Изобретение относится к области биотехнологии, и может найти применение в медицине для создания инновационного биомаркера рассеянного склероза, а также продиагностического и диагностического критерия.
В последнее десятилетие роль В-клеточного ответа, сопровождающегося образованием антител к собственным антигенам, приобретает все большее значение в этиологии и патогенезе аутоиммунных заболеваний. Среди патологий аутоиммунной природы особое место занимает рассеянный склероз (PC) - хроническое нейродегенератавное заболевание, приводящее к разрушению миелиновой оболочки нервных волокон. На данный момент не вызывает сомнений, что аутоантитела, вырабатываемые на мажорный компонент миелиновой оболочки - основной белок миелина (ОБМ), являются одним из важных патогенных факторов, влияющих на деградацию нервных волокон в центральной нервной системе при PC. Диагностика PC и прогностические критерии развития заболевания нуждаются в совершенствовании и дополнительной верификации. Существующий сегодня комплекс функциональных тестов, основанный на клинической картине течения болезни и составивший основу для создания единой шкалы инвалидизации пациентов (EDSS), также нуждается в дополнительном количественном уточнении. Одним из основных компонентов диагностического комплекса являются данные магнитно-резонансной томографии (МРТ) головного мозга пациентов. Однако выявляемые с помощью этого метода очаги демиелинизации не всегда являются абсолютно однозначным свидетельством развития PC, т.к. могут быть вызваны другими нейродегенеративными и посттравматическими процессами.
Известны разнообразные способы получения биосенсора на различные высокотоксичные патогены на основе двухбелковой флуоресцентной системы (патенты США №7495069, опубл. 2009 г., №7183066, опубл. 2007 г.; заявка WO/2000/073437, опубл. 2000 г.). Известен также биосенсор с принципом резонансного переноса энергии на различные высокотоксичные патогены на основе двухбелковой флуоресцентной системы (приведен в заявке РСТ/СА 2000/000620, опубл. 2000 г.). Общим серьезным недостатком приведенных сенсоров является неспособность измерения активности особого типа протеиназ - каталитических антител, специфичных к основному белку миелина, присутствующих в сыворотке крови больных рассеянным склерозом.
Известен наиболее близкий к заявленному биосенсор (Пономаренко Н.А. «Каталитические антитела - протеиназы», М., 2008, автореферат диссертации на соискание ученой степени доктора биологических наук, с.49). Серьезными недостатками данного биосенсора являются недостаточная точность и низкая стабильность.
Задачей изобретения является создание биосенсора на основе генетических конструкций для измерения активности каталитических антител, специфичных к основному белку миелина, присутствующих в сыворотке крови больных рассеянным склерозом.
Поставленная задача решается за счет:
конструирования рекомбинантной плазмидной ДНК pQe30_PS-CFP2/TurboYFP_MBP7 размером 4916 п.о., кодирующей гибридный белок PS-CFP2/TurboYFP_MBP7, содержащий последовательность белков PS-CFP2 и Turbo YFP, соединенных фрагментом основного белка миелина 80-104, и имеющий аминокислотную последовательность SEQ ID NO: 1, где рекомбинантная плазмидная ДНК имеет физическую карту, представленную на фиг.1, и состоит из:
- фрагмента ДНК, содержащего промотор транскрипции Т5 РНК-полимеразы, участок связывания рибосомы и последовательность гибридного белка PS-CFP2/TurboYFP_MBP7;
- фрагмента ДНК плазмиды pQe30, содержащего терминатор транскрипции Т5-РНК-полимеразы бактериофага Т5 и сайт инициации репликации;
- фрагмента ДНК гена β-лактамазы, детерминирующий устойчивость трансформированных плазмидой pQe30_PS-CFP2/TurboYFP_MBP7 клеток Escherichia coli к ампициллину, в качестве генетического маркера;
также за счет штамма Escherichia coli BL21(DE3)/pQe30_PS-CFP2/TurboYFP_MBP7 - продуцента гибридного белка PS-CFP2/TurboYFP_MBP7, полученного путем трансформации клеток штамма Escherichia coli BL21(DE3) рекомбинантной плазмидной ДНК pQe30_PS-CFP2/TurboYFP_MBP7;
и за счет способа получения белка PS-CFP2/TurboYFP_MBP7, включающего культивирование штамма-продуцента Escherichia coli BL21(DE3)/pQe30 PS-CFP2/TurboYFP_MBP7, разрушение клеток и очистку белка PS-CFP2/TurboYFP_MBP7 методом аффинной и гель-фильтрационной хроматографии.
