Изобретение относится к сельскохозяйственной биотехнологии и представляет собой новый штамм эктомикоризообразующего гриба Suillus sibiricus, способного увеличить приживаемость и стимулировать рост сеянцев и саженцев сосны сибирской.
Функционирование одного из важнейших технологических секторов экономики - "Переработка и воспроизводство лесных ресурсов" базируется на объеме лесного фонда той или иной страны. Основная часть бореальных лесов Евразии расположена на территории бывшего Советского Союза - Восточной Европы и Сибири. Главные лесообразующие породы этих лесов - сосновые, еловые, пихтовые, распространены на площади 220 млн га от Балтики до Тихого океана. В целом по Сибири сосна занимает площадь 72582,2 тыс. га; кедр - 36167,1 тыс. га; пихта - 11772,8 тыс. га; лиственница - 213219,0 тыс. га. Интенсивная эксплуатация лесов, главным образом хвойных, ухудшила возрастную структуру лесного фонда, сократила площади, занимаемые ценными сосновыми, кедровыми и еловыми лесами, вызвала снижение внутривидового разнообразия, исчезновение ценных генотипов. Помимо больших объемов вырубок и ветровалов, лесной фонд России истощается также в результате поражения вредителями и пожарами. Например, в Красноярском крае в 2004 г. вспышка массового размножения сибирского шелкопряда уничтожила 1 млн га хвойного леса, лесные пожары только в Богучанском районе этого края уничтожили более 200 тыс. га ценнейших сосновых лесов.
Анализ состояния лесных ресурсов, например, в Западной и Восточной Сибири показывает неблагоприятные тенденции: наблюдается смена пород, исчезают наиболее продуктивные древостои, на значительных площадях семенные насаждения замещаются на леса вегетативного происхождения, происходит увеличение доли искусственных лесонасаждений, наблюдается массовое усыхание отдельных видов хвойных, исчезновение устойчивых клонов. Лесовосстановительные работы в основном включают в себя выделение плюсовых деревьев и насаждений, закладку постоянных лесосеменных плантаций и клоновых архивных участков.
В связи с большой экономической и экологической ролью насаждений хвойных важной становится задача воспроизводства генетических ресурсов главных лесообразующих видов. По данным 2000-2007 гг., в среднем на территории Российской Федерации ежегодно, для возобновления лесных ресурсов необходимо осуществлять посадку темнохвойного леса на территории 598,7 тысяч га, (на территории Томской области - 500 тысяч га). Для этого необходимо вырастить посадочный материал, среднее количество которого для посадки составляет 5-10 тысяч на 1 га. По нормативам выход стандартного посадочного материала в лесопитомниках составляет 800000 на 1 га для кедра (сосны сибирской) и 1-1,2 млн на 1 га для ели, сосны обыкновенной, лиственницы, пихты (для всей территории РФ - 598000 × на 1 млн саженцев). Однако примерно 30-40% таких саженцев не достигает стандартных размеров из-за отсутствия в технологии выращивания сеянцев такого этапа работы, как микоризация [1-5]. Именно по этой причине два основных показателя качества посадочного материала хвойных - высота саженцев и диаметр корневой шейки, оказываются меньше необходимых по ГОСТу.
В настоящее время основной проблемой лесных хозяйств по сохранению генетических ресурсов лесных популяций в России и в мире является разработка технологии ускоренного размножения хвойных для возобновления генетических и семенных заказников [6-8]. На данный момент для репродукции лесных пород используется старая технология посадки в естественных насаждениях с помощью меча Колесова, в лесопитомниках для микоризации растений применяют варварский и неэкологичный способ - ручное разбрасывание снятой с поверхности лесной почвы, либо вообще не проводят микоризацию. Одной из основных тенденций развития данного технологического сектора является переход на систему неистощительного природопользования, что требует применения новых микробных биотехнологий в современной практике искусственного лесовосстановления. В этом смысле современным считается способ выращивания саженцев с закрытой корневой системой, при этом самый важный момент репродукции хвойных представляет деятельность интегрированных растительно-микробных систем.
Настоящим бедствием в лесопитомниках являются эпифитотии инфекционных заболеваний сеянцев, что вызвано нарушением гомеостаза сложившейся экосистемы из-за введения новых агротехнических приемов, монокультуры, смешения ареала культивируемых деревьев и появления новых патогенов. Систематическое применение пестицидов в питомниках изменяет видовой состав микробных ценозов, вследствие чего исчезают многие полезные формы микроорганизмов. Появляются вакантные ниши, которые занимают случайные популяции, что ведет к смене нежелательного грибного компонента в микоризе, обеднению аборигенной микрофлоры, увеличению численности фитопатогенных микроорганизмов и усилению их вирулентных свойств, что ухудшает фитосанитарное состояние питомников. Все это можно предотвратить с помощью мероприятий по сохранению сложившегося биологического равновесия с помощью экологически безопасных микробов-антагонистов [8-12]. В этой связи во всем мире продолжаются работы по поиску полезных форм микроорганизмов, способных длительное время функционировать в искусственной экосистеме, ограничивать рост популяций фитопатогенных бактерий и микромицетов, стимулировать развитие сеянцев и саженцев.
