Настоящая заявка заявляет приоритет Предварительной патентной заявки Соединенных Штатов №60/873,529, зарегистрированной 7 декабря 2006 г., включенной в полном объеме в описание настоящей заявки в качестве ссылки.
Область техники, к которой относится изобретение
Настоящее изобретение предусматривает составы и способы получения сортов сорго, которые устойчивы к гербицидам. В частности, настоящее изобретение предусматривает растения сорго, ткани растения и семена растения, которые содержат измененные гены и белки ацетолактатсинтазы (ALS), которые устойчивы к ингибированию гербицидами, которые в норме ингибируют активность белка (ALS).
Предпосылки создания изобретения
Сорго является вторым по важности кормовым хлебным злаком, выращиваемым в Соединенных Штатах. Производство экономически важно для ферм, функционирующих в пограничных районах с достаточным количеством осадков, из-за способности сорго переносить засуху и жару. Как промышленное животноводство, так и биоэнергетика используют сорго в качестве энергетического субстрата, делая его, таким образом, универсальной сельскохозяйственной культурой.
Во всем мире сорго занимает пятое место среди хлебных злаков (продовольственного зерна). Так как оно устойчиво как к засухе, так и к жаре, оно, несомненно, является наиболее широко выращиваемым хлебным злаком в полупустынных областях Африки к югу от Сахары и в засушливой области центрального полуострова Индии. По существу, сорго используется для потребления человеком в большинстве наиболее засушливых областей мира, что, таким образом, делает его особенно важной продовольственной культурой в этих местах.
Развитие у растений устойчивости к гербицидам сулит значительную выработку и экономические преимущества, использование гербицидов для контролирования сорных растений среди зерновых, по существу, стало едва ли не общим правилом. Однако использование таких гербицидов может также привести к отмиранию или редуцированному росту желательного хлебного злака, делая время и способ применения гербицида критическими или в некоторых случаях невозможными.
Особый интерес для фермеров представляет использование более сильных гербицидов, обладающих эффективностью против широкого спектра сорных растений и быстрым разложением в почве. Растения, ткани и семена растений с устойчивостью к этим соединениям обеспечивают привлекательное решение, давая возможность применять гербициды для контроля роста сорных растений с небольшим риском повреждения сельскохозяйственной культуры. Одним таким классом гербицидов широкого спектра действия являются такие гербициды, которые ингибируют активность фермента ацетолактатсинтазы (ALS) у растения. Ацетолактатсинтаза требуется для продукции незаменимых аминокислот, таких как валин, лейцин и изолейцин, у растений (этот сигнальный путь отсутствует у человека или других животных). Сорго чувствительно ко многим ALS-ингибирующим гербицидам, которые направлены на однодольные виды, что делает использование этих гербицидов для контроля над травянистыми сорными растениями практически невозможным, так как они будут также ингибировать и рост хлебного злака.
Ацетолактатсинтазные гербициды контролируют широкий спектр травянистых и лиственных сорных растений в очень низких дозах. В настоящее время доступно более 56 различных ALS-гербицидов, приготовленных из разных композиций сульфонилкарбомидов (SU), имидозолинонов (IMI), триазолопиримидинов (ТР) и пиримидинилтиобензоатов (РТВ). Было обнаружено, что мутации у некоторых хлебных злаков, например табака, кукурузы (Патент США № 5,767,361) и сои, придают устойчивость к ALS-гербицидам, однако на сегодняшний день не было сделано такого открытия для сорго (в качестве обзора см. Tan et al., 2005, Pest. Manag. Sci. 61:246-247). Вследствие важности сорго на мировом уровне необходимы культуры сорго, которые устойчивы к ингибирующим эффектам ALS-гербицидов, дающие возможность, в связи с этим, получать большую урожайность, когда эти гербициды используются для контролирования травянистых сорных растений. Из-за значимости сорго на мировом уровне необходимыми являются сорта сорго, которые устойчивы к ингибиторным эффектам ALS-гербицидов, позволяющие, таким образом, получить больший урожай сельскохозяйственной культуры при использовании этих гербицидов для контроля травянистых сорных растений.
Сущность изобретения
Настоящее изобретение предусматривает составы и способы получения сортов сорго, которые устойчивы к гербицидам. В частности, настоящее изобретение предусматривает растения сорго, ткани растения и семена растения, которые содержат измененные гены и белки ацетолактатсинтазы (ALS), которые устойчивы к ингибированию гербицидами, которые в норме ингибируют активность белка ALS.
Выращиваемое сорго [Sorghum bicolor (L.) Moench] чувствительно ко многим ALS-ингибирующим гербицидам, которые направлены на однодольные или травянистые виды. Однако, как описано в настоящей заявке, было обнаружено, что генотип сорго проявляет устойчивость к ALS-гербицидам. Генетический анализ выявил генетические различия в зародышевой плазме сорго, которые приводят к фенотипу, устойчивому к ALS-гербицидам.
В одной модификации изобретение предусматривает одно или более растения сорго, зародышевая плазма которых содержит мутацию, которая приводит к устойчивости растения к ALS-гербицидам. Кроме того, в дополнительных модификациях изобретение имеет отношение к потомкам (например, F1, F2, F3 и т.д.) от скрещивания вышеупомянутого растения, при котором зародышевая плазма вышеупомянутых потомков имеет ту же мутацию, что и родительское растение. Следовательно, модификации настоящего изобретения предусматривают гибриды сорго, зародышевая плазма которых содержит такую мутацию, что фенотип растений является устойчивым к ALS-гербицидам.
В одной модификации настоящее изобретение предусматривает гибрид сорго, при котором вышеупомянутая зародышевая плазма придает устойчивость к ингибированию одним или более ацетолактатсинтазными гербицидами при уровнях одного или более вышеупомянутых гербицидов, которые в норме ингибировали бы рост гибрида сорго. В некоторых модификациях вышеупомянутые один или более ацетолактатсинтазные гербициды относятся к группе, состоящей из сульфонилмочевин, имидазолинонов, тразолопиримидинов и пиримидилтиобензоатов. Как обнаружено в АТСС № РТА-7999, в некоторых модификациях вышеупомянутая зародышевая плазма гибрида сорго, которая придает устойчивость к ингибированию одним или более ацетолактатсинтазными гербицидами, содержит мутации в гене ацетолактата. В некоторых модификациях семена от вышеупомянутого гибрида сорго покрыты ацетолактатсинтазным гербицидом.
В одной модификации настоящее изобретение предусматривает способ контролирования сорных растений поблизости от гибрида сорго, как описано здесь, включающий обеспечение одного или более ацетолактатсинтазных гербицидов, применение вышеупомянутых одного или более ацетолактатсинтазных гербицидов на площади, содержащей гибрид сорго, как описано здесь, и контролирование сорных растений поблизости от вышеупомянутого гибрида сорго таким образом, что росту сорных растений наносится ущерб при помощи применения вышеупомянутых одного или более гербицидов, а росту вышеупомянутого гибрида сорго не наносится ущерб. В некоторых модификациях вышеупомянутые один или более ацетолактатсинтазные гербициды относятся к группе, состоящей из сульфонилмочевин, имидазолинонов, тразолопиримидинов и пиримидилтиобензоатов. Как обнаружено в АТСС № РТА-7999, в некоторых модификациях вышеупомянутый гибрид сорго содержит одну или более мутации в гене ацетолактатсинтазы.
В одной модификации настоящее изобретение обеспечивает гибрид сорго, при котором вышеупомянутый гибрид сорго содержит зародышевую плазму, содержащую одну или более мутации в гене ацетолактатсинтазы, такую, что вышеупомянутому гибриду придается устойчивость к одному или более ацетолактатсинтазным гербицидам. В некоторых модификациях вышеупомянутый гибрид сорго создается при помощи интрогрессии зародышевой плазмы сорго, которая содержит вышеупомянутые одну или более мутации для придания устойчивости к одному или более ацетолактатсинтазным гербицидам. В некоторых модификациях вышеупомянутый гибрид сорго создается при помощи включения гетерологичного гена, содержащего одну или более мутации для придания устойчивости к одному или более ацетолактатсинтазным гербицидам.
В одной модификации настоящее изобретение обеспечивает способ для получения гибридной линии растения сорго, устойчивой к одному или более ацетолактатсинтазным гербицидам, включающий определение зародышевой плазмы, придающей вышеупомянутую устойчивость к гербицидам, при котором вышеупомянутая устойчивая к гербицидам зародышевая плазма выводится из устойчивого к гербицидам растения сорго, и внедрение вышеупомянутой зародышевой плазмы в элитную линию растения сорго. В некоторых модификациях вышеупомянутое внедрение вышеупомянутой зародышевой плазмы в вышеупомянутую элитную линию растения сорго производится при помощи интрогрессии. В некоторых модификациях вышеупомянутое внедрение вышеупомянутой зародышевой плазмы в вышеупомянутую элитную линию растения сорго производится при помощи внедрения гетерологичного гена.
В некоторых модификациях настоящее изобретение обеспечивает гибрид сорго, при котором зародышевая плазма вышеупомянутого гибрида обладает полученной устойчивостью к одному или более ацетолактатсинтазным гербицидам и устойчивостью к одному или более соединениям одной или более гербицидных групп, которые не являются ацетолактатсинтазными ингибиторами.