Преимуществами биосенсора является значительно расширенная гамма определяемых патогенных антител за счет введенных фланкирующих аминокислотных остатков и, в то же время, увеличенная стабильность в растворе вследствие значительного уменьшения физического взаимодействия белков-флуорофоров. По сравнению с известным биосенсором расширилась как чувствительность бисенсора, так и его специфичность в отношении пула каталитических антител. Инновационные свойства биосенсора позволяют использовать его для всего круга пациентов с рассеянным склерозом (как для прогрессирующей, так и для ремиттирующей стадий), что невозможно было достигнуть ранее. Внедренные аминокислотные инсерции также значительно повысили физико-химические свойства биосенсора. Пониженная молекулярная интерференция белков-флуорофоров увеличивает стабильность биосенсора и квантовый выход резонансного переноса энергии. Достигнутый технический результат состоит в улучшенной точности, высокой стабильности биосенсора при хранении и процедуре анализа, что, в свою очередь, значительно повысило эффективность и простоту диагностики. Высокая чувствительность и селективность биосенсора снижает количество ложно-положительных результатов и позволяет уверенно различать рассеянный склероз и другие нейродегенеративные заболевания при диагностике.
Преимущество предлагаемого изобретения заключается, во-первых, в том, что в результате осуществления способа получают белок, содержащий фрагмент основного белка миелина между двумя флуоресцентными белками, представляющий собой FRET-пару. Было однозначно показано, что введенные впервые фланкирующие аминокислотные последовательности крайне важны для связывания и последующего гидролиза биосенсора каталитическими патогенными антителами. Введение «якорных» концевых остатков значительно улучшило физико-химические характеристики продукта. Низкая токсичность целевого белка для штамма-продуцента позволяет достичь высоких выходов гибридного белка (около 20% от суммарного клеточного белка). Достоинством способа является простота технологии очистки EPeFRET. Высокий уровень экспрессии и его крайне высокая растворимость и стабильность в водных растворах позволяют осуществлять очистку в две стадии с получением гомогенности продукта более 98%. Для биосинтеза рекомбинантного белка применяются оптимальные регуляторные элементы, контролирующие его экспрессию: промотор и терминатор для высокопроцессивной РНК-полимеразы бактериофага Т5, консенсусный сайт связывания бактериальной рибосомы, стартовый и стоп-кодоны.
Конструирование новой плазмидной ДНК pQe30_PS-CFP2/TwboYFP_MBP7, содержащей флуоресцентные белки PS-CFP2 и TurboYFP, соединенные фрагментом ОБМ80-104 (EPeFRET), производят путем замены исходного линкера одностадийным клонированием с использованием двух олигонуклеотидов. Отбор положительных клонов проводят при помощи ПЦР с использованием специфических праймеров и последующим рестриктным анализом выделенной плазмидной ДНК. Структуру гена, кодирующего гибридный белок, содержащий фрагмент ОБМ 80-104, определяют секвенированием по методу Сэнгера. Экспрессию EPeFRET проводят в штамме E.coli BL21(DE3). При подросте культуры на 37°С после достижения оптической плотности OD600 0.6-0.8 единиц клеточную суспензию переносят на комнатную температуру и инкубируют 24-36 часов для созревания хромофора. Растворимую фракцию белков получают следующим образом. Клеточный лизат наносят на предварительно уравновешенный буфером металлохелатный сорбент Talon Superflow (BD, США) на скорости 1 мл/мин. Затем колонку промывают 100-120 объемами раствора, содержащего 50 мМ фосфата натрия, 300 мМ NaCl, pH 7.0. Элюцию рекомбинантного белка проводят 50 мМ раствором ЭДТА на скорости 0.5 мл/мин. Далее проводят гель-фильтрацию на колонке Superdex 75 в растворе PBS/TBS на скорости 0.5 мл/мин. Полученный раствор белка держат неделю при температуре 10°С с 0.05% азидом натрия для окончательного созревания хромофора. Измерения флуоресценции проводят на флуориметре Саrу Eclipse (Varian) и планшетном ридере Tecan Genios (Tecan) при длине волны возбуждения и испускания 405 и 538 нм соответственно.
