Область техники, к которой относится настоящее изобретение
Изобретение относится к средствам, применяемым при лечении опухолей лимфатических тканей (за исключением миеломы), включающим в качестве активного ингредиента антитела, которые специфически связываются с белками, экспрессируемыми в указанных опухолях лимфатической ткани. Настоящее изобретение относится также к средствам, применяемым при лечении опухолей Т-клеток или опухолей В-клеток (за исключением миеломы). Кроме того, настоящее изобретение относится к антителам, которые специфически связываются с белками, экспрессируемыми в опухолях лимфатических тканей, и которые обладают цитотоксической активностью.
Предпосылки создания изобретения
За иммунитет живых существ отвечают прежде всего лимфатические клетки. К лимфатическим клеткам относятся все клетки, происходящие из одних и тех же гемопоэтических стволовых клеток, которые высвобождаются в периферический кровоток после повторных стадий дифференцировки под действием различных факторов, индуцирующих дифференцировку, включая факторы, называемые иначе ростовыми факторами, имеющиеся в костном мозге или других органах. На основании различий в дифференцировке лимфоциты классифицируются на две больших группы В-клеток и Т-клеток. Считается, что В-клетки обладают способностью продуцировать антитела, тогда как Т-клетки имеют отношение к проявлению антигенности, цитотоксичности и др. свойств. При этом состояние, когда такие клетки по тем или иным причинам подвергаются изменениям, связанным с образованием опухолей, на тех или иных стадиях дифференцировки и начинают бесконтрольно пролиферировать в костном мозге, лимфатической ткани, крови или т. п. , относят к опухоли лимфатической ткани.
Благодаря использованию новой технологии, в частности тех преимуществ, которые дает метод применения моноклональных антител против поверхностных антигенов дифференциации, стало возможным идентифицировать начало и/или стадию дифференцировки лимфатических клеток. В этом случае стало возможно не только определить, из каких: Т-клеток или В-клеток, происходят такие опухолевые клетки, но также идентифицировать степень зрелости опухолевых клеток.
Опухоли лимфатической ткани на основе происхождения и степени зрелости опухолевых клеток подразделяют на два крупных класса: опухоли В-клеток и опухоли Т-клеток. В зависимости от степени зрелости опухолевых клеток опухоли В-клеток далее классифицируют на острый В-лимфолейкоз (В-ОЛЛ), хронический В-лимфолейкоз (В-ХЛЛ), пре-В-лимфому, лимфому Буркитта, фолликулярную лимфому, фолликулярную лимфому коры головного мозга, диффузную лимфому и др. С другой стороны, опухоли Т-клеток, в зависимости от степени зрелости опухолевых клеток, классифицируют на острый В-лимфолейкоз (В-ОЛЛ), хронический В-лимфолейкоз (В-ХЛЛ), заболевание, связанное с человеческим вирусом Т-клеток (АТЛ), лимфому периферических Т-клеток не АТЛ-типа (ПНТЛ) и др. (Zukai Rinso [Gan] (Illustrated Clinical: Cancer), series 17 Leukemia and lymphoma, Takashi Sugimura et al. , Medical View Co. , Ltd. , 1987, В cell tumors, Kiyoshi Takatsuki, Nishimura Shoten, 1991).
Однако, несмотря на последние достижения медицинской технологии, лечение опухолей лимфатических тканей не достигло удовлетворительных результатов. Степень излечивания острого лимфолейкоза (ОЛЛ), например, все еще составляет 20% или ниже, а лимфомы на стадии развитого заболевания около 50%, при этом относительно высокая степень излечивания В-лимфомы связана с прогрессом множественной терапии. Кроме того, Т-лимфома более склонна к проникновению в другие ткани и характеризуется степенью излечивания около 30%, тогда как степень излечивания заболевания, связанного с человеческим вирусом Т-клеток (АТЛ), в настоящее время составляет 10%.
Кроме того, Гото с соавт. (Goto, A. et al. ) сообщили о получении моноклонального антитела (антитела к НМ1.24) при иммунизации мышей миеломными клетками человека (Blood, 1994, 84, 1922-1930). При введении мыши с трансплантированными клетками миеломы человека антитела к НМ1.24, антитело специфически накапливается в опухолевых тканях (Masaakai Kosaka et al. , Nippon Rinsho (Japan Clinical) 1995, 53, 627-635), что дает основание предположить возможность применения антитела к НМ1.24 для диагностики локализации опухоли при радиоизотопном мечении, при терапии частицами, такой как радиоиммунотерапия и др. Однако не было известно данных о том, что антитело к НМ1.24 используется для лечения опухолей лимфатических тканей.
Сущность изобретения
Применяемый в настоящее время метод лечения опухолей лимфатических тканей включает различные виды химиотерапии, рентгенотерапию, трансплантацию костного мозга и др. Однако, как отмечалось выше, ни один из этих методов не дает удовлетворительных результатов при лечении указанных заболеваний и, в этой связи, все еще имеется потребность в прорыве по созданию лекарственных средств и разработке методов, которые могли бы облегчить течение опухолей лимфатических тканей и удлинить продолжительность жизни пациентов.
В связи с вышесказанным целью настоящего изобретения является разработка лекарственного средства для применения в случае опухолей лимфатических тканей, за исключением миеломы.
Для создания такого лекарственного средства авторы изобретения провели обширные исследования in vitro, которые включали анализ методом проточной цитометрии, определение цитотоксической активности, такой как антителообусловленной клеточнозависимой цитотоксичности (ADCC/АОКЦ активности), комплементзависимой цитотоксичности (CDC/КЗЦ цитотоксичности) и др. , исследования in vivo по определению противоопухолевого эффекта с использованием антитела к НМ1.24 (Goto et al. Blood, 1994, 84, 1922-1930), а также работы по выделению белкового антигена, с которым специфически связывается антитело к НМ1.24. В результате проведенных исследований авторы показали, что на лимфатических опухолях экспрессируется белковый антиген, специфически распознаваемый антителом к НМ1.24, при этом указанное антитело к НМ1.24 обладает противоопухолевым действием в отношении опухолей лимфатических тканей, что и составило предмет настоящего изобретения.
В связи с этим, настоящее изобретение относится к лекарственному средству, применяемому при опухолях лимфатических тканей (за исключением миеломы), которое включает в качестве активного ингредиента, антитело, которое специфически связывается с белком, имеющим аминокислотную последовательность, указанную в последовательности SEQ ID NO: 1, и которое обладает цитотоксической активностью (перечни последовательностей приведены в конце описания).
Настоящее изобретение также относится к лекарственному средству, применяемому при опухолях Т-клеток, или к лекарственному средству, применяемому при опухолях В-клеток (за исключением миеломы), включающему в качестве активного ингредиента антитело, которое специфически связывается с белком, имеющим аминокислотную последовательность, указанную в последовательности SEQ ID NO: 1, и которое обладает цитотоксической активностью.
Настоящее изобретение также относится к лекарственному средству, применяемому при опухолях Т-клеток, или к лекарственному средству, применяемому при опухолях В-клеток (за исключением миеломы), включающему в качестве активного ингредиента моноклональное антитело, которое специфически связывается с белком, имеющим аминокислотную последовательность, указанную в последовательности SEQ ID NO: 1, и которое обладает цитотоксической активностью.
Настоящее изобретение также относится к лекарственному средству, применяемому при опухолях Т-клеток, или к лекарственному средству, применяемому при опухолях В-клеток (за исключением миеломы), включающему в качестве активного ингредиента антитело, которое специфически связывается с белком, имеющим аминокислотную, последовательность, указанную в последовательности SEQ ID NO: 1, и которое обладает АОКЦ или КЗЦ активностью в качестве цитотоксической активности.
Настоящее изобретение также относится к лекарственному средству, применяемому при опухолях Т-клеток, или к лекарственному средству, применяемому при опухолях В-клеток (за исключением миеломы), включающему в качестве активного ингредиента антитело, которое специфически связывается с белком, имеющим аминокислотную последовательность, указанную в последовательности SEQ ID NO: 1, и которое в качестве константной области содержит Сγ человеческого антитела.
Настоящее изобретение также относится к лекарственному средству, применяемому при опухолях Т-клеток, или к лекарственному средству, применяемому при опухолях В-клеток (за исключением миеломы), включающему в качестве активного ингредиента химерное антитело или гуманизированное антитело, которое специфически связывается с белком, имеющим аминокислотную последовательность, указанную в последовательности SEQ ID NO: 1, и которое обладает цитотоксической активностью.
Настоящее изобретение также относится к лекарственному средству, применяемому при опухолях Т-клеток, или к лекарственному средству, применяемому при опухолях В-клеток (за исключением миеломы), включающему в качестве активного ингредиента антитело, которое специфически связывается с эпитопом, распознаваемым антителом к НМ1.24.
Настоящее изобретение также относится к лекарственному средству, применяемому при опухолях Т-клеток, или к лекарственному средству, применяемому при опухолях В-клеток (за исключением миеломы), включающему в качестве активного ингредиента антитело к НМ1.24.
Кроме того, настоящее изобретение относится к антителу, которое специфически связывается с белком, экспрессируемым в опухолях лимфатических тканей, и которое обладает цитотоксической активностью.
Краткое описание чертежей
На фиг. 1 показана гистограмма, полученная в результате анализа методом проточной цитометрии указанной В-клеточной линии непрямым методом с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 2 показана гистограмма указанной В-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 3 показана гистограмма указанной В-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 4 показана гистограмма указанной В-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 5 показана гистограмма указанной В-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 6 показана гистограмма указанной В-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 7 показана гистограмма указанной В-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 8 показана гистограмма указанной В-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 9 показана гистограмма указанной В-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 10 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 11 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 12 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 13 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 14 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 15 показана гистограмма указанной Т-клеточной линии, которая была исследована, непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 16 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 17 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 18 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 19 показана гистограмма указанной Т-клеточной линии, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 20 показана гистограмма указанной клеточной линии, не относящейся ни к Т-, ни к В-типу, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 21 показана гистограмма указанной клеточной линии, не относящейся ни к Т-, ни к В-типу, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 22 показана гистограмма указанной клеточной линии, не относящейся ни к Т-, ни к В-типу, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 23 показана гистограмма указанной клеточной линии, не относящейся ни к Т-, ни к В-типу, которая была исследована непрямым методом по способу проточной цитометрии с использованием антитела к НМ1.24 и контрольного мышиного IgG2a.
На фиг. 24 приведен график, показывающий, что антитело к НМ1.24 оказывает цитотоксический эффект на клеточные линии опухолей Т-клеток CCRF-CEM, CCRF-HSB-2 и HPB-MLT зависимым от дозы образом.
На фиг. 25 приведен график, показывающий, что антитело к НМ1.24 оказывает цитотоксический эффект на клеточные линии опухолей В-клеток ЕВ-3, МС116 и CCRF-SB зависимым от дозы образом.
На фиг. 26 приведен график, показывающий, что при введении антитела к НМ1.24 опытной группе мышей с трансплантированной лимфатической опухолью человека, наблюдается подавление увеличения объема опухоли в сравнении с контрольной группой животных, которым вводили IgG2a.
На фиг. 27 приведен график, показывающий, что при введении антитела к НМ1.24 опытной группе мышей с трансплантированной лимфатической опухолью человека, происходит удлинение периода выживания в сравнении с контрольной группой животных, которым вводили IgG2a.
Варианты реализации изобретения
1. Получение антител
1-1. Получение гибридомы
Гибридомы, которые продуцируют антитела, используемые по настоящему изобретению, могут быть сконструированы, в основном, с использованием известных процедур, проведенных ниже. Так, белковый антиген НМ1.24 или клетки, которые экспрессируют антиген НМ1.24, могут использоваться в качестве сенсибилизирующего антигена и далее применяться для иммунизации в рамках традиционного метода иммунизации. Полученные иммунные клетки сливают с известными родительскими клетками в ходе обычного процесса слияния клеток и затем проводят скрининг с помощью традиционного метода скрининга для отбора клеток, которые продуцируют моноклональные антитела.
Конкретно, моноклональные антитела могут быть получены следующим способом. Так, например, в качестве клеток, экспрессирующих антиген НМ1.24, который представляет собой сенсибилизирующий антиген для получения антитела, может использоваться множественная миеломная клеточная линия человека КРММ2 (нерассмотренная заявка на патент Японии (Kokai) 7-236475) или КРС-32 (Goto Т. et. al. , Jpn. J. Clin. Hematol. , 1991, 32, 1400). Альтернативно, в качестве сенсибилизирующего антигена может быть использован белок, имеющий аминокислотную последовательность, указанную ниже в SEQ ID NO: 1, или пентид, или полипептед, содержащие эпитоп, распознаваемый антителом против НМ1.24.
В рамках настоящего описания кДНК, которая кодирует белок, имеющий аминокислотную последовательность, указанную в SEQ ID NO: 1, была вставлена в сайт расщепления XbaI вектора pUC19 с получением плазмиды pRS38-pUC19. E. coli, несущая эту плазмиду, депонирована в Международной Коллекции в соответствии с положением Будапештского Договора как Escherichia coli DH5α (pRS38-pUC19) 5 октября 1993 года Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония) как FERM BP-4434 (см. нерассмотренную заявку на патент Японии (Kokai) 7-196694). Фрагмент кДНК, содержащийся в плазмиде pRS38-pUC19, может быть использован для получения пептида или полипептида, содержащих эпитоп, распознаваемый антителом против НМ1.24, с использованием методов генной инженерии.
Предпочтительно, млекопитающие, которые иммунизируют сенсибилизирующим антигеном, отбирают с учетом совместимости их с родительской клеткой в процедуре клеточного слияния. В основном они включают, не ограничиваясь ими, грызунов, таких как мыши, крысы, хомячки и др.
Иммунизация животных сенсибилизирующим антигеном проводится с использованием известного метода. Общий метод включает, например, внутрибрюшинное или подкожное введение сенсибилизирующего антигена млекопитающему. Специфически, сенсибилизирующий антиген, который был разбавлен и суспендирован в соответствующем количестве фосфатно-буферного раствора (ФБР) или физиологического раствора и др. , смешивают, по желанию, с соответствующим количеством адъюванта Фрейнда. После эмульгирования его предпочтительно вводят млекопитающему несколько раз, каждые 4-21 день. Альтернативно может использоваться подходящий носитель для введения его в момент иммунизации сенсибилизирующим антигеном.
После проведения иммунизации и подтверждения повышения уровня нужного антитела в сыворотке иммунные клетки отбирают из организма млекопитающего и проводят клеточное слияние, в котором предпочтительные иммунные клетки включают, в частности, клетки селезенки.
Миеломные клетки млекопитающих, как пример других родительских клеток, которые подвергают процедуре клеточного слияния с указанными иммунными клетками, включают предпочтительно другие различные клеточные линии, такие как Р3Х63Аg8.653 (J. Immunol. 1979, 123: 1548-1550), P3X63Ag8U. l (Current Topics in Microbiology and Immunology, 1978, 81: 1-7), NS-1 (Kohler, G. and Milstein, C. , Eur. J. Immunol. , 1976, 6: 511-519), MPC-11 (Margulies, D. H. et. al. , Cell, 1976, 8: 405-415), SP2/0 (Shulman, M. et al. , Nature, 1978, 276: 269-270), FO (de St. Groth, S. F. et. al. , J. Immunol. Methods, 1980, 35: I-21), S194 (Trowbridge, I. S. , J. Exp. Med. , 1978, 148: 313-323), R210 (Galfre, G. et. al. , Nature, 1979, 277: 131-133) и др.
Процедура клеточного слияния между указанными иммунными клетками и миеломными клетками может быть проведена, по существу, в соответствии с известным методом, таким как описано Мильштейном с соавт. (Kohler, G. and Milstein, С. , Methods Enzymol. , 1981, 73: 3-46) и др.
Более конкретно указанная процедура слияния клеток проводится в обычном питательном бульоне в присутствии, например, ускорителя слияния клеток. В качестве ускорителя слияния клеток используют, например, полиэтиленгликоль (ПЭГ), Сендаи вирус (HVJ) и, кроме того, может быть, по желанию, добавлен адъювант, такой как диметилсульфоксид и др. , для повышения активности процесса слияния.
Предпочтительно, соотношение использованных иммунных клеток и миеломных клеток соответствует, например, 1-10-кратному количеству иммунных клеток в сравнении с миеломными. Примером культуральных сред, которые могут использоваться для осуществления вышеуказанного процесса слияния клеток является среда RPMI1640 и культуральная среда MEM, пригодные для роста указанных миеломных клеточных линий, и традиционная культуральная среда, которые используются для культивирования указанного типа клеток, кроме того, в них могут быть внесены сывороточные добавки, такие как околоплодная сыворотка теленка (OCT, FSC).
В процессе слияния клеток заданное количество указанных иммунных клеток и миеломных клеток тщательно перемешивают в культуральной среде, в которую добавляют раствор ПЭГ, предварительно нагретый примерно до 37oС, например раствор ПЭГ со средним молекулярным весом от примерно 1000 до 6000, в концентрации от 30 до 60% (вес/объем) и перемешивают с получением нужных клеток слияния (гибридом). Затем, посредством повторного последовательного добавления подходящей культуральной среды и центрифугирования с удалением супернатанта, агенты, необходимые для слияния клеток и др. , которые нежелательны для роста гибридом, могут быть удалены.
Селекцию указанных гибридом проводят при культивировании в традиционной селекционной среде, например в культуральной среде HAT (жидкая культура, содержащая гипоксантин, аминоптерин и тимидин). Культивирование в указанной культуральной среде продолжают, в основном, в течение времени, достаточного для гибели клеток, отличных от желательных гибридом (которые не являются клетками слияния), и, как правило, это составляет от нескольких дней до нескольких недель. Обычно применяют традиционный метод серийных разведений, в рамках которого гибридомы, которые продуцируют желательное антитело, отбирают и подвергают моноклональному клонированию.
