Область техники, к которой относится изобретение
Данное изобретение обеспечивает новый способ приготовления синтетического комплекса инвазивов Invaplex, включающего, по меньшей мере, два инвазивных белка и липополисахарид из инвазивных грамотрицательных бактерий. Синтетический инвазивный комплекс настоящего изобретения может применяться в качестве вакцин, адъюванта для вакцин, биохимических иди других субстанций и в качестве диагностического средства.
Предшествующий уровень техники
Шигеллез является главной причиной острой кишечной инфекции у людей, в особенности в развивающихся странах, где, как предполагается, происходит ежегодно более 163 миллионов случаев заражения, с 1 миллионом летальных исходов [1]. Наиболее распространенными видами Shigella, вызывающими заболевание, являются S.flexneri и S.sonnei [1]. Наличие даже более низкого количества случаев (1,5 миллиона случае/год) заболевания шигеллезом в промышленно развитых странах [1] также показывает, что популяция взрослых иммунологически не защищена и восприимчива. Даже в областях, относительно свободных от шигеллеза, массивное разрушение от войны или стихийных бедствий, таких как землетрясение в 1999 в Kocaeli, Турция, может быстро вызвать вспышку многовидовой, мультифокальной дизентерии [2]. Аналогично, при развертывании войск работники по оказанию помощи или путешественники из развитых стран находятся в областях эпидемии Shigella или в областях, где местная инфраструктура разрушена, то лечение вспышки множественной Shigella spp. может быть значимым. S.dysenteriae 1 вызывает наиболее тяжелую форму шигеллеза и часто ассоциируется с гемолитическим уремическим синдромом [3]. Хотя частота возникновения заболевания S.dysenteriae 1 не так высока, как при S.flexneri или S.sonnei, эпидемии, вызванные S.dysenteriae 1, возникают каждые 10-12 лет [4]. Периодическое возникновение является следствием многих факторов, один из которых, по видимости, связан со снижением иммунитета к S. dysenteriae 1. Наиболее важным является недавнее открытие, что резистентность к антибиотикам фторхинолонового ряда наблюдается при S.dysenteriae 1, выделенной во время последних вспышек [5, 6]. Вероятность следующей эпидемии S.dysenteriae 1 высока и, если она будет вызвана штаммами с резистентностью к антибиотикам широкого спектра действия, включая фторхинолоны, то последствия могут быть тяжелыми.
Виды Shigella являются одними из некоторых подтвержденных агентов биологического терроризма. Очень низкая инфекционная доза (для S.dysenteriae 1 она может составлять 10 бактерий) [7] и простота выращивания организма в простой бактериологической среде являются подходящими факторами для рассмотрения Shigella spp в качестве мощного биологического оружия. Эффективность как агента биологического терроризма подтверждается случаем, когда сотрудники госпиталя были преднамеренно инфицированы S.dysenteriae 2 недовольным сотрудником, который имел "раскрашенные" груди на теле [8]. Другие пищевые вспышки инфекции Shigella хорошо известны - от холодной еды в самолетах до салатов на круизных кораблях [9, 10], что демонстрирует легкость, с которой shigellae могут вызывать заболевание в уязвимой популяции. Предохранение от инфекций Shigella лучше всего выполняется посредством контроля за состоянием окружающей среды, что включает хороший санитарный контроль и обеспечение запасов чистой питьевой воды. С такими усовершенствованиями инфраструктуры не трудно справиться, если shigellae преднамеренно внесена в пищу или напитки. Кроме того, легкость выделения shigellae из природы [5] и простота развития резистентности к антибиотикам штаммов S.dysenteriae 1 в лаборатории подтверждает, что эти агенты являются возможной основной угрозой терроризма. Хотя может быть затруднительным полностью остановить начало вспышки, важным является ограничение вторичного распространения. Эффективные пути профилактики представляют собой вакцины, которые эффективны против shigellae, независимо от резистентности к антибиотикам.
Патогенез Shigella spp. заключается в способности организма к внедрению, внутриклеточной репликации и внутриклеточному распространению в эпителии толстого кишечника. Начальное внедрение shigellae происходит через специализированные интестинальные эпителиальные клетки, называемые М-клетки. Некоторые высококонсервативные, вирулентные кодированные плазмидой белки, называемые плазмидные антигены инвазии (IpaA, IpaB, IpaC и IpaD), являются основными участниками в процессе инвазии и, по всей вероятности, являются ключом эффективности инфекций Shigella. При контакте или прикреплении к клеткам-хозяевам инвазивы Shigella высвобождаются [11] посредством аппарата секреции III типа [12] и индуцируют процесс фагоцитоза, что приводит к захвату и поглощению бактерии клеткой-хозяином [13]. Активные компоненты включают комплекс IpaB:IpaC, который распределяется в мембране клетки-хозяина, формируя канал, по которому другие белки Shigella входят в клетку-хозяина [14]. Недавно мы выделили из интактных, вирулентных клеток Shigella инвазивный комплекс белок-LPS, который служит эффективной вакциной и адъювантом [15-19]. Инвазивный комплекс Shigella (Invaplex) получали путем экстрагирования вирулентной шигеллы водой с последующей анионообменной хроматографией и водной экстракцией. Две фракции, называемые Invaplex 24 и Invaplex 50, содержали основные антигены, включая LPS, IpaB и IpaC. Инвазивный комплекс (Invaplex) прикреплялся к поверхностям культивированных эпителиальных клеток, тогда как аналогичные препараты из неинвазивных шигелл не обеспечивали этого свойства. Сразу после прикрепления Invaplex становился локализованным внутри клетки-хозяина посредством актин-зависимого эндоцитоза, по-видимому, индуцированному Invaplex. Представляется, что IpaC и IpaB играют существенную роль в прикреплении и поглощении, при которых антитела против IpaC или IpaB препятствуют присоединению Invaplex к мембранам клетки-хозяина [20]. Антитела к LPS не изменяют процесс поглощения. Как только поглощенный Invaplex двигается через ранние эндосомы, поздние эндосомы, затем аппарат Гольджи в итоге оказывается свободным в цитоплазме клетки-хозяина. Способность прикрепляться к поверхности эукариотической клетки-хозяина и стимулировать эндоцитоз показывает, что Invaplex сохраняет активную, нативную вирулентную структуру, аналогичную той, которая найдена для инвазивной Shigella.
Исторически, успешные вакцины Shigella усиливают представления об эффективной роли LPS в развитии защиты. Многие подходы используются для вакцин Shigella, включая живые ослабленные вакцины [21, 22], инактивированные вакцины [23, 24] и доставку LPS Shigella или О-полисахаридов с носителями, такими как протеосомы [25], столбнячный анатоксин [26], рибосомы [27] или Invaplex (17, 18; см. ниже). Только живые ослабленные вакцины применяют нативную инвазивность шигелл для доставки LPS и белковых антигенов к мукозной иммунной системе, вероятно, через связанный с фолликулами эпителий [14]. Остаточная патогенность ослабленных вакцинных штаммов может ограничивать этот подход, хотя достигается дополнительное ослабление [28].
Способность к изоляции предполагаемой нативной поверхностной структуры, такой как Invaplex, которая проявляет активности и иммуногенность, аналогичные инвазивной шигелле, имеют значительные сложности при создании вакцин и их разработке. Первое, изолированный нативный комплекс может усиливать доставку к подходящим входным воротам (М-клетки), аналогично с той, которая направляется живыми ослабленными вакцинными штаммами. Прикрепление Invaplex к клеткам-хозяевам (и, возможно, интестинальным М клеткам или М-подобным клеткам в других слизистых тканях) позволяет использовать относительно низкие дозы этого субклеточного комплекса для иммунизации благодаря их эффективности доставки. Аналогично с живыми, ослабленными вакцинами Invaplex содержит все основные антигены Shigella (включая IpaB, IpaC и LPS) и имеет потенциал для стимуляции иммунного ответа, эквивалентного с таковым, получаемым в течение естественной инфекции, включая узнавание эпитопов, обнаруженных только в нативных структурах. Такие эпитопы могут не присутствовать в вакцинах, доставляющих только LPS или О-полисахарид в качестве антигена. Кроме того, консервативные последовательности и иммунологическая перекрестная реактивность белков Ipa, найденных во всех видах Shigella, могут обеспечивать эффективность вакцины, содержащей инвазивы (или другие консервативные антигены), против более чем одного вида Shigella.
Вакцина из комплекса инвазивов Shigella (Invaplex), приготовленная из Shigella flexneri, содержит основные антигены LPS, IpaB и IpaC и является защитной против гомологичного заражения в моделях кератоконъюнктивита у морских свинок и инфицирования легких у мышей [17, 18]. После иммунизации продуцируются антитела к LPS, IpaB и IpaC, которые аналогичны специфике антител, которые наблюдаются после естественной инфекции у людей [29]. Дополнительные исследования показали, что вакцинные продукты Invaplex аналогичной эффективности могут быть выделены из видов Shigella и EIEC [15]. Хотя препараты Invaplex 24 и 50 состоят из многих разных белков, иммунодоминантными антигенами для Invaplex 24 являются LPS, IpaB и IpaC и для Invaplex 50 ключевыми антигенами являются LPS, IpaB, IpaC и 84 кДа (EF-G) и 72 кДа (DnaK) белковые антигены [18]. Другие белки в препаратах Invaplex не иммуногены, что определено с помощью методов вестерн-блоттинга, используя сыворотку, собранную из животных, иммунизированных Invaplex. Invaplex 24 и Invaplex 50 могут быть выделены из диких видов Shigella, хотя зачастую форма Invaplex 24 постоянно сдержит более высокие количества IpaB, IpaC и LPS и некоторые неиммуногенные белки. С вакциной Invaplex высокие титры часто используют против иммунизации Invaplex (Invaplex 24 или Invaplex 50), когда используется в качестве антигена для ELISA. Полагают, что это отражает комбинированный ответ на белки Iра и LPS и ряд коформационных эпитопов, сохраненных комплексным продуктом инвазивов, когда используется как антиген для ELISA.
В результате идентификации и очистки активной части в нативном Invaplex был идентифицирован высокомолекулярный комплекс, который был выделен с помощью гель-фильтрации из препаратов нативного Invaplex 24. Этот высокомолекулярный комплекс, называемый как "высокоочищенный" или HP-Invaplex 24, состоит преимущественно из IpaB, IpaC и LPS. HP-Invaplex 24 является иммуногенным и защитным при уровнях, которые сравнимы с или превышают те, которые проявляются у нативного Invaplex. Никакие другие фракции, полученные гель-фильтрацией, не обладали иммуногенностью и защитной способностью, сравнимой с нативным Invaplex, и по этой причине HP-Invaplex 24 рассматривают как активную часть в нативном Invaplex, ответственную за его иммуногенность и эффективность в качестве вакцины.
Вирулентные шигеллы вызывают заболевание путем проникновения, репликации и распространения в эпителии толстой кишки посредством комплексных серий клеточных и молекулярных событий, контролирующихся рядом плазмид-кодирующих вирулентных факторов, среди которых являются белки Iра [49]. После проникновения в интестинальные эпителиальные клетки слизистой толстой кишки происходит воспалительный ответ в слизистой, характеризующийся повышением провоспалительных цитокинов, что приводит к стимуляции нейтрофилов и макрофагов/моноцитов. В результате заболевание, шигеллоз или бактериальная дизентерия, вызывает развитие от умеренной до тяжелой диареи, лихорадки и интестинальных повреждений. Поскольку тестирование эффективности вакцин на людях и приматах включает заражение вирулентной шигеллой после проведения иммунизации, применение небольших экспериментальных моделей для начального тестирования вакцинных кандидатов снижает риск заболевания добровольцев и приматов. Небольшие экспериментальные модели, такие как модель кератоконъюнктивита морских свинок или легочная модель мышей с экспериментальным инфицированием шигеллой, широко используются для изучения патогенеза и предклинической оценки вакцины [18, 44, 50], при этом мышиная модель часто используется в начальных оценках и модель морских свинок используется для тестирования вакцин, которые являются защитными на примере мышиной модели.
Модель интраназального заражения мышей инфекцией Shigella является подходящей для оценки вакцин Shigella [18, 31, 51, 52]. Патогенез и иммунобиологию наблюдали в легочной модели параллельно с наблюдениями в толстой кишке; при этом вирулентные штаммы Shigella проникают, реплицируются и распространяются в эпителии и впоследствии вызывают иммунный ответ, а также цитокиновый ответ [52]. После инфицирования мыши теряют вес и в конечном итоге умирают, если только у них не развивается защитный иммунитет. Способность к измерению ответа секреторных антител, клеточного иммунного ответа и цитокинового ответа (преимущественно благодаря доступности коммерческих реагентов) делает мышиную модель очень привлекательной для исследований иммунобиологии шигеллеза.
Модель морских свинок является подходящей моделью, которая применяется для изучения вирулентности обоих диких и ослабленных штаммов Shigella и для оценки эффективности потенциальных вакцинных кандидатов [18, 44, 45, 53-56]. Несколько путей иммунизации могут применяться для иммунизации в зависимости от вакцины: пероральные, интраназальные, окулярные и парентеральные иммунизации, все из которых используются для защиты против окулярного заражения. Иммуногенность и эффективность вакцины Shigella на модели кератоконъюнктивита у морских свинок в настоящее время применяют в качестве «трамплина» в первой фазе клинических испытаний.
Существует потребность в химически определенной, синтетической части Invaplex, которая аналогична или превосходит по биологический активности нативного Invaplex. Также есть необходимость в процессе изготовления Invaplex, который может быть быстро переведен в серийное производство и приводить к получению более специфично определенного продукта, в зависимости от соотношений частей отдельных компонентов, которые используются. Также существует необходимость для синтетического Invaplex, который обеспечивается для специфичных применений или имеет специальные функции. Также существует потребность в создании вакцины Invaplex, которая может быть приготовлена быстро из его составных частей, которые могут накапливаться в ожидании будущих потребностей для использования вакцины.
Сущность изобретения
Изобретение относится к синтетическому Invaplex и способу его получения. Синтетические Invaplex (InvaplexAR) аналогичны HP Invaplex 24. HP Invaplex 24 содержит IpaB, IpaC и липополисахарид. Синтетический Invaplex действует как Invaplex, полученный из нативного источника, хотя он может иметь большую активность. Синтетический Invaplex содержит определенные компоненты: комплекс, содержащий инвазивные белки IpaB и IpaC, который дополнительно включает серотип-специфичный липополисахаридный компонент из грамотрицательных бактерий. Липополисахарид является иммуногеном, также как IpaB и IpaC. InvaplexAR может функционировать как мукозный адъювант и вызывать оба сывороточный и мукозный антиген-специфичные иммунные ответы, а также клеточно-опосредованные иммунные реакции. Также возможно использование других инвазивов из других видов бактерий, включая SipB, SipC или вирусные (реовирусные) белки, связанные с эндоцитозом или тканевыми тропизмами.
Липополисахариды, связанные с определенным серотипом, получают из грамотрицательных бактерий, таких как Shigella flexneri, Shigella sonnei, Shigella dysenteriae, Shigella boydii, enteroinvasive E.coli, Yersinia или Salmonella. Более предпочтительные виды включают S.flexneri, S.sonnei, S.boydii, S.dysenteriae или энтероинвазивную Е.coli.
Для приготовления синтетического Invaplex, например InvaplexAR, используют либо высокоочищенные (hp-), либо рекомбинантные (r-) инвазивные белки IpaB и IpaC, при этом очищенный липополисахарид используется в качестве исходного материала. Белки IpaB и IpaC смешивают с образованием комплекса IpaB:IpaC. Этот комплекс смешивают, по меньшей мере, с одним липополисахаридом, ассоциируемым с серотипом грамотрицательных бактерий, с получением синтетического Invaplex. Синтетический Invaplex выделяют из смеси. Обычно, синтетический Invaplex удаляют, основываясь на его заряде. Также используют другие методики очистки. Порядок добавления может изменяться. Липополисахарид может образовывать комплекс с любыми инвазивными белками и затем с другим инвазивным белком с формированием синтетического Invaplex.
Обычно, количество присутствующего IpaC относительно присутствия IpaB подпадает в интервал соотношений от 0,08:1 до 80:1, предпочтительно от 0,8:1 до 20:1, более предпочтительно, по меньшей мере, 8:1. Наиболее предпочтительным для количества IpaC на первом этапе смешивания является его присутствие, по меньшей мере, в десять раз больше относительно IpaB. На втором этапе смешивания липополисахарид и белок (IpaB и IpaC) присутствуют в соотношении от 0,01:1 до 10:1, предпочтительно, по меньшей мере, 1:2. На втором этапе смешивания возможным является включение двух или более типов липополисахаридов, которые приводят к получению синтетического Invaplex, который пригоден для применения в многовалентных иммунных композициях, в виде вакцин. Также является возможным, в дополнении к липополисахариду, включение детектирующей метки, антибиотика, лекарства или биомолекулы, содержащей фермент, белок, полисахарид, РНК или ДНК, или их производных, один из которых способен объединиться с клеткой. Это способствует увеличению возможностей терапевтического и аналитического/диагностического применений.
Синтетические Invaplex изобретения включают те, в которых IpaC и IpaB присутствуют в соотношении, выбранном от 0,08: 1 до 80:1, предпочтительно от 0,8:1 до 20:1, более предпочтительно приблизительно 8:1, и липополисахарид (LPS) присутствует при соотношении, выбранном из интервала от 0,01:1 до 10:1, предпочтительно от 0,5:1 до 5:1, более предпочтительно приблизительно 0,5:1 относительно содержания общего белка (IpaB и IpaC). Предпочтительные синтетические Invaplex включают InvaplexAR S.flexneri, InvaplexAR S.dysenteriae или InvaplexAR S.sonnei.