Рекомбинантная плазмидная ДНК pQe30_PS-CFP2/TurboYFP_MBP7, кодирующая гибридный белок EPeFRET, содержащий пептид ОБМ 80-104, характеризуется следующими признаками:
- имеет размер молекулы 4916 п.о.;
- кодирует гибридный белок EPeFRET, содержащий модифицированную последовательность белков PS-CFP2 и Turbo, соединенных фрагментом основного белка миелина 80-104;
- состоит из фрагмента ДНК, содержащего промотор транскрипции Т5-РНК-полимеразы, участок связывания рибосомы и последовательность гибридного полипептида PS-CFP2/TurboYFP MBP7, содержащего последовательность белков PS-CFP2 и Turbo, соединенных фрагментом основного белка миелина 80-104 и фрагмента ДНК плазмиды рQе30, содержащего терминатор транскрипции Т5-РНК-полимеразы, сайт инициации репликации и ген β-лактамазы;
- имеет уникальную совокупность признаков: промотор и терминатор транскрипции РНК-полимеразы бактериофага Т5; сайт связывания рибосомы; искусственный ген, кодирующий гибридный белок PS-CFP2/TurboYFP MBP7, содержащий последовательность бежов PS-CFP2 и Turbo, соединенных фрагментом основного белка миелина 80-104; ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pQe30_PS-CFP2/TurboYFP_MBP7 клеток к ампициллину.
Для получения штамма-продуцента гибридного белка EPeFRET препаратом плазмидной ДНК pQe30_PS-CFP2/TurboYFP_MBP7 трансформируют компетентные клетки Escherichia coli BL21(DE3) и проводят отбор клонов, обладающих способностью экспрессировать рекомбинантный белок. Наличие и уровень экспрессии рекомбинантных белков контролируют с помощью денатурирующего ПААГ-электрофореза.
Клетки растут при температуре от 4°С до 40°С при оптимуме рН от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы. Клетки проявляют устойчивость к ампициллину (до 500 мкг/мл), обусловленную наличием в плазмиде pQe30_PS-CFP2/TurboYFP_MBP7 гена β-лактамазы (bla).
Преимущества штамма E.coli BL21 (DE3) в качестве основы для создания штамма-продуцента заключаются в использовании бактерий с фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого рекомбинантного белка и загрязнения препарата наиболее активными протеазами E.coli. В хромосомную ДНК BL21 (DЕ3) интегрирован ген Т5-РНК полимеразы, что, совместно с использованием Т5 промотора и Т5 терминатора в плазмиде pQe30 PS-CFP2/TurboYFP_MBP7, обеспечивает быструю и эффективную продукцию белка клетками Е.coli при индукции изопропилтио-β-D-галактозидом или лактозой.
Клетки Е.coli BL21(DE3)/pQe30_PS-CFP2/TurboYFP_MBP7 являются хорошим продуцентом биосенсора вследствие введенной в них уникальной плазмиды pQe30_PS-CFP2/TurboYFP_MBP7. При индукции изопропилтио-β-D-галактозидом происходит эффективный биосинтез гибридного белка EPeFRET, который накапливается в клетках в количестве 20-25% от суммарного белка бактерии.
Биосинтез продукта проводят следующим образом: клетки штамма E.coli BL21(DE3)/pQe30_PS-CFP2/TurboYFP_MBP7 выращивают в богатой среде (например, 2×YT) с добавлением 100-200 мкг/мл ампициллина. При подросте культуры на 37°С после достижения оптической плотности OD600 0.6-0.8 единиц индуцируют синтез белка изопропил-β-D-галактозидом в концентрации 0,1-1,0 мМ, далее клеточную суспензию переносят на комнатную температуру и инкубируют 24-36 часов для созревания хромофора. Увеличение выхода рекомбинантного белка может быть достигнуто с помощью принудительной аэрации культуральной жидкости и использования обогащенных питательных сред (например, с добавлением глюкозы).
Способ получения белка EPeFRET из клеток продуцента включает следующие стадии: отделение бактерий от культуральной среды с помощью центрифугирования, их разрушение одним из обычно применяемых способов (ультразвуковой дезинтеграцией, химическим лизисом с использованием детергентов и хаотропных агентов), отделение от телец включения и водонерастворимых компонентов клетки и очистку целевого белка методом аффинной и гель-проникающей хроматографии.
Идентичность белка EPeFRET заявленному устанавливают методами ПААГ-электрофореза в трис-трициновой системе, MALDI-времяпролетной масс-спектрометрии, секвенированием N-концевой аминокислотной последовательности по методу Эдмана, а также тестированием на предмет расщепления природными протеазами и высокоочищенными каталитическими антителами. Степень очистки EPeFRET определяют методами ПААГ-электрофореза в трис-трициновой системе и MALDI-времяпролетной масс-спектрометрии. Выход EPeFRET, полученного заявленным способом, с чистотой не менее 95% составляет не менее 4 мг/л культуры.