Известно, что для нормального развития любого вида деревьев необходимы специализированные штаммы грибов-макромицетов, образующих эктомикоризу [17-20]. Интерес к феномену симбиоза корней растений с грибами-макромицетами не угасает с описания Франком в 1885 году практически всех типов эктомикориз - ЕсМ. [13; 19]. Известно, что 8000 видов высших растений и 7000-10000 видов грибов планеты образуют ЕсМ, которая рассматривается как реципрокный мутуализм/паразитизм, зависящий от генетических и экологических факторов [21]. Обязательная микотрофность хвойных как основных лесообразующих пород фитоценозов бореальной зоны обусловлена участием эктомикоризных грибов в круговороте биогенных элементов, а также способностью ЕсМ оптимизировать метаболизм растений, усиливать минеральное питание и устойчивость к засухе, засолению, тяжелым металлам, патогенам [11]. В случае мутуалистических отношений ЕсМ-грибы получают от растений от 10 до 50% органического углерода и становятся конкурентнособными в почве, а растения имеют возможность пользоваться коммуникационной подземной сетью из мицелия ЕсМ-грибов и корневых систем разных видов деревьев, по которой передвигаются метаболиты, источники энергии, кофакторы, витамины, гормоны, токсины и, возможно, генетическая информация.
Таким образом, взаимодействующим с окружающей средой компонентом симбиосистемы в эктомикоризах является гриб, который выполняет роль посредника в обменных процессах между растением и почвой. Это мнение признано и не подвергается значительному сомнению исследователями разного профиля: лесоведами, микологами, экологами. Убеждение хорошо соответствует современному уровню знаний о поглощающих органах деревьев, микоризных грибах и микоризах и подтверждается большим объемом фактических данных о закономерностях строения [1; 3; 21]. Таким образом, основная роль столь необходимой для каждого вида дерева микоризы сводится к следующему. Во-первых, увеличивается всасывающая поверхность корней; во-вторых, образуются многие биологически активные вещества, используемые растениями; в-третьих, трудноусваиваемые соединения фосфора и алюмосиликатов почвы переводятся в растворимую форму, доступную растениям; в-четвертых, обеспечивается иммунитет растения к заражению потенциальными почвообитающими паразитами; в-пятых, мицелий микоризных грибов, выходящий из корней разных экземпляров растений в почву, сливается, и по нему мигрируют от одного растения к другому метаболиты, включая питательные вещества, источники энергии, гормоны, токсины, а возможно, и генетическая информация. При этом происходит интеграция популяций и даже разновидовых сообществ растений в единую гигантскую коммуникационную подземную сеть из мицелия грибов-микоризообразователей.
Изучение экологии и физиологии ЕсМ сосредоточено в основном в Европе, Северной Америке и Австралии [13]. В России только в конце и середине прошлого века различными аспектами ЕсМ занимались многие исследователи: Возняковская Ю.М., Худяков Я.П., Шубин В.И., Чумак Н.Ф., Ахромейко А.И., Лобанов Н.В., Гельцер Ф.Ю., Ванин С.И., Зерова М.Я., Селиванов И.А., Еропкин К.И., Шемаханова Н.М., Красовская И.В., Частухин В.Я., Саляев Р.К., Иванова Р.Н., Коротков Г.П., Семенова Л.А., Мишустин Е.Н., Пушкинская О.И., Каратыгин И.В. и другие. В настоящее время ЕсМ изучает Шубин В.И. - в Карелии, Коваленко А.Е., Иванов Д.М. - в Санкт-Петербурге, Воронина Е.Ю. - в Москве, Творожникова Т.А. - в Сыктывкаре, Веселкин Д.В. - в Екатеринбурге, Шкараба Е.М. и Бойко Т.А. - в Перми. Насколько нам известно, на территории Сибири различные виды ЕсМ хвойных сейчас изучает только Д.В. Веселкин (граница Восточного Урала и Западной Сибири), в конце XX века эстонскими учеными была проведена работа по определению ЕсМ в Новосибирской и Иркутской областях, но только образуемых крошечными грибами рода Tomentella, которые плохо культивируются in vitro [22]. Биохимический состав культуральных сред грибов-макромицетов в настоящее время изучается только с точки зрения их использования для фунготерапевтических целей.
В 2004 г. Томском госуниверситете были получены первые практические результаты по систематической идентификации видов ЕСМ-грибов и введению в культуру некоторых видов грибов рода Amanita, Suillus и Cortinarius, Boletus, образующих эктомикоризу с корнями хвойных, и предложена технология ее использования для ускоренного размножения хвойных [22-24]. В дополнение к классическим анатомо-морфологическим описаниям ЕсМ-грибов, томские ученые провели изучение физиологических и биохимических характеристик чистых культур, несмотря на то, что это искусственное и стрессовое состояние для грибов-микоризообразователей [25]. Были введены в культуру Amanita muscaria (L.: Fr) Hook; Amanita pantherina (Fr.) Seer.; Amanita porphyria (Alb. & Schw. ex Fr.); Boletus betulicola (Vasil'kov) Pilat & Dermek; Boletus pinophyllus (Pilat & Dermek); Cortinarius caninus (Fr.) Fr.; Suillus bovinus (Linnaeus: Fr.) O. Kuntze; Suillus luteus (Fries) S.F.Gray, Nat.; Suillus sibiricus (Singer) Singer; Suillus variegatus (Sw.) Kuntze; Tricholoma portentosum (Fr.) Quel, изучены особенности роста на различных средах, ДНК-идентификация и определены содержания глюкозы, белка, фитогормонов и флавоноидов в культуральной жидкости.
