ОБЛАСТЬ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к применению Шаперонина 10 для модуляции передачи сигнала Toll-подобным рецептором, и/или продукции, и/или секреции иммуномодулятора, индуцируемой Toll-подобным рецептором. Более конкретно, данное изобретение относится к модуляции секреции цитокинов и хемокинов, индуцируемой Toll-подобным рецептором, для лечения заболеваний, нарушений и состояний, возникающих вследствие избыточной секреции иммуномодуляторов. Данное изобретение также относится к получению, созданию и/или скринингу агонистов и антагонистов Шаперонина 10.
ПРЕДПОСЫЛКИ К СОЗДАНИЮ ИЗОБРЕТЕНИЯ
Семейство Toll-подобных рецепторов играет важную роль в воспалении и иммунной защите у насекомых, животных и у растений. Toll-подобные рецепторы (TLR) экспрессируются клетками мононуклеарной клеточной линии, в том числе лимфоцитами, макрофагами и дендритными клетками.
TLR2 активируется агонистами TLR2, такими как липотейхоевая кислота и липопептиды, которые могут быть компонентами наружной стенки некоторых бактерий. TLR3 активируется такими агонистами, как двухспиральная РНК вирусного происхождения.
TLR4 активируется липопротеинами, или липополисахаридами (LPS), или эндотоксином, которые являются компонентами наружной стенки грамотрицательных бактерий.
Активация TLR под действием патогенов или полученных из них молекул индуцирует внутриклеточную передачу сигнала, которая сначала приводит к активации фактора транскрипции NF-κΒ (Beg, 2002, Trends Immunol. 2002 23 509-12) и модуляции продукции цитокинов. Однако может быть запущен также и ряд других путей передачи сигнала, включая p38 митоген-активируемую киназу, c-Jun-N-концевую киназу и киназные пути, связанные с внеклеточной передачей сигнала (Flohe, et al., 2003, J Immunol, 170 2340-2348; Triantafilou & Triantafilou, 2002, Trends Immunol, 23 301-304). Профили генной экспрессии, индуцируемые лигированием разных TLR, различаются, но зачастую перекрываются. Например, большая часть генов, активируемых агонистами TLR3 и двухспиральной РНК, также активируются агонистами TLR4 и LPS (Doyle et al., 2002, Immunity, 17 251-263). В результате активации TLR4 под действием LPS в макрофагах происходит секреция TNF-α, IL-12 IL-1β, RANTES и MIP1β (Flohe et al., выше; Jones et al., 2002, J Leukoc Biol, 69 1036-1044).
Шаперонин 10 млекопитающих (также известный как белок теплового шока 10) впервые был описан как митохондриальный белок, участвующий в процессе пространственной упаковки белка, и является гомологом бактериального белка GroES. GroES и шаперонин 10 (Cpn10) олигомеризуется в семичленные кольца, которые связываются в виде крышечки на структуре, подобной чашке, которая включает в себя семь молекул GroEL или Hsp60, которые связывают денатурированные белки в комплекс (Bukau & Horwich, 1998, Cell, 92 351-366; Hartl & Hayer-Hartl, 2002, Science, 295 1852-1858). Cpn10 также часто обнаруживают на поверхности клетки (Belles et al., 1999, Infect Immun, 67 4191-4200; Feng et al., 2001, Blood, 97 3505-3512) и во внеклеточной жидкости (Michael et al., 2003, J Biol Chem, 278 7607-7616; Johnson et al., 2003, Cir Rev Immunol, 23 15-44).
Также было показано, что Cpn10 является фактором супрессии, присутствующим на ранней стадии беременности, и была показана иммуносупрессорная активность при экспериментальном аутоиммунном энцефаломиелите, гиперчувствительности замедленного типа и на моделях отторжения аллотрансплантата (Zhang et al., 2003, J Neurol Sci, 212 37-46; Morton et al., 2000, Immunol Cell Biol, 78 603-607).
Недавнее исследование с использованием Cpn10 Mycobacterium tuberculosis, описанное в публикации международной заявки WO 02/40038, дает возможность предположить, что эта молекула может обладать эффективностью при лечении таких заболеваний, как рак, аллергические реакции и/или состояния, опосредованные Th2-типом иммунных реакций. Предполагается, что этого можно достичь через индукцию цитокинов, таких как TNF-α и IL-6.
Однако механизм действия, которым Cpn10 и, в частности, Cpn10 млекопитающих осуществляет свои иммунорегуляторные функции, остается неясным.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Неожиданно авторы данного изобретения показали, что Cpn10 регулирует стимуляцию секреции иммуномодуляторов, опосредованную агонистом Toll-подобного рецептора.
Более конкретно, Cpn10 отрицательно регулирует индукцию провоспалительных иммуномодуляторов, опосредованную Toll-подобными рецепторами, и положительно регулирует индукцию секреции противовоспалительных иммуномодуляторов, опосредованную Toll-подобными рецепторами.
Таким образом, данное изобретение в широком смысле направлено на модуляцию передачи сигнала Toll-подобным рецептором под действием шаперонина 10 (Cpn10).
В первом аспекте данное изобретение относится к способу регуляции передачи сигнала Toll-подобным рецептором у животного или в полученных от него одной или нескольких клетках, или тканях, или органах, включающему в себя стадию введения Cpn10 или производного Cpn10 животному, в клетки, ткани или органы, тем самым регулируя передачу сигнала Toll-подобном рецептором.
Во втором аспекте, данное изобретение относится к способу регуляции секреции иммуномодулятора у животного или в полученных от него одной или нескольких клетках, или тканях, или органах, включающему в себя стадию введения Cpn10 или производного Cpn10, животному, в клетки, ткани или органы, тем самым регулируя продукцию и/или секрецию иммуномодулятора, индуцируемую Toll-подобным рецептором.
В соответствии с этими аспектами данное изобретение относится к способу регуляции передачи сигнала Toll-подобными рецепторами, и/или продукции, и/или секреции иммуномодуляторов, для модуляции, таким образом, иммунного ответа животного для профилактики или лечения заболевания, нарушения или состояния.
Предпочтительно данное заболевание, нарушение или состояние выбрано из острых или хронических воспалительных заболеваний, таких как септический шок, воспалительное заболевание кишечника, артрит, псориаз, заболевание сердца, атеросклероз, хроническое заболевание легких, кахексия, рассеянный склероз, болезнь «трансплантат против хозяина» (GVHD), трансплантация или рак.
В соответствии с данным изобретением Cpn10 предпочтительно регулирует Toll-подобный рецептор, выбранный из группы, состоящей из TLR2, TLR3 и TLR4.
Более предпочтительно Toll-рецептор выбран из группы, состоящей из TLR2 и TLR4.
Еще более предпочтительно Toll-рецептор представляет собой TLR4.
Соответственно, Cpn10 регулирует передачу сигнала Toll-подобным рецептором и продукцию и/или секрецию иммуномодуляторов, которая стимулируется, активируется или индуцируется под действием агониста Toll-подобного рецептора.
Агонистом Toll-подобного рецептора может быть патоген, полученная из него или продуцируемая им молекула или синтетический агонист Toll-подобного рецептора.
Предпочтительно агонист Toll-подобного рецептора выбран из группы, состоящей из LPS, липопептида и двухспиральной РНК.
В одном варианте осуществления, в котором Toll-подобный рецептор представляет собой TLR4, агонистом предпочтительно является LPS.
В другом варианте осуществления, в котором Toll-подобный рецептор представляет собой TLR3, агонистом предпочтительно является двухспиральная РНК.
Еще в одном варианте осуществления, в котором Toll-подобный рецептор представляет собой TLR2, агонистом предпочтительно является липопептид.
В конкретных вариантах осуществления липопептид может представлять собой PAM3CysSК4.
Соответственно, животное или полученные от него клетки, ткани или органы содержат клетки, экспрессирующие один или несколько Toll-подобных рецепторов.
Предпочтительно клетка представляет собой иммунокомпетентную клетку.
Более предпочтительно, когда иммунокомпетентная клетка представляет собой моноцит, макрофаг, дендритную клетку или лимфоцит.
Предпочтительно, если животное является млекопитающим.
Более предпочтительно животное представляет собой человека.
В соответствии с вышеуказанными аспектами в одном варианте осуществления иммуномодулятор представляет собой провоспалительный цитокин, такой как TNF-α или интерлейкин 6 (IL-6), или провоспалительный хемокин, такой как RANTES, но не ограничивается ими.
В другом варианте осуществления иммуномодулятор представляет собой противовоспалительный цитокин, такой как интерлейкин-10 (IL-10), или противовоспалительный хемокин, такой как TGF-β, но не ограничивается ими.
В тех вариантах осуществления, в которых иммуномодулятор представляет собой провоспалительный цитокин или хемокин, введение Cpn10 предпочтительно вызывает ингибирование, супрессию или иным образом снижает продукцию и/или секрецию указанного иммуномодулятора.
В тех вариантах осуществления, в которых иммуномодулятор представляет собой противовоспалительный цитокин или хемокин, введение Cpn10 предпочтительно повышает, облегчает, способствует или иным образом увеличивает продукцию и/или секрецию указанного иммуномодулятора.
В третьем аспекте данное изобретение относится к выделенному молекулярному комплексу, содержащему Toll-подобный рецептор, агонист Toll-подобного рецептора и Cpn10.
Предпочтительно Toll-подобный рецептор выбран из группы, состоящей из TLR2, TLR3 и TLR4.
Более предпочтительно Toll-рецептор выбран из группы, состоящей из TLR2 и TLR4.
Еще более предпочтительно Toll-подобный рецептор представляет собой TLR4.
В одном варианте осуществления, в котором Toll-подобный рецептор представляет собой TLR4, агонистом является LPS.
В другом варианте осуществления, в котором Toll-подобный рецептор представляет собой TLR3, агонистом является двухспиральная РНК.
Еще в одном варианте осуществления, в котором Toll-подобный рецептор представляет собой TLR2, агонист представляет собой PAM3CysSK4.
В одном конкретном варианте осуществления данное изобретение относится к выделенному молекулярному комплексу, содержащему TLR4, LPS и Cpn10.
В четвертом аспекте данное изобретение относится к способу получения, создания или скрининга агониста Cpn10, включающему в себя стадию определения имитирующего или усиливающего действия предполагаемого агониста на Cpn10 регуляцию передачи сигнала Toll-подобным рецептором, и/или продукцию, и/или секрецию иммуномодулятора, индуцируемого Toll-подобным рецептором.
В пятом аспекте данное изобретение относится к способу получения, создания или скрининга антагониста Cpn10, включающему в себя стадию определения ингибирования, снижения, супрессии или иного понижения под действием Cpn10 регуляции передачи сигнала Toll-подобным рецептором, и/или продукции, и/или секреции иммуномодулятора, индуцируемого Toll-подобным рецептором.
Предпочтительно Toll-подобный рецептор выбран из группы, состоящей из TLR2, TLR3 и TLR4.
Более предпочтительно Toll-рецептор выбран из группы, состоящей из TLR2 и TLR4.
Еще более предпочтительно Toll-подобный рецептор представляет собой TLR4.
В шестом аспекте данное изобретение относится к агонисту Cpn10 или агонисту, полученному, созданному или отобранному при скрининге в соответствии с аспектами, упомянутыми выше.
Будет понятно, что по тексту данного описания «включают в себя», «включает в себя» и «включающий в себя» использованы включительно, а не исключительно, будет понятно, что подразумевается включение установленного целого или группы целого, а не исключение любого другого целого или группы целого.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1 Cpn10 ингибирует LPS-индуцированную активацию клеток RAW264.7 и продукцию провоспалительного медиатора.
(A) Cpn10-опосредованное ингибирование LPS-индуцированной активности NF-κΒ. В 9 отдельных экспериментах 100 мкг/мл Cpn10 (Hsp10 +) или буфера (Hsp10 -) предварительно инкубировали с клетками RAW264-HIV-LTR-LUC в течение 2 ч. Затем добавляли LPS в концентрации 5, 1 и 0,2 нг/мл и через 2 часа измеряли активность люциферазы. Условные единицы свечения (RLU) люциферазы, полученные для 5 нг/мл LPS, принимали за 100% относительной активности люциферазы, а 0% соответствует RLU, полученным в отсутствие LPS. Сам по себе Cpn10 не увеличивал в значительной степени RLU (данные не показаны). Среднее значение процента снижения (± SD) RLU, опосредованного Cpn10, показано для каждой концентрации LPS, и достоверность подсчитывали с использованием парного t-критерия Стьюдента.
(B, C) Cpn10-опосредованное ингибирование LPS-индуцированной секреции RANTES и IL-6. Клетки RAW264.7 инкубировали с указанной концентрацией Cpn10 (Hsp10) в течение 2 ч, с последующим добавлением 1 нг/мл LPS. Через 6 ч трехкратные супернатанты анализировали на наличие RANTES и IL-6 твердофазным иммуноферментным анализом (ELISA). Показано среднее значение процента снижения секреции RANTES и IL-6, опосредованное 100 мкг/мл Cpn10. Достоверность рассчитывали с использованием парного t-критерия Стьюдента.
Фиг.2 Действие Cpn10 на секрецию цитокина в мышиных системах.
(A) Обработка Cpn10 снижала способность перитонеальных макрофагов, стимулированных LPS, продуцировать TNF-α. Мышам C57BL/6 (n=3) вводили Cpn10 (Cpn10 +) или контрольный разбавитель (Cpn10 -). Перитонеальные макрофаги в испытуемой группе от индивидуальных животных собирали путем перитонеального лаважа на 6 день и объединяли. Клетки помещали 2×105/лунку в присутствии LPS (1 мкг/мл). Культуральные супернатанты собирали через 5 часов и уровни TNF-α определяли с помощью ELISA. (Лунки без LPS продуцировали неопределяемое количество TNF-α - данные не показаны). Показаны средние значения ± SE по трем лункам и приведены к уровню продукции на 105 макрофагов на основании CD1 1b-окрашивания. Показаны данные двух идентичных экспериментов, и указан средний процент снижения, с достоверностью, рассчитанной методом ANOVA.
(В) Обработка Cpn10 увеличивала продукцию IL-10 клетками селезенки. Мышам C57BL/6 вводили либо Cpn10, либо контрольный разбавитель, как показано выше. Клетки селезенки собирали на 6 день и объединяли от индивидуальных животных в пределах испытуемой группы и культивировали при 5×105/лунку в присутствии LPS (10 мкг/мл). Культуральные супернатанты собирали через 48 часов и определяли уровни IL-10 с помощью ELISA. Показаны средние значения ± SE по трем лункам. Показан средний процент увеличения со статистическими данными, рассчитанными как для A. (С) Обработка Cpn10 снижала продукцию TNF-α IL-10-/- перитонеальными макрофагами. Мышам IL-10-/- C57BL/6 вводили Cpn10 или контрольный разбавитель, как показано выше, и перитонеальные макрофаги собирали, как показано для A. Через 5 часов культивирования в присутствии LPS (0,1, 1 или 10 мкг/мл) TNF-α определяли в культуральных супернатантах методом ELISA. Показано среднее значение ± SE по трем лункам для одного репрезентативного эксперимента. Уровни TNF-α сравнивали у животных, получавших Cpn10, и контрольных животных, используя непараметрический t-тест.