Изобретение иллюстрируют чертежами
Фиг.1. Физическая карта рекомбинантной плазмиды pQe PS-CFP2/TurboYFP MBP7. BamHI, BglII, HindIII - сайты рестрикции; Т5 promoter - промотор транскрипции; PS-CFP2 - последовательность, кодирующая флуоресцентный циановый белок; YSP -последовательность, кодирующая флуоресцентный желтый белок, MBP7 - последовательность, кодирующая последовательность основного белка миелина 80-104.
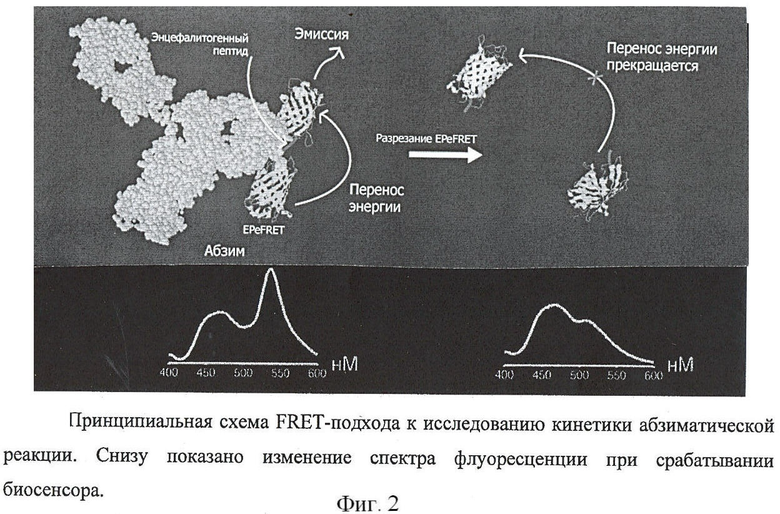
Фиг.2. Принципиальная схема FRET-подхода к исследованию кинетики абзиматической реакции. Снизу показано изменение спектра флуоресценции при срабатывании биосенсора.
Фиг.3. Кривые ингибирования абзиматического и ферментативного гидролиза EPeFRET слитным белком Тrх с фрагментом ОБМ80-104 (А), его мутантом R97A (Б), Сорахоnе (В) и белком-носителем (Г).
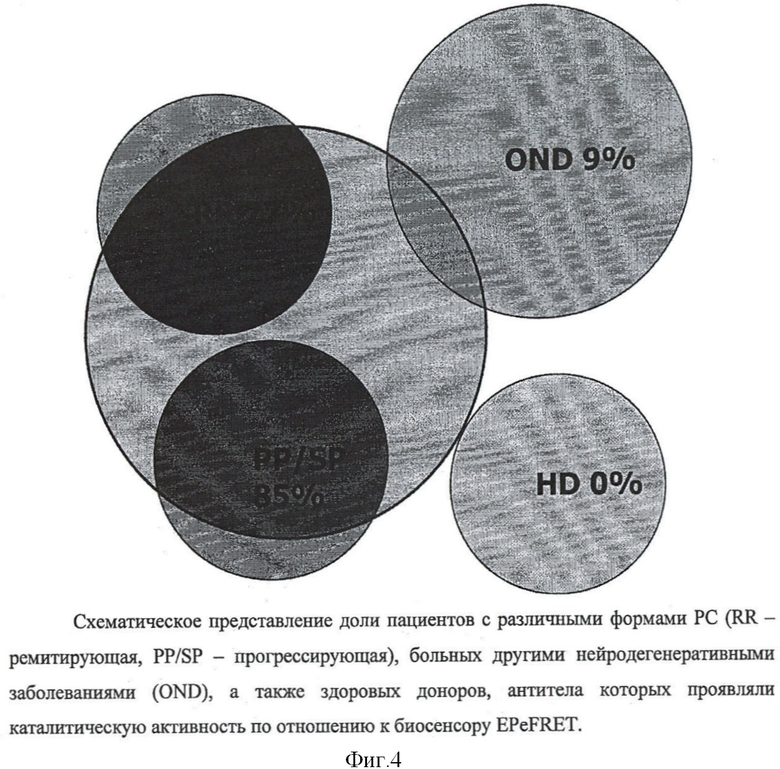
Фиг.4. Схематическое представление доли пациентов с различными формами PC (RR - ремитирующая, PP/SP - прогрессирующая), больных другими нейродегенеративными заболеваниями (OND), а также здоровых доноров, антитела которых проявляли каталитическую активность, измеренную биосенсором EPeFRET.
