По данной заявке испрашивается приоритет предварительной заявки США № 61/026027, поданной 5 февраля 2008, и предварительной заявки США № 61/095429, поданной 9 сентября 2008, обе они приводятся в данный документ в качестве ссылки в полном объеме.
Область техники, к которой относится изобретение
Настоящее описание относится к антителам и их антигенсвязывающим участкам, которые связываются с α5β1. Описание также относится к молекулам нуклеиновых кислот, кодирующим такие антитела и антигенсвязывающие участки, способам создания антител и антигенсвязывающих участков против α5β1, композициям, содержащим данные антитела и антигенсвязывающие участки, и способам применения антител, антигенсвязывающих участков и композиций.
Уровень техники
Интегрин α5β1 представляет собой гетеродимерный белок клеточной поверхности, который связывается с фибронектином и вовлечен в прикрепление клетки и ангиогенез. Этот гетеродимер составлен из субъединицы α5 и субъединицы β1. Интегрин α5β1 обозначают как «классический рецептор фибронектина», который имеет ключевое значение в адгезии к матриксу, миграции, пролиферации, дифференциации и выживаемости. Несмотря на то, что с фибронектином (FN) связывается несколько интегринов, избирательным в отношении FN является интегрин α5β1, поскольку для распознавания лиганда и оптимального взаимодействия ему требуются обе пептидных последовательности на девятом (PHSRN) и десятом (RGDS) повторах типа III фибронектина FN (Danen et al. J. Biol. Chem. 270(37):21612-21618 (1995); Redisk et al. J. Cell Biol. 149(2):521-527 (2000); Takagi et al. EMBO J. 22:4607-4615 (2003)). Интегрин-опосредованная адгезия клетки к FN может запускать приток кальция, активировать тирозин- и серин/треонин протеинкиназы и инозитол-липидный метаболизм и регулировать активность семейства небольших GTP-аз Rho, которое контролирует актиновый цитоскелет и клеточный цикл.
Экспрессия α5β1 наблюдается в большинстве эмбриональных тканей, но после рождения уровень падает в соответствии с терминальной дифференциацией клетки (Muschler & Horwitz Development 113(1):327-337 (1991)). У взрослых мышей дикого типа экспрессия происходит главным образом в сосудистой и соединительной ткани, хотя низкие уровни рецептора широко распространены. Экспрессия как α5β1, так и FN существенно и согласованно повышается на кровеносных сосудах опухолей человека и в тканях, стимулированных факторами роста и цитокинами.
Ангиогенные цитокины, такие как bFGF, VEGF, IL-8, TGF-бета и TNF-α, повышают экспрессию α5β1 на эндотелиальных клетках in-vitro и in-vivo, в то время как в нормальных сосудах и тканях человека эти молекулы экспрессируются в минимальном количестве (Kim et al. Am. J. Path. 156(4):1345-62 (2000); Enaida et al. Fukushima J. Med. Sci. 44(1):43-52 (1998); Klein et al. Mol. Biol. Cell 4(10):973-982 (1993)).
Высокие уровни экспрессии α5β1 не ограничиваются сосудистой системой, поскольку также часто обнаруживают, что экспрессируют α5β1 опухолевые клетки при многих типах рака. Гипоксия опухоли ассоциировалась с повышенной экспрессией α5β1 опухолью (Mousa et al. J. Cell. Biochem. 74:135-143 (1999)). Полагают, что интегрины важны для интравазации опухоли во вновь образованные капилляры и экстравазации в удаленные участки, приводя к распространению опухоли и метастазированию. В целом, паттерн экспрессии α5β1 согласуется с большим числом значений в прогрессировании рака через опосредование активностей как стромальных, так и опухолевых клеток. В большом числе клинических исследований повышение α5β1 на клетках опухоли ассоциировалось с прогрессированием меланомы, оральной плоскоклеточной карциномы и В-клеточных лейкозов человека (Jin & Varner, Br. J. Cancer 90:561-565 (2004); Danen et al. Histopathology 24(3):249-256 (1994); Shinohara et al., Am. J. Clin. Pathol. 111(1):75-88 (1999)).
Сущность изобретения
Настоящее описание относится к выделенным моноклональным антителам, в частности моноклональным антителам человека, у которых наблюдается высокое сродство связывания с интегрином α5β1 человека. Антитела, описанные в настоящем описании, как правило, представляют собой антитела человека, хотя в альтернативных случаях антитела могут представлять собой антитела мыши, химерные антитела или гуманизированные антитела.
В одном из аспектов описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, где антитело: (а) связывается с интегрином α5β1 человека с KD 1×10-7 M или меньше; и (b) способно индуцировать антителозависимую клеточную цитотоксичность. Например, в одном случае антитело принадлежит подклассу, который способен индуцировать ADCC, такому как IgG1 или IgG3. В другом случае ADCC-активность является результатом введения в Fc-область углеводных модификаций, таких как измененные паттерны гликозилирования, по сравнению с нативным углеводным паттерном.
В некоторых случаях антитело связывается с интегрином α5β1 человека с KD 5×10-8 M или меньше, 2×10-8 M или меньше, 1×10-8 M или меньше, 5×10-9 M или меньше, 4×10-9 M или меньше, 3×10-9 M или меньше, или 2,7×10-9 M или меньше.
В дополнительном аспекте настоящее описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, где антитело: (а) связывается с интегрином α5β1 человека с KD 1×10-7 M или меньше; и (b) у него наблюдается повышенная ADCC-активность относительно антитела сравнения. Например, в одном из случаев антитело с повышенной ADCC-активностью содержит, по меньшей мере, одну мутацию в Fc-области по сравнению с Fc-областью дикого типа, и повышенной ADCC-активность является по отношению к тому же антителу, но содержащему Fc-область дикого типа. В некоторых примерах антитело связывается с интегрином α5β1 человека с KD 5×10-8 M или меньше, 2×10-8 M или меньше, 1×10-8 M или меньше, 5×10-9 M или меньше, 4×10-9 M или меньше, 3×10-9 M или меньше, или 2,7×10-9 M или меньше. В дополнительных примерах у антитела наблюдается повышенная ADCC-активность относительно антитела сравнения, которая повышена, по меньшей мере, в 1,1 раз, по меньшей мере, в 1,2 раз, по меньшей мере, в 1,3 раз, по меньшей мере, в 1,5 раз, по меньшей мере, в 2 раза, по меньшей мере, в 3 раза, по меньшей мере, в 5 раз, по меньшей мере, в 10 раз, по меньшей мере, в 20 раз, по меньшей мере, в 30 раз, по меньшей мере, в 40 раз, по меньшей мере, в 50 раз, по меньшей мере, в 100 раз, по меньшей мере, в 200 раз, по меньшей мере, в 500 раз, или, по меньшей мере, в 1000 раз по отношению к антителу сравнения, где антитело сравнения представляет собой то же антитело, но с Fc-областью дикого типа.
В дополнительном аспекте настоящее описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, где антитело: (а) связывается с интегрином α5β1 человека с KD 1×10-7 M или меньше; и (b) содержит, по меньшей мере, одну мутацию в Fc-области по сравнению с Fc-областью дикого типа. Например, в одном из случаев антитело принадлежит к подклассу IgG1, и, по меньшей мере, одна из аминокислот в Fc-области подкласса IgG1 мутирована. В дополнительном примере, по меньшей мере, одна мутация находится в позиции серин 247, аланин 338 или изолейцин 340 в Fc-области подкласса IgG1. В дополнительном примере, по меньшей мере, одну мутацию выбирают из группы, состоящей из S247D, A338L и I340E. В еще одном дополнительном примере антитело содержит мутации S247D, A338L и I340E.
В дополнительном аспекте настоящее описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, где антитело перекрестно конкурирует или конкурирует за связывание с интегрином α5β1 человека с антителом, которое содержит: (а) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:7, или ее консервативные модификации; и (b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8, или ее консервативные модификации.
Дополнительный аспект настоящего описания представляет собой выделенное моноклональное антитело или его антигенсвязывающий участок, содержащие вариабельную область тяжелой цепи, которая представляет собой продукт или производное гена человека VH 4-39, где антитело специфически связывается с интегрином α5β1 человека.
Дополнительный аспект настоящего описания представляет собой выделенное моноклональное антитело или его антигенсвязывающий участок, содержащий вариабельную область легкой цепи, которая представляет собой продукт или производное гена человека VK L6, где антитело специфически связывается с интегрином α5β1 человека.
В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR1, содержащую последовательность SEQ ID NO:1, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR2, содержащую SEQ ID NO:2, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR3, содержащую SEQ ID NO:3, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR1, содержащую SEQ ID NO:4, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR2, содержащую SEQ ID NO:5, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR3, содержащую SEQ ID NO:6, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему:
(а) вариабельную область тяжелой цепи CDR1, содержащую SEQ ID NO:1, или ее консервативные модификации;
(b) вариабельную область тяжелой цепи CDR2, содержащую SEQ ID NO:2, или ее консервативные модификации;
(c) вариабельную область тяжелой цепи CDR3, содержащую SEQ ID NO:3, или ее консервативные модификации;
(d) вариабельную область легкой цепи CDR1, содержащую SEQ ID NO:4, или ее консервативные модификации;
(e) вариабельную область легкой цепи CDR2, содержащую SEQ ID NO:5, или ее консервативные модификации; и
(f) вариабельную область легкой цепи CDR3, содержащую SEQ ID NO:6, или ее консервативные модификации.
В дополнительном аспекте настоящее описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:7, или ее консервативные модификации. Например, описание относится к антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:7. В дополнительном аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8, или ее консервативные модификации. Например, описание относится к антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:8. В дополнительном аспекте настоящее описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:7, или ее консервативные модификации, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8, или ее консервативные модификации. Например, описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:7, и вариабельную область легкой цепи, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:8.
В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR1, содержащую последовательность SEQ ID NO:13, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR2, содержащую последовательность SEQ ID NO:14, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR3, содержащую последовательность SEQ ID NO:15, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR1, содержащую последовательность SEQ ID NO:16, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR2, содержащую последовательность SEQ ID NO:17, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR3, содержащую последовательность SEQ ID NO:18, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему:
(а) вариабельную область тяжелой цепи CDR1, содержащую SEQ ID NO:13, или ее консервативные модификации;
(b) вариабельную область тяжелой цепи CDR2, содержащую SEQ ID NO:14, или ее консервативные модификации;
(c) вариабельную область тяжелой цепи CDR3, содержащую SEQ ID NO:15, или ее консервативные модификации;
(d) вариабельную область легкой цепи CDR1, содержащую SEQ ID NO:16, или ее консервативные модификации;
(e) вариабельную область легкой цепи CDR2, содержащую SEQ ID NO:17, или ее консервативные модификации; и
(f) вариабельную область легкой цепи CDR3, содержащую SEQ ID NO:18, или ее консервативные модификации.
В дополнительном аспекте настоящее описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:19, или ее консервативные модификации. В дополнительном аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:20, или ее консервативные модификации. В дополнительном аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:19, или ее консервативные модификации, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:20, или ее консервативные модификации.
В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR1, содержащую SEQ ID NO:23, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR2, содержащую SEQ ID NO:24, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR3, содержащую SEQ ID NO:25, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR1, содержащую SEQ ID NO:26, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR2, содержащую SEQ ID NO:27, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR3, содержащую SEQ ID NO:28, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему:
(а) вариабельную область тяжелой цепи CDR1, содержащую SEQ ID NO:23, или ее консервативные модификации;
(b) вариабельную область тяжелой цепи CDR2, содержащую SEQ ID NO:24, или ее консервативные модификации;
(c) вариабельную область тяжелой цепи CDR3, содержащую SEQ ID NO:25, или ее консервативные модификации;
(d) вариабельную область легкой цепи CDR1, содержащую SEQ ID NO:26, или ее консервативные модификации;
(e) вариабельную область легкой цепи CDR2, содержащую SEQ ID NO:27, или ее консервативные модификации; и
(f) вариабельную область легкой цепи CDR3, содержащую SEQ ID NO:28, или ее консервативные модификации.
В дополнительном аспекте настоящее описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:29, или ее консервативные модификации. В дополнительном аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:30, или ее консервативные модификации. В дополнительном аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:29, или ее консервативные модификации, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:30, или ее консервативные модификации.
В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR1, содержащую SEQ ID NO:33, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR2, содержащую SEQ ID NO:34, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи CDR3, содержащую SEQ ID NO:35, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR1, содержащую SEQ ID NO:36, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR2, содержащую SEQ ID NO:37, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи CDR3, содержащую SEQ ID NO:38, или ее консервативные модификации. В другом аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему: (а) вариабельную область тяжелой цепи CDR1, содержащую SEQ ID NO:33, или ее консервативные модификации; (b) вариабельную область тяжелой цепи CDR2, содержащую SEQ ID NO:34, или ее консервативные модификации; (c) вариабельную область тяжелой цепи CDR3, содержащую SEQ ID NO:35, или ее консервативные модификации; (d) вариабельную область легкой цепи CDR1, содержащую SEQ ID NO:36, или ее консервативные модификации; (e) вариабельную область легкой цепи CDR2, содержащую SEQ ID NO:37, или ее консервативные модификации; и (f) вариабельную область легкой цепи CDR3, содержащую SEQ ID NO:38, или ее консервативные модификации.
В дополнительном аспекте настоящее описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:39, или ее консервативные модификации. В дополнительном аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:40, или ее консервативные модификации. В дополнительном аспекте описание относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:39, или ее консервативные модификации, и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:40, или ее консервативные модификации.
В дополнительном аспекте предлагается выделенное антитело или его антигенсвязывающий участок, которые связываются с тем же эпитопом на интегрине α5β1 человека, что и любое антитело, описанное в данном документе, и/или конкурируют за связывание с интегрином α5β1 человека с таким антителом.
В одном из вариантов осуществления описание относится к материалу, депонированному в АТСС под номером доступа РТА-9377. В другом варианте осуществления описание относится к материалу, депонированному в АТСС под номером доступа РТА-9378. В другом варианте осуществления описание относится к выделенному антителу, которое содержит вариабельную область тяжелой цепи, депонированную в АТСС под номером доступа РТА-9377. В другом варианте осуществления описание относится к выделенному антителу, которое содержит вариабельную область легкой цепи, депонированную в АТСС под номером доступа РТА-9378. В другом варианте осуществления описание относится к выделенному антителу, содержащему вариабельную область тяжелой цепи, депонированную в АТСС под номером доступа РТА-9377, но у которого в VH-области получены мутации зародышевой линии I30S и N33S. В дополнительном варианте осуществления описание относится к выделенному антителу, которое содержит вариабельные области тяжелой цепи и легкой цепи, депонированные в АТСС под номерами доступа РТА-9377 и РТА-9378, соответственно, или указанному антителу, у которого в VH-области созданы мутации зародышевой линии I30S и N33S. В дополнительном варианте осуществления описание относится к выделенному антителу, которое содержит области CDR1, CDR2 и CDR3 тяжелой цепи вариабельной области тяжелой цепи, депонированной в АТСС под номером доступа РТА-9377, или содержит указанные области CDR1, CDR2 и CDR3, если указанная вариабельная область тяжелой цепи содержит мутации зародышевой линии I30S и N33S. В дополнительном варианте осуществления описание относится к выделенному антителу, которое содержит области CDR1, CDR2 и CDR3 легкой цепи вариабельной области легкой цепи, депонированной в АТСС под номером доступа РТА-9378. В дополнительном варианте осуществления описание относится к выделенному антителу, которое содержит области CDR1, CDR2 и CDR3 легкой цепи и области CDR1, CDR2 и CDR3 тяжелой цепи вариабельных областей тяжелой и легкой цепей, депонированных в АТСС под номерами доступа РТА-9378 и РТА-9378, соответственно, или содержит указанные области CDR1, CDR2 и CDR3, если указанная вариабельная область тяжелой цепи содержит мутации зародышевой линии I30S и N33S.
Антитела по изобретению могут представлять собой, например, полноразмерные антитела, например, подкласса IgG1 или IgG4. Альтернативно, антитела могут представлять собой фрагменты антител, такие как Fab- или Fab'2-фрагменты, или одноцепочечные антитела. В одном из случаев настоящее описание относится к любому антителу, описанному выше, которое представляет собой полноразмерное антитело человека подкласса IgG1, в котором, по меньшей мере, одна аминокислота в Fc-области подкласса IgG1 мутирована. В дополнительном случае, по меньшей мере, одна мутация наблюдается в позиции серин 247, аланин 338 или изолейцин 340. В дополнительном случае, по меньшей мере, одну мутацию выбирают из группы, состоящей из S247D, A338L и I340E. В еще одном дополнительном случае антитело содержит мутации S247D, A338L и I340E.
В дополнительном аспекте описание относится к выделенному моноклональному антителу, содержащему тяжелую цепь, представленную в SEQ ID NO:9, или ее консервативные модификации. Например, антитело содержит тяжелую цепь, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:9. В дополнительном аспекте описание относится к выделенному моноклональному антителу, содержащему легкую цепь, представленную в SEQ ID NO:10, или ее консервативные модификации. Например, антитело содержит легкую цепь, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:10. В дополнительном аспекте описание относится к выделенному моноклональному антителу, содержащему тяжелую цепь, представленную в SEQ ID NO:9, или ее консервативные модификации и легкую цепь, представленную в SEQ ID NO:10, или ее консервативные модификации. Например, антитело содержит тяжелую цепь, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:9, и легкую цепь, которая, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98%, или, по меньшей мере, на 99% идентична аминокислотной последовательности, приведенной в SEQ ID NO:10.
В некоторых вариантах осуществления С-концевой лизин тяжелой цепи какого-либо описанного антитела против α5β1 по изобретению расщепляют и, таким образом, он не представлен. Например, в некоторых вариантах осуществления антитела по настоящему изобретению содержат константную область тяжелой цепи, представленную в SEQ ID NO:43, но где С-концевой лизин не представлен. В большом числе случаев тяжелая и легкая цепи антител против α5β1 необязательно могут включать сигнальную последовательность.
В дополнительном аспекте описание относится к композиции, содержащей любое из антител или их антигенсвязывающих участков, описанных в данном документе, и фармацевтически приемлемый носитель.
В дополнительном аспекте описание относится к иммуноконъюгату, содержащему любое из антител или их антигенсвязывающих участков, описанных в данном документе, связанное с терапевтическим агентом. В одном из случаев терапевтический агент представляет собой цитотоксин или радиоактивный изотоп. В дополнительном аспекте описание относится к композиции, содержащей любой иммуноконъюгат, описанный в данном документе, и фармацевтически приемлемый носитель. Описание также относится к биспецифической молекуле, содержащей антитело или его антигенсвязывающий участок, связанные со вторым функциональным фрагментом, обладающим специфичностью связывания, отличной от специфичности связывания указанного антитела или его антигенсвязывающего участка.
Также предлагаются композиции, содержащие антитело или его антигенсвязывающий участок или иммуноконъюгат, или биспецифическую молекулу по изобретению и фармацевтически приемлемый носитель.
Также изобретением охватываются молекулы нуклеиновых кислот, кодирующие антитела или их антигенсвязывающие участки, а также экспрессирующие векторы, содержащие такие нуклеиновые кислоты, и клетки-хозяева, содержащие такие экспрессирующие векторы. Один из аспектов, например, представляет собой выделенную молекулу нуклеиновой кислоты или экспрессирующий вектор, содержащие последовательность, представленную в SEQ ID NO:11, или ее консервативные модификации. Дополнительный аспект представляет собой выделенную молекулу нуклеиновой кислоты или экспрессирующий вектор, содержащие последовательность, представленную в SEQ ID NO:12, или ее консервативные модификации. Дополнительный аспект представляет собой выделенную молекулу нуклеиновой кислоты или экспрессирующий вектор, содержащие последовательность, выбранную из группы, состоящей из SEQ ID NO:21, 22, 31, 32, 41 и 42, или ее консервативные модификации. Описание также относится к трансгенной мыши, содержащей трансгены тяжелой и легкой цепей иммуноглобулина человека, где мышь экспрессирует антитело по изобретению, а также гибридомам, полученным из такой мыши, где гибридома продуцирует антитело по изобретению.
Описание дополнительно относится к клетке-хозяину, содержащей любой из экспрессирующих векторов, описанных в данном документе.
В дополнительном аспекте описание относится к способу получения антитела против интегрина α5β1, который включает экспрессию антитела в любой клетке-хозяине, описанной в данном документе, и выделение антитела из клетки-хозяина.
В дополнительном аспекте описание относится к способу ингибирования роста опухолевых клеток, экспрессирующих интегрин α5β1, включающему приведение клеток в контакт с антителом или его антигенсвязывающим участком, которые связываются с интегрином α5β1 человека с KD 1×10-7 M или меньше и способны индуцировать антителозависимую клеточную цитотоксичность. В одном из случаев антитело представляет собой полностью антитело человека. В дополнительном случае антитело или его антигенсвязывающий участок создают так, чтобы усилить его способность индуцировать антителозависимую клеточную цитотоксичность. В еще одном дополнительном случае усиление достигается за счет мутации, по меньшей мере, одного аминокислотного остатка в Fc-области.
В дополнительном аспекте изобретение относится к способу ингибирования роста опухолевых клеток, экспрессирующих интегрин α5β1, включающему приведение клеток в контакт с любым из антител или их антигенсвязывающих участков, описанных в данном документе, в количестве, эффективном для ингибирования роста опухолевых клеток.
В дополнительном аспекте изобретение относится к применению любого из антител или их антигенсвязывающих участков, описанных в данном документе, для производства лекарственного препарата для лечения аномального клеточного роста. В еще одном дополнительном аспекте изобретение относится к любому из антител или их антигенсвязывающих участков, описанных в данном документе, для применения в лечении и/или диагностике аномального роста клеток. В одном из вариантов осуществления данного способа аномальный рост клеток представляет собой рак, включая, но ими не ограничиваясь, мезотелиому, рак гепатобилиарной системы (печени и желчевыводящих путей), первичную или вторичную опухоль ЦНС, первичную или вторичную опухоль мозга, рак легких (NSCLC и SCLC), рак костей, рак поджелудочной железы, рак кожи, рак головы или шеи, меланому кожи или внутриглазную меланому, рак яичников, рак толстого кишечника, рак прямой кишки, анальный рак, рак желудка, рак органов желудочно-кишечного тракта (желудка, колоректальный и 12 перстной кишки), рак молочных желез, рак матки, карциному фаллопиевых труб, карциному эндометрия, карциному цервикального канала, карциному вагины, карциному вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечников, саркому мягких тканей, рак мочеиспускательного канала, рак пениса, рак простаты, рак яичка, хронический или острый лейкоз, хронический миелоидный лейкоз, лимфоцитарные лимфомы, рак мочевого пузыря, рак почки или мочеточника, почечно-клеточную карциному, карциному почечной лоханки, новообразования центральной нервной системы (ЦНС), первичную лимфому ЦНС, неходжкинскую лимфому, опухоли спинного мозга, глиому ствола мозга, аденому гипофиза, адренокортикальный рак, рак желчного пузыря, множественную миелому, холангиокарциному, фибросаркому, нейробластому, ретинобластому или сочетание одной или нескольких вышеуказанных злокачественных опухолей.
Описание также относится к способам создания «второго поколения» антител против α5β1 на основании последовательностей антител против α5β1, предложенных в данном документе. Например, описание относится к способу получения антитела против α5β1, содержащему: (а) получение: (i) последовательности вариабельной области тяжелой цепи антитела, содержащей последовательность CDR1, представленную в SEQ ID NO:1, 13, 23 или 33, последовательность CDR2, представленную в SEQ ID NO:2, 14, 24 или 34, и/или последовательность CDR3, представленную в SEQ ID NO:3, 15, 25 или 35; и/или (ii) последовательности вариабельной области легкой цепи антитела, содержащей последовательность CDR1, представленную в SEQ ID NO:4, 16, 26 или 36, последовательность CDR2, представленную в SEQ ID NO:5, 17, 27 или 37, и/или последовательность CDR3, представленную в SEQ ID NO:6, 18, 28 или 38; (b) изменение, по меньшей мере, одного аминокислотного остатка в последовательности вариабельной области тяжелой цепи антитела и/или последовательности вариабельной области легкой цепи антитела для создания, по меньшей мере, одной измененной последовательности антитела; и (с) экспрессию измененной последовательности антитела в виде белка.
Другие особенности и преимущества настоящего описания будут понятны из следующего подробного описания и примеров, которые не должны быть истолкованы как ограничивающие. Содержание всех ссылок, данных Генбанка, патентов и опубликованных патентных заявок, процитированных в данном описании изобретения, прямо приводится в данный документ в качестве ссылок.
Краткое описание чертежей
На фиг.1А представлена последовательность ДНК вариабельной области тяжелой цепи антитела 22В5 (SEQ ID NO:11); на фиг.1В представлена аминокислотная последовательность вариабельной области тяжелой цепи антитела 22В5 (SEQ ID NO:7) - CDR-области подчеркнуты; на фиг.1С представлена последовательность ДНК вариабельной области легкой цепи антитела 22В5 (SEQ ID NO:12); на фиг.1D представлена аминокислотная последовательность вариабельной области легкой цепи антитела 22В5 (SEQ ID NO:8) - CDR-области подчеркнуты; на фиг.1Е представлена последовательность ДНК вариабельной области тяжелой цепи антитела 24С7 (SEQ ID NO:21); на фиг.1F представлена аминокислотная последовательность вариабельной области тяжелой цепи антитела 24С7 (SEQ ID NO:19) - CDR-области подчеркнуты; на фиг.1G представлена последовательность ДНК вариабельной области легкой цепи антитела 24С7 (SEQ ID NO:22); на фиг.1Н представлена аминокислотная последовательность вариабельной области легкой цепи антитела 24С7 (SEQ ID NO:20) - CDR-области подчеркнуты; на фиг.1I представлена последовательность ДНК вариабельной области тяжелой цепи антитела 1D9 (SEQ ID NO:31); на фиг.1J представлена аминокислотная последовательность вариабельной области тяжелой цепи антитела 1D9 (SEQ ID NO:29) - CDR-области подчеркнуты. На фиг.1K представлена последовательность ДНК вариабельной области легкой цепи антитела 1D9 (SEQ ID NO:32); на фиг.1L представлена аминокислотная последовательность вариабельной области легкой цепи антитела 1D9 (SEQ ID NO:30) - CDR-области подчеркнуты; на фиг.1М представлена последовательность ДНК вариабельной области тяжелой цепи антитела 2D2 (SEQ ID NO:41); на фиг.1N представлена аминокислотная последовательность вариабельной области тяжелой цепи антитела 2D2 (SEQ ID NO:39) - CDR-области подчеркнуты; на фиг.1О представлена последовательность ДНК вариабельной области легкой цепи антитела 2D2 (SEQ ID NO:42); на фиг.1Р представлена аминокислотная последовательность вариабельной области легкой цепи антитела 2D2 (SEQ ID NO:40) - CDR-области подчеркнуты; на фиг.1Q представлена аминокислотная последовательность константной области тяжелой цепи IgG1 с подчеркнутыми мутациями S247D, A338L и I340E; и на фиг.1R представлена аминокислотная последовательность константной области легкой цепи IgG1.
На фиг.2 представлено выравнивание вариабельного домена тяжелой цепи (VH) антитела 22В5 с соответствующей последовательностью зародышевой линии. Кроме того, представлено выравнивание вариабельного домена легкой цепи (VK) антитела 22В5 с соответствующей последовательностью зародышевой линии. CDR-области подчеркнуты, идентичные остатки представлены пунктиром, а точки указывают на делеции.
На фиг.3 представлено наложение сенсограмм, полученных введением большого числа концентраций рекомбинантного внеклеточного домена α5β1 поверх иммобилизованного 22В5/DLE. Эти данные получали в присутствии 4,0 мМ CaCl2. Порядок введений был от низкой к высокой концентрации.
На фиг.4 посредством FACS показано дозо-зависимое связывание антитела 22В5/DLE с клетками HUVEC.
На фиг.5 представлено сравнение равновесных констант диссоциации для Fcγ-рецепторов человека и мыши. «wt» обозначает IgG1 дикого типа 22B5; «DLE» обозначает 22В5/DLE.
На фиг.6 представлены результаты анализа блокады адгезии клеток HUVEC. Результаты указывают уровень ингибирования адгезии клеток HUVEC к фибронектину для антитела 22В5 и большого числа вариантов подкласса, а также отрицательного контроля (ВНА2 IgG1). Также представлены рассчитанные величины IC50.
На фиг.7 представлена экспрессия α5 клеток HUVEC и 20 опухолевых клеточных линий, измеренная вестерн-блотом.
На фиг.8 представлена in-vitro ADCC, индуцированная антителом 22В5/DLE по сравнению с IgG1 wt 22B5. На фиг.8A представлен анализ на основе детекции LDH, измеряющий ADCC клеток U87MG в присутствии клеток РВМС человека посредством 22В5/DLE и 22В5 wt IgG1. На фиг.8В представлен анализ на основе детекции ToxiLight, измеряющий ADCC клеток HUVEC в присутствии клеток РВМС человека посредством 22В5/DLE и 22В5 wt IgG1.
На фиг.9 представлены анализы на основе определения LDH, которые указывают на существенное усиление ADCC при использовании антитела 22В5/DLE по сравнению с антителом wt 22В5 IgG1 через широкий диапазон уровней экспрессии антигена.
На фиг.10 представлена ингибирующая активность антитела 22В5/DLE на модели экспериментального метастаза А549-Luc. Фиг.10А: величина метастаза в легкое, измеренная с помощью BLI на 8 неделе (n=11 для контрольной группы, n=14 для группы 22В5 IgG2 и n=12 для группы 22В5/DLE). Фиг.10В: возобновление роста опухолей легких в группе, обработанной 22В5 IgG2, после прекращения лечения. Для сравнения, в группе, обработанной 22В5/DLE, наблюдалось незначительное возобновление роста. Фиг.10С: График Каплана-Мейера степени выживаемости животных из каждой обработанной группы (конечная точка = BLI 1×108 фотонов/секунда), р<0,0001 при сравнении контрольной группы, обработанной носителем, со всеми другими группами, и р<0,05 при сравнении между группами 22В5/DLE и 22В5 IgG2.
На фиг.11 посредством анализа FACS представлено дозо-зависимое связывание антител 1D9, 1D9/DLE, 24C7/DLE, 2D2/DLE, 22B5 и 22В5/DLE с клетками HUVEC.
На фиг.12 представлена in-vitro ADCC, индуцированная антителами 1D9, 1D9/DLE, 2D2/DLE, 22В5/DLE и 24С7/ DLE.
На фиг.13 представлена ADCC-зависимая противоопухолевая эффективность антитела 1D9/DLE на сингенной модели метастатической меланомы. Фиг.13А: Макроскопическая картина легких, резецированных из всех групп. Фиг.13В: Количественная оценка веса легкого (*р<0,05 при сравнении 1D9 IgG1 DLE с 1D9 IgG2). Фиг.13С: Количественная оценка числа метастатических колоний, видимых на поверхности легкого. Статистический анализ посредством дисперсионного анализа ANOVA и теста множественного сравнения Бонферрони (*p<0,05, 1D9 IgG1 DLE по сравнению с 1D9 IgG2; *p<0,05, 1D9 IgG1 DLE по сравнению с IgG2 против KLH).
