Изобретение относится к области клеточной биологии, молекулярной биологии и биотехнологии и может быть использовано при исследовании механизмов сигнальных путей трансформирующих факторов роста (TGF-β), а также для нейтрализации TGF-β при различных заболеваниях, таких как фибропролиферативные нарушения и таких опухолевых заболеваний, где нерегулируемая экспрессия трансформирующего фактора роста-β (TGF-β) играет ключевую роль. Многофункциональный белок, трансформирующий факторы роста-β (TGF-β), играет ключевую роль в процессах эмбрионального развития, иммунного ответа, гематопоэза и развития раковых заболеваний. TGF-бета сигнализация играет центральную роль в развитии опухоли, метастазировании и ангиогенеза. Сигналы TGF-β передаются посредством двух типов рецепторов: типа I и типа II (белки TβRI и TβRII). Существует ряд фибропролиферативных нарушений, когда нерегулируемая экспрессия TGF-β играет ключевую роль. Суперэкспрессия TGF-β наблюдается при гломерулонефрите, пульмонарном фиброзе, циррозе печени и келоидах, что позволяет предположить, что молекулы-антагонисты TGF-β, включая TβRII, могут быть полезными при лечении этих заболеваний. Высокий уровень TGF-β продуцируется многими типами опухолей, включая меланому и рак груди, толстой кишки, пищевода, желудка, печени, легких, поджелудочной железы и простаты, а также и при гематологических злокачественных заболеваниях. Нерегулируемая иммуноподавляющая функция TGF-β провоцирует образование опухолей, в частности, при поздних стадиях болезни, когда опухоль начинает метастазировать. Показано, что рекомбинантный растворимый рецептор TβRII 23-156 обладает антифиброгенной и антиопухолевой активностью (при концентрациях от 2 до 5 мг/кг веса животных) в метастазирующих опухолях путем нейтрализации нежелательного высокого содержания TGF-β1 и TGF-β3 в крови. (Won J., Kim H., Park E.J., et al. Tumorigenicity of mouse thymoma is suppressed by soluble type II transforming growth factor_receptor therapy. Cancer Res 1999; 59:1273-7. Zheng H., Wang J., Koteliansky V.E., Gotwals P.J., Hauer-Jensen M. Recombinant soluble transforming growth factor - type II receptor ameliorates radiation enteropathy in mice. Gastroenterology 2000; 119:1286-96). Высокая специфичность рецептора TβRII к изоформам TGF-β1 и TGF-β3 и низкая к TGF-β2 указывают на то, что TβRII является основным посредником при передаче сигнала от TGF-β1 и TGF-β3.
Молекула TβRII представляет собой трансмембранный белок II типа, содержит 567 аминокислотных остатков, из которых 1-166 составляют внеклеточный или лиганд-связывающий, 167-187 трансмембранный и 187-567 цитоплазматический домен. Последний обогащен сериновыми и треониновыми остатками и обладает серин/треонин - специфической протеинкиназной активностью. Внеклеточный лиганд, связывающий домен, обогащен цистеинами и содержит 6 дисульфидных связей, что затрудняет рефолдинг белка TβRII при экспрессии в бактериях и впоследствии она образует тельца включения.
Известен способ получения белка TβRII в E.coli (Harrie L. Glansbeek, Henk M. van Beuningen, Elly L. Vitters, Peter M. van der Kraan, and Wim B. van den Berg, Expression of recombinant human soluble type II transforming growth factor-b receptor in Pichia pastoris and Escherichia coli: two powerful systems to express a potent inhibitor of transforming growth factor-b// Protein expression and purification. 1998. Vol.12, pp.201-207). В данной работе для экспрессии белка TβRII клетки Escherichia coli M15[pREP4] трансформируют вектором pQE-16/TβRII(3-159 aa) и индуцируют с помощью 2 мМ IPTG (изопропил_β-D-1-тиогалактопиранозид). Белок получают из телец включения, используя процедуру растворения и ренатурации. Максимальная экспрессия целевого белка составляет 1.5 мг с 1 л культуры клеток.