Кроме получения указанной гибридомы посредством иммунизации антигеном животного, отличного от человека, можно также сенсибилизировать человеческие лимфоциты in vitro антигеном НМ1.24 или клеткам, экспрессирующими антитен НМ1.24, а затем полученные сенсибилизированные лимфоциты сливают с миеломными клетками человека, например U266, с получением желательного человеческого антитела, обладающего активностью по связыванию с антигеном НМ1.24 или клетками, экспрессирующими антиген НМ1.24 (см. публикацию по заявке на патент Японии (Kokoku) 1-59878). Кроме того, трансгенное животное, несущее спектр всех генов антител человека, иммунизируют антигеном, например антигеном НМ1.24, или клетками, эспрессирующими антиген НМ1.24, с получением желательного гуманизированного антитела по описанному выше методу (см. заявки на Международный патент WO 93/12227, WO 92/03918, WO 94/02602, WO 94/25585, WO 96/34096 и WO 96/33735).
Сконструированные таким образом гибридомы, продуцирующие моноклональное антитело, могут быть подвергнуты субкультивированию в традиционной культуральной среде или могут храниться в течение длительного периода времени в жидком азоте.
Для получения моноклонального антитела от указанной гибридомы используют метод, в рамках которого указанную гибридому культивируют традиционным способом и получают антитела в супернатанте, или метод, в соответствии с которым гибридому вводят для последующего роста в организм млекопитающего, совместимого с указанной гибридомой, и затем антитела получают из асцитов. Первый способ подходит для получения антител высокой степени чистоты, тогда как последний пригоден для крупномасштабного процесса получения антител.
В частности, гибридома, продуцирующая антитело против НМ1.24, может быть получена с использованием метода Гото (Goto, Т. , et. al. , Blood, 1994, 84: 1922-1930). Она также может быть получена с помощью метода, в рамках которого гибридому, продуцирующую антитело против НМ1.24, которая была депонирована в Международной Коллекции в соответствии с положениями Будапештского Договора как FERM ВР-5233 14 сентября 1995 года Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония) инъецируют внутрибрюшинно мышам линии BALB/C (производство CLEA, Япония) с получением асцитов, из которых выделяют и очищают антитела против НМ1.24, или используется метод, в соответствии с которым указанную гибридому культивируют в подходящей культуральной среде, такой как RPMI1640, содержащей 10% околоплодной сыворотки и 5% BM-Condimed H1 (производство Boehringer Mannheim), среде SFM для выращивания гибридом (производство GIBCO-BRL), среде PFHM-II (производство GIBCO-BRL) и др. , и в этом случае антитело против НМ1.24 может быть выделено и очищено из супернатанта.
1-2. Рекомбинантное антитело
Рекомбинантное антитело, получаемое в соответствии с технологией манипулирования рекомбинантными генами, в рамках которой ген для антитела клонируют из гибридомы и интегрируют в соответствующий вектор, который затем вводят в организм хозяина, может использоваться по настоящему изобретению как моноклональное антитело (см. , например, Carl, А. К. , Вorrebaeck, James, W. Larrick, Therapeutic Monoclonal Antibodies, опубликованная в Великобритании, Macmillan Publishers LTD. , 1990).
В частности, мРНК, кодирующую вариабельную область (V-область) желательного антитела, выделяют из гибридомы, продуцирующей антитело. Для выделения мРНК получают общую РНК с использованием, например, известного метода, такого как метод ультрацентрифугирования в гуанидине (Chirgwin, J. M et al. , Biochemistry, 1979, 18: 5294-5299), the AGPC method (Chomczynski, P. et al. , Analytical Biochemistry, 1987, 162: 156-159) и затем выделяют мРНК из общей массы РНК с использованием набора для очистки мРНК (производство Pharmacia) и др. Альтернативно, мРНК можно непосредственно получить с использованием набора Quick Prep для очистки мРНК производства Pharmacia.
кДНК к V-области антитела может быть синтезирована из полученной указанным способом мРНК с использованием обратной траскриптазы. кДНК может быть синтезирована с использованием включающего обратную транскриптазу набора для синтеза кДНК AMV Reverse Transcriptase First-strand cDNA Synthesis Kit и др. Альтернативно, для синтеза и амплификации кДНК может быть использован набор 5'-Ampli Finder Race Kit (производство Clontech) и метод 5'-Race (Frohman, M. A. et al. , Proc. Natl. Acad. Sci. USA, 1988, 85: 8998-9002; Belyavsky, A. et al. , Nucleic Acids Res. , 1989, 17: 2919-2932), в рамках которой используется полимеразная цепная реакция (ПЦР). Желательный фрагмент ДНК очищают из полученного продукта ПЦР и далее он может быть лигирован с векторной ДНК. Кроме того, на его основе конструируют рекомбинантный вектор и вводят его в E. coli и далее до отбора колоний для получения желательного рекомбинантного вектора. Нуклеотидная последовательность заданной ДНК может быть подтверждена с помощью известного метода, такого как дидезоксиметод.
При получении ДНК, кодирующей V-область желательного антитела, ее можно лигировать с ДНК, кодирующей константную область (С-область) желательного антитела, и затем полученный продукт интегрируют в вектор экспрессии. Альтернативно, ДНК, кодирующая V-область антитела, может быть интегрирована в вектор экспрессии, который уже содержит ДНК, кодирующую С-область антитела.
Для продуцирования антитела, применяемого по настоящему изобретению, ген антитела интегрируют, как будет описано ниже, в вектор экспрессии так, чтобы экспрессия проходила под контролем регуляторной области экспрессии, например энхансера и/или промотора. Впоследствии вектор экспрессии может быть принесен трансформацией в клетку хозяина, где антитело может экспрессироваться.
1-3. Измененное антитело
В соответствии с настоящим изобретением искусственно измененное рекомбинантное антитело, такое как химерное антитело и гуманизированное антитело, может использоваться для цели снижения гетерологичной антигенности в отношении человека. Такие измененные антитела могут быть получены с использованием известных методов.
Химерное антитело может быть получено при лигировании полученной описанным выше способом ДНК, кодирующей V-область антитела, с ДНК, кодирующей С-область антитела человека, и затем полученный продукт встраивают в вектор экспрессии и вводят в организм хозяина для продуцирования в нем антитела (см. заявку на Европейский патент ЕР 125023 и заявку на Международный патент WO 96/02576). Химерное антитело, применяемое по настоящему изобретению, может быть получено с использованием известного метода.
Например, E. coli, несущая плазмиду, которая содержит ДНК, кодирующую L-цепь V-области или Н-цепь V-области химерного антитела против НМ1.24, была депонирована в Международной Коллекции в соответствии с положениями Будапештского Договора как Escherichia coil DH5α (pUC19-1.24L-gk) и Escherichia coil DH5α (pUC19-1.24H-gγl), соответственно, 29 августа 1996 года Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония) как FERM BP-5646 и FERM BP-5644 соответственно (см. заявку на патент Японии 9-271536).
Гуманизированное антитело, которое также называют восстановленным человеческим антителом, получают при трансплантации гипервариабельного участка (ГВУ) антитела млекопитающего, отличного от человека, например антитела мыши, в ГВУ человеческого антитела. В целом, технология, основанная на получении рекомбинантной ДНК, для продуцирования таких антител также известна (см. заявку на Европейский патент ЕР 125023 и заявку на Международный патент WO 96/02576).
Конкретно, методом ПЦР синтезируют последовательность ДНК, которая должна подходить для лигирования ГВУ мышиного антитела с каркасной областью (КО) человеческого антитела с использованием нескольких раздельных олигонуклеотидов, несущих на концах разрезы, перекрывающиеся один с другим. Полученную таким образом ДНК лигируют с ДНК, кодирующей С-область человеческого антитела, и затем встраивают полученный продукт в вектор экспрессии, который затем вводят в организм хозяина для продуцирования антител (см. заявку на Европейский патент ЕР 239400 и заявку на Международный патент WO 96/02576).
Отбирают КО человеческих антител, сшитые через ГВУ так, чтобы участки, определяющие комплементарность, образовывали сайт, благоприятный для связывания с антигеном. При желании, аминокислоты в каркасных областях вариабельной области антитела могут быть замещены так, чтобы гипервариабельный участок восстановленного человеческого антитела мог образовывать подходящий сайт для связывания с антигеном (Sato, К. et al. , Cancer Res. , 1993, 53: 851-856).
Например, E. coli, несущая плазмиду, которая содержит ДНК, кодирующую вариант a (SEQ ID NO: 2) L-цепи V-области и вариант r (SEQ ID NO: 2) L-цепи V-области гуманизированного антитела против НМ1.24, была депонирована в Международной Коллекции в соответствии с положениями Будапештского Договора как Escherichia coli DH5α (pUC19-RVLa-AHM-gk) и Escherichia coli DH5α (рUС19-РVНr-AНМ-gyl) соответственно 29 августа 1996 года Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония) как FERM BP-5645 и FERM BP-5643, соответственно (см. заявку на патент Японии 9-271536). Кроме того, E. coli, несущая плазмиду, которая содержит ДНК, кодирующую вариант s (SEQ ID NO: 4) Н-цепи V-области гуманизированного антитела против НМ1.24, была депонирована в Международной Коллекции в соответствии с положениями Будапештского Договора как Escherichia coli DH5α (pUC19-RVHs-AHM-gyl) 29 сентября 1997 года Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония) как FERM BP-6127 (заявка на патент Японии 9-271536).
В случае химерного или гуманизированного антитела используют С-область человеческого антитела и наиболее предпочтительно в качестве константной области человеческого антитела может быть использована Сγ, такая как Сγ1, Сγ2, Сγ3 и Сγ4. Среди них антитела, содержащие Сγ1 и Сγ3, обладают мощной цитотоксической активностью, т. е. АОКЦ и КЗЦ активностью, и в этой связи они предпочтительно используются по настоящему изобретению.
Химерное антитело включает вариабельную область антитела, происходящего из млекопитающего, отличного от человека, и С-область, происходящую из человеческого антитела, при этом гуманизированное антитело включает гипервариабельные участки антитела, происходящего из млекопитающего, отличного от человека, каркасные области (КО) и С-область антитела, происходящие из человеческого антитела. В соответствии с этим их антигенность в организме человека снижается, так что они могут быть использованы в качестве активного ингредиента лекарственных средств по настоящему изобретению.
Предпочтительный вариант гуманизированного антитела для использования по настоящему изобретению включает гуманизированное антитело против НМ1.24 (см. заявку на патент Японии 9-271536). Предпочтительный вариант L-цепи V-области гуманизированного антитела против НМ1.24 включает тот, в котором имеется аминокислотная последовательность, кодируемая нуклеотидной последовательностью, указанной в SEQ ID NO: 2. Предпочтительный вариант Н-цепи V-области гуманизированного антитела против НМ1.24 включает тот из них, который имеет аминокислотную последовательность, кодируемую последовательностью оснований, указанной в SEQ ID NO: 3 или 4.
1-4. Экспрессия и продуцирование
Гены для антител, сконструированных как указано выше, могут экспрессироваться, в результате чего с использованием известного метода может быть получено антитело. В случае использования клеток млекопитающих экспрессия может проводиться с использованием вектора экспрессии, содержащего обычно применяемый промотор, ген антитела, который должен экспрессироваться, и ДНК, в которой поли А сигнал был оперативно сшит на 3' конце по направлению репликации, или вектор, содержащий указанную ДНК. Примеры промотора/энхансера включают ранний промотор/энхансер цитомегаловируса человека.
Кроме того, в качестве промотора-энхансера, которые могут использоваться для экспрессии антитела по настоящему изобретению, могут найти применение вирусные промоторы/энхансеры, такие как промоторы/энхансеры ретровируса, вируса полиомы, аденовируса и вакуолизирующего обезьяньего вируса 40 (SV40), а также промоторы/энхансеры, полученные из клеток млекопитающих, такие как человеческий фактор элонгации 1α (HEF1α).
Так например, экспрессия может быть легко осуществлена по методу Миллигана с соавт. (Milligan et al. , Nature, 1979, 277, 108) в рамках которого используется промотор/энхансер SV40, или по методу Мицушима с соавт. (Mizushima et al. , Nucleic Acids Res. , 1990, 18, 5322), в котором используется промотор/энхансер HEF1α.
В случае E. coli экспрессия может быть осуществлена при оперативном связывании обычно используемого промотора, сигнальной последовательности для секреции антитела и гена антитела, который подлежит экспрессии, с последующей его экспрессией. В качестве промотора можно, например, упомянуть промотор lacz и промотор аrаВ. В случае использования промотора lacz можно использовать метод Варда с соавт. (Ward et al. , Nature, 1998, 341, 544-546; Faseb J. 1992, 6, 2422-2427), а в случае использования промотора аrаВ может найти применение метод Беттера с соавт. (Better et al. , Science, 1988, 240, 1041-1043).
В качестве сигнальной последовательности для секреции антитела при продуцировании его в периплазме Е. соli может использоваться сигнальная последовательность pelB (Lei, S. P. et al. , J. Bacteriol. , 1987, 169, 4379). После отделения продуцированного в периплазме антитела структура антитела подвергается складыванию перед использованием (см. , например, WO 96/30394).
В качестве сайта начала репликации могут быть использованы такие участки, которые получают из SV40, вируса полиомы, аденовируса, вируса, бычьей папилломы (BPV) и др. Кроме того, для амплификации генных копий в хозяйской клеточной системе вектор экспрессии может включать в качестве селектируемого маркера ген аминогликозидтрансферазы (АРН), ген тимидинкиназы (ТК) и ген ксантингуанинфосфорибозилтрансферазы Е. соli (Ecogpt), ген дигидрофолатредуктазы (dhfr) и др.
Для продуцирования антитела, используемого по настоящему изобретению, можно применять любую систему продуцирования. Система продуцирования антитела может основываться на системе продуцирования in vitro и in vivo. В качестве системы продуцирования in vitro может быть упомянута система продуцирования, в которой используются эукариотические клетки, и система продуцирования, в которой используются прокариотические клетки.
Системы продуцирования на основе эукариотических клеток включают клетки животных, растительные клетки и грибные клетки. Известные клетки животных включают (1) клетки млекопитающих, такие как клетки СНО, клетки COS, миеломные клетки, почечные клетки детенышей хомячков (ВНК), HeLa клетки и Vero клетки, (2) клетки земноводных, такие как ооциты Xenopus или (3) клетки насекомых, такие как sf9, sf21 и Тn5. Известные растительные клетки включают, например, те из них, которые происходят из рода Nicotiana и более конкретно клетки, происходящие из Nicotiana tabacum, которые образуют каллусные культуры. Известные грибные культуры включают дрожжи, такие как представители рода Saccharomyces, более конкретно Saccharomyces cereviceae или нитевидные грибы, такие как представители рода Aspergillus и более конкретно Aspergillus niger.
Системы продуцирования на основе прокариотических клеток включают бактериальные клетки. Известные бактериальные клетки включают Escherichia coli (E. coli) и Bacillus subtilis.
Антитело может быть получено при введении через процесс трансформации гена желательного антитела в эти клетки и последующем культивировании трансформированных клеток in vitro. Культивирование осуществляют известными методами. Так например, могут использоваться культуральные среды, такие как DMEM, MEM, RPMI1640, в которые могут вноситься сывороточные добавки, такие как околоплодная сыворотка теленка (ОСТ). Кроме того, антитела могут продуцироваться in vivo при имплантировании клеток, в которые был введен ген антитела, в брюшной полости животного и др.
В качестве других систем продуцирования можно отметить те из них, в которых используются животные, и те из них, в которых используются растения. Системы продуцирования на основе животных организмов включают клетки млекопитающих и насекомых.
В качестве млекопитающих могут использоваться козы, свиньи, овцы, мыши и крупный рогатый скот (Vicki Glaser, Spectrum Biotechnology Application, 1993). В качестве насекомых может использоваться тутовый шелкопряд.
В случае использования растений может найти применение растение табака.
Ген антитела вводят в организм указанных животных и растений, и в таких животных и растениях продуцируются антитела. Так например, ген антитела вводят в середину гена, кодирующего белок, который продуцируется в молоке, такой как козий β-казеин, для получения слитых генов. Фрагменты ДНК, содержащие слитый ген, в который был вставлен ген антитела, инъецируют в козий эмбрион и затем эмбрион вводят в организм козы. Желательное антитело получают из молока, продуцируемого трансгенной козой, которой стала та коза, которой ввели указанный эмбрион, или ее потомство. Для того чтобы повысить количество молока, содержащего желательное антитело, продуцируемое трансгенной козой, такой трансгенной козе, в случае приемлемости, могут вводиться гормоны (Ebert K. M. et al. , Bio/Technology, 1994, 12, 699-702).
В случае использования тутового шелкопряда для инфицирования его используют бакуловирус, в который встроен ген желательного антитела, при этом желательное антитело может быть получено из жидкостей тела тутового шелкопряда (Susumu, M. et al. , Nature, 1985, 315, 592-594). А в случае использования растений табака ген желательного антитела вставляют в подходящий для растений вектор экспрессии, например рМОN 530, и затем вектор вводят в бактерию, такую как Agrobacterum tumefaciens. Указанную бактерию затем используют для инфицирования растений табака, таких как Nicotiana tabacum, с получением желательного антитела из листьев табака (Julian, К. -С. Ма et al. , Eur. J. Immunol. , 1994, 24, 131-138).