В одной модификации настоящее изобретение обеспечивает способ определения линий растения сорго, устойчивых к ацетолактатсинтазным гербицидам, включающий обеспечение образца нуклеиновой кислоты растения сорго, обеспечение амплификационных праймеров для амплификации у растения сорго участка, соответствующего гену ацетолактатсинтазы, присутствующего в вышеупомянутом образце нуклеиновой кислоты, применение вышеупомянутых амплификационных праймеров к вышеупомянутому образцу нуклеиновой кислоты так, что происходит амплификация вышеупомянутого участка вышеупомянутого гена ацетолактатсинтазы, и определение растений сорго, устойчивых к ацетолоактатсинтазным гербицидам, основанное на присутствии одной или более мутаций, которые придают устойчивость к ацетолактатсинтазным гербицидам, присутствующим в вышеупомянутом амплифицированном образце нуклеиновой кислоты.
В одной модификации настоящее изобретение предусматривает семена сорго, в которых вышеупомянутая зародышевая плазма вышеупомянутых семян содержит мутантный ген ацетолактатсинтазы, такой, что мутация придает устойчивость к ингибированию ацетолактатсинтазными гербицидами. В некоторых модификациях зародышевая плазма вышеупомянутых семян сорго содержит такой мутантный ген ацетолактатсинтазы, как обнаружено в АТСС № РТА-7999. В некоторых модификациях мутантный ген ацетолактатсинтазы является функциональным фрагментом гена, как обнаружено в АТСС № РТА-7999, такого, что этот фрагмент гена кодирует фрагмент белка, достаточный для придания растению сорго устойчивости к ингибированию ацетолактатсинтазными гербицидами. В некоторых модификациях настоящее изобретение предусматривает саженцы сорго, которые выращивают из вышеупомянутых семян, и дополнительные части растения, которые содержат вышеупомянутые саженцы сорго, выращиваемые из вышеупомянутых семян.
В одной модификации настоящее изобретение предусматривает гибридные растения сорго, которые обладают всеми физиологическими и морфологическими качествами вышеупомянутого растения сорго, выращиваемого из вышеупомянутого семени сорго. В дополнительных модификациях настоящее изобретение предусматривает культуры ткани и регенерированные культуры ткани, которые происходят из вышеупомянутого семени сорго или вышеупомянутой части растения сорго, которая содержит мутацию в вышеупомянутом гене ацетолактатсинтазы, как обнаружено в АТСС № РТА-7999.
В некоторых модификациях настоящее изобретение предусматривает гибрид сорго, который содержит ген, который хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен гену устойчивости к ALS-гербициду такому, как обнаруженный в зародышевой плазме KSU 06MN8419, как обнаружено в АТСС № РТА-7999. В некоторых модификациях ген устойчивости к ALS-гербициду, который хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен гену устойчивости к ALS-гербициду такому, как обнаруженный в зародышевой плазме KSU 06MN8419, содержит одну или более замещенные аминокислоты Val531Ile и Trp545Leu, например, как обнаружено в SEQ ID № 1.
В одной модификации настоящее изобретение обеспечивает способ получения семени сорго, включающий скрещивание растения, содержащего мутантный ген ацетолактатсинтазы, как обнаружено в АТСС № РТА-7999, с ним самим или со вторым растением сорго и сбор вышеупомянутых семян от вышеупомянутого скрещивания. В некоторых модификациях методы получения вышеупомянутого семени сорго включают посадку родительской семенной линии сорго, при которой родительская семенная линия содержит зародышевую плазму, которая придает устойчивость к ацетолактатсинтазным гербицидам, с родительской линией-опылителем сорго, при которой зародышевая плазма вышеупомянутой семенной линии-опылителя содержит зародышевую плазму, которая придает устойчивость к ацетолактатсинтазным гербицидам, выращивание вышеупомянутых семенных растений и растений-опылителей сорго вместе, позволяющее вышеупомянутым родительским семенным растениям быть опыленными вышеупомянутыми родительскими растениями-опылителями, и сбор семян, которые получаются от вышеупомянутого опыления.
Описание фигур
Фигура 1 показывает двойную мутацию Val531Ile и Trp545Leu в ALS-гене сорго, которая, как обнаружено, связана с устойчивостью к ALS-гербициду.
Определения
Использованный в настоящей заявке термин «устойчивый» и «толерантный» используется для обозначения растений, например растений сорго, которые способны переносить условия (например, гербициды, такие как ALS-гербициды), вредные для других линий того же вида.
Использованный в настоящей заявке термин «сорт» является синонимом «разновидность» и используется для обозначения хлебных злаков, которые представляют собой группу схожих растений, которые по структурным признакам и характеристикам можно отличить от других сортов того же вида.
Использованный в настоящей заявке термин «гибрид» относится к плодам или потомству генетически неоднородного родительского растения или популяции, полученной в результате контролируемого перекрестного опыления, в противоположность к негибридному семени, полученному в результате естественного опыления.
Использованный в настоящей заявке термин «потомство» относится к поколениям растения, где родословную этого поколения можно установить вплоть до вышеуказанного растения.
Использованный в настоящей заявке термин «производное» устойчивого к гербицидам растения включает как потомство этого устойчивого к гербицидам растения, так и любой мутант, рекомбинант или генно-инженерную производную этого растении, или того же вида, или другого вида, где свойство(а) устойчивости к гербицидам первоначального устойчивого к гербицидам растения было перенесено на растение-производное.
Использованный в настоящей заявке термин «ткань растения» включает дифференцированные и недифференцированные ткани растений, включая те, которые присутствуют в корнях, побегах, листьях, пыльце, семенах и новообразованиях, а также в клетках в культуре (например, отдельные клетки, протопласты, зачатки, каллюсы и т.д.). Ткань растения может быть в растении, в органной культуре, культуре ткани или клеточной культуре.
Использованный в настоящей заявке термин «часть растения» в настоящей заявке относится к растительной структуре или ткани растения, например побегу, семяпочке, ткани, стручку, семени и клетке. В некоторых модификациях настоящего изобретения трансгенными растениями являются хлебные злаки.
Использованный в настоящей заявке термин «поколение F» и «дочернее поколение» относится любым следующим поколениям растений, клеток тканей или организмов после скрещивания двух родительских особей. Поколение, полученное в результате скрещивания двух родительских особей (т.е. двух родителей), является первым дочерним поколением (называемое «F1» или «F1»), что касается семени и его растения, в то время как то, что получается при скрещивании особей F1, является вторым дочерним поколением (называемое «F2» или «F2»), что касается семени и его растения.
Использованный в настоящей заявке термин «зародышевая плазма» относится к любому генетическому материалу растений, который содержит функциональные единицы наследственности. Термин «элитная зародышевая плазма» относится к растению, относящемуся к наследственному материалу с доказанным генетическим преимуществом.
Использованный в настоящей заявке термин «элитное растение» относится к любому растению, которое было получено в результате выведения и селекции по улучшенным агротехническим характеристикам. Например, элитные растения сорго, которые упоминаются в настоящей заявке, нелимитированно включают Тх430, Тх2737, Тх2783, 00MN7645, HP162, Wheatland, Tx3042, OK11, QL41 и Тх643 Bt линии сорго.
Использованный в настоящей заявке термин «признак» или «фенотип» относится к видимым и/или измеримым свойства организма. Например, настоящее изобретение описывает растения, которые являются устойчивыми к ALS-гербицидам.
Использованный в настоящей заявке термин «ALS-гербицид», также известный как AHAS-гербицид, относится к гербициду, который ингибирует активность фермента ацетолактатсинтазы (также известной как синтаза ацетогидроксикислот) у растения. Примеры ALS-гербицидов, описанные в настоящей заявке, нелимитированно включают сульфонилкарбомиды (SU), имидозолиноны (IMI), триазолопиримидины (ТР) и пиримидинилтиобензоаты.
Использованный в настоящей заявке термин «маркер» и «ДНК маркер» и «молекулярный маркер» по отношению к «селекционный маркер» относится к физиологическому или морфологическому признаку, который может быть определен как маркер для его собственной селекции или для селекции других признаков, тесно связанных с этим маркером. Например, таким маркером может быть ген или признак, который ассоциирован с устойчивостью к гербицидам, нелимитированно включая простой последовательный повтор (SSR), полиморфизм одного нуклеотида (SNP), генетические вставки и делеции и тому подобное.
Использованный в настоящей заявке термин «интрогрессировать», «интрогрессионный» и «интрогрессия» относится к традиционной (т.е. классической) методике перекрестного опыления, чтобы внедрить чужеродный генетический материал в линию скрещиваемой популяции. Например, настоящее изобретение предусматривает зерновые растения сорго, интрогрессированные мутантным геном ALS для устойчивости к гербицидам при помощи скрещивания двух поколений растений.
Использованный в настоящей заявке термин «дикий тип», при употреблении по отношению к гену, относится к функциональному гену обычно во всей популяции растений. Функциональный ген дикого типа - это ген, который наиболее часто наблюдается в популяции и, таким образом, арбитражно назначается «нормальной» формой гена или формой гена «дикого типа».
Использованный в настоящей заявке термин «модифицированный», или «мутантный», или «функциональный мутант», при употреблении по отношению к гену или генетическому продукту, относится к гену или генетическому продукту, соответственно, который демонстрирует изменения в последовательности и/или функциональных качествах (т.е. измененные свойства) по сравнению с геном или генетическим продуктом дикого типа. Таким образом, термин «модифицированный» или «мутантный», при употреблении по отношению к последовательности нуклеотидов, относится к последовательности нуклеиновых кислот, которая отличается по одному или более нуклеотидам от другой, как правило, родственной последовательности нуклеиновых кислот, а термин «функциональный мутант», при употреблении по отношению к полипептиду, кодированный при помощи вышеуказанной «модифицированной» или «мутантной» нуклеиновой кислоты, относится к белку или полипептиду, который сохраняет активность. В настоящей заявке мутантный белок ALS или соответствующий «функциональный мутант» является геном ALS, который сохраняет свою первоначальную активность создавать незаменимые аминокислоты. Кроме того, «модифицированная» последовательность нуклеотидов узнается как обнаруженная в вырожденном генетическом коде, как известно специалистам в данной области. Например, генетический код является вырожденным, так как там много вариантов, в которых разные кодоны обозначают одну и ту же аминокислоту; генетический код, в котором некоторые аминокислоты могут кодироваться более чем одним кодоном. Предполагается, что настоящее изобретение включает такую вырожденность (например, при которой гибрид сорго включает ген ALS, который хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен SEQ ID № 1), как обнаружено, например, в зародышевой плазме сорго.