Фиг.3 Обработка Cpn10 мононуклеарных клеток периферической крови (PBMC) человека снижает секрецию TNF-α и IL-6, индуцированную LPS, и не индуцирует толерантность. (A) Cpn10 снижал секрецию TNF-α, индуцированную LPS. К PBMC от восьми различных доноров добавляли Cpn10 (1 и 10 мкг/мл) или буфер (0) за 1 час до добавления 0,04 нг/мл LPS. Супернатаны удаляли через 20 ч и анализировали на наличие TNF-α. Указаны проценты снижения и достоверность, рассчитанная с использованием парного t-критерия Стьюдента. Для всех доноров 10 мкг/мл Cpn10, в отсутствие LPS, не индуцировал уровни TNF-α выше определяемого уровня (31 пг/мл) (данные не показаны). (B) Предварительная обработка с использованием LPS в течение 1 ч не могла индуцировать толерантность к последующей секреции TNF-α, индуцированной LPS. PBMC подвергали воздействию указанных концентраций LPS предварительной обработки. Через 1 ч PBMC стимулировали действием 0,04 нг/мл LPS, и супернатанты анализировали на наличие цитокинов спустя 20 ч. (C) Cpn10 снижал секрецию IL-6, индуцированную LPS. Cpn10 (1 и 10 мкг/мл) или буфер (0) добавляли к PBMC от восьми различных доноров за 1 ч до добавления 0,04 нг/мл LPS. Через 20 ч супернатанты удаляли и анализировали на наличие IL-6. Указаны проценты снижения и достоверность (рассчитанные как для A). Для всех доноров 10 мкг/мл Cpn10, в отсутствие LPS, не индуцировал уровни IL-6 выше определяемого уровня (9 пг/мл) (данные не показаны). (D) Предварительная обработка с использованием LPS в течение 1 ч не могла индуцировать толерантность к последующей секреции IL-6, индуцированной LPS. PBMC подвергали воздействию указанных концентраций LPS предварительной обработки. Через 1 ч PBMC стимулировали 0,04 нг/мл LPS и супернатанты анализировали на наличие цитокина спустя 20 ч.
Фиг.4 Активность Cpn10 в моделях воспаления на мышах.
Cpn10 снижает уровни LPS-индуцированного сывороточного TNF-α и RANTES и повышает уровни IL-10. В 5 различных экспериментах мышам C57BL/6 (n=3 или 4 на группу) вводили буфер (Cpn10 -) или 100 мкг Cpn10 (Cpn10 +) внутривенно за 30 мин до внутривенного введения 10 мкг LPS. Через 1,5 часа животных скарифицировали и определяли сывороточные уровни TNF-α, RANTES и IL-10; (два последних определяли в 3/5 экспериментов). Планки погрешностей представляют стандартные ошибки в каждом эксперименте. Указаны сокращения в процентном выражении TNF-α и RANTES и увеличение IL-10 (± SD), и достоверность рассчитана с использованием ANOVA тестов.
Обработка Cpn10 перед трансплантацией задерживает смертность при GVHD и снижает клиническую тяжесть острого заболевания. Сингенные отрицательные контроли (n=8) (белые кружочки) представляют мышей B6D2F1, которым трансплантировали сингенный костным мозгом B6D2F1 и T-клетки. Аллогенные положительные контроли (n=10) (белые квадратики) представляют мышей реципиентов B6D2F1, которым предварительно вводили разбавитель, трансплантированных клетками мышей доноров B6, которым предварительно вводили разбавитель. Аллогенные + Cpn10 (n=10) (черные квадратики) представляют B6D2F1 реципиентов, получающих трансплантаты костного мозга и Т-клетки от мышей-доноров B6, где и реципиентов, и доноров предварительно перед трансплантацией обрабатывали Cpn10.
Кривые выживания Каплан-Мейера и клинические баллы показаны для трех групп, и аллогенные группы, получавшие Cpn10 и не получавшие Cpn10, сравнивали с помощью логарифмической ранговой статистики и непараметрического t-теста соответственно. Клинические баллы существенно отличались только на 7 день.
Фиг.5 (A) Область подкожной гранулемы у мышей BALB/c, которым вводили полный адъювант Фрейнда (CFA) с Cpn10 и без него. Cpn10 результаты показаны заштрихованными светлым колонками слева от каждой пары колонок по сравнению с буферным контролем (заштрихованы темным, правые колонки). (B-E) PAM3CysSK4-индуцированную активацию макрофагов ингибируют действием Cpn10 в клетках RAW264-HIV-LTR-LUC. (B) Cpn10 или разбавитель добавляли в течение 2 часов, с последующим добавлением PAM3CysSK4 в течение 2 ч, после чего проводят люциферазный анализ. (С) Cpn10 или разбавитель добавляли в течение 2 часов, после чего промывали для удаления Cpn10 или разбавителя, с последующим добавлением PAM3CysSK4 в течение 2 ч, а затем проводили люциферазный анализ. (D) Cpn10 или разбавитель добавляли в течение 2 часов с последующим добавлением PAM3CysSK4 в течение 2 ч, а затем проводили люциферазный анализ. (E) Использовали тот же протокол, что и в (В), за исключением того, что клетки активировали LPS вместо PAM3CysSK4. Верхняя панель показывает данные в условных единицах свечения, а нижняя - процент ингибирования, опосредованного действием Cpn10.
Фиг.6 Cpn10 не связывает TLR4 или TLR2 в отсутствие лиганда. Верхняя панель: TLR4 положительный контроль. MD-2 совместно иммунопреципитирует с TLR4 (но не с TLR2). Нижняя панель: Cpn10 физически не взаимодействует с TLR4 или TLR2 в этих условиях и в отсутствие лиганда.
Фиг.7 Ежедневное подкожное введение Cpn10 уменьшает потерю массы тела при адъювантном артрите у крыс. Среднее значение (± SEM) потери массы тела при адъювантном артрите (n=10 на группу).
Фиг.8 Cpn10-опосредованное воздействие на клетки RAW264.7, стимулированные LPS, не является следствием контаминации LPS (A) в качестве теста на толерантность вследствие LPS контаминации Cpn10, было показано, что предварительная обработка за 2 ч до стимуляции LPS не ингибировала LUC активность. Клетки RAW264-HIV-LTR-LUC предварительно обрабатывали указанными концентрациями LPS в двух параллельных экспериментах; через 2 ч клетки стимулировали действием 5, 1, 0,2 и 0 нг/мл LPS и 2 ч спустя измеряли активность LUC. (B) Обработанный трипсином Cpn10 не ингибировал LPS-индуцированную активность NF-κΒ. Обработка клеток RAW264-HIV-LTR-LUC в двух параллельных экспериментах в течение 2 ч 100 нг/мл Cpn10 (Hsp10) значительно снижала RLU при индукции действием 5, 1 и 0,2 нг/мл LPS по сравнению с клетками, обработанными буфером (Контроль); проценты снижения в RLU после вычитания фонового значения составили 29,7±0,8 (SD), 50±4,6 и 71±7,7 соответственно (p<0,001 с помощью двухфакторного дисперсионного анализа ANOVA, который включал в себя термин для обозначения концентрации LPS). По сравнению с контролем применение Cpn10, обработанного трипсином (Трипсин Hsp10), дало 0,1±8,8, 11,6±4,2 и 21±7,4 а буфера, обработанного трипсином (Трипсиновый буфер) 1,4±2,1, 5,8±1,1 и 14,9±2,4 процент снижения для 5, 1 и 0,2 нг/мл LPS соответственно; (не отличались в значительной степени ни от Контроля, ни друг от друга). Как ожидалось, обработка трипсином стимулирующего LPS не влияла на активность LPS (Трипсин LPS, p>0,05). (С) Cpn10-опосредованное снижение активности LUC, индуцированной LPS, является чувствительным к дозе. Данный эксперимент проводили, как на фиг.1 A, за исключением того, что концентрацию Cpn10 (Hsp10) изменяли, как указано. Для каждой концентрации LPS показан процент ингибирования активности LUC по сравнению с контрольными клетками, предварительно необработанными Cpn10.
Фиг.9 Внутривенная инфузия, а не подкожная инъекция Cpn10 индуцирует изменения величины стимулированного LPS TNF-α ответа клетками PBMC in vitro. A. Стимулированная LPS продукция TNF-α в 0 день (~12 часов перед инфузией) и в 1 день (8 часов после инфузии). B. Данные фиг.9A, графически показанные как изменение стимулированной LPS продукции TNF-α от пре- до пост-инфузии. С. LPS-стимулированная продукция TNF-α в 0 день (~12 часов перед инъекцией) по сравнению с 1 днем (8 часов после подкожной инъекции Cpn10/плацебо).
Фиг.10 Внутривенная инфузия Cpn10 ежедневно в течение 5 дней индуцирует изменения величины LPS-стимулированного TNF-α ответа клетками PBMC in vitro. LPS-стимулированная продукция TNF-α из PBMC, выделенных в 0 день 0 (~12 часов перед инфузией) по сравнению с клетками, выделенными в 1, 4 и 5 день (приблизительно 8 часов после инфузии). Кроме того, PBMC, выделенные на 8 и 12 дни (т.е. 3 дня и 7 дней после конечной инфузии Cpn10 на 5 день соответственно), сравнивали с реакцией, стимулированной LPS, в 0 день. Супернатанты клеточных культур также тестировали на продукцию IL-6 и демонстрировали такую же общую тенденцию, как указано здесь для TNF-α.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение явилось результатом, по меньшей мере, в части, открытия того, что в ряде различных in vitro и in vivo систем человека и мыши Cpn10 ингибируют LPS-опосредованную секрецию провоспалительных цитокинов TNF-α и IL-6 и противовоспалительного цитокина RANTES и увеличивает LPS-индуцированную секрецию противовоспалительного цитокина IL-10 в клетках, в том числе моноцитах, макрофагах и мононуклеарных клетках.
Это противоречит международной публикации WO 02/40038, в которой сделано предположение, что микобактериальный Cpn10 действует для подавления иммунных реакций, опосредованных Th2, путем повышения экспрессии цитокинов, таких как TNF-α и IL-6.
Не привязываясь к какой-либо конкретной теории, в соответствии с настоящим изобретением Cpn10 действует ни через Th1-, ни через Th2-зависимый механизм.
Авторы настоящего изобретения неожиданно продемонстрировали, что Cpn10 влияет на передачу сигнала Toll-подобным рецептором и результирующую секрецию иммуномодулятора в ответ на агонист Toll-подобного рецептора.
Более конкретно, Cpn10 снижает активацию NF-κΒ, стимулированную агонистом Toll-подобного рецептора, и секрецию TNF-α и RANTES и повышает секрецию IL-10 дозозависимым образом.
Кроме того, авторы данного изобретения также продемонстрировали, что Cpn10, Toll-подобные рецепторы и агонисты Toll-подобных рецепторов образуют молекулярный комплекс. Анализ с использованием флуоресцентного резонансного переноса энергии (FRET-анализ) дает возможность предположить, что возможно прямое взаимодействие между Cpn10 и Toll-подобным рецептором или что они находятся, по меньшей мере, в очень тесной пространственной близости (в пределах 1-10 нм друг от друга), в присутствии лиганда TLR (например, LPS).
Будет понятно, что агонист Toll-подобного рецептора может представлять собой патоген (такой как бактерия или вирус), полученную из него молекулу, или продуцируемую патогеном (такую как бактериальный LPS, бактериальный эндотоксин, микобактериальный липоарабиноманнан, липотейхоевая кислота, липопептид или двухспиральная РНК), или может представлять собой синтетический липопептидный агонист Toll-подобного рецептора, такой как пептид/аминокислота, связанная с длинноцепочечными ацилированными насыщенными жирными кислотами, такими как лауриновая и пальмитиновая кислота, например агонист TLR2 PAM3CysSK4.
В данном изобретении под «иммуномодулятором» подразумевается молекулярный медиатор, секретируемый клетками иммунной системы или молекулярный медиатор, который взаимодействует с клетками иммунной системы, который играет роль в активации, поддержании, созревании, ингибировании, супрессии или повышении иммунного ответа.
Под «цитокином» подразумевается молекулярный медиатор, секретируемый клетками иммунной системы, который играет роль в активации, поддержании, созревании, ингибировании, супрессии или повышении иммунного ответа. Неограничивающими примерами цитокинов являются TNF-α, интерлейкин-6 (IL-6), интерлейкин-12 (IL-12), интерлейкин-1β (IL-1β) и интерлейкин-10 (IL-10).
Под «хемокином» подразумевается молекулярный медиатор, который способствует и/или регулирует клеточную миграцию и активацию. Неограничивающими примерами хемокинов являются MIP1α, MIP1β, RANTES и TGF-β.
Под «провоспалительным иммуномодулятором» подразумевается цитокин или хемокин, который играет роль или вовлечен в процесс воспаления или воспалительную реакцию.
Неограничивающими примерами провоспалительных иммуномодуляторов являются IL-6, TNF-α, IL-12 и IL-1β, RANTES и MIP1β.
Под «противовоспалительным иммуномодулятором» подразумевается цитокин или хемокин, который играет роль в ингибировании, супрессии или ином уменьшении воспалительной реакции.
Неограничивающими примерами противовоспалительных иммуномодуляторв являются IL-10 и TGF-β.
Под «выделенным» подразумевается вещество, которое удалили из его природного состояния или иным образом подвергли проводимым людьми манипуляциям. Выделенное вещество может быть по существу или в значительной степени свободно от компонентов, которые обычно сопутствуют ему в природном состоянии, или может быть подвергнуто обработке таким образом, чтобы оно находилось в искусственно созданном состоянии вместе с компонентами, которые обычно сопровождают его в его природном состоянии. Выделенное вещество может быть в природной, химически синтезированной или рекомбинантной форме.
Под «белком» подразумевается аминокислотный полимер. Аминокислоты могут быть природными или неприродными аминокислотами D- или L- аминокислотами, хорошо изученными в данной области.
«Пептид» представляет собой белок, имеющий менее пятидесяти (50) аминокислот.
«Полипептид» представляет собой белок, имеющий более пятидесяти (50) аминокислот.
Используемый здесь термин «нуклеиновая кислота» означает одно- или двухспиральную мРНК, РНК, РНКi и ДНК, в том числе кДНК и геномную ДНК.
Cpn10 и фрагменты, варианты и производные Cpn10
В соответствии с настоящим изобретением «Cpn10» или «шаперонин 10» относится к любому эукариотическому Cpn10, в том числе Cpn10 млекопитающих, например человека, мыши, крысы или другим формам Cpn10.
Предпочтительно Cpn10 представляет собой Cpn10 млекопитающих.
Более предпочтительно Cpn10 представляет собой Cpn10 человека.
Cpn10 белок может содержать приpодную модификацию, такую как гликозилирование или ацетилирование, и/или быть в нативной, химически синтезированной или рекомбинантной форме. Также будет понятно, что Cpn10 также может называться «Hsp10». Они могут рассматриваться как относящиеся к одному и тому же белку.
