ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Данное изобретение относится к антителу к антигену глипикану 3 (т.е. анти-глипикан 3-антителу), в котором цитотоксическая активность, в частности антителозависимая клеточноопосредованная цитотоксичность (ADCC), является усиленной, а также к способу получения этого антитела.
УРОВЕНЬ ТЕХНИКИ
Глипикан 3 (GPC3) является членом семейства гепарансульфат-протеогликанов, присутствующих на поверхности клеток. Было предположено, что GPC3 может участвовать в клеточном делении при развитии и в росте раковых клеток.
Было обнаружено, что тип антитела, который связывается с GPC3, ингибирует рост клеток вследствие активности ADCC (антителозависимой клеточноопосредованной цитотоксичности) и активности CDC (комплементзависимой цитотоксичности) (WO 2003/00883). Кроме того, поскольку GPC3 расщепляется в теле и секретируется в кровь в виде растворимой формы GPC3, было сделано предположение, что рак может диагностироваться с использованием антитела, которое может детектировать растворимую форму GPC3 (WO 2004/022739, WO 2003/100429, WO 2004/018667).
При развитии противоракового агента, который использует цитотоксическую активность антитела, используемое антитело предпочтительно будет иметь высокий уровень активности ADCC. Таким образом, требовалось анти-GPC3-антитело с высоким уровнем цитотоксической активности.
Известно, что модификация сахарных цепей антитела усиливает его ADCC-активность. Например, в WO 99/54342 описано, что ADCC-активность усиливается модификацией гликозилирования антитела. Кроме того, в WO 00/61739 описано, что ADCC-активность регулируется контролированием присутствия или отсутствия фукозы в сахарных цепях антитела. В WO 02/31140 описано получение антитела, имеющего сахарные цепи, которые не содержат α-1,6-фукозы основной структуры (кора) сахарной цепи, продуцированием этого антитела в клетках YB2/0. В WO 02/79255 описано антитело с сахарными цепями, имеющими двурассеченный GlcNAc. Однако анти-GPC3-антитело с повышенной активностью ADCC вследствие модификации сахарной цепи до сих пор не было описано.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение обеспечивает композицию анти-GPC3-антител с повышенной ADCC-активностью, обусловленной изменением его представляющего сахарную цепь компонента, а также способ получения такого антитела.
После многочисленных исследований авторы данного изобретения обнаружили, что антитела против GPC3 с сахарными цепями, лишенными α-1,6-фукозы основной структуры (кора), имеют высокий уровень цитотоксической активности. Таким образом, данное изобретение обеспечивает композицию анти-GPC3-антител, в которых представляющий сахарную цепь компонент антитела был изменен, и, более конкретно, композицию антител с большей фракцией фукоза-недостаточных анти-GPC3-антител. Композиция анти-GPC3-антител с модифицированной сахарной цепью данного изобретения имеет высокий уровень цитотоксической активности и, следовательно, применима в качестве ингибитора роста клеток, такого как противораковый агент.
Данное изобретение обеспечивает также способ получения композиции анти-GPC3-антител, в которых сахарная цепь антитела является модифицированной, предусматривающий стадии: введения нуклеиновой кислоты, кодирующей анти-GPC3-антитело, в клетку-хозяина с уменьшенной способностью присоединения фукозы, такую как клетки YB2/0, и культивирования этой клетки-хозяина для получения этого антитела. Предпочтительно, клеткой с уменьшенной способностью присоединения фукозы к сахарным цепям является клетка, лишенная транспортера фукозы.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фиг. 1 показывает основную структуру N-гликозид-связанных сахарных цепей;
Фиг. 2 показывает активность ADCC химерного антитела, когда мишенями являются клетки НерG2, с использованием моноцитов периферической крови человека (PBMC);
Фиг. 3 показывает активность ADCC химерного антитела, когда мишенью являются клетки HuH-7, с использованием PBMC человека;
Фиг. 4 показывает активность ADCC антител, когда мишенями являются клетки HuH-7, с использованием PBMC человека;
Фиг. 5 показывает хроматограммы ВЖХ с нормальной фазой сахарных цепей, модифицированных α-галактозил-2-АВ, полученных из антител (a, b, c), продуцируемых клетками FT-KO и клетками СНО;
Фиг. 6 показывает предсказанные структуры для пиков G(0)- и G(0)-Fuc, показанных на фиг. 5; и
Фиг. 7 показывает график измерений дифференциальной сканирующей калориметрии (DSC) для антител, продуцируемых клетками FT-KO (а) и клетками СНО (b).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение описывает композицию анти-GPC3-антител, в которых компонент, представляющий сахарную цепь, этого антитела был модифицирован. Известно, что структура сахарной цепи, связанной с антителом, оказывает существенное влияние на проявление цитотоксической активности антитела. Сахарные цепи, которые связаны с антителом, включают в себя N-гликозид-связанные сахарные цепи, которые присоединены к атому азота на боковой цепи остатка аспарагина, и О-гликозид-связанные сахарные цепи, которые присоединены к гидроксильной группе на боковой цепи остатка серина или треонина на молекуле антитела. Данное изобретение сфокусировано на присутствии или отсутствии фукозы в N-гликозид-связанной сахарной цепи.
Фиг. 1 показывает основную структуру N-гликозид-связанных сахарных цепей, присоединенных к антителу. Как показано в основных сахарных цепях IgG (1) и (3) фиг. 1, N-гликозид-связанные сахарные цепи имеют основную структуру (кор), в которой один остаток маннозы (Man) и два остатка N-ацетилглюкозамина (GlcNAc) связаны β-1,4-связями [-Man β1-4GlcNAc β-1-4GlcNAc-]. «GlcNAc» на правой стороне этой структуры называют восстанавливающим концом, а “Man” на левой стороне этой структуры называют невосстанавливающим концом. Когда фукоза связана с восстанавливающим концом, она обычно принимает форму α-связи между положением 6 N-ацетилглюкозамина на восстанавливающем конце и положением 1 фукозы. С другой стороны, в сахарной цепи, показанной в основной сахарной цепи IgG (2) фиг. 1, наряду с вышеупомянутыми двумя сахарными цепями, N-ацетилглюкозамин (GlcNAc) связан с невосстанавливающим концом основной структуры (кора) через β-1,4-связь. Этот тип N-ацетилглюкозамина (GlcNAc) называют «двурассекающим N-ацетилглюкозамином». Сахарной цепью, имеющей двурассекающий N-ацетилглюкозамин, может быть О-гликозид-связанная сахарная цепь или N-гликозид-связанная сахарная цепь, и она образуется переносом N-ацетилглюкозамина к сахарной цепи N-ацетилглюкозаминтрансферазой III (GnTIII). Ген, кодирующий этот фермент, уже был клонирован, и сообщались как аминокислотная последовательность, так и нуклеотидная последовательность ДНК, кодирующей этот фермент (База данных NCBI (ACCESSION D13789)).
В данном изобретении композицией антител с модифицированным или измененным компонентом, представляющим сахарную цепь, (композицией антител с модифицированной сахарной цепью) называют композицию антител, имеющих компонент, представляющий сахарную цепь, которая отличается от композиции антител, продуцируемых клеткой-хозяином, служащей в качестве референсного стандарта.
В данном изобретении можно определить, был или не был представляющий сахарную цепь компонент модифицирован с использованием в качестве референсного стандарта композиции антител, продуцируемых клеткой-хозяином, служащей в качестве референсного стандарта. Если композиция антител имеет представляющий сахарную цепь компонент, отличающийся от композиции антител из референсного стандарта, то эту композицию антител считают композицией антител с модифицированным компонентом, представляющим сахарную цепь.
Клеткой-хозяином, служащей в качестве ссылочного стандарта в данном изобретении, является клетка СНО DG44. Клетка СНО DG44 может быть получена, например, из Invitrogen Corporation.
Примеры композиции антител с модифицированным компонентом, представляющим сахарную цепь, включают в себя, например, композицию антител с увеличенной долей фукоза-недостаточных (например, недостаточных в отношении α-1,6-фукозы кора (основной структуры)) антител в этой композиции антител, и композицию антител с увеличенной долей антител, имеющих присоединенный двурассекающий N-ацетилглюкозамин (GlcNAc) в этой композиции антител.
В предпочтительном варианте осуществления данного изобретения, эта композиция антител имеет более высокую долю фукоза-недостаточных антител, чем композиция антител, используемая в качестве референсного стандарта.
Поскольку некоторые антитела имеют множество N-гликозидных сахарных цепей, фукоза-недостаточное антитело данного изобретения включает в себя не только антитела, в которых фукоза не присоединена, но также антитела, в которых количество фукозных остатков, присоединенных к антителу, является уменьшенным (антитела, имеющие по меньшей мере одну или более сахарных цепей, в которых не присутствует фукоза).
При получении антитела с модифицированной сахарной цепью с использованием клеток-хозяев часто трудно получить композицию, содержащую однородные антитела, в которой все антитела имеют идентичные сахарные цепи. Таким образом, если композиция антител с модифицированным компонентом, представляющим сахарную цепь, является, например, композицией антител с увеличенной долей фукоза-недостаточных антител, то эта композиция антител с модифицированным компонентом, представляющим сахарную цепь, данного изобретения может содержать как антитела, недостаточные в отношении фукозы, так и антитела, не являющиеся недостаточными в отношении фукозы, но общая доля антител, недостаточных в отношении фукозы, будет более высокой, чем в композиции антител, продуцируемых клетками-хозяевами, служащими в качестве референсного стандарта. Данное изобретение особо не ограничивается в отношении конкретной доли фукоза-недостаточных антител в композиции антител с высокой долей фукоза-недостаточных антител данного изобретения, но предпочтительно эта доля равна не менее чем 20%, более предпочтительно, не менее чем 50% и наиболее предпочтительно, не менее чем 90%.
Данное изобретение особо не ограничивается в отношении конкретной доли антител с добавленным двурассекающим N-ацетилглюкозамином в композиции антител, имеющих высокую долю антител с добавленным двурассекающим N-ацетилглюкозамином, но предпочтительно эта доля равна не менее чем 20%, более предпочтительно, не менее чем 50% и наиболее предпочтительно, не менее чем 90%.
Композиция анти-GPC3-антител с модифицированным компонентом, представляющим сахарную цепь, данного изобретения может быть получена способами, известными квалифицированному специалисту в данной области.
Например, фукоза-недостаточное антитело может быть получено экспрессией анти-GPC3-антитела в клетках-хозяевах либо лишенных способности, либо имеющих низкую способность присоединять α-1,6-фукозу кора (основной структуры).
Данное изобретение особо не ограничивается в отношении клеток-хозяев, лишенных способности или имеющих низкую способность присоединять фукозу, но клетки-хозяева без активности фукозотрансферазы или с уменьшенной активностью фукозотрансферазы, клетки-хозяева с более низкой концентрацией фукозы в тельцах Гольджи и т.п. могут быть использованы в данном изобретении. Более конкретно, примеры клеток-хозяев включают в себя клетки миеломы крысы YB2/3HL.P2.G11.16Ag.20 (сокращенно YB2/0) (хранящиеся как АТСС CRL 1662), клетки СНО с нокаутом FTVIII (WO 02/31140), клетки Lec13 (WO 03/035835) и недостаточные в отношении транспортера фукозы клетки (WO 2005/017155).
В данном контексте, термин «недостаточные в отношении транспортера фукозы клетки» относится к клетке, в которой количество транспортера фукозы в клетке меньше, чем в нормальных клетках, или функция транспортера фукозы является ослабленной вследствие отклонения от нормы в структуре транспортера фукозы. Примеры клеток, недостаточных в отношении транспортера фукозы, включают в себя, например, клетки, в которых разрушен ген транспортера фукозы (т.е. клетки с нокаутом этого гена) (далее называемые клетками FT-KO), клетки, в которых либо отсутствует, либо является мутированной часть гена транспортера фукозы, клетки, недостаточные в отношении системы экспрессии гена транспортера фукозы, и т.п. Нуклеотидная последовательность гена, кодирующего транспортер фукозы Китайского хомячка, и его аминокислотная последовательность показаны в SEQ ID NO:126 и 127, соответственно.
Кроме того, можно получить недостаточную в отношении транспортера фукозы клетку данного изобретения с использованием интерференции РНК (RNAi) с применением нуклеотидной последовательности, представленной SEQ ID NO:126. RNAi представляет собой следующий феномен: при введении двухцепочечной РНК (dsRNA) в клетку внутриклеточная мРНК, соответствующая этой последовательности РНК, специфически разрушается и не может экспрессироваться в виде белка. Обычно dsRNA используется с RNAi, но данное изобретение не ограничивается этим, и, например, может быть использована также двухцепочечная РНК, образованная самокомплементарными одноцепочечными молекулами РНК. Что касается районов, образующих двухцепочечную молекулу, эта молекула может быть двухцепочечной во всех районах или может быть одноцепочечной в некоторых районах (например, в одном или обоих концах). Данное изобретение не ограничивается конкретной длиной олиго-РНК, используемого в RNAi. Длина олиго-РНК в данном изобретении может быть равна, например, 5-1000 оснований (или 5-1000 пар оснований в двухцепочечной молекуле), предпочтительно, 10-100 основаниям (или 10-100 парам оснований в двухцепочечной молекуле) и, наиболее предпочтительно, 15-25 основаниям (15-25 парам оснований в двухцепочечной молекуле); однако особенно предпочтительной является длина 19-23 основания (19-23 пар оснований в двухцепочечной молекуле).
Вышеуказанный процесс RNAi использует феномен, в котором dsRNA, состоящая как из смысловой РНК, так и антисмысловой РНК, гомологичных конкретному гену, будет разрушать гомологичную часть транскрипта (мРНК) этого гена. Может быть использована dsRNA, соответствующая всей последовательности гена транспортера фукозы, или более короткая dsRNA (например, 21-23 п.н.), соответствующая части этой последовательности (малая интерферирующая РНК; siRNA). dsRNA может быть непосредственно перенесена в клетку, или может быть получен вектор, продуцирующий dsRNA и перенесен в клетку-хозяина, и затем эта dsRNA может быть продуцирована в этой клетке. Например, вся ДНК или часть ДНК, кодирующей транспортер фукозы, может быть встроена в вектор таким образом, что она образует последовательность обращенного повтора, и этот вектор может быть затем перенесен в клетку-хозяина. Процедура RNAi может проводиться в соответствии с описаниями в следующих ссылках: Fire A. et al., Nature (1998), 391, 806-811; Montgomery M. K. et al., Proc. Natl. Acad. Sci. USA (1998), 95, 15502-15507; Timmons L. et al., Nature (1998), 395, 854; Sanchez A. et al., Proc. Natl. Acad. Sci. USA (1999), 96, 5049-5054; Misquitta L. et al., Proc. Natl. Acad. Sci. USA (1999), 96, 1451-1456; Kennerdell J. R. et al., Cell (1998), 95, 1017-1026; Waterhouse P. M. et al., Proc. Natl. Acad. Sci. USA (1998), 95 13959-13964; and Wianny F. et al., Nature Cell Biol. (2000), 2, 70-75.