Изобретение иллюстрируют следующие примеры.
Пример 1. Конструирование рекомбинантной плазмидной ДНК pQe30_PS-CFP2/TurboYFP_MBP7. В оригинальную плазмиду pQe30 (Qiagen) интегрируют 2 гена белков PS-CFP2 и turboYFP методом Quick change (быстрой замены), как описано в (Но, S.N., Hunt, H.D., Horton, R.M., Pullen, J.K., and Pease, L.R. (1989) Gene 77(1), 51-59).
Вначале проводят стандартную ПЦР с оригинальных плазмид pPS-CFP2-C и pTurboYFP-C, содержащих гены PS-CFP2 и turboYFP соответственно, со следующих праймеров:

Обе ПЦР проводят при следующих условиях:
Т=94°С - 5 минут
1 цикл
Т=94°С - 30 секунд
Т=60°С - 30 секунд
Т=72°С - 1 минута
25 циклов
Используют смесь Taq/Pfu полимераз (Fermentas) в соотношении 10/1.
Далее проводят ПЦР с использованием полученных ранее ПЦР-продуктов без использования праймеров при следующих условиях:
Состав реакционной смеси:
Пцр - продукт гена PS-CFP2 - 2 мкл (0.75 мкг ДНК)
Пцр - продукт гена turboYFP - 2 мкл (0.75 мкг ДНК)
реакционный буфер для herculase полимеразы - 0,6 мкл
высокопроцессивная полимераза - Herculase (Amersham) - 0,1 мкл
дезоксирибонуклеотиды - 0,6 мкл (0,1 мМ каждого)
вода mQ - 0,7 мкл
Режим ПЦР:
Т=94°С - 5 минут
1 цикл
Т=94°С - 30 секунд
Т=55°С - 30 секунд
Т=72°С - 15 минут
5 циклов
Далее проводят стандартный ПЦР с использованием полученного в предыдущем ПЦР
продукта в качестве матрицы в разведении 1/50
Т=94°С - 5 минут
1 цикл
Т=94°С - 30 секунд
Т=60°С - 30 секунд
Т=72°С - 1,5 минуты
25 циклов
Используют смесь Taq/Pfu полимераз (Fermentas) в соотношении
10/1
Полученный ПЦР продукт и плазмиду pQE30 подвергают рестрикции с использованием ферментов BamHI и HindIII с дальнейшим лигированием, результирующая конструкция получила название pQe_PS-CFP2/turboYFP_Bgl.
Далее плазмиду pQe_PS-CFP2/twboYFP_Bgl подвергают рестрикции с помощью фермента BglII и дальнейшему лигированию с олигонуклеотидом, соответствующим фрагменту ОБМ80-104, обработанными предварительно рестрикционными эндонуклеазами BglII и BclI. Конструкция, содержащая энцефалитогенный фрагмент ОБМ 80-104, получила название pQe_PS-CFP2/turboYFP_MBP7.
Пример 2. Экспрессию слитных белков проводят в штамме E.coli BL21(DE3)/pQe30_PS-CFP2/TurboYFP_MBP7 (продуцент гибридного белка PS-CFP2/TurboYFP_MBP7, полученный путем трансформации клеток штамма Escherichia coli BL21(DE3) уникальной рекомбинантной плазмидной ДНК pQe30_PS-CFP2/TurboYFP_MBP7). При подросте культуры на 37°С после достижения оптической плотности OD600 0.7 единиц, клеточную суспензию переносят на комнатную температуру и инкубируют 36 часов для созревания хромофора. Растворимую фракцию белков получают следующим образом. Супернатант наносят на предварительно уравновешенный буфером металлохелатный сорбент Talon Superflow (BD, США) на скорости 1 мл/мин. Затем колонку промывают 120 объемами раствора, содержащего 50 мМ фосфата натрия, 300 мМ NaCl, pH 7.0. Элюцию рекомбинантного белка проводят 50 мМ раствором ЭДТА на скорости 0.5 мл/мин. Далее проводят гель-фильтрацию на колонке Superdex 75 в растворе PBS/TBS на скорости 0.5 мл/мин. Полученный раствор белка держат неделю при температуре 10°С с 0.05% азидом натрия для окончательного созревания хромофора. Измерения флуоресценции проводят на флуориметре Саrу Eclipse (Varian) и планшетном ридере Tecan Genios (Tecan) при длине волны возбуждения и испускания 405 и 538 нм соответственно.
