ИСПРАШИВАНИЕ ПРИОРИТЕТА
По этой заявке испрашивается приоритет по дате подачи временной заявки на патент США с серийным номером 61/015591, поданной 20 декабря 2007 г.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к новым и отличающимся растениям подсолнечника, дающим семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты, а также к соответствующим способам. Настоящее изобретение дополнительно относится к не модифицированным генетически, немутированным растениям подсолнечника, имеющим устойчивость к глифосату, и соответствующим способам.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Культивируемый подсолнечник (Helianthus annuus L.) является основным мировым источником растительного масла. В Соединенных Штатах Америки приблизительно 4 миллиона акров ежегодно засеваются подсолнечником, преимущественно в Дакоте и Миннесоте.
Очень быстрое увеличение количества засеваемых подсолнечником акров за последнее десятилетие в Соединенных Штатах Америки частично является следствием нескольких важных разработок в области селекции подсолнечника и улучшения сортов. Одной из важных разработок было открытие цитоплазматической мужской стерильности и генов для восстановления фертильности, которое позволило получение гибридных растений подсолнечника. Полученные таким образом гибриды были введены в культуру в начале 1970-х гг.
Описание цитоплазматической мужской стерильности (ЦМС) и генетическое восстановление фертильности у подсолнечника представлено в Fick, «Breeding and Genetics» в Sunflower Science and Technology 279-338 (J.F. Carter ed. 1978).
Масло подсолнечника состоит в основном из пальмитиновой (16:0), стеариновой (18:0), олеиновой (18:1), линолевой (18:2) и линоленовой (18:3) кислот. Хотя в растениях присутствуют другие необычные жирные кислоты, пальмитиновая, стеариновая, олеиновая, линолевая и линоленовая кислоты составляют примерно 88% жирных кислот, присутствующих в мировом производстве растительных масел (J.L. Harwood, Plant Acyl Lipids: Structure, Distribution and Analysis, 4 Lipids: Structure and Function, P.K. Stumpf and E.E. Conn ed. (1988)). Пальмитиновая и стеариновая кислоты являются насыщенными жирными кислотами, которые, как было показано в некоторых исследованиях, вносят вклад в увеличение уровня плазматического холестерина, причины коронарной болезни сердца. Согласно последним исследованиям растительные масла с высоким содержанием ненасыщенных жирных кислот, таких как олеиновая и линолевая кислоты, может обладать способностью снижать уровень плазматического холестерина. Насыщенные жирные кислоты также в общем имеют более высокие точки плавления относительно ненасыщенных жирных кислот с таким же числом атомов углерода, что вносит вклад в проблему устойчивости к холоду продуктов питания и может вносить вклад в ощущение воска или жира во рту в ходе приема пищи. Также известно, что пищевые продукты, изготовленные из жиров и масел, имеющих меньше примерно 3% насыщенных жиров будут обычно содержать меньше 0,5 г насыщенных жиров на порцию, и в результате при существующих нормативах маркировки могут быть маркированы, как «не содержащие насыщенных жиров». Следовательно, по ряду причин желательно получать масло подсолнечника с низким содержанием пальмитиновой и стеариновой кислот и высоким содержанием олеиновой или линолевой кислот.
Выведение любой новой желательной растительной зародышевой плазмы состоит из ряда стадий. Селекция растений начинается с анализа и определения проблем и недостатков имеющейся в наличии зародышевой плазмы, установления целей программы и определения конкретных задач скрещивания. Следующей стадией является отбор зародышевой плазмы, обладающей признаками, удовлетворяющими целям программы. Целью является объединение в одном сорте улучшенной комбинации желательных признаков из родительской зародышевой плазмы. Эти важные признаки могут включать высокий выход семян, резистентность к болезням и насекомым, более крепкие стебли и корни, переносимость засухи и жары, и более хорошее агротехническое качество.
Выбор способов селекции и отбора зависит от способа репродукции растения, способности наследовать улучшаемый признак (признаки) и типа используемого в коммерческих целях культивируемого сорта (например, F1-гибрид, чистая линия и т.д.). Для высоконаследуемых признаков будет эффективен выбор превосходных индивидуальных растений, оцениваемых в одном месте, в то время как для признаков с низкой наследуемостью отбор должен быть основан на средних величинах, полученных из повторных оценок семейств родственных растений. Популярные способы селекции обычно включают выведение чистых линий, модифицированное выведение чистых линий, массовый отбор и рекуррентный отбор.
Сложный характер наследования влияет на выбор способа селекции. Возвратное скрещивание используют для переноса одного или нескольких благоприятных генов для высоконаследуемого признака в желаемый культивируемый сорт. Этот подход широко используется для селекции устойчивых к болезням культивируемых сортов. Для улучшения количественно наследуемых признаков, контролируемых рядом генов, используют различные методики рекуррентного отбора. Применение рекуррентного отбора у самоопыляющихся сельскохозяйственных культур зависит от легкости опыления, частоты возникновения успешных гибридов от каждого опыления и числа гибридных потомков от каждого успешного скрещивания.
Каждая программа селекции должна включать периодическую объективную оценку эффективности процедуры селекции. Критерии оценки варьируют в зависимости от цели и задач, но должны включать отдачу от отбора за год исходя из сравнения с соответствующим стандартом, общую ценность усовершенствованных выводимых линий и ряд успешных культивируемых сортов, полученных на единицу на входе (например, в год, на единицу затраченных денежных средств и т.д.).
Перспективные усовершенствованные выводимые линии тщательно тестируют и сравнивают с соответствующими стандартами в условиях окружающей среды, соответствующих коммерческой целевой площади (площадям) в течение трех или более лет. Лучшие линии являются кандидатами для новых коммерческих культивируемых сортов; те, у которых отсутствуют небольшое количество признаков, можно использовать в качестве родительских растений для получения новых популяций для дополнительного отбора.
Эти процессы, которые приводят к конечной стадии продажи и распространения, обычно занимают от восьми до двенадцати лет от начала проведения первого скрещивания. Поэтому выведение новых культивируемых сортов представляет собой длительный процесс, требующий точного долгосрочного планирования, эффективного использования ресурсов и минимального изменения направления.
Наиболее трудной задачей является идентификация индивидуальных растений, которые обладают превосходством на генетическом уровне, поскольку для большинства признаков действительно ценный генотип замаскирован другими соединенными признаками растения или факторами окружающей среды. Одним способом идентификации превосходящего растения является наблюдение за его характеристиками относительно других экспериментальных растений и относительно широко распространенного стандартного культивируемого сорта. Если невозможно сделать выводы по одному наблюдению, то повторные наблюдения обеспечивают лучшую оценку его генетической ценности.
Целью селекции растений является создание новых, уникальных и улучшенных культивируемых сортов и гибридов подсолнечника. Селекционер первоначально отбирает и скрещивает две или несколько родительских линий, с последующими повторными самоопылением и отбором, получая множество новых генетических комбинаций. Теоретически, селекционер может генерировать миллиарды различных генетических комбинаций с помощью скрещивания, самоопыления и мутаций. Селекционер не имеет прямого контроля на клеточном уровне. Поэтому два селекционера никогда не выведут одинаковую линию, или даже очень похожие линии, имеющие одинаковые признаки подсолнечника.
Каждый год селекционер растений отбирает зародышевую плазму для совершенствования следующего поколения. Эту зародышевую плазму выращивают при уникальных и отличающихся географических, климатических и почвенных условиях, а затем проводится дополнительный отбор в течение и в конце вегетационного периода. Выводимые культивируемые сорта являются непредсказуемыми. Непредсказуемость является следствием отбора селекционером, который происходит в уникальном природном окружении без контроля на уровне ДНК (с использованием традиционных процедур скрещивания) и с генерируемыми миллионами различных возможных генетических комбинаций. Обычный селекционер при создании линий не может предсказать полученные конечные линии, кроме как в очень приблизительной и общей форме. Один и тот же селекционер не может получить один и тот же культивируемый сорт дважды, используя точно такие же исходные родительские растения и одинаковые методики отбора. Эта непредсказуемость приводит к расходованию большого количества средств на исследования для выведения новых лучших культивируемых сортов подсолнечника.
Выведение новых культивируемых сортов подсолнечника требует выведения и отбора сортов подсолнечника, скрещивания этих сортов, и отбора лучших гибридов при скрещивании. Гибридные семена получают ручным скрещиванием отобранных родительских растений с фертильными мужскими растениями или используя системы мужской стерильности. Эти гибриды отбирают по некоторым одногенным признакам, таким как цвет стручков, цвет цветков, цвет опушения или резистентность к гербицидам, которые указывают на то, что семя действительно является гибридным. Дополнительные данные по родительским линями, а также фенотип гибрида, влияют на решение селекционера о том стоит ли продолжать работу с конкретным гибридом.
Для выведения культивируемых сортов из скрещиваемых популяций используют способы селекции - выведение чистых линий и рекуррентный отбор. Программы скрещивания объединяют желаемые признаки из двух или нескольких культивируемых сортов или различных разрозненных источников в селекционные пулы, из которых выводят культивируемые сорта самоопылением и отбором желаемых фенотипов. Новые культивируемые сорта оценивают для определения их коммерческого потенциала.
Выведение чистых линий обычно используют для улучшения самоопыляющихся сельскохозяйственных культур. Две родительских линии, которые обладают благоприятными взаимодополняющими признаками скрещивают, получая F1. Популяцию F2 получают самоопылением одного или нескольких F1. Отбор лучших индивидуумов можно начинать на популяции F2; затем, начиная с F3, отбирают лучших индивидуумов в лучших семействах. Для улучшения эффективности отбора по признакам с низким наследованием повторное тестирование семейств можно начинать с поколения F4. На поздних стадиях инбридинга (а именно, F6 и F7) лучшие линии или смеси фенотипически похожих линий тестируют на возможное применение в качестве новых культивируемых сортов.
Массовый и рекуррентный отбор можно использовать для улучшения популяций либо само-, либо перекрестно опыляющихся сельскохозяйственных культур. Генетически вариабельная популяция гетерозиготных индивидуумов либо идентифицируется, либо создается перекрестным скрещиванием нескольких различных родителей. Лучшие растения отбирают исходя из индивидуального превосходства, выдающегося потомства или прекрасной комбинационной способности. Проводят перекрестное скрещивание отобранных растений для получения новой популяции, с которой продолжают дополнительные циклы отбора.
Возвратное скрещивание используют для переноса генов для просто и высоконаследуемого признака в желаемый гомозиготный культивируемый сорт или инбредную линию, которые являются рекуррентными родителями. Источник признака, который необходимо перенести, называется донорным родителем. Ожидается, что полученное растение будет иметь характеристики рекуррентного родителя (например, культивируемого сорта) и желаемый признак, перенесенный из донорного родителя. После первоначального скрещивания отбирают индивидуумов, обладающих фенотипом донорного родителя, и повторно скрещивают (возвратное скрещивание) с рекуррентным родителем. Ожидается, что полученное растение будет иметь характеристики рекуррентного родителя (например, культивируемого сорта) и желаемый признак, перенесенный из донорного родителя.
Метод поколения одного семени в строгом смысле относится к высеванию расщепляющейся популяции, сбору образцов по одному семени на растение и использования этого образца для высевания следующего поколения. Когда популяция продвинется от F2 до желательного уровня инбридинга, каждое из растений, из которых получены линии, можно отследить до различных F2-индивидуумов. Число растений в популяции снижается в каждом поколении, вследствие неспособности некоторых семян к прорастанию или некоторых растений к продукции по меньшей мере одного семени. В результате, не все растения F2, исходно представленные в популяции, будут представлены потомством при завершении процесса улучшения поколения.
В многосеменном методе селекционеры подсолнечника обычно собирают семена из каждого растения в популяции и обмолачивают вместе, получая несортированный материал. Часть материала используют для посева следующего поколения, а часть оставляют на хранение. Метод именуется модифицированным методом поколения одного семени.
