Настоящее изобретение относится к области сельскохозяйственной биотехнологии, в частности к устойчивым к гербицидам растениям подсолнечника, содержащим новые полинуклеотидные последовательности, кодирующие белки большой субъединицы синтазы ацетогидроксикислот устойчивого к гербицидам подсолнечника.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Ацетолактатсинтаза (ALS) или синтаза ацетогидроксикислот (AHAS) является первым ферментом в пути биосинтеза аминокислот с разветвленной цепью валина, лейцина и изолейцина в растениях и микроорганизмах. Пять отдельных семейств соединений ингибируют фермент AHAS, и их используют в качестве неселективных гербицидов широкого спектра действия: сульфонилмочевины (SU), имидазолиноны (IMI), триазолопиримидины (TP), сульфониламинокарбонилтриазолиноны (SCT) и пиримидинилоксибензоаты (POB).
Принимая во внимание их высокую эффективность и низкую токсичность, конкретные гербициды предпочитают для сельскохозяйственного применения. Однако возможность применять некоторые из этих гербицидов в конкретной системе растениеводства зависит от доступности устойчивых сортов интересующего сельскохозяйственного растения. Для получения таких устойчивых сортов селекционерам необходимо разрабатывать генеалогические линии с признаками устойчивости. Соответственно, необходимы устойчивые к гербицидам генеалогические линии и сорта сельскохозяйственных растений, так же как способы и композиции получения и использования устойчивых генеалогических линий и сортов.
Множество сельскохозяйственных культур, включая кукурузу (Zea mays L.), канолу (Brassica napus L.), сахарную свеклу (Beta vulgaris L.), рис (Oryza sativa L.), хлопчатник (Gossypium hirsutum L.) подсолнечник (Helianthus annuus L.), лен (Linum usitatissimum L.), сою [Glycine max (L.) Merr.] и пшеницу (Triticum aestivum L.), устойчивых к AHAS-ингибирующим гербицидам, разработаны посредством множества способов, включая отбор соматических клеток, мутационную селекцию, трансформацию растений и межвидовую гибридизацию (Anderson and Georgeson, 1989; Croughan, 1996; D'Halluin et al., 1992; Newhouse et al., 1991, 1992; Hart et al., 1992; Wright and Penner, 1998; Swanson et al., 1989; Subramanian et al., 1990; Rajasekaran et al., 1996; Sebastian et al., 1989; Pozniak and Hucl, 2004; Al-Khatib and Miller, 2002; Mallory-Smith et al., 1990; McHughen, A. 1989). Устойчивость в большинстве этих случаев обусловлена формой AHAS, которая является менее чувствительной к ингибированию гербицидами из-за уменьшенного связывания с гербицидами, вызванного мутациями в гене(генах), кодирующем каталитическую субъединицу AHAS. Некоторые авторы опубликовали обзоры известных мутаций в генах AHAS, придающих растениям устойчивость к AHAS-ингибирующим гербицидам (Preston and Mallory-Smith; 2001; Tranel and Wright, 2002; Tan et al., 2005). Не опубликовано известных замен аминокислот в регуляторной субъединице, придающих устойчивость к гербицидам.
Обнаружены дикие популяции подсолнечника (Helianthus annuus L.), устойчивые к имидазолинонам (IMI) или сульфонилмочевинам (SU) (Al-Khatib et al 1998, White et al 2002). Проводили интрогрессию признака устойчивости к гербицидам в элитные инбредные линии подсолнечника посредством общепринятых способов селекции с целью разработки и районирования IMI-устойчивых и SU-устойчивых культиваров (Al-Khatib and Miller, 2002; Miller and Al-Khatib, 2002; 2004). На основании молекулярных исследований Kolkman et al. (2004) идентифицировали и охарактеризовали три гена AHAS в подсолнечнике (AHAS1, AHAS2 и AHAS3) и показали, что гены IMI-устойчивости и SU-устойчивости представляли собой аллельные варианты одного и того же локуса (AHASL1). Более того, они показали, что IMI-устойчивый аллель несет мутацию C-на-T в кодоне 205 (номенклатура Arabidopsis thaliana), в то время как SU-устойчивый аллель несет мутацию C-на-T в кодоне 197 (Kolkman et al., 2004).
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Представлены растения подсолнечника, обладающие множественной устойчивостью к гербицидам. Растения являются устойчивыми, по меньшей мере, к гербициду имидазолинону, гербициду сульфонилмочевине, гербициду триазолопиримидину и гербициду пиримидинилоксибензоату и смесям этих гербицидов. Преимущественно, семена растения обладают масличностью более 40%, урожай семян составляет более одной тонны на гектар, и/или растение подсолнечника обладает одиночной головкой. В других вариантах осуществления растения обладают сочетанием этих характеристик. В некоторых вариантах осуществления устойчивые растения обладают геном AHAS, кодирующим мутацию в аминокислоте 574 или эквивалентной ей, которая, по-видимому, придает признак устойчивости, где мутация может представлять собой замену триптофана в этом положении на другую аминокислоту, например лейцин. Конкретные варианты осуществления относятся к растениям, содержащим ген AHASL1, обладающий нуклеотидной последовательностью, указанной в SEQ ID NO:1, или белок AHASL1, обладающий аминокислотной последовательностью, указанной в SEQ ID NO:2.
Представлены также выделенные молекулы нуклеиновой кислоты, кодирующие мутантные гены AHASL1, например, выделенная нуклеиновая кислота, содержащая последовательность SEQ ID NO:1. Представлены также клетки растения, стабильно трансформированные нуклеиновой кислотой, обладающей последовательностью SEQ ID NO:1, и трансгенное растение подсолнечника, содержащее трансген, содержащий SEQ ID NO:1. В других вариантах осуществления растения содержат экспрессирующую кассету с промотором, активным в растениях подсолнечника, который является функционально связанным с выделенной нуклеиновой кислотой, кодирующей белок, обладающий последовательностью, кодируемой SEQ ID NO:1.
Представлены также способы контроля над сорняками поблизости от устойчивого растения подсолнечника описанного выше типа посредством воздействия гербицида на сорняки и растение подсолнечника, где гербицид представляет собой гербицид имидазолинон, гербицид сульфонилмочевину, гербицид триазолопиримидин или гербицид пиримидинилоксибензоат, или смеси этих гербицидов.
Представлены также семена подсолнечника и растение, выращенное из этих семян. Растения обладают устойчивостью к гербицидам против гербицида имидазолинона, гербицида сульфонилмочевины, гербицида триазолопиримидина, гербицида пиримидинилоксибензоата и их смесей. В конкретных вариантах осуществления растения, выращенные из семени, обладают масличностью семени более 40%, урожаем семян, который составляет более одной тонны на гектар, и/или растение подсолнечника обладает одиночной головкой. Дополнительные варианты осуществления включают в себя сочетания вышеупомянутых признаков. Эти растения могут содержать мутантный AHASL1, как описано выше.
В настоящей заявке представлены также способы улучшения толерантности к гербицидам у растения подсолнечника. Предпочтительные варианты осуществления включают в себя регенерацию клетки подсолнечника до растения подсолнечника, где клетку подсолнечника трансформируют конструкцией, обладающей последовательностью нуклеиновой кислоты, кодирующей белок, кодируемый SEQ ID NO:1, регенерируют клетку подсолнечника до растения подсолнечника и отбирают способные к размножению цветущие растения, являющиеся вегетативно и репродуктивно устойчивыми к эффективной дозе гербицида. Гербицид выбран из группы, состоящей из сульфонилмочевины, имидазолинона, пиримидинилоксибензоатов, триазолопиримидина и их смесей.
В настоящей заявке представлены также способы получения устойчивого к гербицидам растения подсолнечника посредством обратного скрещивания. Предпочтительные варианты осуществления включают в себя скрещивание первого растения подсолнечника, обладающего устойчивой к гербицидам активностью AHAS, со вторым растением подсолнечника, не обладающим устойчивой к гербицидам активностью AHAS, где первое растение подсолнечника содержит нуклеиновую кислоту, кодирующую белок, кодируемый SEQ ID NO:1. Преимущественно, отбирают растения-потомки, обладающие устойчивой к гербицидам активностью AHAS. Дополнительные предпочтительные варианты осуществления относятся к семени, обладающему характеристиками устойчивости к гербицидам первого растения подсолнечника.
Конкретный вариант осуществления относится к культивару подсолнечника, обозначенному RW-B. Это семя и соответствующее растение обладает всеми физиологическими характеристиками, характеристиками устойчивости к гербицидам и морфологическими характеристиками, описанными выше. Репрезентативное семя этого культивара подсолнечника депонировано в Американской коллекции типовых культур (ATCC) под инвентарным № PTA-9176. Представлена также культура ткани регенерируемых клеток вышеупомянутых устойчивых растений подсолнечника. Предпочтительные варианты осуществления относится к культурам тканей, где клетки культуры ткани содержат клетки листа, пыльцы, зародыша, семядоли, гипокотиля, меристематические клетки, клетки корня, кончика корня, пыльника, цветка, семени, стебля, семяпочек, побегов, стеблей, цветоножек, сердцевины капсул или шелухи. Устойчивые растения подсолнечника могут являться выращенными из вышеупомянутой культуры ткани и проявлять все морфологические и физиологические характеристики культивара подсолнечника RW-B.
Другие объекты, признаки и преимущества настоящего изобретения очевидны из следующего подробного описания. Следует понимать, однако, что подробное описание и конкретные примеры, в то время как в них указаны предпочтительные варианты осуществления изобретения, приведены только для иллюстрации, поскольку различные изменения и модификации в пределах содержания и объема изобретения очевидны специалистам в данной области из этого подробного описания.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На фиг.1 показано выравнивание частичной последовательности ДНК кодирующей области гена AHASL1 из HA89, линии B770, устойчивой линии RW-B, гена ALS Xanthium sp. и Arabidopsis thaliana.
На фиг.2 показано выравнивание частичной аминокислотной последовательности белка AHASL1 из HA89, линии B770, устойчивой линии RW-B, по гену ALS Xanthium sp. и Arabidopsis thaliana.
На фиг.3 показано наследование IMI устойчивости в RW-B.
На фиг.4 в форме таблицы показан показатель фитотоксичности семи линий подсолнечника, опрысканных различными AHAS-ингибирующими гербицидами.
На фиг.5 показан ответ растений подсолнечника, устойчивого (R) и чувствительного (S), на имазапир в тесте на аллелизм в поколениях F1, F2 и BC1F1.
На фиг.6 показана сегрегация гаплотипа AHAS1 и реакция растений подсолнечника, устойчивого (R) и чувствительного (S), на имазапир, применяемый в дозе 320 г a.i./га.
На фиг.7 показана полная нуклеотидная последовательность гена AHASL1 линии RW-B (SEQ ID NO:1).
На фиг.8 показана полная аминокислотная последовательность для гена AHASL1 линии RW-B (SEQ ID NO:2).
На фиг.9 показано уменьшение высоты (как процент от необработанных контрольных растений) для 4 линий, несущих различные мутантные аллели локуса AHASL1 подсолнечника, через 14 суток после воздействия увеличивающихся доз имазапира.
На фиг.10 показан показатель фитотоксичности для 4 линий подсолнечника, несущих различные мутантные аллели локуса AHASL1 подсолнечника, при испытании увеличивающимися дозами имазапира.
На фиг.11 показаны кривые зависимости от дозы для массы надземной биомассы в сухом состоянии (как процент от необработанных контрольных растений) для 4 линий подсолнечника, несущих различные мутантные аллели локуса AHASL1, при испытании увеличивающимися дозами имазапира.
На фиг.12 показан показатель фитотоксичности для 4 линий подсолнечника, несущих различные мутантные аллели локуса AHASL1 подсолнечника, при испытании увеличивающимися дозами метсульфурона.
На фиг.13 показаны кривые зависимости от дозы для массы надземной биомассы в сухом состоянии (как процент от необработанных контрольных растений) для 4 линий подсолнечника, несущих различные мутантные аллели локуса AHASL1 при испытании увеличивающимися дозами метсульфурона.
ПОДРОБНОЕ ОПИСАНИЕ
Представлены растения подсолнечника, обладающие увеличенной устойчивостью к гербицидам относительно растений подсолнечника дикого типа. Эти растения содержат новую генетическую мутацию в гене AHAS, придающую широкий спектр устойчивости к AHAS-ингибирующим гербицидам, и эта мутация идентифицирована и охарактеризована. Доступность этой новой мутации, в сочетании с применением доступных AHAS-ингибирующих гербицидов, предоставляет систему сельскохозяйственной продукции толерантного к гербицидам подсолнечника и представляет новое и действенное средство контроля над сорняками для выращивающих подсолнечник.
Несколько замен в AHAS могут приводить к устойчивости к ингибиторам AHAS; однако величина устойчивости к различным ингибирующим AHAS гербицидам в широких пределах варьирует среди замен (Saari et al., 1994; Tranel и Wright, 2002). Хотя существуют исключения, устойчивость, вызванную измененным AHAS, можно в основном классифицировать на три типа на основании перекрестной устойчивости: (1) SU-специфическая устойчивость, (2) IMI-специфическая устойчивость, и (3) широкая перекрестная устойчивость. Точечные мутации, приводящие к заменам пролина (Pro)197, придают высокие уровни устойчивости к сульфонилмочевине при только небольшом увеличении устойчивости к имидазолинонам и триазолопиримидинам (Haughn et al., 1988; Lee et al., 1988; Guttieri et al., 1992; Harms et al., 1992; Mourad and King 1992; Guttieri et al., 1995; Wright et al., 1998). С другой стороны, для замен аланина (Ala)205 показали широкую перекрестную устойчивость, однако уровни устойчивости являются умеренными (Saari et al., 1994; Tranel and Wright, 2002). Замены (Ala)122 приводят к устойчивости к IMI, но не к Su (см., например, Sala et al., 2008 a & b).
Признакам и генам устойчивости к AHAS-ингибирующим гербицидам у подсолнечника даны различные наименования. IMI-устойчивый аллель обозначен Imr1 (Bruniard and Miller, 2001), Ar pur или A205V (Kolkman et al. 2004), или Ahasl1-1 (см., например, Sala et al 2008b), и признак обозначен IMISUN. SU-устойчивый аллель обозначен Ar kan или P197L (Kolkman et al. 2004) или AhasL1-2 (см., например, Sala et al., 2008b), и признак известен как SURES. Высоко IMI-устойчивый аллель присутствует в CLHA-Plus, Ahasl1-3 или A122T (см., например, Sala et al., 2008b).
Авторы настоящего изобретения неожиданно обнаружили, что новая мутация в кодоне 574 (номенклатура Arabidopsis thaliana) гена AHASL1 подсолнечника обеспечивает высокие уровни и широкий спектр устойчивости, по меньшей мере, к четырем из пяти семейств AHAS-ингибирующих гербицидов. Эта одиночная мутация, заменяющая триптофан на лейцин в белке AHAS, является ответственной за это заметное расширение устойчивости к гербицидам, где растения подсолнечника по изобретению неожиданно обладают устойчивостью ко всем из гербицидов IMI, SU, TP и POB. Соответственно, эта новая мутация предоставляет ряд технологических преимуществ над всеми приводящими к устойчивости мутациями, в настоящее время известными для подсолнечника. Эти преимущества включают в себя в качестве неограничивающих примеров возможность: применять для сельскохозяйственных растений подсолнечника новые типы гербицидов, которые обычно нельзя применять без вредного воздействия на сам подсолнечник; применять для сельскохозяйственных растений подсолнечника смесь двух или более гербицидов одновременно, которые в другом случае нельзя комбинировать; менять время и тип воздействия гербицидов на подсолнечник; и применять более одного гербицида после появления всходов благодаря устойчивости к примесям, переходящим в новый цикл.
Представлены способы борьбы с нежелательной растительностью или контроля над сорняками посредством обработки семян подсолнечника от устойчивых растений AHAS-ингибирующим гербицидом или смесью гербицидов до посева и/или после предварительного проращивания. Обработанные семена затем высевают, например, в почву в полевых условиях или в горшечную смесь в теплице. Полученные растения также можно обрабатывать AHAS-ингибирующим гербицидом или смесью гербицидов для борьбы с нежелательной растительностью и/или контроля над сорняками в непосредственной близости от растений подсолнечника.
Эксперименты по обратному скрещиванию с использованием маркера позволили разработку коммерчески применимых растений подсолнечника с множественной устойчивостью к гербицидам, описанных в настоящем документе. При обратном скрещивании можно применять прямой отбор, когда генетический локус действует как частично доминантный признак, такой как в случае признака устойчивости к гербицидам. Для отбора растений подсолнечника по изобретению потомство от первоначального скрещивания опрыскивают гербицидом перед обратным скрещиванием. Опрыскивание уничтожает растения, не обладающие желательной характеристикой устойчивости к гербицидам, и растения, обладающие геном устойчивости к гербицидам, используют в последующих обратных скрещиваниях. Этот процесс повторяют для всех дополнительных поколений обратного скрещивания, пока не получат растения, передающие потомству признак множественной устойчивости к гербицидам.
В дополнение к устойчивости к гербицидам, растения подсолнечника также обладают рядом других преимущественных характеристик. Таким образом, семена подсолнечника обладают одной или несколькими из следующих характеристик, масличностью более 40%, урожаем семян более одной тонны на гектар, и растение подсолнечника обладает одиночной головкой. Неожиданно, эти характеристики обнаружены в сочетаниях друг с другом и с широким спектром устойчивости к гербицидам, придаваемым мутацией в кодоне 574.
Поскольку растения подсолнечника, описанные в настоящем документе, являются, примечательно, устойчивыми к четырем или более семействам гербицидов, фермеры могут обрабатывать сельскохозяйственные растения более широким диапазоном гербицидов, чем являлось доступным ранее для применения для растений подсолнечника. Контроль сорняков для сельскохозяйственных растений подсолнечника необходим, чтобы избегать плохого роста и потери урожая, и возможность применять более одного гербицида в одно и то же время или в различные периоды времени на протяжении периода вегетации обеспечивает значительные преимущества контроля над сорняками по сравнению с общепринятыми сортами подсолнечника. Например, поскольку появление растений подсолнечника, выращенных из семян, может занимать вплоть до двух недель, сорняки могут очень легко развиваться и затем затенять сеянцы подсолнечника, что может задерживать рост растений подсолнечника. В отсутствие контроля конкуренция сельскохозяйственных растений подсолнечника с сорняками приводит к основным экономическим потерям в растениеводстве из-за потерь урожая, достигающих настолько высокой величины, как 50%. Растения по настоящему изобретению предотвращают эти опасения, поскольку их устойчивость к более чем одному из существующих семейств гербицидов позволяет использование улучшенных способов применения гербицидов.
Дополнительным преимуществом устойчивости к гербицидам из множества семейств гербицидов является то, что фермеры могут использовать в коммерческих целях сельскохозяйственные растения подсолнечника без дополнительных расходов на разработку новых гербицидов. Разработка новых гербицидов для сельскохозяйственных растений подсолнечника маловероятна из-за высокой стоимости регистрации гербицидов. Множественная устойчивость к известным гербицидам также является преимущественной для сельскохозяйственных растений подсолнечника, чувствительным к повреждению из-за примесей, переходящих в новый цикл. Примеси, переходящие в новый цикл, возникают, когда гербициды после предыдущего сельскохозяйственного растения нарушают рост выращиваемого в настоящее время сельскохозяйственного растения. Наличие устойчивости ко множеству гербицидов уменьшает повреждение сельскохозяйственных растений подсолнечника из-за примесей, переходящих в новый цикл. Кроме того, увеличенная возможность контроля над сорняками для растений подсолнечника по изобретению означает, что будущие проблемы с сорняками для сельскохозяйственных растений, выращиваемых после растений подсолнечника, также являются ограниченными.
В контексте настоящего изобретения термины «толерантный к гербицидам» и «устойчивый к гербицидам» используют взаимозаменяемо, и они обладают эквивалентным значением и эквивалентным объемом. Подобным образом, термины «толерантность к гербицидам» и «устойчивость к гербицидам» используют взаимозаменяемо, и они обладают эквивалентным значением и эквивалентным объемом. Подобным образом, термины для семейств гербицидов, например, «устойчивый к имидазолинону» и «устойчивость к имидазолинону», используют взаимозаменяемо, и они обладают эквивалентным значением и объемом с терминами «толерантный к имидазолинону» и «толерантность к имидазолинону», соответственно.
Представлены устойчивые к гербицидам полинуклеотиды AHASL и устойчивые к гербицидам белки AHASL. Термин «устойчивый к гербицидам полинуклеотид AHASL» означает, для целей этого изобретения, полинуклеотид, кодирующий белок, обладающий активностью устойчивости к гербицидам AHAS. «Толерантный к гербицидам белок AHASL» или «устойчивый к гербицидам белок AHASL» представляет собой белок AHASL, для которого показали измеримо более высокую активность AHAS по сравнению с активностью AHAS белка AHASL дикого типа в присутствии, по меньшей мере, одного гербицида, который, как известно, мешает активности AHAS, и при концентрации или уровне гербицида, которые, как известно, ингибируют AHAS в случае белка AHASL дикого типа. «Толерантное к гербицидам» или «устойчивое к гербицидам» растение представляет собой растение, которое является толерантным или устойчивым, по меньшей мере, к одному гербициду, применяемому в конкретном количестве, которое в норме уничтожает нормальное растение или растение дикого типа или ингибирует его рост. В одном варианте осуществления толерантные к гербицидам растения по настоящему изобретению содержат толерантный к гербицидам или устойчивый к гербицидам белок AHASL. Хотя в недавних публикациях предложены различные обозначения номенклатуры, в данном случае и на всем протяжении настоящего документа термин AHASL используют для представления каталитической или большой (L) субъединицы фермента AHAS. См., например, Duggleby et al., 2008 Plant Physiol. Biochem. 46:309-324.
Кроме того, толерантный к гербицидам или устойчивый к гербицидам белок AHASL можно вводить в растение подсолнечника посредством трансформации растения или его предка нуклеотидной последовательностью, кодирующей толерантный к гербицидам или устойчивый к гербицидам белок AHASL. Такие толерантные к гербицидам или устойчивые к гербицидам белки AHASL кодированы толерантными к гербицидам или устойчивыми к гербицидам AHASL полинуклеотидами. Альтернативно, толерантный к гербицидам или устойчивый к гербицидам белок AHASL может возникать в растении подсолнечника в результате естественно возникшей или индуцированной мутации в эндогенном гене AHASL в геноме растения или его предка. Настоящее изобретение относится к растениям подсолнечника, тканям растения, клеткам растения и клеткам-хозяевам с увеличенной устойчивостью или толерантностью к одному или нескольким гербицидам, включая в качестве неограничивающих примеров гербицид IMI, гербицид SU, гербицид TP, гербицид POB. Преимущественно, растения обладают увеличенной устойчивостью или толерантностью к двум или более из вышеуказанных гербицидов и, более преимущественно, увеличенной устойчивостью или толерантностью к трем или более из вышеуказанных гербицидов. Более преимущественно, растения обладают увеличенной устойчивостью или толерантностью к четырем или более из вышеуказанных гербицидов.
