ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет предварительной заявки США номер 61/057462, озаглавленной «МУЛЬТИВАЛЕНТНАЯ ВАКЦИНА ИЗ НАТИВНЫХ ВЕЗИКУЛ НАРУЖНОЙ МЕМБРАНЫ МЕНИНГОКОККОВ» и поданной 30 мая 2008 г., полное описание и содержание вышеуказанной заявки, таким образом, полностью включено в настоящий документ в качестве ссылки.
Федеральная поддержка исследования или разработки
Правительство США имеет права на настоящее изобретение.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Neisseria meningitidis является основной причиной менингита и септицемии во всем мире. Менингококковый менингит представляет собой воспаление мягких оболочек мозга, оболочки, выстилающей головной мозг и спинной мозг. Как при менингококковой септицемии, так и при менингококковом менингите поражение вызвано неконтролируемым местным или системным воспалительным ответом хозяина. Заболевание, вызываемое менингококками группы B, в настоящее время является причиной по меньшей мере половины всех вызванных менингококками заболеваний во многих странах, включая Северную и Южную Америку и Европу. Возникновение нового вирулентного клона Neisseria meningitidis группы B, известного как ET5, в Норвегии в конце 70-х годов с тех пор стало ответственным за продолжительные эпидемии в Норвегии, на Кубе, в Бразилии и Чили. Эти эпидемии вызвали серьезную опасность для общественного здравоохранения и значительно усилили попытки разработки эффективной вакцины против группы B в нескольких странах с эпидемией. Отсутствие лицензированной в США вакцины против группы B наряду со слабым эффектом вакцин, состоящих из капсульных полисахаридов A и C, на детей младше 18 месяцев препятствует принятию решения о повсеместной вакцинации детей против менингококковых инфекций.
Neisseria meningitidis разделяют на 13 серогрупп, из которых 9 вызывают инвазивное заболевание (A, B, C (C1, C1-), X, Y, W-135, Z, и L). Пять серотипов являются мишенями для разработки вакцин из-за их способности вызывать эпидемии, включая серотипы A, B, C, Y и W135, являющиеся мишенью большого числа исследований для создания вакцин.
Вакцины против серогрупп A, C, Y и W135 Neisseria meningitidis, вызывающих почти все инвазивные менингококковые инфекции, являются доступными и общепринятыми для использования с отличными результатами. Разработка подходящей вакцины против штаммов Neisseria meningitidis группы B более значительно затруднена множеством причин. Например, капсульный полисахарид, определяющий серогруппу, неэффективен и потенциально небезопасен для использования в вакцине, поскольку обладает такой же структурой, как полисиаловая кислота, обнаруженная на определенных клетках человека, а именно клетках крови.
Кроме того, дополнительным фактором невозможности создания подходящей вакцины является антигенная изменчивость и/или непостоянная экспрессия у разных штаммов группы B субкапсульных антигенов, экспонированных на поверхности, таких как белки наружной мембраны и липоолигосахарид (эндотоксин). Не было идентифицировано ни одного антигена, который бы обладал всеми характеристиками, необходимыми для создания эффективной вакцины.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В одном из аспектов настоящее изобретение относится к вакцине, содержащей нативные везикулы наружной мембраны менингококков (NOMV), полученные по меньшей мере из двух штаммов менингококков, генетически модифицированных для обеспечения широкого спектра защиты. Нативные везикулы наружной мембраны менингококков включают три различных набора антигенов на основе PorA, LOS и консервативных белков наружной мембраны; и генетически модифицированные штаммы модифицированы для обеспечения повышения безопасности путем инактивации генов lpxL1, synX и lgtA. Оба штамма менингококков могут экспрессировать LOS с различной коровой структурой LOS и иметь альфа-цепи, состоящие из глюкозы и галактозы. Каждый штамм может экспрессировать по меньшей мере два различных подтипа белков или подтипа эпитопов PorA, которые выбраны на основе подтипов PorA, преобладающих у изолятов группы B. Кроме того, вакцина может дополнительно содержать другой консервативный поверхностный белок, который, как известно, может индуцировать бактерицидные антитела, со сверхэкспрессией в каждом штамме и выбранный из группы, состоящей из вариантов 1 FHBP (GNA1870), вариантов 2 FHBP и вариантов 3 FHBP; NadA; App; NspA; TbpA и TbpB.
В другом аспекте настоящее изобретение относится к комбинации NOMV из трех генетически модифицированных штаммов Neisseria meningitidis, обладающих антигенной изменчивостью. По меньшей мере один из штаммов выбран из (1) H44/76 HOPS-DL, который обладает следующими генетическими модификациями или характеристиками: гены synX, lpxL1 и lgtA инактивированы; инсерция второго гена porA (подтип P1.7-1,1) вместо opaD; повышена экспрессия NadA; и стабилизирована высокая экспрессия Opc и PorA; (2) 8570 HOPS-GAL, обладающего следующими генетическими модификациями или характеристиками: гены synX, lpxL1 и lgtA инактивированы; инсерция второго гена porA вместо opaD; повышена экспрессия варианта 1 белка, связывающего фактор H; и стабилизирована высокая экспрессия PorA и Opc; и/или (3) B16B6 HPS-G2A, обладающего следующими генетическими модификациями или характеристиками: гены synX, lpxL1 и lgtA инактивированы; инсерция второго гена porA вместо opaD; повышена экспрессия варианта 2 белка, связывающего фактор H; и стабилизирована высокая экспрессия PorA и Opc. NOMV получают без обработки детергентом или денатурирующими растворителями из клеточной массы или из истощенной культуральной среды. Вакцину можно комбинировать с одним или несколькими адъювантами и можно вводить внутримышечно и/или интраназально.
В другом аспекте настоящее изобретение относится к композиции вакцины против менингококковой инфекции, более предпочтительно инфекции, вызванной менингококками группы B, содержащей нативные везикулы наружной мембраны менингококков (NOMV) одного или нескольких генетически модифицированных штаммов Neisseria meningitidis. Один или несколько генетически модифицированных штаммов модифицированы посредством: инактивации гена synX, инактивации гена lpxL1, инактивации гена lgtA в каждом штамме, приводящей к экспрессии укороченных или усеченных липоолигосахаридов (LOS) с отсутствием тетрасахарида лакто-N-неотетраозы, и/или инсерции по меньшей мере одного второго гена porA, отличающегося по антигенным свойствам, вместо гена opa. В другом аспекте генетически модифицированный штамм дополнительно обладает повышенной или стабильной экспрессией по меньшей мере одного минорного консервативного белка наружной мембраны, и/или стабилизированной экспрессией по меньшей мере одного белка наружной мембраны. По меньшей мере один второй ген porA, отличающийся по антигенным свойствам, может экспрессировать по меньшей мере один подтип белка или подтип эпитопа PorA, выбранный из наиболее распространенных подтипов PorA изолятов meningitidis группы B.
В другом аспекте настоящее изобретение относится к генетически модифицированному вакцинному штамму Neisseria meningitidis подтипа B. Генетически модифицированный вакцинный штамм может включать штамм H44/76 HOPS-D (B1), штамм 8570 HOS-G1 (B2) и/или штамм B16B6 HPS-G2A (B3).
В другом аспекте настоящее изобретение относится к генетически модифицированному вакцинному штамму Neisseria meningitidis подтипа B, полученному из: штамма H44/76, который имеет следующие генетические модификации: i) ген synX инактивирован, ii) ген lpxL1 инактивирован, iii) ген lgtA инактивирован, iv) инсерция второго гена porA вместо гена opaD, v) экспрессия NadA по сравнению с нативным штаммом повышена и vi) стабилизирована повышенная экспрессия белков Opc и PorA. В некоторых аспектах генетически модифицированный штамм получен из штамма дикого типа ET-5 H44/76 (B:15: P1.7,16: L,3,7:P5.5,C).
В другом аспекте настоящее изобретение относится к генетически модифицированному вакцинному штамму из штамма Neisseria meningitidis подтипа B, полученному из 8570, который обладает следующими генетическими модификациями из: i) ген synX инактивирован, ii) ген lpxL1 инактивирован, iii) ген lgtA инактивирован, iv) инсерция второго гена porA вместо opaD; v) повышена экспрессия варианта 1 белка, связывающего фактор H; и vi) стабилизирована повышенная экспрессия белков PorA и Opc. В некоторых аспектах генетически модифицированный штамм получен из штамма дикого типа ET-5 85 70(B:4: P 1.19,15: L3,7v: P5.5,11,C).
В другом аспекте настоящее изобретение относится к генетически модифицированному вакцинному штамму Neisseria meningitidis подтипа B, полученному из B16B6, который имеет следующие генетические модификации: i) ген synX инактивирован, ii) ген lpxL1 инактивирован, iii) ген lgtA инактивирован, iv) инсерция второго гена porA (подтип P1.22-1,4) вместо opaD; v) повышена экспрессия варианта 2 белка, связывающего фактор H; и vi) стабилизирована повышенная экспрессия белков PorA и Opc. В некоторых аспектах генетически модифицированный штамм получен из штамма дикого типа ET-37 B16B6 (B:2a:P 1.5,2: L2:P5.1,2,5).
В некоторых аспектах настоящее изобретение относится к генетически модифицированному штамму, выращенному на среде с дефицитом железа.
В других аспектах настоящее изобретение относится к генетически модифицированному штамму, где инактивацию гена synX, гена lpxL1 или гена lgtA осуществляют посредством введения гена устойчивости к лекарственному средству в последовательность инактивированного гена.
В другом аспекте изобретение относится к вакцине, содержащей NOMV, полученные из генетически модифицированных штаммов по настоящему изобретению. NOMV получают из клеточной массы или из использованной культуральной среды без обработки детергентом или денатурирующим растворителем. Вакцина может также содержать один или несколько адъювантов. В других аспектах генетически измененный штамм модифицирован так, что экспрессирует белки накопления железа.
В другом аспекте настоящее изобретение относится к вакцине против менингококковой инфекции, содержащей множество нативных везикул наружной мембраны менингококков (NOMV), где по меньшей мере некоторые из NOMV по существу не экспрессируют или не сиалируют липоолигосахарид (LOS), содержат LOS, который включает липид A с пента-ацильной структурой, и имеют повышенный уровень экспрессии по меньшей мере одного минорного консервативного белка наружной мембраны, где минорный консервативный белок наружной мембраны выбран из белков, индуцирующих бактерицидные антитела. Минорный консервативный белок наружной мембраны может быть выбран из группы, состоящей из Nad A, варианта 1 белка, связывающего фактор H (FHBP) и варианта 2 FHBP. В других аспектах по меньшей мере некоторые из NOMV содержат укороченный или усеченный LOS, по существу не имеющий тетрасахарид лакто-N-неотетраозу (LNnT) и/или по меньшей мере некоторые из NOMV содержат два или более различных белков PorA.
В другом аспекте настоящее изобретение относится к способу стимуляции иммунного ответа в ответ на менингококковую инфекцию у животного или человека, включающему введение композиции, содержащей NOMV по меньшей мере из одного генетически измененного штамма N. Meningitidis, животному или человеку для иммунизации против менингококковой инфекции. Вакцину используют для иммунизации против инфекции, вызываемой менингококками группы B.
В другом аспекте настоящее изобретение относится к способу получения генетически модифицированного штамма N. Meningitidis, который можно использовать в вакцине против менингококковой инфекции, включающему стадии: a) выбора штамма менингококка типа B, способного к генетической модификации; b) генетической модификации штамма путем инактивации гена synX, c) генетической модификации штамма путем инактивации гена lpxL1, d) генетической модификации штамма путем инактивации гена lgtA и e) генетической модификации штамма путем повышения экспрессии одного или нескольких консервативных белков наружной мембраны. В других аспектах изобретения способ дополнительно включает генетическую модификацию штамма путем инсерции по меньшей мере одного второго гена porA, отличающегося по антигенным свойствам, в открытую рамку считывания гена opa. В других аспектах изобретения способ дополнительно включает стадию генетической модификации штамма для получения стабильной экспрессии или сверхэкспрессии по меньшей мере одного белка наружной мембраны посредством замены поли-C последовательности внутри промотора или открытой рамки считывания по меньшей мере одного белка наружной мембраны последовательностью, содержащей нуклеотиды G и C.
В другом аспекте настоящее изобретение относится к способу получения вакцины против менингококковой инфекции, включающему стадии: a) культивирования генетически модифицированного штамма Ν. meningitidis, обладающего одной или несколькими модификациями, выбранными из группы, состоящей из ген synX инактивирован, ген lpxL1 инактивирован, ген lgtA инактивирован, введен по меньшей мере один второй ген porA с антигенными отличиями вместо гена opa, экспрессия по меньшей мере одного минорного консервативного белка наружной мембраны, повышена или стабильна, и/или стабильна экспрессия по меньшей мере одного белка наружной мембраны; b) увеличения размеров культуры посредством ферментации, используя культивированный штамм подпункта a), для инокуляции среды в ферментере; c) инактивации ферментированной культуры; d) сбора культивированных клеток N. meningitidis путем центрифугирования с непрерывным потоком и сбора клеточной массы; e) выделения ΝOMV из клеточной массы и f) ресуспендирования ΝOMV в буфере или носителе, подходящем для введения вакцины.
КРАТКОЕ ОПИСАНИЕ ФИГУР
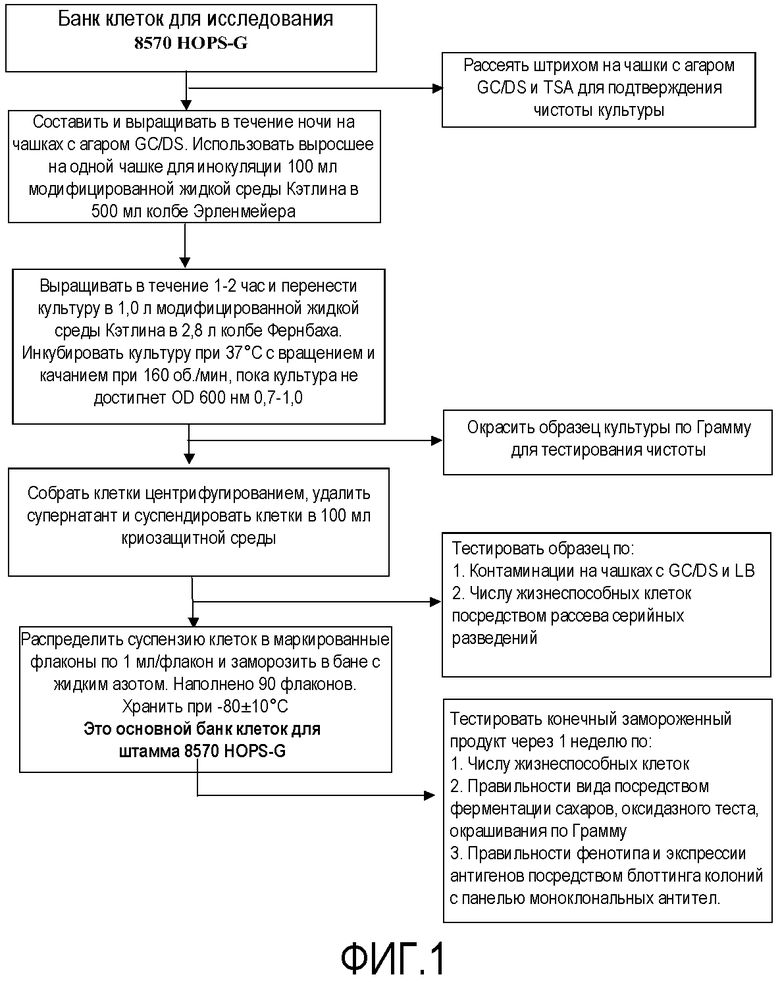
Фигура 1 представляет собой схему технологического процесса, обозначающую получение основного банка клеток из клеток для генетически модифицированных штаммов Neisseria для получения вакцины.