Известен способ получения белка TβRII (Christian С. Boesen, Shawn A. Motyka, Apisit Patamawenu and Peter D. Sun, Development of a recombinant bacterial expression system for the active form of a human transforming growth factor b type II receptor ligand binding domain // Protein expression and purification. 2000. Vol.20, pp.98-104). В работе используют различные подходы к получению активного рекомбинантного рецептора TβRII, включая использование экспрессирующей конструкции слитного белка GST/TβRII (где GST - глутатион S-трансфераза) и очистку целевого белка из телец включения. При использовании системы экспрессии бакуловирусов необходимо дальнейшее дегликозилирование. В случае использования конструкции с GST-тагом получается очень низкий выход активного препарата. При рефолдинге белка TβRII из телец включения выход активного препарата составляет 5 мг с 1 л культуры клеток.
Известен способ получения белка TβRII (Mark S. Fahey, David Dawbam, Shelley J. Alien, Ian C. Paterson and Stephen S. Prime, Expression of recombinant extracellular domain of the type II transforming growth factor-b receptor: utilization in a modified enzyme-linked immunoabsorbent assay to screen TGF-β agonists and antagonists// Analytical biochemistry. 2001. Vol.290, pp.272-276). В этой работе экспрессируют и очищают слитный белок MB-TβRII (где MB - мальтозосвязывающий белок) из растворимой фракции цитоплазматических белков с помощью трансформации плазмиды pMAL-c2TβRII в штамм Escherichia coli XL-1. Выход слитного белка MB-TβRII составляет 15 мг с 1 л культуры клеток, где целевой белок TβRII составляет всего 26% с выходом 3.9 мг.
Все известные способы получения TβRII из Escherichia coli не обеспечивают высокие выходы (1.5 до 5 мг с 1 л культуры клеток). Кроме того, первые два препарата были получены из нерастворимых форм белка (тельца включения), используя различные методы ренатурации, что не обеспечивает однородность препарата и высокую активность. Эти препараты содержат гистидиновый таг (6xHis-rar), что может повлиять на физико-химические и биологическтие свойства природного белка. Полученный препарат слитного белка MB-TpRII не может быть применен в терапии различных заболеваний.
Изобретение решает задачу получения высокоочищенного целевого белка рецептора II типа трансформирующего фактора роста-β человека с нативным N-концом, не содержащим гистидиновый таг, с повышенным (140 мг с 1 л культуры клеток) выходом целевого продукта.
Поставленная задача решается за счет конструирования рекомбинантной плазмидной ДНК pET-32a/TβRII, кодирующей полипептид лиганд-связывающего домена рецептора II типа трансформирующего фактора роста-β человека (6.237 т.п.о.), состоящей из BglII/HmdIII- фрагмента ДНК плазмиды рЕТ-32а длиной 5.838 т.п.о., содержащего lacI-промотор Е.coli, T7 промотор, Trx·Tag кодирующую последовательность, His·Tag кодирующую последовательность, S·Tag кодирующую последовательность, T7 терминатор, lacI-кодирующую последовательность, bla- (ген β-лактамазы) кодирующую последовательность, последовательность f1 участок ori инициации репликации, а также BglII/HindIII фрагмента гена кодирующего ДНК синтетического гена полипептид лиганд-связывающего домена рецептора II типа трансформирующего фактора роста-β человека (TβRII); за счет штамма бактерий Escherichia coli BL21(DE3)/pET-32a/TβRII - продуцента слитного белка тиоредоксин/TβRII; а также за счет способа ренатурации и очистки целевого белка TβRII человека, включающего выделение растворимой фракции цитоплазматических белков из штамма-продуцента Escherichia coli BL21(DE3)/pET-32a/TβRII и ее хроматографическую очистку с последующими ренатурацией слитного белка тиоредоксин/ TβRII при инкубации в течение 72 часов при 4°C, в буфере, содержащем 0.5 М аргинина, в присутствии окисленного и восстановленного глутатиона в соотношении 1:50 мМ соответственно, диализом, расщеплением энтеропептидазой и очисткой целевого белка TβRII от тиоредоксина. Высокая биологическая активность и низкая стоимость полученного препарата позволяют использовать препарат при терапии рака и при фибропролиферативных нарушениях.