Подробное описание
Настоящее описание относится к выделенным моноклональным антителам, в частности, моноклональным антителам человека, которые специфически связываются с α5β1 с высоким сродством. В некоторых случаях антитела по изобретению происходят из конкретных последовательностей тяжелой и легкой цепей зародышевой линии и/или содержат определенные структурные особенности, такие как CDR-области, содержащие конкретные аминокислотные последовательности. Описание относится к выделенным антителам, способам получения таких антител, иммуноконъюгатам и биспецифическим молекулам, содержащим такие антитела, и фармацевтическим композициям, содержащим антитела, иммуноконъюгаты и биспецифические молекулы по изобретению. Описание также относится к способам применения антител, таким как детекция α5β1, а также лечение заболеваний, ассоциированных с экспрессией α5β1, таких как аномальный клеточный рост (например, рак). Соответственно, описание также относится к способам применения антител против α5β1 или их антигенсвязывающих участков для лечения большого числа видов аномального клеточного роста, такого как рак.
С целью облегчения понимания настоящего описания некоторым терминам сначала дается определение. Дополнительные определения приводятся на протяжении подробного описания.
Если иначе не определяется в данном документе, научные и технические термины, используемые в связи с настоящим описанием, должны иметь значения, которые обычно понимаются специалистами в данной области. Кроме того, если контекстом не подразумевается иного, термины в единственном числе должны включать множественное число и термины во множественном числе должны включать единственное число. В целом, терминология, использованная в отношении культивирования клеток и тканей, молекулярной биологии, иммунологии, микробиологии, генетики и химии белков и нуклеиновых кислот и гибридизации, описанных в данном документе, и методики их проведения хорошо известны и широко используются в данной области.
Если иного не указано, способы и техники по настоящему описанию, как правило, осуществляют в соответствии со способами, хорошо известными в данной области и описанными в большом числе общих и более конкретных ссылок, которые приводятся и обсуждаются в настоящем описании изобретения. К таким ссылкам относятся, например, Sambrook and Russell, Molecular Cloning, A Laboratory Approach, Cold Spring Harbor Press, Cold Spring Harbor, NY (2001), Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, NY (2002), and Harlow and Lane Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1990). Ферментативные реакции и методики очистки осуществляют в соответствии с требованиями производителя, как обычно осуществляют в данной области или, как описано в данном документе. Терминология, использованная в отношении аналитической химии, химии органического синтеза и медицинской и фармацевтической химии, описанных в данном документе, и их лабораторные способы и техники хорошо известны и широко используются в данной области. Для химического синтеза, химического анализа, получения, смешивания и доставки лекарственного средства, и лечения пациентов используются стандартные методики.
Используемый в данном документе каждый из следующих терминов имеет значение, ассоциированное с ним в этом разделе.
Как используется в данном документе, для двадцати стандартных аминокислот и их аббревиатур придерживаются принятого употребления. См. Immunology-A Synthesis (2nd Edition, E.S. Golub and D.R. Gren, Eds., Sinauer Associates, Sunderland, Mass. (1991)).
Термины «α5β1» и «интегрин α5β1» используются взаимозаменяемо и включают варианты, изоформы и межвидовые гомологи интегрина α5β1 человека. Нативный α5β1 человека, например, составлен из субъединицы α5 (которая происходит из последовательности предшественника, впоследствии расщепляющейся на две цепи, соединенные дисульфидным мостиком (номер доступа в Генбанке Р08648) и субъединицы β1 (которая происходит из последовательности предшественника, впоследствии процессирующей в зрелую форму) (номер доступа в Генбанке Р05556-1). Известно, что субъединица β1 существует в виде нескольких изоформ, продуцируемых за счет альтернативного сплайсинга (см., например, номера доступа в Генбанке Р05556-2, Р05556-3, Р05556-4 и Р05556-5). Антитела человека против α5β1 по изобретению могут в некоторых случаях перекрестно взаимодействовать с α5β1 видов, отличных от человека. В других случаях антитела могут быть полностью специфичны в отношении α5β1 человека, и может не наблюдаться межвидовой перекрестной реактивности или перекрестной реактивности других типов.
К «иммунному ответу», как должно быть понятно специалисту в данной области, относятся, но ими не ограничиваясь, любая детектируемая антиген-специфическая или аллогенная активация Т-хелперного или цитотоксического Т-клеточного ответа, продукция антител, опосредованная Т-клетками активация аллергических реакций и тому подобное. Термин охватывает действие, например, лимфоцитов, антиген-презентирующих клеток, фагоцитарных клеток, гранулоцитов и растворимых макромолекул, продуцируемых вышеуказанными клетками или печенью (включая антитела, цитокины и комплемент), что приводит к селективному повреждению, деструкции или устранению из организма человека внедрившихся патогенов, клеток или тканей, инфицированных патогенами, раковых клеток, или, в случаях аутоиммунного ответа или патологического воспаления, нормальных клеток или тканей человека.
Под «путем сигнальной трансдукции» понимают биохимическое взаимодействие между большим числом молекул сигнальной трансдукции, которые имеют значение в переносе сигнала из одной части клетки в другую часть клетки. К используемой в данном документе фразе «рецептор клеточной поверхности» относятся, например, молекулы и комплексы молекул, способные получать сигнал и переносить такой сигнал через плазматическую мембрану клетки. Примером «рецептора клеточной поверхности» по настоящему изобретению является интегрин α5β1.
Под термином «антитело», как изложено в данном документе, понимают целые антитела и любой их антигенсвязывающий фрагмент (т.е. «антигенсвязывающий участок») или одиночные цепи. Под «антителом» понимают гликопротеин, содержащий, по меньшей мере, две тяжелых (Н) цепи и две легких (L) цепи, соединенные между собой дисульфидными связями, или его антигенсвязывающий участок. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (обозначаемая в данном документе как VH) и константную область тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов СН1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (обозначаемой в данном документе как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена CL. Области VH и VL могут быть дополнительно подразделены на области гипервариабельности, называемые областями, определяющими комплементарность (CDR), перемежающиеся с областями, которые более консервативны, называемыми каркасными областями (FR). Каждый VH и VL составлен из трех областей CDR и четырех областей FR, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями хозяина или факторами, включающими большое число клеток иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента. Внутри легкой и тяжелой цепей вариабельные и константные области соединены областью «J», состоящей из около 12 или более аминокислот, причем тяжелая цепь также включает область «D», состоящую из около 10 или более аминокислот. См. в целом руководство Fundamental Immunology Ch. 7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989)).
Под термином «антигенсвязывающий участок» антитела (или просто «участок антитела»), используемым в данном документе, понимают один или несколько фрагментов антитела, которые сохраняют способность специфически связываться с антигеном (например, интегрином α5β1). Было показано, что антигенсвязывающую функцию антитела могут выполнять фрагменты полноразмерного антитела. К примерам связывающих фрагментов, охватываемых термином «антигенсвязывающий участок» антитела, относятся (i) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и СН1; (ii) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, соединенных дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и СН1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела; (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH; и (vi) выделенная область, определяющая комплементарность (CDR). Более того, несмотря на то, что два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, они могут быть соединены, используя рекомбинантные техники, с помощью синтетического линкера, который позволит создать их в виде единой белковой цепи, в которой области VL и VH составляют пару с образованием моновалентных молекул (известных как одноцепочечные Fv (scFv); см., например, статьи Bird et al. (1988) Science 242:423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Такие одноцепочечные антитела также охватываются термином «антигенсвязывающий участок» антитела. Эти фрагменты антитела могут быть получены, используя любую подходящую технику, включая принятые техники, известные специалистам в данной области, и фрагменты могут быть скринированы на пригодность таким же образом как интактные антитела.
Под «выделенным антителом», используемым в данном документе, понимают антитело, которое по существу свободно от других антител, обладающих отличными антигенными специфичностями (например, выделенное антитело, которое специфически связывается с α5β1, практически свободно от антител, которые специфически связываются с антигенами, отличными от α5β1). Выделенное антитело, которое специфически связывается с α5β1, может, тем не менее, обладать перекрестной реактивностью с другими антигенами, такими как молекулы α5β1 других видов. Более того, выделенное антитело может быть, по существу, свободно от другого клеточного материала и/или химических веществ.
Под терминами «моноклональное антитело» или «композиция моноклонального антитела», используемыми в данном документе, понимают препарат молекул антитела одного молекулярного состава. У композиции моноклонального антитела наблюдается единственная специфичность связывания и сродство к конкретному эпитопу.
Под терминами «антитело человека» или «полностью антитело человека», используемыми в данном документе, понимают антитела, имеющие вариабельные области, в которых как каркасные области, так и CDR-области происходят из иммуноглобулиновых последовательностей зародышевой линии человека. Более того, если антитело содержит константную область, то константная область также происходит из иммуноглобулиновых последовательностей зародышевой линии человека. Антитела человека по изобретению или их антигенсвязывающие участки могут включать аминокислотные остатки, не кодируемые иммуноглобулиновыми последовательностями зародышевой линии человека (например, мутации, введенные путем случайного или сайт-специфического мутагенеза in vitro или за счет соматической мутации in vivo). Тем не менее, термин «антитело человека», используемый в данном документе, не включает антител, в которых в каркасные последовательности человека трансплантированы последовательности CDR, происходящие из зародышевой линии другого вида млекопитающих, такого как мышь.
Под терминами «моноклональное антитело человека» или «полностью человеческое моноклональное антитело» понимают антитела, у которых наблюдается единственная специфичность связывания, имеющие вариабельные области, в которых как каркасные, так и CDR-области происходят из иммуноглобулиновых последовательностей зародышевой линии человека. В одном из вариантов осуществления моноклональные антитела человека продуцируются гибридомой, которая включает В-клетку, полученную от трансгенного животного, отличного от человека, например трансгенной мыши, имеющей геном, содержащий трансген тяжелой цепи и трансген легкой цепи человека, где В-клетку сливают с иммортализованной клеткой.
Под термином «рекомбинантное антитело человека», используемым в данном документе, понимают все антитела человека, которые получают, экспрессируют, создают или выделяют рекомбинантными способами, такие как (а) антитела, выделенные из животного (например, мыши), которое является трансгенным или трансхромосомным в отношении генов иммуноглобулинов человека, или из гибридомы, полученной на основе этого (дополнительно описано ниже), (b) антитела, выделенные из клетки-хозяина, трансформированной, чтобы экспрессировать антитело человека, например, из трансфектомы, (с) антитела, выделенные из рекомбинантной комбинаторной библиотеки антител человека, и (d) антитела, полученные, экспрессированные, созданные или выделенные любыми другими способами, которые включают сплайсинг генных последовательностей иммуноглобулинов человека с другими последовательностями ДНК. Такие рекомбинантные антитела человека имеют вариабельные области, в которых каркасные и CDR-области происходят из иммуноглобулиновых последовательностей зародышевой линии человека. В некоторых вариантах осуществления, тем не менее, такие рекомбинантные антитела человека могут быть подвергнуты in vitro мутагенезу (или при использовании животного, трансгенного по последовательностям Ig человека, in vivo соматическому мутагенезу) и, таким образом, аминокислотные последовательности VL и VH-областей рекомбинантных антител представляют собой последовательности, которых, несмотря на происхождение и связь с VL и VH-последовательностями зародышевой линии человека, может не существовать в природе в зародышевом наборе антител человека in vivo.
Под используемым в данном документе термином «изотип» или «класс» понимают класс антител (например, IgM или IgG), который кодируется генами константных областей тяжелых цепей. Константные домены антител не вовлекаются в связывание с антигеном, но у них наблюдается большое число эффекторных функций. В зависимости от аминокислотной последовательности константной области тяжелой цепи данное антитело человека или иммуноглобулин может быть отнесено к одному из пяти основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM. Структуры и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны. Известно, что из большого числа классов иммуноглобулинов человека лишь IgG1, IgG2, IgG3, IgG4 и IgM человека активируют комплемент. Известно, что у людей ADCC опосредуется IgG1 и IgG3 человека.
Под используемым в данном документе термином «подкласс» понимают дополнительную детализацию в пределах изотипа гена константной области тяжелой цепи, такую как, например, подклассы IgG1, IgG2, IgG3 или IgG4 в изотипе IgG.
К используемому в данном документе термину «соединение» или «фармацевтическое соединение» относятся антитела, их антигенсвязывающие участки, иммуноконъюгаты и биспецифические молекулы.
Фразы «антитело, распознающее антиген» и «антитело, специфичное для антигена» используются в данном документе взаимозаменяемо с термином «антитело, которое специфически связывается с антигеном».
Под термином «антителозависимая клеточная цитотоксичность» или «ADCC» понимают клеточно-опосредованную реакцию, в которой неспецифические цитотоксические клетки (например, NK-клетки, нейтрофилы, макрофаги и т.д.) распознают антитело, связанное с мишенной клеткой, и после этого вызывают лизис мишенной клетки. Такие цитотоксические клетки, которые опосредуют ADCC, как правило, экспрессируют Fc-рецепторы (FcR). Основные клетки, опосредующие ADCC, (NK-клетки) экспрессируют FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII, FcγRIII и/или FcγRIV. Экспрессия FcR на гематопоэтических клетках обобщается в статье Ravetch и Kinet, Annu. Rev. Immunol., 9:457-92 (1991). Для оценки ADCC-активности молекулы может быть осуществлен in vitro анализ ADCC, такой как анализ, описанный в патенте США 5500362 или 5821337. К используемым для таких анализов эффекторным клеткам относятся мононуклеарные клетки периферической крови (РВМС) и естественные киллеры (NK-клетки). Альтернативно или дополнительно, ADCC-активность интересующих молекул может быть оценена in vivo, например, на модели на животных, такой как модель, описанная в статье Clynes et al., Proc. Natl. Acad. Sci. (USA), 95:652-656 (1998).
Термины «Fc-рецептор» или «FcR» используются для описания рецептора, который связывается с Fc-областью антитела, где Fc-область содержит шарнирную область и домены СН2 и СН3 тяжелой цепи. Например, FcR может представлять собой FcR человека с нативной последовательностью. FcR может представлять собой рецептор, который связывается с IgG-антителом (гамма-рецептор), и к нему относятся рецепторы подклассов FcγRI, FcγRII, FcγRIII и FcγRIV, включая аллельные варианты и альтернативно сплайсированные формы этих рецепторов. К рецепторам FcγRII относятся FcγRIIA («активирующий рецептор») и FcγRIIB («ингибирующий рецептор»), которые имеют схожие аминокислотные последовательности, которые различаются в основном по их цитоплазматическим доменам. Активирующий рецептор FcγRIIA содержит в своем цитоплазматическом домене иммунорецепторный тирозиновый активационный мотив (ITAM). Ингибирующий рецептор FcγRIIB содержит в своем цитоплазматическом домене иммунорецепторный тирозиновый ингибиторный мотив (ITIM) (см. статью Daeron, Annu. Rev. Immunol., 15:203-234 (1997)). Рецепторы FcR рассматриваются в статьях Ravetch и Kinet, Annu. Rev. Immunol., 9:457-92 (1991); Capel et al., Immunomethods, 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med., 126:330-41 (1995). Термином «FcR» в данном документе охватываются и другие FcR, включая не идентифицированные в настоящее время рецепторы. К термину также относится неонатальный рецептор FcRn, который отвечает за перенос материнских IgG плоду (Guyer et al., Immunol., 117:587 (1976) и Kim et al., J. Immunol., 24:249 (1994)). Основной FcR-связывающий сайт на Fc-фрагментах иммуноглобулинов расположен в шарнирной области между доменами СН1 и СН2. Эта шарнирная область взаимодействует с FcR1-3 на различных лейкоцитах и запускает атаку этих клеток на мишень (Wines et al., J. Immunol., 164:5313-5318 (2000)). Шарнирная область охватывает, но ими не ограничиваясь, последовательности, описанные в патенте США 6165476.
Под термином «способность индуцировать антителозависимую клеточную цитотоксичность» понимают способность агента, такого как антитело, демонстрировать ADCC, измеренную посредством анализа(ов), известного(ых) специалистам в данной области. Такая активность обычно отличается связыванием Fc-области с большим числом рецепторов FcR. Не ограничиваясь каким-либо конкретным механизмом, специалист в данной области должен понимать, что способность антитела демонстрировать ADCC может происходить, например, в соответствии с его подклассом (таким как IgG1 или IgG3), за счет мутаций, введенных в Fc-область, или в соответствии с модификациями углеводных паттернов в Fc-области антитела. Такие модификации описываются, например, в патентной публикации США 2007-0092521.
Под термином «производные антитела человека» понимают любую модифицированную форму антитела человека, например, конъюгат антитела и другой агент или антитело.
Под термином «гуманизированное антитело» понимают антитела, в которых в каркасные последовательности человека трансплантированы CDR-последовательности, полученные из зародышевой линии другого вида млекопитающих, такого как мышь. В каркасных последовательностях человека могут быть осуществлены дополнительные модификации каркасной области.
Под термином «химерное антитело» понимают антитела, в которых последовательности вариабельных областей происходят из одного вида, а последовательности константных областей происходят из другого вида, такие как антитело, в котором последовательности вариабельных областей происходят из антитела мыши, а последовательности константных областей происходят из антитела человека.
Под фразой «специфически связывается», используемой в данном документе, понимают соединение, например, белок, нуклеиновую кислоту, антитело и тому подобное, которое распознает и связывается со специфической молекулой, но практически не распознает или не связывается с другими молекулами в образце. Например, антитело или пептидный ингибитор, который распознает и связывается с когнатным лигандом (например, антитело против α5β1, которое связывается со своим когнатным антигеном α5β1) в образце, но практически не распознает или не связывается с другими молекулами в образце. Таким образом, при заданных условиях проведения анализа указанный связывающий фрагмент (например, антитело или его антигенсвязывающий участок) связывается преимущественно с конкретной мишенной молекулой, например, α5β1, и не связывается в значительном количестве с другими компонентами, присутствующими в тестируемом образце. Для отбора антитела, которое специфически связывается с интересующей молекулой, может быть использовано большое число форматов анализов. Например, к числу многих исследований, которые могут быть использованы для идентификации антитела, которое специфически взаимодействует с α5β1, относятся твердофазный иммуноанализ ELISA, иммунопреципитация, BIAcore, FACS и анализ вестерн-блот. Как правило, специфическая или селективная реакция должна быть, по меньшей мере, в два раза выше фонового сигнала или шума и лучше более чем в 10 раз выше фона, даже точнее, говорят, что антитело «специфически связывается» с антигеном, если равновесная константа диссоциации (KD) ≤1 мкМ, например, ≤100 нМ, а также например, ≤10 нМ.
Под используемым в данном документе антителом, которое «специфически связывается с интегрином α5β1 человека» понимают антитело, которое связывается с интегрином α5β1 человека с KD 1×10-7 М или меньше, 5×10-8 М или меньше, 3×10-8 М или меньше, 1×10-8 М или меньше или 5×10-9 М или меньше.
Под термином «kon», используемым в данном документе, понимают скорость прямой реакции или скорость ассоциации конкретного взаимодействия антитело-антиген, тогда как под термином «koff», используемым в данном документе, понимают скорость обратной реакции или скорость диссоциации конкретного взаимодействия антитело-антиген. Под термином «KD», используемым в данном документе, понимают константу диссоциации, которую получают из соотношения koff к kon (т.е. koff/kon) и выражают в виде молярной концентрации (М). Величины KD для антител могут быть определены, используя способы, общепринятые в данной области. Один из способов определения KD антитела представляет собой применение поверхностного плазмонного резонанса, обычно используя биосенсорную систему, такую как система Biacore®.
Под используемым в данном документе термином «высокое сродство» в отношении IgG-антитела понимают антитело, имеющее KD 1×10-7 М или меньше, 5×10-8 М или меньше или 5×10-9 М или меньше для мишенного антигена. Тем не менее, связывание с «высоким сродством» может варьировать для других изотипов антител. Например, под связыванием с «высоким сродством» для изотипа IgM понимают антитело, имеющее KD 10-6 М или меньше, 10-7 М или меньше или 10-8 М или меньше.
Под термином «конкурировать», используемым в данном документе по отношению к антителу, понимают, что если первое антитело или его антигенсвязывающий участок конкурирует за связывание со вторым антителом или его антигенсвязывающим участком, то связывание первого антитела со своим когнатным эпитопом снижается детектируемым образом в присутствии второго антитела по сравнению со связыванием первого антитела в отсутствие второго антитела. Противоположное, связывание второго антитела со своим эпитопом также детектируемым образом снижается в присутствии первого антитела, может быть верно, но необязательно. То есть первое антитело может ингибировать связывание второго антитела со своим эпитопом, но второе антитело может не ингибировать связывание первого антитела со своим соответствующим эпитопом. Тем не менее, если каждое антитело детектируемым образом ингибирует связывание другого антитела со своим когнатным эпитопом или лигандом, либо до такой же, более высокой, либо меньшей степени, говорят, что антитела «перекрестно конкурируют» друг с другом за связывание соответствующего(их) им эпитопа(ов). Например, перекрестно конкурирующие антитела могут связываться с эпитопом или участком эпитопа, с которым связываются антитела, описанные в данном документе. Настоящее изобретение охватывает применение как конкурирующих, так и перекрестно конкурирующих антител. Независимо от механизма, которым проявляется такая конкуренция или перекрестная конкуренция (например, стерическое несоответствие, конформационное изменение или связывание с общим эпитопом или его частью и тому подобное), специалист в данной области должен понимать, основываясь на идеях, предлагаемых в данном документе, что такие конкурирующие и/или перекрестно конкурирующие антитела охватываются изобретением и могут быть использованы для способов, описанных в данном документе.
К термину «эпитоп» относится любая белковая детерминанта, способная специфически связываться с иммуноглобулином или Т-клеточным рецептором. Эпитопные детерминанты обычно состоят из химически активных поверхностных группировок молекул, таких как аминокислоты или боковые цепи сахаров, и обычно имеют характеристики специфической трехмерной структуры, а также специфические характеристики заряда. Конформационные и неконформационные эпитопы различаются тем, что связывание с первыми, но не с последними, снижается при наличии денатурирующих растворителей.
Под «гликоформой» понимают комплексную олигосахаридную структуру, содержащую сцепление большого числа углеводных единиц. Такие структуры описываются, например, в руководстве Essentials of Glycobiology Varki et al., eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1999), в котором также предлагается обзор стандартных гликобиологических обозначений. К таким гликоформам относятся, но ими не ограничиваясь, G2, G1, G0, G-1 и G-2 (см., например, международную патентную публикацию WO 99/22764).
«Паттерн гликозилирования» определяется как паттерн углеводных единиц, которые ковалентно соединены с белком (например, гликоформа), а также как сайт(ы), по которым гликоформа(ы) ковалентно присоединена(ы) к пептидному остову белка, конкретнее к белку иммуноглобулина.
По всей видимости, антитела, экспрессируемые различными клеточными линиями или в трансгенных животных, будут обладать разными гликоформами и/или паттернами гликозилирования по сравнению друг с другом. Тем не менее, все антитела, кодируемые молекулами нуклеиновых кислот по данному описанию или содержащие аминокислотные последовательности по данному описанию, являются частью настоящего описания вне зависимости от гликозилирования таких антител.
Под используемым в данном документе термином «индивид» понимают любого человека или животное. Под термином «животное» понимают всех позвоночных, например, млекопитающих и животных, отличных от млекопитающих, таких как приматы, овцы, собаки, кошки, лошади, коровы, курицы, земноводные, рептилии и т.д.
Под используемым в данном документе термином «лечить» понимают снижение частоты, с которой пациент испытывает симптомы заболевания (т.е. рост опухоли и/или метастазирование или другой эффект, опосредованный через количество и/или активность иммунных клеток, и тому подобное). К термину относится введение соединений или агентов по настоящему изобретению для предотвращения или задержки появления симптомов, осложнений или биохимических признаков заболевания, смягчения симптомов или купирования или ингибирования дальнейшего развития заболевания, состояния или расстройства. Лечение может быть профилактическим (для предотвращения или задержки возникновения заболевания или для предотвращения проявления его клинических или субклинических симптомов) или терапевтической супрессией или смягчением симптомов после выявления заболевания.
Более подробно большое число аспектов изобретения описывается в последующих подразделах.
Антитела против α5β1
Антитела по изобретению отличаются определенными функциональными особенностями или свойствами антител. Например, антитела специфически связываются с α5β1 человека. Предпочтительно, чтобы антитело по изобретению связывалось с α5β1 с высоким сродством, например, с KD 1×10-7 M или меньше.
Предпочтительно, чтобы антитело связывалось с α5β1 человека с KD 5×10-8 M или меньше, 2×10-8 M или меньше, 5×10-9 M или меньше, 4×10-9 M или меньше, 3×10-9 M или меньше или 2,7×10-9 M или меньше. В данной области известны анализы для оценки связывающей способности антител к α5β1, к ним относятся, например, ELISA, вестерн-блот, RIA и анализ проточной цитометрии. Подходящие анализы подробно описываются в примерах. Кинетический анализ связывания (например, сродство связывания) антител также может быть оценен с помощью анализов, известных в данной области, таких как анализ Biacore.
Антитела против α5β1 по настоящему изобретению также способны индуцировать антителозависимую клеточную цитотоксичность (ADCC). Такая функциональность может быть достигнута, например, путем применения специфических подклассов (например, IgG1, IgG2, IgG3), включающих мутации Fc-домена, которые могут дополнительно повысить уровень ADCC-активности антитела. Такие мутации и способы измерения ADCC дополнительно описываются в примерах.
Моноклональное антитело 22В5
Одно из наглядных антител по изобретению представляет собой моноклональное антитело человека 22В5, описанное в примерах 1 и 2. Аминокислотная последовательность VH антитела 22В5 представлена на фиг.1B и приводится в SEQ ID NO:7. Аминокислотная последовательность VL антитела 22В5 представлена на фиг.1D и приводится в SEQ ID NO:8. Как показано на фиг.1В и фиг.2, вариабельная область тяжелой цепи антитела 22В5 содержит две мутации по сравнению с генной последовательностью зародышевой линии человека. А именно, 22В5 содержит одну мутацию изолейцина в серин по аминокислотному остатку номер 30 (I30S) и одну мутацию аспарагина в серин по аминокислотному остатку номер 33 (N33S). Под используемым в данном документе обозначением «22В5» понимают антитело, в котором были сделаны указанные мутации (I30S и N33S) вариабельной области тяжелой цепи зародышевой линии.
Учитывая, что 22В5 может связываться с α5β1, последовательности VH и VL могут быть «смешаны и совмещены» с другими антителами против α5β1 для создания дополнительных связывающих молекул против α5β1 по изобретению. Связывание с α5β1 таких «смешанных и совмещенных» антител может быть протестировано, используя анализы связывания, описанные выше и в примерах (например, анализы ELISA). В одном из случаев при смешивании и совмещении VH и VL-цепей VH-последовательность определенной пары VH/VL замещается подобной в структурном отношении VH-последовательностью. Аналогично, в другом случае VL-последовательность определенной пары VH/VL замещается подобной в структурном отношении VL-последовательностью.
В другом аспекте изобретение относится к антителам, которые содержат области CDR1, CDR2 и CDR3 тяжелых и легких цепей антитела 22В5. Аминокислотная последовательность CDR1-области области VH антитела 22В5 представлена в SEQ ID NO:1. Аминокислотная последовательность CDR2-области области VH антитела 22В5 представлена в SEQ ID NO:2. Аминокислотная последовательность CDR3-области области VH антитела 22В5 представлена в SEQ ID NO:3. Аминокислотная последовательность CDR1-области VL-области антитела 22В5 представлена в SEQ ID NO:4. Аминокислотная последовательность CDR2-области VL-области антитела 22В5 представлена в SEQ ID NO:5. Аминокислотная последовательность CDR3-области VL-области антитела 22В5 представлена в SEQ ID NO:6. CDR-области определяют, используя систему Кабат (Kabat E.A. et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication № 91-3242).
Учитывая, что антитело 22В5 связывается с α5β1 и что антигенсвязывающая специфичность обеспечивается преимущественно областями CDR1, CDR2 и CDR3, последовательности CDR1, CDR2 и CDR3 VH-области и последовательности CDR1, CDR2 и CDR3 VL-области могут быть «смешаны и совмещены» (т.е. CDR-области различных антител против α5β1 могут быть смешаны и совмещены, хотя каждое антитело будет содержать, как и положено, CDR1, CDR2 и CDR3 области VH и CDR1, CDR2 и CDR3 области VL) для создания дополнительных связывающих молекул против α5β1 по изобретению. Связывание с α5β1 таких «смешанных и совмещенных» антител может быть протестировано, используя анализы связывания, описанные выше и в примерах (например, анализы ELISA, анализ Biacore). В одном из случаев при смешивании и совмещении CDR-последовательностей области VH последовательность CDR1, CDR2 и/или CDR3 из определенной последовательности VH замещается структурно подобной(ыми) последовательностью(ями) CDR. Аналогично, при смешивании и совмещении последовательностей CDR области VL последовательность CDR1, CDR2 и/или CDR3 из определенной последовательности VL, как правило, замещается структурно подобной(ыми) последовательностью(ями) CDR. Специалисту в данной области будет нетрудно понять, что новые последовательности VH и VL могут быть созданы путем замещения одной или нескольких последовательностей CDR-областей областей VH и/или VL структурно подобными последовательностями исходя из CDR-последовательностей, описанных в данном документе.
Соответственно, по другому аспекту изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему: (а) вариабельную область тяжелой цепи CDR1, содержащую аминокислотную последовательность SEQ ID NO:1; (b) вариабельную область тяжелой цепи CDR2, содержащую аминокислотную последовательность SEQ ID NO:2; (c) вариабельную область тяжелой цепи CDR3, содержащую аминокислотную последовательность SEQ ID NO:3; (d) вариабельную область легкой цепи CDR1, содержащую аминокислотную последовательность SEQ ID NO:4; (e) вариабельную область легкой цепи CDR2, содержащую аминокислотную последовательность SEQ ID NO:5; и/или (f) вариабельную область легкой цепи CDR3, содержащую аминокислотную последовательность SEQ ID NO:6; где антитело специфически связывается с α5β1, предпочтительно с α5β1 человека.
Моноклональные антитела 2D2, 24C7 и 1D9
Другое наглядное антитело по изобретению представляет собой моноклональное антитело человека 2D2, описанное в примерах 1 и 8. Аминокислотная последовательность области VH антитела 2D2 представлена в SEQ ID NO:39. Аминокислотная последовательность области VL антитела 2D2 представлена в SEQ ID NO:40.
Другое наглядное антитело по изобретению представляет собой моноклональное антитело человека 24С7, описанное в примерах 1 и 8. Аминокислотная последовательность области VH антитела 24С7 представлена в SEQ ID NO:19. Аминокислотная последовательность области VL антитела 24С7 представлена в SEQ ID NO:20.
Другое наглядное антитело по изобретению представляет собой моноклональное антитело человека 1D9, описанное в примерах 1 и 8. Аминокислотная последовательность области VH антитела 1D9 представлена в SEQ ID NO:29. Аминокислотная последовательность области VL антитела 1D9 представлена в SEQ ID NO:30.
Учитывая, что антитела 2D2, 24С7 и 1D9 могут связываться с α5β1, последовательности VH и VL этих антител могут быть «смешаны и совмещены» с другими антителами против α5β1 для создания дополнительных связывающих молекул против α5β1 по изобретению. Связывание с α5β1 таких «смешанных и совмещенных» антител может быть протестировано, используя анализы связывания, описанные выше и в примерах (например, анализы ELISA,). В одном из случаев при смешивании и совмещении цепей VH и VL последовательность VH определенной пары VH/VL замещается структурно подобной последовательностью VH. Аналогично, в другом случае последовательность VL определенной пары VH/VL замещается структурно подобной последовательностью VL.