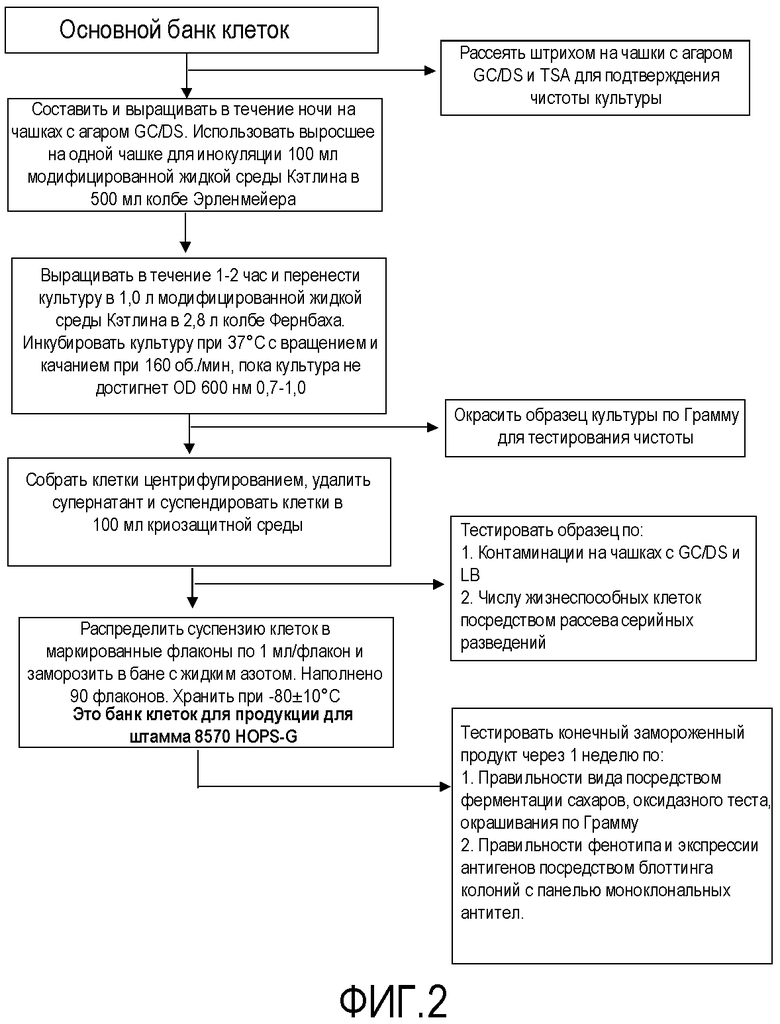
Фигура 2 представляет собой схему технологического процесса, обозначающую получение препарата банка клеток, применяемого для продукции генетически модифицированных штаммов Neisseria для получения вакцины.
Фигура 3 представляет собой схему технологического процесса, обозначающую ферментацию Neisseria, используемую для получения генетически модифицированных штаммов Neisseria для получения вакцины.
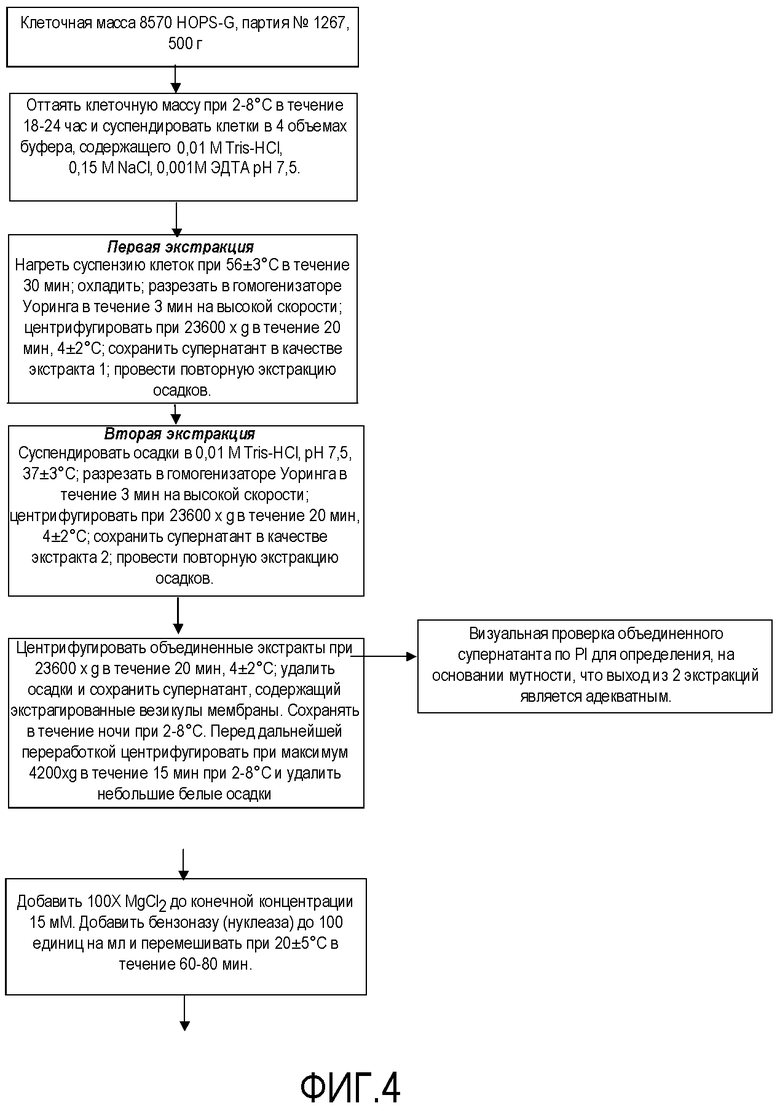
Фигура 4 представляет собой схему технологического процесса, обозначающую очистку NOMV из генетически модифицированных штаммов Neisseria для получения вакцины.
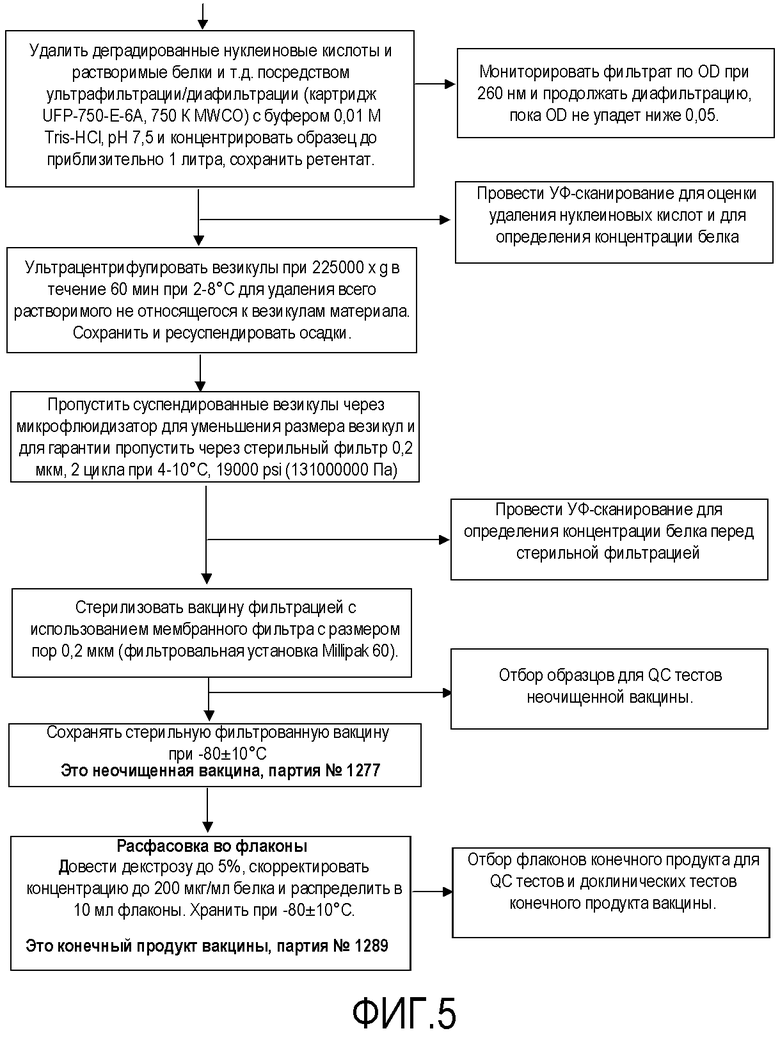
Фигура 5 представляет собой продолжение схемы технологического процесса фигуры 4.
Фигура 6 представляет собой изображение геля, окрашенного Кумасси синим, показывающего белковое содержание стандартного маркера (дорожка 1), контрольного препарата NOMV 8570 HOPS-G (дорожка 2), фильтрованной неочищенной вакцины (дорожка 3) и конечного продукта вакцины (дорожка 4).
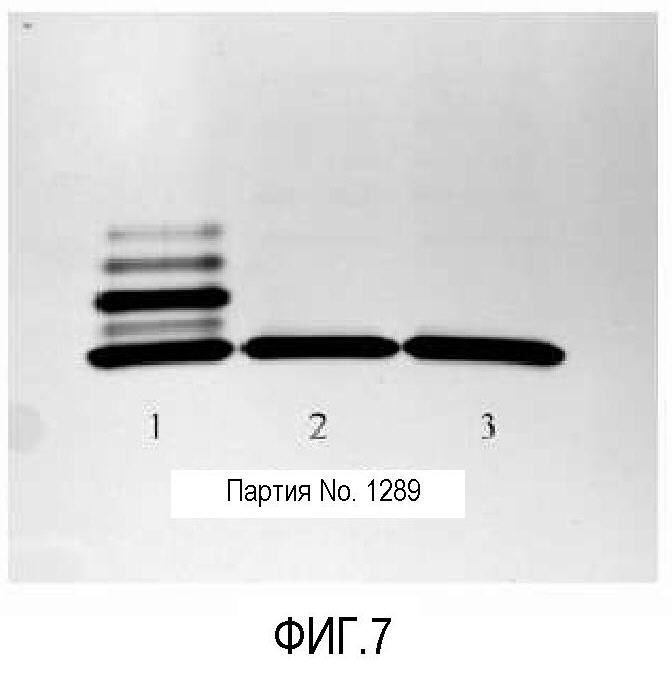
Фигура 7 представляет собой гель, окрашенный серебром, показывающий содержание липоолигосахарида в вакцине. Дорожка 1 представляет собой контрольный ML5 LPS, дорожка 2 представляет собой фильтрованную неочищенную вакцину, и дорожка 3 представляет собой конечный продукт вакцины. Пятнадцать мкл разведения 1:2 100 мкг/мл вакцины разделяли в геле (20 мкл из 100 мкл/мл 1:2 разведения контроля).
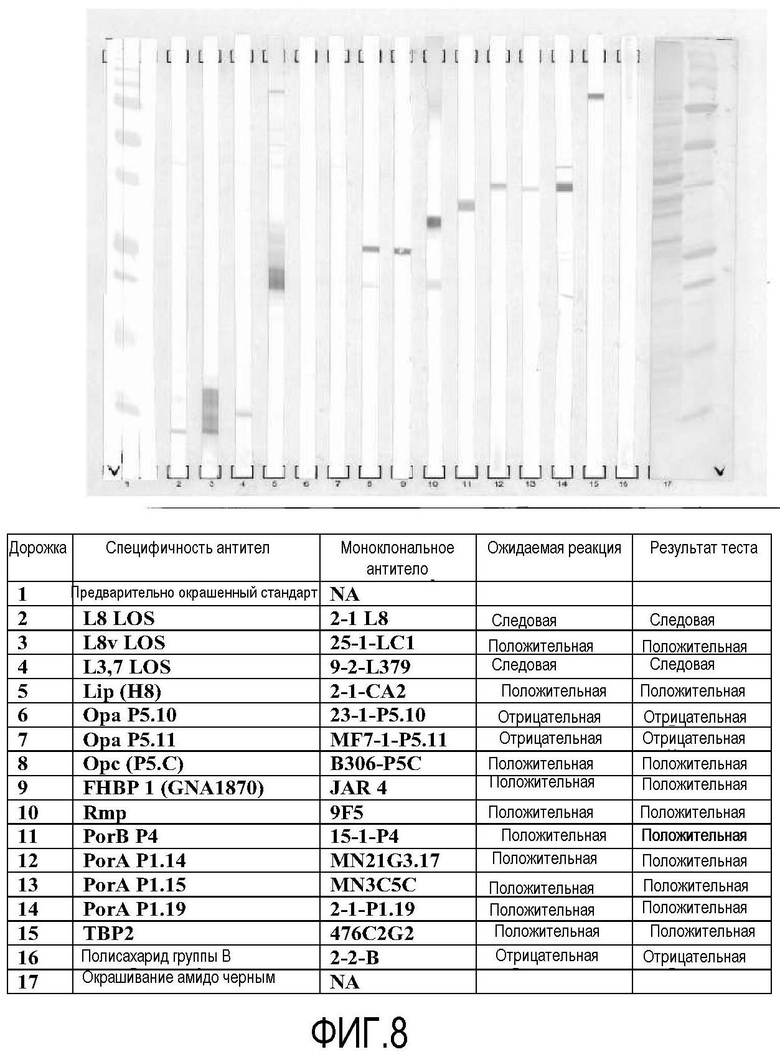
Фигура 8 представляет собой изображение окрашенного антителом Вестерн-блота, показывающего идентичность и состав белков, обнаруженных в вакцине 8570 HOPS-G NOMV.