В соответствии с данным изобретением также могут быть использованы фрагменты Cpn10.
В одном варианте осуществления «фрагмент» включает в себя аминокислотную последовательность, которая составляет менее 100%, но по меньшей мере 30%, предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 80% или еще более предпочтительно по меньшей мере 90%, 95% или 98% белка Cpn10.
Термин «фрагмент» включает в себя и охватывает «биологически активный фрагмент», который сохраняет биологическую активность белка Cpn10. Например, биологически активный фрагмент Cpn10, способный регулировать передачу сигнала Toll-подобным рецептором и/или секрецию иммуномодулятора, может быть использован в соответствии с данным изобретением. Биологически активный фрагмент составляет по меньшей мере более 50% биологической активности целого белка Cpn10, предпочтительно по меньшей мере более 60% биологической активности, более предпочтительно по меньшей мере более 75% биологической активности, еще более предпочтительно по меньшей мере более 80% биологической активности и преимущественно по меньшей мере 90% или 95% биологической активности Cpn10.
Используемый здесь термин «вариантные» белки представляют собой белки, в которых одна или несколько аминокислот заменены другими аминокислотами. Варианты белков Cpn10, которые сохраняют биологическую активность нативного Cpn10 или Cpn10 дикого типа, могут быть использованы в соответствии с данным изобретением. В данной области хорошо известно, что некоторые аминокислоты могут быть изменены на другие с приблизительно сходными свойствами без изменения характера активности белка (консервативные замены). В основном замены, которые вероятно вызывают наибольшие изменения свойств полипептидов, представляют собой замены, в которых (a) гидрофильный остаток (например, Ser или Thr) замещен гидрофобным остатком или на гидрофобный остаток (например, Leu, Ile, Phe или Val); (b) цистеин или пролин замещен любым другим остатком или на любой другой остаток; (c) остаток, имеющий электроположительную боковую цепь (например, Arg, His или Lys), замещен электроотрицательным остатком, или на электроотрицательный остаток (например, Glu или Asp), или (d) остаток, имеющий объемную боковую цепь (например, Phe или Trp) замещен остатком или на остаток, имеющий меньшую боковую цепь, (например, Ala, Ser) или без боковой цепи (например, Gly).
В отношении вариантов Cpn10, их можно получить с помощью мутагенеза белка Cpn10 или мутагенезом кодирующей нуклеиновой кислоты, как, например, случайным мутагенезом или сайт-направленным мутагенезом. Примеры методов мутагенеза нуклеиновой кислоты приведены в Главе 9 публикации CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, Ausubel et al., supra, которая включена здесь в качестве ссылки.
Используемые здесь «производные» белки Cpn 10 данного изобретения включают в себя белки, которые были изменены, например, конъюгацией или образованием комплекса с другими химическими частями молекулы или с использованием методик пост-трансляционной модификации, хорошо известных в данной области, в том числе слитые клеточные белки.
Другие производные, рассматриваемые данным изобретением, включают в себя, но не только, пегилирование, модификацию по боковым цепям, включение неприродных аминокислот и/или их производных во время синтеза белка и применение кросс-линкеров и другие способы, которые накладывают конформационные ограничения на белок Cpn10. Примеры модификаций боковых цепей, рассматриваемых настоящим изобретением, включают в себя модификации аминогрупп, например, ацилированием с использованием уксусного ангидрида; ацилирование аминогрупп янтарным ангидридом и тетрагидрофталевым ангидридом; амидирование метилацетимидатом; карбамоилирование аминогрупп цианатом; пиридоксилирование лизина пиридоксаль-5-фосфатом с последующим восстановлением с использованием NaBН4; восстановительное алкилирование взаимодействием с альдегидом с последующим восстановлением с использованием NaBН4; и тринитробензилирование аминогрупп 2,4,6-тринитробензолсульфоновой кислотой (TNBS).
Карбоксильная группа может быть модифицирована активацией карбодиимидом через образование O-ацилизомочевины, за которым следует последовательное получение производных, например, до соответствующего амида.
Гуанидиновая группа остатков аргинина может быть модифицирована образованием гетероциклических конденсированных продуктов с такими реактивами, как 2,3-бутандион, фенилглиоксаль и глиоксаль.
Сульфгидрильные группы могут быть модифицированы такими способами, как окисление надмуравьиной кислотой до цистеиновой кислоты; образование производных ртути с использованием 4-хлормеркурифенилсульфоновой кислоты, 4-хлормеркурибензоата; 2-хлормеркури-4-нитрофенола, хлорида фенилртути и других соединений ртути; образование смешанных дисульфидов с другими тиольными соединениями; реакция с имидом малеиновой кислоты, малеиновым ангидридом или другим замещенным имидом малеиновой кислоты; карбоксиметилирование иодуксусной кислотой или иодацетамидом; и карбамоилирование цианатом при щелочном pH.
Остатки триптофана могут быть модифицированы, например, алкилированием индольного кольца 2-гидрокси-5-нитробензилбромидом или сульфонилгалидами или окислением N-бромсукцинимидом.
Остатки тирозина могут быть модифицированы нитрованием тетранитрометаном до образования 3-нитро производного тирозина.
Имидазольное кольцо остатка гистидина может быть модифицировано N-карбэтоксилированием диэтилпирокарбонатом или алкилированием производными иодуксусной кислоты.
Примеры включения неприродных аминокислот и производных во время синтеза белка включают в себя, но не только, применение 4-аминомасляной кислоты, 6-аминокапроновой кислоты, 4-амино-3-гидрокси-5-фенилпентановой кислоты, 4-амино-3-гидрокси-6-метилгептановой кислоты, т-бутилглицина, норлейцина, норвалина, фенилглицина, орнитина, саркозина, 2-тиенилаланина и/или D-изомеров аминокислот.
Производные также могут включать в себя слитые клетки и короткие пептидные последовательности эпитопа (эпитоп-тэги). Хорошо известные примеры слитых клеток включают в себя, но не только, глутатион-S-трансферазу (GST), Fc часть IgG человека, белок, связывающий мальтозу (MBP) и гексагистидин (HIS6), которые особенно применимы для выделения слитого белка методом аффинной хроматографии. Для очистки слитого полипептида методом аффинной хроматографии подходящими матрицами для аффинной хроматографии являются глутатион-, амилоза- и никель- или кобаль-конъюгированные смолы соответственно. Многие из этих матриц доступны в виде «набора», такого как QIAexpress™ system (Qiagen), используемые с (HIS6) слитыми клетками, и система очистки Pharmacia GST.
Один конкретный пример слитой клетки представляет собой GST, описанный у Morton el al., 2000, Immunol Cell Biol 78 603-607. В некоторых случаях слитые клетки имеют сайты расщепления протеазой, например, для Фактора Xa или тромбина, которые дают возможность соответствующей протеазе частично переваривать слитый полипептид по данному изобретению и тем самым высвобождать из него рекомбинантный белок Cpn10. Высвобожденный белок Cpn10 затем может быть выделен из слитой клетки последовательным хроматографическим разделением. При расщеплении GST-Cpn10 тромбином, например, образуется производный белок GSM-Cpn10.
Слитые клетки в соответствии с данным изобретением также включают в своем объеме «эпитоп-тэги», которые обычно представляют собой короткие пептидные последовательности для каждого доступного специфичного антитела. Хорошо известными примерами «эпитоп-тэгов», для которых специфические моноклональные антитела легкодоступны, включают в себя c-myc, гемагглютинин и FLAG-тэги.
Белки Cpn10 в соответствии с данным изобретением (в том числе фрагменты, варианты, производные и гомологи) могут быть получены с помощью любого подходящего способа, известного специалистам в данной области, в том числе химическим синтезом и рекомбинантной экспрессией.
Предпочтительно Cpn10 представляет собой Cpn10.
Например, рекомбинантный белок Cpn10 может быть получен способом, включающим стадии:
(i) получения экспрессионной конструкции, которая включает в себя выделенную нуклеиновую кислоту, кодирующую Cpn10, функционально связанную с одной или несколькими регуляторными нуклеотидными последовательностями в экспрессионном векторе;
(ii) трансфектирования или трансформирования подходящей клетки-хозяина данной экспрессионной конструкцией; и
(iii) экспрессии рекомбинантного белка в указанной клетке-хозяине.
Данный способ, описанный у Morton et al., 2000, supra является примером способа получения рекомбинантного белка Cpn10.
Способы лечения и фармацевтические композиции
Настоящее изобретение относится к способам, в соответствии с которыми Cpn10-опосредованная регуляция передачи сигнала Toll-подобным рецептором и, более конкретно секреция иммуномодулятора, может быть использована для профилактического или терапевтического лечения заболеваний, нарушений или состояний, чувствительных к такому лечению.
Такие заболевания, нарушения или состояния могут быть вызваны чрезмерными уровнями провоспалительных цитокинов и хемокинов и, следовательно, могут быть чувствительными к ингибированию, супрессии или иному уменьшению передачи сигнала Toll-подобным рецептором.
Альтернативно или дополнительно, такие заболевания, нарушения или состояния могут быть чувствительны к увеличению, облегчению, стимуляции или иному усилению передачи сигнала Toll-подобным рецептором и секреции противовоспалительного цитокина и хемокина.
Например, Cpn10 уменьшает продукцию TNF-α и RANTES и увеличивает продукцию IL-10 в in vivo модели нелетальной эндотоксинемии у мышей. Cpn10 также обладает значительной иммуносупрессивной активностью в in vivo трансплантационной модели у мышей, и лечение с использованием Cpn10 увеличивает коэффициент выживаемости мышей с болезнью «трансплантат против хозяина».
Cpn10 также снижает кахексию у мышей с адъювантным артритом. Повышенные уровни воспалительных цитокинов связаны с кахексией при ряде заболеваний, таких как рак и ревматоидный артрит.
Кроме того, Cpn10 улучшает заживление ран в in vivo модели у мышей.
Cpn10, введенный в виде однократной внутривенной дозы людям in vivo, заметно снижает ответ провоспалительных цитокинов после LPS-стимуляции ex vivo дозозависимым образом, ясно показывая, что Cpn10 оказывает иммуномодулирующее действие у пациентов в клинических испытаниях на людях.
Чрезмерные воспалительные или неконтролируемые иммунные реакции наносят вред хозяину, поскольку ряд систем отрицательной обратной связи образовались для торможения продукции провоспалительных медиаторов. Один такой механизм обратной связи включает в себя IL-10, важный иммунорегуляторный цитокин, секретируемый мононуклеарными клетками и моноцитами, который принимает участие в ограничении воспалительных реакций и индукции иммунологической толерантности.
Cpn10 ингибирует, но не прекращает секрецию TNF-á, IL-6 или RANTES. Это благоприятное свойство, поскольку полное удаление TNF-α (например, анти-TNF-α антителами) в результате может приводить к аномальному иммунитету, предрасположенности пациентов инфекции и снижению противоопухолевого иммунитета, что может предрасполагать пациентов к увеличению частоты возникновения опухолей.
Способность Cpn10 снижать продукцию и/или секрецию провоспалительных иммуномодуляторов указывает на то, что Cpn10 найдет терапевтическое применение при состояниях, при которых избыточная секреция провоспалительных иммуномодуляторов приводит к заболеванию.
Многие заболевания связаны с чрезмерным или хроническим воспалением, следовательно, модуляция передачи сигнала TLR рецептором, в результате приводящая к модуляции секреции цитокинов, может иметь широкое клиническое применение. Например, чрезмерная секреция провоспалительных цитокинов, таких как TNF-α, является одной из ведущих причин смертности при острых состояниях, таких как септический шок, и является одним из основных факторов, способствующих непрерывному повреждению тканей при хронических воспалительных заболеваниях, таких как воспалительное заболевание кишечника (IBD), артрит, псориаз, застойное заболевание сердца, рассеянный склероз и хроническое обструктивное заболевание легких.
При трансплантации ткани или органа лимфоциты хозяина или донора могут распознавать антигены клеток донора или хозяина, соответственно, как чужеродные, и высвобождение цитокинов, которые активируют клетки врожденной иммунной системы, в результате приводит к отторжению трансплантированной ткани или органа или болезни «трансплантат против хозяина». Иммуносупрессирующие лекарственные средства играют большую роль в терапевтическом лечении и лечении отторжения трансплантата или болезни «трансплантат против хозяина». Однако лекарственные средства вызывают тяжелые побочные эффекты у пациентов, являются очень дорогими и у некоторых пациентов они малоэффективны.
Cpn10, в том числе варианты, фрагменты и производные Cpn10, можно вводить животным in vivo или вводить in vitro в одну или несколько клеток, тканей или органов, полученных от животного для регуляции, таким образом, передачи сигнала Toll-подобным рецептором, и/или продукции, и/или секреции иммуномодулятора, индуцируемого Toll-подобным рецептором.
Обычно, но не исключительно, Cpn10 включая варианты, производные и фрагменты, можно доставлять в виде фармацевтических композиций, которые дополнительно содержат соответствующий фармацевтически приемлемый носитель, разбавитель или эксципиент.
Под «фармацевтически приемлемым носителем, разбавителем или эксципиентом» подразумевается твердый или жидкий наполнитель, разбавитель или инкапсулирующее вещество, которое можно безопасно использовать при системном введении. В зависимости от конкретного пути введения можно использовать различные носители, хорошо известные в данной области. Эти носители могут быть выбраны из группы, включающей в себя сахара, крахмалы, целлюлозу и ее производные, солод, желатин, тальк, сульфат кальция, растительные масла, синтетические масла, полиолы, альгиновую кислоту, фосфатные буферные растворы, эмульгаторы, физиологический раствор и соли, такие как соли неорганической кислоты, в том числе гидрохлориды, бромиды и сульфаты органических кислот, такие как ацетаты, пропионаты и малонаты и апирогенную воду.
Полезной ссылкой, описывающей фармацевтически приемлемые носители, разбавители и эксципиенты, является Remington's Pharmaceutical Sciences (Mack Publishing Co. N.J. USA, 1991), которая включена здесь как ссылка.
Для обеспечения пациента композицией по данному изобретению может быть использован любой безопасный путь введения. Например, могут быть использованы пероральный, ректальный, парентеральный, сублингвальный, буккальный, внутривенный, внутрисуставной, внутримышечный, внутрикожный, подкожный, ингаляционный, внутрибрюшинный, интрацеребровентрикулярный, чрескожный и подобные. Внутримышечная и подкожная инъекции подходят, например, для введения иммуногенных композиций, вакцин и ДНК вакцин.
Лекарственные формы включают в себя таблетки, дисперсии, суспензии, инъекции, растворы, сиропы, пастилки, капсулы, суппозитории, аэрозоли чрескожные пластыри и подобное. Эти лекарственные формы также могут включать в себя инъекционное введение или имплантацию с устройством для контролируемого высвобождения, созданные специально для этой цели, или другие формы имплантатов, модифицированные для работы дополнительно таким образом. Контролируемое высвобождение терапевтического агента может осуществляться покрытием его, например, гидрофобными полимерами, в том числе акриловыми смолами, восками, высшими алифатическими спиртами, полимолочной и полигликолевой кислотами и некоторыми производными целлюлозы, такими как гидроксиметилцеллюлоза. Кроме того, контролируемое высвобождение можно осуществить с использованием других полимерных матриц, липосом и/или микросфер.