На мировом рынке присутствуют 5 компаний, производящих препараты на основе микоризы: Mykoplant AG, Rootgrow (США), Mycosym (Швейцария), INOQ (Германия), Simbiom (Чехия). В России производятся препараты Мицефит и Симбионт, не содержащие микоризу, а только вытяжку из нее.
При изготовлении препаратов для стимуляции развития древесных пород, как правило, это споры грибов и бактерий, смешивают с основой - глиной или другим сорбентом. Основным недостатком всех этих сухих форм препаратов является трудность и непредсказуемость прорастания гиф грибов из спор. Для сокращения сроков укоренения и приживаемости черенков в искусственном субстрате используются, как правило, искусственные стимуляторы роста растений, требующие многочасового выдерживания черенков в растворе.
Наиболее близким к предлагаемому является штамм Suillus sibiricus (Singer, 1938) который был получен из гриба Suillus sibiricus произрастающего в Бурятии, где нет кедровых лесов. Зарегистрирован и сдан на хранение в Коллекцию культур базидиомицетов LE(BIN) Учреждения Российской академии наук Ботанического института им. В.Л.Комарова РАН с присвоением регистрационного №LE(BIN) 1086 в 1938 году. В литературных источниках не обнаружено какой-либо информации о свойствах этого штамма и его использовании. Можно предположить, что в настоящее время свойства штамма изменились ввиду того, что во время хранения, штамм необходимо пересевать, в результате чего, через три года он теряет свою биологическую активность.
При создании изобретения ставилась задача - получение нового штамма эктомикоризообразующего гриба, способного увеличить приживаемость и стимулировать рост сеянцев и саженцев сосны сибирской.
Указанная задача решается новым штаммом эктомикоризообразующего гриба Suillus sibiricus.
Полученный штамм был принят на хранение в Коллекцию культур базидиомицетов LE(BIN) Учреждения Российской академии наук Ботанического института им. В.Л.Комарова РАН с присвоением регистрационного №LE(BIN)2178 19.04.2007.
Полученный штамм выделен из плодового тела Suillus sibiricus в чистую культуру традиционным методом.
Описание культуры Suillus sibiricus (Singer) Singer штамм для депонирования в коллекции Ботанического Института им. В.Л. Комарова РАН (Санкт-Петербург)
Вид базидиомицета - Suillus sibiricus (Singer) Singer
1. Его номер в Вашей коллекции: №6.
2. Где: Россия, Томск, Томская область, п.Старое Лоскутово, припоселковый кедрач (описание см. ниже).
3. Когда: 27 августа 2006 года.
4. Кем: доц. Биологического Института Томского госуниверситета О.Б.Вайшля; н.с. сектора общей микробиологии ОСП «НИИ биологии и биофизики Томского госуниверситета» Ведерниковой А.А.
5. Как был получен изолят: На месте сбора плодовое тело надрезали в нижней части ножки вдоль волокон прокаленным скальпелем и затем расщепляли в направлении разреза вместе со шляпкой. Из участка перехода ножки в шляпку вырезали кусочки трамы длиной несколько миллиметров и помещали на агаризованную питательную среду.
6. Подробный состав среды, на которую он был выделен: сусло-агар; картофельный агар.
7. Среда, на которой вы его храните: Среда Худякова-Возняковской.
8. Кто определял исходное плодовое тело: н.с. ОСП «НИИ биологии и биофизики Томского госуниверситета» Агафонова Н.Н.
9. Где (в каком гербарии) хранится ваучерный образец, если он был сохранен: Коллекционный материал находится в научном фондохранилище грибов музейного типа ОСП «НИИ биологии и биофизики Томского госуниверситета».
10. Географические координаты сборов (если есть): координаты определены с помощью GPS: 56° 23' 37.3'' северной широты; 085° 08' 34.1'' восточной долготы.
Штамм Suillus sibiricus характеризуется следующими культурально-морфологическими и физико-биохимическими признаками.
Культурально-морфологические. У молодой недельной культуры, выращенной с использованием твердой питательной среды мицелий имеет белый цвет, далее по мере культивирования мицелий приобретает розовато-кремовую окраску и иногда выделяет пигментированные экссудаты. Колонии ватные, воздушный мицелий высокий, отдельные мицелиальные нити переплетены во всех направлениях. При микроскопировании видны множественные пряжки. При выращивании данного штамма на различных питательных средах его колонии имеют различный цвет, форму и структуру мицелия.
Для штамма, выращиваемого на жидкой питательной среде, характерен рост в виде обрывков гиф или мицелиальных конгломератов неправильной или шаровидной формы, от которых отходят разной длины одиночные гифы. Рост мицелия происходит по периферии колоний. По мере роста культуры происходит изменение цвета и консистенции среды от светло желтого жидкого до коричневого вязкого
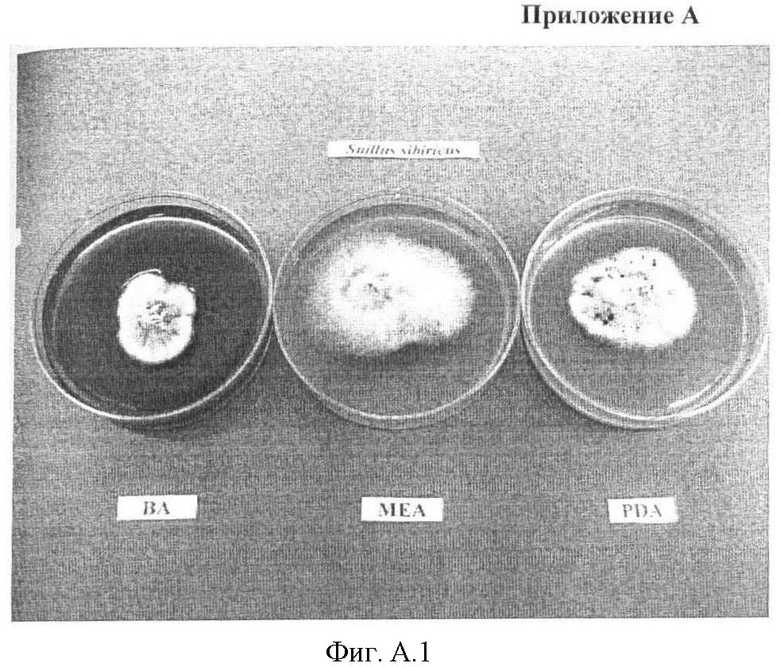
Приложение А. Фиг А.1 - Внешний вид колоний гриба Suillus sibiricus, выращенного на твердых питательных средах: ВА - голодный агар, МЕА - мальц-экстракт, РDА - картофельно-декстрозный агар.
При этом логарифмическая кривая роста, измеренная по уменьшению содержания глюкозы или по увеличению концентрации водорастворимого белка в культуральной жидкости, имеет классическую S-образную форму
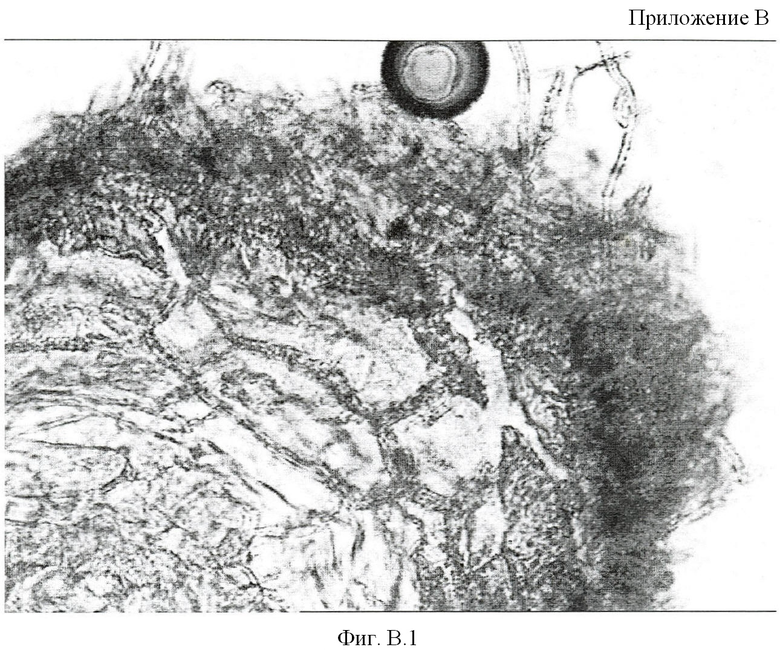
Приложение Б. Микрофотография среза микоризованного среза сосны сибирской, Фиг В.1 - Поперечный срез корня сосны сибирской, микоризованной культурой макромицета Suillus sibiricus, увеличение 15×40. Видна сеть Гартига, образованная мицелием гриба и пряжки на грифах.
Изобретение поясняется следующим конкретным примером получения штамма Suillus sibiricus.
Пример выращивания на твердой питательной среде:
Для масштабирования гриба проводят пересев из исходной пробирки с чистой матричной культурой на чашки Петри со стерильной агаризованной питательной средой. На питательную среду обожженным в пламени горелки копьем помещают кусочек агара с мицелием размером 1 см × 1 см; 1 см × 2 см. Засеянные чашки Петри выдерживают в термостате при температуре плюс 24°С в течение месяца. Готовыми к использованию считаются культуры, образовавшие густой воздушный мицелий, занимающий не менее 80% площади поверхности питательной среды. У молодой недельной культуры мицелий имеет белый цвет, далее по мере культивирования мицелий приобретает розовато-кремовую окраску и иногда выделяет пигментированные экссудаты. Колонии ватные, воздушный мицелий высокий, отдельные мицелиальные нити переплетены во всех направлениях. При микроскопировании видны множественные пряжки. При выращивании данного штамма на различных питательных средах его колонии имеют различный цвет, форму и структуру мицелия.
Приложение А. ФигА.2 - Поверхностный рост мицелия гриба Suillus sibiricus на жидкой питательной среде Худяковой-Вознесенской.
Для выращивания в жидкой питательной среде используют модифицированную среду Худякова-Возняковской:
Состав среды Худякова-Возняковской в расчете на 1 литр [26]:
глюкоза - 15 г;
NH4Cl - 0,6 г;
гидролизованный казеин - 5,0 мл;
КН2РО4 - 1,0 г;
MgSO4 · 7Н2O - 0,5 г;
NaCl - 0,1 г;
СаСl2 - 0,1 г;
FeCl3 1,0 мкг;
дрожжевой автолизат - 0,2 г; агар - 15 г;
дистиллированная вода - 1000 мл, pH (4,0-5,0).
Стерилизация 30 мин при 0,5 атм.
Микроэлементы для питательной среды в расчете на 1 литр:
CuSO4 - 0,23 мг;
Na2MoO4 - 0,043 мг;
ZnSO4 - 4,3 мг;
Na2B4O7 - 0,045 мг;
МnСl2 - 0,046 мг.
Витамины для питательной среды в расчете на 1 литр (добавлять после плавления агара):
фолиевая кислота - 500 мкг;
пиридоксин - 500 мкг;
тиамин - 500 мкг;
никотиновая кислота - 500 мкг;
рибофлавин - 500 мкг;
селенат натрия 570 мкг.
Жидкую культуру грибов выращивают на модифицированной питательной среде Худякова-Возняковской [26] с помощью термостатируемого шейкера при режиме работы 180-220 оборотов в минуту, температуре плюс 25-28 градусов по Цельсию в течение 14-16 суток. Соотношение объема питательной среды к объему качалочной колбы составляет 1:10. Затравка для масштабирования биомассы - 1-2% инокулята от объема среды.
Для штамма выращиваемого на жидкой питательной среде характерен рост в виде обрывков гиф или мицелиальных конгломератов неправильной или шаровидной формы, от которых отходят разной длины одиночные гифы. Рост мицелия происходит по периферии колоний. По мере роста культуры происходит изменение цвета и консистенции среды от светло желтого жидкого до коричневого вязкого (Приложение А). При этом логарифмическая кривая роста, измеренная по уменьшению содержания глюкозы или по увеличению концентрации водорастворимого белка в культуральной жидкости, имеет классическую S-образную форму.
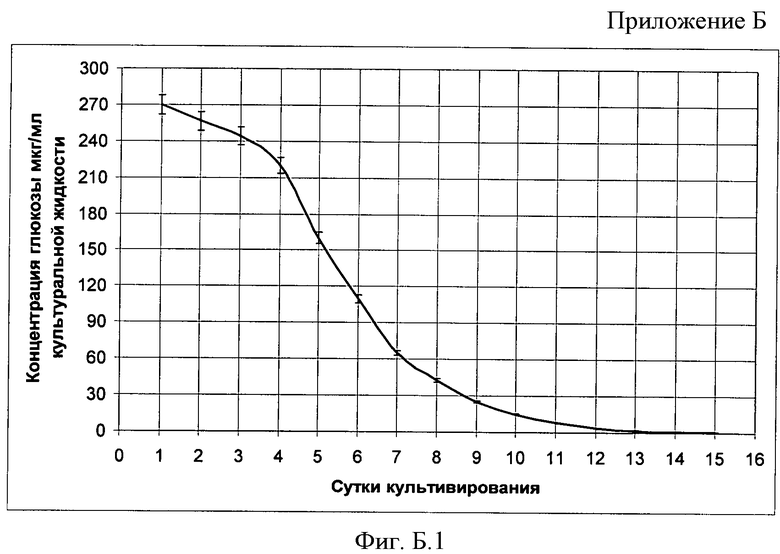
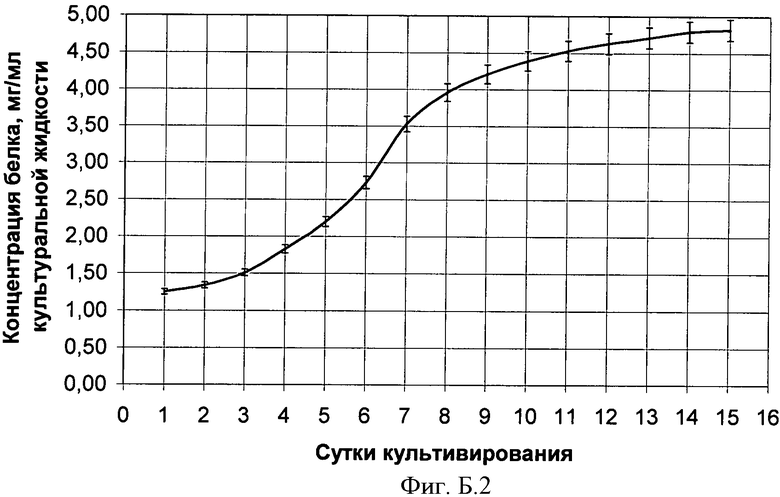
В Приложении Б представлена динамика роста штамма в культуральной жидкости, на Фиг Б.1 - Изменение концентрации глюкозы в процессе культивирования in vitro эктомикоризообразующего гриба Suillus sibiricus, на Фиг Б.2 - Изменение концентрации водорастворимого белка в процессе культивирования in vitro эктомикоризообразующего гриба Suillus sibiricus.
Полученный таким образом штамм имеет специфические пряжки в структуре гиф и характеризуется повышенной способностью образовывать активную сеть Гартига в составе микоризы с корнями сосны сибирской (Приложение В).
Далее приведены результаты исследований - изучения способности данного штамма Suillus sibiricus эфективно увеличивать приживаемость хвойных.
В качестве грибов, способных образовывать эктомикоризу с корнями хвойных пород деревьев, использовались следующие макромицеты: масленок сибирский (Suillus sibiricus), козляк (Suillus bovinus).