Многосеменной метод используют для экономии труда при сборе семян. Значительно быстрее удалять семена машинным способом, чем удалять одно семя из каждого растения вручную в случая односеменного метода. Многосеменной метод также делает возможным высевать одинаковое число семян популяции каждого поколения инбридинга. Собирается достаточное число семян, чтобы скомпенсировать влияние тех растений, которые не прорастают или не дают семян.
Описание других способов селекции, обычно используемых для различных признаков и сельскохозяйственных культур, можно найти в одном из нескольких справочных источников (например, Allard, 1960; Simmonds, 1979; Sneep et al., 1979; Fehr, 1987).
Правильное тестирование должно детектировать основные недостатки и уровень превосходства или улучшения над существующими культивируемыми сортами. В дополнение к демонстрации улучшенных характеристик должен существовать спрос на новый культивируемый сорт, который совместим с промышленными стандартами, или который создает новый рынок. Введение нового культивируемого сорта может внести дополнительные затраты производителю семян, производителю растений, переработчику и потребителю из-за специальной рекламы и продаж, измененных семян и технологий коммерческого производства, и использования нового продукта. В тестировании, предшествующему выпуску нового культивируемого сорта, следует учитывать затраты на исследование и выведение, а также техническое превосходство конечного культивируемого сорта. Для размножаемых семенами культивируемых сортов должно быть возможном легкое и экономичное производство семян.
Подсолнечник, Helianthus annuus L., является важной и ценной полевой сельскохозяйственной культурой. Поэтому постоянной целью селекционеров растений является выведение стабильных высокоурожайных культивируемых сортов подсолнечника, которые являются агротехнически устойчивыми. Текущей целью является максимизация количества семян, полученных с используемой площади, и снабжение питанием как животных, так и человека. Для достижения этой цели селекционер подсолнечника должен отбирать и выводить растения подсолнечника, имеющие признаки, которые дают в результате лучшие культивируемые сорта.
Предполагается, что нижеследующие примеры из данной области техники и связанные с ними ограничения, являются иллюстративными, а не исключающими. Другие ограничения области техники будут очевидны специалистам в данной области при прочтении спецификации.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Следующие варианты осуществления изобретения описаны совместно с системами, инструментами и способами, которые подразумеваются типовыми и иллюстративными и не ограничивают объем изобретения. В различных вариантах осуществления изобретения одна или несколько из вышеописанных проблем уменьшаются или устраняются, в то время как другие варианты осуществления направлены на другие усовершенствования.
Изобретение относится к новому растению подсолнечника, дающему семена, имеющие низкое содержание насыщенных жиров. Частично это изобретение относится к семенам подсолнечника, имеющим низкое содержание насыщенных жиров, к растениям или частям растений подсолнечника, дающим семена, имеющие низкое содержание насыщенных жиров, и к способам получения растения подсолнечника, полученного скрещиванием растений подсолнечника, дающим семена с низким содержанием насыщенных жиров, самих с собой или с другим культивируемым сортом подсолнечника, и к созданию с помощью мутагенеза или трансформации вариантов растений подсолнечника, дающих семена с низким содержанием насыщенных жиров.
Аспекты изобретения относятся к новым растениям подсолнечника, дающим семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты. Это изобретение частично относится к семенам подсолнечника, имеющим низкое содержание насыщенных жиров и высокое содержание линолевой кислоты, к растениям или частям растений, подсолнечника, дающим семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты, и к способам получения растения подсолнечника, полученного скрещиванием растений подсолнечника, дающим семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты, самих с собой или с другим культивируемым сортом подсолнечника, и к созданию с помощью мутагенеза или трансформации вариантов растений подсолнечника, дающих семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты.
Примеры семян, имеющих низкое содержание насыщенных жиров, включают, но не ограничены этим, семена, имеющие примерно 2,8% или меньше, примерно 2,9% или меньше, примерно 3% или меньше, примерно 3,1% или меньше, примерно 3,2% или меньше или примерно 3,3% или меньше общего содержания вместе взятых пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0).
Примеры семян, имеющих низкое содержание насыщенных жиров и высокое содержание линолевой кислоты (18:2), включают, но не ограничены этим, семена, имеющие примерно 4,1% или меньше, примерно 5% или меньше, примерно 6% или меньше, примерно 7% или меньше, примерно 8% или меньше, примерно 9% или меньше, примерно 10% или меньше, примерно 11% или меньше или примерно 12% или меньше общего содержания вместе взятых пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0) и имеющие примерно 15%, примерно 20%, примерно 25%, примерно 30%, примерно 35%, примерно 40%, примерно 45%, примерно 50%, примерно 55%, примерно 60%, примерно 65%, примерно 70% или примерно 74% или больше линолевой кислоты (18:2).
Поэтому любые такие способы с использованием частей подсолнечника, которые дают семена, имеющие низкое содержание насыщенного жира и, необязательно, высокое содержание линолевой кислоты, являются частью этого изобретения (например, самоопыление, возвратное скрещивание, получение гибридов, скрещивание с популяциями и т.п.). Все растения, полученные с использованием растений, дающих семена, имеющих в качестве родителя растение с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты, входят в объем этого изобретения. Предпочтительно, растение подсолнечника можно использовать в скрещивании с другими, отличающимся растениями подсолнечника для получения первого поколения (F1) гибридных семян и растений подсолнечника с улучшенными характеристиками.
В другом аспекте настоящее изобретение относится к измененым по одному или множеству генов растениям подсолнечника, дающим семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты. Перенесенный ген (гены) может, предпочтительно, быть доминантным или рецессивным аллелем. Перенесенный ген (гены) может придавать такие признаки, как резистентность к гербицидам, резистентность к насекомым, резистентность к бактериальным, грибковыми или вирусным заболеваниям, мужская фертильность, мужская стерильность, улучшенные питательные качества и промышленная применимость. Ген может представлять собой природный ген подсолнечника или трансген, введенный с помощью генно-инженерных методик.
В другом аспекте настоящее изобретение относится к регенерируемым клеткам для использования в культуре тканей растений подсолнечника, дающих семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты. Культура тканей может быть способна к регенерации растений, имеющих физиологические и морфологические характеристики вышеупомянутого растения подсолнечника, дающего семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, и к регенерации растений, имеющих по существу такой же генотип, как вышеупомянутое растение подсолнечника. Способные к регенерации клетки в таких культурах тканей могут представлять собой зародыши, протопласты, клетки меристемы, каллюс, пыльцу, листья, пыльники, корни, кончики корней, цветки, семена, стручки или стебли. Кроме того, настоящее изобретение относится к растениям подсолнечника, регенерированным из культур тканей по изобретению.
В другом аспекте настоящее изобретение относится к способу введения желаемого признака в растения подсолнечника, дающих семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, причем способ включает: скрещивание растения подсолнечника, дающего семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, с растением другого культивируемого сорта подсолнечника (который содержит желаемый признак) для получения потомства F1, причем желаемый признак выбран из группы, состоящей из мужской стерильности, резистентности к гербицидам, резистентности к насекомым и резистентности к бактериальному заболеванию, грибковому заболеванию или вирусному заболеванию; отбор одного или нескольких растений из потомства, которые имеют желаемый признак, для получения отбираемых растений потомства; скрещивание отбираемых растений потомства с растениями подсолнечника, дающими семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, для получения растений потомства от возвратного скрещивания; отбор растений потомства от возвратного скрещивания, которые имеет желаемый признак и физиологические и морфологические характеристики растений подсолнечника, дающих семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, для получения отбираемых растений потомства при возвратном скрещивании; и повторение этих стадий для получения отбираемых растений первого поколения или следующих поколений потомства при возвратном скрещивании, которые содержат желаемый признак и все физиологические и морфологические характеристики растений подсолнечника, дающих семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты.
В дополнение к типовым аспектам и вариантам осуществления изобретения, описанным выше, при изучении следующих описаний будут очевидны дополнительные аспекты и варианты осуществления.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показано детальное картирование QTL (локусов количественных признаков) низкого содержания стеариновой кислоты в интервале НА1875-НА1865 хромосомы LG17 (А: карты LG17 с новыми маркерами (выделены голубым цветом) и В: детальное картирование низкого содержания стеариновой кислоты в интервале НА1875-ORS565).
На фиг.2 показано выравнивание последовательностей гена KASII-2 из двух родительских линий, на котором показаны SNP и вставки и делеции (идентификационные номера (ID): 333.1 (SEQ ID NO:38) и 333.2 (SEQ ID NO:39) представляют клоны из ампликонов OND163R, а 332.4 (SEQ ID NO:40) и 332.5 (SEQ ID NO:41) из ампликонов H280R[1]/687R-1-8-1).
На фиг.3 показана колокализация QTL низкого содержания пальмитиновой кислоты (А) и гена жирных кислот KASIII-2 (B) на LG 5.
СПОСОБЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В нижеследующих описании и таблицах используется ряд терминов. Для ясного и единого понимания спецификации и формулы изобретения, включая объем изобретения, обеспечиваемый этими терминами, приведены следующие определения.
Аллель. Аллель представляет собой любую одну или несколько альтернативных форм гена, которые все относятся к одному признаку или характеристике. В диплоидной клетке или организме, две аллели данного гена занимают соответствующие локусы на паре гомологичных хромосом.
Возвратное скрещивание. Возвратным скрещиванием является процесс, в котором селекционер повторно скрещивает гибридное потомство с одним из родителей, например, гибрид первого поколения F1 с одним из родительских генотипов гибрида F1.
Элитный подсолнечник. Культивируемый сорт подсолнечника, который был получен стабильным по некоторым коммерчески важным агротехническим признакам, включающим стабильный выход, составляющий примерно 100% или выше, относительно выхода контрольных сортов в той же местности в то же время и при тех же условиях. В одном варианте осуществления «элитный подсолнечник» обозначает культивируемый сорт подсолнечника, который был получен стабильным по некоторым коммерчески важным агротехническим признакам, включающим стабильный выход, составляющий примерно 110% или выше, относительно выхода контрольных сортов в той же местности в то же время и при тех же условиях. В другом варианте осуществления «элитный подсолнечник» обозначает культивируемый сорт подсолнечника, который был получен стабильным по некоторым коммерчески важным агротехническим признакам, включающим стабильный выход, составляющий примерно 115% или выше, относительно выхода контрольных сортов, в той же местности в то же время и при тех же условиях.
Зародыш. Зародышем является маленькое растение, которое содержится в зрелом семени.
FAME-анализ. FAME-анализ (анализ метиловых эфиров жирных кислот) представляет собой способ, позволяющий точное количественное определение жирных кислот, которые составляют классы сложных липидов.
Имидазолиноновая резистентность (Imi). Резистентность и/или толерантность, придаваемая одним или несколькими генами, которые изменяют ацетолактат-синтетазу (ALS), также известную как синтетаза ацетогидроксикислот (AHAS), что позволяет ферменту сопротивляться действию имидазолинона.
Мутагенез. Мутагенез относится к мутагенезу растения или части растения с использованием мутагена (например, химического или физического агента, который увеличивает частоту мутаций в целевом растении или части растения). В качестве неограничивающего примера для получения мутантных аллелей в эндогенных генах растений можно использовать методику двойного химического мутагенеза по Konzak, описанную в патенте США №6696294.
Содержание масла. Его измеряют как процент от целых высушенных семян, и оно отличается для различных сортов. Его можно определить с использованием различных аналитических методик, таких как ЯМР, NIR (отражения в ближнем ИК-диапазоне) и экстракция по Сокслету.
Общее процентное соотношение жирных кислот. Его определяют, экстрагируя из семени образец масла, получая метиловые эфиры жирных кислот, присутствующих в этом образце масла, и анализируя соотношения различных жирных кислот в образце с помощью газовой хроматографии. Композиция жирных кислот также может служить отличительной характеристикой сорта.
Измененный (изменение) по одному гену. Измененное по одному гену растение относится к растениям, которые были выведены с помощью методики селекции растений, называемой возвратным скрещиванием, или методами генной инженерии, в котором восстановлены практически все из желаемых морфологических и физиологических характеристик сорта в дополнение к перенесенному в сорт одному гену с помощью возвратного скрещивания или генной инженерии.