Использованный в настоящей заявке термин «гетерологичный», при употреблении по отношению к гену или нуклеиновой кислоте, относится к гену, над которым производились какие-либо манипуляции.
Использованный в настоящей заявке термин «часть» или «функциональный фрагмент», при употреблении по отношению к белку (как в «во фрагменте данного белка» или «белковый фрагмент»), относится к фрагментам белка. Фрагменты могут варьироваться по размеру от четырех аминокислотных остатков до полной последовательности аминокислот минус одна аминокислота. В настоящем изобретении белковый фрагмент предпочтительно является функциональным так, что белковый фрагмент придает устойчивость к ингибированию ацетолактатсинтазными гербицидами приведенному растению.
Подробное описание изобретения
В одной модификации настоящее изобретение обеспечивает гены, кодирующие измененные гены и белки ALS. В некоторых модификациях настоящее изобретение предусматривает использование гербицидов, которые не ингибируют фермент ALS у сорго, который содержит измененный фермент ALS, чтобы снизить численность однодольных и двудольных сорных растений, присутствующих на ниве.
В одной модификации настоящее изобретение обеспечивает ALS-устойчивую зародышевую плазму растения сорго, полученную, например, в результате скрещивания двух родительских растений, дикого сорго «Tailwind или Tw» и линии элитного растения-опылителя Tx2783, собирая урожай поколения F1 и проводя последующие два обратных скрещивания (BC2F3:F4), получая в результате семена, сохраненные под АТСС №: РТА-7999, обозначенные KSU 06MN8419.
В одной модификации настоящее изобретение обеспечивает зародышевую плазму сорго, которая придает устойчивость к ингибированию ALS-гербицидами, и, кроме того, придает устойчивость к насекомым, к пятнистому стеблевому пилильщику Chilo partellus (Girijashankar et al., 2005, Plant Cell Rep. 24:513-522, полное описание которого включается в текст настоящей заявки). Например, гибрид сорго, зародышевая плазма которого содержит синтетический ген cryl Ac Bacillus thuringiensis (Bt), интогрессируется в линию сорго, зародышевая плазма которого придает устойчивость к ALS-гербицидам. Также осуществляется внедрение устойчивости к ALS-гербицидам и устойчивости к насекомым путем переноса генов растения в тот же гибрид сорго. Специалист в данной области обнаружит различные способы, описанные в настоящей заявке, которые можно применять для внедрения двух или более признаков устойчивости в один и тот же гибрид сорго.
В одной модификации ген устойчивости к ALS-гербицидам, как найдено у сорго, включающего ALS зародышевую плазму KSU 06MN8419, сохраненную под АТСС №: РТА-7999, вставляют в элитные разновидности сорго путем выведения растений и селекции, предусматривая, таким образом, развитие устойчивых к гербицидам у разновидностей сельскохозяйственной культуры, которые будут переносить использование ALS-ингибирующих гербицидов для контролирования сорных растений. В некоторых модификациях ген устойчивости к ALS-гербицидам хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен гену устойчивости к ALS-гербициду такому, как обнаружено в зародышевой плазме KSU 06MN8419. В некоторых модификациях ген устойчивости к ALS-гербицидам, который хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен гену устойчивости к ALS-гербицидам такому, как обнаружено в зародышевой плазме KSU 06MN8419, содержит одну или более замещенные аминокислоты Val531Ile и Trp545Leu. Внедрение признака устойчивости к гербицидам в вышеупомянутую разновидность хлебного злака позволяет использовать эти гербициды для контролирования однодольных и двудольных сорных растений, которые произрастают в присутствии этих сельскохозяйственных культур. В некоторых модификациях внедрение ALS-устойчивой зародышевой плазмы в элитные линии происходит способом интрогрессии или классической селекции. В некоторых модификациях внедрение гена устойчивости к ALS-гербицидам в элитные линии происходит путем гетерологичного переноса генов. В некоторых модификациях изобретение обеспечивает гибрид сорго, при котором хотя бы один предок вышеупомянутого гибрида сорго получен от ALS-устойчивой зародышевой плазмы, обозначенной KSU 06MN8419, сохраненной под АТСС №: РТА-7999. В некоторых модификациях настоящее изобретение обеспечивает гибрид сорго, при котором хотя бы один предок вышеупомянутого гибрида сорго содержит ген ацетолактатсинтазы, который обладает устойчивостью к ALS-гербицидам, как обнаружено в зародышевой плазме KSU 06MN8419, сохраненной под АТСС №: РТА-7999. В некоторых модификациях ген устойчивости к ALS-гербицидам, как обнаружено у гибрида сорго, хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен гену устойчивости к ALS-гербициду такому, как обнаружено в зародышевой плазме KSU 06MN8419. В некоторых модификациях ген устойчивости к ALS-гербицидам, который хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен гену устойчивости к ALS-гербицидам такому, как обнаружено в зародышевой плазме KSU 06MN8419, содержит одну или более замещенные аминокислоты Val531Ile и Trp545Leu.
В некоторых модификациях ALS-устойчивую зародышевую плазму интрогрессируют в элитную линию сорго при помощи способа классической селекции. Примеры способов классической селекции для сорго можно найти, например, у Sleper and Poehman, 2006, Breeding Field Crops, Fifth Edition, Blackwell Publishing, который в полном объеме включен в описание настоящей заявки в качестве ссылки.
В одной модификации ALS-устойчивую зародышевую плазму интрогрессируют в растение сорго, которое обеспечивает продукцию для потребления человеком. В некоторых модификациях ALS-устойчивую зародышевую плазму интрогрессируют в растение сорго, которое обеспечивает продукцию для домашнего скота (например, домашней птицы, крупного рогатого скота, свиней, овей и т.п.). В одной модификации ген устойчивости к ALS-гербицидам внедряют в геном растения путем переноса генов, при помощи векторов, известных специалистам в данной области технологий.
В одной модификации настоящее изобретение обеспечивает способы контролирования сорных растений на поле с хлебными злаками сорго. В некоторых модификациях контролирование сорных растений включает такое применение ALS-гербицида на поле с хлебными злаками сорго, что рост сорных растений ингибируется, но ущерб росту растений сорго не наносится. В некоторых модификациях применяемый ALS-гербицид относится к семейству сульфонилкарбомидных гербицидов, включающих один или более таких активных компонентов, как амидосульфурон, азимсульфурон, бенсульфурон-метил, хлоримурон-этил, хлорсульфурон, циносульфурон, циклосульфамурон, этаметсульфурон-метил, этоксисульфурон, флазасульфурон, флупирсульфурон-метил-натрий, форамсульфурон, галосульфурон-метил, имазолсульфурон, йодосульфурон-метил-натрий, мезосульфурон-метил, метсульфурон-метил, никосульфурон, оксасульфурон, примисульфурон-метил, пираксосульфурон-этил, римсульфурон, сульфометурон-метил, сульфосульфурон, тифенсульфурон-метил, триасульфурон, трибенурон-метил, трифлоксисульфурон-натрий, трифлусульфурон-метил, триофенсульфурон и тритосульфурон. В некоторых модификациях применяемый ALS-гербицид относится к семейству имидазолиноновых гербицидов, включающих один или более таких активных компонентов, как имазаметабенз-метил, имазамокс, имазапик, имазапир, имазаквин и имазетапир. В некоторых модификациях применяемый ALS-гербицид относится к семейству пиримидилтиобензоатных гербицидов, включающих один или более таких активных компонентов, как биспирибак-натрий, пирибензоксим, пирифталид, пириминобак-метил и пиритиобак-натрий. В некоторых модификациях применяемый ALS-гербицид относится к семейству триазолопиримидиновых гербицидов, включающих один или более таких активных компонентов, как клорансулам-метил, диклусолам, флорасулам, флуметсулам, метосулам и пенокссулам. В некоторых модификациях применяемый ALS-гербицид включает сочетание активных компонентов одного или более семейства ALS-гербицидов, как описано в настоящей заявке. Однако данная публикация не ограничивается использованными ALS-гербицидами, и специалист в данной области примет во внимание, что новые химические вещества, которые ингибируют фермент ALS, открываются постоянно.
В одной модификации настоящее изобретение обеспечивает использование трансгенов, содержащих гетерологичный ген, как ген, кодирующий мутантный белок ALS, для обеспечения селективного сельскохозяйственного признака устойчивости к ALS-гербицидам. В одной модификации трансген содержит мутантный ген ALS, как обнаружено в зародышевой плазме KSU 06MN8419, сохраненной под АТСС №: РТА-7999. В некоторых модификациях ген ALS хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен мутантному гену ALS такому, как обнаружено в зародышевой плазме KSU 06MN8419. В некоторых модификациях мутантный ген ALS, который хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен мутантному гену такому, как обнаружено в зародышевой плазме KSU 06MN8419, содержит одну или более замещенные аминокислоты Val531Ile и Trp545Leu.