Недостаточные в отношении транспортера фукозы клетки, полученные процедурой RNAi, могут быть подвергнуты скринингу, как указано, посредством активности транспортера фукозы. Скрининг может также проводиться на основе транскрипции и экспрессии гена транспортера фукозы с использованием Вестерн-блоттинга или Нозерн-блоттинга.
Антитело с двурассекающим N-ацетилглюкозамином (GlcNAc), добавленным к сахарной цепи, может быть получено экспрессией анти-GPC3-антитела в клетке-хозяине, способной образовывать структуру двурассекающего N-ацетилглюкозамина (GlcNAc) на сахарной цепи.
Способ получения антитела с сахарной цепью, имеющей добавленный N-ацетилглюкозамин, уже известен (WO 02/79255). Клетка-хозяин, способная образовывать структуру двурассекающего N-ацетилглюкозамина (GlcNAc) на сахарной цепи, особо не ограничивается в данном изобретении, но может включать в себя, например, клетку-хозяина, имеющую экспрессирующий вектор, содержащий ДНК, кодирующую GnTIII. Таким образом, анти-GPC3-антитело, имеющее сахарную цепь с добавленным двурассекающим N-ацетилглюкозамином, может быть получено с использованием клетки-хозяина, содержащей как экспрессирующий вектор с ДНК, кодирующей GnIII, так и экспрессирующий вектор, кодирующий анти-GPC3-антитело. Как ДНК, кодирующая GnIII, так и ген, кодирующий анти-GPC3-антитело, могут присутствовать на одном и том же векторе или могут присутствовать на разных векторах.
Другим способом увеличения доли композиции фукоза-недостаточных антител или антител с присоединенным двурассекающим N-ацетилглюкозамином в этой композиции антител является увеличение доли этих антител в композиции очисткой фукоза-недостаточных антител или антител с присоединенным двурассекающим N-ацетилглюкозамином.
Анализ сахарных цепей может проводиться любыми способами, известными квалифицированным в данной области специалистам. Например, сахарная цепь может быть высвобождена из антитела реакцией этого антитела с N-гликозидазой F (Roche) и т.п. Затем эти сахарные цепи могут быть обессолены твердофазной экстракцией с использованием целлюлозного картриджа (Shimizu Y. et al., Carbohydrate Research 332 (2001), 381-388), сконцентрированы и высушены, и помечены флуоресцентно 2-аминопиридином (Kondo A. et al., Agricultural and Biological Chemistry 54:8 (1990), 2169-2170). Реагент удаляют из пиридиламиносахарных цепей (РА-сахарных цепей) твердофазной экстракцией с использованием целлюлозного картриджа, затем сахарные цепи концентрируют центрифугированием с получением очищенных РА-сахарных цепей. Эти сахарные цепи могут анализироваться ВЖХ с обращенной фазой с использованием колонки с октадецилсиланом (ODS). Полученные таким образом РА-сахарные цепи могут анализироваться двухмерным картированием с использованием комбинирования ВЖХ-анализа с обращенной фазой с ODS-колонкой и ВЖХ-анализа с нормальной фазой с амино-колонкой.
Анти-GPC3-антитело с модифицированной сахарной цепью данного изобретения не ограничивается какими-либо конкретными антителами при условии, что оно связывается с GPC3. Предпочтительно, связывание с GPC3 может быть специфическим. Предпочтительные анти-GPC3-антитела данного изобретения включают в себя антитела, которые имеют последовательность определяющего комплементарность района (CDR), показанную в таблице 1 ниже.
Таблица 1
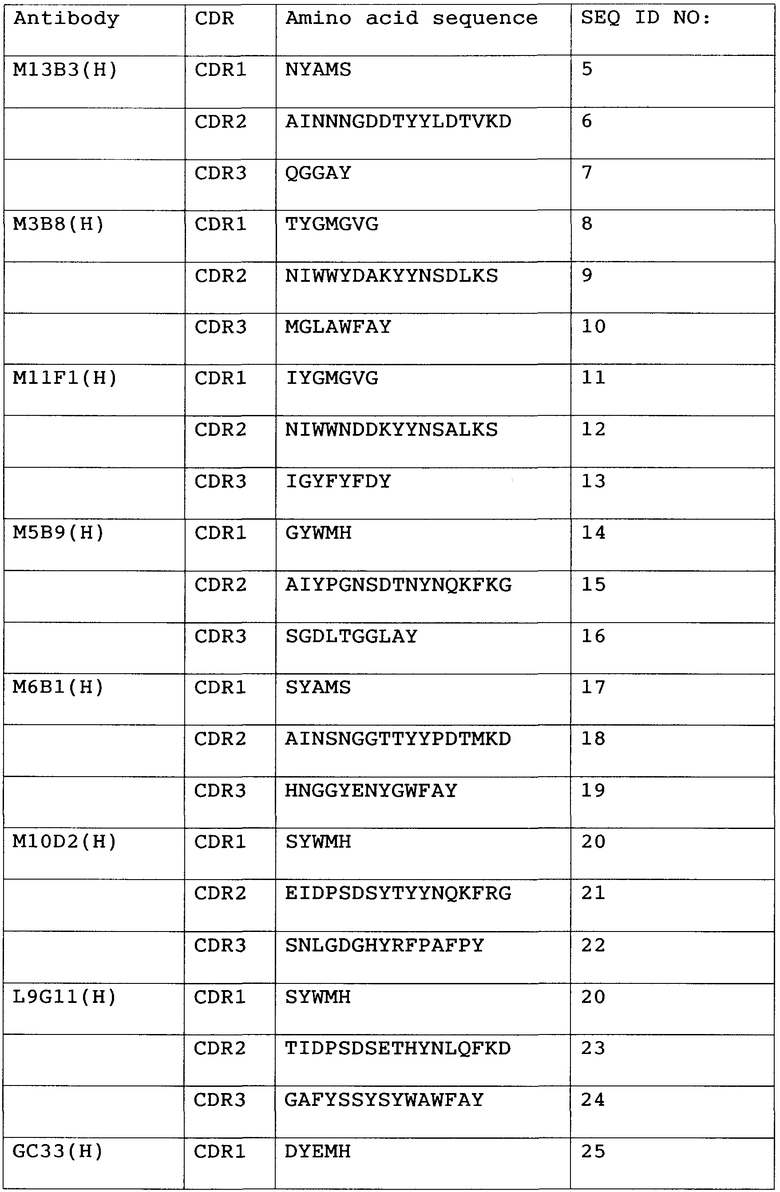
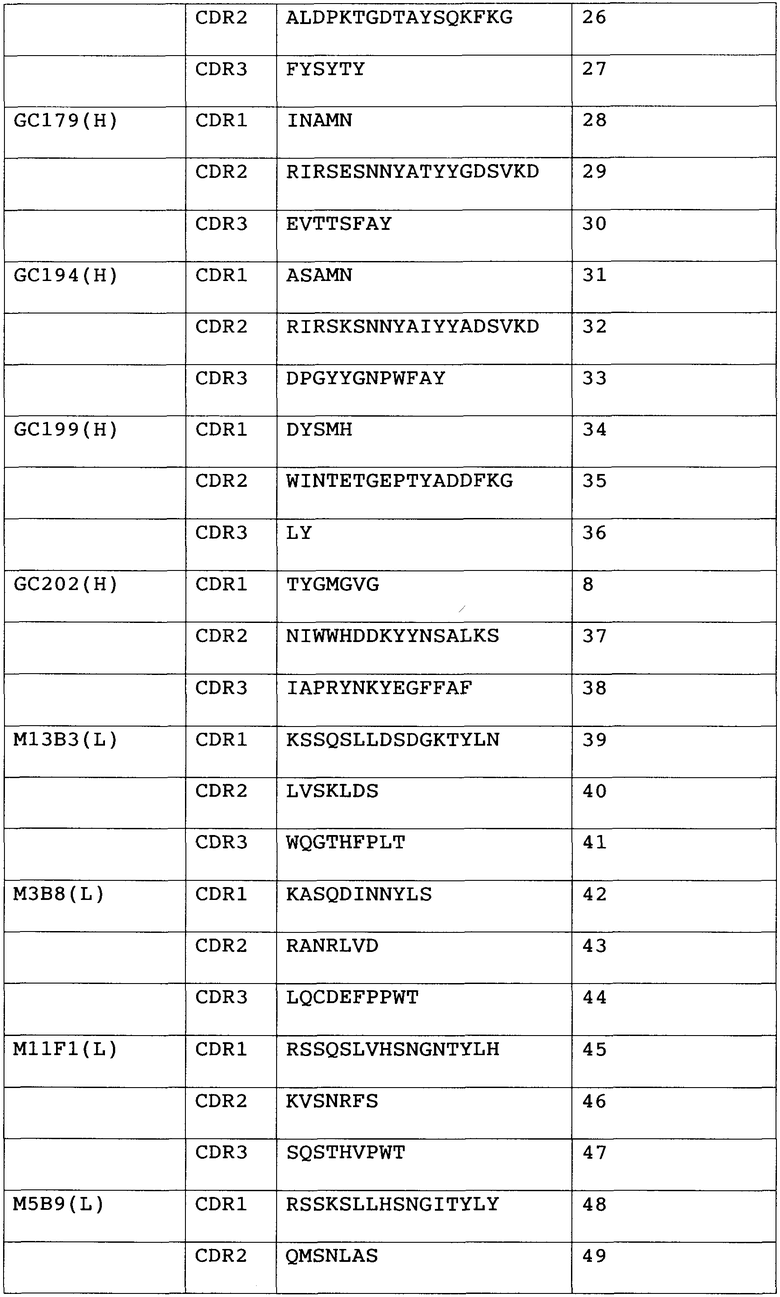
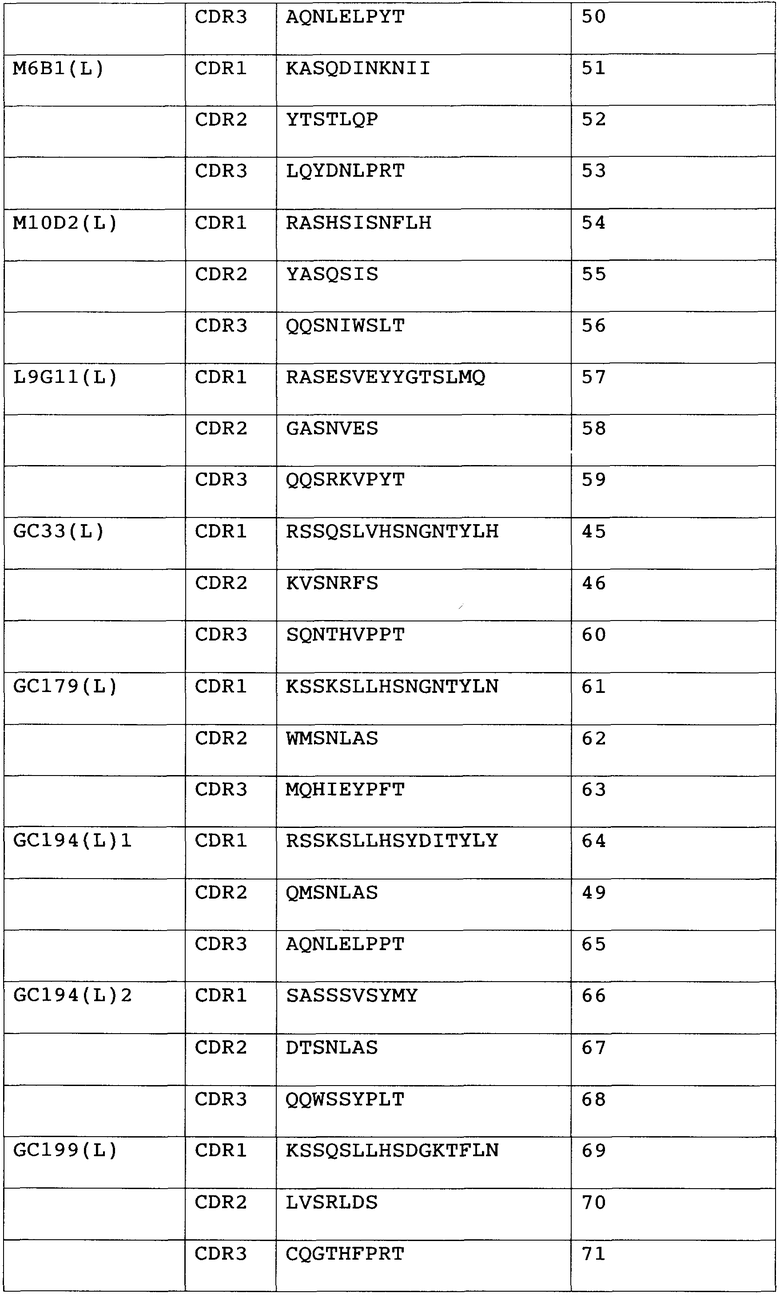

1) Антитело, 2) Аминокислотная последовательность
Антитела с последовательностью CDR из списка, приведенного в таблице выше, имеют высокий уровень цитотоксической активности. Антитела с последовательностью CDR из списка, приведенного в таблице выше, узнают эпитопы аминокислот 524-563 на GPC3. Поскольку антитела, которые узнают эпитопы аминокислот 524-563, имеют высокий уровень цитотоксической активности, они являются предпочтительными в качестве анти-GPC3-антител данного изобретения.
В одном предпочтительном варианте осуществления данного изобретения, композиция антител, имеющих модифицированный компонент, представляющий сахарную цепь, данного изобретения, характеризуется проявлением повышенной активности ADCC. В данном изобретении присутствие или отсутствие повышенной активности ADCC можно определить сравнением активности ADCC композиции антител данного изобретения с активностью ADCC композиции антител референсного стандарта. Если композиция антител данного изобретения обнаруживает более высокую активность ADCC, чем референсный стандарт, то считают, что активность ADCC является повышенной.