Стабилизированный. Воспроизводимо передаваемый из одного поколения в другое поколение инбредных растений одного сорта.
Общее количество насыщенных жиров (TOTSAT). Общее процентное содержание масла из насыщенных жиров в семени, включая C12:0, C14:0, C16:0, C18:0, C20:0, C22:0 и C24:0.
По одному конкретному варианту осуществления изобретение относится к новому растению подсолнечника, дающему семена, имеющие низкое содержание насыщенных жиров. Этот вариант осуществления относится к семенам подсолнечника, имеющим низкое содержание насыщенных жиров, к растениям или частям растений подсолнечника, дающим семена, имеющие низкое содержание насыщенных жиров, и к способам получения растения подсолнечника, полученного скрещиванием растения подсолнечника, дающего семена с низким содержанием насыщенных жиров, самих с собой или с другим культивируемым сортом подсолнечника, и к созданию вариантов мутагенезом или трансформацией растений подсолнечника, дающих семена с низким содержанием насыщенных жиров.
Другие аспекты изобретения относятся к новым растениям подсолнечника, дающим семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты. Этот вариант осуществления изобретения относится к семенам подсолнечника, имеющим низкое содержание насыщенных жиров и высокое содержание линолевой кислоты, к растениям или частям растений подсолнечника, дающим семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты, и к способам получения растения подсолнечника, полученного скрещиванием растений подсолнечника, дающих семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты, самих с собой или с другим культивируемым сортом подсолнечника, и к созданию вариантов мутагенезом или трансформацией растений подсолнечника, дающих семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты.
Примеры семян, имеющих низкое содержание насыщенных жиров, включают, но не ограничены этим, семена, имеющие общее содержание вместе взятых пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0) примерно 2,8% или меньше, примерно 2,9% или меньше, примерно 3% или меньше, примерно 3,1% или меньше, примерно 3,2% или меньше или примерно 3,3% или меньше.
Примеры семян, имеющих низкое содержание насыщенных жиров и высокое содержание линолевой кислоты (18:2), включают, но не ограничены этим, семена, имеющие общее содержание вместе взятых пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0) примерно 6% или меньше, примерно 4,1% или меньше, примерно 5% или меньше, примерно 6% или меньше, примерно 7% или меньше, примерно 8% или меньше, примерно 9% или меньше, примерно 10% или меньше, примерно 11% или меньше или примерно 12% или меньше, и имеющие примерно 15%, примерно 20%, примерно 25%, примерно 30%, примерно 35%, примерно 40%, примерно 45%, примерно 50%, примерно 55%, примерно 60%, примерно 65%, примерно 70% или примерно 74% или больше линолевой кислоты (18:2).
Поэтому любые такие способы с использованием растений подсолнечника, которые дают семена, имеющие низкое содержание насыщенного жира и, необязательно, высокое содержание линолевой кислоты, являются частью этого изобретения (например, самоопыление, возвратное скрещивание, получение гибридов, скрещивание с популяциями и т.п.). Все растения, полученные с использованием растений, которые дают семена, имеющих низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, входят в объем этого изобретения. Предпочтительно, растение подсолнечника можно использовать в скрещивании с другими, отличающимся растениями подсолнечника для получения семян и растений гибридов подсолнечника первого поколения (F1) с улучшенными характеристиками.
В другом аспекте настоящее изобретение относится к измененым по одному или множеству генов растениям подсолнечника, дающим семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты. Перенесенный ген (гены) может, предпочтительно, быть доминантным или рецессивным аллелем. Перенесенный ген (гены) может придавать такие признаки, как резистентность к гербицидам, резистентность к насекомым, бактериальную резистентность, грибковую резистентность или резистентность к вирусным заболеваниям, мужская фертильность, мужская стерильность, улучшенные питательные качества и промышленная применимость. Ген может представлять собой природный ген подсолнечника или трансген, введенный с помощью генно-инженерных методик.
В другом аспекте настоящее изобретение относится к регенерируемым клеткам для использования в культуре тканей растений подсолнечника, дающих семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты. Культура тканей может быть способна к регенерации растений, имеющих физиологические и морфологические характеристики вышеупомянутого растения подсолнечника, дающего семена, имеющих низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, и к регенерации растений, имеющих по существу такой же генотип, как вышеупомянутое растение подсолнечника. Способные к регенерации клетки в таких культурах тканей могут представлять собой зародыши, протопласты, клетки меристемы, каллюс, пыльцу, листья, пыльники, корни, кончики корней, цветки, семена, стручки или стебли. Кроме того, настоящее изобретение относится к растениям подсолнечника, регенерированным из культур тканей по изобретению.
В другом аспекте настоящее изобретение относится к способу введения желаемого признака в растения подсолнечника, дающие семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, причем способ включает: скрещивание растения подсолнечника, дающего семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, с растением другого культивируемого сорта подсолнечника, которое содержит желаемый признак, для получения потомства F1, причем желаемый признак выбран из группы, состоящей из мужской стерильности, резистентности к гербицидам, резистентности к насекомым и резистентности к бактериальному заболеванию, грибковому заболеванию или вирусному заболеванию; отбор одного или нескольких растений из потомства, которые имеют желаемый признак, для получения отбираемых растений потомства; скрещивание отбираемых растений потомства с растениями подсолнечника, дающими семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, для получения растений потомства от возвратного скрещивания; отбор растений потомства от возвратного скрещивания, которые имеют желаемый признак и физиологические и морфологические характеристики растений подсолнечника, дающих семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты, для получения отбираемых растений потомства от возвратного скрещивания; и повтор этих стадий для получения отбираемых растений первого поколения или следующих поколений потомства от возвратного скрещивания, которые содержат желаемый признак и все физиологические и морфологические характеристики растений подсолнечника, дающих семена, имеющие низкое содержание насыщенных жиров и, необязательно, высокое содержание линолевой кислоты.
Пригодные способы включают, но не ограничены этим, экспрессионные векторы, введенные в ткани растения с использованием способа прямого переноса, такого как опосредованная микрочастицами доставка, инъекции ДНК, электропорация и т.п. Экспрессионные векторы можно ввести в ткани растения с помощью опосредованной микрочастицами доставки с использованием устройства Biolistic, трансформацией агробактериями. Предполагается, что растения-трансформанты, полученные с использованием протоплазмы по изобретению, входят в объем этого изобретения.
С приходом методик молекулярной биологии, которые позволяют выделять и характеризовать гены, кодирующие определенные белковые продукты, ученые в области биологии растений были сильно заинтересованы в изменении генома растений относительно введения и экспрессии чужеродных генов, или дополнительных или модифицированных версий нативных или эндогенных генов (иногда под контролем других промоторов), с целью определенного изменения признаков растения. Такие чужеродные дополнительные и/или модифицированные гены в настоящем описании собирательно именуются «трансгенами». За последние пятнадцать-двадцать лет были разработаны несколько способов получения трансгенных растений, и настоящее изобретение в частных вариантах осуществления также относится к трансформированным версиям заявленных в формуле изобретения сорта или культивируемого сорта.
Трансформация растений включает конструирование экспрессионного вектора, который будет функционировать в клетках растений. Такой вектор содержит ДНК, которая включает ген под контролем регуляторного элемента или функционально связанный с ним (например, промотору). Экспрессионный вектор может содержать один или несколько таких функционально связанных комбинаций гена и регуляторного элемента. Вектор (векторы) могут находиться в плазмидной форме и их можно использовать по одиночке или в комбинации с другими плазмидами для получения трансформированных растений подсолнечника с использованием описанных ниже способов трансформации для включения трансгенов в генетический материал растения (растений) подсолнечника.
Экспрессионные векторы для трансформации подсолнечника: маркерные гены
Экспрессионные векторы включают по меньшей мере один генетический маркер, функционально связанный с регуляторным элементом (например, промотором), который позволяет выделить трансформированные клетки, содержащие маркер, либо отрицательной селекцией (например, ингибированием роста клеток, которые не содержат ген селектируемого маркера) либо положительной селекцией (т.е. скринингом продукта, кодируемого генетическим маркером). В области, связанной с трансформацией, хорошо известно множество часто используемых генов селектируемых маркеров для трансформации растений, и они включают, например, гены, кодирующие ферменты, которые метаболически обезвреживают селективный химический агент, который может представлять собой антибиотик или гербицид, или гены, которые кодируют измененную мишень, нечувствительную к ингибитору. В данной области также известны несколько способов положительной селекции.
Одним часто используемым геном селектируемого маркера для трансформации растений является ген неомицин-фосфотрансферазы II (nptII) под контролем регуляторных сигналов растений, который придает резистентность к канамицину. Смотрите, например, Fraley et al., Proc. Natl. Acad. Sci. U.S.A., 80:4803 (1983). Другим часто используемым геном селектируемого маркера является ген гигромицин-фосфотрансферазы, который придает резистентность к антибиотику гигромицину. Смотрите, например, Vanden Elzen et al., Plant Mol. Biol., 5:299 (1985).
Дополнительные гены селектируемых маркеров бактериального происхождения, которые придают резистентность к антибиотикам, включают гены гентамицин-ацетилтрансферазы, стрептомицин-фосфотрансферазы, аминогликозид-3'-аденил-трансферазы и детерминанту, определяющую резистентность к блеомицину. Смотрите Hayford et al., Plant Physiol. 86:1216 (1988); Jones et al., Mol. Gen. Genet., 210:86 (1987); Svab et al., Plant Mol. Biol. 14:197 (1990); Hille et al., Plant Mol. Biol. 7:171 (1986). Другие гены селектируемых маркеров придают резистентность к гербицидам, таким как глифосат, глюфозинат или бромоксинил. Смотрите Comai et al., Nature 317:741-744 (1985); Gordon-Kamm et al., Plant Cell 2:603-618 (1990); и Stalker et al., Science 242:419-423 (1988).
Другие гены селектируемых маркеров для трансформации растений имеют небактериальное происхождение. Эти гены включают, например, гены мышиной дигидрофолат-редуктазы, растительной 5-енолпирувилшикимат-3-фосфат-синтазы и растительной ацетолактат-синтазы. Смотрите Eichholtz et al., Somatic Cell Mol. Genet. 13:67 (1987); Shah et al., Science 233:478 (1986); Charest et al., Plant Cell Rep. 8:643 (1990).
Другой класс маркерных генов для трансформации растений требует скрининга предположительно трансформированных растительных клеток, вместо прямого генетического отбора трансформированных клеток на резистентность к токсическому веществу, такому как антибиотик. Эти гены особенно полезны для количественной оценки или визуализации пространственной картины экспрессии гена в определенных тканях и часто именуются репортерными генами, поскольку они могут быть слиты с геном или регуляторной последовательностью гена для исследования экспрессии гена. Часто используемые гены для скрининга предположительно трансформированных клеток включают гены β-глюкуронидазы (GUS), β-галактозидазы, люциферазы и хлорамфеникол-ацетил-трансферазы. Смотрите R.A. Jefferson, Plant Mol. Biol. Rep. 5:387 (1987); Teeri et al., EMBO J. 8:343 (1989); Koncz et al., Proc. Natl. Acad. Sci U.S.A. 84:131 (1987); DeBlock et al., EMBO J. 3:1681 (1984).
Не так давно стали доступны способы визуализации активности GUS in vivo, которые не требуют разрушения ткани растений. Molecular Probes publication 2908, Imagene, T.M. Green, p. 1-4 (1993); и Naleway et al., J. Cell Biol. 115:151a (1991). Однако не было доказано, что эти способы визуализации активности GUS in vivo полезны для получения трансформированных клеток вследствие своей низкой чувствительности, высокой фоновой флуоресценции и ограничений, соответствующих с использованием генов люцифераз в качестве селектируемых маркеров.