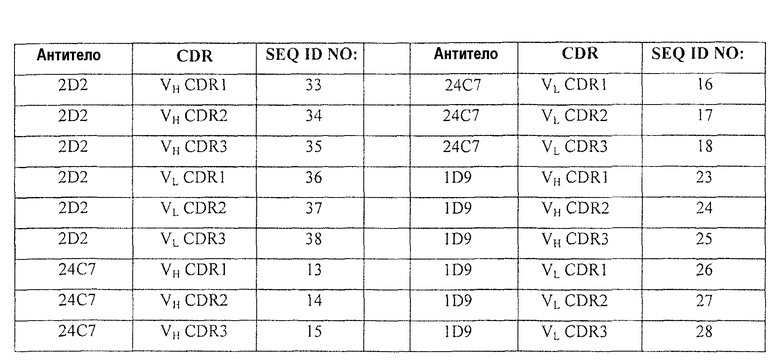
В другом аспекте изобретение относится к антителам, которые содержат области CDR1, CDR2 и CDR3 тяжелых и легких цепей антител 2D2, 24С7 и 1D9. Соответствующие аминокислотные последовательности этих CDR-областей указаны ниже.
CDR-области определяют, используя систему Кабат (Kabat E.A. et al. (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication № 91-3242).
Учитывая, что антитела 2D2, 24С7 и 1D9 связываются с α5β1 и что антигенсвязывающая специфичность обеспечивается преимущественно областями CDR1, CDR2 и CDR3, последовательности CDR1, CDR2 и CDR3 области VH и последовательности CDR1, CDR2 и CDR3 области VL могут быть «смешаны и совмещены» (т.е. CDR-области различных антител против α5β1 могут быть смешаны и совмещены, хотя каждое антитело будет содержать, как и положено, CDR1, CDR2 и CDR3-области области VH и CDR1, CDR2 и CDR3-области области VL) для создания дополнительных связывающих молекул против α5β1 по изобретению. Связывание с α5β1 таких «смешанных и совмещенных» антител может быть протестировано, используя исследования связывания, описанные выше и в примерах (например, анализы ELISA, анализ Biacore). В одном из случаев при смешивании и совмещении CDR-последовательностей области VH последовательность CDR1, CDR2 и/или CDR3 определенной последовательности VH замещается структурно подобной(ыми) последовательностью(ями) CDR. Аналогично, при смешивании и совмещении CDR-последовательностей области VL последовательность CDR1, CDR2 и/или CDR3 определенной последовательности VL, как правило, замещается структурно подобной(ыми) последовательностью(ями) CDR. Специалисту в данной области будет нетрудно понять, что новые последовательности VH и VL могут быть созданы замещением одной или нескольких последовательностей CDR- области VH и/или VL структурно подобными последовательностями из CDR-последовательностей, описанных в данном документе.
Соответственно, по другому аспекту изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему: (а) вариабельную область тяжелой цепи CDR1, содержащую аминокислотную последовательность SEQ ID NO:33; (b) вариабельную область тяжелой цепи CDR2, содержащую аминокислотную последовательность SEQ ID NO:34; (c) вариабельную область тяжелой цепи CDR3, содержащую аминокислотную последовательность SEQ ID NO:35; (d) вариабельную область легкой цепи CDR1, содержащую аминокислотную последовательность SEQ ID NO:36; (e) вариабельную область легкой цепи CDR2, содержащую аминокислотную последовательность SEQ ID NO:37; и/или (f) вариабельную область легкой цепи CDR3, содержащую аминокислотную последовательность SEQ ID NO:38; где антитело специфически связывается с α5β1, предпочтительно α5β1 человека.
В другом аспекте изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему: (а) вариабельную область тяжелой цепи CDR1, содержащую аминокислотную последовательность SEQ ID NO:13; (b) вариабельную область тяжелой цепи CDR2, содержащую аминокислотную последовательность SEQ ID NO:14; (c) вариабельную область тяжелой цепи CDR3, содержащую аминокислотную последовательность SEQ ID NO:15; (d) вариабельную область легкой цепи CDR1, содержащую аминокислотную последовательность SEQ ID NO:16; (e) вариабельную область легкой цепи CDR2, содержащую аминокислотную последовательность SEQ ID NO:17; и/или (f) вариабельную область легкой цепи CDR3, содержащую аминокислотную последовательность SEQ ID NO:18; где антитело специфически связывается с α5β1, предпочтительно α5β1 человека.
В другом аспекте изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему: (а) вариабельную область тяжелой цепи CDR1, содержащую аминокислотную последовательность SEQ ID NO:23; (b) вариабельную область тяжелой цепи CDR2, содержащую аминокислотную последовательность SEQ ID NO:24; (c) вариабельную область тяжелой цепи CDR3, содержащую аминокислотную последовательность SEQ ID NO:25; (d) вариабельную область легкой цепи CDR1, содержащую аминокислотную последовательность SEQ ID NO:26; (e) вариабельную область легкой цепи CDR2, содержащую аминокислотную последовательность SEQ ID NO:27; и/или (f) вариабельную область легкой цепи CDR3, содержащую аминокислотную последовательность SEQ ID NO:28; где антитело специфически связывается с α5β1, предпочтительно α5β1 человека.
Антитела, имеющие определенные последовательности зародышевой линии
В некоторых аспектах антитело по изобретению содержит вариабельную область тяжелой цепи из определенного гена тяжелой цепи иммуноглобулина зародышевой линии и/или вариабельную область легкой цепи из определенного гена легкой цепи иммуноглобулина зародышевой линии.
Например, в одном из аспектов изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, которая представляет собой продукт или производное гена человека VH 4-39, где антитело специфически связывается с α5β1. В другом аспекте изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, которая представляет собой продукт или производное гена человека VH 3-30.3, где антитело специфически связывается с α5β1. В еще одном аспекте изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область легкой цепи, которая представляет собой продукт или производное гена человека VK L6, где антитело специфически связывается с α5β1. В еще одном наглядном аспекте изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, где антитело:
(а) содержит вариабельную область тяжелой цепи, которая представляет собой продукт или производное гена человека VH 4-39 (этот ген кодирует аминокислотную последовательность, приведенную в SEQ ID NO:7) или гена человека 3-30.3 (этот ген кодирует аминокислотную последовательность, приведенную в SEQ ID NO:19, 29 или 39);
(b) содержит вариабельную область легкой цепи, которая представляет собой продукт или производное гена человека VK L6 (этот ген кодирует аминокислотную последовательность, приведенную в SEQ ID NO:8, 20, 30 или 40); и
(c) специфически связывается с α5β1, предпочтительно α5β1 человека.
Примером антитела, имеющего VH и VL-последовательности генов VH 4-39 и VK L6 соответственно, является 22В5. Примерами антител, имеющих VH и VL-последовательности генов VH 3-30.3 и VK L6 соответственно, являются 24С7, 2D2 и 1D9.
Используемое в данном документе антитело человека содержит вариабельные области тяжелой или легкой цепи, которые представляют собой «продукт» или «производное» определенной последовательности зародышевой линии, если вариабельные области антитела получают из системы, в которой используются иммуноглобулиновые гены зародышевой линии человека. К таким системам относятся иммунизация трансгенной мыши, несущей гены иммуноглобулинов человека, с помощью интересующего антигена или скрининг библиотеки иммуноглобулиновых генов человека в формате фагового дисплея с помощью интересующего антигена. Антитело человека, которое представляет собой «продукт» или «производное» иммуноглобулиновой последовательности зародышевой линии человека, может быть идентифицировано в силу этого путем сравнения аминокислотной последовательности антитела человека с аминокислотными последовательностями иммуноглобулинов зародышевой линии человека и отбора иммуноглобулиновой последовательности зародышевой линии человека, которая наиболее близка по последовательности (т.е. имеет самый высокий % идентичности) к последовательности антитела человека. Антитело человека, которое представляет собой «продукт» или «производное» определенной иммуноглобулиновой последовательности зародышевой линии человека, может содержать аминокислотные отличия по сравнению с последовательностью зародышевой линии, вследствие, например, природных соматических мутаций или преднамеренного введения сайт-специфической мутации. Тем не менее, выбранное антитело человека, как правило, по меньшей мере, на 90% идентично по аминокислотной последовательности с аминокислотной последовательностью, кодируемой иммуноглобулиновым геном зародышевой линии человека, и содержит аминокислотные остатки, которые идентифицируют антитело человека как человеческое при сравнении с аминокислотными последовательностями иммуноглобулинов зародышевой линии других видов (например, последовательностями зародышевой линии мыши). В некоторых случаях антитело человека может быть, по меньшей мере, на 95% или даже, по меньшей мере, на 96%, 97%, 98% или 99% идентично по аминокислотной последовательности с аминокислотной последовательностью, кодируемой иммуноглобулиновым геном зародышевой линии. В некоторых случаях антитело человека идентично по аминокислотной последовательности с аминокислотной последовательностью, кодируемой Ig геном зародышевой линии. Как правило, у антитела человека, происходящего из определенной последовательности зародышевой линии человека, будет наблюдаться не более 10 аминокислотных отличий от аминокислотной последовательности, кодируемой иммуноглобулиновым геном зародышевой линии человека. В некоторых случаях у антитела человека может наблюдаться не более 5 или даже не более 4, 3, 2 или 1 аминокислотного отличия от аминокислотной последовательности, кодируемой иммуноглобулиновым геном зародышевой линии.
Гомологичные антитела
В еще одном аспекте антитело по изобретению содержит вариабельные области тяжелой и легкой цепей, содержащие аминокислотные последовательности, которые гомологичны аминокислотным последовательностям наглядных антител, описанных в данном документе, и где антитела сохраняют желаемые функциональные свойства антител против α5β1 по изобретению.
Например, изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(а) вариабельная область тяжелой цепи содержит аминокислотную последовательность, которая, по меньшей мере, на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:7, 19, 29 и 39;
(b) вариабельная область легкой цепи содержит аминокислотную последовательность, которая, по меньшей мере, на 80% гомологична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:8, 20, 30 и 40; и у антитела наблюдаются одно или несколько из следующих свойств:
(i) антитело связывается с α5β1 человека с KD 1×10-7 M или меньше;
(ii) антитело способно индуцировать антителозависимую клеточную цитотоксичность.
В большом числе примеров антитело может представлять собой, например, антитело человека, гуманизированное антитело или химерное антитело.
В других примерах аминокислотные последовательности VH и/или VL могут быть на 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологичны последовательностям, приведенным выше. Антитело, имеющее области VH и VL, обладающие высокой (т.е. 80% или больше) гомологией с областями VH и VL последовательностей, приведенных выше, может быть получено мутагенезом (например, сайт-специфическим или ПЦР-опосредованным мутагенезом) молекул нуклеиновых кислот, кодирующих SEQ ID NO:7, 19, 29, 39, 8, 20, 30 и/или 40, с последующим тестированием кодируемого измененного антитела в отношении сохраненной функции (т.е. свойств, приведенных в (i) и/или (ii) выше), используя функциональные анализы, описанные в данном документе.
Используемый в данном документе процент гомологии между двумя аминокислотными последовательностями равен проценту идентичности между двумя последовательностями. Процент идентичности между двумя последовательностями представляет собой функцию от числа идентичных позиций, принадлежащих последовательностям (т.е. % гомологии = # идентичных позиций/общее # позиций × 100), принимая в расчет число разрывов и длину каждого разрыва, которые необходимо ввести для оптимального выравнивания двух последовательностей. Сравнение последовательностей и определение процента идентичности между двумя последовательностями может быть осуществлено, используя математический алгоритм, описанный в неограничивающих примерах ниже.
Процент идентичности между двумя аминокислотными последовательностями может быть определен, используя алгоритм E. Meyers и W. Miller (Comput. Appl. Biosci., 4:11-17 (1988)), который встроен в программу ALIGN (версия 2.0), используя таблицу остатков весовых коэффициентов РАМ120, штраф за продолжающийся гэп 12 и штраф за гэп 4. Кроме того, процент идентичности между двумя аминокислотными последовательностями может быть определен, используя алгоритм Needleman и Wunsch (J. Mol. Biol. 48:444-453 (1970)), который встроен в программу GAP в пакете программ GCG (доступен в ), используя либо матрицу Blossum 62, либо матрицу РАМ250, и штраф за открытие гэпа 16, 14, 12, 10, 8, 6 или 4 и за его продолжение 1, 2, 3, 4, 5 или 6.
Дополнительно или альтернативно, белковые последовательности по настоящему изобретению дополнительно могут использоваться в качестве «поисковой последовательности» для осуществления поиска в опубликованных базах данных, например, для идентификации родственных последовательностей. Такой поиск может быть осуществлен, используя программу XBLAST (версия 2.0) авторов Altschul et al. (1990) J. Mol. Biol. 215:403-10. Поиск белков посредством BLAST может быть осуществлен с помощью программы XBLAST, вес выравнивания = 50, длина слова = 3 для получения аминокислотных последовательностей, гомологичных молекулам антител по изобретению. Для получения выравниваний с разрывами с целями сравнения может быть использована программа Gapped BLAST, описанная в статье Altschul et al. (1997) Nucleic Acids Res. 25(17):3389-3402. При использовании программ BLAST и Gapped BLAST могут быть использованы параметры по умолчанию соответствующих программ (например, XBLAST и NBLAST). См. http://www.ncbi.nlm.nih.gov.
Антитела с консервативными модификациями
В некоторых случаях антитело по изобретению содержит вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, где одна или несколько из этих CDR-последовательностей содержат определенные аминокислотные последовательности на основе наглядных антител, описанных в данном документе (например, 22В5, 1D9, 24C7 и 2D2) или их консервативные модификации, и где антитела сохраняют одно или несколько желаемых функциональных свойств антител против α5β1 по изобретению. Соответственно, изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, где:
(а) последовательность вариабельной области тяжелой цепи CDR3 содержит аминокислотную последовательность, выбранную из группы, состоящую из SEQ ID NO:3, 15, 25 и 35 и их консервативных модификаций;
(b) последовательность вариабельной области легкой цепи CDR3 содержит аминокислотную последовательность, выбранную из группы, состоящую из SEQ ID NO:6, 18, 28 и 38 и их консервативных модификаций; и у антитела наблюдаются одно или несколько из следующих свойств:
(i) антитело связывается с α5β1 человека с KD 1×10-7 M или меньше;
(ii) антитело способно индуцировать антителозависимую клеточную цитотоксичность.
В других случаях последовательность вариабельной области тяжелой цепи CDR2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:2, 14, 24 и 34 и их консервативных модификаций; и последовательность вариабельной области легкой цепи CDR2 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:5, 17, 27 и 37 и их консервативных модификаций. В другом случае последовательность вариабельной области тяжелой цепи CDR1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 13, 23 и 33 и их консервативных модификаций; и последовательность вариабельной области легкой цепи CDR1 содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:4, 16, 26 и 36 и их консервативных модификаций.
Под используемым в данном документе термином «консервативная модификация» понимают аминокислотные модификации, которые существенно не влияют или не изменяют характеристики связывания антитела, содержащего аминокислотную последовательность. К таким консервативным модификациям относятся замены, добавки и делеции аминокислот. Модификации могут быть введены в антитело по изобретению стандартными техниками, известными в данной области, такими как сайт-специфический мутагенез и ПЦР-опосредованный мутагенез. Консервативная модификация данной последовательности может включать такие последовательности, которые, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 97%, по меньшей мере, на 98% или, по меньшей мере, на 99% идентичны этой последовательности. Консервативные замены аминокислот представляют собой замены, при которых аминокислотный остаток замещается аминокислотным остатком, имеющим аналогичную боковую цепь. В данной области определены семейства аминокислотных остатков, имеющие аналогичные боковые цепи. К этим семействам относятся аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или несколько аминокислотных остатков в CDR-областях антитела по изобретению могут быть замещены другими аминокислотными остатками из того же семейства боковых цепей и измененное антитело может быть протестировано в отношении сохраненной функции (т.е. свойств, приведенных в (i) и (ii) выше), используя функциональные анализы, описанные в данном документе. Другой тип аминокислотной модификации представляет собой устранение пар аспарагин-глицин, которые образуют сайты потенциального дезамидирования, путем изменения одного или обоих остатков. В другом примере расщепили С-концевой лизин тяжелой цепи антитела против α5β1 по изобретению и он, таким образом, не представлен. Расщепление С-концевого лизина может быть разработано заранее или может быть результатом условий, использованных для экспрессии и очистки антитела. В большом числе случаев тяжелые и легкие цепи антител против α5β1 могут необязательно включать сигнальную последовательность.
Антитела, которые связываются с тем же эпитопом, что и наглядные антитела против α5β1 по изобретению
В другом аспекте изобретение относится к антителам, которые связываются с тем же эпитопом на α5β1 человека, что и любое наглядное моноклональное антитело против α5β1 по изобретению (т.е. антителам, которые способны перекрестно конкурировать за связывание с α5β1 с любым моноклональным антителом по изобретению). Например, референсное антитело для исследований перекрестного конкурирования может представлять собой моноклональное антитело 22В5 (имеющее последовательности VH и VL, представленные в SEQ ID NO:7 и 8 соответственно) или моноклональное антитело 24С7 (имеющее последовательности VH и VL, представленные в SEQ ID NO:19 и 20 соответственно), или моноклональное антитело 1D9 (имеющее последовательности VH и VL, представленные в SEQ ID NO:29 и 30 соответственно), или моноклональное антитело 2D2 (имеющее последовательности VH и VL, представленные в SEQ ID NO:39 и 40 соответственно). Такие перекрестно конкурирующие антитела могут быть идентифицированы исходя из их способности перекрестно конкурировать с антителами 22В5, 24С7, 1D9 или 2D2 в стандартных анализах связывания с α5β1. Например, для демонстрации перекрестной конкуренции с наглядными антителами по настоящему изобретению могут быть использованы анализ BIAcore, анализы ELISA или проточная цитометрия. Способность тестируемого антитела ингибировать связывание, например, антител 22В5, 24С7, 1D9 или 2D2 с α5β1 человека указывает на то, что тестируемое антитело может конкурировать с антителами 22В5, 24С7, 1D9 или 2D2 за связывание с α5β1 человека и, таким образом, связывается с тем же эпитопом на α5β1 человека, что и антитела 22В5, 24С7, 1D9 или 2D2. В одном из случаев антитело, которое связывается с тем же эпитопом на α5β1 человека, что и антитела 22В5, 24С7, 1D9 или 2D2, представляет собой моноклональное антитело человека. Такие моноклональные антитела человека могут быть получены и выделены, как описано, например, в примерах.
Созданные и модифицированные антитела
Антитело или его антигенсвязывающий участок по изобретению дополнительно может быть получено, используя антитело, имеющее одну или несколько последовательностей VH и/или VL, описанных в данном документе, в качестве исходного материала для создания модифицированного антитела, причем модифицированное антитело может обладать измененными свойствами по отношению к исходному антителу. Антитело может быть создано модификацией одного или нескольких остатков в одном или обеих вариабельных областях (т.е. VH и/или VL), например, в одном или нескольких CDR-областях и/или в одном или нескольких каркасных областях. Дополнительно или альтернативно, антитело может быть создано модификацией остатков в константной(ых) области(ях), например, с целью изменения эффекторной(ых) функции(й) антитела.
Один из типов создания вариабельной области, которая может быть осуществлена, представляет собой пересадку CDR. Антитела взаимодействуют с мишенными антигенами преимущественно по аминокислотным остаткам, которые расположены в шести областях, определяющих комплементарность, тяжелой и легкой цепи (CDR-областях). Поэтому аминокислотные последовательности в CDR-областях больше различаются между отдельными антителами, чем последовательности вне CDR-областей. Поскольку CDR-последовательности отвечают за большинство взаимодействий антитело-антиген, возможно экспрессировать рекомбинантные антитела, которые имитируют свойства специфических природных антител, путем конструирования экспрессирующих векторов, которые включают CDR-последовательности из специфического природного антитела, пересаженные в каркасные последовательности отличного антитела с отличающимися свойствами (см., например, статьи Riechmann L. et al. (1998) Nature 332:323-327; Jones P. et al. (1986) Nature 321:522-525; Queen C. et al. (1989) Proc. Natl. Acad. Sci. U.S.A. 86:10029-10033; патент США 5225539 автора Winter и патенты США 5530101, 5585089, 5693762 и 6180370 авторов Queen et al.)
Соответственно, другой аспект изобретения относится к выделенному моноклональному антителу или его антигенсвязывающему участку, содержащему вариабельную область тяжелой цепи, содержащую последовательности CDR1, CDR2 и CDR3, содержащие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 13, 23 и 33, SEQ ID NO:2, 14, 24 и 34 и SEQ ID NO:3, 15, 25 и 35 соответственно, и вариабельную область легкой цепи, содержащую последовательности CDR1, CDR2 и CDR3, содержащие аминокислотную последовательность, выбранную из группы, состоящую из SEQ ID NO:4, 16, 26 и 36, SEQ ID NO:5, 17, 27 и 37, и SEQ ID NO:6, 18, 28 и 38, соответственно. Поэтому такие антитела содержат CDR-последовательности областей VH и VL моноклональных антител 22В5, 24С7, 1D9 или 2D2, но могут содержать каркасные последовательности, отличные от каркасных последовательностей этих антител.
Такие каркасные последовательности могут быть получены из опубликованных баз данных ДНК или опубликованных ссылок, которые включают генные последовательности антитела зародышевой линии. Например, последовательности ДНК зародышевой линии для генов вариабельных областей тяжелой и легкой цепей человека могут быть найдены в базе данных последовательностей зародышевой линии человека «VBase» (доступной через интернет ), а также в руководстве Kabat E.A. et al. (1991), Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication № 91-3242; статьях Tomlinson I.M. et al. (1992), J. Mol. Biol. 227:776-798; и Cox J.P.L. et al. (1994), Eur. J. Immunol. 24:827-836; содержание которых прямо приводится в данном документе в качестве ссылки). В качестве другого примера, ДНК-последовательности зародышевой линии генов вариабельных областей тяжелой и легкой цепей человека могут быть найдены в базе данных Genbank. Например, следующие последовательности тяжелой цепи зародышевой линии, обнаруженные у мыши HCo7 HuMAb, доступны в сопроводительных номерах доступа базы данных Genbank: 1-69 (NG_0010109, NT_024637 и BC070333), 3-33 (NG_0010109 и NT_024637) и 3-7 (NG_0010109 и NT_024637). В качестве другого примера, следующие последовательности тяжелой цепи зародышевой линии, обнаруженные у мыши HCo12 HuMAb, доступны в сопроводительных номерах доступа базы данных Genbank: 1-69 (NG_0010109, NT_024637 и BC070333), 5-51 (NG_0010109 и NT_024637) и 4-34 (NG_0010109 и NT_024637), 3-30.3 (Х92283) и 3-23 (AJ406678).
К каркасным последовательностям, используемым в антителах по изобретению, относятся, но ими не ограничиваясь, последовательности, которые в структурном отношении аналогичны каркасным последовательностям, используемым в выбранных антителах по изобретению, например, аналогичны каркасным последовательностям VH 4-39 и/или VH 3-30.3 и/или каркасным последовательностям VK L6, используемым в наглядных моноклональных антителах по изобретению. Например, последовательности CDR1, CDR2 и CDR3 области VH и последовательности CDR1, CDR2 и CDR3 области VL могут быть пересажены в каркасные области, которые имеют последовательность, идентичную последовательности, обнаруженной в иммуноглобулиновом гене зародышевой линии, из которого происходит каркасная последовательность, или CDR-последовательности могут быть пересажены в каркасные области, которые содержат одну или несколько мутаций по сравнению с последовательностями зародышевой линии. Например, было обнаружено, что в некоторых случаях полезно мутировать остатки в каркасных областях, чтобы сохранить или усилить антигенсвязывающую способность антитела (см., например, патенты США 5530101; 5585089; 5693762 и 6180370 авторов Queen et al.).
Другой тип модификации вариабельной области представляет собой мутирование аминокислотных остатков в областях CDR1, CDR2 и/или CDR3 областей VH и/или VL с целью улучшения одного или нескольких свойств связывания (например, сродства) интересующего антитела. Для введения мутации(й) может быть осуществлен сайт-специфический мутагенез или ПЦР-опосредованный мутагенез, и влияние на связывание антитела, или другое интересующее функциональное свойство может быть оценено в анализах in vitro или in vivo, описанных в данном документе и предложенных в примерах. Как правило, вводят консервативные модификации (описанные выше). Мутации могут представлять собой аминокислотные замены, добавки или делеции. Более того, как правило, изменяют не более одного, двух, трех, четырех или пяти остатков в CDR-области.
Соответственно, в другом варианте осуществления изобретение относится к выделенным моноклональным антителам против α5β1 или их антигенсвязывающим участкам, содержащим вариабельную область тяжелой цепи, содержащую: (а) CDR1-область области VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO:1, 13, 23 и 33, и аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавок по сравнению с последовательностями SEQ ID NO:1, 13, 23 или 33; (b) CDR2-область области VH, содержащую аминокислотную последовательность, выбранную из группы, состоящую из последовательностей SEQ ID NO:2, 14, 24 и 34, и аминокислотную последовательность, имеющую одну, две, три, четыре или пять амнокислотных замен, делеций или добавок по сравнению с последовательностями SEQ ID NO:2, 14, 24 или 34; (c) CDR3-область области VH, содержащую аминокислотную последовательность, выбранную из группы, состоящую из последовательностей SEQ ID NO:3, 15, 25 и 35, и аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавок по сравнению с последовательностями SEQ ID NO:3, 15, 25 или 35; (d) CDR1-область области VL, содержащую аминокислотную последовательность, выбранную из группы, состоящую из последовательностей SEQ ID NO:4, 16, 26 и 36, и аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавок по сравнению с последовательностями SEQ ID NO:4, 16, 26 или 36; (e) CDR2-область области VL, содержащую аминокислотную последовательность, выбранную из группы, состоящую из последовательностей SEQ ID NO:5, 17, 27 и 37, и аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавок по сравнению с последовательностями SEQ ID NO:5, 17, 27 или 37; и (f) CDR3-область области VL, содержащую аминокислотную последовательность, выбранную из группы, состоящую из последовательностей SEQ ID NO:6, 18, 28 и 38, и аминокислотную последовательность, имеющую одну, две, три, четыре или пять аминокислотных замен, делеций или добавок по сравнению с последовательностями SEQ ID NO:6, 18, 28 или 38.
К созданным антителам по изобретению относятся антитела, в которых модификации осуществлены по каркасным остаткам в области VH и/или VL, например, для улучшения свойств антитела. Обычно такие модификации каркасных областей осуществляют с целью снижения иммуногенности антитела. Например, один из способов представляет собой «возвратное мутирование» одного или нескольких каркасных остатков к соответствующей последовательности зародышевой линии. Конкретнее, антитело, которое подверглось соматической мутации, может содержать каркасные остатки, которые отличаются от последовательности зародышевой линии, из которой антитело происходит. Такие остатки могут быть идентифицированы сравнением каркасных последовательностей антитела с последовательностями зародышевой линии, из которых антитело происходит. Для возврата последовательностей каркасной области к их зародышевой конфигурации соматические мутации могут быть «возвратно мутированы» к последовательности зародышевой линии с помощью, например, сайт-специфического мутагенеза или ПЦР-опосредованного мутагенеза.
Другой тип модификации каркасной области включает мутирование одного или нескольких остатков в каркасной области или даже в одном или нескольких CDR-областях для удаления Т-клеточных эпитопов, чтобы таким образом снизить потенциальную иммуногенность антитела. Этот подход также обозначают как «деиммунизация», и он более подробно описывается в патентной публикации США 20030153043 авторами Carr et al.
Помимо или в противоположность к модификациям, созданным в каркасных или CDR-областях, антитела по изобретению могут быть созданы включением модификаций в Fc-область, как правило, с целью изменения одного или нескольких функциональных свойств антитела, таких как период полувыведения из сыворотки, фиксация комплемента, связывание с Fc-рецептором и/или антигензависимая клеточная цитотоксичность. Более того, антитело по изобретению может быть химически модифицировано (например, к антителу могут быть присоединены один или несколько фрагментов) или модифицировано с целью изменения его гликозилирования, опять же для изменения одного или нескольких функциональных свойств антитела. Каждый из этих аспектов более подробно описывается ниже. Нумерация остатков в Fc-области соответствует индексу EU по Kabat.
В одном из случаев шарнирную область домена СН1 модифицируют таким образом, что изменяют количество цистеиновых остатков в шарнирной области, например, увеличивают или уменьшают. Этот подход дополнительно описывается в патенте США 5677425 авторами Bodmer et al. Количество цистеиновых остатков в шарнирной области СН1 изменяют, например, для содействия сборке легкой и тяжелой цепей или для повышения или снижения стабильности антитела.
В другом случае подвергают мутированию шарнирную область Fc-фрагмента антитела для снижения биологического периода полувыведения антитела. Конкретнее, в пограничную область доменов СН2-СН3 Fc-шарнирного фрагмента вводят одну или несколько аминокислотных мутаций, так что связывание антитела со стафилококковым белком А (SpA) ослабевает по сравнению со связыванием SpA нативным Fc-шарнирным доменом. Этот подход более подробно описывается в патенте США 6165745 авторами Ward et al.
В другом случае антитело модифицируют с целью повысить его биологический период полувыведения. Возможно большое число подходов. Например, могут быть встроены одна или несколько из следующих мутаций: T252L, T254S, T256F, как описывается в патенте США 6277375 автором Ward. Альтернативно, для повышения биологического периода полувыведения антитело может быть изменено в СН1-домене или CL-области таким образом, чтобы содержать эпитоп, связывающий рецептор спасения, взятый из двух петель СН2-домена Fc-области иммуноглобулина IgG, как описано в патентах США 5869046 и 6121022 авторами Presta et al.
В других случаях для изменения эффекторной(ых) функции(й) антитела изменяют Fc-область путем замещения, по меньшей мере, одного аминокислотного остатка отличным аминокислотным остатком. Например, одна или несколько аминокислот, выбранных из аминокислотных остатков 234, 235, 236, 237, 297, 318, 320 и 322, могут быть заменены отличным аминокислотным остатком, так что антитело будет обладать измененным сродством в отношении эффекторного лиганда, но сохранит антигенсвязывающую способность родительского антитела. Эффекторный лиганд, сродство к которому изменяют, может представлять собой Fc-рецептор или С1-компонент комплемента. Этот подход более подробно описывается в патентах США 5624821 и 5648260 авторами Winter et al.
В другом случае одна или несколько аминокислот, выбранных из аминокислотных остатков 329, 331 и 322, могут быть заменены отличным аминокислотным остатком, так что антитело будет обладать измененным связыванием C1q и/или сниженной или отсутствующей комплементзависимой цитотоксичностью (CDC). Этот подход более подробно описывается в патенте США 6194551 авторами Idusogie et al.
В другом примере изменяют один или несколько аминокислотных остатков в позициях аминокислот 231 и 239, таким образом, изменяя способность антитела фиксировать комплемент. Этот подход дополнительно описывается в РСТ публикации WO 94/29351 авторами Bodmer et al.
В еще одном примере Fc-область модифицируют, чтобы повысить способность антитела опосредовать антителозависимую клеточную цитотоксичность (ADCC) и/или повысить сродство антитела к Fcγ-рецептору, посредством модификации одной или нескольких аминокислот в следующих позициях: 238, 239, 248, 249, 252, 254, 255, 256, 258, 265, 267, 268, 269, 270, 272, 276, 278, 280, 283, 285, 286, 289, 290, 292, 293, 294, 295, 296, 298, 301, 303, 305, 307, 309, 312, 315, 320, 322, 324, 326, 327, 329, 330, 331, 333, 334, 335, 337, 338, 340, 360, 373, 376, 378, 382, 388, 389, 398. 414, 416, 419, 430, 434, 435, 437, 438 или 439.