Все хроматографические стадии выделения проводят на приборах для скоростной и высокоэффективной жидкостной хроматографии (FPLC, HPLC) фирм Gilson и Waters (Millipore).
Выделение поликлональных IgG-антител из сывороток крови людей и мышей проводят по следующей схеме. Первоначально фракцию антител осаждают 50%-ным сульфатом аммония на 24 часа. Процедуру переосаждения повторяют трижды. Белки, полученные таким образом, разводят 1:10 по отношению к объему исходной сыворотки раствором PBS и наносят на скорости 0.5 мл/мин на колонку HiTrap Protein-G Sepahrose (Amresham, США). Затем колонку промывают 100 объемами буфера нанесения на скорости 2 мл/мин для отделения неспецифически сорбированных сывороточных белков. Фракцию иммуноглобулинов класса G элюируют с колонки 100 мМ раствором глицин-НСl рН 2.6 и незамедлительно нейтрализуют 2 М раствором трис-основания до рН 7.5.
Выделение ОБМ-связывающих антител проводят методом аффинной хроматографии с использованием в качестве сорбента иммобилизованного на сефарозе основного белка миелина. Для этого тотальную фракцию IgG наносят на иммобилизованный ОБМ на скорости 0.05 мл/мин в PBS. Далее колонку промывают 90 объемами буфера нанесения на скорости 2 мл/мин. Элюцию ОБМ-связывающих антител с сорбента проводят 100 мМ раствором глицин-НСl рН 2.6 и нейтрализуют раствором трис-основания до рН 7.5.
Идентификацию препаратов антител и оценку их чистоты проводят с помощью метода денатурирующего электрофореза в полиакриламидном геле с последующим окрашиванием серебром и иммуноферментным анализом.
Кинетические измерения проводят с использованием различных концентраций EPeFRET (от 0.25 до 30 мкМ) в растворе PBS для антител или TBS-CaCl2 в случае ферментов. Кислотный буфер для катепсина D неприменим вследствие резкого уменьшения выхода флуоресценции субстрата в таких условиях. Конечные концентрации антител, трипсина и металлопротеазы матрикса-3 80, 40 и 30 нМ соответственно. Катепсин D неактивен в подобных концентрациях. Ингибирование осуществляют при постоянной концентрации EPeFRET 1 мкМ и изменяющейся концентрации ингибиторов от 0.001 до 100 мкМ приблизительно через порядок. Концентрацию антител берут 80 нМ, трипсина и ММР-3 - 10 нМ. Реакцию проводят в объеме 120 мкл в черных 96-ти луночных плашках, измерение ведут при 405/535 нм соответственно.
Кинетические константы гидролиза EPeFRET аутоантителами, трипсином и металлопротеазой матрикса - 3, а также ингибирование данной реакции слитными белками и Сорахоnе, известным лекарственным средством при PC, представляющем собой сополимер лизина, глутаминовой кислоты, аланина и тирозина, представлены в таблице 1 и на фиг.2 и 3.
(ceк-1M-1)
Каталитическая эффективность изученных биокатализаторов уменьшалась примерно на порядок по ряду трипсин - ММР-3 - абзим. Характер ингибирования расщепления EPeFRET слитными белками тиоредоксина с фрагментом ОБМ80-104, его мутантом, белком-носителем и Сорахоnе радикально отличается в случае катализа абзимами и модельными протеазами. Если для каталитических антител способность к ингибированию в ряду Тrх-ОБМ80-104, Тrх-ОБМ80-104(R97А), Сорахоnе, Тrх падает со значения IC50 7.0·10-3 до >100 мкМ, то для ферментов рассчитанные значения IC50 находятся в диапазоне от 3 до 20 мкМ независимо от типа ингибитора. Эффективное действие Сорахоnе на трипсин по-видимому объясняется высоким (до 29%) содержанием остатков лизина в этом сополимере.
