ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к системам и способам, которые измеряют быстро затухающую Т2*-релаксацию, для эффективного определения количества меченых клеток при помощи магнитно-резонансного изображения. Открытые системы и способы полезны в различных применениях, включая транспорт клеток и клеточную терапию.
УРОВЕНЬ ТЕХНИКИ
Клеточная терапия, использующая стволовые клетки и иммунные клетки, с целью восстановления и реваскуляризации все более часто применяются в клинических испытаниях. Успех или неудача могут зависеть от точной доставки клеток к заданным органам. Определение количества клеток, доставленных к заданной ткани(тканям), имеет огромное значение для расчета оптимальной дозы и времени клеточной терапии. Чтобы метить клетки ex vivo, использовались агенты суперпарамагнитной окиси железа (SPIO), обеспечивающие исследователям возможность контролировать миграцию, пролиферацию и хоминг этих клеток с помощью магнитно-резонансного (МР) изображения. Мечение SPIO вызывает эффект релаксации c высокой скоростью (R2*), который возрастает линейно с возрастанием концентрации железа. R2* определяется как 1/Т2*.
Т2*-релаксометрия обычно достигается получением множества градиент-эхо изображений. В тканях, содержащих клетки, меченные железом с высокой концентрацией, Т2* может быть ультракороткой. В приведенных в качестве примера случаях Т2* находится ниже 1-2 миллисекунд, хотя точный период Т2* меняется от применения к применению. Время эхо для градиентного эхо как правило ограничивается настройками аппаратуры. На практике обычно ультракороткого времени эхо является недостижимым. Поэтому затухание сигнала в тканях с ультракоротким Т2* является как правило слишком быстрым для нормального градиент-эхо изображения.
Несмотря на все современные достижения, остается потребность в системах и/или способах, которые преодолеют трудности, связанные с традиционными техниками определения количества клеток. Кроме того, остается потребность в таких техниках определения количества клеток, которые не требуют модификации аппаратуры и/или специализированных расчетов РЧ импульса. Кроме того, нужны системы и способы для эффективного и надежного контроля и/или определения количества меченых клеток, например меченых стволовых клеток, в различных применениях, включая клеточную терапию и подобные. Эти и другие нужды удовлетворяются раскрытыми системами и способами.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение обеспечивает системы и способы для измерения и/или определения количества клеток в различных применениях, например транспорте клеток или клеточной терапии. Иллюстративные варианты осуществления раскрытых систем и способов включают использование клеток, которые были помечены ex vivo контрастирующим агентом, или других идентифицирующих характеристик. Меченые клетки контролируются с помощью МР изображения, чтобы оценить миграцию, пролиферацию и/или хоминг меченых клеток. Обычно контрастирующий агент представляет собой SPIO, хотя альтернативные контрастирующие агенты могут использоваться без отклонения от сущности и объема настоящего изобретения.
Согласно настоящему изобретению Т2*-релаксометрия преимущественно используется при измерении концентрации клеток в различных применениях, связанных с клетками. Поскольку в клетках, меченных железом с высокой концентрацией, Т2* является ультракороткой, в настоящем описании раскрыты предпочтительные системы и способы для измерения Т2*-релаксометрии, такие системы и способы используют последовательность спин-эхо изображений вместо стандартных градиент-эхо изображений для достижения желаемых результатов. В приведенных в качестве примера случаях Т2* не превышает 1-2 миллисекунд, хотя раскрытые системы и способы полезны в применении в широком диапазоне значений Т2*, такие значения Т2* как правило отличаются от применения к применению. Раскрытые системы и способы индуцируют нормальный спин-эхо сигнал, генерирующий первое спин-эхо изображение с последующим индуцированием множества спин-эхо сигналов, генерирующих серию дополнительных спин-эхо изображений от соответствующих смещений эхо, до вышеупомянутого затухания Т2* и затем получением схем Т2* при помощи подбора экспоненты.
Спин-эхо сигналы, выходящие из клеток, формируются для МР изображений первыми радиочастотными (РЧ) импульсами, за которыми следуют вторые радиочастотные импульсы, соответственно. При помощи спин-эхо сигналов строится кривая T2, где для клеток, меченных частицами/наночастицами SPIO, T2 гораздо длиннее, чем Т2*, и определяется через Msse-t/T. Кривая затухания Т2* спин-эхо затем определяется через Msse-TE/T2e-(t-TE)/T2*. Множество спин-эхо изображений берется на разных интервалах, определяемых шагом смещения эхо, который может быть меньше 1 мс. Схема ультракороткого Т2* строится по первому спин-эхо изображению и множеству спин-эхо изображений с подходящим смещением эхо при помощи подбора экспоненты. Полная схема Т2* строится при помощи наложения схемы ультракороткого Т2* на схему нормального Т2*.
Дополнительные свойства, функции и преимущества раскрытых систем и способов будут ясны из последующего описания, в частности при чтении с прилагаемыми чертежами.