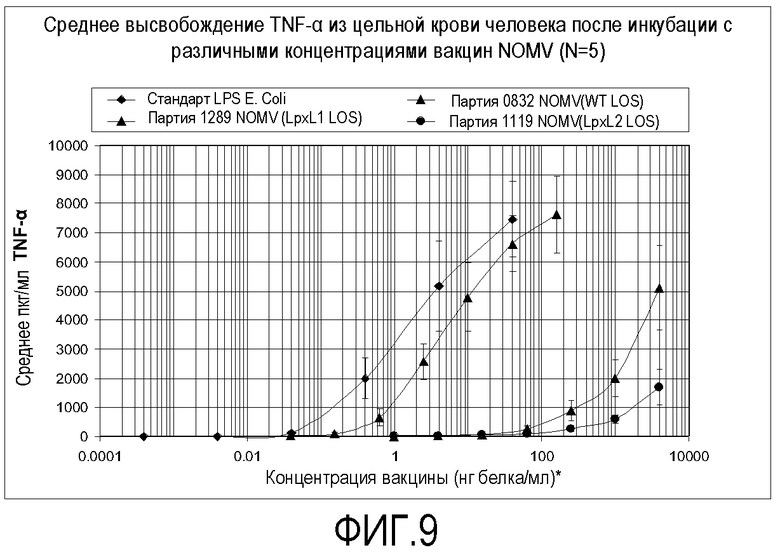
Фигура 9 представляет собой график, изображающий TNF-α, высвобождаемый из крови человека после инкубации с различными концентрациями вакцины.
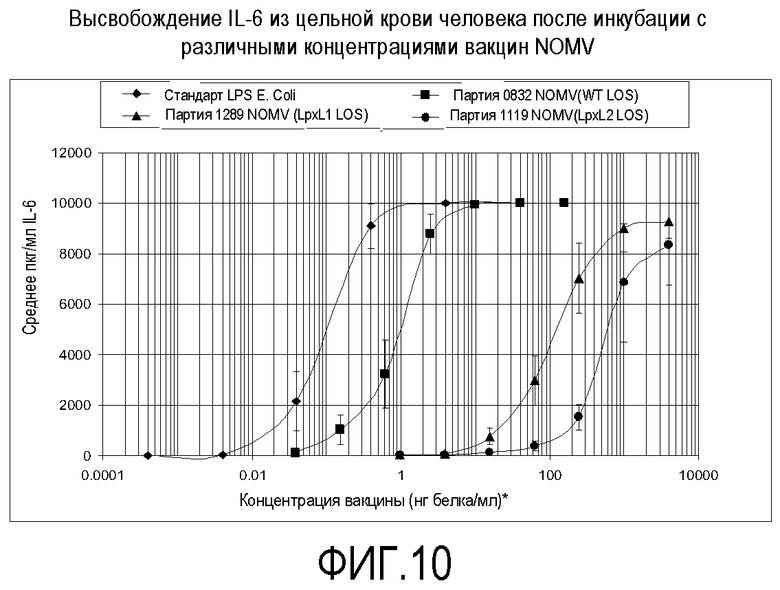
Фигура 10 представляет собой график, на котором показано высвобождение IL-6 из крови человека после инкубирования с различными концентрациями генетически модифицированной вакцины NOMV.
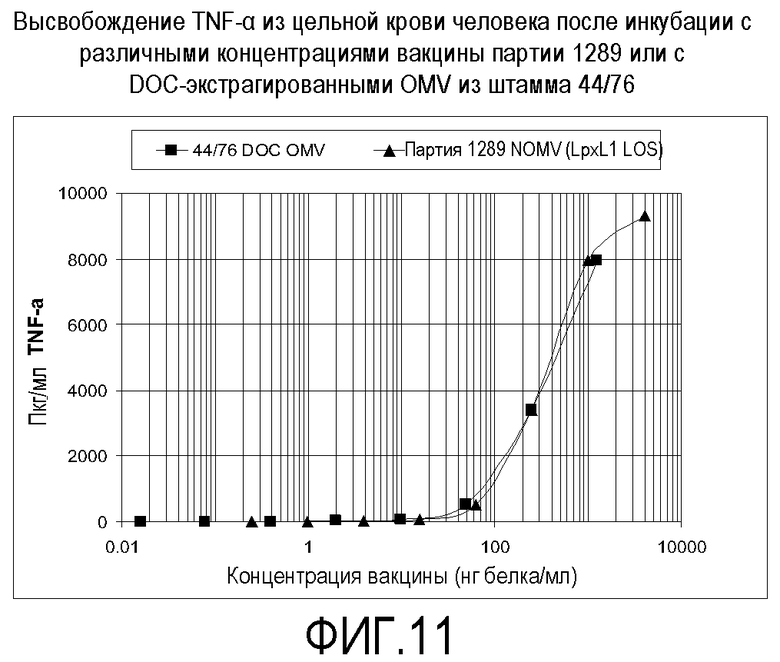
Фигура 11 представляет собой график, на котором показан TNF-α, высвобождаемый из крови человека после инкубации с различными концентрациями генетически модифицированной вакцины по сравнению с DOC-экстрагированными OMV из штамма 44/76.
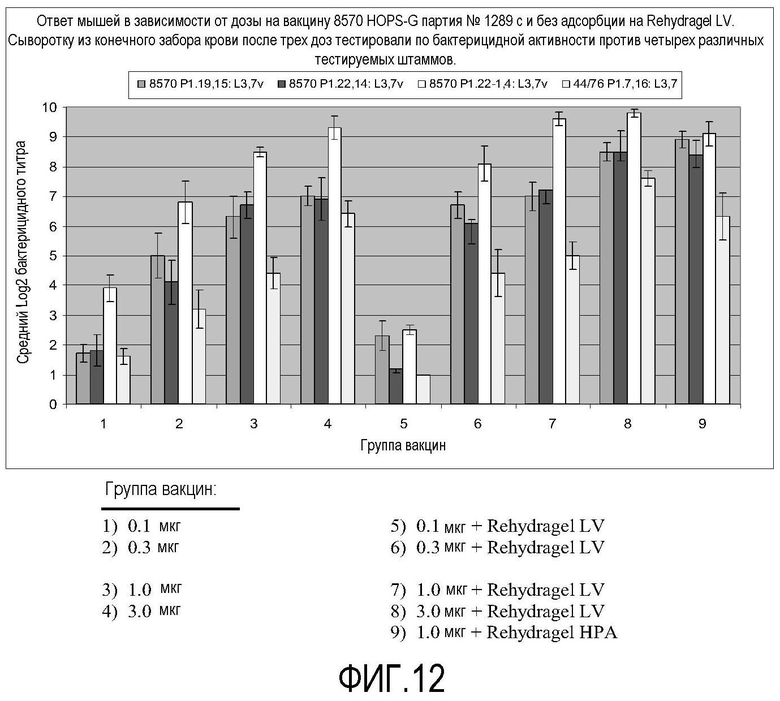
Фигура 12 представляет собой столбчатую диаграмму, на которой показаны бактерицидные титры у мышей, вакцинированных различными концентрациями вакцины 8570 HOPS-G с адъювантом или без.
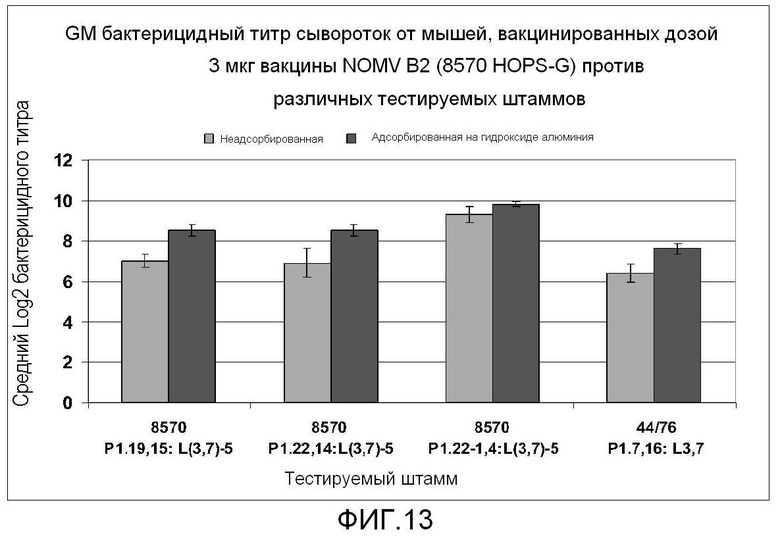
Фигура 13 представляет собой столбчатую диаграмму, на которой показаны бактерицидные титры у мышей, вакцинированных вакциной 8570 HOPS-G, против различных тестируемых штаммов.
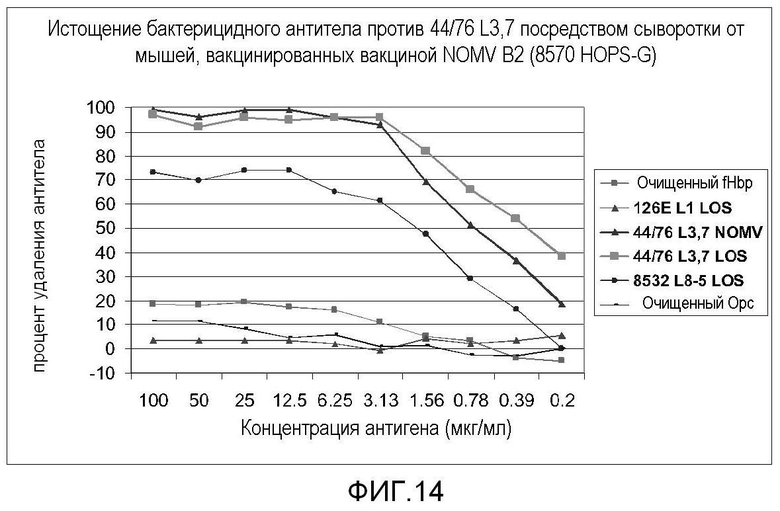
Фигура 14 представляет собой график, на котором показаны результаты анализа истощения бактерицидных антител для антигенов LOS, GNA1870, NOMV и Opc.
На фигуре 15 показан ответ антител у кроликов, вакцинированных вакциной 8570 HOPS-G NOMV с адъювантом и без.
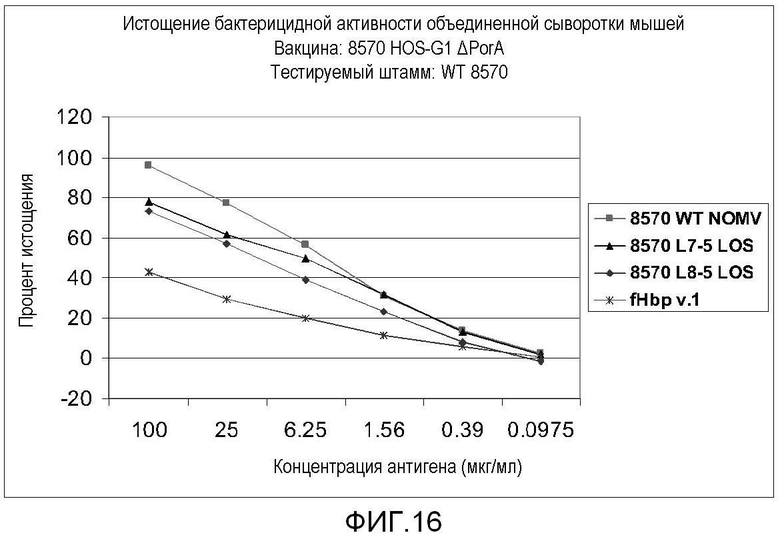
Фигура 16 представляет собой график, на котором показаны результаты анализа бактерицидного истощения для тестируемых штаммов против вакцины 8570 HOPS-G1 NOMV.
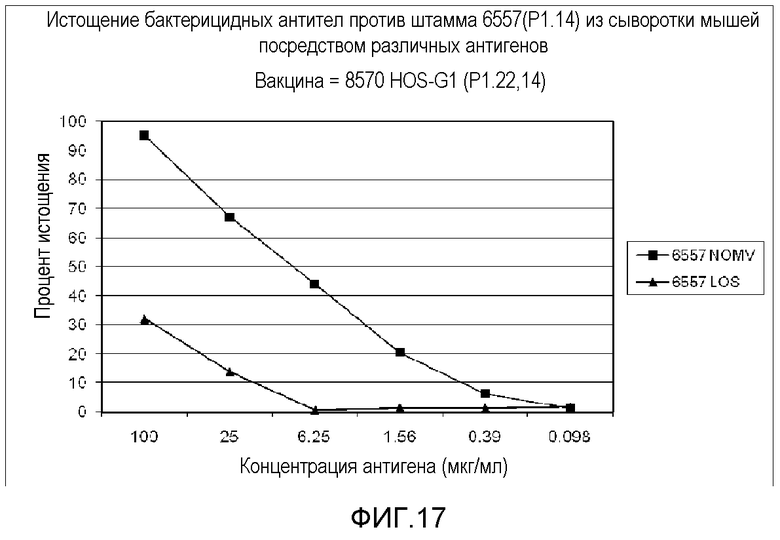
Фигура 17 представляет собой график, на котором показаны результаты анализа бактерицидного истощения для антигенов LOS и FHBP для штамма 8570 HOPS-G1 с нокаутом PorA.
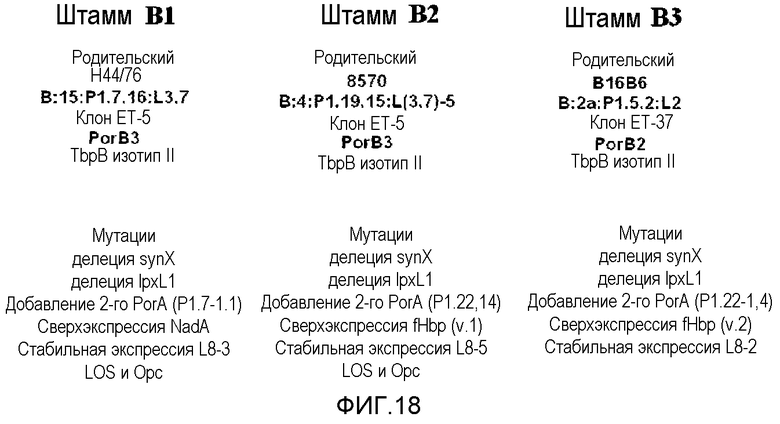
На фигуре 18 представлен фенотип трех генетически модифицированных штаммов Neisseria (A=B1, B=B2 и C=B3) по настоящему изобретению.
На фигуре 19 представлены плазмиды, используемые для конструирования генетически модифицированных штаммов Neisseria: a) плазмида, сконструированная для нокаута lgtA, b) плазмида для экспрессии второго PorA, c) плазмида для сверхэкспрессии fHbp под контролем ортологичного (Ptac в случае E. coli) промотора и d) плазмида для сверхэкспрессии NadA под контролем гомологичного промотора (промотор PorA N. meningitidis) и e) характерная схема трансформации N. meningitidis плазмидой для сверхэкспрессии fHbp (вариант 1 и 2) и ΝadA посредством замены гена NspA гомологичной рекомбинацией.