Этот подход дополнительно описывается в РСТ публикации WO 00/42072 автором Presta. Более того, сайты связывания на IgG1 человека для FcγRI, FcγRII, FcγRIII и FcRn были картированы, и были описаны варианты с улучшенным связыванием (см. Shields R.L. et al. (2001) J. Biol. Chem. 276:6591-6604). Показано, что специфические мутации в позициях 256, 290, 298, 333, 334 и 339 улучшают связывание с FcγRIII. Кроме того, показано, что следующие сочетанные мутации улучшают связывание с FcγRIII: Т256А/S298A, S298A/E333A, S298A/K224A и S298A/E333A/K334A. Например, антитело 22В5/DLE, описанное в данном документе, имеет подкласс IgG1 изотипа IgG, но с введенными путем сайт-специфического мутагенеза следующими мутациями (по сравнению с подклассом IgG1 дикого типа): S247D; A338L и I340E. Аналогично, как более подробно описано ниже, такие мутации S247D, A338L и I340E были введены в моноклональные антитела 24С7, 1D9 и 2D2. Как дополнительно описывается в примерах, такие мутации могут повысить сродство антитела к Fcγ-рецепторам и, таким образом, повысить его эффекторную функцию. Поэтому изобретение относится к антителу, которое содержит, по меньшей мере, одну мутацию в Fc-области и обладает детектируемо большим ADCC-ответом, чем в других отношениях идентичное антитело, не содержащее, по меньшей мере, одной мутации.
В еще одном примере модифицируют гликозилирование антитела. Например, может быть создано агликозилированное антитело (т.е. антитело с недостатком гликозилирования). Гликозилирование может быть изменено с целью, например, повышения сродства антитела к антигену. Такие углеводные модификации могут быть осуществлены, например, путем изменения одного или нескольких сайтов гликозилирования в последовательности антитела. Например, могут быть осуществлены одна или несколько аминокислотных замен, которые приводят к устранению одного или нескольких сайтов гликозилирования каркасной области вариабельной области, благодаря чему устраняется гликозилирование в этом сайте. Такое агликозилирование может повышать сродство антитела к антигену. Такой подход более подробно описан в патентах США 5714350 и 6350861 авторами Co et al.
Дополнительно или альтернативно, может быть создано антитело, которое обладает измененным типом гликозилирования, такое как гипофукозилированное антитело, имеющее уменьшенные количества фукозильных остатков, или антитело, имеющее увеличенное количество бисекторных структур GlcNac. Было показано, что такие измененные паттерны гликозилирования повышают ADCC-способность антител. Такие углеводные модификации могут быть осуществлены, например, путем экспрессии антитела в клетке-хозяине с измененным механизмом гликозилирования. В данной области были описаны клетки с измененным механизмом гликозилирования, и они могут быть использованы в качестве клеток-хозяев, в которых экспрессируют рекомбинантные антитела по изобретению, таким образом продуцируя антитело с измененным гликозилированием. Например, в клеточных линиях Ms704, Ms705 и Ms709 отсутствует ген фукозилтрансферазы FUT8 (альфа (1,6) фукозилтрансфераза), поэтому у антител, экспрессируемых на клеточных линиях Ms704, Ms705 и Ms709, отсутствует фукоза у их углеводов. Клеточные линии Ms704, Ms705 и Ms709 FUT8-/- создавали путем прицельного разрушения гена FUT8 в клетках СНО/DG44, используя два замещающих вектора (см. патентную публикацию США 20040110704 авторов Yamane et al. и статью Yamane-Ohnuki et al. (2004) Biotechnol. Bioeng 87:614-22). В качестве другого примера, в ЕР 1176195 авторами Hanai et al. описывается клеточная линия с функционально разрушенным геном FUT8, который кодирует фукозилтрансферразу, так что у антител, экспрессируемых в такой клеточной линии, наблюдается гипофукозилирование за счет снижения или устранения фермента, соответствующего альфа 1,6-связи. Авторами Hanai et al. также описываются клеточные линии, которые обладают низкой ферментативной активностью в отношении добавления фукозы к N-ацетилглюкозамину, который связывается с Fc-областью антитела, или не обладают данной ферментативной активностью, например, миеломная клеточная линия крысы YB2/0 (ATCC CRL 1662). В РСТ публикации WO 03/035835 автором Presta описывается вариант клеточной линии СНО, клетки Lec13, со сниженной способностью присоединять фукозу к Asn(297)-связанным углеводам, что также приводит к гипофукозилированию антител, экспрессируемых в такой клетке-хозяине (см. также статью Shields R.L. et al. (2002) J. Biol. Chem. 277:26733-26740). В РСТ публикации WO 99/54342 авторами Umana et al. описываются клеточные линии, созданные для экспрессии гликопротеин-модифицированных гликозилтрансфераз (например, бета(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII), так что у антител, экспрессируемых в созданных клеточных линиях, наблюдаются увеличенные бисекторные структуры GlcNac, что приводит к повышенной ADCC-активности антител (см. также Umana et al. (1999) Nat. Biotech. 17:176-180). Альтернативно, фукозные остатки антитела могут быть отщеплены, используя фермент фукозидазу. Например, фукозидаза альфа-L-фукозидаза удаляет с антител фукозильные остатки (Tarentino A.L. et al. (1975) Biochem. 14:5516-23).
Другая модификация антител по изобретению представляет собой пегилирование. Антитело может быть пегилировано с целью, например, повысить биологический (например, сывороточный) период полувыведения антитела. Для пегилирования антитела антитело или его фрагмент, как правило, приводят в контакт с полиэтиленгликолем (PEG), таким как реакционноспособный сложный эфир или альдегидное производное PEG, в условиях, в которых одна или несколько групп PEG присоединяются к антителу или фрагменту антитела. Как правило, пегилирование осуществляют через реакцию ацилирования или реакцию алкилирования с реакционноспособной молекулой PEG (или аналогичным реакционноспособным водорастворимым полимером). Под используемым в данном документе термином «полиэтиленгликоль» понимают любую форму PEG, которая использовалась для получения производных других белков, такую как моно (С1-С10) алкокси- или арилоксиполиэтиленгликоль или полиэтиленгликольмалеимид. В некоторых случаях пегилируемое антитело представляет собой агликозилированное антитело. В данной области известны способы пегилирования белков, и они могут быть применены к антителам по изобретению. См., например, ЕР 0154316 авторов Nishimura et al. и ЕР 0401384 авторов Ishikawa et al.
Способы создания антител
Как описано выше, антитела против α5β1, имеющие последовательности VH и VL, описанные в данном документе, могут быть использованы для создания новых антител против α5β1 путем модификации последовательностей VH и/или VL, или константной(ых) области(ей), соединенной(ых) с ними. Таким образом, в другом аспекте изобретения структурные особенности антитела против α5β1 по изобретению, например антитела 22В5, 24С7, 1D9 или 2D2, используют для создания родственных в структурном отношении антител против α5β1, у которых сохраняется, по меньшей мере, одно функциональное свойство антитела по изобретению, такое как связывание с α5β1 человека. Например, одна или несколько CDR-областей антител 22В5, 24С7, 1D9 или 2D2 или их мутаций могут быть объединены с помощью рекомбинантных техник с известными каркасными областями и/или другими CDR-областями для создания дополнительных, созданных с помощью рекомбинантных техник антител против α5β1 по изобретению, описанных выше. К другим типам модификаций относятся модификации, описанные в предыдущем разделе. Исходный материал для способа создания представляет собой одну или несколько последовательностей VH и/или VL по данному изобретению или один или несколько их CDR-областей. Для создания нового антитела необязательно на самом деле получать (т.е. экспрессировать в виде белка) антитело, имеющее одну или несколько последовательностей VH и/или VL по данному изобретению или один или несколько их CDR-областей. Достаточно, чтобы информацию, содержащуюся в последовательности(ях), можно было использовать в качестве исходного материала для создания последовательности(ей) «второго поколения», полученной(ых) из исходной(ых) последовательности(ей), и затем последовательность(и) «второго поколения» получают и экспрессируют в виде белка.
Соответственно, в другом аспекте изобретение относится к способу получения антитела против α5β1, содержащему:
(а) получение: (i) последовательности вариабельной области тяжелой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящую из SEQ ID NO:1, 13, 23 и 33, последовательность CDR2, выбранную из группы, состоящую из SEQ ID NO:2, 14, 24 и 34, и/или последовательность CDR3, выбранную из группы, состоящую из SEQ ID NO:3, 15, 25 и 35; и/или (ii) последовательности вариабельной области легкой цепи антитела, содержащей последовательность CDR1, выбранную из группы, состоящей из SEQ ID NO:4, 16, 26 и 36, последовательность CDR2, выбранную из группы, состоящую из SEQ ID NO:5, 17, 27 и 37, и/или последовательность CDR3, выбранную из группы, состоящей из SEQ ID NO:6, 18, 28 и 38;
(b) изменение, по меньшей мере, одного аминокислотного остатка в последовательности вариабельной области тяжелой цепи антитела и/или последовательности вариабельной области легкой цепи антитела с целью создания, по меньшей мере, одной измененной последовательности антитела; и
(c) экспрессию измененной последовательности антитела в виде белка.
Для получения и экспрессии измененной последовательности антитела могут быть использованы стандартные молекулярно-биологические техники.
Предпочтительно, чтобы антитело, кодируемое измененной(ыми) последовательностью(ями) антитела, представляло собой антитело, которое сохранило бы одно, несколько или все функциональные свойства антител против α5β1, описанных в данном документе, к этим функциональным свойствам относятся, но ими не ограничиваясь:
(i) связывание с α5β1 человека с KD 1×10-7 M или меньше;
(ii) способность индуцировать ADCC.
Функциональные свойства измененных антител могут быть оценены, используя стандартные анализы, доступные в данной области и/или описанные в данном документе, такие как анализы, приведенные в примерах (например, проточную цитометрию, анализы связывания).
В некоторых аспектах способов создания антител по изобретению мутации могут быть введены случайным образом или избирательно вдоль всей или части последовательности, кодирующей антитело против α5β1, и полученные модифицированные антитела против α5β1 могут быть скринированы в отношении связывающей активности и/или других функциональных свойств, описанных в данном документе. Способы мутирования описаны в данной области. Например, в РСТ публикации WO 02/092780 автором Short описаны способы создания и скринирования мутаций антител, используя насыщающий мутагенез, сборку искусственным лигированием или их сочетание. Альтернативно, в РСТ публикации WO 03/074679 авторами Lazar et al.описываются способы применения компьютерных методов скрининга с целью оптимизации физико-химических свойств антител.
Молекулы нуклеиновых кислот, кодирующие антитела по изобретению
Другой аспект изобретения относится к молекулам нуклеиновых кислот, которые кодируют антитела по изобретению. Нуклеиновые кислоты могут присутствовать в целых клетках, в клеточном лизате или в частично очищенной,или практически чистой форме. Нуклеиновую кислоту «выделяют» или «делают практически чистой», очищая от других клеточных компонентов или других контаминантов, например, других нуклеиновых кислот или белков клетки, с помощью любых подходящих техник, включая обработку щелочью/SDS, CsCl-бэндинг, колоночную хроматографию, электрофорез в агарозном геле и другие. См. руководство F. Ausubel et al., ed. (1987) Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York. Нуклеиновая кислота по изобретению может представлять собой, например, ДНК или РНК и может содержать или не содержит интронные последовательности. Как правило, нуклеиновая кислота представляет собой молекулу кДНК.
Нуклеиновые кислоты по изобретению могут быть получены, используя любые подходящие техники молекулярной биологии. В отношении антител, экспрессируемых гибридомами (например, гибридомами, полученными от трансгенных мышей, несущих гены иммуноглобулина человека, дополнительно описанными ниже), молекулы кДНК, кодирующие легкую и тяжелую цепи антитела, созданного гибридомой, могут быть получены с помощью ПЦР-амплификации или техник клонирования кДНК. В отношении антител, полученных из библиотеки генов иммуноглобулинов (например, используя техники фаговых дисплеев), нуклеиновая кислота, кодирующая антитело, может быть получена из библиотеки.
К молекулам нуклеиновых кислот по изобретению относятся, например, молекулы, кодирующие последовательности VH и VL моноклонального антитела 22В5. Последовательность ДНК, кодирующая последовательность VH антитела 22В5, представлена в SEQ ID NO:11. Последовательность ДНК, кодирующая последовательность VL антитела 22В5, представлена в SEQ ID NO:12. Другие наглядные молекулы нуклеиновых кислот, описанные в данном документе, представляют собой молекулы, кодирующие последовательности VH и VL моноклональных антител 24С7, 1D9 или 2D2. Последовательность ДНК, кодирующая последовательность VH антитела 24С7, представлена в SEQ ID NO:21. Последовательность ДНК, кодирующая последовательность VL антитела 24С7, представлена в SEQ ID NO:22. Последовательность ДНК, кодирующая последовательность VH антитела 1D9, представлена в SEQ ID NO:31. Последовательность ДНК, кодирующая последовательность VL антитела 1D9, представлена в SEQ ID NO:32. Последовательность ДНК, кодирующая последовательность VH антитела 2D2, представлена в SEQ ID NO:41. Последовательность ДНК, кодирующая последовательность VL антитела 2D2, представлена в SEQ ID NO:42.
После получения ДНК-фрагментов, кодирующих сегменты VH и VL, эти ДНК-фрагменты могут быть дополнительно обработаны с помощью любых подходящих техник рекомбинантных ДНК, например, для превращения генов вариабельных областей в гены цепей полноразмерного антитела, в гены Fab-фрагментов или в ген scFv. В этих манипуляциях ДНК-фрагмент, кодирующий VL или VH, функционально связывают с другим ДНК-фрагментом, кодирующим другой белок, таким как константная область антитела или гибкий линкер. Под термином «функционально связывают», используемым в данном контексте, понимают, что два ДНК-фрагмента соединяют таким образом, что аминокислотные последовательности, кодируемые двумя ДНК-фрагментами, остаются в рамке считывания.
Выделенная ДНК, кодирующая VH-область, может быть превращена в ген полноразмерной тяжелой цепи путем функционального связывания VH-кодирующей ДНК с другой молекулой ДНК, кодирующей константные области тяжелой цепи (СН1, СН2 и СН3). Последовательности генов константных областей тяжелой цепи человека известны в данной области (см., например, руководство Kabat, E.A. et al. (1991) Sequences of Proteins of Immunological Interest, Fifth edition, U.S. Department of Health and Human Services, NIH Publication № 91-3242), и ДНК-фрагменты, охватывающие эти области, могут быть получены стандартной ПЦР-амплификацией. Константная область тяжелой цепи может представлять собой константную область иммуноглобулинов IgG1, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, но наиболее предпочтительно, если представляет собой константную область IgG1 или IgG4. Последовательность константной области IgG1 может представлять собой любой из большого числа аллелей или аллотипов, как известно, обнаруживаемых среди различных индивидуумов, таких как Gm(1), Gm(2), Gm(3) и Gm(17). Эти аллотипы представляют природную аминокислотную замену в константных областях IgG1. В отношении гена тяжелой цепи Fab-фрагмента, ДНК, кодирующая VH, может быть функционально связана с другой молекулой ДНК, кодирующей лишь константную область тяжелой цепи СН1.
Выделенная ДНК, кодирующая VL-область, может быть превращена в ген полноразмерной легкой цепи (а также ген легкой цепи Fab-фрагмента) путем функционального связывания VL-кодирующей ДНК с другой молекулой ДНК, кодирующей константную область легкой цепи CL. Последовательности генов константных областей легкой цепи человека известны в данной области (см., например, руководство Kabat E.A. et al. (1991) Sequences of Proteins of Immunological Interest, Fifth edition, U.S. Department of Health and Human Services, NIH Publication № 91-3242), и ДНК-фрагменты, охватывающие эти области, могут быть получены стандартной ПЦР-амплификацией. Константная область легкой цепи может представлять собой константную область легкой цепи каппа или лямбда.
Для создания гена scFv VH- и VL-кодирующие ДНК-фрагменты функционально связывают с другим фрагментом, кодирующим гибкий линкер, например, кодирующим аминокислотную последовательность (Gly4-Ser)3, так что последовательности VH и VL могут быть экспрессированы в виде непрерывного одноцепочечного белка, причем VL и VH-области соединены гибким линкером (см., например, статьи Bird et al. (1988) Science 242:423-426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883; McCafferty et al., (1990) Nature 348:552-554).
Продукция моноклональных антител по изобретению
Моноклональные антитела (mAb) по настоящему изобретению могут быть получены большим числом техник, включающих методологию, принятую для моноклональных антител, например, стандартную технику гибридизации соматических клеток авторов Kohler и Milstein (1975) Nature 256:495. Для продукции моноклонального антитела также могут быть использованы и другие техники, например, вирусная или онкогенная трансформация В-лимфоцитов.
Предпочтительной системой на животных для получения гибридом является система на мышах. Продукция гибридомы у мыши является общепринятой процедурой. Протоколы иммунизации и техники выделения иммунизированных спленоцитов для слияния известны в данной области. Партнеры по слиянию (например, миеломные клетки мыши) и процедуры слияния также известны.
Химерные или гуманизированные антитела по настоящему изобретению могут быть получены, основываясь на последовательности моноклонального антитела мыши, полученного, как описано выше. ДНК, кодирующая иммуноглобулины, содержащие тяжелую и легкую цепи, может быть получена из интересующей гибридомы мыши и создана таким образом, чтобы содержать иммуноглобулиновые последовательности, отличные от мышиных (например, человека), используя подходящие молекулярно-биологические техники. Например, для создания химерного антитела вариабельные области мыши могут быть связаны с константными областями человека, используя способы, известные в данной области (см., например, патент США 4816567 авторов Cabilly et al.). Для создания гуманизированного антитела CDR-области мыши могут быть встроены в каркасные области человека, используя способы, известные в данной области (см., например, патент США 5225539 автора Winter и патенты США 5530101; 5585089; 5693762 и 6180370 авторов Queen et al.).
В некоторых случаях антитела по изобретению представляют собой моноклональные антитела человека. Такие моноклональные антитела человека, направленные против α5β1, могут быть получены, используя трансгенных или трансхромосомных мышей, несущих участки иммунной системы человека вместо иммунной системы мыши. К этим трансгенным или трансхромосомным мышам относятся мыши, обозначаемые в данном документе как HuMAb Mouse® и KM Mouse®, соответственно, и совместно обозначаются в данном документе как «мыши с Ig человека».
Мышь HuMAb® (продукция Medarex®, Inc.) содержит минилокусы иммуноглобулиновых генов человека, которые кодируют неперестроенные последовательности тяжелой (µ и γ) и легкой κ цепей иммуноглобулина человека наряду с направленными мутациями, которые инактивируют эндогенные локусы µ и κ-цепей (см., например, статью Lonberg et al. (1994) Nature 368(6474):856-859). Соответственно, у мышей наблюдается сниженная экспрессия IgM или κ-цепи мыши, и в ответ на иммунизацию встроенные трансгены тяжелой и легкой цепей человека испытывают переключение класса и соматическую мутацию, что приводит к получению высокоаффинных моноклональных IgGk человека (Lonberg N. et al. (1994), выше; рассматривается в обзоре Lonberg, N. (1994) Handbook of Experimental Pharmacology 113:49-101; Lonberg N. и Huszar D. (1995) Intern. Rev. Immunol. 13:65-93, и Harding F. и Lonberg N. (1995) Ann. N.Y. Acad. Sci. 764:536-546). Получение и применение мыши HuMAb® и геномные модификации, которые несут такие мыши, дополнительно описаны в статьях Taylor L. et al. (1992) Nucleic Acids Research 20:6287-6295; Chen J. et al. (1993) International Immunology 5:647-656; Tuaillon et al. (1993) Proc. Natl. Acad. Sci. USA 90:3720-3724; Choi et al. (1993) Nature Genetics 4:117-123; Chen, J. et al. (1993) EMBO J. 12:821-830; Tuaillon et al. (1994) J. Immunol. 152:2912-2920; Taylor L. et al. (1994) International Immunology 6:579-591; and Fishwild D. et al. (1996) Nature Biotechnology 14; 845-851. См. дополнительно патенты США 5545806; 5569825; 5625126; 5633425; 5789650; 5877397; 5661016; 5814318; 5874299 и 5770429 авторов Lonberg и Kay; патент США 5545807 авторов Surani et al.; РСТ публикации WO 92/03918, WO 93/12227, WO 94/25585, WO 97/13852, WO 98/24884 и WO 99/45962 авторов Lonberg и Kay; и РСТ публикацию WO 01/14424 авторов Korman et al.
В другом случае антитела человека по изобретению могут быть получены, используя мышь, которая несет иммуноглобулиновые последовательности человека на трансгенах и трансхромосомах, такую как мышь, которая несет трансген тяжелой цепи человека и трансхромосому легкой цепи человека. Такие мыши, обозначаемые в данном документе как «мыши КМТМ», подробно описаны в РСТ публикации WO 02/43478 авторов Ishida et al.
Кроме того, в данной области доступны альтернативные системы трансгенных животных, экспрессирующие гены иммуноглобулинов человека, и они могут быть использованы для получения антител против α5β1 по изобретению. Например, может быть использована альтернативная трансгенная система, обозначаемая как ксеномышь (Abgenix, Inc.); такие мыши описаны, например, в патентах США 5939598; 6075181; 6114598; 6150584 и 6162963 авторов Kucherlapati et al.
Более того, в данной области доступны альтернативные системы трансхромосомных животных, экспрессирующие гены иммуноглобулинов человека, и они могут быть использованы для получения антител против α5β1 по изобретению. Например, могут быть использованы мыши, несущие как трансхромосому тяжелой цепи человека, так и трансхромосому легкой цепи человека, обозначаемые как «мыши ТС»; такие мыши описаны в статье Tomizuka et al. (2000) Proc. Natl. Acad. Sci. USA 97:722-727. Более того, в данной области описаны коровы, несущие трансхромосомы тяжелой и легкой цепей человека (Kuroiwa et al. (2002) Nature Biotechnology 20:889-894), и они могут быть использованы для получения антител против α5β1 по изобретению.
Моноклональные антитела человека по изобретению также могут быть получены, используя способы фагового дисплея для скрининга библиотек иммуноглобулиновых генов человека. Такие способы фагового дисплея для выделения антител человека хорошо известны в данной области. См., например, патенты США 5223409; 5403484 и 5571698 авторов Ladner et al.; патенты США 5427908 и 5580717 авторов Dower et al.; патенты США 5969108 и 6172197 авторов McCafferty et al.; и патенты США 5885793; 6521404; 6544731; 6555313; 6582915 и 6593081 авторов Griffiths et al.
Моноклональные антитела человека по изобретению также могут быть получены, используя мышей SCID, в которых были восстановлены иммунные клетки человека, так что при иммунизации может быть получен антительный ответ человека. Такие мыши описаны, например, в патентах США 5476996 и 5698767 авторов Wilson et al.
Иммунизация мышей с Ig человека
При использовании мышей с Ig человека для получения антител человека по изобретению такие мыши могут быть иммунизованы с помощью очищенного или обогащенного препарата антигена α5β1 и/или рекомбинантного α5β1 или слитого белка α5β1, как описано в статьях Lonberg N. et al. (1994) Nature 368(6474):856-859; Fishwild D. et al. (1996) Nature Biotechnology 14:845-851; и РСТ публикациях WO 98/24884 и WO 01/14424. Предпочтительно, чтобы при первой инфузии брались мыши в возрасте 6-16 недель. Например, очищенный или рекомбинантный препарат (5-50 мкг) антигена α5β1 может быть использован для внутрибрюшинной иммунизации мышей с Ig человека. Более того, для иммунизации мышей могут быть использованы полипептидные фрагменты соответствующих белков, например, α5 и/или β1. Например, полипептидные фрагменты могут быть конъюгированы с молекулой носителя для повышения их иммуногенности. Такие молекулы носителя хорошо известны в данной области, и к ним относятся среди прочих гемоцианин лимфы улитки, бычий сывороточный альбумин, тиреоглобулин, дифтерийный токсоид и столбнячный токсоид.
Подробные процедуры получения полностью человеческих моноклональных антител против α5β1 описаны в примере 1 ниже. Суммирующий эксперимент с большим числом антигенов показал, что трансгенные мыши отвечали при исходной внутрибрюшинной иммунизации (IP) антигеном в полном адъюванте Фрейнда с последующими IP иммунизациями раз в две недели (в итоге до 6) антигеном в неполном адъюванте Фрейнда. Тем не менее, также обнаружено, что эффективны и другие наполнители, отличные от адъюванта Фрейнда. Кроме того, обнаружено, что высоко иммуногенны и целые клетки в отсутствие наполнителя. Иммунный ответ может быть проконтролирован в течение курса протокола иммунизации с помощью образцов плазмы, получаемых с помощью ретроорбитальных кровотечений. Плазма может быть скринирована с помощью ELISA (как описано выше), и мыши с достаточными титрами иммуноглобулина человека против α5β1 могут быть использованы для слияний. Мыши могут быть стимулированы внутривенным введением антигена за 3 дня до забоя и изъятия селезенки. Предполагается, что может потребоваться 2-3 слияния для каждой иммунизации. Каждым антигеном обычно иммунизируют от 6 до 24 мышей. Обычно используют оба штамма НСо7 и НСо12. Кроме того, оба трансгена и НСо7, и НСо12 могут быть совмещены вместе в одной мыши, имеющей два различных трансгена тяжелой цепи человека (НСо7/НСо12). Альтернативно или дополнительно может быть использован штамм КМ Mouse®, как описано в примере 1.
Получение гибридом, продуцирующих моноклональные антитела человека
Для получения гибридом, продуцирующих моноклональные антитела человека по изобретению, спленоциты и/или клетки лимфатических узлов иммунизированных мышей могут быть выделены и слиты с подходящей иммортализованной клеточной линией, такой как миеломная клеточная линия мыши. Полученные гибридомы могут быть скринированы в отношении продукции антигенспецифических антител. Например, суспензии одиночных клеток лимфоцитов селезенки иммунизованных мышей могут быть слиты с клетками несекретирующей миеломы мыши Р3Х63-Ag8.653 (ATCC, CRL 1580) в количестве один к шести с помощью 50% PEG. Клетки помещают в количестве приблизительно 2×105 в плоскодонный планшет для микротитрования с последующей инкубацией в течение двух недель в селективной среде, содержащей 20% сыворотку FetalClone, 18% кондиционированные среды «653», 5% origen (продукция фирмы IGEN), 4 мМ L-глютамин, 1 мМ пируват натрия, 5 мМ HEPES, 0,055 мМ 2-меркаптоэтанол, пенициллин в концентрации 50 Ед/мл, стрептомицин в концентрации 50 мг/мл, гентамицин в концентрации 50 мг/мл и 1-кратную НАТ-добавку (продукция фирмы Sigma; НАТ добавляют через 24 часа после слияния). Примерно через две недели клетки могут быть культивированы в среде, в которой НАТ заменяют на НТ. Отдельные лунки затем могут быть скринированы посредством ELISA в отношении моноклональных антител человека IgM и IgG. После появления распространенного роста гибридомы за средой можно вести наблюдение обычно через 10-14 дней. Гибридомы, секретирующие антитело, могут быть повторно помещены, вновь скринированы, и если все еще позитивны в отношении IgG человека, моноклональные антитела могут быть субклонированы, по меньшей мере, дважды с помощью предельного разведения. Устойчивые субклоны затем могут быть культивированы in vitro для получения небольших количеств антитела в тканевой культуральной среде для характеристики.
Для очистки моноклональных антител человека отобранные гибридомы могут быть выращены в двухлитровых роллерных колбах для очистки моноклональных антител. Супернатанты могут быть отфильтрованы и сконцентрированы перед аффинной хроматографией с помощью белок А-сефарозы (продукция фирмы Pharmacia, Piscataway, N.J.). Для гарантии чистоты элюированный IgG может быть проверен с помощью гель-электрофореза и высокоэффективной жидкостной хроматографии. Буферный раствор может быть заменен на PBS, и концентрация может быть определена по OD280, используя коэффициент гашения 1,43. Моноклональные антитела могут быть разлиты по аликвотам и оставлены на хранение при -80°С.
Получение трансфектом, продуцирующих моноклональные антитела
Антитела по изобретению также могут быть получены в трансфектоме клетки-хозяина, используя, например, сочетание техник рекомбинантных ДНК и способов трансфекции генов, хорошо известные в данной области (например, Morrison S. (1985) Science 229:1202).
Например, для экспрессии антител или их фрагментов молекулы ДНК, кодирующие фрагментарные или полноразмерные легкую и тяжелую цепи, могут быть получены стандартными молекулярно-биологическими техниками (например, ПЦР-амплификацией или клонированием кДНК, используя гибридому или фаг, которые экспрессируют интересующее антитело), и молекулы ДНК могут быть встроены в экспрессирующие векторы, так что гены функционально связываются с последовательностями, контролирующими транскрипцию и трансляцию. В этом контексте под термином «функционально связанный» понимают, что ген антитела лигируют в вектор, так что последовательности, контролирующие транскрипцию и трансляцию в векторе, выполняют намеченную функцию регуляции транскрипции и трансляции гена антитела. Экспрессирующий вектор и последовательности, контролирующие экспрессию, выбирают так, чтобы они были совместимы с используемой экспрессирующей клеткой-хозяином. Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть встроены в отдельные векторы или чаще оба гена встраивают в один и тот же экспрессирующий вектор. Гены антитела встраивают в экспрессирующий вектор любыми подходящими способами (например, лигированием комплементарных сайтов рестрикции на генном фрагменте антитела и векторе или лингированием тупых концов при отсутствии сайтов рестрикции). Вариабельные области легкой и тяжелой цепей антител, описанных в данном документе, могут быть использованы для создания генов полноразмерного антитела любого изотипа и подкласса антитела путем встраивания их в экспрессирующие векторы, уже кодирующие константные области тяжелой и легкой цепей желаемого изотипа и подкласса, таким образом, что VH-сегмент функционально связывается с СН-сегментом(ами) в векторе и VK-сегмент функционально связывается с CL-сегментом в векторе. Дополнительно или альтернативно рекомбинантный экспрессирующий вектор может кодировать сигнальный пептид, который способствует секреции цепи антитела из клетки-хозяина. Ген цепи антитела может быть клонирован в вектор, таким образом, что сигнальный пептид связывается внутри рамки считывания с аминоконцом гена цепи антитела. Сигнальный пептид может представлять собой сигнальный пептид иммуноглобулина или гетерологичный сигнальный пептид (т.е. сигнальный пептид из белка, отличного от иммуноглобулина).
Помимо генов цепей антитела рекомбинантные экспрессирующие векторы по изобретению, как правило, несут регуляторные последовательности, которые контролируют экспрессию генов цепей антитела в клетке-хозяине. Под термином «регуляторная последовательность» понимают промоторы, энхансеры и другие элементы, контролирующие экспрессию (например, сигналы полиаденилирования), которые контролируют транскрипцию или трансляцию генов цепей антитела. Такие регуляторные последовательности описываются, например, в руководстве Goeddel (Gene Expression Technology. Methods in Enzymology 185, Academic Press, san Diego, CA (1990)). Специалисту в данной области будет понятно, что создание экспрессирующего вектора, включая отбор регуляторных последовательностей, может зависеть от таких факторов, как выбор трансформируемой клетки-хозяина, уровень экспрессии желаемого белка и т.д. К предпочтительным регуляторным последовательностям для экспрессии в клетке-хозяине млекопитающего относятся вирусные элементы, которые приводят к высоким уровням экспрессии белка в клетках млекопитающих, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV), вируса обезьян 40 (SV40), аденовируса (например, основной поздний промотор аденовируса (AdMLP)) и вируса полиомы. Альтернативно, могут быть использованы невирусные регуляторные последовательности, такие как убиквитиновый промотор или β-глобиновый промотор. Более того, регуляторные элементы составлены из последовательностей от различных источников, такие как промоторная система SR, которая содержит последовательности из раннего промотора SV40 и длинный терминальный повтор вируса Т-клеточного лейкоза человека типа 1 (Takebe Y. et al. (1988) Mol. Cell. Biol. 8:466-472).