При продуцировании антитела в системах продукции in vitro и in vivo, как указано выше, ДНК, кодирующая тяжелую цепь (Н-цепь) или легкую цепь (L-цепь) антитела, может быть раздельно введена в вектор экспрессии, которыми потом одновременно трансформируют хозяйские клетки, в другом случае ДНК, кодирующие Н-цепь и L-цепь, могут быть интегрированы в едином векторе экспрессии и использованы для трансформации хозяйского организма (см. заявку на Международный патент WO 94/11523).
Антитело, продуцируемое по описанному выше методу, может быть связано с различными молекулами, такими как полиэтиленгликоль (ПЭГ), для получения с целью последующего использования модифицированного антитела. "Антитело" в контексте настоящего описания включает такие модифицированные антитела. Для того чтобы получить такое модифицированное антитело, антитело подвергают химической модификации. Такие методы уже имеются на достигнутом уровне техники.
2. Разделение и очистка антител
2-1. Разделение и очистка антитела
Антитела, продуцируемые и экспрессируемые, как было описано выше, могут быть отделены от внуренней или внешней среды клетки или выделены из организма хозяина и далее они могут быть очищены до гомогенности. Выделение и очистка антитела с целью последующего использования по настоящему изобретению могут быть осуществлены с использованием афинной хроматографии. В качестве колонки, используемой в такой афинной хроматографии, может применяться колонка с белком А и колонка с белком G. Примерами носителей, применяемых в колонке А, являются Hyper D. , Poros, Sepharose F. F. и др.
Альтернативно, без всяких ограничений могут использоваться традиционные методы выделения и очистки белков. Выделение и очистка антитела для использования его далее по настоящему изобретению могут быть проведены при связывании надлежащим способом в процессе хроматографирования, отличном от указанной выше афинной хроматографии, при фильтровании, ультрафильтровании, высаливании, диализе и др. Хроматография включает, например, ионообменную хроматографию, гидрофобную хроматографию, гель-фильтрацию и др. Такие виды хроматографии могут быть введены в систему ВЭЖХ. Альтернативно, может быть использована хроматография с обращением фаз.
2-2. Определение концентрации антитела
Концентрация антитела, полученного в указанном выше разделе 2-1, может быть определена измерением поглощения или при помощи иммуноферментного твердофазного анализа (ИФТФА) и др. Так, при использовании для измерения уровня поглощения, антитело по настоящему изобретению или образец, содержащий антитело, разбавляют соответствующим образом ФБР(-) и затем измеряют поглощение при 280 нм, после чего проводят расчет с использованием коэффициента поглощения (оптической плотности), равного 1,35, при концентрации 1 мг/мл. При использовании метода ИФТФА измерения проводят следующим образом. 100 мкл козьего против человеческого IgG (производство Bio Source) разбавляют до концентрации 1 мкг/мл в 0,1 М бикарбонатном буфере, рН 9,6, вносят на 96-гнездную микротитрационную планшету (производство Nunc) и инкубируют в течение ночи при температуре 4oС для иммобилизации антитела.
После блокирования 100 мкл каждого разбавленного соответствующим образом антитела по настоящему изобретению или образец, содержащий антитело, или 100 мкл человеческого IgG известной концентрации, взятого в качестве стандарта, добавляют в ячейку и проводят инкубирование при комнатной температуре в течение 1 часа. После промывания добавляют 100 мкл разбавленного в 5000 раз антитела против IgG человека, меченного щелочной фосфатазой (производство Bio Source), и проводят инкубацию при комнатной температуре в течение 1 часа. После промывания добавляют субстратный раствор и инкубируют и затем, после измерения поглощения при 405 нм, с использованием Microplate reader, модель 3550 (производство Bio-Rad), проводят подсчет концентрации желательного антитела.
3. Анализ методом проточной цитометрии
Реакционная способность антител, используемых по настоящему изобретению в отношении опухолевых клеток лимфатической ткани, может быть исследована методом проточной цитометрии. Используемые клетки могут представлять собой установленные клеточные линии или свежевыделенные клетки. В качестве установленных клеточных линий может использоваться, например, линия Т-клеток RPMI 8402 (АТСС CRL-1994), CCRF-CEM (АТСС CCL-119), полученная из клеток при остром лимфобластном лейкозе, HPB-ALL (FCCH1018), полученная из клеток при остром лимфолейкозе, HPB-MLT (FCCH1019), полученная из клеток при Т-лимфоме, JM (FCCH1023), полученная из клеток при остром лимфолейкозе, MOLT-4 (АТСС CRL-1582), полученный из клеток при остром лимфобластном лейкозе, Jurkat (FCCH1024), полученная из клеток при остром лимфолейкозе, CCRF-HSB-2 (АТСС CCL-120.1), полученная из клеток при остром лимфобластном лейкозе, МТ-1 (FCCH1043), полученная из клеток при заболевании, связанном с человеческим вирусом Т-клеток, КТ-3, полученная из клеток при лимфоме Леннерта (Shimizu, S. et al. , Blood, 1988, 71, 196-203) и др. ; а в качестве линии В-клеток линия (АТСС TIB-190), трансформированная вирусом ЕВ, положительная на вирус ЕВ линия В-клеток SKW 6.4 (АТСС TIB-215), МС116 (ATCC CRL-1649), полученная из клеток с В-лимфомой, CCRF-SB (ATCC CCL-120), полученная из клеток при остром лимфобластном лейкозе, линия В-клеток RPMI 6410 (FCCH6047), полученная от пациента с острой миелоцитарной лейкемией, Daudi (ATCC CCL-213), полученная из клеток при лимфоме Буркитта, ЕВ-3 (ATCC CCL-85), полученная из клеток при лимфоме Буркитта, Jijoye (ATCC CCL-87), полученная из клеток при лимфоме Буркитта, Raji (ATCC CCL-86), полученная из клеток при лимфоме Буркитта, и в качестве клеточной линии, не относящейся ни к Т-, ни к В-типу, линия HL-60 (ATCC CCL-240), полученная из клеток при острой миелоцитарной лейкемии, ТНР-1 (ATCC TIB-202), полученная из клеток при острой моноцитарной лейкемии, U-937 (ATCC CRL-1593), полученная из клеток при гистиоцитарной лимфоме, К-562 (ATCC CCL-243), полученная из клеток при хронической миелоцитарной лейкемии и др.
После промывания вышеуказанных клеток в ФБР(-) к ним добавляют 100 мкл антитела или контрольного антитела, разбавленного до концентрации 25 мкг/мл в FACS буфере ФБР(-), содержащем 2% околоплодной сыворотки теленка и 0,1% азида натрия, и затем инкубируют полученную смесь на льду в течение 30 минут. После промывания в FACS буфере добавляют 100 мкл противомышиного козьего антитела, меченного ФИТЦ (GAM, производство Becton Dickinson), в концентрации 25 мкг/мл и затем инкубируют смесь на льду в течение 30 минут. После промывания в FACS буфере клетки суспендируют в 600 мкл или 1 мл FACS буфера и измеряют интенсивность флуоресценции всех клеток с помощью прибора FACScan (производство Becton Dickinson).
На основании полученного для каждого типа клеток значения интенсивности флуоресценции вычисляется реакционная способность антитела, с точки зрения использования по настоящему изобретению, с каждым видом клеток. Так, на основании значения интенсивности флуоресценции для каждого типа клеток можно определить, экспрессируется ли антиген НМ1.24 на клетках каждого вида (позитивных или негативных) или может быть определена степень экспрессии. Данные по наличию и интенсивности экспрессии антигена НМ1.24 в опухолевых клетках лимфатической ткани приведены ниже в примере 2.2, посвященном анализу метода проточной цитометрии.
Опухолевые клетки при опухолях лимфатической ткани, которые могут стать мишенью для лечения по настоящему изобретению, экспрессируют антиген НМ1.24. Более конкретно, опухолевые клетки в опухоли лимфатической ткани предпочтительно относятся к тем клеткам, которые являются позитивными в отношении антигена НМ1.24 и в которых процент проявления указанного антигена не ниже 5%. Более конкретно, опухолевые клетки в опухолях лимфатической ткани предпочтительно представляют собой те из них, в которых позитивный процент проявления антигена НМ1.24 составляет 20% или выше.
Более конкретно, опухолевые клетки в опухолях лимфатической ткани представляют собой предпочтительно те из них, в которых позитивный процент проявления антигена НМ1.24 составляет 50% или выше. Более конкретно, опухолевые клетки в опухолях лимфатической ткани предпочтительно представляют собой те из них, в которых позитивный процент проявления антигена НМ1.24 составляет 80% или выше.
4. Цитотоксическая активность
4-1. Измерение КЗЦ активности
Антитело, применяемое по настоящему изобретению, представляет собой такое антитело, которое обладает, например, КЗЦ активностью в качестве цитотоксической активности.
КЗЦ активность лекарственного средства, применяемого при опухолях лимфатических тканей по настоящему изобретению, может быть измерена следующим образом. Вначале готовят клетки-мишени в концентрации 4•105 клеток/мл в соответствующей среде, например в среде RPMI1640, содержащей 10% околоплодной сыворотки теленка (производство Gibco-BRL). В качестве клеток-мишеней могут быть использованы CCRF-CEM (ATCC CCL-119), CCRF-HSB-2 (ATCC CCL-120.1), HPB-MLT (FCCH1019), ЕВ-3 (ATCC CCL-85), MC116 (ATCC CRL-1649), CCRF-SB (ATCC CCL-120), К-562 (ATCC CCL-243) и др. Пятьдесят мкл указанных клеток вносят на 96-гнездную микротитрационную планшету с плоским дном (производство Falcon) и планшету инкубируют в инкубаторе с подачей СО2 при температуре 37oС в течение ночи.
Затем добавляют антитело, КЗЦ активность которого предстоит измерить, инкубируют все в течение 60 минут и затем добавляют соответствующим образом разбавленный комплемент, например комплемент детеныша кролика (Baby Rabbit Complement, производство Cedarlane) и проводят инкубацию в течение 2 часов. Затем к каждой ячейке добавляют 10 мкл Alamar Bule (производство Bio Source) и проводят инкубацию в течение 4-х часов и затем измеряют интенсивность флуоресценции (длина волны возбуждения 530 нм, длина волны эмиссии 590 нм) с использованием системы измерения флуоресценции CytoFluor 2350 (производство Millipor). Цитотоксическая активность (%) может быть вычислена по формуле (А-С)/(В-С)•100, где А обозначает интенсивность флуоресценции при инкубации в присутствии антитела, В обозначает интенсивность флуоресценции при инкубации в среде, не содержащей антитело, и С представляет собой интенсивность флуоресценции ячейки, не содержащей клеток.
4-2. Измерение АОКЦ активности
Антитело, используемое по настоящему изобретению, представляет собой такое антитело, которое обладает, например, АОКЦ активностью в качестве цитотоксической активности.
АОКЦ активность лекарственного средства, используемого при лечении опухолей лимфатических тканей по настоящему изобретению, может быть измерена следующим образом. Вначале, моноядерные клетки выделяют в качестве эффекторных клеток из периферической крови человека или из костного мозга при гравитационном центрифугировании. В качестве клеток-мишеней берут CCRF-CEM (АТСС CCL-119), CCRF-HSB-2 (ATCC CCL-120.1), HPB-MLT (FCCH1019), ЕВ-3 (АТСС CCL-85), МС116 (АТСС CRL-1649), CCRF-SB (АТСС CCL-120), К-562 (АТСС CCL-243) или др. , которые метят 51Сr с получением препаратов В-клеток-мишеней. Затем к меченым клеткам-мишеням добавляют антитело, активность которого предстоит измерить, и проводят инкубацию. Эффекторные клетки в подходящем соотношении добавляют затем к клеткам-мишеням и проводят инкубацию.
После инкубации собирают супернатант и измеряют радиоактивность с помощью гамма-счетчика, при этом для измерения максимума высвобожденной радиоактивности может быть использован 1% NP-40. Цитотоксическую активность (%) вычисляют по формуле (А-С)/(В-С)•100, где А обозначает радиоактивность (имп/мин), высвобождаемую в присутствии антитела, В обозначает радиоактивность (имп/мин), высвобождаемую при наличии NP-40, и С обозначает радиоактивность (имп/мин), высвобождаемую в среде, не содержащей антитело.
4-3. Повышение цитотоксической активности
Для достижения цитотоксической активности, такой как АОКЦ активность и КЗЦ активность, предпочтительно использовать Сγ, в частности Сγ1 и Сγ3, в качестве константной области (С-области) антитела человека. Кроме того, усиленная АОКЦ активность или КЗЦ активность может быть индуцирована при добавлении, изменении или модификации части аминокислот в С-области антитела.
В качестве примера можно указать на конструирование IgM-подобного полимера IgG при замещении аминокислот (Smith, R. I. F. and Morrison, S. L. , Bio/Technology, 1994, 12, 683-688), конструирование IgM-подобного полимера IgG при добавлении аминокислот (Smith, R. I. F. et al. , J. Immunology, 1995, 154, 2226-2236), экспрессию тандемно-лигированного гена, кодирующего L-цепь (Shuford, W. , et al. , Science, 1991, 252, 724-727), димеризацию IgG при замещении аминокислот (Саron, Р. С. et al. , J. Exp. Med. , 1992, 176, 1191-1195; Shopes, В. , J. Immunology, 1992, 148, 2918-2922), димеризацию IgG при химической модификации (Wolff, E. A. et al. , Cancer Res. , 1993, 53, 2560-2565) и введение эффекторной функции при изменении аминокислот(ы) в шарнирной области антитела (Norderhaug, L. , et al. , Eur. J. Immunol. , 1991, 21, 2379-2384) и др. Все эти процедуры могут быть осуществлены с применением сайт-специфичного мутагенеза с использованием праймера, при добавлении нуклеотидной последовательности в сайте расщепления рестрикционных ферментов при использовании химических модифицирующих веществ, которые создают ковалентную связь.
5. Подтверждение терапевтического эффекта
Терапевтический эффект лекарственного средства, используемого по настоящему изобретению, при опухолях лимфатических тканей может быть подтвержден при введении антитела, используемого по настоящему изобретению, животным, которым трансплантировали клетки опухоли лимфатических тканей с последующей оценкой противоопухолевого эффекта средства на животных.
В качестве лимфатических опухолевых клеток, вводимых животному, могут использоваться установленные клеточные линии или свежевыделенные клетки. В качестве установленной клеточной линии может быть использована CCRF-CEM (ATCC CCL-119), HPB-MLT (FCCH1019), MOLT-4 (ATCC CRL-1582), CCRF-HSB-2 (ATCC CCL-120.1) и др. в качестве Т-клеточной линии и CESS (ATCC TIB-190), SKW 6.4 (ATCC TIB-215), CCRF-SB (ATCC CCL-120), RPMI 6410 (FCCH6047), EB-3 (ATCC CCL-85) и др. в качестве В-клеточной линии.
Животные, которым провели трансплантацию, предпочтительно относятся к тем, у которых иммунологические функции снижены или отсутствуют. Например, могут использоваться мыши, лишенные волосяного покрова, мыши SCID, бежевые мыши, крысы, лишенные волосяного покрова, и др. Определенный противоопухолевый эффект может быть подтвержден измерением объема и веса опухолей или на основании длительности выживания животных и др.
Как показано в приведенных ниже примерах, введение антитела против НМ1.24 приводит к подавлению роста объема опухоли и, кроме того, к увеличению периода выживаемости мышей с трансплантированной опухолью. Эти факты указывают на то, что антитело против НМ1.24 обладает противоопухолевым воздействием на опухоли лимфатической ткани.
6. Способ введения и фармацевтические препараты
Лекарственные средства для лечения опухолей лимфатических тканей по настоящему изобретению могут быть введены либо системно, либо местно, парентеральным способом, например путем внутривенной инъекции, такой как капельная инфузия, внутримышечной инъекции, внутрибрюшинной инъекции и подкожной инъекции. Метод введения может быть выбран по показанию в зависимости от возраста и состояния пациента. Эффективную дозировку выбирают в диапазоне от 0,01 мг до 100 мг на килограмм веса тела на введение. Альтернативно, может быть выбрана дозировка, в диапазоне от 1 до 1000 мг, предпочтительно от 5 до 50 мг на пациента.
Лекарственные средства для лечения опухолей лимфатической ткани по настоящему изобретению могут содержать фармацевтически приемлемые носители или добавки, в зависимости от применяемого способа введения. Примеры таких носителей или добавок включают воду, фармацевтически приемлемый органический растворитель, коллаген, поливиниловый спирт, поливинил-пирролидон, карбоксивиниловый полимер, натрий-карбоксиметил-целлюлозу, натриевую соль полиакриловой кислоты, альгинат натрия, водорастворимый декстран, натрий-карбоксиметиловый крахмал, пектин, метилцеллюлозу, этилцеллюлозу, ксантановую камедь, аравийскую камедь, казеин, желатин, агар, диглицерин, пропиленгликоль, полиэтиленгликоль, вазелин, парафин, стеариловый спирт, стеариновую кислоту, человеческий сывороточный альбумин (ЧСА), маннит, сорбит, лактозу, фармацевтически приемлемое поверхностно-активное вещество и др. Используемые добавки выбирают из указанных средств или их сочетаний, не ограничиваясь ими, в зависимости от формы применяемой дозировки.