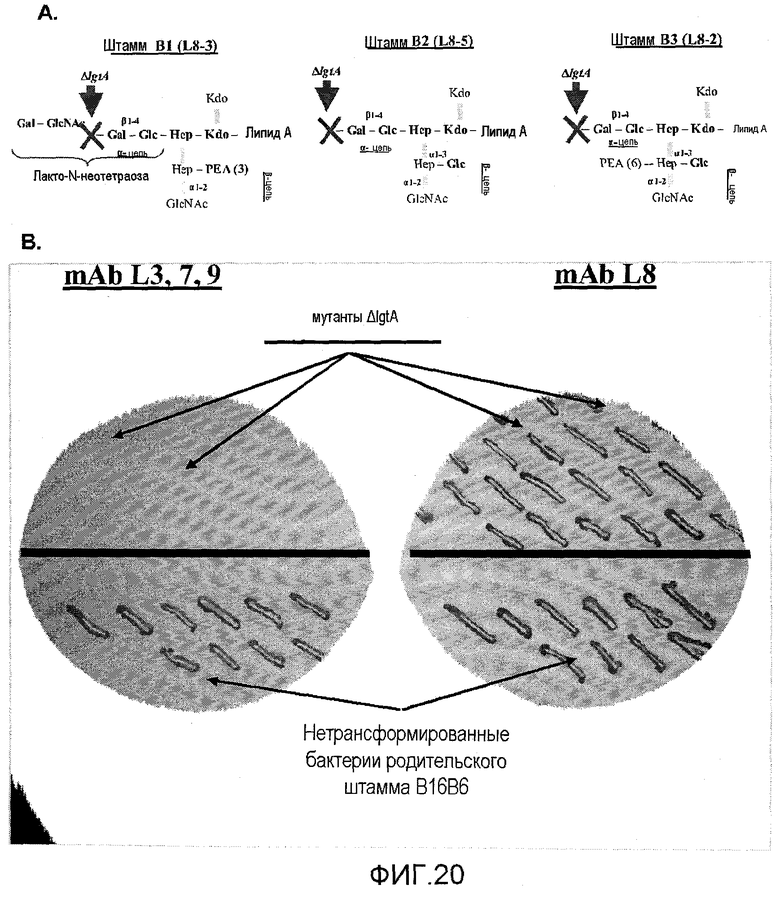
Фигура 20a представляет собой изображение стабилизации иммунотипа с усеченным LOS вакцинного штамма ΝOMV посредством нокаута гена lgtA трех генетически модифицированных штаммов. На фигуре 20b представлен иммуноблоттинг экспрессии альфа-цепи LOS генетически измененным штаммом B2 и родительским штаммом (B16B6) с моноклональными антителами против L3.7,9 (слева) и L8 (справа).
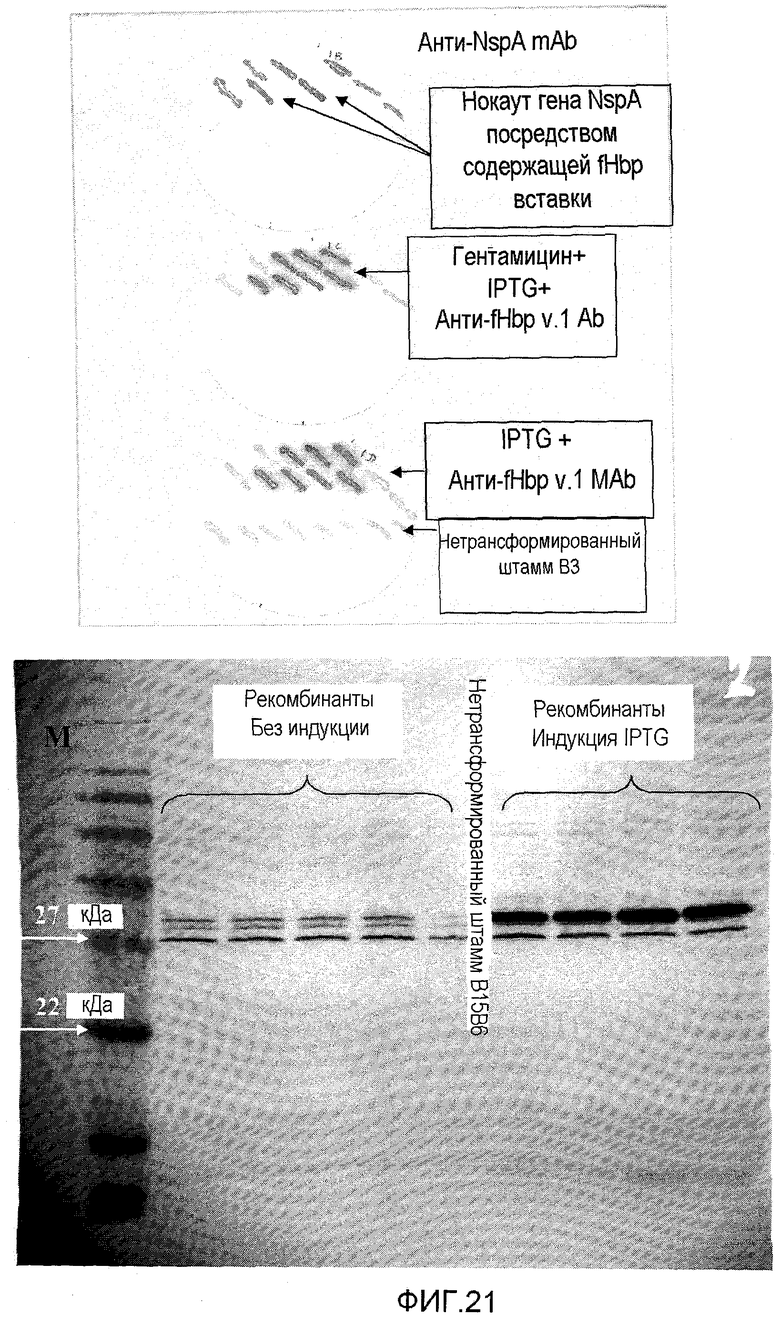
На фигуре 21 графически показана экспрессия варианта 2 fHbp в генетически модифицированном штамме B3. На фигуре 21a показан выбор штамма, содержащего устойчивый к гентамицину рекомбинант, содержащий сверхэкспрессированный fHbp, посредством иммуноблоттинга, и фигура 21b представляет собой Вестерн-блоттинг с использованием моноклонального антитела против fHbp JAR4, показывающий повышенную экспрессию fHBp.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к композиции вакцины с широким спектром защиты, используемой для иммунизации против инфекции, вызываемой менингококками, более предпочтительно подгруппой Neisseria meningitidis типа B. В одном из вариантов осуществления настоящее изобретение относится к композиции вакцины, содержащий нативные везикулы наружной мембраны менингококков (NOMV) по меньшей мере одного, предпочтительно по меньшей мере двух, более предпочтительно по меньшей мере трех генетически модифицированных штаммов Neisseria meningitidis. Нативные везикулы наружной мембраны менингококков, называемые также пузырьками, представляют собой везикулы, сформированные или полученные из фрагментов наружной мембраны грамотрицательной бактерии, которые естественным образом отделяются во время роста и которые можно получить из культуральной среды или из клеток неагрессивными способами, в которых не применяются детергенты или денатурирующие растворители. Эти NOMV, как правило, содержат белки наружной мембраны (OMP), липиды, фосфолипиды, периплазматический материал и липополисахарид (LPS), в том числе липоолигосахариды. Грамотрицательные бактерии, особенно патогены, подобные N. meningitidis, часто передают ΝOMV при вирулентном инфицировании процессом, известным как пузырение. В соответствии с настоящим изобретением ΝOMV представляют собой везикулы, полученные из наружной мембраны бактерий без использования способов химической денатурации и полученные из генетически модифицированных штаммов, которые обладают антигенным разнообразием и каждый из которых генетически модифицирован с целью повышения безопасности, антигенной стабильности и расширения защитного иммунного ответа.
В одном из вариантов осуществления настоящее изобретение относится к композиции вакцины, содержащей нативные везикулы наружной мембраны менингококков (NOMV), полученные по меньшей мере из двух или более генетически модифицированных штаммов N. meningitidis, предпочтительно по меньшей мере из трех различных генетически модифицированных штаммов.
Некоторые варианты осуществления настоящего изобретения относятся к штаммам N. meningitidis с антигенным разнообразием, предпочтительно подтипа B, содержащим по меньшей мере три генетических модификации в бактериальном геноме, более предпочтительно по меньшей мере пять генетических модификаций, более целесообразно по меньшей мере шесть генетических модификаций. Генетические модификации могут включать одну или несколько из следующих модификаций: 1) инактивацию гена synX, необходимого для биосинтеза сиаловой кислоты, что приводит к потере способности экспрессировать или сиалировать липоолигосахарид (LOS) в капсуле; 2) инактивацию гена lpxL1, что приводит к значительно меньшей токсичности LOS, имеющего липид A с пента-ацильной структурой; 3) инсерцию второго гена porA, отличающегося по антигенным свойствам, вместо генов opa (OpaC или OpaD); 4) повышение экспрессии по меньшей мере одного минорного консервативного белка наружной мембраны, где минорный консервативный белок наружной мембраны обладает способностью индуцировать бактерицидные антитела (в качестве неограничивающих примеров, ΝadA, вариант 1 белка, связывающего фактор H (FHBP), и вариант 2 (FHBP); 5) инактивацию гена lgtA в каждом штамме, что приводит к экспрессии укороченного или усеченного LOS, у которого отсутствует тетрасахарид лакто-N-неотетраозы (LNnT); и/или 6) стабилизированной экспрессии конкретных белков наружной мембраны, таких как Opc и PorA, ответственных за фазовые варианты штаммов дикого типа.
Настоящее изобретение относится к генетически модифицированным штаммам, обеспечивающим как повышенную безопасность использования, так и увеличение широты защитного ответа антител на менингококковые инфекции. В одном из вариантов осуществления генетически модифицированные штаммы обеспечивают повышенную безопасность посредством включения по меньшей мере одной из следующих мутаций в бактериальный геном: делеции гена synX, что блокирует синтез сиаловой кислоты капсида и приводит к формированию отрицательного капсульного фенотипа NOMV, делеции гена lpxL1, уменьшающей активность эндотоксина, приводя к пента-ацильной структуре липида A, и/или делеции гена lgtA, блокирующей биосинтез лакто-N-неотетраозы на липоолигосахариде (LOS), что стабилизирует усеченную структуру LOS; более предпочтительно генетически модифицированные штаммы предоставляют две из этих мутаций, наиболее предпочтительно генетически модифицированные штаммы предоставляют все три эти мутации. В другом варианте осуществления настоящего изобретения генетически модифицированные штаммы обладают увеличенной широтой защитного ответа антител из-за нацеливания по меньшей мере на один из трех наборов возможных защитных антигенов, содержащихся в NOMV. Три возможных для нацеливания антигена включают по меньшей мере одно из следующего: белок PorA, по меньшей мере один консервативный минорный белок и/или коровая структура LOS, и включают любое их сочетание. В более предпочтительных вариантах осуществления генетически модифицированный штамм нацелен по меньшей мере на два из возможных защитных антигенов, наиболее предпочтительно нацелен на все три возможных защитных антигена.
В некоторых вариантах осуществления настоящего изобретения мутацию synX- (инактивация гена synX) вводили в генетически модифицированный штамм способом, как описано в патенте США 6558677, полное содержание которого приведено в настоящем документе в качестве ссылки. В кратком изложении, плазмиду на основе pUC19, содержащую ген synX, в котором последовательность 200 п.о. заменена на ген устойчивости к канамицину, использовали для трансформации генетически модифицированного штамма. Устойчивые к Kan трансформанты отбирали и тестировали посредством ПЦР по присутствию поврежденного гена synX и по отрицательному капсульному фенотипу. Этот synX-мутант сконструирован на основании результатов и информации о последовательности, опубликованной Swartley и Stephens (Swartley and Stephens (1994) J. Bacteriol. 176: 1530-1534), которые показали, что вставка транспозона в ген synX приводит к отрицательному капсульному фенотипу. Такую же или эквивалентную мутацию можно вводить в любой поддающийся трансформации штамм N. meningitidis. Подходящая плазмида для использования для трансформации менингококков была сконструирована с использованием следующего способа. Три последовательности ДНК соединяли с использованием слияния способом полимеразной цепной реакции (ПЦР) с элонгацией перекрывающихся фрагментов (SOE) (Horton et al. (1989) Gene 77: 61-65). Три последовательности включали, в следующем порядке, начиная с 5'-конца, synXB, основания 67-681; ген устойчивости к канамицину из pUC4K (Pharmacia LKB Biotech Co.), 671-1623; и synxB, основания 886-1589. Кроме того, на 5'-конце добавляли предположительную последовательность для накопления, ACCGTCTGAA, посредством включения ее на конце праймера для ПЦР, применяемого для амплификации последовательности оснований 67-691 synXB. Полную конструкцию амплифицировали посредством ПЦР, очищали и лигировали по тупым концам в pUC19. pUC19 использовали для трансформации Escherichia coli DH5α и отбирали на LB с агаром с 50 мкг канамицина. Отбирали устойчивую к канамицину колонию, ДНК выделяли, очищали и расщепляли XbaI. Другую копию предположительной последовательности для накопления лигировали в полилинкер и полученную плазмиду опять использовали для трансформации E. coli DH5α, и проводили скрининг устойчивых к канамицину колоний посредством ПЦР по присутствию дополнительной последовательности для накопления. Плазмидную ДНК выделяли из отобранной колонии и использовали в качестве матрицы для ПЦР с использованием праймеров, амплифицирующих только часть вставки из плазмиды, исключая ген устойчивости к ампициллину, который не следует вводить в N. meningitidis. Затем амплифицированную ДНК очищали и использовали для трансформации генетически модифицированного штамма Ν. meningitidis. Мутант synX(-) Ν. meningitides отбирали по устойчивости к канамицину и подтверждали амплификацией ПЦР модифицированной области.
В некоторых вариантах осуществления настоящего изобретения ген lpxL1 инактивировали в генетически модифицированных штаммах для получения LOS с уменьшенной эндотоксичностью, экспрессированного на ΝOMV в композициях вакцины. Липид A из LOS N. meningitidis обычно представляет собой гекса-ацильную структуру и является ответственным за эндотоксические свойства LOS. Две вторичные жирные кислоты, связанные ацил-окси-ацилом, присутствующие в липиде A, являются важными для эндотоксической активности. Генетически модифицированный штамм включает мутант lpxL1, как описано van der Ley и соавторами (van der Ley, P., Steeghs, L., Hamstra, HJ., van Hove, J., Zomer, B., and van Alphen, L. Modification of lipid A biosynthesis in Neisseria meningitidis lpxL mutants: influence on lipopolysaccharide structure, toxicity, and adjuvant activity. Infection and Immunity 69(10), 5981-5990, 2001). Делеция гена lpxL1 приводила к экспрессии нормальных уровней пента-ацильного LOS с сильно уменьшенной эндотоксичностью, как тестировали и посредством теста на пирогенность у кроликов, и посредством анализа высвобождения цитокинов с использованием моноцитов человека из цельной крови. Другие способы нарушения функции гена lpxL1 предусмотрены в нижеприведенных вариантах осуществления настоящего способа для разработки генетически модифицированных штаммов.