Помимо генов цепей антител и регуляторных последовательностей рекомбинантные экспрессирующие векторы по изобретению могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках-хозяевах (например, точки начала репликации) и гены селектирующих маркеров. Ген селектирующего маркера облегчает селекцию клеток-хозяев, в которые вектор был встроен (см., например, патенты США 4399216, 4634665 и 5179017 авторов Axel et al.). Например, обычно ген селектирующего маркера придает устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клетке-хозяину, в которую вектор был встроен. К генам селектирующих маркеров относятся ген дигидрофолатредуктазы (DHFR) (для применения в клетках-хозяевах dhfr- с селекцией/амплификацией на фоне метотрексата) и ген neo (для селекции на фоне G418).
Для экспрессии легкой и тяжелой цепей экспрессирующий(е) вектор(ы), кодирующие тяжелую и легкую цепи, трансфицируют в клетку-хозяина с помощью любых подходящих техник. Под большим числом форм термина «трансфекция» понимают широкий диапазон техник, обычно используемых для встраивания экзогенной ДНК в прокариотическую или эукариотическую клетку-хозяина, например, электропорацию, преципитацию фосфатом кальция, трансфекцию DEAE-декстраном и тому подобное. Несмотря на возможность экспрессировать антитела по изобретению либо в прокариотических, либо в эукариотических клетках-хозяевах, наиболее распространена экспрессия антител в эукариотических клетках и обычно в клетках-хозяевах млекопитающих.
К клеткам-хозяевам млекопитающих для экспрессии рекомбинантных антител по изобретению относятся, например, клетки яичника китайского хомячка (СНО) (включая клетки СНО dhfr-, описанные в статье Urlaub и Chasin, (1980) Proc. Natl. Acad. Sci. USA 77:4216-4220, используемые с селектирующим маркером DHFR, например, как описано в статье R.J. Kaufman и P.A. Sharp (1982) Mol. Biol. 159:601-621), миеломные клетки NS0, клетки COS и клетки Sp2. В частности, для применения с миеломными клетками NS0 или клетками СНО другая экспрессирующая система представляет собой экспрессирующую систему гена GS (глутаминсинтетазы), описанную в WO 87/04462, WO 89/01036 и EP 333841. При встраивании рекомбинантных экспрессирующих векторов, кодирующих гены антитела, в клетки-хозяева млекопитающих продуцируются антитела посредством культивирования клеток-хозяев в течение периода времени, достаточного для экспрессии антитела в клетках-хозяевах или секреции антитела в культуральную среду, в которой клетки-хозяева выращиваются. Антитела могут быть извлечены из культуральной среды, используя любые подходящие способы очистки белков.
Характеристика связывания антитела с антигеном
Антитела или их антигенсвязывающие участки по изобретению могут быть протестированы на связывание с α5β1, например, с помощью стандартного анализа ELISA. Вкратце, плашки для микротитрования покрывают очищенным α5β1 в концентрации 0,25 мкг/мл в PBS и затем блокируют с помощью 5% бычьего сывороточного альбумина в PBS. В каждую лунку добавляют разведения антитела (например, разведения плазмы, полученной от α5β1-иммунизированных мышей) и инкубируют в течение 1-2 часов при 37°С. Плашки отмывают с помощью PBS/Tween и затем инкубируют с вторичным реагентом (например, для антител человека с Fc-специфичным поликлональным реагентом Ig козы против человека), конъюгированным с щелочной фосфатазой, в течение 1 часа при 37°С. После отмывки плашки отрабатывают субстратом pNPP (1 мг/мл) и анализируют при OD 405-650. Предпочтительно, для слияний должны использоваться мыши, у которых создаются самые высокие титры.
Анализ ELISA, описанный выше, также может использоваться для скрининга гибридом, у которых наблюдается положительная реакционноспособность с иммуногеном α5β1. Гибридомы, которые связываются с α5β1 с высокой авидностью, субклонируют и дополнительно характеризуют. Один клон каждой гибридомы, у которого сохраняется реакционноспособность родительских клеток (с помощью ELISA), может быть отобран для создания банка клеток в объеме 5-10 ампул, хранящихся при -140°С, и для очистки антитела.
Для очистки антител против α5β1 отобранные гибридомы можно выращивать в двухлитровых роллерных колбах для очистки моноклональных антител. Супернатанты могут быть профильтрованы и сконцентрированы перед аффинной хроматографией с помощью белок А-сефарозы (продукция фирмы Pharmacia, Piscataway, N.J.). Чтобы гарантировать чистоту, элюированный IgG может быть проверен с помощью гель-электрофореза и высокоэффективной жидкостной хроматографии. Буферный раствор может быть заменен на PBS, и концентрация может быть определена по OD280, используя коэффициент гашения 1,43. Моноклональные антитела могут быть разлиты по аликвотам и оставлены на хранение при -80°С.
Дополнительно, эпитоп, связывающийся с антителом, может быть охарактеризован стандартными способами, известными в данной области. К таким способам относятся продукция набора перекрывающихся пептидных фрагментов α5 и/или β1 и оценка связывания антитела с большим числом фрагментов. Альтернативно, в пептид α5 и/или β1 могут быть введены мутации, например, с помощью аланин-сканирующего мутагенеза, при котором каждая аминокислота замещается аланиновым остатком, и связывание антитела с мутантным пептидом может быть сравнено со связыванием антитела с белком дикого типа, таким образом, идентифицируя сайты, где мутация(и) влияет(ют) на связывание. См., например, статью Cunningham et al., (1989) Science 244:1081-1085.
Для определения, связываются ли отобранные моноклональные антитела против α5β1 или их антигенсвязывающий участок с уникальными эпитопами, каждое антитело может быть биотинилировано, используя коммерчески доступные реагенты (продукция фирмы Pierce, Rockford, IL). Могут быть осуществлены сравнительные исследования, используя немеченые моноклональные антитела и биотинилированные моноклональные антитела, с применением планшетов для проведения ELISA, покрытых α5β1, как описано выше. Связывание биотинилированных mAb может быть детектировано с помощью зонда стрептавидин-щелочная фосфатаза.
Для определения изотипа очищенных антител могут быть осуществлены анализы ELISA на определение изотипов, используя реагенты, специфичные для антител определенного изотипа. Например, для определения изотипа моноклонального антитела человека лунки планшетов для микротитрования могут быть покрыты иммуноглобулином против человека в концентрации 1 мкг/мл в течение при 4°С. После блокирования с помощью 1% BSA планшеты приводят в контакт с тестируемыми моноклональными антителами или контролями очищенных изотипов в концентрации 1 мкг/мл или меньше при температуре окружающей среды в течение от одного до двух часов. Лунки затем могут быть приведены в контакт либо с IgG1-, либо с IgM человека-специфичными зондами, конъюгированными с щелочной фосфатазой. Планшеты обрабатывают и анализируют, как описано выше.
IgG-иммуноглобулины человека против α5β1 могут быть дополнительно протестированы в отношении взаимодействия с антигеном α5β1 с помощью вестерн-блота. Вкратце, интегрин α5β1 может быть получен и подвергнут электрофорезу в полиакриламидном геле с додецилсульфатом натрия. После электрофореза разделенные антигены переносят на нитроцеллюлозные мембраны, блокируют с помощью 10% фетальной телячьей сыворотки и обрабатывают тестируемыми моноклональными антителами. Связывание IgG человека может быть детектировано, используя IgG против человека, конъюгированный со щелочной фосфатазой, и обработав таблетками субстрата BCIP/NBT (продукция фирмы Sigma Chem. Co., St. Louis, Mo.).
Иммуноконъюгаты
В другом аспекте настоящее изобретение относится к антителу против α5β1 или его фрагменту, конъюгированному с терапевтическим фрагментом, таким как цитотоксин, лекарственное средство (например, иммуносупрессант) или радиотоксин. Такие конъюгаты обозначаются в данном документе как «иммуноконъюгаты». Иммуноконъюгаты, которые включают один или несколько цитотоксинов, обозначаются как «иммунотоксины». К цитотоксину или цитотоксическому агенту относится любой агент, который вреден для клеток (например, уничтожает их). К примерам относятся таксол, цитохалазин В, грамицидин D, этидиумбромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пиромицин и их аналоги или гомологи. К терапевтическим агентам также относятся, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацилдекарбазин), алкилирующие агенты (например, меклоретамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнитол, стрептозотоцин, митомицин С и цис-дихлордиамин платина (II) (DDP) (цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (АМС)) и антимитотические агенты (например, винкристин и винбластин).
К другим примерам терапевтических цитотоксинов, которые могут быть конъюгированы с антителом или его антигенсвязывающим участком по изобретению, относятся дуокармицины, калихемицины, майтансины и ауристатины и их производные. Один из примеров конъюгата антитела с калихемицином коммерчески доступен (MylotargTM; Wyeth-Ayerst).
Цитотоксины могут быть конъюгированы с антителами по изобретению или их антигенсвязывающими участками, используя большое число технологий с применением линкеров. К примерам типов линкеров, которые были использованы для конъюгирования цитотоксина с антителом, относятся, но ими не ограничиваясь, гидразоны, тиоэфиры, сложные эфиры, дисульфиды и пептидсодержащие линкеры. Линкер может быть выбран таким образом, чтобы, например, он был чувствителен к расщеплению низкими величинами рН в лизосомальном компартменте или чувствителен к расщеплению протеазами, такими как протеазы, преимущественно экспрессируемые в опухолевой ткани, такие как катепсины (например, катепсины В, С, D).
Для дополнительного рассмотрения типов цитотоксинов, линкеров и способов конъюгирования терапевтических агентов с антителами см. также статьи Saito G. et al. (2003) Adv. Drug Deliv. Rev. 55:199-215; Trail P.A. et al. (2003) Cancer Immunol. Immunother. 52:328-337; Payne G. (2003) Cancer Cell 3:207-212; Allen T.M. (2002) Nat. Rev. Cancer 2:750-763; Pastan I. and Kreitman R.J. (2002) Curr. Opin. Investig. Drugs 3:1089-1091; Senter P.D. and Springer, C.J. (2001) Adv. Drug Deliv. Rev. 53:247-264.
Антитела или их антигенсвязывающие участки по настоящему изобретению также могут быть конъюгированы с радиоактивным изотопом для получения цитотоксических радиоактивных лекарственных препаратов, также обозначаемых как радиоиммуноконъюгаты. К примерам радиоактивных изотопов, которые могут быть конъюгированы с антителами для диагностического или терапевтического применения, относятся, но ими не ограничиваясь, йод131, индий111, иттрий90 и лютеций177. Способы получения радиоиммуноконъюгатов известны в данной области. Некоторые радиоиммуноконъюгатов коммерчески доступны, включая ZevalinTM (продукция фирмы IDEC Pharmaceuticals) и BexxarTM (продукция фирмы Corixa Pharmaceuticals), и для получения радиоиммуноконъюгатов могут быть использованы аналогичные способы, используя антитела по изобретению.
Конъюгаты антител по изобретению могут быть использованы для модификации данного биологического ответа, и фрагмент лекарственного средства не должен быть истолкован как ограниченный классическими химическими терапевтическими агентами. Например, фрагмент лекарственного средства может представлять собой белок или полипептид, обладающий желаемой биологической активностью. К таким белкам могут относиться, например, ферментативно активный токсин или его активный фрагмент, такой как абрин, рицин А, псевдомонадный экзотоксин или дифтерийный токсин; белок, такой как фактор некроза опухолей или интерферон γ; или модификаторы биологического ответа, такие как, например, лимфокины, интерлейкин-1 («IL-1»), интерлейкин-2 («IL-2»), интерлейкин-6 («IL-6»), гранулоцитоарно-макрофагальный колониестимулирующий фактор («GM-CSF»), гранулоцитоарный колониестимулирующий фактор («G-CSF») или другие факторы роста.
Методики конъюгирования такого терапевтического фрагмента с антителом известны, см., например, Arnon et al., "Monoclonal Antibodies for Immunotargeting of Drugs in Cancer Therapy", in Monoclonal Antibodies and Cancer Therapy, Reisfeld et al. (eds.), pp.243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al, "Antibodies for Drug Delivery", in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp.623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody Carriers of Cytotoxic Agents in Cancer Therapy: A Review", in Monoclonal Antibodies '84: Biological and Clinical Applications, Pinchera et al. (eds.), pp.475-506 (1985); "Analysis, Results, and Future Prospective of The Therapeutic Use of Radiolabeled Antibody in Cancer Therapy", in Monoclonal Antibodies for Cancer Detection and Therapy, Baldwin et al. (eds.), pp.303-16 (Academic Press 1985), and Thorpe et al. "The Preparation and Cytotoxic Properties of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-58 (1982).
Биспецифические молекулы
В другом аспекте настоящее изобретение относится к биспецифическим молекулам, содержащим антитело против α5β1 или его фрагмент по изобретению. Антитело по изобретению или его антигенсвязывающие участки могут образовывать производное или связываться с другой функциональной молекулой, например, другим пептидом или белком (например, другим антителом или лигандом рецептора) с образованием биспецифической молекулы, которая связывается, по меньшей мере, с двумя различными сайтами связывания или мишенными молекулами. Антитело по изобретению может на самом деле образовывать производное или связываться с более чем одной другой функциональной молекулой с образованием мультиспецифических молекул, которые связываются с более чем двумя различными сайтами связывания и/или мишенными молекулами; такие мультиспецифичные молекулы также охватываются термином «биспецифическая молекула», используемым в данном документе. Для создания биспецифической молекулы по изобретению антитело по изобретению может быть функционально связано (например, с помощью химического сочетания, генетического слияния, нековалентной ассоциации или иным образом) с одной или несколькими другими связывающими молекулами, такими как другое антитело, фрагмент антитела, пептид или связывающий миметик, в результате чего получается биспецифическая молекула.
Соответственно, настоящее изобретение относится к биспецифическим молекулам, содержащим, по меньшей мере, одну первую специфичность связывания для α5β1 и вторую специфичность связывания для второго мишенного эпитопа. В конкретном аспекте изобретения второй мишенный эпитоп представляет собой Fc-рецептор, например, FcγRI человека (CD64) или Fcγ-рецептор (CD89). Таким образом, изобретение относится к биспецифическим молекулам, способным связываться как с FcγR или эффекторными клетками, экспрессирующими FcγR (например, моноцитами, макрофагами или полиморфноядерными клетками (PMN), так и с мишенными клетками, экспрессирующими α5β1. Эти биспецифические молекулы нацеливают α5β1-экспрессирующие клетки на эффекторную клетку и запускают активности эффекторной клетки, опосредованные Fc-рецептором, такие как фагоцитоз α5β1-экспрессирующих клеток, антителозависимая клеточно-опосредованная цитотоксичность (ADCC), высвобождение цитокинов или выработка супероксид-аниона.
В одном из аспектов изобретения, в котором биспецифическая молекула является мультиспецифичной, молекула может дополнительно включать третью специфичность связывания помимо специфичности связывания против Fc и специфичности связывания против α5β1. В одном из случаев третья специфичность связывания представляет собой участок против фактора усиления (EF), например, молекулы, которая связывается с поверхностным белком, вовлеченным в цитотоксическую активность, и за счет этого повышает иммунный ответ против мишенной клетки. «Участок против фактора усиления» может представлять собой антитело, функциональный фрагмент антитела или лиганд, который связывается с данной молекулой, например, антигеном или рецептором, и таким образом приводит к усилению влияния детерминант связывания для Fc-рецептора или антигена мишенной клетки. «Участок против фактора усиления» может связываться с Fc-рецептором или антигеном мишенной клетки. Альтернативно, участок против фактора усиления может связываться с объектом, который отличается от объекта, с которым связываются первая и вторая специфичности связывания. Например, участок против фактора усиления может связываться с цитотоксической Т-клеткой (например, через CD2, CD3, CD8, CD28, CD4, CD40, ICAM-1) или другой иммунной клеткой, что приводит к повышенному иммунному ответу против мишенной клетки).
В одном из случаев биспецифические молекулы по изобретению содержат в качестве специфичности связывания, по меньшей мере, одно антитело или фрагмент этого антитела, включая, например, Fab, Fab', F(ab')2, Fv или одноцепочечный Fv. Антитело также может представлять собой димер легких или тяжелых цепей, или любой его минимальный фрагмент, такой как Fv или одноцепочечная конструкция, как описано в патенте США 4946778 авторов Ladner et al.
В одном из случаев специфичность связывания Fcγ-рецептора обеспечивается моноклональным антителом, связывание которого не блокируется иммуноглобулином G человека (IgG). Под используемым в данном документе термином «IgG-рецептор» понимают любой из восьми генов цепи γ, локализованных на хромосоме 1. Эти гены кодируют в целом двенадцать трансмембранных или растворимых изоформ рецептора, которые сгруппированы в три класса Fcγ-рецепторов: FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16). В одном из случаев Fcγ-рецептор является высокоаффинным FcγRI человека. FcγRI человека представляет собой молекулу размером 72 кДа, у которой наблюдается высокое сродство для мономерного IgG (108-109 М-1).
Продукция и характеристика некоторых моноклональных антител против Fcγ описаны авторами Fanger et al. в РСТ публикации WO 88/00052 и в патенте США 4954617. Эти антитела связываются с эпитопом FcγRI, FcγRII или FcγRIII в сайте, который отличается от Fcγ-связывающего сайта рецептора и таким образом их связывание по существу не блокируется физиологическими уровнями IgG. Специфические антитела против FcγRI, используемые по настоящему описанию, представляют собой mAb 22, mAb 32, mAb 44, mAb 62 и mAb 197. Гибридома, продуцирующая mAb 32, доступна из American Type Culture Collection, ATCC под номером доступа НВ9469. В других случаях антитело против Fcγ-рецептора представляет собой гуманизированную форму моноклонального антитела 22 (Н22). Продукция и характеристика антитела Н22 описаны в статье Graziano R.F. et al. (1995) J. Immunol. 155 (10):4996-5002 и РСТ публикации WO 94/10332. Клеточную линию, продуцирующую антитело Н22, депонировали в American Type Culture Collection под наименованием HA022CL1 и присвоили номер доступа CRL 11177.
В других случаях специфичность связывания Fc-рецептора обеспечивается антителом, которое связывается с IgA-рецептором человека, например, Fc-альфа-рецептором (FcαRI (CD89)), связывание которого обычно не блокируется иммуноглобулином А человека (IgA). Под термином «IgA-рецептор» понимают генный продукт одного α-гена (FcαRI), локализованного на хромосоме 19. Известно, что этот ген кодирует несколько альтернативно сплайсируемых трансмембранных изоформ размером от 55 до 110 кДа. FcαRI (CD89) конститутивно экспрессируется на моноцитах/макрофагах, эозинофилах и нейтрофилах, но не на клеточных популяциях, не являющихся эффекторными. FcαRI обладает средним сродством (≈ 5×107 М-1) как к IgA1, так и к IgA2, которое возрастает при экспозиции с цитокинами, такими как G-CSF или GM-CSF (Morton H.C. et al. (1996) Critical Reviews in Immunology 16:423-440). Было описано четыре FcαRI-специфичных моноклональных антитела, идентифицируемых как А3, А59, А62 и А77, которые связываются с FcαRI вне домена, связывающего лиганд IgA (Monteiro R.C. et al. (1992) J. Immunol. 148:1764).
FcαRI и FcγRI представляют собой наглядные триггерные рецепторы для применения в биспецифических молекулах по изобретению, поскольку они (1) экспрессируются в основном на иммунных эффекторных клетках, например, моноцитах, клетках PMN, макрофагах и дендритных клетках; (2) экспрессируются в высоких уровнях (например, 5000-100000 на клетку); (3) представляют собой медиаторы цитотоксической активности (например, ADCC, фагоцитоза); (4) опосредуют усиленную антигенную презентацию антигенов, включая сами антигены, мишенные для них.
Несмотря на то, что предпочтительны моноклональные антитела человека, другими антителами, которые могут использоваться в биспецифических молекулах по изобретению, являются мышиные, химерные и гуманизированные моноклональные антитела.
Биспецифические молекулы по настоящему описанию могут быть получены конъюгированием составляющих специфичностей связывания, например, специфичностей связывания с FcR и с α5β1, используя любые подходящие способы. Например, каждая специфичность связывания биспецифической молекулы может быть получена отдельно и затем конъюгирована одна с другой. Если специфичности связывания представляют собой белки или пептиды, то для ковалентного конъюгирования может быть использовано большое число сочетающих или перекрестно связывающих агентов. К примерам перекрестно связывающих агентов относятся белок А, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 5,5'-дитиобис (2-нитробензойная кислота) (DTNB), орто-фенилендималеимид (oPDM), N-сукцинимидил-3-(2-пиридилдитио) пропионат (SPDP) и сульфосукцинимидил 4-(N-малеимидометил) циклогексан-1-карбоксилат (сульфо-SMCC) (см., например, статьи Karpovsky et al. (1984) J. Exp. Med. 160:1686; Liu, MA et al. (1985) Proc. Natl. Acad. Sci. USA 82:8648). К другим способам относятся способы, описанные в статьях Paulus (1985) Behring Ins. Mitt. No. 78, 118-132; Brennan et al. (1985) Science 229:81-83) и Glennie et al. (1987) J. Immunol. 139:2367-2375). К подходящим конъюгирующим агентам относятся SATA и сульфо-SMCC, оба являются продукцией фирмы Pierce Chemical Co. (Rockford, IL).
Если специфичности связывания представляют собой антитела, они могут быть конъюгированы посредством сульфгидрильной связи двух тяжелых цепей в области С-концов шарнирных областей. В одном из случаев шарнирную область модифицируют так, что бы она перед конъюгацией содержала нечетное количество сульфгидрильных остатков, такое как один остаток.
Альтернативно, обе специфичности связывания могут кодироваться в одном и том же векторе и экспрессироваться и собираться в одной и той же клетке-хозяине. Этот способ можно использовать, в частности, если биспецифическая молекула представляет собой слитый белок mAb × mAb, mAb × Fab, Fab × F(ab')2 или лиганд × Fab. Биспецифическая молекула по изобретению может представлять собой одноцепочечную молекулу, содержащую одно одноцепочечное антитело и одну детерминанту связывания, или одноцепочечную биспецифическую молекулу, содержащую две детерминанты связывания. Биспецифические молекулы могут содержать, по меньшей мере, две одноцепочечных молекулы. Способы получения биспецифических молекул описаны, например, в патенте США 5260203; патенте США 5455030; патенте США 4881175; патенте США 5132405; патенте США 5091513; патенте США 5476786; патенте США 5013653; патенте США 5258498 и патенте США 5482858.
Связывание биспецифических молекул со своими специфическими мишенями может быть подтверждено с помощью, например, фермент-связанного иммуносорбентного анализа (ELISA), радиоиммуноанализа (RIA), анализа FACS, биоанализа (например, ингибирования роста) или анализа вестерн-блот. В каждом из этих анализов, как правило, детектируется наличие особо интересующих комплексов белок-антитело, используя меченый реагент (например, антитело), специфичное для интересующего комплекса. Например, комплексы FcR-антитело могут быть детектированы, используя, например, антитело, связанное с ферментом, или фрагмент антитела, который распознает и специфически связывается с комплексами антитело-FcR. Альтернативно, комплексы могут быть детектированы, используя любой другой иммуноанализ. Например, антитело может быть радиоактивно мечено и использовано в радиоиммуноанализе (RIA) (см., например, Weintraub, B., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986). Радиоактивный изотоп может быть детектирован такими способами, как применение γ-счетчика или сцинтилляционного счетчика или авторадиографией.
Фармацевтические композиции
В другом аспекте настоящее описание относится к композиции, например, фармацевтической композиции, содержащей одно или комбинацию моноклональных антител или их антигенсвязывающего(их) участка(ов) по настоящему изобретению, находящихся в смеси наряду с фармацевтически приемлемым носителем. Такие композиции могут включать одно или комбинацию (например, два или больше различных) антител, или иммуноконъюгатов или биспецифических молекул по изобретению. Например, фармацевтическая композиция по изобретению может содержать комбинацию антител (или иммуноконъюгатов или биспецифических молекул), которые связываются с различными эпитопами на мишенном антигене или которые обладают взаимодополняющими активностями.
Фармацевтические композиции по изобретению также могут применяться в комбинированной терапии, например, в сочетании с другими агентами. Например, комбинированная терапия может включать антитело против α5β1 по настоящему изобретению в сочетании, по меньшей мере, с одним другим противовоспалительным или иммуносупрессивным агентом. Примеры терапевтических агентов, которые могут быть использованы в комбинированной терапии, более подробно описаны ниже в разделе о применении антител по изобретению.
Под используемым в данном документе «фармацевтически приемлемым носителем» понимают любой и все растворители, дисперсионные среды, покрывающие вещества, антибактериальные и противогрибковые агенты, изотоничные и замедляющие всасывание агенты и тому подобное, которые являются физиологически совместимыми. Как правило, носитель подходит для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от пути введения активное соединение, т.е. антитело, его антигенсвязывающий участок, иммуноконъюгат или биспецифическая молекула, может быть покрыто материалом, защищающим соединение от действия кислот и других естественных условий, которые могут инактивировать соединение.
В некоторых вариантах осуществления антитела по настоящему изобретению могут находиться в нейтральной форме (включая цвиттерионные формы) или в виде положительно или отрицательно заряженных соединений. В некоторых случаях антитела могут находиться в комплексе с контр-ионом, образуя фармацевтически приемлемую соль. Таким образом, фармацевтические соединения по изобретению могут включать одну или несколько фармацевтически приемлемых солей.
Под «фармацевтически приемлемой солью» понимают соль, которая сохраняет желаемую биологическую активность исходного соединения (например, антитела) и не оказывает нежелательных токсикологических эффектов (см., например, статью Berge, S.M., et al. (1977) J. Pharm. Sci. 66:1-19). Например, к термину «фармацевтически приемлемая соль» относится комплекс, содержащий одно или несколько антител и один или несколько контр-ионов, где контр-ионы происходят из фармацевтически приемлемых неорганических и органических кислот и оснований.
К примерам таких солей относятся кислотно-аддитивные соли и основно-аддитивные соли. К кислотно-аддитивным солям относятся соли, полученные из нетоксичных неорганических кислот, таких как соляная, азотная, фосфорная, серная, бромистоводородная, йодистоводородная, фосфористая и тому подобное, а также из нетоксичных органических кислот, таких как алифатические моно- и дикарбоновые кислоты, фенил-замещенные алкановые кислоты, гидроксиалкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты и тому подобное. К основно-аддитивным солям относятся соли, происходящие из щелочноземельных металлов, таких как натрий, калий, магний, кальций и тому подобное, а также из нетоксичных органических аминов, таких как N,N'-дибензилэтилендиамин, N-метилглюкамин, хлорпрокаин, холин, диэтаноламин, этилендиамин, прокаин и тому подобное.
Более того, к фармацевтически приемлемым неорганическим основаниям относятся ионы металлов. К ионам металлов относятся, но ими не ограничиваясь, соответствующие соли щелочных металлов, соли щелочноземельных металлов и другие физиологически приемлемые ионы металлов. К солям, происходящим из неорганических оснований, относятся соли алюминия, аммония, кальция, кобальта, никеля, молибдена, ванадия, марганца, хрома, селена, олова, меди, окисного железа, закисного железа, лития, магния, соли марганца трехвалентного, марганца двухвалентного, калия, рубидия, натрия и цинка и в своей обычной валентности.
Фармацевтически приемлемые кислотно-аддитивные соли антител по настоящему изобретению могут быть получены из следующих кислот, включая без ограничения муравьиную, уксусную, ацетамидобензойную, адипиновую, аскорбиновую, борную, пропионовую, бензойную, камфорную, карбоновую, цикламиновую, дегидрохолиевую, малоновую, этилендиаминтетрауксусную, этилсерную, fendizoic, метафосфорную, янтарную, гликолевую, глюконовую, молочную, яблочную, винную, таниновую, лимонную, азотную, аскорбиновую, глюкуроновую, малеиновую, фолиевую, фумаровую, пропионовую, пировиноградную, аспарагиновую, глутаминовую, бензойную, соляную, бромистоводородную, йодистоводородную, лизин, изолимонную, трифторуксусную, памовую, пропионовую, ортоаминобензойную, мезиловую, оротовую, щавелевую, щавелевоуксусную, олеиновую, стеариновую, салициловую, аминосалициловую, кремниевую, пара-гидроксибензойную, никотиновую, фенилуксусную, миндальную, эмбоновую, сульфоновую, метансульфоновую, фосфорную, фосфониевую, этансульфоновую, этандисульфоновую, аммоний, бензолсульфоновую, пантотеновую, нафталенсульфоновую, толуолсульфоновую, 2-гидроксиэтансульфоновую, сульфаниловую, серную, азотную, азотистую, монометиловый эфир серной кислоты, циклогексиламиносульфоновую, β-гидроксибутировую, глицин, глицилглицин, глутаминовую, кокадилат, диаминогексановую, камфорсульфоновую, глюконовую, тиоциановую, оксоглютаровую, пиридоксаль-5-фосфат, хлорфеноксиуксусную, ундекановую, N-ацетил-L-аспарагиновую, муциновую и галактуроновую кислоты.
К фармацевтически приемлемым органическим основаниям относятся триметиламин, диэтиламин, N,N'-дибензилэтилендиамин, хлорпрокаин, холин, дибензиламин, диэтаноламин, этилендиамин, меглюмин (N-метилглюкамин), прокаин, циклические амины, катионы четвертичного аммония, аргинин, бетаин, каффеин, клемизол, 2-этиламиноэтанол, 2-диэтиламиноэтанол, 2-диметиламиноэтанол, этандиамин, бутиламин, этаноламин, этилендиамин, N-этилморфолин, N-этилпиперидин, этилглюкамин, глюкамин, глюкозамин, гистидин, гидрабамин, имидазол, изопропиламин, метилглюкамин, морфолин, пиперазин, пиридин, пиридоксин, неодимий, пиперидин, полиаминовые смолы, прокаин, пурины, теобромин, триэтиламин, трипропиламин, триэтаноламин, трометамин, метиламин, таурин, холат, 6-амино-2-метил-2-гептанол, 2-амино-2-метил-1,3-пропандиол, 2-амино-2-метил-1-пропанол, алифатические моно- и дикарбоновые кислоты, фенил-замещенные алкановые кислоты, гидроксиалкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты, стронций, трицин, гидразин, фенилциклогексиламин, 2-(N-морфолино)этансульфоновая кислота, бис(2-гидроксиэтил)амино-трис(гидроксиметил)метан, N-(2-ацетамидо)-2-аминоэтансульфоновая кислота, 1,4-пиперазиндиэтансульфоновая кислота, 3-морфолино-2-гидроксипропансульфоновая кислота, 1,3-бис[трис (гидроксиметил)метиламино]пропан, 4-морфолинпропансульфоновая кислота, 4-(2-гидроксиэтил)пиперазин-1-этансульфоновая кислота, 2-[(2-гидрокси-1,1-бис(гидроксиметил)этил)амино]этансульфоновая кислота, N,N-бис(2-гидроксиэтил)-2-аминоэтансульфоновая кислота, 4-(N-морфолино)бутансульфоновая кислота, 3-(N,N-бис[2-гидроксиэтил]амино)-2-гидроксипропансульфоновая кислота, 2-гидрокси-3-[трис(гидроксиметил)метиламино]-1-пропансульфоновая кислота, 4-(2-гидроксиэтил)пиперазин-1-(2-гидроксипропансульфоновая кислота), пиперазин-1,4-бис(2-гидроксипропансульфоновая кислота)дигидрат, 4-(2-гидроксиэтил)-1-пиперазинпропансульфоновая кислота, N,N-бис (2-гидроксиэтил)глицин, N-(2-гидроксиэтил)пиперазин-N'-(4-бутансульфоновая кислота), N-[трис(гидроксиметил)метил]-3-аминопропансульфоновая кислота, N-трис(гидроксиметил)метил-4-аминобутансульфоновая кислота, N-(1,1-диметил-2-гидроксиэтил)-3-амино-2-гидроксипропансульфоновая кислота, 2-(циклогексиламино)этансульфоновая кислота, 3-(циклогексиламино)-2-гидрокси-1-пропансульфоновая кислота, 3-(циклогексиламино)-1-пропансульфоновая кислота, N-(2-ацетамидо)иминодиуксусная кислота, 4-(циклогексиламино)-1-бутансульфоновая кислота, N-[трис(гидроксиметил)метил]глицин, 2-амино-2-(гидроксиметил)-1,3-пропандиол и трометамол.