Получение экспрессирующей конструкции pET-32a/TβRII для экспрессии в клетках Е.coli в виде слитного белка с несущим белком в виде тиоредоксина увеличивает растворимость слитного белка, позволяя чистить целевой белок непосредственно из фракции цитоплазматических белков с простой ренатурацией на колонке.
Последовательность ДНК (см. SEQ ID NO 1), кодирующая белок TβRII (4-136) аминокислотных остатков) (см. SEQ ID NO 2), синтезируют из 20 олигонуклеотидов (длиной 35-42 нуклеотидов каждый) с помощью полимеразной цепной реакции (ПЦР). Кодоны, имеющие низкую эффективность экспрессии в прокариотах, замененяют согласно таблице встречаемости прокариотических кодонов (см. SEQ ID NO 1). Амплифицированную последовательность ДНК расщепляют с помощью BglII и HindIII и клонируют в плазмиду рЕТ-32а по сайтам рестрикции BglII/HmdIII вслед за геном партнера по слиянию тиоредоксина, непосредственно после последовательности ДНК, кодирующей сайт расщепления энтеропептидазы (Asp)4-Lys. Последовательность гена подтверждают автоматическим секвенированием с помощью ABI Prism-310 Genetic Analyzer (Applied Biosystems, США).
Плазмидной ДНК pET-32a/TβRII трансформируют компетентные бактериальные клетки штамма Escherichia coli BL21(DE3). Штамм BL21(DE3) (Invitrogen) предназначен для экспрессии рекомбинантных белков. Обозначение DE3 означает, что штамм содержит лизоген фага λ DE3, который несет ген для Т7 РНК-полимеразы под контролем lacUV5-промотора. Для индукции экспрессии Т7 РНК-полимеразы необходим IPTG (изопропил_β-D-1-тиогалактопиранозид). Штамм не содержат lon и OmpT протеазы, что уменьшает деградацию экспрессированных в штамме разнородных белков. 100 мкл компетентные клетки штамма BL21(DE3) трансформируют с 15 нг плазмидной ДНК pET-32a/TβRII. Трансформированную культуру клеток наносят на чашку со средой LB (Лурия-Бертони), содержащей 100 мкг/мл ампициллина и растят при 37°C в течение 18 часов. Выросшие колонии бактерий являются штаммом-продуцентом BL21(DE3)/pET-32a/TβRII, так как только трансформированные клетки резистентны к ампициллину. Штамм Escherichia coli BL21(DE3)/pET-32a/TβRII обеспечивает конститутивный синтез белка тиоредоксин/TβRII, соответственно уровень экспрессии составляет не менее примерно 1 г из 1 л клеточной культуры. Выделение и очистку целевого белка проводят с помощью хроматографических методов, в частности аффинной хроматографии. Растворимый слитный белок очищают и ренатурируют на колонке с носителем Ni-NTA агароза. При ренатурации подбирают соотношение концентрации окисленного и восстановленного глутатиона как 1:50. Ренатурированный слитный белок тиоредоксин/TβRII расщепляют с помощью рекомбинантной каталитической субъединицы энтеропептидазы. Подбирают оптимальное соотношение белок-фермент как 1:40000. После расщепления целевой белок ТβRII отделяют от тиоредоксина на Ni-NTA агарозе. 140 мг высокоочищенного TβRII без N- и С-концевых тагов получают с 1 л клеточной культуры.