Striga, широко известное как witchweed, род растений-паразитов травянистых и однодольных зерновых культур, таких как кукуруза, пшеница и сорго. Сильное засорение Striga поля с хлебными злаками повреждает растения-хозяева, вызывая замедление роста, хлороз, увядание и, в конечном счете, смерть растения-хозяина, особенно, в его естественной среде обитания, где может быть недостаточно воды. Родиной Striga являются полупустынные и степные области Африки и Азии, однако оно может разрастаться за пределы своей естественной области обитания, и данные засорения присутствуют на сельскохозяйственных площадях в Соединенных Штатах. Засорения широко распространены в Африке, поэтому Striga является постоянной проблемой для фермеров в полупустынных и полупустынных областях к югу от Сахары, где сорго является основной продовольственной культурой. Растение-паразит тяжело истребить, так как его семена лежат в состоянии покоя в почве вплоть до десяти лет до прорастания.
Биологические сигналы, стимулирующие прорастание семян Striga, включают стимуляцию выделениями корней растения-хозяина. Семена Striga прорастают при помощи направления к хозяину инфицирующих структур, которые прикрепляются к корням хозяина и инвазируют сосудистую систему хозяина, отнимая, таким образом, у хозяина воду, минеральные вещества и углеводы. Как только появился первый контакт с хозяином, инициирование Striga нарастает, и начинают формироваться дополнительные корни, а отъем у растения-хозяина возрастает. В естественных местах обитания, где недостаточно воды, Striga эффективно выкачивает из растения-хозяина минеральные вещества и воду.
Паразитизм Striga ингибируют ALS-гербицидами. Настоящее изобретение не ограничивается конкретным механизмом. На самом деле, понимание механизма не требуется, чтобы применять на практике настоящее изобретение. Несмотря на это, заражение корней сорго Striga устраняется при помощи покрытия семян сорго ALS-гербицидами перед посадкой. В одной модификации семена гибрида сорго содержат зародышевую плазму, которая придает устойчивость к ALS-гербицидам вышеупомянутому гибриду сорго, если покрыть одним или более ALS-гербицидами перед посадкой. В некоторых модификациях покрытые семена сажают на сельскохозяйственную площадь, где произрастает вид растения-паразита Striga. В некоторых модификациях гибридные растения сорго, которые вырастают из вышеупомянутых покрытых семян, устойчивы к инфекции Striga.
В одной модификации настоящее изобретение предусматривает гибрид сорго, зародышевая плазма которого придает устойчивость к ALS-гербицидам и устойчивость к одному или более дополнительным гербицидам из одной или более различных групп гербицидов. Например, дополнительные группы гербицидов, используемых для ингибирования роста сорных растений, нелимитированно включают ингибиторы синтеза липидов (например, арилоксифеноксипропионаты, циклогексанодеионы, бензофураны, хлоро-карбоновые кислоты, фосфородитиоаты, тиокарбаматы), ингибиторы фотосинтеза в фотосистеме II (например, фенил-карбаматы, пиридазиноны, триазины, триазиноны, триазолиноны, урацилы, амиды, карбамиды, бензотиадиазиноны, нитрилы, фенил-пиридины), ингибиторы фотосинтеза в фотосистеме I (например, бипиридилиумы), ингибиторы протопорфириноген оксидазы (например, дифенилэфиры, N-фенилфталимиды, оксадиазолы, оксизолидиндионы, фенилпиразолы, пиримидиндионы, тиадиазолы), ингибиторы биосинтеза каротиноидов (например, пиридазиноны, пиридинкарбоксамиды, изоксазолидиноны, триазолы), ингибиторы 4-гидроксифенил-пируват-диоксигеназы (например, каллистемоны, изоксазолы, пиразолы, трикетоны), ингибиторы EPSP-синтазы (например, глицины), ингибиторы глютамин синтетазы (например, фосфиновые кислоты), ингибиторы дигидроптероат синтазы (например, карбаматы), ингибиторы сборки микротрубочек (например, бензамиды, бензойная кислота, динитроанилины, фосфороамидаты, пиридины), ингибиторы клеточного движения (например, ацетамиды, хлороацетамиды, оксиацетамиды), ингибиторы синтеза клеточной стенки (например, нитрилы, триазолокарбоксамиды) и ингибиторы транспорта ауксина (например, фталаматы, семикарбазоны).
Классическая селекция сорго
Полевые культуры классически выводили при помощи методики, которая позволяла получить выгоду от способа(ов) опыления растений. Растение считают «самоопыляющимся», если пыльца от одного растения может переноситься на тот же самый или другой цветок, тогда как растения считают «перекрестноопыляющимся», если пыльца должна прилететь с цветка другого растения, чтобы произошло опыление.
Самоопыляющиеся растения и отбираемые во многих поколениях, по большей части, если не все, становятся гомозиготными по их генным локусам, производя, таким образом, единообразную популяцию гомозиготных потомков. Скрещивание между двумя гомозиготными растениями разных сред или двух разных гомозиготных линий произведет единообразную популяцию гибридных растений, которая, более чем вероятно, будет гетерозиготной по некоторому количеству генных локусов. Скрещивание двух родителей, каждый из которых гетерозиготен по некоторому числу генных локусов, произведет поколение гибридных растений, которые генетически отличаются и не единообразны.
Растения сорго являются самоопыляющимися растениями, но они также могут быть отобраны при помощи перекрестного опыления. Развитие гибридов сорго нуждается в развитии родителей-опылителей (восстановителей фертильности) и семенных родительских инцухтов при помощи использования цитоплазматической мужской системы восстановления стерильности-фертильности, скрещивая семенных родителей и родителей-опылителей и оценки скрещиваний. Программы племенного отбора комбинируют желаемые признаки; в данной публикации желаемым признаком является устойчивость растения к ALS-гербицидам. Этот признак вводится в селекционный пул одной или более линий, так что новые инцухтированные линии создаются при помощи скрещивания с последующей селекцией растений с желаемым признаком, за которыми следуют дополнительные скрещивания и т.д. Новые инцухты скрещивают с другими инцухтированными линиями (например, элитными линиями растений, как те, которые описаны в настоящей заявке).
Племенной отбор начинается со скрещивания двух генотипов, таких как Tailwind и элитная линия сорго (например, Тх430, Тх2737, Тх2783, 00MN7645, НР162, Wheatland, Tx3042, OK11, QL41, Tx643 и Bt сорго). Если два первичных родителя не обеспечивают всех желаемых свойств, тогда в селективную популяцию могут быть подключены другие источники. Например, если требуется получить такой гибрид, что желательны как устойчивость к ALS-гербицидам, так и к другим гербицидам, описанным в настоящей заявке, тогда растение со всеми этими качествами можно скрестить при помощи классической селекционной методики. Способом племенного отбора высшие растения самоопыляют и отбирают в последовательных поколениях. В последовательных поколениях в результате самоопыления и селекции гетерозиготное состояние уступает гомозиготным линиям. Обычно на практике в способе племенного отбора самоопыление и селекция применяются к пяти или более поколениям (например, S1, S2, S3, S4, S5 и т.д.).
Обратное скрещивание используют для улучшения растительной линии. Обратное скрещивание осуществляет перенос специфического желаемого признака от одного источника к другому, у которого этот признак отсутствует. Это осуществляется, например, при помощи скрещивания донора (например, Tailwind) с инбредной линией (например, Тх2783 элитная линия опылителя, как описано в настоящей заявке). Потомство от этого скрещивания затем обратно скрещивается (т.е. обратное скрещивание) с элитной инбредной линией с последующим отбором по желаемому признаку (например, устойчивости к к ALS-гербицидам) в полученном в результате потомстве. Следующие пять или более поколений, полученных путем обратного скрещивания с отбором потомства по желаемому признаку, обычно гетерозиготны по локусу (локусам), контролирующим желаемый фенотип, но будут похожи на элитного родителя по другим генетическим признакам. Последнее обратное скрещивание обычно происходит путем самоопыления, чтобы дать чистое отобранное потомство для передачи генов.
В текущих гибридных интрогрессивных программах селекции сорго развивают новые родительские линии, чтобы они были либо семенными родительскими линиями (например, Wheatland, Tx3042, N223, 01MN1589, 03MN954, OK11, QL41 и Тх643), либо линиями родителей-опылителей (например, Tx430, Tx2737, Tx2783, R45, 00MN7645 и НР162), в зависимости от того, содержат ли они гены-восстановители фертильности или нет; у семенных родительских линий нет генов-восстановителей фертильности, и конкретные из них обладают цитоплазматической мужской стерильностью (также известные как растения «А-линии»), а другие обладают цитоплазматической мужской фертильностью (также известные как растения «В-линии»), при этом линии родителей-опылителей не обладают мужской стерильностью и содержат гены-восстановители фертильности (также известные как растения «R-линии»). Семенные родительские линии обычно создают так, чтобы они обладали цитоплазматической мужской стерильностью так, что пыльники у этих растений минимальны или их нет совсем, таким образом, требуется перекрестное опыление. Семенные родительские линии будут только производить семя, а цитоплазма передается только через яйцо. Пыльца для перекрестного опыления предоставляется линиями родителей-опылителей, которые содержат гены, необходимые для полного восстановления фертильности у гибридов F1, а скрещивание соединяет с семенным родителем, обладающим цитоплазматической мужской стерильностью, чтобы произвести высокоурожайный простой гибрид с хорошим качеством зерен. Примеры R-линии и В-линии, которые нашли применение в настоящем изобретении, включают, но не ограничиваются описанными в Таблице 1.
