Измерения опухоли выполняли в день 0 (опухоли в начале лечения имели объем приблизительно 125 мм3), и в дни 3, 6, 10, 13, 17 и 20 после инъекции антител. Мышей подвергали эвтаназии, когда опухоли достигали конечной точки (1500 мм3) или когда мыши обнаруживали большую, чем 15%, потерю массы.
Все 11 мышей в группе IgG достигали конечной точки опухоли приблизительно в день 17 (фиг.24А). В группе только с анти-CTLA-4-антителом семь из 11 мышей достигали конечной точки опухоли приблизительно в день 13 и две мыши не имели опухолей (фиг.24С). В группе с комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела одна мышь достигала конечной точки опухоли приблизительно в день 17, одна мышь достигала конечной точки опухоли приблизительно в день 45 и девять мышей не содержали опухоли в день 45 (фиг.24D).
На фиг.25 показано, что средний объем опухоли, измеренный в день 10, составлял приблизительно 1485 мм3 для контрольной IgG-группы; приблизительно 1010 мм3 для группы только с анти-CTLA-4-антител, приблизительно 695 мм3 для группы только с анти-PD-1-антителом, и приблизительно 80 мм3 для группы с комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела. На фиг.26 показано, что медиана объема опухоли, измеренного в день 10 была приблизительно на 1365 мм3 для группы IgG; приблизительно 1060 мм3 для группы только с анти-CTLA-4-антителом; приблизительно 480 мм3 для группы только с анти-PD-1-антителом; и приблизительно 15 мм3 для группы с комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела (которая снижалась до 0 мм3 в день 17).
Данное исследование показывает, что на мышиной модели лечение комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела имеет значительно большее действие на рост опухолей, чем любое антитело по отдельности, даже когда опухоль уже хорошо образовалась.
Пример 15
Титрование дозы комбинированной терапии (анти-CTLA-4-антитела и анти-PD-1-антитела) на рост установленной опухоли
Раковые клетки MC38 ободочной кишки (PD-L1-) имплантировали мышам C57BL/6 (2×106 клеток/мышь) в течение времени, достаточного (приблизительно 6-7 дней) для образования опухолей, как описано в примере 3. Группы из 10 мышей инъецировали IP в дни 0, 3, 6 и 10 следующим образом: группа (A) IgG мыши (контроль, 20 мг/кг), группа (B) моноклональное анти-PD-1-антитело 4H2 (10 мг/кг) и IgG мыши (10 мг/кг), группа (C) моноклональное анти-CTLA-4-антитело 9D9 (10 мг/кг) и IgG мыши (10 мг/кг), группа (D) моноклональное анти-CTLA-4-антитело 9D9 (10 мг/кг) и моноклональное анти-PD-1-антитело 4H2 (10 мг/кг), группа (E) моноклональное анти-CTLA-4-антитело 9D9 (3 мг/кг) и моноклональное анти-PD-1-антитело 4H2 (3 мг/кг), или группа (F) моноклональное анти-CTLA-4-антитело 9D9 (1 мг/кг) и моноклональное анти-PD-1-антитело 4H2 (1 мг/кг). С использованием электронного штангенциркуля опухоли измеряли в трех измерениях (высота×ширина×длина) и рассчитывали объем опухоли. Измерения опухоли выполняли в начале лечения (т.е. в день 0 опухоли имели средний объем приблизительно 90 мм3) и в дни 3, 6, 10, 13, 17 и 20 после лечения антителами. Мышей подвергали эвтаназии, когда опухоли достигали конечной точки (конкретного объема опухоли, например, 1500 мм3, и/или когда мыши обнаруживали большую, чем приблизительно 15% потерю массы).
На фиг.27A показано, что все 10 контрольных мышей достигали конечной точки опухоли. На фиг.27B показано, что группа, получавшая 10 мг/кг анти-PD-1-антитела (группа B), имела 6 мышей, которые достигали конечной точки опухоли, и 4 мыши с опухолями, имеющими объем приблизительно 750 мм3 или менее. На фиг.27C показано, что группа, получавшая 10 мг/кг анти-CTLA-4-антитела (группа C), имела 3 мыши, которые достигали конечной точки опухоли, и 7 мышей с опухолями, имеющими объем приблизительно 1000 мм3 или менее. На фиг.27D показано, что группа, получавшая комбинацию 10 мг/кг анти-PD-1-антитела с 10 мг/кг анти-CTLA-4-антитела (группа D) имела 2 мыши с опухолями, имеющими объем приблизительно 1000 мм3 или менее, и 8 мышей, которые не имели опухолей. На фиг.27E показано, что группа, получавшая комбинацию 3 мг/кг анти-PD-1-антитела с 3 мг/кг анти-CTLA-4-антитела (группа E), имела одну мышь, которая достигла конечной точки опухоли, 7 мышей с опухолями, имеющими объем приблизительно 500 мм3 или менее, и 2 мыши, которые не имели опухолей. На фиг.27F показано, что группа, получавшая комбинацию 1 мг/кг анти-PD-1-антитела с 1 мг/кг анти-CTLA-4-антитела (группа F), имела 4 мыши, которые достигли конечной точки опухоли, 5 мышей с опухолями, имеющими объем приблизительно 1100 мм3 или менее, и одну мышь, которая не имела опухоли.
На фиг.27G и 27H показаны объемы опухолей у мышей получавших последовательно анти-PD-1-антитело первым и анти-CTLA-4-антитело вторым, и наоборот. Мыши фиг.27G сначала получали 10 мг/кг анти-CTLA-4-антитела в каждый из дней 0 и 3, и затем получали 10 мг/кг анти-PD-1-антитела в каждый из дней 6 и 10. Мыши фиг.27H сначала получали 10 мг/кг анти-PD-1-антитела в каждый из дней 0 и 3, и затем получали 10 мг/кг анти-CTLA-4-антитела в каждый из дней 6 и 10. Что касается группы G, в день 27 8 мышей достигали конечной точки опухоли, одна мышь имела очень маленькую опухоль (которая после значительной задержки в конечном счете исчезала) и одна мышь не имела опухоли. Что касается группы Н, в день 27 8 мышей достигали конечной точки опухоли и 2 мыши не имели опухоли.
На фиг.28 показано, что средний объем опухоли, измеренный в день 10, составлял приблизительно 1250 мм3 для группы IgG-контроля; приблизительно 470 мм3 для анти-PD-1-антитела с IgG-контролем; приблизительно 290 мм3 для анти-CTLA-4-антитела с IgG-контролем (измеренный в день 6); приблизительно 40 мм3 для группы комбинации анти-CTLA-4-антитела (10 мг/кг) и анти-PD-1-антитела (10 мг/кг); приблизительно 165 мм3 для группы комбинации анти-CTLA-4-антитела (3 мг/кг) и анти-PD-1-антитела (3 мг/кг); и приблизительно 400 мм3 для группы комбинации анти-CTLA-4-антитела (1 мг/кг) и анти-PD-1-антитела (1 мг/кг). На фиг.29 показано, что медиана объема опухоли, измеренного в день 13, была приблизительно 1680 мм3 для группы IgG-контроля; приблизительно 400 мм3 для анти-PD-1-антитела с IgG-контролем; приблизительно 660 мм3 для анти-CTLA-4-антитела с IgG-контролем; 0 мм3 для группы комбинации анти-CTLA-4-антитела (10 мг/кг) и анти-PD-1-антитела (10 мг/кг); приблизительно 90 мм3 для группы комбинации анти-CTLA-4-антитела (3 мг/кг) и анти-PD-1-антитела (3 мг/кг); и приблизительно 650 мм3 для группы комбинации анти-CTLA-4-антитела (1 мг/кг) и анти-PD-1-антитела (1 мг/кг). Для комбинированного лечения анти-PD-1-антителом с анти-CTLA-4-антителом количество мышей на группу, которые не имели опухолей в день 27 данного исследования, было равно 8/10 (10 мг/кг), 2/10 (3 мг/кг) и 1/10 (1 мг/кг) (данные не показаны).
Данное исследование показывает, что на мышиной модели опухоли лечение комбинацией анти-CTLA-4-антитела и анти-PD-1-антитела действует зависимым от дозы образом и имеет значимо большее действие на рост опухоли, чем оба антитела по отдельности, даже при более низкой дозе и даже, когда опухоль уже хорошо развилась. Кроме того, антитела могут вводиться последовательно (анти-CTLA-4-антитело первым и анти-PD-1-антитело вторым или наоборот), и эта комбинация является все еще превосходящей монотерапии с использованием антител.
Пример 16
Эффективность in vivo комбинированной терапии (анти-CTLA-4-антитела и анти-PD-1-антитела) в отношении возникновения (установления) и роста фибросаркомы
Клетки SA1/N фибросаркомы (PD-L1-) (Leach et al. (1996) Science 271:1734-1736) имплантировали подкожно в мышей A/J (2×106 клеток/мышь) в день 0. В дни 1, 4, 7 и 11 после имплантации, мышей инъецировали IP следующим образом: группа (A) только PBS (называемый “носителем”); группа (B) IgG мыши (контроль, 10 мг/кг на мышь), группа (C) моноклональное анти-PD-1-антитело 4H2 (10 мг/кг на мышь), группа (D) моноклональное анти-CTLA-4-антитело 9D9 (10 мг/кг или 0,2 мг/кг на мышь) и группа (E) моноклональное анти-PD-1-антитело 4H2 (10 мг/кг на мышь) в комбинации с моноклональным анти-CTLA-4-антителом 9D9 (0,2 мг/кг на мышь). Данное исследование длилось 41 день и измерения опухолей выполняли в разные дни по ходу всего исследования (см. фиг.29). Объем опухоли рассчитывали измерением опухолей в трех измерениях (высота×ширина×длина) с использованием электронного штангенциркуля. Мышей подвергали эвтаназии, когда опухоли достигали указанной конечной точки опухоли - объема 1500 мм3 и/или мыши развивали изъязвленные опухоли.
На фиг.30A и 30B показано, что 19 из 20 контрольных (9/10 в группе A и 10/10 в группе B) мышей либо достигали конечной точки опухоли либо развивали изъязвленные опухоли. На фиг.30C показано, что группа, получавшая 10 мг/кг анти-PD-1-антитела (группа C), имела 6 мышей, которые достигали конечной точки опухоли (2 с объемом, большим, чем 1500 мм3, и 4 с изъязвленной опухолью), и 4 мыши, которые не имели опухоли. На фиг.30D показано, что группа, получавшая 10 мг/кг анти-CTLA-4-антитела (группа D), имела 5 мышей, которые достигали конечной точки опухоли (2 с объемом, большим, чем 1500 мм3, и 3 с изъязвленной опухолью), одну мышь с небольшой опухолью (объем приблизительно 70 мм3) и 4 мыши, которые не имели опухоли. На фиг.30E показано, что группа, получавшая 0,2 мг/кг анти-CTLA-4-антитела (группа E), имела 10 мышей, которые достигали конечной точки опухоли (6 с объемом, большим, чем 1500 мм3, и 4 с изъязвленной опухолью). На фиг.30F показано, что группа, получавшая комбинацию 10 мг/кг анти-PD-1-антитела с 0,2 мг/кг анти-CTLA-4-антитела (группа F), имела 2 мышей, которые достигали конечной точки опухоли (одну с объемом опухоли, большим, чем 1500 мм3, и одну с изъязвленной опухолью и 8 мышей, которые не имели опухоли.
На фиг.31 и 32 показаны средний объем опухоли и медиана объема опухоли, которые развивались у получавших лечение и не получавших лечения мышей по ходу всего исследования. Ингибирование роста опухолей у мышей, получавших данные антитела, в сравнении с мышами, получавшими мышиный IgG в качестве контрольного антитела, суммировано в таблице 6.
† Группы, как они определены на фиг.30. A=носитель (PBS); B=IgG мыши; C=анти-PD-1-антитело, 10 мг/кг; D=анти-CTLA-4-антитело, 10 мг/кг; E=анти-CTLA-4-антитело, 0,2 мг/кг и F=анти-PD-1-антитело, 10 мг/кг с анти-CTLA-4-антителом, 0,2 мг/кг
Полученные результаты дополнительно показывают, что комбинированная терапия, включающая анти-PD-1 и анти-CTLA-4-антитела, является значительно более эффективной, чем лечение любым из антител по отдельности. Действительно, комбинация является еще более эффективной, чем терапии только с одним антителом, даже в том случае, когда комбинированная терапия включает субтерапевтическую дозу анти-CTLA-4-антитела. Приведенные данные показывают также, что неожиданно присутствие или отсутствие PD-L1 на опухоли может не влиять на эффективность лечения комбинацией антител, хотя присутствие PD-L1 может влиять на эффект монотерапии с использованием антител таким образом, что экспрессия PD-L1 на опухоли может приводить к ингибированию противоопухолевых Т-клеточных реакций (см. фиг.40).
Пример 17
Эффективность in vivo и титрование дозы комбинированной терапии (анти-CTLA-4-антитела и анти-PD-1-антитела) на рост фибросаркомы PD-L1-
Клетки SA1/N фибросаркомы (PD-L1-) имплантировали подкожно мышам A/J (2×106 клеток/мышь) в день 0 в течение времени, достаточного (приблизительно 7 дней) для установления опухоли. В дни 7, 10, 13 и 16 после имплантации, десять групп из 8 мышей, имеющих средний объем опухоли 110 мм3, инъецировали IP следующим образом: группа (A) только PBS (называемый “носителем”); группа (B) IgG мыши (контроль, 10 мг/кг на мышь), группа (C) моноклональное анти-CTLA-4-антитело 9D9 (0,25 мг/кг на мышь), группа (D) моноклональное анти-CTLA-4-антитело 9D9 (0,5 мг/кг или 0,2 мг/кг на мышь), группа (E) моноклональное анти-CTLA-4-антитело 9D9 (5 мг/кг на мышь); группа (F) моноклональное анти-PD-1-антитело 4Н2 (3 мг/кг на мышь); группа (G) моноклональное анти-PD-1-антитело 4Н2 (10 мг/кг на мышь); группа (Н) моноклональное анти-PD-1-антитело 4Н2 (10 мг/кг на мышь); группа (I) моноклональное антитело 4Н2 (10 мг/кг на мышь) в комбинации с моноклональным анти-CTLA-4-антителом 9D9 (0,6 мг/кг на мышь); и группа (J) моноклональное анти-PD-1-антитело 4Н2 (3 мг/кг на мышь) в комбинации с моноклональным анти-CTLA-4-антителом 9D9 (0,5 мг/кг на мышь).
В дни 10, 13, 16 и 19 после имплантации две группы из 6 мышей, имеющих средний объем опухоли 255 мм3, инъецировали IP следующим образом: группа (К) IgG мыши (контроль, 10 мг/кг на мышь) и группа (L) моноклональное анти-PD-1-антитело 4H2 (10 мг/кг на мышь) в комбинации с моноклональным анти-CTLA-4-антителом 9D9 (1 мг/кг на мышь). Данное исследование длилось 51 день и измерения опухолей выполняли в разные дни по ходу всего исследования (см. фиг.33-38). Объем опухоли рассчитывали измерением опухолей в трех измерениях (высота×ширина×длина) с использованием электронного штангенциркуля. Мышей подвергали эвтаназии, когда опухоли достигали указанной конечной точки опухоли - объема 1500 мм3 и/или развивали изъязвленную опухоль.
На фиг.33 показана реакция на лечение иммуностимулирующими антителами у мышей с опухолями, имеющими начальный объем приблизительно 110 мм3 (т.е. в момент обработки первым антителом). На фиг.33А и 33В показано, что все 16 контрольных мышей (группы А и В) достигали конечной точки опухоли (15 с объемом опухоли, большим, чем 1500 мм3 и 1 с изъязвленной опухолью). На фиг.33С-33Е показано, что несущие опухоль мыши отвечают на лечение анти-CTLA-4-антителом зависимым от дозы образом (например, группа С, получающая 0,25 мг/кг, имела 7/8 мышей, достигших конечной точки опухоли, и одну мышь с объемом опухоли, меньшим, чем 200 мм3, тогда как группа Е, получающая 5 мг/кг, имела 6/8 мышей, достигших конечной точки опухоли, и две мыши не имели опухоли). На фиг.33F и 33G показано, что мыши отвечали приблизительно одинаково независимо от дозы анти-PD-1-антитела (группа F получала 3 мг/кг и группа G получала 10 мг/кг). В противоположность этому, мыши, получавшие комбинированное лечение 10 или 3 мг/кг анти-PD-1-антитела с 0,25 или 0,5 мг/кг анти-CTLA-4-антитела (группы Н, I и J), показали значительное уменьшение роста опухоли. Например, на фиг.33J показано, что группа, получавшая комбинацию 3 мг/кг анти-PD-1-антитела с 0,5 мг/кг анти-CTLA-4-антитела (группа J), имела 2 мыши, которые имели изъязвленные опухоли, 2 мыши с объемом опухоли, меньшим, чем 500 мм3, и 4 мыши, которые не имели опухоли. Это неожиданное синергическое действие анти-PD-1-антитела, комбинированного с анти-CTLA-4-антителом, вместе с неожиданной эффективностью субтерапевтических уровней анти-CTLA-4-антитела в данной комбинации, показаны на фиг.32 (средний объем опухоли) и 35 (медиана объема опухоли).
На фиг.36 показана реакция на лечение иммуностимулирующими антителами у мышей с более крупными опухолями, имеющими начальный объем приблизительно 250 мм3 (т.е. в момент обработки первым антителом). На фиг.36А показано, что все 6 контрольных мышей (группа К) достигали конечной точки опухоли (4 с опухолями, большими, чем 1500 мм3, и 2 с изъязвленной опухолью). На фиг.36В показано, что группа, получавшая комбинацию 10 мг/кг анти-PD-1-антитела с 1 мг/кг анти-CTLA-4-антитела (группа L), имела одну мышь с изъязвленной опухолью, 4 мыши с объемом опухоли, большим, чем 1500 мм3, и одну мышь, которая не имела опухоли. Средние объемы опухоли и медианы объемов опухолей показаны на фиг.37 и 38.
Ингибирование роста опухоли у мышей, получавших данные антитела, в сравнении с мышами, получавшими IgG мыши в качестве контрольного антитела, суммировано в таблице 7 и на фиг.39.
(день 51)
Вместе приведенные данные показывают, что комбинированная терапия, включающая анти-PD-1 и анти-CTLA-4-антитела, является значительно более эффективной, чем лечение каждым антителом по отдельности. Кроме того, неожиданно доза каждого антитела может быть уменьшена без влияния на синергическую эффективность данной комбинации иммуностимулирующих терапевтических антител. По-видимому, комбинированная терапия является более эффективной даже в том случае, когда масса опухоли является более зрелой (т.е. большей).
Пример 18
Опухолевый иммунитет у мышей после лечения анти-PD-1-антителами и повторной иммунизации клетками фибросаркомы (PD-L1-)
Мышей, которые выживали без опухолей при иммунизации опухолевыми клетками и лечении анти-PD-1-антителом (т.е. обработки, сходной с исследованиями эффективности, описанными в примерах 5 и 6), затем повторно иммунизировали опухолевыми клетками для исследования иммунитета к образованию опухоли после такого лечения. Кратко, в начальной иммунизации клетки фибросаркомы SA1/N (PD-L1-) имплантировали подкожно мышам A/J (1×106 клеток/мышь) в день 0. В дни 1, 4, 7, 10, 14, 17 и 20 после имплантации группы мышей инъецировали IP либо IgG мыши (контроль, 10 мг/кг на мышь), либо одной из различных доз анти-PD-1-антитела 4H2 (30, 10, 3, 1 и 0,3 мг/кг на мышь). Образование и объем опухолей подвергали мониторингу с использованием прецизионного электронного штангенциркуля два раза в неделю до завершения данного исследования. Группы из 8 мышей не имели опухолей после лечения анти-PD-1-антителами (4, которых лечили с использованием 30 мг/кг, 2 с 3 мг/кг, одна с 1 мг/кг и одна с 0,3 мг/кг).
Восемь получавших лечение не содержащих опухоли мышей A/J повторно иммунизировали подкожной имплантацией 1×106 клеток фибросаркомы SA1/N на мышь. В качестве контроля девяти необработанным мышам подкожно имплантировали с использованием 1×106 клеток фибросаркомы SA1/N на мышь. Образование и объем опухолей подвергали мониторингу с использованием прецизионного электронного штангенциркуля два раза в неделю до дня 62 после имплантации. Все девять необработанных мышей (контроль) достигали конечной точки опухоли в день 22 после имплантации клетками фибросаркомы. В противоположность этому, у восьми не содержащих опухоли мышей, повторно иммунизированных клетками фибросаркомы, не развивались опухоли до дня 62 после имплантации. На фиг.47 показан средний объем опухолей для необработанных и повторно обработанных мышей. Полученные результаты демонстрируют, что лечение иммуностимулирующим антителом, таким как анти-PD-1-антитело, обеспечивает прошедшего лечение субъекта иммунитетом в отношении последующего образования опухоли, даже в присутствии клеток, способных образовывать опухоль.