Предпочтительное количество или концентрация гербицида, применяемое для растений или семян, как описано в настоящем описании, представляет собой «эффективное количество» или «эффективную концентрацию». Термины «эффективное количество» и «эффективная концентрация» обозначают, для целей этого изобретения, количество и концентрацию, соответственно, достаточные для уничтожения или ингибирования роста сходных, относящихся к дикому типу, растений, тканей растения, клеток растения или клеток-хозяев, но означают, что указанное количество не уничтожает устойчивые к гербицидам растения, ткани растения, клетки растения и клетки-хозяева по настоящему изобретению или не ингибирует их рост настолько серьезно. Как правило, эффективное количество гербицида представляет собой количество, общепринятое для применения в системах сельскохозяйственного производства для уничтожения интересующих сорняков. Такое количество известно обычным специалистам в данной области. Относящиеся к дикому типу растение, ткань растения, клетка растения или клетка-хозяин представляют собой растение, ткань растения, клетку растения или клетку-хозяина, соответственно, у которых отсутствуют характеристики устойчивости к гербицидам и/или конкретный полинуклеотид по изобретению. Термин «дикий тип» не подразумевает, что у растения, ткани растения, клетки растения или другой клетки-хозяина отсутствует рекомбинантная ДНК в геноме и/или что они не обладают характеристиками устойчивости к гербицидам, отличными от описанных в настоящем документе. Термин «растение» предназначен для обозначения растения на любой стадии развития, так же как любой части или частей растения, которые могут являться связанными с целым интактным растением или отделенными от него. Такие части растения включают в себя в качестве неограничивающих примеров органы, ткани и клетки растения. Примеры конкретных частей растения включают в себя стебель, лист, корень, соцветие, цветок, отдельный цветок соцветия, плод, цветоножку, плодоножку, тычинку, пыльник, рыльце, столбик, завязь, лепесток, чашелистик, плодолистик, кончик корня, корневой чехлик, корневой волосок, волосок листа, бородку зерна, пыльцевое зерно, микроспору, семядолю, гипокотиль, эпикотиль, ксилему, флоэму, паренхиму, эндосперм, клетку-спутник, замыкающую клетку и любые другие известные органы, ткани и клетки растения. Более того, признают, что семя представляет собой растение.
Настоящее изобретение относится к растению подсолнечника, обладающему увеличенной устойчивостью к гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смеси по сравнению с сортом растения дикого типа. В настоящем документе представлено подробное описание селекции и отбора растений подсолнечника, обладающих такой увеличенной устойчивостью к гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смесям. Одно из растений, полученных этими способами, депонировано в ATCC (инвентарный номер патентного депонирования PTA-9176) и обозначено в настоящем документе как сорт подсолнечника RW-B. Депонирование 2500 семян сорта подсолнечника RW-B в Американской коллекции типовых культур выполнено 25 апреля 2008 г. Это депонирование выполнено согласно положениям и условиям Будапештского Соглашения о депонировании микроорганизмов. Депонирование выполнено на срок, по меньшей мере, тридцать лет и, по меньшей мере, на пять лет после поступления в ATCC наиболее недавнего запроса о предоставлении образца.
Растения подсолнечника по настоящему изобретению включают в себя как нетрансгенные растения, так и трансгенные растения. «Нетрансгенное растение» представляет собой растение, в геноме которого отсутствует рекомбинантная ДНК. «Трансгенное растение» представляет собой растение, содержащее рекомбинантную ДНК в геноме, и его можно получить введением рекомбинантной ДНК в геном растения. Когда такую рекомбинантную ДНК вводят в геном трансгенного растения, потомство растения также может содержать рекомбинантную ДНК. Растение-потомок, содержащее, по меньшей мере, часть рекомбинантной ДНК, по меньшей мере, одного трансгенного растения-предка, также является трансгенным растением. Примером нетрансгенного растения подсолнечника, обладающего увеличенной устойчивостью к гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смесям, является растение подсолнечника RW-B или генетически модифицированное производное растения подсолнечника RW-B, или любое потомство растения подсолнечника RW-B, или растение, обладающее характеристиками толерантности к гербицидам растения подсолнечника RW-B.
Растения подсолнечника можно размножать способами культивирования тканей и клеток. По существу любую ткань растения с клетками, способными к делению клеток, можно использовать для размножения растения способами культивирования тканей и клеток. Культуры можно начинать из листа, пыльцы, зародыша, семядоли, гипокотиля, меристематических клеток, корня, кончика корня, пыльника, цветка, семени, стебля, семяпочек, побегов, стеблей, цветоножек, сердцевины капсул или шелухи. Ткани, взятые из сосудистых областей стеблей и корней, являются особенно подходящими. В патентах США № 4670391, 4670392, 4673648, 4681849, 4687743 и 5030572 описаны способы регенерации растений подсолнечника из культур клеток, полученных из тканей подсолнечника. Существующий уровень техники является таким, что иллюстративные способы получения растений подсолнечника из культур клеток и культур тканей в настоящее время являются хорошо известными обычному специалисту в данной области. Множество способов культивирования растений можно использовать для регенерации целых растений, такие как описанные в Gamborg and Phillips, «Plant Cell, Tissue and Organ Culture, Fundamental Methods», Springer Berlin, 1995); Evans et al. «Protoplasts Isolation and Culture», Handbook of Plant Cell Culture, Macmillian Publishing Company, New York, 1983; или Binding, «Regeneration of Plants, Plant Protoplasts», CRC Press, Boca Raton, 1985; или в Klee et al., Ann. Rev. of Plant Phys. 38:467 (1987). Подробные описания систем культивирования Helianthus annuus можно найти в Chapter 11, Sunflower Biotechnology, Bidney, D. L. and Scelonge, C. J., pp. 559-593 и процитированных в этом документе ссылках, Sunflower Technology and Production, edited by A. A. Schneiter, Agronomy 35, publishers, American Society of Agronomy Inc. 1997, Transformation of Sunflower, содержание всех из которых приведено в настоящем документе в качестве ссылки.
Устойчивые к гербицидам растения подсолнечника можно получать с использованием способов культивирования тканей для отбора клеток растений, содержащих придающие устойчивость к гербицидам мутации, и затем регенерации из них устойчивых к гербицидам растений. См., например, патенты США № 5773702 и 5859348, полное содержание обоих из которых приведено в настоящем документе в качестве ссылки. Дополнительные подробности селекции мутаций можно найти в «Principals of Cultivar Development» Fehr, 1993 Macmillan Publishing Company, содержание которого включено в настоящее описание в качестве ссылки.
Мутантный ген и кодируемый им белок придают растениям подсолнечника широкий спектр устойчивости к гербицидам
Авторы настоящего изобретения обнаружили, что замена триптофана в положении аминокислоты 574, или в эквивалентном положении (номенклатура Arabidopsis thaliana), в белке AHASL может вызывать у растения подсолнечника, экспрессирующего этот белок, увеличенную устойчивость к гербицидам, в частности, к гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смесям. Более конкретно, этот мутантный белок придает растениям множественную устойчивость к гербицидам.
Соответственно, устойчивые к гербицидам растения подсолнечника, описанные в настоящем документе, включают в себя в качестве неограничивающих примеров, растения подсолнечника, имеющие в геномах, по меньшей мере, одну копию полинуклеотида AHASL, кодирующую устойчивый к гербицидам белок AHASL, имеющий лейциновую замену в положении аминокислоты 574 (номенклатура Arabidopsis thaliana). Более того, специалисту в данной области известно, что такие положения аминокислот могут варьировать в зависимости от того, добавляют ли или удаляют ли аминокислоты, например, на N-конце аминокислотной последовательности. Таким образом, варианты осуществления настоящего изобретения, включенные в настоящую заявку, включают замены аминокислот в указанном положении или эквивалентном положении (например, «положении аминокислоты 574 или эквивалентном положении»). «Эквивалентное положение» представляет собой положение, которое находится в пределах той же самой области, как приведенное в качестве примера положение аминокислоты. Такие области известны в данной области знаний, или их можно определять выравниваниями множества последовательностей, как описано в настоящем описании, или посредством способов, известных в данной области.
Одиночная замена аминокислоты, которую можно использовать для получения устойчивых к гербицидам белков AHASL подсолнечника, представлена вместе с полинуклеотидами, кодирующими такие белки, и устойчивыми к гербицидам растениями, тканями растений, клетками растений и семенами, несущими эту мутацию. Представлены также выделенные полинуклеотиды подсолнечника AHASL1, AHASL2 и AHASL3, кодирующие устойчивые к гербицидам белки AHASL1, AHASL2 и AHASL3, соответственно. Каждый из таких устойчивых к гербицидам белков AHASL1, AHASL2 и AHASL3 может содержать аминокислоту, отличную от триптофана в положении 574 или эквивалентном положении (номенклатура Arabidopsis thaliana). Предпочтительно, в таких устойчивых к гербицидам белках AHASL1, AHASL2 и AHASL3, аминокислота в положении 574 или эквивалентном положении представляет собой лейцин.
Кроме того, представлены выделенные полипептиды, кодирующие белки AHASL. Выделенные полипептиды кодируют аминокислотную последовательность, обладающую аминокислотными последовательностями, указанными на фиг.8 (аминокислотная последовательность для гена AHASL1 линии RW-B, SEQ ID NO:2), аминокислотными последовательностями, кодируемыми нуклеотидными последовательностями, указанными на фиг.7 (нуклеотидная последовательность гена AHASL1 линии RW-B, SEQ ID NO:1), и функциональные фрагменты и варианты аминокислотных последовательностей, кодирующих полипептид AHASL, обладающий активностью AHAS.
Не ожидают, что делеции, вставки и замены белковых последовательностей, включенные в настоящую заявку, приводят к радикальным изменениям характеристик белка. Однако, когда трудно предсказать точный эффект замены, делеции или вставки до их осуществления, специалисту в данной области известно, что эффект можно оценивать общепринятыми анализами скрининга. То есть, активность можно оценивать анализами активности AHAS. См., например, Singh et al. (1988) Anal. Biochem. 171:173-179, содержание которого приведено в настоящем документе в качестве ссылки. Кроме того, белки, описанные в настоящем документе, включают как встречающиеся в природе белки, так и их варианты и модифицированные формы. Такие варианты продолжают обладать желательной активностью AHAS.
Изобретение относится также к выделенным или очищенным полинуклеотидам и белкам. «Выделенные» или «очищенные» молекула полинуклеотида или белок, или его биологически активная часть, являются по существу или в основном свободными от компонентов, которые в норме сопутствуют молекуле полинуклеотида или белка или взаимодействуют с ними, как обнаружено в их естественном природном окружении. Таким образом, выделенные или очищенные молекула полинуклеотида или белок являются по существу свободными от другого клеточного материала или культуральной среды при получении рекомбинантными способами, или по существу свободными от химических предшественников или других химических веществ при химическом синтезе. Предпочтительно, «выделенная» нуклеиновая кислота является свободной от последовательностей (предпочтительно, кодирующих белок последовательностей), естественным образом фланкирующих нуклеиновую кислоту (т.е. последовательностей, локализованных на 5'- и 3'-концах нуклеиновой кислоты) в геномной ДНК организма, из которого получена нуклеиновая кислота. Например, в различных вариантах осуществления выделенная молекула полинуклеотида может содержать менее приблизительно 5 т.п.о., 4 т.п.о., 3 т.п.о., 2 т.п.о., 1 т.п.о., 0,5 т.п.о., 0,4 т.п.о., 0,3 т.п.о., 0,2 т.п.о. или 0,1 т.п.о. из нуклеотидной последовательности, в природе фланкирующей молекулу полинуклеотида в геномной ДНК клетки, из которой получена нуклеиновая кислота. Белок, который является по существу свободным от клеточного материала, включает в себя препараты белка, имеющие менее приблизительно 30%, 25%, 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% (по сухой массе) контаминирующего белка. При рекомбинантном получении белка по настоящему изобретению или его биологически активной части, предпочтительно, культуральная среда представляет менее приблизительно 30%, 25%, 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% (по сухой массе) химических предшественников или не относящихся к интересующему белку химических веществ.
Представлены выделенные молекулы полинуклеотида, обладающие нуклеотидными последовательностями, кодирующими белки AHASL, и соответствующие белки. Представлены также нуклеотидные последовательности, кодирующие устойчивый к гербицидам белок AHASL из устойчивого к гербицидам растения подсолнечника. В одном варианте осуществления белки AHASL устойчивого к гербицидам подсолнечника обладают заменой триптофана на лейцина в положении аминокислоты 574 или эквивалентном положении (номенклатура Arabidopsis thaliana) по сравнению с соответствующей аминокислотной последовательностью дикого типа. В частности, представлены молекулы полинуклеотида, содержащие нуклеотидные последовательности, кодирующие аминокислотные последовательности, показанные на фиг.8, и их фрагменты и варианты, кодирующие полипептиды, обладающие активностью AHAS. Кроме того, представлены полипептиды, обладающие аминокислотной последовательностью, кодируемой молекулами полинуклеотида, описанными в настоящем документе, например молекулами, указанными на фиг.7, и их фрагментами и вариантами, кодирующими полипептиды, обладающие активностью AHAS.
Кроме того, специалисту в данной области дополнительно известно, что посредством мутаций можно вводить изменения в нуклеотидные последовательности по изобретению, таким образом, вызывая изменения в аминокислотной последовательности кодируемых белков AHASL без изменения биологической активности белков AHASL. Таким образом, выделенную молекулу полинуклеотида, кодирующую белок AHASL, обладающий последовательностью, отличающейся от последовательности из SEQ ID NO:2, можно получать введением одной или нескольких замен, добавлений или делеций нуклеотидов в соответствующую нуклеотидную последовательность, описанную в настоящем документе, так что одну или несколько замен, добавлений или делеций аминокислот вводят в кодируемый белок. Мутации можно вводить общепринятыми способами, такими как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез.
Выделенные устойчивые к гербицидам молекулы полинуклеотидов AHASL по изобретению можно использовать в полинуклеотидных конструкциях для трансформации растений, в частности сельскохозяйственных растений, для увеличения устойчивости растений к гербицидам, в частности к гербицидам, известным как ингибирующие активность AHAS, более конкретно, к гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смесям. Такие полинуклеотидные конструкции можно использовать в экспрессирующих кассетах, экспрессирующих векторах, трансформирующих векторах, плазмидах и т.п. Для трансгенных растений, полученных после трансформации такими полинуклеотидными конструкциями, показали увеличенную устойчивость к AHAS-ингибирующим гербицидам, например, таким как гербицид IMI, гербицид SU, гербицид TP, гербицид POB и/или их смеси.
Выделенные полинуклеотидные молекулы, обладающие нуклеотидной последовательностью, кодирующей мутантный белок AHASL, как описано выше, можно использовать в векторах для трансформации растений, так что полученные растения обладают увеличенной устойчивостью к гербицидам, в частности гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смесям. Выделенные полинуклеотидные молекулы AHASL по настоящему изобретению можно использовать в векторах отдельно или в сочетании с нуклеотидной последовательностью, кодирующей малую субъединицу фермента AHAS (AHASS), для придания растениям устойчивости к гербицидам. См. патент США № 6348643, содержание которого приведено в настоящем документе в качестве ссылки.
Молекулы полинуклеотидов, отличающиеся от нуклеотидных последовательностей, описанных в настоящем документе, также включены в настоящее изобретение. Нуклеотидные последовательности по настоящему изобретению включают последовательности, кодирующие белки AHASL, описанные в настоящем документе, но обладающие консервативными заменами из-за вырожденности генетического кода. Эти встречающиеся в природе аллельные варианты можно идентифицировать с использованием хорошо известных способов молекулярной биологии, таких как способы полимеразной цепной реакции (ПЦР) и гибридизации, как описано ниже. Нуклеотидные последовательности включают в себя также синтетически полученные нуклеотидные последовательности, которые получены, например, с использованием сайт-направленного мутагенеза, но которые все еще кодируют белок AHASL. В общем, варианты нуклеотидной последовательности по настоящему изобретению могут обладать, по меньшей мере, приблизительно 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с конкретной нуклеотидной последовательностью, описанной в настоящем документе. Нуклеотидная последовательность AHASL может кодировать белок AHASL, соответственно, обладающий аминокислотной последовательностью, обладающей, по меньшей мере, приблизительно 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичностью последовательности с аминокислотной последовательностью белка AHASL, описанной в настоящем документе.
Варианты осуществления по изобретению относятся также к экспрессирующему вектору для растений, обладающему промотором, управляющим экспрессией в растении, функционально связанным с полинуклеотидной молекулой по настоящему изобретению. Выделенная полинуклеотидная молекула содержит нуклеотидную последовательность, кодирующую белок AHASL, или его функциональный фрагмент и вариант. Экспрессирующие векторы для растений из вариантов осуществления настоящего изобретения не зависят от конкретного промотора, только в том, что такой промотор является способным управлять экспрессией генов в клетке растения. Предпочтительные промоторы включают в себя конститутивные промоторы и предпочтительные для определенных тканей промоторы.
Трансформирующие векторы можно использовать для получения растений, трансформированных интересующим геном. Трансформирующий вектор может иметь ген селективного маркера по настоящему изобретению и интересующий ген, подлежащий введению, и, как правило, экспрессии в трансформированном растении. Такой ген селективного маркера обладает устойчивым к гербицидам полинуклеотидом AHASL по настоящему изобретению, функционально связанным с промотором, управляющим экспрессией в клетке-хозяине. Для применения в растениях и клетках растений, трансформирующий вектор обладает устойчивым к гербицидам полинуклеотидом AHASL по настоящему изобретению, функционально связанный с промотором, который управляет экспрессией в клетке растения. Интересующие гены по настоящему изобретению варьируют в зависимости от желательного исхода. Например, различные изменения фенотипа могут являться интересующими, включая модифицирующие состав жирных кислот в растении, изменение содержания аминокислот в растении, изменение механизмов защиты растения от насекомых и/или патогенов, и т.п. Этих результатов можно достигать обеспечением экспрессии гетерологичных продуктов или увеличенной экспрессии эндогенных продуктов в растениях. Альтернативно, результатов можно достигать обеспечением уменьшения экспрессии одного или нескольких эндогенных продуктов, в частности ферментов или кофакторов в растении. Эти изменения приводят к изменению фенотипа трансформированного растения.
В одном варианте осуществления интересующие гены включают в себя гены устойчивости к насекомым, например, такие как гены белка-токсина Bacillus thuringiensis (патенты США № 5366892; 5747450; 5736514; 5723756; 5593881; и Geiser et al. (1986) Gene 48:109). Белки или полипептиды AHASL, описанные в настоящем документе, можно очищать, например, из растений подсолнечника. Выделенную молекулу полинуклеотида, кодирующую белок AHASL по настоящему изобретению, можно использовать для экспрессии белка AHASL по настоящему изобретению в микроорганизме, таком как E. coli или дрожжи. Экспрессированный белок AHASL можно очищать из экстрактов E. coli или дрожжей любым способом, известным обычным специалистам в данной области.
Варианты осуществления настоящего изобретения относятся к молекулам полинуклеотида AHASL и их фрагментам и вариантам. Для целей по этому изобретению термин «фрагменты и варианты» обозначает фрагменты и варианты приведенных в качестве примеров полипептидов, обладающих активностью AHAS. В конкретных вариантах осуществления способы включают в себя применение толерантных к гербицидам или устойчивых к гербицидам растений. Фрагмент нуклеотидной последовательности AHASL по настоящему изобретению может кодировать биологически активную часть белка AHASL, или он может представлять собой фрагмент, который можно использовать в качестве зонда для гибридизации или праймера для ПЦР с использованием способов, описанных ниже. Биологически активную часть белка AHASL можно получать выделением части одной из нуклеотидных последовательностей AHASL по настоящему изобретению, экспрессии кодируемой части белка AHASL (например, посредством рекомбинантной экспрессии in vitro), и оценки активности кодированной части белка AHASL1. Молекулы полинуклеотида, представляющие собой фрагменты нуклеотидной последовательности AHASL, содержат, по меньшей мере, приблизительно 15, 20, 25, 30, 40, 50, 75, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1050 или 1100 нуклеотидов, или вплоть до числа нуклеотидов, присутствующих в полноразмерной нуклеотидной последовательности, описанной в настоящем документе, в зависимости от предназначенного применения.
Фрагмент нуклеотидной последовательности AHASL, кодирующей биологически активную часть белка AHASL по настоящему изобретению, кодирует, по меньшей мере, приблизительно 15, 20, 25, 30, 40, 50, 75, 100, 125, 150, 175, 200, 225, 250, 275, 300, 325 или 350 непрерывных аминокислот, или вплоть до общего числа аминокислот, присутствующих в полноразмерном белке AHASL по настоящему изобретению. Фрагменты нуклеотидной последовательности AHASL, применимые в качестве зондов для гибридизации и праймеров для ПЦР, не обязательно кодируют биологически активную часть белка AHASL.
Варианты нуклеотидных последовательностей AHASL можно получать введением мутаций случайным образом по всей или части кодирующей последовательности AHASL, например, насыщающим мутагенезом, и можно проводить скрининг полученных мутаций по активности AHAS для идентификации мутантов, сохраняющих активность AHAS, включая активность устойчивой к гербицидам AHAS. После мутагенеза кодируемый белок можно экспрессировать рекомбинантным способом, и активность белка можно определять с использованием общепринятых способов анализа. Таким образом, нуклеотидные последовательности по изобретению включают в себя последовательности, описанные в настоящем документе, так же как их фрагменты и варианты. Нуклеотидные последовательности AHASL по настоящему изобретению, и их фрагменты и варианты, можно использовать в качестве зондов и/или праймеров для идентификации и/или клонирования гомологов AHASL в других растениях, такие зонды можно использовать для детекции транскриптов или геномных последовательностей, кодирующих такие же или идентичные белки. Таким образом, способы, такие как ПЦР, гибридизация, и т.п., можно использовать для идентификации таких последовательностей, обладающих значительной идентичностью с последовательностями по настоящему изобретению. См., например, Sambrook et al. (1989) Molecular Cloning: Laboratory Manual (2d ed., Cold Spring Harbor Laboratory Press, Plainview, NY) и Innis, et al. (1990) PCR Protocols: A Guide to Methods and Applications (Academic Press, NY). Нуклеотидные последовательности AHASL, выделенные на основании идентичности их последовательности с нуклеотидными последовательностями AHASL, указанными в настоящем документе, или их фрагментами и вариантами, включены в настоящее изобретение.