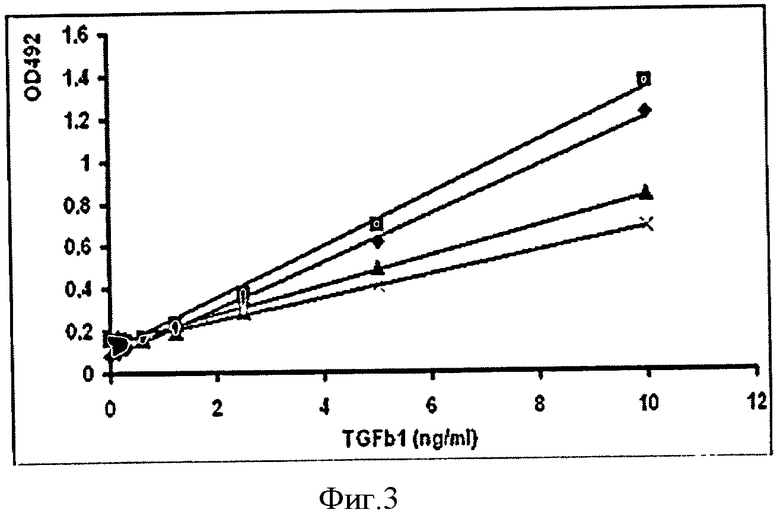
Для сравнительного анализа биологической активности полученного высокоочищенного препарата TβRII используют метод иммуноферментного анализа (enzyme-linked immunosorbent assay, ELISA) (фиг.3), измеряя связывание с лигандом TGF-β1. Сравнивают активность с коммерческим рекомбинантным препаратом TβRII (фирмы R&D Systems), полученным из клеточной линии миеломы мышей. Полученный нами высокоочищенный белок TβRII связывает TGF-β1 более высоким сродством, по сравнению с коммерческим препаратом.
Изобретение иллюстрируют чертежи.
Фиг.1. Генетическая конструкция для экспрессии слитного белка Trx/TβRII. С помощью рестриктаз HindIII и BglII синтетический ген целевого белка клонирован в плазмидную ДНК рЕТ-32а по сайтам рестрикции BglII/HindIII вслед за геном партнера по слиянию тиоредоксина, непосредственно после последовательности ДНК, кодирующей сайт расщепления энтеропептидазы (Asp)4-Lys.
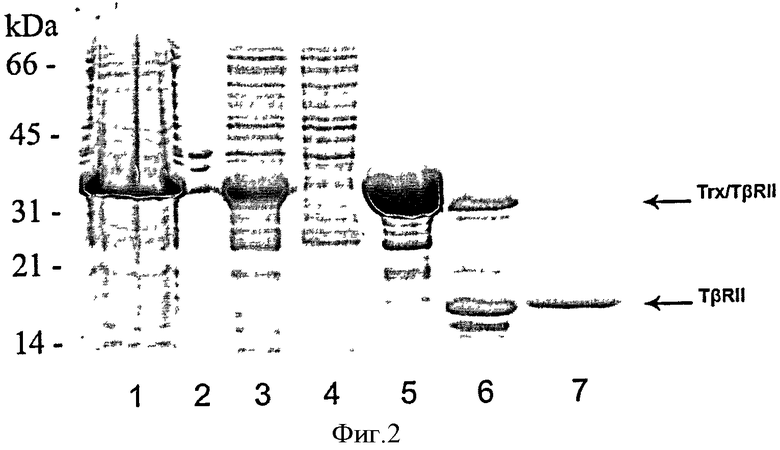
Фиг.2. Экспрессия и очистка рекомбинантного белка TβRII человека. Пробы, содержащие по 10 мкг белка, анализируют на 15% полиакриламидном геле в денатурирующих условиях (Ds-Na-ПААГ).
1 - Экспрессия Trx/TβRII в клетках Е.coli, 2 - фракция нерастворимых белков; 3 - фракция цитоплазматических белков; 4 - фракция белков, не связанных с Ni-NTA агарозой; 5 - слитный белок Trx/TβRII после очистки на Ni-NTA агарозе; 6 - препарат слитного белка Trx/TβRII после расщепления энтеропептидазой; 7 - целевой белок TβRII после очистки на Ni-NTA агарозе.
Фиг.3. Сравнительный анализ связывания TβRII с TGFβ1 с помощью иммуноферментного анализа: ■ - очищенный рекомбинантный TβRII 1 мг/мл; ♦ - коммерческий препарат TβRII 1 мг/мл; ▲ - очищенный рекомбинантный TβRII 0.5 мг/мл; × - коммерческий препарат TβRII 0.5 мг/мл.
Изобретение иллюстрируют примеры.
Пример 1.
Получение экспрессирующей конструкции pET-32a/TβRII.