Пример 19
Опухолевый иммунитет у мышей после терапии единственным антителом (анти-PD-1-антителом) или комбинированной терапии (анти-CTLA-4-антителом и анти-PD-1-антителом), повторно иммунизированных клетками рака ободочной кишки PD-L1-
Мышей, которые выживали без опухолей при иммунизации опухолевыми клетками и лечении либо одним анти-PD-1-антителом, либо анти-PD-1-антителом, комбинированным с анти-CTLA-4-антителом (т.е. обработки, сходной с исследованиями эффективности, описанными в примерах 2-4), затем повторно иммунизировали опухолевыми клетками для исследования иммунитета к образованию опухоли после такого лечения. Кратко, в начальной иммунизации клетки, клетки МС38 рака ободочной кишки (PD-L1-) имплантировали мышам C57BL/6 (2×106 клеток/мышь) в день 0. В дни 0, 3, 6 и 10 после имплантации, группы мышей инъецировали IP с использованием одной из следующих обработок: (1) IgG мыши (контроль, 10 мг/кг на мышь), (2) моноклональное анти-PD-1-антитело 4H2 или (3) моноклональное анти-PD-1-антитело 4H2 в комбинации с анти-CTLA-4-антителом 9D9. Рост опухоли подвергали мониторингу с использованием прецизионного электронного штангенциркуля, как описано в примере 15. Группа из 11 мышей не имела опухолей после лечения анти-PD-1-антителом (всего 2) или комбинированного лечения анти-PD-1-антителом/анти-CTLA-4-антителом (всего 9).
11 обработанных, не содержащих опухоли мышей C57BL/6 повторно иммунизировали имплантацией 2×107 клеток MC38 рака ободочной кишки на мышь (т.е. дозы клеток, в 10 раз большей, чем при начальной иммунизации). В качестве контроля, семь необработанных мышей имплантировали с использованием 2×107 клеток MC38 рака ободочной кишки на мышь. Рост и объем опухоли подвергали мониторингу с использованием прецизионного электронного штангенциркуля по ходу всего эксперимента по повторной иммунизации (по меньшей мере 20 дней). На фиг.48 показано, что все семь необработанных (контрольных) мышей развивали опухоли и достигали конечной точки опухоли в день 18 после имплантации клеток рака ободочной кишки. В противоположность этому, все 11 не имеющих опухоли мышей, повторно иммунизированных клетками рака ободочной кишки, не развивали опухоли до дня 18 после имплантации. На фиг.49 показан средний объем опухоли для необработанных и повторно иммунизированных мышей. Приведенные данные показывают, что, подобно монотерапии с использованием антител, комбинированная терапия антителами, приводящая к блокаде PD-1 и CTLA-44, продуцирует стойкий иммунитет к рецидиву опухоли.
Пример 20
Эффективность in vivo комбинированной терапии (с анти-CTLA-4- и анти-PD-1-антителами) в отношении роста установленной опухоли
Клетки CT26 рака ободочной кишки имплантировали мышам Balb/c (2×106 клеток/мышь) в течение времени, достаточного (приблизительно 10 дней) для образования опухолей. В день 10 после имплантации, выполняли измерения опухолей и мышей рандомизировали на основе среднего объема опухоли (приблизительно 250 мм3) на 5 групп для последующей терапии с использованием антител. В день 1 (т.