Позднее в качестве маркера для экспрессии генов в прокариотических и эукариотических клетках использовали ген, кодирующий зеленый флуоресцентный белок (GFP). Смотрите Chalfie et al., Science 263:802 (1994). GFP и мутанты GFP можно использовать в качестве пригодных для скрининга маркеров.
Экспрессионные векторы для трансформации подсолнечника: промоторы
Включенные в экспрессионные векторы гены должны управляться нуклеотидной последовательностью, содержащей регуляторный элемент, например промотор. В области трансформации в настоящий момент хорошо известны несколько типов промоторов, а также другие регуляторные элементы, которые можно использовать поодиночке или в комбинации с промоторами.
Используемый в настоящем описании термин «промотор» относится к области ДНК, которая расположена выше начала транскрипции (в 5'-области от начала транскрипции), и которая вовлечена в узнавание и связывание РНК-полимеразы и других белков для инициации транскрипции. «Растительным промотором» является промотор, способный инициировать транскрипцию в растительных клетках. Примеры промоторов, деятельность которых зависит от стадии развития, включают промоторы, которые предпочтительно инициируют транскрипцию в определенных тканях, таких как листья, корни, семена, волокна, сосуды ксилемы, трахеиды или склеренхима. Такие промоторы именуются «тканепредпочтительными». Промоторы, которые инициируют транскрипцию только в определенных тканях именуются «тканеспецифичными». «Промотор, специфичный к типу клеток» преимущественно управляет экспрессией в определенных типах клеток в одном или нескольких органах, например, клетках сосудов в корнях или листьях. «Индуцируемый» промотор представляет собой промотор под контролем окружающей среды. Примеры условий окружающей среды, которые могут воздействовать на транскрипцию под контролем индуцируемых промоторов включают анаэробные условия или присутствие света. Тканеспецифичные, тканепредпочтительные, специфичные к типу клеток и индуцируемые промоторы составляют класс «неконститутивных» промоторов. «Конститутивным» промотором является промотор, который активен при большинстве условий окружающей среды.
А. Индуцируемые промоторы
Индуцируемый промотор функционально связан с геном для экспрессии в подсолнечнике. Необязательно, индуцируемый промотор функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая функционально связана с геном для экспрессии в подсолнечнике. С индуцируемым промотором скорость транскрипции увеличивается в ответ на индуцирующий агент.
В настоящем изобретении можно использовать любой индуцируемый промотор. Смотрите Ward et al., Plant Mol. Biol. 22:361-366 (1993). Типовые индуцируемые промоторы включают, но не ограничены этим: промоторы из ACEI-системы, которая отвечает на медь (Mett et al., PNAS 90:4567-4571 (1993)); ген In2 из маиса, который отвечает на антидот для бензолсульфонамидных гербицидов (Hershey et al., Mol. Gen. Genetics 227:229-237 (1991); и Gatz et al., Mol. Gen. Genetics 243:32-38 (1994)); и Tet-репрессор из Tn10 (Gatz et al., Mol. Gen. Genetics 227:229-237 (1991)). Особенно предпочтительным индуцируемым промотором является промотор, отвечающий на индуцирующий агент, на который растения в норме не отвечают. Примером индуцируемого промотора является индуцируемый промотор из гена стероидного гормона, транскрипционную активность которого индуцирует глюкокортикостероидный гормон. Schena et al., Proc. Natl. Acad. Sci. U.S.A. 88:0421 (1991).
Б. Конститутивные промоторы
Конститутивный промотор функционально связан с геном для экспрессии в подсолнечнике или конститутивный промотор функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая функционально связана с геном для экспрессии в подсолнечнике.
В настоящем изобретении можно использовать различные конститутивные промоторы. Типовые конститутивные промоторы включают, но не ограничены этим: промоторы из вирусов растений, такие как промотор 35S из CaMV (Odell et al., Nature 313:810-812 (1985)); промоторы актиновых генов риса (McElroy et al., Plant Cell 2:163-171 (1990)); убиквитиновый промотор (Christensen et al., Plant Mol. Biol. 12:619-632 (1989), и Christensen et al., Plant Mol. Biol. 18:675-689 (1992)); pEMU (Last et al., Theor. Appl. Genet. 81:581-588 (1991)); MAS (Velten et al., EMBO J. 3:2723-2730 (1984)); и промотор гистона Н3 маиса (Lepetit et al., Mol. Gen. Genetics 231:276-285 (1992), и Atanassova et al., Plant Journal 2 (3):291-300 (1992)). ALS-промотор, фрагмент XbaI/NcoI в 5'-области структурного гена ALS3 из Brassica napus (или нуклеотидная последовательность, аналогичная фрагменту XbaI/NcoI), представляет особенно пригодный конститутивный промотор. Смотрите заявку РСТ WO 96/30530.
В. Тканеспецифичные или тканепредпочтительные промоторы
Тканеспецифичный промотор функционально связан с геном для экспрессии в подсолнечнике. Необязательно, тканеспецифичный промотор функционально связан с нуклеотидной последовательностью, кодирующей сигнальную последовательность, которая функционально связана с геном для экспрессии в подсолнечнике. Растения, трансформированные представляющим интерес геном, функционально связанным с тканеспецифичным промотором могут продуцировать белковый продукт трансгена исключительно или преимущественно в определенной ткани.
В настоящем изобретении можно использовать любой тканеспецифичный или тканепредпочтительный промотор. Типовые тканеспецифичные или тканепредпочтительные промоторы включают, но не ограничены, предпочтительно экспрессирующийся в корнях промотор, такой как промотор гена фазеолина (Murai et al., Science 23:476-482 (1983) и Sengupta-Gopalan et al., Proc. Natl. Acad. Sci. U.S.A. 82:3320-3324 (1985)); специфично экспрессирующийся в листьях и индуцируемый светом промотор, такой как промотор генов cab или rubisco (Simpson et al., EMBO J. 4(11):2723-2729 (1985) и Timko et al., Nature 318:579-582 (1985)); специфически экспрессирующийся в пыльнике промотор, такой как промотор из LAT52 (Twell et al., Mol. Gen. Genetics 217:240-245 (1989)); специфически экспрессирующийся в пыльце промотор, такой как промотор из Zm13 (Guerrero et al., Mol. Gen. Genetics 244:161-168 (1993)) или предпочтительно экспрессирующийся в микроспорах промотор, такой как промотор из apg (Twell et al., Sex. Plant Reprod. 6:217-224 (1993)).
Транспорт продуцируемого трансгенами белка в клеточные компартменты, такие как хлоропласты, вакуоли, пероксисомы, глиоксомы, клеточная стенка или митохондрии, или для секреции в апопласты, можно осуществить посредством функционального связывания нуклеотидной последовательности, кодирующей сигнальную последовательность с 5'- и/или 3'-областью гена, кодирующего представляющий интерес белок. В ходе белкового синтеза и процессинга направляющие последовательности на 5'- и/или 3'-концах структурного гена могут определять, в какой компартмент в конце концов переместится кодируемый белок.
Присутствие сигнальной последовательности направляет полипептид либо во внутриклеточную органеллу, либо в клеточный компартмент, или на секрецию в апопласты. В данной области известны множество сигнальных последовательностей. Смотрите, например, Becker et al., Plant Mol. Biol. 20:49 (1992); P.S. Close, Master's Thesis, Iowa State University (1993); C. Knox et al., "Structure and Organization of Two Divergent Alpha-Amylase Genes from Barley," Plant Mol. Biol. 9:3-17 (1987); Lerner et al., Plant Physiol. 91:124-129 (1989); Fontes et al., Plant Cell 3:483-496 (1991); Matsuoka et al., Proc. Natl Acad. Sci. 88:834 (1991); Gould et al, J. Cell. Biol. 108:1657 (1989); Creissen et al., Plant J. 2:129 (1991); Kalderon, et al., A short amino acid sequence able to specify nuclear location, Cell 39:499-509 (1984); Steifel, et al., Expression of a maize cell wall hydroxyproline-rich glycoprotein gene in early leaf and root vascular differentiation, Plant Cell 2:785-793 (1990).
Гены чужеродных белков и агротехнические гены
С использованием трансгенных растений по настоящему изобретению можно получать чужеродный белок в промышленных количествах. Поэтому методики отбора и размножения трансформированных растений, хорошо изученные в данной области, дают множество трансгенных растений, которые собирают стандартным образом, а затем чужеродный белок можно выделить из представляющей интерес ткани или из всей биомассы. Выделение белка из растительной биомассы можно осуществить известными способами, которые обсуждаются, например, в Heney and Orr, Anal. Biochem. 114:92-6 (1981).
В аспектах изобретения трансгенное растение, предоставленное для коммерческого получения чужеродного белка, является растением подсолнечника. В других аспектах представляющей интерес биомассой являются семена. Для относительно маленького числа трансгенных растений, у которых наблюдается высокий уровень экспрессии, генетическую карту можно получить главным образом с помощью стандартных методов анализа RFLP (полиморфизма длины рестрикционных фрагментов), ПЦР и SSR (повторов коротких последовательностей), которые идентифицируют приблизительное положение встроенной молекулы ДНК на хромосоме. Примеры методик в этом отношении смотрите в Glick and Thompson, Methods in Plant Molecular Biology and Biotechnology, CRC Press, Boca Raton 269:284 (1993). Информация о картировании в отношении хромосомной локализации полезна для защиты собственности на данное трансгенное растение. При проведении несанкционированного разведения и скрещивания с другой зародышевой плазмой карту области встраивания можно сравнить с аналогичными картами проверяемых растений для того, чтобы определить имеют ли последние общих родителей с данным растением. Сравнение карт может включать гибридизацию, RFLP, ПЦР, SSR и секвенирование, которые являются стандартными методиками.
Аналогичным образом агротехнические гены могут экспрессироваться в трансформированных растениях. Более конкретно, с помощью методов генной инженерии можно создать растения для экспрессии различных фенотипов, представляющих агротехнический интерес. Примеры генов, которые можно использовать в этом отношении, включают, но не ограничены, классифицированные ниже гены.
1. Гены, которые придают резистентность к вредителям или болезням, и которые кодируют:
А) Гены резистентности к болезням растений. Защита растений часто активируется специфическим взаимодействием между продуктом гена резистентности к болезни (R) в растении и продуктом соответствующего невирулентного гена (Avr) в патогене. Сорт растения можно трансформировать клонированными генами резистентности для создания растений, резистентных к специфическим штаммам патогенов. Смотрите, например, Jones et al., Science 266:789 (1994) (клонирование гена Cf-9 томата, придающего резистентность к Cladosporium fulvum); Martin et al., Science 262:1432 (1993) (ген Pto томата, придающего резистентность к Pseudomonas syringae pv. tomato кодирует протеинкиназу); Mindrinos et al., Cell 78: 1089 (1994) (ген RSP2 из Arabidopsis, придающий резистентность к Pseudomonas syringae).
Б) Ген, придающий резистентность к вредителям, таким как соевая цистообразующая нематода. Смотрите, например, заявку РСТ WO 96/30517; заявку РСТ WO 93/19181.
В) Белок из Bacillus thuringiensis либо его производные, либо смоделированный на его основе синтетический полипептид. Смотрите, например, статью Geiser et al., Gene 48:109 (1986), в которой раскрыты клонирование и нуклеотидная последовательность гена Bt δ-эндотоксина. Более того, молекулы ДНК, кодирующие гены δ-эндотоксина, можно приобрести в Американской коллекции типовых культур (АТСС), Мананассас, Вирджиния, например, под номерами доступа в АТСС: 40098, 67136, 31995 и 31998.
Г) Лектин. Смотрите, например, описание, данное в Van Damme et al., Plant Molec. Biol. 24:25 (1994), в котором раскрыты нуклеотидные последовательности нескольких генов маннозосвязывающих лектинов из Clivia miniata.
Д) Витамин-связывающий белок, такой как авидин. Смотрите заявку РСТ US93/06487. В заявке излагается применение авидина и авидиновых гомологов в качестве ларвицидов против насекомых-вредителей.