КРАТКОЕ ОПСИАНИЕ ЧЕРТЕЖЕЙ
Чтобы помочь специалистам в данной области техники в создании и использовании раскрытых систем и способов, приведена ссылка на прилагаемые чертежи, на которых:
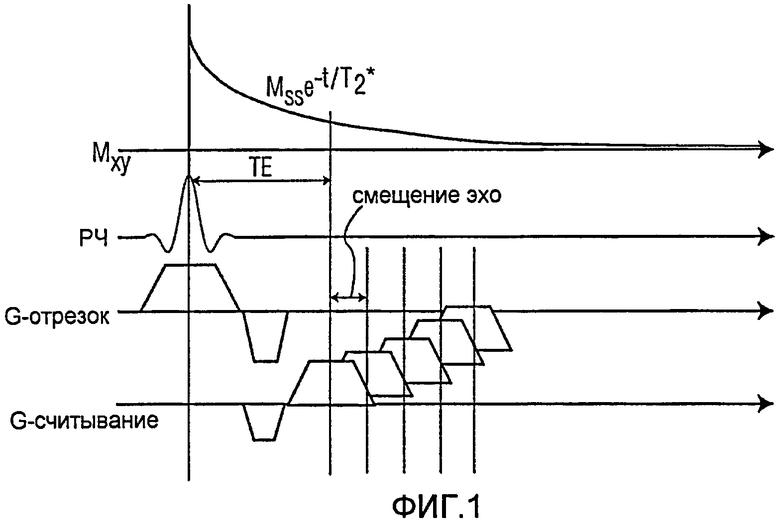
Фиг.1 представляет собой схематическую последовательность стандартной Т2* релаксометрии, получаемой при помощи многократного градиентного эхо;
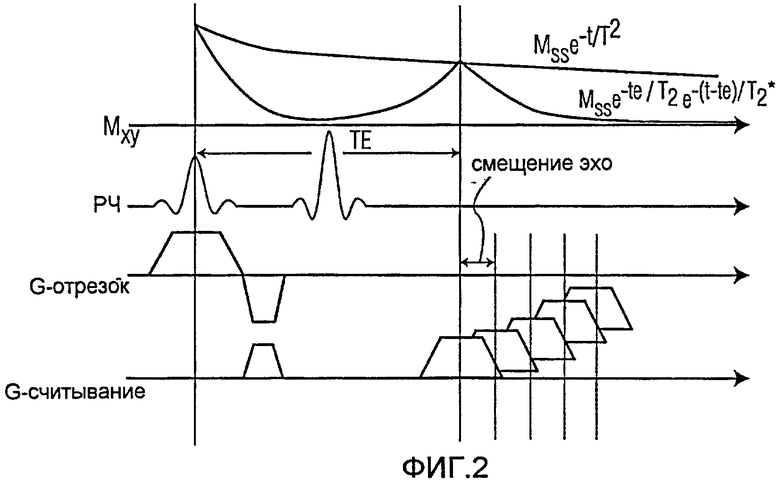
Фиг.2 представляет собой схематическую последовательность ультракороткой Т2* релаксометрии;
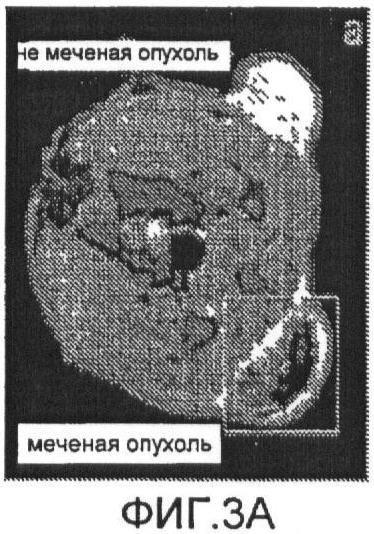
Фиг.3a представляет собой аксиальное градинет-эхо изображение крысы с опухолью;
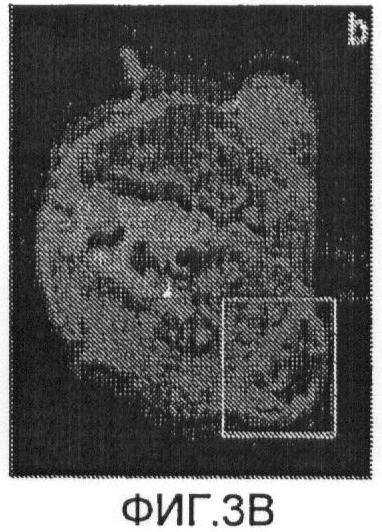
Фиг.3b представляет собой аксиальное спин-эхо изображение со смещением 0,8;
Фиг.3с представляет собой plussian blue-деформированный срез опухоли;
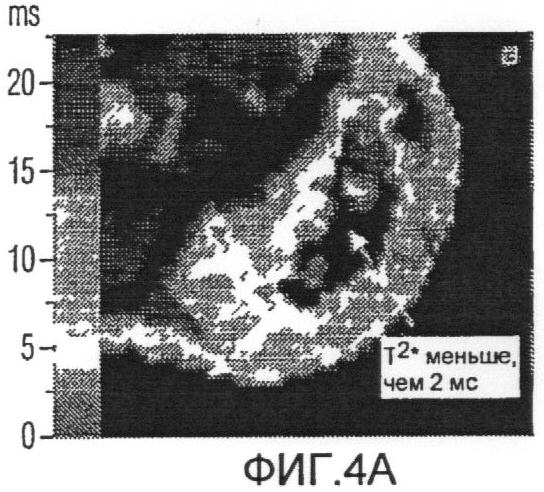
Фиг.4а представляет собой схему нормального Т2*, скрытого пороговым сигналом для устранения шума;
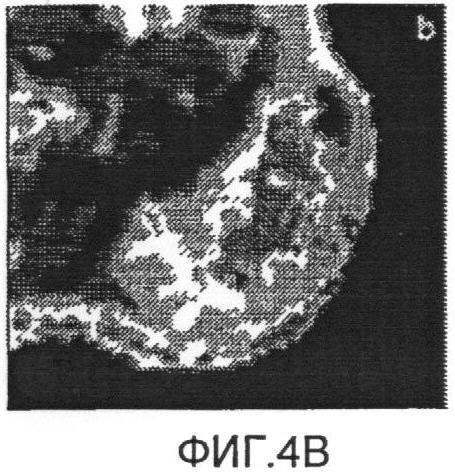
Фиг.4b представляет собой схему ультракороткого Т2*, наложенного на схему нормального Т2*;
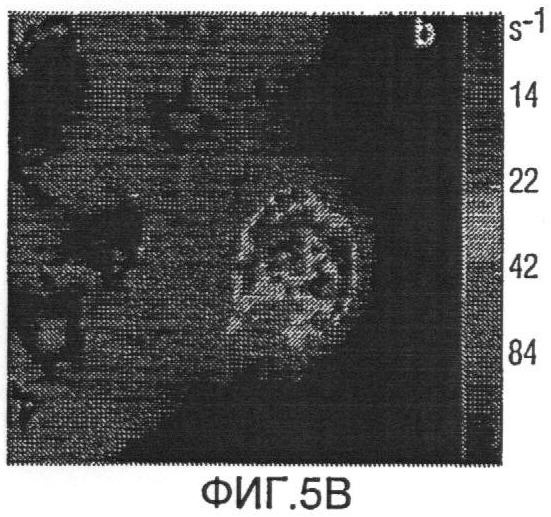