Заболевания, подлежащие лечению по способу настоящего изобретения, представляют собой опухоли лимфатической ткани (за исключением миеломы), которые несут антиген на опухолевых клетках и с которым связывается антитело, применяемое по настоящему изобретению. В качестве конкретных заболеваний такого рода можно отметить острый В-лимфолейкоз (В-ОЛЛ), хронический В-лимфолейкоз (В-ХЛЛ), пре-В-лимфому, лимфому Буркитта, фолликулярную лимфому, фолликулярную лимфому коры головного мозга, диффузную лимфому, острый Т-лимфолейкоз (Т-ОЛЛ), хронический Т-лимфолейкоз (Т-ХЛЛ), заболевание, связанное с человеческим вирусом Т-клеток (АТЛ), периферическую Т-лимфому, не связанную с человеческим вирусом Т-клеток (ПНТЛ) и др. Лекарственные средства по настоящему изобретению используются в качестве лекарственных средств при лечении опухолей лимфатических тканей.
Примеры
Приведенные ниже примеры служат для более детального описания настоящего изобретения. Следует отметить, что настоящее изобретение не ограничивается указанными примерами никоим образом.
Пример 1. Конструирование антитела против НМ1.24
1. Получение мышиных асцитов, содержащих антитело против НМ1.24
Получают гибридомы, продуцирующие антитела против НМ1.24 в соответствии с методом Гото с соавт. (Goto, Т. et al. , Blood, 1994, 84, 1922-1930).
Мышам линии BALB/с (получаемые от фирмы CLEA, Япония), которым предварительно ввели внутрибрюшинно 500 мкл 2,6,10,14-тетраметилпентадекан (производство Wako Pure Chemical Industries, Ltd. ), за 11 и за 3 дня до эксперимента инъецируют внутрибрюшинно 5•106 гибридомных клеток. Начиная с 10 дня после инъекции гибридомных клеток, собирают асциты, которые накопились в брюшной полости мыши, с помощью постоянной иглы 19 размера Наррусаз (производство Medikit). Собранные асциты центрифугируют дважды со скоростью 1000 и 3000 об/мин с использованием низкоскоростной центрифуги RLX-131 (производство Tomy Seiko) для удаления гибридом и контаминантов, таких как клетки крови и др.
2. Очистка антитела против НМ1.24 из мышиных асцитов
Очистку антитела против НМ1.24 из указанных выше мышиных асцитов проводят следующим образом. После добавления равного количества ФБР(-) к мышиным асцитам смесь фильтруют с использованием волоконного фильтра Mediaprep (производство Millipore) и затем подвергают афинной очистке с использованием высокоскоростного прибора для очистки антител ConSep LC100 (производство Millipore) и колонки Hyper D Protein A (колонка объемом 20 мл производства Nihon Gaisi), ФБР(-) в качестве адсорбционного буфера и 0,1 М натрий-цитратного буфера (рН 4) в качестве элюирующего буфера, в соответствии с прилагаемыми инструкциями. Элюируемые фракции сразу же доводят до значения рН 7,4 при добавлении 1 М Трис-НСl (рН 8,0), и затем подвергают концентрированию и замещают буфер на ФБР(-) с использованием ультрафильтрационного концентратора Centriprep 10, с последующим стерильным фильтрованием через мембранный филар Millex-GV (производство Millipore) с размером пор 0,22 мкм с получением очищенного антитела против НМ1.24.
3. Определение концентрации антитела
Концентрацию очищенного антитела определяют при измерении поглощения. Так, очищенное антитело разбавляют в ФБР(-), измеряют поглощение при 280 нм и вычисляют концентрацию с использованием коэффициента пересчета, составляющего: 1,35 единиц оптической плотности = 1 мг/мл.
Пример 2. Изучение реакционной способности антитела против НМ1.24 с лимфатическими опухолевыми клетками
1. Очистка контрольного мышиного IgG2a.
Контрольный мышиный IgG2a очищают следующим образом. Коммерчески доступный IgG2a (KAPPA) (UpC 10) из асцитов (производство Cappel) растворяют в очищенной воде и ФБР(-). Раствор фильтруют с использованием мембранного фильтра Acrodisc (производство Gelman Sciences) с размером пор 0,2 мкм, и затем очищают афинным способом с помощью высокоскоростного прибора для очистки антител ConSep LC 100 (производство Millipore), колонки Hyper D Protein А (колонка объемом 20 мл, производство Nihon Gaisi), ФБР(-) в качестве адсорбционного буфера и 0,1 М натрий-цитратного буфера (рН 4) в качестве элюирующего буфера, в соответствии с прилагаемыми инструкциями.
Элюируемые фракции сразу же доводят до значения рН 7,4 при добавлении 1 М Трис-НСl (рН 8,0) и затем подвергают концентрированию и замещают буфером ФБР(-) с использованием центрифужного ультрафильтрационного концентратора Centriprep 10 с последующим стерильным фильтрованием через мембранный фильтр Millex GV (производство Millipore) с размером пор 0,22 мкм с получением очищенного контрольного мышиного IgG2a.
Определение концентрации контрольного мышиного IgG2a проводят в соответствии с процедурой, приведенной ранее в разделе 3. Определение концентрации антитела.
2. Анализ по методу проточной цитометрии
Реакционную способность антитела против НМ1.24 с лимфатическими опухолевыми клетками исследуют методом проточной цитометрии. После промывания в ФБР(-) Т-клеточной линии RPMI 8402 (АТСС CRL-1995), CCRF-CEM (ATCC CRL-119), полученной из клеток при остром лимфобластном лейкозе HPB-ALL (FCCH1018), полученной из клеток при остром лимфолейкозе, HPB-MLT (FCCH1019), полученной из клеток при Т-лимфоме, JM (FCCH1023), полученной из клеток при остром лимфолейкозе, MOLT-4 (АТСС CRL-1582), полученной из клеток при остром лимфобластном лейкозе, Jurkat (FCCH1024), полученной из клеток при остром лимфолейкозе, CCRF-HSB-2 (АТСС CCL-120.1), полученной из клеток при остром лимфобластном лейкозе, МТ-1 (FCCH1043), полученной из клеток при заболевании, связанном с человеческим вирусом Т-клеток, и КТ-3, полученной из клеток при лимфоме Леннерта (Shimizu, S. et al. , Blood, 1988, 71, 196-203), а в качестве В-клеточной линии клеток CESS (ATCC TIB-190), трансформированных вирусом ЕВ, положительных на ЕВ вирус В-клеток SKW 6.4 (АТСС TIB-215), MC116 (ATCC CRL-1649), полученных при В-лимфоме, CCRF-SB (ATCC CCL-120), полученной из клеток при остром лимфобластном лейкозе, В-клеток RPMI 6410 (FCCH6047), полученных от пациента с острой миелоцитарной лейкемией, Daudi (ATCC CCL-213), полученной из клеток при лимфоме Буркитта, ЕВ-3 (АТСС CCL-85), полученной из клеток при лимфоме Буркитта, Jijoye (ATCC CCL-87), полученной из клеток при лимфоме Буркитта, Raji (ATCC CCL-86), полученной из клеток при лимфоме Буркитта и в качестве клеточной линии, не относящейся ни к Т-, ни к В-типам, HL-60 (ATCC CCL-240), полученной из клеток при острой миелоцитарной лейкемии, ТНР-1 (ATCC TIB-202), полученной из клеток при острой моноцитарной лейкемии, U-937 (ATCC CRL-1593), полученной из клеток при гистиоцитарной лимфоме, и К-562 (ATCC CCL-243), полученной из клеток при хронической миелоцитарной лейкемии, добавляют 100 мкл антитела против НМ1.24 или очищенное контрольное мышиное антитело IgG2, разбавленное до концентрации 25 мкг/мл в FACS буфере (ФБР(-), содержащий 2% околоплодной сыворотки теленка и 0,1% азида натрия), после чего все инкубируют на льду в течение 30 минут.
После промывания в FACS буфере добавляют 100 мкл козьего противомышиного антитела (GAM), меченного ФИТЦ, в концентрации 25 мкг/мл и затем инкубируют смесь на льду в течение 30 минут. После промывания в FACS буфере клетки суспендируют в 600 мкл или 1 мл FACS буфера и в каждой клеточной суспензии измеряют интенсивность флуоресценции с помощью прибора FACScan (производство Becton Dickinson). Результаты, представленные на фиг. 1-23, подтверждают, что все Т-клеточные линии и все В-клеточные линии (за исключением Daudi и Raji, которые не реагируют), реагируют с антителом против НМ1.24 и в высокой степени экспрессируют антиген НМ1.24. С другой стороны, ни одна из клеточных линий, не относящихся к Т- или В-типу, не реагирует с антителом против НМ1.24 и не экспрессирует антиген.
На гистограммах, приведенных на фиг. 1-23, маркеры показывают, что при окрашивании контрольным мышиным IgG2a негативные клетки составляют 98% и позитивные клетки составляют 2%. На основании картины, полученной с указанными гистограммными маркерами, был вычислен процент позитивных по антигену НМ1.24 клеток при использовании антитела против НМ1.24, и результат представлен в таблице. По проценту позитивных по антигену НМ1.24 клеток уровень экспрессии антигена НМ1.24 подразделили на 5 уровней: -, +/-, +, ++ и +++. В результате было подтверждено, что все Т-клеточные линии и В-клеточные линии (за исключением Daudi и Raji) в высокой степени экспрессируют антиген НМ1.24, что аналогично результатам, показанным на фиг. 1-23. При этом во всех случаях использования клеточных линий, не относящихся к Т- и В-типам, процент позитивных по антигену НМ1.24 клеток был отрицательным или составлял менее 5%, что указывает на то, что экспрессия антигена отсутствует или находится на очень низком уровне.
Пример 3. Определение КЗЦ активности
КЗЦ активность антитела против НМ1.24 по отношению к лимфатическим опухолевым клеткам определяется следующим образом:
1. Получение клеток-мишеней
В качестве клеток-мишеней готовят взвесь CCRF-CEM (АТСС CCL-119), полученную из клеток при остром лимфолейкозе, CCRF-HSB-2 (АТСС CCL-120.1), полученную из клеток при остром лимфобластном лейкозе, HPB-MLT (FCCH1019), полученную из клеток при Т-лимфоме, ЕВ-3 (АТСС CCL-85), полученную из клеток при лимфоме Буркитта, МС 116 (АТСС CRL-1649), полученную из клеток при лимфоме Буркитта, CCRF-SB (АТСС CCL-120), полученную из клеток при остром лимфолейкозе, и К562 (АТСС CCL-243), полученную из клеток при хронической миелоцитарной лейкемии, в концентрации 4•105 клеток/мл в среде RPMI1640 (производство Gibco BRL), содержащей 10% околоплодной сыворотки теленка (Gibco BRL). Пятьдесят мкл каждой из этих клеточных суспензий вносят на 96-гнездную микротитрационную планшету с плоским дном (производство Falcon) и проводят инкубацию в инкубаторе с высокой влажностью в атмосфере 5% СO2 (производство Tabai) в течение ночи при 37oС.
2. Получение антитела против НМ1.24
Очищенное антитело против НМ1.24, полученное в указанном выше Примере 1, готовят в концентрациях 0, 0,2, 2 и 20 мкг/мл в среде RPMI1640, содержащей 10% околоплодной сыворотки теленка (производство Gibco BRL), и 50 мкл смеси добавляют к 96-гнездному микротитрационному планшету, подготовленному, как указано выше в Разделе 1. После инкубирования планшеты в инкубаторе с высокой влажностью в атмосфере 5% СО2 (производство Tabai) при температуре 37oС в течение 60 минут, проводят центрифугирование на низкоскоростной центрифуге 05PR-22 (производство Hitachi) при 1000 об/мин в течение 5 минут и отбирают 50 мкл супернатанта.
3. Приготовление комплемента
Комплемент детенышей кролика (Baby Rabbit) (производство Cedarlane) растворяют в очищенной воде из расчета 1 мл на ампулу, и далее разбавляют в 5 мл среды RPMI1640 (производство Gibco-BRL), не содержащей околоплодной сыворотки теленка. Пятьдесят мкл указанной смеси распределяют по 96-гнездному микротитрационному планшету с плоским дном, подготовленному, как указано выше, в Разделе 2, и проводят инкубацию в инкубаторе с высокой влажностью в атмосфере 5% СО2 (производство Tabai) в течение 2-х часов при 37oС.
4. Определение КЗЦ активности
После завершения инкубации добавляют 10 мкл Alamar Bule (производство Bio Source) к каждой ячейке 96-гнездного микротитрационного планшета с плоским дном, описанного в Разделе 3, и проводят инкубацию в инкубаторе с высокой влажностью в атмосфере 5% СО2 (производство Tabai) в течение 4-х часов при 37oС. Затем каждую ячейку промеряют на интенсивность флуоресценции (длина волны возбуждения 530 нм, длина волны эмиссии 590 нм) с использованием системы измерения флуоресценции CytoFluor 2350 (производство Millipore). Цитотоксическую активность (в процентах) вычисляют по формуле (A-C)/(В-С)•100, где А обозначает интенсивность флуоресценции при инкубировании в присутствии антитела, В обозначает интенсивность флуоресценции при инкубировании только в среде без добавления антитела, С обозначает интенсивность флуоресценции ячейки, не содержащей клеток.
Результаты показали, как видно из фиг. 24 и 25, что К562, которая не реагирует с антителом против НМ1.24 по результатам анализа методом проточной цитометрии, не продемонстрировала цитотоксичности даже при добавлении антитела против НМ1.24, тогда как CCRF-CEM, CCRF-HSB-2, HPB-MLT, ЕВ-3, МС-116 и CCRF-SB, которые взаимодействуют с антителом против НМ1.24, демонстрируют цитотоксичносгь способом, зависимым от концентрации антитела против НМ1.24. Этот результат указывает, что антитело против НМ1.24 проявляет КЗЦ активность в отношении лимфатической опухоли, несущей на клеточной поверхности белковый антиген, с которым антитело против НМ1.24 специфически связывается.
Пример 4. Противоопухолевое действие антитела против НМ1.24 в отношении мышей с трансплантированной лимфатической опухолью человека
1. Получение антитела для введения
1-1. Получение антитела против НМ1.24
Очищенное антитело против НМ1.24, полученное в приведенном выше примере 1, готовят в концентрации 1 мг/мл и 200 мкг/мл в стерильно отфильтрованном ФБР(-) и используют в следующих экспериментах.
1-2. Получение контрольного мышиного IqG2a
Очищенное антитело, полученное в указанном выше примере 2, готовят в концентрации 1 мг/мл в стерильно отфильтрованном ФБР(-) и используют в следующих экспериментах.
2. Противоопухолевый эффект антитела против НМ1.24 на мышах с трансплантированной лимфатической опухолью человека
2-1. Получение мышей с трансплантированной лимфатической опухолью человека
Мышей с трансплантированной лимфатической опухолью человека получают следующим способом. Клетки CCRF-HSB-2 (АТСС CCL-120.1), полученные при остром лимфобластном лейкозе и которые субкультивировали in vivo с использованием мышей SCID (Clea, Япония), готовят в виде взвеси в концентрации 1•108 клеток/мл в среде RPMI1640, содержащей 10% околоплодной сыворотки теленка (производство Gibco-BRL). Клеточные суспензии, полученные как указано выше, инъецируют подкожно в брюшную полость мышей SCID (6-недельные самцы), на следующий день после введения им внутрибрюшинно 100 мкл антиаксиального препарата GM1 (Waco Pure Chemical Industries).
2-2. Введение антитела
На 7 день после трансплантации опухоли измеряют циркулем диаметр опухоли, образовавшейся в том месте, куда указанным мышам трансплантировали CCRF-HSB-2 с лимфатической опухолью человека. После определения объема опухоли животных группируют так, чтобы в каждой группе среднее значение объема опухоли было примерно одинаковым (по 8 животных на группу, всего 3 группы). Начиная с одного и того же дня, животным в каждой группе вводят внутрибрюшинно 100 мкл антитела против НМ1.24 в концентрации 1 мг/мл или 200 мкг/мл, или 1 мкг/мл контрольного мышиного IgG2a, подготовленного как указано выше в Разделе 1. Введение проводят дважды в неделю, всего 19 раз аналогичным образом. В течение этого периода измеряют циркулем диаметр опухоли дважды в неделю и подсчитывают объем опухоли.
2-3. Оценка противоопухолевого эффекта антитела против НМ1.24 на мышей с трансплантированной лимфатической опухолью человека
Противоопухолевый эффект антитела против НМ1.24 оценивают по изменению объема опухоли и периода выживаемости мышей. В результате, как показано на фиг. 26, было установлено, что увеличение объема опухоли подавляется при введении животным антитела против НМ1.24 в сравнении с контрольной группой животных, которым вводили мышиный IgG2a. При этом, что также видно из Фиг. 27, при введении группе животных антитела против НМ1.24 наблюдается увеличение периода выживаемости животных в сравнении с контрольной группой животных, которым вводят мышиный IgG2a. Эти факты указывают на то, что антитело против НМ1.24 скрывают противоопухолевое действие на мышей с трансплантированной лимфатической опухолью человека.
Ссылочный пример 1. Получение гибридом, которые продуцируют мышиное моноклональное антитело против НМ1.24
По методу Гото с соавт. (Goto, Т. et al. , Blood, 1994, 84, 1922-1930) получают гибридомы, которые продуцируют мышиное моноклональное антитело против НМ1.24.
Плазменную клеточную линию КРС-32 (1•107), полученную из костного мозга пациента с множественной миеломой человека (Goto, Т. et al. , Jpn. J. Clin. Hematol. , 1991, 32, 1400) инъецируют дважды в брюшную полость мышей линии BALB/c (производство Charles River) каждые шесть недель.
За 3 дня до умерщвления животного в селезенку мыши инъецируют 1,5•106 клеток КРС-32 для усиления антителообразующей способности мыши (Goto, Т. et al. , Tokushima J. Exp. Med. , 1990, 37, 89). После умерщвления животного экстрагируют селезенку, и в экстрагированном органе проводят процедуру клеточного слияния с миеломной клеткой SP2/0 (по методу Groth, de St. and Schreidegger, Cancer Research, 1981, 41, 3465).