Активность ADCC может быть измерена способом, известным квалифицированным в данной области специалистам, например, смешиванием анти-GPC3-антитела с эффекторными клетками и клетками-мишенями и затем определением уровня ADCC. Более конкретно, клетки селезенки мыши, моноциты человека, выделенные из периферической крови (PBMC) и костного мозга и т.п., могут быть использованы в качестве эффекторных клеток, а клетки человека, экспрессирующие GPC3, такие как линия клеток HuH-7 печеночно-клеточного рака человека, могут быть использованы в качестве клеток-мишеней. Сначала эти клетки-мишени метят 51Сr, добавляют анти-GPC3-антитело, клетки инкубируют и затем добавляют эффекторные клетки в подходящем соотношении с клетками-мишенями и их инкубируют вместе. После инкубации супернатант собирают и активность ADCC измеряют счетом радиоактивности в этом супернатанте.
Анти-GPC3-антитело
Анти-GPC3-антитело может быть получено способом, известным квалифицированным в данной области специалистам. Например, это антитело может быть получено с использованием GPC3 в качестве сенсибилизирующего антигена для иммунизации в соответствии с обычным способом иммунизации, слиянием этих иммунных клеток с известными исходными клетками при помощи обычной процедуры слияния клеток и скринингом на продуцирующее моноклональное антитело клеток при помощи обычного способа скрининга. Более конкретно, моноклональные антитела могут быть получены следующим образом. Сначала GPC3 для использования в качестве сенсибилизирующего антигена получают экспрессией GPC3 (MXR7) на основе последовательности гена/аминокислотной последовательности, описанных в Lage, H. et al., Gene 188 (1997), 151-156. Другими словами, последовательность гена, кодирующую GPC3, встраивают в известный экспрессирующий вектор. После трансформации подходящих клеток-хозяев этим вектором белок-мишень глипикан 3 человека очищают из клеток-хозяев или супернатанта культуральной среды известным способом. Затем очищенный белок GPC3 используют в качестве сенсибилизирующего антигена. Альтернативно, в качестве сенсибилизирующего антигена может быть использован частичный пептид GPC3. В таком способе этот частичный пептид может быть получен химическим синтезом в соответствии с аминокислотной последовательностью GPC3 человека. Эпитопы на молекуле GPC3, узнаваемые анти-GPC3-антителом данного изобретения, не ограничиваются, и анти-GPC3-антитело данного изобретения может узнавать любой эпитоп, присутствующий на молекуле GPC3. Это объясняется тем, что анти-GPC3-антитело проявляет ингибирующую рост клеток активность через его ADCC-активность, CDC-активность или ингибирование активности фактора роста, и тем, что рост клеток также ингибируется действием цитотоксического вещества, такого как радиоактивный изотоп, химиотерапевтическое лекарственное средство, бактериальный токсин, присоединенные к анти-GPC3-антителу. Таким образом, антигеном для получения анти-GPC3-антитела данного изобретения может быть любой фрагмент GPC3 при условии, что он содержит эпитоп, присутствующий на молекуле GPC3.
В особенно предпочтительном варианте осуществления, в качестве сенсибилизирующего антигена для генерирования антитела, которое узнает эпитоп из аминокислот 524-563 GPC3, может быть использован пептид, содержащий аминокислоты 524-563.
Млекопитающее, используемое для иммунизации сенсибилизирующим антигеном, особо не ограничивается в данном изобретении, но предпочтительно оно должно быть выбрано с учетом совместимости с исходными клетками, которые должны быть использованы в слиянии клеток, и может включать в себя грызуна, например мышь, крысу или хомяка, или кролика, обезьяну и т.п. Это животное может быть иммунизировано сенсибилизирующим антигеном с использованием известного способа. Обычно, например, млекопитающее может быть инъецировано внутрибрюшинно или подкожно сенсибилизирующим антигеном. Более конкретно, сенсибилизирующий антиген может быть разбавлен и суспендирован в подходящем количестве забуференного фосфатом солевого раствора (ЗФР) или физиологического солевого раствора, смешан с подходящим количеством общепринятого адъюванта, такого как полный адъювант Фрейнда, если желательно, эмульгирован и введен млекопитающему многократно каждые 4 дня - 21 день. Кроме того, при иммунизации сенсибилизирующим антигеном может быть использован подходящий носитель.
После иммунизации млекопитающего описанным выше образом и детектирования уровня желаемого антитела в сыворотке иммунные клетки собирают из этого млекопитающего и подвергают клеточному слиянию. Клетки селезенки являются особенно предпочтительными иммунными клетками для слияния клеток. Клетки миеломы млекопитающих используют в качестве исходных клеток-партнеров для слияния с иммунными клетками. Известные клеточные линии, подходящие для использования в качестве клеток миеломы, включают в себя P3 (P3x63Ag8.653) (J. Immnol. (1979) 123, 1548-1550), P3x63Ag8U.l (Current Topics in Microbiology and Immunology (1978) 81, 1-7), NS-1 (Kohler. G. and Milstein, C. Eur. J. Immunol. (1976) 6, 511-519), MPC-11 (Margulies. D.H. et al., Cell (1976) 8, 405-415), SP2/0 (Shulman, M. et al., Nature (1978) 276, 269-270), FO (de St. Groth, S. F. et al., J. Immunol. Methods (1980) 35, 1-21), S194 (Trowbridge, I. S. J. Exp. Med. (1978) 148, 313-323), R210 (Galfre, G. et al., Nature (1979) 277, 131-133).
Слияние иммунных клеток и клеток миеломы в основном может проводиться в соответствии с известным способом, например способом, описанным Kohler and Milstein (Kohler, G. And Milstein, C., Methods Enzymol. (1981) 73, 3-46). Более конкретно, слияние клеток может проводиться, например, в общепринятой жидкой культуральной среде, содержащей стимулятор слияния клеток. Примеры стимулирующих слияние клеток химических веществ включают в себя полиэтиленгликоль (PEG) и вирус Сендай (HVJ) и т.п. Если желательно, для увеличения эффективности слияния может быть добавлен дополнительный агент, такой как диметилсульфоксид и т.п. Отношение иммунных клеток к клеткам миеломы может быть установлено произвольно. Например, предпочтительной является установка отношения иммунных клеток к клеткам миеломы 1:1-10:1. Общепринятая жидкая культуральная среда, используемая для культивирования этих типов клеток, такая как жидкая среда RPMI-1640, жидкая среда МЕМ или другая жидкая среда, подходящая для роста линии клеток миеломы, может быть использована в качестве жидкой среды в процедуре слияния клеток. Вместе со средой может быть также использована сывороточная добавка, такая как фетальная телячья сыворотка. В процедуре слияния клеток указанные количества иммунных клеток и миеломных клеток тщательно смешивают в жидкой культуральной среде и затем раствор PEG (например, со средней молекулярной массой приблизительно 1000-6000), который был нагрет до 37ºС, добавляют обычно при концентрации 30-60% (масса/объем) и перемешивают для создания возможности образования слитых клеток (гибридом). Затем добавляют подходящую жидкую среду и центрифугируют для удаления супернатанта. Посредством повторения этой процедуры удаляют любые химикалии, использованные для клеточного слияния, неблагоприятные для роста гибридом. Полученные таким образом гибридомы отбирают культивированием их в общепринятой жидкой селективной среде, такой как среда НАТ (культуральная среда, содержащая гипоксантин, аминоптерин и тимидин). Культивирование в среде НАТ продолжают в течение достаточного периода времени (обычно в течение нескольких дней - нескольких недель), пока клетки, другие, чем гибридомы-мишени (неслитые клетки), не погибнут. Затем, проводят общепринятую процедуру лимитирующего разведения с последующим скринингом и моноклонированием гибридом, которые продуцируют антитело-мишень. Наряду с иммунизацией животного, не являющегося человеком, антигеном для получения гибридом, как описано выше, желаемые антитела человека, имеющие GPC3-связывающую активность, могут быть получены сенсибилизацией лимфоцитов человека GPC3 in vitro и затем слиянием этих сенсибилизированных лимфоцитов с иммортализованными клетками миеломы человека (см. Публикацию патента Японии с номером Н1-59878). Кроме того, можно ввести GPC3 в качестве антигена трансгенному животному, имеющему полный репертуар генов антител человека, для генерирования клеток, продуцирующих анти-GPC3-антитела, и собрать антитела к GPC3 человека из иммортализованных клеток (см. Международные заявки на патент с номерами WO 94/25585, WO 93/12227, WO 92/03918 и WO 94/02602). Гибридомы, продуцирующие моноклональные антитела, полученные вышеуказанным образом, могут быть субкультивированы в общепринятой жидкой культуральной среде и могут храниться в течение продолжительного периода времени в жидком азоте.
Рекомбинантные антитела
Моноклональное антитело, использованное в данном изобретении, является рекомбинантным моноклональным антителом, которое может быть получено клонированием гена этого антитела из гибридомы, встраиванием этого гена в подходящий вектор и интеграцией этого вектора в клетку-хозяина (например, см. Vandamme, A.M. et al., Eur. J. Biochem. (1990) 192: 767-775, 1990). Более конкретно, мРНК, кодирующую вариабельную (V) область анти-GPC3-антитела, выделяют из гибридомы, продуцирующей анти-GPC3-антитело. мРНК может быть выделена с использованием известного способа, такого как способ центрифугирования с гуанидином (Chirgwin, J.M. et al., Biochemistry (1979) 18, 5294-5299) и AGPC-способ (Chomczynski, P., et al., Anal. Biochem. (1987) 162, 156-159), для получения тотальной РНК и затем получения мРНК с использованием набора для очистки мРНК (Pharmacia) и т.п. мРНК может быть также получена непосредственно с использованием набора для получения мРНК QuickPrep (Pharmacia). Затем синтезируют кДНК вариабельной (V) области этого антитела из полученной таким образом мРНК с использованием обратной транскриптазы. кДНК может быть синтезирована с использованием набора для синтеза кДНК первой цепи обратной транскриптазой AMV (Seikagaku Corporation), и т.п. Может быть также использован 5'-RACE-способ с использованием набора 5'-AmpliFINDER RACE Kit (Clontech) и ПЦР для синтеза и амплификации кДНК (Frohman, M.A. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 8998-9002, Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-2932). ДНК-фрагмент-мишень очищают из продукта ПЦР и лигируют с ДНК-вектором. Желаемый рекомбинантный вектор готовят встраиванием этих векторов. Его вводят в E. coli и желаемую колонию отбирают для приготовления желаемого рекомбинантного вектора. Нуклеотидную последовательность ДНК-мишени подтверждают известным способом, таким как способ терминации цепи с использованием дидезоксинуклеотида. После получения ДНК, кодирующей вариабельную (V) область анти-GPC3-антитела-мишени, ее встраивают в экспрессирующий вектор, содержащий ДНК, кодирующую константную область желаемого антитела (С-область). Для получения анти-GPC3-антитела, используемого в данном изобретении, ген этого антитела встраивают в этот экспрессирующий вектор таким образом, что он будет экспрессироваться под контролем района регуляции экспрессии, такого как энхансер, промотор и т.п. Затем это антитело экспрессируют трансформацией клетки-хозяина этим экспрессирующим вектором. Ген этого антитела может быть экспрессирован в клетках-хозяевах встраиванием ДНК, кодирующей тяжелую цепь (Н-цепь) и легкую цепь (L-цепь) антитела, в отдельные экспрессирующие векторы и одновременной трансформацией клеток-хозяев или встраиванием ДНК, кодирующей как Н-цепь, так и L-цепь, в единый экспрессирующий вектор и трансформацией клеток-хозяев (см. WO 94/11523). Кроме того, рекомбинантное антитело может быть получено не только с использованием вышеупомянутых клеток-хозяев, но также с использованием трансгенного животного. Например, ген антитела может быть встроен в середину гена, кодирующего белок, продуцируемый специфически в молоке (такой как β-казеин козы), с получением слитого гена. Затем ДНК-фрагмент, содержащий ген этого антитела, инъецируют в зародыш козы и этот зародыш имплантируют в козу. Желаемое антитело может быть получено из молока, продуцируемого трансгенной козой, рожденной козой, имплантированной этим зародышем, и ее потомством. Кроме того, могут быть использованы подходящие гормоны в трансгенной козе для увеличения количества молока, содержащего желаемое антитело, которой продуцируется этой трансгенной козой (Ebert, K.M. et al., Bio/Technology (1994) 12, 699-702).
Измененные антитела
Кроме описанных выше антител, в данном изобретении может быть использовано искусственно измененное рекомбинантное антитело, например химерное антитело, гуманизированное антитело и т.п., с целью уменьшения ксеноантигенности относительно людей. Такие модифицированные антитела могут быть получены в соответствии с известными способами. Химерное антитело может быть получено лигированием ДНК, кодирующей V-область антитела, полученной, как описано выше, с ДНК, кодирующей С-область антитела человека, и затем встраиванием этих ДНК в экспрессирующий вектор. Вектор, в котором встроены эти ДНК, интегрируют в клетки-хозяева для получения этого антитела. Химерное антитело, применимое в данном изобретении, может быть получено с использованием такого общепринятого способа. Гуманизированное антитело, также называемое реконструированным антителом человека, содержит CDR антитела из млекопитающего не человека, такого как мышь, трансплантированный на CDR антитела человека. Общие способы генной инженерии для получения гуманизированных антител известны в данной области (см. ЕР 125023 и WO 96/02576). Более конкретно, ДНК-последовательность, предназначенную для связывания CDR мышиного антитела и каркасной области (FR) антитела человека, синтезируют при помощи ПЦР с использованием в качестве праймеров множества олигонуклеотидов, приготовленных таким образом, что они перекрывают концевые районы CDR и FR (этот способ описан в WO 98/13388). Каркасную область антитела, которая должна быть связана через CDR, выбирают таким образом, что этот CDR будет образовывать подходящий сайт связывания антитела. Если необходимо, аминокислоты этой каркасной области в вариабельном районе антитела могут быть заменены таким образом, что этот реконструированный CDR антитела человека будет образовывать подходящий сайт связывания антитела (Sato, K. et al., Cancer res. (1993) 53, 851-856). Для С-области химерного антитела и гуманизированного антитела используют С-область антитела человека. Например, Cy1, Cy2, Cy3 и Cy4 могут быть использованы в Н-цепи, а Сκ и Сλ могут быть использованы в L-цепи. Кроме того, С-область антитела человека может быть модифицирована для улучшения стабильности или производительности этого антитела. Химерное антитело содержит вариабельную область антитела из млекопитающего не человека и константную область из антитела человека. С другой стороны, гуманизированное антитело содержит CDR антитела из млекопитающего не человека и каркасную область и С-область из антитела человека. Поскольку гуманизированное антитело имеет более низкую антигенность в теле человека, оно является более применимым в качестве активного ингредиента в терапевтическом агенте данного изобретения.
Модифицированные антитела
Антитело, используемое в данном изобретении, не ограничивается целой молекулой антитела и может быть фрагментом антитела или модифицированной формой антитела, при условии, что оно связывается с GPC3 и ингибирует активность GPC3. Данное изобретение включает в себя также бивалентные антитела, а также моновалентные антитела. Примеры фрагмента антитела включают в себя Fab, F(ab')2, Fv, Fab/с, имеющий один Fab и полный Fc, или одноцепочечный Fv (scFv), где Fv Н-цепи или L-цепи связан подходящим лигандом. Более конкретно, для получения фрагмента антитела это антитело может быть обработано ферментом, таким как папаин или пепсин, или ген, кодирующий такой фрагмент антитела, может быть сконструирован, встроен в экспрессирующий вектор и экспрессирован в подходящей клетке-хозяине (см., например, Co, M.S. et al., J. Immunol. (1994) 152, 2968-2976, Better, M. & Horwitz, A. H. Methods in Enzymology (1989) 178, 476-496, Academic Press, Inc., Plueckthun, A. & Skerra, A. Methods in Enzymology (1989) 178, 476-496, Academic Press, Inc., Lamoyi, E., Methods in Enzymology (1989) 121, 652-663, Rousseaux, J. et al., Methods in Enzymology (1989) 121, 663-669, Bird, R. E. et al., TIBTECH (1991) 9, 132-137). scFv может быть получен соединением V-области Н-цепи и V-области L-цепи антитела. В scFv V-область Н-цепи и V-область L-цепи соединяют посредством линкера, предпочтительно пептидного линкера (Huston, J.S. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 5879-5883). V-область Н-цепи и V-область L-цепи в scFv могут происходить из любых описанных здесь антител. Любой одноцепочечный пептид, содержащий 12-19 аминокислотных остатков, может быть использован в качестве пептидного линкера, соединяющего эти V-области. ДНК, кодирующая scFv, может быть получена амплификацией фрагмента при помощи ПЦР с использованием в качестве матрицы части ДНК, кодирующей всю или желаемую аминокислотную последовательность, из последовательностей ДНК, кодирующей Н-цепь или V-область Н-цепи вышеупомянутого антитела, и ДНК, кодирующей L-цепь или V-область L-цепи вышеупомянутого антитела, с парой праймеров, которая определяет оба его конца. Затем этот фрагмент амплифицируют с комбинацией ДНК, кодирующей пептидную линкерную часть, и парой праймеров, которая определяет оба конца, которые должны быть лигированы с Н-цепью и L-цепью. После получения ДНК, кодирующей scFv, могут быть получены в соответствии со стандартным способом экспрессирующий вектор, содержащий эту ДНК, и клетка-хозяин, трансформированная этим экспрессирующим вектором. scFv может быть получен из такого хозяина в соответствии со стандартным способом. Эти фрагменты антител могут быть продуцированы в хозяине посредством получения их гена таким же образом, как описано выше, и его экспрессией. В данном изобретении термин «антитело» включает в себя также фрагмент этого антитела. Анти-GPC3-антитело, присоединенное к различным молекулам, таким как PEG и т.п., может быть использовано в качестве модифицированного антитела. В данном изобретении термин «антитело» включает в себя также такое модифицированное антитело. Модифицированное антитело может быть получено химической модификацией антитела, полученного, как описано выше. Способы модификации антител уже созданы в данной области.
Кроме того, антитело, используемое в данном изобретении, может быть биспецифическим антителом. Биспецифическое антитело может быть антителом, имеющим антигенсвязывающий сайт, который узнает другой эпитоп на молекуле GPC3, или оно может быть антителом, в котором один антигенсвязывающий сайт узнает GPC3, а другой антигенсвязывающий сайт связывания узнает цитотоксическое вещество, такое как химиотерапевтическое лекарственное средство и происходящий из клетки токсин. В таком случае это цитотоксическое вещество будет действовать непосредственно на клетки, экспрессирующие GPC3, и, конкретно, на опухолевые клетки повреждения, и подавлять рост опухолевых клеток. Биспецифическое антитело может быть получено связыванием двух типов HL-пар антител. Оно может быть также получено получением слитой клетки, продуцирующей биспецифическое антитело, посредством слияния гибридом, которые продуцируют различные моноклональные антитела. Биспецифическое антитело может быть также получено способами генной инженерии.
Экспрессия и получение рекомбинантных антител или модифицированных антител
Ген антитела, сконструированный, как отмечалось выше, может быть экспрессирован или получен известными способами. В случае клеток млекопитающих, обычный применимый промотор, ген, который должен быть экспрессирован, и поли-А-сигнальная последовательность справа от 3'-конца могут быть функционально связаны вместе и экспрессированы. Например, в качестве промотора/энхансера могут быть использованы немедленно ранние промотор/энхансер цитомегаловируса человека. Кроме того, другие промоторы/энхансеры, которые могут быть использованы для экспрессии антитела данного изобретения, включают в себя вирусные промоторы/энхансеры ретровируса, полиомавируса, аденовируса и вируса 40 обезьяны (SV40) или промоторы/энхансеры из клеток млекопитающих, такие как фактор элонгации 1а человека (HEF-1a). Антитела могут быть легко экспрессированы по способу Mulligan et al. (Nature (1979) 277, 108), когда используется промотор/энхансер SV40, и по способу Mizuschima et al., (Nucleic Acids Res. (1990) 18, 5322), когда используется промотор/энхансер HTF1α.
Антитело данного изобретения может быть получено с использованием эукариотической системы экспрессии, способной присоединять сахарную цепь к экспрессируемому антителу. Эукариотические клетки включают в себя, например, установленные клеточные линии млекопитающих и клеточные линии насекомых, клетки животных, клетки грибов и клетки дрожжей.
Предпочтительно, антитело данного изобретения экспрессируют в клетках млекопитающих, например в клетках СНО, COS, клетках миеломы, ВНК, Vero или HeLa. Желаемое антитело получают культивированием трансформированных клеток-хозяев in vitro или in vivo. Клетки-хозяева могут культивироваться с использованием известных способов. Например, в качестве культуральной среды могут быть использованы DMEM, МЕМ, RPMI-1640 или IMDM, и может быть добавлен сывороточный комплемент, например фетальная телячья сыворотка (ФТС).
Выделение и очистка антитела
Антитело, экспрессированное и полученное описанным выше образом, может быть выделено из клеток-хозяев или животных-хозяев и очищено до гомогенности. Антитело данного изобретения может быть выделено и очищено с использованием аффинной колонки, например белок А-колонки, такой как Hyper D, POROS и Sepharose F.F. (Pharmacia). Кроме того, в данном изобретении могут быть использованы любые общепринятые способы. Например, антитело может быть выделено и очищено подходящим образом посредством выбора и комбинирования аффинных колонок, таких как Белок А-колонка, с хроматографическими колонками, процедурами фильтрации, ультрафильтрации, высаливания и диализа (Antibodies: A Laboratory Manual, Ed. Harlow, David Lane, Cold Spring Harbor Laboratory, 1988). Антитело, имеющее желаемую сахарную цепь, может быть выделено с использованием колонки с лектином способом, известным в данной области, и этот способ описан в WO 02/30954.
Определение активности антител
Антигенсвязывающая активность (Antibodies: A Laboratory Manual, Ed. Harlow, David Lane, Cold Spring Harbor Laboratory, 1988) и ингибирование связывания лиганд-рецептор (Harada, A. et al., International Immunology (1993) 5, 681-690) антигена, используемого в данном изобретении, могут быть измерены известными способами. ELISA (твердофазный иммуноферментный анализ), EIA (ферментный иммуноанализ), RIA (радиоиммуноанализ) или способ флуресцентного антитела могут быть использованы для измерения антигенсвязывающей активности анти-GPC3-антитела данного изобретения. Например, EIA проводят следующим образом. Пробу, содержащую анти-GPC3-антитело, такую как культуральный супернатант продуцирующих анти-GPC3-антитело клеток или очищенное антитело, добавляют в чашку, покрытую GPC3. Добавляют вторичное антитело, меченное ферментом, таким как щелочная фосфатаза, эту чашку инкубируют и промывают и затем добавляют субстрат фермента, такой как п-нитрофенилфосфат, и определяют оптическую плотность для оценки антигенсвязывающей активности.
Фармацевтическая композиция
Данное изобретение обеспечивает фармацевтическую композицию, содержащую анти-GPC3-антитело с модифицированным компонентом, представляющим сахарную цепь, данного изобретения.
Фармацевтическая композиция, содержащая композицию антител данного изобретения, применима для предупреждения и/или лечения заболеваний, ассоциированных с ростом клеток, таких как рак, и особенно применима для предупреждения и/или лечения рака печени. Фармацевтическая композиция, содержащая антитело данного изобретения, может быть приготовлена способами, известными квалифицированным в данной области специалистам. Эта фармацевтическая композиция может вводиться парентерально в форме инъекционного препарата, содержащего стерильный раствор или суспензию в воде или другой фармацевтически приемлемой жидкости. Например, эта фармацевтическая композиция может быть приготовлена подходящим образом посредством объединения этого антитела с фармацевтически приемлемыми носителями и средами, такими как стерильная вода и физиологический раствор, растительное масло, эмульгатор, суспендирующий агент, поверхностно-активное вещество, стабилизатор, ароматизирующий эксципиент, разбавитель, носитель, консервант, связывающий агент, с последующим смешиванием в унифицированной стандартной форме, требуемой для обычно приемлемой фармацевтической практики. Количество активного ингредиента, включенного в эти фармацевтические препараты, является таким, что обеспечивается подходящая доза в пределах предписанного диапазона.
Эта стерильная композиция для инъекции может быть приготовлена в соответствии с общепринятой фармацевтической практикой с использованием дистиллированной воды для инъекции в качестве носителя.
Например, физиологический раствор или изотонический раствор, содержащий глюкозу или другие добавки, такие как D-сорбит, D-манноза, D-маннит и хлорид натрия, могут быть использованы в качестве водного раствора для инъекции, необязательно в комбинации с подходящим солюбилизирующим агентом, например спиртом, таким как этанол и полиспирт, такой как пропиленгликоль или полиэтиленгликоль, и неионогенным поверхностно-активным веществом, таким как полисорбат 80™, HCO-50 и т.п.
Примеры маслянистой жидкости включают в себя кунжутное масло и соевое масло, и оно может быть объединено с бензилбензоатом или бензиловым спиртом в качестве солюбилизирующего агента. Другими компонентами, которые могут быть включены, являются фосфатный буфер или натрий-ацетатный буфер, болеутоляющее средство, такое как гидрохлорид прокаина, стабилизатор, такой как бензиловый спирт или фенол, и антиоксидант. Приготовленный инъекционный раствор будет обычно упакован в подходящую ампулу.
Способ введения является обычно парентеральным, например введение посредством инъекции, трансназальное введение, чреслегочное введение, чрескожное введение. Введение может быть системным или локальным, посредством внутривенного введения, внутримышечной инъекции, внутрибрюшинной инъекции, подкожной инъекции.
Подходящие способы введения могут быть выбраны в соответствии с возрастом и состоянием пациента. Однократная доза фармацевтической композиции, содержащей антитело или полинуклеотид, кодирующий это антитело, может быть выбрана из диапазона 0,001-1000 мг/кг массы тела. С другой стороны, можно также выбирать дозу в диапазоне 0,001-100000 мг/тело, но данное изобретение ни в какой мере не ограничивается этими численными диапазонами. Доза и способ введения будут зависеть от массы, возраста, состояния и т.п. пациента и может быть подходящим образом выбрана квалифицированными в данной области специалистами.
Содержание всех патентов и ссылочных документов, специально цитируемых в описании этой заявки, включено тем самым в качестве ссылки в их полном объеме. Кроме того, содержание описания и рисунки заявки на патент Японии 2004-311356, которая является основой для заявления приоритета этой заявки, включены тем самым в качестве ссылки в их полном виде.
ПРИМЕРЫ
Данное изобретение объясняется подробно при помощи следующих примеров, но никоим образом не ограничивается этими примерами.
Пример 1
Получение мышиного анти-GPC3-антитела
Растворимый белок GPC3, лишенный гидрофобного района С-конца (аминокислот 564-580), получали в качестве иммунизирующего белка для получения анти-GPC3-антитела для иммунизации. В качестве животного для иммунизации использовали мышь MRL/MpJUmmCrj-lpr/lpr (далее называемую мышью MRL/lpr, покупаемой из Charles River Japan), которая является мышью с аутоиммунным заболеванием. Иммунизацию начинали, когда возраст мыши был 7 или 8 недель, и препарат для начальной иммунизации корректировали до дозы 100 мкг/мышь растворимого GPC3. Эмульсию готовили с использованием полного адъюванта Фрейнда (FCA, Bekton Dickinson) и инъецировали подкожно. После серии из пяти иммунизаций дозу конечной иммунизации разбавляли в ЗФР до 50 мкг/мышь и инъецировали внутривенно через хвостовую (каудальную) вену. В день 4 после конечной иммунизации клетки селезенки извлекали, смешивали с клетками миеломы мыши Р3-Х63Ag8U1 (далее называемыми Р3U1, покупаемыми из АТСС) в соотношении 2:1, и проводили слияние клеток постепенным добавлением PEG-1500 (Roche Diagnostics). Гибридомы подвергали скринингу при помощи ELISA с использованием иммунопланшетов с иммобилизованным растворимым белком GPC3 основной структуры (кора). Положительные клоны моноклонировали процедурой предельных разведений. В результате получали 11 клонов антител с сильной активностью связывания GPC3 (М3С11, М13В3, М1Е7, М3В8, М11F1, L9G11, M19B11, M6B1, M18D4, M5B9 и М10D2).
Среди полученных анти-GPC3-антител особенно сильную CDC-активность проявляли M11F1 и М3В8. Таким образом, GST-слитый белок, содержащий эпитопы M11F1 и М3В8 (GC-3), который является слитым белком, содержащим пептид от 524 Ala до 563 Lys GPC3 и GST, использовали в качестве иммуногенов для иммунизации 3 мышей Balb/c (Charles River Japan) и 3 мышей MRL/lpr. Для первой иммунизации препарат GC-3 в концентрации 100 мкг/мышь эмульгировали с ФТС и инъецировали подкожно. Спустя две недели препарат 50 мкг/мышь эмульгировали с неполным адъювантом Фрейнда (FIA) и инъецировали подкожно. После пяти иммунизаций конечную иммунизацию (50 мкг/мышь) инъецировали внутривенно всем мышам через каудальную вену и подвергали слиянию клеток. Гибридомы подвергали скринингу при помощи ELISA с использованием иммунопланшетов с иммобилизованным растворимым белком GPC3 кора, лишенным гидрофобного района С-конца (аминокислот 564-580). Положительные клоны моноклонировали с использованием процедуры предельных разведений. В результате получали 5 клонов антител с сильной активностью связывания GPC3 (GC199, GC202, GC33, GC179 и GC194.)