Фиг.5a представляет R2* схемы меченых боковых опухолей;
Фиг.5b представляет R2* схемы не меченых боковых опухолей;
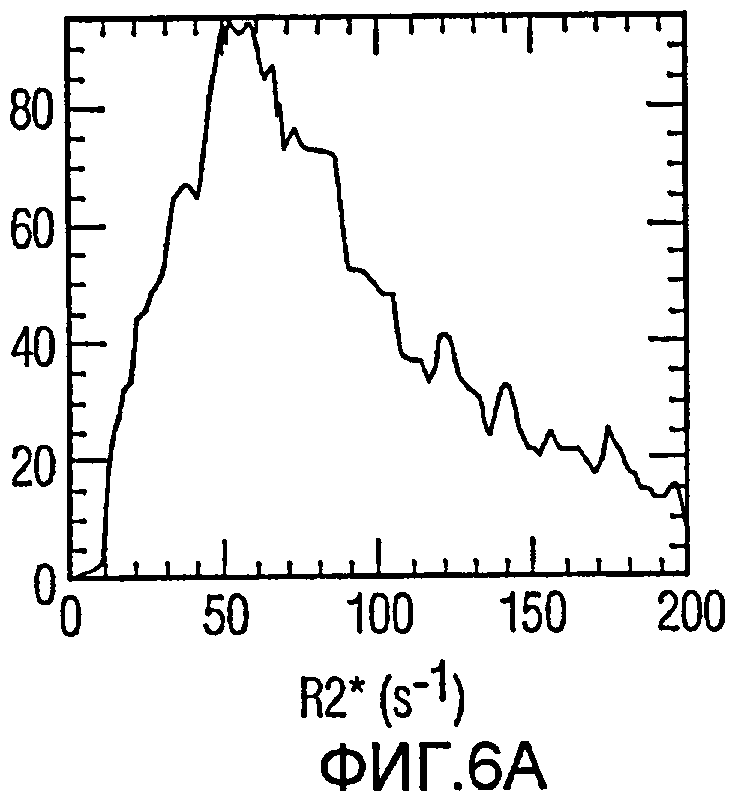
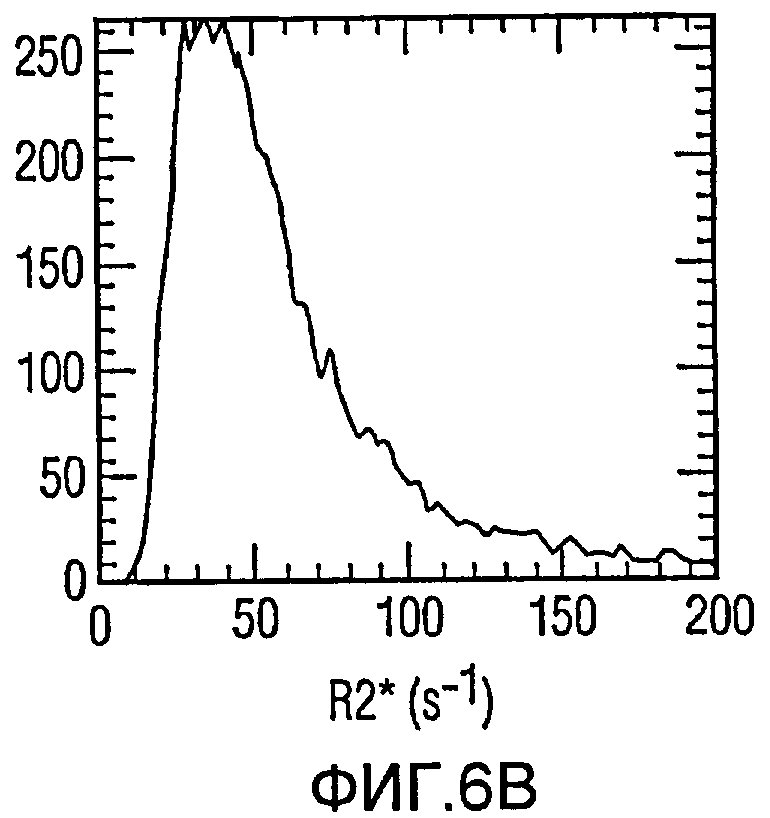
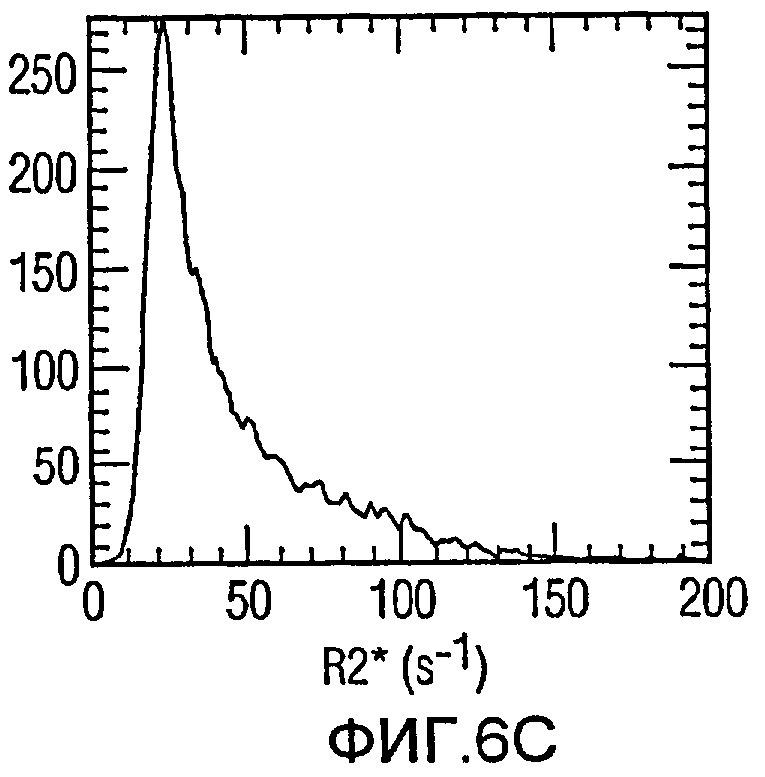
Фиг.6(а)-6(с) представляет собой гистограммы опухолей с разным количеством меченых железом клеток; и
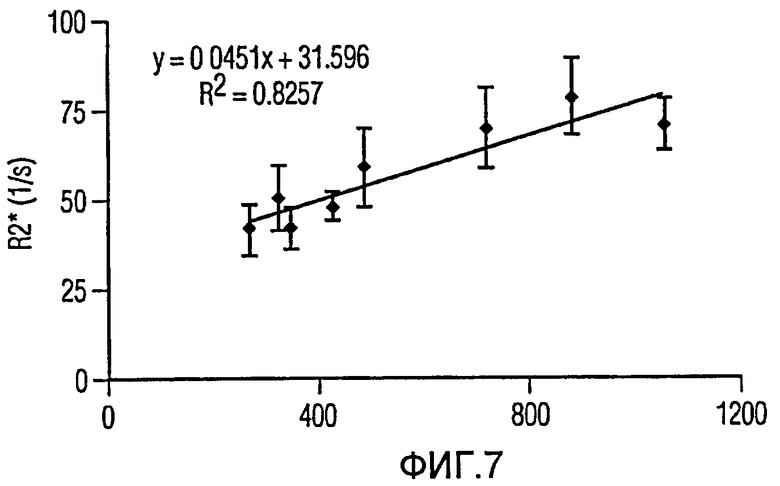
Фиг.7 представляет собой график, иллюстрирующий линейную корреляцию R2* с количеством клеток/мм3.
ОПИСАНИЕ ИЛЛЮСТРАТИВНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Раскрыты системы и способы для измерения и/или определения количества клеток, не требующие модификаций аппаратуры и/или разработок специализированного РЧ сигнала. Раскрытые системы/способы имеют широкий спектр применений, включающий применения для транспорта клеток и клеточной терапии. Меченые клетки контролируются при помощи генерации МР изображений для того, чтобы отследить их миграцию, пролиферацию и хоминг. Периоды быстро затухающей Т2* - релаксации измеряются путем генерации МР изображений, чтобы эффективно определить количество меченых клеток, как здесь описано.
Агенты SPIO влияют на период Т1, Т2 и Т2*-релаксации. Влияние SPIO в компартментах клетки на Т2*-релаксацию в десять раз больше, чем на Т2-релаксацию. В результате в клетках, меченых SPIO, Т2 намного длиннее, чем Т2*. Раскрытые системы и способы используют сравнительно длинное Т2 затухание с помощью получения серии спин-эхо изображений, чтобы преимущественно облегчить определение значения Т2*, несмотря на значительное ослабление сигнала, связанное с затуханием ультракороткого Т2*.
На Фиг.1 показана основная схема нормальной Т2*-релаксометрии, используя последовательность многократного градиентного эхо. Сигнал индуцируется РЧ импульсом с небольшим поворотом угла. Вслед за импульсом возбуждения применяется градиентное считывание, чтобы произвести эхо. Время между РЧ импульсом и серединой градиента считывания определяется как «ТЕ». Очевидно, что временной интервал ТЕ ограничивается РЧ импульсом и формой волны градиента для градиента выбора отрезков, и градиентом считывания. Таким образом, ТЕ ограничивается настройками аппаратуры, включая, в частности, силу градиента и время нарастания градиента.