Обычно эта система восстановления цитоплазматической мужской стерильности-фертильности применяется для получения гибридных семян при помощи высаживания блоков рядов, обладающих цитоплазматической мужской стерильностью (семенных родительских) растений и блоков рядов растений-восстановителей фертильности (родителей-опылителей), так что семенные родительские растения опыляются ветром пыльцой от растения родителя-опылителя. Этот процесс производит жизнестойкий простой гибрид, с которого потребители собирают урожай и сажают.
Обладающие цитоплазматической мужской стерильностью семенные родительские растения также могут быть получены при помощи генетической селекции рецессивных ядерных генов мужской стерильности в определенную популяцию, однако, обычно, система восстановления цитоплазматической мужской стерильности-фертильности является той системой, которая используется для селекции гибридного сорго. Sleper and Poehlman, 2006, Breeding Field Crops, Fifth Ed., Blackwell Publishing обеспечивает хороший отчет данных процедур для селекции сорго и в полном объеме включен в описание настоящей заявки.
Настоящее изобретение не ограничивается перечисленными элитными родительскими линиями сорго, и специалист в данной области отметит, что любая элитная линия сорго является в равной степени подходящей для составов и способов, описанных в настоящей заявке.
Растительная трансгеника
Гетерологичные гены, предназначенные для экспрессии в растениях, сначала собирают в экспрессионные векторы, содержащие гетерологичный ген и подходящие транскрипционные и трансляционные контрольные элементы, такие способы хорошо известны специалистам в данной области. Способы включают технологию in vitro ДНК-гибридизации, технологию синтеза и генетической рекомбинации in vivo. Примеры технологий подробно описаны в данной области (см., например, Sambrook et al. (1989) Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Press, Plainview, N.Y. и Ausubel, F.M. et. al. (1989) Current Protocols in Molecular Biology, John Wiley & Sons, New York, N.Y., включенные в описание настоящей заявки в качестве ссылки).
Как правило, эти векторы содержат последовательность нуклеиновых кислот, кодирующую гетерологичный ген, функционально связанный с промотором, и другие регуляторные последовательности (например, энхансеры, сигналы полиаденилирования и т.д.), необходимые для экспрессии в растении.
Промоторы неограниченно включают конститутивные промоторы, тканево-, органно- и эволюционно-специфичные промоторы и индуцируемые промоторы. Примеры промоторов неограниченно включают конститутивный промотор 35S вируса мозаики цветной капусты; индуцируемый нанесением раны промотор из томата, лейцин аминопептидазу (Chao et al., 1999, Plant Physiol 120:979-992, которая в полном объеме включена в описание настоящей заявки в качестве ссылки); химически-индуцируемый промотор из табака, Patigenesis-Related 1 (индуцируемый при помощи салициловой кислоты и S-метиловым эфиром бензотиадиазол-7-карботиокислоты); хит-шок промотор (Патент США № 5,187,267, который в полном объеме включен в описание настоящей заявки в качестве ссылки); и семя-специфичные промоторы.
Модуль экспрессии может также включать любые последовательности, необходимые для экспрессии мРНК. Такие последовательности нелимитированно включают терминаторы транскрипции, энхансеры, такие как интроны, вирусные последовательности и последовательности, предназначенные для таргетинга генов к специфическим органеллам и клеточным компартментам.
Множество терминаторов транскрипции доступно для использования в экспрессии последовательностей при помощи таких промоторов, как описанные в настоящей заявке. Терминаторы транскрипции ответственны за завершение транскрипции после транскрипта и его правильное полиаденилирование. Подходящие терминаторы транскрипции, для которых известно, что они функционируют в растениях, нелимитированно включают терминатор CaMV 35S, терминатор tml, терминатор pea rbc E9 и терминаторы нопалин и октопин синтаз (Odell et al., 1985, Nature 313:810; Rosenberg et al., 1987, Gene, 56:125; Guerineau et al., 1991, Mol. Gen. Genet. 262:141; Proudfoot, 1991, Cell, 64:671; Sanfacon et al., 1990, Genes Dev. 5:141; Mogen et al., 1990, Plant Cell, 2:1261; Munroe et al., 1990, Gene, 91:151; Ballas et al., 1989, Nucleic Acids Res. 17:7891; Joshi et al., 1987, Nucleic Acid Res., 15:9627, которые в полном объеме включены в описание настоящей заявки в качестве ссылок).
В некоторых модификациях конструкты для экспрессии интересующего гетерологичного гена включают одну или более последовательности, которые, как было обнаружено, усиливают экспрессию гена изнутри транскрипционной единицы. Эти последовательности могут использоваться в сочетании с интересующей нас последовательностью нуклеиновых кислот, чтобы увеличить экспрессию у растений. Было показано, что различные интронные последовательности усиливают экспрессию, особенно в клетках однодольных. Интронные последовательности традиционно вставляют в растительные трансформирующие векторы, обычно внутрь нетранслируемой лидерной последовательности.
В некоторых модификациях конструкт для экспрессии интересующей гетерологичной последовательности нуклеиновых кислот также включает регуляторную последовательность, такую как сигнал ядерной локализации (Kalderon et al., 1984, Cell 39:499; Lassner et al., 1991, Plant Molecular Biology 17:229), растительная трансляционная консенсусная последовательность (Joshi, 1987, Nucleic Acids Research 15:6643), интрон (Luehrsen and Walbot, 1991, Mol Gen. Genet. 225:81) и тому подобное, функционально связанные с последовательностью нуклеиновых кислот, кодирующей гетерологичный ген.
В приготовление конструкта, содержащего последовательность нуклеиновой кислоты, кодирующую гетерологичный ген, или кодирующую последовательность, сконструированную, чтобы снизить экспрессию гетерологичного гена, различные фрагменты ДНК могут использоваться так, чтобы обеспечить последовательности ДНК желаемой ориентации (например, смысловые и антисмысловые), а соответственно, и желаемой рамкой считывания. Например, адапторы и линкеры могут применяться, чтобы объединить фрагменты ДНК, или могут использоваться другие манипуляции, чтобы обеспечить удобные сайты рестрикции, удаление избыточной ДНК, удаление сайтов рестрикции и тому подобное. Для этой цели предпочтительно применяется in vitro мутагенез, восстановление при помощи праймеров, рестрикция, гибридизация, резекция, сшивание и тому подобное, при этом имеют место вставки, делеции или замещения (например, транзиции и трансверсии).
Многочисленные трансформирующие векторы подходят для трансформации растений. Выбор вектора для использования будет зависеть от предпочтительного способа трансформации и видов-мишеней трансформации. Для конкретных видов-мишеней предпочтительны разные антибиотические или гербицидные селекционные маркеры. Селекционные маркеры, традиционно используемые при трансформации, включают ген nptII, который придает устойчивость к канимицину и родственным антибиотикам (Messing and Vierra, 1982, Gene 19:259; Bevan et al., 1983, Nature 304:184, которые включены в описание настоящей заявки в качестве ссылок), ген bar, который придает устойчивость к гербициду фосфинотрицину (White et al., 1990, Nucl Acids Res. 18:1062; Spencer et al., 1990. Theor. Appl. Genet. 79:625, которые в полном объеме включены в описание настоящей заявки в качестве ссылок), ген hph, который придает устойчивость к антибиотику гигромицину (Blochlinger and Diggelmann, 1984, Mol. Cell. Biol. 4:2929, которые в полном объеме включены в описание настоящей заявки в качестве ссылок), и ген dhfr, который придает устойчивость к метотрексату (Bourouis et al., 1983 EMBO J., 2:1099, которая в полном объеме включена в описание настоящей заявки в качестве ссылки).
В некоторых модификациях плазмидный вектор Ti (T-ДНК) адаптирован для использования в Agrobacterium опосредованном процессе трансфекции, как в Патенте США 6,369,298 (сорго) и Патентах США 5,981,839, 6,051,757, 5,981,840, 5 824,877 и 4,940,838, которые в полном объеме включены в описание настоящей заявки в качестве ссылок. Конструирование рекомбинантных плазмид Ti и Ri в основном следует способам, типично используемым с более обычными векторами, такими как pBR322. Дополнительное использование может основываться на вспомогательных генетических элементах, обнаруженных вместе с исходными плазмидами и иногда собранными из чужеродных последовательностей. Они могут нелимитированно включать структурные гены для устойчивости к антибиотикам в качестве селекционных генов.
Сейчас применяются две системы рекомбинантных плазмидных векторов Ti и Ri. Первая система называется «коинтегрирующая» система. В этой системе челночный вектор, содержащий интересующий ген, при помощи генетической рекомбинации вставляют в неонкогенную плазмиду Ti, которая содержит как действующий в цис-положении, так и действующий в транс-положении элементы, необходимые для трансформации растения, такие как, например, в челночном векторе pMLJ1, и неонкогенную плазмиду Ti pGV3850. Использование T-ДНК в качестве фланкирующей области в конструкте для интеграции в Ti- или Ri-плазмиду было описано в ЕРО № 116,718 и РСТ Публикациях №№ WO 84/02913, 02919 и 02920; Herrera-Estrella, 1983, Nature 303:209-213; Fraley et al., 1983, Proc. Natl. Acad. Sci, USA 80:4803-4807; Horsch et al., 1984, Science 223:496-498; и DeBlock et al., 1984, EMBO J. 3:1681-1689, которые в полном объеме включены в описание настоящей заявки в качестве ссылок.
Вторая система называется «бинарной» системой, в которой используются две плазмиды, и интересующий ген вставляют в челночный вектор, содержащий элементы, действующие в цис-положении, необходимые для трансформации растений. Другие полезные функции обеспечиваются в транс при помощи неонкогенной Ti-плазмиды, как показано на примере челночного вектора pBIN19 и неонкогенной Ti-плазмиды PAL4404. Некоторые из этих векторов имеются в продаже.