Последовательность ДНК (см. SEQ ID NO 1), кодирующая белок TβRII (4-159 аминокислотных остатков) синтезируют из 20 олигонуклеотидов (длиной 35-42 нуклеотидов каждый) с помощью полимеразной цепной реакции (ПЦР). Для улучшения экспрессии гена в бактериях плохо экспрессируемые в прокариотических клетках эукариотические кодоны заменяют на соответствующие прокариотические. В ген добавляют сайт рестрикции для рестриктаз BglII, а также сайт расщепления энтеропептидазой на 3'-конце и сайт рестрикции для HindIII на 5'-конце. С помощью рестриктаз HindIII и BglII ген клонируют в экспрессирующую плазмиду рЕТ-32а по сайтам рестрикции BglII/HindIII вслед за геном тиоредоксина (фиг.1).
Пример 2.
Экспрессия и очистка белка TβRII.
Экспрессия слитного белка Trx/TβRII, находящегося в составе рекомбинантного плазмидного вектора рЕТ-32а, проводится в бактериальном штамме Escherichia coli BL21(DE3). Клетки трансформируют плазмидной ДНК pET32a/TβRII и клетки выращивают в жидкой питательной среде LB с ампициллином (100 мкг/мл) при 37°C и интенсивном перемешивании (230 об/мин) в течение 18 часов. После этого клетки инокулируют в жидкую среду ТВ (1 мл ночной культуры на 100 мл среды) с ампициллином (100 мкг/мл) и индуцируют экспрессию добавлением 0.02 мМ IPTG (изопропил_β-D-1-тиогалактопиранозид). Культуру клеток выращивают при 27°C в течение 21 часа. После этого клетки осаждают центрифугированием при 4000 g и 4°C в течение 15 минут, промывают в буфере, содержащем 300 мМ NaCl и 25 мМ NaH2PO4 (рН 7.4), снова осаждают и после удаления надосадка замораживают при -80°C для хранения.
Разрушение клеток проводят в буфере, содержащем 10 мМ имидазола, 300 мМ NaCl и 25 мМ NaH2PO4 (рН 8.0), продавливанием под давлением на установке «French press» (Spectronic Instruments, Inc., США). Разрушенную клеточную массу осаждают центрифугированием при 28000 об/мин в течение 45 мин. Растворимую фракцию цитоплазматических белков наносят на хроматографическую колонку Ni-NTA. Промывают колонку 3-4 объемами 35 мМ имидазола, после чего переносят промытую смолу со связанным белком в 200 мл буфер для ренатурации, содержащий 0.5 М L-аргинина, 60 мМ Tris-HCl (рН 8.0), 2 мМ восстановленного глутатиона, 0.04 мМ окисленного глутатиона. Инкубируют раствор в течение 3 суток при +4°C при постоянном медленном перемешивании на магнитной мешалке. После этого переносят раствор белка со смолой в пустую колонку и собирают проскок. Концентрируют белок до концетрации 1 мг/мл с помощью целлюлозной мембраны YM 30 в ячейке для концентрации под давлением газа азота и диализуют против буфера, содержащего 100 мМ NaCl и 80 мМ Трис, (рН 7.5) при 4°C в течение 18-20 часов. После диализа слитный белок Trx/TβRII расщепляют рекомбинантной легкой цепью человеческой энтеропептидазы при соотношении фермент-субстрат 1:100000 в течение 18 часов при комнатной температуре. После расщепления остаточную активность энтеропептидазы удаляют на колонке с STI (soybean trypsin inhibitor, соевый ингибитор трипсина)-агарозой. Белок TβRII отделяют от тиоредоксина на колонке с никелевой агарозой. На последней стадии препарат диализуют против буфера, содержащего 50 мМ фосфата натрия (рН 7.5) и 150 мМ NaCl, после чего стерилизуют через фильтр и хранят при 4°C для дальнейшего использования. N-концевую последовательность TβRII определяют после электрофореза в полиакриламидном геле и трансблотинга на мембрану поливинилидентифторида (PVDF).
На всех этапах очистки белок анализируют спектрофотометрически и с помощью электрофореза в 12% полиакриламидном геле. Результатом проделанной работы является получение высокоочищенного рекомбинантного лиганд-связывающего домена трансформирующего фактора роста-β человека TβRII (SEQ ID NO 2) (фиг.2).