е. спустя 10 дней после имплантации клеток CT26), мышей инъецировали IP (1) IgG мыши (контроль), (2) моноклональным анти-CTLA-4-антителом 9D9, (3) моноклональным анти-PD-1-антителом 4H2 или (4) моноклональным анти-CTLA-4-антителом и 9D9 и моноклональным анти-PD-1-антителом 4H2, в концентрации 10 мг/кг на мышь. Инъекции антител вводили также в дни 3, 6 и 10. Использованные композиции антител имели низкие уровни эндотоксина и по существу не агрегировались. С использованием электронного штангенциркуля опухоли измеряли в трех измерениях (высота×ширина×длина) и рассчитывали объем опухоли. Измерения опухолей выполняли в день 0 (опухоли в начале лечения имели объем приблизительно 125 мм3) и в дни 3, 6, 10, 13, 17 и 20 после инъекции антител. Мышей подвергали эвтаназии, когда опухоли достигали указанной конечной точки (конкретного объема опухоли, например, 1500 мм3, и/или когда мыши обнаруживали большую, чем приблизительно 15% потерю массы). Полученные результаты показаны на фиг.50. Данное исследование показывает, что на мышиной модели опухоли лечение комбинацией CTLA-4-антитела и PD-1-антитела имеет значительно большее действие на рост опухоли, чем каждое антитело по отдельности, даже в том случае, когда опухоль уже хорошо установилась.
Пример 21
Действие анти-PD-1-антитела человека на функцию регуляторных Т-клеток
Регуляторные T-клетки являются лимфоцитами, которые подавляют иммунную реакцию. В данном примере регуляторные Т-клетки испытывали на их ингибирующую функцию в отношении пролиферации и секреции IFN-гамма CD4+CD25- Т-клетками в присутствии или в отсутствие моноклонального анти-PD-1-антитела.
Регуляторные Т-клетки очищали из PBMC с использованием набора для выделения регуляторных CD4+CD25+ Т-клеток (Miltenyi Biotec). Регуляторные T-клетки добавляли в реакцию смешанных лимфоцитов (см. выше), содержащую очищенные CD4+CD25- T-клетки и аллогенные дендритные клетки в соотношении 2:1 CD4+CD25- Т-клеток к регуляторным T-клеткам. Моноклональное анти-PD-1-антитело 5C4 добавляли при концентрации 10 мкг/мл. В качестве отрицательного контроля служил вариант без антитела или изотипически сходное контрольное антитело. Культуральные супернатанты собирали в день 5 для измерения цитокинов с использованием системы детектирования цитокинов Beadlyte (Upstate). Клетки метили 3H-тимидином, культивировали еще в течение 18 часов и анализировали на пролиферацию клеток. Результаты показаны на фиг.51А (пролиферация Т-клеток) и 51В (секреция IFN-гамма). Добавление моноклонального анти-PD-1-антитела человека 5C4 частично снимало ингибирование, вызываемое регуляторными Т-клетками (Treg-клетками), в отношении пролиферации и секреции IFN-гамма CD4+CD25- T-клетками, что свидетельствует о том, что анти-PD-1-антитела влияют на регуляторные Т-клетки.
Пример 22
Действие анти-PD-1-антитела человека на активацию Т-клеток
В данном примере исследовали действие блокады PD-1-пути анти-PD-1-антителом 5С4 на активацию Т-клеток. Очищенные CD4+ Т-клетки человека (набор Dynal для очистки CD4 Т-клеток) активировали 1 мкг/мл растворимого анти-CD3-антителом (BD) в присутствии аутологичных моноцитов или полученных из моноцитов дендритных клеток (DC). Моноциты очищали с использованием набора для очистки CD4 моноцитов Miltenyi и DC генерировали in vitro после культивирования моноцитов с GM-CSF и IL-4 (PeproTech) в течение 7 дней. Спустя три дня активации в присутствии или в отсутствие разведенного анти-PD-1-антитела или постороннего изотопически сходного контрольного mAb, культуральные супернатанты собирали для анализа ELISA секреции IFN-гамма, добавляя тритированный тимидин во время последних 18 часов данного анализа для измерения пролиферации Т-клеток. Результаты, показанные на фиг.52A и 52B, демонстрируют, что блокада PD-1 анти-PD-1-антителом приводила к усиленной пролиферации Т-клеток и усиленной секреции IFN-гамма. Наблюдали также синергическое действие анти-PD-1-антитела и анти-CTLA-4-антитела на активацию Т-клеток (конкретно, на секрецию IFN-гамма) в присутствии моноцитов.