Вышеуказанные композиции можно вводить способом, совместимым с лекарственной формой, и в таком количестве, которое является фармацевтически эффективным. Доза, вводимая пациенту, в контексте настоящего изобретения, должна быть достаточной, чтобы вызвать благоприятный ответ у пациента на протяжении соответствующего периода времени. Количество вводимого агента (агентов) может зависеть от пациента, которому проводят лечение, в том числе от возраста, пола, массы тела и общего состояния здоровья, факторов, которые будут зависеть от мнения лечащего врача.
Фармацевтические композиции и способы лечения в соответствии с данным изобретением могут подходить для медицинского и/или ветеринарного использования и, соответственно, применяться на практике у людей и животных, не относящихся к человеку, в том числе таких млекопитающих, как люди, домашний скот, домашние животные и служебные животные, но не только.
Агонисты и антагонисты Cpn10
Настоящее изобретение рассматривает способы разработки, создания, скрининга или иного получения агониста или антагониста Cpn10.
Пояснение действия Cpn10 при передаче сигнала Toll-подобным рецептором и секреции иммуномодуляторов обеспечивает новую и неожиданную возможность, при которой агонисты и антагонисты Cpn10 могут быть специфически созданы или подвергнуты скринингу в соответствии с их воздействием на передачу сигнала Toll-подобным рецептором и секрецию иммуномодуляторов. В принципе, эта иммунорегуляторная активность может не зависеть от активности шаперона, которой также обладает Cpn10.
«Агонист» относится к молекуле, которая усиливает активность другой молекулы или рецепторного сайта.
Обычно агонист Cpn10 супрессирует, снижает или иным образом ингибирует секрецию провоспалительного цитокина или хемокина, как правило, индуцируемую агонистом Toll-подобного рецептора.
Обычно агонист Cpn10 увеличивает, облегчает или иным образом повышает секрецию противовоспалительного цитокина или хемокина, как правило, индуцируемую агонистом Toll-подобного рецептора.
«Антагонист» относится к молекуле, которая блокирует или ингибирует активность другой молекулы или рецепторного сайта.
Обычно антагонист Cpn10 супрессирует, снижает или иным образом ингибирует способность Cpn10 к отрицательной регуляции секреции провоспалительного цитокина или хемокина, индуцированной агонистом Toll-подобного рецептора.
Обычно антагонист Cpn10 ингибирует способность Cpn10 положительно регулировать индукцию секреции противовоспалительного цитокина или хемокина агонистом Toll-подобного рецептора.
В одном варианте осуществления агонист или антагонист может быть миметиком Cpn10, хотя данное изобретение не ограничивается агонистами и/или антагонистами, имеющими структурное сходство с Cpn10.
Используемый здесь «миметик» относится к молекуле, которая имеет сходство с одной или несколькими конкретными структурами и/или функциональными областями или доменами Cpn10 и включает в себя модифицированные формы Cpn10, обладающие агонистической или антагонистической активностью.
В одной конкретной форме настоящее изобретение относится к способу, в котором агонисты и/или антагонисты Cpn10 могут быть разработаны, созданы, подвергнуты скринингу или получены иным образом путем определения, регулирует ли возможный агонист и/или антагонист Cpn10 передачу сигнала Toll-подобным рецептором или секрецию иммуномодуляторов, индуцируемую Toll-подобным рецептором.
Будет понятно, что в соответствии с данным изобретением передачу сигнала Toll-подобным рецептором и продукцию и/или секрецию иммуномодуляторов можно измерить или определить на уровне генной экспрессии (например, продукцию РНК эндогенного цитокина или хемокина), путем измерения или определения внутриклеточно или внеклеточно секретируемого белка, анализами репортерного гена и анализами, которые определяют или оценивают внутриклеточные сигнальные молекулы.
В одном конкретном варианте осуществления данное изобретение относится к способу, в котором передачу сигнала Toll-подобным рецептором оценивают или определяют в соответствии с активностью NF-kB in vitro.
В другом конкретном варианте осуществления данное изобретение относится к способу, в котором передачу сигнала Toll-подобным рецептором оценивают или определяют по секреции одного или нескольких иммуномодуляторов, таких как IL-6, TNF-α, RANTES, IL-10, IL-12 и продукции и/или секреции IL-1α.
В другом конкретном варианте осуществления данное изобретение относится к способу, в котором передачу сигнала Toll-подобным рецептором оценивают или определяют по c-Jun-N-концевой киназе и/или внеклеточному сигналу, связанному с передачей сигнала киназой.
В соответствии с данным изобретением выделенный молекулярный комплекс, содержащий Cpn10, Toll-подобный рецептор и агонист Toll-подобного рецептора может быть использован для получения, создания или скрининга агониста или антагониста Cpn10.
В отдельной форме выделенный молекулярный комплекс содержит Cpn10, TLR-4 и LPS.
Только для примера, возможный агонист может быть идентифицирован по способности образовывать молекулярный комплекс с Toll-подобным рецептором и агонистом Toll-подобного рецептора.
Только для примера, возможный антагонист может быть идентифицирован по способности предотвращать или разрушать образование молекулярного комплекса, содержащего Cpn10, Toll-подобный рецептор и агонист Toll-подобного рецептора.
С учетом вышеизложенного будет понятно, что существует несколько методик, которые могут облегчать получение агонистов и/или антагонистов Cpn10 в соответствии с данным изобретением.
Неограничивающие примеры включают в себя скрининг молекулярных библиотек, таких как библиотеки синтетических химических соединений, в том числе комбинаторные библиотеки, такими способами, которые описаны у Nestler & Liu, 1998, Comb. Chem. High Throughput Screen. 1, 113 и Kirkpatrick et al., 1999, Comb. Chem. High Throughput Screen 2 211.
Также предполагается, что библиотеки природных молекул могут быть подвергнуты скринингу по такой методике, которая рассмотрена у Kolb, 1998, Prog. Drug. Res. 51 185.
Больше структурных подходов может быть использовано при скрининге структурных баз данных с использованием компьютера, компьютерного моделирования и/или дизайна или более традиционных биофизических методик, которые определяют взаимодействия молекулярных связей, которые хорошо известны в данной области.
Поиск по структурным базам данных с использованием компьютера, моделирование и дизайн все больше и больше используются в качестве методики для разработки молекул агонистов и антагонистов. Примеры способов поиска по базам данных можно найти в международной публикации WO 94/18232 (направлена на получение миметиков антигенов ВИЧ), патенте Соединенных Штатов № 5752019 и международной публикации WO 97/41526 (направлена на идентификацию миметиков EPO), каждая из которых включена здесь в качестве ссылки.
В основном, другие подходящие способы включают в себя любую из разнообразных биофизических методик, которые идентифицируют молекулярные взаимодействия. Способы, подходящие для потенциально полезных методик, таких как методы конкурентного радиолигандного связывания, аналитическое ультрацентрифугирование, микрокалориметрия, поверхностный плазмонный резонанс и оптические способы на основе биосенсоров представлены в Части 20 CURRENT PROTOCOLS IN PROTEIN SCIENCE Eds. Coligan et al., (John Wiley & Sons, 1997), включенной здесь в качестве ссылки.
Для лучшего понимания настоящего изобретения и практического осуществления специалисту рекомендуется обратиться к следующим неограничивающим примерам.
ПРИМЕРЫ
Пример 1
Модуляция Cpn10 цитокинов и хемокинов, стимулированных LPS
Материалы и методы
Получение и очистка Cpn10
Рекомбинантный Cpn10 человека (GenBank поступление № X75821) получали в E. Coli, в основном, как описано Ryan et al. (Ryan et al., 1995, J Biol Chem, 270 22037-22043). Кроме того, вещество, которое не связывало Macro-Prep High Q (BioRad), в дальнейшем очищали S-Сефарозой, а затем гельфильтрацией хроматографировано в геле (Superdex 200, Amersham Biosciences). Очищенный Cpn10 в 50 мМ Трис-HCl (pH 7,6) и 150 мМ NaCl буфере фильтровали через Acrodisc с 0,2 мм мембраной Mustang E в соответствии с инструкциями производителя (Pall Corporation, Ann Arbor, MI. Cat No. MSTG5E3) для удаления остаточных эндотоксинов и хранили при -70ºC. Определяли чистоту Cpn10, которая составила >97%, с помощью SDS-PAGE. Аликвоты размораживали один раз перед использованием. Все пробы Cpn10 проявляли такую же молярную активность, как E. coli GroES в методах рефолдинга роданезы, опосредованного GroEL (Brinker et a/.,2001, Cell, 107 223-233) (данные не показаны). Загрязнение Cpn10 липополисахаридом определяли методом амебоцитного лизата (Limulus Amebocyte Lysate) (BioWhittaker, Walkersville, MD), составляет <1 EU/мг в очищенном Cpn10 белке.
Опухолевые клеточные линии
Было показано, что клеточные линии K562 (эритролейкоз человека), Mono Mac 6 (моноцитарная линия человека), U937 (гистиоцитарная лимфома человека), P815 (мастоцитома мыши), EL4 (Т-клеточная лимфома мыши), Jurkat (Т-клеточный лейкоз человека), RAW 264,7 (ATCC TIB 71, макрофаг мыши), L929 (фибросаркома мыши), В16 (меланома мыши), HeLa (карцинома шейки матки человека) и MCA-2 (фибросаркома мыши) являются негативными по микоплазме. Клетки выращивали в питательной среде, свободной от эндотоксинов, содержащей RPMI 1640 (Gibco Labs, Life Technologies, Grand Islin, N.Y., USA), 10% эмбриональную сыворотку теленка (Life Technologies), 2 мМ глютамина (Sigma), 10 мМ HEPES (Sigma), 100 мкг/мл стрептомицина и 100 МЕ/мл пенициллина (CSL Ltd, Melbourne, Australia).
Биоанализ RAW264-HIV-LTR-LUC
Клетки RAW264-HIV-LTR-LUC культивировали в присутствии G418 (200 мкг/мл) в течение одной недели после восстановления из жидкого азота и выращивали как суспензионные культуры в 25 см3 флаконах (Greiner Labortechnik, Frickenhausen, Германия). Клетки RAW264-HIV-LTR-LUC дезагрегировали повторным пипетированием и высеивали в концентрации 2,5×105 клеток на лунку в 24-луночные планшеты и инкубировали в течение ночи (37ºC и 5% СО2). LPS из E. coli (Sigma L-6529. Штамм 055:B5, Sigma) растворяли в стерильной дистиллированной воде и хранили при 4ºC в стеклянных пробирках по 1 мг/мл. Непосредственно перед применением раствор интенсивно встряхивали перед взятием аликвот. Cpn10 добавляли к клеточным культурам в течение двух часов с последующим добавлением LPS в указанных концентрациях, а затем дополнительно 2 ч адгезивные клетки подвергали люциферазному исследованию (Luciferase Assay System, Promega, Madison, WI). Люциферазную активность считывали в течение 15 с на Люменометре Turner Designs TD 20/20.
Исследование RAW264.7 IL-6 и RANTES
RAW264.7 клетки высеивали 2,5×105 клеток на лунку в 24-луночные планшеты и культивировали в течение ночи при 37ºC и 5% CО2. Cpn10 или буфер добавляли к клеткам троекратно в течение 2 ч с последующим добавлением LPS (1 нг/мл). Через шесть часов супернатанты собирали и 3-кратно анализировали продукцию RANTES и IL-6 с использованием набора Duoset ELISA (R & D Systems). Оптическую плотность (при 450 нм) каждого образца определяли с помощью считывающего устройства для микропланшетов (Magellan 3, Sunrise - Tecan, Durham, NC).
Продукция цитокинов клетками селезенки и макрофагами, полученными от животных, обработанных Cpn10
Мышей C57BL/6 (H-2b, Ly-5,2+) приобретали в Австралийском Исследовательском Центре (Perth, Западная Австралия, Австралия), а мыши линии C57BL/6 IL-10-/- (H-2b, Ly-5,2+) предоставлены Австралийским Государственным Университетом (Canberra, Австралия). Используемая культуральная среда представляла собой 10% FCS/IMDM (JRH Biosciences, Lenexa, KS), дополненную 50 ед/мл пенициллина, 50 мкг/мл стрептомицина, 2 мМ L-глутамином, 1 мМ пируватом натрия, 0,1 мМ неприродной аминокислотой, 0,02 мМ β-меркаптоэтанолом и 10 мМ HEPES, и клетки культивировали при pH 7,75, 37ºC и 5% CО2. Мышам линии C57BL/6 (n=3 на группу) проводили подкожные инъекции Cpn10 (100 мкг) или разбавителя ежедневно в течение 5 дней, перитонеальные макрофаги собирали на следующий день перитонеальным лаважем и объединяли от отдельных животных внутри испытуемой группы. Клетки высеивали трижды 2×l05 на лунку в присутствии LPS (1 мкг/мл). Культуральные супернатанты собирали через 5 часов и уровни TNF-α определяли методом ELISA (см. ниже). Результаты приводили к продукции на 105 макрофагов, исходя из CD11b окрашивания методом FACS поглощенных клеток. Для определения IL-10 клетки селезенки собирали от тех же животных и объединяли, как показано выше, и культивировали трехкратно 5×105 на лунку в отсутствие (не показано) или присутствии LPS (10 мкг/мл). Культуральные супернатанты собирали через 48 часов и определяли уровень IL-10 методом ELISA (см. ниже).
Исследования цитокинов мышиных клеток, стимулированных LPS in vitro
Пары моноклональных антител, используемых в ELISA анализе TNF-α и IL-10, приобретали на фирме PharMingen (San Diego, CA) и использовали в концентрациях, рекомендованных производителем. Супернатанты разводили в питательной среде 1:1 IL-10 и TNF-α. Цитокины захватывали захватывающие антитела, и определяли прямым биотин-меченным детектирующим антителом. Меченную стрептавидином пероксидазу хрена (Kirkegaard и Perry laboratories, Gaithersburg, MD) и питательную среду (Sigmafast OPD) в дальнейшем использовали для количественного определения иммобилизованного биотина. Планшеты считывали при 492 нм с помощью микропланшетного ридера Spectraflour Plus (Tecan). Рекомбинантные цитокины (PharMingen) использовали в качестве стандартов для ELISA анализа. Стандарты анализировали в двух параллелях, и чувствительность методов составила 15 пг/мл для IL-10 и TNF-α.
Исследование TNF-α и IL-6 PBMC человека
Мононуклеарные клетки периферической крови человека (PBMC) выделяли из гепаринизированной крови от здоровых добровольцев путем центрифугирования в градиенте плавучей плотности на Ficoll-Hypaque. PBMC распределяли 106 жизнеспособных клеток на мл в 200 мкл в 96-луночных планшетах для тканевых культур (Greiner). Затем добавляли Cpn10 и планшеты инкубировали в течение 1 ч, с последующим добавлением LPS и дополнительной инкубацией 20 ч при 37ºC, 5% CO2, после чего супернатанты собирали и параллельные пробы анализировали на продукцию TNF-á и IL-6 (Duoset набор для ELISA; R&D Systems). Чувствительность этих анализов составила 31 пг/мл для TNF-α и 9 пг/мл для IL-6.