По методу ИФТФА на клетках (Posner, M. R. et al. , J. Immunol. Methods, 1982, 48, 23) с использованием КРС-32 проводят скрининг культурального супернатанта гибридом на наличие антитела. 5•104 клеток КРС-32 суспендируют в 50 мл ФБР, аликвоту переносят на 96-гнездную микротитрационную планшету (U-образное гранулированное дно, производство Iwaki) и затем высушивают на воздухе при температуре 37oС в течение ночи. После блокирования реакции добавлением ФБР, содержащим 1% бычьего сывороточного альбумина (БСА), вносят культуральный супернатант гибридом и проводят инкубацию при 4oС в течение 2-х часов. Затем проводят реакцию при 4oС в течение 1 часа козьего антитела против мышиного IgG, меченного пероксидазой (производство Zymed). После промывания проводят реакцию с раствором о-фенилекдиамина (производство Sumitomo Bakelite) при комнатной температуре в течение 30 минут.
Реакцию останавливают добавлением 2 н. серной кислоты и измеряют поглощение при 492 нм с использованием счетчика ИФТФД (производство Bio-Rad). Для отбора гибридом, которые продуцируют антитела против человеческого иммуноглобулина, культуральный супернатант позитивных гибридом вначале адсорбируют на человеческую сыворотку, после чего проводят скрининг реакционной способности в отношении других клеточных линий с помощью метода ИФТФА. Отбирают позитивные гибридомы и их реакционную способность к различным клеткам определяют с помощью метода проточной цитометрии. Последний отобранный гибридомный клон клонируют дважды и инъецируют в брюшную полость мышей линии BALB/c, обработанных пристаном, и далее собирают из них асциты.
Из асцитов мыши получают моноклональные антитела и очищают их осаждением сульфатом аммония и с использованием набора для афинной хроматографии белка A (Ampure PA, производство Amersham). Очищенные антитела метят ФИТЦ с использованием набора Quick Tag FITC biding kit (производство Boehringer Mannheim).
В результате исследований показано, что моноклональные антитела, продуцируемые 30 гибридомными клонами, реагируют с КРС-3 и RPMI 8226. После клонирования исследуют реакционную способность клеточного супернатанта от указанных гибридом с другими клеточными линиями или моноядерными клетками периферической крови.
Из них 3 клона продуцируют моноклональные антитела, которые специфически реагируют с плазменной клеткой. Из указанных 3-х клонов был отобран и обозначен как НМ1.24 гибридомный клон, который наиболее приемлем для анализа методом проточной цитометрии и который обладает КЗЦ активностью к RPMI 8226.
Подкласс моноклональных антител, продуцируемых указанными гибридомами, идентифицируют методом ИФТФА с использованием противомышиного антитела кролика, специфичного к данному подклассу (производство Zymed). Антитело против НМ1.24 относится к подклассу IgG2a. Гибридома НМ1.24, которая продуцирует антитело против НМ1.24, была депонирована в Международной Коллекции в соответствии с положениями Будапештского Договора как FERM ВР-5233 14 сентября 1995 года Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония).
Ссылочный пример 2. Получение гуманизированного антитела против НМ1.24
Гуманизированное антитело против НМ1.24 получают в соответствии со следующим методом.
Из гибридомы НМ1.24, полученной в ссылочном примере 1, получают общую РНК с использованием традиционного метода. На ее основе синтезируют кДНК, кодирующую V-область мышиного антитела, и амплифицируют по методу с использованием полимеразной цепной реакции (ПЦР) и по методу 5'-RACE. Получают фрагмент ДНК, содержащий ген, кодирующий мышиную V-область, который лигируют с вектором клонирования на основе плазмиды pUC и затем вводят в компетентные клетки Е. соli с получением трансформанта Е. соli. Указанную плазмиду получают из трансформанта. Определяют традиционным методом нуклеотидную последовательность кодирующего участка кДНК в плазмиде и идентифицируют гипервариабельный участок (ГВУ) в каждой V-области.
Для конструирования вектора, экспрессирующего химерное антитело против НМ1.24, в HEF вектор вставляют кДНК, кодирующую V-область каждой из L-цепи и Н-цепи мышиного антитела против НМ1.24. Далее для конструирования гуманизированного антитела против НМ1.24 ГВУ V-области мышиного антитела против НМ1.24 пересаживают на человеческое антитело по методу пересадки ГВУ. L-цепь человеческого антитела REI используют в качестве L-цепи антитела человека, для каркасных областей (КО) 1-3 Н-цепи человеческого антитела используют КО 1-3 человеческого антитела HG3 и КO4 из человеческого антитела JH6 используют для КО4. Некоторые аминокислоты в КО из V-области Н-цепи замещают так, чтобы ГВУ-трансплантированное антитело могло сформировать подходящий сайт для связывания антигена.
Для экспрессии гена L-цепи и Н-цепи сконструированного таким образом гуманизированного антитела против НМ1.24 в клетки млекопитающего каждый ген отдельно вводят в вектор HEF для конструирования вектора, который будет экспрессировать L-цепь или Н-цепь гуманизированного антитела против НМ1.24 соответственно.
При одновременном введении указанных 2-х векторов экспрессии в СНО клетки устанавливается клеточная линия, которая продуцирует гуманизированное антитело против НМ1.24. Антигенсвязывающая активность и ингибирование связывающей активности по отношению к клеточной линии WISH клеток амниона человека гуманизированного антитела против НМ1.24, полученного при культивировании указанной клеточной линии,
были исследованы клеточным методом ИФТФА. Полученный результат показывает, что гуманизированное антитело против НМ1.24 имеет антигенсвязывающую активность, равную таковой у химерного антитела, и в том, что касается активности по ингибированию связывания с использованием биотинсодержащего мышиного антитела против НМ1.24, оно также проявляет активность, равную активности химерного антитела или мышиного антитела.
Е. coli, несущая плазмиду, которая содержит ДНК, кодирующую V-область L-цепи, и ДНК, кодирующую V-областъ Н-цепи химерного антитела против НМ1.24, была депонирована в Международной Коллекции в соответствии с положениями Будапештского Договора как Escherichia coli DH5α (pUC19-1.24L. -gk) и Escherichia coli DH5α (pUC19-1.24H-gγl) 29 августа 1996 г. Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония) как FERM BP-5646 и FERM BP-5644 соответственно. Кроме того, E. coli, несущая плазмиду, которая содержит ДНК, кодирующую вариант a (SEQ ID NO: 2) V-области L-цепи или вариант r (SEQ ID NO: 3) V-области Н-цепи гуманизированного антитела против НМ1.24, была депонирована в Международной Коллекции в соответствии с положениями Будапештского Договора как Escherichia coli DH5α (pUC19-RVLa-AHM-gk) и Escherichia coli DH5α (pUC19-RVHr-gγl), соответственно, 29 августа 1996 г. Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония) как FERM BP-5645 и FERM BP-5643, соответственно. Далее, E. coli, несущая плазмиду, которая содержит ДНК, кодирующую вариант s (SEQ ID NO: 4) V-области Н-цепи гуманизированного антитела против НМ1.24, была депонирована в Международной Коллекции в соответствии с положениями Будапештского Договора как Escherichia coil DH5α (pUC19-RVHs-AHM-gγl) 29 сентября 1997 г. Национальным Институтом биологических наук и гуманитарных технологий (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, of 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония) как FERM BP-6127.
Ссылочный пример 3. Клонирование кДНК, кодирующей белковый антиген НМ1.24
Клонируют кДНК, кодирующую белковый антиген НМ1.24, специфически распознаваемый антителом против НМ1.24.
1. Конструирование библиотеки кДНК
1) Получение общей РНК
Из клеточной линии множественной миеломы человека КРММ2 получают общую РНК по методу Чиргвина с соавт. (Chirgwin et al. , Biochemistry, 18, 5294, 1979). В соответствии с указанным методом, 2,2•108 клеток КРММ2 полностью гомогенизируют в 20 мл 4 M гуанидинотиоцианата (производство Nacalai Tesque Inc. ). Гомогенат наслаивают на 5,3 М раствор хлорида цезия в центрифужной пробирке, которую затем центрифугируют с использованием ротора Beckman SW40 со скоростью 31000 об/мин при 20oС в течение 24 часов для осаждения РНК. Осадок РНК промывают 70% этанолом и затем растворяют в 300 мкл 10 мМ Трис-НСl (рН 7,4), содержащем 1 М ЭДТА и 0,5% ДСН. Добавляют проназу (производство Boehringer) до концентрации 0,5 мг/мл и затем проводят инкубацию при температуре 37oС в течение 30 минут. Смесь экстрагируют фенолом и хлороформом и РНК осаждают этанолом. Осадок РНК растворяют в 200 мкл 10 мМ Трис-НСl (рН 7,4), содержащем 1 мМ ЭДТА.
2) Получение поли(А)+РНК
Поли(А)+РНК очищают с использованием в качестве исходного материала 500 мкг обшей РНК, полученной по описанному выше методу с помощью набора для выделения мРНК Fast Track 2,0 mRNA Isolation Kit (производство Invitrogen) в соответствии с прилагаемой к набору инструкцией.
3. Конструирование библиотеки ДНК
Синтезируют двуцепочечную кДНК с использованием в качестве исходного материала 10 мкг указанной выше поли-(А)+РНК с помощью набора для синтеза кДНК TimeSaver cDNA Synthesis Kit (производство Pharmacia), в соответствии с прилагаемой к набору инструкцией, и затем проводят лигирование с EcoRI адаптером, поставляемым в наборе, с использованием Directional Cloning Toolbox (производство Pharmacia), в соответствии с прилагаемой к набору инструкцией. Кэширование и обработку рестрикционным ферментом NotI адаптера EcoRI проводят в соответствии с инструкцией, прилагаемой к набору. Далее двуцепочечную кДНК, прилагаемую к адаптеру, имеющую размер 500 нп или более, отделяют и очищают с использованием 1,5% низкоплавкого агарозного геля (производство Sigma) с получением 40 мкл добавленной к адаптеру цепочечной кДНК.
Полученную таким образом добавленную к адаптеру двуцепочечную кДНК лигируют с использованием вектора pCOSl (заявка на патент Японии 8-255196) и ДНК-лигазы Т4 (производство Gibco-BRL), которая была предварительно обработана рестриктазами EcoRI и NotI и щелочной фосфатазой (производство Takara Shuzo) для создания библиотеки кДНК. Сконструированную библиотеку кДНК с помощью трансдукции вводят в штамм Е. coli DH5α (производство Gibco-BRL), при этом впоследствии было установлено, что это независимый клон, имеющий общий размер около 2,5•106.
2. Клонирование методом прямой экспрессии
1) Траксфекция COS-7 клеток
Примерно 5•105 клонов указанного выше штамма трансдуцированной E. coli культивируют в среде 2-YT (Molecular Cloning: A Laboratory Manual, Sambrook et al. , Cold Spring Harbor Laboratory Press, 1989), содержащей 50 мкг/мл ампициллина, для амплификации кДНК, которую подвергают обработке по щелочному методу (Molecular Cloning: A Laboratory Manual, Sambrook et al. , Cold Spring Harbor Laboratory Press, 1989), с восстановлением плазмидной ДНК из Е. соli. Полученную таким образом плазмидную ДНК переносят методом трансфекции в COS-7 клетки методом электропорации с использованием прибора Gene Pulser (производство Bio-Rad).
В соответствии с указанным методом, 10 мкг очищенной плазмидной ДНК добавляют к 0,8 мл раствора COS-7 клеток, в котором клетки были суспендированы в ФБР в концентрации 1•107 клеток/мл, и полученную смесь подвергают воздействию импульсов в 1500 V при емкости 25 мкФ. По окончании 10-минутного восстановительного периода при комнатной температуре клетки после электропорации культивируют в культуральной среде DMEM (производство Gibco-BRL), содержащей 10% околоплодной сыворотки теленка (производство Gibco-BRL) при температуре 37oС в атмосфере 5% СО2 в течение 3-х дней.
2) Подготовка кюветы
Кювету, на которую наносят мышиное антитело против НМ1.24, готовят по методу Сида с соавт. (В. Seed et al. , Proc. Natl. Acad. Sci. USA, 84, 3365-3369, 1987). В соответствии с указанным методом мышиное антитело против НМ1.24 добавляют к 50 мМ Трис-НСl (рН 9,5) до концентрации 10 мкг/мл. Три миллилитра приготовленного таким образом раствора антитела добавляют к чашке с клеточной культурой диаметром 60 мм и инкубируют при комнатной температуре в течение 2-х часов. После промывания 3 раза 0,15 М раствором NaCl к чашке добавляют ФБР, содержащий 5% околоплодной сыворотки теленка, 1 мМ ЭДТА и 0,02% NаN3. Смесь после блокирования используют для последующего клонирования.
3) Клонирование библиотеки кДНК
COS-7 клетки, подвергшиеся трансфекции по описанному выше методу, обрабатывают ФБР, содержащим 5 мМ ЭДТА. После однократного промывания клеток ФБР, содержащим 5% околоплодной сыворотки теленка, их суспендируют в ФБР, содержащем 5% околоплодной сыворотки теленка и 0,02% NaN3 до концентрации примерно 1•106 клеток/мл. Суспензию вносят в кювету, подготовленной как указано выше, и проводят инкубацию при комнатной температуре в течение примерно 2-х часов. После осторожного трехкратного промывания ФБР, содержащим 5% околоплодной сыворотки теленка и 0,02% NаN3, выделяют плазмидную ДНК из клеток, связанных с кюветой, с использованием раствора, содержащего 0,6% ДСН и 10 мМ ЭДТА.
Восстановленную плазмидную ДНК переносят методом трансдукции в E. coli DH5α. После амплификации по описанному выше методу плазмидную ДНК восстанавливают щелочном методом. Восстановленную плазмидную ДНК переносят путем трансфекции в COS-7 клетки с помощью электропорации и восстановленную из клеток плазмидную ДНК связывают, как описано выше. Повторяют один раз аналогичную процедуру и восстановленную плазмидную ДНК расщепляют рестриктазами. EcoRI и NotI, что подтверждает факт наличия вставки, имеющей размер около 0,9 тыс. нм. Клетки E. coli, в которых часть восстановленной плазмидной ДНК была перенесена путем трансдукции, инокулируют на чашку с 2-YT агаром, содержащим 50 мкг/мл ампициллина. После культивирования в течение ночи выделяют из одной колонии плазмидную ДНК. Ее расщепляют рестриктазами EcoRI и NotI с получением клона р3.19, в котором размер вставки составляет примерно О, 9 тыс. нп.
Проводят реакцию указанного клона с использованием набора для секвенирования PRISM, Dye Terminator Cycle Sequencing kit (производство Perkin Elmer), в соответствии с прилагаемой к набору инструкцией и определяют нуклеотидную последовательность с использованием секвенатора ДНК ABI 373А DNA Sequencer (производство Perkin Elmer). Указанная нуклеотидная последовательность и соответствующая аминокислотная последовательность представлены в SEQ ID NO: 1.
Промышленная применимость
Результаты анализа методом проточной цитометрии показали, что антитело против НМ1.24 активно взаимодействует с большинством клеток, полученных из опухолей лимфатических тканей, указывая на то, что в случае, многих опухолей лимфатических тканей полипептид, имеющий эпитоп, распознаваемый антителом против НМ1.24, экспрессируется в высокой степени. Кроме того, введение мышам с трансплантированной лимфатической опухолью человека, которая реагирует с антителом НМ1.24, антитела против НМ1.24 приводит к подавлению роста объема опухоли, а также к увеличению периода выживания. Указанные факты показывают, что антитело против НМ1.24 или антитела, распознаваемые, полипептидом, имеющим эпитоп, распознаваемый антителом против НМ1.24, обладают цитотоксической активностью против многих опухолей лимфатических тканей и это дает основание предполагать, что указанное антитело может использоваться для лечения больных с опухолями лимфатических тканей.
Ссылка на микроорганизмы, депонированные в соответствии с договором по Патентному сотрудничеству (Patent Cooperation Treaty), Положение 13-2 и наименование Депонирующего Института.
Депонирующий Институт
Наименование: Национальный Институт биологических наук и гуманитарных технологий. Агентство промышленной науки и технологии (National Institute of Bioscience and Human-Technology, Agency-of Industrial Science and Technology)
Адрес: 1-3, Higashi 1-chome, Tsukuba city, Ibaraki pref. , Япония
Микроорганизм (1)
Наименование: Escherichia coli DH5α (pRS38-pUC19)
Номер хранения: FERM BP-4434
Дата депонирования: 5 октября 1993 года
Микроорганизм (2)
Наименование: Гибридома НМ1.24
Номер хранения: FERM BP-5233
Дата депонирования: 14 сентября 1995 г.
Микроорганизм (3)
Наименование: Escherichia coil DH5α (pUC19-RVHr-AHM-gγl)
Номер хранения: FERM BP-5643
Дата депонирования: 29 августа 1996 г.
Микроорганизм (4)
Наименование: Escherichia coil DH5α (pUC19-l. 24H-gγl)
Номер хранения: FERM BP-5644
Дата депонирования: 29 августа 1996 г.
Микроорганизм (5)
Наименование: Escherichia coli DH5α (pUC19-RVLa-AHM-gk)
Номер хранения: FERM BP-5645
Дата депонирования: 29 августа 1996 г.
Микроорганизм (6)
Наименование: Escherichia coli DH5α (pUC19-l. 24L-gk)
Номер хранения: FERM BP-5646
Дата депонирования: 29 августа 1996 г.