Пример 23
Оценка активности ADCC анти-PD-1-антитела
В данном примере выполняли анализ антителозависимой клеточной цитотоксичности (ADCC) для оценки, может ли анти-PD-1-антитело индуцировать ADCC в отношении клеток-мишеней. Две версии 5С4, одну с Fc-областью IgG1 человека (5С4-IgG1) и другую с Fc-областью IgG4 человека (5С4-IgG4), испытывали в данном анализе. Набор Delfia клеточной цитотоксичности из Perkin Elmer использовали для данного анализа. Кратко, очищенные CD4+ Т-клетки человека (набор для очистки CD4 Т-клеток Dynal) активировали связанным с планшетом анти-CD3-антителом (BD) для индукции экспрессии PD-1. Затем активированные CD4 Т-клетки-мишени метили реагентом BATDA. Меченые CD4 T-клетки добавляли в 96-луночный планшет с V-образным дном с последующим добавлением PBMC человека (отношение эффекторных клеток к клеткам-мишеням (Е/Т) 50:1) и указанного антитела. После инкубирования в течение 1 часа при 37°С планшет центрифугировали. Супернатант переносили в плоскодонный 96-луночный планшет и планшет считывали с использованием планшет-ридера RubyStar. Результаты показали, что 5С4-IgG4 не опосредовал ADCC на активированных CD4 Т-клетках, тогда как 5С4-IgG1 действительно опосредовал ADCC на активированных CD4 Т-клетках (фиг.53), что свидетельствовало о том, что ADCC-активность связана с Fc-областью анти-PD-1-антитела.
Пример 24
Оценка комплементзависимой цитотоксичности анти-PD-1-антитела
В данном примере испытывали комплементзависимую цитотоксичность (CDC) анти-PD-1-антитела. Две версии 5С4, одну с Fc-областью IgG1 человека (5С4-IgG1) и другую с Fc-областью IgG4 человека (5С4-IgG4), испытывали в данном анализе. Кратко, очищенные CD4+ Т-клетки человека (набор для очистки CD4 Т-клеток Dynal) активировали связанным с планшетом анти-CD3-антителом (BD) для индукции экспрессии PD-1. Серийные разведения анти-PD-1-антитела (5C4) и контрольных антител от 50 мкг/мл до 640 пг/мл испытывали на CDC в присутствии комплемента человека (Quidel-A113). Для измерения цитотоксичности использовали краситель аламаровый синий (Biosource International). Планшет считывали на флуоресцентном планшет-ридере (EX530 EM590). Количества жизнеспособных клеток являются пропорциональными единицам флуоресценции. Результаты показали, что ни 5C4-IgG1, ни 5C4-IgG4 не опосредовал CDC на активированных CD4 T-клетках, тогда как антитело положительного контроля (анти-HLA-ABC-антитело) опосредовало CDC (фиг.54).
Пример 25
Оценка экспрессии PD-1 на Т-клетках человека
В данном примере PBMC человека от различных доноров испытывали на экспрессию PD-1 на различных популяциях клеток при помощи FACS. В данном анализе использовали биотинилированное анти-PD-1-антитело, которое проявляло гораздо более высокую чувствительность, чем коммерчески доступное анти-PD-1-антитело при детектировании молекул PD-1 на поверхности клеток. Связанное антитело детектировали с использованием РЕ-конъюгированного стрептавидина. Проточные цитометрические анализы выполняли с использованием проточной цитометрии FACScan (Becton Dickinson) и программного обеспечения Flowjo (Tree Star). Экспрессию PD-1 детектировали на некоторых периферических Т-клетках человека, но не на В-клетках или моноцитах. Дополнительное испытание субпопуляций Т-клеток указывает на то, что PD-1 экспрессируется на CD4+ и CD8+ Т-клетках памяти и эффекторных Т-клетках, но отсутствуют на «необученных» CD4 или CD8 Т-клетках.
