ПРИТЯЗАНИЕ НА ПРИОРИТЕТ
По настоящей заявке испрашивается приоритет согласно 35 U.S.C. § 119(e) временной заявки США № 61/445426, поданной 22 февраля 2011 года, "CANOLA GERMPLASM EXHIBITING SEED COMPOSITIONAL ATTRIBUTES THAT DELIVER ENHANCED CANOLA MEAL NUTRITIONAL VALUE HAVING OMEGA-9 TRAITS".
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к зародышевой плазме и сортам канолы. В некоторых вариантах осуществления изобретение относится к зародышевой плазме, имеющей признаки состава муки (например, сниженные уровни антипитательных факторов и увеличенные уровни белка), которые модифицированы независимо от цвета оболочки семян. Конкретные варианты осуществления относятся к зародышевой плазме канолы, проявляющей темный цвет семян в комбинации, например, со сниженными уровнями антипитательных факторов (например, кислотное детергентное волокно (ADF) и полифенольные соединения) и увеличенными уровнями белка и фосфора.
УРОВЕНЬ ТЕХНИКИ
"Канола" относится к рапсу (Brassica spp.), который имеет содержание эруковой кислоты (C22:1) не более 2 процентов по массе (по сравнению с общим содержанием жирных кислот в семенах) и который обеспечивает (после перемалывания) высушенную воздухом муку, содержащую менее 30 микромоль (мкмоль) глюкозинолатов на грамм обезжиренной (не содержащей масел) муки. Эти типы рапса отличаются их пригодностью в пищу по сравнению с более традиционными сортами этого вида. Масло канолы является превосходным пищевым маслом вследствие его низких уровней насыщенных жирных кислот.
Хотя в рапсовой муке содержание белка является относительно высоким, содержание в ней волокон снижает ее усвояемость и ее ценность в качестве корма для животных. По сравнению с соевой мукой, мука канолы и масличного рапса содержит более высокое количество диетической клетчатки и более низкий процент белка. Вследствие ее более высокого содержания пищевых волокон, мука канолы имеет приблизительно на 20% меньше метаболизируемой энергии (ME), чем соевая мука. В результате, ценность муки остается низкой относительно муки из других семян масличных культур, таких как соевая мука, в частности, в рационах свиней и птиц. Rakow (2004a) Canola meal quality improvement through the breeding of yellow-seeded varieties-an historical perspective, AAFC Sustainable Production Systems Bulletin. Кроме того, присутствие глюкозинолатов в некоторых типах муки канолы также снижает ее ценность вследствие вредоносных эффектов, которые эти соединения оказывают на рост и размножение домашнего скота.
Сорта канолы отличаются частично их цветом оболочки семян. Цвет оболочки семян, как правило, подразделяется на два основных класса: желтый и черный (или темно-коричневый). Также наблюдаются различные оттенки этих цветов, такие как красновато-коричневый и желтовато-коричневый. Часто наблюдается, что сорта канолы с более светлым цветом оболочки семян имеют более тонкую оболочку и, таким образом, меньше волокна и больше масла и белка, чем сорта с темной оболочкой семян. Stringam et al. (1974) Chemical and morphological characteristics associated with seed coat color in rapeseed, Proceedings of the 4th International Rapeseed Congress, Giessen, Germany, pp. 99-108; Bell and Shires (1982) Can. J. Animal Science 62:557-65; Shirzadegan and Robbelen (1985) Gotingen Fette Seifen Anstrichmittel 87:235-7; Simbaya et al. (1995) J. Agr. Food Chem. 43:2062-6; Rakow (2004b) Yellow-seeded Brassica napus canola for the Canadian canola Industry, AAFC Sustainable Production Systems Bulletin. Одно из возможных объяснений этому состоит в том, что растение канолы может тратить больше энергии на продуцирование белков и масел, если ему не требуется эта энергия для продуцирования компонентов волокон оболочки семян. Также описано, что линии канолы с желтыми семенами имеют более низкое содержание глюкозинолата, чем линии канолы с черными семенами. Rakow et al. (1999b) Proc. 10th Int. Rapeseed Congress, Canberra, Australia, Sep. 26-29, 1999, Poster #9. Таким образом, исторически следуют по пути выведения сортов канолы с желтыми семенами в качестве потенциального пути увеличения пищевой ценности муки канолы. Bell (1995) Meal and by-product utilization in animal nutrition, Brassica oilseeds, production and utilization. Eds. Kimber and McGregor, Cab International, Wallingford, Oxon, OX108DE, UK, pp. 301-37; Rakow (2004b), выше; Rakow & Raney (2003).