Микроорганизм (7)
Наименование: Escherichia coli DH5α (pUC19-RVHs-AHM-gγl)
Номер хранения: FERM BP-6127
Дата депонирования: 29 сентября 1997 г.
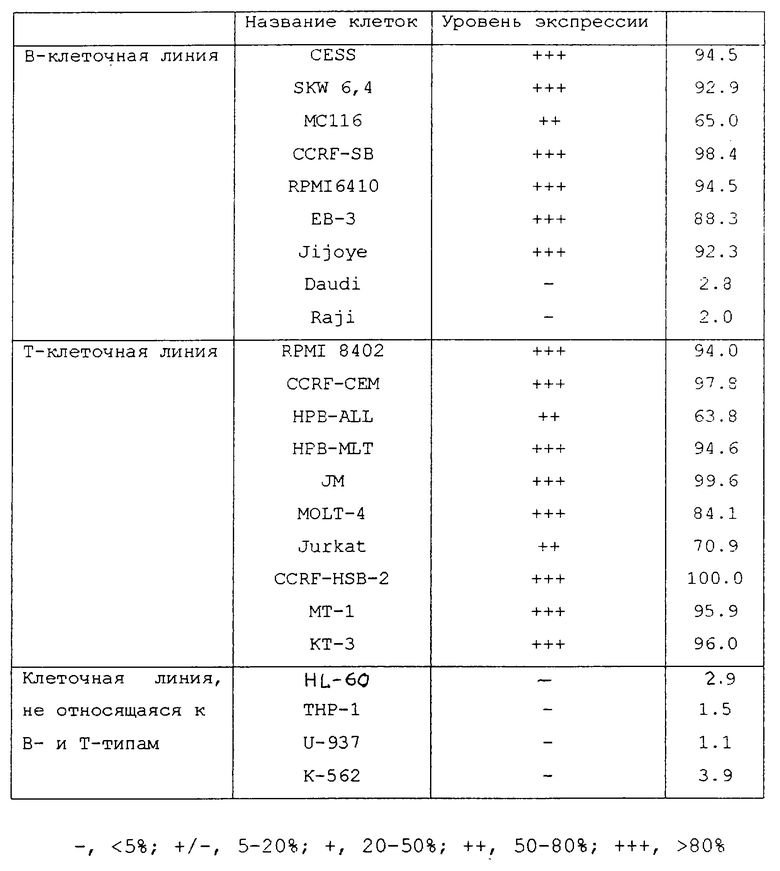










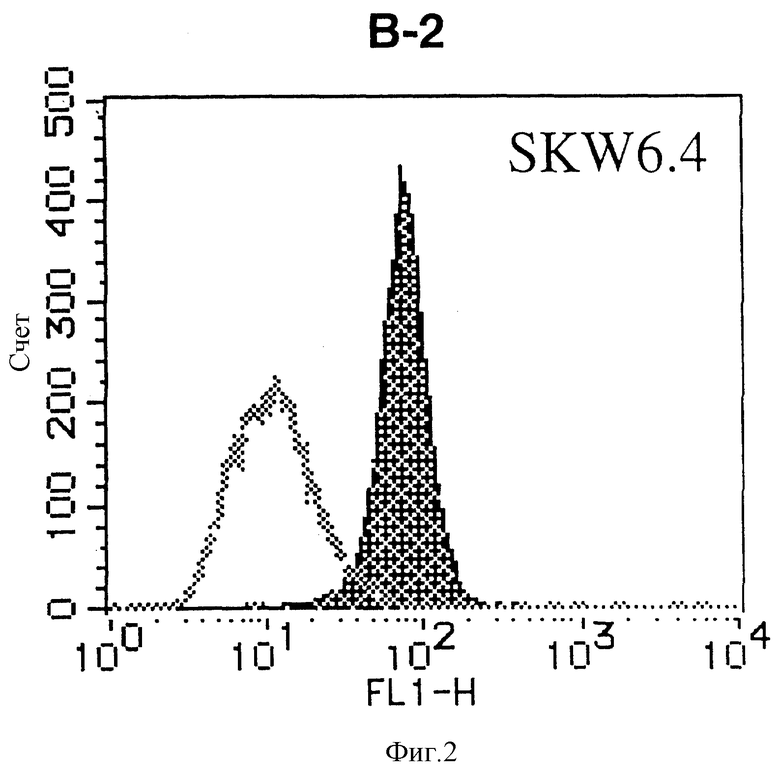
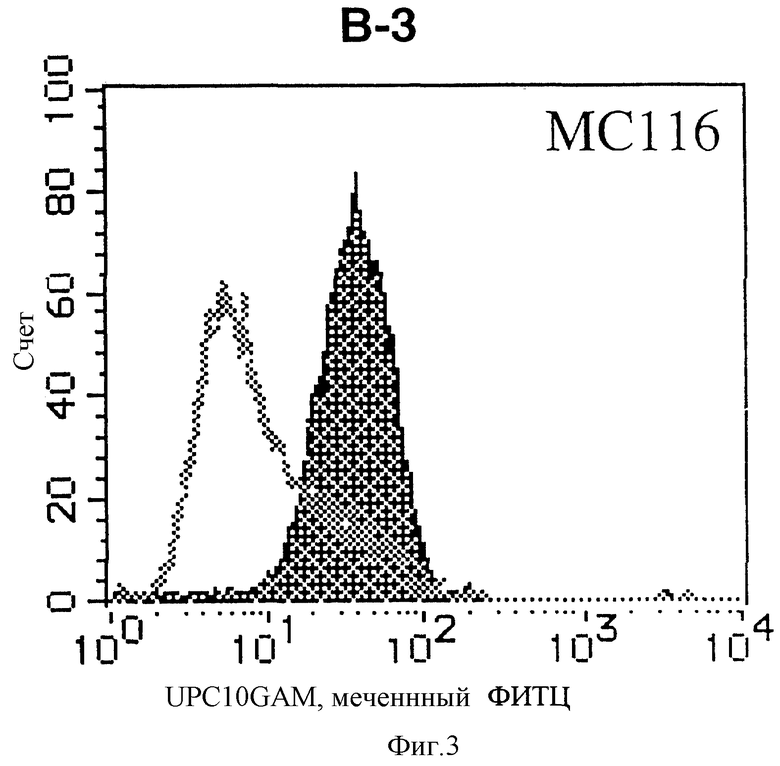
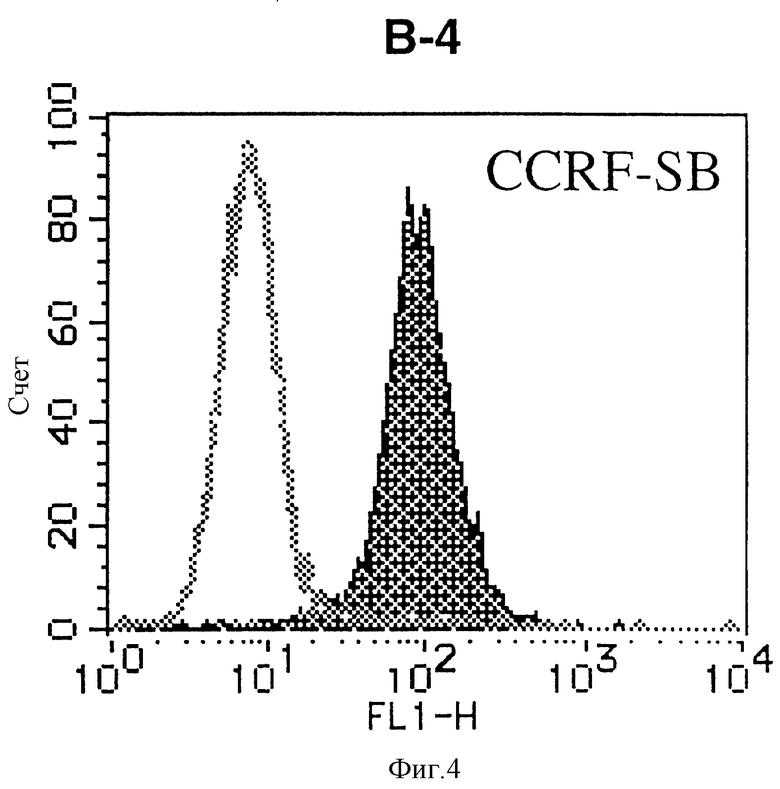
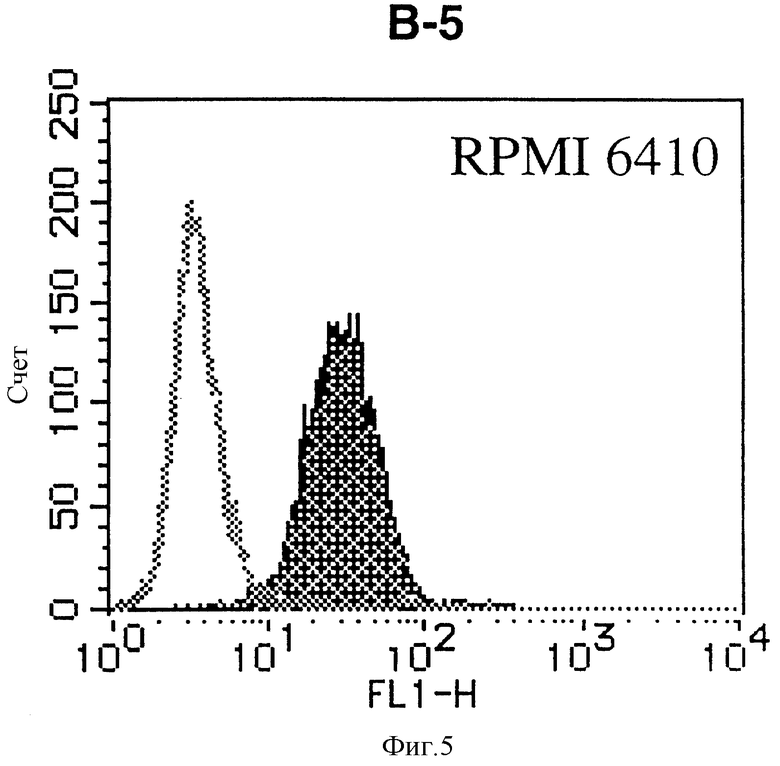
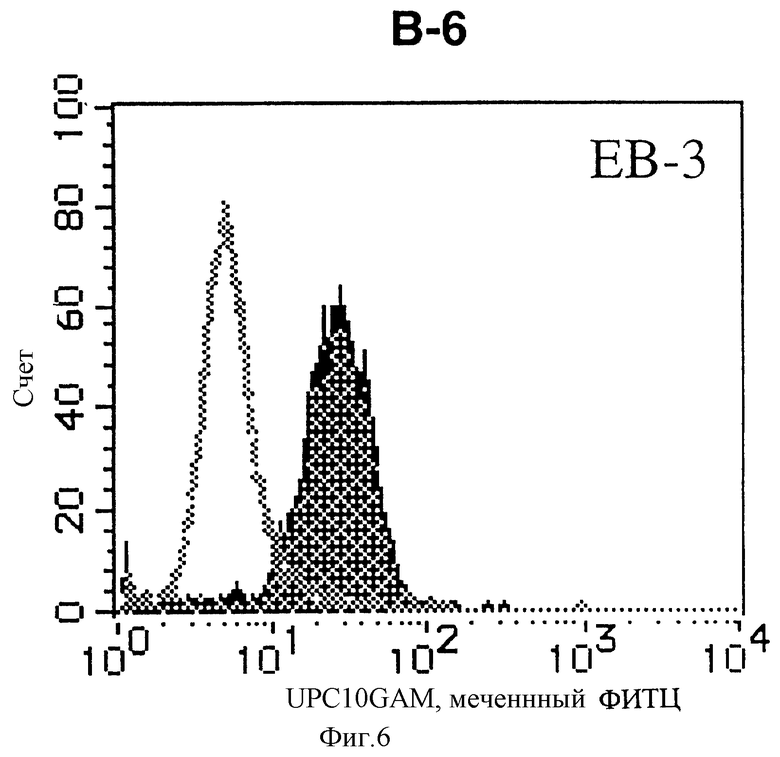
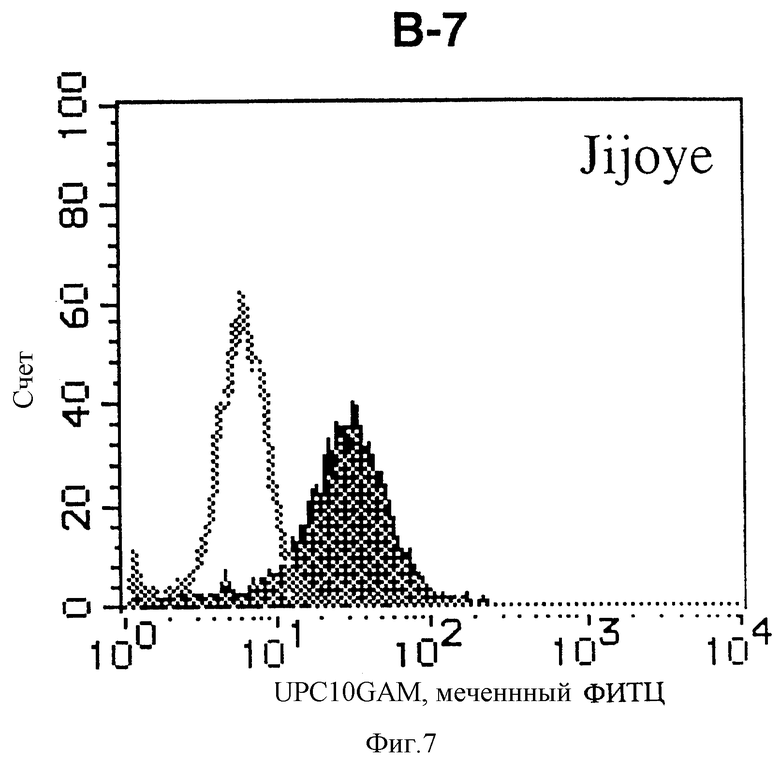
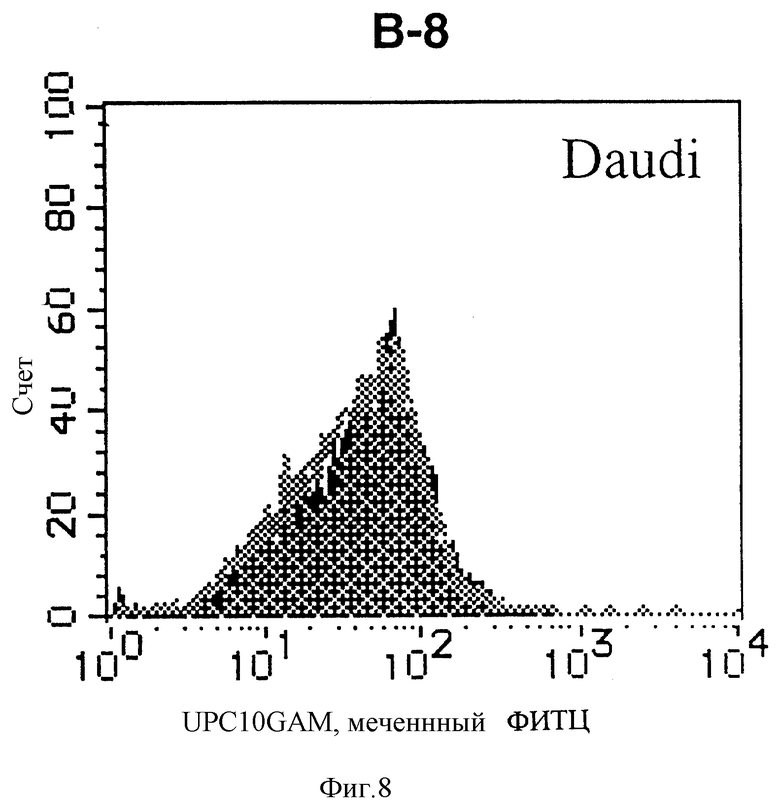
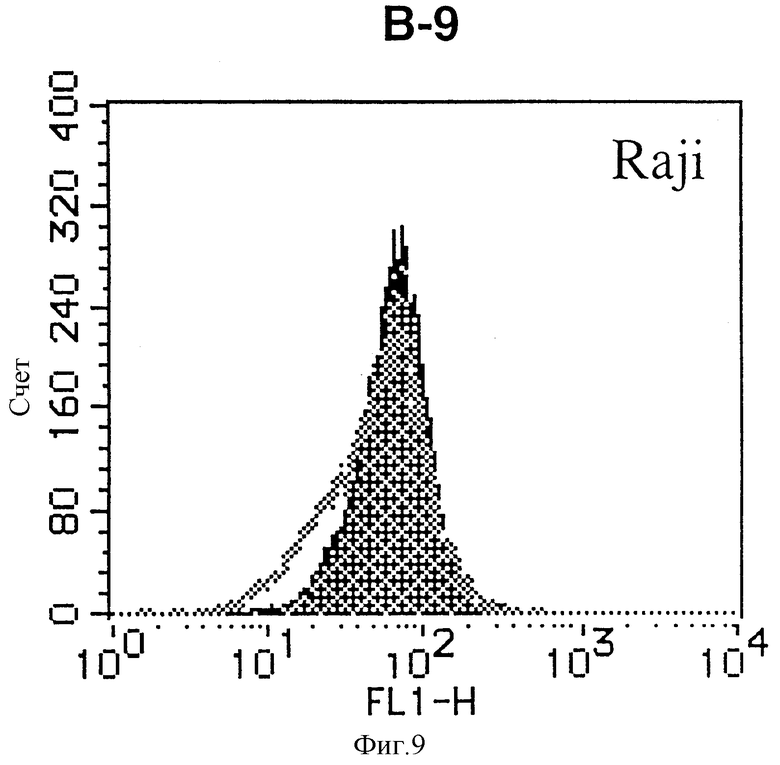
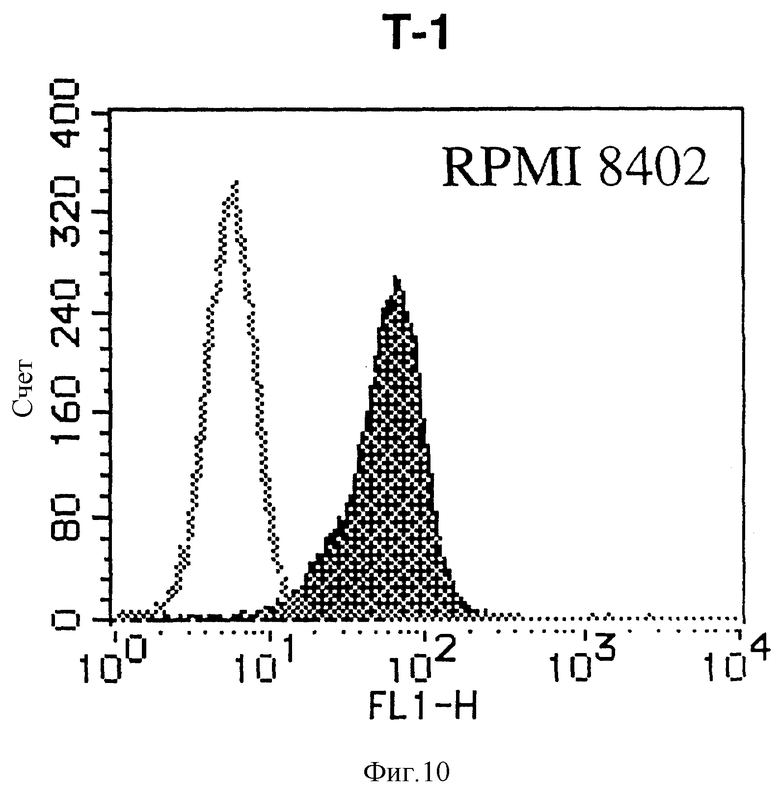
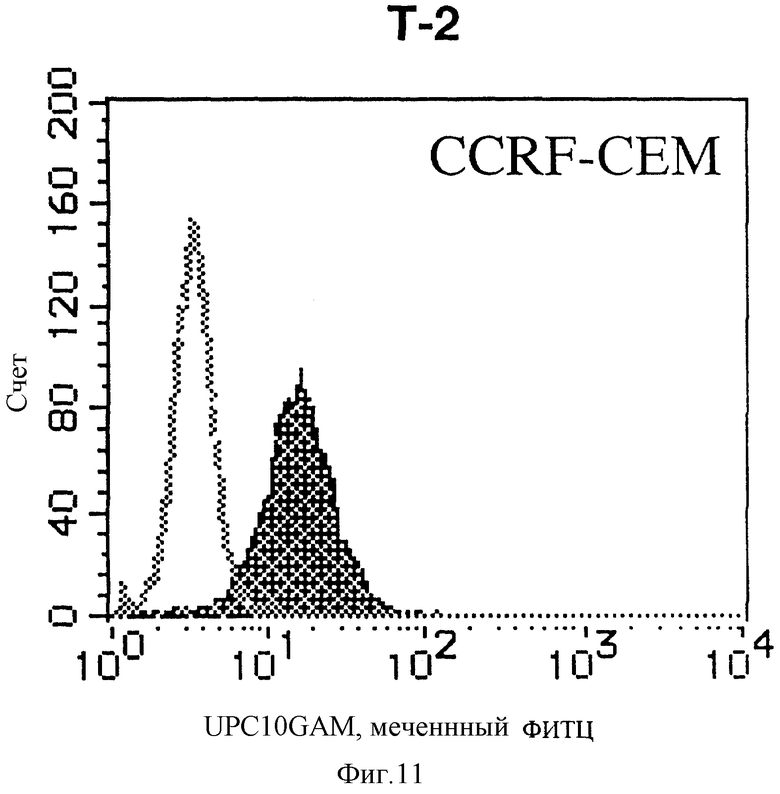
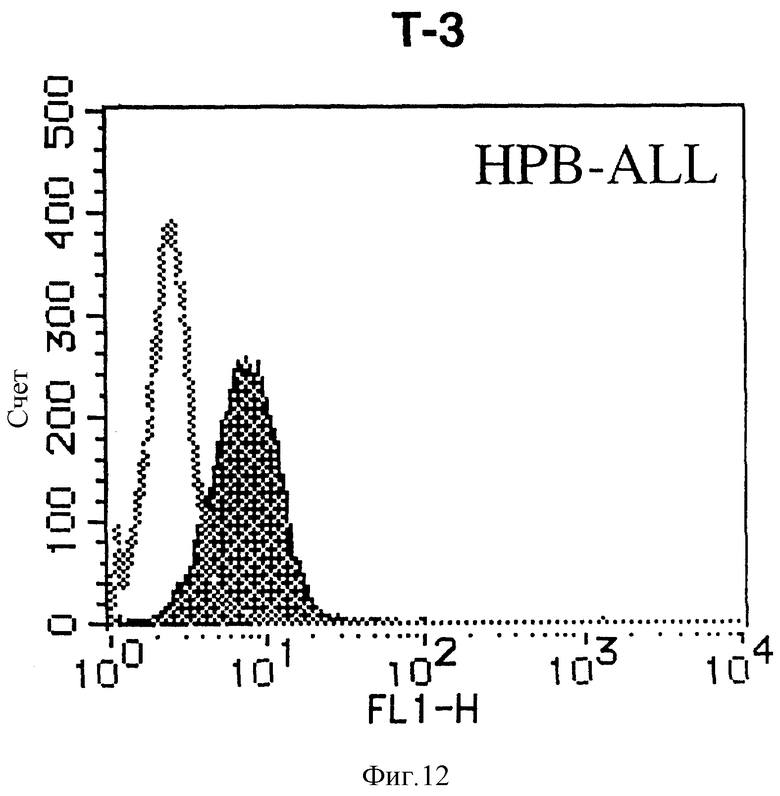
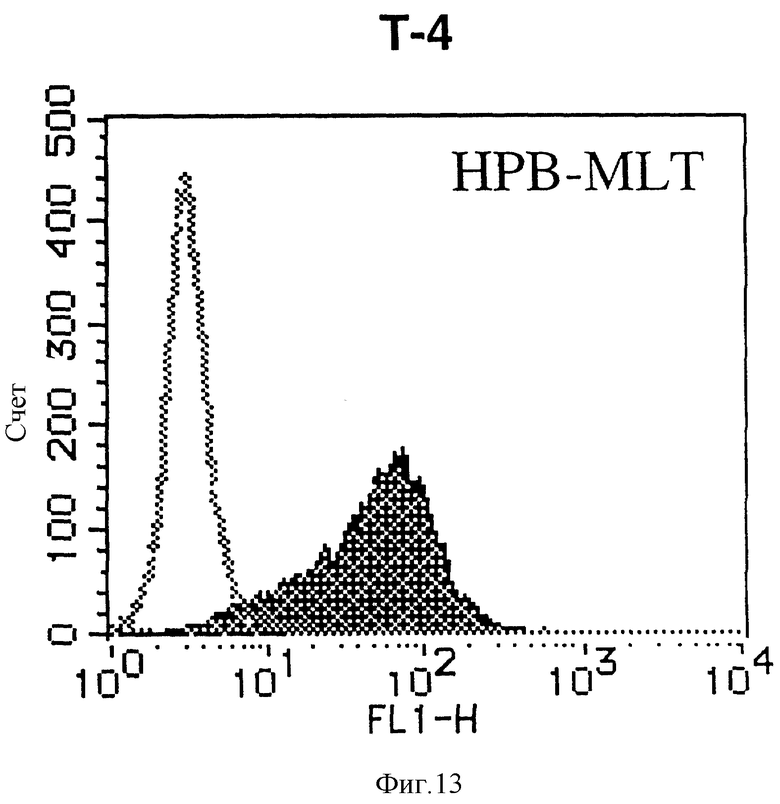
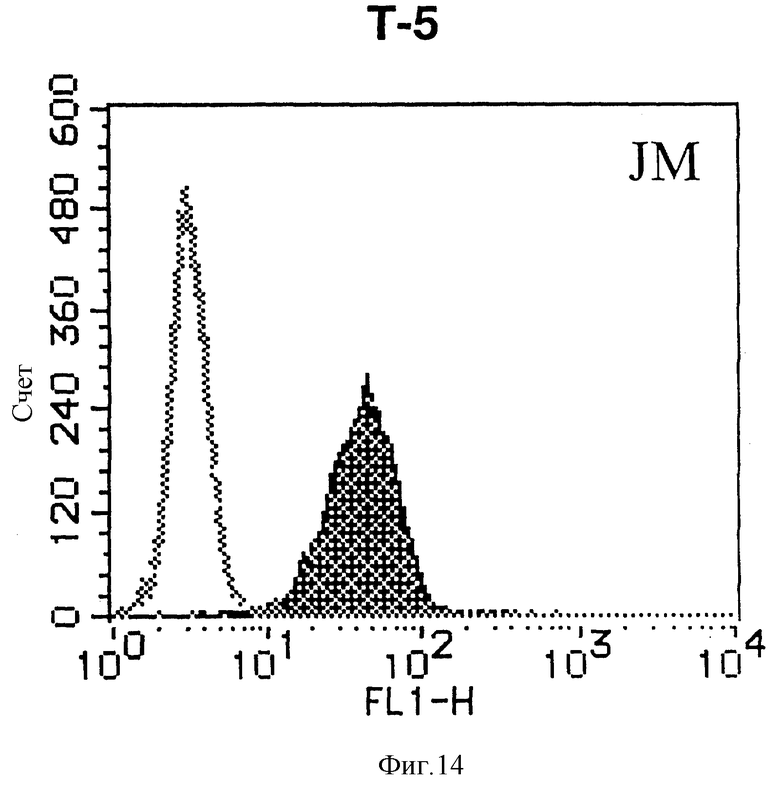