Настоящее изобретение не ограничивается объемом конкретных описанных в данном описании вариантов осуществления. Действительно, различные модификации настоящего изобретения, наряду с описанными выше, будут очевидными для специалистов в данной области из приведенного выше описания и сопутствующих фигур. Предполагается, что такие модификации входят в объем прилагаемой формулы изобретения. Таким образом, настоящее изобретение должно ограничиваться только прилагаемой формулой изобретения вместе с полным объемом эквивалентов, на которые дает право настоящая формула изобретения.























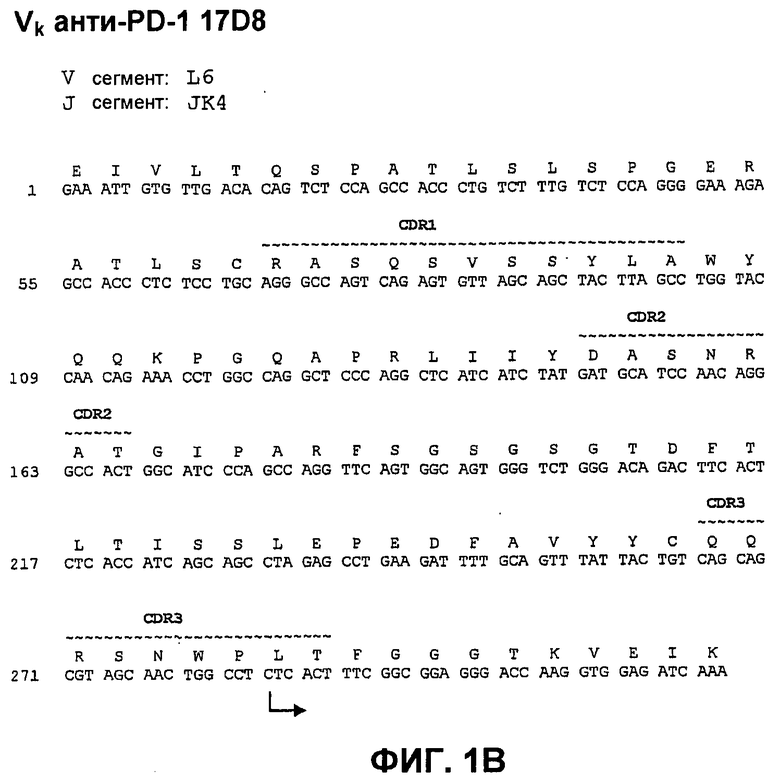
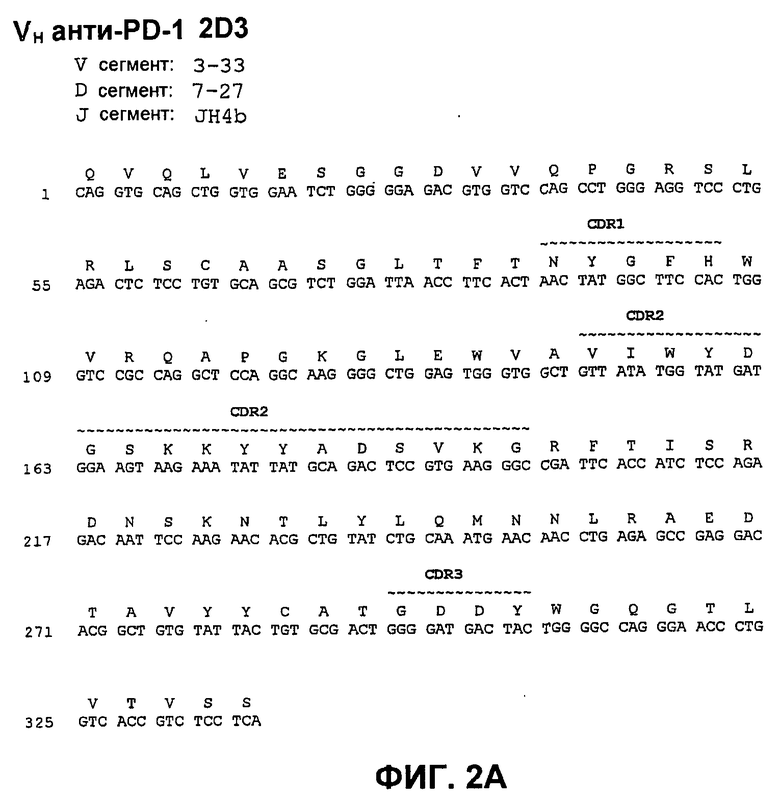

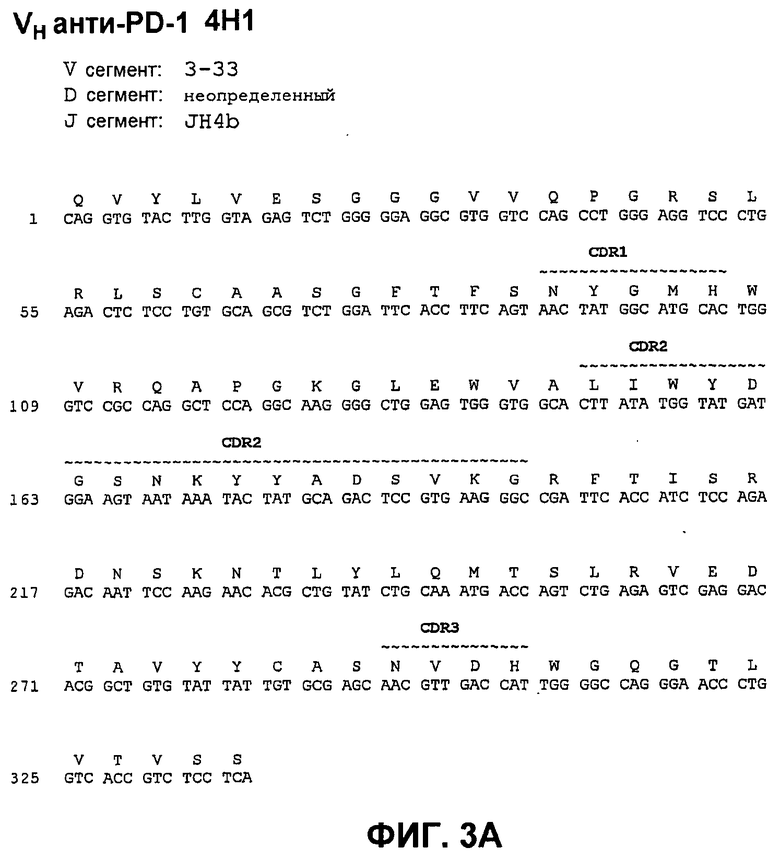
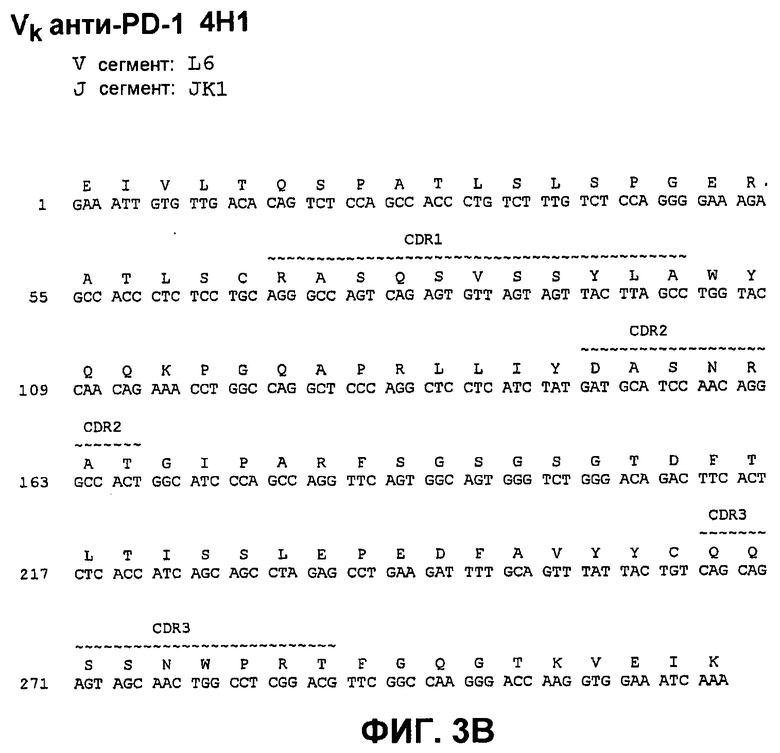
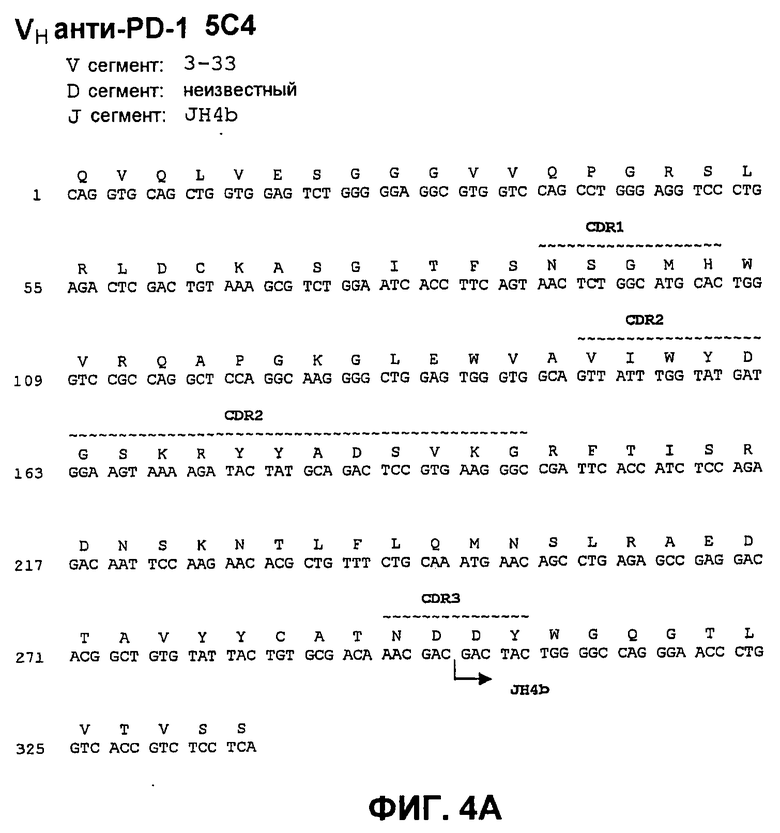


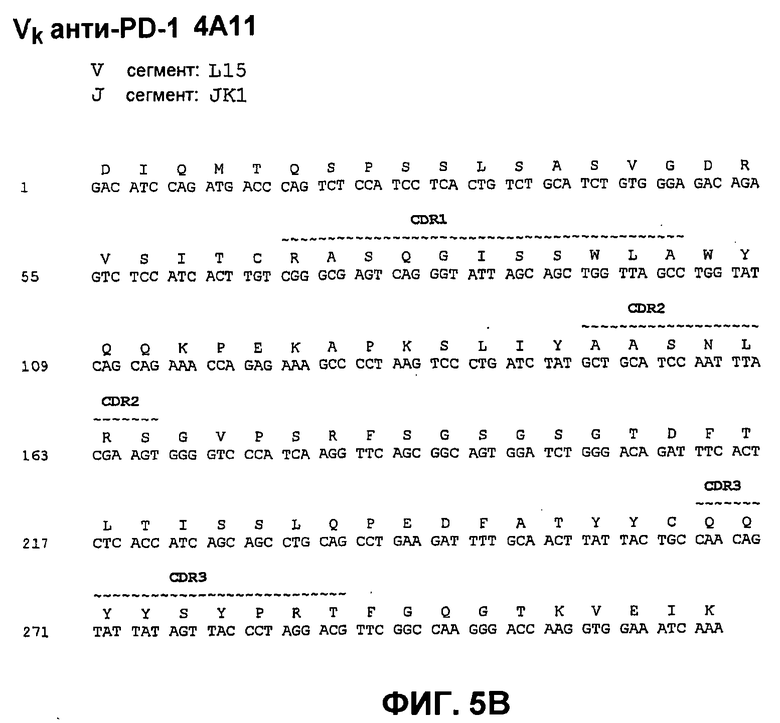




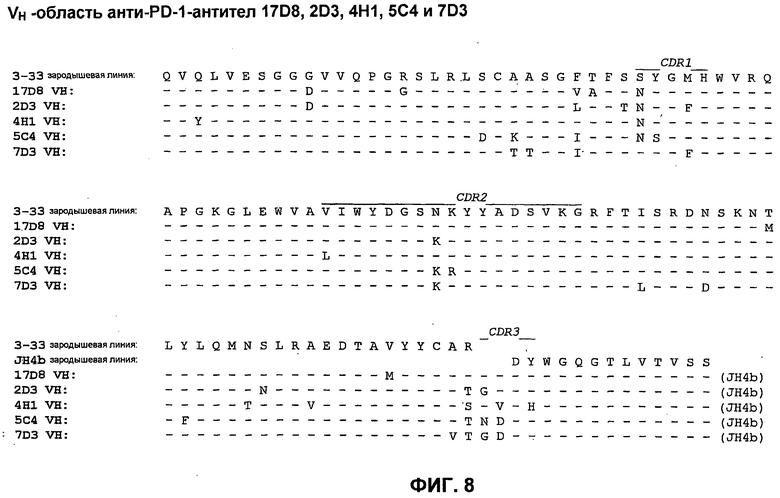
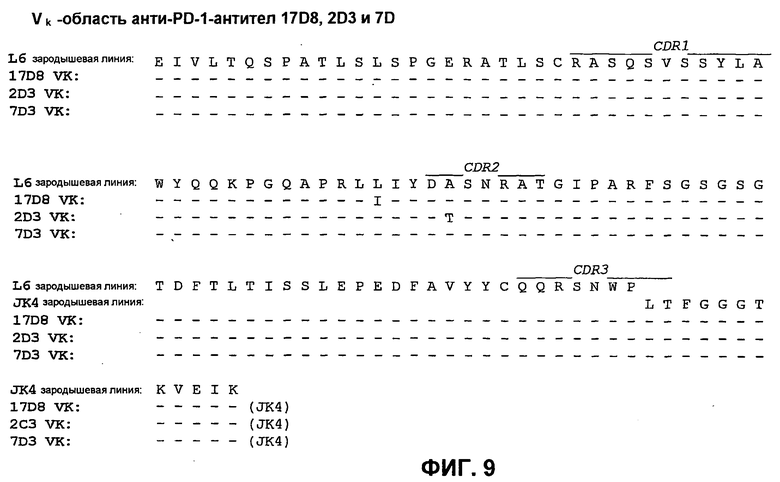
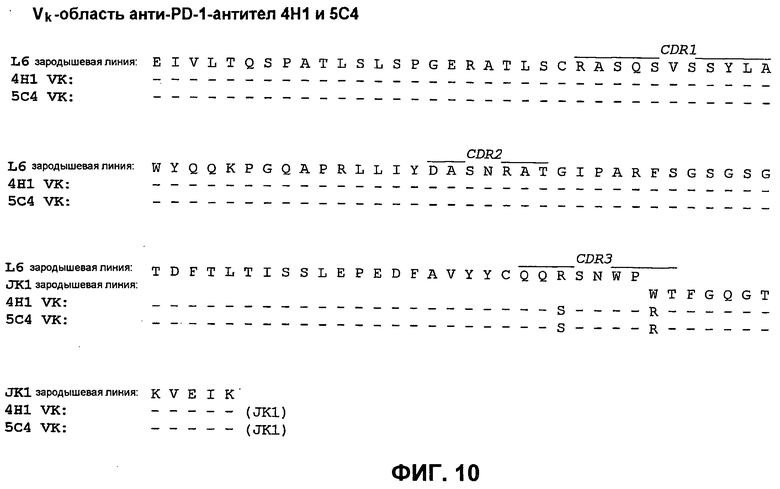
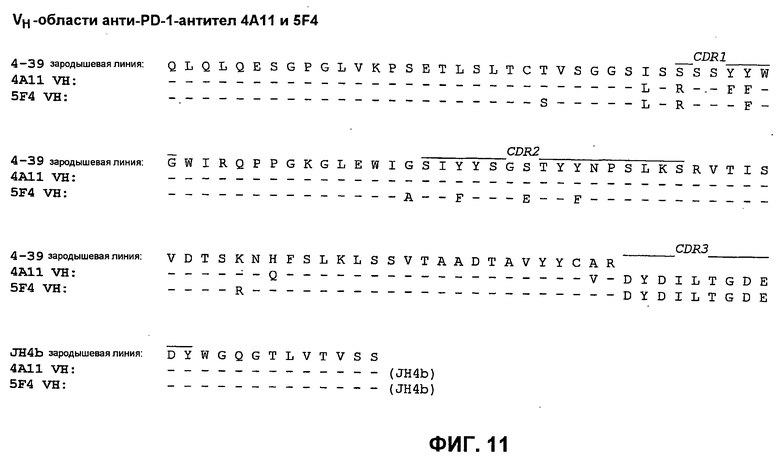
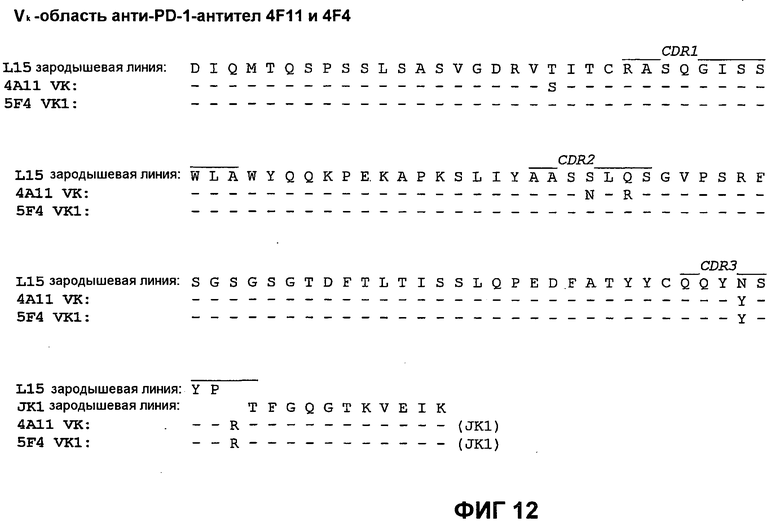
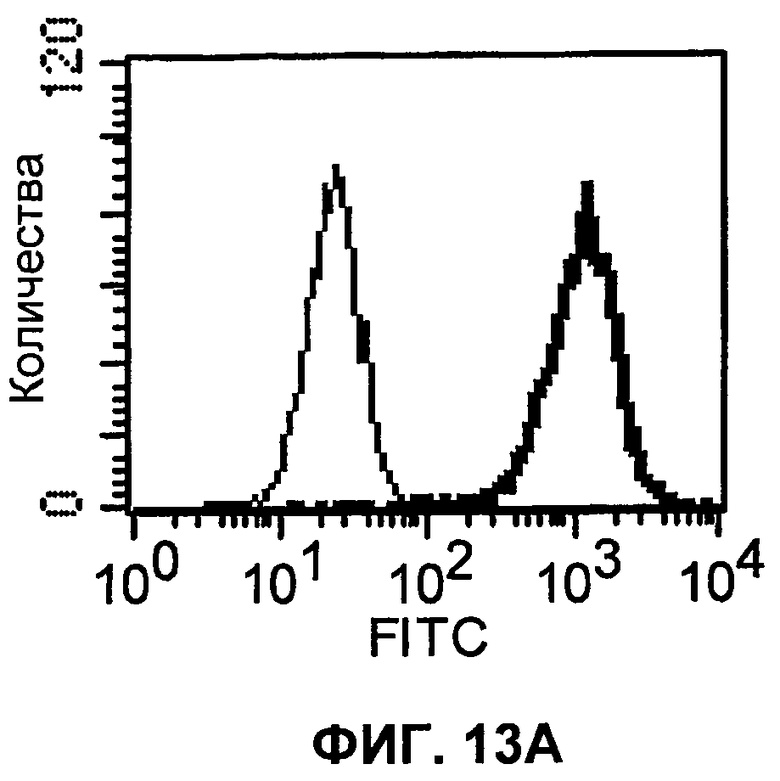
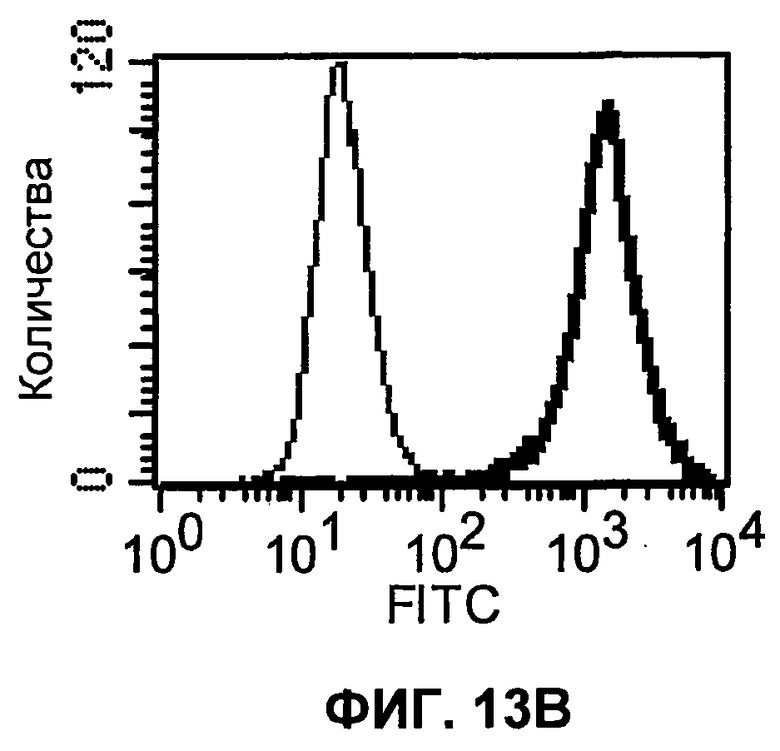
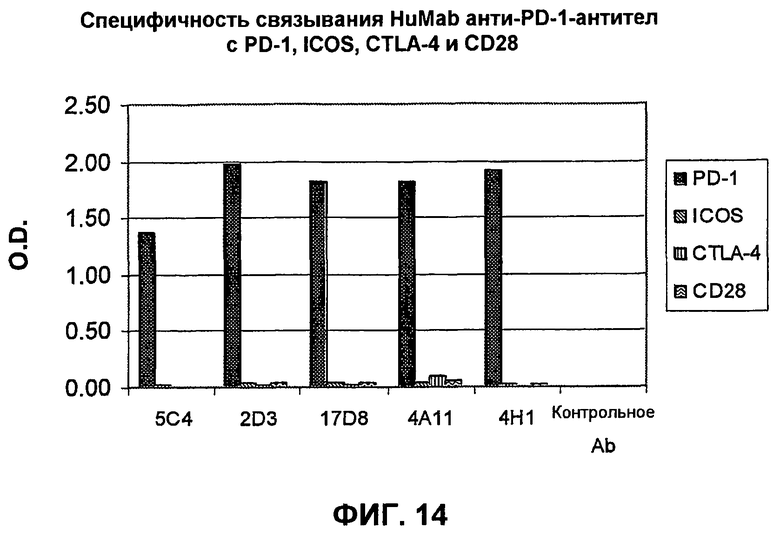
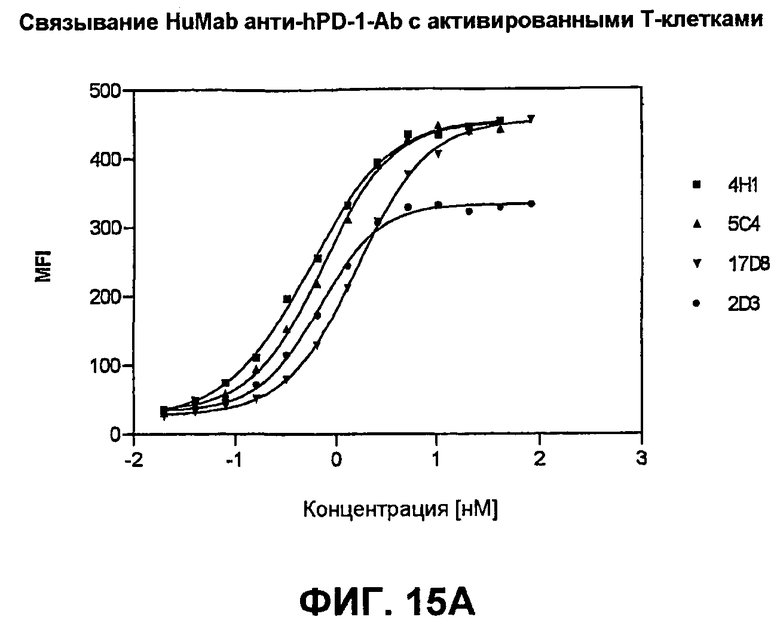
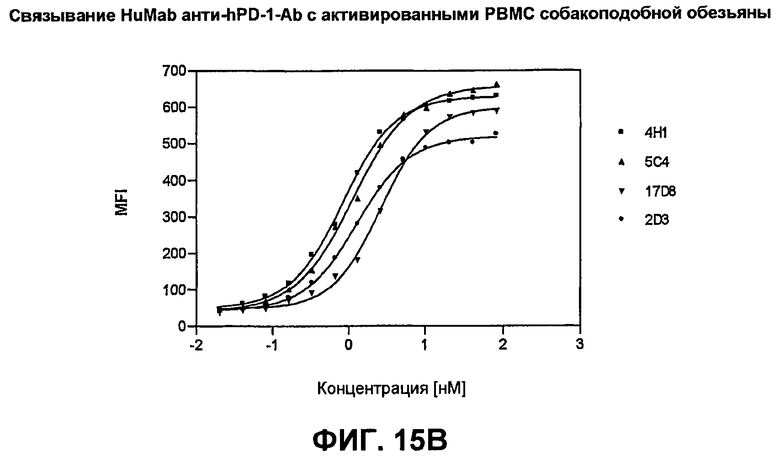
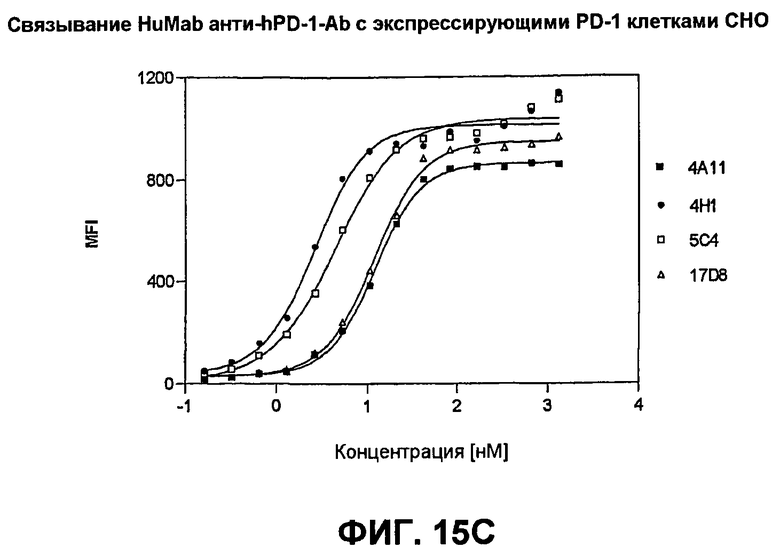
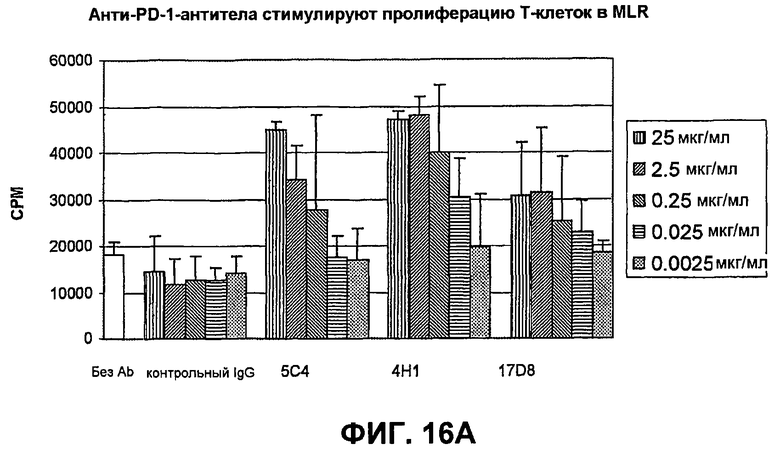
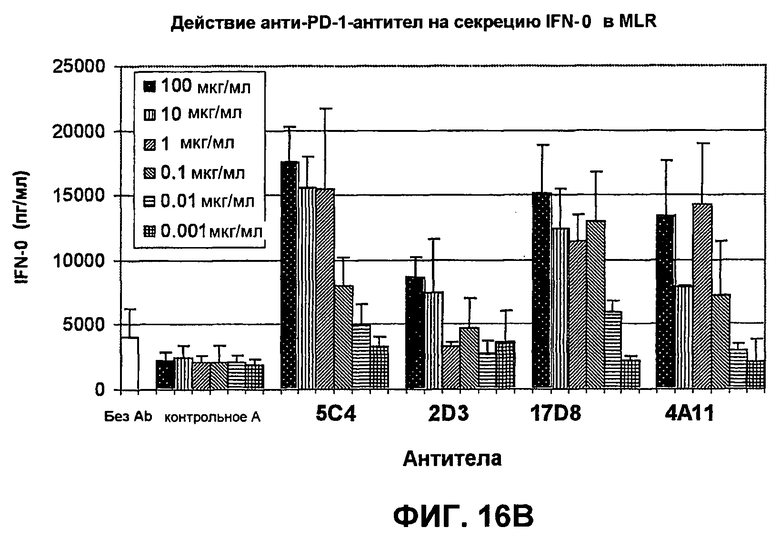
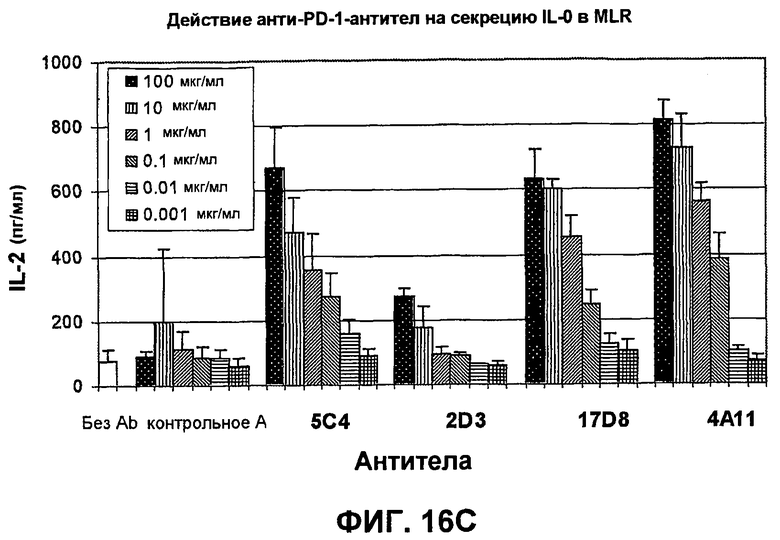
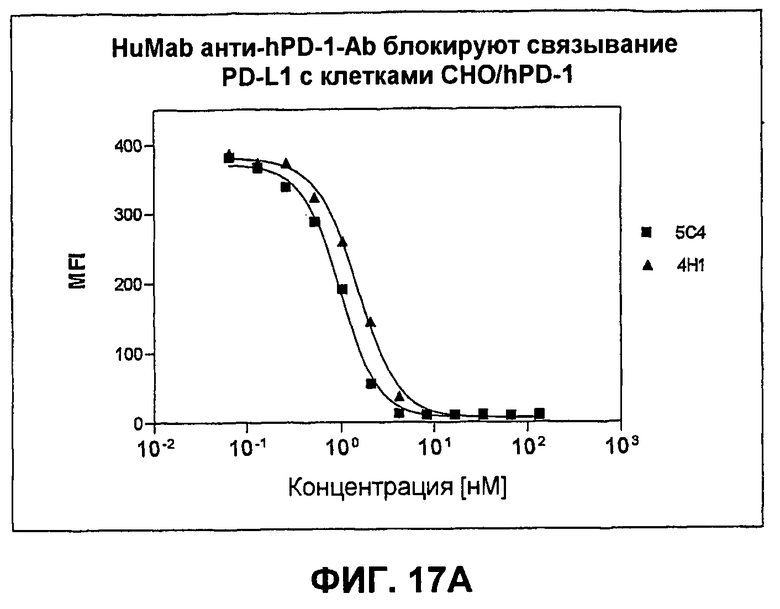
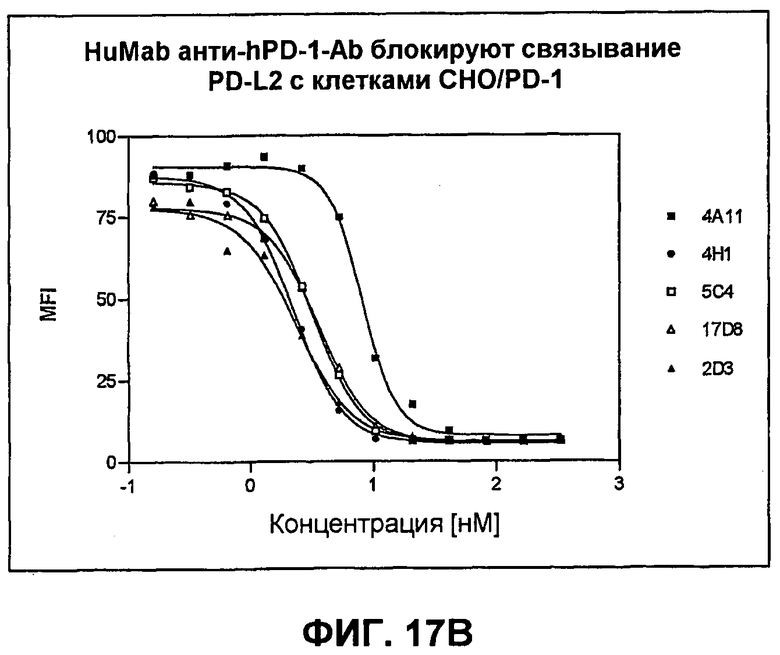
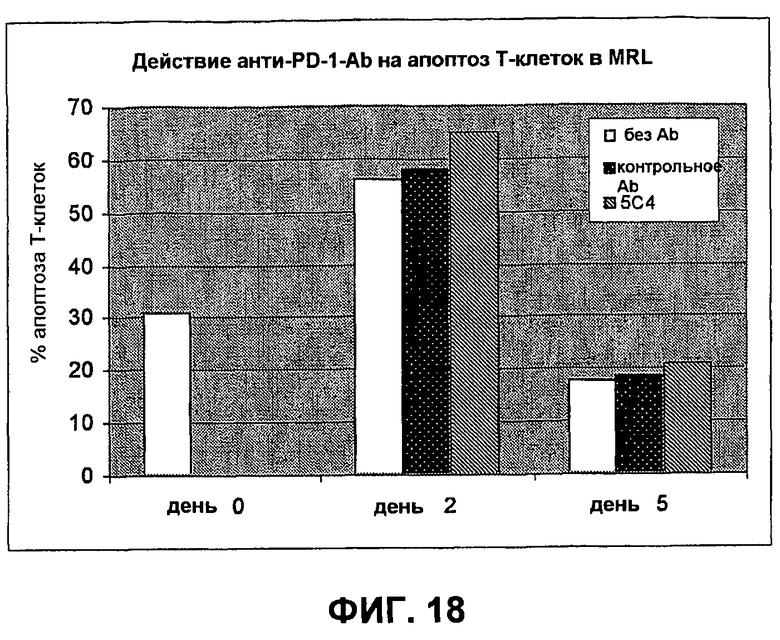
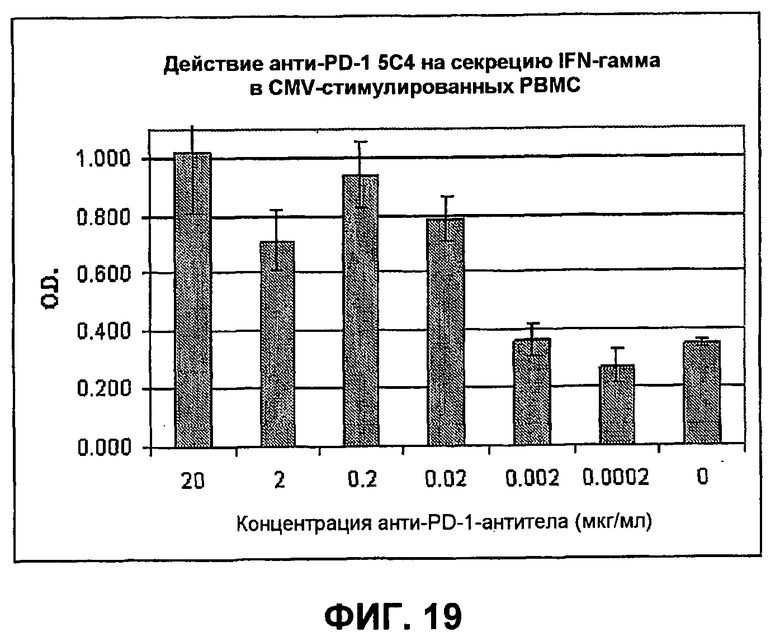
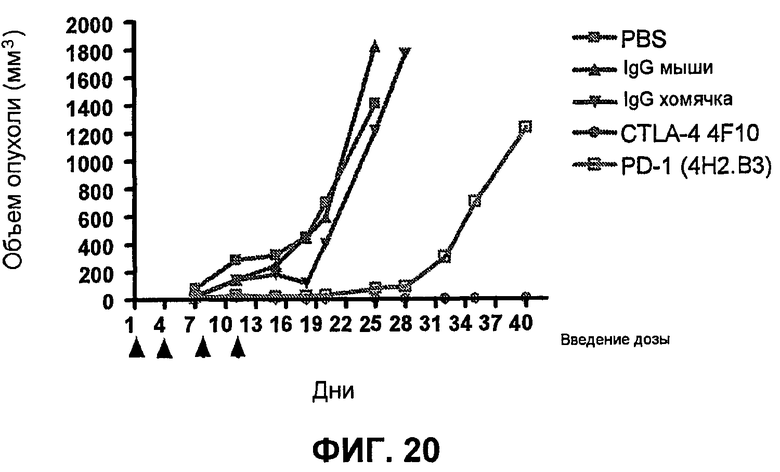
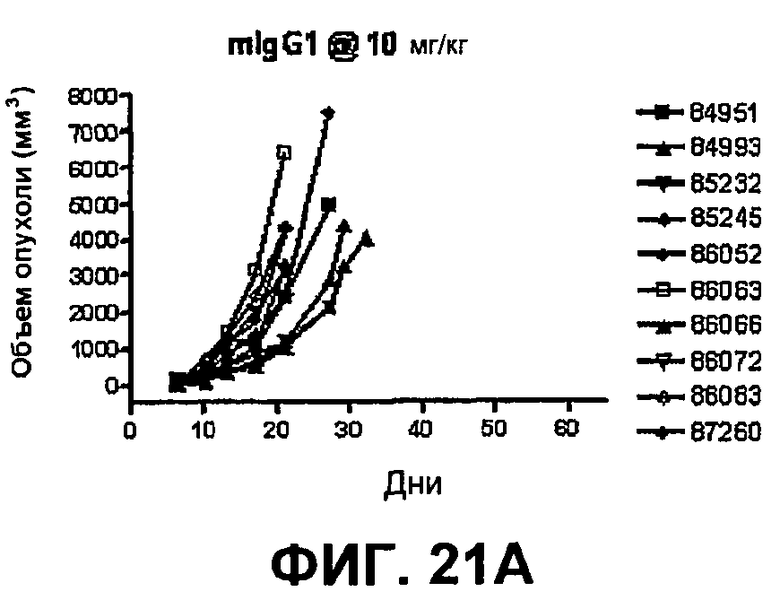
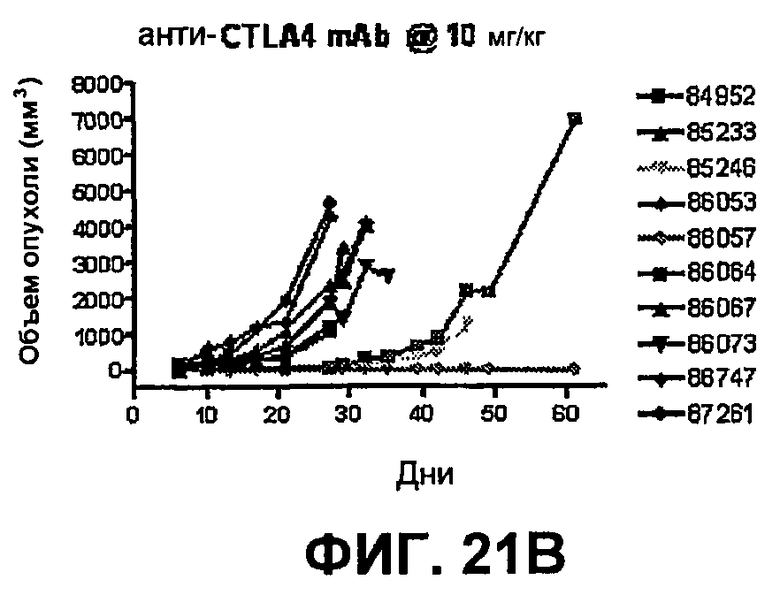
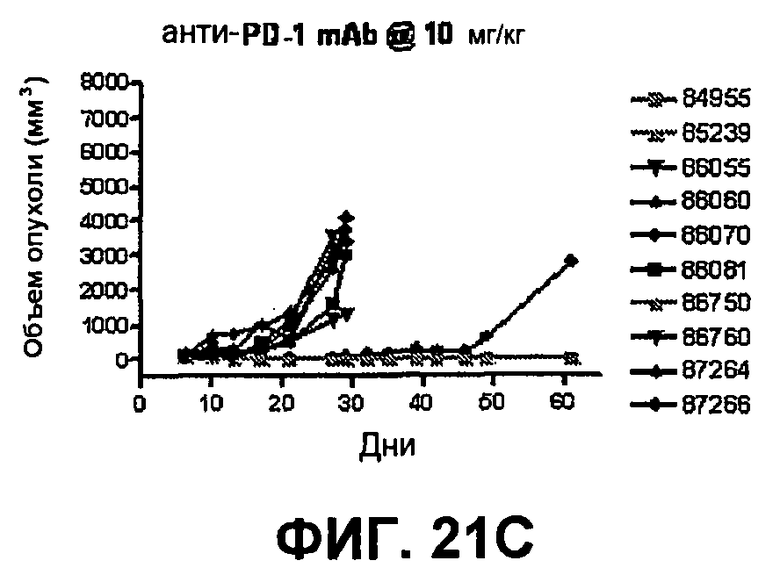
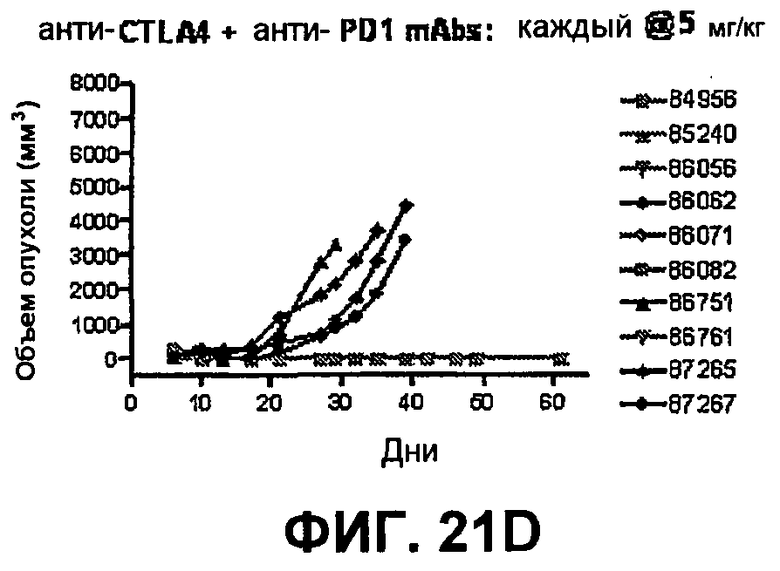

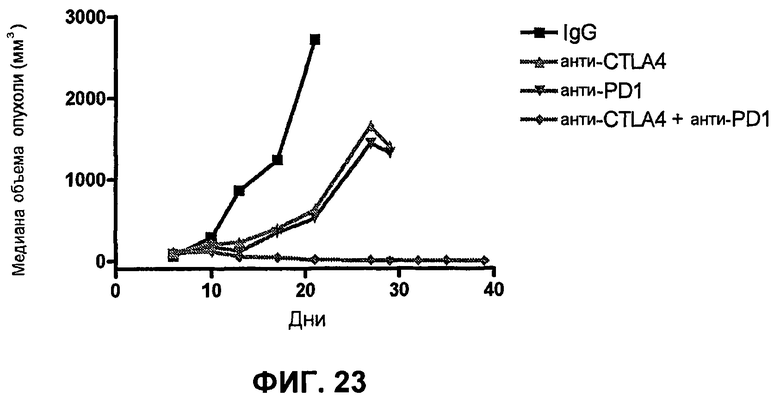
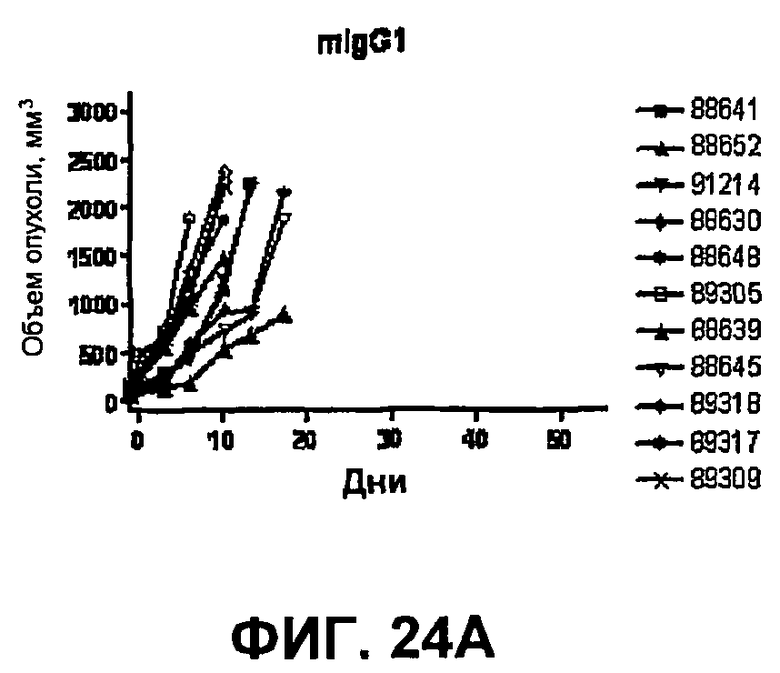
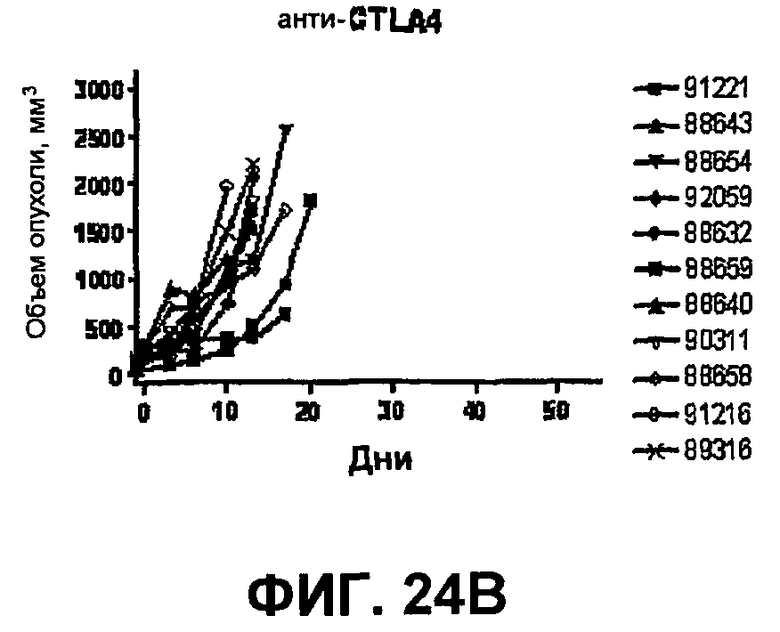
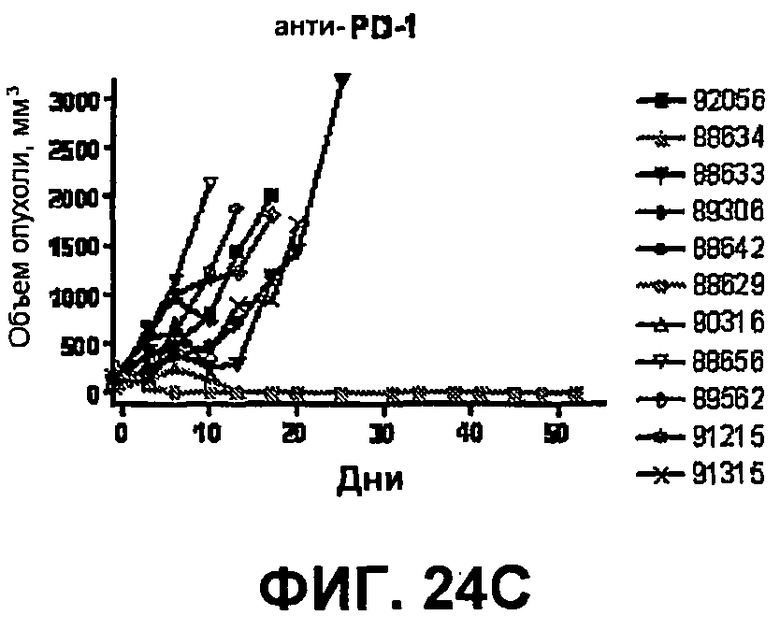
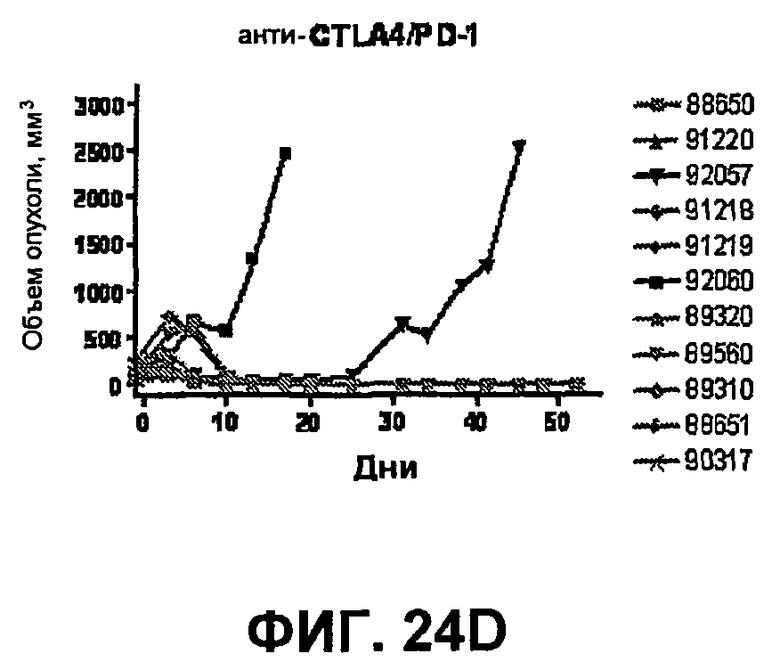
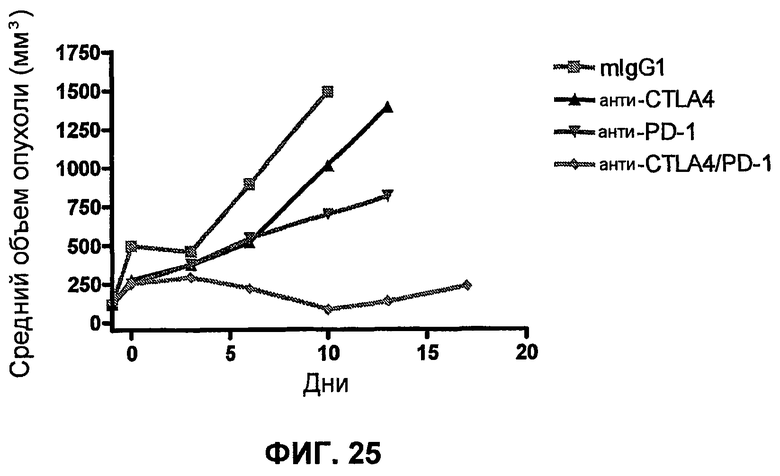
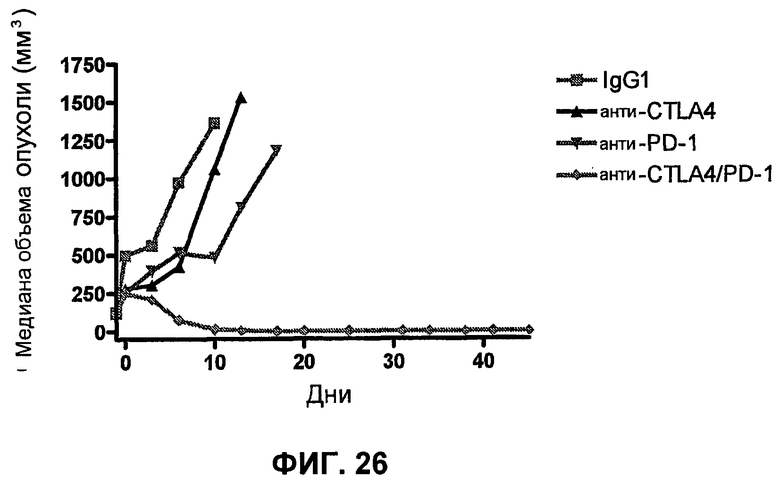
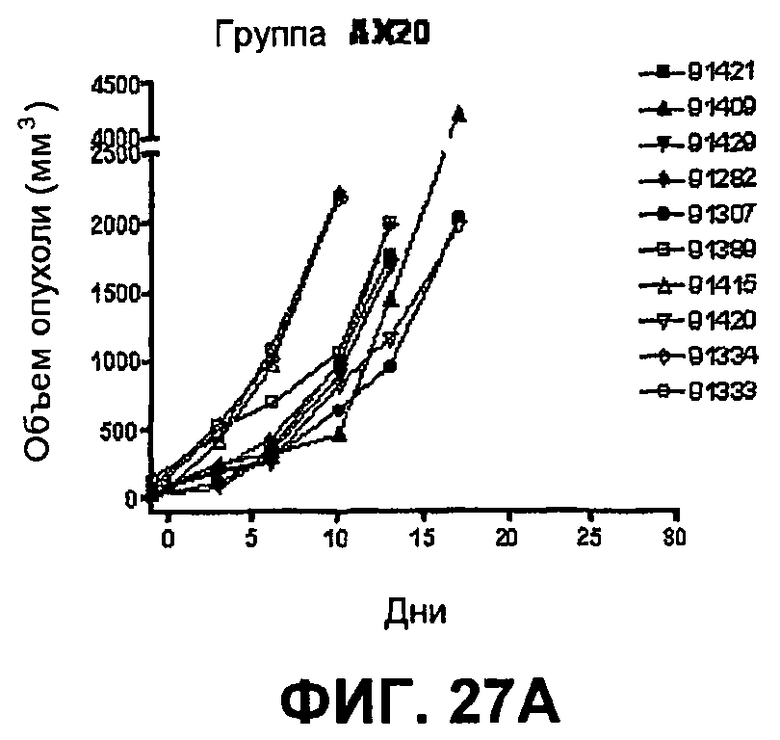
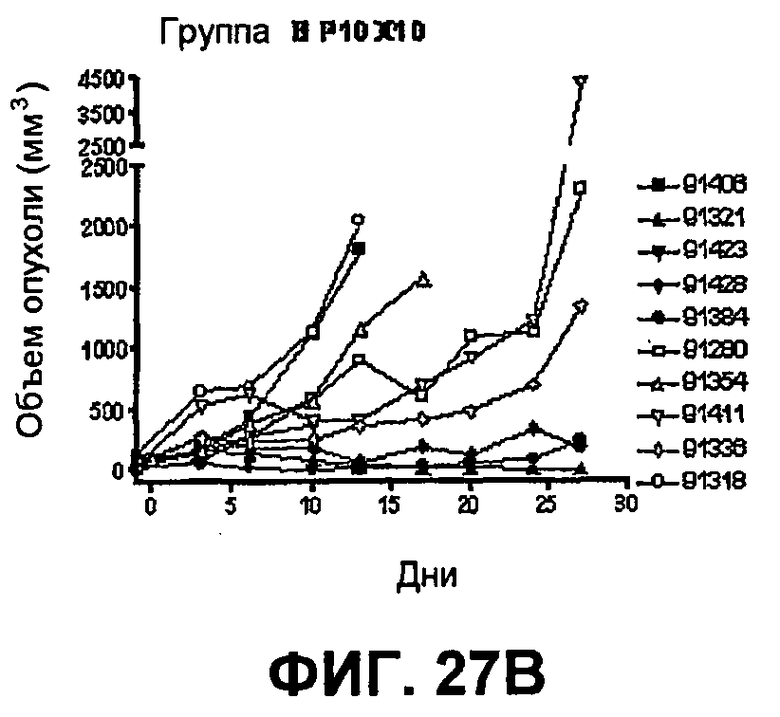
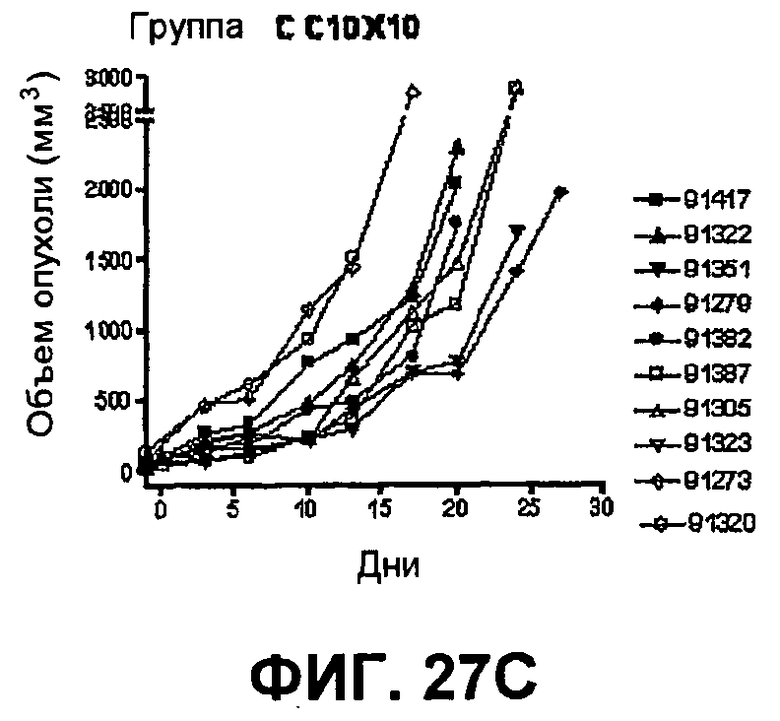
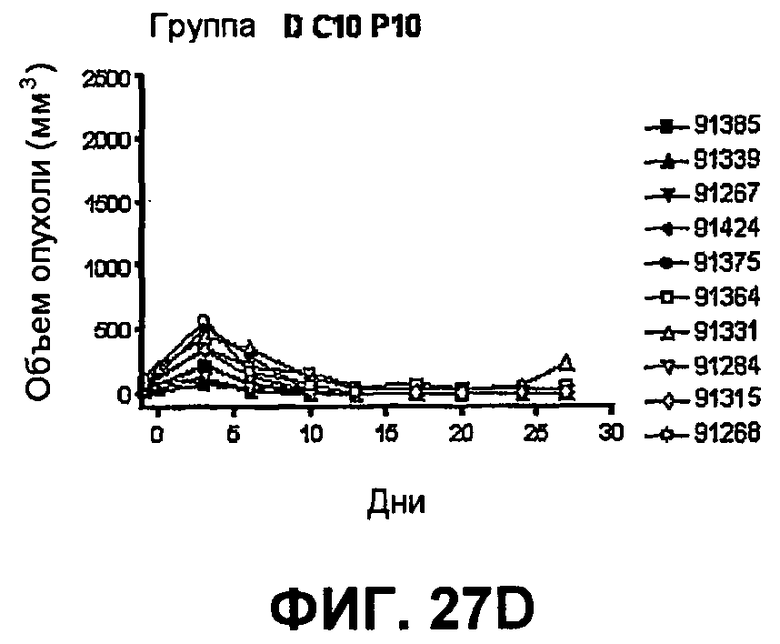
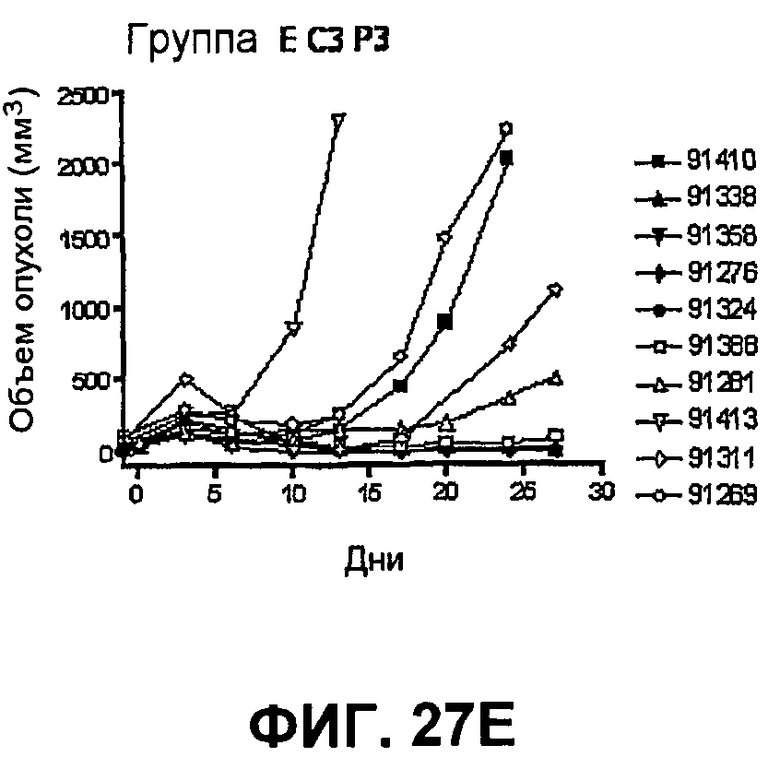
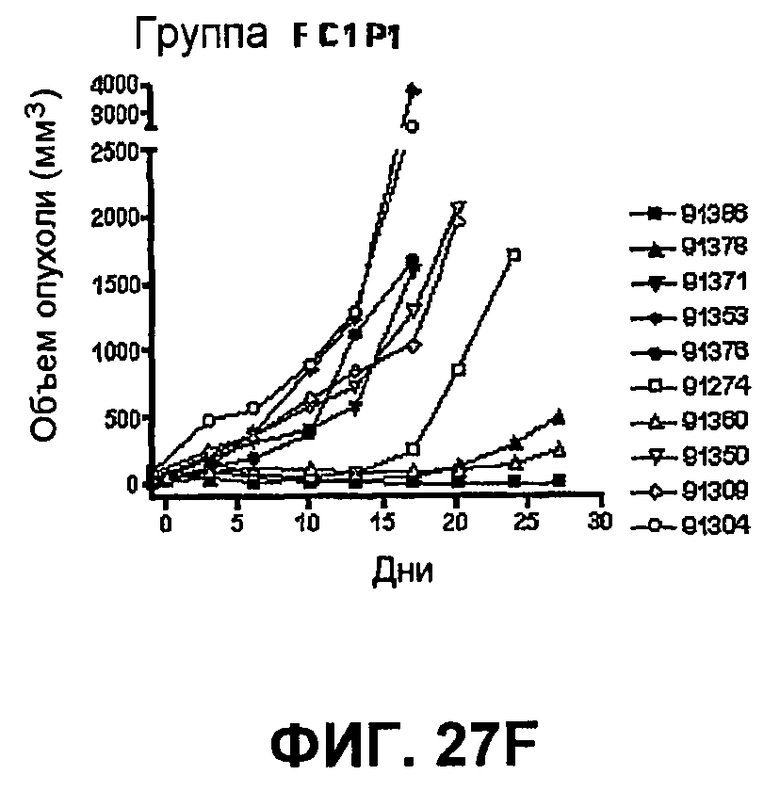
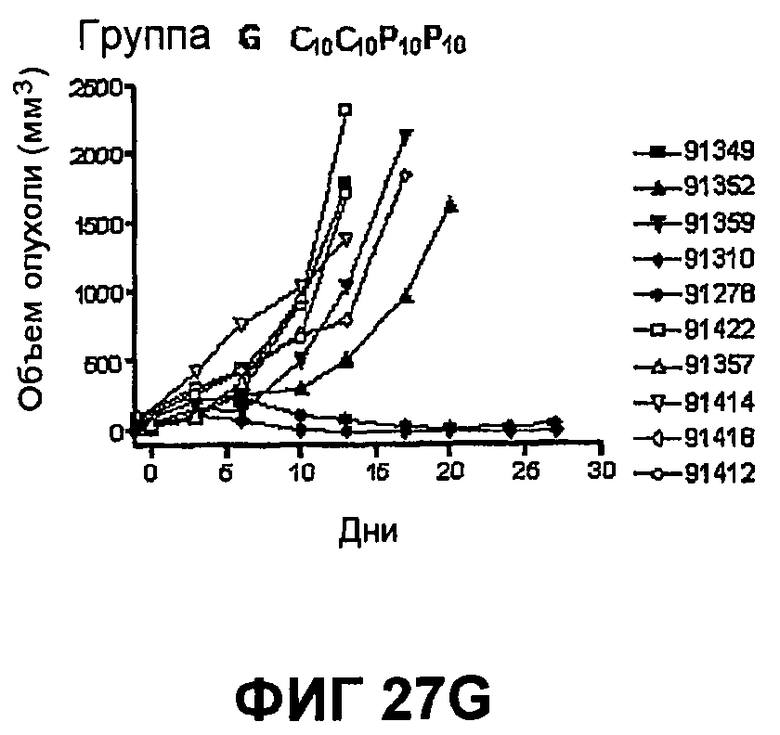
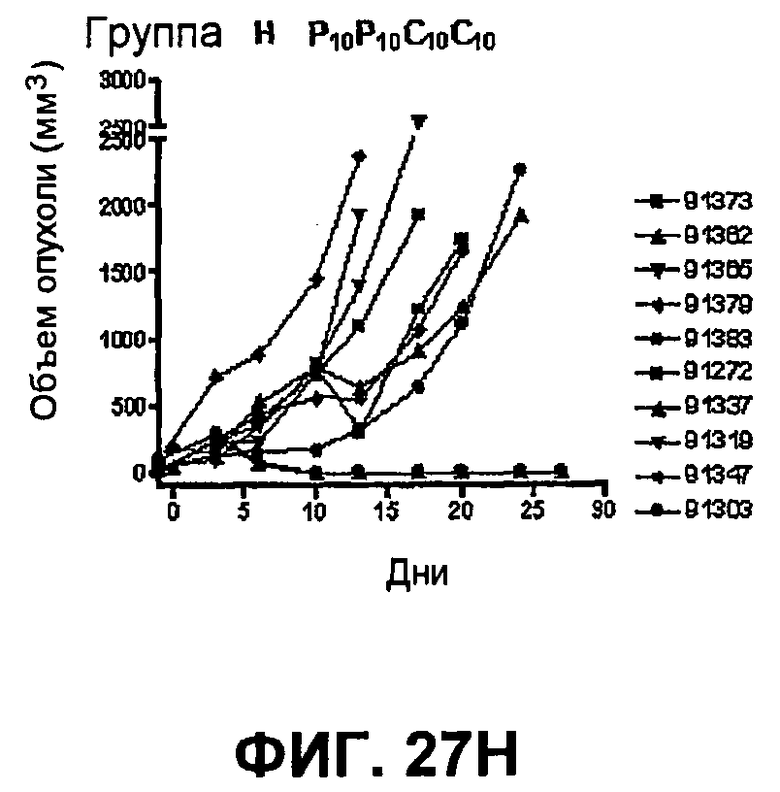
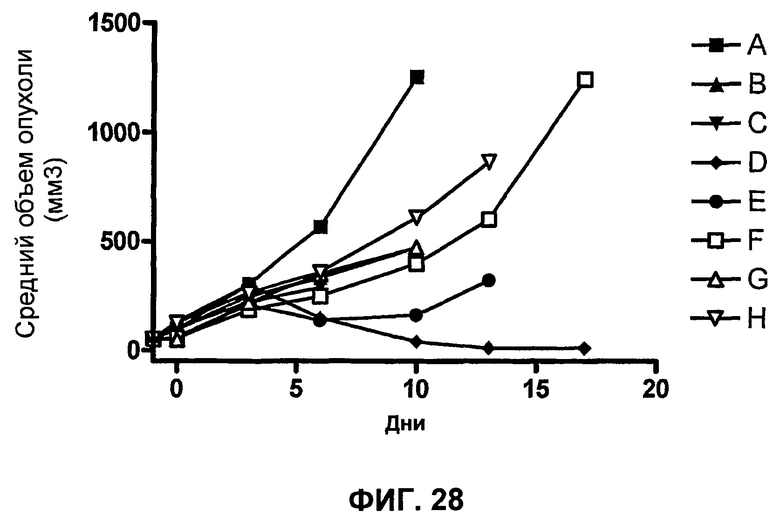
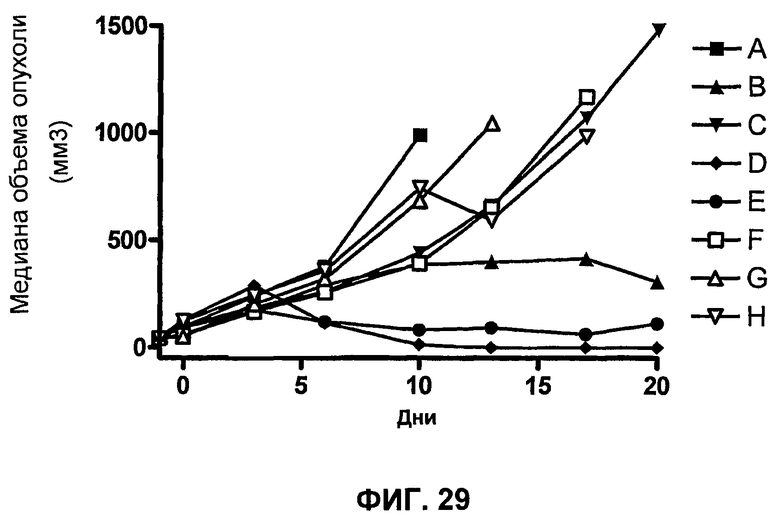
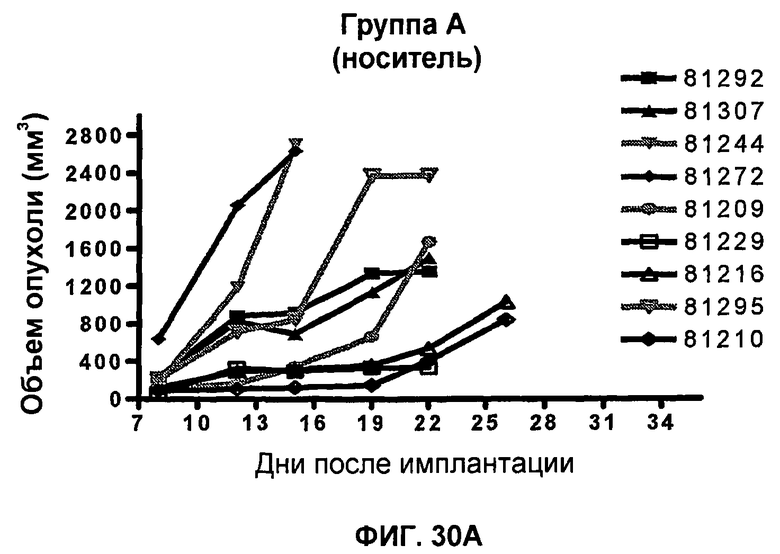
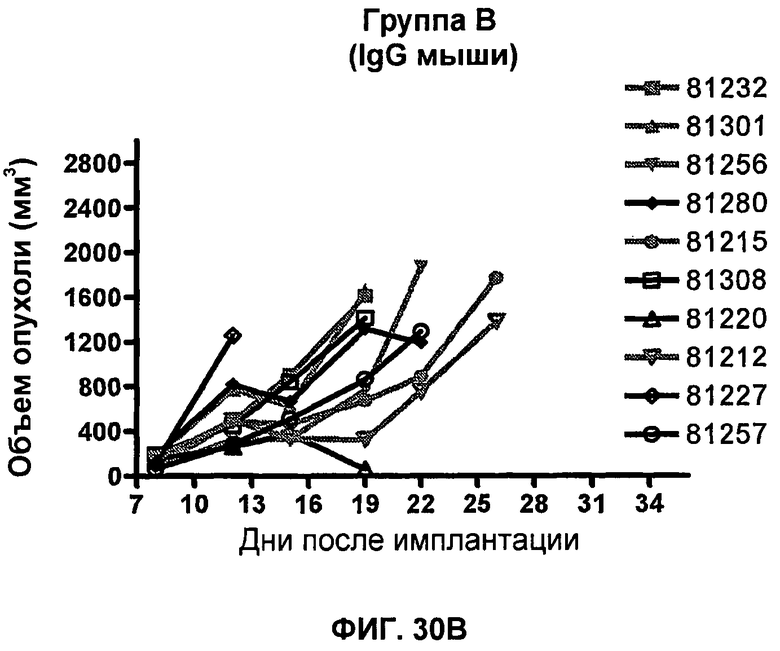

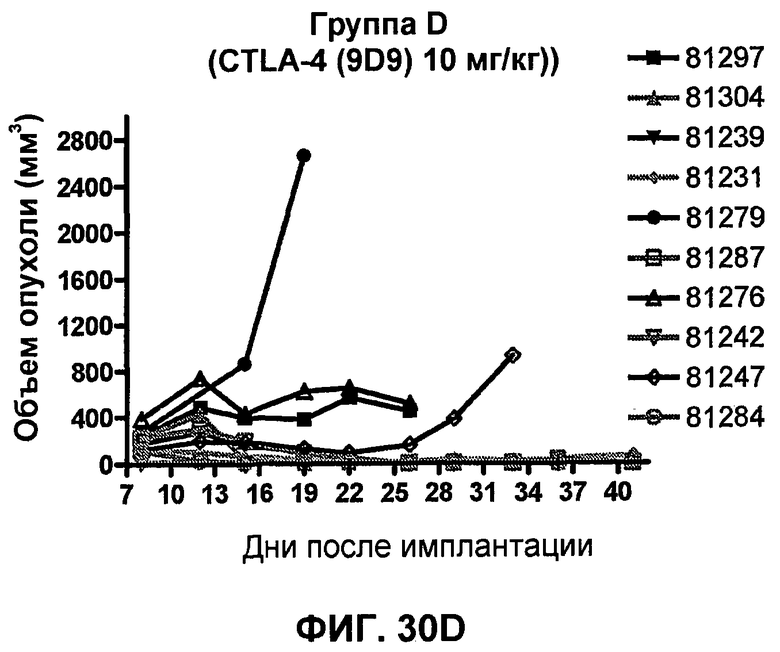
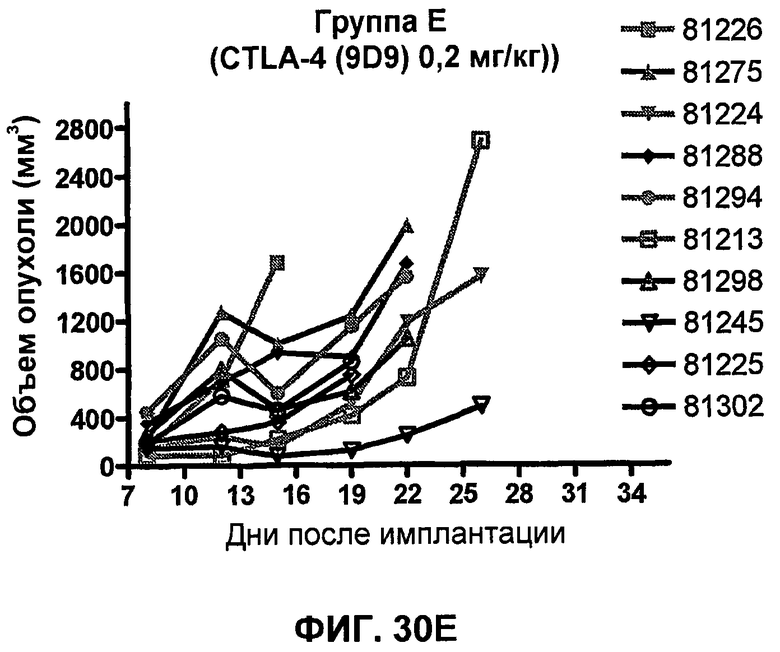
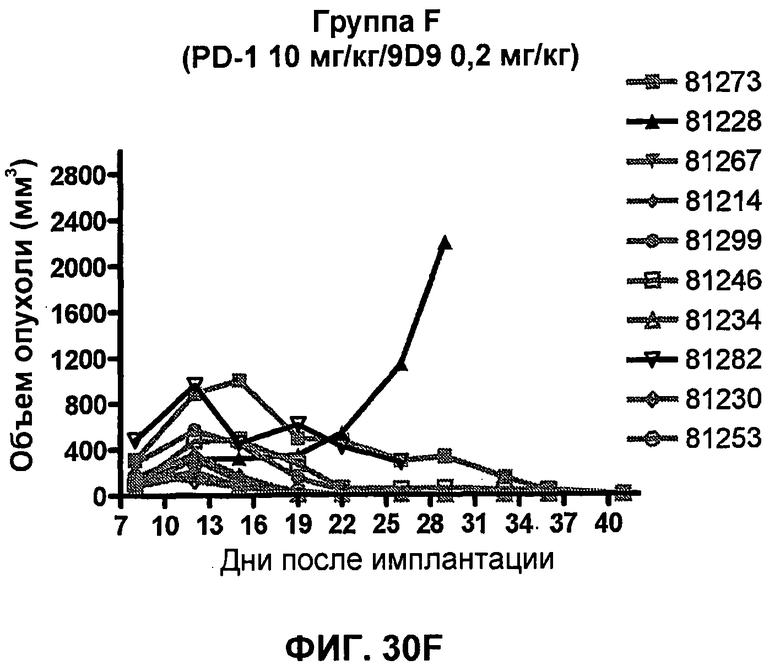
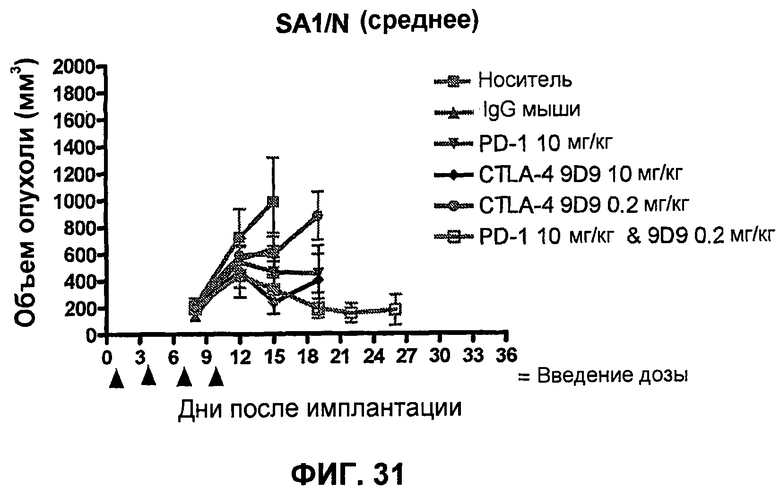
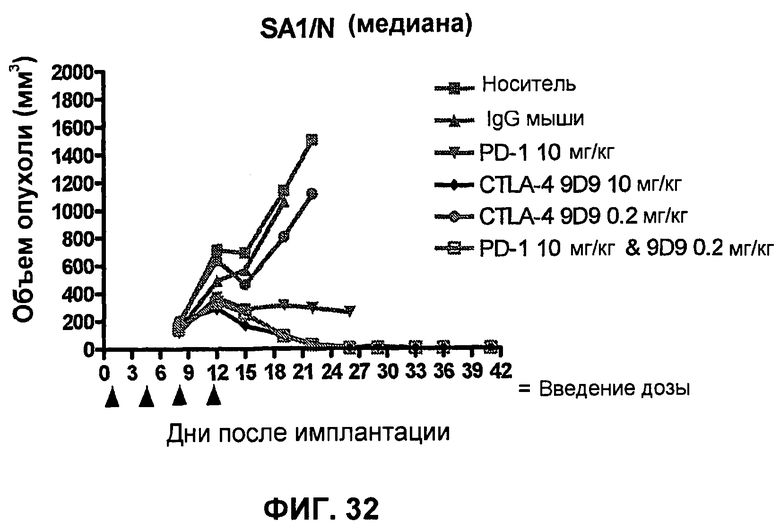
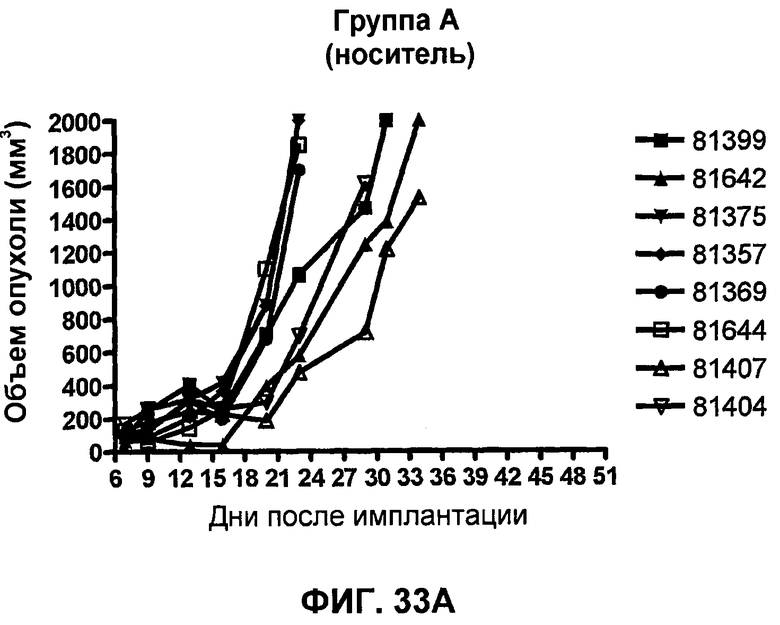
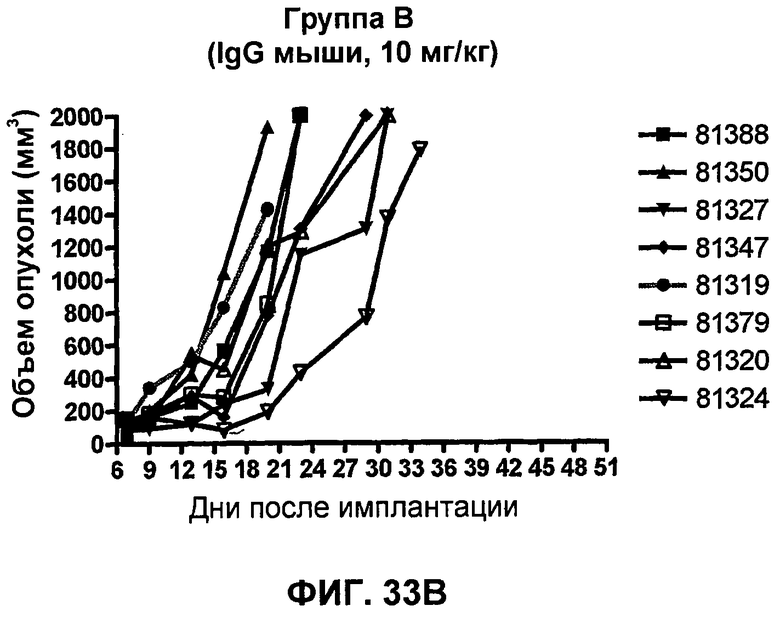
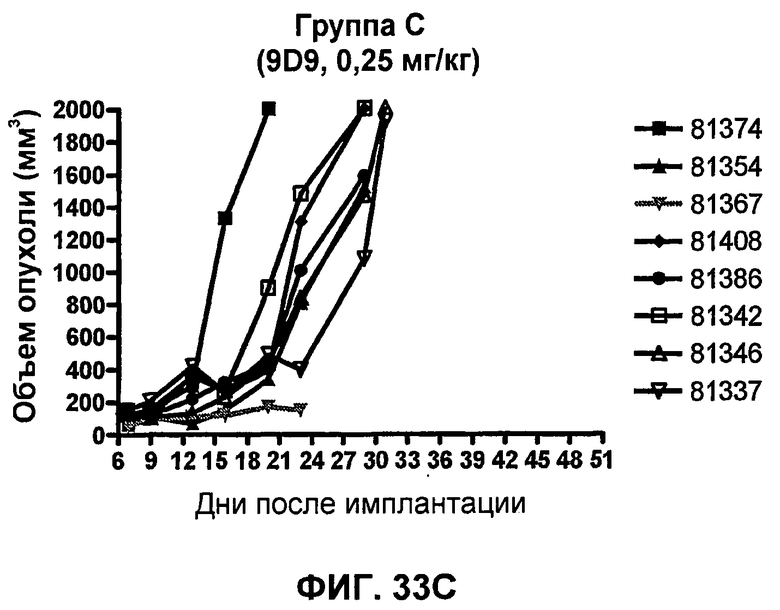