Выход белка TβRII из 1 л культуры клеток составляет 140 мг (Табл.1).
(15.1 кДа)
Пример 3.
Тестирование биологической активности рекомбинантного препарата TβRII.
Для сравнительного анализа биологической активности полученного высокоочищенного препарата TRRII и коммерческого препаратов TβRII используют метод иммуноферментного анализа (enzyme-linked immunosorbent assay, ELISA) (фиг.3), измеряя связывание с трансформирующим фактором роста 1 (TGF-β1). На 96-луночную плату (Costar, USA) наносят рекомбинантный белок TβRII с концентрацией 1 мкг/мл в фосфатно-солевом буфере (по 100 мкл в лунку) и инкубируют в течение 16 часов при комнатной температуре. Трехкратно промывают лунки фосфатно-солевым буфером (PBS), содержащим 0.05% Tween-20. Далее плату инкубируют 1 час при комнатной температуре в буфере, содержащем 50 мМ Tris-HCl (рН 7.5), 100 мМ NaCl, 1.4% БСА (бычий сывороточный альбумин), 0.05% Tween-20 для блокирования мест неспецифического связывания белков. Затем плату однократно отмывают буфером для промывки и наносят различные количества (0.156, 0.312, 0.625, 1.25, 2.5, 5, 10, 100 нг/мл) коммерческого препарата рекомбинантного TGF-β1 (CHEMICON). Плату инкубируют при комнатной температуре в течение 2 часов с последующей трехкратной отмывкой. Затем наносят биотинилированные поликлональные антитела к TGF-β1 (R&D Systems) с концентрацией 1 мкг/мл по 100 мкл в лунку, инкубируют в течение 2 часов при комнатной температуре и отмывают. Для окрашивания добавляют по 100 мкл стрептавидина, конъюгированного с пероксидазой хрена, инкубируют в течение 20 минут при комнатной температуре. Повторяют процедуру отмывки. Взаимодействия антител с TGFβ1 проявляют с помощью 100 мкл субстратного буфера, содержащего 100 мМ цитрат натрия, 50 мМ фосфат натрия (рН 5.0), 0.5 мг/мл тетраметилбензидин и 0.05% H2O2, инкубируя в течение 30 минут при комнатной температуре. Для остановки реакции в каждую лунку добавляют по 50 мкл раствора 1М H2SO4. Реакцию детектируют на вертикальном спектрофотометре Multiscan (Labsystems, Финляндия) при длине волны 450 нм.


| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ВЫДЕЛЕНИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ | 2011 |
|
RU2499052C2 |
| Способ получения рекомбинантного противоопухолевого модифицированного белка DR5-B человека | 2018 |
|
RU2687435C1 |
| СПОСОБ ПОЛУЧЕНИЯ МУТАНТНОГО БЕЛКА TRAIL ЧЕЛОВЕКА | 2009 |
|
RU2405038C1 |
| ПЛАЗМИДНЫЙ ВЕКТОР pE-Trx-Aur, ШТАММ ESCHERICHIA COLI ДЛЯ ЭКСПРЕССИИ АНТИМИКРОБНОГО ПЕПТИДА АУРЕЛИНА И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО ПЕПТИДА | 2009 |