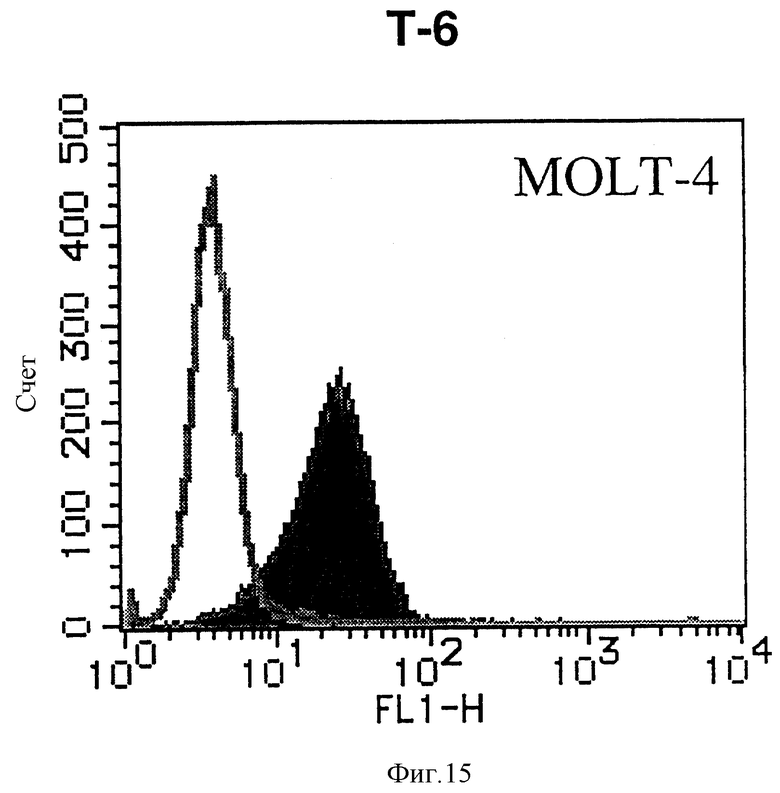
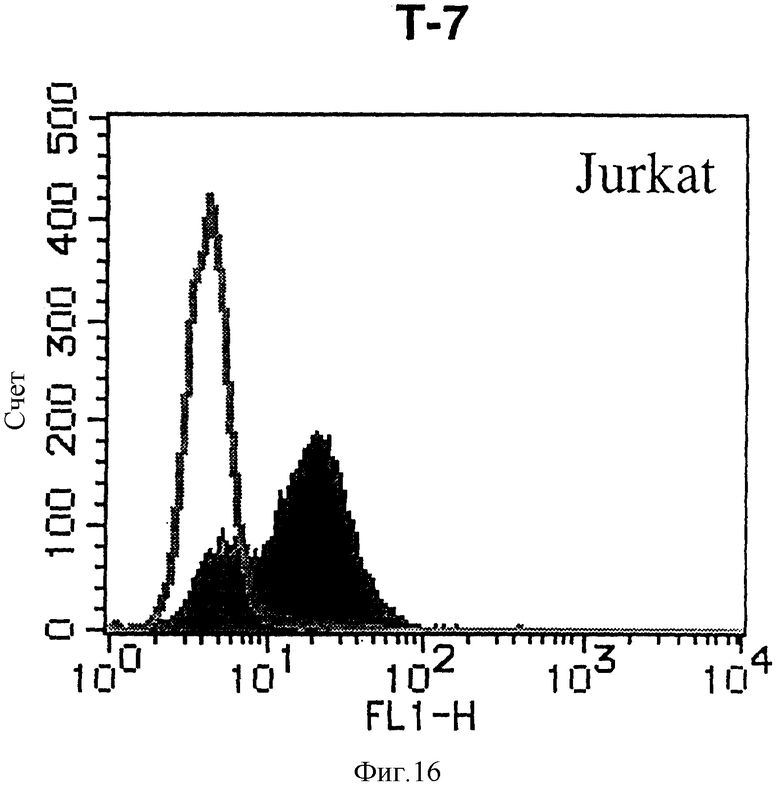
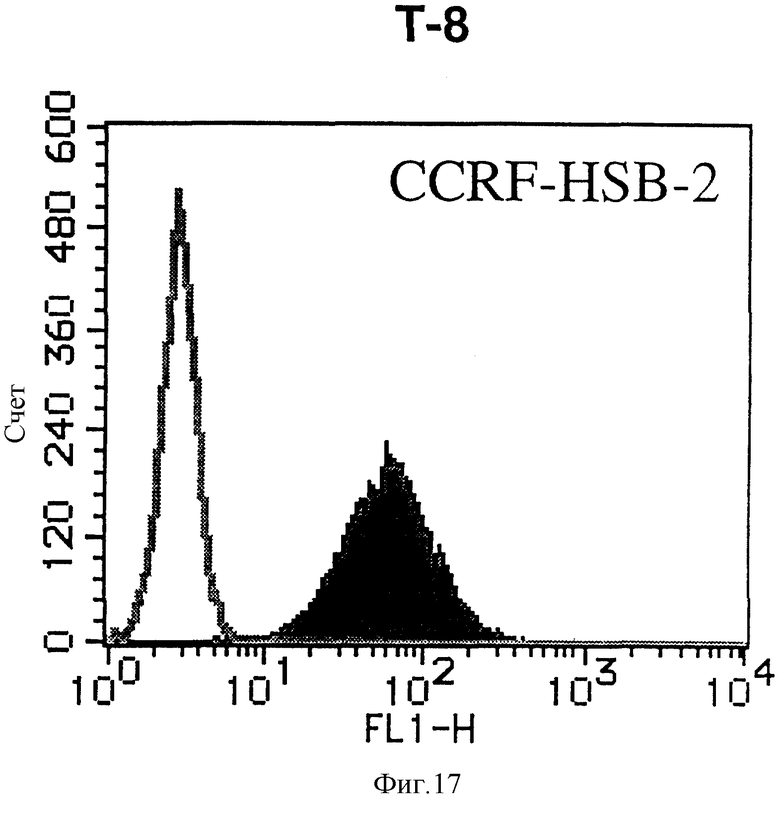
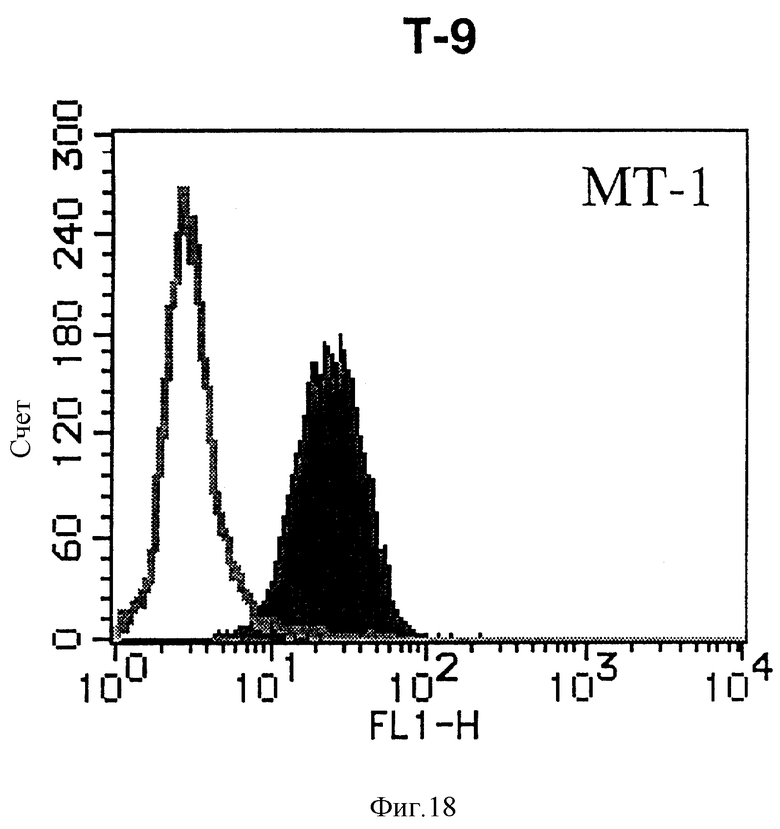
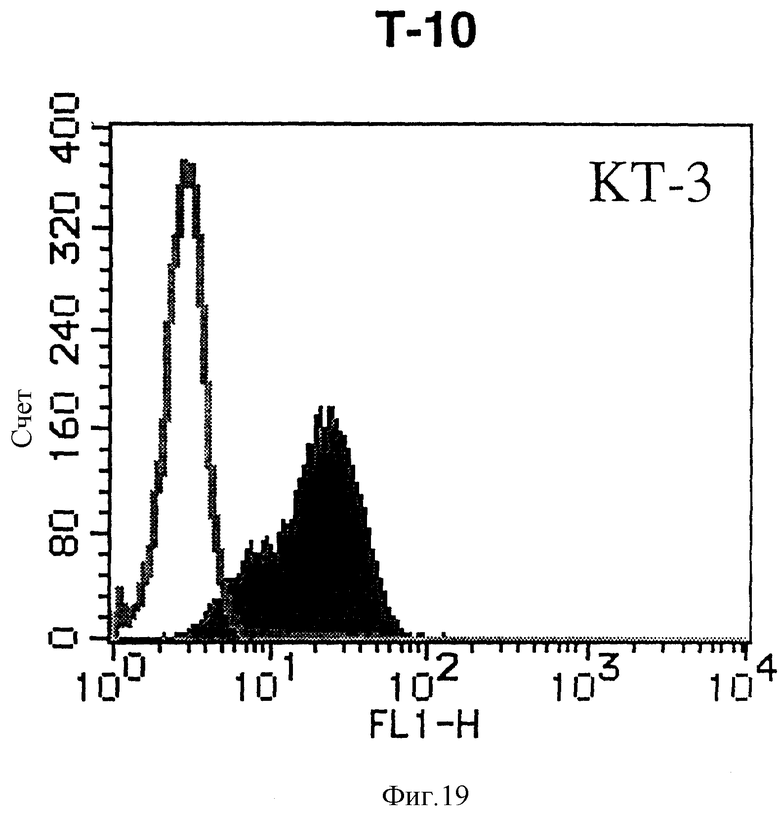
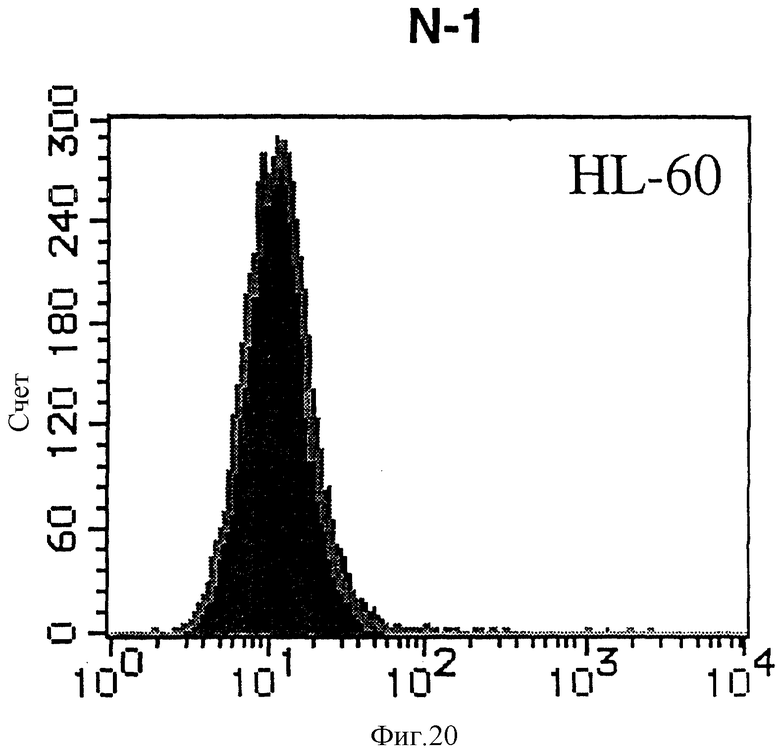
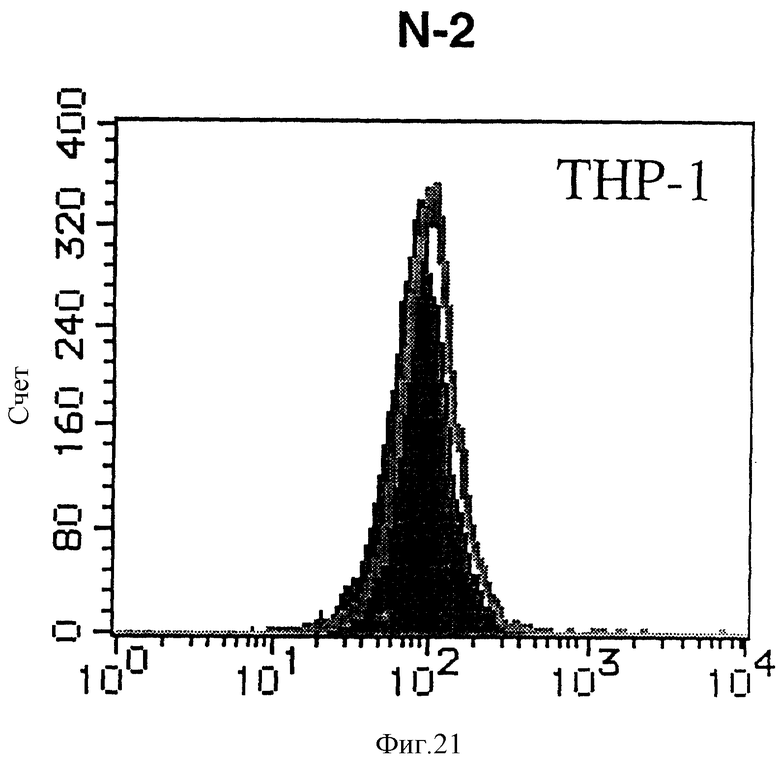
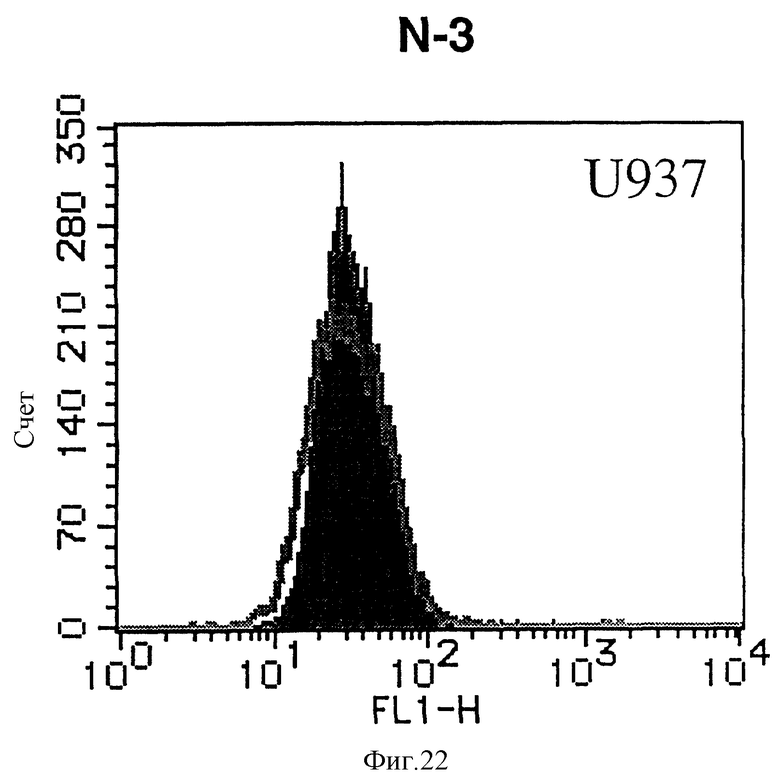
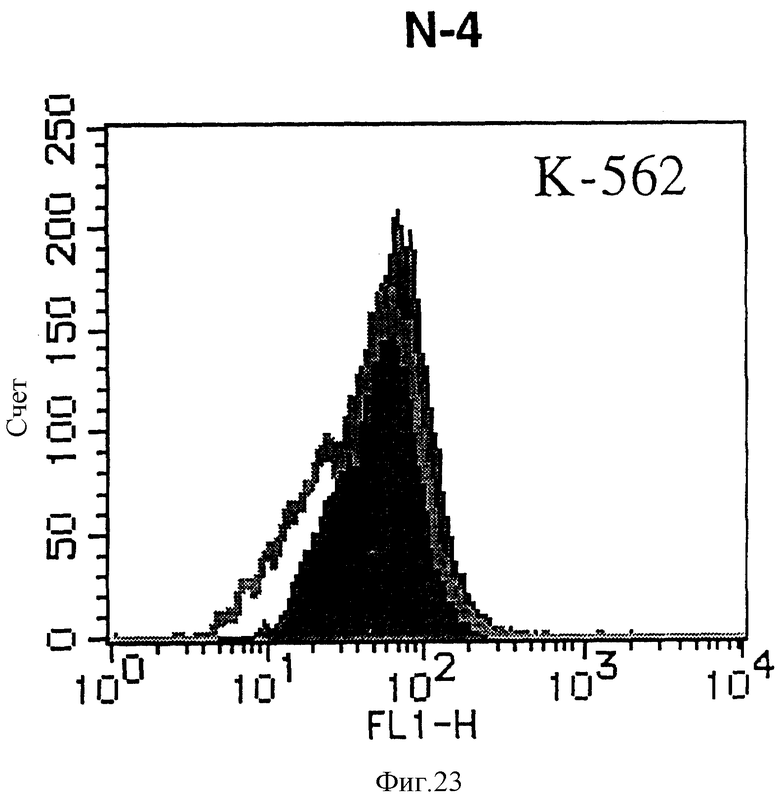
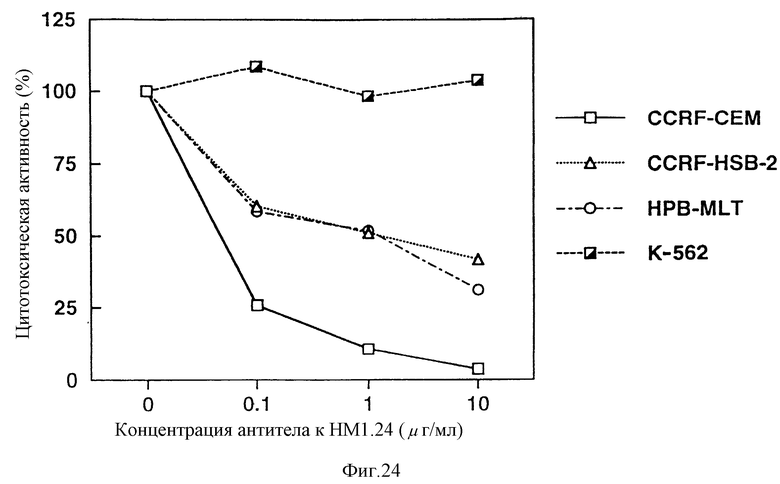
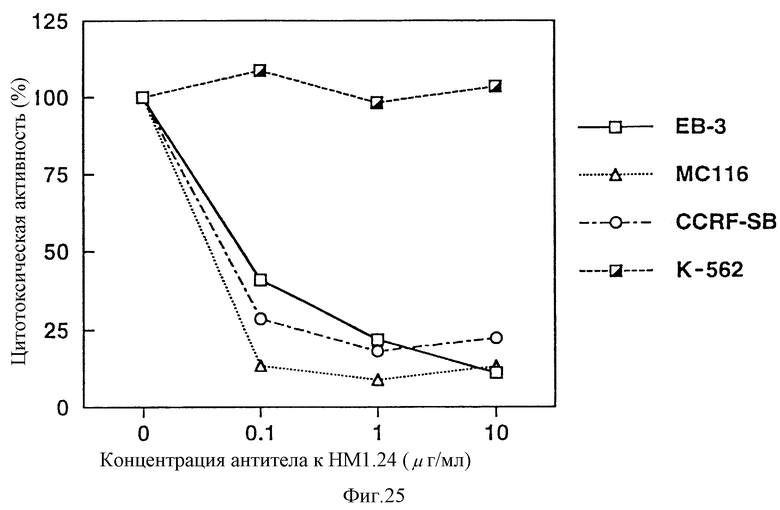
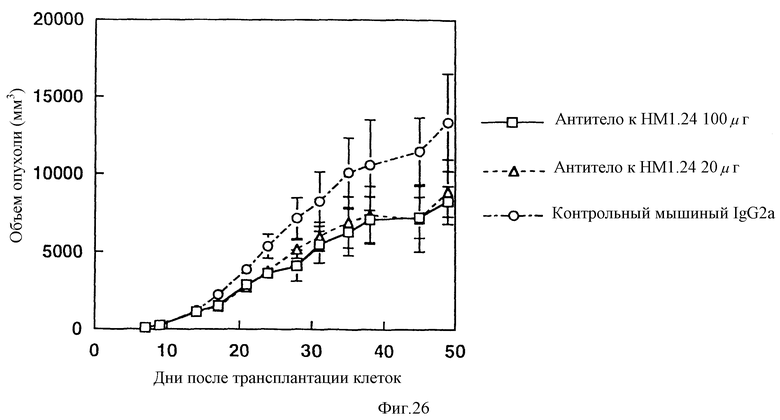
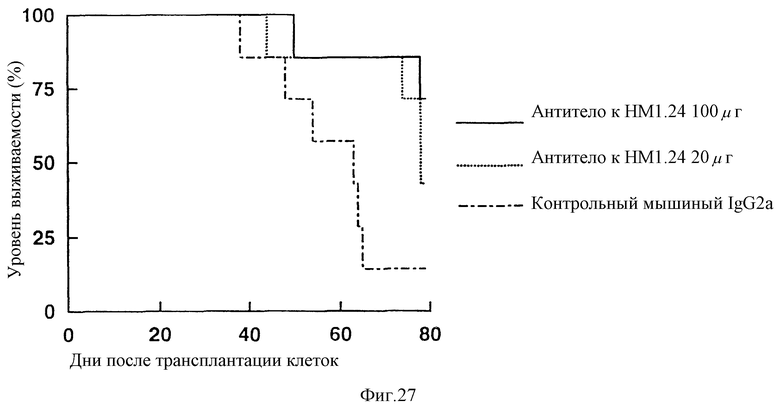
Изобретение относится к медицине. Сущность его составляет лекарственное средство, применяемое при лечении опухолей лимфатической ткани (за исключением миеломы), включающее в качестве активного