Было показано, что некоторые формы видов Brassica с желтыми семенами, близкородственные B. napus (например, B. rapa и B. juncea) имеют более низкие уровни волокон в их семенах, а затем в муке. Создание зародышевой плазмы B. napus с желтыми семенами продемонстрировало, что количество волокон в B. napus может быть уменьшено путем введения генов, контролирующих пигментацию семян, из родственных видов Brassica. Однако введение генов, контролирующих пигментацию семян, из родственных видов Brassica в ценные масличные сорта Brassica, такие как сорта канолы, усложняется тем фактом, что в наследование желтых оболочек семян в доступных в настоящее время линиях с желтыми семенами вовлечено множество рецессивных аллелей. Более того, "скручивание стручков" также является проблемой, часто встречающейся в ходе переноса желтого цвета оболочки семян из других видов Brassica, таких как juncea и carinata.
Доступно очень мало информации в отношении того, какова вариабельность для волокон в зародышевой плазме B. napus с темными семенами, и отсутствуют сообщения о выведении линий канолы с темными семенами, которые содержат уменьшенные уровни антипитательных факторов (например, волокна и полифенольные соединения) и увеличенные уровни белка.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В настоящем описании описаны перекрестноопыляющиеся сорта (CL044864, CL065620) и гибриды (CL166102H, CL121460H и CL121466H) канолы (Brassica napus), содержащие зародышевую плазму, обеспечивающую новую комбинацию изменений состава муки канолы, которые, как было показано, влияют на питательную ценность. В некоторых вариантах осуществления растения канолы, содержащие зародышевую плазму по изобретению, могут продуцировать семена, например, с новыми комбинациями уровней белка, волокон и фосфора, так что эти компоненты семян являются независимыми от цвета оболочки семян. В конкретных вариантах осуществления такие растения могут продуцировать семена с более высоким содержанием белка и более низким содержанием волокон, чем стандартные типы канолы, а также уровни фосфора, которые сходны с или превышают уровни фосфора в стандартных типах канолы. Инбредные линии и гибриды канолы, содержащие зародышевую плазму по изобретению, в некоторых вариантах осуществления могут обеспечивать свойства повышенной питательности муки при использовании непосредственно в качестве ингредиента кормов или пищи и/или при использовании в качестве сырья для обработки изолятов и концентратов белка. Такие семена могут быть темными (например, черными, темными и крапчатыми) или светлыми.
Таким образом, в настоящем описании описана зародышевая плазма Brassica, которую можно использовать для получения растений канолы, имеющих желаемые признаки состава семян независимо от цвета семян. В некоторых вариантах осуществления растения, содержащие такую зародышевую плазму, можно использовать для получения муки канолы с желаемыми питательными качествами. В конкретных вариантах осуществления предусмотрены инбредные линии канолы (и их растения), содержащие зародышевую плазму по изобретению. В следующих вариантах осуществления предусмотрены гибридные линии канолы (и их растения), имеющие инбредное растение канолы, содержащее зародышевую плазму по изобретению, в качестве исходного растения. Сорта канолы по изобретению включают, например, но не ограничиваясь ими: CL044864; CL065620; CL166102H; CL121460H и CL121466H.