Пример 3. Для характеристики субстратной специфичности антител FRET-сенсоры инкубируют с иммуноглобулинами (суммарная IgG фракция), изолированными из сыворотки крови 26 больных рассеянным склерозом, 22 пациентов с другими нейродегенеративными заболеваниями (other neurological diseases, OND), 11 здоровых доноров (healthy donors, HD) и мышей аутоиммунной линии SJL с ЕАЕ, индуцированным иммунизацией ОБМ. Замороженные образцы сывороток предоставлены Московским областным научно-исследовательским институтом (МОНИКИ) и Клинической больницей РАН. Характеризация каталитической и связывающей активности аутоантител осуществлена для 26 больных рассеянным склерозом, 22 пациентов с другими нейродегенеративными заболеваниями и 11 здоровых доноров. Возраст пациентов с PC варьировался от 16 до 72 лет (среднее 39.5 лет, интерквартильное значение от 28 до 48 лет). Параметр EDSS изменялся от 0 до 6.5 единиц (среднее 3.2±1.8). Клинический диагноз верифицировался в соответствии с международными критериями по классификации Позера (Poser, C.M. and V.V.Brinar, Diagnostic criteria for multiple sclerosis. Clin Neurol Neurosurg, 2001. 103(1); p.1-11) и подтверждался данными лабораторных исследований. Когорты пациентов с другими нейродегенеративными заболеваниями и здоровые доноры имели среднее значение по возрасту 65 лет (интерквартильи от 39 до 72 лет) и 33.5 лет (интерквартильи от 26 до 47 лет) соответственно. Необходимые разрешения получены от каждого пациенты в соответствии с нормативными документами Министерства здравоохранения Российской Федерации. Статистический анализ проводили при помощи программы SPSS 13 с использованием непараметрического U теста Mann-Whitney и ранговой корреляции Spearman.
Мыши аутоиммунной линий SJL получены из питомника Harlan (UK) и содержались в виварии филиала Института Биоорганической Химии РАН в г.Пущине в стерильных условиях, минимизирующих контакт иммунной системы с внешними антигенами (Specific Pathogen-Free статус). Возраст мышей, используемых в экспериментах, варьируется от 5 недель до 12 месяцев. Индукцию ЕАЕ у модельных мышей проводят по стандартной методике {Current protocols on CD. [cited; computer laser optical discs], Coligan, J.E. and National Institutes of Health (U.S.), Current protocols in immunology. 1992, New York: Greene Pub. Associates and Wiley-Interscience. 5 v.). Мышей в возрасте от 6 до 8 недель иммунизируют введением эмульсии 50 мкг антигена (ОБМ быка) в полном адъюванте Френда, содержащем 4 мг/мл инактивированного нагреванием М. tuberculosis. Иммунизацию проводят дважды с недельным интервалом. За день до и через два после первой иммунизации мышам интраперитонеально вводят 400 нг pertussis токсина. Мониторинг мышей на предмет развития ЕАЕ ведут ежедневно, сбор крови производят методом декапитации через 2-3 недели.
Каталитические свойства проявили антитела, изолированные из 77% пациентов с прогрессирующим и 85% с ремитирующим типом течения PC. Среди больных нейродегенеративными заболеваниями и здоровых доноров доля положительного каталитического ответа составляет 9% и 0% соответственно (фиг.4). Исключение составляют два случая остеохондроза, при которых было отмечено наличие каталитической деградации пептида ОБМ под действием аутоантител. Причем эти больные характеризуются значительной длительностью патологического процесса и серьезными нейрональными поражениями. Контроль специфичности гидролиза осуществляют деградацией рекомбинантных белков бычим трипсином, катепсином D и металлопротеазой матрикса 3 (ММР-3). 