ингредиента антитело, которое специфически связывается с белком, имеющим аминокислотную последовательность, указанную в SEQ ID NO: 1, в описании, и которое обладает цитотоксической активностью. Указанное антитело содержит констатную область Cγ или представляет собой химерное антитело против НМ1. Технический результат - расширение арсенала средств против опухолей лимфатической ткани. 2 с. и 17 з. п. ф-лы, 27 ил. , 1 табл.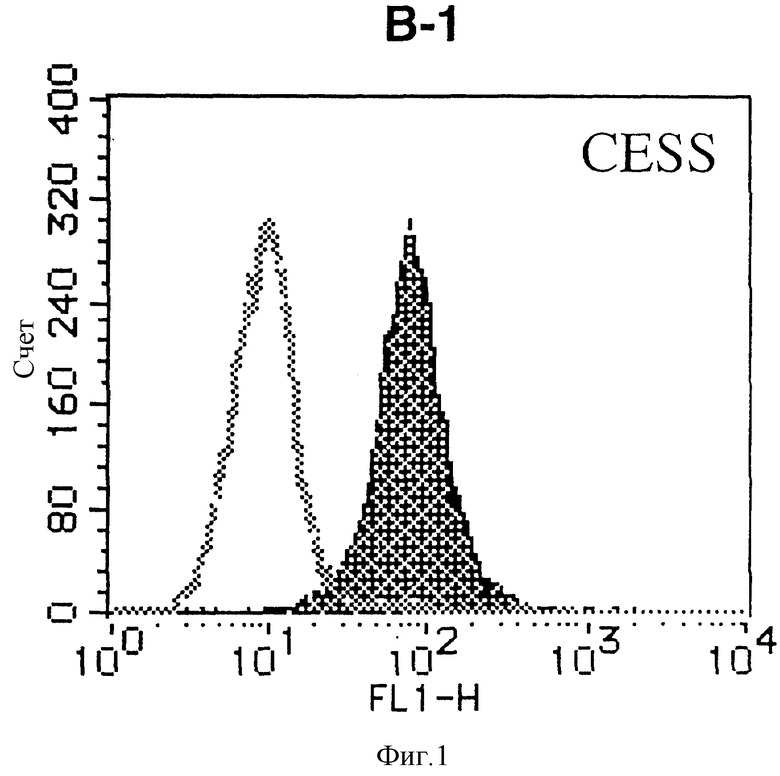
7. Лекарственное средство по п. 4, отличающееся тем, что указанное антитело содержит константную область Сγ из человеческого антитела.
| Торфодобывающая машина с вращающимся измельчающим орудием | 1922 |
|
SU87A1 |
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |