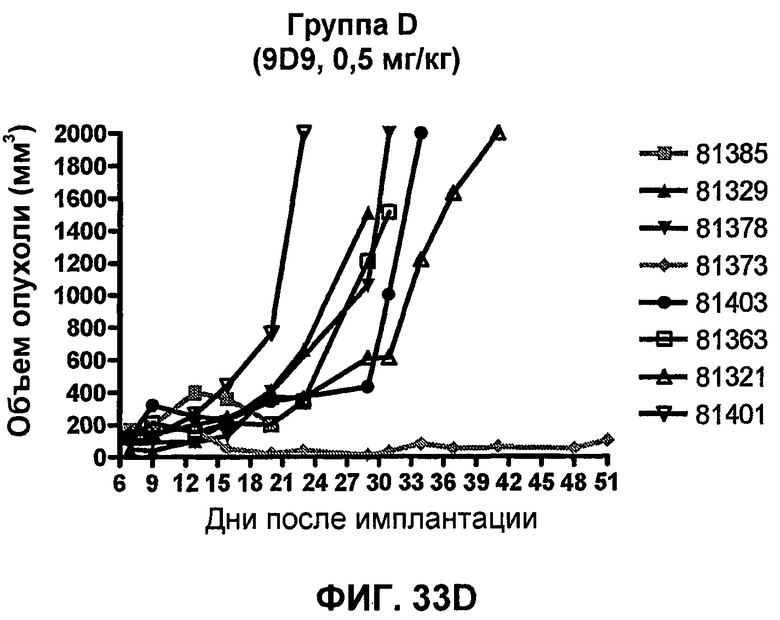
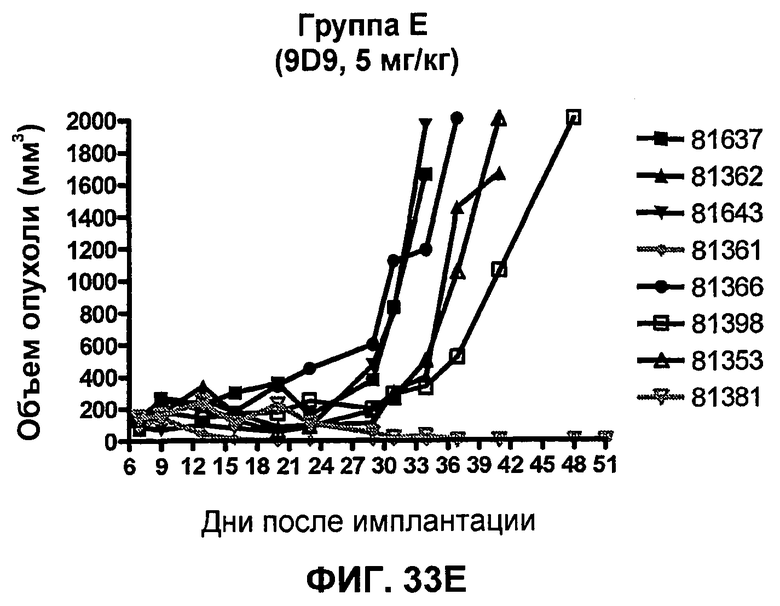
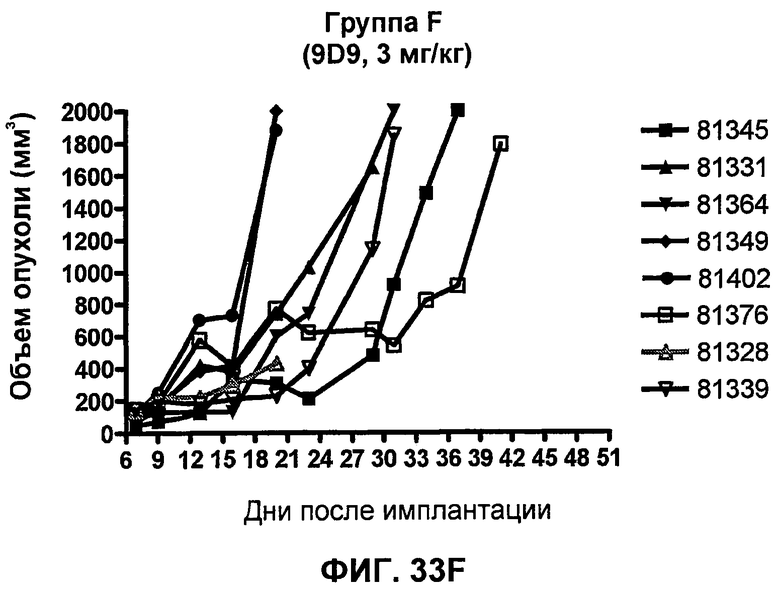
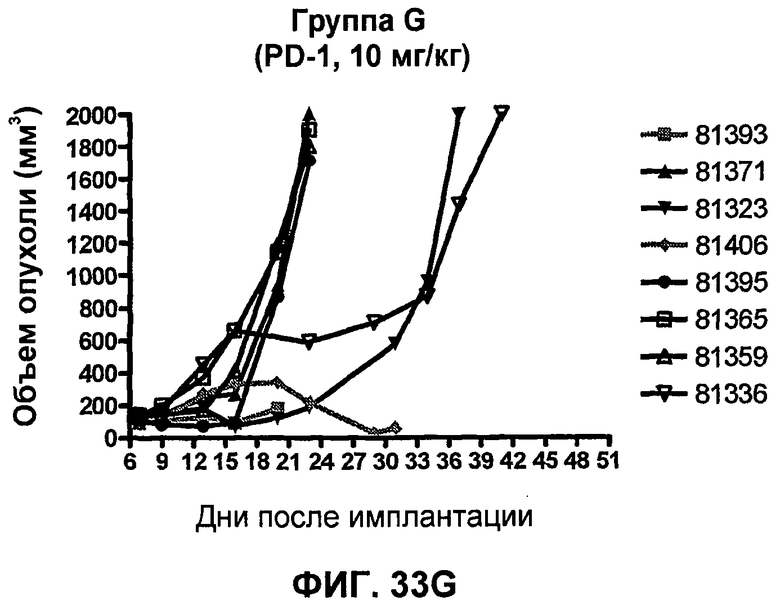
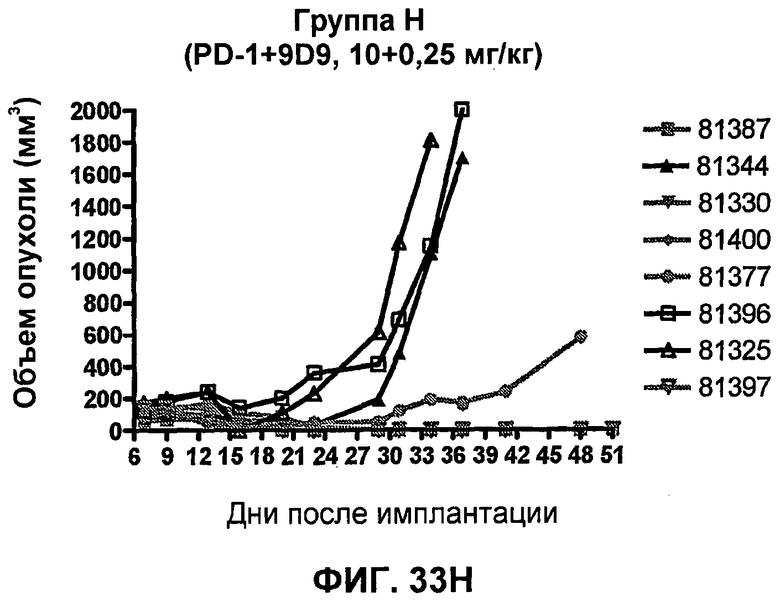
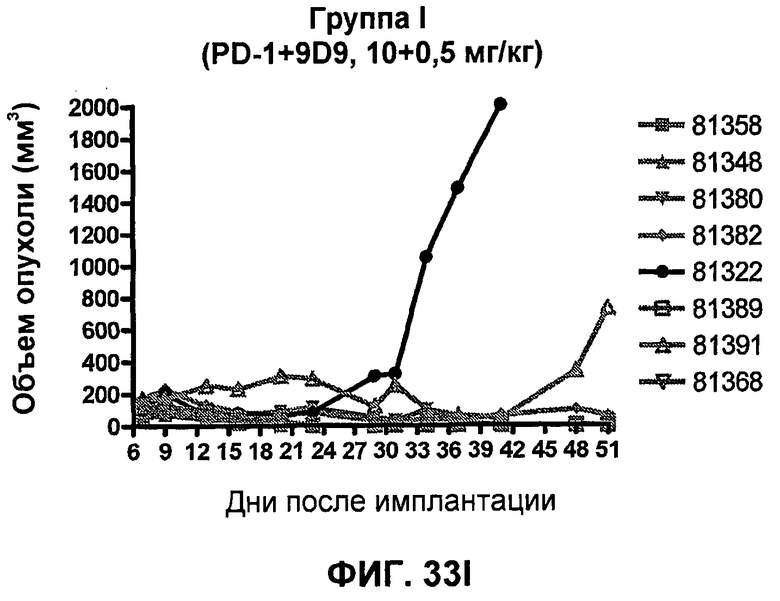
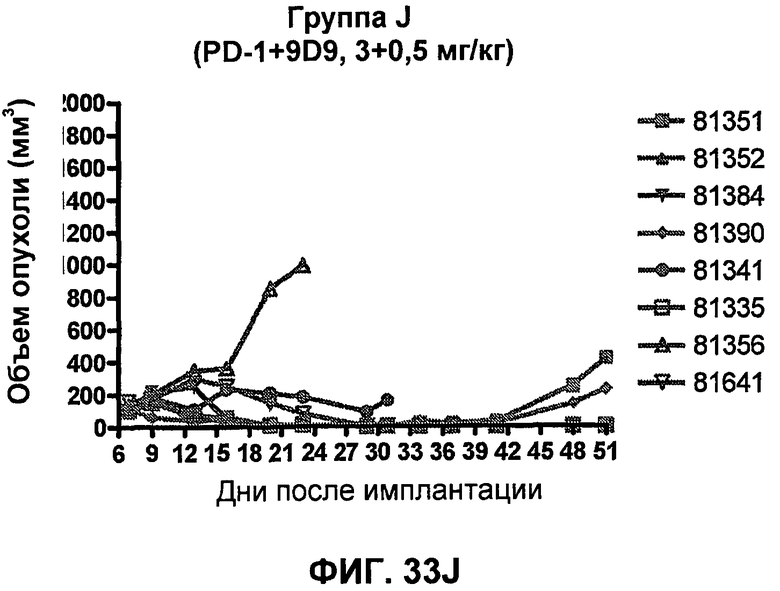

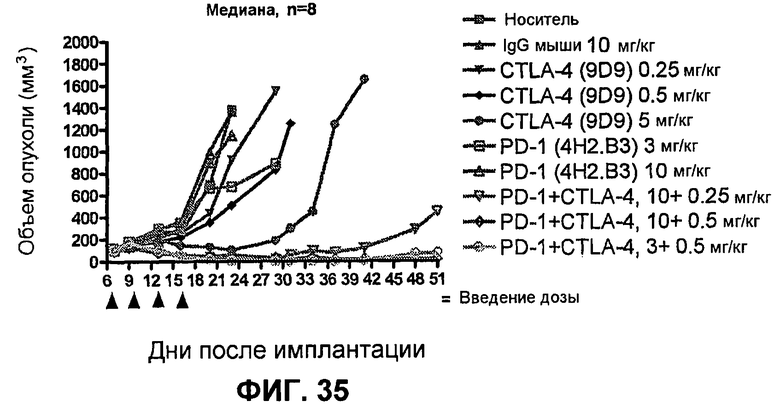

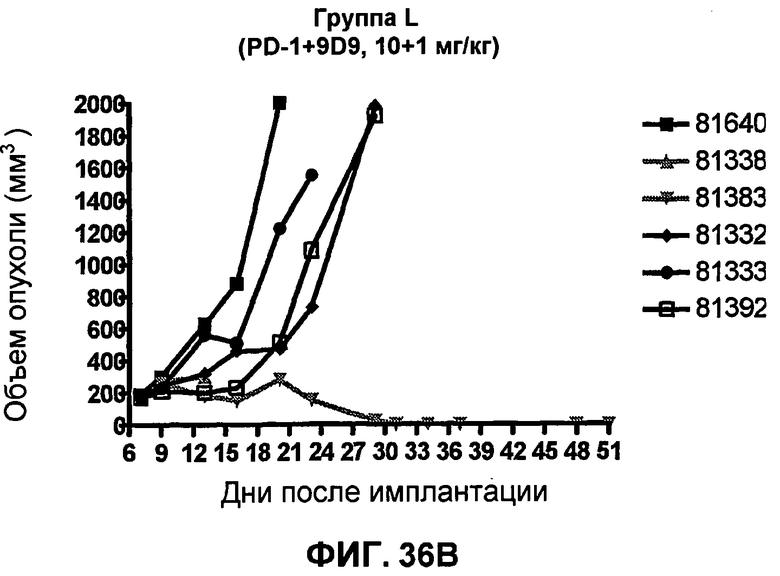

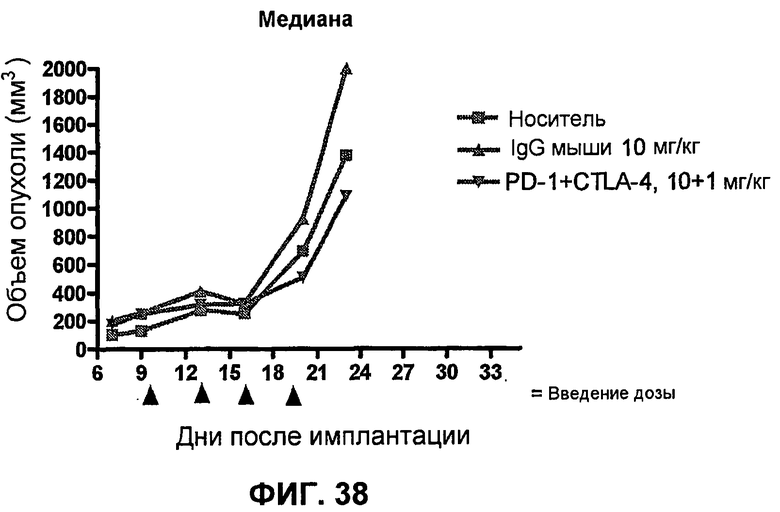
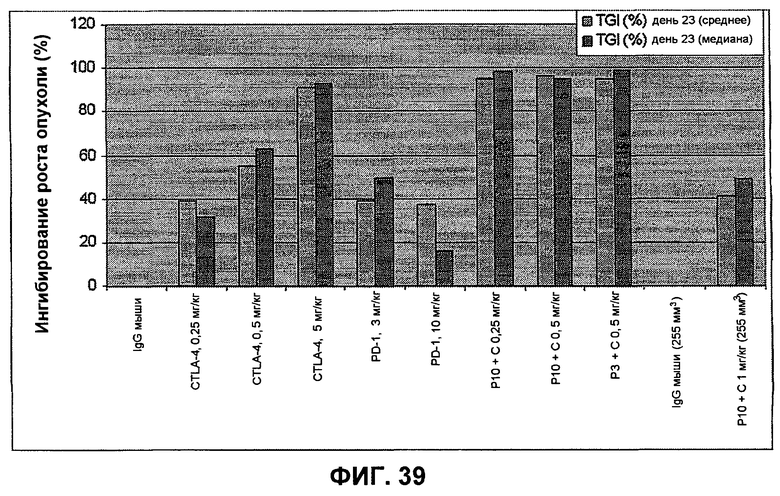
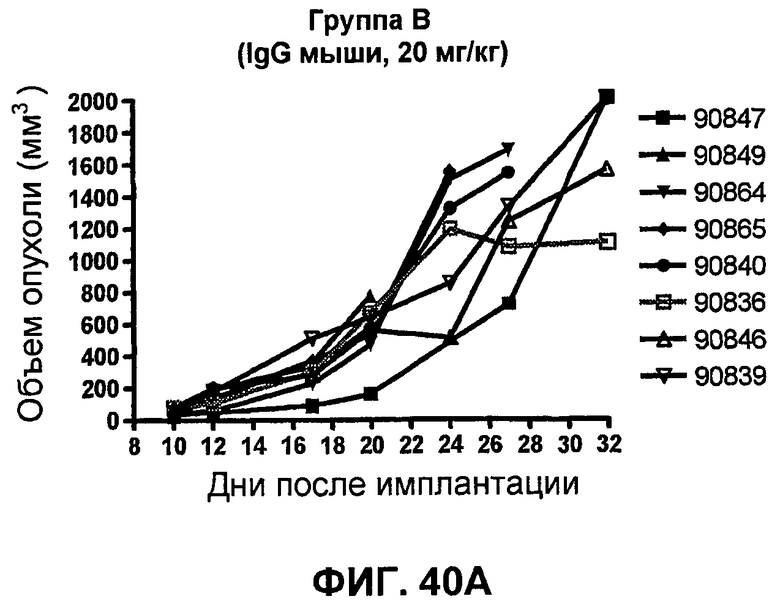
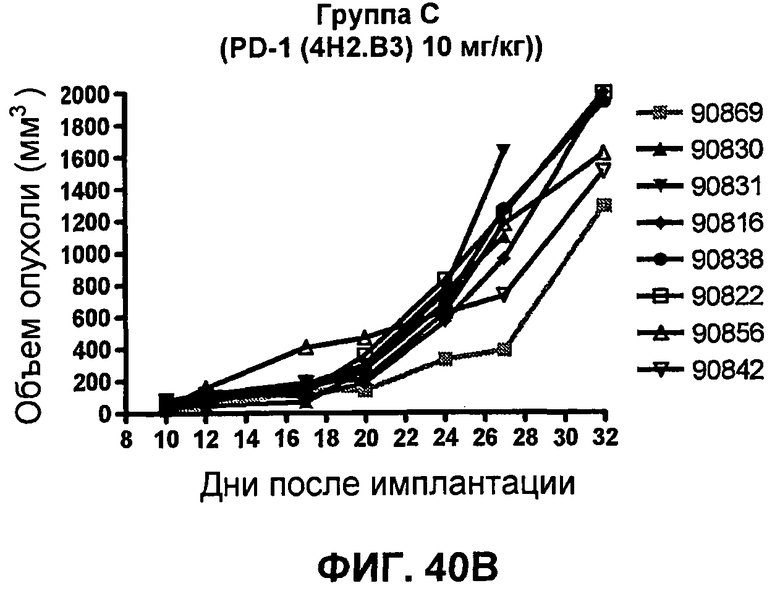
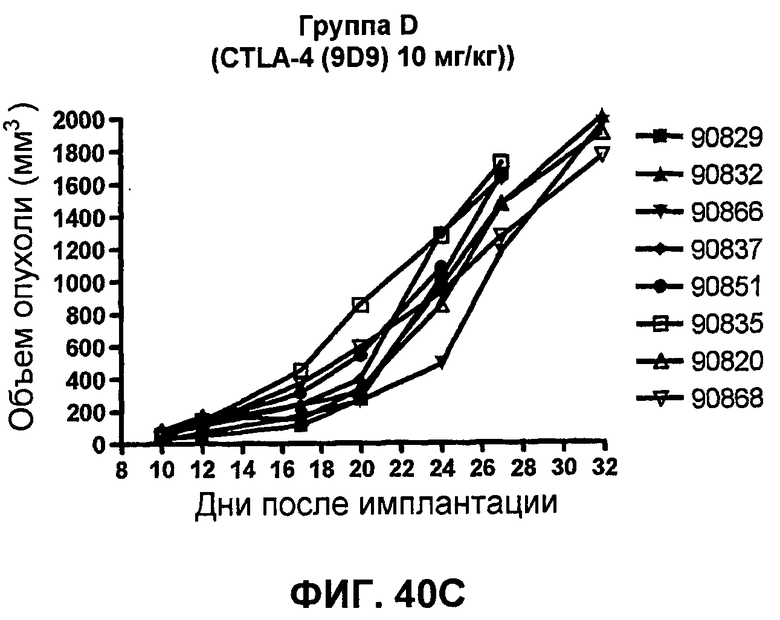
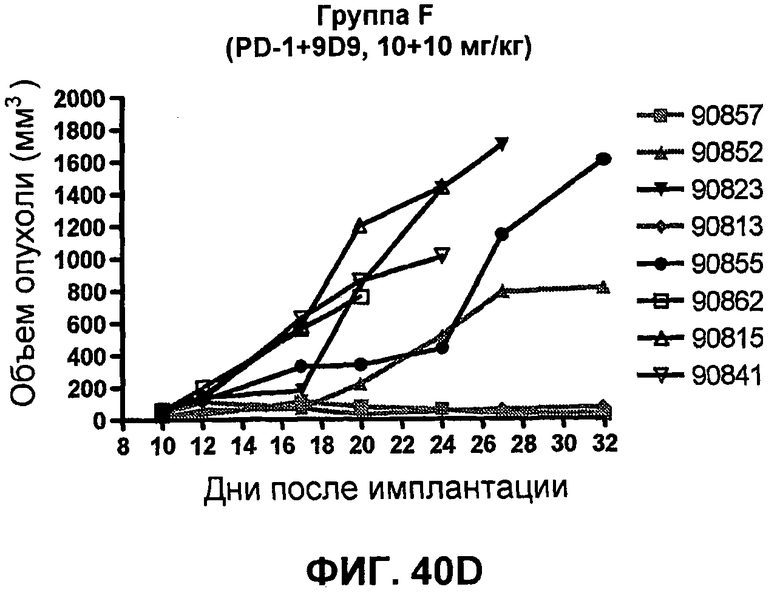
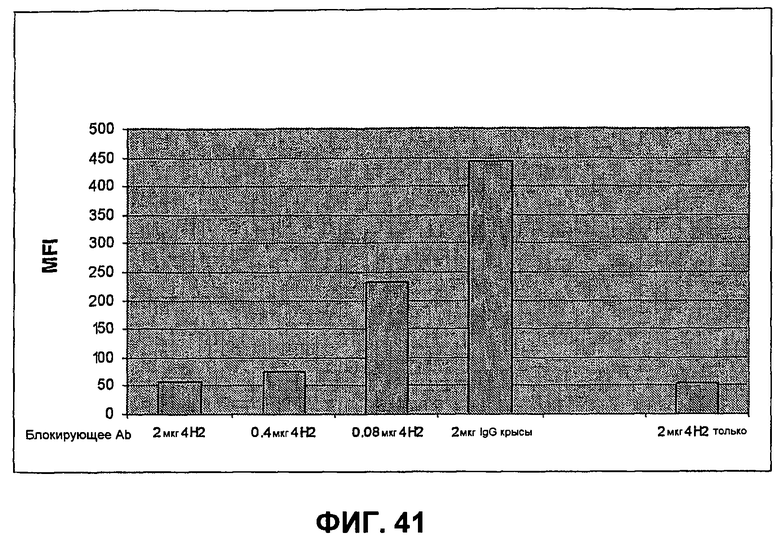
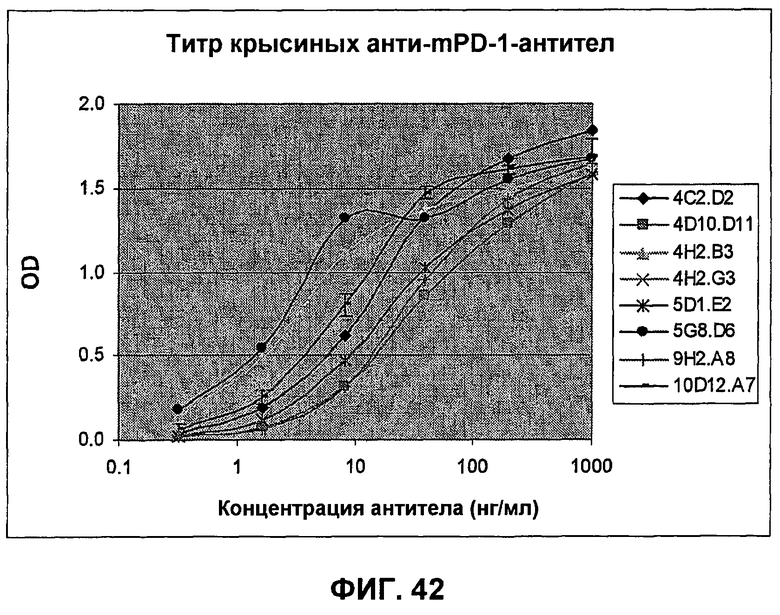
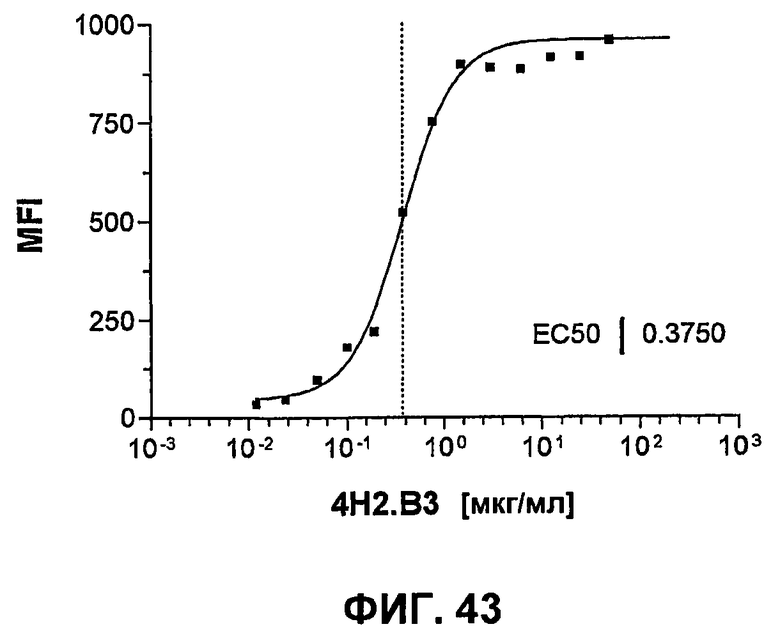
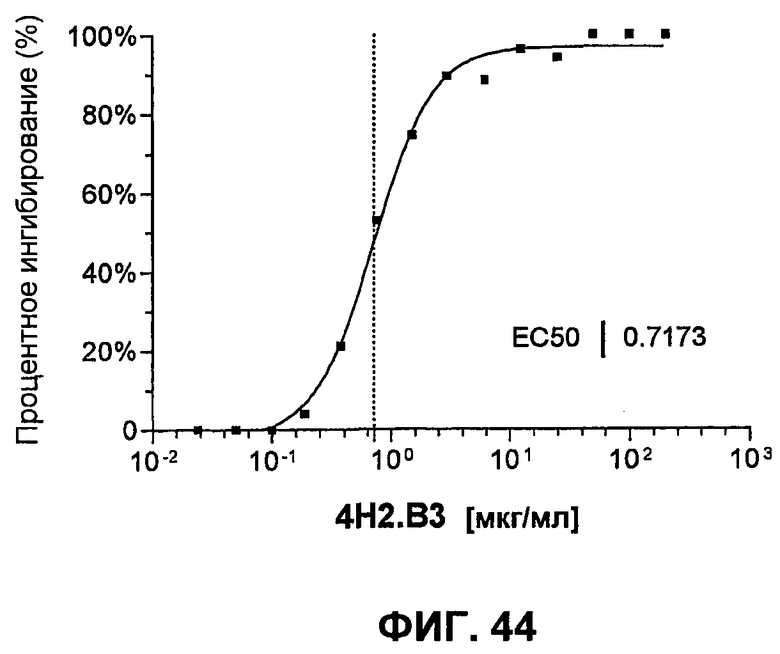
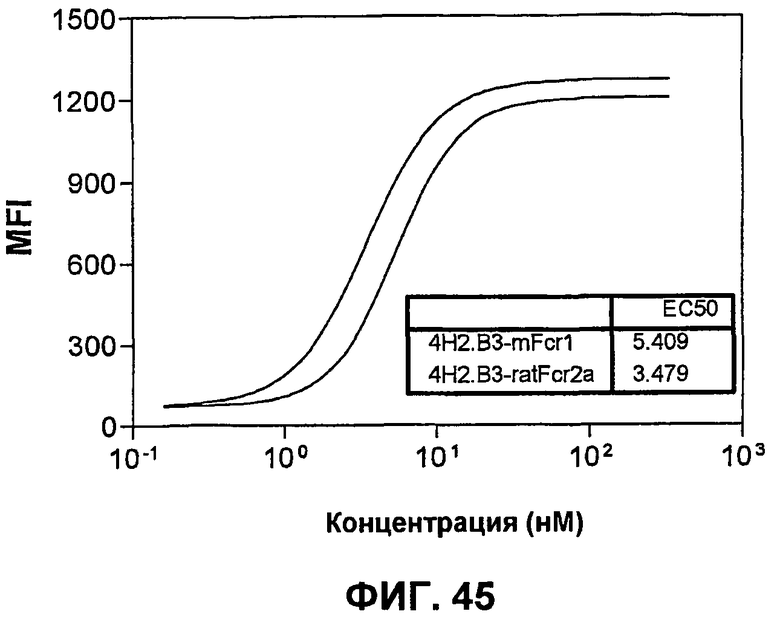
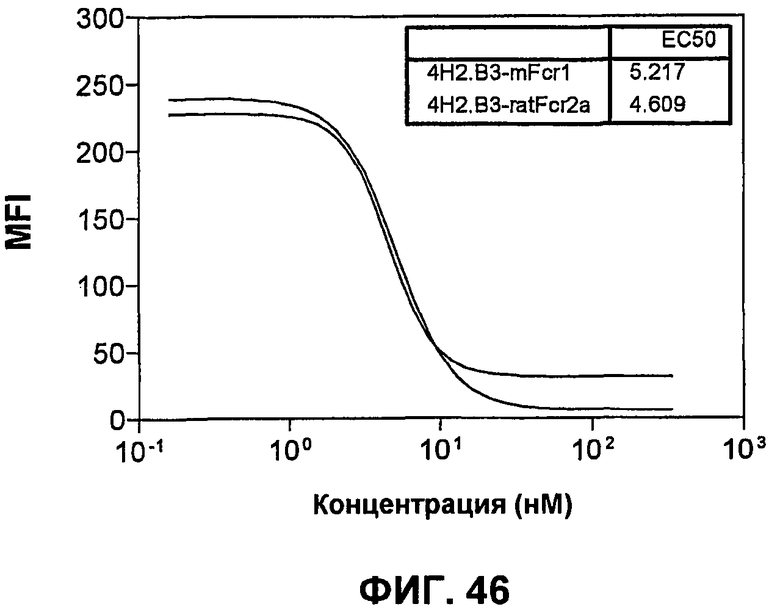
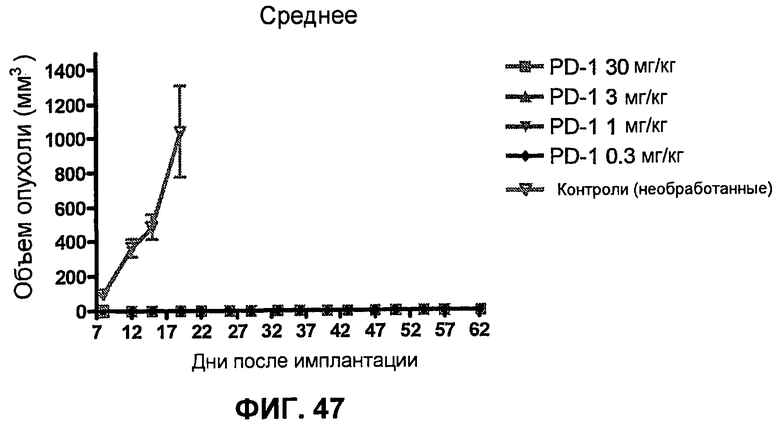
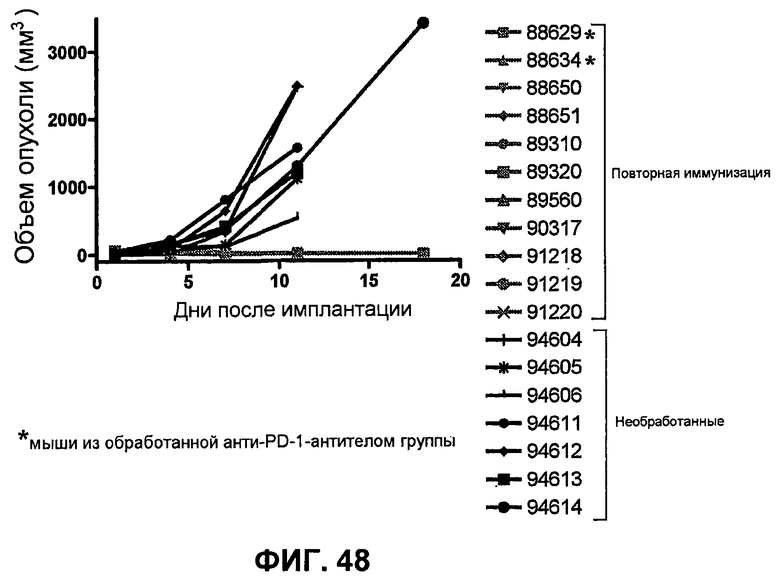
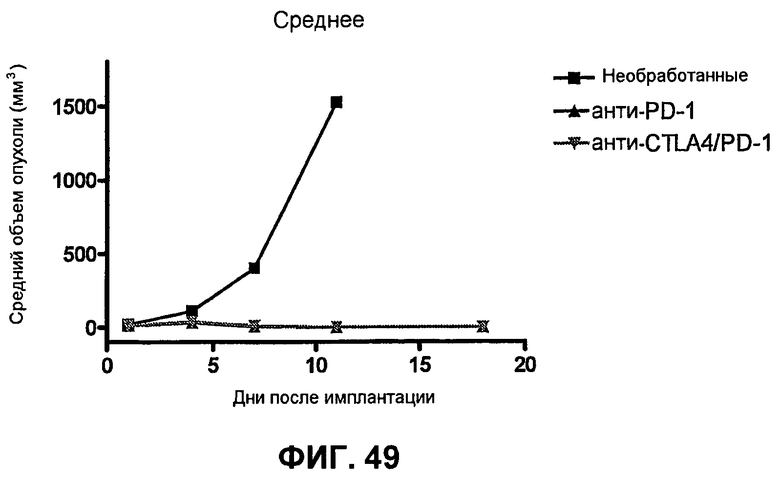
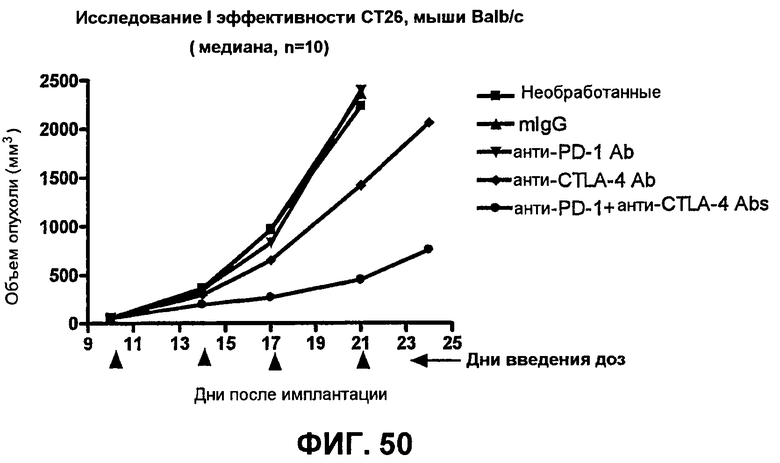
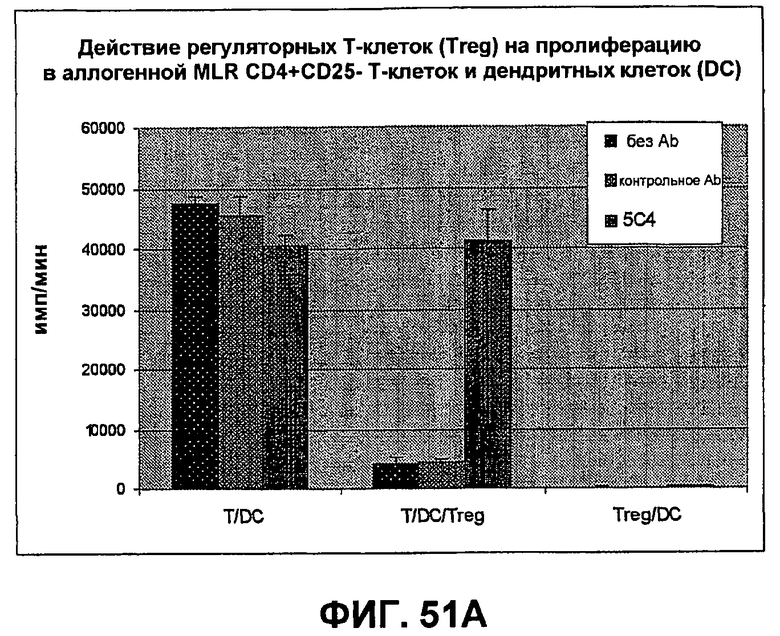
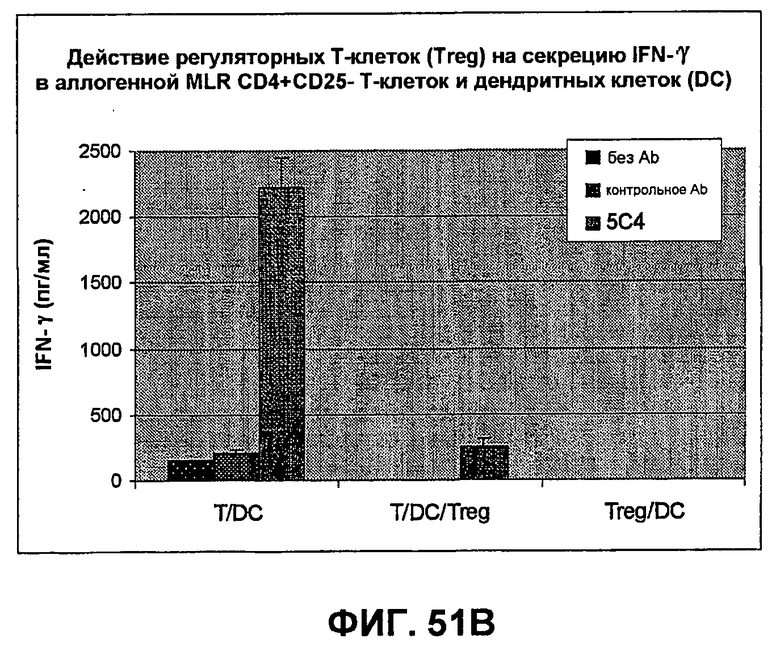

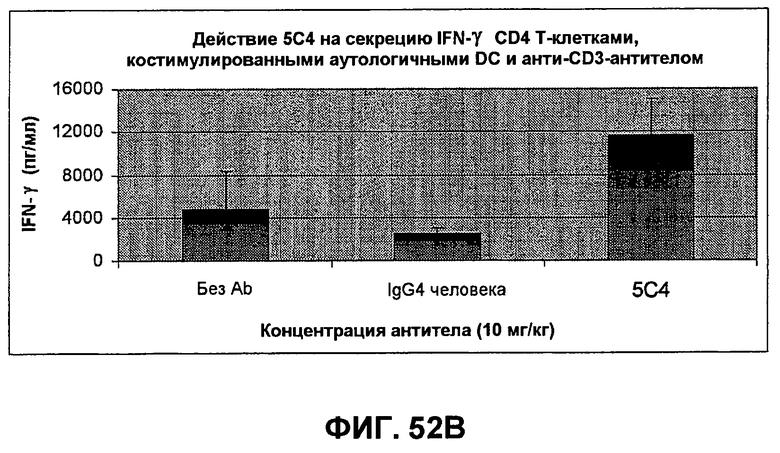
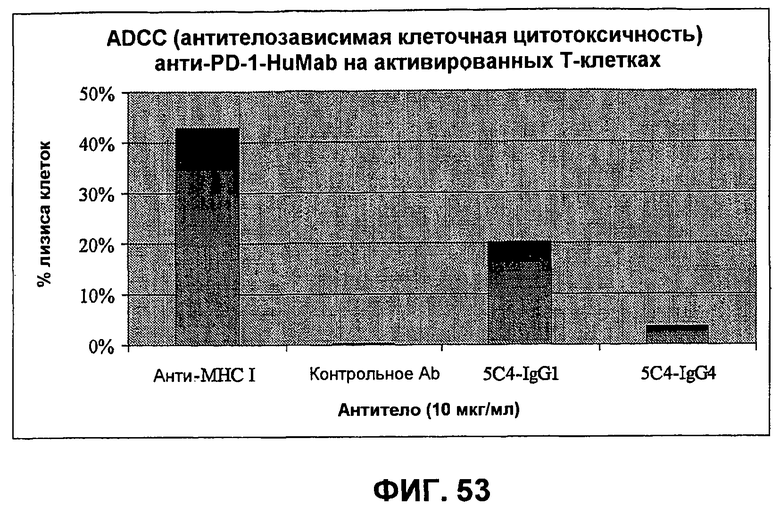
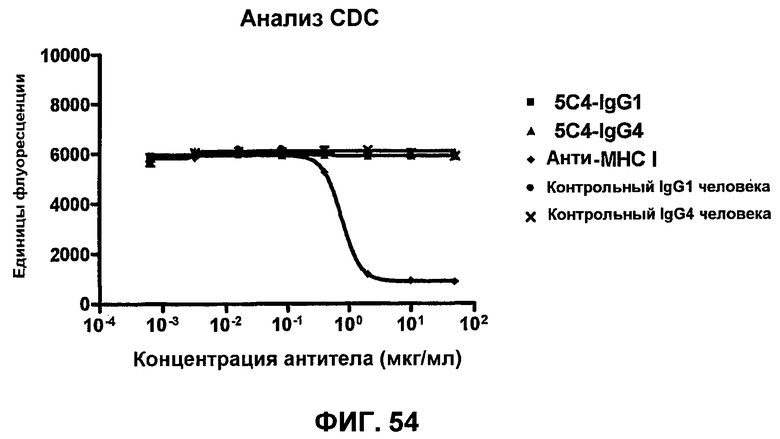
Изобретение относится к области биотехнологии и иммунологии. Представлены: способ ингибирования роста опухолевых клеток у индивидуума и способ усиления иммунного ответа у индивидуума, включающие введение индивидууму моноклонального антитела против PD-1 и моноклонального антитела против CTLA-4 10D1. При этом моноклональное антитело против PD-1 имеет следующие свойства: связывается с PD-1 человека со значением KD, равным 1×10-8 М или меньше; не связывается ни с CD28 человека, ни с CTLA-4, ни с ICOS; способно усиливать пролиферацию Т-клеток в анализе реакции лимфоцитов в смешанной культуре (MLR); способно усиливать продукцию интерферона гамма в анализе MLR и способно усиливать секрецию интерлейкина-2 (IL-2) в анализе MLR. Изобретение обеспечивает синергическое действие при совместном использовании указанных антител. 2 н. и 8 з.п. ф-лы, 54 ил., 7 табл., 25 пр.
1. Способ ингибирования роста опухолевых клеток у индивидуума, включающий введение индивидууму:
(a) моноклонального антитела против PD-1, и
(b) моноклонального антитела против CTLA-4 10D1,
где моноклональное антитело против PD-1 имеет следующие свойства:
(i) связывается с PD-1 человека со значением KD, равным 1·10-8 М или меньше;
(ii) не связывается ни с CD28 человека, ни с CTLA-4, ни с ICOS;
(iii) способно усиливать пролиферацию Т-клеток в анализе реакции лимфоцитов в смешанной культуре (MLR);
(iv) способно усиливать продукцию интерферона гамма в анализе MLR; и
(v) способно усиливать секрецию интерлейкина-2 (IL-2) в анализе MLR.
2. Способ по п.1, где опухолевыми клетками являются клетки злокачественной опухоли, выбранной из группы, состоящей из меланомы, рака почки, рака предстательной железы, рака молочной железы, рака толстой кишки и рака легкого.