Синтетические Invaplex изобретения могут быть составлены с образованием композиций, пригодных для использования в качестве иммуногенных композиций или вакцин. Invaplex может использоваться как иммуноген и/или как адъювант. Композиции будут включать, в дополнение к синтетическому Invaplex, по меньшей мере, фармацевтически приемлемый носитель. Invaplex присутствует в количестве, эффективном для желаемого действия и в, по меньшей мере, первую очередь, определяется эмпирически. Предполагается, что это количество аналогично таковому для нативного Invaplex, например Invaplex 24, используемого в аналогичной цели.
Дополнено, синтетические инвазивные комплексы способствуют транспорту молекулы и связанных материалов через клеточную мембрану. Эти молекулы могут располагаться в непосредственной близости к представляющей интерес клетке и синтетическому Invaplex или могут присутствовать в и/или на синтетическом Invaplex. Молекулами могут быть детектирующие метки, антибиотики, лекарства или биомолекулы, содержащие белки, ферменты, РНК или ДНК, липополисахариды, полисахариды и тому подобные.
Синтетические Invaplex могут применяться в способах, в которых применяются нативные Invaplex, например способах иммунизации, способах улучшения транспорта биомолекул, например ДНК, РНК, белков, через мембраны клеток и др. Синтетический Invaplex может создаваться как многовалентный или иметь большую активность относительно нативного Invaplex. Это должно повышать эффективность существующего способа и его применимость. InvaplexAR, изготовленный из рекомбинантных IpaB, IpaC и либо LPS S.sonnei либо LPS S.flexneri 2a, вызывал иммунные ответы, которые сравнимы или выше по сравнению с иммунным ответом, вызванным нативным Invaplex. Ответы IpaB и IpaC при иммунизации InvaplexAR были постоянно выше, чем те, которые были вызваны после иммунизации нативным Invaplex. Иммунизация InvaplexAR предоставляет сравнимые или более высокие уровни защиты против заражения на моделях мышей и морских свинок по сравнению с иммунизацией нативным Invaplex. Иммунизации InvaplexAR индуцирует клеточный иммунитет, больший, чем Shigella-специфичный ответ, вызванный нативным Invaplex. См, например, таблицы 7 и 8.
Краткое описание чертежей
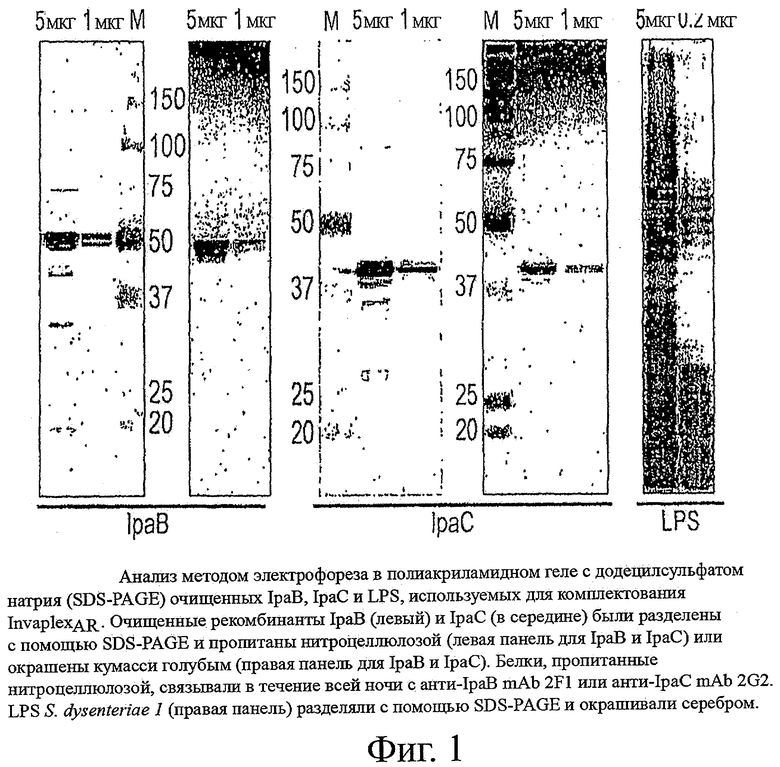
Фиг.1 показывает анализ методом электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) очищенных IpaB, IpaC и LPS, используемых для комплектования InvaplexAR;
фиг.2 показывает очистку InvaplexAR S.flexneri 2а (панель A), S.sonnei (панель В) и S.dysenteriae 1 (панель C);
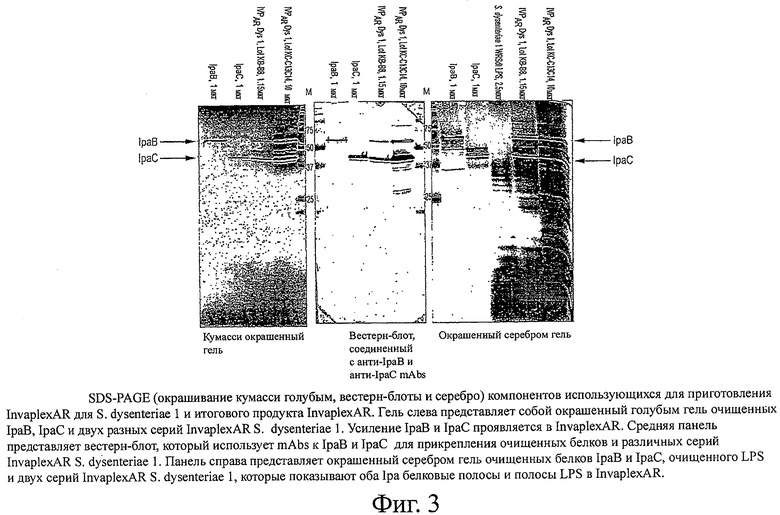
фиг.3 показывает SDS-PAGE (окрашивания кумасси голубым, вестерн-блоты и серебром) компонентов, использующихся для приготовления InvaplexAR для S.dysenteriae 1 и конечного продукта InvaplexAR;
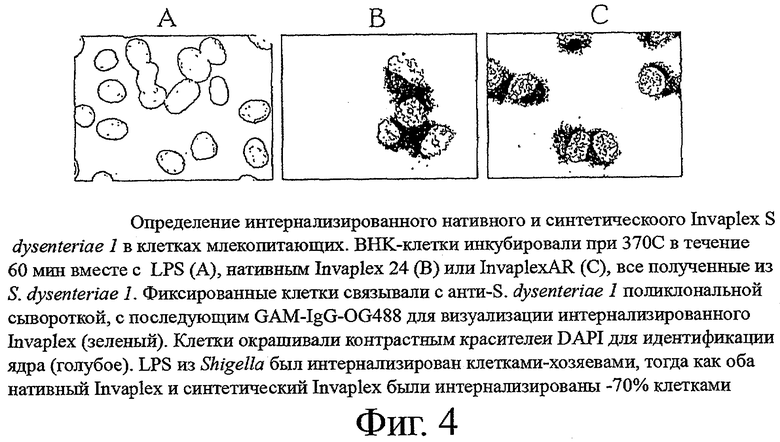
фиг.4 показывает определение интернализированного нативного и синтетического Invaplex S.dysenteriae 1 в клетках млекопитающих;
фиг.5 показывает титры Invaplex 50, Invaplex 24 и LPS-специфичных сывороточных IgG и IgA в конечных точках на день 42 после интраназальной иммунизации нативным Invaplex 24 S.flexneri 2а, InvaplexAR, очищенными IpaB, IpaC или LPS S.flexneri 2a;
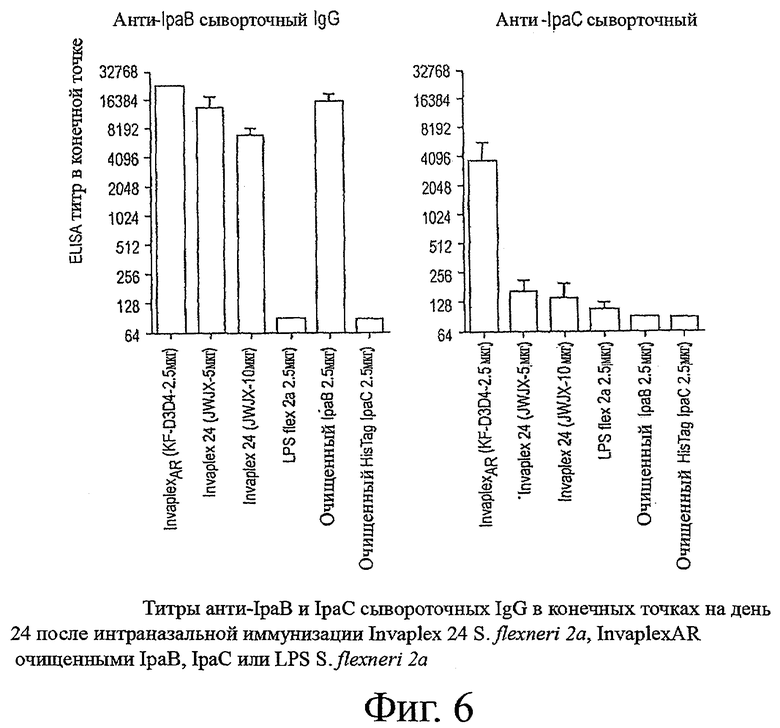
фиг.6 показывает титры анти-IpaB и IpaC сывороточных IgG в конечных точках на день 24 после интраназальной иммунизации Invaplex 24 S. flexneri 2а, InvaplexAR, очищенными IpaB, IpaC или LPS S.flexneri 2a;
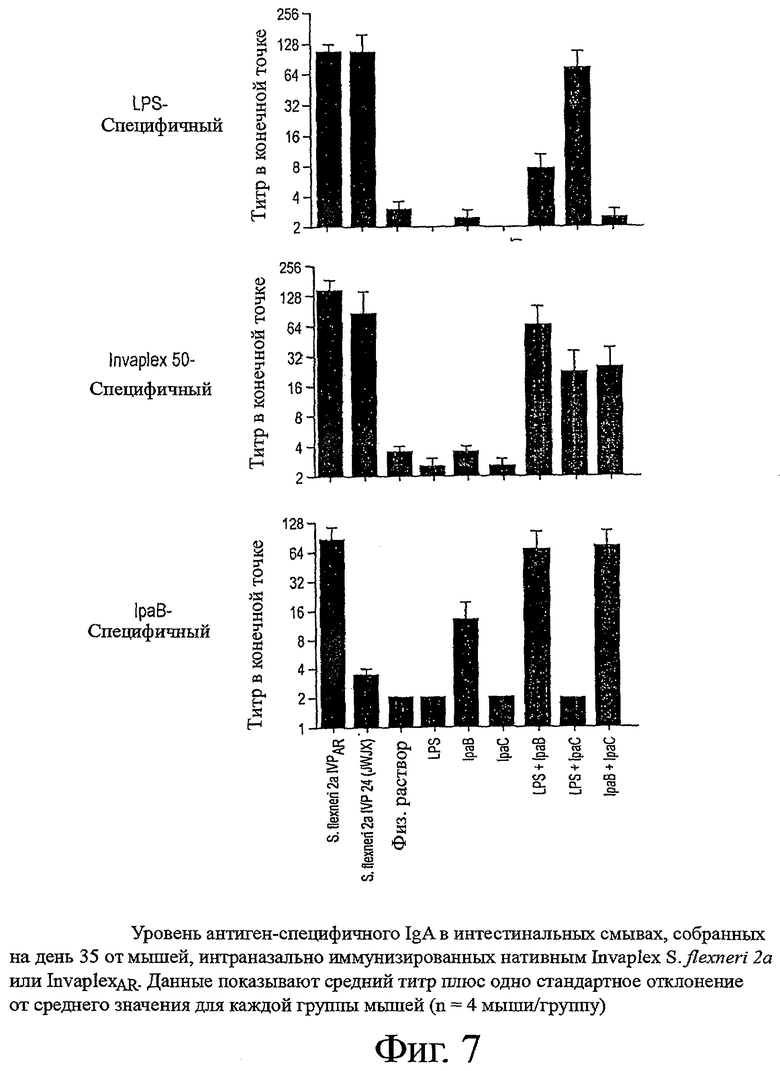
фиг.7 показывает уровень антиген-специфичного IgA в интестинальных смывах, собранных на день 35 от мышей, интраназально иммунизированных нативным Invaplex S.flexneri 2а или InvaplexAR;
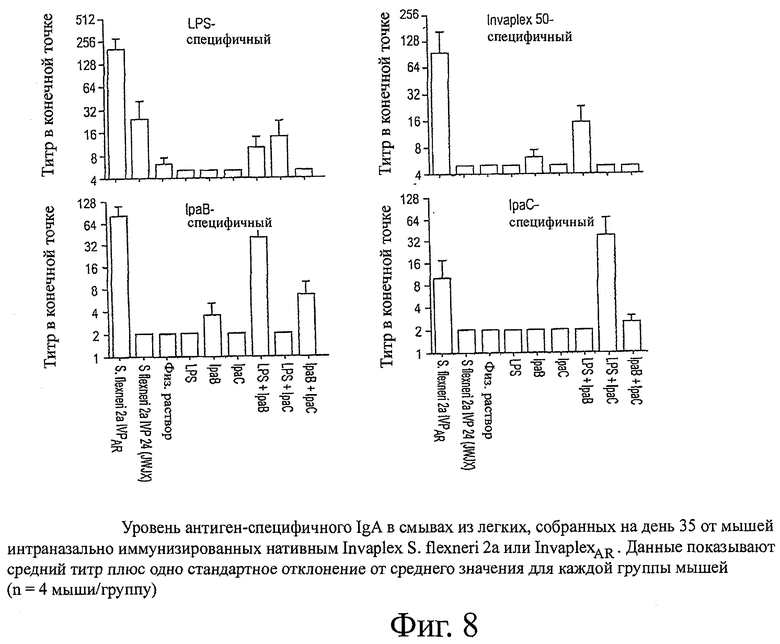
фиг.8 показывает уровень антиген-специфичного IgA в смывах из легких, собранных на день 35 от мышей, интраназально иммунизированных нативным Invaplex S.flexneri 2а или InvaplexAR;
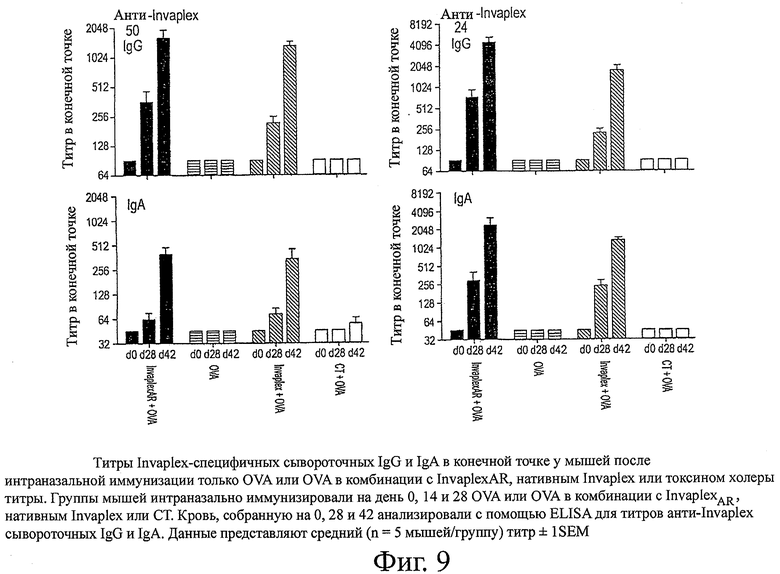
фиг.9 показывает титры Invaplex-специфичных сывороточных IgG и IgA в конечной точке у мышей после интраназальной иммунизации только OVA или OVA в комбинации с InvaplexAR, нативным Invaplex или токсином холеры;
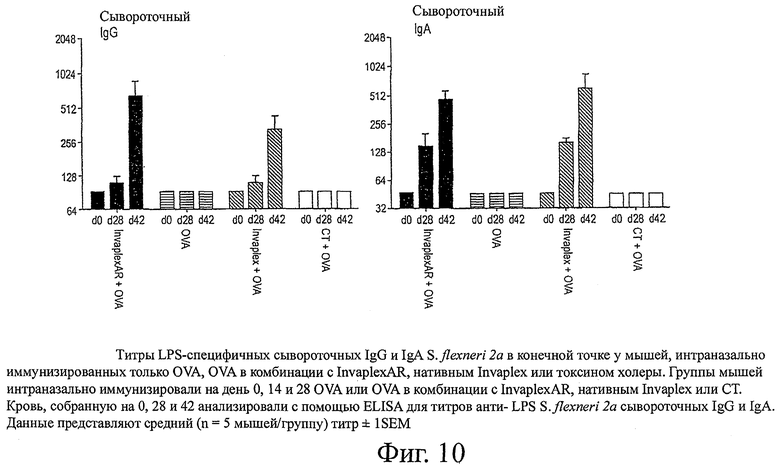
фиг.10 показывает титры LPS-специфичных сывороточных IgG и IgA S.flexneri 2а в конечной точке у мышей, интраназально иммунизированных только OVA, OVA в комбинации с InvaplexAR, нативным Invaplex или токсином холеры;
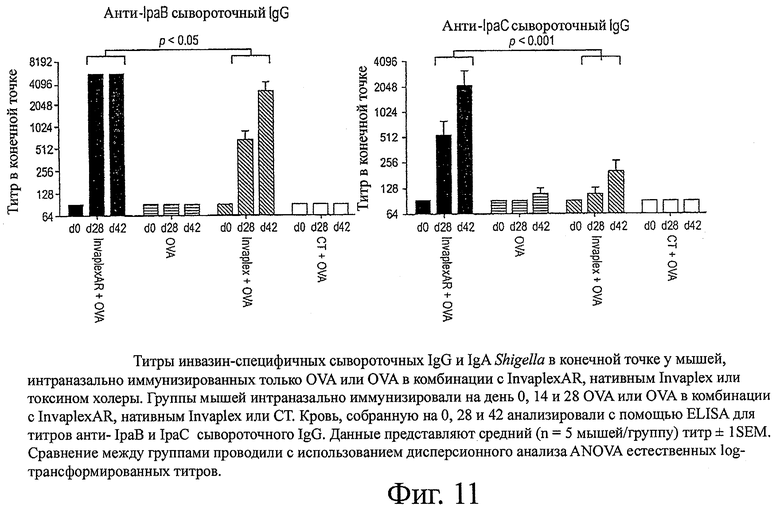
фиг.11 показывает титры инвазив-специфичных сывороточных IgG и IgA Shigella в конечной точке у мышей, интраназально иммунизированных только OVA или OVA в комбинации с InvaplexAR, нативным Invaplex или токсином холеры;
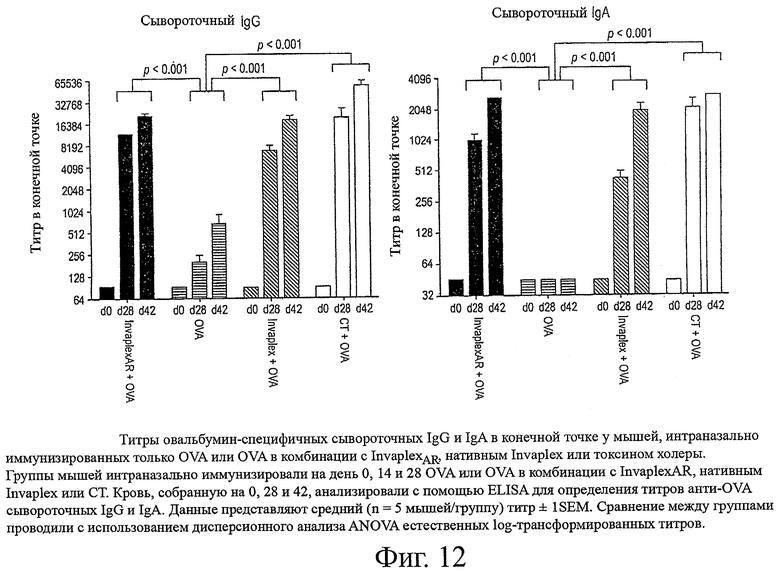
фиг.12 показывает титры овальбумин-специфичных сывороточных IgG и IgA в конечной точке у мышей, интраназально иммунизированных только OVA или OVA в комбинации с InvaplexAR, нативным Invaplex или токсином холеры;
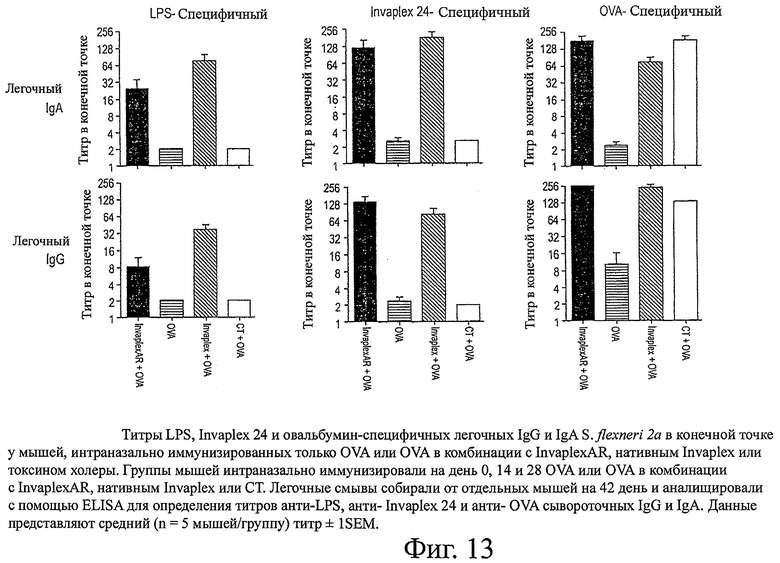
фиг.13 показывает титры LPS, Invaplex 24 и овальбумин-специфичных легочных IgG и IgA S.flexneri 2а в конечной точке у мышей, интраназально иммунизированных только OVA или OVA в комбинации с InvaplexAR, нативным Invaplex или токсином холеры.
Подробное описание изобретения
Биохимия синтетического Invaplex
Получение и очистка рекомбинантных IpaB и IpaC и нативного LPS S.flexneri, S.sonnei и S.dysenteriae 1
Очистка белков Ipa
Белки Ipa являются высокостабильными во всех Shigella spp [32, 33]. Рекомбинантная E.coli, экспрессирующая или IpaB, или IpaC, ранее была описана. Для очистки рекомбинантных белков Ipa существуют два подхода. Меченный гистидином (HisTag) рекомбинантный IpaC очищают с помощью аффинной хроматографии, используя никелевые колонки. Действие гистидиновых остатков на Ipa белковые биологические и иммунологические функции или способность белков формировать Invaplex не изучено, однако полагают, что HisTag-IpaC сохраняет биологическую активность [30]. Альтернативная процедура очистки используется для получения рекомбинантного IpaB. Этот способ пользуется преимуществом нативным сродством IpaB к шаперону IpgC [34, 35]. Посредством коэкспрессии IpaB совместно с HisTag-шаперон IpgC в том же рекомбинантном организме возможным является очистка комплекса IраВ/шаперон. После очистки белковый комплекс HisTag-IpaC/IpaB денатурируют, что приводит к образованию мономерного белка IpaB и HisTag-IpgC, который позволяет шаперону, меченному гистидином, селективно извлекаться на никелевой колонке, пока свободный белок Ipa проходит через колонку.
Очистка HisTag-IpaC
Рекомбинантный IpaC экспрессировали в E.coli с использованием индуцибельной системы экспрессии IPTG (плазмида рЕТ). Ген ipaC был клонирован в плазмиду pET15b (Novagen) и экспрессирован в векторе BL21(DE3)pLysS E.coli [30]. Рекомбинантный белок IpaC содержал несколько гистидиновых остатков на N-конце, что обеспечивает возможность очистки на никелевой колонке [30]. Экспрессирующийся белок IpaC располагается в тельцах включения и требует солюбилизации мочевиной. Удаление мочевины часто приводит к потере растворимости, если концентрация IpaC слишком высокая (>1 мг/мл). Таким образом, IpaC, элюированный с никелевой колонки, сохранялся в мочевине с сохранением растворимости. Используя в малом масштабе культуры (3 л), возможным является получение приблизительно 75 мг очищенного IpaC с выходом приблизительно 8 мг/л. Продукт является более чем на 90% чистым и активен с анти-IpaC mAb 2G2 [32] (фиг.1). Очищенный блок растворим и может храниться замороженным. Пример очищенного IpaC представлен на фиг.1.
Очистка IpaB
Рекомбинантный IpaB экспрессировали в Е.coli BL21(DE3). Ген ipgC клонировали в pET15b; ген ipaB клонировали отдельно в pACYC - дуэтный вектор. После введения IPTG комплекс HisTag IpgC/IpaB растворяли и очищали на никелевой аффинной колонке. Комплекс HisTag-IpgC/IpaB элюировали в присутствии EDTA. Для повторной очистки на никелевой аффинной колонке удаляли EDTA и добавляли неионный детергент ОРОЕ (N-октил-олиго-оксиэтилен) до конечной концентрации 1% объем/объем. OPOE разрушает комплекс HisTag-IpgC/IpaB, обеспечивая тем самым возможность свободному белку HisTag-IpgC связываться с никелевой (Ni-Сефароза) колонкой, а свободному от метки IpaB проходить через колонку. Фракции из свободного объема исследовали на наличие IpaB с помощью анти-IpaB mAb 2F1. Положительные фракции анализировали с помощью SDS-PAGE (окрашенной) для определения наличия загрязненного HisTag-IpgC. В случае загрязнения материал снова обрабатывают ОРОЕ и повторно наносят на никелевую колонку для удаления остаточного HisTag-IpgC. Конечный растворимый продукт IpaB практически не загрязнен HisTag-IpgC, приблизительно на 90% чистый и взаимодействует с анти-IpaB mAb 2F1 [32] (фиг.1). Выход IpaB на литр исходной культуры составлял примерно 3,5 мг/л.
Очистка LPS S.flexneri 2а, S.sonnei или S.dysenteriae 1
LPS S.flexneri 2a, S.sonnei и S.dysenteriae 1 был получен по процедуре Вестфола [36], которая включает экстрагирование шигелл горячим фенолом/водой. В качестве источника LPS могут использоваться вирулентные или ослабленные штаммы Shigella, поскольку экспрессируется гладкий фенотип LPS. В вышеописанных экспериментах использовались дикие типы S.flexneri 2а (штамм 2457Т) и S.sonnei (Mosely). Для S.dysenteriae 1 использовался ослабленный штамм WRSdI для снижения риска инфицирования персонала лаборатории. WRSdI является virG, нокаут stx, ранее полученный на WRAIR [37].
Экстракция, очистка и характеристика LPS
LPS экстрагируют методом Вестфола [36]. Вкратце, осадки бактериальной клетки суспендируют в горячей (68°C) дистиллированной воде (5 мл воды на каждый грамм осадка). Добавляют такое же количество фенола, нагретого до 68°C, и раствор тщательно встряхивают в течение 15 минут. Затем пробирки охлаждают до приблизительно 10°C±5°C. Образцы центрифугируют, удаляют водную фазу и хранят при 4°C. Экстракцию клеточной массы осуществляют дважды и все водные фазы собирают. Водную фазу подвергают диализу против дистиллированной воды в течение двух дней и затем центрифугируют (8000×g, 30 мин) для удаления посторонних продуктов распада клеток. Этот супернатант подвергают ультра-центрифугированию (90000×g) в течение 2 часов и осадок сохраняют. Осадок промывают стерильной дистиллированной водой, ресуспендируют в стерильной дистиллированной воде при 4°C в течение ночи, объединяют и лиофилизируют. Конечный лиофилизованный продукт взвешивают и затем берут небольшую порцию (<10 мг), растворяют в 1 мл воды, свободной от токсинов, и проводят биохимический анализ (см. ниже).
Определение эндотоксина очищенного LPS выполняют с помощью хромогенного лизата амебоцитов мечехвоста (LAL). Эндотоксин E.coli служит контрольным реагентом для этого анализа. Все результаты представляли в международных единица эндотоксина (EU). Очищенный LPS также анализировали с помощью SDS-PAGE с окрашиванием серебром для определения наличия типичного многополосного профиля гладкого LPS. Фиг.1 показывает окрашенный серебром гель очищенного LPS S.dysenteriae 1, обнаруживая типичный многополосный характер гладкого LPS. Конечный продукт LPS имеет следовое содержание белков (<5%, определенное с помощью анализа Брэдфорда) и ДНК (<5%, определенное с помощью окрашивания Хехстом) и вступает в реакцию с серотип-специфичными антителами к LPS (анти-S.dysenteriae 1 LPS mAb MAB753 (Chemicon International) для LPS S.dysenteriae 1; mAb 2E8 for S.flexneri 2a LPS; MAB755 (Chemicon) для LPS S.sonnei), определенное с помощью вестерн-блоттинга или ELISA.
Количественное определение содержания IpaB и IpaC в InvaplexAR с помощью иммунологического анализа
Количество IpaB и IpaC в Invaplex (синтетическом или нативном) определяли, используя модифицированную процедуру ELISA. В ELISA использовали очищенные рекомбинантные белки IpaB или IpaC для получения стандартных графиков определения количества антигенов в препаратах Invaplex. Планшеты для проведения ELISA Immulon IB (ThermoLab Systems) сенсибилизировали в течение ночи при 4°C либо 50 мкл рекомбинантного IpaB, рекомбинантного IpaC либо InvaplexAR. Антиген титровали (трижды), используя 2-кратные серийные разведения в карбонатном сенсибилизирующем буфере (0,2 М карбоната, pH 9,8) при начальных концентрациях 125 нг/мл (IpaB), 200 нг/мл (IpaC) и 10 мкг/мл (Invaplex). После промывания и блокирования казеином аффинно-очищенные моноклональные антитела, специфичные для IpaB (2F1) или IpaC (2G2) [32], инкубировали с планшетами, сенсибилизированными антигеном в течение 2 часов. После промывания детектировали антиген-специфичное антитело, используя антимышиный иммуноглобулин G (IgG), конъюгированный со щелочной фосфатазой (Kirkegaard & Perry). В качестве субстрата использовали пара-нитрофенилфосфат и измеряли оптическую плотность при длине волны 405 нм (OD405), используя планшет-ридер ELISA (Molecular Devices, Menlo Park, CA) через 60 минут после инкубации с субстратом. Используя программу Softmax Pro 4.5 (Molecular Devices), строили стандартные кривые, нанося OD405 против концентрации (нг/мл). Концентрацию неизвестных образцов затем интерполировали из стандартной кривой.
InvaplexAR создавали таким, чтобы он имел концентрации IpaB и IpaC и соотношение IpaC/IpaB, аналогичные Invaplex 24 HP. Определяли соотношение количества IpaC к IpaB для InvaplexAR и сравнивали с HP-Invaplex 24, наиболее чистой формой Invaplex. Молярное соотношение IpaC/IpaB в HP-Invaplex составляло приблизительно 8. Это определяли с помощью денситометрии гелей SDS-PAGE и анализов определения антител для IpaB и IpaC. В дополнение, предполагали, что содержание LPS соответствует относительной массе (приблизительно 0,5-0,6 мг LPS на каждый 1 мг белка), которая была найдена для HP-Invaplex.
Измерения LPS в Invaplex
Содержание LPS в препаратах Invaplex измеряли с помощью определения количества 2- кето-3-деоксиоктоната (KDO) в каждом препарате [43] или с помощью использования анализа амебоцитного лизата «лимулуса» (Cambrex Inc.).
Способ формирования синтетического Invaplex. Приготовление InvaplexAR для S.flexneri 2а, S.sonnet и S.dysenteriae 1
Очищенные компоненты перемешивали в соотношении, аналогичном для высокоочищенного нативного Invaplex, с образованием синтетического Invaplex. Анализ HP-Invaplex S.flexneri 2а показал, что молярное соотношение IpaC/IpaB составляло приблизительно 8,0 и соотношение LPS к общему белку составляло приблизительно 0,56 мг LPS/мг общего белка. Используя эти параметры в качестве критерия для восстановления Invaplex из очищенных IpaB, IpaC и LPS, проводили серии экспериментов для создания InvaplexAR. Синтетический Invaplex очищали сразу после образования с помощью ионообменной жидкостной хроматографии быстрого разрешения (FPLC).
Очищенные, растворимые IpaB и IpaC, каждый в своем соответствующем конечном буфере, смешивали друг с другом в молярном соотношении IpaC/IpaB 8. После перемешивания IpaB и IpaC раствор медленно добавляли к сухому порошку LPS (соотношение LPS к общему белку составляет 0,56). Может быть использован LPS из любого вида Shigella, для описываемых экспериментов использовали LPS S.flexneri 2а, S.sonnei или S.dysenteriae 1. Смесь инкубировали при 37°C приблизительно в течение 2 часов при встряхивании. Затем смесь белок/LPS разбавляли 20 мМ Tris-HCl, 0,10 М NaCl и 1,2 М мочевины для снижения концентрации NaCl перед выполнением ионообменной хроматографии. Разбавленную смесь затем очищали на анионообменной колонке HiTrap(TM) Q HP.
Эксперименты по сборке: формирование InvaplexAR S.flexneri
Предварительные эксперименты по сборке компенсировали нерастворимость IpaC путем сохранения обоих белков IpaC и IpaB в буфере, содержащем ≥4М мочевины. Как только смесь IpaC/IpaB добавили к LPS, растворимость не являлась более проблемной и допускала возможность удаления мочевины. Очищенный типичный IpaC сам по себе осаждается при разбавлении. В предварительных экспериментах молярное соотношение IpaC/IpaB 8 было сохранено путем добавления 8 мкМ HisTaglpaC к 1 мкМ IpaB в конечном объеме 1 мл. Оба белка изначально готовили в 20 мМ Tris-HCl, 0,5 М NaCl, 6 М мочевины, рН 7,9. Белки смешивали в стеклянной пробирке и инкубировали при температуре 37°C без встряхивания в течение 15 мин. Сухой LPS (230 мкг) в отдельной стеклянной пробирке также инкубировали при 37°C без перемешивания в течение 15 мин. Через 15 мин инкубации смесь IpaB+IpaC использовали для солюбилизации предварительно нагретого LPS путем медленного добавления белковой смеси по стенке пробирки с последующим интенсивным перемешиванием. Не наблюдалось заметного образования комочков и осаждение. Смесь IpaB/IpaC/LPS затем инкубировали при 37°C со встряхиванием (200 rpm) в течение 2 ч. Для ионообменной хроматографии (IEC) смесь разводили в пять раз предварительно нагретым буфером 20 мМ Tris, pH 9,0 для снижения концентрации соли. Осаждения смеси при охлаждении до комнатной температуры не наблюдалось. Для окончательной очистки разбавленную смесь IpaB/IpaC/LPS наносили на анионообменную колонку (HiTrap(TM) Q HP), для хроматографии использовали буфер A, содержащий 20 мМ Tris-HCl, рН 9,0, и буфер B, содержащий 1 М NaCl, 20 мМ Tris-HCl, рН 9,0, с элюцией ступенчатым градиентом от 0% (5 объемов колонки) до 24% (10 объемов колонки) до 50% (6 объемов колонки) до 100% Буфер B (5 объемов колонки). Собирали 1 мл фракций из 1 мл колонки HiTrap(TM) Q HP при скорости элюирования 1 мл/мин. Фракции анализировали на наличие IpaB, IpaC и LPS с помощью спотблоттинга. InvaplexAR S.flexneri элюированный в 50% В (фиг.2) содержал наибольшее количество всех трех компонентов (IpaB, IpaC и LPS), как было определено с помощью вестерн-блоттинга и окрашенных серебром гелей (фиг.2А). Комплекс IpaB/IpaC/LPS элюировался между стандартами 1 МДа и 669 кДа, то есть имел аналогичный интервал размеров, что и HP-Invaplex.
Крупномасштабное производство синтетического Invaplex S.dysenteriae 1
Повторяли вышеприведенный эксперимент, только с десятикратным увеличением количества реагентов для повышения выхода InvaplexAR. В дополнение, вместо LPS S.flexneri использовали LPS S.dysenteriae 1. Также 3,28 мг IpaC в 20 мМ Tris-HCl, 0,5 М NaCl, 6 М Мочевины, рН 7,9, смешивали с 0,62 мг IpaB в 20 мМ Tris-HCl, 0,5 М NaCl, рН 7,9, в общем объеме 5 мл. Белковую смесь и отельную пробирку с LPS (2,1 мг) предварительно нагревали до 37°C. Смесь IpaB+IpaC добавляли к пробирке с LPS и тщательно перемешивали для растворения LPS. Осаждения не наблюдалось. Реакционную смесь инкубировали при 37°C при встряхивании (200 rpm) в течение 2 часов. Затем предварительно нагретый Буфер А для IEC (20 мМ Tris-HCl, рН 9,0) добавляли для разбавления соли 1:5, получая конечный объем 25 мл. После охлаждения до комнатной температуры разбавленную реакционную смесь помещали в 1 мл колонку HiTrap(TM) Q HP для IEC и элюировали ступенчатым градиентом для очистки Invaplex (см. выше). InvaplexAR S.dysenteriae 1 элюировался в 50% Буфере B. Он содержал IpaB, IpaC и LPS S.dysenteriae 1 (фиг.2C). Анализ пиков 0,24 М и 1,0М NaCl не определил значимые количества IpaB, IpaC или LPS. Выход полученного InvaplexAR составлял приблизительно 10-20% от общего количества белка, использовавшегося для получения комплекса.
Аналогичные эксперименты по сборке проводились для S.sonnei, приводя к получению InvaplexAR S.sonnei (см. фиг.2B).
Крупномасштабное производство InvaplexAR Shigella flexneri 2а
Пятнадцатикратное увеличение содержания реагентов применяли для получения большего количества InvaplexAR для специальных исследований. Используя молярное соотношение 8 IpaC/1 IpaB, 16,4 мг IpaC (в 20 мМ Tris-HCl, 0,5 М NaCl, 6 М Мочевины, рН 7,9) смешивали с 3,1 мг IpaB (в 20 мМ Tris-HCl, 0,5 М NaCl, рН 7,9) в общем объеме 20 мл. Белковую смесь и отдельную пробирку с LPS Shigella flexneri 2а (11,4 мг) предварительно нагревали до 37°C. Смесь IpaB+IpaC добавляли к пробирке с LPS и тщательно перемешивали для растворения LPS. Реакционную смесь инкубировали при 37°C при встряхивании (200 rpm) в течение 2 часов. Затем предварительно нагретый Буфер A для IEC (20 мМ Tris-HCl, рН 9,0) добавляли для разбавлении соли 1:5, получая конечный объем 100 мл. После охлаждения до комнатной температуры разбавленную реакционную смесь наносили на 5 мл колонку HiTrap(TM) Q HP для IEC и элюировали ступенчатым градиентом для очистки Invaplex (см. выше). InvaplexAR S.flexneri 2а элюировался в 50% Буфере B. Он содержал IpaB, IpaC и LPS S.flexneri 2a. Анализ пиков 0,24 M и 1,0М NaCl не определил значимые количества IpaB, IpaC или LPS. Выход полученного InvaplexAR составлял приблизительно 9% от общего количества белка, использующегося для получения комплекса.
Анализ препаратов InvaplexAR с помощью электрофореза и вестерн-блоттинга для IpaB и IpaC
Общие белки препаратов InvaplexAR определяли с помощью SDS-PAGE окрашенных гелей. Вестерн-блоттинги использовали для определения присутствия, размера и относительных концентраций IpaB и IpaC. Окрашенные серебром гели использовали для оценки профиля LPS [18]. Моноклональные антитела, специфичные либо к белку IpaB, либо к IpaC, использовали для анализа нитроцеллюлозных блотов [32]. После инкубации с протеином A, конъюгированным со щелочной фосфатазой, блоты обрабатывали ASMX фосфатом (Sigma) и красителем Fast red TR (Sigma). Видеоизображения блотов анализировали на компьютере Macintosh, используя видеокамеру CCD (Cohu) и программное обеспечение NIH Image (версия 1.62). Денситометрия цифровых изображений осуществлялась с помощью аналитической программы GelPro Image (версия 2.0.10, Media Cybernetics, Inc.). Профиль геля, содержащего общий белок, и содержание IpaB и IpaC должны совпадать с таковыми для HP-Invaplex 24 или HP-Invaplex 50.
Гель-фильтрация
Гель-фильтрацию использовали для определении, действительно ли компоненты InvaplexAR, обнаруженные при ионообменной хроматографии, образуют комплекс. Это тот же способ, который используется для получения HP-Invaplex. На колонку Superose 6 HR 10/30 (Amersham Pharmacia), откалиброванную голубым декстраном 200 (2 МДа), тиреоглобулином (669 кДа) и овальбумином (43 кДа), наносили InvaplexAR (приблизительно 3 мг). Фракции (0,5 мл, при скорости потока 0,25 мл/мин) собирали и анализировали с помощью иммуно-спот блотинга на наличие IpaB, IpaC и LPS. Все три компонента InvaplexAR (IpaB, IpaC и LPS) элюировались во фракции между 2 МДа и 669 кДа, демонстрируя, что они связаны в комплекс, так как они элюировались с размером, намного большим, чем отдельные компоненты (43 кДа для IpaC; 62 кДа для IpaB). Размер SEC InvaplexAR аналогичен размеру, при котором элюировался HP-Invaplex.
Оценка InvaplexAR S.dysenteriae 1 в тканевой модели поглощения
Invaplex прикрепляется к и стимулирует эндоцитоз в культивированных клетках млекопитающих. Это свойство зависит от присутствия инвазивов IpaB и IpaC и применения актина клетки-хозяина в течение процесса эндоцитоза [38]. Анализ поглощения Invaplex является моделью in vitro, используемой для измерения функциональной активности препаратов Invaplex. Анализ моделировали после анализа бляшкообразования Shigella [39] и анализа поглощения Shigella [40, 41]. Анализ включал инкубирование InvaplexAR с фибробластами ВНК-21 (60-70% слипание) на предметном стекле. После инкубации в течение 5, 15, 30 или 60 минут клетки промывали трижды PBS и фиксировали формалином. Нарушали проницаемость мембран фиксированных клеток PBS, дополненного 0,1% Triton Х-100 и 0,1% BSA (промывочный буфер) и исследовали с помощью поликлональных мышиных сывороточных антител, специфичных к LPS, IpaB и IpaC. Связанные поликлональные мышиные антитела детектировали с помощью GAM-IgG-Oregon Green 488 (Molecular Probes), с последующим контрастным окрашиванием DAPI для идентификации ядра. Клетки, обработанные Invaplex, исследовали под флуоресцентными микроскопом с фиксированием цифрового изображения. Оба нативный и синтетический Invaplex S.dysenteriae 1 поглощались клетками ВНК с одинаковым видом окрашенной цитоплазмы для обоих препаратов, подтверждая, что InvaplexAR сохраняет способность стимулировать поглощение клетками млекопитающих (фиг.4). Это показывает, что объединение рекомбинантных IpaB и IpaC с LPS S.dysenteriae 1 в комплекс, выделенный с помощью ионообменной FPLC, приводит к получению синтетического продукта, который сохраняет такую же биологическую активность, что и нативный комплекс.
Иммуногенность, адъювантность и защитная эффективность вакцин, содержащих нативный и синтетический Invaplex
Модели легочной инфекции мышей [31] и кератоконъюнктивита морских свинок [44] использовали для определения, стимулирует ли интраназальная иммунизация с InvaplexAR S.flexneri 2а защитный иммунный ответ, который эффективен против заражения гомологичным S.flexneri 2а. Отдельные исследования на мышах применялись для определения, может ли защитить иммунизация с InvaplexAR S.sonnei от заражения гомологичным S.sonnei и от заражения гетерологичным S.flexneri 2а. Дополнительные исследования проводили для оценки адъювантной активности InvaplexAR, используя овальбумин (OVA) в качестве иммуногена. В экспериментах на адъювантность иммунные ответы, вызванные после иммунизации только OVA, сравнивали с иммунными ответами, вызванными OVA в комбинации с или с нативным Invaplex, InvaplexAR или с токсином холеры установленного мощного мукозного адъюванта. Эксперименты на морских свинках и мышах проводили в WRAIR при использовании протокола IACUC.
Эксперименты на мышах
Четыре отдельных эксперимента на животных были выполнены для оценки иммуногенности, адъювантности и защитной эффективности синтетического Invaplex. Использовали традиционные способы в каждом эксперименте и описывали в следующих параграфах. Методы и материалы, специфичные для каждого исследования, указаны в отдельных разделах.