Фармацевтическая композиция по изобретению также может включать фармацевтически приемлемый антиоксидант. К примерам фармацевтически приемлемых антиоксидантов относятся: (1) водорастворимые антиоксиданты, такие как аскорбиновая кислота, цистеингидрохлорид, бисульфат натрия, метабисульфит натрия, сульфит натрия и тому подобное; (2) жирорастворимые антиоксиданты, такие как аскорбилпальмитат, бутилированный гидроксианизол (ВНА), бутилированный гидрокситолуол (ВНТ), лецитин, пропилгаллат, альфа-токоферол и тому подобное; и (3) хелирующие агенты, связывающие ионы металлов, такие как лимонная кислота, этилендиаминтетрауксусная кислота (EDTA), сорбитол, винная кислота, фосфорная кислота и тому подобное.
К примерам подходящих водных и неводных носителей, которые могут быть использованы в фармацевтических композициях по изобретению, относятся вода, этанол, многоатомные спирты (такие как глицерин, пропиленгликоль, полиэтиленгликоль и тому подобное) и их подходящие смеси, растительные масла, такие как оливковое масло, и сложные органические эфиры для инъекций, такие как этилолеат. Присущее состояние текучести можно сохранить, например, используя покрывающие материалы, такие как лецитин, поддерживая желаемый размер частиц в случае дисперсий и применяя поверхностно-активные вещества.
Эти композиции также могут содержать наполнители, такие как консерванты, увлажняющие агенты, эмульгирующие агенты и диспергирующие агенты. Предохранение от микроорганизмов может быть обеспечено как с помощью процедур стерилизации, выше, так и путем включения большого числа антибактериальных и противогрибковых агентов, например, парабена, хлорбутанола, фенолсорбиновой кислоты и тому подобного. В композиции также может быть желательно включить изотоничные агенты, такие как сахара, хлорид натрия и тому подобное. Кроме того, длительная абсорбция инъецируемой фармацевтической формы может быть обусловлена включением агентов, которые задерживают абсорбцию, таких как моностеарат алюминия и желатин.
К фармацевтически приемлемым носителям относятся стерильные водные растворы или дисперсии и стерильные порошки для экстемпорального приготовления стерильных растворов или дисперсии для инъекций. Применение таких сред и агентов для фармацевтически активных веществ известно в данной области. За исключением случаев, когда какие-либо принятые среды или агент несовместимы с активным соединением, предполагается применение их в фармацевтических композициях по изобретению. В композиции также могут быть введены дополнительные активные соединения.
Терапевтические композиции, как правило, должны быть стерильными и устойчивыми в условиях производства и хранения. Композиция может находиться в смеси в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для создания высокой концентрации лекарственного средства. Носитель может представлять собой растворитель или дисперсионную среду, содержащую, например, воду, этанол, многоатомный спирт (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и тому подобное) и их подходящие смеси. Собственная текучесть может быть сохранена, например, используя покрытие, такое как лецитин, сохраняя желаемый размер частиц в случае дисперсии и применяя поверхностно-активные вещества. Во многих случаях будет предпочтительно включать в композицию изотоничные агенты, например, сахара, многоатомные спирты, такие как маннитол, сорбитол или хлорид натрия. Длительная абсорбция инъецируемых композиций может быть обусловлена включением в композицию агента, который задерживает абсорбцию, например, солей моностеаратов и желатина.
Стерильные растворы для инъекций могут быть получены введением активного соединения в желательном количестве в подходящем растворителе с одним или комбинацией желательных ингредиентов, перечисленных выше, с последующей стерилизацией микрофильтрацией. В целом, дисперсии получают путем введения активного соединения в стерильный носитель, который содержит основную дисперсионную среду и желательные другие ингредиенты, перечисленные выше. В случае стерильных порошков для получения стерильных растворов для инъекций, к способам получения относятся, но ими не ограничиваясь, высушивание вакуумом и сушка с замораживанием (лиофилизация), которые приводят к получению порошка активного ингредиента плюс любой дополнительный желательный ингредиент из их раствора, заранее простерилизованного фильтрацией.
Количество активного ингредиента, которое может быть скомбинировано с материалом носителя для продукции единичной лекарственной формы, будет изменяться в зависимости от индивидуума, получающего лечение, и конкретного способа введения. Количество активного ингредиента, которое может быть скомбинировано с материалом носителя для продукции единичной лекарственной формы, будет в целом составлять то количество композиции, которое создает терапевтический эффект. В целом, из ста процентов это количество будет находиться в диапазоне от около 0,01 процента до около девяноста девяти процентов активного ингредиента, предпочтительно от около 0,1 процента до около 70 процентов, самое предпочтительное от около 1 процента до около 30 процентов активного ингредиента в сочетании с фармацевтически приемлемым носителем.
Схемы приема доводят для получения оптимального желаемого ответа (например, терапевтического ответа). Например, может быть введена единственная таблетка, может быть введено несколько дробных доз в течение периода времени или доза может быть пропорционально снижена или увеличена, как укажет тяжесть терапевтической ситуации. Для легкости введения и однотипности дозы особенно предпочтительно составлять парентеральные композиции в стандартной лекарственной форме. Под стандартной лекарственной формой, используемой в настоящем описании, понимают физически дискретные единицы, подходящие в качестве однократных доз для индивидуумов, получающих лечение; каждая единица содержит заранее определенное количество активного соединения, рассчитанное для продукции желаемого терапевтического эффекта в ассоциации с желаемым фармацевтическим носителем. Спецификация стандартных лекарственных форм по изобретению определяется и непосредственно зависит от (а) уникальных характеристик активного соединения и конкретного достигаемого терапевтического эффекта и (b) ограничений, существующих в данной области приготовления соединений, таких как активное соединение для лечения чувствительности у индивидуумов.
Для введения антитела доза находится в диапазоне от около 0,0001 до 100 мг/кг и чаще от 0,01 до 5 мг/кг массы тела хозяина. Например, дозы могут составлять 0,3 мг/кг массы тела, 1 мг/кг массы тела, 3 мг/кг массы тела, 5 мг/кг массы тела или 10 мг/кг массы тела, или находиться в диапазоне от 1 до 10 мг/кг. Характерная схема приема включает введение один раз в неделю, один раз в две недели, один раз в три недели, один раз в четыре недели, один раз в месяц, один раз в 3 месяца или один раз каждые от трех до 6 месяцев. Схемы приема антитела против α5β1 или его антигенсвязывающего участка по изобретению включают, например, 1 мг/кг массы тела или 3 мг/кг массы тела путем внутривенного введения, причем антитело получают, используя одну из следующих схем приема: (i) шесть доз один раз в четыре недели, затем раз в три месяца; (ii) раз в три недели; (iii) однократно 3 мг/кг массы тела, после чего 1 мг/кг массы тела раз в три недели.
В некоторых способах два или несколько моноклональных антител с различными специфичностями связывания вводятся одновременно, в этом случае доза каждого вводимого антитела снижается в указанных диапазонах. Антитело обычно вводят большое число раз. Интервалы между единичными дозами могут составлять, например, раз в неделю, раз в месяц, раз в три месяца или раз в год. Интервалы также могут быть нерегулярными, указываемыми измерением в крови уровней антитела для мишенного антигена у пациента. В некоторых способах дозу доводят до достижения концентрации антитела в плазме от около 1 до 1000 мкг/мл и в некоторых способах от около 25 до 300 мкг/мл.
Альтернативно, антитело может быть введено в виде препаративной формы длительного высвобождения, в этом случае требуется меньшая частота введения. Доза и частота варьируют в зависимости от периода полувыведения антитела у пациента. В целом, у антител человека наблюдается самый длинный период полувыведения, далее следуют гуманизированные антитела, химерные антитела и антитела, отличные от человеческих. Доза и частота введения могут варьировать в зависимости от того, является ли лечение профилактическим или терапевтическим. При профилактическом применении вводится относительно низкая доза с относительно длительными интервалами в течение длительного периода времени. Некоторые пациенты продолжают получать лечение на протяжении всей жизни. При терапевтическом применении иногда требуется относительно высокая доза при относительно коротких промежутках до снижения или прекращения прогрессирования заболевания, и предпочтительно до тех пор, пока у пациента не будет наблюдаться частичного или полного смягчения симптомов заболевания. После чего пациент может получать профилактическую схему.
Фактические уровни дозировки активных ингредиентов в фармацевтических композициях по настоящему описанию могут варьировать таким образом, чтобы получить количество активного ингредиента, которое эффективно для достижения желаемого терапевтического ответа у конкретного пациента, композиции и пути введения, не являясь токсичным для пациента. Выбранный уровень дозировки будет зависеть от большого числа фармакокинетических факторов, включающих активность используемых конкретных композиций по настоящему описанию или их сложного эфира, соли или амида, способ введения, время введения, скорость выведения конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, используемые в сочетании с конкретными используемыми композициями, возраст, пол, вес, состояние, общее состояние здоровья и предшествующий анамнез пациента, получающего лечение, и подобные факторы, хорошо известные в данных областях медицины.
«Терапевтически эффективная доза» антитела против α5β1 по изобретению предпочтительно приводит к снижению тяжести симптомов заболевания, увеличению частоты и продолжительности периодов с отсутствием симптомов заболевания или предотвращению ухудшения или нарушения вследствие тяжести заболевания. Например, при лечении α5β1-положительных опухолей «терапевтически эффективная доза» предпочтительно ингибирует клеточный рост или рост опухоли, по меньшей мере, на около 20%, более предпочтительно, по меньшей мере, на около 40%, еще более предпочтительно, по меньшей мере, на около 60% и самое предпочтительное, по меньшей мере, на около 80% относительно индивидуумов, не получавших лечение. Способность соединения ингибировать опухолевый рост может быть оценена на модельной системе на животных, предсказывающей эффективность для опухолей человека. Альтернативно, это свойство композиции может быть оценено по способности соединения ингибировать, такое ингибирование in vitro может быть оценено посредством анализов, известных специалистам-практикам. Терапевтически эффективное количество терапевтического соединения может уменьшать размер опухоли или иным образом облегчать симптомы у индивидуума. Специалист в данной области смог бы определить такие количества на основании таких факторов как величина индивидуума, тяжесть симптомов, наблюдаемых у индивидуума, и выбранные конкретная композиция или способ введения.
Композиция по настоящему описанию может быть введена одним или несколькими путями введения, используя один или несколько из большого числа способов, известных в данной области. Как будет понятно специалисту в данной области, путь и/или способ введения будут меняться в зависимости от желаемых результатов. К путям введения антител или их антигенсвязывающих участков по изобретению относятся внутривенный, внутримышечный, внутридермальный, внутрибрюшинный, подкожный, спинальный или другие парентеральные пути введения, например, с помощью инъекции или инфузии. Под фразой «парентеральное введение», используемой в данном документе, понимают способы введения, отличные от энтерального и местного введения, обычно посредством инъекции, и к ним относятся без ограничения внутривенная, внутримышечная, внутриартериальная, внутриоболочечная, внутрикапсулярная, интраорбитальная, внутрисердечная, внутридермальная, внутрибрюшинная, транстрахеальная, подкожная, субкутикулярная, внутрисуставная, субкапсулярная, субарахноидальная, внутриспинальная, эпидуральная и надчревная инъекция и инфузия.
Альтернативно, антитело или его антигенсвязывающий участок по изобретению может быть введено путем, отличным от парентерального, таким как местный, эпидеральный путь введения или путь введения через слизистую оболочку, например, интраназально, перорально, вагинально, ректально, сублингвально или местно.
Активные соединения могут быть получены с носителями, которые будут защищать соединение от быстрого высвобождения, такими как препаративная форма с контролируемым высвобождением, включающая импланты, трансдермальные пластыри и микроинкапсулированные системы доставки. Могут быть использованы биодеградируемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры, полимолочная кислота. Многие способы получения таких препаративных форм запатентованы или в основном известны специалистам в данной области. См., например, Sustained and Controlled release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Терапевтические композиции могут быть введены с помощью медицинских устройств, известных в данной области. Например, терапевтическая композиция по изобретению может быть введена с помощью безигольного устройства для гиподермальной инъекции, такого как устройства, описанные в патентах США 5399163; 5383851; 5312335; 5064413; 4941880; 4790824 или 4596556. К примерам хорошо известных имплантов и модулей по настоящему описанию относятся: патент США 4487603, в котором описывается имплантируемая микроинфузионная помпа для введения лекарственного препарата с контролируемой скоростью; патент США 4486194, в котором описывается терапевтическое устройство для введения лекарственных препаратов через кожу; патент США 4447233, в котором описывается инфузионная помпа для доставки лекарственного препарата с точной скоростью инфузии; патент США 4447224, в котором описывается имплантируемый аппарат для инфузии с изменяемым потоком для непрерывной доставки лекарственного средства; патент США 4439196, в котором описывается осмотическая система доставки лекарственного средства, имеющая многокамерные отделы; и патент США 4475196, в котором описывается осмотическая система доставки лекарственного средства. Специалистам в данной области известны и многие другие такие импланты, системы доставки и модули.
В некотором случае моноклональные антитела человека или их антигенсвязывающие участки по изобретению могут быть смешаны для достижения подходящего распределения in vivo. Например, гематоэнцефалический барьер (ВВВ) не пропускает многие высокогидрофильные соединения. Для прохождения терапевтических соединений по изобретению через ВВВ (при необходимости) они могут быть смешаны, например, в липосомах. В отношении способов производства липосом, см., например, патенты США 4522811; 5374548 и 5399331. Липосомы могут содержать один или несколько фрагментов, которые избирательно транспортируются в специфические клетки или органы, таким образом усиливая доставку нацеленного лекарственного средства (см., например, статью V.V. Ranade (1989) J. Clin. Pharmacol. 29:685). К типичным нацеленным фрагментам относятся фолат или биотин (см., например, патент США 5416016 авторов Low et al.); маннозиды (Umezawa et al., (1988) Biochem. Biophys. Res. Commun. 153:1038); антитела (P.G. Bloeman et al. (1995) FEBS Lett. 357:140; M. Owais et al. (1995) Antimicrob. Agents Chemother. 39:180); рецептор сурфактантного белка А (Briscoe et al. (1995) Am. J. Physiol. 1233:134); p120 (Schreier et al. (1994) J. Biol. Chem. 269:9090); также см. K. Keinanen; M.L. Laukkanen (1994) FEBS Lett. 346:123; J.J. Killion; I.J. Fidler (1994) Immunomethods 4:273.
Применение и способы по изобретению
Антитела, в частности антитела человека, композиции антител и способы по настоящему описанию имеют большое число in vitro и in vivo диагностических и терапевтических применений, включая диагностику и лечение α5β1-опосредованных расстройств. Например, эти молекулы могут быть введены в клетки в культуре in vitro или ех vivo, или человеку, например, in vivo, для лечения, профилактики и диагностики большого числа заболеваний. Под используемым в данном документе термином «индивидуум» понимают человека и животных. К животным относятся все позвоночные, например, млекопитающие и животные, отличные от млекопитающих, такие как приматы, овцы, собаки, кошки, коровы, лошади, курицы, земноводные и рептилии. К предпочтительным индивидуумам относятся люди, имеющие расстройства, опосредованные активностью α5β1. Способы особенно подходят для лечения людей, страдающих расстройством, ассоциированным с аномальной экспрессией α5β1. При введении антител против α5β1 наряду с другим агентом они могут быть введены в любом порядке или одновременно.
Принимая во внимание специфическое связывание антител по изобретению с α5β1, антитела по изобретению могут быть использованы для специфического определения экспрессии α5β1 на поверхности клеток и, более того, могут быть использованы для очистки α5β1 путем иммуноаффинной очистки.
Более того, принимая во внимание экспрессию α5β1 на большом числе опухолевых клеток (см., например, пример 6, фиг.7) и его вовлеченность в ангиогенез, антитела человека, композиции антител и способы по настоящему описанию могут быть использованы для лечения индивидуума с аномальным клеточным ростом, например, страдающего расстройством, отличающимся наличием опухолевых клеток, экспрессирующих α5β1, включая, например, мезотелиому, рак гепатобилиарной системы (печени и желчевыводящих путей), первичную или вторичную опухоль ЦНС, первичную или вторичную опухоль мозга, рак легких (NSCLC и SCLC), рак костей, рак поджелудочной железы, рак кожи, рак головы или шеи, меланому кожи или внутриглазную меланому, рак яичников, рак толстого кишечника, рак прямой кишки, анальный рак, рак желудка, рак органов желудочно-кишечного тракта (желудка, колоректальный и 12 перстной кишки), рак молочных желез, рак матки, карциному фаллопиевых труб, карциному эндометрия, карциному цервикального канала, карциному вагины, карциному вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечников, саркому мягких тканей, рак мочеиспускательного канала, рак пениса, рак простаты, рак яичка, хронический или острый лейкоз, хронический миелоидный лейкоз, лимфоцитарные лимфомы, рак мочевого пузыря, рак почки или мочеточника, почечно-клеточную карциному, карциному почечной лоханки, новообразования центральной нервной системы (ЦНС), первичную лимфому ЦНС, неходжкинскую лимфому, опухоли спинного мозга, глиому ствола мозга, аденому гипофиза, адренокортикальный рак, рак желчного пузыря, множественную миелому, холангиокарциному, фибросаркому, нейробластому, ретинобластому или сочетание одной или нескольких вышеуказанных злокачественных опухолей.
В одном из случаев антитела (например, моноклональные антител человека, мультиспецифические и биспецифические молекулы и композиции) или их антигенсвязывающие участки по изобретению могут быть использованы для детекции уровней α5β1 или уровней клеток, которые содержат α5β1 на своей мембранной поверхности, причем эти уровни затем могут быть связаны с некоторыми симптомами заболевания. Альтернативно, антитела могут быть использованы для ингибирования или блокирования функции α5β1, что, в свою очередь, может быть связано с предотвращением или смягчением некоторых симптомов заболевания, таким образом, принимая α5β1 в качестве медиатора заболевания. Этого можно достичь приведением образца и контрольного образца в контакт с антителом против α5β1 в условиях, которые способствуют образованию комплекса между антителом и α5β1. Любые комплексы, образованные между антителом и α5β1, детектируют и сравнивают в образце и контроле.
В другом случае антитела (например, антитела человека, мультиспецифические и биспецифические молекулы и композиции) или их антигенсвязывающие участки по изобретению могут быть изначально протестированы в отношении связывающей активности, ассоциированной с терапевтическим или диагностическим применением in vitro. Например, композиции по изобретению могут быть протестированы, используя анализы проточной цитометрии, описанные в примерах ниже.
Антитела (например, антитела человека, мультиспецифические и биспецифические молекулы, иммуноконъюгаты и композиции) или их антигенсвязывающие участки по изобретению дополнительно используют в лечении и диагностике заболеваний, связанных с α5β1. Например, моноклональные антитела человека, мультиспецифические или биспецифические молекулы и иммуноконъюгаты могут быть использованы для выявления in vivo или in vitro одной или нескольких из следующих биологических активностей: ингибирование роста и/или уничтожение клетки, экспрессирующей α5β1; опосредование фагоцитоза или ADCC клетки, экспрессирующей α5β1, при наличии эффекторных клеток человека, или блокирование связывания лиганда α5β1 с α5β1.
В конкретном случае антитела (например, антитела человека, мультиспецифические и биспецифические молекулы и композиции) или их антигенсвязывающие участки используют in vivo для лечения, профилактики или диагностики большого числа заболеваний, связанных с α5β1. К примерам заболеваний, связанных с α5β1, относятся в числе прочих аномальный клеточный рост, такой как рак. В одном из вариантов осуществления аномальный клеточный рост представляет собой рак, включая, но ими не ограничиваясь, мезотелиому, рак гепатобилиарной системы (печени и желчевыводящих путей), первичную или вторичную опухоль ЦНС, первичную или вторичную опухоль мозга, рак легких (NSCLC и SCLC), рак костей, рак поджелудочной железы, рак кожи, рак головы или шеи, меланому кожи или внутриглазную меланому, рак яичников, рак толстого кишечника, рак прямой кишки, анальный рак, рак желудка, рак органов желудочно-кишечного тракта (желудка, колоректальный и 12 перстной кишки), рак молочных желез, рак матки, карциному фаллопиевых труб, карциному эндометрия, карциному цервикального канала, карциному вагины, карциному вульвы, болезнь Ходжкина, рак пищевода, рак тонкого кишечника, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечников, саркому мягких тканей, рак мочеиспускательного канала, рак пениса, рак простаты, рак яичка, хронический или острый лейкоз, хронический миелоидный лейкоз, лимфоцитарные лимфомы, рак мочевого пузыря, рак почки или мочеточника, почечно-клеточную карциному, карциному почечной лоханки, новообразования центральной нервной системы (ЦНС), первичную лимфому ЦНС, неходжкинскую лимфому, опухоли спинного мозга, глиому ствола мозга, аденому гипофиза, адренокортикальный рак, рак желчного пузыря, множественную миелому, холангиокарциному, фибросаркому, нейробластому, ретинобластому или сочетание одной или нескольких вышеуказанных злокачественных опухолей.
Подходящие пути введения композиций антител (например, моноклональных антител человека, мультиспецифических и биспецифических молекул и иммуноконъюгатов) или их антигенсвязывающих участков по изобретению in vivo и in vitro хорошо известны в данной области и могут быть выбраны специалистом в данной области. Например, композиции антител могут быть введены с помощью инъекции (например, внутривенной или подкожной). Подходящие дозы используемых молекул будут зависеть от возраста и веса индивидуума и концентрации и/или состава композиции антител.
Ранее описанные антитела человека против α5β1 или их антигенсвязывающие участки по изобретению могут быть введены совместно с одним или несколькими другими терапевтическими агентами, например, цитотоксическим агентом, радиотоксическим агентом или иммуносупрессивным агентом. Антитело может быть связано с агентом (в виде иммунокомплекса) или может быть введено отдельно от агента. В последнем случае (раздельное введение) антитело может быть введено до, после или одновременно с агентом или может быть введено совместно с другими известными способами лечения, например, противоракового лечения, например, облучением. К таким терапевтическим агентам относятся среди прочих антинеопластические агенты, такие как доксорубицин (адриамицин), цисплатин, сульфат блеомицина, кармустин, хлорамбуцил и циклофосфамид гидроксимочевина, которые сами по себе эффективны лишь при уровнях, токсичных или субтоксичных для пациента. Цисплатин может быть введен внутривенно по 100 мг/доза раз в четыре недели, и адриамицин вводят внутривенно по 60-75 мг/мл дозы один раз в 21 день. Сочетанное введение антител человека против α5β1 или их антигенсвязывающих фрагментов по настоящему изобретению с химиотерапевтическими агентами предоставляет два противораковых агента, которые влияют через различные механизмы, что приводит к цитотоксическому эффекту для опухолевых клеток человека. Такое сочетанное введение может решить проблемы, вызванные развитием устойчивости к лекарственным средствам или изменением антигенности опухолевых клеток, что сделало бы их невосприимчивыми к антителу.
Мишень-специфичные эффекторные клетки, например, эффекторные клетки, связанные с композициями (например, антителами человека, мультиспецифическими и биспецифическими молекулы),по изобретению также могут быть использованы в качестве терапевтических агентов. Эффекторные клетки для нацеливания могут представлять собой лейкоциты человека, такие как макрофаги, нейтрофилы или моноциты. К другим клеткам относятся эозинофилы, естественные клетки-киллеры и другие клетки, несущие рецепторы IgG или IgA. При необходимости эффекторные клетки могут быть получены от индивидуума, получающего лечение. Мишень-специфичные эффекторные клетки могут быть введены в виде суспензии клеток в физиологически приемлемом растворе. Количество вводимых клеток может составлять порядка от 108 до 109, но будет меняться в зависимости от лечебной цели. В целом, количество должно быть достаточным для достижения локализации на мишенной клетке, например, опухолевой клетке, экспрессирующей α5β1, и для эффекта уничтожения клетки посредством, например, фагоцитоза. Пути введения также могут варьировать.
Лечение с помощью мишень-специфичных эффекторных клеток может быть осуществлено наряду с другими техниками удаления мишеневых клеток. Например, противоопухолевая терапия с применением композиций (например, антител человека, мультиспецифических и биспецифических молекул) по изобретению и/или эффекторных клеток, усиленных этими композициями, может быть использована наряду с химиотерапией. Кроме того, комбинированная иммунотерапия может быть использована для направления двух различных популяций цитотоксических эффекторных клеток на отторжение опухолевых клеток. Например, антитела против α5β1, связанные с рецептором анти-Fc-гамма RI или анти-CD3, могут быть использованы наряду с агентами, специфически связывающимися с рецептором IgG или IgA.
Биспецифические и мультиспецифические молекулы по изобретению также могут быть использованы для модуляции FcγR или уровней FcγR на эффекторных клетках, такой как кэппинг и устранение рецепторов на поверхности клетки. Для этой цели также могут быть использованы смеси анти-Fc-рецепторов.
При наличии комплемента также могут быть использованы композиции (например, антитела человека, гуманизированные или химерные антитела, мультиспецифические и биспецифические молекулы и иммуноконъюгаты) по изобретению, которые имеют сайты связывания комплемента, такие как участки из IgG1, -2 или -3, или IgM, которые связывают комплемент. В одном из случаев обработка ex vivo популяции клеток, содержащих мишенные клетки, с помощью связывающего агента по изобретению и подходящих эффекторных клеток может быть дополнена добавлением комплемента или сыворотки, содержащей комплемент. Фагоцитоз мишенных клеток, покрытых связывающим агентом по изобретению, может быть улучшен за счет связывания белков комплемента. В другом случае мишенные клетки, покрытые композициями (например, антителами человека, мультиспецифическими и биспецифическими молекулами) по изобретению также могут быть лизированы с помощью комплемента. В еще одном случае композиции по изобретению не активируют комплемент.
Композиции (например, антитела человека, гуманизированные или химерные антитела, мультиспецифические и биспецифические молекулы и иммуноконъюгаты) по изобретению также могут быть введены наравне с комплементом. Соответственно, изобретение охватывает композиции, содержащие антитела человека, мультиспецифические или биспецифические молекулы и сыворотку или комплемент. Эти композиции обладают преимуществом, поскольку комплемент находится в непосредственной близости к антителам человека, мультиспецифическим или биспецифическим молекулам. Альтернативно, антитела человека, мультиспецифические или биспецифические молекулы по изобретению и комплемент или сыворотка могут быть введены раздельно.
Также настоящее изобретение охватывает наборы, содержащие композиции антител по изобретению (например, антител человека, биспецифических или мультиспецифических молекул или иммуноконъюгатов) и инструкции по применению. Набор может дополнительно содержать один или несколько дополнительных реагентов, таких как иммуносупрессивный реагент, цитотоксический агент или радиотоксический агент, или одно или несколько дополнительных антител человека или их антигенсвязывающих участков по изобретению (например, антитело человека, обладающее добавочной активностью, которое связывается с эпитопом на антигене α5β1, отличным от эпитопа, распознаваемого первым антителом человека).
Соответственно, пациентам, получающим композиции антител по изобретению, может быть дополнительно введен (до, одновременно или после введения антитела человека по изобретению) другой терапевтический агент, такой как цитотоксический или радиотоксический агент, который усиливает или дополняет терапевтический эффект антител человека.
В других случаях индивидуум может дополнительно получать агент, который модулирует, например, усиливает или ингибирует, экспрессию или активность Fcγ или Fcγ-рецепторов, посредством, например, лечения индивидуума с помощью цитокина. К цитокинам для введения в процессе лечения мультиспецифичной молекулой относятся гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), интерферон γ (IFNγ) и фактор некроза опухолей (TNF).
Композиции (например, антитела человека, мультиспецифические и биспецифические молекулы) по изобретению также могут быть использованы для нацеливания клеток, экспрессирующих FcγR или α5β1, например, для мечения таких клеток. Для такого применения связывающий агент может быть связан с молекулой, которая может быть детектирована. Таким образом, изобретение относится к способам локализации ex vivo или in vitro клеток, экспрессирующих Fc-рецепторы, такие как Fcγ или α5β1. Детектируемая метка может представлять собой, например, радиоизотоп, флуоресцентное соединение, фермент или кофактор фермента.
В конкретном случае изобретение относится к способам детекции наличия антигена α5β1 в образце или измерения количества антигена α5β1, включающим приведение в контакт образца и контрольного образца с моноклональным антителом человека или его антигенсвязывающим участком, который специфически связывается с α5β1, в условиях, которые позволяют образовать комплекс между антителом или его частью и α5β1. Образование комплекса затем детектируют, причем образование различных комплексов между сравниваемым образцом и контрольным образцом указывает на наличие антигена α5β1 в образце.
В других случаях изобретение относится к способам лечения α5β1-опосредованного расстройства у индивидуума, например, аномального клеточного роста, такого как рак, путем введения индивидууму антител человека или их антигенсвязывающих участков, описанных выше. Такие антитела и их производные используются для ингибирования α5β1-индуцированных активностей, ассоциированных с некоторыми расстройствами, например, ангиогенеза, пролиферации и дифференциации. При контакте антитела с α5β1 (например, путем введения антитела индивидууму) способность α5β1 индуцировать такие активности ингибируется и, таким образом, лечение ассоциированного расстройства оказывается эффективным. Композиция антитела может быть введена самостоятельно или вместе с другим терапевтическим агентом, таким как цитотоксический или радиотоксический агент, который действует наравне или синергично с композицией антитела для лечения или профилактики α5β1-опосредованного заболевания.
В еще одном случае иммуноконъюгаты по изобретению могут быть использованы для нацеливания соединений (например, терапевтических агентов, меток, цитотоксинов, радиотоксинов, иммуносупрессантов и т.д.) на клетки, которые имеют рецепторы клеточной поверхности α5β1, путем связывания таких соединений с антителом. Таким образом, изобретение также относится к способам локализации ex vivo или in vivo клеток, экспрессирующих α5β1 (например, с помощью детектируемой метки, такой как радиоизотоп, флуоресцентное соединение, фермент или кофактор фермента). Альтернативно, иммуноконъюгаты могут быть использованы для уничтожения клеток, которые имеют рецепторы клеточной поверхности α5β1, путем нацеливания цитотоксинов или радиотоксинов на α5β1.