В некоторых вариантах осуществления генетически модифицированный штамм содержит инсерцию второго гена porA, отличающегося по антигенным свойствам, вместо одного из генов opa (OpaC или OpaD). Мажорный белок наружной мембраны, порин A, или PorA, Neisseria meningitidis, является продуктом гена porA. PorA обладает широким антигенным разнообразием и является объектом фазовой изменчивости для уклонения от иммуноселективного давления; таким образом, он не всегда перекрестно взаимодействует с другими подтипами. Для повышения реактивности композиций вакцины в отношении различных подтипов PorA по меньшей мере один дополнительный ген porA встраивают в ген opaC или opaD генетически измененного штамма. Серотип PorA, отобранный для вставки, выбирают на основе наиболее преобладающих форм PorA, обнаруженных при инфекциях, вызываемых менингококками подтипа B. Подходящие серотипы PorA включают в качестве неограничивающих примеров: P1.7-1 (из штамма M1080); P1.22,14 (из штамма M4410); P1.22,1,4; или другие подходящие серотипы PorA, что понятно специалисту в данной области или как описано в современной литературе, например, как описано в Sacchi et al., Diversity and prevalence of PorA types in Neisseria meningitidis serogroup B in the United States, 1992-1998, J Infect Dis. 2000 Oct; 182(4): 1169-76. Вторые гены PorA могут находиться под контролем любого подходящего сильного промотора, обеспечивающего экспрессию белка PorA, например промотора PorA подходящих штаммов, например, штамма H44/76. Подходящие способы клонирования гена porA в генетически измененный штамм известны специалисту в данной области и могут включать в качестве неограничивающих примеров гомологичную рекомбинацию. Например, ген porA можно амплифицировать с помощью ПЦР с ДНК бактериальной хромосомы, клонировать в клонирующий вектор и затем клонировать в соответствующим образом сконструированную плазмиду, например pUC19, с использованием слияния генов посредством модификации способа ПЦР с элонгацией перекрывающихся фрагментов. Эту сконструированную плазмиду можно вводить в бактериальный геном посредством гомологичной рекомбинации, так чтобы заменить ген opa. Трансформанты можно отбирать блоттингом колоний с моноклональными антителами против порина. Эти способы известны специалисту в данной области.
В следующих вариантах осуществления настоящего способа модифицированные штаммы обладают стабильной и/или повышенной экспрессией по меньшей мере одного минорного белка наружной мембраны. Подходящие минорные белки наружной мембраны обладают способностью индуцировать бактерицидные антитела (например, в качестве неограничивающих примеров, NadA, вариант 1 белка, связывающего фактор H (FHBP), и вариант 2 (FHBP). Не ограничиваясь рамками какой-либо теории, стабилизация и/или повышенная экспрессия высоко консервативных экспонированных на поверхности минорных белков наружной мембраны, идентифицированных посредством геномного анализа, как обладающие потенциалом индуцировать защитные антитела, могут приводить к увеличению перекрестного защитного иммунного ответа. Подходящие консервативные минорные белки включают в качестве неограничивающих примеров, NadA, вариант 1 и 2 FHBP и Opc. Способы стабилизации и/или сверхэкспрессии минорного белка наружной мембраны (OMP) включают использование экспрессирующих плазмид и гомологичной рекомбинации или других подходящих способов, известных специалисту в данной области. Минорные OMP могут находиться под контролем сильного промотора, в качестве неограничивающих примеров, промотора PorA N. meningitidis или индуцируемого IPTG промотора ptac E. Coli.
Как описано в примерах ниже, сконструированные плазмиды использовали для установления повышенной экспрессии fHbp 1 и fHbp 2 в генетически модифицированных штаммах, где сверхэкспрессированный белок выглядит правильно процессированным, липидированным и транслоцированным к поверхности наружной мембраны. Например, экспрессия v.1 под контролем индуцируемого IPTG промотора Ptac E. coli в штамме 8570 HOPS-G (B2) была приблизительно в 4 раза выше, чем в родительском штамме 8570, а экспрессия v.2 в штамме B16B2 HPS-G2A (B3) была в 32-64 раза выше, чем в родительском штамме B16B6 (см. фигуру 20). Альтернативно, систему экспрессии с использованием промотора PorA можно использовать для стабилизации/сверхэкспрессии минорных консервативных белков.
В следующих вариантах осуществления настоящего способа генетически модифицированные штаммы обладают инактивацией гена lgtA, что приводит к экспрессии укороченного или усеченного LOS с отсутствием тетрасахарида лакто-N-неотетраозы (LNnT).
Важной характеристикой LOS менингококков является фазовая изменчивость, которая происходит из-за мутаций с высокой частотой в гомополимерных отрезках нуклеотидных остатков в lgtA и других генах Neisseria. Эти мутации включают или выключают экспрессию трансферазы LgtA, которая опосредует сборку α-цепи LOS (изменяя конфигурацию заместителей гептозы два). Эта активация гена lgtA с фазовой изменчивостью может приводить к нежелательному удлинению α-цепи LOS, дающему в результате лакто-N-неотетраозу, обладающую структурным сходством с антигенами клеток крови человека. Генетически модифицированные штаммы по настоящему изобретению обладают геном lgtA, нокаутированным посредством нарушения нативного гена с помощью маркера устойчивости к антибиотику или другого подходящего маркера (для использования для скрининга по изменениям в гене), в качестве неограничивающего примера, гена устойчивости к зеомицину. Способы нокаута гена lgtA известны специалисту в данной области, включая в качестве неограничивающих примеров конструирование плазмид и гомологичную рекомбинацию или трансформацию. Подвергнутый мутагенезу ген ΔlgtA являлся неактивным во всех модифицированных штаммах на протяжении по меньшей мере 22 наблюдаемых пассажей и присутствовала стабилизированная усеченная форма коровой структуры LOS. Делеция гена lgtA стабилизирует усеченную α-коровую структуру LOS, например, обеспечивая усеченные коровые структуры, как изображено на фигуре 19, где показаны три иллюстративных модифицированных штамма, например штамм B3 содержит альфа-цепь L8 с коровой структурой L3. Генетически модифицированные штаммы по настоящему изобретению содержат специфические коровые структуры LOS, соответствующие иммунотипам L3, L5 и L2, обеспечивая стабилизированные коровые структуры, на которые можно вызвать иммунный ответ, и в примерах ниже показаны усеченные L8-подобные LOS (L8-3, L8-5 и L8-2), которые способны уничтожать штаммы дикого типа, экспрессирующие полноразмерный LOS. Штаммы ΔlgtA способны стимулировать антитела, распознающие как усеченные, так и полноразмерные формы структуры LOS, без перекрестной реакции с олигосахаридами лакто-N-неотетраозы, обнаруженными на клетках крови человека.
В следующих вариантах осуществления настоящего изобретения генетически модифицированные штаммы N. meningitidis обладают стабилизированной экспрессией белков наружной мембраны, которые обычно являются чувствительными к фазовой изменчивости в штаммах дикого типа, например, в качестве неограничивающих примеров, Opc и PorA. Экспрессию этих белков можно стабилизировать способами, известными в данной области и включающими способ замены последовательности полимерных повторов либо в промоторах, либо внутри рамки считывания гена, подлежащего стабилизации, не повторяющейся последовательностью оптимальной длины, вызывая максимальную экспрессию. Например, часть последовательности поли-C или поли-G в промоторе этих генов можно заменять последовательностью такой же длины, содержащей как C, так и G нуклеотиды, например, последовательность поли-G 12 п.о. из промотора opcA (см. SEQ ID NO:1) заменяли новой последовательностью такой же длины, содержащей как C, так и G нуклеотиды, и участок Not I (см. SEQ ID NO:2, участок Not I подчеркнут). В других подходящих вариантах осуществления последовательность поли-G в промоторе PorA (например, см. SEQ ID NO:3) можно заменять новой последовательностью, содержащей как C, так и G нуклеотиды.
Дополнительные варианты осуществления настоящего изобретения относятся к вакцинным штаммам, выращенным в жидкой среде, содержащей низкий уровень железа, чтобы индуцировать белковую экспрессию белков, вовлеченных в накопление железа, например, связывающий трансферрин белок A и B. В некоторых вариантах осуществления используемая среда не содержит специфической добавки хелаторов железа, таких как десферол. Одна подходящая среда является модифицированной из среды, опубликованной BW Catlin (Catlin BW. (1973) J. Infec. Dis. 128: 178-194), посредством замены нескольких индивидуальных аминокислот на 1% казаминокислоты (сертифицированные, Difco Laboratories). Среда содержала на литр: 0,4 г NH4Cl, 0,168 г KCl, 5,85 г NaCl, 1,065 г Na2HPO4, 0,17 г KH2PO4, 0,647 г цитрат натрия, 6,25 г лактата натрия (60% сироп), 0,037 г CaCl2·2H2O, 0,0013 г MnSO4·H2O, 5 г глицерина, 0,02 г цистеина, 10 г казаминокислот, 0,616 г MgSO4 и дистиллированную воду до одного литра. Такую же среду с дефицитом железа использовали для исходных флаконов и для флаконов или ферментеров с конечной культурой.
Композиция вакцины по настоящему изобретению, содержащая NOMV по меньшей мере из трех различных генетически модифицированных штаммов подгруппы B, может обеспечивать три потенциальных уровня защиты или три типа антигенов, каждый из которых потенциально индуцирует защитный ответ антител. Три антигена представляют собой белок PorA (шесть различных подтипов PorA присутствуют в вакцине, два в каждом из трех вакцинных штаммов); липоолигосахариды (три различных коровых структуры LOS присутствуют в вакцине, одна из каждого штамма) и консервативные минорные белки NadA, варианты 1 и 2 FHBP, и Opc, сверхэкспрессированные в вакцинных штаммах. Хотя PorA обладает относительно высоким уровнем антигенной изменчивости с идентифицированными сотнями различных вариантов последовательности, конкретные сероподтипы PorA встречаются намного более часто, чем другие, и умеренное число различных сероподтипов потенциально может защищать против более чем половины заболеваний, вызванных группой B. Наличие более одного антигена, способного индуцировать бактерицидные антитела, в вакцине является важным, поскольку показано, что когда поверхностная плотность антигена является низкой, антитела против него могут не являться способными инициировать событие опосредованного комплементом лизиса. Но если присутствуют антитела против двух или более таких антигенов, антитела вместе могут инициировать опосредованный комплементом лизис. Генетически модифицированные штаммы по настоящему изобретению включают в качестве неограничивающих примеров три штамма, изображенные на фигуре 18, включая штаммы B1 (44/76 HOPS-D), B2 (8570 HOPS-G1) и B3 (B16B6 HPS-G2).
Настоящее изобретение относится к вакцине, обеспечивающей широкий спектр защиты против менингококковой инфекции, особенно менингококковых инфекций, вызываемых Neisseria meningitidis подгруппы B. Композицию вакцины по настоящему изобретению можно комбинировать с существующей тетравалентной вакциной A, C, Y и W-135 для обеспечения защиты против большинства патогенных серогрупп N. meningitidis. Не ограничиваясь рамками какой-либо конкретной теории, вакцина по настоящему изобретению может также обеспечивать вспомогательную защиту против других патогенных серогрупп, так же как против минорных серогрупп менингококков, поскольку субкапсульные антигены, на которых она основана, распространены среди всех серогрупп менингококков.
В предпочтительных вариантах осуществления настоящего изобретения интересующие гены или интересующую ДНК доставляют и интегрируют в бактериальную хромосому посредством гомологичной и/или сайт-специфической рекомбинации. Интегративные векторы, используемые для доставки таких генов и/или оперонов, могут представлять собой репликативные в определенных условиях или суицидальные плазмиды, бактериофаги, транспозоны или линейные фрагменты ДНК, полученные рестрикционным гидролизом или амплификацией ПЦР, как известно специалисту в данной области. В некоторых вариантах осуществления интеграция нацелена на хромосомные области, необязательные для роста in vitro. В других вариантах осуществления интересующий ген или интересующую ДНК можно доставлять в бактерию посредством эписомальных векторов, таких как кольцевые/линейные репликативные плазмиды, космиды, плазмиды, лизогенные бактериофаги или бактериальные искусственные хромосомы. Отбор по событиям рекомбинации можно осуществлять посредством селективных генетических маркеров, таких как гены, придающие устойчивость к антибиотикам (например, канамицину, зеомицину, эритромицину, хлорамфениколу, гентамицину и т.д.), гены, придающие устойчивость к тяжелым металлам и/или токсическим соединениям, или гены, комплементирующие ауксотрофные мутации. Альтернативно, можно проводить скрининг посредством амплификации ПЦР, секвенирования, рестрикционного расщепления или других способов, известных специалисту в данной области.
«Вакцина», как указано в настоящем документе, определена как фармацевтическая или терапевтическая композиция, используемая для инокуляции животного для иммунизации против инфекции организмом, предпочтительно патогенным организмом. Вакцины, как правило, содержат один или несколько антигенов, полученных из одного или нескольких организмов, которые при введении животному могут стимулировать активный иммунитет и защищать этого животного от инфекции этими или родственными патогенными организмами.
Очищенные NOMV получают для введения млекопитающим, соответственно, человеку, мышам, крысам или кроликам, способами, известными в данной области, которые могут включать фильтрацию для стерилизации раствора, разведение раствора, добавление адъюванта и стабилизацию раствора.
Вакцины по настоящему изобретению можно вводить человеку или животному множеством способов, включая в качестве неограничивающих примеров, например, парентеральный (например, внутримышечный, чрескожный), интраназальный, пероральный, местный или другие способы, известные специалисту в данной области. Термин парентеральный, как используют в настоящем документе далее, включает способы внутривенной, подкожной, внутрикожной, внутримышечной, внутриартериальной инъекции или инфузии. Вакцина может быть в форме препарата для однократной дозы или представлена во флаконах для множественных доз, которые можно использовать для программ массовой вакцинации. Подходящие способы получения и использования вакцин можно найти в Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Pa., Osol (ed.) (1980) and New Trends in Developments in Vaccines, Voller et al. (eds.), University Park Press, Baltimore, Md. (1978), содержание которого приведено в настоящем документе в качестве ссылки.
Композицию вакцины по настоящему изобретению, как правило, вводят парентерально в единичных дозированных составах, содержащих общепринятые, хорошо известные нетоксичные физиологически приемлемые носители, адъюванты и/или наполнители.