Иммунизации и заражение
Мышей иммунизировали в 0, 14 и 28 день. Общий объем антигена 25 мкл вводили по 5-6 небольших капель, которые закапывали в наружные ноздри с помощью микропипетки. Кровь брали из хвоста, отбирая у мышей на дни 0, 28 и 42 и через 2 недели после заражения (день 63). На день 35 или 42 мышам делали эвтаназию с последующим немедленным сбором мукозных смывов и удаляли селезенку для анализа пролиферации и образования цитокинов in vitro InvaplexAR S.flexneri 2a. Через три недели после конечной иммунизации мышей (15 на группу) заражали интраназально летальной дозой (1,6×107 коэ) S.flexneri 2а (2457Т) или S.sonnei (Mosely, 8×106 коэ), как описано для модели инфицирования легких мышей [34]. До интраназальной иммунизации или заражения мышей анестезировали смесью гидрохлорида кетамина (40 мг/кг) и ксилазина (12 мг/кг). Зараженных мышей ежедневно контролировали в течение двух недель для определения потери веса, взъерошенной шерсти и сонливости, а также смерти, использующейся в качестве конечной точки. Выжившие мыши кровоточили 14 дней после заражения (день 63).
Исследование иммуногенности и защиты InvaplexAR S.flexneri 2а
Отдельные группы мышей-самок Balb/c (10-15 мышей/группу) иммунизировали интраназально на день 0, 14 и 28 либо 5 мкг нативного Invaplex S.flexneri 2а (серия JWJX) или 2,5 мкг синтетического InvaplexAR S.flexneri 2а (Серия KF-D3D4). Контрольными животными являлись животные, привитые физиологическим раствором. Кровь собирали на день 0, 28, 42 и 63. Животных заражали интраназально на день 49.
Исследование иммуногенности и защиты InvaplexAR S.flexneri 2а II
Отдельные группы мышей-самок Balb/c (15-20 мышей/группу) иммунизировали интраназально на день 0, 14 и 28 либо 5 мкг нативного Invaplex S.flexneri 2а (серия JWJX) или 2,5 мкг синтетического InvaplexAR S.flexneri 2а (серия KR-C5). Контрольными животными являлись животные, привитые физиологическим раствором. Кровь собирали на день 0, 28, 42 и 63. Смывы из легких и кишечника собирали на день 42 у 4 животных из группы. Оставшихся животных заражали интраназально на день 49.
Изучение адъювантности InvaplexAR S.flexneri 2а
Способность InvaplexAR функционировать как мукозный адъювант определяли у мышей, используя овальбумин (OVA) в качестве модели антигена протеина. Мышей Balb/cByJ (5 мышей/группу) интраназально иммунизировали на день 0, 14 и 28 только с OVA (5 мкг) или OVA (5 мкг) в комбинации либо с InvaplexAR S.flexneri 2а (серия KF-D3D4; 2,5 мкг), нативным Invaplex S.flexneri 2а (Серия JWJX; 5 мкг) либо с холерным токсином (СТ; 5 мкг). Кровь собирали на день 0, 28 и 42. Контрольных животных прививали физиологическим раствором. Смывы из легких и кишечника собирали на день 42. Сывороточные титры в конечной точке на день 0, 28 и 42 со специфичностью для Invaplex 24, Invaplex 50, LPS, OVA, IpaB и IpaC определяли с помощью ELISA. Антиген-специфичные антитела в смывах со слизистых оболочек также оценивали.
Исследование иммуногенности и защиты InvapIexAR S.sonnet
Иммуногенность и защитную эффективность InvaplexAR S.sonnei определяли у мышей, используя гомологичное (S.sonnei 53G) и гетерологичное (S.flexneri 2а 2457Т) заражения. Мышей Balb/cByJ (10-15 мышей/группу) интраназально иммунизировали на день 0, 14 и 28 InvaplexAR S.sonnei (серия KJ-D3D6; 2,5 мкг), нативным Invaplex S.sonnei (Серия JOJP; 5 мкг) или нативным Invaplex S.flexneri 2а (Серия JWJX; 5 мкг). Контрольных животных прививали физиологическим раствором. Кровь собирали на день 0, 28 и 42.
Эксперименты на морских свинках
Морских свинок (вид Hartley, 6-10 на группу) иммунизировали интраназально вакцинами, содержащими синтетический или нативный Invaplex (25 мкг/дозу). Дозовый объем (100 мкл) распределяли равномерно между каждой ноздрей. Физиологический раствор использовался для иммунизации контрольных животных. Морских свинок иммунизировали на дни 0, 14 и 28, и кровь из латеральной ушной вены кровоточила на дни 0, 28, 42 и в течение 14 дней после заражения. Перед интраназальной иммунизацией морских свинок анестезировали смесью из кетамина (40 мг/мг) и ксилазина (4 мг/кг). Через три недели после третьей иммунизации морских свинок заражали интраокулярно S.flexneri 2а (штамм 2457Т) и за ними наблюдали ежедневно в течение 5 дней для оценки степени воспаления и кератоконъюнктивита, как ранее описывалось [44].
ELISA
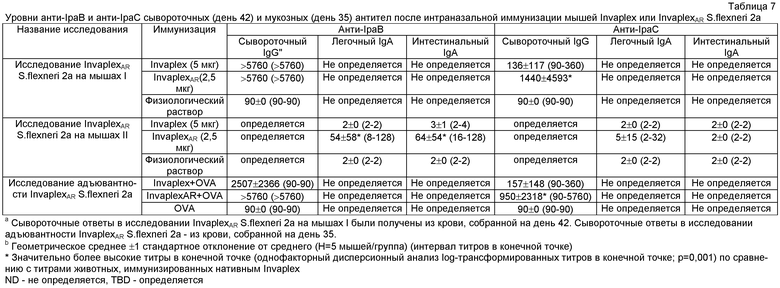
Антиген-специфичные IgA и IgG антитела в сыворотке и выделениях со слизистых оболочек, включая легочные и интестинальные лаважи и каловые выделения от иммунизированных и/или зараженных мышей и морских свинок [18, 29, 46], определяли с помощью ELISA. На 96-луночные планшеты Immunlon TJB для ELISA (ThermoLab Systems) сенсибилизировали антигенами, включая очищенный LPS S.flexneri 2а, LPS S.sonnei, экстрагированные водой антигены Shigella, очищенные белки IpaB и IpaC и нативный Invaplex S.flexneri 2а и OVA в течение ночи при 4°C. Планшеты блокировали 2% казеином (в буфере Tris-физиологический раствор, рН 7,5) и серию двукратных разведений сыворотки и смывов со слизистых инкубировали сенсибилизированными антигеном планшетами в течение 4 часов при 25°C. После промывания PBS, содержащим 0,05% Tween 20, антиген-специфическое связывание антитела определяли с помощью анти-мышиных IgG или анти-мышиных IgA антител, конъюгированных с щелочной фосфотазой (Kirkegaard & Perry). Активность щелочной фосфатазы определяли с использованием п-нитрофенилфосфата в качестве субстрата (1 мг/мл). После 30-мин инкубации с субстратом измеряли оптическую плотность (OD405), используя планшет-ридер для ELISA (Molecular Devices, Menlo Park, CA). Для каждого животного определяли титры в конечной точке и использовали для вычисления средних геометрических титров для каждой группы в каждый момент времени. Обычно животные, интраназально иммунизированные Invaplex, отвечали 4-8-кратным повышением сывороточных (IgG и IgA) и мукозных (легкие и кишечник) IgA тиров на антигены Shigella по сравнению с преиммунными образцами или контрольными животными, иммунизированными физиологическим раствором.
Анализ общего IgA
Концентрацию общего IgA в мукозных образцах определяли с помощью иммуноферментного анализа. Концентрации образцов определяли по стандартным кривым, используя очищенный мышиный IgA, анализируемый параллельно. Специфичную активность мукозного IgA рассчитывали путем деления значения титра в конечной точке для каждого отдельного мукозного образца на значение концентрации общего IgA в том же образце [47].
Антигенная стимуляция культивированных лимфоцитов из мышей, иммунизированных InvaplexAR
Пролиферацию лимфоцитов при стимуляции антигена определяли, используя спленоциты, выделенные из Invaplex-иммунизированных мышей и нативных мышей. Мононуклеарные клетки (2×105 на лунку) инкубировали с Invaplex, IpaB или IpaC или препаратами LPS S.flexneri 2а. Одновременно в отдельных лунках спленоциты стимулировали конканавалином A в качестве положительного контроля. Отрицательные контроли включали клетки, инкубированные в полной среде, и клетки от нативных мышей, стимулированные антигеном. После инкубации антигена в течение 4-7 дней клеточную пролиферацию измеряли путем нерадиоактивного определения активности дегидрогеназы, используя MTS (Promega) [48]. Наблюдалась сильная корреляция с увеличением оптической плотности индикаторов и количества жизнеспособных клеток на лунку. Для определения цитокинов до измерения пролиферации на 3 и 5 дни собирали супернатанты клеточных культур (см. ниже). Индексы стимуляции (SI) рассчитывали путем деления средней оптической плотности антиген-стимулированных клеток на среднюю оптическую плотность клеток, стимулированных только средой. SI клеток из мышей, иммунизированных Invaplex, сравнивали с SI клеток из неиммунизированных мышей.
Антиген-специфичный системный иммунный ответ после интраназальной иммунизации InvaplexAR
Иммуногенность синтетического Invaplex (InvaplexAR) S.flexneri 2а, изготовленного из отдельных очищенных компонентов, а не вирулентного организма (нативный Invaplex) оценивали у мышей. Группы мышей (n=6-10) были интраназально иммунизированы на день 0, 14 и 28 нативным Invaplex 24 S.flexneri 2а (5 или 10 мкг), InvaplexAR (2,5 мкг), очищенным IpaB (2,5 мкг), очищенным IpaC (2,5 мкг), LPS (2,5 мкг) или физиологическим раствором. Через три недели после третьей иммунизации (день 49) мышей заражали Shigella flexneri 2а (2457Т). Кровь, отобранная на день 0, 28, 42 и 63, анализировали с помощью ELISA для определения реакций сывороточных IgG и IgA на Invaplex 50, Invaplex 24, очищенного LPS, IpaB и IpaC. Фиг.5 и 6 отображают титры сывороточных IgG и IgA в конечной точке, определенные в крови, отобранной на день 42 (через две недели после третьей иммунизации).
Привитые физиологическим раствором мыши и преиммунная сыворотка от иммунизированных мышей не содержали определяемые уровни антиген-специфичных антител (данные не показаны). Иммунизация InvaplexAR S.flexneri 2а вызывала иммунные ответы, определяемые по сывороточным IgG и IgA, на Invaplex 50 и Invaplex 24 на сопоставимом уровне с той реакцией, которая была индуцирована нативным Invaplex 24 (фиг.5), и были значительно выше (P<0,001) по сравнению с теми, которые были вызваны физиологическим раствором. Двукратное увеличение количества нативного Invaplex (серия JWJX), использующегося для иммунизации, не привело к увеличению уровня Invaplex 50, LPS или Invaplex 24-специфичного сывороточного IgG или IgA ответов, измеренных на день 42 (фиг.5). Иммунизация очищенным IpaB вызывала сильный IpaB-специфичный (фиг.6) и Invaplex 24-специфичный ответ (GMT>5760 и 3800 соответственно) и умеренный ответ на Invaplex 50 (GMT 950). Иммунизация очищенным LPS не вызывала определяемый по сывороточным IgG или IgA ответ на любой антиген, использующийся в анализе ELISA, у большинства (5/6) мышей в вакцинируемой группе. Аналогично, иммунизация очищенным IpaC также не вызывала определяемый иммунный ответ на любой анализируемый антиген, включая IpaC-специфичный ELISA. Примечательно, мыши, иммунизированные InvaplexAR, имели среди прочих наивысшие IpaC-специфичные титры в конечной точке (фиг.6 и 8), значительно выше (p<0,001), чем те, которые вызывали после иммунизации нативного Invaplex.
Защита мышей от летального заражения S.flexneri после интраназальной иммунизации InvaplexAR
Летальная модель легких включает интраназальную прививку мышам летальной дозы Shigellae, которая вызывает развитие инфекции в легких, приводя к резкой потере веса, пневмонии и в конечном счете смерти через двухнедельный период наблюдения. Через три недели после интраназальной иммунизации InvaplexAR S.flexneri 2а или нативным Invaplex мышей заражали летальной дозой S.flexneri 2a (штамм 2457Т). У нативных мышей (которым вводили физиологический раствор) 11 из 13 мышей умерли с максимальной потерей веса в среднем на 31,4% от веса перед заражением (см. таблицу 1). Все мыши, иммунизированные InvaplexAR S.flexneri 2а (p<0,001) или нативным Invaplex (p<0,001), пережили летальное заражение S.flexneri 2а. Что касается потери веса (который является наиболее чувствительным индикатором защитного иммунитета), то мыши, иммунизированные InvaplexAR, потеряли меньше от их веса, измеренного перед заражением (21,3%) по сравнению с потерей веса на 23,8%-26,0% у нативных Invaplex-иммунизированных мышей. Кроме того, день восстановления веса (показатель восстановления после заражения) был днем 7 для InvaplexAR и днем 13 (нативный Invaplex, 5 мкг) или днем 7 (нативный Invaplex, 10 мкг).