Е) Ингибитор фермента, например, ингибитор протеазы или протеиназы или ингибитор амилазы. Смотрите, например, J. Biol. Chem. 262:16793 (1987) (нуклеотидная последовательность ингибитора цистеиновой протеазы из риса); Huub et al., Plant Molec. Biol. 21:985 (1993) (нуклеотидная последовательность кДНК, кодирующая протеиназный ингибитор I из табака); Sumitani et al., Biosci. Biotech. Biochem. 57:1243 (1993) (нуклеотидная последовательность ингибитора альфа-амилазы из Streptomyces nitrosporeus); и патент США №5494813 (авторы Hepher и Atkinson, выдан 27 февраля 1996 г.).
Ж) Специфичные для насекомых гормон или феромон, такой как экдистероидный или ювенильный гормон, их вариант, миметик на их основе или их антагонист или агонист. Смотрите, например, описание в Hammock et al., Nature 344:458 (1990) бакуловирусной инфекции клонированной эстеразы ювенильного гормона, которая его инактивирует.
З) Специфичные для насекомых пептид или нейропептид, которые при экспрессии разрушают физиологию пораженного насекомого. Например, смотрите описания в Regan, J. Biol. Chem. 269:9 (1994) (экспрессионное клонирование позволило получить ДНК, кодирующую рецептор диуритеческого гормона насекомых) и Pratt et al., Biochem. Biophys. Res. Comm. 163:1243 (1989) (идентификация аллостатина в Diploptera puntata). Смотрите также патент США № 5266317 авторов Tomalski et al., в котором раскрыты гены, кодирующие специфичные для насекомых, парализующие нейротоксины.
И) Специфичный для насекомых яд, в природе продуцируемый змеями, осами и т.д. Например, смотрите Pang et al., Gene 116:165 (1992), где описана гетерологичная экспрессия в растениях гена, кодирующего токсичный для насекомых пептид из скорпиона.
К) Фермент, отвечающий за избыточное накопление монотерпена, сесквитерпена, стероида, гидроксамовой кислоты, фенилпропаноидного производного или другой небелковой молекулы с инсектицидным действием.
Л) Фермент, вовлеченный в модификацию, в том числе пост-трансляционную модификацию, биологически активной молекулы; например, гликолитический фермент, протеолитический фермент, липолитический фермент, нуклеазу, циклазу, трансаминазу, эстеразу, гидролазу, фосфатазу, киназу, фосфорилазу, полимеразу, эластазу, хитиназу и глюканазу, либо природного, либо искусственного происхождения. Смотрите заявку РСТ WO 93/02197 авторов Scott et al., в которой раскрыта нуклеотидная последовательность гена каллазы. Молекулы ДНК, которые содержат кодирующие хитиназу последовательности, можно получить, например, из АТСС под номерами доступа 39637 и 67152. Смотрите также Kramer et al., Insect Biochem. Molec. Biol. 23:691 (1993), в которой изложена нуклеотидная последовательность кДНК, кодирующая хитиназу гусеницы табачного бражника, и Kawalleck et al., Plant Molec. Biol. 21:673 (1993), где приведена нуклеотидная последовательность полиубиквитинового гена ubi4-2 из петрушки.
М) Молекулу, которая стимулирует передачу сигнала. Например, смотрите раскрытые в статье Botella et al., Plant Molec. Biol. 24:757 (1994), нуклеотидные последовательности для клонов кДНК кальмодулина бобов мунг, и статью Griess et al., Plant Physiol. 104:1467 (1994), в которой приведена нуклеотидная последовательность клона кДНК кальмодулина маиса.
Н) Пептид с гидрофобным моментом. Смотрите заявку РСТ WO 95/16776 (в которой раскрыты пептидные производные тахиплезина, которые ингибируют грибковые патогены растений) и заявку РСТ WO 95/18855 (в которой описаны синтетические антимикробные пептиды, придающие резистентность к болезням).
О) Мембранную пермеазу, которая образует или блокирует каналы. Например, смотрите раскрытую в статье Jaynes et al., Plant Sci. 89:43 (1993) гетерологичную экспрессию аналога секропин-β-литического пептида, который наделяет трансгенные растения табака резистентностью к Pseudomonas solanacearum.
П) Вирус-инвазивный белок или полученный из него комплексный токсин. Например, накопление белков оболочки вируса в трансформированных растительных клетках придает резистентность к вирусной инфекции и/или развитию болезни, вызванной вирусом, из которого получен ген белка оболочки, а также вызванной родственными вирусами. Смотрите Beachy et al., Ann. Rev. Phytopathol. 28:451 (1990). Опосредованная белком оболочки резистентность у трансформированных растений была получена к вирусу мозаики люцерны, вирусу мозаики огурцов, вирусу полосатости табака, вирусу Х картофеля, вирусу Y картофеля, вирусу гравировки табака, вирусу погремковости табака и вирусу мозаики табака. Id.
Р) Специфичное к насекомым антитело или полученный из него иммунотоксин. Поэтому антитело, имеющее своей мишенью критическую метаболическую функцию в кишечнике насекомых, будет инактивировать затронутый фермент, убивая насекомое. Смотрите также Taylor et al., Abstract #497, Seventh Int'l Symposium on Molecular Plant-Microbe Interactions (Edinburgh, Scotland) (1994) (энзиматическая инактивация в трансгенном табаке с помощью продукции одноцепочечных фрагментов антител).
С) Специфичное к вирусу антитело. Смотрите, например, статью Tavladoraki et al., Nature 366:469 (1993), в которой показано, что трансгенные растения, экспрессирующие гены рекомбинантных антител, защищены от атаки вируса.
Т) Останавливающий развитие белок, продуцируемый в природе патогеном или паразитом. Таким образом, грибковые эндо-α-1,4-D-полигалактуроназы способствуют колонизации и высвобождению питательных веществ из растения в результате солюбилизации клеточной стенки гомо-α-1,4-D-галактуроназой. Смотрите Lamb et al., Bio/Technology 10:1436 (1992). Клонирование и характеристика гена, который кодирует ингибирующий эндополигалактуроназу белок из фасоли, описаны в статье Toubart et al., Plant J. 2:367 (1992).
У) Останавливающий развитие белок, продуцируемый в природе растениями. Например, в статье Logemann et al., Bio/Technology 10:305 (1992) было показано, что трансгенные растения, экспрессирующие инактивирующий рибосому ген ячменя, обладали повышенной резистентностью к грибковым заболеваниям.
2. Гены, которые придают резистентность к гербицидам:
А) Гербициду, который ингибирует точку роста или меристему, такому как имидазолинон или сульфонилмочевина. Типовые гены в этой категории кодируют мутантные ферменты ALS и AHAS, описанные, например, в Lee et al., EMBO J. 7:1241 (1988) и Miki et al., Theor. Appl Genet. 80:449 (1990), соответственно.
Б) Гербициду, который ингибирует фотосинтез, такому как триазин (гены psbA и gs+) или бензонитрил (ген нитрилазы). В статье Przibila et al., Plant Cell 3:169 (1991) описана трансформация Chlamydomonas плазмидами, кодирующими мутантный гены psbA. Нуклеотидные последовательности для генов нитрилаз раскрыты в патенте США № 4810648 автора Stalker, а молекулы ДНК, содержащие эти гены, доступны под номерами доступа АТСС - 53435, 67441 и 67442. Клонирование и экспрессия ДНК, кодирующей глутатион-S-трансферазу, описаны в Hayes et al., Biochem. J. 285:173 (1992).
3. Гены, которые придают положительный признак или вносят вклад в положительный признак, такой как:
А) Модифицированный метаболизм жирных кислот, например, в результате трансформации растения антисмысловым геном стеарил-АСР-десатуразы для увеличения содержания стеариновой кислоты в растении. Смотрите Knultzon et al., Proc. Natl. Acad. Sci. U.S.A. 89:2624 (1992).
Б) Снижение содержания фитата: 1) Введение кодирующего фитазу гена будет усиливать деградацию фитата, добавляя больше свободного фосфата трансформированному растению. Например, смотрите Van Hartingsveldt et al., Gene 127:87 (1993), где раскрыта нуклеотидная последовательность гена фитазы из Aspergillus niger. 2) Можно ввести ген, который снижает содержание фитата. Например, в маисе это можно осуществить клонированием и затем повторным введением ДНК, ассоциированной с одним аллелем, который отвечает за мутантные растения маиса, отличающиеся низким содержанием фитовой кислоты. Смотрите Raboy et al., Maydica 35:383 (1990).
В) Модифицированная композиция углеводов, получаемая, например, трансформацией растений геном, кодирующим фермент, который изменяет структуру ветвления крахмала. Смотрите Shiroza et al., J. Bacteol. 170:810 (1988) (нуклеотидная последовательность гена фруктозилтрансферазы из Streptococcus mutants); Steinmetz et al., Mol. Gen. Genet. 20:220 (1985) (нуклеотидная последовательность гена левансукразы из Bacillus subtilis); Pen et al., Bio/Technology 10:292 (1992) (получение трансгенных растений, которые экспрессируют α-амилазу из Bacillus lichenifonnis); Elliot et al., Plant Molec. Biol. 21:515 (1993) (нуклеотидная последовательность генов инвертаз томата); Sogaard et al., J. Biol. Chem. 268:22480 (1993) (сайт-направленный мутагенез гена α-амилазы ячменя); и Fisher et al., Plant Physiol. 102:1045 (1993) (фермент ветвления крахмала II из эндоспермы маиса).
Способы трансформации подсолнечника
Для трансформации растений были разработаны множество способов, включая протоколы биологической и физической трансформации растений. Смотрите, например, Miki et al., "Procedures for Introducing Foreign DNA into Plants" в Methods in Plant Molecular Biology and Biotechnology, B.R. Glick and J.E. Thompson, Eds. (CRC Press, Inc., Boca Raton, 1993), страницы 67-88. В дополнение доступны экспрессионные векторы и способы культивирования in vitro для трансформации растительных клеток или тканей и регенерации растений. Смотрите, например, Gruber et al., "Vectors for Plant Transformation" в Methods in Plant Molecular Biology and Biotechnology, B.R. Glick and J.E. Thompson, Eds. (CRC Press, Inc., Boca Raton, 1993), с. 89-119.
А) Опосредованная агробактериями трансформация. Один способ введения экспрессионного вектора в растения основан на природной системе трансформации агробактерий. Смотрите, например, Horsch et al., Science 227:1229 (1985). A. tumefaciens и A. rhizogenes являются патогенными для растений почвенными бактериями, которые генетически трансформируют растительные клетки. Плазмиды Ti и Ri из A. tumefaciens и A. rhizogenes, соответственно, несут гены, отвечающие за генетическую трансформацию растения. Смотрите, например, C.I. Kado, Crit. Rev. Plant Sci. 10:1 (1991). Описание векторной системы на основе агробактерий и способов опосредованного агробактериями переноса генов приведены в Gruber et al., supra, Miki et al., supra, и Moloney et al., Plant Cell Reports 8:238 (1989). Смотрите также патент США №5563055 (авторов Townsend и Thomas), выданный 8 октября 1996 г.
Б) Прямой перенос генов. В качестве альтернативы для опосредованной агробактериями трансформации был разработан ряд способов трансформации растений, в совокупности именуемых прямым переносом генов. В общем применимым способом трансформации растений является трансформация, опосредованная бомбардировкой микрочастицами, в которой ДНК несут на своей поверхности микрочастицы с размером от 1 до 4 мкм. Экспрессионный вектор вводят в ткани растения с помощью биолистического устройства, которое придает микрочастицам ускорение до 300-600 м/с, что достаточно для проникновения через клеточную стенку и мембраны. Sanford et al., Part. Sci. Technol. 5:27 (1987); J.C. Sanford, Trends Biotech. 6:299 (1988); Klein et al., Bio/Technology 6:559-563 (1988); J.C. Sanford, Physiol. Plant 7:206 (1990); Klein et al., Biotechnology 10:268 (1992). Смотрите также патент США №5015580 (авторов Christou et al.), выданный 14 мая 1991 г.; патент США №5322783 (авторов Tomes et al.), выданный 21 июня 1994 г.