Композиции вакцины по настоящему изобретению могут дополнительно содержать один или несколько адъювантов. «Адъювант» представляет собой вещество, которое служит для усиления, ускорения или продления антиген-специфического иммунного ответа на антиген при использовании в комбинации со специфическими антигенами вакцины, но не стимулирует иммунный ответ при самостоятельном использовании. Подходящие адъюванты включают неорганические или органические адъюванты. Подходящие неорганические адъюванты включают в качестве неограничивающих примеров соль алюминия, такую как гель гидроксида алюминия (квасцы) или фосфат алюминия (предпочтительно гидроксид алюминия), но могут также представлять собой соль кальция (в частности, карбонат кальция), железа или цинка, или могут представлять собой нерастворимую суспензию ацилированного тирозина, или ацилированных сахаров, дериватизированных катионами или анионами полисахаридов или полифосфазенов. Другие подходящие адъюванты известны специалисту в данной области. Можно использовать также системы адъювантов Th1, и они включают в качестве неограничивающих примеров, например, монофосфориллипид А, другие нетоксичные производные LPS и комбинацию монофосфориллипида A, такого как 3-де-O-акрилированный монофосфориллипид A (#D-MPL), вместе с солью алюминия.
Другие подходящие адъюванты включают в качестве неограничивающих примеров MF59, MPLA, Mycobacterium tuberculosis, Bordetella pertussis, бактериальные липополисахариды, соединения аминоалкилглюкозаминфосфата (AGP), или их производные или аналоги, которые являются доступными из Corixa (Hamilton, Mont.) и которые описаны в патенте США No. 6113918; например, 2-[(R)-3-тетрадеканоилокситетрадеканоиламино]этил, 2-дезокси-4-O-фосфоно-3-O-[(R)-3-тетрадеканоиокситетрадеканоил]-2-[(R)-3-тетрадеканоиокситетрадеканоиламино]-b-D-глюкопиранозид, MPL™ (3-O-деацилированный монофосфориллипид A) (доступный из Corixa), описанный в патенте США No. 4912094, синтетические полинуклеотиды, такие как олигонуклеотиды, содержащие мотив CpG (патент США No. 6207646), COG-ODN (CpG олигодезоксинуклеотиды), полипептиды, сапонины, такие как Quil A или STIMULON™ QS-21 (Antigenics, Framingham, Mass.), описанный в патенте США No. 5057540, токсин коклюша (PT), или термолабильный токсин E. coli (LT), в частности LT-K63, LT-R72, CT-S109, PT-K9/G129; см., например, Международные публикации патентов No. WO 93/13302 и WO 92/19265, холерный токсин (в форме дикого типа или в мутантной форме). Альтернативно, различные масляные составы, такие как стеарилтирозин (ST, см. патент США No. 4258029), дипептид, известный как MDP, сапонин, субъединица B холерного токсина (CTB), термолабильный энтеротоксин (LT) из E. coli (разработан генетически токсоидированный мутант LT) и эмульсомы (Pharmos, LTD., Rehovot, Israel). Различные цитокины и лимфокины являются подходящими для использования в качестве адъювантов. Одним таким адъювантом является гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), обладающий нуклеотидной последовательностью, как описано в патенте США No. 5078996. Цитокин интерлейкин-12 (IL-12) представляет собой другой адъювант, описанный в патенте США No. 5723127. Показано, что другие цитокины или лимфокины обладают иммуномодулирующей активностью, включая, в качестве неограничивающих примеров, интерлейкины 1-α, 1-β, 2, 4, 5, 6, 7, 8, 10, 13, 14, 15, 16, 17 и 18, интерфероны-α, β и γ, гранулоцитарный колониестимулирующий фактор и факторы некроза опухоли α и β, и являются подходящими для использования в качестве адъювантов.
Композиции вакцины можно лиофилизировать для получения вакцины против N. meningitidis в сухую форму для простоты транспортировки и хранения. Кроме того, вакцину можно получать в форме смешанной вакцины, содержащей NOMV, содержащие белки из генетически измененных штаммов, описанных выше, и по меньшей мере один другой антиген, при условии, что добавленный антиген не мешает эффективности вакцины и побочные эффекты и неблагоприятные реакции не увеличиваются аддитивно или синергически. Вакцину можно связывать с химическими группами, которые могут улучшать растворимость, поглощение, биологическое время полужизни вакцины и т.д. Альтернативно, группы могут уменьшать токсичность вакцины, прекращать или ослаблять какой-либо нежелательный побочный эффект вакцины и т.д. Группы, способные опосредовать такие эффекты, описаны в Remington's Pharmaceutical Sciences (1980). Способы присоединения таких групп к молекуле хорошо известны в данной области.
Вакцину можно хранить в запечатанном флаконе, ампуле или т.п. Вакцину по настоящему изобретению, как правило, можно вводить в форме спрея для интраназального введения или посредством капель в нос, ингаляций, тампонов на миндалины или капсул, жидкостей, суспензий или эликсиров для перорального введения. В случае когда вакцина находится в сухой форме, вакцину растворяют или суспендируют в стерилизованной дистиллированной воде перед введением. Предпочтительно используют любой инертный носитель, такой как солевой раствор, фосфатно-солевой буфер, или любой такой носитель, в котором вакцина NOMV обладает подходящей растворимостью.
Композиции вакцины по настоящему изобретению могут включать носитель. В случае раствора или жидкой аэрозольной суспензии подходящие носители могут включать в качестве неограничивающих примеров солевой раствор, раствор сахарозы или другие растворы фармацевтически приемлемых буферов. Аэрозольные растворы могут дополнительно содержать поверхностно-активное вещество.
Подходящими наполнителями и растворителями, которые можно использовать, являются вода, раствор Рингера и изотонический раствор хлорида натрия, включая солевые растворы, забуференные фосфатом, лактатом, Tris и т.п. Кроме того, стерильные нелетучие масла являются общепринятыми в качестве растворителя или суспендирующей среды, включая, в качестве неограничивающих примеров, синтетические моно- или ди-глицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, находят применение в получении подходящих для инъекции лекарственных средств.
Подходящие для инъекции препараты, например стерильные подходящие для инъекции водные или масляные суспензии, составляют, как известно в данной области, с использованием подходящих диспергирующих или увлажняющих веществ и суспендирующих веществ. Стерильные подходящие для инъекции препараты представляют собой также стерильный подходящий для инъекции раствор или суспензию в нетоксичном, подходящем для парентерального введения разбавителе или растворителе, например, в форме раствора в 1,3-бутандиоле.
Описанный в настоящее время способ и его преимущества будут лучше понятны со ссылкой на следующие примеры. Эти примеры представлены для описания конкретных вариантов осуществления настоящего изобретения. Предоставлением этих конкретных примеров заявители не ограничивают объем и содержание настоящего способа. Специалистам в данной области понятно, что полный объем описанного в настоящее время способа включает объект, определенный формулой изобретения, приложенной к этому описанию, и любые изменения, модификации или эквиваленты этих пунктов формулы изобретения.
ПРИМЕРЫ
Пример 1: Разработка генетически модифицированного вакцинного штамма N. meningitidis и получение NOMV, содержащих белки наружной мембраны генетически модифицированного вакцинного штамма.
Генетически модифицированный штамм 8570 HOPS-G1 модифицировали посредством пяти генетических модификаций из родительского штамма 8570, который анализировали мультилокусным ферментативным электрофорезом в лаборатории, из которой получен этот штамм, и определили, что он принадлежит к клональному комплексу ET-5 (Caugant, et al.). Вариабельные области PorA были типированы по последовательности, и иммунотип LOS был верифицирован перед выполнением генетических модификаций. Штамм 8570 представлял собой клон ET-5 4:P1.19, 15:L7v, ProB3 (ST4) Tbp2 типа II. Серии из пяти последовательных генетических модификаций выполняли для штамма, как описано ниже:
1) Второй, отличный ген porA вставляли в локус opaD, нокаутируя ген opaD. Основанную на pUC 19 плазмиду pA 18.4, не имеющую маркера устойчивости к антибиотику во вставке, использовали для инсерции второго гена porA в хромосому в локус opaD, инактивируя ген opaD заменой последовательности 100 п.о. в середине гена на вставку. Инсерция содержала новый ген porA, взятый из штамма M4410 (B:15:PI.22,14) и помещенный под промотор porA, взятый из штамма H44/76. Полученный тип porA представлял собой P1.19,15: P1.22,14, содержащий два гена порина A.
2) Начиная со штамма, полученного из 1, со вторым экспрессированным PorA, экспрессию белка наружной мембраны OpcA стабилизировали заменой последовательности поли-C 12 п.о. в промоторе opcA новой последовательностью такой же длины, содержащей как C, так и G нуклеотиды. Оригинальную последовательность промотора (SEQ ID NO:1) (последовательность поли-G выделена курсивом и жирным шрифтом) 5'..CATAGTTAAAACCTCTAAAATTTGGATTGTAGTCGGATATGGTAACATAACGTAAATAATCGTTACGCTTACAATTATATTCTTAAGCTTTC GGGGGGGGGGGG ATTT..3' заменяли модифицированной последовательностью промотора (SEQ ID NO:2), содержащей как C, так и G нуклеотиды, с участком Not I (подчеркнут)
5'..CATAGTTAAAACCTCTAAAATTTGGATTGTAGTCGGATATGGTAACATAACGTAAATAATCGTTACGCTTACAATTATATTCTTAAGCTTTC GCGCGGCCGCGC ATTTT.3' Выбрана последовательность для замены, содержащая участок NotI, чтобы позволить верификацию присутствия замененной последовательности. Плазмида, используемая для трансформации, представляла собой pOpc79 (SEQ ID NO:4). Вставка плазмиды не содержит маркера устойчивости к антибиотику. Отбор трансформантов был основан на блоттинге колоний с моноклональным антителом против OpcA. Выбирали штамм, подлежащий трансформации, являющийся отрицательным по OpcA фазовым вариантом, и сильные положительные по OpcA клоны идентифицировали блоттингом колоний. Истинные трансформанты отличали от положительных по OpcA фазовых вариантов посредством ПЦР и анализа рестрикционным ферментом (Not I).
3) Начиная со штамма, полученного из 2, ген lpxL1, представляющий собой ацилтрансферазу, ответственную за связывание одной из двух связанных ацил-окси-ацилом жирных кислот с липидом A из LOS, инактивировали заменой последовательности 260 п.о. в середине гена lpxL1 вставкой, содержащей ген устойчивости к антибиотику tetM. Ген tetM получали из плазмиды pJS1934, полученной из транспозона Tn916 (Swartley, et al. 1993. Mol. Microbiol. 10:299-310). Плазмида, используемая для инактивации гена lpxL1, представляла собой pMn5 (SEQ ID NO:5). Присутствие вставки гена lpxL1 верифицировали посредством ПЦР, дающей ампликон 3,3 т.п.о. с использованием праймеров в начале и в конце гена lpxL1.
4) Начиная со штамма, полученного со стадии 3, экспрессию консервативного белка наружной мембраны GNA 1870 (вариант 1) (FHBP v.l) увеличивали вставкой второй копии варианта 1 гена GNA 1870 в локус nspA, с нокаутом экспрессии NspA. Вновь вставленный ген составлял часть вставки, содержащей ген устойчивости к антибиотику гентамицину, lac оперон E. coli с индуцируемым IPTG промотором Ptac, вариант 1 гена GNA 1870 и терминатор rrnB, использованная плазмида изображена на фигуре 19c и SEQ ID NO:6. Плазмиду на основе PUC19, pBE/GNA1870/101, использовали для трансформации и гомологичной рекомбинации для вставки варианта 1 гена GNA 1870 в модифицированный штамм. Плазмиду pBE/GNA1870/101 (7687 п.о.) сконструировали с элементами, как описано в таблице 1 (последовательность можно найти в SEQ ID NO:6).
5) Штамм, полученный на стадии 4, трансформировали плазмидой на основе pUC19, содержащей ген synX, в котором последовательность 200 п.о. заменяли на ген устойчивости к канамицину. Отбирали устойчивые к Kan трансформанты и тестировали посредством ПЦР на присутствие инактивированного гена synX и отрицательный капсульный фенотип. Результаты верифицировали нокаут гена synX.
6) Штамм, полученный на стадии 4, трансформировали плазмидой pBE-501, содержащей ген устойчивости к зеомицину, нокаутирующий ген lgtA (SEQ ID NO:9). Плазмида pBE-501 содержала элементы, которые можно найти в таблице 2. Нокаутом гена lgtA получали экспрессию укороченного или усеченного LOS с отсутствием тетрасахарида лакто-N-неотетраозы (LNnT) (см. фигуру 20).
Этот генетически модифицированный штамм тестировали, чтобы убедиться в сохранении всех пяти мутаций и экспрессии всех ожидаемых антигенов.
Пример 2: Получение вакцинных количеств генетически модифицированного штамма
Затем генетически модифицированные штаммы использовали для получения основного банка клеток и банка клеток для продукции для использования для изготовления вакцины, как подробно описано на схемах технологического процесса на фигурах 1-5 для получения композиции NOMV. Культуру NOMV тестируют по экспрессии белков наружной мембраны и LOS.
Пример 3: Характеризация вакцин
Конечный продукт, полученный из примера 2, подвергали тестированию для контроля качества и доклиническому тестированию безопасности и иммуногенности на мышах и кроликах.
Состав конечного продукта вакцины представлял собой:
Белок 200 лаг/мл
Липоолигосахарид 36 тг/мл
Нуклеиновая кислота 2,5 мкг/мл
Хлорид натрия 0,9%
Буфер Tris-HCl 0,01 M pH 7,6
Композицию вакцины далее анализировали электрофорезом в полиакриламидном геле с додецилсульфатом натрия и Вестерн-блоттингом. На фигуре 6 изображен гель, окрашенный Кумасси синим, показывающий белковое содержание в вакцине (дорожка 4) по сравнению с контролем (дорожка 2) и партией фильтрованной неочищенной вакцины (дорожка 3). На фигуре 7 изображен гель, окрашенный серебром, показывающий липоолигосахаридный компонент вакцины (дорожка 3) по сравнению с контролем (ML5 LPS, дорожка 1) и партией фильтрованной неочищенной вакцины (дорожка 2). На фигуре 8 изображены результаты тестирования идентичности вакцины по главным компонентам вакцины NOMV в соответствии с антителами, как перечислено в таблице 3.
Результаты, которые можно найти на фигурах 6, 7 и 8, показывают белки, обнаруженные в вакцинах NOMV из генетически модифицированного штамма 8570 HOPS-G NOMV, содержат белки и LOS, как описано.
Пример 4: Тест общей безопасности вакцины
Вакцину тестировали в Тесте общей безопасности вакцины, как предписано в 21 CFR 610.11. Результаты для вакцины приведены в таблице 4.
Пример 5: Тест пирогенности на кроликах*
Результаты теста пирогенности на кроликах по активности эндотоксина приведены в таблице 3 для генетически модифицированной вакцины 8570 HOPS-G NOMV отдельно и для вакцины, адсорбированной на адъюванте гидроксиде алюминия. Приведенные значения представляют собой наивысшие тестированные количества, которые не вызывали лихорадку у кроликов (увеличение температуры >0,5°C), результаты можно найти в таблице 5.