Трансгенез
В настоящем документе описаны способы получения трансгенного растения, которое является устойчивым к гербицидам. Способы включают в себя трансформацию растения экспрессирующим вектором для растений, обладающим промотором, управляющим экспрессией в растении, функционально связанным с выделенной молекулой полинуклеотида по изобретению. Выделенная молекула полинуклеотида обладает нуклеотидной последовательностью, кодирующей белок AHASL или функциональный фрагмент и вариант аминокислотных последовательностей.
Варианты осуществления настоящего изобретения относятся к растениям, органам растений, тканям растений, клеткам растений, семенам и не относящимся к человеку клеткам-хозяевам, трансформированным, по меньшей мере, одной молекулой полинуклеотида, экспрессирующей кассетой или трансформирующим вектором по изобретению. Такие трансформированные растения, органы растения, ткани растений, клетки растений, семена и не относящиеся к человеку клетки-хозяева обладают увеличенной толерантностью или устойчивостью, по меньшей мере, к одному гербициду, при уровнях гербицида, которые уничтожают нетрансформированное растение, ткань растения, клетку растения или не относящуюся к человеку клетку-хозяина, или ингибируют их рост, соответственно.
Полинуклеотиды AHASL, описанные в настоящем документе, применяют в способах увеличения устойчивости толерантных к гербицидам растений. В одном варианте осуществления толерантные к гербицидам растения обладают толерантным к гербицидам или устойчивым к гербицидам белком AHASL. Толерантные к гербицидам растения включают в себя как растения, трансформированные толерантными к гербицидам нуклеотидными последовательностями AHASL, так и растения, имеющие в геномах эндогенный ген, кодирующий толерантный к гербицидам белок AHASL. Нуклеотидные последовательности, кодирующие толерантные к гербицидам белки AHASL и толерантные к гербицидам растения, имеющие эндогенный ген, кодирующий толерантный к гербицидам белок AHASL, включают в себя полинуклеотиды и растения, описанные в настоящем документе, и известные в данной области. См., например, патенты США № 5013659, 5731180, 5767361, 5545822, 5736629, 5773703, 5773704, 5952553 и 6274796; содержание всех из которых приведено в настоящем документе в качестве ссылки. Такие способы увеличения устойчивости толерантных к гербицидам растений включают в себя трансформацию толерантного к гербицидам растения, по меньшей мере, одной полинуклеотидной конструкцией, обладающей промотором, управляющим экспрессией в клетке растения, функционально связанным с устойчивым к гербицидам полинуклеотидом AHASL по изобретению, в частности полинуклеотидом, кодирующим устойчивый к гербицидам белок AHASL, и фрагментами и вариантами указанных полинуклеотидов, кодирующими полипептиды, обладающие устойчивой к гербицидам активностью AHAS. Однако ни один из белков AHASL до настоящего изобретения не являлся способным придавать обладающую широким спектром и сильную устойчивость к гербицидам, полученную с использованием гена AHASL по настоящему изобретению.
Доступны многочисленные векторы для трансформации растений и способы трансформации растений. См., например, An, G. et al. (1986) Plant Physiol. 81:301-305; Fry, J., et al. (1987) Plant Cell Rep. 6:321-325; Block, M. (1988) Theor. Appl Genet. 76:767-77; Hinchee, et al. (1990) Stadler. Genet. Symp. 203212.203-212; Cousins, et al. (1991) Aust. J. Plant Physiol, 18:481-494; Chee, P. P. and Slightom, J. L. (1992) Gene 118:255-260; Christou, et al. (1992) Trends. Biotechnol. 10:239-246; D'Ηalluin, et al. (1992) Bio/Technol. 10:309-314; Dhir, et al. (1992) Plant Physiol. 99:81-88; Casas et al. (1993) Proc. Nat. Acad. Sci. USA 90:11212-11216; Christou, P. (1993) In Vitro Cell. Dev. Biol-Plant; 29P:119-124; Davies, et al. (1993) Plant Cell Rep. 12:180-183; Dong, J. A. and Mchughen, A. (1993) Plant Sci. 91:139-148; Franklin, C. I. and Trieu, T. N. (1993) Plant Physiol. 102:167; Golovkin, et al. (1993) Plant Sci. 90:41-52; Guo Chin ScL Bull. 38:2072-2078; Asano, et al. (1994) Plant Cell Rep. 13; Ayeres N. M. and Park, W. D. (1994) Crit. Rev. Plant. Sci. 13:219-239; Barcelo, et al. (1994) Plant. J. 5:583-592; Becker, et al., (1994) Plant. J. 5:299-307; Borkowska et al. (1994) Acta. Physiol. Plant 16:225-230; Christou, P. (1994) Agro. Food. Md Hi Tech. 5:17-27; Eapen et al. (1994) Plant Cell Rep. 13:582-586; Hartman, et al. (1994) Bio-Technology 12:919923; Ritala, et al. (1994) Plant. Mol. Biol. 24:317-325; и Wan, Y. C. and Lemaux, P. G. (1994) Plant Physiol. 104:3748.
Варианты осуществления способов по изобретению включают в себя введение полинуклеотидной конструкции в растение. Термин «введение» означает презентацию растению полинуклеотидной конструкции таким образом, что конструкция получает доступ к внутренней части клетки растения. Способы не зависят от конкретного способа введения полинуклеотидной конструкции в растение, полинуклеотидная конструкция должна только получать доступ к внутренней части, по меньшей мере, одной клетки растения. В данной области известны способы введения полинуклеотидных конструкций в клетки растений, включая, в качестве неограничивающих примеров, способы стабильной трансформации, способы временной трансформации и опосредованные вирусами способы. Термин «стабильная трансформация» означает, что полинуклеотидная конструкция, введенная в растение, интегрирует в геном растения и является способной наследоваться его потомством. Термин «временная трансформация» означает, что полинуклеотидная конструкция, введенная в растение, не интегрирует в геном растения.
Для трансформации растений и клеток растений нуклеотидные последовательности по изобретению вставляют с использованием общепринятых способов в любой вектор, известный в данной области, пригодный для экспрессии нуклеотидных последовательностей в растении или клетке растения. Выбор вектора зависит от предпочтительного способа трансформации и растения-мишени, подлежащего трансформации. В варианте осуществления настоящего изобретения нуклеотидная последовательность AHASL функционально связана с промотором растения, известным как дающий высокий уровень экспрессии в клетке растения, и эту конструкцию затем вводят в растение, которое является чувствительным к одному или несколькими из следующих гербицидов, к гербициду IMI, гербициду SU, гербициду TP, или гербициду POB, и трансформированное растение регенерируют. Трансформированное растение является толерантным к воздействию уровня гербицида IMI, гербицида SU, гербицида TP гербицида POB, и/или их смесей, которые могут уничтожать или значительно повреждать нетрансформированное растение. Этот способ можно применять к любым видам растений; однако, предпочтительно, он является преимущественным при применении для растений подсолнечника.
Способы конструирования экспрессирующих кассет для растений и введения чужеродных нуклеиновых кислот в растения в основном известны в данной области и описаны ранее. Например, чужеродную ДНК можно вводить в растения с использованием индуцирующих опухоль (Ti) плазмидных векторов. Другие способы, применяемые для доставки чужеродной ДНК, включают в себя использование опосредованной PEG трансформации протопластов, электропорации, волокон для микроинъекции, и биолистики или бомбардировки микрочастицами для прямого поглощения ДНК. Такие способы известны в данной области (патент США № 5405765 Vasil et al.; Bilang et al. (1991) Gene 100:247-250; Scheid et al., (1991) Mol. Gen. Genet., 228:104-112; Gnerche et al., (1987) Plant Science 52:111-116; Neuhause et al., (1987) Theor. Appl Genet. 75:30-36; Klein et al., (19S7) Nature 327:70-73; Howell et al. (1980) Science 208:1265; Horsch et al. (19S5) Science 227:1229-1231; DeBlock et al. (1989) Plant Physiology 91:694-701; Methods for Plant Molecular Biology (Weissbach and Weissbach, eds.) Academic Press, Inc. (1988) и Methods in Plant Molecular Biology (Schuler and Zielinski, Eds.) Academic Press, Inc. (1989). Способ трансформации зависит от клетки растения, подлежащей трансформации, стабильности используемых векторов, уровня экспрессии продуктов генов и других параметров.
Другие пригодные способы введения нуклеотидных последовательностей в клетки растений и последующей вставки в геном растения включают в себя микроинъекцию, как в Crossway et al. (1986) Biotechniques 4:320-334, электропорацию, как описано в Riggs et al. (1986) Proc. Natl. Acad. Sci. USA 83:5602-5606, опосредованную агробактериями трансформацию, как описано в Townsend et al., патенте США № 5563055, Zhao et al., патенте США № 5981840, прямой перенос генов, как описано в Paszkowski et al. (1984) EMBO J. 3:2717-2722, и баллистическое ускорение частиц, как описано, например, в Sanford et al., патенте США № 4945050; Tomes et al., патенте США № 5879918; Tomes et al., патенте США № 5886244; Bidney et al., патенте США № 5932782; Tomes et al. (1995) «Direct DNA Transfer into Intact Plant Cells via Micro projectile Bombardment», in Plant Cell, Tissue, and Organ Culture: Fundamental Methods, ed. Gamborg and Phillips (Springer-Verlag, Berlin); McCabe et al. (1988) Biotechnology) 6:923-926); и трансформацию Lec1 (WO 00/28058). См. также De Wet et al. (1985) in The Experimental Manipulation of Ovule Tissues, ed. Chapman et al. (Longman, New York), pp. 197-209 (pollen); Kaeppler et al. (1990) Plant Cell Reports 9:415-418 и Kaeppler et al. (1992) Theor. Appl. Genet. 84:560-566 (опосредованная волокнами трансформация); D'Halluin et al. (1992) Plant Cell 4:1495-1505 (электропорация); Li et al. (1993) Plant Cell Reports 12:250-255; содержание всех из которых приведено в настоящем документе в качестве ссылки.
Полинуклеотиды из вариантов осуществления настоящего изобретения можно вводить в растения посредством приведения растений в контакт с вирусом или вирусными нуклеиновыми кислотами. Как правило, такие способы включают в себя введение полинуклеотидной конструкции по настоящему изобретению в составе молекулы вирусной ДНК или РНК. Известно, что белок AHASL по настоящему изобретению можно первоначально синтезировать в виде части вирусного полибелка, который затем можно подвергать процессингу посредством протеолиза in vivo или in vitro для получения желательного рекомбинантного белка. Кроме того, известно, что промоторы по настоящему изобретению также включают в себя промоторы, используемые для транскрипции посредством вирусных РНК-полимераз. Способы для введения полинуклеотидных конструкций в растения и экспрессии белка, кодируемого в них, включающие в себя молекулы вирусной ДНК или РНК, известны в данной области. См., например, патенты США № 5889191, 5889190, 5866785, 5589367 и 5316931; содержание которых приведено в настоящем документе в качестве ссылки.
Клетки, которые были трансформированы, можно выращивать до растений общепринятыми способами. См., например, McCormick et al. (1986) Plant Cell Reports 5:81-84. Затем эти растения можно выращивать и опылять с помощью того же самого трансформированного штамма или других штаммов, и идентифицировать полученный гибрид, обладающий конститутивной экспрессией желательной фенотипической характеристики. Можно выращивать два или более поколений, чтобы убедиться, что эта экспрессия желательной фенотипической характеристики стабильно сохраняется и наследуется, и что получены семена, собранные, чтобы убедиться в экспрессии желательной фенотипической характеристики. Таким образом, варианты осуществления настоящего изобретения относятся к трансформированному семени (обозначаемому также «трансгенное семя»), обладающему полинуклеотидной конструкцией по настоящему изобретению, например экспрессирующей кассетой по настоящему изобретению, стабильно встроенной в геном. Устойчивые к гербицидам растения, описанные в настоящем документе, находят применение в способах контроля над сорняками. Таким образом, варианты осуществления по изобретению включают в себя способы контроля над сорняками поблизости от устойчивого к гербицидам растения по настоящему изобретению. Способы включают в себя воздействие эффективного количества гербицида на сорняки и на устойчивое к гербицидам растение, где растение обладает увеличенной устойчивостью к одному или нескольким из следующих гербицидов, гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смесям, по сравнению с растением дикого типа.
Варианты осуществления настоящего изобретения относятся к нетрансгенным и трансгенным семенам с увеличенной толерантностью к одному или нескольким гербицидам, в частности AHAS-ингибирующему гербициду, более конкретно, гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смесям. Такие семена включают в себя, например, нетрансгенные семена подсолнечника, обладающие характеристиками толерантности к гербицидам растения подсолнечника RW-B, и трансгенные семена, содержащие полинуклеотидную молекулу по настоящему изобретению, кодирующую устойчивый к гербицидам белок AHASL.
Тестирование на присутствие мутантного гена AHASL
Нуклеотидные последовательности из вариантов осуществления настоящего изобретения можно использовать для выделения соответствующих последовательностей из других организмов, в частности других растений, более конкретно, других двудольных растений. Таким образом, такие способы, как ПЦР, гибридизация и т.п., можно использовать для идентификации таких последовательностей на основании гомологии их последовательностей с последовательностями, указанными в настоящем документе. Последовательности, выделенные на основании идентичности их последовательности с полными полинуклеотидными последовательностями AHASL, указанными в настоящем документе, или с их фрагментами, включены в настоящее изобретение. Таким образом, выделенные полинуклеотидные последовательности, кодирующие белок AHASL и гибридизующиеся в строгих условиях с последовательностями, описанными в настоящем документе, или с их фрагментами, включены в настоящее изобретение.
В способе ПЦР можно сконструировать олигонуклеотидные праймеры для применения в реакциях ПЦР для амплификации соответствующих последовательностей ДНК из кДНК или геномной ДНК, выделенной из любого интересующего растения. Способы конструирования праймеров для ПЦР и клонирования после ПЦР в общем известны в данной области. См. Innis and Gelfand, eds. (1995) PCR Strategies (Academic Press, New York); и Innis and Gelfand, eds. (1999) PCR Methods Manual (Academic Press, New York). Известные способы ПЦР включают в себя в качестве неограничивающих примеров способы с использованием парных праймеров, гнездовых праймеров, одиночных специфических праймеров, вырожденных праймеров, специфических для гена праймеров, специфических для вектора праймеров, праймеров с частичными несовпадениями и т.п. Полинуклеотидные последовательности AHASL по настоящему изобретению представлены в экспрессирующих кассетах для экспрессии в интересующем растении. Кассета может включать 5'- и 3'-регуляторные последовательности, функционально связанные с полинуклеотидной последовательностью AHASL по настоящему изобретению. «Функционально связанный» представляет собой функциональную связь между промотором и второй последовательностью, где промоторная последовательность инициирует и опосредует транскрипцию последовательности ДНК, соответствующей второй последовательности. Как правило, функционально связанный означает, что связанные последовательности нуклеиновой кислоты являются непрерывными и, при необходимости соединения двух кодирующих белок областей, непрерывными и лежащими в одной рамке считывания. Кроме того, кассета может содержать, по меньшей мере, один дополнительный ген для совместной трансформации организма. Альтернативно, дополнительный ген(ы) можно предоставлять на множестве экспрессирующих кассет.
Такую экспрессирующую кассету предоставляют со множеством участков рестрикции для вставки полинуклеотидной последовательности AHASL, чтобы она находилась под контролем регуляции транскрипции регуляторных областей. Кроме того, экспрессирующая кассета может содержать гены селективных маркеров. Экспрессирующая кассета может содержать в 5'-3' направлении транскрипции область инициации транскрипции и трансляции (т.е. промотор), полинуклеотидную последовательность AHASL по настоящему изобретению и область терминации транскрипции и трансляции (т.е. область терминации), функциональные в растениях. Промотор может являться нативным или аналогичным, или чужеродным или гетерологичным, для растения-хозяина и/или для полинуклеотидных последовательностей AHASL по изобретению. Кроме того, промотор может представлять собой природную последовательность или, альтернативно, синтетическую последовательность. Когда промотор является «чужеродным» или «гетерологичным» для растения-хозяина, подразумевают, что промотор не обнаружен в природном растении, в которое вводят промотор. Когда промотор является «чужеродным» или «гетерологичным» для полинуклеотидной последовательности AHASL, подразумевают, что промотор не является нативным или встречающимся в природе промотором для функционально связанной полинуклеотидной последовательности AHASL. Как применяют в настоящем документе, химерный ген содержит кодирующую последовательность, функционально связанную с областью инициации транскрипции, которая является гетерологичной для кодирующей последовательности.
В то время как устойчивые к гербицидам полинуклеотиды AHASL, описанные в настоящем документе, можно использовать в качестве генов селективных маркеров для трансформации растений, экспрессирующие кассеты, содержащие эти полинуклеотиды, могут содержать другой ген селективного маркера для отбора трансформированных клеток. Гены селективных маркеров используют для отбора трансформированных клеток или тканей. Маркерные гены включают в себя в качестве неограничивающих примеров гены, кодирующие устойчивость к антибиотикам, такие как гены, кодирующие неомицин-фосфотрансферазу II (NEO) и гигромицин-фосфотрансферазу (HPT), так же как гены, придающие устойчивость к гербицидным соединениям 5, таким как глюфосинат аммония, бромоксинил, имидазолиноны, и 2,4-дихлорфеноксиацетат (2,4-D). См., в общем, Yarranton (1992) Curr. Opin. Biotech. 3:506-511; Christopherson et al. (1992) Proc. Natl. Acad. Sci. USA 89:6314-6318; Yao et al. (1992) Cell 1:63-72; Reznikoff (1992) Mol. Microbiol. 6:2419-2422; Barkley et al. (1980) in The Operon, pp. 177-220; Hu et al. (1987) Cell 48:555-566; Brown et al. (1987) Cell 10 49:603-612; Figge et al. (1988) Cell 52:713-722; Deuschle et al. (1989) Proc. Natl. Acad. Sci. USA 86:5400-5404; Fuerst et al. (1989) Proc. Natl. Acad Sci. USA 86:2549-2553; Deuschle et al. (1990) Science 248:480-483; Gossen (1993) Ph.D. Thesis, University of Heidelberg; Reines et al. (1993) Proc. Natl. Acad. Sci. USA 90:1917-1921; Labow et al. (1990) Mol. Cell. Biol. 10:3343-3356; Zambretti et al. (1992) Proc. Natl. Acad. Sci. USA 15 89:3952-3956; Baim et al. (1991) Proc. Natl. Acad, Sci. USA 88:5072-5076; Wyborski et al. (1991) Nucleic Acids Res. 19:4647-4653; Hillenand-Wissman (1989) Topics Mol. Struc. Biol. 10:143-162; Degenkolb et al. (1991) Antimicrob. Agents Chemother. 35:1591-1595; Kleinschr M et al. (1988) Biochemistry 27:1094-1104; Bonin (1993) PhD. Thesis, University of Heidelberg; Gossen et al. (1992) Proc. Natl. Acad Sci. USA 89:5547-5551; Oliva et al. (1992) Antimicrob. Agents Chemother. 36:913-919; Hlavka et al. (1985) Handbook of Experimental Pharmacology, Vol. 78 (Springer-Verlag, Berlin); Gill et al. (1988) Nature 334:721-724. Содержание таких настоящих изобретений приведено в настоящем документе в качестве ссылки. Вышеуказанный список генов селективных маркеров не предназначен, чтобы являться ограничивающим, и можно использовать любой ген селективного маркера по необходимости.
В способе гибридизации всю или часть известной нуклеотидной последовательности AHASL можно использовать для скрининга библиотек кДНК или геномных библиотек. Способы конструирования библиотек кДНК и геномных библиотек в общем известны в данной области. Так называемые зонды для гибридизации могут представлять собой фрагменты геномной ДНК, фрагменты кДНК, фрагменты РНК или другие олигонуклеотиды, и могут являться меченными поддающейся детекции группой, такой как 32P, или любым другим поддающимся детекции маркером, таким как другие радиоактивные изотопы, флуоресцентное соединение, фермент или кофактор фермента. Зонды для гибридизации можно получать мечением синтетических олигонуклеотидов, основанных на известной нуклеотидной последовательности AHASL, описанной в настоящем документе. Кроме того, можно использовать вырожденные праймеры, сконструированные на основании консервативных нуклеотидных или аминокислотных остатков известной нуклеотидной последовательности AHASL или кодируемой аминокислотной последовательности. Зонд, как правило, содержит область нуклеотидной последовательности, которая гибридизуется в строгих условиях, по меньшей мере, приблизительно с 12, предпочтительно, приблизительно с 25, более предпочтительно, приблизительно с 50, 75, 100, 125, 150, 175, 200, 250, 300, 350, 400, 500, 600, 700, 800, 900, 1000, 1100 или более последовательными нуклеотидами из нуклеотидной последовательности AHASL по настоящему изобретению или ее фрагмента или варианта. Получение зондов для гибридизации в общем известно в данной области.