Настоящее описание дополнительно иллюстрируется следующими примерами, которые не должны быть истолкованы, как ограничивающие. Содержание всех фигур и всех ссылок, патентов и опубликованных патентных заявок, процитированных на протяжении этого описания, прямо приводится в данном документе в качестве ссылок в полном объеме.
ПРИМЕРЫ
Пример 1
Получение гибридомы, продуцирующей антитело против α5β1
Наглядные антитела по изобретению получали, отбирали и анализировали следующим образом:
Иммунизация и создание гибридомы:
Для получения гибридомы использовались следующие иммуногены: очищенный рекомбинантный белок интегрин α5-Fc человека; интегрин α5-His (продукция фирмы R&D Systems, на заказ); клетки NIH3T3, трансфицированные для экспрессии α5 человека; клеточная линия Jurkat, клеточная линия U-937 (ATCC Cat № CRL-1593) и клеточная линия K-562 (ATCC Cat № CCL-243), которые в естественных условиях экспрессируют интегрин α5β1 человека.
Очищенный рекомбинантный белок интегрин α5-Fc человека представляет собой химерную конструкцию внеклеточного домена интегрина α5 человека (аминокислоты 42-995), слитого с Fc-доменом крысы, которую клонировали в вектор pSecTag2 и экспрессировали, используя систему 293-FreeStyle (продукция фирмы Invitrogen). Клетки NIH3T3, трансфицированные для экспрессии α5 человека, продуцировали согласно следующему. кДНК полноразмерного интегрина α5 человека (продукция фирмы Invitrogen, Cat. № FL1002, клон ID:3629647) клонировали из клона MGC полноразмерного интегрина α5 человека (Invitrogen) и субклонировали в ретровирусный экспрессирующий вектор (pBabe). Вирусные частицы получали и использовали для заражения клеток NIH3T3 (ATCC Cat. CRL-1658). Для отбора положительных стабильных клонов клеток добавляли пиромицин. Для отбора стабильных клонов клеток с высоким уровнем экспрессии осуществляли анализы экспрессии белка человека α5 вестерн-блот и FACS.
Полностью человеческие моноклональные антитела против интегрина α5β1 человека получали, используя штамм трансгенных по Ig человека мышей Нсо7/Нсо12, а также штамм трансхромосомных/трансгенных по генам человека мышей КМ (продукция фирмы Medarex, Inc). Все эти штаммы экспрессируют полностью человеческие антитела, которые неотличимы от антител, выделенных от людей. В этих штаммах мышей эндогенный ген легкой каппа цепи мыши был разрушен гомозиготным образом, как описано в статье Chen et al. (1993) EMBO J. 12:811-820, и эндогенный ген тяжелой цепи мыши был разрушен гомозиготным образом, как описано в примере 1 РСТ публикации WO 01/09187. Каждый из этих штаммов мышей несет трансген легкой каппа цепи человека КСо5, описанный в статье Fishwild et al. (1996) Nature Biotechnology 14:845-851. Штамм НСо7 несет трансген тяжелой цепи человека НСо7, описанный в патентах США 5545806, 5625825 и 5545807. Штамм НСо12 несет трансген тяжелой цепи человека НСо12, описанный в примере 2 РСТ публикации WO 01/09187. Штамм НСо7/НСо12 несет оба трансгена тяжелой цепи НСо7 и НСо12. Штамм КМ несет минихромосому человека, описанную в статье Ishida et al., (2002), Cloning and Stem Cells, 4:91-102.
Для получения полностью человеческих моноклональных антител против α5β1 мышей HuMab штаммов НСо7/НСо12 и КМ иммунизировали рекомбинантным белком интегрин α5-Fc человека или интегрин α5-His, клетками NIH3T3, трансфицированными для экспрессии α5 человека и клеточной линией Jurkat, клеточной линией U-937 и клеточной линией K-562, которые в естественных условиях экспрессируют α5β1 человека. Общие схемы иммунизации мышей HuMab описываются в статьях Lonberg N. et al.(1994) Nature 368(6474): 856-859; Fishwild, D. еt al. (1996) Nature Biotechnology 14: 845-851 и РСТ публикации WO 98/24884. При первой инфузии антигена брались мыши в возрасте 6-16 недель. Для внутрибрюшинной (IP) и подкожной (Sc) иммунизации мышей HuMab использовали очищенный рекомбинантный препарат антигена интегрин α5-Fc человека или интегрин α5-his (в количестве 15-20 мкг), препарат трансфицированных клеток NIH3T3 или клеток Jurkat, клеток U-937 и клеток К-562 (1×107 клеток).
Трансгенных мышей иммунизировали антигеном в адъюванте Ribi внутрибрюшинно и подкожно с интервалами 1-4 недели (всего до 16 иммунизаций). Иммунный ответ наблюдали в крови, взятой посредством ретроорбительных кровопусканий. Сыворотку скринировали посредством анализа FACS (описанного ниже), и мышей с достаточными титрами иммуноглобулина человека против α5β1 использовали для слияний. Мышей стимулировали внутривенным введением антигена за 3 и 2 дня до забоя и извлечения селезенки и/или лимфатических узлов. Как правило, для каждого антигена осуществляли 10-20 слияний. Всего иммунизировали 60 мышей НСо7/НСо12 и КМ. Каждым антигеном иммунизировали несколько дюжин мышей.
Отбор мышей HuMab, продуцирующих антитела против α5β1:
Для отбора мышей HuMab, продуцирующих антитела, которые связываются с α5β, сыворотку иммунизированных мышей скринировали посредством проточной цитометрии (FACS) в отношении связывания с клеточной линией, экспрессирующей полноразмерный интегрин α5β1 человека, и не связывания с контрольной клеточной линией, не экспрессирующей α5β1. Вкратце, клетки NIH3T3, экспрессирующие α5, инкубировали с сывороткой иммунизированных мышей, разбавленной 1:20. Клетки отмывали, и специфическое связывание антитела детектировали с помощью Ab против IgG человека, меченого FITC. Анализы проточной цитометрии осуществляли на приборе проточном цитометре FACS (продукция фирмы Becton Dickinson, San Jose, CA). Мышей, у которых вырабатывались самые высокие титры антител против α5β1, использовали для слияний. Слияния осуществляли, как описано ниже, и гибридомные супернатанты тестировали с помощью FACS в отношении активности против α5β1.
Получение гибридом, продуцирующих моноклональные антитела человека против α5β1:
Спленоциты и/или лимфоциты из лимфатических узлов мыши, выделенные от мышей HuMab, сливали, используя электрослияние (Е-слияние, технология Cyto PulseTM, Cyto PulseTM Sciences, Inc., Glen Burnie, MD), с миеломной клеточной линией мышей Sp2/0 (ATCC, CRL-1581, Manassas, VA), используя протоколы, рекомендованные производителем. Вкратце, суспензии одиночных клеток лимфоцитов селезенки и/или лимфатических узлов иммунизированных мышей сливали с эквивалентным количеством несекретирующих клеток миеломы мышей Sp2/0, используя Е-слияние. Клетки помещали в плоскодонные планшеты для микротитрования в количестве примерно 2×104 спленоцитов/лунка и инкубировали в течение от 10 до 14 дней в селективной среде, содержащей 10% эмбриональную бычью сыворотку, 10% кондиционированную клетками P388D1 (ATCC, CRL-TIB-63) среду, от 3 до 5% (IGEN) в среде DMEM (продукция фирмы Mediatech, Herndon, VA, Cat. № CRL 10013) при высоком содержании глюкозы, L-глютамина и пирувата натрия), 5 мМ HEPES, 0,055 мМ 2-меркаптоэтанол, 50 мг/мл гентамицин и 1-кратный НАТ (продукция фирмы Sigma, Cat. № CRL -P-7185). Через 1-2 недели клетки культивировали в среде, в которой НАТ заменяли на НТ. Примерно через 10-14 дней после помещения клеток супернатанты из отдельных лунок скринировали вначале в отношении содержания в них антител с гамма, каппа-цепями человека. Супернатанты, которые отмечались как позитивные в отношении антител с гамма- каппа-цепями человека, затем последовательно скринировали посредством FACS (описанного выше) в отношении антител человека против α5β1.
Моноклональные антитела IgG:
Гибридомы, секретирующие антитела, переносили в 24-луночные планшеты, вновь скринировали и при подтверждении позитивности в отношении IgG-моноклональных антител человека против α5β1 субклонировали, по меньшей мере, дважды путем предельного разведения. Устойчивые субклоны затем культивировали in vitro для получения небольших количеств антитела в тканевой культуральной среде с целью дополнительной характеристики. Процедуры, описанные выше, использовали для продукции нескольких моноклональных антител против α5β1, включая антитела, обозначенные как «22В5», «24С7», «1D9» и «2D2», которые описаны в данном документе.
Пример 2
Структурная характеристика моноклонального антитела человека 22В5
Последовательности кДНК, кодирующие вариабельные области тяжелой и легкой цепей моноклонального антитела 22В5, получали из гибридомы 22В5, используя стандартные техники ПЦР, и секвенировали, используя стандартные техники секвенирования ДНК.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи антитела 22В5 показаны на фиг.1А и 1В и в SEQ ID NO:11 и 7 соответственно. Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи антитела 22В5 показаны на фиг.1С и 1D и в SEQ ID NO:12 и 8 соответственно.
Исходная молекула 22В5 относилась к подклассу IgG4. Подкласс IgG переключали на IgG1 для обеспечения связывания с FcγR и эффекторных функций, таких как ADCC. Для дополнительного повышения связывания с рецепторами FcγR и активности эффекторных функций в константную область IgG1 вводили три мутации (S247D, A338L и I340E) посредством сайт-специфичного мутагенеза. Антитело 22В5 с подклассом IgG1 и содержанием этих трех мутаций обозначают как «22B5/DLE» или «22B5 IgG1 DLE». Две мутации вариабельного домена тяжелой цепи возвращали к зародышевой линии для минимизации иммуногенности (I30S и N33S). В вариабельной последовательности легкой цепи мутаций обнаружено не было. Тяжелая цепь 22B5/DLE представлена в виде SEQ ID NO:9, тогда как легкая цепь 22B5/DLE представлена в виде SEQ ID NO:10.
Сравнение последовательности тяжелой цепи иммуноглобулина 22В5 с известными последовательностями тяжелых цепей иммуноглобулинов зародышевой линии человека показало, что в тяжелой цепи антитела 22В5 используется сегмент VH из гена VH 4-39 зародышевой линии человека, сегмент D из гена 3-10 зародышевой линии человека и сегмент JH из гена JH 6b зародышевой линии человека. Выравнивание последовательности VH антитела 22В5 с последовательностью VH 4-39 зародышевой линии представлено на фиг.2. Дополнительный анализ последовательности VH антитела 22В5, используя систему Kabat определения области CDR, привело к изображению областей CDR1, CDR2 и CDR3 тяжелой цепи, представленному на фиг.1В и в SEQ ID NO:1, 2 и 3 соответственно.
Сравнение последовательности легкой цепи иммуноглобулина 22В5 с известными последовательностями легких цепей иммуноглобулинов зародышевой линии человека показало, что в легкой цепи антитела 22В5 используется сегмент VL из гена VK L6 зародышевой линии человека и сегмент JK из гена JK 4 зародышевой линии человека. Выравнивание последовательности VL антитела 22В5 с последовательностью VK L6 зародышевой линии представлено на фиг.2. Дополнительный анализ последовательности VL антитела 22В5, используя систему Kabat определения области CDR, привело к изображению областей CDR1, CDR2 и CDR3 легкой цепи, представленному на фиг.1D и в последовательностях SEQ ID NO:4, 5 и 6 соответственно.
Пример 3
Характеристика специфичности связывания и кинетический анализ связывания моноклонального антитела человека 22В5
В этом примере сродство связывания и кинетический анализ связывания антитела человека против α5β1 22В5 с мутациями IgG1 DLE (22B5/DLE) оценивали посредством анализа Biacore. Также специфичность связывания оценивали с помощью проточной цитометрии (FACS).
Сродство связывания и кинетический анализ связывания
Кинетический анализ связывания и номинальную авидность антитела 22В5/DLE к внеклеточному домену интегрина α5β1 определяли, используя анализ Biacore (Biacore АВ, Uppsala, Sweden). Для осуществления кинетических анализов BIAcore антитело 22В5/DLE иммобилизовали на биосенсорном чипе, и большое число концентраций внеклеточного домена рекомбинантного α5β1 человека пропускали через поверхность при 25,0°С (см. фиг.3). Данные по связыванию подводили в целом под модель связывания одного-к-одному образца с дрейфующим исходным уровнем. 22В5/DLE обратимо связывается с α5β1. Константа связывания (KD) для внеклеточного домена рекомбинантного α5β1 человека находилась в диапазоне от 2,7 до 4,1 нМ. KD оказалась выше при измерении в отсутствие двухвалентных катионов, согласуясь с катион-зависимой активацией интегрина. Кинетические параметры связывания находились в диапазоне от 3,4×10-5 до 4,5×10-5 М-1 с-1 для скорости прямой реакции и от 1,2×10-3 до 1,4×10-3 с-1 для скорости обратной реакции (см. фиг.3 и таблицу 1).
Для измерения номинальной авидности внеклеточный домен рекомбинантного α5β1 человека иммобилизовали на биосенсорном чипе, и большое число концентраций 22В5/DLE пропускали через поверхность при 25,0°С. Величина номинальной авидности составляла 0,05 пМ в 20 мМ HEPES, pH 7,4; 150 мМ NaCl; 0,005% P20 с 4,0 мМ MgCl2.
Итоговые данные по связыванию и кинетическому анализу, полученные на основании исследований BIAcore
Под CaCl2 понимают буфер, содержащий 10 мМ HEPES, pH 7,4; 150 мМ NaCl; 0,005% P20 с 4,0 мМ CaCl2; под MgCl2 понимают буфер, содержащий 10 мМ HEPES, pH 7,4; 150 мМ NaCl; 0,005% P20 с 4,0 мМ MgCl2.
Определение с помощью FACS сродства к клеткам, экспрессирующим α5β1
Антитело 22В5/DLE тестировали на его сродство связывания с интегрином клеточной поверхности α5, используя анализ FACS, с применением клеток, экспрессирующих эндогенный интегрин α5β1 человека (HUVEC). Вкратце, клетки выделяли, используя трипсин-EDTA, и отмывали с помощью охлажденного PBS. После внесения аликвот в 96-луночные планшеты клетки блокировали сывороткой и инкубировали с различными концентрациями специфического mAb в течение 1 часа при 4°С. Затем клетки отмывали и инкубировали со вторичным антителом против κ-цепи человека, конъюгированным с флуорофором R-PE, и анализировали, используя проточный цитометр FACSCalibur. Для каждого образца собирали 10000 событий, не применяя какого-либо гейтирования. Для определения KD рассчитывали среднее геометрическое гистограммы каждого образца и выражали как функцию от концентрации mAb. KD рассчитывали после подведения под равновесную модель с двумя состояниями. Антитело 22В5/DLE связывается с эндогенным α5β1 с KD 2,15 нМ (n=4) в клетках HUVEC (см. фиг.4).
Пример 4
Сродство к Fcγ-рецепторам, определенное с помощью анализа Biacore
Способность антитела 22В5/DLE, которое содержит три мутации IgG1 DLE (описанные ранее в примере 2), усиливать связывание с рецепторами FcγR оценивали в эксперименте связывания Biacore. Сродство связывания с FcγR антитела 22В5/DLE сравнивали со сродством связывания IgG1 дикого типа (wt). Было протестировано три класса рецепторов FcγR: 1) высокоаффинный рецептор FcγRI; 2) два полиморфных варианта низкоаффинного рецептора FcγRIIa/131Н и FcγRIIa/131R; и 3) два полиморфных варианта рецептора со средним сродством FcγRIIIa/158F и FcγRIIIa/158V. Результаты (представленные на фиг.5 и суммированные в таблице 2) показывают примерно 6-кратное увеличение для высокоаффинного рецептора FcγRI. Наблюдалось 122- и 70-кратное усиление связывания для рецепторов со средней аффинностью FcγRIIIa/158F и 158V соответственно. Сравнение сродства связывания антитела 22В5/DLE и IgG1 дикого типа также оценивали против Fcγ-рецепторов мыши. Максимальное усиление связывания наблюдалось для mFcγRIV, затем для mFcγRI и mFcγRIII (см. фиг.5).
Связывание с рецепторами FcγR в анализе Biacore (KD, кратность усиления относительно wt IgG1
Пример 5
Клеточная функциональная активность антитела 22В5/DLE
Блокада клеточной адгезии
Была протестирована способность антитела 22В5/DLE нарушать адгезию клеток, опосредованную интегрином α5β1. Анализы клеточной адгезии осуществляли путем предварительной инкубации клеток HUVEC с антителом (22В5/DLE, 22В5 IgG1, вариантами подкласса IgG2 и IgG4 или mAb отрицательного контроля (ВНА2 IgG1)) и затем помещения в планшеты, покрытые фибронектином (FN) или коллагеном. Клетки HUVEC (15000) смешивали с антителом в буфере для адгезии (Hepes-забуференный солевой раствор, содержащий глюкозу и бычий сывороточный альбумин) в течение 20 минут при комнатной температуре. Клетки добавляли в лунки 96-луночного планшета, покрытые фибронектином или коллагеном, и оставляли для адгезии к планшету в течение одного часа при 37°С/5% СО2. Неадгезировавшие клетки удаляли отмывкой каждой лунки три раза. Измеряли количество адгезировавших клеток, оставшихся в каждой лунке. Адгезировавшие клетки лизировали путем добавления буфера, содержащего краситель CyQUANT GR, и флуоресценцию измеряли при Ex/Em: 485/535 нм с помощью плашечного ридера. Как можно видеть на фиг.6, mAb, функционально блокирующие интегрин α5β1, избирательно ингибируют адгезию клеток HUVEC к фибронектину, но не к коллагену (отрицательный контроль). Как показано на фиг.6, величины IC50 для 22В5/DLE и вариантов его подкласса (IgG1 дикого типа, IgG1 DLE, IgG2 и IgG4) оказались схожими в этом одночасовом анализе. Эти данные указывают на то, что связывание антитела с α5 и/или α5β1 ингибирует связывание интегрина с фибронектином. Эти данные дополнительно указывают на то, что антитело связывается с α5β1, если он экспрессируется на поверхности клеток. То есть эпитоп, распознаваемый антителом, доступен, если цепи α5 и β1 ассоциированы, и эпитоп доступен для связывания, если интегрин экспрессируется на клеточной поверхности.
Пример 6
ADCC-эффекторная функция in vitro
Активности эффекторных функций оценивали путем анализов ADCC в присутствии мононуклеарных клеток периферической крови человека (РВМС) против α5β1-позитивных мишенных клеток, используя стандартные способы. Уровни экспрессии интегрина α5 для ряда клеточных линий измеряли с помощью анализа вестерн-блот в соответствии со следующим. Клетки лизировали в лизирующем буфере RIPA (продукция фирмы Upstate), содержащем ингибиторы протеазы (продукция фирмы Roche) и фосфатазы (продукция фирмы Calbiochem). Осуществляли электрофорез лизатов (по 3 мкг общего белка в каждом) в геле SDS-PAGE и иммуноблоттинг с антителами против интегрина α5. Иммунореактивные полосы идентифицировали и анализировали посредством инфракрасной системы изображения Odyssey фирмы LI-COR Bioscience. Пример ряда экспрессии показан на фиг.7.
Для измерения ADCC эффекторные клетки (РВМС) выделяли, используя центрифугирование в градиенте фиколла, из образцов банка крови Сан-Диего согласно стандартным способам. Десять тысяч мишенных клеток (HUVEC или опухолевых линий) предварительно инкубировали с антителом в ростовой среде при комнатной температуре в течение 20 минут, после чего добавляли один миллион клеток РВМС человека (Е:Т=100:1), и смесь инкубировали при 37°С/5% СО2 в течение 4 часов. ADCC-опосредованный клеточный лизис измеряли, используя детекцию высвобождения лактатдегидрогеназы (LDH) (Roche) или набор для биоанализа ToxiLight (Cambrex) согласно стандартным протоколам.
Антитело 22В5/DLE связывается с опухолью или интегрином α5β1, экспрессируемым эндотелием, и стимулирует ADCC (до 80% лизиса мишенных клеток) с величиной ЕС50 0,04 нМ по сравнению с 0,23 нМ для wt 22B5 IgG1. Эти данные указывают на то, что мутации DLE усиливали ADCC-активность по сравнению с IgG1 такого типа. В четырехчасовом анализе ни для 22В5 IgG2, ни для 22В5 IgG4 ADCC-активности обнаружено не было. В качестве mAb отрицательного контроля использовали KLH IgG1. Примеры ADCC в клетках HUVEC и типичной опухолевой линии (U87MG) показаны на фиг.8 (отражает оба способа детекции).
Как показывают данные, представленные на фиг.9, клеточная цитотоксичность, индуцируемая 22В5/DLE и wt 22D5 IgG1, коррелирует с уровнем экспрессии α5β1 клеточной линией. У клеток с самой высокой экспрессией α5β1 (панель А, U87MG) наблюдается самый высокий уровень клеточной цитотоксичности, тогда как у клеточных линий со средней экспрессией (фиг.9, Hs578T и 3T3, 5) наблюдается промежуточная цитотоксичность. Отмечено, что антитело 22В5/DLE усилило ADCC-эффекты в клетках M24met, которые экспрессировали низкий уровень α5β1 (примеры в нижней панели фиг.9). Эти данные указывают на то, что ADCC опосредуется, по меньшей мере, частично, связыванием антитела с α5β1, экспрессируемым клеткой.
Пример 7
Антиметастатическая эффективность in vivo
Помимо его активностей пролиферации, миграции и выживаемости в эндотелиальных и опухолевых клетках, интегрин α5β1 также участвует в метастазировании опухоли за счет стимуляции экстравазации опухолевых клеток и миграции в отдаленные органы. Следующее исследование предназначено продемонстрировать антиметастатическую эффективность 22В5/DLE в предклинической модели.
Ингибирование экспериментального метастазирования А549-Luc в легкое
А549-Luc представляет собой клеточную линию немелкоклеточного рака легкого человека (NSCLC), трансфицированную геном люциферазы. При внутривенной имплантации клетки А549-Luc предпочтительно локализуются в легком, что может быть измерено посредством биолюминесцентного изображения (BLI). В этом исследовании клетки А549-Luc вводили (в количестве 3×106 на животного) через хвостовую вену в мышей SCID BALB/c, которые получили предварительную дозу соответствующего антитела за два дня до инъекции. После имплантации животные получали дозы один раз в неделю до 8 недели. Биолюминесценцию каждого животного измеряли один раз в неделю до 20 недели.
У антитела 22В5/DLE наблюдалась наибольшая эффективность против метастазирования опухоли с TGI (ингибированием роста опухоли) 97% на 8 неделе (конечная неделя введения дозы), что является статистически значимым (р<0,01) по сравнению с контрольной группой. Введение дозы антитела 22В5 IgG1, которое, как предполагается, не опосредует ADCC в этой модели, также приводило к существенному TGI, составившему 89%, на 8 неделе (фиг.10А). Величины р сравнения между 22В5/DLE и 22В5 IgG2 на 5, 6, 7 и 8 неделях составили 0,07; 0,03; 0,08 и 0,17, соответственно. Все способы лечения прекращали после 8 недели, тогда как ежедневный мониторинг нежелательных физиологических признаков и еженедельные измерения посредством BLI животных продолжали.
Опухоли в группе, обработанной 22В5/DLE, остались супрессированными через 13 недель, тогда как в группе, обработанной 22В5 IgG2, начали быстро расти примерно через 2 недели после отмены лекарственного средства (фиг.10В). Конечную точку исследования полагали достигнутой при достижении BLI опухоли 1×108 фонотов/секунда, в это время мышей забивали. На графике Каплана-Мейера (фиг.10С) показано, что антитело 22В5/DLE существенно удлиняло среднее время выживаемости животных до 20 недель по сравнению с 14 неделями для животных, обработанных 22В5 IgG2, и 10 неделями для контрольной группы.
Эти данные указывают на то, что в этой модели противоопухолевый эффект антитела 22В5 не опосредуется исключительно ADCC-активностью. То есть 22В5-IgG2 ингибировало рост/инвазию опухоли, хотя неизвестно, опосредует ли IgG2 человека эффекторные функции, включая клеточную цитотоксичность, а также опосредованные антителом противоопухолевые эффекты. Кроме того, данные показали, что рост опухоли, оставшийся супрессированным после введения 22В5-IgG1-DLE, прекратился. Эти данные указывают на то, что блокирование связывания α5β1 с FN опосредует противоопухолевый эффект (22В5-IgG2), который дополнительно усиливается ADCC-активностью (22В5-IgG1/DLE).
Пример 8
Структурная характеристика моноклональных антител человека 24С7, 1D9 и 2D2
Последовательности кДНК, кодирующие вариабельные области тяжелой и легкой цепей моноклональных антител 24С7, 1D9 и 2D2, получали из гибридом 24С7, 1D9 и 2D2 (соответственно), используя стандартные техники ПЦР, и секвенировали, используя стандартные техники секвенирования ДНК.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи антитела 24С7 показаны на фиг.1Е и 1F и в последовательностях SEQ ID NO:21 и 19, соответственно. Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи антитела 24C7 показаны на фиг.1G и 1H и в последовательностях SEQ ID NO:22 и 20, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи антитела 1D9 показаны на фиг.1I и 1J и в последовательностях SEQ ID NO:31 и 29, соответственно. Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи антитела 1D9 показаны на фиг.1K и 1L и в последовательностях SEQ ID NO:32 и 30, соответственно.
Нуклеотидная и аминокислотная последовательности вариабельной области тяжелой цепи антитела 2D2 показаны на фиг.1M и 1N и в последовательностях SEQ ID NO:41 и 39, соответственно. Нуклеотидная и аминокислотная последовательности вариабельной области легкой цепи антитела 1D9 показаны на фиг.1O и 1P и в последовательностях SEQ ID NO:42 и 40, соответственно.
Исходные молекулы 24С7, 1D9 и 2D2 относились к подклассу IgG1. В отношении подкласса IgG1, для дополнительного повышения связывания с рецепторами FcγR и активности эффекторных функций в константную область IgG1 вводили три мутации (S247D, A338L и I340E) посредством сайт-специфичного мутагенеза. Таким образом, антитело 24С7, относящееся к подклассу IgG1 и содержащее эти три мутации, обозначают как «24С7/DLE» или «24С7 IgG1 DLE». Подобным образом, аналогичные антитела с этими тремя мутациями обозначают в данном документе как «1D9/DLE» или «1D9 IgG1 DLE» и «2D2/DLE» или «2D2 IgG1 DLE». Аналогично, антитела с двумя мутациями S247D и I340E обозначают в данном документе как мутанты «DE», например, «24C7/DE» или «24C7 IgG1 DE». Аналогичные наименования любых антител, описанных в данном документе, и с любой одной, двумя или тремя указанными выше мутациями обозначают в данном документе подобным же образом. Например, под «1D9/DE» понимают антитело 1D9, описанное в данном документе, относящееся к подклассу IgG1, со следующими мутациями в Fc-области - S247D и I340E. Аналогично, «2D2/L» относилось бы к антителу 2D2, описанному в данном документе, с изотипом IgG1 и содержащему мутацию A338L и так далее.
Сравнение последовательности тяжелой цепи иммуноглобулина 24С7 с известными последовательностями тяжелых цепей иммуноглобулинов зародышевой линии человека показало, что в тяжелой цепи 24С7 используется сегмент VH из гена VH 3-30.3 зародышевой линии человека, сегмент D из гена 7-27 зародышевой линии человека и сегмент JH из гена JH 6b зародышевой линии человека. Анализ последовательности VH антитела 24С7, используя систему Kabat определения CDR-области, привело к изображению областей CDR1, CDR2 и CDR3 тяжелой цепи, представленному на фиг.1F и в последовательностях SEQ ID NO:13, 14 и 15, соответственно.
Сравнение последовательности легкой цепи иммуноглобулина 24C7 с известными последовательностями легких цепей иммуноглобулинов зародышевой линии человека показало, что в легкой цепи 24C7 используется сегмент VL из гена VK L6 зародышевой линии человека и сегмент JK из гена JK 2 зародышевой линии человека. Анализ последовательности VL антитела 24C7, используя систему Kabat определения CDR-области, привело к изображению областей CDR1, CDR2 и CDR3 легкой цепи, представленному на фиг.1H и в последовательностях SEQ ID NO:16, 17 и 18, соответственно.
Сравнение последовательности тяжелой цепи иммуноглобулина 1D9 с известными последовательностями тяжелых цепей иммуноглобулинов зародышевой линии человека показало, что в тяжелой цепи 1D9 используется сегмент VH из гена VH 3-30.3 зародышевой линии человека и сегмент JH из гена JH 6b зародышевой линии человека. Сегмент D не мог быть сопоставлен с геном зародышевой линии из-за распространенных мутаций. Анализ последовательности VH антитела 1D9, используя систему Kabat определения CDR-области, привело к изображению областей CDR1, CDR2 и CDR3 тяжелой цепи, представленному на фиг.1J и в последовательностях SEQ ID NO:23, 24 и 25, соответственно.
Сравнение последовательности легкой цепи иммуноглобулина 1D9 с известными последовательностями легких цепей иммуноглобулинов зародышевой линии человека показало, что в легкой цепи 1D9 используется сегмент VL из гена VK L6 зародышевой линии человека и сегмент JK из гена JK 1 зародышевой линии человека. Анализ последовательности VL антитела 1D9, используя систему Kabat определения CDR-области, привело к изображению областей CDR1, CDR2 и CDR3 легкой цепи, представленному на фиг.1L и в последовательностях SEQ ID NO:26, 27 и 28, соответственно.
Сравнение последовательности тяжелой цепи иммуноглобулина 2D2 с известными последовательностями тяжелых цепей иммуноглобулинов зародышевой линии человека показало, что в тяжелой цепи 2D2 используется сегмент VH из гена VH 3-30.3 зародышевой линии человека, сегмент D из гена 7-27 зародышевой линии человека и сегмент JH из гена JH 6b зародышевой линии человека. Анализ последовательности VH антитела 2D2, используя систему Kabat определения CDR-области, привело к изображению областей CDR1, CDR2 и CDR3 тяжелой цепи, представленному на фиг.1N и в последовательностях SEQ ID NO:33, 34 и 35, соответственно.
Сравнение последовательности легкой цепи иммуноглобулина 2D2 с известными последовательностями легких цепей иммуноглобулинов зародышевой линии человека показало, что в легкой цепи 2D2 используется сегмент VL из гена VK L6 зародышевой линии человека и сегмент JK из гена JK 3 зародышевой линии человека. Анализ последовательности VL антитела 2D2, используя систему Kabat определения CDR-области, привело к изображению областей CDR1, CDR2 и CDR3 легкой цепи, представленному на фиг.1P и в последовательностях SEQ ID NO:36, 37 и 38, соответственно. Ниже в таблице 3 представлено использование генов в моноклональных антителах 22В5, 2D2, 24С7 и 1D9.