В Таблице 1 приведены данные физиолого-биохимической характеристики культуры эктомикоризообразующего Suillus sibiricus, параметры, описывающие кривую роста. Изменение содержания глюкозы и водорастворимого белка при глубинном культивировании микоризообразующих грибов Suillus sibiricus и Suillus bovinus. На основании скрининга отбирались культуры, имеющие эти параметры, поскольку только эти культуры обладали способностью продуцировать гормоны-стимуляторы роста растений и выделять флавоноиды, сигнальные вещества, позволяющие грибу колонизировать растение и образовывать с ним симбиоз.
В Таблице 2 приведены данные содержания индивидуальных цитокининов, индолилуксусной кислоты и флавоноидов в 10-дневной культуре Suillus sibiricus.
Мониторинг эффекта микоризации на физиологическое состояние сеянцев сосны сибирской
Проверку эффективности микоризации данным штаммом проводили на сеянцах и саженцах сосны сибирской в условиях защищенного и открытого грунта. Для микоризации сеянцев использовали методику, разработанную В.Я.Частухиным. Внесение микоризы проводилось двумя способами: 1) культуральной жидкостью гриба (гелевая форма); 2) кусочками твердой питательной среды с мицелием гриба (твердая форма). Мицелий вносился в почву, в районе междурядий, на глубину 5-10 см. Степень микоризации определяли по индексу микоризации, одновременно проводя микроскопический контроль с помощью микроскопа A-plan (Carl Zeiss).
Основными показателями физиолого-биохимического благополучия микоризованных растений считаются такие параметры, как высота растений и содержание фотосинтетических пигментов - хлорофиллов и каротиноидов.
Пигментный фонд. Показано, что после микоризации в хвое сеянцев увеличился уровень каротиноидов. В Таблице. 3 представлены данные пигментного фонда хвои сеянцев сосны сибирской после 45 дней микоризации корней культурой гриба Suillus sibiricus.
Наиболее сильно это проявляется при микоризации сосны штаммом, выращенным на основе сусло-агара. Микоризованные растения через два месяца после проведения опыта показали, что уровень каротиноидов в зеленой части у сеянцев сосны сибирской, инокулированных культурой гриба, выращенной на сусло-агаре, вырос в 1,7 раза и в 2,9 раза при микоризации почвенным препаратом.
Морозоустойчивость
Одной из причин гибели сеянцев хвойных при лесовосстановлении являются морозобоины - наружные продольные трещина ствола. Обусловливаются неодинаковым сжатием наружных и внутренних слоев древесины при заморозках за счет накопления воды в древесине. Известно, что грибной компонент микоризы синтезирует такие криопротекторные вещества, как трегалоза, что препятствует образованию центров кристаллизации воды и последующему за этим разрушению клеток. Индекс микоризации (ИМ) - соотношение суммы микоризных окончаний к сумме всех поглощающих корней корневой системы растения. 23-50%.
Таблица 5 - Индекс микоризации и устойчивость сеянцев сосны сибирской к низким температурам после инокуляции штаммом Suillus sibiricus.
Результаты таблицы 5 показывают, что у микоризованных растений сосны сибирской, вследствие того, что ЕСМ-грибы выделяют в область ризосферы сеянцев фитогормоны-стимуляторы роста, происходит более раннее одревеснение стволиков и, соответственно, число морозобоин после отрицательных температур уменьшается. При этом индекс микоризации на таких сеянцах почти в два-три раза выше, чем в контроле. Все это свидетельствует о криопротекторном эффекте микоризации сеянцев, при котором короткие молекулы углеводной природы препятствуют образованию центров кристаллизации внутри клеток.
Скорость роста контрольных и микоризованных сеянцев
В физиологии лесных культур скорость роста изучают по наиболее интегральному показателю высоты растений. В каждом варианте измеряли не менее 30-50 сеянцев сосны горной сосны обыкновенной и ели колючей, результаты приведены в таблицах 8-11 и свидетельствуют о достоверности обнаруженных различий к концу летнего сезона [17-18]. При этом микоризованные растения достоверно обгоняют по скорости роста контрольные растения, независимо от того, гелевой или твердой формой препарата они были обработаны и где выращены - в открытом или закрытом грунте.
В Таблице 5 представлены данные о высоте сеянцев сосны сибирской при выращивании в условиях открытого грунта, мм
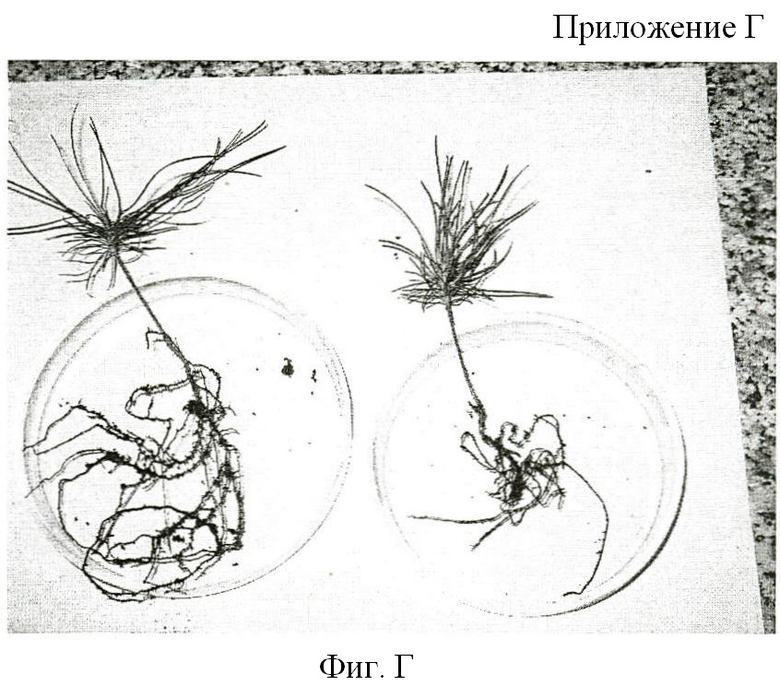
На фигуре Приложения Г, отчетливо видна разница в степени развития корневой системы контрольных и микоризованных сеянцев сосны сибирской.