Конкретные варианты осуществления изобретения включают зародышевую плазму канолы, придающую семенам канолы признаки высокого содержания белка и низкого содержания волокон, где растение канолы продуцирует семена, имеющие, в среднем, по меньшей мере 68% олеиновой кислоты (C18:1) и менее 3% линоленовой кислоты (C18:3). В других вариантах осуществления растение канолы включает зародышевую плазму канолы. Также описаны семена, продуцируемые растением канолы. Дополнительные варианты осуществления включают растение-потомка, выращенное из семян растения канолы. Также описаны способы внесения в сорт канолы по меньшей мере одного желаемого признака, выбранного из группы, состоящей из высокого содержания белка, низкого содержания волокон, по меньшей мере 68% олеиновой кислоты (C18:1) и менее 3% линоленовой кислоты (C18:3), независимо от цвета оболочки семян.
Также в настоящем описании описаны растительные товарные продукты, получаемые из инбредных растений или гибридов канолы, содержащих зародышевую плазму по изобретению. Конкретные варианты осуществления включают муку канолы или семена, получаемые из такого инбредного растения или гибрида канолы.
Также описаны способы повышения питательной ценности муки канолы. Например, описаны способы интрогрессии комбинации характеристик состава муки канолы в зародышевую плазму Brassica независимо от цвета семян. В конкретных вариантах осуществления зародышевую плазму по изобретению можно комбинировать с зародышевой плазмой канолы, которая характеризуется желтой оболочкой семян, с получением зародышевой плазмы, которая способна обеспечивать муку канолы с улучшенными желаемыми характеристиками, сообщенными каждой из зародышевых плазм.
Указанные выше и другие признаки станут более понятными из следующего подробного описания нескольких вариантов осуществления, которое следует далее с отсылкой на прилагаемые чертежи.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 представлены изображения нескольких сортов канолы, имеющих темный цвет оболочки семян.
На фиг.2 представлены данные анализа состава семян определенных инбредных линий и гибридов B. napus. Образцы семян взяты из повторных испытаний в западной Канаде. Данные о составе семян были предсказаны на основе NIR, а затем подтверждены с использованием эталонных химических способов.
СПОСОБ(Ы) ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
I. Обзор нескольких вариантов осуществления
Мука канолы представляет собой фракцию семян канолы, остающуюся после процесса экстракции масла. Мука канолы является источником белка, и, таким образом, ее используют в нескольких применениях, включая составление кормов для животных и выделение высокоценных концентратов и изолятов белка. Волокна в оболочке семян, семядоле и зародыше, которые попадают в муку, ограничивают частоту включения муки канолы в корм моногастрических видов животных, и, таким образом, мука канолы, как правило, не обеспечивает той же пищевой ценности, что и мука, полученная из других источников (например, соя). Было показано, что формы с желтыми семенами в видах, близкородственных B. napus (например, B. rapa и B. juncea), имеют более низкие уровни волокон в их семенах и, следовательно, в муке. Это наблюдение стало причиной попыток введения признака низкого содержания волокон в семенах в B. napus зависимым от желтого цвета семян образом. Разработка полученной зародышевой плазмы B. napus с желтыми семенами продемонстрировала, что количество волокон может быть снижено в B. napus с помощью этого подхода.
До настоящего изобретения не предполагали, что сорта канолы с темными семенами могут проявлять содержание волокон в семенах, настолько же низкое, как наблюдали в сортах с желтыми семенами. Более того, линии канолы с темными семенами, содержащие сниженные уровни антипитательных факторов (например, волокна и полифенольные соединения) и увеличенные уровни белка и фосфора, которые могут быть источниками улучшенной муки канолы, не были описаны. В некоторых вариантах осуществления зародышевая плазма канолы, описанная в настоящем описании, обеспечивает комбинации нескольких ключевых признаков улучшенного состава муки, которые экспрессируются независимо от цвета оболочки семян. В конкретных вариантах осуществления может быть достигнута более высокая частота включения муки канолы, полученной из семян канолы, содержащих зародышевую плазму по изобретению, в пищевой рацион, например, свиней и птиц.