Сигнал, полученный от градиентного эхо, определяется через Mss -TE/T2*, где Mss представляет собой намагниченность в устойчивом состоянии. В тканях, содержащих клетки, меченные железом с высокой концентрацией, Т2* может быть ниже 1-2 миллисекунд. Следовательно, сигнал может затухать до уровня шума со временем эхо в пару миллисекунд. Приложенные ранее усилия по уменьшению ТЕ включали модификацию оборудования или большой объем работ над разработкой последовательности, и не смогли стать оптимальными или полезными для множества традиционных применений.
На Фиг.2 схематично изображены различные параметры, связанные с иллюстративным вариантом осуществления настоящего изобретения. Согласно раскрытым системам и способам, спин-эхо используется, чтобы получить изображение. Использование спин-эхо заменяет традиционное использование градиентного эхо. В иллюстративном варианте осуществления настоящего изобретения, спин-эхо формируется 90-градусным РЧ сигналом, за которым следует 180-градусный РЧ сигнал. Интенсивность сигнала за ТЕ определяется отношением Mss -TE/T2. Поскольку в клетках, меченных SPIO, Т2 гораздо длиннее, сигнал, полученный от спин-эхо гораздо сильнее сигнала, полученного от градиентного эхо, что помогает избежать негативных последствий, связанных с большой потерей сигнала на изображении. Схема ультракороткой Т2*-релаксации может затем быть наложена на схему нормального Т2* для создания конечной Т2*-схемы в зоне обзора.
Измерение ультракороткой Т2*-релаксации может быть достигнуто получением серий спин-эхо изображений, как показано на Фиг.2. Первое эхо получается как нормальное спин-эхо изображение. Следующие изображения получаются путем смещения считывания по направлению к кривой затухания Т2* используя шаги, подходящие для смещения эхо, которые могут быть ниже 1 миллисекунды. Этот способ позволяет получить образец кривой затухания Т2*, построенной по спин-эхо сигналу. Затем могут быть получены схемы Т2* путем подбора экспоненты.
Обращаясь к Фиг.2, получают серию изображений с последовательностью спин-эхо. Первое сканирование получается в виде стандартного спин-эхо изображения. Последующие сканирования (сканирование 2- сканирование 5) получаются со смещением эхо по кривой затухания Т2*, определяемой отношением: Msse-TE/T2e-(t-TE)/T2*. Как показано на фиг.2, раскрытые системы и способы эффективны в преодолении ограничений, связанных с быстрым затуханием Т2*, путем успешного применения спин-эхо.
Для дальнейшей иллюстрации применений и преимуществ, связанных с раскрытыми системами и способами, сделана ссылка на следующие примеры. Однако нужно понимать, что данные примеры не являются ограничивающими объем настоящего изобретения, а только демонстрирующими иллюстративные варианты осуществления и/или выгоды изобретения:
Пример 1:
Чтобы облегчить измерение быстро затухающей Т2*-релаксации в тканях, содержащих клетки, меченные железом с высокой концентрацией, где затухание Т2* слишком быстрое для составления нормальной схемы многократного градиентного эхо, использовалась следующая методика. МР эксперименты in vivo на крысах с опухолями, меченными железом, использовались для демонстрации того, что раскрытая методика может быть использована для количественной оценки ультракороткой Т2*, не превышающей 1-2 миллисекунд или ниже. В сочетании с составлением схемы регулярной Т2*, раскрытая техника может быть использована, чтобы обеспечить in vivo количественную оценку и контроль тканей, содержащих клетки, меченные большим количеством железа.
Наночастицы SPIO широко используются, чтобы влиять на периоды релаксации T1, Т2 и Т2* меченых тканей и клеток. Период релаксации Т2* представляет собой наиболее чувствительный параметр для обнаружения клеток, меченных SPIO, и, основываясь на преимуществах раскрытых в настоящем описании систем и способов, Т2*-релаксометрия может быть успешно использована для количественной оценки и контроля меченных стволовых клеток в клеточной терапии. Как упоминалось выше, Т2*-релаксометрию как правило получают построением множества градиент-эхо изображений. Однако в тканях, содержащих клетки, меченные железом с высокой концентрацией, Т2* может быть ниже 2 миллисекунд, и, следовательно, затухание сигнала происходит слишком быстро для периода нормального градиентного эхо. Используя преимущества сравнительно долгого затухания Т2 клеток, связанных со SPIO, раскрытая система/способ используется для измерения быстро затухающей Т2*-релаксации при помощи серии спин-эхо изображений. В иллюстративном примере, была проведена количественная оценка in vivo короткого Т2* на крысах с опухолями, меченными железом.
Последовательность действий: измерение ультракороткой Т2* было достигнуто с помощью получения серий спин-эхо изображений, как показано на Фиг.2. Первое эхо было получено в виде нормального спин-эхо изображения. Следующие изображения были получены при помощи смещения считывания по направлению к кривой Т2*, используя шаги, не превышающие 1 миллисекунду. Это позволило получить кривую затухания Т2* из спин-эхо сигнала.