Другим способом физической доставки ДНК в растения является озвучивание целевых клеток. Смотрите Zhang et al., Bio/Technology 9:996 (1991). В качестве альтернативы, для введения экспрессионных векторов в растения используют слияние липосом и сферопластов. Смотрите Deshayes et al., EMBO J, 4:2731 (1985); Christou et al., Proc Natl. Acad. Sci. U.S.A. 84:3962 (1987). Также было опубликовано прямое поглощение ДНК протопластами с использованием CaCl2-преципитации, поливинилового спирта или поли-L-орнитина. Hain et al., Mol. Gen. Genet. 199:161 (1985), и Draper et al., Plant Cell Physiol. 23:451 (1982). Также была описана электропорация протопластов и целых клеток и тканей. Смотрите Donn et al., в рефератах VIIth International Congress on Plant Cell and Tissue Culture IAPTC, A2-38, с. 53 (1990); D'Halluin et al., Plant Cell 4:1495-1505 (1992), и Spencer et al., Plant Mol. Biol. 24:51-61 (1994).
После трансформации целевых тканей подсолнечника экспрессия вышеописанных генов селектируемых маркеров позволяет осуществить предпочтительный отбор трансформированных клеток, тканей и/или растений с использованием способов регенерации и отбора, хорошо известных в данной области.
Вышеописанные способы трансформации обычно будут использоваться для получения трансгенного сорта. Затем трансгенный сорт можно скрещивать с другим (нетрансформированным или трансформированным) сортом для получения нового трансгенного сорта. В качестве альтернативы, генетический признак, который был введен методами генной инженерии в отдельный культивируемый сорт подсолнечника с использованием вышеописанных методик трансформации, можно перенести в другой культивируемый сорт с использованием традиционных методик возвратного скрещивания, которые хорошо известны в области селекции растений. Например, возвратное скрещивание можно использовать для перенесения полученного генно-инженерным путем признака из общедоступного неэлитного сорта в элитный сорт или из сорта, содержащего чужеродный ген в своем геноме, в сорт или сорта, которые не содержат этот ген. Используемый в настоящем описании термин «скрещивание» может относится к простому скрещиванию Х на Y или к процессу возвратного скрещивания в зависимости от контекста.
Культура тканей подсолнечника
Дальнейшее получение растения подсолнечника, дающего семена с низким содержанием насыщенных жиров и, необязательно, с высоким содержанием линолевой кислоты, может происходить в результате самоопыления или с помощью культивирования тканей и регенерации. Культивирование различных тканей подсолнечника и регенерация из них растений являются известными. Например, размножение культивируемого сорта подсолнечника с помощью культуры тканей описано в патенте США 6998516.
Дальнейшее воспроизводство сорта можно осуществить с помощью культуры тканей и регенерации. Культивирование различных тканей соевых бобов и регенерация из них растений хорошо известны и опубликованы в множестве источников. В качестве примера можно привести патент США 6998516. Поэтому в другом аспекте это изобретение относится к клеткам, которые при росте и дифференцировке продуцируют растения подсолнечника, имеющие семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты.
Используемый в настоящем описании термин «культура ткани» указывает на композицию, содержащую выделенные клетки одного и того же или различных типов, или коллекцию таких клеток, упорядоченных в части растения. Примеры типов культур тканей включают протопласты, клетки каллюса, скопления клеток растений и растительные клетки, который генерируют культуру тканей, интактную в растениях или частях растений, таких как зародыши, пыльца, цветки, семена, стручки, листья, стебли, корни, кончики корней, пыльники и т.п. Средства получения и поддержания культуры тканей растений хорошо известны в данной области. В качестве примера, для получения регенерированных растений использовали культуру тканей, содержащую органы. В патентах США №№ 5959185, 5973234 5977445 и 6998516 описаны некоторые методики.
Измененные по одному гену растения
При использовании термина «растение подсолнечника» в контексте настоящего изобретения он также включает любое изменение гена этого сорта. Используемый в настоящем описании термин «изменение гена у растения» относится к тем растениям подсолнечника, которые были выведены с помощью методики селекции, называемой возвратным скрещиванием, или с помощью генной инженерии, в которых восстановлены практические все желаемые морфологические и физиологические характеристики сорта в дополнение к одному гену, перенесенному в сорт с помощью методики возвратного скрещивания. Способы возвратного скрещивания можно использовать с настоящим изобретением для улучшения характеристики сорта или введения характеристики в сорт. Используемый в настоящем описании термин «возвратное скрещивание» относится к повторному скрещиванию гибридного потомства с рекуррентным родителем (например, возвратное скрещивание 1, 2, 3, 4, 5, 6, 7, 8 или больше раз с рекуррентным родителем). Родительское растение подсолнечника, которое вносит ген для желаемой характеристики, называют «нерекуррентным» или «донорным родителем». Эта терминология отражает факт, что в протоколе возвратного скрещивания нерекуррентного родителя используют один раз и, соответственно, к нему не возвращаются. Родительское растение подсолнечника, в которое переносят ген или гены из нерекуррентного родителя, известно как рекуррентный родитель, поскольку его используют в нескольких раундах протокола возвратного скрещивания (Poehlman & Sleper, 1994; Fehr, 1987). В типичном протоколе возвратного скрещивания исходный сорт, представляющий интерес (рекуррентного родителя) скрещивают со вторым сортом (нерекуррентным родителем), который несет один представляющий интерес ген, который необходимо перенести. Потомство, полученное от этого скрещивания, затем скрещивают снова с рекуррентным родителем и процесс повторяют до получения роастения подсолнечника, где в конвертированном растении восстановлены практически все желаемые морфологические и физиологические характеристики рекуррентного родителя в дополнение к одному гену, перенесенному из нерекуррентного родителя.
Отбор подходящего рекуррентного родителя является важной стадией для успешной процедуры возвратного скрещивания. Целью протокола возвратного скрещивания является изменение или замещение одного признака или характеристики в исходном сорте. Для осуществления этого один ген рекуррентного сорта модифицируют или замещают желаемым геном нерекуррентного родителя, в тоже время сохраняя практически всю остальную желаемую генетическую и поэтому желаемую физиологическую и морфологическую структуру исходного сорта. Выбор конкретного нерекуррентного родителя будет зависеть от целей возвратного скрещивания. Одной из основных целей является добавление некоторого желательного с коммерческой точки зрения, агротехнически важного признака в растение. Точный протокол возвратного скрещивания будет зависеть от изменяемых характеристики или признака для определения соответствующего протокола тестирования. Хотя способы возвратного скрещивания упрощаются, если переносимая характеристика представляет собой доминантный аллель, также можно переносить рецессивный аллель. В этом случае может быть необходимо ввести тестирование потомства для определения успешного переноса желаемой характеристики.
Были идентифицированы множество одногенных признаков, которые обычно не отбираются при выведении нового сорта, но которые могут быть улучшены методиками возвратного скрещивания. Одногенные признаки могут быть трансгенными или нет, примеры этих признаков включают, но не ограничены, мужскую стерильность, восковой крахмал, резистентность к гербицидам, резистентность к бактериальным, грибковым и вирусным болезням, резистентность к насекомым, мужская фертильность, улучшенные питательные качества, промышленное применение, стабильная урожайность и увеличение урожайности. Эти гены обычно наследуются через ядро. Несколько из этих одногенных признаков описаны в патентах США №№ 5959185, 5973234 и 5977445.
Это изобретение также направлено на способы получения растения подсолнечника скрещиванием первого родительского растения подсолнечника со вторым родительским растением подсолнечника, в которых первое или второе родительское растения подсолнечника представляет собой растение подсолнечника, дающее семена с низким содержанием насыщенных жиров, и, необязательно, высоким содержанием линолевой кислоты. Кроме того, оба первое и второе родительские растения подсолнечника могут иметь свое происхождение от растения подсолнечника, дающего семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты. Поэтому любые такие способы с использованием растения подсолнечника, дающего семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты, являются частью этого изобретения (т.е. самоопыление, возвратное скрещивание, получение гибридов, скрещивание с популяциями и т.п.). Все растения, полученные с использованием растения подсолнечника, дающего семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты, в качестве родителя входят в объем этого изобретения, включая те, что выведены из сортов, полученных из растения подсолнечника, дающих семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты. Предпочтительно, чтобы растение подсолнечника можно было использовать в скрещивании с другими, отличающимся растениями подсолнечника для получения семян и растений гибридов подсолнечника первого поколения (F1) с улучшенными характеристиками. Сорт по изобретению можно также использовать для трансформации, в которой экзогенные гены вводятся и экспрессируются сортом по изобретению. Подразумевается, что генетические варианты, созданные или с помощью традиционных способов селекции с использованием растения подсолнечника, дающего семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты, или с помощью трансформации растения подсолнечника, дающего семена с низким содержанием насыщенных жиров и, необязательно, высоким содержанием линолевой кислоты, посредством любого из ряда протоколов, известных специалистам в данной области, входят в объем этого изобретения.
ПРИМЕРЫ
Настоящее изобретение дополнительно описано в следующих примерах, которые приведены для иллюстрации, и не предполагается, что они каким-либо образом ограничивают изобретение.
Пример 1: Растения подсолнечника, дающие семена с низким содержанием насыщенных жиров
Посредством обычных селекционных методик была выведена зародышевая плазма подсолнечника с необычно низким содержанием насыщенных жиров. Содержание масел в семенах культивируемых сортов подсолнечника приведено в таблице 1.
Пример 2: Растения подсолнечника, дающие семена с низким содержанием насыщенных жиров и высоким содержанием линолевой кислоты
С помощью обычных селекционных методик была выведена зародышевая плазма подсолнечника с необычно низким содержанием насыщенных жиров. Содержание масел в семенах культивируемых сортов подсолнечника приведено в таблице 2.
Пример 3: Растения подсолнечника, дающие семена с низким содержанием насыщенных жиров
С помощью обычных селекционных методик была выведена зародышевая плазма подсолнечника с необычно низким содержанием насыщенных жиров. Содержание масел в семенах культивируемых сортов подсолнечника приведено в таблице 3.
Как видно из таблицы 3, данные показывают, что масло семян имеет общее содержание насыщенных жиров, сниженное практически до 2,33% при высоком содержании олеиновой кислоты (>80%), отсутствие насыщенных жиров (<3,5%) в NuSun (55-50% олеиновой кислоты), содержание олеиновой кислоты до 95,30%; содержание стеариновой кислоты, сниженное практически до 0,25%, содержание пальмитиновой кислоты, сниженное практически до 1,47%, и низкое содержание насыщенных жиров (<7,0%) в присутствии линолевой кислоты (<55% олеиновой кислоты).
Пример 4: Растения подсолнечника, дающие семена с низким содержанием насыщенных жиров, стеариновой кислоты и пальмитиновой кислоты
С помощью обычных селекционных методик была выведена зародышевая плазма подсолнечника с необычно низким содержанием насыщенных жиров. Содержание масел в семенах культивируемых сортов подсолнечника приведено в таблице 4.
Как видно из таблицы 4, этот набор данных включает низкие значения для содержания стеариновой (0,23%), пальмитиновой (1,37%) и общего содержания насыщенных масел (2,28%).
Пример 5: Разработка маркера низкого содержания стеариновой и пальмитиновой кислот
Стратегия разработки маркера была создана, как описано в настоящем документе. Сначала идентифицировали маркеры из целевых QTL-областей, разработанные в Dow AgroSciences, а также из общедоступных ресурсов, и проводили их скрининг на полиморфизм между родительскими линиями соответствующих картируемым популяциям. Затем проводили скрининг полиморфных маркеров в картируемых популяциях. Для мономорфных (неинформативных) маркеров, были разработаны праймеры для амплификации их соответствующих геномных локусов и определяли нуклеотидные последовательности ампликонов для идентификации однонуклеотидного полиморфизма (SNP) (при наличии такового) между родительскими линиями. Для идентифицированных SNP были разработаны анализы для аллельной дискриминации TaqMan MGB и они были картированы в соответствующих популяциях. Во-вторых, исходя из последовательностей генов-кандидатов для жирных кислот были разработаны праймеры, фланкирующие интроны, для выделения последовательностей генов жирных кислот из родительских линий. На основе нуклеотидного полиморфизма были разработаны маркеры, исходя из их полиморфной природы, и затем проведен их скрининг в картируемых популяциях. Для новых маркеров использовали JoinMap 3.0 (Van Ooijen, 2004a), а для детального картирования QTL использовали MapQTL 5 (Van Ooijen, 2004b).