Например, полную последовательность AHASL, описанную в настоящем документе, или одну или несколько ее частей можно использовать в качестве зонда, способного к специфической гибридизации в строгих условиях с соответствующими последовательностями и матричными РНК AHASL. Способы гибридизации включают в себя скрининг посредством гибридизации рассеянных библиотек ДНК (бляшек или колоний). Гибридизацию таких последовательностей можно проводить в строгих условиях. Термины «строгие условия» или «строгие условия гибридизации» означают, для целей по этому изобретению, условия, при которых зонд может гибридизоваться с его последовательностью-мишенью до поддающейся детекции более высокой степени, чем с другими последовательностями (например, по меньшей мере, в 2 раза выше фона). Строгие условия зависят от последовательности и могут являться различными при различных обстоятельствах. Как правило, строгие условия являются такими, при которых концентрация соли представляет собой менее приблизительно 1,5 M ион Na, как правило, концентрация иона Na (или других солей) приблизительно 0,01-1,0 M при pH 7,0-8,3 и температура составляет, по меньшей мере, приблизительно 30°C для коротких зондов (например, 10-50 нуклеотидов) и, по меньшей мере, приблизительно 60°C для длинных зондов (например, более 50 нуклеотидов). Строгих условий можно также достигать добавлением дестабилизаторов, таких как формамид. Иллюстративные условия низкой строгости включают в себя гибридизацию в буферном растворе из 30-35% формамида, 1 M NaCl, 1% SDS (додецилсульфата натрия) при 37°C, и отмывку в 1X-2X SSC (20X SSC=3,0 M NaCl/0,3 M цитрат тринатрия) при 50-55°C. Иллюстративные условия умеренной строгости включают в себя гибридизацию в 40-45% формамиде, 1,0 M NaCl, 1% SDS при 37°C, и отмывку в 0,5X-1X SSC при 55-60°C. Иллюстративные условия высокой строгости включают в себя гибридизацию в 50% формамиде, 1 M NaCl, 1% SDS при 37°C, и отмывку в 0,1X SSC при 60-65°C. Необязательно, буферы для отмывки могут содержать от приблизительно 0,1% до приблизительно 1% SDS. Продолжительность гибридизации, как правило, менее приблизительно 24 часов, обычно от приблизительно 4 до приблизительно 12 часов.
Специфичность, как правило, является функцией от отмывок после гибридизации, где критическими факторами являются ионная сила и температура раствора для конечной отмывки. Для гибридов ДНК-ДНК Tm можно аппроксимировать из уравнения Meinkoth и Wahl (1984) Anal. Biochem. 138:267-284: Tm=81,5°C+16,6 (log M) + 0,41 (%GC) - 0,61 (% form) - 500/L; где M представляет собой молярность моновалентных катионов, %GC представляет собой процент нуклеотидов гуанозина и цитозина в ДНК, % form представляет собой процентное содержание формамида в растворе для гибридизации, и L представляет собой длину гибрида в парах оснований. Tm представляет собой температуру (при определенной ионной силе и pH) при которой 50% комплементарной последовательности-мишени гибридизуется с полностью совпадающим зондом. Tm уменьшают приблизительно на 1°C для каждого 1% несоответствия; таким образом, Tm, условия гибридизации и/или отмывки можно регулировать для гибридизации с последовательностями желаемой идентичности. Например, при поиске последовательностей с идентичностью >90% Tm можно уменьшать на 10°C. Как правило, строгие условия выбирают приблизительно на 5°C ниже температурной точки плавления (Tm) для конкретной последовательности и комплементарной ей при определенной ионной силе и pH. Однако для чрезвычайно строгих условий можно использовать гибридизацию и/или отмывку на 1, 2, 3 или 4°C ниже температурной точки плавления (Tm); для умеренно строгих условий можно использовать гибридизацию и/или отмывку на 6, 7, 8, 9 или 10°C ниже температурной точки плавления (Tm); для условий с низкой строгостью можно использовать гибридизацию и/или отмывку на 11, 12, 13, 14, 15 или 20°C ниже температурной точки плавления (Tm). Обычному специалисту в данной области понятно, что с использованием уравнения, составов для гибридизации и отмывки и желаемой Tm по существу описывают варианты строгости гибридизации и/или растворов для отмывки. Если желаемая степень несоответствия приводит к Tm менее 45°C (раствор в воде) или 32°C (раствор в формамиде), предпочтительным является увеличение концентрации SSC, чтобы можно было использовать более высокую температуру. Обширное руководство по гибридизации нуклеиновых кислот обнаружено в Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, Part I, Chapter 2 (Elsevier, New York); and Ausubel et al., eds. (1995) Current Protocols in Molecular Biology, Chapter 2 (Greene Publishing and Wiley-Interscience, New York).
Трансгенез и оптимизация экспрессии
Настоящее изобретение относится к способам увеличения активности AHAS в растении посредством трансформации растения полинуклеотидной конструкцией, обладающей промотором, функционально связанным с нуклеотидной последовательностью AHASL1. Способы включают в себя введение полинуклеотидной конструкции по изобретению, по меньшей мере, в одну клетку растения и регенерацию трансформированного растения с использованием промотора, способного управлять экспрессией гена в клетке растения. Предпочтительно, такой промотор представляет собой конститутивный промотор или предпочтительный для ткани промотор. Способы находят применение для увеличения устойчивости или толерантности растения, по меньшей мере, к одному гербициду, и преимущественно двум, трем, или четырем или более гербицидов, мешающим каталитической активности фермента AHAS, в частности, к гербициду IMI, гербициду SU, гербициду TP, гербициду POB и/или их смесям.
Когда это целесообразно, ген(ы) можно оптимизировать для увеличенной экспрессии в трансформированном растении. То есть, гены можно синтезировать с использованием предпочтительных для растения кодонов для улучшения экспрессии. См., например, в Campbell and Gowri (1990) Plant Physiol 92:1-11 обсуждение предпочтительной для хозяина частоты использования кодонов. В данной области доступны способы синтеза предпочтительных для растения генов. См., например, патенты США № 5380831 и 5436391 и Murray et al. (1989) Nucleic Acids Res. 17:477-498, содержание которых приведено в настоящем документе в качестве ссылки.
Известны дополнительные модификации последовательности для увеличения экспрессии генов в клетках хозяина. Они включают в себя исключение последовательностей, кодирующих случайные сигналы полиаденилирования, сигналы участков соединения экзон-интрон, подобные транспозонам повторы, и других таких хорошо охарактеризованных последовательностей, которые могут являться неблагоприятными для экспрессии генов. Содержание G-C последовательности можно регулировать до уровня, среднего для данных клеток хозяина, как рассчитано по отношению к известным генам, экспрессируемым в клетке-хозяине. Когда возможно, последовательность модифицируют для удаления предсказанных вторичных шпилечных структур мРНК. Нуклеотидные последовательности для усиления экспрессии гена также можно использовать в экспрессирующих векторах для растений. Они включают в себя интроны Adh1 кукурузы, ген интрона 1 (Callis et al. Genes and Development 1:1183-1200, 1987), и лидерные последовательности, (W-последовательность) из вируса табачной мозаики (TMV), вируса хлоратической пятнистости кукурузы и вируса мозаики люцерны (Gallie et al. Nucleic Acid Res. 15:8693-8711, 1987 и Skuzeski et al. Plant Mol. Biol. 15:65-79, 1990). Показано, что первый интрон локуса shrunken-1 кукурузы увеличивает экспрессию генов в химерных генетических конструкциях. В патентах США № 5424412 и 5593874 описано применение конкретных интронов в конструкциях для экспрессии генов, и Gallie et al. (Plant Physiol. 106:929-939, 1994) показали также, что интроны являются применимыми для регуляции экспрессии генов на тканеспецифической основе. Для дополнительного усиления или для оптимизации экспрессии гена большой субъединицы AHAS, экспрессирующие векторы для растений по настоящему изобретению могут также содержать последовательности ДНК, содержащие участки прикрепления к ядерному матриксу (MAR). Клетки растения, трансформированные с помощью таких модифицированных систем экспрессии, затем могут обладать сверхэкспрессией или конститутивной экспрессией нуклеотидной последовательности по настоящему изобретению.
Экспрессирующие кассеты могут, кроме того, содержать 5'-лидерные последовательности в конструкции экспрессирующей кассеты. Такие лидерные последовательности могут действовать для усиления трансляции. Трансляционные лидеры известны в данной области и включают в себя: лидеры пикорнавирусов, например лидер EMCV (5'-некодирующая область энцефаломиокардита) (Elroy-Stein et al. (1989) Proc. Natl Acad. Sci. USA 86:6126-6130); лидеры потивирусов, например, лидер TEV (вируса гравировки табака) (Gallie et al. (1995) Gene 165(2):233-238), лидер MDMV (вирус карликовой мозаики кукурузы) (Virology 154:9-20), и связывающий тяжелую цепь иммуноглобулина белок человека (BiP) (Macejak et al. (1991) Nature 353:90-94); нетранслируемый лидер из мРНК белка оболочки вируса мозаики люцерны (AMV РНК 4) (Jobling et al. (1987) Nature 325:622-625); лидер вируса табачной мозаики (TMV) (Gallie et al. (1989) in Molecular Biology of RNA, ed. Cech (Liss, New York), pp. 237-256); и лидер вируса хлоратической пятнистости кукурузы (MCMV) (Lommel et al. (1991) Virology 81:382-385). См. также Della-Cioppa et al. (1987) Plant Physiol. 84:965-968. Можно использовать также другие способы, известные для усиления трансляции, например, интроны и т.п. При получении экспрессирующей кассеты можно манипулировать различными фрагментами ДНК, так чтобы предоставлять последовательности ДНК в правильной ориентации и, по необходимости, в правильной рамке считывания. С этой целью можно применять адаптеры или линкеры для соединения фрагментов ДНК или других манипуляций, которые можно применять для предоставления подходящих участков рестрикции, удаления избыточной ДНК, удаления участков рестрикции или т.п. Для этой цели можно применять мутагенез in vitro, репарацию праймеров, рестрикцию, отжиг, повторные перестановки, например транзиции и трансверсии.
Экспрессирующие кассеты для экспрессии полинуклеотидов в растениях, тканях растений, клетках растений и других клетках-хозяевах. Экспрессирующие кассеты обладают промотором, поддающимся экспрессии в растении, ткани растения, клетке растения или других интересующих клетках-хозяевах, функционально связанных с полинуклеотидом по изобретению, обладающим нуклеотидной последовательностью, кодирующей либо полноразмерный (т.е. включая транзитный пептид хлоропластов) или зрелый белок AHASL (т.е. без транзитного пептида хлоропластов). Если желательна экспрессия в пластидах или хлоропластах растений или клетках растений, экспрессирующая кассета может также обладать функционально связанной нацеленной на хлоропласты последовательностью, кодирующей транзитный пептид хлоропластов. Такие экспрессирующие кассеты можно использовать в способе увеличения толерантности к гербициду растения или клетки-хозяина. Способ включает в себя трансформацию растения или клетки-хозяина экспрессирующей кассетой, содержащей промотор, поддающийся экспрессии в интересующем растении или клетке-хозяине, и промотор является функционально связанным с полинуклеотидом из варианта осуществления настоящего изобретения, содержащим нуклеотидную последовательность, кодирующую устойчивый, например, к гербициду IMI, гербициду SU, гербициду TP или гербициду POB белок AHASL.
Использование термина «полинуклеотидные конструкции» не ограничено ДНК. Специалисту в данной области известно, что полинуклеотидные конструкции, в частности, полинуклеотиды и олигонуклеотиды, состоящие из рибонуклеотидов и комбинаций рибонуклеотидов и дезоксирибонуклеотидов, также можно применять в способах, описанных в настоящем документе. Такие дезоксирибонуклеотиды и рибонуклеотиды включают в себя как природные молекулы, так и синтетические аналоги. Полинуклеотидные конструкции по изобретению включают в себя также все формы полинуклеотидных конструкций, включая, в качестве неограничивающих примеров, одноцепочечные формы, двухцепочечные формы, шпильки, структуры стебель-и-петля, и т.п. Более того, специалисту в данной области понятно, что каждая нуклеотидная последовательность, описанная в настоящем документе, включает в себя также последовательность, комплементарную приведенной в качестве примера нуклеотидной последовательности.
В способах по изобретению можно применять полинуклеотидную конструкцию, способную управлять, в трансформированном растении, экспрессией, по меньшей мере, одного белка, или, по меньшей мере, одной РНК, например, такой как антисмысловая РНК, которая является комплементарной, по меньшей мере, части мРНК. Как правило, такая полинуклеотидная конструкция состоит из последовательности, кодирующей белок или РНК, функционально связанной с 5'- и 3'-регуляторными областями транскрипции. Альтернативно, можно использовать полинуклеотидную конструкцию, не способную управлять, в трансформированном растении, экспрессией белка или РНК. Для экспрессии полинуклеотида по настоящему изобретению в интересующей клетке-хозяине, полинуклеотид, как правило, является функционально связанным с промотором, способным управлять экспрессией гена в интересующей клетке-хозяине. Способы экспрессии полинуклеотидов в клетках-хозяевах не зависят от конкретного промотора и включают в себя применение любого промотора, который является известным в данной области и который является способным управлять экспрессией гена в интересующей клетке-хозяине.
В то время как может являться предпочтительным экспрессировать полинуклеотиды AHASL с использованием гетерологичных промоторов, можно использовать нативные промоторные последовательности. Такие конструкции могут изменять уровни экспрессии белка AHASL в растении или клетке растения. Таким образом, фенотип растения или клетки растения является измененным. Область терминации может являться нативной для области инициации транскрипции, может являться нативной для функционально связанной интересующей последовательности AHASL, может являться нативной для растения-хозяина или может являться полученной из другого источника (т.е. чужеродной или гетерологичной для промотора, интересующей полинуклеотидной последовательности AHASL, растения-хозяина или любой их комбинацией). Подходящие области терминации доступны из Ti-плазмиды A. tumefaciens, такие как области терминации октопин-синтазы и нопалин-синтазы. См. также Guerineau et al. (1991) Mol. Gen. Genet. 262:141-144; Proudfoot (1991) Cell 64:671-674; Sanfacon et al. (1991) Genes Dev. 5:141-149; Mogen et al. (1990) Plant Cell 2:1261-1272; Munroe et al. (1990) Gene 91:151-158; Ballas et al. (1989) Nucleic Acid Res. 17:7891-7903; и Joshi et al. (1987) Nucleic Acid Res. 15:9627-9639.
Ряд промоторов можно использовать в практическом осуществлении изобретения, и их можно выбирать на основании желательного исхода. Нуклеиновые кислоты можно комбинировать с конститутивными, предпочтительными для тканей, или другими промоторами для экспрессии в растениях. Такие конститутивные промоторы включают в себя, например, коровый промотор из промотора Rsyn7 и другие конститутивные промоторы, описанные в WO 99/43838 и патенте США № 6072050; коровый промотор CaMV 35S (Odell et al. (1985) Nature 313:810-812); промотор актина риса (McElroy et al. (1990) Plant Cell 2:163-171), убиквитина (Christensen et al. (1989) Plant Mol. Biol. 12:619-632 и Christensen et al. (1992) Plant Mol. Biol. 18:675-689), pEMU (Last et al. (1991) Theor. Appl. Genet. 81:581-588), MAS (Velten et al. (1984) EMBO J. 3:2723-2730), ALS (патент США № 5659026), и т.п. Другие конститутивные промоторы включают в себя, например, промоторы из патентов США № 5608149; 5608144; 5604121; 5569597; 5466785; 5399680; 5268463; 5608142 и 6177611. Предпочтительные для тканей промоторы можно использовать для нацеливания на усиленную экспрессию AHASL в конкретной ткани растения. Такие предпочтительные для тканей промоторы включают в себя в качестве неограничивающих примеров предпочтительные для листа промоторы, предпочтительные для корня промоторы, предпочтительные для семени промоторы и предпочтительные для стебля промоторы. Предпочтительные для тканей промоторы описаны, например, в Yamamoto et al. (1997) Plant J. 12(2):255-265; Kawamata et al. (1997) Plant Cell Physiol. 38(7):792-803; Hansen et al. (1997) Mol. Gen. Genet. 254(3):337-343; Russell et al. (1997) Transgenic Res. 6(2):157-168; Rinehart et al. (1996) Plant Physiol. 112(3):1331-1341; Van Camp et al. (1996) Plant Physiol. 112(2):525-535; Canevascini et al. (1996) Plant Physiol. 112(2):513-524; Yamamoto et al. (1994) Plant Cell Physiol. 35(5):773-778; Lam (1994) Results Prob I. Cell Differ. 20:181-196; Orozco et al. (1993) Plant Mol. Biol. 23(6):1129-1138; Matsuoka et al. (1993) Proc. Natl. Acad. Sci. USA 90(20):9586-9590; and Guevara-Garcia et al. (1993) Plant J. 4(3):495-505. Такие промоторы можно модифицировать, если необходимо, для слабой экспрессии.
В одном варианте осуществления интересующие нуклеиновые кислоты нацелены на хлоропласт для экспрессии. Таким образом, когда интересующая нуклеиновая кислота напрямую не встроена в хлоропласт, экспрессирующая кассета, кроме того, содержит нацеленную на хлоропласт последовательность, содержащую нуклеотидную последовательность, кодирующую транзитный пептид хлоропластов для направления интересующего продукта гена в хлоропласты. Такие транзитные пептиды известны в данной области. По отношению к нацеленным на хлоропласт последовательностям, «функционально связанный» означает, что последовательность нуклеиновой кислоты, кодирующая транзитный пептид (т.е. нацеленная на хлоропласт последовательность), является связанной с полинуклеотидом AHASL, так что две последовательности являются непрерывными и находятся в одной и той же рамке считывания. См., например, Von Heijne el al. (1991) Plant Mol. Biol. Rep. 9:104-126; Clark et al. (1989) J. Biol. Chem. 264:17544-17550; Della-Cioppa et al. (1987) Plant Physiol. 84:965-968; Romer et al. (1993) Biochem. Biophys. Res. Commun. 196:1414-1421; and Shah et al. (1986) Science 233:478-481.
В то время как белки AHASL, описанные в настоящем документе, могут включать в себя нативный транзитный пептид хлоропластов, любой транзитный пептид хлоропластов, известный в данной области, можно сливать с аминокислотной последовательностью зрелого белка AHASL по настоящему изобретению посредством функционального связывания нацеленной на хлоропласт последовательности с 5'-концом нуклеотидной последовательности, кодирующей зрелый белок AHASL.
Нацеленные на хлоропласт последовательности известны в данной области и включают в себя малую субъединицу рибулоза-1,5-бисфосфат-карбоксилазы (Rubisco) хлоропластов (de Castro Silva Filho et al. (1996) Plant Mol. Biol. 30:769-780; Schnell et al. (1991) J. Biol. Chem. 266(5):3335-3342); 5-(енолпирувил)шикимат-3-фосфатсинтазу (EPSPS) (Archer et al. (1990) J Bioenerg. Biomemb. 22(6):789-810); триптофансинтазу (Zhao et al. (1995) J. Biol. Chem. 270(11):6081-6087); пластоцианин (Lawrence et al. (1997) J Biol. Chem. 272(33):20357-20363); хоризматсинтазу (Schmidt et al. (1993) J Biol. Chem. 268(36):27447-27457); и связывающий поглощающий свет хлорофилл a/b белок (LHBP) (Lamppa et al. (1988) J. Biol. Chem. 263:14996-14999). См. также Von Heijne et al. (1991) Plant Mol. Biol. Rep. 9:104-126; Clark et al. (1989) J Biol. Chem. 264:17544-17550; Della-Cioppa et al. (1987) Plant Physiol. 84:965-968; Romer et al. (1993) Biochem. Biophys. Res. Commun. 196:1414-1421; и Shah et al. (1986) Science 233:478-481.
Способы трансформации хлоропластов известны в данной области. См., например, Svab et al. (1990) Proc. Natl. Acad. Sci. USA 87:8526-8530; Svab and Maliga (1993) Proc. Natl. Acad. Sci. USA 90:913-917; Svab and Maliga (1993) EMBO J. 12:601-606. Способ основан на доставке с помощью генной пушки ДНК, содержащей селективный маркер, и нацеливании ДНК на геном пластид посредством гомологичной рекомбинации. Кроме того, трансформацию пластид можно осуществлять посредством трансактивации молчащего происходящего из пластид трансгена посредством предпочтительной для ткани экспрессией кодируемой в ядре и нацеленной на пластиды РНК-полимеразы. Такая система опубликована в McBride et al. (1994) Proc. Natl. Acad. Sci. USA 91:7301-7305.
Интересующие нуклеиновые кислоты для нацеливания на хлоропласты можно оптимизировать для экспрессии в хлоропластах, чтобы учитывать различия в частоте использования кодонов между ядром растения и этой органеллой. Таким образом, интересующие нуклеиновые кислоты можно синтезировать с использованием предпочтительных для хлоропластов кодонов. См., например, патент США № 5380831, содержание которого приведено в настоящем документе в качестве ссылки.
Как описано в настоящем описании, нуклеотидные последовательности AHASL из вариантов осуществления настоящего изобретения находят применение для усиления толерантности к гербицидам растений, содержащих в геномах ген, кодирующий толерантный к гербицидам белок AHASL. Такой ген может представлять собой эндогенный ген или трансген. Кроме того, в конкретных вариантах осуществления последовательности нуклеиновой кислоты по настоящему изобретению можно комплектовать с любой комбинацией интересующих полинуклеотидных последовательностей для получения растений с желаемым фенотипом. Например, полинуклеотиды можно комплектовать с любыми другими полинуклеотидами, кодирующими полипептиды, обладающие пестицидной и/или инсектицидной активностью, например, такие как белки-токсины Bacillus thuringiensis (описанные в патентах США № 5366892; 5747450; 5737514; 5723756; 5593881; и Geiser et al. (1986) Gene 48:109). Полученные комбинации могут также содержать множественные копии любого из интересующих полинуклеотидов.
Нуклеотидные последовательности, описанные в настоящем документе, можно также использовать в смысловой ориентации для супрессии экспрессии эндогенных генов в растениях. Способы супрессии экспрессии генов в растениях с использованием нуклеотидных последовательностей в смысловой ориентации известны в данной области. Способы, как правило, включают в себя трансформацию растений конструкцией ДНК, содержащей промотор, управляющий экспрессией в растении, функционально связанный, по меньшей мере, с частью нуклеотидной последовательности, соответствующей транскрипту эндогенного гена. Как правило, такая нуклеотидная последовательность обладает значительной идентичностью последовательности с последовательностью транскрипта эндогенного гена, предпочтительно, более чем приблизительно 65% идентичностью последовательности, более предпочтительно, более чем приблизительно 85% идентичностью последовательности, наиболее предпочтительно, более чем приблизительно 95% идентичностью последовательности. См. патенты США № 5283184 и 5034323, содержание которых приведено в настоящем документе в качестве ссылки.