Таким образом, предлагаемый штамм гриба-макромицета, образующий эктомикоризу с корнями сосны сибирской и другими хвойными породами деревьев, проявляет высокую степень микоризации и стимулирующего действиия на рост сеянцев вследствие того, что он обладает способностью синтезировать гормоны-стимуляторы роста растений и флавоноиды, способствующие образованию прочного симбиоза корней сосны сибирской с мицелием Suillus sibiricus. Штамм гриба вносится только один раз, полностью исключает необходимость предварительной обработки посадочного материала и предназначен для образования долгосрочного симбиоза корневой системы хвойных с микроорганизмами почвы, что даст возможность ускорения воспроизводства и круглогодичной пересадки саженцев на любом этапе развития. Штамм перспективен для получения препарата с помощью которого возможно преодоление трудностей, связанных с гетеровегетативным размножением и пересадкой ценнейших пород хвойных деревьев, с увеличением приживаемости хвойных на безлесных территориях и урбанизированных землях, на обедненных почвах в ландшафтно-озеленительных работах, при укреплении дамб и озеленении угольных карьеров.
Источники информации
1. Веселкин Д.В. Реакция эктомикориз Pinus sylvestris L. на техногенное загрязнение различных типов // Сиб. экол. журн. 2005. №4. С.753-761.
2. Каратыгин И.В. Коэволюция грибов и растений. СПб: Гидрометеоиздат, 1993. 115 с.
3. Селиванов И.А. Микосимбиотрофизм как форма консортивных связей в растительном покрове Советского Союза. М.: Наука, 1981. 232 с.
4. Чумак Н.Ф. Микоризы, образуемые грибами рода Suillus у сосны обыкновенной // Микология и фитопатология. 1981. Т.15. Вып.3. С.202-207.
5. Шемаханова Н.М. Микотрофия древесных пород. М.: Изд-во АН СССР, 1962. 374 с.
6. Chalot М., Javelle A., Blaudez D., Lambilliote R., Cooke R., Sentenac H., Wipf D., Botton B. An update on nutrient transport processes in ectomycorrhizas // Plant and Soil. 2002. Vol.244. №1-2. P.165-175.
7. Cornelissen J.H.C., Aert R., Cerabolini В., WergerM.J.A., Heijden van der M.G.A. Carbon cycling traits of plant species are linked with mycorrhizal strategy // Oecologia. 2001. Vol.129. №4. P.611-619.
8. Harley J.L., Smith S.E. Mycorrhizal symbiosis. London-New-York: Academic Press, 1983. 483 p.
9. Godbout C, Fortin J.A. Synthesized ectomycorrhizae of aspen: fungal genus level of structural characterization // Can. J. Bot. 1984. Vol.63. №2. P.252-256.
10. Qian X.M., Kottke I., Oberwinkler F. Activity of different ectomycorrhizal types studied by vital fluorescence // Plant and Soil. 1998. Vol.199. P.91-98.
11. Smith S.E., Read D.J. Mycorrhizal Symbiosis. - London: Academic Press Limited. - 1997. - 514 p.
12. Woo K.-S., Fins L., McDonald G.I., Wenny D.L., Eramian A. Effects of nursery environment on needle morphology of Pinus monticola Dougl. and implications for tree improvement programs // New Forests. 2002. Vol.24. №2. P.113-129.
13. Roman., Claveria V., Miguel M. A revision of the descriptions of ectomycorrhizas published since 1961 // Mycology research. The British Mycological Society. UK, 2005. - T.109 (10). - P.1063-1104.
14. Giraud M. Prelevement et analyse de mycorhizes. In La truffe, FNPT 10. - 1987. - P.49-63.
15. Hutchinson L.J. Description and identification of cultures of ectomycorrhizal fungi found in North America // Mycotaxon, 1991. - V.XLII. - P.387-504.
16. Sellin A. Morphological and stomatal responses of Norway spruce foliage to irradiance within a canopy depending on shoot age // Env. and Exp. Bot. 2001. Vol.45. №3. P.115-131.
17. Agerer R. (ed.). Colour Atlas of Ectomycorrhizae. Einhorn-Verlag, Schwabisch Gmund. - 1996-2008. - V.I-VI.
18. Agerer R. Ectomycorrhizae of Tomentella albomarginata (Thelephoraceae) on Scots pine // Mycorrhiza. 1996. Vol.6. №1. P.1-7.
19. Agerer R. Fungal relationships and structural identity of their ectomycorrhizae // Mycol Progress. 2006. Vol.5. N2. P.67-107.
20. Read D.J. Plant-microbe mutualisms and community structure // Biodiversity and ecosystem function. Ed.: Schulze E.D., Mooney H.A. Springer-Verlag. Berlin Heidelberg. 1999. P.181-209.