А) Разработка маркеров для низкого содержания стеариновой кислоты
Разработка SSR-маркера: Проводили скрининг восьми SSR-маркеров на полиморфизм между родительскими линиями ONN687R и H757B/LS10760B-B-17-3-23-5 из картируемой популяции ONN687R×757B/LS10760B-B-17-3-23-5, которую раньше использовали для картирования QTL низкого содержания стеариновой кислоты (смотрите таблицу 5). Четыре SSR маркера - HA0442, CRT22, ORS565 и ORS732 - были полиморфными. Нуклеотидные последовательности ампликонов HA0442 и CRT22 из ONN687R и H757B/LS10760B-B-17-3-23-5 были определены на секвенаторе ABI 3730, и их длина составляла 163 п.о. и 165 п.о., соответственно, для маркера HA0442, и 290 п.о. и 261 п.о., соответственно, для CRT22. Ампликоны ORS565 и ORS732 из ONN687R и H757B/LS10760B-B-17-3-23-5 разделяли на 3%-х гелях Metaphor. Генотипирование соответствующих картируемых популяций ONN687R×H757B/LS10760B-B-17-3-23-5 проводили с HA0442, CRT22, ORS565 и ORS732, используя следующие ПРЦ-праймеры и условия реакции.
HA0442 прямой праймер: 5'-HEX-TGGAACTGTAAATGGACCCAAG-3' (SEQ ID NO:1)
HA0442 обратный праймер: 5'-GCACTGCACCATTTATGAGAAG-3' (SEQ ID NO:2)
CRT22 прямой праймер: 5'-HEX-TCGAGATGAAACCGAATGAAGAAA-3' (SEQ ID NO:3)
CRT22 обратный праймер: 5'-GTTTCTTGGGACTGATATTGCCAAGTGGG-3' (SEQ ID NO:4)
ORS565 прямой праймер: 5'-TGGTCAACGGATTTAGAGTCAA-3' (SEQ ID NO:5)
ORS565 обратный праймер: 5'-TCCAGTTTGGTCTTGATTTGG-3' (SEQ ID NO:6)
ORS732 прямой праймер: 5'-GCACGGAACTCCTCAAATGT-3' (SEQ ID NO:7)
ORS732 обратный праймер: 5'-GCACGGGAAACAAAGAGTCA-3' (SEQ ID NO:8)
Состав реакционной смеси для ПЦР:
ДНК - 4 нг
1ЧПЦР-буфер (Qiagen, Valencia, California)
Прямой праймер - 0,25 мкМ
Обратный праймер - 0,25 мкМ
MgCl2 - 1 мМ
dNTP - 0,1 мМ каждый
PVP - 0,4%
0,04 единицы HotStar Taq ДНК-полимеразы (Qiagen, Valencia, California)
Общий объем: 4,8 мкл
Настройки термоциклера:
Стадия 1: 94°C - 12 минут
Стадия 2: 94°С - 30 секунд
Стадия 3: 55°С - 30 секунд
Стадия 4: 72°С - 30 секунд
Стадия 5: повтор стадий 2, 3 и 4 - 35 циклов
Стадия 6: 72°С - 30 минут
Разработка SNP-маркера: Для разработки SNP-маркеров использовали восемь пар праймеров для амплификации восьми геномных локусов как на ONN687R, так и на H757B/LS10760B-B-17-3-23-5 (таблица 6). Три пары праймеров (ZVG76snpF/R, ZVG77snpF/R и ZVG78snpF/R) были разработаны на основе последовательностей из проб полиморфизма длины рестрикционных фрагментов (RFLP) - ZVG76, ZVG77 и ZVG78 (Kolkman et al. 2007). Последовательности праймеров для HT57F/R, HT64F/R, HT131F/R, HT134F/R и HT210F/R взяты из Lai et al. (2005). SNP был обнаружен в ампликонах из HT64F/R, HT210F/R и ZVG78snpF/R. Анализ аллельной дискриминации TaqMan MGB был разработан для одного SNP-локуса в ампликоне HT64F/R и одного SNP-локуса в ампликоне ZVG78snpF/R (смотрите ниже), и было проведено генотипирование картируемых популяций ONN687R×H757B/LS10760B-B-17-3-23-5 с этими двумя SNP-маркерами с использованием разработанных TaqMan-анализов.
Существует четыре SNP-локуса (отмечены полужирным шрифтом) в ампликонах HT64F/R из ONN687R и H757B/LS10760B-B-17-3-23-5. TaqMan-анализ был разработан для R-локуса. Последовательностями прямого праймера, обратного праймера, пробы 1 и пробы 2 являются: 5'-CCGGCTGCTTCTAGACCTTATAAG-3' (SEQ ID NO:9), 5'-TCGTCGGTGGGACACACA-3' (SEQ ID NO:10), 5'-6FAM-ACTGTTGGATCGGTTC-3' (SEQ ID NO:11) и 5'-VIC-CACTGTTGGATCGATT-3' (SEQ ID NO:12), соответственно.

Также существует четыре SNP-локуса (отмечены полужирным шрифтом) в ампликонах ZVG78snpF/R из ONN687R и H757B/LS10760B-B-17-3-23-5. TaqMan-анализ был разработан для R-локуса на 5'-конце. Последовательностями прямого праймера, обратного праймера, пробы 1 и пробы 2 являются: 5'-GTCCATCTTTCCTCAACGACTTG-3' (SEQ ID NO:14), 5'-CCTAAACGCCTCGAAAAAGCT-3' (SEQ ID NO:15), 5'-6FAM-TTACCATGTCTATAATGC-3' (SEQ ID NO: 16) и 5'-VIC-ATTACCATGTCTGTAATGC-3' (SEQ ID NO: 17), соответственно.

Для двух SNP-маркеров использовали следующие условия ПЦР.
Состав реакционной смеси для ПЦР в реальном времени:
ДНК - 25 нг
1X Taqman Universal PCR Master Mix
Прямой праймер - 22,5 мкМ
Обратный праймер - 22,5 мкМ
Проба 1 - 5 мкМ
Проба 2 - 5 мкМ
Общий объем: 25 мкл
Настройки амплификатора (Bio-Rad iCycler) для проведения ПЦР в реальном времени:
Стадия 1: 95°С - 15 минут
Стадия 2: 94°С - 30 секунд
Стадия 3: 60°С - 1 минута
Стадия 4: повтор стадий 2 и 3 - 65 циклов
Стадия 5: 4°С постоянно
Разработка маркеров вставок-делеций: Были созданы праймеры для амплификации и секвенирования 32 генов, связанных с жирными кислотами, из двух родительских линий ONN687R и H757B/LS10760B-B-17-3-23-5. Семь генов имели полиморфизм, четыре гена слабо амплифицировались, а все другие были мономорфными (таблица 6). Проводили скрининг картируемой популяции ONN687R×H757B/LS10760B-B-17-3-23-5 на все идентифицированные полиморфизмы.
Картирование новых маркеров и детальное картирование QTL низкого содержания стеариновой кислоты: использовали JoinMap 3.0 (Van Ooijen, 2004a) для картирования всех новых идентифицированных полиморфных маркеров. Маркер CRT22 давал значительное нарушение сегрегации и не был картирован. Шесть маркеров, разработанных на основе генов-кандидатов, были картированы на хромосомах отличных от целевой хромосомы 17 (таблица 6). Семь маркеров - HA0442, ORS565, HT64, ZVG78, KASI-2, KASI-4 и ORS732 - были картированы на хромосоме 17. Гены жирных кислот KASI-2 и KASI-4 были картированы на 17-й хромосоме, но не близко к целевому QTL низкого содержание стеариновой кислоты (фиг.1). С использованием новых картированных маркеров было получено детальлное картирование QTL низкого содержания стеариновой кислоты с помощью MapQTL 5 (Van Ooijen, 2004b) в интервале Ha1875-ORS565, который простирается на 27 сМ в верхней теломерной области LG17. Детально картированный QTL имеет значительный показатель сцепления генов 23,2 и объясняет 50,8% вариаций содержания стеариновой кислоты. Новые картированные маркеры можно использовать для облегчения селекции по низкому содержанию стеариновой кислоты в селекционных программах.
Б) Разработка и картирование маркера вставок-делеций для QTL пальмитиновой кислоты
SNP в вставки-делеции наблюдались в последовательностях ампликонов родительских линий H280R[1]/687R-1-8-1 и OND163R с парой праймеров для гена жирных кислот KASIII-2 (таблица 6, фиг.2). Проводили скрининг картируемой популяции H280R[1]/687R-1-8-1×OND163R с этой парой праймеров, и ампликоны разделяли на 3%-ном геле Metaphor. Этот маркер вставки-делеции был локализован внутри QTL низкого содержания пальмитиновой кислоты на группе сцепления 5 (фиг.3) с помощью программы картирования JoinMap 3.0 (Van Ooijen, 2004a).
Список маркеров, исследованных для заполнения QTL-области
низкого содержания стеариновой кислоты
Изученные гены жирных кислот
Хотя это изобретение описано в определенных вариантах осуществления настоящее изобретение можно дополнительно модифицировать в границах сущности и объема этого изобретения. Поэтому предполагается, что эта заявка покрывает любые вариации, применение или адаптации изобретения, использующие его основные принципы. Кроме того, предполагается, что эта заявка покрывает отклонения от настоящего описания, возникающие в границах известной или общепринятой практики в области, к которой относится изобретение, и которые попадают в границы приложенной формулы изобретения.