|
RU2412999C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PTRX-TEVRS-РТН, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК, СПОСОБНЫЙ К ПРОТЕОЛИТИЧЕСКОМУ РАСЩЕПЛЕНИЮ С ОБРАЗОВАНИЕМ ФРАГМЕНТА ЭНДОГЕННОГО ЧЕЛОВЕЧЕСКОГО ПАРАТИРЕОИДНОГО ГОРМОНА (1-34), ШТАММ ESCHERICHIA COLI BL21(DE3)/PTRX-TEVRS-РТН - ПРОДУЦЕНТ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО PTH (1-34) | 2019 |
|
RU2700452C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pЕ-Trx-Lc-def, ШТАММ Escherichia coli ДЛЯ ЭКСПРЕССИИ АНТИМИКРОБНОГО ПЕПТИДА ДЕФЕНСИНА ЧЕЧЕВИЦЫ Lens culinaris И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО ПЕПТИДА | 2010 |
|
RU2456345C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ ГИБРИДНЫЙ БЕЛОК L-HEP, ШТАММ ESCHERICHIA COLI ПРОДУЦЕНТ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА | 2019 |
|
RU2716975C1 |
| Способ очистки рекомбинантного белка, содержащего в своем составе последовательности миелопептидов | 2016 |
|
RU2630302C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pET22b(+)/slurp-2, КОДИРУЮЩАЯ БЕЛОК SLURP-2, И ШТАММ БАКТЕРИЙ Escherichia coli BL21(DE3) pET22b(+)/slurp-2- ПРОДУЦЕНТ БЕЛКА SLURP-2 ЧЕЛОВЕКА | 2015 |
|
RU2583307C2 |
| ПЛАЗМИДНЫЙ ВЕКТОР pE-Lc-LTP, ШТАММ БАКТЕРИИ Escherichia coli ДЛЯ ЭКСПРЕССИИ ЛИПИД-ТРАНСПОРТИРУЮЩИХ БЕЛКОВ ЧЕЧЕВИЦЫ Lens culinaris И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННЫХ БЕЛКОВ | 2009 |
|
RU2415940C1 |
Изобретение относится к биотехнологии. Описана рекомбинантная плазмидная ДНК pET-32a/TβRII. Плазмида состоит из BglII/HindIII-фрагмента ДНК плазмиды рЕТ-32а, содержащего lacI-промотор E.coli, T7 промотор, Trx·Tag кодирующую последовательность, His·Tag кодирующую последовательность, S·Tag кодирующую последовательность, T7 терминатор, lacI-кодирующую последовательность, bla-кодирующую последовательность, последовательность f1 участок ori инициации репликации, а также BglII/HindIII фрагмента гена, кодирующего ДНК синтетического гена TβRII. Представлен штамм бактерий Escherichia coli BL21 (DE3)/pET-32a/TβRII - продуцент слитного белка тиоредоксин/TβRII. Предложен способ выделения целевого белка TβRII человека. Растворимую фракцию цитоплазматических белков выделяют из штамма-продуцента Escherichia coli BL21(DE3)/pET-32a/TβRII. Проводят хроматографическую очистку с последующей ренатурацией слитного белка тиоредоксин/ TβRII. Далее белок инкубируют в течение 72 часов при 4°С в буфере, содержащем 0.5 