Кроме того, представлены способы увеличения устойчивости к гербицидам растения подсолнечника посредством общепринятой селекции растений, включающей половое размножение. Способы включают в себя скрещивание первого растения подсолнечника, которое является устойчивым к гербициду, со вторым растением подсолнечника, которое может являться или может не являться устойчивым к гербициду или может являться устойчивым к другому гербициду или гербицидам, чем первое растение. Первое растение может представлять собой любое из устойчивых к гербицидам растений, описанных в настоящем документе, включая, например, RW-B. Второе растение может представлять собой любое растение, способное производить жизнеспособные растения-потомки (т.е. семена) при скрещивании с первым растением. Как правило, но не обязательно, первое и второе растения принадлежат к одному и тому же виду подсолнечника. Растения-потомки, полученные этим способом, обладают увеличенной устойчивостью к гербицидам по сравнению с первым или вторым растением, или с обоими. Когда первое и второе растения являются устойчивыми к разным гербицидам, растения-потомки обладают комбинированными характеристиками толерантности к гербицидам первого и второго растения. Эти способы могут дополнительно включать в себя одно или несколько поколений обратного скрещивания растений-потомков первого скрещивания с растением той же линии или генотипа, как либо первое, либо второе растение. Альтернативно, потомство первого скрещивания или любого последующего скрещивания можно скрещивать с третьим растением из другой линии или с другим генотипом по сравнению с первым или вторым растением. Кроме того, эти способы могут включать в себя отбор растений, обладающих характеристиками толерантности к гербицидам первого растения, второго растения, или и первого, и второго растения.
Способы контроля над сорняками
Сельскохозяйственные растения подсолнечника часто меняют в севообороте с зерновыми, такими как пшеница. Гербициды SU часто применяют как часть системы контроля над сорняками для пшеницы. В дополнение к гербицидам SU, гербициды IMI можно использовать как часть системы контроля над сорняками для пшеницы при выращивании IMI пшеницы. Применение гербицидов в смене пшеница/подсолнечник в севообороте приводит к повреждению из-за примесей, переходящих в новый цикл. Повреждение подсолнечника из-за примесей гербицидов, переходящих в новый цикл, происходит, когда остатки гербицида, применяемого для предыдущего сельскохозяйственного растения, в этом примере SU или IMI, применяемого для пшеницы, присутствуют в почве при посеве сельскохозяйственного растения подсолнечника. Действительно, некоторые гербициды могут вызывать повреждение подсолнечника из-за примесей, переходящих в новый цикл, включая многие AHAS-ингибиторы (Blarney et al., 1997). Повреждение сельскохозяйственных растения подсолнечника можно уменьшать с использованием устойчивых к гербицидам культиваров подсолнечника.
Устойчивые к гербицидам культивары подсолнечника, однако, обладают высокой специфичностью и их использование позволяет только опрыскивание тем же самым семейством гербицидов после появления всходов. Например, чтобы защитить сельскохозяйственное растение подсолнечника от повреждения из-за примесей SU, переходящих в новый цикл, можно использовать SU-устойчивые культивары. Использование SU-устойчивых культиваров позволяет распыление только гербицидов SU над сельскохозяйственным растением подсолнечника. К настоящему времени показаны только SU-устойчивые или IMI-устойчивые специфические культивары - не описаны культивары, обладающие перекрестной устойчивостью к различным AHAS-ингибиторам. Удивительно и неожиданно, присутствие гена, описанного в настоящем документе, и экспрессия соответствующего белка AHAS защищает сельскохозяйственные растения подсолнечника от потенциального повреждения из-за примесей, переходящих в новый цикл, вызванного AHAS-ингибирующими гербицидами и следовательно позволяет распыление множества AHAS-ингибирующих гербицидов для контроля над сорняками после появления всходов. Эта возможность использования множества гербицидов для контроля над сорняками в случае сельскохозяйственного растения подсолнечника обеспечивает растениеводам намного увеличенную возможность контролировать зарастание сорняками сельскохозяйственных растений подсолнечника.
Неожиданное открытие белка AHAS, обладающего устойчивостью ко множеству гербицидов, обеспечивает:
1. Возможность применения новых типов AHAS-ингибирующих гербицидов для контроля над сорняками в случае сельскохозяйственного растения подсолнечника (т.е.: POB и TP) или регистрации для подсолнечника хорошо известных гербицидов, применяемых для других сельскохозяйственных растений.
2. Смешивание двух или более гербицидов для применения в одно и то же время для сельскохозяйственного растения подсолнечника из-за высоких уровней перекрестной устойчивости к различным семействам AHAS-ингибирующих гербицидов. Кроме того, перекрестная устойчивость позволяет дизайн и разработку новых составов гербицидов, которые могут комбинировать различные AHAS-ингибирующие гербициды с целью увеличения спектра контроля над сорняками в случае подсолнечника.
3. Большую гибкость в организации использования гербицидов для контроля над сорняками в случае подсолнечника, то есть более одного типа гербицидов можно выбирать и применять в различные периоды времени одного и того же сезона в зависимости от типа сорняков, подлежащих контролю.
4. Устойчивость к повреждению из-за примесей гербицидов, переходящих в новый цикл после предыдущего сельскохозяйственного растения, и, в то же самое время, устойчивость к другому AHAS-ингибирующему гербициду, применяемому после появления всходов.
Контроль над нежелательной растительностью понимают как обозначающий уничтожение сорняков и/или иным образом замедление или ингибирование нормального роста сорняков. Сорняки, в самом широком смысле, понимают как обозначающие все растения, растущие в тех местах, где они являются нежелательными. Неограничивающие примеры сорняков включают в себя двудольные и однодольные сорняки. Двудольные сорняки включают в себя в качестве неограничивающих примеров сорняки из родов: Sinapis, Lepidium, Galium, Stellaria, Matricaria, Anthemis, Galinsoga, Chenopodium, Urtica, Senecio, Amaranthus, Portulaca, Xanthium, Convolvulus, Ipomoea, Polygonum, Sesbania, Ambrosia, Cirsium, Carduus, Sonchus, Solanum, Rorippa, Rotala, Lindernia, Lamium, Veronica, Abutilon, Emex, Datura, Viola, Galeopsis, Papaver, Centaurea, Trifolium, Ranunculus и Taraxacum. Однодольные сорняки включают в себя в качестве неограничивающих примеров, сорняки из родов: Echinochloa, Setaria, Panicum, Digitaria, Phleum, Poa, Festuca, Eleusine, Brachiaria, Lolium, Bromus, Avena, Cyperus, Sorghum, Agropyron, Cynodon, Monochoria, Fimbristyslis, Sagittaria, Eleocharis, Scirpus, Paspalum, Ischaemum, Sphenoclea, Dactyloctenium, Agrostis, Alopecurus и Apera.
Кроме того, сорняки могут включать в себя, например, сельскохозяйственные растения, растущие в нежелательном месте. Например, выросшее самосевом растение кукурузы, присутствующее на поле, где преимущественно присутствуют растения подсолнечника, можно рассматривать как сорняк, если растение кукурузы является нежелательным на поле с растениями подсолнечника.
Варианты осуществления настоящего изобретения относятся к способам увеличения толерантности или устойчивости растения, ткани растения, клетки растения или другой клетки-хозяина к одному или нескольким гербицидам, мешающим активности фермента AHAS. Предпочтительно, такой гербицид представляет собой гербицид IMI, гербицид SU, гербицид TP, гербицид POB и/или их смеси. Преимущественно увеличивается устойчивость или толерантность к двум или более из вышеуказанных гербицидов. Более преимущественно, увеличивается устойчивость или толерантность к трем или более из вышеуказанных гербицидов. Наиболее преимущественно, увеличивается устойчивость или толерантность к четырем или более из вышеуказанных гербицидов. Гербициды IMI могут включать в себя в качестве неограничивающих примеров, PURSUIT® (имазетапир), CADRE® (имазапик), RAPTOR® (имазамокс), SCEPTER® (имазаквин), ASSERT® (imazethabenz), ARSENAL® (имазапир), производное любого из вышеупомянутых гербицидов, и смесь двух или более из вышеупомянутых гербицидов, например, имазапир/имазамокс (ODYSSEY®). Гербицид IMI может являться выбранным, в качестве неограничивающих примеров, из 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-никотиновой кислоты, [2-(4-изопропил)-4-J b метил-5-оксо-2-имидазолин-2-ил)-3-хинолинкарбоновой] кислоты, [5-этил-2-(4-изопропил-]-4-метил-5-оксо-2-имидазолин-2-ил)-никотиновой кислоты, 2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)-никотиновой кислоты, [2-(4-изопропил-4-метил-5-оксо-2-J имидазолин-2-ил)-5-метилникотиновой кислоты и смеси метил-[6-(4-изопропил-4-]метил-5-оксо-2-имидазолин-2-ил)-м-толуата и метил-[2-(4-изопропил-4-метил-5-]оксо-2-имидазолин-2-ил)-п-толуата. Применение 5-этил-2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-ил)-никотиновой кислоты и [2-(4-изопропил-4-метил-5-оксо-2-имидазолин-2-]-ил)-5-(метоксиметил)-никотиновой кислоты является предпочтительным. Применение [2-(4-изопропил-4-]-метил-5-оксо-2-имидазолин-2-ил)-5-(метоксиметил)-никотиновой кислоты является особенно предпочтительным.
Гербициды SU могут включать в себя в качестве неограничивающих примеров хлорсульфурон, метсульфурон-метил, сульфометурон-метил, хлоримурон-этил, тифенсульфурон-метил, трибенурон-метил, бенсульфурон-метил, никосульфурон, этаметсульфурон-метил, римсульфурон, трифлусульфурон-метил, триасульфурон, примисульфурон-метил, циносульфурон, амидосульфурон, флузасульфурон, имазосульфурон, пиразосульфурон-этил, галосульфурон, азимсульфурон, циклосульфурон, этоксисульфурон, флазасульфурон, флупирсульфурон-метил, форамсульфурон, иодосульфурон, оксасульфурон, мезосульфурон, просульфурон, сульфосульфурон, трифлоксисульфурон, тритосульфурон, производные любого из вышеуказанных гербицидов и смесь двух или более из вышеуказанных гербицидов.
Гербициды TP могут включать в себя в качестве неограничивающих примеров клорансулам, диклосулам, флорасулам, флуметсулам, метосулам и пенокссулам.
Гербициды POB могут включать в себя в качестве неограничивающих примеров биспирибак, пиритиобак, пириминобак, пирибензоксим и пирифталид.
Известно, что гербициды POB являются близко родственными пиримидинилтиобензоатным гербицидам и являются обобщенными под заголовком последнего наименования в Американском обществе по изучению сорняков. Соответственно, гербициды могут включать в себя пиримидинилтиобензоатные гербициды, включая, в качестве неограничивающих примеров, гербициды POB, описанные выше. В свете вышеизложенного, заявители используют термины пиримидинилтиобензоат и пиримидинилоксибензоат взаимозаменяемо с аббревиатурой POB.
Применение гербицидов, перечисленных в настоящем документе, не предназначено для ограничения настоящего изобретения конкретным гербицидом или конкретным семейством гербицидов. Специалистам в данной области понятно, что можно применять также другие гербициды.
Посредством предоставления растений, обладающих увеличенной устойчивостью к гербицидам, можно применять широкое множество составов для защиты растений подсолнечника от сорняков, так чтобы улучшать рост растений подсолнечника и уменьшать конкуренцию за питательные вещества. Гербицид сам по себе можно применять для контроля над сорняками в областях, окружающих растения, описанные в настоящем документе, до появления всходов, после появления всходов, до высадки и при высадке. В одном варианте осуществления можно использовать состав гербицида IMI, содержащий другие добавки. В другом варианте осуществления можно использовать состав гербицида SU, содержащий другие добавки. В другом варианте осуществления можно использовать состав гербицида TP, содержащий другие добавки. В другом варианте осуществления можно использовать состав гербицида POB, содержащий другие добавки. Гербицид(ы) можно использовать также для обработки семян. Добавки, обнаруженные в составах гербицида IMI, гербицида SU, гербицида TP, гербицида POB и/или их смесей, могут включать в себя другие гербициды, детергенты, адъюванты, усиливающие растекание средства, усиливающие прилипание средства, стабилизаторы или т.п. Состав гербицида может представлять собой влажный или сухой препарат и может включать в себя, но не ограничиваться ими, сыпучие порошки, эмульгируемые концентраты и жидкие концентраты. Гербицид и составы гербицидов можно применять согласно общепринятым способам, например посредством опрыскивания, орошения, распыления или т.п. В этих способах AHAS-ингибирующий гербицид можно применять любым способом, известным в данной области, включая, в качестве неограничивающих примеров, обработку семян, обработку почвы и внекорневую обработку.
Перед применением AHAS-ингибирующий гербицид можно переводить в общепринятые составы, например растворы, эмульсии, суспензии, пыль, порошки, пасты и гранулы. Форма применения зависит от конкретной намеченной цели; в каждом случае она должна обеспечивать хорошее и равномерное распределение соединения в соответствии с настоящим изобретением.
Составы получают известным способом (см., например, обзор в US 3060084, EP-A 707 445 (для жидких концентратов), Browning, «Agglomeration», Chemical Engineering, Dec. 4, 1967, 147-48, Perry's Chemical Engineer's Handbook, 4th Ed., McGraw-Hill, New York, 1963, pages 8-57 и et seq. WO 91/13546, US 4172714, US 4144050, US 3920442, US 5180587, US 5232701, US 5208030, GB 2095558, US 3299566, Klingman, Weed Control as a Science, John Wiley and Sons, Inc., New York, 1961, Hance et al., Weed Control Handbook, 8th Ed., Blackwell Scientific Publications, Oxford, 1989 and Mollet, H., Grubemann, A., Formulation technology, Wiley VCH Verlag GmbH, Weinheim (Germany), 2001, 2. D. A. Knowles, Chemistry and Technology of Agrochemical Formulations, Kluwer Academic Publishers, Dordrecht, 1998 (ISBN 0-7514-0443-8), например, посредством дополнения активного соединения вспомогательными веществами, подходящими для составления агрохимикатов, такими как растворители и/или носители, если желательно, эмульгаторы, поверхностно-активные вещества и диспергирующие средства, консерванты, противовспенивающие средства, средства против замерзания, в случае составов для обработки семян, также, необязательно, окрашивающие и/или связующие вещества и/или желирующие средства.
Примеры подходящих растворителей представляют собой воду, ароматические растворители (например, ароматические фракции нефти, ксилол), парафины (например, фракции минерального масла), спирты (например, метанол, бутанол, пентанол, бензиловый спирт), кетоны (например, циклогексанон, гамма-бутиролактон), пирролидоны (NMP, NOP), ацетаты (диацетат гликоля), гликоли, диэтиламиды жирных кислот, жирные кислоты и сложные эфиры жирных кислот. В принципе, можно использовать также смеси растворителей.
Примеры пригодных носителей представляют собой измельченные природные минералы (например, каолины, глины, тальк, мел) и размолотые синтетические минералы (например, высокодисперсный диоксид кремния, силикаты). Пригодные эмульгаторы представляют собой неионные и анионные эмульгаторы (например, эфиры жирного спирта и полиоксиэтилена, алкилсульфонаты и арилсульфонаты).
Примеры диспергирующих средств представляют собой отработанный лигнин-сульфитный щелок и метилцеллюлозу.
Применяемые пригодные поверхностно-активные вещества представляют собой соли щелочных металлов, щелочноземельных металлов и аммония лигнинсульфоновой кислоты, нафталинсульфоновой кислоты, фенолсульфоновой кислоты, дибутилнафталинсульфоновой кислоты, алкиларилсульфонаты, алкилсульфаты, алкилсульфонаты, сульфаты жирных спиртов, эфиры жирных кислот и сульфатированных гликолей жирных спиртов, кроме того, продукты реакции конденсации сульфированного нафталина и производных нафталина с формальдегидом, продукты реакции конденсации нафталина или нафталинсульфоновой кислоты с фенолом и формальдегидом, эфир полиоксиэтилена и октилфенола, этоксилированный изооктилфенол, октилфенол, нонилфенол, эфир алкилфенола и полигликоля, эфир трибутилфенила и полигликоля, эфир тристеарилфенила и полигликоля, алкилариловые полиэфиры спиртов, продукты реакции конденсации спирта и жирного спирта и этиленоксида, этоксилированное касторовое масло, алкиловые эфиры полиоксиэтилена, этоксилированный полиоксипропилен, ацеталь эфира лаурилового спирта и полигликоля, сложные эфиры сорбита, отработанный лигнин-сульфитный щелок и метилцеллюлозу.
Вещества, пригодные для получения поддающихся непосредственному распылению растворов, эмульсий, паст или масляных дисперсий, представляют собой фракции минерального масла с температурой кипения от средней до высокой, такие как керосин или дизельное масло, кроме того, каменноугольные масла и масла растительного или животного происхождения, алифатические, циклические и ароматические углеводороды, например толуол, ксилол, парафин, тетрагидронафталин, алкилированные нафталины или их производные, метанол, этанол, пропанол, бутанол, циклогексанол, циклогексанон, изофорон, высокополярные растворители, например диметилсульфоксид, N-метилпирролидон или воду.
А также средства против замерзания, такие как глицерин, этиленгликоль, пропиленгликоль, и бактерицидное средство можно добавлять в состав. Пригодными противовспенивающие средствами являются, например, противовспенивающие средства на основе силикона или стеарата магния.
Пригодные консерванты представляют собой, например, дихлорофен и гемиформаль бензилового спирта. Составы для обработки семян подсолнечника могут, кроме того, содержать связующие вещества и, необязательно, окрашивающие вещества. Связующие вещества можно добавлять для улучшения адгезии активных веществ на семенах после обработки. Пригодные связующие вещества представляют собой блок-сополимеры поверхностно-активных веществ EO/PO, но также поливинилспирты, поливинилпирролидоны, полиакрилаты, полиметакрилаты, полибутены, полиизобутилены, полистирол, полиэтиленамины, полиэтиленамиды, полиэтиленимины (Lupasol®, Polymin®), полиэфиры, полиуретаны, поливинилацетат, тилозу и сополимеры, полученные из этих полимеров.
Необязательно, в состав можно включать окрашивающие вещества. Пригодные окрашивающие вещества или красители для составов для обработки семян представляют собой Родамин B, C.I. пигмент красный 112, C.I. растворитель красный 1, пигмент синий 15:4, пигмент синий 15:3, пигмент синий 15:2, пигмент синий 15:1, пигмент синий 80, пигмент желтый 1, пигмент желтый 13, пигмент красный 112, пигмент красный 48:2, пигмент красный 48:1, пигмент красный 57:1, пигмент красный 53:1, пигмент оранжевый 43, пигмент оранжевый 34, пигмент оранжевый 5, пигмент зеленый 36, пигмент зеленый 7, пигмент белый 6, пигмент коричневый 25, основной фиолетовый 10, основной фиолетовый 49, кислый красный 51, кислый красный 52, кислый красный 14, кислый синий 9, кислый желтый 23, основной красный 10, основной красный 108.
Примером пригодного желирующего средства является карраген (Satiagel®). Порошки, материалы для распределения и пригодные для распыления продукты можно получать смешиванием или одновременным измельчением активных веществ с твердым носителем. Гранулы, например гранулы с покрытием, гранулы с вкраплениями и гомогенные гранулы, можно получать связыванием активных соединений с твердыми носителями. Примеры твердых носителей представляют собой природные ископаемые, такие как силикагели, силикаты, тальк, каолин, аттаглина, известняк, известь, мел, железистая известковистая глина, лесс, глина, доломит, диатомовая земля, сульфат кальция, сульфат магния, оксид магния, измельченные синтетические материалы, удобрения, например, такие как сульфат аммония, фосфат аммония, нитрат аммония, мочевины, и продукты растительного происхождения, такие как мука зерновых, мука из коры деревьев, древесная мука и мука из ореховой скорлупы, целлюлозные порошки и другие твердые носители.
Как правило, составы содержат от 0,01 до 95% по массе, предпочтительно, от 0,1 до 90% по массе, AHAS-ингибирующего гербицида. В этом случае AHAS-ингибирующие гербициды применяют с чистотой от 90% до 100% по массе, предпочтительно, 95%-100% по массе (согласно спектру NIVIR). Для целей обработки семян подсолнечника соответствующие составы можно разводить в 2-10 раз, что приводит к концентрациям в готовых для использования препаратах 0,01-60% активного соединения по массе, предпочтительно, 0,1-40% по массе.
AHAS-ингибирующие гербициды можно использовать как таковые, в форме их составов или полученных из них форм для использования, например в форме поддающихся прямому распрыскиванию растворов, порошков, суспензий или дисперсий, эмульсий, масляных дисперсий, паст, поддающихся распылению продуктов, материалов для распределения или гранул, посредством опрыскивания, разбрызгивания, распыления, распределения или заливки. Формы для использования полностью зависят от предназначенных целей; они предназначены для обеспечения в каждом случае наилучшего возможного распределения AHAS-ингибирующего гербицида в соответствии с настоящим изобретением.
Водные формы для использования можно получать из концентратов эмульсий, паст или смачиваемых порошков (поддающихся распылению порошков, масляных дисперсий) добавлением воды. Для получения эмульсий, паст или масляных дисперсий вещества сами по себе или растворенные в масле или растворителе можно гомогенизировать в воде посредством смачивателя, повышающего клейкость вещества, диспергирующего средства или эмульгатора. Однако возможно также получать концентраты, состоящие из активного вещества, смачивателя, повышающего клейкость вещества, диспергирующего средства или эмульгатора и, если необходимо, растворителя или масла, и такие концентраты являются пригодными для разведения водой.
Концентрации активного соединения в готовых для использования препаратах могут меняться в относительно широких диапазонах. Как правило, они составляют 0,0001-10%, предпочтительно от 0,01 до 1% по массе.
В одном варианте осуществления настоящего изобретения дополнительным объектом настоящего изобретения является способ обработки почвы посредством применения, в частности, в сеялке: либо гранулярного состава, содержащего AHAS-ингибирующий гербицид в форме композиции/состава (например, гранулярного состава) необязательно с одним или несколькими твердыми или жидкими применимыми в области сельского хозяйства носителями и/или, необязательно, с одним или несколькими применимыми в области сельского хозяйства поверхностно-активными веществами. Этот способ преимущественно применяют для предпосевной почвообработки для подсолнечника.
Варианты осуществления настоящего изобретения относятся также к семенам подсолнечника, покрытым составом для обработки семян и/или содержащим состав для обработки семян, содержащим, по меньшей мере, один AHAS-ингибирующий гербицид, выбранный из группы, состоящей из амидосульфурона, азимсульфурона, бенсульфурона, хлоримурона, хлорсульфурона, циносульфурона, циклосульфамурона, этаметсульфурона, этоксисульфурона, флазасульфурона, флупирсульфурона, форамсульфурона, галосульфурона, имазосульфурона, иодосульфурона, мезосульфурона, метсульфурона, никосульфурона, оксасульфурона, примисульфурона, просульфурона, пиразосульфурона, римсульфурона, сульфометурона, сульфосульфурона, тифенсульфурона, триасульфурона, трибенурона, трифлоксисульфурона, трифлусульфурон, тритосульфурона, имазаметабенза, имазамокса, имазапика, имазапира, имазаквина, имазетапира, клорансулама, диклосулама, флорасулама, флуметсулама, метосулама, пенокссулама, биспирибака, пириминобака, пропоксикарбазона, флукарбазона, пирибензоксима, пирифталида и пиритиобака.