В некоторых модификациях интересующая последовательность нуклеиновой кислоты направлена на определенный локус в растительном геноме. Сайт-специфическая интеграция этой интересующей последовательности нуклеиновой кислоты в геном растительной клетки может достигаться, например, при помощи гомологичной рекомбинации с использованием полученных из Agrobacterium последовательностей. Обычно растительные клетки инкубируют со штаммом Agrobacterium, который содержит таргетный вектор, в котором последовательности, которые гомологичны последовательности ДНК внутри локуса-мишени, фланкируются последовательностями перенесенной-ДНК (Т-ДНК) Agrobacterium, как описано ранее (Патент США № 5,501,967, который в полном объеме включен в описание настоящей заявки в качестве ссылки). Специалист в данной области знает, что гомологичная рекомбинация может быть достигнута использованием таргетных векторов, которые содержат последовательности, которые гомологичны любой части растительного гена-мишени, который принадлежит к регуляторным элементам гена или кодирующим участкам гена. Гомологичная рекомбинация может быть достигнута на любом участке растительного гена, при условии, что известна последовательности нуклеиновых кислот участков, фланкирующая сайт-мишень. Agrobacterium tumefaciens является обыкновенной почвенной бактерией, которая вызывает заболевание корончатый галл при помощи переноса части своей ДНК в растение-хозяина. Перенесенная ДНК (Т-ДНК) стабильно интегрируется в геном растения, где ее экспрессия приводит к синтезу растительных гормонов и, таким образом, к опухолевому росту клеток. Предполагаемый макромолекулярный комплекс формируется в процессе переноса Т-ДНК из бактериальной клетки в растительную клетку.
В некоторых модификациях нуклеиновые кислоты, как описанные в настоящей заявке, утилизируют, чтобы сконструировать векторы, полученные из (+)РНК вирусов растений (например, вирус мозаичной болезни костра, вирус мозаичной болезни табака, вирус мозаичной болезни люцерна, вирус мозаичной болезни огурца, вирус мозаичной болезни томата и их сочетаниями и гибридами). Обычно интересующий гетерологичный полинуклеотид можно экспрессировать с этих векторов в качестве химерного белка (например, химерных белков оболочки) или с его собственного субгеномного промотора или другого промотора. Способы конструирования и использования таких вирусов описаны в Патентах США №№ 5,846,795; 5,500,360; 5,173,410; 5,965,794; 5,977,438 и 5,866,785, которые в полном объеме включены в описание настоящей заявки в качестве ссылок.
В некоторых модификациях интересующая нас гетерологичная последовательность нуклеиновой кислоты, включающая мутантный трансген ALS, как обнаруженный в зародышевой плазме KSU 06MN8419, сохраненной под АТСС №: РТА-7999, вводится прямо в растение. В некоторых модификациях этот мутантный трансген ALS хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен гену ALS такому, как обнаружено в зародышевой плазме KSU 06MN8419. В некоторых модификациях мутантный ген ALS, который хотя бы на 70% гомологичен, хотя бы на 80% гомологичен, хотя бы на 85% гомологичен, хотя бы на 90% гомологичен, хотя бы на 95% гомологичен, хотя бы на 97% гомологичен или хотя бы на 99% гомологичен гену устойчивости к ALS-гербициду такому, как обнаружено в зародышевой плазме KSU 06MN8419, содержит одну или более замещенных аминокислот Val531Ile и Trp545Leu. Один вектор, используемый для технологии прямого переноса генов, в сочетании с селекцией при помощи гербицида Basta (или фосфинотрицин) представляет собой модифицированную версию плазмиды pCIB246 с промотором CaMV 35S, функционально связанным с геном E. Coli GUS и терминатором транскрипции CaMV 35S (WO 93/07278, включенный в описание настоящей заявки в качестве ссылки).
Если последовательность нуклеиновых кислот, кодирующая гетерологичный ген, функционально связана с соответствующим промотором и вставлена в подходящий вектор для использования в определенной технологии трансформации (например, один из векторов, описанных выше), описанная выше рекомбинантная ДНК может быть введена в растительную клетку несколькими известными в данной области способами. Специалисты в данной области оценят, что выбор метода зависит от типа растения, являющегося мишенью трансформации. В некоторых модификациях вектор сохраняют эписомально. В некоторых модификациях вектор интегрируют в геном. В некоторых модификациях для интродукции вектора в растительную клетку используется прямая трансформация в геном пластид (например, см. Патенты США №№ 5,451,513; 5,545,817; 5,545,818; РСТ публикация WO 95/16783, которые в полном объеме включены в описание настоящей заявки в качестве ссылок).
Базовая технология трансформации хлоропласта затрагивает вставку участков клонированной ДНК пластиды, фланкирующей селективный маркер вместе с нуклеиновой кислотой, кодирующей интересующие нас последовательности, в подходящую ткань-мишень (например, биолистика или трансформация протопласта хлоридом кальция или ПЭГ). От 1 до 1,5 фланкирующих участков, называемых таргетными последовательностями, способствуют гомологичной рекомбинации при помощи генома пластиды, и допускают, таким образом, замещение или модификацию специфических участков пластома. Сначала точечные мутации в 16S рРНК и rps12 генов хлоропласта, придающие устойчивость к спектиномицину и/или стрептомицину, используют в качестве селективных маркеров для трансформации (Svab et al., 1990, Proc. Natl. Acad. Sci., 87:8526; Staub and Maliga, 1992, Plant Cell, 4:39, которые включены в описание настоящей заявки в качестве ссылок). Присутствие сайтов клонирования между этими маркерами позволяет вводить в вектор, мишенью которого являются пластиды, чужеродные молекулы ДНК (Staub and Maliga, 1993, EMBO J., 12:601). Существенные увеличения частоты трансформации получают при помощи замещения рРНК или р-белка рецессивных генов устойчивости к антибиотикам доминантными селективными маркерами, бактериальным геном aadA, кодирующим спектриномицин-детоксифицирующий фермент аминоглосозид-3'-аденилтрансферазу (Svab and Maliga, 1993, Proc. Natl. Acad. Sci. 90:913). В данной области известны другие селективные маркеры, пригодные для трансформации пластид, и они охвачены в рамках настоящего изобретения. Получают растения, гомоплазмические по геному пластид, содержащему две последовательности нуклеиновых кислот, разделенных при помощи промотора по настоящему изобретению, и они предпочтительно допускают высокую экспрессию РНК, кодированных при помощи молекулы ДНК.
В одной модификации векторы, используемые на практике в настоящем изобретении, микроинъецируют прямо в растительные клетки (Crossway, 1985, Mol. Gen. Genet, 202:179). В некоторых модификациях вектор трансфицируют в растительную клетку, используя полиэтиленгликоль (Krens et al., 1982, Nature, 296:72; Crossway et al., 1986, BioTechnique, 4:320), слияние протопластов с другими объектами, такими как миниклетки, клетки, лизосомы или другие способные к слиянию тела с липидной поверхностью (Fraley et al., 1982, Proc. Natl. Acad. Sci., USA, 79:1859); и трансформацию протопластов (ЕР 0 292 435); прямой перенос генов (Paszkowski et al., 1984, EMBO J., 3:2717; Hayashimoto et al., 1990, Plant Physiol. 93:857).
В некоторых модификациях вектор может быть введен в растительные клетки при помощи электропорации (Fromm et al., 1985, Proc. Natl. Acad. Sci. USA 82:5824; Riggs et al., 1986, Proc. Natl. Acad. Sci. USA 83:5602). В этой технологии растительные протопласты электропорируют в присутствии плазмид, содержащих генетический конструкт. Электрические импульсы поля высокой напряженности обратимо пермеабилизируют биомембраны, позволяя введение плазмид. Электропорированные растительные протопласты восстанавливают клеточную стенку, делятся и формируют растительный каллюс. В дополнение к прямой трансформации в некоторых модификациях векторы, содержащие последовательность нуклеиновых кислот, кодирующую гетерологичный ген, трансфицируют, используя Agrobacterium опосредованную трансформацию (Hinchee et al., 1988, Biotechnology, 6:915; Ishida et al., 1996, Nature Biotechnology 14:745, которые включены в описание настоящей заявки в качестве ссылок). Agrobacterium является репрезентативным родом граммотрицательного семейства Rhizobiaceae. Его разновидности ответственны за такие опухоли растений, как корончатый галл и заболевание «бородатый корень». В дедифференцированной ткани, что характерно для опухолей, продуцируются и катаболизируются аминокислотные производные, известные как опины. Бактериальные гены, ответственные за экспрессию опинов, являются подходящим источником контрольных элементов для химерных полигенных экспрессирующихся кластеров. Гетерологичные генетические последовательности (например, последовательности нуклеиновых кислот, функционально связанные с промотором по настоящему изобретению) могут быть вставлены в соответствующие растительные клетки при помощи Ti-плазмиды Agrobacterium tumerfaciens (описанной ранее). Ti-плазмида передается растительной клетке посредством инфекции при помощи Agrobacterium tumerfaciens, и стабильно интегрируется в растительный геном (Schell, Science, 237:1176). Виды, которые подвержены инфекции Agrobacterium, могут быть трансформированы in vitro. Способы трансформации для получения трансгенных растений сорго с использованием Agrobacterium-опосредованной трансформации, приведены в Патенте США 6,369,298.