Зародышевую плазму по изобретению можно использовать (например, через селекцию) для выведения канолы, имеющей желаемые признаки состава семян с одним или несколькими дополнительными желаемыми признаками (например, улучшенный состав масла, увеличенное продуцирование масла, модифицированный состав семян, увеличенное содержание белка, устойчивость к заболеваниям, устойчивость к паразитам, устойчивость к гербицидам и т.д.). Зародышевую плазму по изобретению можно использовать в качестве исходной зародышевой плазмы, посредством которой могут быть внесены дополнительные изменения в состав семян так, чтобы можно было вывести линии и гибриды канолы, которые обеспечивают муку канолы, имеющую более значительные усовершенствования описанного в настоящем описании типа.
II. Сокращения
ADF кислотное детергентное волокно
ADL кислотный детергентный лигнин
AID кажущаяся усвояемость в подвздошной кишке
AME кажущаяся метаболизирующаяся энергия
BSC канола с черными семенами
CP процент неочищенного белка
DM концентрация сухого вещества
ECM улучшенная мука канолы по настоящему изобретению
FAME жирные кислоты/метиловые сложные эфиры жирных кислот
GE валовая энергия
HT "высокотемпературная" обработка
LT "низкотемпературная" обработка
NDF нейтральное детергентное волокно
ЯМР ядерный магнитный резонанс
NIR спектроскопия в ближней инфракрасной области
SAE сложный эфир синоповой кислоты
SBM соевая мука
SER растворимый экстрагируемый остаток
SID стандартизированная усвояемость в подвздошной кишке
TAAA истинная доступность аминокислот
TDF общее содержание пищевых волокон
TME истинная метаболизируемая энергия
WF белые хлопья
III. Термины
Обратное скрещивание: способы обратного скрещивания можно использовать для введения последовательности нуклеиновой кислоты в растения. Способ обратного скрещивания широко используют на протяжении десятилетий для введения новых признаков в растения. Jensen, N., Ed. Plant Breeding Methodology, John Wiley & Sons, Inc., 1988. В типичном протоколе обратного скрещивания исходный представляющий интерес сорт (рекуррентное родительское растение) скрещивают со вторым сортом (нерекуррентное родительское растение), который содержит представляющий интерес ген, подлежащий трансфекции. Затем полученное потомство от этого скрещивания вновь скрещивают с рекуррентным родительским растением и процесс повторяют до тех пор, пока не получают растение, где по существу все из желаемых морфологических и физиологических характеристик рекуррентного растения восстановлены в преобразованном растении, в дополнение к перенесенному гену из нерекуррентного родительского растения.
Масло канолы: масло канолы относится к маслу, экстрагированному из коммерческих сортов рапса. Для получения масла канолы семена, как правило, сортируют и смешивают в зерновых элеваторах с получением единообразного продукта. Затем смешанные семена измельчают, и масло обычно экстрагируют гексаном, а затем рафинируют. Затем полученное масло можно продавать для применения. Содержание масла обычно измеряют в качестве процента от цельных высушенных семян, и содержание конкретных масел является характеристикой различных сортов канолы. Содержание масел можно легко и стандартным образом определить с использованием различных аналитических способов, например, и не ограничиваясь ими: ЯМР; NIR; экстракции Сокслета, или другими способами, широко доступными специалистам в данной области. См. Bailey, Industrial Oil & Fat Products (1996), 5th Ed. Wiley Interscience Publication, New York, New York. Процентное содержание общих жирных кислот, как правило, определяют путем экстракции образца масла из семян, получения метиловых сложных эфиров жирных кислот, присутствующих в образце масла, и анализа соотношений различных жирных кислот в образце с использованием газовой хроматографии. Состав жирных кислот также может быть отличительной характеристикой конкретных сортов.