М аргинина, в присутствии окисленного и восстановленного глутатиона в соотношении 1:50 мМ соответственно. Затем проводят диализ, расщепление каталитической субъединицы рекомбинантной энтеропептидазой и очистку целевого белка TβRII от тиоредоксина. Изобретение позволяет получать повышенный выход высокоочищенного TβRII белка человека с нативным N-концом (140 мг с одного литра культуры клеток). 3 н.п. ф-лы, 3 ил., 1 табл.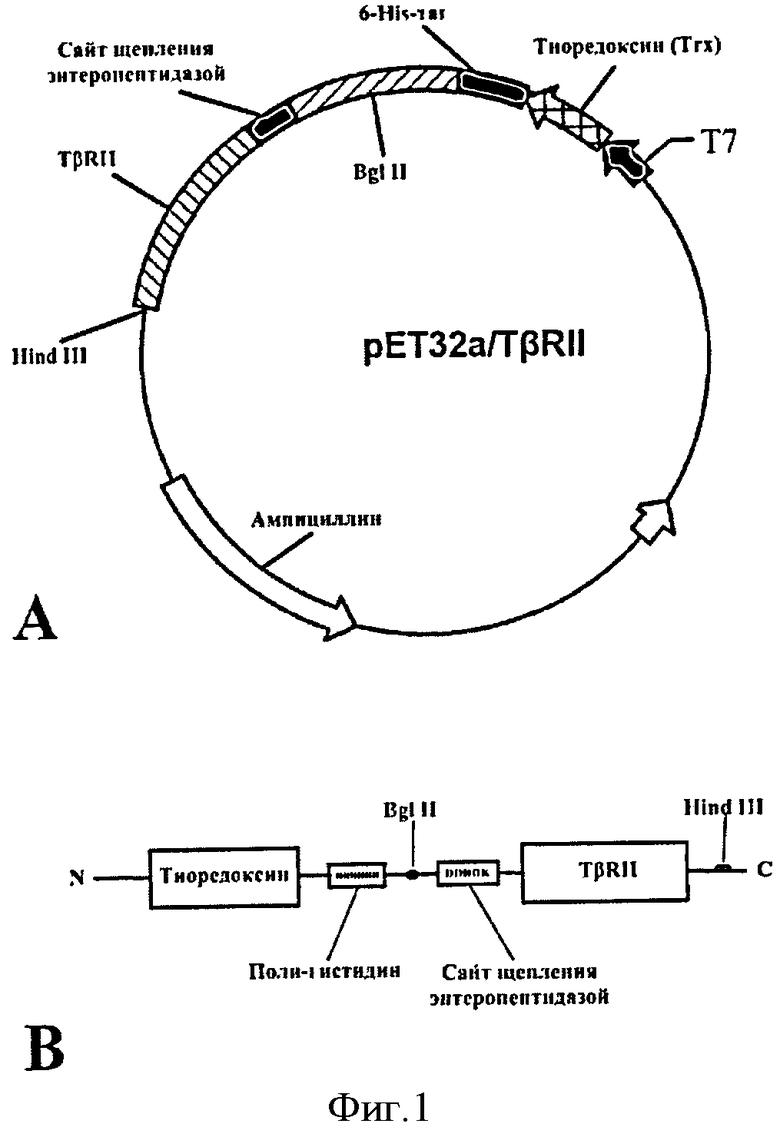
1. Рекомбинантная плазмидная ДНК pET-32a/TβRII, кодирующая полипептид лиганд-связывающего домена рецептора II типа трансформирующего фактора роста-β человека (6.237 т.п.о.), состоящая из BglII/HindIII - фрагмента ДНК плазмиды рЕТ-32а длиной 5.838 т.п.о., содержащего lacl-промотор E.coli, T7 промотор, Trx·Tag кодирующую последовательность, His·Tag кодирующую последовательность, S·Tag кодирующую последовательность, T7 терминатор, laсI - кодирующую последовательность, bla- (ген β-лактамазы) кодирующую последовательность, последовательность fl участок ori инициации репликации, а также BglII/HindIII фрагмента гена, кодирующего ДНК синтетического гена полипептид лиганд-связывающего домена рецептора II типа трансформирующего фактора роста-β человека (TβRII).
2. Штамм бактерий Escherichia coli BL21 (DE3)/pET-32a/TβRII - продуцент слитного белка тиоредоксин/TβRII.
3. Способ выделения целевого белка TβRII человека, включающий выделение растворимой фракции цитоплазматических белков из штамма-продуцента Escherichia coli BL21(DE3)/pET-32a/TβRII, хроматографическую очистку с последующими ренатурацией слитного белка тиоредоксин/ TβRII, инкубируя белок в течение 72 ч при 4°С в буфере, содержащем 0,5 М аргинина, в присутствии окисленного и восстановленного глутатиона в соотношении 1:50 мМ соответственно, диализом, расщеплением каталитической субъединицей рекомбинантной энтеропептидазой и очисткой целевого белка TβRII от тиоредоксина.
| Christian С | |||
| BOESEN et al | |||
| Development of a recombinant bacterial expression system for the active form of a human transforming growth factor b type II receptor ligand binding domain | |||
| Protein expression and purification | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| ПЕПТИД, СПОСОБНЫЙ СВЯЗЫВАТЬСЯ С TGF-β 1 И ИНГИБИРОВАТЬ БИОЛОГИЧЕСКУЮ АКТИВНОСТЬ TGF-β 1 IN VITRO И/ИЛИ IN VIVO, ПРИМЕНЕНИЕ ПЕПТИДА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, КОНСТРУКЦИЯ ДНК, ВЕКТОР ЭКСПРЕССИИ, КЛЕТКА-ХОЗЯИН И СПОСОБ ПОЛУЧЕНИЯ ПЕПТИДА | 2004 |
|
RU2333917C2 |