21. Tedersoo Leho. Ectomycorrhizal fungi: diversity and community structure in Estonia, Seyshelles and Australia / Dissertationes Biologicae Universitatis Tartuensis. - Tartu Ulikooli Kirjastus. - 2007. - 54 p.
22. Вайшля О.Б., Ведерникова A.A., Данченко A.M., Фолин A.M., Фролов B.B. Биологически активные вещества макромицетов, образующих эктомикоризу с хвойными, при культивировании in vitro // Лесное хозяйство и зеленое строительство в Западной Сибири: Матер. IV междунар. Интернет-семинара. Томск: Изд-во Томского госуниверситета, 2009, 268 с. - С.34-48.
23. Вайшля О.Б., Фролов В.В. Экспресс-технология восстановления хвойных лесов на проблемных землях // Матер. VI Всероссийской научно-практической конференции с международным участием «Инновационные технологии». - Томск: Издательство Томского политехнического университета, 2008. - С.331-335.
24. Вайшля О.Б., А.А. Ведерникова, А.П. Бондаренко. Микробиологические аспекты гипергенеза. - Томск: ТМЛ - Пресс, 2007. - 288 с.
25. Вайшля О.Б., А.А.Ведерникова. Культивирование in vitro, идентификация и биохимический состав некоторых видов Homobasidiomycetidae, образующих эктомикоризу с хвойными // Хвойные бореальной зоны, 2009. Том XXVI, №1. - С.58-61.
26. Худяков Я.П., Возняковская Ю.М. Чистые культуры микоризных грибов // Микробиология. М.: Изд-во АН СССР 1951. Т.ХХ. Вып.1. С.14-19.
Приложение
зид, фМ
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ повышения приживаемости саженцев сосны обыкновенной | 2023 |
|
RU2824516C1 |
| СПОСОБ ПОДГОТОВКИ МИЦЕЛИЯ БАЗИДИОМИЦЕТОВ ДЛЯ МИКОРИЗАЦИИ ПОЧВЫ ПРИ ИСКУССТВЕННОМ ВЫРАЩИВАНИИ ХВОЙНЫХ РАСТЕНИЙ | 2020 |
|
RU2751481C1 |
| СПОСОБ ЭКСПРЕСС-ОПРЕДЕЛЕНИЯ СИМБИОТИЧЕСКОЙ ЭФФЕКТИВНОСТИ ГРИБОВ АРБУСКУЛЯРНОЙ МИКОРИЗЫ И ИНДЕКСОВ МИКОРИЗАЦИИ | 2012 |
|
RU2528864C2 |
| Биотехнологический способ оптимизации производства привитых саженцев винограда на основе применения гриба Glomus intraradices Shenck & Smith, штамм RCAM02146 | 2017 |
|
RU2672381C2 |
| СПОСОБ ЗАСЕЛЕНИЯ КОРНЕЙ РАСТЕНИЙ КЛЮКВЫ МИКРОМИЦЕТОМ OIDIODENDRON MAIUS BARRON | 2022 |
|
RU2821889C2 |
| Способ оценки качества биопрепаратов на основе грибов арбускулярной микоризы | 2021 |
|
RU2776652C1 |
| Способ периодического глубинного культивирования мицелия базидиальных грибов | 2024 |
|
RU2821927C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПОСЕВНОГО МАТЕРИАЛА ЧЕРНОГО ТРЮФЕЛЯ | 2023 |
|
RU2833962C1 |
| СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСНОГО МИКРОБИОЛОГИЧЕСКОГО УДОБРЕНИЯ | 2006 |
|
RU2318784C2 |
| ТЕХНОЛОГИЯ ВОССТАНОВЛЕНИЯ ЛЕСНЫХ ЭКОСИСТЕМ НА ТЕХНОГЕННО НАРУШЕННЫХ ТЕРРИТОРИЯХ ЕВРОПЕЙСКОГО СЕВЕРО-ВОСТОКА РОССИИ | 2007 |
|
RU2343692C1 |

Изобретение относится к сельскохозяйственной биохимии. Средство представляет собой штамм эктомикоризообразующего гриба Suillus sibiricus (6) №LE(BIN)2178. Штамм депонирован в Коллекции культур базидиомицетов LE(BIN) Учреждения Российской академии наук Ботанического института им. В.Л.Комарова РАН с присвоением регистрационного №LE(BIN)2178. Изобретение позволяет преодолеть трудности, связанные с гетеровегетативным размножением и пересадкой ценнейших пород хвойных деревьев. Также позволяет увеличить приживаемость хвойных на безлесных территориях и урбанизированных землях, на обедненных почвах в ландшафтно-озеленительных работах, при укреплении дамб и озеленении угольных карьеров. 6 ил., 5 табл.
Средство для увеличения приживаемости и стимуляции роста сеянцев и саженцев сосны сибирской, представляющее собой штамм эктомикоризообразующего гриба Suillus sibiricus (6) №LE(BIN)2178.
| ВАЙШЛЯ О.Б | |||
| и др | |||
| Культивирование in vitro, идентификация и биохимический состав некоторых видов Homobasidiomycetidae, образующих эктомикоризу с хвойными, Хвойные бореальной зоны 2009, №1, с.58-61 | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| Стимулятор роста сеянцев древесных культур | 1989 |
|
SU1681751A1 |