Эксперимент in vivo: клетки меланомы С8161 были помечены (Feridex)-протамин сульфатными (FEPro) комплексами с помощью процедур мечения, которые известны в данной области техники. 2х106 меченых FEPro или немеченых (контрольных) клеток меланомы были билатерально подкожно имплантированы в бок 5 голых крыс. Приблизительно через две недели после заражения опухолевыми клетками выполнялось ОМР на 3Т Intera whole-body scaner (Philips Medical System) при помощи специализированной 7-сантиметровой РЧ-катушки в форме соленоида для крыс. Схема нормальной Т2* была получена последовательностью многократного градиентного эхо (MGES) [TR/TE = 1540/16 мс, 13 эхо, матрица 256×256, 17 отрезков, толщина отрезков = 1,0 мм, зона обзора= 80 мм, NEX = 4]. Чтобы измерить короткую Т2*, были получены пять наборов спин-эхо изображений с шагом считывания эхо 0 мс, 0,4 мс, 0,8 мс, 1,2 мс, 2,3 мс соответственно, со следующими параметрами: TR/TE = 1000/6,4, матрица 144×144, 17 отрезков, толщина отрезков = 1,5 мм, зона обзора = 80 мм, NEX = 4.
Анализ данных: анализ данных выполнялся при помощи инструментов программного обеспечения IDL. Схемы Т2* были получены при помощи подбора экспоненты. Оба набора данных (то есть схема нормальной Т2* и схема короткой Т2*) были совмещены и показаны как схема Т2*.
Схема ультракороткой Т2*-релаксометрии и схема стандартной MGES Т2* были получены для 4 крыс. На Фиг.3а показано аксиальное градиент-эхо изображение от боковых опухолей крысы. Отсутствие сигнала в меченых опухолях было вызвано клетками, меченными железом с высокой концентрацией, как показано на Фиг.3с. Однако спин-эхо изображение той же опухоли (Фиг.3b) меньше подвержено затуханию сигнала, обусловленному сравнительно долгим временем Т2-релаксации клеток, связанных со SPIO. Схема Т2*, измеренная при помощи MGES (Фиг.4а), показывает площадь высоких значений Т2* на границе опухоли, показывающих последовательное ослабление FEPro-мечения по мере роста опухоли. На схеме MGES T2* нельзя обнаружить никакого сигнала вследствие быстрого затухания T2*, вызванного клетками, меченными железом с высокой концентрацией, в центре опухоли. Для сравнения, схема ультракороткой T2* (Фиг.4b) демонстрирует значения Т2* в центре опухоли, cоставляющие приблизительно ≤1 мс, что соответствует областям клеток, меченных железом с высокой концентрацией, согласно Фиг.3а.
Заключение: Этот эксперимент демонстрирует эффективное измерение времени ультракороткой T2*-релаксации в клетках и тканях. МР эксперименты in vivo демонстрируют, что этим способом можно измерять значения ультракороткого T2* вплоть до 1 мс или ниже в клетках, меченных железом с высокой концентрацией. В сочетании с традиционной схемой Т2*, раскрытая техника может использоваться для обеспечения количественной оценки и контроля тканей, содержащих клетки, меченные большим количеством железа, in vivo.
Эксперимент 2
Оценка количества меченых стволовых клеток в заданных тканях в экспериментальных моделях чрезвычайно важна, чтобы подобрать оптимальную дозу и время для клеточной терапии в клинических испытаниях. Агенты SPIO используются для мечения клеток, чтобы контролировать их миграцию, пролиферацию и хоминг при помощи МР изображения. Скорость релаксации R2* (1/T2*) является чувствительным параметром для количественного определения внутриклеточных SPIO.
В этом иллюстративном примере было изучено количественное отношение между числом меченных железом клеток и скоростью релаксации R2* у крысы с опухолью. Более конкретно, было изучено количественное соотношение между меченными железом клетками и скоростью тканевой релаксации R2* у крысы с опухолью. Эксперименты in vivo показали хорошую линейную корреляцию между количеством меченных железом клеток и тканевой R2*. В дальнейшем данные показали, что измерение R2* является надежным и чувствительным способом определения количества меченых клеток in vivo.
Клети меланомы С8161 и клетки глиомы С6 были помечены (Feridex)-протамин сульфатными (FEPro) комплексами с помощью известных процедур. 2×106 меченных FEPro или немеченых (контрольных) клеток меланомы (n=4) или 1×106 меченных FEPro или немеченых клеток глиомы (n=4) были билатерально подкожно имплантированы голым крысам. Приблизительно через две недели после заражения опухолевыми клетками выполнялось ОМР на 3Т Intera whole-body scaner (Philips Medical System) при помощи специализированной 7-сантиметровой РЧ-катушки в форме соленоида для крыс. Схема нормальной R2* была получена последовательностью многократного градиентного эхо (MGES) [TR/TE = 1540/16 мс, 13 эхо, матрица 256×256, 17 отрезков, толщина отрезков = 1,0 мм, зона обзора = 80 мм, NEX = 4]. Чтобы измерить скорость релаксации R2* в тканях с высокой концентрацией меченых клеток, были получены пять наборов спин-эхо изображений с шагом считывания эхо 0 мс, 0,4 мс, 0,8 мс, 1,2 мс, 2,3 мс соответственно, со следующими параметрами: TR/TE = 1000/6,4, матрица 144×144, 17 отрезков, толщина отрезков = 1,5 мм, зона обзора = 80 мм, NEX = 4. Скорость релаксации R2* была подсчитана путем подбора экспоненты при помощи инструментов программного обеспечения IDL. Оба набора данных (то есть схема регулярной R2* и схема тканевой R2* с высокой концентрацией меченых клеток) были совмещены. R2*-релаксация опухоли была подсчитана по пикселям как среднее от R2*-релаксации по всему объему опухоли.
Результаты: Мечение железом не изменило рост опухоли. Не было статистически значимой разницы в размерах опухоли меченых и немеченых опухолей. Размеры меченых опухолей находились в диапазоне от 1890 мм3 до 4950 мм3 на время построения изображения, что означает от 325 до 1056 меченых клеток на мм3 в восьми опухолях.