В некоторых модификациях вектор вводят посредством баллистического ускорения частиц (U.S. Patent No. 4,945,050; Casas et al., 1993, Proc. Natl. Acad. Sci. USA 90:11212, которые в полном объеме включены в описание настоящей заявки в качестве ссылок).
В некоторых модификациях после селекции материала трансформированных растений, которые могут экспрессировать гетерологичный ген, кодирующий гетерологичный белок или его варианты, целые растения регенерируются. Регенерация растения из культивируемых протопластов описана Evans et al., Handbook of Plant Cell Cultures, Vol. 1: (MacMillan Publishing Co. New York, (1983); Vasil I.R. (ed.), Cell Culture and Somatic Cell Genetics of Plants, Acad. Press, Orlando, Vol. I, (1984) и Vol. III, (1986), которые в полном объеме включены в описание настоящей заявки в качестве ссылок. Известно, что многие растения могут быть регенерированы из культивируемых клеток или тканей, нелимитированно включая все основные виды сахарного тростника, сахарной свеклы, хлопка, фруктовых и других деревьев, бобовых и овощей, и однодольных (например, растений, описанных выше). Способы регенерации варьируются от вида к виду, но в основном сначала обеспечивается суспензия трансформированных протопластов, содержащих копии гетерологичного гена. Формируется ткань каллюса, а побег может быть индуцирован из каллюса и впоследствии укоренен.
В качестве альтернативы формирование зачатка может быть индуцировано из суспензии протопластов. Эти зачатки созревают и формируют взрослые растения. Культуральная среда обычно содержит различные аминокислоты и гормоны, такие как ауксин и цитокинины. Побеги и корни в норме развиваются одновременно. Эффективная регенерация будет зависеть от среды, от генотипа и от истории культуры. Репродуктивность регенерации зависит от контролирования этих переменных.
Примеры
Пример 1 - Устойчивость к гербицидам в генотипе сорго
Семена сорго с диким генотипом, которые, как было показано, имели устойчивый к ALS-гербициду фенотип, обозначенные «Tailwind», и сорго с чувствительным к гербицидам генотипом, обозначенные 90SN7, высаживали в 12-литровые горшки в теплице. Несколько растений каждого типа опрыскивали либо 1) Lightring Herbicide (сочетание Имазетапира и Имазапира) в количестве 2,56 унций на акр-1 (2х количество гербицида), либо 2) Steadfast ® Herbicide (DuPont™; сочетание Никосульфурона и Римсульфурона) в количестве 1,50 унций на акр-1 (2х количество гербицида), либо 3) сочетание Lightring Herbicide в количестве 2,56 унций на акр-1 и Steadfast ® Herbicide в количестве 1,50 унций на акр-1. Для каждой обработки растения Tailwind не показывали существенных повреждений через 12 дней применения гербицида, в то время как растения 90SN7 погибли, показывая, что Tailwind обладали перекрестной устойчивостью к IMI (Lightring Herbicide) и SU (Steadfast ® Herbicide) классам ALS-ингибирующих гербицидов.
Пример 2 - Скрещивания Tailwind с элитными родительскими линиями сорго
Tailwind скрещивали с различными элитными родителями, включая Тх430 и Wheatland. Популяции F2, полученные от скрещиваний с этими родителями, оценивали при помощи сегрегационного анализа, чтобы определить количество генов, участвующих в экспрессии толерантности. Популяции растений выращивали в теплице и опрыскивали 1х и 3х количествами Accent® Herbicide (DuPont™Никосульфурон), Option® Herbicide (BayerCropScience; Форамсульфурон) и Steadfast ® Herbicide. Подсчет в популяции живых/мертвых особей позволял провести генетический анализ.
Сегрегационный анализ показал один основной, частично доминантный ген в популяции, полученной от Тх430 для обработки каждым гербицидом. Похожий анализ популяций, полученных от скрещиваний с Wheatland, показал один основной, частично доминантный ген, а также еще потенциально два или три гена-модификатора, которые оказывали влияние на относительную экспрессию признака толерантности.
Попытки селекции растений были инициированы при помощи обратного скрещивания признака толерантности с коммерчески значимыми элитными родителями-опылителями сорго Тх430, Тх2737, Тх2783, 00MN7645 и НР162, а также с коммерчески значимыми элитными семенными родителями сорго, включая Wheatland, Тх3042, ОК11, QL41 и Тх643, с селекцией на толерантность к гербицидам в каждом поколении. Полученное в результате скрещивания Tailwind с Тх2783 (BC2F3:F4) семя было сохранено в АТСС для публичного доступа.
Пример 3 - Генетическое секвенирование для гена ALS-устойчивости
Попытки генетического секвенирования были инициированы, чтобы определить, существует ли мутация таргетного сайта для фенотипа толерантности к гербицидам. Поиск последовательности, произведенный с использованием National Institute of Health's Basic Local Alignment Search Tool (BLAST) в Институте исследования генома (TIGR) в базе данных Собрания растительных транскриптов, определил Набор транскриптов, представляющий ALS-ген сорго (ТА3960_4558; SEQ IN NO: 1). Были созданы амплификационные праймеры для полимеразной цепной реакции (ПЦР), F4r-CACATCACCCTTGTACCAGCTC (SEQ ID NO: 3) и B5-GATTGTGCACATTGATATTGATCC (SEQ ID NO: 4), чтобы амплифицировать участки гена сорго, аналогичные участкам гена AHAS Arabidopsis thaliana, чтобы повлиять на экспрессию толерантности к ALS-гербицидам (A. thaliana: Ala122, Pro197, Ala205, TRp574 и Ser653; Tan et al., 2005).
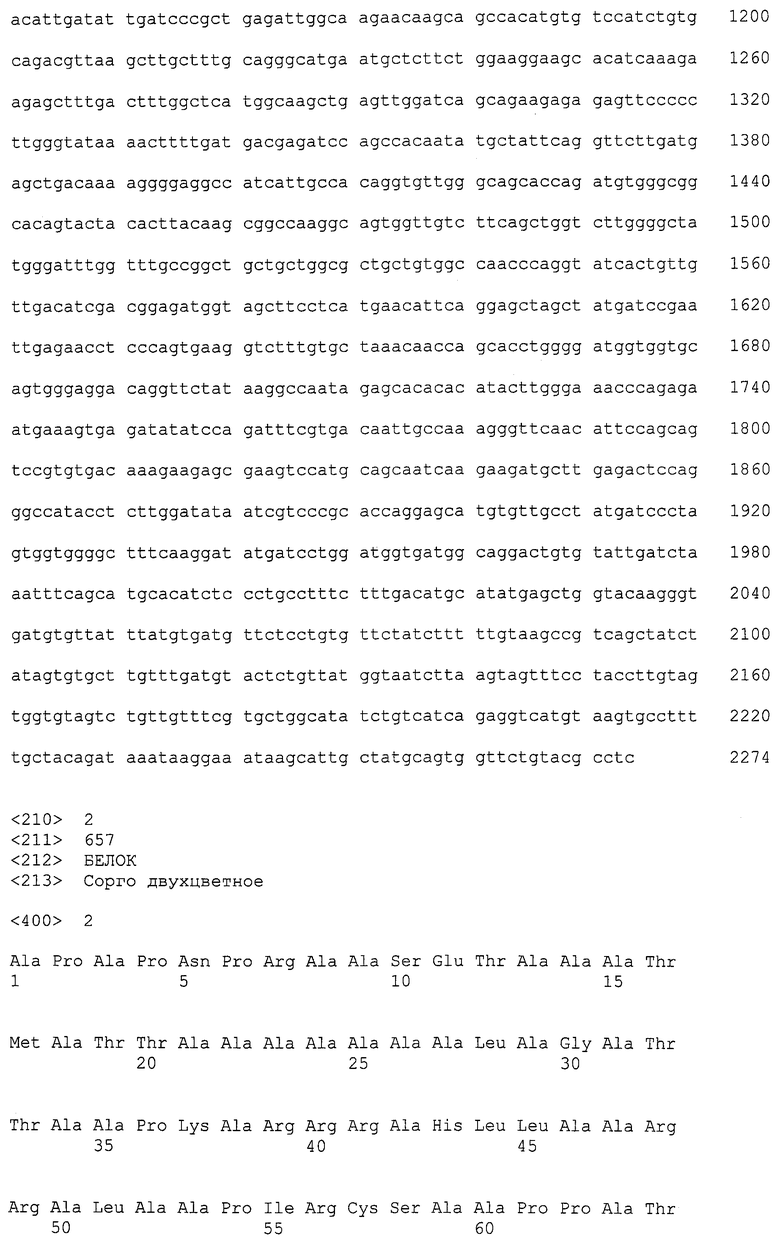
Амплификационные праймеры полимеразной цепной реакции успешно использовали, чтобы амплифицировать участок гена в устойчивых (S1-1, S1-2 и S1-3) и чувствительных (Тх623 и Тх430) к гербицидам генотипах сорго, используя следующие условия термоциклирования: денатурация при 94ºС 60 секунд, отжиг при 62ºС 45 секунд и наращивание при 72ºС 45 секунд. Продукты амплификации ПЦР очищали, используя QIAquick PCR Purification Kit (QIAGEN), и секвенировали на оборудовании для секвенирования Государственного университета Канзаса. Выведенная последовательность аминокислот (SEQ ID NO: 2) имеет мутации в Val531Ile (GTC на АТС), соответствующую A. thaliana Val560Ile, и Trp545Leu (TGG на TTG), соответствующую A. thaliana Trp574Leu, в генотипах, устойчивых к гербицидам (Фигура 1).