Вариабельные области Н-цепи и L-цепи клонировали и каждую последовательность определяли стандартным способом. Кроме того, CDR-районы определяли сравнением с базой данных аминокислотных последовательностей известного антитела и проверяли на гомологию. Последовательности этих CDR-районов показаны в таблицах 1 и 2.
Пример 2
Получение химерного антитела анти-GPC3-антитела мышь-человек
Последовательности вариабельной области Н-цепи и L-цепи анти-GPC3-антитела GC33 лигировали с последовательностями константной области IgG1 и κ-цепи человека. ПЦР проводили с использованием синтетического олигонуклеотида, комплементарного 5'-концевой нуклеотидной последовательности вариабельной области Н-цепи антитела, имеющей последовательность Козака, и синтетического олигонуклеотида, комплементарного 3'-концевой нуклеотидной последовательности, имеющей сайт NheI. Полученный таким образом продукт ПЦР клонировали в вектор pB-CH, в котором константная область IgG1 человека была встроена в вектор pBluescript KS+ (Toyоbo Co., Ltd.). Вариабельную область Н-цепи мыши и константную область Н-цепи человека (γ1-цепь) лигировали через сайт NheI. Полученный таким образом фрагмент гена Н-цепи клонировали в экспрессирующий вектор pCXND3. Кроме того, проводили ПЦР с использованием синтетического олигонуклеотида, комплементарного 5'-концевой нуклеотидной последовательности вариабельной области L-цепи антитела, имеющей последовательность Козака, и синтетического олигонуклеотида, комплементарного 3'-концевой нуклеотидной последовательности, имеющей сайт BsiWI. Полученный таким образом продукт ПЦР клонировали в вектор pB-CL, в котором константная область κ-цепи человека была встроена в вектор pBluescript KS+ (Toyоbo Co., Ltd.). Вариабельную область и константную область L-цепи человека лигировали через сайт BsiWI. Полученный таким образом фрагмент гена L-цепи клонировали в экспрессирующий вектор pUCAG. Вектор pUCAG является вектором, полученным расщеплением pCXN (Niva et al., Gene, 1991, 108, 193-200) рестриктазой BamHI с получением фрагмента 2,6 т.п.н., который лигировали с сайтом рестриктазы BamHI вектора pUC19 (Toyobo Co., Ltd.).
Для получения экспрессирующего вектора химерного анти-GPC3-мышь-человек-антитела фрагмент гена получали расщеплением вектора pUCAG, в котором был вставлен фрагмент гена L-цепи, рестриктазой HindIII (Takara Shuzo Co., Ltd.). Этот фрагмент гена лигировали с сайтом расщепления рестриктазы HindIII pCXND3, содержащей ген Н-цепи, и затем клонировали. Полученная таким образом плазмида экспрессировала ген устойчивости к неомицину, ген DHFR и ген химерного анти-GPC3-мышь-человек-антитела в клетках животного. Аминокислотная последовательность вариабельной области Н-цепи показана в SEQ ID NO:3, а аминокислотная последовательность вариабельной области L-цепи показана в SEQ ID NO:4.
Пример 3
Получение химерного анти-GPC3-антитела с низким содержанием фукозы
Сначала клетки YB2/0 (ATCC, CRL-1662) культивировали в качестве клеток-хозяев в среде RPMI-1640, содержащей 10% ФТС. Затем 25 мкг экспрессирующего химерное анти-GPC3-антитело вектора, полученного, как описано в примере 2, вводили в клетки YB2/0 (ATCC CRT-1662) электропорацией в концентрации 7,5×106 клеток/0,75 мл ЗФР(-) при 1,4 кВ и 25 мкФ. После периода восстановления клеток в течение 10 мин при комнатной температуре эти клетки, обработанные электропорацией, суспендировали в 40 мл среды RPMI-1640, содержащей 10% ФТС. Готовили 10-кратное разведение с использованием той же самой среды. После культивирования в течение 24 часов в СО2-термостате (5% СО2) добавляли генетицин (Invitrogen Corp.) в концентрации 0,5 мг/мл и эти клетки культивировали в течение 2 недель. Клеточные линии с высоким уровнем экспрессии химерного антитела подвергали скринингу с использованием сэндвич-ELISA с антителом против IgG человека и устанавливали клеточные линии, стабильно экспрессирующие это антитело. Каждое химерное анти-GPC3-мышь-человек-антитело очищали с использованием Hi Trap Protein G HP (Amersham).
Пример 4
Измерение ADCC-активности с использованием PBMC из периферической крови человека
Получение раствора PBMC человека
Периферическую кровь с добавленным гепарином, собранную из здорового взрослого человека, разбавляли 2-кратно ЗФР(-) и наносили на Ficoll-Paque™ PLUS (Amersham). После центрифугирования (500×g, 30 минут, 20ºС) выделяли промежуточный слой, который является фракцией мононуклеарных клеток. После промывки этого слоя 3 раза эти клетки суспендировали в 10% ФТС/RPMI для получения раствора PBMC человека.
Получение клеток-мишеней
Клетки НерG2 (АТСС) и клетки HuH-7 (Health Science Research Resources Bank), культивируемые в среде 10% ФТС/RPMI-1640, отделяли от чашки с использованием буфера для диссоциации клеток (Invitrogen), наносили в виде аликвот в 96-луночный U-донный планшет (Falcon) при концентрации 1×104 клеток на лунку и культивировали в течение одного дня. После культивирования добавляли 5,55 МБк 51Cr и клетки культивировали 1 час при 37ºС в термостате с 5% газообразным СО2. Клетки промывали один раз культуральной средой и добавляли 50 мкл среды 10% ФТС/RPMI-1640 для получения клеток-мишеней.
Анализ высвобождения хрома (активности ADCC)
Объем 50 мкл раствора антитела, приготовленного в различных концентрациях, добавляли к клеткам-мишеням и давали реагировать на льду в течение 15 минут. Затем добавляли 100 мкл раствора PBMC человека (5×105 клеток на лунку) и эти клетки инкубировали в течение 4 часов при 37ºС в термостате с 5% газообразным СО2. После культивирования этот планшет центрифугировали и радиоактивность в 100 мкл культурального супернатанта измеряли с использованием гамма-счетчика. Удельную скорость высвобождения хрома определяли по следующей формуле.
Удельная скорость высвобождения хрома (%)=(А-С)×100/(В-С)
В этой формуле А представляет среднюю величину радиоактивности (имп/мин) в каждой лунке; В представляет среднюю величину радиоактивности (имп/мин) в лунке, в которой к клеткам-мишеням добавляли 100 мкл 2% водного раствора NP-40 (Нонидет Р-40, Номер кода 252-23, Nacalai Tesque) и 50 мкл среды 10% ФТС/RPMI; и С представляет среднюю величину радиоактивности (имп/мин) в лунке, в которой к клеткам-мишеням добавляли 150 мкл среды 10% ФТС/RPMI. Анализ проводили в трех повторностях и рассчитывали средние величины и стандартные отклонения ADCC-активности (%).
Фиг. 2 и 3 показывают ADCC-активность химерного анти-GPC3-антитела, измеренную с использованием PBMC. На этих фигурах вертикальная ось представляет цитотоксическую активность, а горизонтальная ось представляет концентрацию (мкг/мл) добавленного антитела. Фиг. 2 показывает результаты, когда в качестве клеток-мишеней использовали клетки HepG2, а фиг. 3 показывает результаты для клеток HuH-7. Белые кружки показывают активность химерного антитела GC33, продуцируемого клетками СНО, а черные кружки показывают активность химерного антитела GC33, продуцируемого клетками YB2/0. Химерное антитело GC33 с низкой фукозой, продуцируемое клетками YB2/0, обнаруживает более сильную активность ADCC, чем химерное антитело GC33, продуцируемое клетками СНО, ясно свидетельствуя о том, что ADCC-активность анти-GPC3-антитела усиливается модификацией сахарной цепи.
Пример 5
Создание антитело-продуцирующих клеток
Гигромицин В добавляли в среду SFMII(+) в конечной концентрации 1 мг/мл и клеточную линию, недостаточную в отношении транспортера фукозы (клон 3F2), субкультивировали в этой среде. Готовили суспензию клеток 3F2 в фосфатном буфере Дульбекко (8×106 клеток/0,8 мл). К этой суспензии клеток добавляли 25 мкг экспрессирующего антитело вектора (Ссылочные примеры 1 и 2) и эту суспензию клеток переносили в Gene Pulser Cuvette. После оставления этой кюветы на льду в течение 10 минут этот вектор вводили в клетки электропорацией с использованием GENE-PULSER II при 1,5 кВ и 25 мкФ. Эти клетки суспендировали в 40 мл среды SFMII(+) и переносили в 96-луночный плоскодонный планшет (Iwaki) при 100 мкл на лунку. После инкубирования этого планшета в СО2-термостате в течение 24 часов при 37ºС добавляли генетицин (Invitrogen Cat. No. 10131-027) в конечной концентрации 0,5 мг/мл. Количество антитела, продуцируемого устойчивыми к лекарственному средству клетками, измеряли для установления клеточных линий, продуцирующих гуманизированное анти-GPC3-антитело.
Пример 6
Очистка антител
Супернатант из экспрессирующей антитело клеточной линии собирали и наносили на колонку Hitrap™ rProtein A (Pharmacia Cat. No. 17-5080-01) с использованием насоса Р-1 (Pharmacia). После промывания этой колонки буфером для связывания (20 мМ фосфат натрия, рН 7,0)) этот белок элюировали буфером для элюции (0,1 М глицин-HCl (рН 2,7)). Элюат сразу же нейтрализовали нейтрализующим буфером (1 М Трис-HCl (рН 9,0)). Фракции элюции антитела собирали с использованием DC-анализа белка (BIO-RAD Cat. No. 500-0111) и объединяли и концентрировали до приблизительно 2 мл с использованием Centriprep-YM10 (Millipore Cat. No. 4304). Затем эти антитела разделяли гель-фильтрацией с использованием колонки Superdex 200 26/60 (Pharmacia), уравновешенной 20 мМ ацетатным буфером с 150 мМ NaCl (рН 6,0). Пики фракции мономера собирали, концентрировали с использованием Centriprep-YM10 и фильтровали через фильтрующий элемент MILLEX-GW (Millipore Cat. No. SLGV 013SL) и затем хранили при 4оС. Измеряли поглощение при 280 нм и концентрацию очищенного антитела рассчитывали из коэффициента молярного поглощения.
Пример 7
Активность ADCC in vitro гуманизированного анти-GPC3-антитела, продуцируемого клетками FT-KO
Фиг. 4 показывает активность ADCC in vitro анти-GPC3-антитела, продуцируемого клетками FT-KO, при использовании PBMC человека. Этот способ является способом, описанным в примере 4. На этой фигуре вертикальная ось представляет цитотоксическую активность (%), а горизонтальная ось представляет концентрацию (мкг/мл) добавленного антитела. В качестве клеток-мишеней использовали клетки HuH-7. Белые кружки показывают активность анти-GPC3-антитела, продуцируемого клетками СНО дикого типа, а черные кружки показывают активность анти-GPC3-антитела, продуцируемого клетками FT-KO. Анти-GPC3-антитело с низкой фукозой, продуцируемое клетками FT-KO, обнаруживает более сильную активность ADCC, чем анти-GPC3-антитело, продуцируемое клетками СНО дикого типа, что ясно указывает на то, что активность ADCC анти-GPC3-антитела, продуцируемого клетками FT-KO, является повышенной.
Пример 8
Анализ сахарных цепей гуманизированного анти-GPC3-антитела, продуцируемого клетками FT-KO
1. Получение 2-аминобензамид-меченых сахарных цепей (2АВ-меченых сахарных цепей)
Антитело, продуцируемое клетками FT-KO данного изобретения, и антитела, продуцируемые клетками СНО, в качестве контрольной пробы обрабатывали N-глюкозидазой F (Roche Diagnostics) для высвобождения этих сахарных цепей из белка (Weitzhandler M. et al., Journal of Pharmaceutical Sciences 83:12 (1994), 1670-1675). После удаления белка этанолом (Schenk B. et al., The Journal of Clinical Investigation 108:11 (2001), 1687-1695) сахарные цепи концентрировали и сушили и флуоресцентно метили 2-аминопиридином (Bigge J.C. et al., Analitical Biochemistry 230:2 (1995), 229-238). Реагент удаляли из 2-АВ-меченых сахарных цепей твердофазной экстракцией с использованием целлюлозного картриджа и после концентрирования центрифугированием получали 2-АВ-меченые сахарные цепи. Затем эти очищенные 2-АВ-меченые сахарные цепи обрабатывали β-галактозидазой (Seikaguku Corp.) с получением α-галактозил-2-АВ-меченых сахарных цепей.
2. Анализ α-галактозил-2-АВ-меченых сахарных цепей с использованием ВЖХ с нормальной фазой
Антитела, продуцируемые клетками FT-KO данного изобретения, и антитела, продуцируемые клетками СНО, в качестве контрольной пробы, получали в виде α-галактозил-2-АВ-меченых сахарных цепей в соответствии с описанным выше способом и анализировали ВЖХ с нормальной фазой с использованием амидной колонки (Tosoh Corp. TSKgel Amide-80) и хроматограммы сравнивали. В антителах, продуцируемых клетками СНО, основным компонентом является G(0), а G(0)-Fuc ответственен за приблизительно 4% площади пика. С другой стороны, в антителах, продуцируемых клетками FT-KO, основным компонентом является G(0)-Fuc, и он присутствует по меньшей мере в 90% площади пика в каждой из клеточных линий (фиг. 5 и таблица 2). Фиг. 6 показывает предположительные структуры для пиков G(0) и G(0)-Fuc.
Пример 9
Анализ термостабильности гуманизированного анти-GPC3-антитела, продуцируемого клетками FT-KO
1. Приготовление раствора пробы для DSC-измерения
Наружный диализный раствор был 20 моль/л натрий-ацетатным буфером (рН 6,0), содержащим 200 ммоль/л хлорида натрия. Диализную мембрану, наполненную 700 мкг-эквивалентами раствора антитела, диализовали погружением в наружный диализный раствор в течение ночи для получения раствора пробы.