Мечение FEPro значимо удлинило скорость релаксации R2* в опухоли. Фиг.5a и 5b иллюстрируют схемы R2* меченой и немеченой опухоли соответственно. Влияние мечения железом на R2*-релаксацию может быть также доказано при помощи R2*-гистограммы опухоли с 1056 мечеными клетками/мм3 (Фиг.6а) и 325 мечеными клетками/мм3 (Фиг.6b). Меченые опухоли показали гораздо большее распространение R2* по сравнению с контрольной опухолью (Фиг.6с). Средняя R2* опухоли демонстрирует очень хорошую линейную корреляцию с количеством меченых клеток на мм3 (Фиг.7), с коэффициентом корреляции 0,91 (р<0,01).
Заключение: в этом иллюстративном примере было исследовано количественное соотношение между меченными железом клетками и скоростью тканевой релаксации R2*. Несмотря на то, что были использованы две различных линии опухолевых клеток, данные in vivo демонстрируют хорошую линейную корреляцию между количеством меченных железом клеток и тканевой R2*. Экспериментальные данные также показывают, что измерение R2* представляет собой надежный и чувствительный инструмент для количественной оценки меченных железом клеток. Согласно этому раскрытые системы и способы могут использоваться для эффективной неинвазивной количественной оценки меченных железом клеток in vivo.
В целом, системы и способы настоящего изобретения предлагают значительно улучшенные техники для МР измерений меченых клеток в различных применениях. Действительно, современные исследования в области транспорта клеток и клеточной терапии начинаются с инъекций большого количества клеток, меченных SPIO, в заданный участок, что приводит к очень короткому Т2* в меченых и окружающих клетках. Открытые системы и способы способствуют значительному улучшению в количественной оценке меченых клеток, несмотря на ультракороткое затухание Т2*, с которым приходится сталкиваться в таких тканевых системах. Раскрытые системы и способы могут также применяться для измерения ультракороткой Т2* других контрастирующих агентов, которые вызывают значимую разницу между Т2 и Т2*-релаксацией.
Хотя настоящее изобретение было описано со ссылкой на иллюстративные варианты осуществления и их реализации, раскрытые системы и способы не ограничены такими иллюстративными вариантами осуществления. Напротив, как будет видно специалистам в данной области техники из приведенного здесь описания, раскрытые системы и способы допускают модификации, изменения и усовершенствования без отклонения от сущности и объема настоящего изобретения. Следовательно, настоящее изобретение определенно охватывает такие модификации, изменения и усовершенствования в пределах его объема.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПИН-ЭХО МР-ВИЗУАЛИЗАЦИЯ | 2015 |
|
RU2702843C2 |
| КОНТРАСТНОЕ СРЕДСТВО ДЛЯ T И/ИЛИ T МАГНИТНО-РЕЗОНАНСНОГО СКАНИРОВАНИЯ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2011 |
|
RU2471502C1 |
| Способ количественной оценки степени перегрузки железом печени у детей | 2018 |
|
RU2701771C1 |
| МР ТОМОГРАФИЯ С НУЛЕВОЙ ЭХО-ЗАДЕРЖКОЙ | 2015 |
|
RU2686879C2 |
| СПОСОБ НЕИНВАЗИВНОГО МОНИТОРИНГА БИОДЕГРАДАЦИИ ПОЛИМЕРНОГО СКАФФОЛДА | 2024 |
|
RU2832295C1 |
| ПАРАЛЛЕЛЬНАЯ МР-ТОМОГРАФИЯ С КАРТИРОВАНИЕМ ЧУВСТВИТЕЛЬНОСТИ С ПОМОЩЬЮ РЧ-КАТУШКИ | 2016 |
|
RU2683605C1 |
| ДИНАМИЧЕСКАЯ КОНТРАСТНАЯ УЛУЧШЕННАЯ МР ВИЗУАЛИЗАЦИЯ С РЕКОНСТРУКЦИЕЙ СЖАТОГО ИЗМЕРЕНИЯ | 2011 |
|
RU2557334C2 |
| СИСТЕМЫ И СПОСОБЫ ОЦЕНКИ ПЕРЕНОСА ГАЗА В ЛЕГКИХ, ИСПОЛЬЗУЯ MPT С ГИПЕРПОЛЯРИЗОВАННЫМ Xe | 2007 |
|
RU2452372C2 |
| МАГНИТНО-РЕЗОНАНСНАЯ ВИЗУАЛИЗАЦИЯ С ПОДАВЛЕНИЕМ АРТЕФАКТОВ ПОТОКА | 2012 |
|
RU2605524C2 |
| ЯМР СПЕКТРОСКОПИЯ С ИСПОЛЬЗОВАНИЕМ СРЕДСТВА ЯМР С ГРАДИЕНТНЫМ ПОЛЕМ | 2003 |
|
RU2251097C2 |


Группа изобретений относится к медицине, способам и системам магнитно-резонансного (МР) измерения ультракороткой Т2*-релаксации (Р), использующим получение спин-эхо, для контроля количества, миграции, пролиферации и/или хоминга клеток, меченных большим количеством железа, при транспорте клеток, клеточной терапии, в том числе in vivo. Генерируют последовательность МР изображений и измеряют Т2*-Р с кривой затухания Т2*, полученной серией спин-эхо изображений, включая этапы: индуцирования первого спин-эхо сигнала (СЭС), генерирующего первое спин-эхо изображение, и множества СЭС, генерирующих серию дополнительных спин-эхо изображений из подходящих смещений затухания Т2*; получения Т2* изображения при помощи подбора экспоненты. При этом первый СЭС и второй СЭС образуются при помощи первого радиочастотного импульса (РЧИ), за которым следует второй РЧИ, отличный от первого. СЭС индуцируется РЧИ с небольшим углом наклона. Формируют эхо, используя градиентное считывание. В тканях, содержащих клетки, меченные железом с высокой концентрацией, при Т2 ниже 1 мсек сигнал может затухать до уровня шума со временем эхо 2 мсек. Поскольку Т2 гораздо длиннее в клетках, меченных суперпарамагнитной окисью железа, и сигнал, полученный с помощью спин-эхо, гораздо сильнее сигнала, полученного с помощью градиентного эхо, данный способ и система для его осуществления позволяют избежать негативных эффектов, связанных со значительной потерей сигнала на изображении, используя сравнительно длинное Т2 затухание при получении серии спин-эхо изображений, облегчая определение Т2*, несмотря на значительное ослабление сигнала, связанное с затуханием ультракороткого Т2*, тем самым обеспечивают улучшение количественной оценки меченых клеток; позволяют измерять ультракороткую Т2* других контрастирующих агентов, вызывающих значительную разницу между Т2 и Т2*-Р. 2 н. и 10 з.п. ф-лы, 7 ил.