** Количество гидроксида алюминия/кг, предназначенное для использования в исследовании на человеке для всех составов (доз).
В общем, вакцина отдельно прошла тест при 0,4 пг/кг, адъювант гидроксид алюминия прошел тест при 15 пг/кг (наибольшее количество на кг, подлежащее использованию в клиническом исследовании), и вакцина, адсорбированная на гидроксиде алюминия, прошла тест при 0,5 лаг/кг, но не прошла при 1,0 пг/кг. Экстраполяция этих результатов на основе пг/кг позволяет предполагать, что адсорбированная вакцина может является не пирогенной для человека вплоть до дозы в диапазоне 25-50 мкг.
Пример 6: Высвобождение цитокинов из цельной крови человека
Вакцину тестировали на содержание эндотоксина посредством измерения ее способности индуцировать провоспалительные цитокины TNF-альфа и IL-6 из свежей цельной крови человека. Результаты показаны на фигурах 9 и 10. Данные представляют собой среднее из 3 (стандарт LPS E. Coli и вакцина NOMV партии 1119 с lpxL2 LOS) или 5 (партия 0832 NOMV с LOS дикого типа и вакцина партии 1289 NOMV с lpxL1 LOS) тестов. Планки погрешностей представляют собой стандартную ошибку среднего. Концентрация NOMV основана на белке, но стандарт LPS E. Coli основан на LPS по массе. Не ограничиваясь рамками какой-либо конкретной теории, эти результаты позволяют предполагать, что настоящая вакцина может иметь профиль безопасности у добровольцев-людей, сходный с тем, какой наблюдали для вакцины, содержащей lpxL2 LOS (Менингококковая вакцина 44/76 MOS 5D NOMV, партия № 1119, BB-IND 12687).
Активность вакцины 8570 HOPS-G NOMV партии № 1289 сравнивали с активностью экстрагированных дезоксихолатом везикул наружной мембраны (OMV). Везикулы получали с использованием основного способа, описанного Fredriksen JH, et al. NIPH Annals, 14:67-80, 1991, за исключением того, что на протяжении процедуры использовали 0,5% дезоксихолат (DOC) вместо использования 1,2% DOC для ресуспендирования осадков после ультрацентрифугирования. Результаты этого сравнения показаны на фигуре 11.
Пример 7: Иммуногенность у мышей и ответ бактерицидных антител
Мышам вводили три дозы генетически модифицированного вакцинного штамма 8570 HOPS-G с интервалами в четыре недели с адсорбцией или без на адъюванте гидроксиде алюминия (Rehydragel LV). Группы из 10 мышей вакцинировали внутрибрюшинно на 0, 4 и 8 недели с помощью 0,1, 0,3, 1,0 или 3,0 мкг NOMV, где группы вакцин перечислены в таблице 6. Сыворотку отбирали на 0, 7 и 10 неделе. Сыворотку тестировали на бактерицидные антитела против четырех различных штаммов, родительского штамма вакцины и нескольких родственных штаммов с использованием нормальной сыворотки человека в качестве источника комплемента. В сыворотке до вакцинации одинаково отсутствовала бактерицидная активность.
Результаты, полученные с помощью 10-недельной сыворотки (три дозы вакцины), показаны на фигуре 12, показывающей бактерицидный титр различных групп вакцины для генетически модифицированных штаммов. Два из тестируемых штаммов являлись изогенными с родительским штаммом вакцины. Они получены из родительского штамма заменой гена porA измененным porA, обладающим отличной специфичностью сероподтипа. Два из белков PorA, экспрессированных в этих тестируемых штаммах, присутствуют в вакцине (P1.19,15 и P1.22,14), но третий (P1.22-1,4) не присутствует. Четвертый штамм, 44/76, обладает отличным PorA, отличным PorB и отличной коровой структурой LOS по сравнению с вакцинным штаммом. Неожиданно, в отличие от опубликованных исследований, в которых для экстрагированных дезоксихолатом везикулярных вакцин показывают, что антиген PorA, как правило, является доминантным антигеном, для полученных по настоящему изобретению вакцин показали, что основная часть бактерицидной активности не зависела от сероподтипа штамма-мишени, и таким образом, не направлена против PorA.
Для бактерицидных антител, индуцированных у мышей посредством вакцины 8570 HOPS-G NOMV, не показали специфичности для сероподтипа, но, по-видимому, они в основном являются независимыми от сероподтипа и серотипа (Фигура 13). Обнаружено, что антитела, уничтожающие штамм 44/76, в основном направлены против LOS. Планки представляют собой стандартную ошибку среднего. Вакцину вводили с и без адсорбции на адъюванте гидроксиде алюминия Rehydragel LV.
Анализ специфичности ответа бактерицидных антител против гетерологичного штамма 44/76 проводили истощением бактерицидной активности с помощью различных выделенных антигенов. Сыворотку мышей после вакцинации разводили до бактерицидной конечной точки (-50% уничтожение) и инкубировали в лунках 96-луночного планшета для микротитрования, покрытых различными концентрациями нескольких антигенов. После 4-часовой инкубации сыворотку тестировали по бактерицидной активности и определяли процент удаления бактерицидного антитела. Очищенный LOS, полученный из штамма-мишени, (иммунотип L3,7) являлся способным удалять почти все антитело. Очищенный LOS (иммунотип L8v), полученный из вакцинного штамма, являлся способным удалять приблизительно 70% антитела. Консервативный белок GNA 1870 (очищенный, рекомбинантный белок), по-видимому, удалял приблизительно 20% бактерицидной активности, что, без связи с какой-либо конкретной теорией, может указывать на некоторое кооперативное уничтожение, вовлекающее как анти-LOS антитело, так и анти-GNA1870 антитело, как показано на фигуре 14.
Пример 8: Иммуногенность у кроликов
Вакцину тестировали также по иммуногенности у кроликов. Группы из четырех кроликов вакцинировали внутримышечно с помощью различных доз вакцины, с адсорбцией или без на адъюванте гидроксиде алюминия. Три дозы вводили с интервалами по шесть недель и кровь отбирали через две недели после последней инъекции. Определяли ответ бактерицидного антитела у кроликов на четыре тестируемых штамма. Тестируемые штаммы включали 3 изогенных варианта 8570, экспрессирующие различные белки PorA и L3,7v LOS, и штамм 44/76, обладающий гетерологичным PorA и LOS с отличной коровой структурой. Белки PorA P1.19,15 и P1.22,14 присутствовали в вакцине, но P1.22-1,4 не присутствовал. Результаты бактерицидных тестов приведены на фигуре 15. Анализ перекрестно реактивной бактерицидной активности против штамма 44/76 проводили таким же способом, как для сыворотки мышей, и результаты являлись в основном такими же. Большинство перекрестно-реактивных бактерицидных антител можно было удалить посредством очищенного LOS, гомологичного тестируемому штамму.
Пример 9: Получение и тестирование на животных лабораторной партии полной мультивалентной NOMV вакцины.
В дополнение к штамму 8570 HOPS-G1, который описан в примерах выше, два дополнительных вакцинных штамма были выбраны и генетически модифицированы. Первый представлял собой штамм B 16B6 (B:2a:P 1.5,2:L2). Этот штамм принадлежит к генетической группе ET-37 и обладает белком PorB класса 2 и связывающим трансферрин белком B типа I. Второй представлял собой штамм 44/76 (B:15:P1.7,16:L3,7), который принадлежит к генетической группе ET-5 и является представителем эпидемического штамма, ответственного за эпидемию, вызванную менингококками группы B в Норвегии в 1970 гг. и 1980 гг. Он экспрессирует белок PorB класса 3 и связывающий трансферрин белок B типа II.
Штамм B16B6 генетически модифицировали в основном таким же способом, как описано для штамма 8570 HOPS-G1. Два гена инактивировали, synX и lpxLl, для предотвращения синтеза капсулы и сиалирования LOS и для уменьшения токсичности LOS. Второй ген porA (подтип Pl.22-4) вставляли вместо гена opaD. Вариант 2 GNA 1870 (FHBP) с индуцируемым IPTG промотором Ptac E. coli вставляли вместо гена nspA в качестве второй копии с использованием плазмиды pBE-201 (SEQ ID NO:7). Плазмиду pBE-201 (7687 п.о. для дополнительной экспрессии fHBP (вариант 2)) сконструировали с элементами, как описано в таблице 7.
Фазовый вариант полученного штамма, экспрессирующий усеченную альфа-цепь, состоящую из глюкозы и галактозы, L2 LOS, отбирали блоттингом колоний. Полученный генетически модифицированный штамм обозначили B16B6 HPS-G2, см. фигуру 18.
Штамм 44/76 также генетически модифицировали по тому же принципу, как описано для штамма 8570 HOPS-G1. Два гена, synX и lpxL1, инактивировали инсерционным мутагенезом, второй ген porA (подтип Pl.7-1, 1) вставляли вместе с его промотором вместо гена opaD и вторую копию nadA вставляли под промотором porA вместо гена nspA. Плазмиду pBE-311 использовали для гомологичной рекомбинации для вставки гена NadA, плазмиду 3-11 сконструировали с элементами, как описано в таблице 8, и последовательность можно найти в SEQ ID NO:8.
**) Участок поли G из 14G промотора 44-76 модифицировали заменой оптимальными для экспрессии 11 G.
Кроме того, экспрессию OpcA стабилизировали исправлением фазовой изменчивости, связанной с его геном. Это выполнено, как описано для штамма 8570 HOPS-G1, посредством прерывания поли-G участка в его промоторе в примере 1. Ген lgtA прерывали, как в примере 1, получая усеченный LOS. Фазовый вариант полученного штамма, экспрессирующий иммунотип L8, отбирали блоттингом колоний с помощью специфических для L8 моноклональных антител. Этот генетически модифицированный штамм обозначили 44/76 HOPS-D, как показано на фигуре 18. Два дополнительных штамма охарактеризовали для подтверждения стабильности всех генетических модификаций и культуры каждого для хранения заморозили.
Пример 10: Получение NOMV вакцины из штаммов B16B6 HPS-G2 и 44/76 HOPS-D.
Три генетически модифицированных штамма использовали для получения лабораторных партий композиций вакцин NOMV. Штаммы выращивали в модифицированной среде Кэтлина в виде культур по одному литру в колбах Фернбаха с вращением и качанием. Клетки собирали центрифугированием, взвешивали и клеточную массу замораживали. Клеточную массу оттаивали и использовали для получения NOMV, следуя в основном такому же способу, как описано для клинической партии вакцины из штамма 8570 HOPS-G1, как описано в примере 2. Процесс доводили до меньшего масштаба и ультрацентрифугирование проводили дважды при 225000×g в течение 60 мин при 2-8°C для удаления нуклеиновых кислот и всего растворимого, не относящегося к везикулам материала.
Пример 11: Иммунизация мышей полной мультивалентной вакциной
Группы из десяти мышей CD-1 вакцинировали внутрибрюшинно двумя пг вакцины NOMV из каждого генетически модифицированного вакцинного штамма (всего 6 пг для комбинированной вакцины с NOMV из трех штаммов). Три дозы вводили на 0, 4 и 8 недели. Кровь отбирали перед вакцинацией и через 2 недели после последней вакцинации (на 10 недель).
Сыворотку от индивидуальных мышей тестировали на бактерицидные антитела против гомологичных штаммов и объединенную сыворотку от каждой группы из 10 мышей тестировали против панели из 14 гетерологичных штаммов группы B и штамма группы 1 C, экспрессирующих широкий диапазон различных субкапсульных антигенов.
Комбинированная мультивалентная вакцина индуцировала титр со средним геометрическим 1:256 против каждого из трех вакцинных штаммов и 4-кратным или большим увеличением количества бактерицидных антител против 13 из 15 гетерологичных штаммов. Два из тестируемых штаммов не подвергались уничтожению, несмотря на обладание антигеном, общим с одним из вакцинных штаммов. Наблюдаемые бактерицидные титры против панели штаммов приведены в таблице 9.
Бактерицидные титры объединенной сыворотки мышей против разнообразной панели тестируемых штаммов
Обозначение вакцин: B1=44/76 HOPS-D NOMV
B2=8570 HOS-G1 NOMV
B3=B16B6 HPS-G2 NOMV
Эти результаты показывают способность комбинированной вакцины индуцировать бактерицидные (защитные) антитела против широкого диапазона штаммов группы B и, потенциально, также против штаммов других серогрупп.
Анализом бактерицидных антител с использованием бактерицидного истощения показали, что антитела против всех трех наборов антигенов вовлечены в уничтожение по меньшей мере некоторых из тестируемых штаммов. В некоторых случаях, по-видимому, антитела против более чем одного антигена были вовлечены и действовали совместно для получения бактерицидной активности против данного штамма.
Дополнительные группы мышей вакцинировали вакциной NOMV, полученной из изогенных мутантов штамма 8570 HOPS-G1. Мутантные штаммы отличались по экспрессии PorA. Два мутанта экспрессировали один PorA (один или другой из двух в мультивалентном вакцинном штамме) и третий представлял собой мутант с нокаутом PorA, не экспрессирующий белка PorA. Бактерицидные титры, индуцируемые каждым из четырех штаммов против нескольких различных тестируемых штаммов, показаны в таблице 10.