Все публикации и патенты, упомянутые в настоящей заявке, включены в ее описание в качестве ссылок. Различные модификации и изменения описанных способов и составов по изобретению будут очевидны для специалистов в данной области без отступления от объема и сущности изобретения. Кроме того, изобретение было описано применительно к специфическим предпочтительным модификациям, следует понимать, что, как заявлено, изобретение не следует неоправданно ограничивать до таких специфических модификаций. На самом деле, различные модификации описанных способов осуществления изобретения, которые очевидны для специалистов в соответствующих областях, следует включать в объем следующей патентной формулы.
| название | год | авторы | номер документа |
|---|---|---|---|
| СОРГО, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ, ДЕЙСТВУЮЩИМ НА АЦЕТИЛ-КоА КАРБОКСИЛАЗУ | 2008 |
|
RU2457674C2 |
| ПОДСОЛНЕЧНИК С НИЗКИМ СОДЕРЖАНИЕМ НАСЫЩЕННЫХ ЖИРОВ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ | 2008 |
|
RU2489849C2 |
| ПОДСОЛНЕЧНИК С НИЗКИМ СОДЕРЖАНИЕМ НАСЫЩЕННЫХ ЖИРОВ И СООТВЕТСТВУЮЩИЕ СПОСОБЫ | 2013 |
|
RU2639561C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОЛУЧЕНИЯ УКОРОЧЕННЫХ РАСТЕНИЙ КУКУРУЗЫ | 2016 |
|
RU2745987C2 |
| ЗАРОДЫШЕВАЯ ПЛАЗМА КАНОЛЫ, ПРОЯВЛЯЮЩАЯ ПРИЗНАКИ СОСТАВА СЕМЯН, КОТОРЫЕ ОБЕСПЕЧИВАЮТ УВЕЛИЧЕННУЮ ПИТАТЕЛЬНУЮ ЦЕННОСТЬ МУКИ КАНОЛЫ, ИМЕЮЩЕЙ ПРИЗНАКИ ОМЕГА-9 | 2012 |
|
RU2622086C2 |
| УСТОЙЧИВЫЕ К ГЕРБИЦИДАМ РАСТЕНИЯ ПОДСОЛНЕЧНИКА | 2009 |
|
RU2551781C2 |
| ВЫСОКОМАСЛИЧНЫЙ УСТОЙЧИВЫЙ К ИМИДАЗОЛИНОНУ ПОДСОЛНЕЧНИК | 2006 |
|
RU2420059C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| НОВАЯ МУТАЦИЯ, ВОВЛЕЧЕННАЯ В ПОВЫШЕННУЮ ТОЛЕРАНТНОСТЬ РАСТЕНИЙ К ИМИДАЗОЛИНОНОВЫМ ГЕРБИЦИДАМ | 2005 |
|
RU2525933C2 |
| ГЕНЕТИЧЕСКИЕ МАРКЕРЫ, АССОЦИИРОВАННЫЕ С ЗАСУХОУСТОЙЧИВОСТЬЮ КУКУРУЗЫ | 2010 |
|
RU2575605C2 |




Изобретение относится к биотехнологии. Описан гибрид сорго, в котором зародышевая плазма гибрида сорго придает устойчивость к ингибированию одним или более ацетолактатсинтазным гербицидом, где указанная зародышевая плазма гибрида сорго включает последовательность, приведенную в описании, которая дополнительно включает одно или боле нуклеотидных замещений, выбранных из группы, состоящей из замены гуанина на аденин в позиции 1641 и замены гуанина на тимидин в позиции 1641. Раскрыт способ выращивания описанного гибрида сорго в присутствии сорных растений. Описан способ получения гибридной линии растения сорго, устойчивой к одному или нескольким ацетолактатсинтазным гербицидам, включающий введение зародышевой плазмы в элитную линию растения сорго путем интродукции гетерологичного гена. Раскрыт способ определения линий растения сорго, устойчивых к ацетолактатсинтазным гербицидам. Представлено семя для выращивания растения, устойчивого к ацетолактатсинтазным гербицидам. Изобретение позволяет получать гибрид сорго, устойчивый к ингибированию одним или более ацетолактатсинтазным гербицидом при уровнях вышеуказанных гербицидов, которые в норме ингибировали бы рост гибрида сорго. 5 н. и 9 з.п. ф-лы, 1 ил., 1 табл., 3 пр.
1. Гибрид сорго, в котором зародышевая плазма вышеуказанного гибрида сорго придает устойчивость к ингибированию одним или более ацетолактатсинтазными гербицидами при уровнях одного или более вышеуказанных гербицидов, которые в норме ингибировали бы рост гибрида сорго, где указанная зародышевая плазма вышеуказанного гибрида сорго, которая придает устойчивость к ингибированию одним или несколькими ацетолактатсинтазными гербицидами, включает последовательность, содержащую SEQ ID NO:1, которая дополнительно включает одно или более нуклеотидных замещений, выбранных из группы, состоящей из замены гуанина на аденин в позиции 1641 и замены гуанина на тимидин в позиции 1641.
2. Гибрид сорго по п.1, где вышеуказанные один или более ацетолактатсинтазные гербициды относятся к группе, состоящей из сульфонилмочевин, имидазолинонов, триазолопиримидинов и пиримидилтиобензоатов.
3. Гибрид сорго по п.1, где вышеуказанная устойчивость к ингибированию одним или несколькими ацетолактатсинтазными гербицидами введена в зародышевую плазму вышеуказанного гибрида сорго при помощи интрогрессии.
4. Гибрид сорго по п.1, где семена от вышеуказанного гибрида сорго покрыты ацетолактатсинтазным гербицидом.
5. Способ выращивания гибрида сорго по любому из пп.1-4 в присутствии сорных растений вблизи него, включающий:
a) обеспечение одного или нескольких ацетолактатсинтазных гербицидов, выбранных из группы, состоящей из сульфонилмочевин, имидазолинонов, триазолопиримидинов, пиримидилтиобензоатов и их комбинации,
b) нанесение вышеуказанного одного или нескольких ацетолактатсинтазных гербицидов на площади, содержащих гибрид сорго,
и
c) уничтожение сорных растений вблизи вышеуказанного гибрида сорго таким образом, что росту сорных растений наносится ущерб за счет применения вышеуказанного одного или нескольких гербицидов, а росту вышеуказанного гибрида сорго не наносится ущерб.
6. Способ по п.5, при котором вышеуказанный гибрид сорго создан при помощи включения гетерологичного гена, содержащего одну или более вышеуказанные мутации, для придания вышеуказанному гибриду сорго устойчивости к одному или нескольким ацетолактатсинтазным гербицидам.
7. Способ получения гибридной линии растения сорго, устойчивой к одному или нескольким ацетолактатсинтазным гербицидам, включающий:
a) определение зародышевой плазмы, придающей вышеуказанную устойчивость к гербицидам, при котором вышеуказанная устойчивая к гербицидам зародышевая плазма происходит из устойчивого к гербицидам растения сорго по любому из пп.1-4; и
b) введение вышеуказанной зародышевой плазмы в элитную линию растения сорго путем интродукции гетерологичного гена.
8. Способ по п.7, при котором вышеуказанное введение вышеуказанной зародышевой плазмы в вышеуказанную элитную линию растения сорго производится при помощи интрогрессии.
9. Способ по п.7, при котором вышеуказанная устойчивая к гербицидам зародышевая плазма обладает устойчивостью к одному или нескольким ацетолактатсинтазным гербицидам и устойчивостью к одному или нескольким соединениям одной или нескольких гербицидных групп, которые не являются ацетолактатсинтазными ингибиторами.
10. Способ определения линий растения сорго, устойчивых к ацетолактатсинтазным гербицидам, включающий:
a) обеспечение образца нуклеиновой кислоты из растения сорго, где указанный образец нуклеиновой кислоты включает последовательность, содержащую SEQ ID NO:1, которая дополнительно включает одно или боле нуклеотидных замещений, выбранных из группы, состоящей из замены гуанина на аденин в позиции 1641 и замены гуанина на тимидин в позиции 1641,
b) обеспечение амплификационных праймеров для амплификации у растения сорго участка, соответствующего гену ацетолактатсинтазы, присутствующего в вышеуказанном образце нуклеиновой кислоты,
c) применение вышеуказанных амплификационных праймеров к вышеуказанному образцу нуклеиновой кислоты так, что происходит амплификация вышеуказанного участка вышеуказанного гена ацетолактатсинтазы, и
d) определение растений сорго, устойчивых к ацетолактатсинтазным гербицидам, основанное на присутствии одной или более мутаций, которые придают устойчивость к ацетолактатсинтазным гербицидам, присутствующим в вышеуказанном амплифицированном образце нуклеиновой кислоты.
11. Семя для выращивания растения, устойчивого к ацетолактатсинтазным гербицидам, где семя представляет собой семя гибрида сорго по любому из пп.1-4.
12. Гибрид сорго по п.2, где ацетолактатсинтазный гербицид представляет собой сульфонилмочевинный гербицид, выбранный из группы, состоящей из никосульфурона, римсульфурона и сульфометурон-метила.
13. Способ по п.5 или 6, где один или несколько ацетолактатсинтазных гербицидов выбраны из группы, состоящей из сульфонилмочевин, имидазолинонов, триазолопиримидинов и пиримидилтиобензоатов.
14. Способ по п.13, где ацетолактатсинтазный гербицид представляет собой сульфонилмочевинный гербицид, выбранный из группы, состоящей из никосульфурона, римсульфурона и сульфометурон-метила.
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| СПОСОБ ПОЛУЧЕНИЯ МУТАНТНОЙ ZEA MAYS | 1990 |
|
RU2128426C1 |