Исследование TNF-α, RANTES и IL-10 в сыворотке мышей после инъекционного введения LPS
Самок мышей линии BALB/c в возрасте 8-10 недель (Animal Resource Centre, Perth, Австралия) помещали под нагревательную лампу приблизительно на 10 минут, затем фиксировали и внутривенно вводили Cpn10 в определенных дозах. Через 30 минут внутривенно вводили 10 мкг LPS, используя тот же протокол. Через 1,5 часа после введения LPS путем сердечной пункции кровь собирали в 1 мл пробирки, ускоряющие коагуляцию (MiniCollect, Interpath), и хранили при 4ºС для определения сывороточного TNF-α и RANTES с использованием набора для ELISA (R&D Systems). Продукцию IL-10 в сыворотке измеряли с помощью специфического ELISA OptEIA IL-10 мыши (BD Biosciences Pharmingen).
Пересадка костного мозга и болезнь "трансплантат против хозяина" (GVHD)
Самок мышей возрастом 8-14 недель линий C57BL/6 (B6, H-2b, Ly-5,2+), B6 Ptprca Ly-5a(H-2b, Ly-5,1+) и B6D2F1 (H-2b/d, Ly-5.2+) приобретали в Австралийском Исследовательском Центре (Perth, Западная Австралия, Австралия). Cpn10 (100 мкг на животного) или контрольный разбавитель вводили животным донорам и реципиентам подкожно ежедневно на протяжении пяти дней до пересадки. Мышей размещали в стерильные микроизоляционные клетки и им давали стерильную подкисленную воду (pH 2,5) и обычный корм в течение первых двух недель после трансплантации. Мышам производили трансплантацию в соответствии со стандартным протоколом, описанным ранее (Hill et al., 1997, Blood, 90 3204-3213; Hill et al., 1998, J Clin Invest, 102, 115-123). Вкратце, в 1 день мышам линии B6D2F1 проводили суммарное облучение тела 1300 сГр (137Cs источник при 108 сГр/мин), которое разделяли на две дозы с промежутком в 3 ч, для сведения к минимуму желудочно-кишечной токсичности. Донорский костный мозг (5×106) и донорские T-клетки селезенки (2×106), очищенные на нейлоновой вате, ресуспендировали в 0,25 мл среды Leibovitz L-15 (Gibco BRL, Gaithersburg MD) и путем инъекции вводили каждому реципиенту. Ежедневно контролировали выживаемость и еженедельно проводили клиническую оценку GVHD. Степень системной GVHD определяли с помощью системы количественных показателей, которая суммирует изменения по пяти клиническим параметрам; потеря массы тела, положение тела (согнутое), активность, качество шерсти и целостность кожи (максимальный показатель=10) (Hill et al., 1997, supra; Hill et al., 1998, J Clin Invest, 102 115-123; Cook et al., 1996, Blood, 88 3230-3239). Отдельным мышам проставляли на уши метку и сортировали от 0 до 2 по каждому критерию без информации об обработанных группах. Животных с тяжелой клинической формой GVHD (показатели >6) скарифицировали в соответствии с этическими нормами и день смерти считали следующим днем.
Статистический анализ
Статистическую обработку полученных результатов проводили с использованием одномерного дисперсионного анализа (ANOVA), t-критерия Стьюдента или логарифмической зависимости ранговой статистики с использованием SPSS для Windows 11.5.0 (SPSS Вc.).
Результаты
Ингибирование передачи сигнала LPS, под действием Cpn10, с использованием клеток-индикаторов RAW264-HIV-LTR-LUC
Для исследования роли Cpn10 как иммуносупрессирующего агента изучали способность Cpn10 ингибировать активацию NF-кВ, опосредованную LPS. Клетки линии RAW264-HTV-LTR-LUC представляют собой клеточную линию макрофагов мыши (RAW264.7), стабильно экспрессирующих репортерный ген люциферазы с HIV промотором длинного концевого повтора, которые сильно быстро реагируют на стимуляцию NF-кВ. Данные клетки обеспечивают чувствительный биологический анализ путей передачи сигнала TLR4 в макрофагах, стимулированных бактериальным LPS (Sweet & Hume, 1995, J Inflamm, 45 126-135). Во избежание применения сверхфизиологических уровней LPS установили диапазон титрования для концентрации LPS, составляющий приблизительно 80%, 50% и 20% от максимальной активности люциферазы, стимулированной LPS (5, 1 и 0,2 нг соответственно) (данные не представлены). Предварительная инкубация репортерных клеток со 100 мкг/мл Cpn10 в течение 2 часов могла значительно ингибировать активность люциферазы, стимулированную LPS, на 30-50% при этих концентрациях LPS (фиг.1A). Более короткое время предварительной инкубации обеспечивало менее воспроизводимое ингибирование, а время предварительной инкубации свыше 18 часов не обеспечивало ингибирующего действия (данные не показаны).
Ингибирование продукции IL-6 и RANTES, опосредованное Cpn10, в клетках RAW264.7, стимулированных LPS
Для иллюстрации того, что опосредованное Cpn10 ингибирование активации NF-кВ, индуцированной действием LPS, осуществлялось за счет уменьшения секреции провоспалительных медиаторов, исследовали способность Cpn10 ингибировать продукцию провоспалительного цитокина IL-6 и провоспалительного хемокина RANTES, индуцированную LPS. Применяли LPS в дозе 1 нг, что вызывало продукцию приблизительно 50% от максимальной продукции RANTES в данном исследовании (данные не показаны). 2 ч период предварительной инкубации (сходный с используемым выше) был неспособен вызывать толерантность в этих исследованиях независимо от используемых доз LPS (данные не приведены). Cpn10 опосредовал дозозависимое уменьшение секреции, индуцированной LPS, как RANTES (фиг.1B), так и IL-6 (фиг.1С), показывая, что ингибирование активации NF-кВ в данной системе ведет к уменьшению секреции провоспалительного медиатора.
Cpn10-опосредованное ингибирование TNF-α, индуцированное LPS, не зависело от IL-10
Для определения действия Cpn10 в более физиологической клеточной популяции мышам вводили Cpn10 и отделяли их перитонеальные макрофаги и стимулировали LPS in vitro. Обработка Cpn10 значительно уменьшает секрецию TNF-α этими клетками, индуцированную LPS (фиг.2A), иллюстрируя, что Cpn10 опосредует похожие воздействия на макрофаги, обработанные in vivo, наблюдаемые с клетками RAW264.7, обработанными in vitro.
IL-10 является мощным иммунодепрессирующим цитокином, способным ингибировать передачу сигнала TLR4 (Berlato et al., 2002, J Immunol, 168 6404-6411; Suhrbier & Linn, 2003, Trends Immunol, 24 165-168), и в тех случаях, когда клетки селезенки животных, обработанных Cpn10, стимулировали действием LPS, значительное увеличение продукции IL-10 наблюдалось по сравнению с контрольными животными (фиг.2B). Однако опосредованное Cpn10 снижение продукции TNF-α, индуцированной LPS (фиг.2A), не требовало IL-10, поскольку подобные уменьшения секреции TNF-α наблюдали в тех случаях, когда перитонеальные макрофаги от мышей IL-10-/-, обработанных Cpn10, стимулировали LPS in vitro (фиг.1С). Таким образом, сниженная секреция TNF-α и повышенная продукция IL-10, по-видимому, являются независимыми следствиями Cpn10 обработки.
Cpn10-обработка мононуклеарных клеток периферической крови человека (PBMC)
Для определения того, является ли Cpn10 также активным в отношении первичных клеток человека, PBMC от здоровых доноров предварительно обрабатывали Cpn10 или буфером в течение 1 ч, а затем стимулировали 0,04 нг/мл LPS в течение 20 ч. Эту дозу LPS принимали за наиболее низкую дозу, способную достоверно стимулировать значительную секрецию TNF-α (Johnson et al., 2004, J Biol Chem, в печати) и соответствующую дозе у человека, которая вызывает развитие умеренного транзиторного синдрома, сходного с клиническим сепсисом (Granowitz et al., 1993, J Immunol, 151 1637-1645; Lynn et al., 2003, J Infect Dis, 187 631-639). В PBMC от восьми доноров 1 мкг/мл Cpn10 опосредовал в среднем снижение на 23,7%, а 10 мкг/мл - в среднем на 23,3% снижение секреции TNF-α, индуцированной LPS (фиг.3А), что иллюстрирует, что Cpn10 также уменьшает секрецию TNF-α, индуцированную LPLS, из PBMC. Не наблюдалось значительного уменьшения секреции TNF-α при применении 0,1 мкг/мл Cpn10 (данные не показаны). Для иллюстрации того, что индукция толерантности в данной системе не осуществлялась, PBMC предварительно обрабатывали в диапазоне концентраций LPS и 1 ч спустя их стимулировали 0,04 нг/мл LPS в течение 20 ч. Предварительная обработка LPS в течение 1 ч не ингибировала секрецию TNF-α, стимулированную повторной обработкой LPS (фиг.3В), иллюстрируя, что LPS толерантность не является причиной активности Cpn10.
IL-6 представляет собой другой хорошо известный воспалительный цитокин, индуцируемый LPS. Для определения ингибировал ли Cpn10 секрецию IL-6, индуцированную LPS, PBMC от восьми доноров обрабатывали 1-10 мкг/мл Cpn10 или буфером в течение 1 ч, с последующей стимуляцией 0,04 нг/мл LPS в течение 20 ч. Среднее 18,6 и 24,4% уменьшение секреции IL-6, индуцированной LPS, наблюдали для 1 или 10 мкг/мл Cpn10 соответственно (фиг.3С). Значительного уменьшения секреции IL-6 не отмечалось при использовании 0,1 мкг/мл Cpn10 (данные не представлены). Предварительная обработка LPS в течение 1 ч не ингибировала секрецию IL-6, стимулированную вторичной обработкой LPS, еще раз иллюстрируя, что индукции толерантности в данной системе не происходит (фиг.3D).
Обработка Cpn10 ингибировала секрецию TNF-α, индуцированную LPS, in vivo
Модифицированную модель эндотоксемии использовали для определения, способен ли Cpn10, доставленный in vivo, ингибировать секрецию TNF-α in vivo, индуцированную LPS. Мышам линии BALB/c вводили 100 мкг Cpn10 внутривенно за 30 минут до внутривенного введения 10 мкг LPS и взятие крови осуществляли через 1,5 часа. В результате введения Cpn10 уровень TNF-α в сыворотке снижался в среднем на 47,6%, уровень RANTES в сыворотке снижался в среднем на 40,1%, и уровень IL-10 в сыворотке повышался в среднем на 43,3% в нескольких повторных экспериментах (фиг.4А). Ежедневное, на протяжении пяти дней, предварительное введение Cpn10 не могло значительно увеличить уровень ингибирования TNF-α (данные не представлены). Эти данные согласуются с предыдущими экспериментами на культурах тканей и демонстрируют in vivo эффективность Cpn10 в снижении продукции TNF-α и увеличении продукции IL-10 после стимуляции LPS.
Cpn10 ослаблял острые симптомы болезни "трансплантат против хозяина" (GVHD)
Острая GVHD, после аллогенной трансплантации костного мозга (ВМТ), представляет собой опосредованное Т-клетками заболевание, при котором донорские Т-клетки распознают алло-антигены реципиента и дифференцируются в Th1-клетки доминирующего типа. В результате Th1-цитокины Т-клеточного происхождения (главным образом IFN-γ) вызывают высвобождение цитопатических количеств воспалительных цитокинов (например, TNF-α) мононуклеарными клетками донора, в тех случаях, когда они стимулированы LPS, который проникает через поврежденную ионизирующим излучением слизистую оболочку желудочно-кишечного тракта. Эти цитокины и алло-реактивные Т-клетки в дальнейшем способствуют усилению гастроинтестинального повреждения и проникновению LPS. Смертность от GVHD в моделях ВМТ предотвращают в тех случаях, когда в мононуклеарных клетках донора отсутствует TLR4, LPS эффективно блокируется (терапевтическими антагонистами) (Cooke et ah, 2001, J Clin Invest, 107 1581-1589), или сам TNF-α нейтрализован. Изучали возможность применения Cpn10 во время перитрансплантатного периода для облегчения течения GVHD. Введение Cpn10 донорам трансплантата и реципиентам до трансплантации в значительной степени задерживало летальность от GVHD (фиг.4B). Кроме того, тяжесть GVHD, определяемая по клиническим показателям, также быстро уменьшалась после ВМТ (фиг.4B). Несмотря на то, что Cpn10 был способен задерживать GVHD и снижать процент ранней смертности, в конечном итоге животные погибали от GVHD, в связи с неспособностью Cpn10 прекратить секрецию TNF-α или воздействовать на пролиферацию Т-клеток и секрецию IFNγ (данные не показаны). Лечение животных Cpn10 после трансплантации не воздействует в значительной степени на GVHD (данные не представлены).
Обсуждение
Cpn10 опосредовал 23-56% ингибирование секреции TNF-α в зависимости от используемых систем и дозы LPS и Cpn10. Cpn10 повышал секрецию IL-10, индуцированную LPS, приблизительно на 40-200%, в зависимости от системы, но ингибирование секреции TNF-α не зависело от повышения продукции IL-10.
Поскольку LPS, полученный из E. Coli, представляет собой хорошо описанный агонист для TLR4, исследования показывают, что Cpn10 модулирует передачу сигнала TLR4 через NF-KB. Однако, вероятно, что Cpn10 модулирует секрецию цитокинов через другие пути, стимулированные комплексом TLR4 и TLR2.
Ингибирующие эффекты Cpn10 опосредуются чрезвычайно быстро, в интервале от 30 минут (фиг.4A) до 2 часов (фиг.1A). Это подразумевает ингибирование ранних событий в передаче сигнала или активацию быстрых механизмов отрицательной обратной связи, подобных PI3K, а не поздней фазы механизмов обратной связи, в которые вовлечены IRAK или SOCS (Fukao & Koyasu, 2003, Trends Immunol, 24 358-363).
Специфический ингибитор PI3K, вортманнин, не оказывал определяемого воздействия на активность Cpn10 (данные не представлены), что означает, что Cpn10 не влияет на PI3K путь передачи сигнала.
Пример 2
Cpn10 и TLR2
Материалы и Методы
Мышам линии BALB/c подкожно вводили овальбумин (10 мкг) (Sigma), эмульгированный в CFA (Sigma). CFA содержит экстракты клеточных стенок микобактерий, которые предположительно содержат липопептидные агонисты TLR2 (Lim et al., 2003, Int Immunopharmacol, 3 115-118; Tsuji et al., 2000, Infect Immun; 68 6883-6890; Kirschning & Schumann, 2002, Curr Top Microbiol Immunol, 270 121-44). Хорошо известно, что CFA вызывает развитие гранулем (Bergeron et al., 2001, Eur Respir J, 18 357-361; Shah et al., 2001, J Assoc Physicians India, 49 366-368).