2. Измерение температуры термодеструкции с использованием DSC (дифференциальной сканирующей калориметрии)
После тщательного дегазирования раствора пробы и ссылочного раствора (наружного диализного раствора) каждый их них помещали в калориметр и термически уравновешивали при 20ºС. Затем проводили DSC-измерение от 20ºС до 100ºС при скорости сканирования приблизительно 1 К/мин. Этот результат представлен наклоном пика разложения как функции температуры (фиг. 7). Было обнаружено, что температура термодеструкции антител, продуцируемых клетками СНО, и антител, продуцируемых клетками FT-KO, является эквивалентной.
Ссылочный пример 1
Гуманизация GC33
Информацию о последовательности антитела получали из публично доступной базы данных Кабата (ftp://ftp.ebi.ac.uk.pub/databases/kabat/) и вариабельную область Н-цепи и вариабельную область L-цепи по отдельности подвергали поиску гомологии. Было обнаружено, что вариабельная область Н-цепи имеет высокий уровень гомологии с DN13 (Smithson et al., Mol. Immunol. 1999; 36: 113-124). Было также обнаружено, что вариабельная область L-цепи имеет высокий уровень гомологии с мРНК IgК Homo sapiens в отношении VLJ-области легкой цепи каппа, частичных CDS иммуноглобулина, клона: К64 с Accession Number AB064105. Сигнальную последовательность Accession Number S40357, которая имеет высокий уровень гомологии с АВ064105, использовали в качестве сигнальной последовательности L-цепи. Затем CDR трансплантировали в FR этих антител для получения гуманизированного антитела.
Более конкретно, синтетические олиго-ДНК с длиной приблизительно 50 оснований конструировали таким образом, что приблизительно 20 оснований из них гибридизовались друг с другом, и эти синтетические олиго-ДНК собирали при помощи ПЦР для получения гена, кодирующего каждую вариабельную область. Их расщепляли в последовательности HindIII, встроенной на конце 5'-конца синтетического ДНК-олиго, и в последовательности BamHI, встроенной на конце 3'-конца олиго-ДНК, и этот синтетический ДНК-олиго клонировали в экспрессирующий вектор HEFgγ1, когда клонировали константную область IgG1 человека, или экспрессирующий вектор HEFgκ, когда клонировали константную область κ-цепи человека (Sato et al., Mol. Immunol., 1994; 371-381). Н-цепь и L-цепь этого гуманизированного GC33, сконструированные, как описано выше, каждую, обозначали ver.a. Гуманизированное антитело GC33 (ver.a/ver.a), в котором обе цепи, Н-цепь и L-цепь, были ver.a, имели более низкую активность связывания, чем антитело с вариабельными областями GC33 мыши (мышь/мышь). Химерные антитела получали объединением мышиных последовательностей GC33 и последовательностей ver.a для Н-цепей и L-цепей (мышь/ver.a, ver.a/мышь) и оценивали активность связывания. Более низкая активность связывания была обнаружена с антителом ver.a, что свидетельствует о том, что уменьшение активности связывания обусловлено заменой аминокислот, которая была связана с Н-цепью. Затем готовили модифицированные Н-цепи, названные ver.с, ver.f, ver.h, ver.i, ver.j и ver.k. Все гуманизированные антитела GC33 проявляли тот же самый уровень активности связывания, что и химерное антитело, имеющее вариабельные области мышиного GC33. Нуклеотидные последовательности вариабельных областей Н-цепи гуманизированного GC33 ver.a, ver.с, ver.f, ver.h, ver.i, ver.j и ver.k показаны в SEQ ID NO:74, 75, 76, 78, 79 и 80, а все их аминокислотные последовательности показаны в SEQ ID NO:81, 82, 83, 84, 85, 86 и 87, соответственно. Нуклеотидная последовательность вариабельной области ver.a L-цепи гуманизированного GC33 показана в SEQ ID NO:88, а ее аминокислотная последовательность показана в SEQ ID NO:89, соответственно. В вариабельных областях ver.i, ver.j и ver.k Н-цепи гуманизированного GC33 шестая глутаминовая кислота заменена глутамином. Эти антитела проявляли явно увеличенную термостабильность.
Ссылочный пример 2
Изменение L-цепи гуманизированного GC33
Что касается деамидирования белка, было известно, что константа скорости реакции деамидирования зависит от первичной последовательности. Известно также, что последовательность Asn-Gly особенно чувствительна к деамидированию (Rocinson et al., Proc. Natl. Acad. Sci. USA 2001; 98: 944-949). Поскольку Asn 33 в CDR1 вариабельной области ver.a L-цепи гуманизированного GC33 SEQ ID NO:88 имеет первичную последовательность Asn-Gly, предсказывается, что этот остаток является чувствительным к деамидированию.
Для оценки действия деамидирования Asn 33 на активность связывания этого антитела получали модифицированное антитело, в котором Asn 33 был заменен Asp. Набор Quick Change Site-Directed Mutagenesis Kit (Stratagene) использовали для введения точковой мутации. Более конкретно, 50 мкл реакционного раствора, содержащего 125 нг смыслового праймера (CTT GTA CАC AGT GAC GGA AAC ACC TAT: SEQ ID NO:124), 125 нг антисмыслового праймера (ATA GGT GTT TCC GTC ACT GTG TAC AAG: SEQ ID NO:125), 5 мкл 10х реакционного буфера, 1 мкл смеси dNTP, 10 нг вектора HEFgκ, в который была клонирована L-цепь ver.a гуманизированного GC33, и 1 мкл ДНК-полимеразы Pfu Turbo, подвергали 12 циклам, состоящим из 30 сек при 95ºС, 1 мин при 55ºС и 9 мин при 68ºС. Продукт реакции расщепляли рестриктазой DpnI в течение 2 часов при 37ºС и вводили в XL1-Blue компетентные клетки для получения трансформантов. Вариабельную область вырезали из клонов, содержащих корректную мутацию, и опять клонировали в экспрессирующий вектор HEFgκ. Экспрессирующий вектор HEFgy1, содержащий Н-цепь ver.k гуманизированного GC33, вводили в клетки COS7 с использованием Fugene 6 (Roche). Супернатант культуральной среды собирали из клеток, временно экспрессирующих это модифицированное антитело. Концентрацию антитела количественно определяли сэндвич-ELISA с использованием антитела против IgG человека и активность связывания этого модифицированного антитела оценивали при помощи ELISA с использованием планшета, покрытого растворимым белком-кором GPC3. Активность связывания терялась в модифицированном антителе (N33D), в котором Asn 33 был заменен Asp, что позволяет предположить, что на активность связывания значимо влияет деамидирование в Asn 33.
Сообщалось, что деамидирование Asn 33 подавляется заменой Gly 34 другим аминокислотным остатком (WO 03057881 А1). В соответствии с этим способом, получали ряд модифицированных антител заменой G34 17 другими аминокислотными остатками за исключением Cys и Met с использованием набора Quick Change Site-Directed Mutagenesis Kit с получением G34A, G34D, G34E, G34F, G34H, G34N, G34P, G34Q, G34I, G34K, G34L, G34V, G34W, G34Y, G34R, G34S и G34T. Активность связывания этих антител оценивали с использованием культурального супернатанта клеток COS7, временно экспрессирующих эти антитела. Было обнаружено, что активность связывания сохранялась, даже если G34 заменяют другим аминокислотным остатком, иным чем Pro (G34P) и Val (G34V).
Аминокислотные последовательности CDR1 L-цепи модифицированных антител представлены SEQ ID NO: 90 (G34A), SEQ ID NO: 91 (G34D), SEQ ID NO: 92 (G34E), SEQ ID NO: 93 (G34F), SEQ ID NO: 94 (G34H), SEQ ID NO: 95 (G34N), SEQ ID NO: 96 (G34T), SEQ ID NO: 97 (G34Q), SEQ ID NO: 98 (G34I), SEQ ID NO: 99 (G34K), SEQ ID NO: 100 (G34L), SEQ ID NO: 101 (G34S), SEQ ID NO: 102 (G34W), SEQ ID NO: 103 (G34Y), SEQ ID NO: 104 (G34R), SEQ ID NO: 105 (G34V) и SEQ ID NO: 106 (G34P), соответственно. Аминокислотные последовательности вариабельных областей L-цепи модифицированных антител представлены SEQ ID NO: 107 (G34A), SEQ ID NO: 108 (G34D), SEQ ID NO: 109 (G34E), SEQ ID NO: 110 (G34F), SEQ ID NO: 111 (G34H), SEQ ID NO: 112 (G34N), SEQ ID NO: 113 (G34T), SEQ ID NO: 114 (G34Q), SEQ ID NO: 115 (G34I), SEQ ID NO: 116 (G34K), SEQ ID NO: 117 (G34L), SEQ ID NO: 118 (G34S), SEQ ID NO: 119 (G34W), SEQ ID NO: 120 (G34Y), SEQ ID NO: 121 (G34R), SEQ ID NO: 122 (G34V) и SEQ ID NO: 123 (G34P), соответственно.
Ссылочный пример 3
Деструкция гена транспортера фукозы в клетках СНО
1. Конструктирование нацеливающего вектора
(1) Получение вектора КО1
Ген устойчивости к гигромицину (Hygr) конструировали при помощи ПЦР с праймерами Hyg5-BH и Hyg3-NT из плазмиды pcDNA3.1/Hygro (Invitrogen), которая имеет последовательность, идентичную 5'-части стартового кодона гена транспортера фукозы, присоединением сайта BamHI и последовательности TGCGC к 5'-части стартового кодона и сайта NotI, добавленного к 3'-части, содержащей этот район, до сигнала присоединения полиА SV40, и Hygr-фрагмент вырезали.
Прямой праймер
Hyg5-BH 5'-GGA TCC TGC GCA TGA AAA AGC CTG AAC TCA CC-3' (SEQ ID NO:128)
Обратный праймер
Hyg3-NT 5'-GCG GCC GCC TAT TCC TTT GCC CTC GGA CG-3' (SEQ ID NO:129)
Нацеливающий на транспортер фукозы вектор ver.1 (далее называемый вектором КО1) конструировали встраиванием 5'-части нуклеотидной последовательности транспортера фукозы (от SmaI при основании № 2780 до BamHI при основании № 4323 нуклеотидной последовательности, показанной в SEQ ID NO:126), 3'-части (от основания № 4284 до SacI при основании № 10934) и Hygr-фрагмента в вектор pMC1DT-A (Yagi T, Proc. Natl. Acad. Sci. USA, Vol. 87, p. 9918-9922, 1990). Характеристикой этого вектора КО1 является то, что этот Нygr-фрагмент будет экспрессироваться от промотора транспортера фукозы, когда имеет место гомологичная рекомбинация, так как к этому Hygr-фрагменту не присоединен промотор. Однако Hygr не всегда экспрессируется до такой степени, что образуется устойчивость к гигромицину В, если только одна копия этого вектора инсертируется в клетку гомологичной рекомбинацией. Вектор КО1 расщепляли NotI и вводили в клетку. Ожидается, что транспортер фукозы будет терять 41 пару оснований экзона 1, в том числе стартовый кодон, вследствие введения вектора КО1, что будет приводить к потере его функции.
(2) Получение pBSK-pgk-1-Hygr
Вектор pBSK-pgk-1 получали отрезанием промотора гена pgk-1 мыши от вектора pKJ2 (Popo H, Biochemical Genetics, Vol. 28, p. 299-308, 1990) рестриктазами EcoRI-PstI и клонированием его в сайт EcoRI-PstI pBluescript (Stratagene). При помощи ПЦР с праймерами Hyg5-AV и Hyg3-BH из pcDNA3.1/Hygro сайт EcoT22I и последовательность Козака присоединяли к 5'-части Hygr, а сайт BamHI добавляли к 3'-части, содержащей район до сигнала присоединения поли А SV40, и затем вырезали Hygr-фрагмент.
Прямой праймер
Hyg5-AV ATG CAT GCC ACC ATG AAA AAG CCT GAA CTC ACC-3' (SEQ ID NO:130)
Обратный праймер
Hyg3-BH 5'-GGA TCC CAG GCT TTA CAC TTT ATG CTT C-3' (SEQ ID NO:131)
Вектор pBSK-pgk-1-Hygr получали встраиванием Hygr (EcoT22I-BamHI)-фрагмента в сайт PstI-BamHI pBSK-pgk-1.
(3) Получение вектора КО2
Нацеливающий на транспортер фукозы вектор ver.2 (далее называемый вектором КО2) конструировали встраиванием 5'-части нуклеотидной последовательности транспортера фукозы (от SmaI при основании № 2780 до BamHI при основании № 4323 нуклеотидной последовательности, показанной в SEQ ID NO:126), 3'-части (от основания № 4284 до SacI при основании № 10934) и pgk-1-Hygr-фрагмента в вектор pMC1DT-A. В отличие от вектора КО1 вектор КО2 будет придавать устойчивость к гигромицину В, даже если только одна копия этого вектора встроена гомологичной рекомбинацией, так как промотор гена pgk-1 присоединен к Hygr. Вектор КО2 расщепляли NotI и вводили в клетки. Ожидается, что транспортер фукозы будет терять 46 пар оснований экзона 1, в том числе стартовый кодон, вследствие введения вектора КО2, что будет приводить к потере его функции.
(4) Получение pBSK-pgk-1-Puror
Вектор pBSK-pgk-1-Puror получали расщеплением вектора pPUR (BD Biosciences) PstI и BamHI и встраиванием этого расщепленного фрагмента (Puror) в сайт PstI-BamHI pBSK-pgk-1.
(5) Получение вектора КО3
Нацеливающий на транспортер фукозы вектор ver.3 (далее называемый вектором КО3) конструировали встраиванием 5'-части нуклеотидной последовательности транспортера фукозы (от SmaI при основании № 2780 до BamHI при основании № 4323 нуклеотидной последовательности, показанной в SEQ ID NO:126), 3'-части (от основания № 4284 до SacI при основании № 10934) и pgk-1-Puror-фрагмента в вектор pMC1DT-A. Кроме того, к 3'-концу pgk-1-Puror присоединяли последовательность для связывания с праймером для описанного ниже скрининга. Вектор КО3 расщепляли NotI и встраивали в клетки. Ожидается, что транспортер фукозы будет терять 46 пар оснований экзона 1, в том числе стартовый кодон, вследствие введения вектора КО3, что будет приводить к потере его функции.
Обратный праймер
RSGR-A 5'-GCT GTC TGG AGT ACT GTG CAT CTG C-3' (SEQ ID NO:132)
Описанные выше три вида нацеливающих векторов использовали для нокаута гена транспортера фукозы.