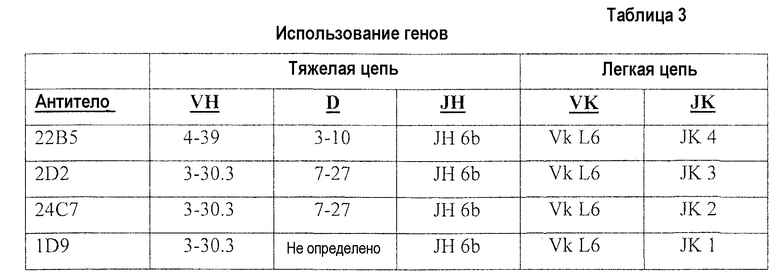
Пример 9
Характеристика специфичности связывания и кинетический анализ связывания моноклональных антител человека 22В5, 24С7, 1D9 и 2D2
В этом примере специфичность связывания антител человека против α5β1 22В5, 24С7, 1D9 и 2D2 подкласса IgG1 при наличии и в отсутствие мутаций IgG1 DLE (т.е. «22B5 G1 DLE», «22B5 G1», «1D9 G1 DLE», «1D9 G1» и т.д.) оценивали посредством проточной цитометрии, используя сортировку флуоресцентно-активированных клеток (FACS).
Определение сродства к клеткам, экспрессирующим α5β1, с помощью FACS
Антитела 22В5/DLE, 24С7/DLE, 1D9/DLE и 2D2/DLE тестировали в отношении их сродства связывания к интегрину клеточной поверхности α5, используя анализ FACS с применением клеток, экспрессирующих эндогенный интегрин α5β1 человека (HUVEC). Вкратце, клетки выделяли, используя трипсин-EDTA, и отмывали с помощью охлажденного PBS. После внесения аликвот в 96-луночные планшеты клетки блокировали сывороткой и инкубировали с различными концентрациями специфического mAb в течение 1 часа при 4°С. Затем клетки отмывали и инкубировали со вторичным антителом против κ-цепи человека, конъюгированным с флуорофором R-PE, и анализировали, используя проточный цитометр FACSCalibur. Для каждого образца собирали 10000 событий без применения какого-либо гейтирования. Для определения KD рассчитывали среднее геометрическое гистограммы каждого образца и выражали как функцию от концентрации mAb. KD рассчитывали после подведения под равновесную модель с двумя состояниями. Результаты представлены на фиг.11А и 11В.
Пример 10
In vitro ADCC-эффекторная функция моноклональных антител человека 22В5, 24С7, 1D9 и 2D2
Активности эффекторных функций оценивали посредством анализов антителозависимой клеточно-опосредованной цитотоксичности (ADCC), описанных ранее в примере 6. Величины ЕС50 в отношении стимуляции ADCC (до 80% лизиса мишенных клеток) в типичной опухолевой линии (U87MG) представлены на фиг.12 для антител 1D9, 1D9/DLE, 2D2/DLE, 22B5/DLE и 24C7/DLE.
Пример 11
In vivo ADCC-зависимая противоопухолевая эффективность в сингенной модели метастатической меланомы
Для демонстрации ADCC-специфического ответа моноклональное антитело человека 1D9/DLE (которое связывается с α5β1 человека и мыши, но функционально не нейтрализует α5β1) испытывали на модели ингибирования опухолевого роста (TGI) у сингенных мышей с интактными иммунными эффекторными клетками. Конкретно, ADCC-опосредованную противоопухолевую активность оценивали на модели сингенных мышей, у которой клетки меланомы мыши B16F10 экспрессировали α5β1. Линия меланомы мыши B16F10, экспрессирующая интегрин α5, является высоко метастатичной и при внутривенном введении первично колонизирует легкое. Опухолевые клетки вводили внутривенно в иммунокомпетентных мышей C57BL/6. Клетки B16F10 (2×105 на животное) вводили через хвостовую вену в сингенных мышей-хозяев C57BL/6, которые предварительно за 1 день до инъекции получали антитела (10 мг/кг, n=10 на группу). Животные получали подкожно антитело один раз в неделю в течение всего 3 недель. На 21 день животных забивали и резецировали легкие.
Как показано результатами на фиг.13, антитело 1D9/DLE продуцировало существенную противоопухолевую эффективность in vivo, указывая на то, что для продукции TGI достаточно одного ADCC-ответа. Для отображения специфического эффекта ADCC в модели животных обрабатывали с помощью 1D9 IgG2 (нет данных об опосредовании ADCC) и 1D9 IgG1 DLE (изотип IgG1 с усилением Fc за счет мутаций DLE). Большую противоопухолевую эффективность наблюдали с 1D9 IgG1 DLE, чем с 1D9 IgG2. Поскольку пары изотипов имеют идентичные аминокислотные последовательности в антигенсвязывающем домене и in vitro активности, данные указывают на то, что за повышенную эффективность 1D9 IgG1 DLE была ответственна усиленная ADCC в результате мутаций DLE.
Информация о депонировании
Заявители депонировали вариабельные области тяжелой и легкой цепей антитела, обозначенного в данном документе как 22В5, в American Type Culture Collection (ATCC) Manassas, VA 20110-2209 U.S.A. VH-область антитела 22В5 депонировали 16 июля 2008 и присвоили депозитный номер АТСС РТА-9377. VL-область антитела 22В5 депонировали 16 июля 2008 и присвоили депозитный номер АТСС РТА-9377. Эти депозиты сделаны в соответствии с положениями Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры и правилами на основании этого договора (Будапештский договор). Эти депозиты должны поддерживаться без ограничения в депозитарии АТСС в течение периода 30 лет или 5 лет после последнего запроса или в течение срока действия патента независимо от его длительности, и должны быть заменены, если депозиты станут нежизнеспособными в течение этого периода. Доступность депонированных материалов не должна быть истолкована как разрешение на выполнение любых аспектов настоящего изобретения в нарушение прав, данных по уполномочию любого правительства в соответствии с его законами патентования. Предполагается, что выше изложенного описания должно быть достаточно, чтобы специалист в данной области смог выполнить все аспекты настоящего изобретения. Настоящее изобретение не должно быть ограничено в объеме депонированными материалами, поскольку депонированный вариант осуществления понимается как одна из иллюстраций некоторых аспектов изобретения, и любые конструкции, которые функционально эквивалентны, охватываются данным изобретением. Депонирование материала в данном документе не является признанием, что изложенного в данном документе описания недостаточно, чтобы осуществить любой аспект изобретения, включая его лучший способ, и оно не должно быть истолковано как ограничивающее объем формулы изобретения конкретными иллюстрациями, которые оно представляет. На самом деле, специалистам в данной области из вышеизложенного описания должно быть понятно большое число модификаций изобретения помимо представленных и описанных в данном документе, и они охватываются прилагаемой формулой изобретения.





















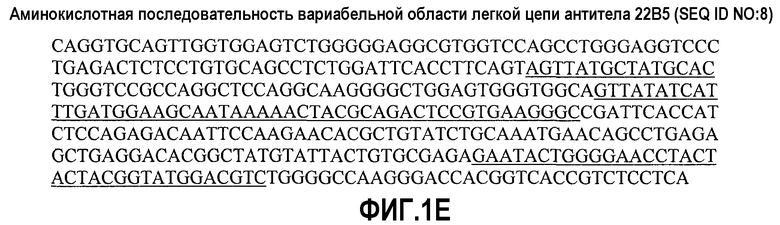



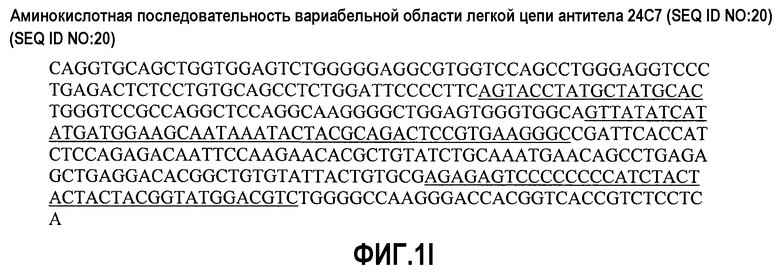





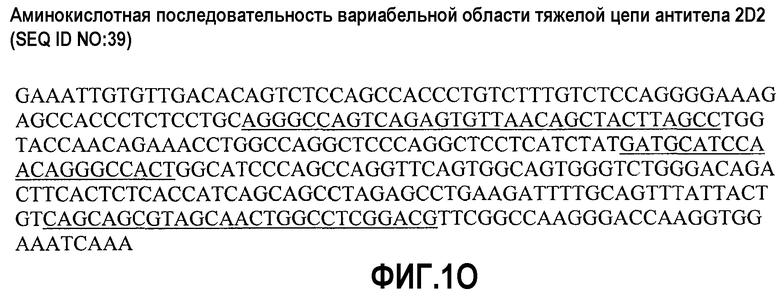




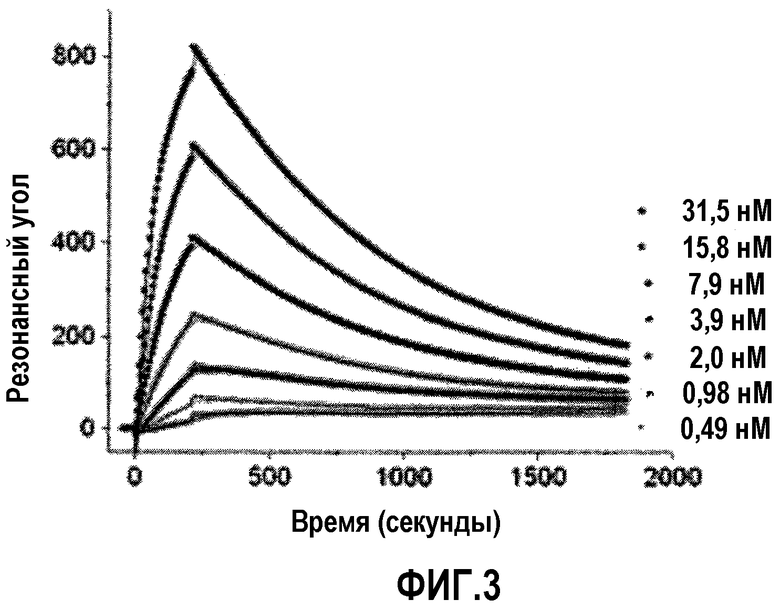
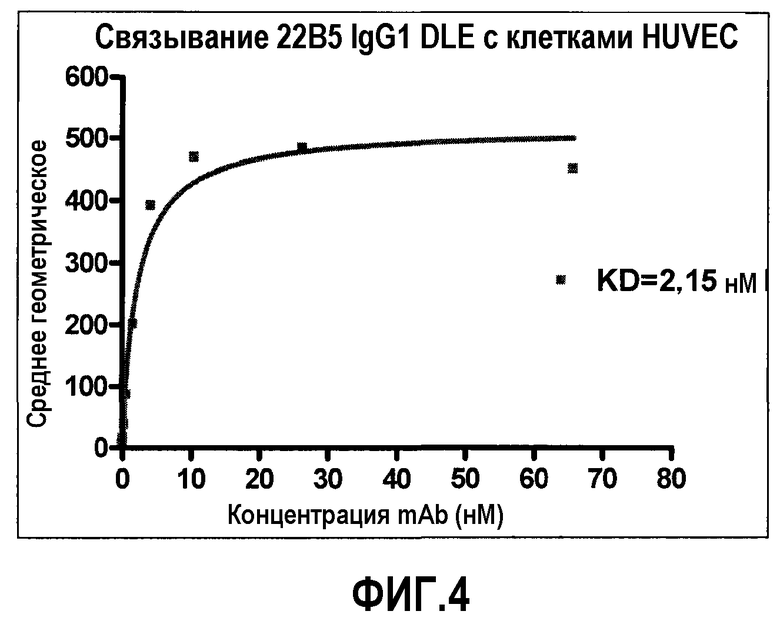
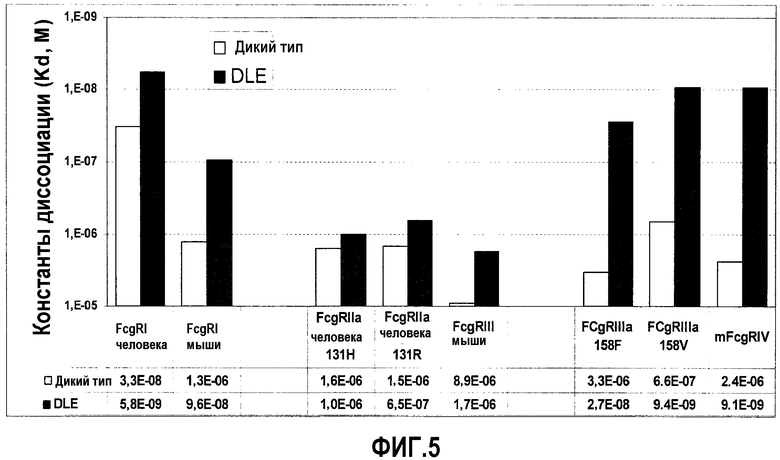


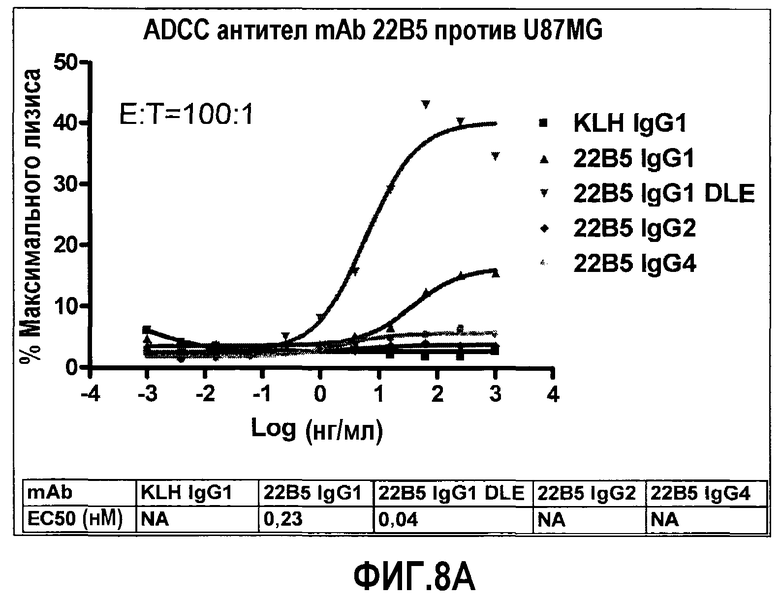
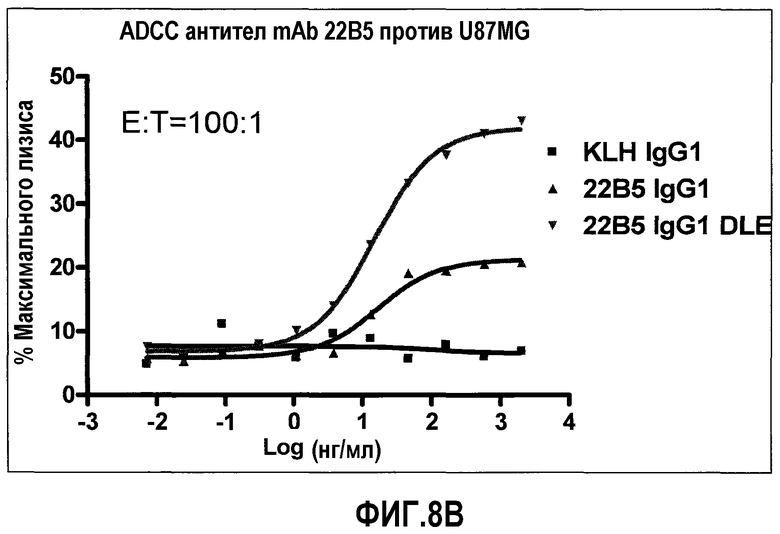
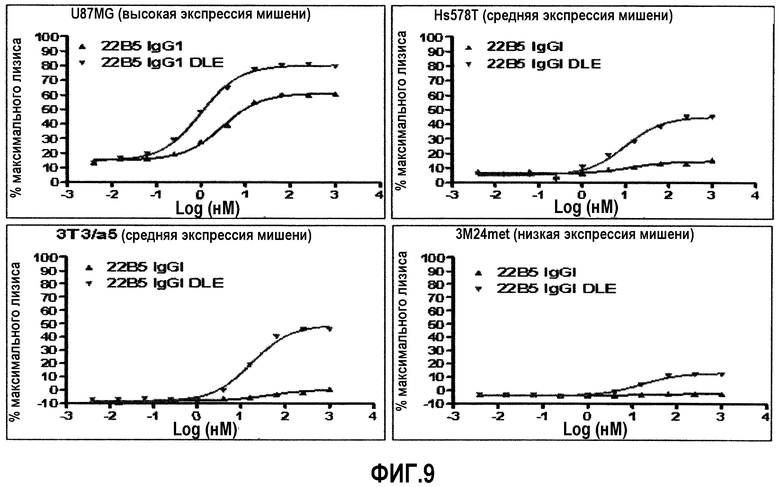
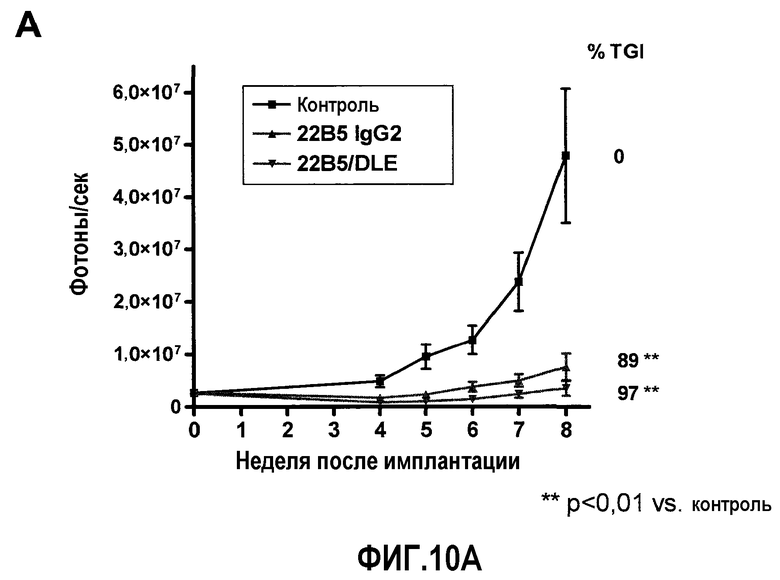
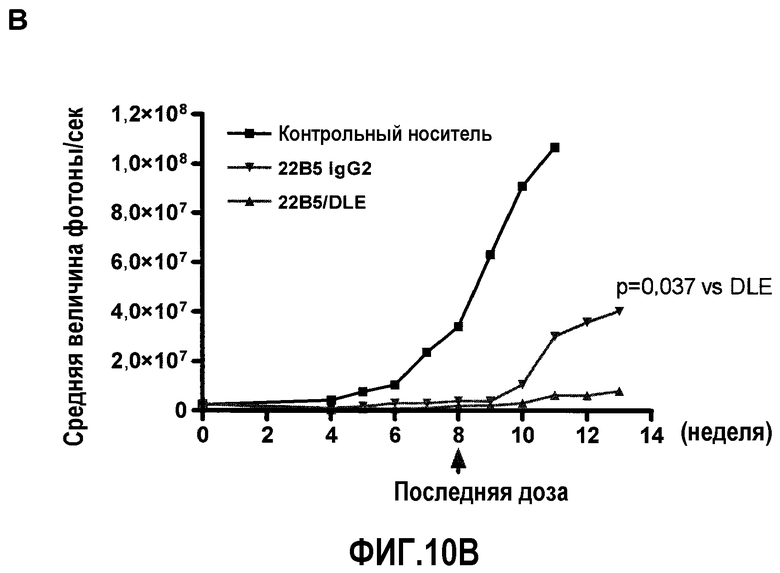
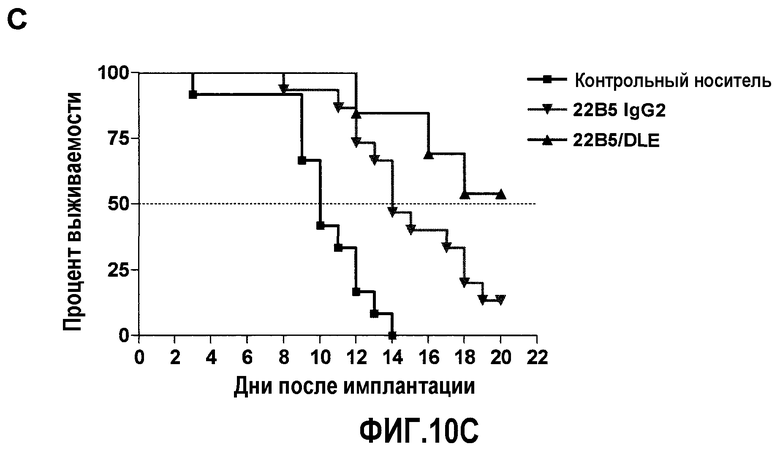

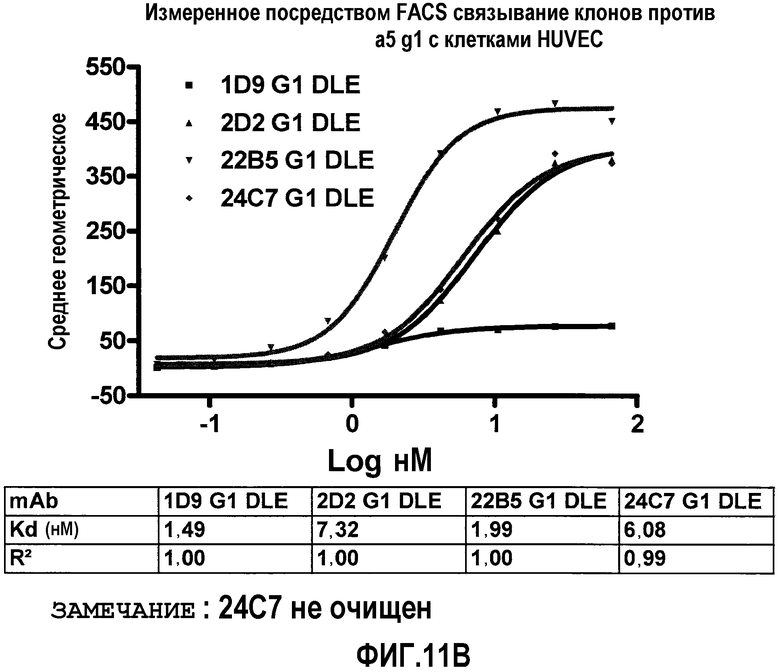
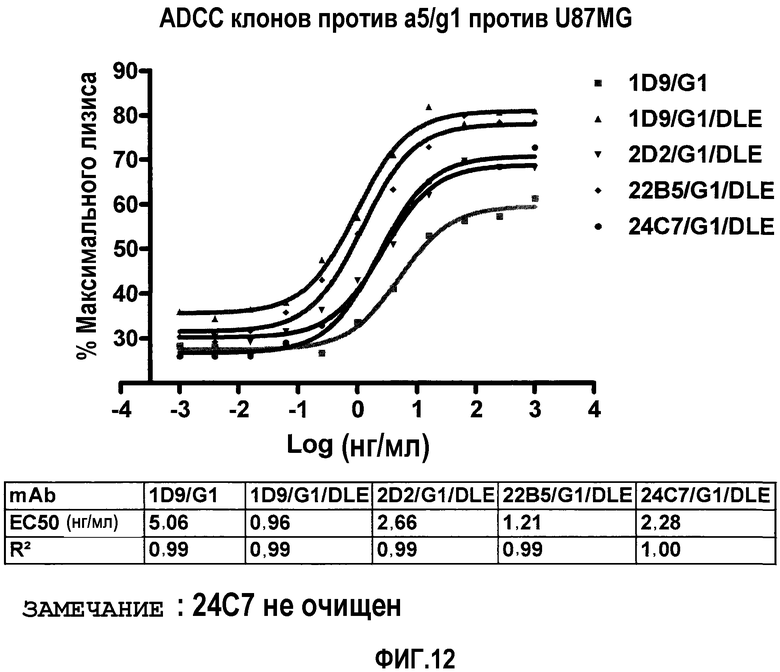
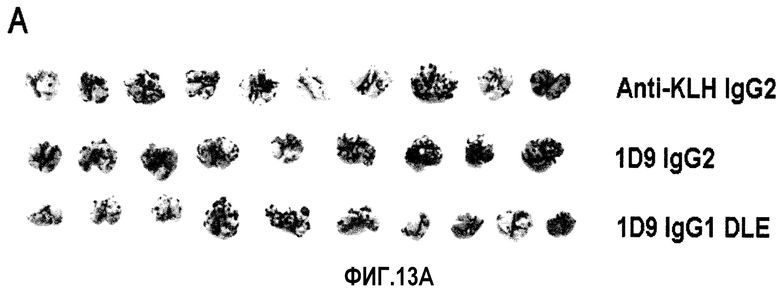
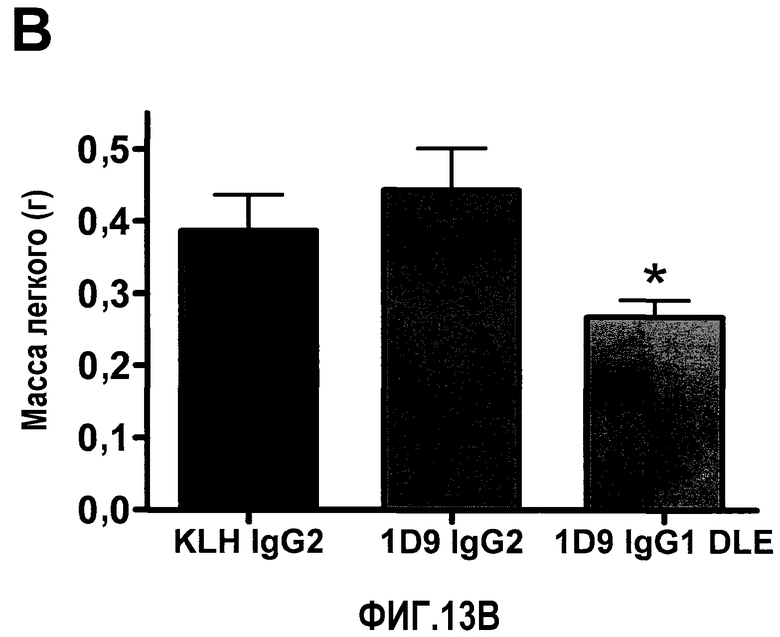
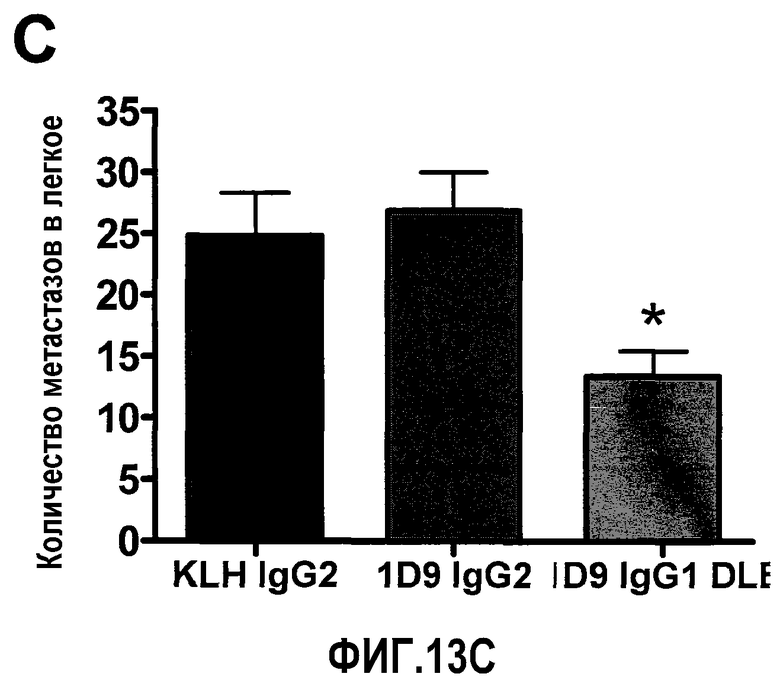
Настоящее изобретение относится к области иммунологии. Предложено выделенное моноклональное антитело к интегрину α5β1 человека. Антитело характеризуется тем, что содержит 6 CDR, 3 CDR из легкой цепи и 3 CDR из тяжелой цепи. Описана нуклеиновая кислота (НК), кодирующая антитело по изобретению, экспрессирующий вектор, содержащий молекулу НК; клетка-хозяин, содержащая вектор; и способ получения антитела на основе клетки. Раскрыты: композиция и способ для ингибирования роста опухолевых клеток, экспрессирующих интегрин α5β1 человека, на основе антитела. Описан вариант способа ингибирования роста опухолевых клеток, экспрессирующих интегрин α5β1 человека, использующий композицию. Изобретение обеспечивает новые антитела с высоким (порядка нм, как измерено FACS) сродством связывания с интегрином α5β1 человека, что может найти применение в медицине в терапии опухолей, опосредованных экспрессией интегрина α5β1. 7 н. и 6 з.п. ф-лы, 36 ил., 3 табл., 11 пр.
1. Выделенное моноклональное антитело к интегрину α5β1 человека, содержащее:
(a) CDR1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:1;
(b) CDR2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:2;
(c) CDR3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:3;
(d) CDR1 вариабельной области легкой цепи, содержащую SEQ ID NO:4;
(e) CDR2 вариабельной области легкой цепи, содержащую SEQ ID NO:5; и
(f) CDR3 вариабельной области легкой цепи, содержащую SEQ ID NO:6.
2. Антитело по п.1, содержащее:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO:7; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO:8.
3. Антитело по п.1 или 2, где указанное антитело представляет собой полноразмерное IgGl-антитело человека и где по меньшей мере одна аминокислота в Fc-области указанного антитела мутирована, и дополнительно, где у указанного антитела наблюдается ADCC больше, чем у идентичного антитела, у которого указанная по меньшей мере одна аминокислота не мутирована.
4. Антитело по п.3, в котором по меньшей мере одна мутация происходит в аминокислотном положении серин 247, аланин 338 или изолейцин 340.
5. Антитело по п.3, где по меньшей мере одну мутацию выбирают из группы, состоящей из мутаций: серин 247 на аспарагиновую кислоту (S247D), аланин 338 на лейцин (A338L) и изолейцин 340 на глутаминовую кислоту (I340E).
6. Антитело по п.5, где антитело содержит мутации S247D, A338L и I340E.
7. Антитело по п.1, где указанное антитело содержит тяжелую цепь, состоящую из аминокислотной последовательности, приведенной в SEQ ID N0:9, и легкую цепь, состоящую из аминокислотной последовательности, приведенной в SEQ ID NO:10.
8. Композиция для ингибирования роста опухолевых клеток, экспрессирующих интегрин α5β1 человека, содержащая эффективное количество антитела по любому из пп.1-7 и фармацевтически приемлемый носитель.
9. Выделенная молекула нуклеиновой кислоты, кодирующая антитело по любому из пп.1-7.
10. Экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты по п.9.
11. Клетка-хозяин для экспрессии антитела по любому из пп.1-7, содержащая экспрессирующий вектор по п.10.
12. Способ получения антитела против α5β1, включающий экспрессию антитела в клетке-хозяине по п.11.
13. Способ ингибирования роста опухолевых клеток, экспрессирующих интегрин α5β1 человека, включающий приведение указанных клеток в контакт с антителом по любому из пп.1-7 или композицией по п.8 в количестве, эффективном для ингибирования роста опухолевых клеток.