Для первых пяти тестируемых штаммов в таблице 10 экспрессия PorA не оказывала эффекта на титр бактерицидных антител, индуцированных вакциной. Для последних двух штаммов, которые оба экспрессировали P1.14, присутствие эпитопа P1.14 в вакцине коррелировало со способностью соответствующей сыворотки уничтожать штамм. Это показывает, что антитела против PorA вовлечены в наблюдаемое уничтожение некоторых штаммов. Для других штаммов, таких как гомологичный штамм и штамм 44/76, другие антигены являются ответственными за большую часть бактерицидной активности. Это показано анализом с исследованием бактерицидного истощения. Результаты одного такого анализа приведены на фигуре 16. Результаты, показанные на фигуре 17, показывают, что антитела против LOS и FHBP (GNA1870) были вовлечены в уничтожение штамма 8570 антисывороткой против мутанта с нокаутом PorA 8570 HOPS-G1.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ТРАНСФЕРРИН-СВЯЗЫВАЮЩИЙ БЕЛОК И HSF ИЗ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ | 2003 |
|
RU2359696C2 |
| МЕНИНГОКОККОВЫЕ ПОЛИПЕПТИДЫ fHBP | 2009 |
|
RU2475496C2 |
| СПОСОБ БЕЗДЕТЕРГЕНТНОГО ПОЛУЧЕНИЯ ВЕЗИКУЛ НАРУЖНОЙ МЕМБРАНЫ ГРАМОТРИЦАТЕЛЬНОЙ БАКТЕРИИ | 2012 |
|
RU2607021C2 |
| МЕНИНГОКОККОВЫЕ ПОЛИПЕПТИДЫ fHBP | 2012 |
|
RU2567003C2 |
| ВАКЦИНЫ С ИСПОЛЬЗОВАНИЕМ ВЕЗИКУЛ НА ОСНОВЕ GNA 1870 ШИРОКОГО СПЕКТРА ДЕЙСТВИЯ ДЛЯ ПРОФИЛАКТИКИ ЗАБОЛЕВАНИЙ, ВЫЗЫВАЕМЫХ NEISSERIA MENINGITIDIS | 2006 |
|
RU2432962C2 |
| НАБОР ДЛЯ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ ПРОТИВ Neisseria meningitidis СЕРОЛОГИЧЕСКОЙ ГРУППЫ В | 2008 |
|
RU2498815C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДОСТАВКИ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА | 1994 |
|
RU2174845C2 |
| ВАРИАНТЫ ФАКТОР Н-СВЯЗЫВАЮЩЕГО БЕЛКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714248C2 |
| ПЕПТИД NEISSERIA MENINGITIDIS ДЛЯ ТЕРАПЕВТИЧЕСКОГО И ДИАГНОСТИЧЕСКОГО ПРИМЕНЕНИЯ | 2001 |
|
RU2313535C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ САХАРИД ПНЕВМОКОККА СЕРОТИПА 14 | 2010 |
|
RU2549438C2 |

























Группа изобретений относится к области медицины, а именно к биофармацевтики, и может быть использована для приготовления вакцин. Для этого вакцина содержит: нативные везикулы наружной мембраны (NOMV), полученные по меньшей мере из двух штаммов менингококков, которые были генетически модифицированы для широкого спектра защиты, где нативные везикулы наружной мембраны включают различные наборы антигенов, где различные наборы антигенов содержат один или несколько белков РоrА, один или несколько LOS и один или несколько консервативных белков наружной мембраны; где генетически модифицированные штаммы были модифицированы для повышения безопасности путем инактивации генов lpxL1, synX и lgtA и где по меньшей мере один генетически модифицированный штамм экспрессирует по меньшей мере два различных подтипа белков или подтипа эпитопов РоrА. Также предложен способ стимуляции иммунного ответа на менингококковую инфекцию, включающий в себя введение вакцинной композиции по настоящему изобретению. Группа изобретений обеспечивает защитный иммунитет против менингококковой инфекции, предпочтительно инфекции, вызываемой менингококками подтипа В. 4 н. и 45 з.п. ф-лы, 11 пр., 21 ил., 10 табл.
1. Вакцина, содержащая нативные везикулы наружной мембраны (NOMV), полученные по меньшей мере из двух штаммов менингококков, которые были генетически модифицированы для широкого спектра защиты, где нативные везикулы наружной мембраны включают различные наборы антигенов, где различные наборы антигенов содержат один или несколько белков РоrА, один или несколько LOS и один или несколько консервативных белков наружной мембраны; где генетически модифицированные штаммы были модифицированы для повышения безопасности путем инактивации генов lpxL1, synX и lgtA; и где по меньшей мере один генетически модифицированный штамм экспрессирует по меньшей мере два различных подтипа белков или подтипа эпитопов РоrА.
2. Вакцина по п.1, где LOS, экспрессированный каждым штаммом, имеет отличающуюся коровую структуру LOS и имеет альфа-цепь, состоящую из глюкозы и галактозы.
3. Вакцина по п.1, где каждый подтип белков или подтип эпитопов РогА содержит по меньшей мере один, наиболее преобладающий подтип РогА из изолятов группы В.
4. Вакцина по п.1, где в каждом штамме сверхэкспрессирован другой консервативный поверхностный белок с известной способностью индуцировать бактерицидные антитела и он выбран из группы, состоящей из варианта 1 FHBP, варианта 2 FHBP и варианта 3 FHBP; NadA; App; NspA; TbpA и TbpB.
5. Вакцина по п.1, где NOMV получены без обработки детергентом или денатурирующими растворителями из клеточной массы или культуральной среды, собранной из бактериальной культуры.
6. Вакцина по п.1, где вакцину суспендируют в 5%-ной глюкозе, используемой в качестве наполнителя.
7. Вакцина по п.1, где NOMV комбинируют с одним или несколькими адъювантами, например гидроксидом алюминия или фосфатом алюминия, MF 59, CPG-ODN или MPLA.
8. Вакцина по п.1, где вакцина против менингококковой инфекции содержит нативные везикулы наружной мембраны менингококков (NOMV) одного или нескольких генетически модифицированных штаммов Neisseria meningitidis, где один или несколько генетически модифицированных штаммов модифицированы путем:
i. инактивации гена synX,
ii. инактивации гена lpxL1,
iii. инактивации гена lgtA в каждом штамме, что приводит к экспрессии укороченных или усеченных липоолигосахаридов (LOS) без тетрасахарида лакто-N-неотетраозы, и
iv. инсерции по меньшей мере одного второго гена роrА, отличающегося антигенными свойствами, вместо гена ора.
9. Вакцина по п.8, где генетически модифицированный штамм дополнительно стабильно экспрессирует по меньшей мере один минорный консервативный белок наружной мембраны или его экспрессия повышена.
10. Вакцина по п.8, где генетически модифицированный штамм дополнительно стабильно экспрессирует по меньшей мере один белок наружной мембраны, где белок наружной мембраны выбран из группы, содержащей Орс и РоrА.
11. Вакцина по п.8, где по меньшей мере один второй ген роrА, отличающийся антигенными свойствами, экспрессирует по меньшей мере один подтип белка или подтип эпитопа РоrА, выбранный из распространенных подтипов РоrА изолятов Meningitidis группы В.
12. Вакцина по п.9, где по меньшей мере один минорный консервативный белок наружной мембраны выбран из группы, состоящей из: вариантов 1 FHBP (GNA 1870), варианта 2 FHBP, варианта 3 FHBP; NadA; App, NspA, TbpA и В.
13. Вакцина по п.1, где вакцина содержит генетически модифицированный вакцинный штамм из штамма Neisseria meningitidis подтипа В, включая штамм Н44/76 HOPS-D.
14. Вакцина по п.13, где вакцина содержит генетически модифицированный вакцинный штамм Neisseria meningitidis подтипа В, полученный из штамма Н44/76, который имеет следующие генетические модификации:
i) ген synX инактивирован,
ii) ген lpxL1 инактивирован,
iii) ген lgtA инактивирован,
iv) проведена инсерция второго гена рогА вместо гена opaD,
v) повышена экспрессия NadA по сравнению с нативным штаммом, и
vi) отмечается стабильная повышенная экспрессия белков Орс и РоrА.
15. Вакцина по п.13, где генетически модифицированный штамм Н44/76 HOPS-DL получен из штамма дикого типа ЕТ-5 Н44/76.
16. Вакцина по п.1, где вакцина содержит генетически модифицированный вакцинный штамм Neisseria meningitidis подтипа В, включая штамм 8570 HOS-G1.
17. Вакцина по п.16, где генетически модифицированный вакцинный штамм Neisseria meningitidis подтипа В получен из 8570, который имеет следующие генетические модификации:
i) ген synX инактивирован,
ii) ген lpxL1 инактивирован,
iii) ген lgtA инактивирован,
iv) проведена инсерция второго гена роrА вместо opaD;
v) повышена экспрессия варианта 1 белка, связывающего фактор Н; и
vi) отмечается стабильная повышенная экспрессия белков РоrА и Орс.
18. Вакцина по п.16, где генетически модифицированный штамм получен из штамма дикого типа ЕТ-5 8570.
19. Вакцина по п.1, где вакцина содержит генетически модифицированный вакцинный штамм Neisseria meningitidis подтипа В, включая штамм В16 В6 HPS-G2A.
20. Вакцина по п.19, где генетически модифицированный штамм получен из штамма дикого типа ЕТ-37 В16В6.
21. Вакцина по п.1, где генетически модифицированный вакцинный штамм Neisseria meningitidis подтипа В получен из В16В6, который имеет следующие генетические модификации:
i) ген synX инактивирован,
ii) ген lpxL1 инактивирован,
iii) ген lgtA инактивирован,
iv) проведена инсерция второго гена роrА (подтип Р1.22-1,4) вместо opaD;
v) повышена экспрессия варианта 2 белка, связывающего фактор Н; и
vi) отмечается стабильная повышенная экспрессия белков РоrА и Орс.
22. Вакцина по любому из пп.1-21, где NOMV получают из клеточной массы или из культуральной среды без обработки детергентом или денатурирующим растворителем.
23. Вакцина по любому из пп.1-21, где композицию вакцины суспендируют в 5%-ной глюкозе, используемой в качестве наполнителя.
24. Вакцина по любому из пп.1-21, где NOMV комбинируют с одним или несколькими адъювантами.
25. Вакцина по любому из пп.1-21, где генетически измененный штамм модифицирован так, что экспрессирует белки накопления железа.
26. Вакцина по любому из пп.13-21, где штамм выращивают на среде с дефицитом железа.
27. Вакцина по п.26, где NOMV получают из клеточной массы или из культуральной среды без обработки детергентом или денатурирующим растворителем.
28. Вакцина по п.26, где композицию вакцины суспендируют в 5%-ной глюкозе, используемой в качестве наполнителя.
29. Вакцина по п.26, где NOMV комбинируют с одним или несколькими адъювантами.
30. Вакцина по п.26, где генетически измененный штамм модифицирован так, что экспрессирует белки накопления железа.
31. Вакцина по любому из пп.13-21, где инактивацию гена synX, гена lpxL1 или гена lgtA осуществляют посредством вставки гена устойчивости к лекарственному средству в последовательность инактивированного гена.
32. Вакцина по п.31, где NOMV получают из клеточной массы или из культуральной среды без обработки детергентом или денатурирующим растворителем.
33. Вакцина по п.31, где композицию вакцины суспендируют в 5%-ой глюкозе, используемой в качестве наполнителя.
34. Вакцина по п.31, где NOMV комбинируют с одним или несколькими адъювантами.
35. Вакцина по п.31, где генетически измененный штамм модифицирован так, что экспрессирует белки накопления железа.
36. Комбинация нативных везикул наружной мембраны (NOMV) из трех генетически модифицированных штаммов Neisseria meningitides, обладающих антигенной изменчивостью, где три генетически модифицированных штамма содержат:
(1) штамм Н44/76 HOPS-DL со следующими генетическими модификациями или характеристиками:
инактивацией генов synX, lp×L1 и lgtA;
инсерцией второго гена роrА, подтип Р1.7-1,1, вместо opaD;
повышенной экспрессией NadA по сравнению с диким типом;
стабильной эксперссией L8-6 LOS; и стабильной высокой экспрессией Орс и РоrА;
(2) штамм 8570 HOPS-GiL со следующими генетическими модификациями или характеристиками:
инактивацией генов synX, lp×L1 и lgtA;
инсерцией второго гена роrА, подтип Р1.22.1,4, вместо opaD;
повышенной экспрессией варианта 1 белка, связывающего фактор Н;
стабильной эксперссией L8-6 LOS; и стабильной высокой экспрессией РоrА и Орс;
(3) штамм В16В6 HPS-GaA со следующими генетическими модификациями или характеристиками:
инактивацией генов synX, lp×L1 и lgtA;
инсерцией второго гена рогА, подтип Р 1.22.1,4, вместо opaD;
повышенной экспрессией варианта 2 белка, связывающего фактор Н;
стабильной эксперссией L8-6 LOS; и стабильной высокой экспрессией РоrА и Орс.
37. Комбинация NOMV п.36, где штамм Н44/76 HOPS-DL получен из штамма дикого типа ЕТ-5 Н44/76.
38. Комбинация NOMV по п.36, где штамм 8570 HOPS-GIL получен из штамма дикого типа ЕТ-5 8570.
39. Комбинация NOMV по п.36, где штамм В16В6 HPS-G2L получен из штамма дикого типа ЕТ-37 В16В6.
40. Композиция вакцины, содержащая нативные везикулы наружной мембраны (NOMV) из одного или нескольких генетически модифицированных штаммов Neisseria meningitidis подтипа В, выбранных из группы, состоящей из:
a) генетически модифицированного вакцинного штамма Neisseria meningitidis подтипа В, полученного из: штамма Н44/76, содержащего генетические модификации:
i) ген synX инактивирован,
ii) ген lp×L1 инактивирован,
iii) ген lgtA инактивирован,
iv) встроен второй ген роrА вместо гена opaD,
v) повышена экспрессия NadA по сравнению с природным штаммом, и
vi) стабилизирована повышенная экспрессия белков Орс и РоrА;
b) генетически модифицированного вакцинного штамма Neisseria meningitidis подтипа В, полученного из 8570, содержащего следующие генетические модификации:
i) ген synX инактивирован,
ii) ген lp×L1 инактивирован,
iii) ген lgtA инактивирован,
iv) встроен второй ген роrА (подтип Р1.22-1,4) вместо гена opaD,
v) повышена экспрессия варианта 2 белка, связывающего фактор Н; и
vi) стабилизирована повышенная экспрессия белков РоrА и Орс; и
c) генетически модифицированного вакцинного штамма Neisseria meningitidis подтипа В, полученного из В16В6, содержащего следующие генетические модификации:
i) ген synX инактивирован,
ii) ген lp×L1 инактивирован,
iii) ген lgtA инактивирован,
iv) проведена инсерция второго гена роrА вместо opaD;
v) повышена экспрессия варианта 2 белка, связывающего фактор Н; и
vi) отмечается стабильная повышенная экспрессия белков РоrА и Орс.
41. Композиция вакцины по п.40, где композиция вакцины содержит NOMV из двух или более генетически модифицированных штаммов.
42. Композиция вакцины по п.40, где композиция вакцины содержит NOMV из трех или более генетически модифицированных штаммов.
43. Вакцина по любому из пп.40-42, где NOMV получают из клеточной массы или из культуральной среды без обработки детергентом или денатурирующим растворителем.
44. Вакцина по любому из пп.40-42, где композицию вакцины суспендируют в 5%-ной глюкозе, используемой в качестве наполнителя.
45. Вакцина по любому из пп.40-42, где NOMV комбинируют с одним или несколькими адъювантами.
46. Вакцина по любому из пп.40-42, где генетически измененный штамм модифицирован так, что экспрессирует белки накопления железа.
47. Способ стимуляции иммунного ответа на менингококковую инфекцию у животного или человека, включающий введение вакцины по любому из пп.1-35 или композиции вакцины по любому из пп.40-42 животному или человеку для иммунизации против менингококковой инфекции.
48. Способ по п.47, где вакцину используют для иммунизации против инфекции, вызываемой менингококками группы В.
49. Способ по п.47, где вакцину вводят внутримышечно и/или интраназально для иммунизации против менингококковой инфекции.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Устройство для исследования скважин и пластов | 1984 |
|
SU1208214A1 |
| Способ получения средства, обладающего иммуномодулирующей активностью | 1990 |
|
SU1797894A1 |
| US 200300594444 A1, 27.03.2003. | |||