Термин «семя» относится к семенам и пропагулам растений всех типов, включая в качестве неограничивающих примеров истинные семена, части семян, корневые побеги, клубнелуковицы, луковицы, плоды, клубнеплоды, зерна, черенки, побеговые черенки и т.п., и обозначает, в предпочтительном варианте осуществления, истинные семена.
Термин «покрытый и/или содержащий», как правило, указывает на то, что активный ингредиент по большей части находится на поверхности продукта размножения на время применения, хотя большая или меньшая часть ингредиента может проникать в продукт размножения, в зависимости от способа применения. Когда продукт размножения (повторно) высаживают, он может абсорбировать активный ингредиент.
Обрабатывающее нанесение на семена подсолнечника AHAS-ингибирующего гербицида или состава, содержащего AHAS-ингибирующий гербицид, проводят опрыскиванием или опылением семян до посева растений и до появления растений.
При обработке семян подсолнечника соответствующие составы наносят обработкой семян эффективным количеством AHAS-ингибирующего гербицида или состава, содержащего AHAS-ингибирующий гербицид. В настоящей заявке дозы для применения, как правило, составляют от 0,1 г до 10 кг активного ингредиента («a.i.») (или смеси a.i. или состава) на 100 кг семян, предпочтительно, от 1 г до 5 кг на 100 кг семян, в частности, от 1 г до 2,5 кг на 100 кг семян. Следующие примеры предложены для иллюстрации, а не для ограничения.
Идентификация устойчивых растений
300 семян из дикой популяции подсолнечника, известной как толерантная к IMI, высевали в условиях теплицы в Nidera Experimental Station в Venado Tuerto, Santa Fe, Argentina, вместе с контрольными линиями HA89, B770 (Чувствительный - «S»), BTI-M1 и RHA426 (Устойчивый - «R»). На стадии V4 все растения опрыскивали имазапиром в дозе 80 г a.i./га. Через две недели после обработки все растения оценивали по повреждениям посредством визуального осмотра. S линии HA89 и B770 были уничтожены обработкой. Для R линии RHA426 показали признаки хлороза, и в R линии BTI-M1 не присутствовало каких-либо признаков повреждения. Всего 159 растений из дикой популяции выдержало обработку гербицидами, и 129 были уничтожены (отнесены к категории S). Кроме того, для 18 из 159 растений, переживших обработку гербицидами, не показали симптомов повреждения гербицидами, как и для мутантной R линии BTI-M1, и они отнесены к категории R.
Генетика устойчивости
Одно R растение (RW-73) использовали в качестве мужского в скрещиваниях с культивируемыми чувствительными линиями HA89 и B770. От десяти до 20 F1-растений после каждого скрещивания опрыскивали имазапиром в дозе 80 г a.i./га, с использованием B770 (S) и BTI-M1 (R) в качестве контроля. Ни для одного из F1-растений не показали симптомов повреждения гербицидами, что являлось сходным с растениями BTI-M1. Однако обработка имазапиром уничтожала растения B770.
Проводили обратное скрещивание одного F1-растения из скрещивания HA89/RW-73 с чувствительным родителем. А также проводили обратное скрещивание одного растения из скрещивания B770/RW-73 с B770. F2-семена получали опылением F1-растений среди них.
Полученные BC1F1 и F2-семена высевали в условиях теплицы вместе с B770 и BTI-M1, используемыми в качестве чувствительного и устойчивого контроля, соответственно. На стадии V3-V4 все растения опрыскивали имазапиром в дозе 80 г a.i./га. Растения через 14 суток после обработки относили к категориям R, S или I (промежуточное). Растения относили к категории R, если для них не показывали повреждения гербицидом, S, если они погибали, или I, если для них показывали уменьшение высоты или хлороз. Результаты наследования устойчивости к имидазолинону в RW-73 показаны в таблице 1, показанной на фиг.3.
В частности, в таблице 1 показан ответ растений подсолнечника [устойчивое (R), промежуточное (I), чувствительное (S)] к имазапиру, применяемому в дозе 80 г a.i./га в популяциях F1, F2 и BC1F1, полученных в результате скрещивания между устойчивой линией, RW-73, и одной из каждой из двух чувствительных линий, B770 и HA89, и проводили тесты χ2 на однолокусной модели для контроля устойчивости.
Для F2-популяций, полученных в результате скрещиваний чувствительный Ч устойчивый, показали хорошее соответствие соотношению 1:2:1 R:I:S, что указывает на сегрегацию одного гена устойчивости к имазапиру. Для подтверждения этих результатов по F2-популяциям проводили тестовое скрещивание F1-растений с чувствительными родителями, и полученное потомство оценивали по реакции на имазапир. Для популяций BC1F1 получили хорошее соответствие соотношению 1:1 I:S, подтверждающее гипотезу одного локуса (таблица 1).
Результаты генетического исследования показывают, что устойчивость в RW-73 наследуется как частично доминантный признак, придаваемый одним кодируемым в ядре геном. Этот характер наследования согласуется с другими обнаружениями, где отмечен генетический контроль устойчивости к ингибирующим AHAS гербицидам.
Разработка линии подсолнечника, являющейся гомозиготной по характеристике устойчивости к гербицидам RW-73
Гетерогенное и гетерозиготное состояние дикой популяции подсолнечника, анализированной выше, препятствует ее использованию для сравнений относительного уровня устойчивости по отношению к культивируемым линиям, обладающим признаками устойчивости к гербицидам. С использованием обратного скрещивания с использованием маркера разработали устойчивую линию, близко совпадающую с B770, закодированную как RW-B. RW-B являлась гомозиготной по гену устойчивости, поскольку ее потомство, полученное посредством самовоспроизводства, не сегрегирует по чувствительности после воздействия имазапира.
Устойчивость RW-B к различным AHAS-ингибирующим гербицидам
Линии подсолнечника: Следующие генетические линии подсолнечника использовали для определения устойчивости AHAS: две S линии («B770» и «HA89»), две линии, устойчивые к SU, полученные из популяции SURES (SuBL и SuRL), две линии, устойчивые к IMI, полученные из популяции IMISUN («Rha 426» и «B770imi») и линию «RW-B». HA89 и Rha426 являются инбредными линиями, выпущенными USDA. SURES и IMISUN представляют собой селекционные популяции, выпущенные USDA. B770 представляет собой патентованную линию из Nidera. B770imi представляет собой патентованную линию с фоном B770, несущую ген устойчивости Imr1. RW-B получали, как описано выше.
Метод: По восемь семян из каждой из семи линий высевали в горшки 20Ч20Ч30 см. После появления всходов сеянцы прореживали вручную, оставляя 4 сеянца на горшок. По пять горшков из каждой линии обрабатывали различными гербицидами или дозами. В каждый эксперимент включали необработанный контрольный горшок из каждой линии. Растения выращивали в теплице в условиях естественного дневного света, дополненного галогенными лампами по 400 В для обеспечения длительности дня 14 ч. Дневная/ночная температуры составляли 25 и 20°C, соответственно. На стадии развития V4 (Schneiter and Miller 1981) растения опрыскивали тремя гербицидами SU (хлорсульфурон, никосульфурон и форамсульфурон), тремя IMI (имазамокс, имазапир и имазапик), POB (биспирибак-Na) и TP (клорансулам-метил). Через четырнадцать суток после обработки растения оценивали фенотипически, с использованием показателя фитотоксичности (PI). PI представляет собой фенотипическую шкалу от 0 до 9, по которой оценивали каждый горшок посредством визуального осмотра. Растения без каких-либо симптомов регистрировали как «0», увеличенные уровни низкорослости и пожелтения по отношению к необработанным контрольным растениям регистрировали как «1»-«4», увеличенные уровни аномалий листа и некроза листа регистрировали от «5» до «8», и погибшие растения с полным некрозом апекса регистрировали как «9». Данные PI для каждого эксперимента анализировали посредством теста ANOVA, средние сравнивали посредством теста LSD.
Результаты показаны в таблице 2 из фиг.4, в которой показан показатель фитотоксичности семи линий подсолнечника, опрысканных различными AHAS-ингибирующими гербицидами; таблица 2. Ответ линий подсолнечника (B770, HA89, SuBL, SuRL, Rha426, B770imi и RW-B) на четыре различных семейства ингибирующих AHAS гербицидов. Данные представляют MS среди среднеквадратической суммы для линий, *** P<0,001, LSD значения наименьшей значимой разности. Средние значения с одинаковой буквой показывают, что они не являются статистически различными (P<0,01). Повреждение измеряли с использованием показателя фитотоксичности (PI).
Линии S HA89 и B770 были уничтожены при воздействиях всех гербицидов. SU-устойчивые линии SuBL и SuRL являлись устойчивыми к трем использованным SU, но были уничтожены всеми другими гербицидами. IMI-устойчивые линии Rha426 и B770imi являлись умеренно устойчивыми к трем IMI и являлись высоко чувствительными к SU, POB и TP. Примечательно, что для RW-B, показали высокие уровни устойчивости ко всем четырем семействам гербицидов. Неожиданно, для линии RW-B показали такой же уровень устойчивости к SU, как для SU-устойчивых линий SuBL и SuRL, и лучший уровень устойчивости к IMI, чем у IMI-устойчивых линий Rha426 и B770imi. Кроме того, по другому неожиданному результату, для RW-B показали высокий уровень устойчивости к POB и TP. Ни для одной из других линий не показали этого типа устойчивости.
Тест на аллелизм
RW-B растения скрещивали с RHA426, чтобы определить, является ли ген устойчивости в RW-B аллельным по отношению к Imr1, мутации, ранее описанной в устойчивых к имидазолинону инбредных линиях. При этом скрещивании F1-гибридные растения подвергали самоопылению и обратному скрещиванию с RHA426 для получения F2- и BC1F1-поколений, соответственно. Растения из родительских и из F1, F2 и BC1F1 испытывали с помощью 160 г a.i./га и 320 г a.i./га имазапира на стадии V2-V4. После испытания растениям приписывали баллы, как описано выше. Не наблюдали чувствительных растений в популяциях F2 и BC1F1, полученных в результате этого скрещивания, когда потомство оценивали при более низкой дозе гербицида, что указывает на то, что гены устойчивости в RW-B и RHA426 представляют собой аллели одного и того же локуса. Эти результаты показаны в таблице 3 на фиг.5. В частности, в таблице 3 показан ответ растений подсолнечника (т.е. как устойчивых (R) и чувствительных (S)) на имазапир, применяемый в дозах 80 г a.i./га и 320 г a.i./га в поколениях F1, F2 и BC1F1, полученных в результате скрещиваний между RW-B и инбредной линией, несущей ген устойчивости к имидазолинону Imr1, RHA 426.
С другой стороны, когда популяции F2 и BC1F1 оценивали после применения гербицида в более высокой дозе (320 г a.i./га), можно было наблюдать сегрегацию по чувствительности. Детектировали только два фенотипических класса, устойчивый класс, состоящий из растений без каких-либо признаков повреждения или только со слабыми симптомами, и чувствительный фенотип, который был уничтожен при этой дозе примерно одинаково с контрольной линией RHA426. Наблюдаемые степени сегрегации среди 508 подвергнутых скринингу F2-растений значимо не отличались от степени сегрегации 3:1 (см. таблицу 3). Для подтверждения этих результатов проводили обратное скрещивание F1-растений с RHA426 и проводили скрининг полученных BC1F1-растений при 320 г a.i./га имазапира. Для наблюдаемых степеней сегрегации получили хорошее соответствие соотношению 1:1 R:S, подтверждая, что для гена устойчивости в RW-B показана полная доминантность над геном устойчивости в RHA426, и что оба гена устойчивости представляют собой аллели одного и того же локуса, AHAS1.
Косегрегация гаплотипа AHAS1 с устойчивостью к гербицидам
Материалы и методы: ДНК выделяли из молодых листьев линий B770, RW-B и RHA426, так же как из индивидуальных F1- и F2-растений после скрещивания RW-B/RHA426 с использованием способа Dellaporta (1983). Фрагменты гена AHAS1 амплифицировали с помощью ПЦР с использованием праймеров p-AHAS 18 и p-AHAS 19 (Kolkman et al., 2004). Продукты ПЦР амплифицировали в 15-мкл реакционной смеси, содержащей 1 ед. Taq ДНК-полимеразы (Biotools), 70 нг геномной ДНК подсолнечника, 25 мкг бычьего сывороточного альбумина (BSA), с конечной концентрацией 100 мкМ каждый dNTP, 0,25 мкМ каждый праймер, 90 мМ Трис-HCl pH 8, 20 мМ (NH4)2SO4 и 2,5 мМ MgCl2. Программа ПЦР состояла из начальной стадии денатурации при 94°C в течение 2 мин, с последующими 40 циклами 30 с при 94°C, 30 с при 56°C и 30 с при 72°C, с последующей финальной стадией элонгации при 72°C в течение 10 мин. Продукты амплификации (3 мкл/дорожку) разделяли в стандартном секвенирующем геле, содержащем 6% полиакриламид, 8 M мочевину и 1X TBE, при постоянной мощности 60 В в течение 2-3 ч и детектировали окрашиванием нитратом серебра (Silver Sequence™ Promega Biotech, USA). Размер каждого аллеля SSR определяли с использованием маркера молекулярной массы и продуктов стандартной реакции секвенирования в соседних дорожках геля.
Результаты: в гене AHAS1 подсолнечника присутствует полиморфизм простых повторов последовательности (SSR), позволяющий отличить линии, несущие аллель Imr1, от любого другого генотипа подсолнечника (Kolkman et al., 2004). Амплификацией ПЦР фрагмента гена AHAS1, содержащего этот SSR, с использованием праймеров p-AHAS18 и p-AHAS19 получали продукт 321 п.о. для RW-B и фрагмент 312 п.о. для RHA426. Этот полиморфизм варианта длины, детектируемый в RW-B и RHA426, использовали для исследования сегрегации в популяциях F2 и BC1F1, полученных в результате скрещивания обеих линий. Сто одиннадцать растений из популяции F2 и 126 растений из популяции BC1F1 выбирали случайным образом, отбирали образцы для выделения ДНК, проводили испытание имазапиром при применении дозы 320 г a.i./га и генотипирование с использованием этого маркера. В популяции F2 30 растений были уничтожены гербицидом (S), и для 81 не показали каких-либо симптомов или слабого повреждения (R). Эти данные показаны в таблице 4. В частности, в таблице 4 показана реакция растений подсолнечника (т.е. как устойчивых (R) и чувствительных (S)) на имазапир, применяемый в дозе 320 г a.i./га и сегрегация гаплотипа AHAS1 (A = гаплотип RW-B, B = гаплотип RHA426) в популяциях F2 и BC1F1, полученных в результате скрещивания RW-B/RHA426.
Наблюдаемая степень сегрегации для устойчивости значимо не отличалась от ожидаемой степени сегрегации для полностью доминантного фактора, сегрегирующего в F2 (3R:1S). Наблюдаемая сегрегация для маркера SSR AHAS1 (26 A/A: 55 A/B: 30 B/B) соответствует ожидаемой степени сегрегации для кодоминантного маркера, сегрегирующего в F2 (1:2:1). Все чувствительные растения, генотипированные по SSR AHAS1, являлись гомозиготными по гаплотипу RHA426 (B/B), где R-растения являлись гетерозиготными (A/B) или гомозиготными по гаплотипу RW-B (A/A) (см. таблицу 4). Косегрегацию фенотипов устойчивости к гербицидам и гаплотипов AHAS1 дополнительно оценивали в 126 потомках BC1F1, сегрегирующих по устойчивости. Наблюдаемые степени сегрегации для устойчивости соответствовали соотношению 1:1, как ожидали для сегрегации одного локуса в BC1F1. Гаплотипы SSR AHAS1 полностью косегрегировали с фенотипами по реакции на гербициды, 64 A/B: 62 B/B. Чувствительное потомство являлось гомозиготным по гаплотипу RHA426 (B/B), в то время как устойчивое потомство являлось гетерозиготным по гаплотипам RHA426 и RW-B (A/B). Эти результаты подтверждают, что ген устойчивости в RW-B является отличным от гена устойчивости в RHA426, и что оба гена являются аллельными вариантами локуса AHAS1.
Амплификация ПЦР и секвенирование гена AHAS1 подсолнечника из RW-B
Геномную ДНК выделяли из ткани листа подсолнечника с использованием набора DNeasy 96 Plant Qiagen (каталожный № 69181). Проводили амплификацию ПЦР гена AHAS1 в трех фрагментах и прямое секвенирование в Macrogen USA. Амплификацию ПЦР осуществляли с помощью Taq ДНК-полимеразы Qiagen's Hotstart и сопутствующих реагентов (каталожный № 203205). Праймеры для ПЦР для трех фрагментов являлись следующими:
AHAS1Fl (прямой праймер на парах оснований 1-19 общедоступной последовательности подсолнечника AY541451) 5'ΑTGGCGGCTCCTCCCAACC3',
AHAS1R1 (обратный праймер на парах оснований 757-777 общедоступной последовательности подсолнечника AY541451) 5'CGGTAACCTCATCGGTTCATC3',
AHAS1F2 (прямой праймер на парах оснований 757-777 общедоступной последовательности подсолнечника AY541451) 5'GATGAACCGATGAGGTTACCG3',
AHAS1R2 (обратный праймер на парах оснований 1794-1814 общедоступной последовательности подсолнечника AY541451) 5'TCCGCCTTTTGGGTCACTCGAS',
AHAS1F3 (прямой праймер на парах оснований 1248-1269 общедоступной последовательности подсолнечника AY541451) 5'GGTGACTAATCTTGATTTTTCG3' и
AHAS1R3 (обратный праймер на парах оснований 1949-1968 общедоступной последовательности подсолнечника AY541451) 5'TCAATATTTCGTTCTGCCAT3'.
Три фрагмента ПЦР перекрывают все участки мутаций, известных как придающие устойчивость к гербицидам IMI. Получили выравнивание нуклеотидов, и в полученных хроматограммах исследовали полиморфизмы между чувствительной линией B770 и устойчивой линией RW-B.
Результаты секвенирования гена AHASL1 подсолнечника
Продукты ПЦР секвенировали для получения последовательностей AHASL1 для линий подсолнечника RW-B и B770. Выравнивание этих нуклеотидных последовательностей и нуклеотидных последовательностей гена AHAS Xanthium sp. (Genbank инв. № U16280) показано на фиг.1.
В частности, на фиг.1 показано выравнивание частичной последовательности ДНК кодирующей области гена AHASL1 из HA89 (GenBank инв. № AY541451) (SEQ ID NO:3), линии B770 (SEQ ID NO:4), устойчивой линии RW-B (SEQ ID NO:5), гена ALS Xanthium sp. (GenBank инв. № U16280) (SEQ ID NO:6) и гена AHAS Arabidopsis thaliana (GenBank инв. № AY124092) (SEQ ID NO:7). Звездочкой указана одиночная мутация, обнаруженная в RW-B, замена этого основания отвечает за замену Trp-на-Leu в положении аминокислоты 574 (на основании последовательности Arabidopsis thaliana). Положения в нуклеотидной последовательности указаны стрелками и пронумерованы согласно последовательности A. thaliana.
Выравнивание выявило, что ген AHASL1 из RW-B обладает транзицией G-на-T относительно AHASL1 из B770.
Выравнивание предсказанных аминокислотных последовательностей из нуклеотидных последовательностей AHASL1 из RW-B, B770 и Xanthium sp представлено на фиг.2.
В частности, на фиг.2 показано выравнивание частичной аминокислотной последовательности белка AHASL1 из HA89 (GenBank инв. № AY541451) (SEQ ID NO:8), линии B770 (SEQ ID NO:9), устойчивой линии RW-B (SEQ ID NO:10), гена ALS Xanthium sp. (GenBank инв. № U16280) (SEQ ID NO:11) и гена AHAS Arabidopsis thaliana (GenBank инв. № AY124092) (SEQ ID NO:12). Звездочкой указана замена аминокислоты в кодоне 574 (на основании последовательности Arabidopsis thaliana).
По сравнению с аминокислотной последовательностью AHASL1 из B770, аминокислотная последовательность AHASL1 из RW-B обладает заменой триптофан-на-лейцин в положении аминокислоты 574 в полноразмерной аминокислотной последовательности, кодируемой нуклеотидной последовательностью AHASL Arabidopsis thaliana. По этой причине этот новый аллель локуса Ahasl1 обозначен в настоящей заявке W574L.
Сравнение агрономических характеристик (урожай семян и масличность) гибридов подсолнечника, несущих аллель W574L, по сравнению с общепринятыми гибридами (дикий тип) и гибридами, несущими аллель A205V (признак IMISUN)
Цель: Этот эксперимент проводили для количественной оценки и сопоставления агрономической продуктивности гибридов, несущих аллель W574L, с общепринятыми гибридами (дикий тип) и гибридами imisun (аллель A205V).
Материалы: Линию RW-B переводили в мужскую стерильную линию посредством обратного скрещивания с цитоплазматической мужской стерильной линией (cms B770). После 4 обратных скрещиваний полученную стерильную линию обозначили «cms RW-B».
Линию RW-B использовали также как источник устойчивости к гербицидам для превращения двух различных общепринятых восстанавливающих линий (дикого типа): R20 и R54. Полученные почти изогенные линии обозначены R20-574 и R54-574.
F1-гибриды между общепринятыми линиями, линиями, полученными из IMISUN и RW-B, получены в полевых условиях. Их генеалогическая схема и случаи мутаций показаны в следующей таблице.
представляют собой два зарегестрированных коммерческих гибрида,
известных как «Paraiso 33» и «Paraiso 103 CL», соответственно
Методы: Семена высевали в полевых условиях (Venado Tuerto, Santa Fe, Argentina) в дизайне полностью рандомизированного блока с четырьмя повторами. Каждый участок состоял из трех рядов длиной 6 метров и разделенных 0,7 м. Семена в ряду были разделены 0,30 м.
Следующие переменные регистрировали для каждого участка: сутки до цветения, высота при цветении, урожай (кг/га) и процент масла в семенах (%). С этими переменными проводили тест anova, и средние для различных случаев и гибридов сравнивали с помощью критерия Тьюки.