1. Способ контроля меченых клеток, включающий в себя:
мечение клеток ex vivo контрастирующим агентом;
контроль миграции, пролиферации и/или хоминга упомянутых меченных клеток при помощи генерации последовательности магнитно-резонансных (МР) изображений;
измерение Т2*-релаксометрии с кривой затухания Т2*, полученной серией спин-эхо изображений, включая этапы, на которых:
(a) индуцируют первый спин-эхо сигнал, генерирующий первое спин-эхо изображение;
(b) индуцируют множество спин-эхо сигналов, генерирующее серию дополнительных спин-эхо изображений из подходящих смещений указанного затухания Т2*; и
(c) получают Т2* изображения при помощи подбора экспоненты,
при этом упомянутый первый спин-эхо сигнал и упомянутый второй спин-эхо сигнал образуются при помощи первого радиочастотного (РЧ) импульса, за которым соответственно следует второй РЧ импульс, причем первый РЧ импульс отличается от второго РЧ импульса.
2. Способ по п.1, в котором упомянутый контрастирующий агент представляет собой суперпарамагнитную окись железа (SPIO).
3. Способ по п.1, в котором Т2* является ультракоротким.
4. Способ по п.3, в котором Т2* изменяется от применения к применению, и в некоторых применениях меньше или равно 2 мс.
5. Способ по п.1, в котором упомянутый первый РЧ импульс представляет собой 90° РЧ импульс, а второй РЧ импульс представляет собой 180° РЧ импульс.
6. Способ по п.1, в котором кривая затухания Т2 определяется через Мssе-t/T, где Mss представляет собой намагниченность в устойчивом состоянии.
7. Способ по п.1, в котором упомянутая кривая затухания Т2* определяется через: Msse-TE/T2e-(t-TE)/T2, где Mss представляет собой намагниченность в устойчивом состоянии.
8. Способ по п.1, в котором упомянутое подходящее смещение эхо выполняют с шагами, не превышающими 1 или 2 мс.
9. Способ по п.1, в котором упомянутые Т2* изображения совмещены и показаны в виде общего Т2* изображения.
10. Способ по любому одному из предшествующих пунктов, в котором меченые клетки измеряют применительно к транспорту клеток или клеточной терапии.
11. Система для контроля меченых клеток, содержащая:
средство для мечения клеток ex vivo контрастирующим агентом;
средство для контроля миграции, пролиферации и/или хоминга упомянутых меченых клеток при помощи генерации последовательности магнитно-резонансных (МР) изображений;
средство для измерения Т2*-релаксометрии с кривой затухания Т2*, полученной серией спин-эхо изображений, включая этапы, на которых:
(a) индуцируют первый спин-эхо сигнал, генерирующий первое спин-эхо изображение;
(b) индуцируют множество спин-эхо сигналов, генерирующее серию дополнительных спин-эхо изображений из подходящих смещений указанного затухания Т2*; и
(c) получают Т2* изображения при помощи подбора экспоненты,
при этом упомянутый первый спин-эхо сигнал и упомянутый второй спин-эхо сигнал образуются при помощи первого радиочастотного (РЧ) импульса, за которым соответственно следует второй РЧ импульс, причем первый РЧ импульс отличается от второго РЧ импульса.
12. Система по п.11, в которой первый РЧ импульс представляет собой 90° РЧ импульс, а второй РЧ импульс представляет собой 180° РЧ импульс.
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ НАСЫЩЕНИЯ ГЕМОГЛОБИНА КРОВИ КИСЛОРОДОМ | 1993 |
|
RU2070325C1 |
| СРЕДСТВО КОНТРАСТИРОВАНИЯ ПРИ МАГНИТНО-РЕЗОНАНСНОМ ТОМОГРАФИЧЕСКОМ ИССЛЕДОВАНИИ | 1992 |
|
RU2087157C1 |
| Способ измерения времен Т @ продольной ядерной магнитной релаксации | 1988 |
|
SU1578608A1 |
| Heike E | |||
| Daldrup-Link et al | |||
| Targeting of Hematopoietic Progenitor Cells with MR Contrast Agents // Radiology. | |||