В дополнение, защитная способность отдельных компонентов (IpaB, IpaC и LPS), используемых для конструирования InvaplexAR, также оценивали на мышиной летальной модели легких (см. таблицу 1). Иммунизация очищенным LPS и очищенным IpaC не защищала мышей от смерти (оба P>0,05) и, хотя иммунизация IpaB защищала 5 из 6 мышей от летального заражения, мыши никогда не восстанавливали вес и оставались симптоматичными (низкий вес, взъерошенная шерсть) до конца периода наблюдений. Максимальная потеря веса после заражения в среднем составляла 29,9% для LPS, 31,0% для IpaB и 33,9% для IpaC, все из которых были очень близки к значению потери веса у нативных мышей - 31,4%.
Результаты заражения мышей, иммунизированных InvaplexAR, смертельной дозой S.flexneri 2а показывают, что InvaplexAR стимулирует степень защиты, которая сравнима с или превышает таковую для нативного Invaplex. Кроме того, представляется, что комплекс из IpaB, IpaC и LPS необходим для таких отдельных компонентов, которые не способны стимулировать полностью эффективный защитный иммунный ответ.
Антиген-специфичный мукозный иммунный ответ после интраназальной иммунизации InvaplexAR
Смывы со слизистых кишечника и легких, собранные на день 35 от мышей, иммунизированных InvaplexAR S.flexneri 2а или нативным Invaplex, анализировали с помощью ELISA для определения уровней антиген-специфичных IgA. Иммунизация InvaplexAR индуцировала уровни LPS и Invaplex-специфичного интестинального IgA, которые были сравнимы с уровнями, индуцированными иммунизацией нативным Invaplex (фиг.7), и были значительно выше уровней (p<0,001) IpaB-специфичного IgA. Минимальный уровень IpaC-специфичного интестинального IgA устанавливался после иммунизации с любыми вакцинными препаратами Invaplex (таблицы 7 и 8).
Интраназальная иммунизация InvaplexAR вызывала сильную иммунную реакцию в легких, направленную на LPS, Invaplex 50, IpaB и IpaC (фиг.8 и таблицы 7 и 8). Минимальные уровни LPS-специфичного IgA индуцировались в легких после иммунизации нативным Invaplex с неопределяемыми уровнями антител, специфичных для Invaplex 50, IpaB и IpaC.
Антиген-специфичный клеточный пролиферативный ответ и профили секретируемых цитокинов после интраназальной иммунизации InvaplexAR
Спленоциты, собранные от иммунизированных мышей на день 35, стимулировали in vitro с использованием Invaplex 24, IpaB или IpaC для оценки индукции антиген-специфичных клеточно-опосредованных ответов. Пролиферация клеток после инкубации с антигеном указывала на предварительную память о воздействии и иммунологическую память. Конкавалин A (ConA), который неспецифично активирует Т-клеточную пролиферацию, использовался в качестве положительного контроля для демонстрации уровней жизнеспособных клеток. Индексы стимуляции (SIs) после стимуляции ConA варьировали от 13,8 до 15,9 (таблица 2). Клетки от животных, привитых физиологическим раствором, не пролиферировали после инкубации с Invaplex, IpaB или IpaC. Спленоциты от животных (4/4), иммунизированных InvaplexAR, пролиферировали после инкубации с Invaplex (SI=10,2), IpaB (SI=8,7) и IpaC (SI=6,9), показывая иммунологическую память к присутствующим антигенам. IpaB и IpaC-специфичные пролиферативные ответы в группах, иммунизированных InvaplexAR, были значительно выше (P<0,01), чем пролиферативные ответы в группах, иммунизированных нативным Invaplex. Спленоциты от 4/4 животных, иммунизированных нативным Invaplex, пролиферировали после инкубации с Invaplex (SI=6,8). Низкие до неопределяемых уровни пролиферации возникали в спленоцитах от мышей, иммунизированных нативным Invaplex, после ex vivo инкубации с IpaB (1/4 мышей) или IpaC (0/4 мышей).
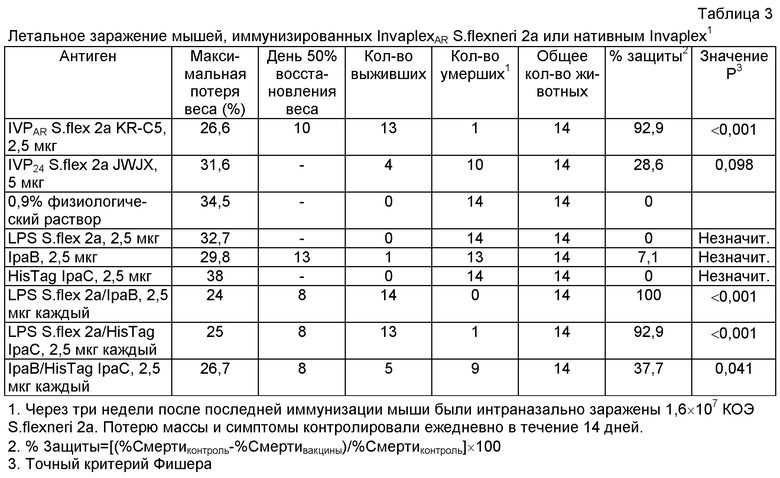
Подтверждение защиты мышей от летального заражения S.flexneri после интраназальной иммунизации InvaplexAR с использованием другой серии InvaplexAR
Эксперимент, оценивающий защитную способность InvaplexAR в мышиной летальной модели легких, повторяли с использованием другой серии InvaplexAR S.flexneri 2а. В дополнение, потенциальную оценку очищенных компонентов повторяли на мышах, иммунизированных смесью из двух очищенных компонентов (IpaB+IpaC; IpaB+LPS; IpaC+LPS). Для такого заражения мышей прививали интраназально с немного более высокой дозой заражения (1,6×106 КОЭ) S.flexneri 2а (штамм 23457Т). У нативных мышей (которым вводили физиологический раствор) 14 из 14 мышей умерли с максимальной потерей веса в среднем на 34,5% от веса до заражения (см. таблицу 3). Мыши, иммунизированные InvaplexAR S.flexneri 2а (13 из 14 выживших; р<0,001), пережили летальное заражение S.flexneri, тогда как мыши, иммунизированные нативным Invaplex, имели значительно более низкий уровень выживаемости для дозы заражения, использующейся в этом эксперименте (см. таблицу 3). Что касается потери веса у мышей, иммунизированных InvaplexAR, то они потеряли меньше от их веса до заражения (26,6%) по сравнению с 31,6% потери веса у мышей, иммунизированных нативным Invaplex, и 34,5% у мышей, привитых физиологическим раствором.
Защитная способность отдельных компонентов (IpaB, IpaC и LPS) или пары очищенных компонентов, используемых для конструирования InvaplexAR, подтвердила предыдущие результаты, что IpaC и LPS не являются защитными. Очищенный IpaB также не являлся защитным (см. таблицу 3). Смеси из двух очищенных компонентов приводили к защите в комбинации IpaB+LPS и комбинации IpaC+LPS, тогда как комбинация IpaB+IpaC была не полностью защитной.
Результаты второго эксперимента, оценивающего защитную способность InvaplexAR, четко показывают, что InvaplexAR стимулирует степень защиты, которая превышает степень защиты нативного Invaplex. Кроме того, представляется, что отдельные компоненты (IpaB, IpaC или LPS) неспособны стимулировать полностью эффективный защитный иммунный ответ.
Исследование иммуногенности InvaplexAR S.sonnei и защиты у мышей
Сывороточные иммунные ответы против Invaplex 50 S.sonnei, LPS S.sonnei, IpaB и IpaC определяли с помощью ELISA. Мыши, привитые физиологическим раствором (отрицательный контроль), не имели определяемых уровней антиген-специфичных сывороточных IgG или IgA на день 42 (таблица 4). Аналогично, титры сывороточных IgG и IgA, специфичных к Invaplex 50 S.sonnei и LPS, в конечной точке достигали значений после иммунизации InvaplexAR S.sonnei и нативным Invaplex (таблица 4). Группы мышей, иммунизированные InvaplexAR S.sonnei, имели более высокие уровни IpaB-специфичных сывороточных IgG (GMT>5760, P<0,001) по сравнению с группами мышей, иммунизированными нативным Invaplex S.sonnei (GMT 546). Животные, иммунизированные InvaplexAR, имели средний титр анти-IpaC сывороточных IgG 1091 (Р<0,001), тогда как животные, иммунизированные нативным Invaplex или физиологическим раствором, имели неопределяемые уровни IраС-специфичных IgG.
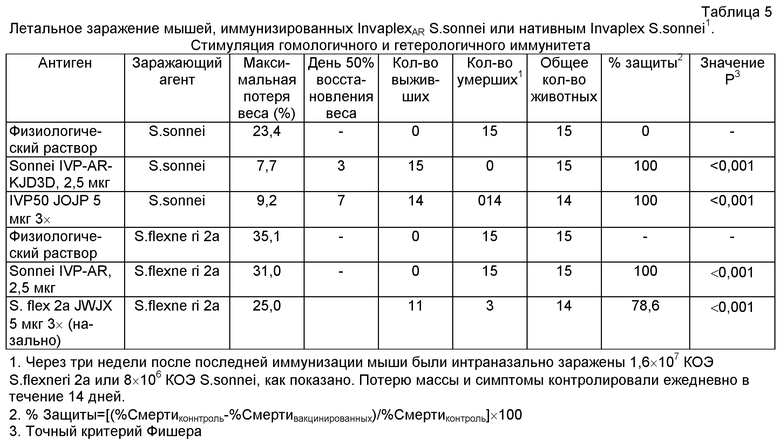
Защита мышей от летального заражения S.sonnei или гетерологичного S.flexneri после интраназальной иммунизации InvaplexAR S.sonnei
Защитная способность InvaplexAR была также оценена для InvaplexAR, изготовленного из очищенного LPS S.sonnei и рекомбинантных IpaB и IpaC, используя мышиную летальную модель легких. Ее сравнивали с защитной способностью нативного Invaplex S.sonnei. В дополнение, также оценивали способность InvaplexAR S.sonnei защищать от заражения гетерологичным S.flexneri. У нативных мышей (которым вводили физиологический раствор) 15 из 15 мышей умерли с максимальной потерей веса в среднем на 23,4% от веса перед заражением (см. таблицу 5). Мыши, иммунизированные InvaplexAR S.sonnei, пережили (15 из 15 выживших; р<0,001) летальное заражение S.sonnei, тогда как мыши, иммунизированные нативным Invaplex, также проявляли твердую защиту (14 из 14 выживших, Р<0,001) (см. таблицу 5). Что касается потери веса, то мыши, иммунизированные InvaplexAR S. sonnei, потеряли 7,7% от их веса перед заражением по сравнению с потерей веса на 9,2% у мышей, иммунизированных нативным Invaplex S.sonnei, и на 23,4% у мышей, привитых физиологическим раствором.
Примечательно, InvaplexAR S.sonnei также обеспечивал значительную защиту (15 из 15 зараженных мышей выжили, (Р<0,001)) против заражения гетерологичным S.flexneri 2а, наводя на мысль, что иммунный ответ на либо IpaB, либо IpaC может отражать в значительной степени защитный иммунный ответ.
Результаты этих экспериментов, оценивающих защитную способность InvaplexAR S.sonnei, четко показывают, что InvaplexAR среди других видов Shigella стимулирует степень защиты, которая сравнима со степенью защиты нативного Invaplex.
Защита морских свинок InvaplexAR S.flexneri 2а, используя модель кератоконъюнктивита у морских свинок
Модель кератоконъюнктивита у морских свинок используется для оценки вакцин Shigella и часто используется в качестве предшественников клинических испытаний на людях. Модель включает инфицирование глаз морских свинок Shigella, которая вызывает развитие инфекции в эпителии роговицы. Это осложнение (тяжелый кератоконъюнктивит) является результатом инвазии эпителия роговицы шигеллами и последующего воспалительного ответа хозяина, почти такого же, какой наблюдается в кишечном тракте человека.
В этом эксперименте морских свинок иммунизировали три раза интраназально либо InvaplexAR S.flexneri 2а либо нативным Invaplex. Также использовали контрольную группу, получавшую физиологический раствор. Через три недели после последней иммунизации животных заражали окулярно S.flexneri 2а. Оба InvaplexAR (90% защита, Р<0,001) и нативный Invaplex (100% защита, Р<0,001) обеспечивали твердую защиту (см. таблицу 6), демонстрируя, что InvaplexAR сравним с нативным Invaplex.