Cpn10 (100 мкг) вводили дважды в день в течение 5 дней с двумя дозами предыдущей инъекции CFA. Подкожные гранулемы измеряли в указанное время.
Результаты
Для определения, способен ли Cpn10 воздействовать на активность образования CFA гранулемы, мышам, обработанным Cpn10 и контрольным буферным раствором, инъекционно вводили CFA. Обработка Cpn10 значительно уменьшала размер индукции гранулемы (фиг.5A).
Заключение
Так же как CFA стимулирует TLR2 и CFA индуцирует образование гранулемы, так и уменьшение образование гранулемы, опосредованное введением Cpn10, свидетельствует о том, что Cpn10 также ингибирует передачу сигнала TLR2.
Пример 3
Cpn10 ингибирует активацию NF-кВ агонистом TLR2 PAM 3 CYS-SK 4
PAM3CysSK4 (липопептид) представляет собой хорошо известный агонист TLR2 (Agrawal et al., J Immunol, 2003,171 4984-9) и способен стимулировать HIV LTR (фиг.5B-D), что, как предполагается, активирует такие факторы транскрипции, как NF-кВ (Lee et al., J Immunol. 2002, 168 4012-4017).
Материалы и Методы
PAM3CysSK4 приобретали у EMC Microcollection GmbH и растворяли в качестве разведения матричного раствора 1 мг/мл в воде. Cpn10 и аналитическую систему RAW264-HIV-LTR-LUC были выполнены, как описано ранее для LPS. Вкратце, клетки RAW-LUC высевали 2,5×105 клеток/мл в 24-луночные планшеты и инкубировали в течение ночи при 37ºC. Cpn10 добавляли к клеткам в концентрации 120 мкг/мл и инкубировали в течение 2 ч при 37ºC. Затем добавляли PAM3Cys-SK4 (10 нг/мл, или 2 нг/мл, или 0 нг/мл) в течение 2 ч до проведения люциферазного анализа. LPS в концентрации 1 нг/мл либо 0,2 нг/мл использовали в качестве положительного контроля.
Результаты
Cpn10 ингибировал активацию HIV LTR под действием TLR2 агониста PAM3CysSK4 (фиг.5В). Это ингибирование сохранялось независимо от того, изменялась ли питательная среда перед добавлением Cpn10 (фиг.5C) или перед добавлением PAM3CysSK4 (фиг.5D).
Заключение
Cpn10 способен ингибировать сигналы активации провоспалительного медиатора в макрофагах, стимулированных агонистом TLR2.
Пример 4
Иммунопреципитация Cpn10 Toll-подобным рецептором в присутствии лиганда
Материалы и Методы
Иммунопреципитация
TLR4 был представлен в виде слитой структуры внеклеточного домена (ECD) TLR4 и тяжелой цепи Fc мыши (T4:Fc) в кондиционной среде из секретирующих клеток (10 мл/точку). T4:Fc собирали центрифугированием с использованием 20 мкл упакованных бус белка А (PAS, CL4B, Sigma) и осадок промывали 3х лизирующим буфером перед разделением методом SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия). Белки переносили путем электропереноса на Hybond-C нитроцеллюлозные мембраны. Мембраны блокировали в 5% растворе сухого молока в PBS и 0,1% Tween 20 в течение 30 минут при 37ºC и зондировали в течение дополнительных 30 минут при 37ºC антимышиным антителом, конъюгированным с пероксидазой хрена (Visintin et al., 2003, J. Biol. Chem. 278 48313-48320.)
Для связывания 1 мкг бакуловирусного MD-2 или 10 мкг Cpn10 добавляли к смеси T4:Fc/PAS и инкубировали в течение ночи при 4ºC. TLR2 использовали в качестве контроля. Кроме того, 10 мкг Cpn10 иммунопреципитировали используя 2 мкг анти-Cpn10 поликлонального антитела. В качестве контроля Cpn10 не добавляли к IP (для определения неспецифичных фоновых полос антитела). Как MD-2 (1 мкг), так и Cpn10 (10 мкг) нагружали в дорожку, чтобы показать общее загруженное количество (INPUT).
Результаты
Эксперимент по коиммунопреципитации проводили для определения, взаимодействует ли рекомбинантный Cpn10 непосредственно с рекомбинантными TLR2: или TLR4:Fc слитыми белками. Рекомбинантый MD-2 использовали в качестве контроля (который взаимодействует с TLR4, но не с TLR2).
Фиг.6 (верхняя панель) демонстрирует, что MD-2 коиммунопреципитирует с TLR4 (но не с TLR2). Нижняя панель фиг.6 показывает, что Cpn10 физически не взаимодействует ни с TLR4, ни с TLR2 в отсутствие лиганда.
Пример 5
FRET анализ взаимодействия Cpn10 с Toll-подобным рецептором в присутствии лиганда
Введение
Плазматическая мембрана клеток состоит из латеральных неоднородных структур, пятен и микродоменов. Эти мембранные микродомены или липидные бляшки богаты гликосфинголипидами и холестерином и вовлечены в клеточные процессы, такие как мембранный сортинг и трансдукция сигнала. Исследовали значение образования липидных бляшек в естественном иммунном распознавании бактерий с использованием биохимических и флуоресцентных методов воспроизведения изображений. Было обнаружено, что рецепторные молекулы, вовлеченные в LPS-клеточную активацию, в том числе CD14, Hsp70, Hsp90, хемокиновый рецептор 4 (CXCR4), фактор роста и дифференцировки 5 (GDF5) и TLR4, присутствуют в микродоменах после стимуляции LPS. Целостность липидных бляшек является необходимым для LPS-клеточной активации, поскольку лекарственные средства, разрушающие бляшки, такие как нистатин или MCD, ингибируют секрецию TNF-α, индуцированную LPS. Эти результаты дают возможность предположить, что целостная бактериальная система распознавания лежит в основе связывания CD14 бактериальными компонентами и рекрутинга многочисленных сигнальных молекул, таких как Hsp70, Hsp90, CXCR4, GDF5 и TLR4, в сайте связывания CD14-LPS, в липидных бляшках (Triantafuou et al., 2002, J Cell Sci 115 2603).
Cpn10 дозозависимым образом уменьшает интенсивность сигнала LPS-стимуляции клеток человека и мыши как in vivo, так и in vitro. Таким образом было сделано предположение, что Cpn10 может локализоваться на липидных бляшках во время группировки сигнальных LPS, путем связывания либо непосредственно с TLR4, либо с одним из других участников кластера для прерывания передачи сигнала.
Представленные здесь результаты представляют данные, полученные FRET анализом с использованием Cpn10 и кроличьего поликлонального анти-Cpn10 антитела.
Материалы и Методы
FRET измерения
Флуоресцентный резонансный перенос энергии (FRET) является неинвазивным методом воспроизведения изображения, используемым для определения близости молекул. FRET может происходить на расстоянии свыше 1-10 нм и эффективно увеличивает разрешение оптического микроскопа до молекулярного уровня. Данный метод включает в себя нерадиационный перенос энергии от возбужденного состояния донорских молекул к соответствующему акцептору. Скорость энергии переноса является величиной, обратно пропорциональной показателю расстояния между донором и акцептором в шестой степени.
В этом FRET анализе образцы метили донор- и акцептор-конъюгированными антителами и энергию переноса определяли как увеличение флуоресценции донора (разгорание) после полного фотообесцвечивания молекулы акцептора. FRET изображения рассчитывали по увеличению флуоресценции донора после фотообесцвечивания акцептора. Здесь FRET использовали для определения концентрации рецепторных молекул, вовлеченных в клеточную активизацию, индуцированную LPS (клетки линии MonoMac6), в липидных бляшках.
Результаты
На основании проведенных экспериментов с использованием FRET можно изучить, являлся ли Cpn10 локализованным совместно с липидными бляшками. FRET измеряли в величинах разгорания флуоресценции донора после полного фотообесцвечивания флуорофора акцептора. Увеличение флуоресценции донора после деструкции акцептора указывает на то, что флуоресценция донора была погашена в присутствии акцептора вследствие переноса энергии.
Исследуя эффективность переноса энергии, с использованием положительного контроля, т.е. переноса энергии между моноклональными антителами и различными эпитопами молекул GM1 (ганглиозид, молекула, ассоциированная с бляшками), было показано, что максимальная эффективность энергии переноса (E%) составляла 37±1,0.
Также использовали отрицательный контроль между FITC-GM1 и родамином-MHC, который не выявил значительного переноса энергии (3±0,4). Считается, что это фоновое значение FRET вызвано случайным FRET, поскольку оба типа присутствуют в высоких концентрациях.
Данные, представленные в таблице 1, демонстрируют, что CD14 находится в липидных бляшках, как описано в недавней публикации (Triantafilou et ah, J Cell Science 2002, supra), однако, не было обнаружено, что TLR4 и Cpn10 связаны с липидными бляшками до стимуляции LPS, и они, по-видимому, попадают туда после стимуляции LPS.
Данные в таблице 2 описывают близость Cpn10 к кластеру TLR4, снова с использованием FRET. В таблице 2 положительным контролем был сам TLR4, и давал максимальную эффективность переноса энергии (E%) 36±2,0.
Как показано в таблице 2, Cpn10 не был связан с Hsp70, Hsp90 или CXCR4 в отсутствие LPS, но демонстрировал связь с этими членами кластера LPS-активации после стимуляции LPS. Хотя эти данные, демонстрирующие наличие ассоциации Cpn10 с TLR4 до стимуляции LPS, здесь не показаны, существует устойчивая ассоциация Cpn10 с TLR4 при стимуляции LPS.
Пример 6
Cpn10 и кахексия
Материалы и Методы
Индукция кахексии адъювантом
Целью настоящего исследования было определение, приводит ли введение Cpn10 крысам во время развития адъювантного артрита к снижению потери веса. Самкам крыс Dark Agouti (n=30, 150-160 г) подкожно вводили 0,1 мл CFA в основание хвоста. Адъювант состоял из неполного адъюванта Фрейнда (Difco, Michigan, USA), к которому добавляли 10 мг/мл термоинактивированных Mycobacterium tuberculosis H37RA (Difco). Проявление выявляемого артрита в данной модели происходит в основном на 8-10 день после инъекции CFA.
Обработка Cpn10
Крысам вводили подкожно Cpn10 0,25 мг/кг (n=10), или 2,5 мг/кг (n=10), или разбавитель в качестве контроля (Tris/солевой буфер) (n=10) ежедневно со 2 по 13 день и ежедневно взвешивали.
Статистическая обработка
Тестировали разницу в массе тела у крыс, получающих Cpn10 или разбавитель в качестве контроля, на достоверность, используя одномерный дисперсионный анализ (ANOVA).
Результаты
Наблюдалась полная потеря веса во всех группах после введения CFA, которая оказалась более значительной в контрольной группе (фиг.7).
Когда данные по потере массы тела для групп, получавших 0,25 и 2,5 мг/кг Cpn10, объединили и сравнили с контрольной группой, наблюдалось статистически достоверное различие (p=0,027) в потере массы тела. Объединенные данные от двух групп, получавших лечение, являются обоснованными в этом случае, вследствие сходных значений потери массы тела в двух группах (p=0,94) на протяжении этого периода времени.
Заключение
Адъювантный артрит приводит к изменениям в составе организма и продукции цитокинов, что имитирует провоспалительную кахексию, запускаемую цитокинами, при хроническом воспалительном артрите (Mayer, 1997, Arthritis Rheum, 40 534-539). Введение Cpn10 in vivo либо in vitro уменьшает продукцию провоспалительных цитокинов клетками, стимулированными LPS и другими агонистами.
Действия Cpn10 исследовали в двух дозах у крыс, у которых кахексия была индуцирована экспериментально однократной инъекцией CFA. Разбавляющий контроль и Cpn10 вводили подкожно животным со сходными массой тела и возрастом. В результате введения CFA происходило значительное снижение массы тела. При сравнении групп, получавших Cpn10, и группы, получавшей контроль, наблюдалось статистически достоверное снижение потери массы тела у крыс, получавших Cpn10.
Известно, что повышенные уровни воспалительных цитокинов, в том числе TNF-α, IL-1β и IL-6, коррелируют с кахексией при некоторых заболеваниях, в том числе раке и ревматоидном артрите (Argiles, 2003. Curr Opin Clin Nutr Metab Care, 6 401-406; Walsmith, 2002, Int J Cardiol, 85 89-99). Авторами изобретения было показано, что введение Cpn10 снижает продукцию TNF-α, IL-6 и RANTES и увеличивает продукцию противовоспалительного цитокина IL-10 в мышиных моделях эндотоксемии и болезни "трансплантат против хозяина" и при in vitro LPS стимуляции свежевыделенных РВМС и моноцитарных клеточных линий.
Пример 7
Cpn10 не вызывает толерантность
Материалы и Методы
Биологический анализ RAW264-HIV-LTR-LUC
Клетки линии RAW264-HIV-LTR-LUC культивировали и высеивали для анализа, как описано выше. Клетки инкубировали с LPS, Cpn10 или контрольными буферными растворами в течение 2 ч, с последующим добавлением стимулирующего LPS в указанных концентрациях. После дополнительной 2 ч инкубации адгезивные клетки обрабатывали для люциферазного анализа (Luciferase Assay System, Promega, Madison, WI). Люциферазную активность считывали в течение 15 секунд на люминометре Turner Designs Luminometer TD 20/20.
Получение и очистка Cpn10
Рекомбинантный Cpn10 человека получали и очищали, как описано выше.
Обработка Cpn10 трипсином
2,5% трипсин (Gibco) дважды фильтровали через Acrodisc, как указано выше, и добавляли в концентрации 40 мкг/мл к Cpn10 (2-3 мг/мл). После инкубации при 37ºC в течение ночи раствор трипсин/Cpn10 нагревали до 90ºC в течение 15 минут для дезактивации трипсина перед добавлением к биопробам. После обработки трипсином Cpn10 не обнаруживался методами SDS-PAGE и Вестерн-блоттингом, и вещество находилось в неактивном состоянии при анализе методом рефолдинга роденезы (данные не показаны).
Статистическая обработка
Статистическую обработку (одномерный дисперсионный анализ -ANOVA, t-критерий Стьюдента или логарифмическая ранговая статистика) проводили, используя программу SPSS для Windows 11.5.0 (SPSS Inc.).
Результаты
Индукция толерантности не отвечала за ингибирование передачи сигнала LPS в клетках линии RAW264-HIV-LTR-LUC
Толерантность к LPS представляет собой хорошо известный феномен, при котором снижена реакция на повторную стимуляцию LPS. LPS толерантность обычно индуцируется в том случае, если временной интервал между двумя воздействиями LPS превышает 3 ч и концентрация LPS во время первичного воздействия является достаточно высокой для стимуляции макрофагов (West & Heagy, 2002, Crit Care Med 30 S64-S73; Fujihara et al., 2003, Pharmacol. Ther. 100 171-194).