Изобретение относится к области биотехнологии, конкретно к получению рекомбинантных биомаркеров полипептидной природы, и может быть использовано в диагностике рассеянного склероза. Для получения белка PS-CFP2/TurboYFP_MBP7 конструируют рекомбинантную плазмидную ДНК PS-CFP2/TurboYFP_MBP7 размером 4916 п.о., кодирующую гибридный белок, содержащий последовательность белков PS-CFP2 и Turbo YFP, соединенных фрагментом основного белка миелина 80-104. В состав плазмидной ДНК также входит промотор транскрипции Т5 РНК-полимеразы, участок связывания рибосомы; фрагмент ДНК плазмиды ген β-лактамазы, детерминирующий устойчивость клеток Escherichia coli к ампициллину, в качестве генетического маркера. Полученной плазмидной ДНК трансформируют клетки штамма Escherichia coli BL21(DE3) с получением штамма-продуцента гибридного белка PS-CFP2/TurboYFP_MBP7. Для получения белка PS-CFP2/TurboYFP_MBP7 проводят культивирование штамма-продуцента Escherichia coli BL21(DE3)/pQe30_PS-CFP2/TurboYFP_MBP7, разрушают клетки и очищают целевой белок методом аффинной и гель-фильтрационной хроматографии. Изобретение позволяет повысить чувствительность и стабильность биосенсора, а также расширить его специфичность в отношении пула каталитических антител. 3 н.п. ф-лы, 7 ил., 1 табл.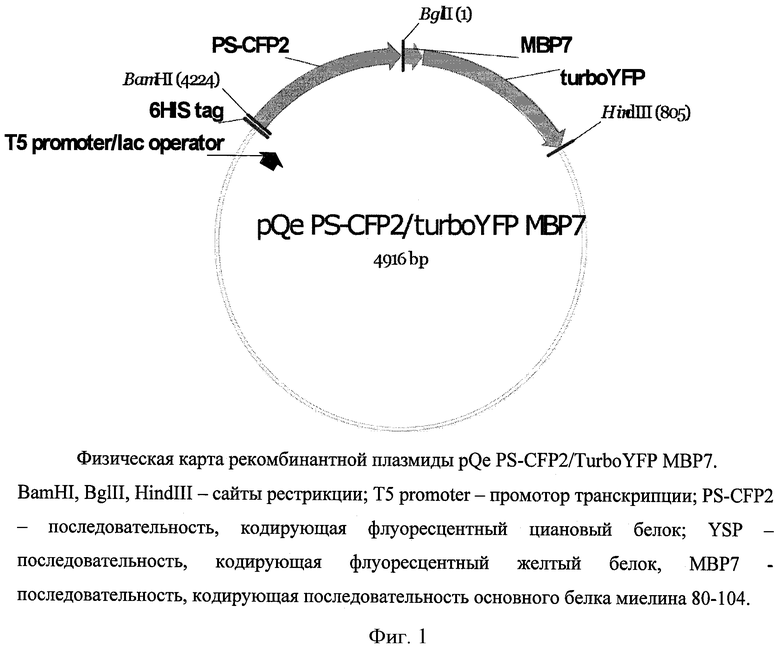
1. Рекомбинантная плазмидная ДНК pQe30_PS-CFP2/TurboYFP_MBP7 размером 4916 п.о., кодирующая гибридный белок PS-CFP2/TurboYFP_MBP7, содержащий последовательность белков PS-CFP2 и Turbo YFP, соединенных фрагментом основного белка миелина 80-104, и имеющий аминокислотную последовательность SEQ ID NO:1, где рекомбинантная плазмидная ДНК имеет физическую карту, представленную на фиг.1, и состоит из:
фрагмента ДНК, содержащего промотор транскрипции Т5 РНК-полимеразы, участок связывания рибосомы и последовательность гибридного белка PS-CFP2/TurboYFP_MBP7;
фрагмента ДНК плазмиды pQe30, содержащего терминатор транскрипции Т5-РНК-полимеразы бактериофага Т5 и сайт инициации репликации;
фрагмента ДНК гена β-лактамазы, детерминирующего устойчивость трансформированных плазмидой pQe30_PS-CFP2/TurboYFP_MBP7 клеток Escherichia coli к ампициллину, в качестве генетического маркера.
2. Штамм Escherichia coli BL21(DE3)/pQe30_PS-CFP2/TurboYFP_MBP7 - продуцент гибридного белка PS-CFP2/TurboYFP_MBP7, полученный путем трансформации клеток штамма Escherichia coli BL21(DE3) рекомбинантной плазмидной ДНК pQe30_PS-CFP2/TurboYFP_MBP7 по п.1.
3. Способ получения белка PS-CFP2/TurboYFP_MBP7, включающий культивирование штамма-продуцента Escherichia coli BL21(DE3)/pQe30_PS-CFP2/TurboYFP_MBP7 по п.2, разрушение клеток и очистку белка PS-CFP2/TurboYFP_MBP7 методом аффинной и гель-фильтрационной хроматографии.
| ПОНОМАРЕНКО Н.А | |||
| Каталитические антитела - протеазы, 2008: Автореферат диссертации на соискание ученой степени доктора биологических наук, с.49 | |||
| BELOGUROV А.А | |||
| et al | |||
| Recognition and Degradation of Myelin Basic Protein Peptides by Serum Autoantibodies: Novel Biomarker for Multiple Sclerosis, The Journal of Immunology, 2008, v.180, n.2, p.1258-1267 | |||
| EP 1674478 A1, 28.06.2006. |