Изучение адъювантности (иммуногенности) InvaplexAR S.flexneri 2a на мышах
Интраназальная иммунизация только с OVA или OVA, комбинированного с СТ, или предварительно иммунизированные образцы от иммунизированных животных (день 0) не вызывали образование определяемых уровней сывороточных IgG или IgA со специфичностью к Invaplex 50 S.flexneri 2а, Invaplex 24 S.flexneri 2а (фиг.9), LPS S.flexneri 2а (фиг.10), IpaB или IpaC (фиг.11). Иммунизация с OVA, комбинированным либо с InvaplexAR, либо с нативным Invaplex, вызывала аналогичные титры сывороточных IgG и IgA, специфичных к Invaplex 50, Invaplex 24 и LPS, в конечной точке (фиг.9 и 10). Титры в конечной точке против очищенных IpaB (p<0,05) и IpaC (р<0,001) были выше у мышей, иммунизированных OVA+InvaplexAR, по сравнению с мышами, иммунизированными OVA+нативный Invaplex (фиг.11). Сравнимые уровни OVA-специфичных сывороточных IgG и IgA были индуцированы у мышей, иммунизированных OVA в комбинации с InvaplexAR, нативным Invaplex или СТ, на день 42 и были значительно выше (р<0,001), чем ответы, вызванные у мышей после иммунизации только с OVA (фиг.12).
Антиген-специфичные антитела в смывах легких также были проанализированы с помощью ELISA для изучения мукозных иммунных ответов, индуцированных после иммунизации (фиг.13). Иммунизация с OVA в комбинации с InvaplexAR вызывала аналогичные уровни LPS- и Invaplex 24-специфичных IgG и IgA в смывах из легких по сравнению с ответами после иммунизации с OVA в комбинации с нативным Invaplex и более высокие, чем уровни, индуцированные после иммунизации только с OVA или OVA в комбинации с CT. Аналогичные уровни OVA-специфичных легочных IgG и IgA были вызваны после иммунизации с OVA в комбинации с InvaplexAR, нативным Invaplex или CT, которые были значительно (Р<0,001) выше, чем те, которые были вызваны после иммунизации только с OVA. Средние уровни LPS- и Invaplex-специфичных IgA в интестинальных смывах (данные не представлены) были обнаружены в смывах от мышей, иммунизированных OVA в комбинации с InvaplexAR или нативным Invaplex, и были неопределяемыми у мышей, иммунизированных только OVA или OVA в комбинации с CT. OVA-специфичные интестинальные IgA ответы были ниже уровней обнаружения во всех исследуемых образцах.
Способ формирования и оценки синтетического Invaplex, соединенного с антибиотиком или другой лекарственной молекулой для внутриклеточной доставки
Многие лекарственные средства, включая антибиотики, часто неэффективны против внутриклеточной мишени, поскольку они не способны проникать через мембраны клеток млекопитающих или поскольку они требуют высоких внеклеточных концентраций для достижения терапевтических концентраций внутри клеток млекопитающих. Синтетический Invaplex обеспечивает механизм для создания комплекса из инвазивов, LPS и антибиотика путем смешивания компонентов во время стадии сборки для создания синтетического Invaplex. При завершении сборки нативные свойства Invaplex позволяют ему транспортировать прикрепленный антибиотик или терапевтическую молекулу в клетки млекопитающих.
Очищенные, растворенные IpaB и IpaC, каждый в своем соответствующем финальном буфере, смешивали друг с другом в молярном соотношении IpaC/IpaB 8. После перемешивания IpaB и IpaC раствор антибиотика, например гентамицина при 5-100 мкг/мл, добавляли к смеси. Затем IpaB, IpaC и раствор антибиотика медленно добавляли к сухому порошку LPS (соотношение LPS к общему белку составляет 0,56). LPS из любого вида Shigella может быть использован; для описываемых экспериментов использовали LPS S. flexneri 2а, S. sonnei или S. dysenteriae 1. Смесь инкубировали при 37°С приблизительно в течение 2 часов при встряхивании. Затем смесь белок/LPS/антибиотик разбавляли 20 мМ Tris-HCl, 0,10 М NaCl и 1,2 М мочевины для снижения концентрации NaCl перед выполнением ионообменной хроматографии. Для финальной очистки разбавленную смесь IpaB/IpaC/LPS/антибиотик смесь очищали на анионообменной колонке (HiTrap(TM) Q HP) с использованием Буфера А, состоящего из 20 мМ Tris-HCl, рН 9,0, и Буфера В, состоящего из 1 М NaCl, 20 мМ Tris-HCl, рН 9,0, и ступенчатого градиента от 0% (5 объемов колонки) до 24% (10 объемов колонки) до 50% (6 объемов колонки) до 100% Буфер В (5 объемов колонки). Собирали 1 мл фракций из 1 мл колонки HiTrap(TM) Q HP при скорости элюирования 1 мл/мин. Фракции из каждого этапа анализировали на наличие IpaB, IpaC и LPS с помощью спотблоттинга. Фракции, содержащие IpaB, IpaC и LPS, представляли собой фракции синтетического Invaplex.
Эффективность комплекса синтетический Invaplex-антибиотик будет оценена по его способности убивать внутриклеточные микроорганизмы, такие как шигеллы, в культивируемых клетках. Комплекс синтетический Invaplex-антибиотик инкубировали с Shigella-инфицированными культивируемыми клетками. Через 2-24 часа количество внутриклеточной Shigella определяют путем нанесения лизатов обработанных клеток (и необработанных контрольных клеток) на планшеты с триптиказно-соевым агаром для определения уровня внутриклеточного лизиса в клетках, которым вводили комплекс синтетический Invaplex-антибиотик.
Дополнительные варианты и модификации вышеуказанного будут очевидны для специалиста в данной области техники и охватываются заявленной формулой изобретения.







Изобретение относится к медицине, а именно фармацевтике. Разработан химически стандартизированный синтетический трехкомпонентный комплекс Invaplex, включающий инвазивные белки IpaB и IpaC и липополисахарид из грамотрицательных бактерий. При этом серотип-специфичный липополисахаридный компонент может включать в себя два или более липополисахаридов, выбранных, по меньшей мере, из двух различных серотипов грамотрицательных бактерий. Разработан способ получения комплекса, включающий комбинирование IpaB и IpaC, по меньшей мере, с одним липополисахаридом, связанным с серотипом грамотрицательных бактерий, причем этап А) выполняется как в два отдельных этапа смешивания, включая смешивание IpaB и IpaC с формированием комплекса IраВ:IраС и смешивание комплекса IраВ:IраС, по меньшей мере, с одним липополисахаридом (LPS) с формированием синтетического Invaplex и его извлечением. Синтетический Invaplex применяется для иммунизации субъекта, транспортирования молекулы через мембрану клетки и обладает улучшенными качествами за счет постоянства состава и биологической активности. 8 н. и 26 з.п.ф-лы, 8 табл., 13 ил.
1. Синтетический Invaplex, включающий комплекс инвазивных белков, состоящий из комплекса IраВ:IраС, связанного с серотип-специфичным липополисахаридным компонентом из грамотрицательных бактерий, в котором инвазивный белковый комплекс получен путем смешивания высокоочищенного или рекомбинантного IpaB и IpaC с образованием комплекса IраВ:IраС, в котором IpaC и IpaB присутствуют в соотношении от 0,08:1 до 80:1, полученный путем смешивания комплекса IраВ:IраС, по меньшей мере, с одним липополисахаридом (LPS), где LPS присутствует относительно общего белка (IpaB и IpaC) в соотношении от 0,01:1 до 10:1.
2. Синтетический Invaplex по п.1, в котором серотип-специфичный липополисахаридный компонент является определяющим серотип грамотрицательных бактерий, выбранных из S.flexneri, S.sonnei, S.dysenteriae, S.boydii, enteroinvasive E.coli, Yersinia или Salmonella.
3. Синтетический Invaplex по п.2, в котором серотип-специфичный липополисахаридный компонент является определяющим серотип грамотрицательных бактерий, выбранных из S.flexneri, S.sonnei, S.boydii, S.dysenteriae или enteroinvasive E.coli.
4. Синтетический Invaplex по п.3, в котором грамотрицательные бактерии выбирают из S.flexneri, S.sonnei, S.boydii или S.dysenteriae.
5. Синтетический Invaplex no п.1, в котором LPS связан с серотипом Shigella.
6. Синтетический Invaplex no п.1, в котором IpaB является очищенным нативным IpaB или рекомбинантным IpaB.
7. Синтетический Invaplex по п.1, в котором IpaC является очищенным нативным IpaC или рекомбинантным IpaC.
8. Синтетический Invaplex по п.1, в котором синтетический Invaplex является InvaplexAR.
9. Синтетический Invaplex по п.8, в котором InvaplexAR представляет InvaplexAR S.flexneri, InvaplexAR S.dysenteriae или InvaplexAR S.sonnet.
10. Синтетический Invaplex по п.8, в котором содержание инвазивов и липополисахарида в InvaplexAR соответствует содержанию высоко очищенных из нативного Invaplex 24.
11. Синтетический Invaplex no п.8, в котором серотип-специфичный липополисахаридный компонент включает два или более липополисахаридов, выбранных из, по меньшей мере, двух различных серотипов грамотрицательных бактерий.
12. Синтетический Invaplex по п.1, в котором соотношение составляет от 0,8:1 до 20:1.
13. Синтетический Invaplex по п.12, в котором соотношение составляет приблизительно 8:1.
14. Синтетический Invaplex по п.1, в котором соотношение составляет от 0,5:1 до 5:1.
15. Синтетический Invaplex по п.14, в котором соотношение составляет от приблизительно 0,5:1.
16. Синтетический Invaplex по п.1, дополнительно содержащий метку, антибиотик, лекарство или биомолекулу.
17. Синтетический Invaplex по п.16, в котором биомолекула включает белки, ферменты, полисахарид или нуклеиновые кислоты.
18. Способ получения синтетического Invaplex, включающий
А) комбинирование IpaB и IpaC, по меньшей мере, с одним липополисахаридом, связанным с серотипом грамотрицательных бактерий, причем этап А) выполняется как два отдельных этапа смешивания, включая смешивание IpaB и IpaC с формированием комплекса IраВ:IраС и смешивание комплекса IpaB:IpaC, по меньшей мере, с одним липополисахаридом (LPS) с формированием синтетического Invaplex; и В) извлечение синтетического Invaplex.
19. Способ по п.18, в котором IpaB представляет собой r-IpaB и IpaC представляет собой r-IраС.
20. Способ по п.18, в котором IpaC и IpaB присутствуют в соотношении, выбранном из диапазона от 0,08:1 до 80:1.
21. Способ по п.18, в котором LPS относительно количества общего белка (IpaB и IpaC) присутствует в соотношении от 0,01:1 до 10:1.
22. Композиция, содержащая синтетический Invaplex по п.1 и фармацевтически приемлемый носитель.
23. Композиция по п.22, дополнительно содержащая иммуноген.
24. Композиция по п.22, в которой синтетический Invaplex присутствует в достаточном количестве для повышения или развития иммунного ответа на иммуноген.
25. Композиция по п.22, в которой количество присутствующего синтетического Invaplex достаточно для индуцирования защитного иммунного ответа.
26. Композиция по п.22, в которой синтетический Invaplex дополнительно содержит меченый детектор, антибиотик, лекарство или биомолекулу.
27. Композиция по п.26, в которой биомолекула включает белки, ферменты, полисахарид или нуклеиновые кислоты.
28. Композиция, содержащая синтетический Invaplex по п.11 и фармацевтически приемлемый носитель.
29. Применение синтетического Invaplex по п.1 для иммунизации субъекта.
30. Применение синтетического Invaplex по п.11 для иммунизации субъекта.
31. Применение синтетического Invaplex по п.1 для транспортирования молекулы через мембрану клетки.
32. Применение по п.31, в котором молекула является меченым детектором, антибиотиком, лекарством или биомолекулой.
33. Применение синтетического Invaplex по п.16 для транспортирования метки, антибиотика, лекарства или биомолекулы через мембрану клетки.
34. Применение по п.33, в котором детектируют присутствие метки, антибиотика, лекарства или биомолекулы в клетке.
| СТРУННЫЙ ОСЦИЛЛЯТОР | 1928 |
|
SU18355A1 |
| TURBYFILL ROSS К | |||
| et | |||
| al Isolation and characterization of Shigella flexneri invasin complex subunit vaccine | |||
| Infection and immunity | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| СПОСОБ ПАЙКИ ТЕЛЕСКОПИЧЕСКИХ КОНСТРУКЦИЙ | 1996 |
|
RU2094190C1 |
| Фазовый указатель для синхронно работающих аппаратов | 1929 |
|
SU23462A1 |