3. Способ по п.1, где опухолевыми клетки являются клетки злокачественной опухоли, выбранной из группы, состоящей из рака костей, рака поджелудочной железы, рака кожи, рака головы и шеи, кожной или внутриглазной злокачественной меланомы, рака матки, рака яичника, рака прямой кишки, рака анальной области, рака желудка, рака яичек, рака матки, рака фаллопиевых труб, рака эндометрия, рака шейки матки, рака влагалища, рака вульвы, болезни Ходжкина, не-Ходжкинской лимфомы, рака пищевода, рака тонкой кишки, рака эндокринной системы, рака щитовидной железы, рака паращитовидной железы, рака надпочечников, саркомы мягких тканей, рака мочеиспускательного канала, рака пениса, хронического или острого лейкоза, включая острый миелоидный лейкоз, хронический миелоидный лейкоз, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, солидных опухолей детей, лимфоцитарной лимфомы, рака мочевого пузыря, рака почек или мочеточника, рака почечных лоханок, неоплазмы центральной нервной системы (ЦНС), первичной лимфомы ЦНС, опухолевого ангиогенеза, опухоли спинного мозга, глиомы ствола мозга, аденомы гипофиза, саркомы Капоши, эпидермоидного рака, плоскоклеточного рака, Т-клеточной лимфомы, злокачественных заболеваний, связанных с влиянием окружающей среды, включая злокачественные заболевания, вызванные асбестом, и комбинаций указанных типов злокачественных заболеваний.
4. Способ по п.1, где моноклональное антитело против PD-1 содержит:
а) вариабельную область тяжелой цепи CDR1 с последовательностью SEQ ID NO:15;
вариабельную область тяжелой цепи CDR2 с последовательностью SEQ ID NO:22;
вариабельную область тяжелой цепи CDR3 с последовательностью SEQ ID NO:29;
вариабельную область легкой цепи CDR1 с последовательностью SEQ ID NO:36;
вариабельную область легкой цепи CDR2 с последовательностью SEQ ID NO:43; и
вариабельную область легкой цепи CDR3 с последовательностью SEQ ID NO:50;
b) вариабельную область тяжелой цепи CDR1 с последовательностью SEQ ID NO:16;
вариабельную область тяжелой цепи CDR2 с последовательностью SEQ ID NO:23;
вариабельную область тяжелой цепи CDR3 с последовательностью SEQ ID NO:30;
вариабельную область легкой цепи CDR1 с последовательностью SEQ ID NO:37;
вариабельную область легкой цепи CDR2 с последовательностью SEQ ID NO:44; и
вариабельную область легкой цепи CDR3 с последовательностью SEQ ID NO:51;
c) вариабельную область тяжелой цепи CDR1 с последовательностью SEQ ID NO:17;
вариабельную область тяжелой цепи CDR2 с последовательностью SEQ ID NO:24;
вариабельную область тяжелой цепи CDR3 с последовательностью SEQ ID NO:31;
вариабельную область легкой цепи CDR1 с последовательностью SEQ ID NO:38;
вариабельную область легкой цепи CDR2 с последовательностью SEQ ID NO:45; и
вариабельную область легкой цепи CDR3 с последовательностью SEQ ID NO:52;
d) вариабельную область тяжелой цепи CDR1 с последовательностью SEQ ID NO:18;
вариабельную область тяжелой цепи CDR2 с последовательностью SEQ ID NO:25;
вариабельную область тяжелой цепи CDR3 с последовательностью SEQ ID NO:32;
вариабельную область легкой цепи CDR1 с последовательностью SEQ ID NO:39;
вариабельную область легкой цепи CDR2 с последовательностью SEQ ID NO:46; и
вариабельную область легкой цепи CDR3 с последовательностью SEQ ID NO:53;
e) вариабельную область тяжелой цепи CDR1 с последовательностью SEQ ID NO:19;
вариабельную область тяжелой цепи CDR2 с последовательностью SEQ ID NO:26;
вариабельную область тяжелой цепи CDR3 с последовательностью SEQ ID NO:33;
вариабельную область легкой цепи CDR1 с последовательностью SEQ ID NO:40;
вариабельную область легкой цепи CDR2 с последовательностью SEQ ID NO:47; и
вариабельную область легкой цепи CDR3 с последовательностью SEQ ID NO:54;