Для формального учета индукции толерантности, ответственной за результаты наблюдений, приведенных на фиг.1A, клетки RAW264-HIV-LTR-LUC предварительно обрабатывали LPS в диапазоне концентраций в течение 2 ч, с последующей стимуляцией 5, 1 и 0,2 нг/мл LPS. Предварительная обработка концентрациями LPS в диапазоне от 1 до 0,0005 нг/мл не ингибировала индукцию активности LUC за секунду экспозиции LPS (фиг.8A). Таким образом, для системы RAW264-HIV-LTR-LUC двух часовой период предварительной обработки оказался недостаточным для индукции толерантности. Более того, субстимулирующие дозы (0,005-0,0005 нг/мл), очень схожие с теми, которые были обнаружены в препаратах Cpn10, также не способны ингибировать активность LUC, опосредованную LPS. Таким образом, маловероятно, что толерантность к LPS ответственна за результаты, представленные на фиг.1A, поскольку (i) 2 ч интервал между воздействиями LPS был недостаточным для индукции толерантности к LPS в системе RAW-264-HIV-LTR-LUC, и (ii) неопределяемые (или субстимулирующие) уровни LPS, которые возможно контаминируют препараты Cpn10, не способны опосредовать толерантность к LPS.
Cpn10 является чувствительным к гидролизу трипсином
Для иллюстрации того, что ингибирующая активность Cpn10 (фиг.1A) утрачивалось после протеолитического расщепления, Cpn10 подвергали гидролизу трипсином и активность трипсина подавляли нагреванием перед добавлением к биологическим пробам. Деструкцию Cpn10 трипсином подтверждали методами SDS-PAGE/Вестерн-блоттингом и методом рефолдинга роданезы (данные не представлены). (Только нагревание не влияло в значительной степени на активность Cpn10 в анализах LUC или рефолдинга роданезы - данные не представлены). Предварительная обработка Cpn10 снова значительно ингибировала люциферазную активность, опосредованную LPS (фиг.8B, контроль по сравнению с Hsp10, p<0,001, см. пояснение к фигуре). Однако трипсин/термообработанный Cpn10 не ингибировал передачу сигнала LPS (фиг.8B, Трипсин Hsp10), давая значения, сходные со значениями для трипсина/термообработанных контрольных буферных растворов (фиг.8B, Трипсин Буфер). Как ожидалось, обработка трипсином (после нагревания при 90ºC в течение 15 мин) не воздействовала существенным образом на активность LPS E. coli (фиг.8B, Трипсин LPS). Таким образом, протеолитическое разрушение Cpn10 в результате приводит к утрате ингибирующего действия препаратами Cpn10, демонстрируя, что резистентные к трипсину примеси (например, LPS) препаратов Cpn10 не отвечали за ингибирующую активность.
Опосредованное Cpn10 снижение активности LUC, стимулированной LPS, в клетках RAW264-HIV-LTR-LUC зависело от дозы
Процент снижения активности LUC, показанные на фиг.1A, получали с использованием 100 мкг/мл Cpn10. Для определения, зависит ли активность Cpn10 от дозы, клетки RAW264-HIV-LTR-LUC обрабатывали диапазоном концентраций Cpn10 перед добавлением LPS. (Обработка с 100 мкг/мл таким образом представляет собой повтор экспериментов, показанных на фиг.1A). Четкая зависимость доза-ответ наблюдалась с повышением уровней ингибирования наглядно от 2 до 100 мкг Cpn10, с ингибированием, проявляющимся до выравнивания после 100 мкг/мл (фиг.8C).
Пример 8
Действие Cpn10 у людей в клинических испытаниях
Ia фаза клинического испытания
Материалы и Методы
Девятнадцать здоровых добровольцев в возрасте 18-55 лет регистрировали для I фазы 14-дневного испытания Cpn10 для оценки фармакокинетики и безопасности Cpn10, введенного однократной внутривенной инфузией или подкожной инъекцией в двойном слепом плацебо контролируемом исследовании. После скрининга и составления информированного согласия пациенты голодали в течение ночи перед введением Cpn10 в дозах 1, 2,5, 5 или 10 мг, вводимого путем 10 минутной внутривенной инфузии, или 5 мг, вводимого подкожно. Образцы крови (50 мл) для выделения (РВМС) собирали до введения дозы (приблизительно за 12 часов), через 8 часов после введения и на 6 день после введения Cpn10. За пациентами осуществляли мониторинг в течение 14 дней после лечения для оценки побочных эффектов, с взятием крови в интервалах для стандартных гематологических и биохимических исследований и определения появления анти-Cpn10 антител.
Выделение и хранение PBMC
PBMC выделяли из гепаринизированной крови путем центрифугирования в градиенте плотности Ficoll-Hypaque Plus (Amersham), используя протокол производителя. После двух стадий промывания клетки ресуспендировали в замораживающей среде (10% DMSO в эмбриональной сыворотке теленка [FBS]) и замораживали, используя метод ступенчатого замораживания при -70ºC. Клетки транспортировали в сухом льду, а затем хранили в жидком азоте до использования.
Стимуляция PBMC
PBMC размораживали и центрифугировали в FBS, с последующим промыванием и ресуспендированием в RPMI с 10% FBS. Аликвоты клеток с конечной плотностью 1×106 жизнеспособных клеток/мл помещали в 24-луночные планшеты для тканевых культур в присутствии или отсутствие LPS. После 20 часовой инкубации при 37ºC с 5% CO2 собирали супернатанты клеточных культур и исследовали на содержание TNF-α, используя коммерческий TNF-α человека методом ELISA (R & D Systems).
Результаты
Сравнили запускаемый LPS ответ на 1 день по сравнению с 0 днем (например, после введения Cpn10 vs. до введения Cpn10) в группах добровольцев, получавших 1 мг (n=1), 2,5 мг (n=3), 5 мг(n=3) и 10 мг Cpn10 (n=3) или плацебо (n=3). (Примечание: PBMC испытуемых 001 и 004 (группа людей, получавших 1 мг Cpn10) были нежизнеспособны вследствие гемолиза образцов крови. Следовательно, данные по PBMC в данной группе включают только данные от одного участника). Фиг. 9A и 9B демонстрируют дозозависимое изменение, опосредованное Cpn10, продукции TNF-α PBMC, стимулированными диапазоном концентраций LPS in vitro. То есть снижалась запускаемая LPS реакция на 1 день по сравнению с 0 днем у 1 из 3 испытуемых в группе, получавших 2,5 мг, у 3-х из 3 испытуемых в группе, получавших 5 мг, и у 1 из 3 испытуемых в группе, получавших 10 мг. Являющиеся недостаточными из-за небольшого размера групп эти данные, по-видимому, указывают слабую реверсию направления, наблюдаемого в группе 5 мг, при увеличении дозы до 10 мг. У испытуемых, получавших 5 мг Cpn10 путем подкожной инъекции, Cpn10-опосредованное действие на продукцию TNF-α не проявляется (фиг.9C), однако размер группы также слишком мал для предоставления убедительных данных.
Выводы
В качестве предиктора изменений в иммунной активности, опосредованных Cpn10, во время I фазы клинического испытания собирали мононуклеарные клетки периферической крови (РВМС) приблизительно за 12 часов до и 8 часов после однократной внутривенной инфузии или подкожной инъекции Cpn10 (или плацебо). Эти клетки стимулировали диапазоном концентраций LPS in vitro в отсутствие экзогенного Cpn10 для оценки уровня продукции TNF-α. Данные дают возможность предположить, что Cpn10 может воздействовать на снижение провоспалительного ответа при введении в диапазоне доз от 2,5 до 10 мг. Однако численность групп в данном исследовании была очень маленькой, и будут накоплены другие данные для подтверждения гипотезы биологического действия Cpn10 in vivo. Данные по группе, получавшей дозу 10 мг, по-видимому, не согласуются с направлением TNF-α ответа по группам, получавшим 2,5 и 5 мг Cpn10, хотя малочисленность группы не дает возможности проанализировать эти данные помимо рассуждения.
Отсутствие эффекта в этом испытании in vitro в PBMC, выделенных от испытуемых, получавших 5 мг Cpn10 подкожно, наиболее вероятно связано с относительно небольшим количеством белка, который определяется (и, следовательно, является биодоступным) в кровотоке методом ELISA.
Отмечено, что PBMC были выделены в момент времени (8 часов после введения дозы), в который Cpn10 больше не определялся в сыворотке (данные не показаны), подтверждая точку зрения, что хотя tl/2 этого рекомбинантного белка короткое (~1 ч), его биологические эффекты могут быть долгосрочными. Также важно указать, что изменение (т.е. увеличение) LPS ответа на 1 день по сравнению с 0 днем (например, у испытуемых, получавших плацебо) является феноменом, хорошо подтвержденным документально, как у грызунов, так и у человека, и предположительно связан с реакцией на стресс (Granowitz et al., 1993, J Immunol 151 1637). То есть увеличение ответа, запускаемого LPS, PBMC на 1 день по сравнению с 0 днем может быть вследствие стресса во время испытания, т.е. введение непроверенного лекарственного средства, многократное взятие крови и т.д. Cpn10 (член группы стрессовых белков) может принимать участие в снижении обострения воспалительной реакции, связанного со стрессом.
lb фаза клинического испытания
Материалы и Методы
Десять добровольцев в возрасте 18-65 лет с рассеянным склерозом, на данный момент не получающие иммуномодулирующее лечение, были зарегистрированы на 28-и дневное исследование Ib фазы для оценки безопасности, переносимости и фармакокинетики Cpn10, вводимого путем многократных внутривенных инфузий. Данное исследование представляло собой плацебо-контролируемое двойное слепое испытание с подъемом многократных доз. Аналогично фазе Ia испытуемым вводили дозы Cpn10 (или плацебо) 5 или 10 мг путем внутривенной инфузии. Образцы крови для выделения PBMC собирали приблизительно за 12 часов перед введением дозы и через 8 часов после введения дозы на 1 и 5 день. Для оценки длительности биологического действия Cpn10 брали кровь для выделения PBMC на 3 и 7 дни после последнего введения соединения (на 5 день).
РВМС выделяли, хранили и стимулировали, как описано для фазы 1a клинического испытания.
Результаты
Сравнивали ответ, стимулируемый LPS, на 1, 4, 5, 8 и 12 дни по сравнению с 0 днем (т.е. после введения Cpn10 vs. перед введением Cpn10) в группах добровольцев, получавших 5 ежедневных внутривенных инфузий 2,5 (n=4), или 5 мг (n=4) Cpn10, или плацебо (n=2). На фиг.10 показано дозозависимое Cpn10-опосредованное изменение продукции TNF-α PBMC, стимулированными LPS in vitro. Следует отметить, что данные представлены как разница в процентах продукции TNF-α в ответ на стимуляцию LPS на D1 относительно D0, и D4 относительно D0, и т.д. То есть ответ, стимулированный LPS, на D1 относительно D0 был снижен у всех 4 испытуемых в группе, получавших 5 мг Cpn10 на 38-94%. Сравнивая величину TNF-α в D4 и D0 внутри данной группы, процент снижения продукции TNF-α составил 36-72% и 38-59% по сравнению с продукцией TNF-α в D5 и D0. Не было снижения TNF-α, стимулированного LPS, в испытуемой группе 2,5 мг Cpn10 во всех временных точках в ходе исследования «on drug», т.е. на 1, 4, и 5 дни во время испытания.
Заключение
В качестве предиктора изменения иммунной активности, опосредованного Cpn10, во время Ib фазы клинического испытания на добровольцах с рассеянным склерозом, собирали мононуклеарные клетки периферической крови (РВМС) приблизительно за 12 часов до и примерно 8 часов после внутривенной инфузии Cpn10 (или плацебо) на 1, 4 и 5 дни на протяжении пятидневного ежедневного протокола инфузионного введения. Кроме того, PBMC выделяли на 3 и 7 дни, после конечной инфузии Cpn10. Эти клетки стимулировали LPS in vitro в отсутствие экзогенного Cpn10 для определения уровня продукции TNF-α. Эти данные дают возможность предположить, что может быть эффект Cpn10 при уменьшении провоспалительного ответа, при введении 5 мг/день. То есть, у всех четырех исследуемых из Группы B, получавших 5 мг/день Cpn10, наблюдалось значительное уменьшение TNF-α, стимулированного LPS, на 1, 4 и 5 дни. Напротив, у исследуемых, которые получали 2,5 мг/день Cpn10, не было отмечено снижения продукции TNF-α, индуцированной LPS, во время испытания «on drug», т.е. на 1, 4 и 5 день. Также не было снижения продукции TNF-α, стимулированной LPS, в клетках от двух исследуемых, которым инфузионно вводили плацебо (Cpn10 носитель). Однако две группы в данном исследовании были очень малочисленными, и другие данные будут накоплены для подтверждения гипотезы о биологическом действии Cpn10 in vivo.
Как и в Ia фазе испытания, отмечено, что PBMC выделяли в момент времени (8 ч после введения дозы), в который Cpn10 больше не определялся в сыворотке, подтверждая точку зрения, что хотя tl/2 этого рекомбинантного белка короткое (~1 ч), его биологические эффекты могут быть долгосрочными. Однако исходя из этих нескольких экспериментальных результатов невозможно приписать длительному действию Cpn10 на продукцию воспалительных цитокинов больше 12 часов.
Как отмечалось во время Ia фазы исследования, также важно указать, что изменение (т.е. повышение) ответа на LPS на 1 день по сравнению с 0 днем (например, у испытуемых, получавших плацебо) представляет собой хорошо подтвержденный документально феномен как у грызунов, так и у людей, и предполагается, что это связано с реакцией на стресс (Granowitz et al., 1993, supra).
Продукция TNF-α клетками, выделенными на 8 и 12 дни от испытуемых, получавших плацебо, а также и некоторых испытуемых, получавших Cpn10, может быть связана с тем фактом, что исследуемые являлись амбулаторными больными в тот период времени и, следовательно, могли быть менее взволнованными и напряженными. Как ранее предполагалось, Cpn10 (член группы стрессовых белков) может принимать участие в уменьшении обострения воспалительной реакции, связанной со стрессом.
ВЫВОД
Помимо ключевой роли в пространственной упаковке белка внутри митохондрий, оказывается, Cpn10 обладает внеклеточной функцией в модуляции специфических воспалительных процессов. В ряду различных in vitro систем человека и мыши и в двух мышиных моделях заболеваний Cpn10 последовательно ингибировал индуцированную LPS секрецию провоспалительных цитокинов TNF-α и IL-6 и/или провоспалительного хемокина RANTES и повышал LPS индуцированную секрецию противовоспалительного цитокина IL-10. Опосредованное Cpn10 уменьшение секреции TNF-α не было абсолютным; вместо этого Cpn10 опосредовал уменьшение уровней TNF-α приблизительно на 25-60%, в зависимости от системы и дозы LPS и Cpn10. Cpn10 увеличивал секрецию IL-10, индуцированную LPS, приблизительно на 40-200%, в зависимости от системы, но уменьшение секреции TNF-α не зависело от этого увеличения IL-10.