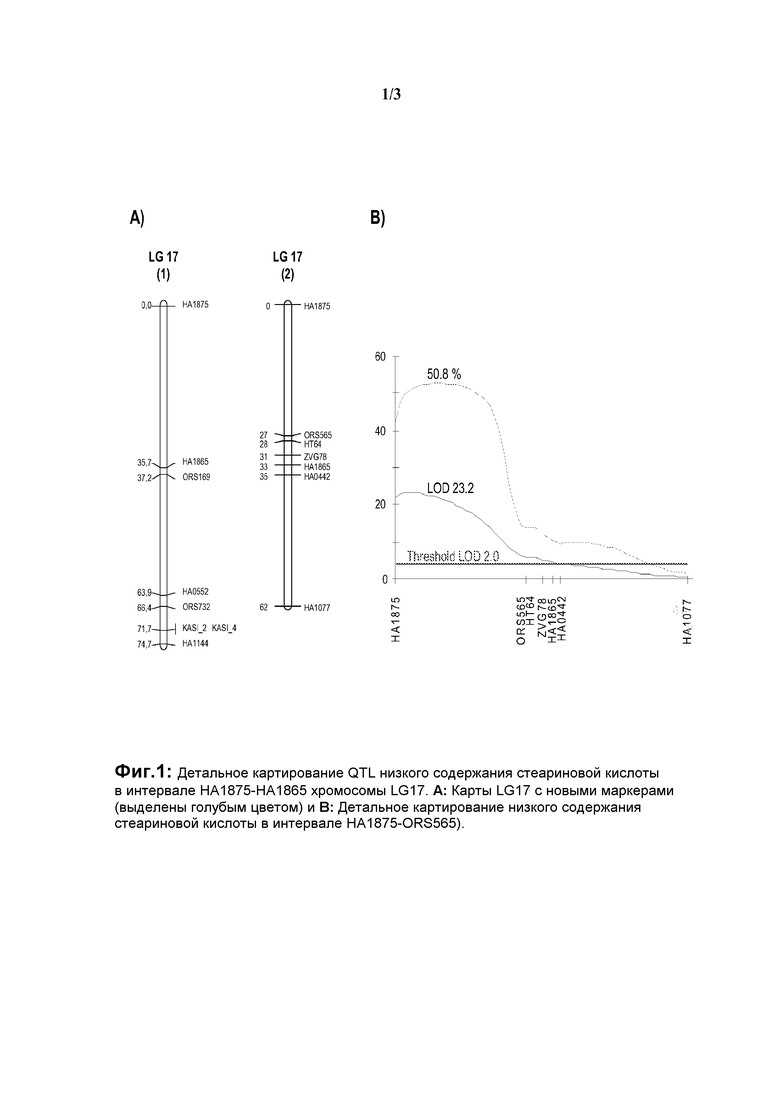

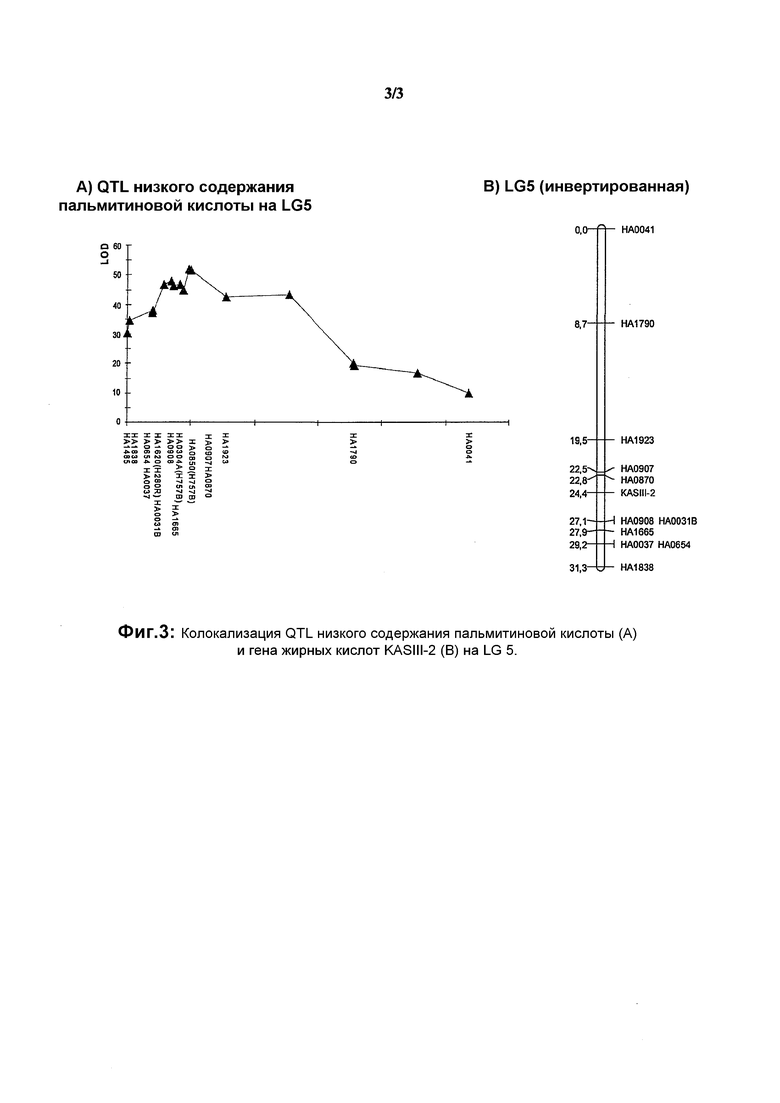
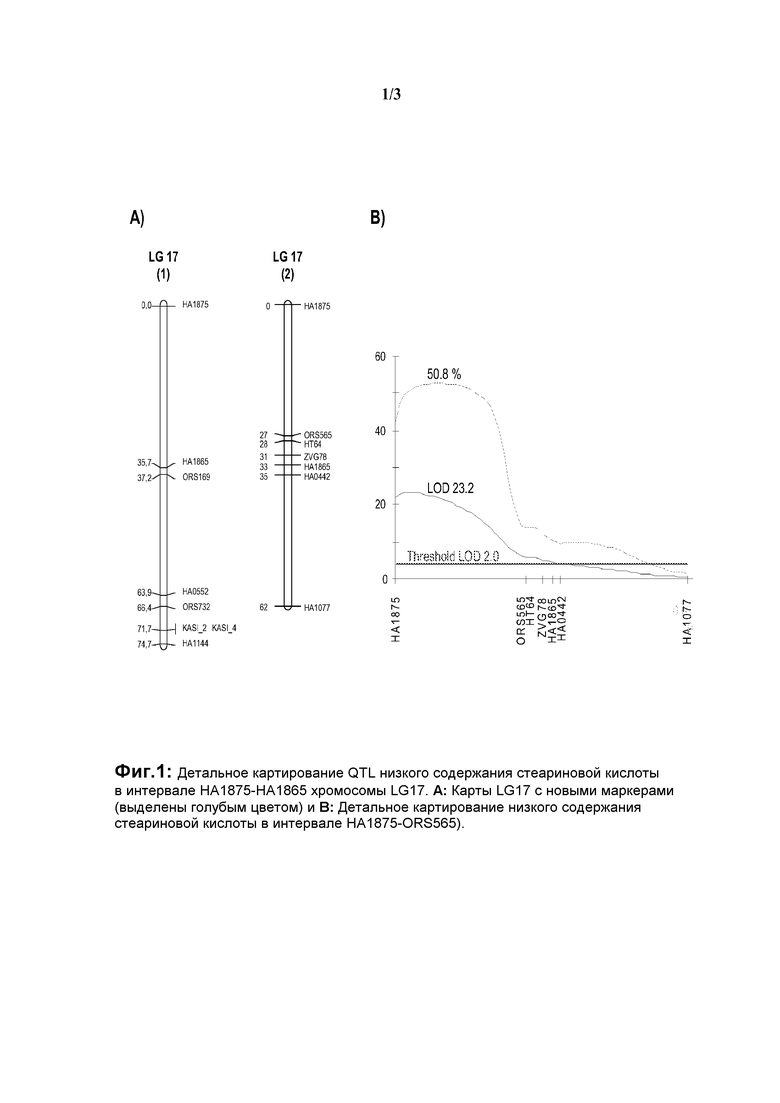

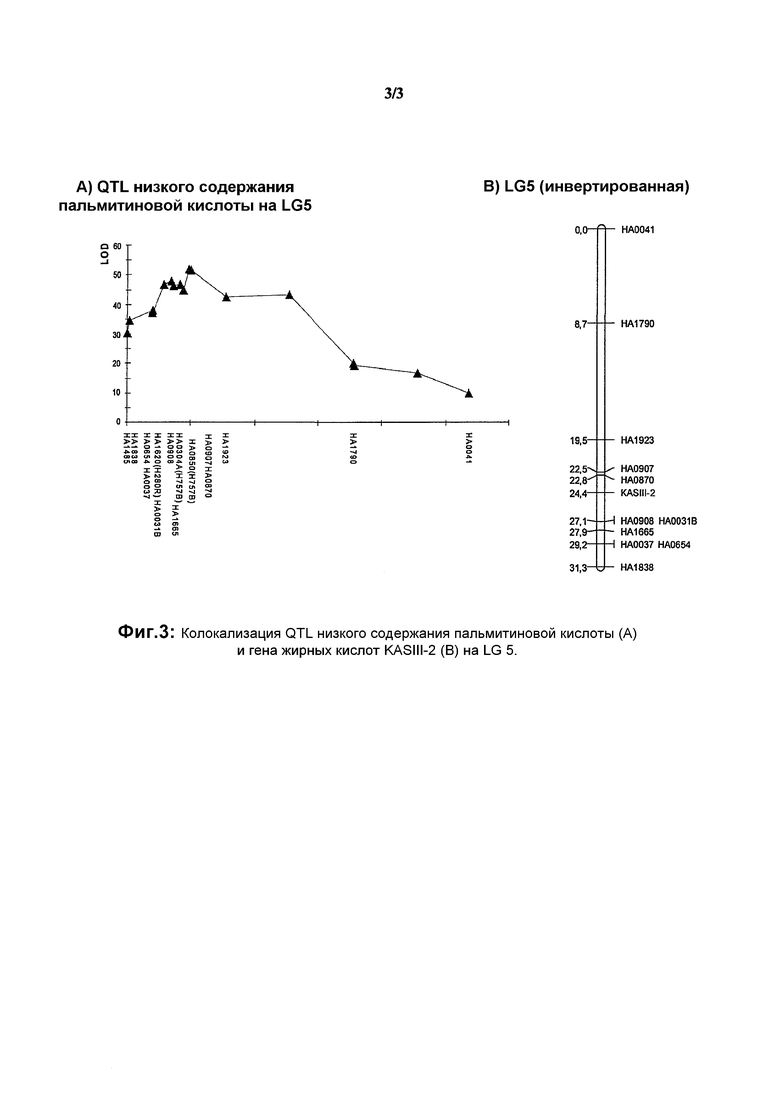
Изобретение относится к области биохимии, в частности к маслу подсолнечника, включающему от 2,64 до 6,02% общего содержания взятых вместе пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0) и от 15,99 до 74,43% содержания линолевой кислоты (18:2), а также к пищевому продукту, его содержащему. Изобретение позволяет эффективно получать масло, характеризующееся определенным содержанием жирных кислот. 2 н. и 5 з.п. ф-лы, 3 ил., 6 табл., 5 пр.
1. Масло подсолнечника, экстрагированное из семян сорта подсолнечника, стабилизированного по признаку масла семян профиля жирных кислот, включающее от 2,64 до 6,02% общего содержания взятых вместе пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0) и от 15,99 до 74,43% содержания линолевой кислоты (18:2).
2. Масло подсолнечника по п. 1, в котором профиль жирных кислот масла подсолнечника включает общее содержание взятых вместе пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0) от 2,64 до 5,2% и общее содержание линолевой кислоты (18:2) от 58,62 до 74, 43%.
3. Масло подсолнечника по п. 1, в котором профиль жирных кислот масла подсолнечника включает от 2,64 до 4,56% общего содержания взятых вместе пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0).
4. Масло подсолнечника по п. 1 или 2 для пищевого применения.
5. Пищевой продукт для пищевого применения, содержащий масло подсолнечника по любому из пп. 1-3, экстрагированного из семян подсолнечника, стабилизированного по признаку масла семян профиля жирных кислот, включающего от 2,64 до 6,02% общего содержания взятых вместе пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0) и от 15,99 до 74,43% линолевой кислоты (18:2).
6. Пищевой продукт по п. 5, где профиль жирных кислот масла подсолнечника включает от 2,64 до 5,2% общего содержания взятых вместе пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0), и от 58,62 до 74,43% содержание линолевой кислоты (18:2).
7. Пищевой продукт по п. 5, где профиль жирных кислот масла подсолнечника включает от 2,64 до 4,56% общего содержания взятых вместе пальмитиновой кислоты (16:0) и стеариновой кислоты (18:0).
| Устройство для регистрации промышленной частоты в энергосистемах | 1974 |
|
SU496504A1 |
| CA 2058849 A1, 10.07.1992 | |||
| COLE G | |||
| et al., New sunflower and soybean cultivars for novel vegetable oil types, Lipid / Fett, 1998, v | |||
| Облицовка комнатных печей | 1918 |
|
SU100A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Кулисный парораспределительный механизм | 1920 |
|
SU177A1 |
| СЕМЕНА ПОДСОЛНЕЧНИКА И МАСЛО, ИМЕЮЩЕЕ ВЫСОКОЕ СОДЕРЖАНИЕ СТЕАРИНОВОЙ КИСЛОТЫ | 1995 |
|
RU2147407C1 |
| EP 1598422 А2, 23.11.2005. | |||