2. Введение векторов в клетки СНО
НТ-добавку (100х) (Invitrogen Cat. No. 11067-030) и пенициллин-стрептомицин (Invitrogen Cat. No. 15140-122) добавляли к СНО-S-FMII HT (Invitrogen Cat. No. 12052-098), каждый в объеме 1/100 относительно объема СНО-S-FMII HT. Клетки СНО DXB11 субкультивировали в культуральной среде (далее называемой SEMII(+)), и эту среду SEMII(+) использовали для культивирования этих клеток после переноса гена. Клетки СНО суспендировали в фосфатном буфере Дульбекко (далее называемом ЗФР, Invitrogen cat. No. 14190-144) при концентрации (8×106 клеток/0,8 мл). Затем к этой суспензии клеток добавляли 30 мкг нацеливающего вектора и эту суспензию клеток переносили в Gene Pulser Cuvette (4 мм) (Bio-Rad, Cat. No. 1652088). После оставления этой кюветы на льду в течение 10 минут этот вектор вводили в клетки электропорацией с использованием GENE-PULSER II (Bio-Rads, Code No. 340BR) при 1,5 кВ и 25 мкФ. После введения этого вектора клетки суспендировали в 200 мл среды SFMII(+) и переносили в двадцать 96-луночных плоскодонных планшетов (Iwaki, Cat. No. 1860-096) при 100 мкл на лунку. После инкубирования этих планшетов в СО2-термостате в течение 24 часов при 37оС добавляли реагент.
3. Стадия нокаута 1
Либо вектор КО1, либо вектор КО2 вводили в клетки СНО и после 24 часов эти клетки подвергали отбору с использованием гигромицина В (Invitrogen, Cat. No. 10687-010). Гигромицин В растворяли в SFMII(+) до концентрации 0,3 мг/мл и добавляли при 100 мкл на лунку.
4. Скрининг на гомологичные рекомбинанты при помощи ПЦР
(1) Приготовление ПЦР-пробы
Скрининг на гомологичные рекомбинанты проводили при помощи ПЦР. Клетки СНО, используемые в скрининге, культивировали в 96-луночных планшетах. После удаления супернатанта добавляли 50 мкл на лунку буфера для цитолиза и эти клетки сначала нагревали при 55оС в течение 2 часов и затем при 95оС в течение 15 минут для инактивации протеазы К для получения ПЦР-матрицы. Буфер для цитолиза состоял из 5 мкл 10×LA-буфер II (Takara Bio Inc., дополненный La Taq), 2,5 мкл 10% NP-40 (Roche, Cat. No. 1 332 473), 4 мкл протеиназы К (20 мг/мл, Takara Bio Inc. Cat. No. 9033) и 38,5 мкл дистиллированной воды (Nacalai Tesque Cat. No. 36421-35) на лунку.
(2) Условия ПЦР
Реакционная смесь ПЦР содержала 1 мкл вышеуказанной пробы ПЦР, 5 мкл 10 х LA-буфера II, 5 мкл MgCl2 (25 мМ), 5 мкл dNTP (2,5 мМ), 2 мкл праймера (10 мкМ каждого), 0,5 мкл LA Taq (5 МЕ/мкл, Cat. No. RR002B) и 29,5 мкл дистиллированной воды (общий объем 50 мкл). Для скрининга клеток, содержащих вектор КО1, в качестве ПЦР-праймеров использовали ТР-F4 и THygro-R1, а для скрининга клеток, содержащих вектор КО2, в качестве праймеров использовали ТР-F4 и THygro-F1.
Условия ПЦР для клеток, содержащих вектор КО1, состояли из предварительного нагревания при 95ºС в течение 1 минуты, проведения 40 циклов, состоящих из цикла амплификации 30 секунд при 95ºС, 30 секунд при 60ºС и 2 минут при 60ºС, и затем повторного нагревания при 72ºС в течение 7 минут. Условия ПЦР для клеток, содержащих вектор КО2, состояли из предварительного нагревания при 95ºС в течение 1 минуты, проведения 40 циклов, состоящих из цикла амплификации 30 секунд при 95ºС и 3 минут при 70ºС, и затем повторного нагревания при 70ºС в течение 7 минут.
Используемые праймеры приведены ниже. В пробах клеток, в которых происходит гомологичная рекомбинация с вектором КО1 или вектором КО2, будет амплифицироваться ДНК приблизительно 1,6 т.п.н. или 2,0 т.п.н., соответственно. Праймер ТР-F4 был сконструирован для 5'-района генома транспортера фукозы снаружи от вектора, а THygro-F1 и THygro-R1 были сконструированы для гена Hygr внутри этого вектора.
Прямой праймер (КО1, КО2)
ТР-F4 5'-GGA ATG CAG CTT CCT CAA GGG ACT CGC-3' (SEQ ID NO:133)
Обратный праймер (КО1)
THygro-R1 5'-TGC ATC AGG TCG GAG ACG CTG TCG AAC-3' (SEQ ID NO:134)
Обратный праймер (КО2)
THygro-F1 5'-GCA CTC GTC CGA GGG CAA AGG AAT AGC-3' (SEQ ID NO:135)
5. Результаты ПЦР-скрининга
В целом анализировали 918 клеток, содержащих вектор КО1, и одна клетка была, по-видимому, гомологичным рекомбинантом (степень гомологичной рекомбинации: приблизительно 0,1%). В целом анализировали 537 клеток, содержащих вектор КО2, и 17 клеток, по-видимому, являются гомологичными рекомбинантами (степень гомологичной рекомбинации: приблизительно 3,2%).
6. Блот-анализ по Саузерну
Гомологичную рекомбинацию дополнительно подтверждали блоттингом по Саузерну. В целом 10 мкг геномной ДНК получали из культивированных клеток согласно стандартному способу для анализа в блоттинге по Саузерну. ПЦР проводили с использованием двух праймеров, приведенных ниже, для получения зонда 387 п.н., соответствующего району оснований с номерами 2113-2500 нуклеотидной последовательности, показанной в SEQ ID NO:126, который использовали в блоттинге по Саузерну. Эту геномную ДНК расщепляли BglII.
Прямой праймер
Bgl-F: 5'-TGT GCT GGG AAT TGA ACC CAG GAC-3' (SEQ ID NO:136)
Обратный праймер
Bgl-R: 5'-CTA CTT GTC TGT GCT TTC TTC C-3' (SEQ ID NO:137)
В результате расщепления BglII этот блот будет показывать полосу приблизительно 30 т.п.н. из хромосомы транспортера фукозы, приблизительно 4,6 т.п.н. из хромосомы, в которой произошла гомологичная рекомбинация с вектором КО1, и приблизительно 5,0 т.п.н. из хромосомы, в которой произошла гомологичная рекомбинация с вектором КО2. Этот эксперимент содержал 1 клетку из гомологичной рекомбинации с вектором КО1 и 7 клеток из гомологичной рекомбинации с вектором КО2. Эта единственная клетка, полученная из гомологичной рекомбинации с вектором КО1, была сначала названа 5С1, но более поздний анализ выявил, что эта клетка состояла из популяций множественных клеток. Таким образом, эти клетки клонировали лимитирующим разведением перед использованием в этом эксперименте. Одна из клеток, полученная с вектором КО2, была названа 6Е2.
7. Стадия нокаута 2
Эти три вектора использовали для создания клеточных линий, полностью дефектных по гену транспортера фукозы, из клеток, в которых имела место гомологичная рекомбинация с векторами КО1 и КО2. Комбинации векторов и клеток были следующими: Способ 1 объединял вектор КО2 и клетки 5С1 (КО1), Способ 2 объединял вектор КО2 и клетки 6Е2 (КО2) и Способ 3 объединял вектор КО3 и клетки 6Е2 (КО2). Каждый вектор вводили в подходящие клетки и после 24 часов начинали отбор с использованием гигромицина В и пуромицина (Nacalai Tesque, Cat. No. 294-12). Конечная концентрация гигромицина В была установлена на 1 мг/мл в Способе 1 и 7 мг/мл в Способе 2. В Способе 3 гигромицин добавляли при конечной концентрации 0,15 мг/мл, а пуромицин - в конечной концентрации 8 мкг/мл.
8. Скрининг на гомологичные рекомбинанты при помощи ПЦР
Для скрининга клеток из Способа 1 ПЦР проводили для детектирования клеток, имеющих гомологичную рекомбинацию как с вектором КО1, так и с вектором КО2. Для клеток из Способа 2 были сконструированы следующие праймеры ПЦР: TPS-F1 был образован в районе оснований с номерами 3924-3950 SEQ ID NO:126, а SHyhro-R1 был образован в районе оснований с номерами 4248-4274. Эти праймеры будут амплифицировать 350 п.н. района гена транспортера фукозы, содержащего делецию вследствие вектора КО2. Таким образом, в ПЦР-скрининге в Способе 2 клетки, не обеспечивающие продукта амплификации 350 п.н., считаются полностью лишенными гена транспортера фукозы. Условия ПЦР состояли из предварительного нагревания в течение 1 минуты при 95ºС, 35 циклов, состоящих из цикла амплификации 30 секунд при 95ºС и 1 минуты при 70ºС, и повторного нагревания 7 минут при 70ºС.
Прямой праймер
TPS-F1: 5'-CTC GAC TCG TCC CTA TTA GGC AAC AGC-3' (SEQ ID NO:138)
Обратный праймер
SHygro-R1: 5'-TCA GAG GCA GTG GAG CCT CCA GTC AGC-3' (SEQ ID NO:139)
Для способа 3 ТР-F4 использовали в качестве прямого праймера и RSGR-A использовали в качестве обратного праймера. Условия ПЦР состояли из предварительного нагревания в течение 1 минуты при 95ºС, 35 циклов цикла амплификации 30 секунд при 95ºС, 30 секунд при 60ºС и 2 минуты при 72ºС, и повторного нагревания 7 минут при 72ºС. В пробе клеток, имеющих гомологичную рекомбинацию с вектором КО3, будет амплифицироваться ДНК приблизительно 1,6 т.п.н. Эта процедура ПЦР будет детектировать клетки, имеющие гомологичную рекомбинацию с вектором КО3, а также клетки, все еще имеющие гомологичную рекомбинацию с вектором КО2.
9. Результаты ПЦР-скрининга
В Способе 1 анализировали в целом 616 клеток, и 18 клеток, по-видимому, были гомологичными рекомбинантами (степень гомологичной рекомбинации: 2,9%). В способе 2 анализировали в целом 524 клетки, и 2 клетки, по-видимому, были гомологичными рекомбинантами (степень гомологичной рекомбинации: 0,4%). Кроме того, в Способе 3 анализировали в целом 382 клетки, и 7 клеток, по-видимому, были гомологичными рекомбинантами (степень гомологичной рекомбинации: 1,8%).
10. Блот-анализ по Саузерну
Блоттинг по Саузерну проводили в соответствии с описанным выше способом. Среди анализированных клеток была обнаружена 1 клетка, полностью лишенная гена транспортера фукозы. В стадии нокаута 1 результаты анализа ПЦР и блоттинга по Саузерну согласуются, но в стадии нокаута 2 они не согласуются. Возможные причины являются следующими. 1. В Способе 1 смешивали вместе клетки, имеющие гомологичную рекомбинацию независимо от КО1 или КО2. 2. Ген транспортера фукозы присутствовал не в виде одной пары (2 генов), а в виде множественных пар (или в виде более, чем 3 генов). 3. Во время культивирования клеточных линий, созданных в стадии нокаута 1, количество копий гена транспортера фукозы, остающееся в субкультивируемых клетках, увеличивалось.
11. Анализ экспрессии фукозы
Экспрессию фукозы анализировали при помощи ПЦР в 26 клетках, которые, как было обнаружено, являются гомологичными рекомбинантами. В целом 1×106 клеток окрашивали на льду в течение 1 часа 100 мкл ЗФР, содержащего 5 мкг/мл агглютинина Lens culinaris, конъюгата FITC (Vector Laboratories, Cat. No. FL-1041), 2,5% ФТС и 0,02% азид натрия (далее называемого FACS-раствором). Затем эти клетки промывали 3 раза FACS-раствором и анализировали с использованием FACSСalibur (Becton Dickinson). Эти результаты ясно показали, что экспрессия фукозы уменьшается только в клетках, которые, как было обнаружено в блоттинг-анализе по Саузерну, лишены гена транспортера фукозы.
Приведенные выше результаты обнаружили следующее.