Результаты: Средние значения для каждого случая для 4 зарегистрированных переменных приведены в таблице 6. Как можно видеть, три случая обладали сходными средними значениями урожая. Анализы показали различия в масличности семян среди случаев, поскольку для гибридов с аллелем A205V получили более низкую масличность, чем для двух других типов гибридов. Сутки до цветения и высота растения также отличались между случаями, поскольку гибриды, несущие аллель W574L были немного меньшими и более ранними, чем два других типа гибридов. В заключение, для двух гибридов, несущих аллель W574L, показали одинаковый урожай и сравнимую масличность семян с общепринятыми гибридами (дикого типа) и гибридами IMISUN, которые включают в себя коммерческие гибридные сорта.
Средние для каждого гибрида для каждой из зарегистрированных переменных, и результаты анализа переменных приведены в таблице 7.
и масличности для 3 различных генотипов (случаев мутации) для
локуса AHASL1. Среднее значение для каждого генотипа или случая
мутации представляет собой среднее значение для двух различных
гибридов
значимых различий между случаями.
2 CV: коэффициент изменчивости
3 MS: среднее квадратичное среди случаев
4 p: значение вероятности F-величины.
5 MSD: минимальное значимое различие по Тьюки
подсолнечника, несущих три различных генотипа (случая мутации) для локуса AHASL1
Аллель
±SD1
±SD
± SD
±SD
гибридами: одинаковые буквы указывают на отсутствие статистически значимых различий.
2 CV: коэффициент изменчивости
3 MS: среднее квадратичное среди гибридов
4 p: значение вероятности F-величины.
5 MSD: минимальное значимое различие по Тьюки.
Толерантность линии RW-B к 4 различным смесям AHAS-ингибирующих гербицидов
Цель: сравнение продуктивности (оцененной как показатель фитотоксичности) линии, несущей аллель W574L (RW-B), и линий, несущих аллели P197L (SURES), A205V (IMISUN), и аллели дикого типа (общепринятые).
Материалы: Использовали следующие инбредные линии подсолнечника: две чувствительные к имидазолинону линии («B770» и «HA89»), две линии, устойчивые к SU гербицидам, полученные из популяции SURES («SuBL» и «SuRL»), две линии, устойчивые к IMI гербицидам, полученные из IMISUN («RHA426» и «B770imi»), и линии «RW-B», несущие аллель W574L. HA89 и RHA426 представляют собой общедоступные инбредные линии, выпущенные USDA. B770 и B770imi представляют собой запатентованные линии от Nidera S.A., в то время как B770imi обладает генетическим фоном B770, несущим ген устойчивости Imr1 (= аллель A205V) для устойчивости к имидазолинонам. RW-B получали, как описано выше.
Метод: По восемь семян из каждой из семи линий высевали в горшки 20Ч20Ч30 см. После появления всходов сеянцы прореживали вручную, оставляя 4 сеянца на горшок. По пять горшков из каждой линии обрабатывали различными смесями гербицидов. В каждый эксперимент включали необработанный контрольный горшок из каждой линии. Растения выращивали в теплице в условиях естественного дневного света, дополненного галогенными лампами по 400 В для обеспечения длительности дня 14 ч. Дневная/ночная температуры составляли 25 и 20°C, соответственно. На стадии развития V4 (Schneiter и Miller 1981) растения опрыскивали четырьмя различными смесями, содержащими два различных AHAS-ингибирующих гербицида, сульфонилмочевину (Метсульфурон) и имидазолинон (Имазапир). Концентрации гербицида для каждой из четырех смесей гербицидов являлись следующими: Смесь A представляла собой 20 г a.i./га имазапира и 1,25 г a.i./га метсульфурона, Смесь B представляла собой 40 г a.i./га имазапира и 2,5 г a.i./га метсульфурона, Смесь C представляла собой 80 г a.i./га имазапира и 5 г a.i./га метсульфурона, и Смесь D представляла собой 160 г a.i./га имазапира и 10 г a.i./га метсульфурона.
Через четырнадцать суток после обработки растения оценивали фенотипически, с использованием показателя фитотоксичности (PI). PI представляет собой фенотипическую шкалу от 0 до 9, по которой оценивали каждый горшок посредством визуального осмотра. Растения без каких-либо симптомов регистрировали как «0», увеличенные уровни низкорослости и пожелтения по отношению к необработанным контрольным растениям регистрировали как «1»-«4,» увеличенные уровни аномалий листа и некроза листа регистрировали от «5» до «8», и погибшие растения с полным некрозом апекса регистрировали как «9». Данные PI для каждого эксперимента анализировали посредством теста ANOVA, средние сравнивали посредством критерия Тьюки.
Результаты показаны в таблице 8. Линии HA89 и B770 были уничтожены при воздействиях всех смесей гербицидов. Для SU-устойчивых линий SuBL и SuRL показали некоторый уровень толерантности к Смеси A и Смеси B, но достигали высокого уровня некроза листьев и низкорослости при увеличении доз гербицидов, например, для Смеси C. Все растения были уничтожены Смесью D с наибольшими дозами активных ингредиентов. IMI-устойчивые линии RHA426 и B770imi являлись умеренно устойчивыми к Смеси A, Смеси B и Смеси C, даже несмотря на то, что для них показали увеличенное повреждение при увеличении дозы гербицидов. При воздействии Смеси D на эти линии все растения были уничтожены. Примечательно, что для RW-B показана толерантность ко всем из тестированных смесей, даже к смеси, содержащей наивысшую концентрацию активных ингредиентов.
Заключение: Для RW-B показали не только более высокий уровень толерантности к каждому из ингибирующих AHAS гербицидов, тестированным по отдельности, но также наивысший уровень толерантности к комбинации гербицидов, тестированных одновременно в смеси.
несущих различные случаи мутации в локусе AHASL1 при испытании 4
смесями из двух ингибирующих AHAS гербицидов
(г a.i./га)
(г a.i./га)
2 p: вероятность
3 MSD: Минимальное значимое различие
Ответ линий подсолнечника, гомозиготных по аллелю W574L, по сравнению с линиями, гомозиготными по любому из аллелей A205V и P197L, и гаплотип чувствительности к имазапиру на уровне целого растения
Цель: Этот эксперимент проводили для количественной оценки и сопоставления чувствительности к имазапиру линий подсолнечника, несущих гомозиготный аллель W574L, по сравнению с линиями, гомозиготными по аллелям P197L и A205V, на уровне целого растения.
Материалы:
В таблице 9 показаны семена различных линий подсолнечника, полученные в полевых условиях
Семена высевали в чашки Петри и после прорастания сеянцы пересаживали в горшки диаметром 10 см в горшечную смесь, состоящую из равных частей вермикулита, почвы и песка. Растения выращивали в теплице в условиях естественного света, дополненного лампами по 400 В с галогенидом натрия для обеспечения длительности дня 16 ч. Дневная/ночная температуры составляли 25 и 20°C, соответственно. На стадии V2-V4 (Schneiter & Miller, 1981) по 10 растений каждого генотипа случайным образом регистрировали для каждой обработки, состоящей из восьми доз имазапира (0, 40, 80, 160, 320, 480, 640 и 800 г a.i./га, без обработки, 0,5Ч, 1Ч, 2Ч, 4Ч, 6Ч, 8Ч и 10Ч, соответственно), и определения биомассы в нулевой момент времени. Эксперимент организовывали в дизайне рандомизированного блока с полнофакторной (линия подсолнечника Ч обработка) организацией обработок и 10 повторами.
В день обработки гербицидами по десять растений каждого генотипа срезали по семядольному узлу и высушивали при 60°C в течение 48 ч для определения массы в сухом состоянии в нулевой момент времени.
Оставшиеся растения поддерживали 14 суток после обработки имазапиром (DAT), и определяли их высоту, показатель фитотоксичности (PI) и надземную сухую биомассу. Высоту определяли как расстояние между семядольным узлом и апексом каждого растения. Данные о надземной сухой биомассе для каждой линии переводили в накопление биомассы после воздействия посредством вычитания соответствующей средней биомассы в нулевой момент времени для каждого образца. Данные о сухой биомассе переводили в проценты от необработанных контрольных растений внутри каждой линии, чтобы позволить прямое сравнение между группами. PI представляет собой фенотипическую шкалу от 0 до 9, по которой оценивали каждое растение посредством визуального осмотра. Растения без каких-либо симптомов регистрировали как «0», увеличенные уровни низкорослости и хлороза по отношению к необработанным контрольным растениям регистрировали как «1»-«4», увеличенные уровни аномалий листа и некроза листа регистрировали от «5» до «8», и погибшие растения с полным некрозом апекса регистрировали как «9».
Результаты:
Высота. После воздействия имидазолинона высота растений являлась сильно уменьшенной в чувствительной (B770) и толерантной к SU (SuBL) линиях. Фактически, даже при пониженной дозе распыления гербицида (0,5Ч), высота являлась уменьшенной от 100% для необработанных контрольных растений до 35,0% и 42,7% в чувствительной и толерантной к SU линиях, соответственно. Для толерантной к IMI линии (B770IMI) показали значимое уменьшение высоты только после 2Ч дозы воздействия гербицида, и затем при увеличении дозы гербицида увеличение снижения высоты было более сильным, приближаясь к такому же уровню, как для чувствительной линии при 10Ч (32,8%).
В отличие от этого, для линии подсолнечника RW-B показали меньшее снижение высоты при испытании увеличивающихся доз имидазолинонов. Это снижение высоты достигало только 75% от необработанных контрольных растений для воздействия гербицидов в дозах от 4Ч до 10Ч (таблица 10 и фиг.9).

Показатель фитотоксичности
Для четырех тестированных мутантов показали большие различия в симптомах после воздействия увеличивающихся доз имазапира. Для линии RW-B подсолнечника показали только слабое уменьшение размера листьев и более светлую зеленую окраску по сравнению с контрольными растениями при увеличении доз гербицида (Таблица 11). В отличие от этого, даже несмотря на то, что для растений, несущих аллель A205V (B770IMI), не показали какого-либо повреждения при дозе 0,5Ч, у них присутствовал увеличенный уровень повреждений (хлороз, деформация листьев и некроз листьев) при дозах имазапира, увеличивающихся от 1Ч до 10Ч. Для Su-толерантных и чувствительных линий показали почти идентичную реакцию: полную чувствительность при воздействии гербицидов в дозах 0,5Ч-10Ч.

Масса надземной биомассы в сухом состоянии
Кривые зависимости от дозы для массы в сухом состоянии мутантов W574L, P197L и 205V показаны на фиг.11. Масса надземной биомассы в сухом состоянии в случае W574L в гомозиготном состоянии являлась уменьшенной по отношению к необработанным контрольным растениям только при воздействии гербицидов в дозах 6Ч, 8Ч и 10Ч. Масса в сухом состоянии растений RW-B при дозе 10Ч составляла 78% от необработанного контроля. При этом масса в сухом состоянии для всех других мутантов и линии дикого типа значимо отличалась от массы для линии RW-B в каждой из тестированных доз (таблица 12).

Заключение: В линии RW-B, несущей случай мутации W574L в гомозиготном состоянии в локусе AHASL1 подсолнечника, присутствовал более высокий уровень толерантности ко всем тестированным дозам имазапира, чем в Su-толерантной линии и в уже известных толерантных к IMI линиях.
Ответ мутантного аллеля W574L по сравнению с мутациями A205V и P197L и чувствительный к метсульфурону гаплотип на уровне целого растения.
Цель: Этот эксперимент проводили для количественной оценки и сопоставления чувствительности к метсульфурону на уровне целого растения, для линий подсолнечника, несущих аллель W574L в гомозиготном состоянии, с гомозиготными линиями, несущими аллели P197L и A205V.
Материалы:
В таблице 13 показаны семена различных линий подсолнечника, полученные в полевых условиях
Семена высевали в чашки Петри и после прорастания сеянцы пересаживали в горшки диаметром 10 см в горшечную смесь, состоящую из равных частей вермикулита, почвы и песка. Растения выращивали в теплице в условиях естественного света, дополненного лампами по 400 В с галогенидом натрия для обеспечения длительности дня 16 ч. Дневная/ночная температуры составляли 25 и 20°C, соответственно. На стадии V2-V4 (Schneiter & Miller, 1981) по 10 растений каждого генотипа случайным образом регистрировали для каждой обработки, состоящей из восьми доз имазапира (0, 2,5, 5, 10, 15, 20 г a.i./га, без обработки, 0,5Ч, 1Ч, 2Ч, 3Ч и 4Ч, соответственно), и определения биомассы в нулевой момент времени. Эксперимент организовывали в дизайне рандомизированного блока с полнофакторной (линия подсолнечника x обработка) организацией обработок и 10 повторами.
В день обработки гербицидами по десять растений каждого генотипа срезали по семядольному узлу и высушивали при 60°C в течение 48 ч для определения массы в сухом состоянии в нулевой момент времени.
Оставшиеся растения поддерживали 14 суток после обработки имазапиром (DAT) и определяли их показатель фитотоксичности (PI) и надземную сухую биомассу. Данные о надземной сухой биомассе для каждой линии переводили в накопление биомассы после воздействия посредством вычитания соответствующей средней биомассы в нулевой момент времени для каждого образца. Данные о сухой биомассе переводили в проценты от необработанных контрольных растений внутри каждой линии, чтобы позволить прямое сравнение между группами. PI представляет собой фенотипическую шкалу от 0 до 9, по которой оценивали каждое растение посредством визуального осмотра. Растения без каких-либо симптомов регистрировали как «0», увеличенные уровни низкорослости и хлороза по отношению к необработанным контрольным растениям регистрировали как «1»-«4», увеличенные уровни аномалий листа и некроза листа регистрировали от «5» до «8», и погибшие растения с полным некрозом апекса регистрировали как «9».
Результаты
Показатель фитотоксичности: Для мутантов показали большие различия в ответе на воздействие увеличивающихся доз метсульфурона (таблица 13 и фиг.12). Общепринятый генотип (B770) был уничтожен даже при более низких дозах метсульфурона. Для линии подсолнечника, несущей мутацию W574L в гомозиготном состоянии, показали только слабое уменьшение размера растений и более светлую зеленую окраску по сравнению с контрольными растениями при увеличении доз гербицида от 0,5 до 3Ч (таблица 13). Фактически, для них показали только хлороз и деформации при воздействии 4Ч дозы метсульфурона. В отличие от этого, для линии, несущей аллель A205V (B770IMI), показали увеличенный уровень повреждений при дозе от 0,5Ч до 4Ч. Фактически, при воздействии метсульфурона в дозе 2Ч, почти все растения этой линии были уничтожены гербицидом.
Для SU-толерантной линии (SuBL), несущей уже известный мутантный аллель для толерантности подсолнечника к сульфонилмочевине, не показали симптомов или показали слабый хлороз при воздействии метсульфурона в дозах 0,5Ч и 1Ч, но потом уровень их повреждения увеличивался, достигая балла 9 (все растения были уничтожены гербицидом) при 4Ч дозе метсульфурона.
AHASL1 подсолнечника, при испытании увеличивающимися дозами метсульфурона
2 p: вероятность
3 MSD: Минимальное значимое различие
Результаты:
Масса надземной биомассы в сухом состоянии: Кривые зависимости от дозы для массы в сухом состоянии мутантов W574L, P197L и A205V показаны на фиг.5 и в таблице 14. Для толерантных и чувствительных к IMI линий показали немедленное уменьшение массы в сухом состоянии после воздействия гербицидов. Биомасса SU-толерантной линии уменьшалась от 83,3% до 30,3% от необработанных контрольных растений при увеличении доз метсульфурона от 0,5 до 4Ч.
Интересно, что для линии RW-B не показали значимого уменьшения массы в сухом состоянии при воздействии метсульфурона в дозе 0,5 и 1Ч. При увеличении доз наблюдали значимое уменьшение массы в сухом состоянии. Однако для этой линии показали значимо более высокую массу в сухом состоянии, чем для любой из других линий в каждой из тестированных доз.
для 4 линий подсолнечника, несущих различные мутантные аллели локуса AHASL1 подсолнечника, при
испытании увеличивающимися дозами метсульфурона
2 p: вероятность
3 MSD: Минимальное значимое различие
Заключение: В линии RW-B, несущей случай мутации W574L в гомозиготном состоянии в локусе AHASL1 подсолнечника, присутствовал более высокий уровень толерантности ко всем тестированным дозам метсульфурона, чем в IMI-толерантной линии и в уже известных SU-толерантных линиях.Список литературы
Al-Khatib K., J.R. Baumgartner, D.E. Peterson, and R.S. Currie. 1998. Imazethapyr resistance in common sunflower (Helianthus annuus). Weed Sci. 46:403-407.
Al-Khatib K., and J.F. Miller. 2002. Registration of four genetic stocks of sunflower resistant to imidazolinone herbicides. Crop Sci. 40:869-870.
Anderson, P. C. and M. Georgeson. 1989. Herbicide-tolerant mutants of corn. Genome 34:994-999.
Ashburner, M. 1990. Drosophila, A Laboratory Handbook. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Bently, A., B. Mac Lennan, J. Calvo, and C.R. Dearolf. 2000. Targeted recovery of mutations in Drosophila. Genetics 156:1169-1173.
Bernasconi, P., A.R. Woodworth, B.A. Rosen, M.V. Subramanian, and D.L. Siehl. 1995. A naturally occurring point mutation confers broad range tolerance to herbicides that target acetolactate synthase. J. Biol. Chem. 270:17381-17385.
Bradford, M.M. 1976. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254.
Bruniard J.M. and J.F. Miller. 2001. Inheritance of imidazolinone herbicide resistance in sunflower. Helia 24:11-16.
Chong, C.K. and J.D.Choi. 2000. Amino acid residues conferring herbicide tolerance in tobacco acetolactate synthase. Biochem. Biophys. Res. Commun. 279:462-467.
Croughan, T. P. 1996. Herbicide resistant rice. U.S. patent 5,545,822.
Devine, M. D., M.A.S. Marles, and L. M. Hall. 1991. Inhibition of acetolactate synthase in susceptible and resistant biotypes of Stellaria media. Pestic. Sci. 31:273-280.
D'Halluin, K. M., M. Bossut, E. Bonne, B. Mazur, J. Leemans, and J. Botterman. 1992. Transformation of sugarbeet (Beta vulgaris L.) and evaluation of herbicide resistance in transgenic plants. Bio/Technology 10:309-314.
Duggleby, R. G. and S. S. Pang. 2000. Acetohydroxyacid synthase. J. Biochem. Mol. Biol. 33:1-36.
Grula J.W., R.L. Hudspeth, S.L. Hobbs and D.M. Anderson. 1995. Organization, inheritance and expression of acetohydroxyacid synthase genes in the cotton allotetraploid Gossypium hirsutum. Plant Mol. Biol. 28:837-846.
Guttieri, M.J., C.V. Eberlein, C.A. Mallory-Smith, D.C. Thill, and D.L. Hoffman. 1992. DNA sequence variation in Domain A of the acetolactate synthase genes of herbicide-resistant and -susceptible weed biotypes. Weed Sci. 40:670-676.
Guttieri, M.J., C.V. Eberlein, and D.C. Thill. 1995. Diverse mutations in the acetolactate synthase allele confer chlorsulfuron resistance in Kochia scoparia biotypes. Weed Sci. 43:175-178.
Harms, C.T., S.L. Armour, J.J. DiMaio, L.A. Middlesteadt, D. Murray, D.V. Negrotto, H. Thompson-Tyler, K. Weymann, A.L. Montoya, R.D. Shillito, and G.C. Jen. 1992. Herbicide resistance due to amplification of a mutant acetohydroxyacid synthase allele. Mol. Gen. Genet. 233:427-435.
Hart, S.E., J.W. Saunders, and D. Penner. 1992. Chlorsulfuron resistant sugar beet: Cross-resistance and physiological basis of resistance. Weed Sci. 40:378-383.
Haughn, G.W., J. Smith, B. Mazur, and C. Somerville. 1988. Transformation with a mutant Arabidopsis acetolactate synthase gene renders tobacco resistant to sulfonylureas. Mol. Gen. Genet. 211:266-271.
Inukai, T., A. Sako, H.Y. Hirano, and Y. Sano. 2000. Analysis of intragenic recombination at wx in rice: Correlation between molecular and genetic maps within the locus. Genome 43:589-596.
Jander, G., S.R. Baerson, J.A. Hudak, K.A. Gonzalez, K.J. Gruys, and R.L. Last. 2003. Ethylmethanesulfonate saturation mutagenesis in Arabidopsis to determine frequency of herbicide resistance. Plant Physiol. 131:139-146.
Kolkman, J.M., M.B. Slabaugh, J.M. Bruniard, S. Berry, B.S. Bushman, C. Olungu, N. Maes, G. Abratti, A. Zambelli, J.F. Miller, A. Leon, and S.J. Knapp. 2004. Acetohydroxyacid synthase mutations conferring resistance to imidazolinone or sulfonylurea herbicides in sunflower. Theor. Appl. Genet. 109:1147-1159.
Kozik A., R.W. Michelmore, S.J. Knapp, M.S. Matvienko, L. Rieseberg, H. Lin, M. van Damme, D. Lavelle, P. Chevalier, J. Ziegle, P. Ellison, J.M. Kolkman, M.B. Slabaugh, K. Livingston, L.Z. Zhou, S. Church, S. Edberg, L. Jackson, R. Kesseli and K. Bradford. 2002. Sunflower and lettuce ESTs from the compositae genome project. http://compgenomics.ucdavis.edu
Lee, Y.T. and R.G. Duggleby. 2001. Identification of the regulatory subunit of Arabidopsis thaliana acetohydroxyacid synthase and reconstitution with its catalytic subunit. Biochemistry 40:6836-6844.
Lee, Y.T. and R.G. Duggleby. 2002. Regulatory interactions in Arabidopsis thaliana acetohydroxyacid synthase. FEBS Lett. 512:180-184.
Lee, K.Y., J. Townsend, J. Tepperman, M. Black, C.F. Chui, B. Mazur, P. Dunsmuir, and J. Bedbrook. 1988. The molecular basis of sulfonylurea resistance in tobacco. EMBO J. 7:1241-1248.
Mallory-Smith C.A., D.C. Thill and M.J. Dial. 1990. Identification of sulfonylurea herbicide-resistant prickly lettuce (Lactuca serriola). Weed Technol. 4:163-168.