f) вариабельную область тяжелой цепи CDR1 с последовательностью SEQ ID NO:20;
вариабельную область тяжелой цепи CDR2 с последовательностью SEQ ID NO:27;
вариабельную область тяжелой цепи CDR3 с последовательностью SEQ ID NO:34;
вариабельную область легкой цепи CDR1 с последовательностью SEQ ID NO:41;
вариабельную область легкой цепи CDR2 с последовательностью SEQ ID NO:48; и
вариабельную область легкой цепи CDR3 с последовательностью SEQ ID NO:55; и
g) вариабельную область тяжелой цепи CDR1 с последовательностью SEQ ID NO:21;
вариабельную область тяжелой цепи CDR2 с последовательностью SEQ ID NO:28;
вариабельную область тяжелой цепи CDR3 с последовательностью SEQ ID NO:35;
вариабельную область легкой цепи CDRI с последовательностью SEQ ID NO:42;
вариабельную область легкой цепи CDR2 с последовательностью SEQ ID NO:49; и
вариабельную область легкой цепи CDR3 с последовательностью SEQ ID NO:56.
5. Способ по п.4, где моноклональное антитело против PD-1 выбрано из группы, состоящей из:
a) антитела, содержащего вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:1; и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:8;
b) антитела, содержащего вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:2; и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:9;
c) антитела, содержащего вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:3; и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:10;
d) антитела, содержащего вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:4; и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:11;
e) антитела, содержащего вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:5; и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:12; или
f) антитела, содержащего вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:6; и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:13; и
g) антитела, содержащего вариабельную область тяжелой цепи с аминокислотной последовательностью SEQ ID NO:7; и вариабельную область легкой цепи с аминокислотной последовательностью SEQ ID NO:14.
6. Способ по любому из пп.1-5, где моноклональное антитело против PD-1 является химерным, гуманизированным или антителом человека или его антигенсвязывающим фрагментом.
7. Способ по п.6, где моноклональное антитело против PD-1 является антителом человека или его антигенсвязывающим фрагментом.
8. Способ по п.6, где моноклональное антитело против PD-1 является антителом изотипа IgG1 или IgG4.
9. Способ по п.4 или 5, где моноклональное антитело против PD-1 является 5С4.
10. Способ усиления иммунного ответа у индивидуума, включающий введение индивидууму:
(a) моноклонального антитела против PD-1 и
(b) моноклонального антитела против CTLA-4 10D1,
где моноклональное антитело против PD-1 имеет следующие свойства:
(i) связывается с PD-1 человека со значением KD, равным 1·10-8 М или меньше;
(ii) не связывается ни с CD28 человека, ни с CTLA-4, ни с ICOS;
(iii) способно усиливать пролиферацию Т-клеток в анализе реакции лимфоцитов в смешанной культуре (MLR);
(iv) способно усиливать продукцию интерферона гамма в анализе MLR; и
(v) способно усиливать секрецию интерлейкина-2 (IL-2) в анализе MLR.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| US 2004175380 А1, 09.09.2004 | |||
| MARGALIT В | |||
| MOKYR, et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