На основании того факта, что только одна клетка из 616 подвергнутых скринингу клеток имеет полную делецию гена транспортера фукозы, частота гомологичной рекомбинации была очень низкой, приблизительно 0,16%. Как отмечалось выше, имеются несколько возможных причин того, что результаты ПЦР и блоттинг-анализа по Саузерну не согласовались. Однако клеточные линии, полученные в Способе 3, могут не содержать смеси клеток, имеющих гомологичную рекомбинацию независимо от векторов КО2 и КО3, так как отбор выполняли с использованием двух типов лекарственных средств. Кроме того, вряд ли все другие клеточные линии, имеющие гомологичную рекомбинацию с использованием ПЦР, содержат многочисленные популяции клеток. Как отмечалось выше, если присутствуют 3 или более генов транспортера фукозы, нацеливание на этот ген в клетках было бы особенно трудным. Гомологичные рекомбинанты могли бы быть получены только с использованием такого вектора, как вектор КО1, где ген Hygr едва ли экспрессируется, и скринингом большого количества клеток.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ ГЛИПИКАНА 3 | 2005 |
|
RU2427588C2 |
| АНТИТЕЛО ПРОТИВ ГЛИПИКАНА 3 | 2011 |
|
RU2611751C2 |
| АНТИТЕЛО ПРОТИВ ГЛИПИКАНА-3 С УЛУЧШЕННЫМИ КИНЕТИЧЕСКИМИ ПОКАЗАТЕЛЯМИ В ПЛАЗМЕ | 2008 |
|
RU2445366C2 |
| ЛЕКАРСТВЕННОЕ СРЕДСТВО ДЛЯ ЛЕЧЕНИЯ РАКА ПЕЧЕНИ | 2009 |
|
RU2523897C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2011 |
|
RU2774414C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2011 |
|
RU2663123C2 |
| ВЫДЕЛЕННОЕ АНТИ-CD30 АНТИТЕЛО (ВАРИАНТЫ), ХОЗЯЙСКАЯ КЛЕТКА, СПОСОБ ПОЛУЧЕНИЯ ХИМЕРНОГО ИЛИ ГУМАНИЗИРОВАННОГО ВАРИАНТА АНТИ-CD30 АНТИТЕЛ (ВАРИАНТЫ), СПОСОБ ИНГИБИРОВАНИЯ РОСТА КЛЕТОК CD30+ И СПОСОБ ИНГИБИРОВАНИЯ РОСТА ОПУХОЛЕВЫХ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ CD30 | 2006 |
|
RU2492186C2 |
| БЕЛОК СЛИЯНИЯ НАЦЕЛЕННОГО НА GPC3 АНТИТЕЛА И ИНТЕРФЕРОНА α И ЕГО ПРИМЕНЕНИЕ | 2022 |
|
RU2836245C2 |
| НАЦЕЛЕННОЕ НА GPC3 ЛЕКАРСТВЕННОЕ СРЕДСТВО, КОТОРОЕ ВВОДЯТ ПАЦИЕНТУ, ЧУВСТВИТЕЛЬНОМУ К НАЦЕЛЕННОЙ НА GPC3 ЛЕКАРСТВЕННОЙ ТЕРАПИИ | 2013 |
|
RU2707882C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ПРОТИВ СПЕЦИФИЧЕСКОГО МЕМБРАННОГО АНТИГЕНА ПРОСТАТЫ (PSMA) И СПОСОБ ИНГИБИРОВАНИЯ РОСТА КЛЕТОК, ЭКСПРЕССИРУЮЩИХ PSMA | 2006 |
|
RU2421466C2 |
























































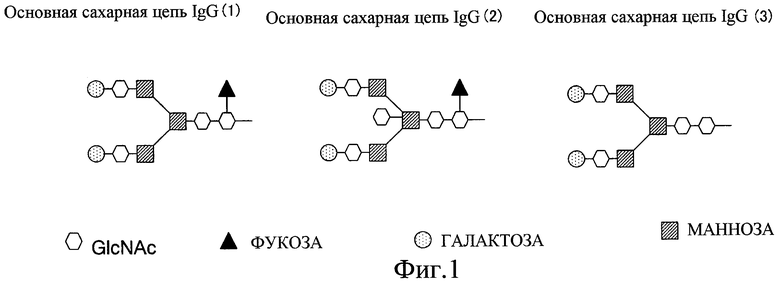
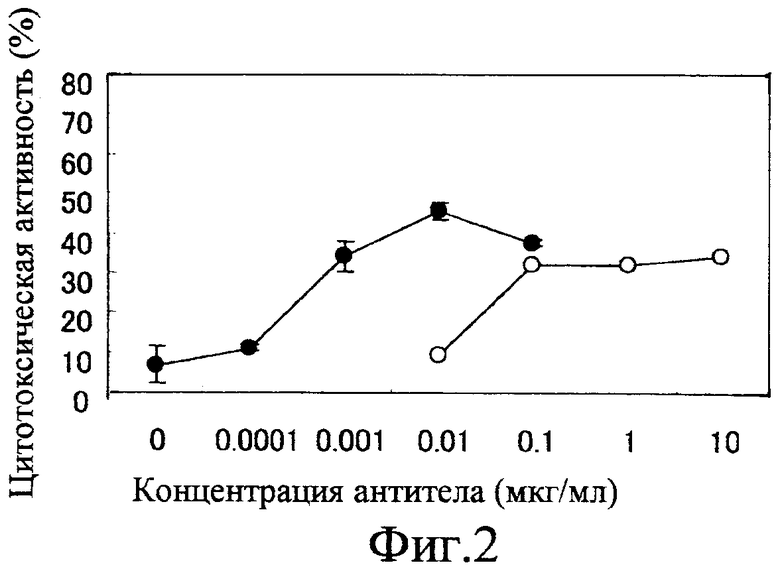
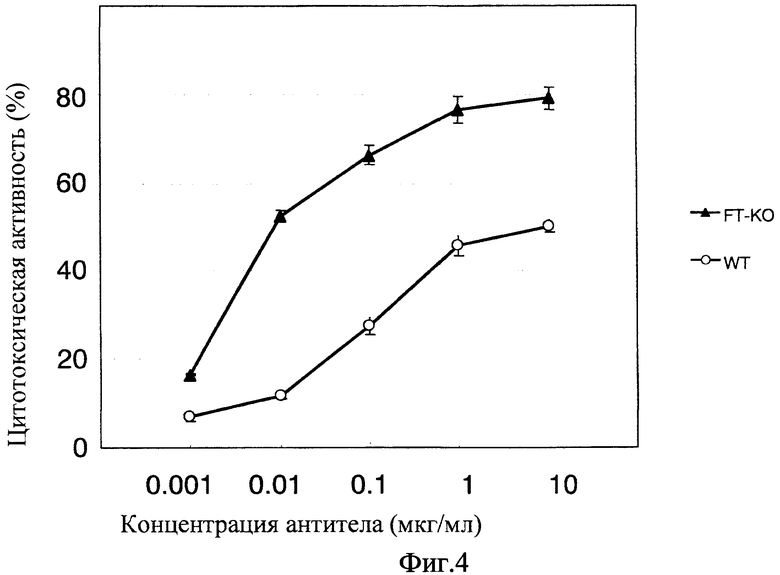
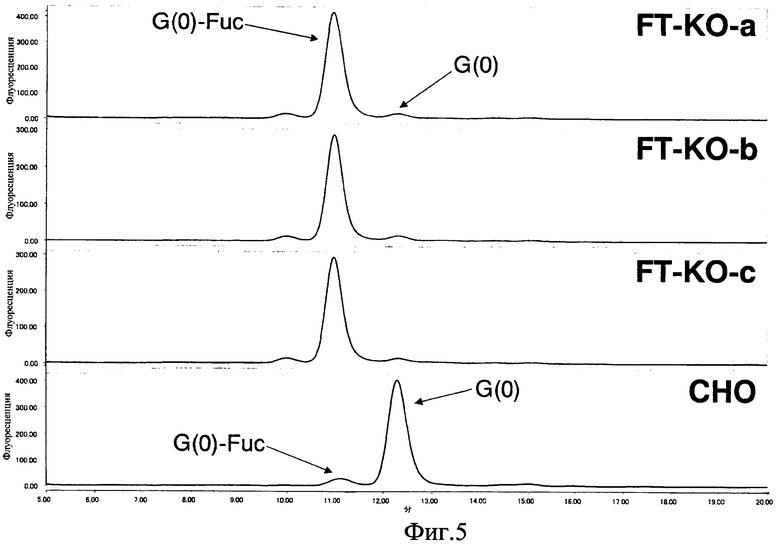
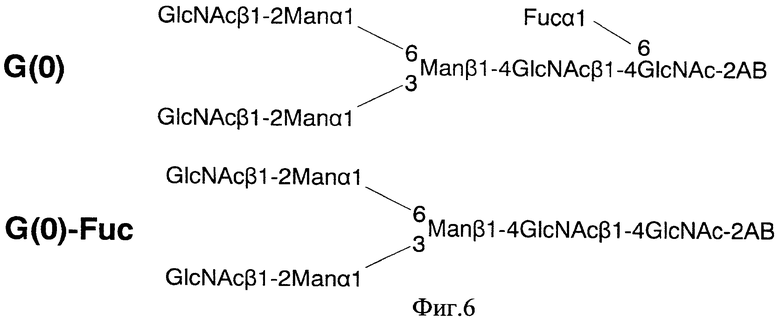

Настоящее изобретение относится к области иммунологии. Описаны варианты композиций антител класса IgGl, направленных к человеческому глипикану 3. Каждая композиция содержит в эффективном количестве смесь антител к человеческому глипикану 3. Смесь состоит из антител стандартного строения и либо более 20% антител, лишенных фукозы, либо антител, имеющих двурассекающй N-ацетилглюкозамид. Каждое из антител характеризуется определенной аминокислотной последовательностью. В одном варианте композиции антитело содержит три CDR легкой и три CDR тяжелой цепи. В другом варианте каждое из антител характеризуется наличием вариабельных областей тяжелой и легкой цепей. Описаны варианты способа получения антитела к глипикану 3 человека с использованием клетки с уменьшенной способностью присоединения фукозы к сахарным цепям, в которую введен ген, кодирующий антитело из композиции. Предложено противораковое средство на основе композиции антител. Использование изобретения обеспечивает увеличение ADCC цитотоксической активности, что может найти применение в терапии рака. 5 н. и 5 з.п. ф-лы, 7 ил., 2 табл., 9 пр.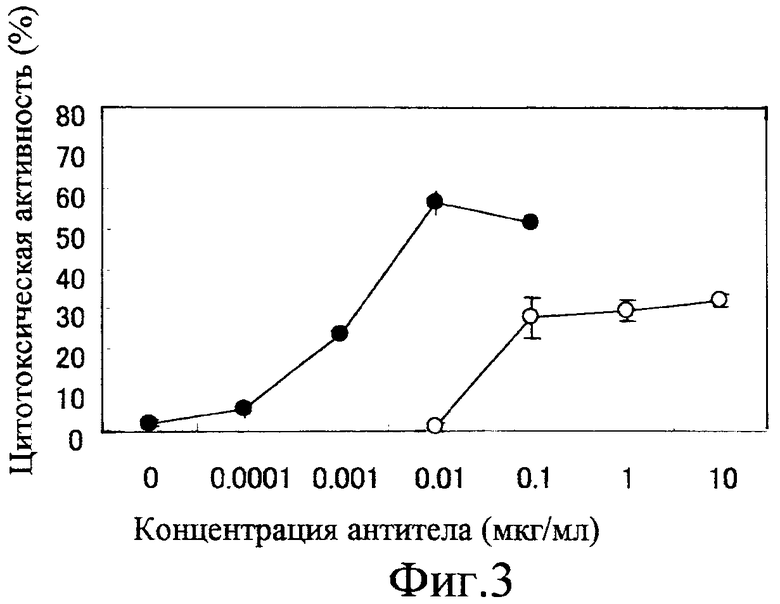
1. Композиция антител к человеческому глипикану 3, обладающая повышенной ADCC активностью в отношении опухолевых клеток, содержащая эффективное количество смеси полноразмерных антител к человеческому глипикану 3 класса IgGl, где смесь состоит из антител к глипикану 3 и модифицированного антитела к глипикану 3, выбранного из лишенного фукозы антитела к глипикану 3 или антитела к глипикану 3, имеющего двурассекающий N-ацетилглюкозамид (G1cNAc), где смесь имеет долю антител, лишенных фукозы, более чем 20%, и антитела имеют:
вариабельную область тяжелой цепи, содержащую CDR1, имеющий аминокислотную последовательность, представленную в SEQ ID No: 25, CDR2, имеющий аминокислотную последовательность, представленную в SEQ ID No: 26, и CDR3, имеющий аминокислотную последовательность, представленную в SEQ ID No: 27, и
вариабельную область легкой цепи, содержащую CDR1, имеющий аминокислотную последовательность, представленную в SEQ ID No: 45, 90-103 или 104, CDR2, имеющий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR3, имеющий аминокислотную последовательность, представленную в SEQ ID No: 60.
2. Композиция антител к человеческому глипикану 3, обладающая повышенной ADCC активностью в отношении опухолевых клеток, содержащая эффективное количество смеси полноразмерных антител к глипикану 3 класса IgGl, где смесь состоит из антитела к глипикану 3 и модифицированного антитела к глипикану 3, выбранного из лишенного фукозы антитела к глипикану 3 или антитела к глипикану 3, имеющего двурассекающий N-ацетилглюкозамид (G1cNAc), где смесь имеет долю антител, лишенных фукозы, более чем 20%, и антитела имеют одну из перечисленных (1) - (7) вариабельных областей тяжелой цепи:
(1) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID No: 81;
(2) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID No: 82;
(3) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID No: 83;
(4) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID No: 84;
(5) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID No: 85;
(6) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID No: 86;
или
(7) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID No: 87
и
вариабельную область легкой цепи, содержащую аминокислотную последовательность, представленную в SEQ ID No: 92.
3. Композиция антител к человеческому глипикану 3 по п.1, содержащая антитела, имеющие вариабельную область легкой цепи, содержащую одно из перечисленных (1)-(15) CDR 1, 2 и 3:
(1) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 90, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(2) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 91, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(3) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 92, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(4) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 93, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(5) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 94, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(6) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 95, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(7) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 96, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(8) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 97, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(9) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 98, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(10) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 99, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(11) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 100, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(12) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 101, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(13) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 102, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(14) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 103, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60;
(15) CDR 1, содержащий аминокислотную последовательность, представленную в SEQ ID No: 104, CDR 2, содержащий аминокислотную последовательность, представленную в SEQ ID No: 46, и CDR 3, содержащий аминокислотную последовательность, представленную в SEQ ID No: 60.
4. Композиция антител по п.1, где антитела содержат вариабельную область легкой цепи, выбранную из (1)-(15):
(1) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 107;
(2) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 108;
(3) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 109;
(4) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 110;
(5) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 111;
(6) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 112;
(7) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 113;
(8) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 114;
(9) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 115;
(10) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 116;
(11) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 117;
(12) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 118;
(13) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 119;
(14) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 120;
и
(15) вариабельная область легкой цепи, содержащая аминокислотную последовательность, представленную в SEQ ID No: 121.
5. Композиция антител по любому из пп.1-4, имеющая долю антител, лишенных фукозы, более чем 50%.
6. Композиция антител по любому из пп.1-4, имеющая долю антител, лишенных фукозы, более чем 90%.
7. Способ получения антитела к глипикану 3 человека с использованием клетки, где ген, кодирующий указанное антитело, которое является действующим веществом композиции, определенной в любом из пп.1-4, был введен в указанную клетку, имеющую уменьшенную способность присоединения фукозы к сахарным цепям.
8. Способ получения антитела по п.7, где клетка, имеющая уменьшенную способность присоединения фукозы к сахарным цепям, является клеткой, лишенной транспортера фукозы.
9. Способ получения антитела к глипикану 3 человека, которое является действующим веществом композиции, определенной в любом из пп.1-4, включающий стадии:
(a) введения гена, кодирующего указанное антитело, определенное в любом из пп.1-4, в клетку, имеющую уменьшенную способность присоединения фукозы к сахарным цепям; и
(b) культивирования этой клетки.
10. Противораковое лекарственное средство, имеющее в качестве активного ингредиента композицию антител по любому из пп.1-6.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| SHINKAWA et al., "The absence of fucose but not the presense of galactose or bisecting N-acetylglucosamine of human IgGl complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity", The Journal of Biological Chemistry, 2003, v.278, №5, с.3466-3473 | |||
| СРЕДСТВО, ПРИМЕНЯЕМОЕ ПРИ ЛЕЧЕНИИ ОПУХОЛЕЙ ЛИМФАТИЧЕСКОЙ ТКАНИ | 1998 |
|
RU2177805C2 |