Marshall M.W., K. Al-Khatib, and T. Loughin. 2001. Gene flow, growth, and competitiveness of imazethapyr-resistant common sunflower. Weed Sci. 49:14-21.
Massinga RA, K. Al-Khatib, P. St. Amand and J.F. Miller. 2003. Gene flow from imidazolinone-resistant domesticated sunflower to wild relatives. Weed Sci. 51:854-862.
McCourt, J.A. and R.G. Duggleby. 2006. Acetohydroxyacid synthase and its role en the biosynthetic pathway for branched-chain amino acids. Amino Acids 31(2):173.
McCourt, J.A., S.S. Pang, J. King-Scott, L.W. Guddat and R.G. Duggleby. 2006. Herbicide-binding sites revealed in the structure of plant acetohydroxyacid synthase. PNAS 103(3): 569-573.
McHughen, A. 1989. Agrobacterium mediated transfer of chlorsulfuron resistance to commercial flax cultivars. Plant Cell Rep. 8:445-449.
McNaughton, K.E., J. Letarte, E.A. Lee and F.J. Tardif. 2005. Mutations in ALS confer herbicide resistance in redroot pigweed (Amaranthus retroflexus) and Powell amaranth (Amaranthus powellii). Weed Sci. 53:17-22.
Miller J.F., and K. Al-Khatib. 2002. Registration of imidazolinone herbicide-resistant sunflower maintainer (HA425) and fertility restorer (RHA426 and RHA427) germplasms. Crop Sci. 42:988-989.
Miller J.F., and K. Al-Khatib. 2004. Registration of two oilseed sunflower genetic stocks, SURES-1 and SURES-2, resistant to tribenuron herbicide. Crop Sci. 44:1037-1038.
Milliman, L.D., D.E. Riechers, L.M. Wax and F.W. Simmons. 2003. Characterization of two biotypes of imidazolinone-resistant eastern black nightshade (Solanum ptycanthum). Weed Sci. 51:139-144.
Mourad, G., and J. King. 1992. Effect of four classes of herbicides on growth and acetolactate synthase activity in several variants of Arabidopsis thaliana. Planta 188:491-497.
Newhouse, K., B.K. Singh, D.L. Shaner, and M. Stidham. 1991. Mutations in corn (Zea mays L.) conferring resistance to imidazolinones. Theor. Appl. Genet. 83:65-70.
Newhouse, K., W. Smith, M. Starrett, T. Schaefer, and B.K. Singh. 1992. Tolerance to imidazolinone herbicides in wheat. Plant Physiol. 100:882-886.
Ouellet T., R.G. Rutledge and B.L., Miki. 1992. Members of the acetohydroxyacid synthase multigene family of Brassica napus have divergent patterns of expression. Plant J. 2:321-330.
Powles S.B. and J.A.M. Holtum. 1994. Herbicide resistance in plants: biology and biochemistry. Lewis Publishers. Boca Ratуn, Fla.
Pozniak C.J., and P.J. Hucl. 2004. Genetic analysis of imidazolinone resistance in mutation-derived lines of common wheat. Crop Sci. 44:23-30.
Preston, C. and C.A. Mallory-Smith. 2001. Biochemical mechanisms, inheritance, and molecular genetics of herbicide resistance in weeds. In: S.B. Powles and D.L. Shaner (eds). Herbicide Resistance and World Grains, CRC Press, Boca Raton, FL, pp 23-60.
Rajasekaran, K., J.W. Grula, and D.M. Anderson. 1996. Selection and characterization of mutant cotton (Gossypium hirsutum L.) cell lines resistant to sulfonylurea and imidazolinone herbicides. Plant Sci. 199:115-124.
Ray, T.B. 1984. Site of action of chlorsulfuron. Inhibition of valine and isoleucine biosynthesis in plants. Plant Physiol. 75:827-831.
Saari, L. L., J. C. Cotterman, and D. C. Thill. 1994. Resistance to acetolactate synthase inhibiting herbicides. Pages 83-139 in S. B. Powles and J.A.M. Holtum, eds. Herbicide Resistance in Plants: Biology and Biochemistry. Ann Arbor, MI: Lewis.
Sala, C.A, Bulos, M. & Echarte, A.M. 2008a. Genetic analysis of an induced mutation conferring imidazolinone resistance in sunflower. Crop Science 48:1817-1822.
Sala, C.A., M. Bulos, M. Echarte, S.R. Whitt, R. Ascenzi. 2008b. Molecular and biochemical characterization of an induced mutation conferring imidazolinone resistance in sunflower. Theoretical and Applied Genetics 108 (1): 115-112.
Santel, H.J., B.A. Bowden, V.M. Sorenson, K.H. Mueller, and J. Reynolds. 1999. Flucarbazone-sodium-a new herbicide for grass control in wheat. Proc. West. Soc. Weed Sci. 52:124-125.
Schneiter, A.A., and J.F. Miller. 1981. Description of sunflower growth stages. Crop Sci. 21:901-903.
Sebastian, S.A., G.M. Fader, J.F. Ulrich, D.R. Forney, and R.S. Chaleff. 1989. Semi-dominant soybean mutation for resistance to sulfonylurea herbicides. Crop Sci. 29:1403-1408.
Seefeldt, S.S., J.E. Jensen and E.P. Fuerst. 1995. Log-logistic analysis of herbicide dose response relationships. Weed Technol. 9:218-227.
Shaner D.L. 1991. Physiological effects of the imidazolinone herbicides. In: Shaner DL, O'Connor SL (eds) The imidazolinone herbicides. Lewis, Ann Arbor, Mich., pp 129-138.
Shaner, D.L., P.C. Anderson, and M.A. Stidham. 1984. Imidazolinones: Potent inhibitors of acetohydroxyacid synthase. Plant Physiol. 76:545-546.
Singh, B.K. 1999. Biosynthesis of valine, leucine and isoleucine. P. 227-247. In B.K. Singh (ed.) Plant amino acids. Marcel Dekker Inc., New York.
Singh, B.K., M.A. Stidham, and D.L. Shaner. 1988. Assay of acetohydroxyacid synthase. Ann. Biochem. 171:173-179.
Snow, E.T., R.S. Foote, and S. Mitra. 1984. Base-pairing properties of O-6-Methylguanine in template DNA during in vitro DNA replication. J. Biol. Chem. 259:8095-8100.
Subramanian, M.V., and B.C. Gerwick. 1989. Inhibition of acetolactate synthase by triazolopyrimidines: a review of recent developments. P. 277-288. In J.R. Whitaker, P.E. Sonnet, eds, Biocatalysis in Agricultural Biotechnology. American Chemical Society, Washington, DC.
Subramanian, M.V., H.Y. Hung, J.M. Dias, V.W. Miner, J.H. Butler, and J.J. Jachetta. 1990. Properties of mutant acetolactate synthases resistant to triazolopyrimidine sulfonanilide. Plant Physiol. 94:239-244.
Swanson, E.B., M.J. Hergesell, M. Arnoldo, D.W. Sippell, and R.S.C. Wong. 1989. Microspore mutagenesis and selection: Canola plants with field tolerance to imidazolinones. Theor. Appl. Genet. 78:525-530.
Tan, S., R.R. Evans, M.L. Dahmer, B.K. Singh, and D.L. Shaner. 2005. Imidazolinone-tolerant crops: history, current status and future. Pest Manag. Sci. 61:246-257.
Tranel, P.J. and T.R. Wright. 2002. Resistance of weeds to AHAS inhibiting herbicides: what have we learned? Weed Sci 50:700-712.
Trucco, F., A.G. Hager and P.J. Tranel. 2006. Acetolactate synthase mutation conferring imidazolinone-specific herbicide resistance in Amaranthus hybridus. J. Plant Physiol. 163:475-479.
Umbarger, H.E. 1978. Amino acid biosynthesis and its regulation. Annu. Rev. Biochem. 47:533-606.
White A.D., M.D. Owen, R.G. Hartzler, and J. Cardina. 2002. Common sunflower resistance to acetolactate-inhibiting herbicides. Weed Sci. 50:432-437.
Wright, T.R., N.F. Bascomb, S.F. Sturner, and D. Penner. 1998. Biochemical mechanism and molecular basis for ALS-inhibiting herbicide resistance in sugar beet (Beta vulgaris) somatic cell selections. Weed Sci. 46:13-23.
Wright, T.R., and D. Penner. 1998. Cell selection and inheritance of imidazolinone resistance in sugar beet (Beta vulgaris). Theor. Appl. Genet. 96:612-620.









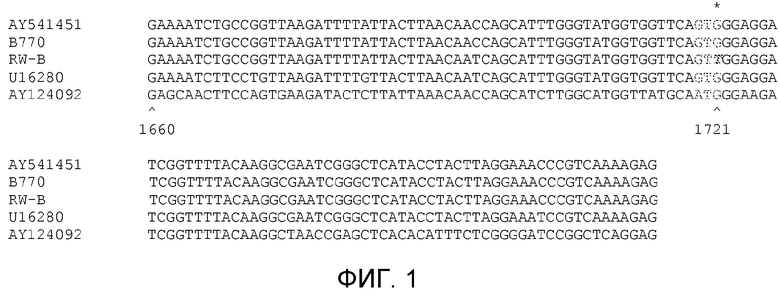
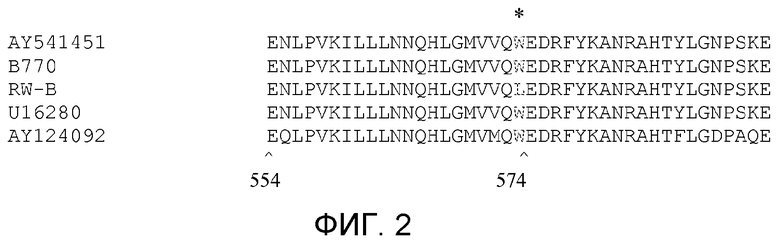
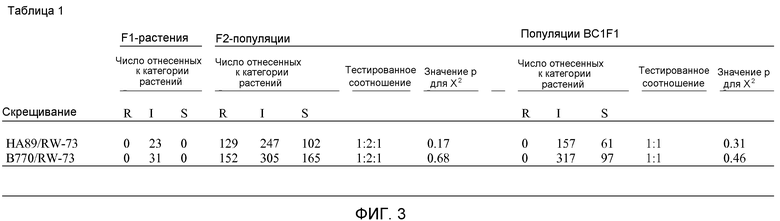
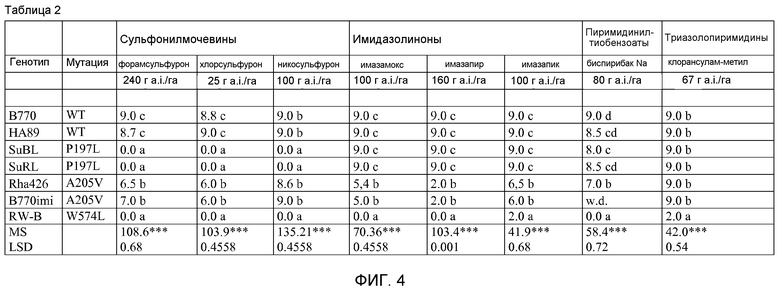
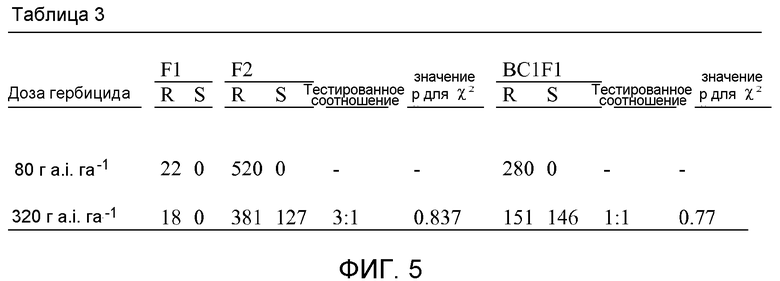



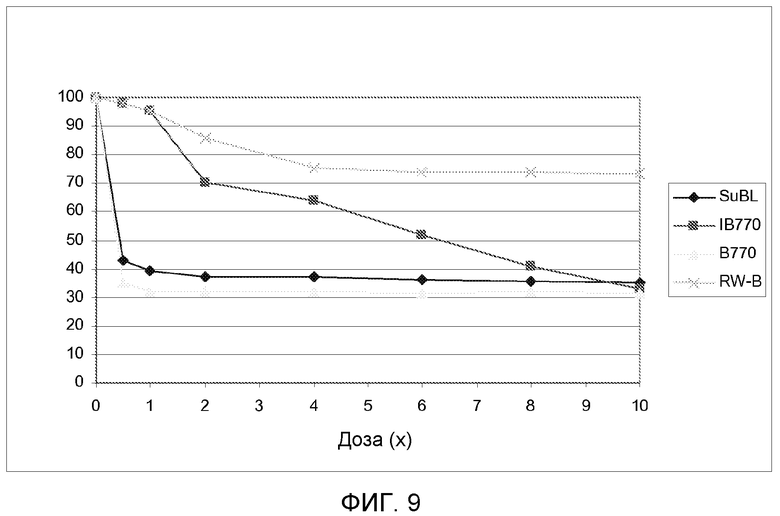
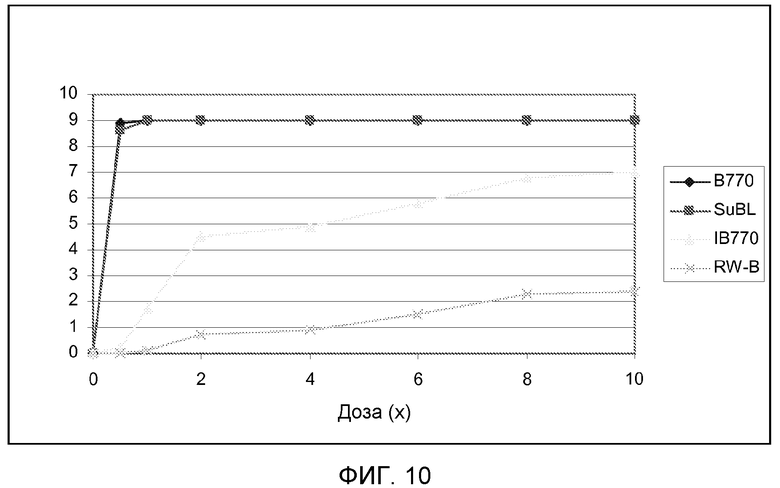
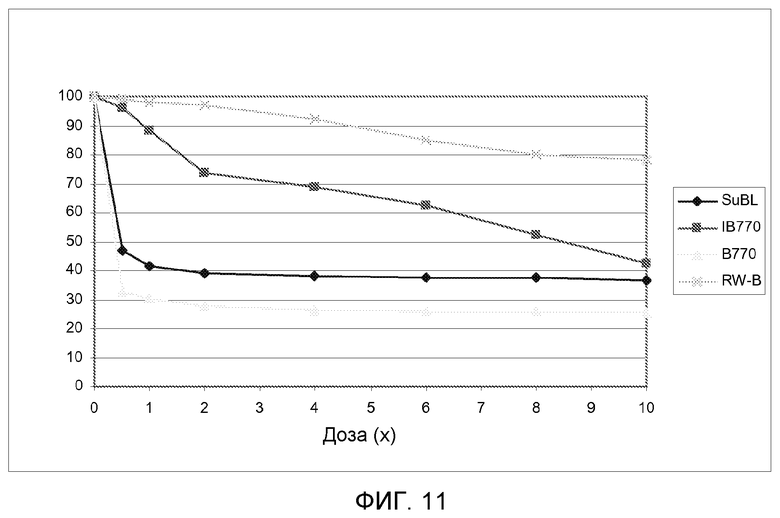
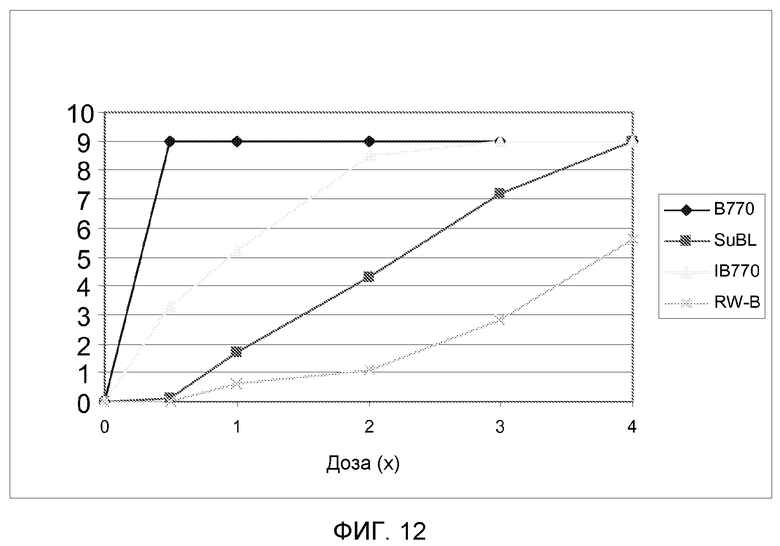
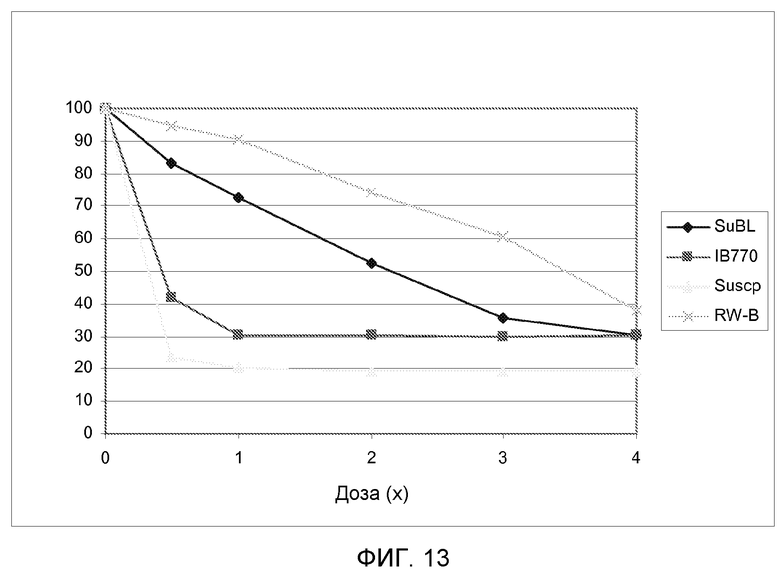
Изобретение относится к области биохимии, в частности к способу борьбы с сорняками поблизости от растения подсолнечника. При этом способ включает нанесение на сорняки и растение подсолнечника эффективного количества любого из гербицидов, выбранных из группы, состоящей из гербицида имидазолинона, гербицида сульфонилмочевины, гербицида триазолопиримидина, гербицида пиримидинилоксибензоата и их смесей. Указанное растение подсолнечника содержит в своем геноме по крайней мере одну копию по крайней мере гена синтазы ацетогидроксикислоты (AHAS), кодирующего устойчивый к гербициду белок, имеющий аминокислотную последовательность, представленную SEQ ID NO:2. Изобретение позволяет эффективно бороться с сорняками поблизости от растения подсолнечника. 8 з.п. ф-лы, 13 ил., 14 табл.
1. Способ борьбы с сорняками поблизости от растения подсолнечника, включающий
нанесение на сорняки и растение подсолнечника эффективного количества любого из гербицидов, выбранных из группы, состоящей из гербицида имидазолинона, гербицида сульфонилмочевины, гербицида триазолопиримидина, гербицида пиримидинилоксибензоата и их смесей,
где растение подсолнечника содержит в своем геноме по крайней мере одну копию по крайней мере гена синтазы ацетогидроксикислоты (AHAS), кодирующего устойчивый к гербициду белок, имеющий аминокислотную последовательность, представленную SEQ ID NO:2.
2. Способ по п.1, где растение подсолнечника имеет повышенную устойчивость к гербициду имидазолинона, гербициду сульфонилмочевины, гербициду триазолопиримидина, гербициду пиримидинилоксибензоата и их смесям по сравнению с растением подсолнечника, которое не содержит в своем геноме по крайней мере одну копию по крайней мере гена синтазы ацетогидроксикислоты (AHAS), кодирующего устойчивый к гербициду белок, имеющий аминокислотную последовательность, представленную SEQ ID NO:2.
3. Способ по п.1, где ген AHAS имеет нуклеотидную последовательность, представленную SEQ ID NO:1.
4. Способ по п.1, где растение подсолнечника является:
(а) растением подсолнечника линии RW-B, где репрезентативное семя депонировано в АТСС под инвентарным № РТА-9176;
(b) потомством линии RW-B;
(с) мутантом, рекомбинантом, генетически модифицированным производным линии RW-B, или
(d) растением, являющимся потомком по крайней мере одного из растений по подпунктам (а)-(с).
5. Способ по п.1, где растение подсолнечника обладает одиночной головкой.
6. Способ по п.1, где растение подсолнечника содержит в своем геноме по крайней мере одну копию выделенной нуклеиновой кислоты, имеющей последовательность SEQ ID NO:1.
7. Способ по п.1, где растение подсолнечника содержит в своем геноме по крайней мере одну копию экспрессирующей кассеты, содержащей промотор, активный в растениях подсолнечника, функционально связанный с выделенной нуклеиновой кислотой, кодирующей белок, имеющий последовательность, кодируемую SEQ ID NO:1.
8. Способ по п.1, где растение подсолнечника выращено из семени растения подсолнечника, содержащего в своем геноме по крайней мере одну копию гена AHAS, кодирующего устойчивый к гербициду белок, имеющий аминокислотную последовательность, представленную SEQ ID NO:2.
9. Способ по п.8, где ген AHAS имеет нуклеотидную последовательность, представленную SEQ ID NO:1.
| WO2007054555 A2, 18.05.2007 | |||
| Реактивный двигатель внутреннего горения | 1926 |
|
SU7322A1 |
| WO2006024694 A2, 09.03.2006 | |||
| TAN S | |||
| et al., Imidazolinone-tolerant crops: history, current status and future, Pest Manag Sci, 2005, vol | |||
| Устройство для сортировки каменного угля | 1921 |
|
SU61A1 |
| Котел | 1921 |
|
SU246A1 |
| Устройство для измерения напряженности магнитного поля | 1985 |
|
SU1347056A1 |
| EP0492113 А2, 01.07.1992 | |||
| СПОСОБ ПОЛУЧЕНИЯ МУТАНТНОЙ ZEA MAYS | 1990 |
|
RU2128426C1 |