С учетом того, что LPS, полученный из E.coli, является хорошо описанным агонистом рецептора TLR4, описанные здесь эксперименты указывают на то, что Cpn10 может отрицательно модулировать передачу сигнала TLR4. Не ясно, как именно Cpn10 опосредует свою ингибирующую активность, хотя представляется, что ингибирующий эффект очень быстрый, от 30 минут (фиг.4A) до 2 часов (фиг.1A). Что подразумевает ингибирование ранних сигнальных событий или активацию быстрых механизмов отрицательной обратной связи, таких как фосфоинозитид 3-киназа (PI3K). Однако не удалось препятствовать активности Cpn10 специфически ингибитором PI3K, вортманнином, предполагая, что данный каскад реакций не вовлечен в механизм действия Cpn10.
Способность снижать, но не подавлять полностью секрецию TNF-α будет отличать Cpn10 от других противовоспалительных лекарственных средств, в частности основанных на анти-TNF-α антителах, которые могут осуществлять эффективное удаление TNF-α. Однако такое подавление не всегда целесообразно. Например, было показано, что лечение антителами к TNF-α в конечном счете усиливает тяжесть течения рассеянного склероза (MS; Wiendl & Hohlfeld, 2002, BioDrugs 16, 183-200). MS был предложен в качестве возможной терапевтической мишени для Cpn10, поскольку сообщалось, что Cpn10 уменьшает клинические признаки и задерживает начало болезни в мышиной модели MS (экспериментальный аутоиммунный энцефаломиелит; Zhang et al., 2003, J Neurol Sci 212, 37-46).
Как описано здесь, способность Cpn10 уменьшать экспрессию воспалительных медиаторов указывает на то, что Cpn10 может найти терапевтическое применение при состояниях, в которых избыточная передача сигнала LPS, TLR4, и/или TLR2 ведет к патологии.
Кроме того, такой способ действия осуществляется не через индукцию толерантности или селективную активацию/супрессию Th1/Th2 ответов.
Целью данного описания являлось изложение предпочтительных вариантов осуществления изобретения, без ограничения данного изобретения до какого-либо варианта его осуществления или определенной совокупности признаков. Следовательно, специалисту в данной области понятно, что в свете данного описания различные модификации и изменения могут быть сделаны в конкретных вариантах осуществления, представленных в качестве примеров, не отклоняясь от объема настоящего изобретения.
Все компьютерные программы, алгоритмы, патенты и научная литература, упомянутые здесь, включены здесь в качестве ссылок.
Энергию переноса между различными парами определяли по повышению флуоресценции донора после акцепторного фотообесцвечивания. Данные представлены как среднее ± стандартное отклонение в ряду независимых экспериментов.
Энергию переноса между различными парами определяли по повышению флуоресценции донора после акцепторного фотообесцвечивания. Данные представлены как среднее ± стандартное отклонение в ряду независимых экспериментов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ОБНАРУЖЕНИЯ КОНТАМИНАНТОВ ПОЛИМЕРОВ ГЛЮКОЗЫ | 2012 |
|
RU2557995C2 |
| КОМПОЗИЦИЯ ДЛЯ СТИМУЛИРОВАНИЯ СОЗРЕВАНИЯ ДЕНДРИТНЫХ КЛЕТОК, СОДЕРЖАЩАЯ ГИБРИДНЫЙ БЕЛОК Rv2299c/ESAT-6 | 2017 |
|
RU2733886C2 |
| АГЕНТЫ И СПОСОБЫ, ОСНОВАННЫЕ НА ПРИМЕНЕНИИ ДОМЕНА EDA ФИБРОНЕКТИНА | 2006 |
|
RU2430738C2 |
| Применение симбиотического консорциума штаммов пробиотических лактобацилл, выделенных из женского молока, оказывающего бактерицидное действие на мастит-индуцирующие патогены, для изготовления лекарственных средств для комплексной профилактики и лечения лактационного мастита | 2022 |
|
RU2801119C1 |
| БИОЛОГИЧЕСКИЕ МАТЕРИАЛЫ И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2508296C2 |
| ИНДУКЦИЯ АПОПТОЗА В ОПУХОЛЕВЫХ КЛЕТКАХ, ЭКСПРЕССИРУЮЩИХ TOLL-ПОДОБНЫЙ РЕЦЕПТОР | 2005 |
|
RU2401661C9 |
| ПРИМЕНЕНИЕ ТЕНЕЙ БАКТЕРИЙ ДЛЯ ОПОСРЕДОВАННОЙ СТИМУЛЯЦИИ ВРОЖДЕННЛГО ИММУНИТЕТА | 2011 |
|
RU2564110C2 |
| СРЕДСТВО ДЛЯ БЛОКИРОВАНИЯ ЭФФЕКТОВ ЛИПОТЕЙХОЕВЫХ КИСЛОТ - АГОНИСТОВ ТОЛЛ-ПОДОБНОГО РЕЦЕПТОРА 2 | 2022 |
|
RU2781012C1 |
| АНТАГОНИСТЫ ТОЛЛ-ПОДОБНОГО РЕЦЕПТОРА 4 И ПРИМЕНЕНИЕ ПРИ АУТОИММУННЫХ ЗАБОЛЕВАНИЯХ ПЕЧЕНИ | 2016 |
|
RU2707560C2 |
| ИММУНОМОДУЛИРУЮЩИЕ ЭКСТРАКТЫ ИЗ БАКТЕРИЙ Lactobacillus И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2008 |
|
RU2500412C2 |


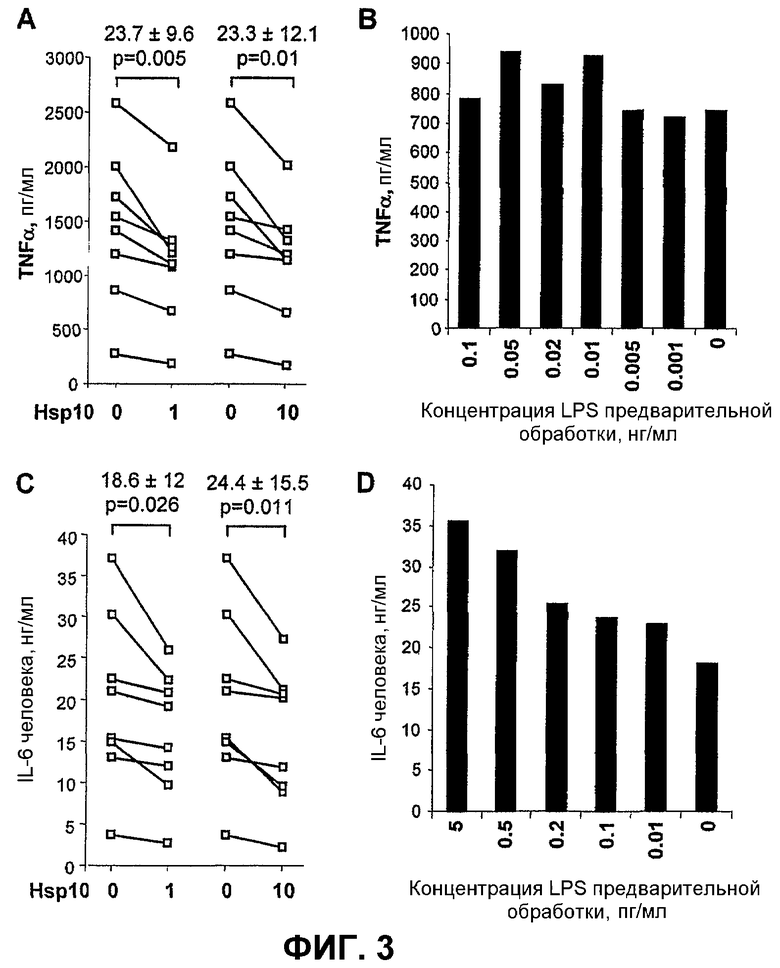








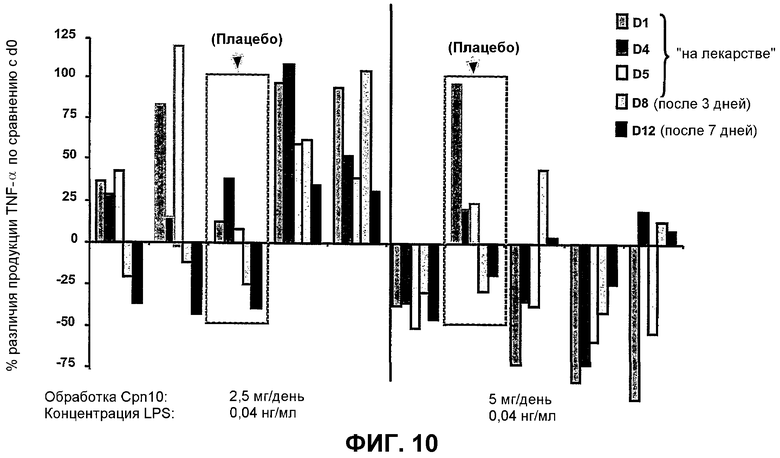
Изобретение относится к области медицины. Предложены способ регуляции передачи сигнала Toll-подобным рецептором (TLR), способ регуляции индуцируемой агонистом TLR2 или TLR4 продукции и/или секреции иммуномодулятора, а также способ лечения заболеваний, чувствительных к регуляции передачи сигнала TLR2 или TLR4 у животного, включающие стадию введения животному, в клетки, ткани или органы шаперонина 10 (Cpn10). Также предложен молекулярный комплекс для обнаружения агониста или антагониста Cpn10, содержащий TLR, агонист TLR и Cpn10, и способ скрининга антагониста Cpn10 с использованием указанного комплекса. Изобретение обеспечивает возможность профилактического или терапевтического лечения заболеваний, нарушений или состояний, связанных с сердцем, чувствительных к регуляции передачи сигнала TLR2 или TLR4. 5 н. и 22 з.п. ф-лы, 10 ил., 2 табл., 8 пр.
1. Способ регуляции передачи сигнала Toll-подобным рецептором у животного или в одной или нескольких клетках, или тканях, или органах, полученных от него, включающий в себя стадию введения Срn10 животному, в клетки, ткани или органы, для регуляции тем самым передачи сигнала, индуцированного агонистом Toll-подобного рецептора, где указанный Toll-подобный рецептор представляет собой Toll-подобный рецептор 2 (TLR2) или Toll-подобный рецептор 4 (TLR4).
2. Способ по п.1, где агонист представляет собой патоген или получен из него.
3. Способ по п.2, где агонист выбран из LPS или липопептида.
4. Способ по п.1, где животное представляет собой млекопитающее.
5. Способ по п.4, где млекопитающим является человек.
6. Способ регуляции индуцируемой агонистом Toll-подобного рецептора продукции и/или секреции иммуномодулятора у животного или в одной или нескольких клетках, или тканях, или органах, полученных от него, включающий стадию введения Срn10 или производного Срn10 животному, в клетки, ткани или органы, для регуляции тем самым продукции и/или секреции иммуномодулятора, индуцируемой агонистом Toll-подобного рецептора, где указанный Toll-подобный рецептор представляет собой Toll-подобный рецептор 2 (TLR2) или Toll-подобный рецептор 4 (TLR4).
7. Способ по п.6, где агонист представляет собой патоген или получен из него.
8. Способ по п.7, где агонист выбран из LPS или липопептида.
9. Способ по п.6, где продукция и/или секреция иммуномодулятора отрицательно регулируется Срn10.
10. Способ по п.9, где иммуномодулятор представляет собой провоспалительный цитокин или хемокин.
11. Способ по п.10, где провоспалительный цитокин представляет собой IL-6 или TNF-α.
12. Способ по п.10, где провоспалительный хемокин представляет собой RANTES.
13. Способ по п.6, где продукция и/или секреция иммуномодулятора положительно регулируется Срn10.
14. Способ по п.13, где иммуномодулятор представляет собой противовоспалительный цитокин или хемокин.
15. Способ по п.14, где противовоспалительный цитокин представляет собой IL-10 или TGF-β.
16. Способ по п.6, где животное представляет собой млекопитающее.
17. Способ по п.16, где млекопитающим является человек.
18. Способ профилактического или терапевтического лечения заболевания, нарушения или состояния, связанных с сердцем, чувствительных к регуляции передачи сигнала Toll-подобным рецептором, у животного, включающий стадию введения Срn10 указанному животному, для регуляции тем самым передачи сигнала, индуцированной агонистом Toll-подобного рецептора, у указанного животного, где указанный Toll-подобный рецептор представляет собой Toll-подобный рецептор 2 (TLR2) или Toll-подобный рецептор 4 (TLR4).
19. Способ по п.18, где агонист представляет собой патоген или получен из него.
20. Способ по п.18, где животное представляет собой млекопитающее.
21. Способ по п.20, где млекопитающим является человек.
22. Выделенный молекулярный комплекс для обнаружения агониста или антагониста Срn10, где комплекс содержит Toll-подобный рецептор, агонист Toll-подобного рецептора и Срn10, где комплекс образован путем приведения агониста Toll-подобного рецептора и Срn10 в контакт с Toll-подобным рецептором, где указанный Toll-подобный рецептор представляет собой Toll-подобный рецептор 2 (TLR2) или Toll-подобный рецептор 4 (TLR4).
23. Выделенный молекулярный комплекс по п.22, где Toll-подобный рецептор представляет собой TLR4, а агонистом является LPS.
24. Выделенный молекулярный комплекс по п.22, где подобный рецептор представляет собой TLR2, а агонистом является липопептид.
25. Способ скрининга антагониста Срn10, включающий стадии образования молекулярного комплекса, включающего Toll-подобный рецептор, агонист Toll-подобного рецептора и Срn10:
a) в присутствии антагониста Срn10-кандидата и
b) в отсутствие антагониста Срn10-кандидата; и обнаружения того, предотвращается или нарушается ли образование молекулярного комплекса присутствием антагониста Срn10-кандидата, где антагониста Срn10-кандидата, в отношении которого обнаружено, что он предотвращает или нарушает образование молекулярного комплекса, идентифицируют как антагониста Срn10, где указанный Toll-подобный рецептор представляет собой Toll-подобный рецептор 2 (TLR2) или Toll-подобный рецептор 4 (TLR4).
26. Способ по п.25, где обнаружение того, предотвращает или нарушает антагонист Срn10 образование молекулярного комплекса, включает обнаружение ингибирования продукции и/или секреции иммуномодулятора, индуцируемого Toll-подобным рецептором.
27. Способ по п.25, где обнаружение того, предотвращает или нарушает антагонист Срn10 образование молекулярного комплекса, включает обнаружение ингибирования передачи сигнала Toll-подобным рецептором.
| BEG AA "Endogenous ligands of Toll-like receptors: implications for regulating inflammatory and immune responses" Trends in immunology, 2002 Nov; 23(11): 509-512 | |||
| ZHANG В et al | |||
| "Early pregnancy factor suppresses experimental autoimmune encephalomyelitis induced in Lewis rats with myelin basic protein and in SJL/J mice with myelin proteolipid |