Коммерчески полезный: как используют в рамках изобретения, термин "коммерчески полезный" относится к линиям и гибридам растений, которые имеют достаточную растительную мощность и плодородие, чтобы культуру линии или гибрида растения могли получить фермеры с использованием общепринятого сельскохозяйственного оборудования. В конкретных вариантах осуществления из растений или растительных материалов коммерчески полезного сорта могут быть экстрагированы товарные продукты с описанными компонентами и/или качествами. Например, масло, содержащее желаемые компоненты масла, может быть экстрагировано из семян коммерчески полезной линии или гибрида растений с использованием общепринятого оборудования для измельчения и экстракции. В определенных вариантах осуществления коммерчески полезная линия растений представляет собой инбредную линию или гибридную линию. "Агрономически элитные" линии и гибриды, как правило, имеют желаемые агрономические характеристики; например, и не ограничиваясь ими: увеличенный выход по меньшей мере одного товарного продукта; зрелость; устойчивость к заболеваниям; и устойчивость к полеганию.
Элитная линия: любая растительная линия, которая является результатом скрещивания и селекции в целях улучшения агрономических характеристик. Элитное растение представляет собой любое растение из элитной линии.
Улучшенная мука канолы: как используют в рамках изобретения, термин "улучшенная мука канолы" означает муку канолы с улучшенным составом, получаемую путем обработки семян канолы, которые имеют увеличенные уровни белка и уменьшенные уровни по меньшей мере некоторого антипитательного компонента. Улучшенная мука канолы в настоящем описании, в рамках настоящего изобретения, может по-разному называться "ECM", "ECM канолы с черными семенами", "BSC ECM" или "DAS BSC ECM". Однако настоящее изобретение не ограничивается только ECM канолы с зародышевой плазмой с черными семенами.
По существу производный: в некоторых вариантах осуществления манипуляции с растениями, семенами или их частями могут приводить к созданию по существу производных сортов. Как используют в рамках изобретения, термин "по существу производный" соответствует соглашению, принятому The International Union for the Protection of New Varieties of Plants (UPOV):
[A] сорт считается по существу производным из другого сорта ("исходный сорт"), когда
(i) он преимущественно происходит из исходного сорта или из сорта, который сам по себе преимущественно происходит из исходного сорта, с сохранением проявления неотъемлемых характеристик, которые являются результатом генотипа или комбинации генотипов исходного сорта;
(ii) отчетливо отличается от исходного сорта; и
(iii) за исключением отличий, которые являются результатом преобразования, он соответствует исходному сорту в проявлении неотъемлемых характеристик, которые являются результатом генотипа или комбинации генотипов исходного сорта.
UPOV, Sixth Meeting with International Organizations, Geneva, Oct. 30, 1992 (документ, подготовленный Office of the Union).
Растительный товарный продукт: как используют в рамках изобретения, термин "растительный товарный продукт" относится к товарам, изготавливаемым из конкретного растения или части растения (например, растения, содержащего зародышевую плазму по изобретению, и части растения, полученной из растения, содержащего зародышевую плазму по изобретению). Товарный продукт может представлять собой, например, и не ограничиваясь ими: зерно; муку; фураж; белок; выделенный белок; муку мелкого помола; масло; измельченные или цельные зерна или семена; любой пищевой продукт, содержащий любую муку, масло или измельченные или цельные зерна; или силос.
Линия растений: как используют в рамках изобретения, "линия" относится к группе растений, которые проявляют небольшое генетическое варьирование (например, отсутствие генетического варьирования) между индивидуумами по меньшей мере по одному признаку. Инбредные линии могут быть созданы с помощью нескольких поколений самоопыления и селекции или, альтернативно, путем вегетативного размножения из исходного родителя с использованием способов культивирования тканей или клеток. Как используют в рамках изобретения, термины "культивар", "сорт" и "тип" являются синонимами, и эти термины относятся к линии, которую используют для промышленного производства.
Растительный материал: как используют в рамках изобретения, термин "растительный материал" относится к любому обработанному или необработанному материалу, происходящему, целиком или частично, из растения. Например, и не ограничиваясь этим, растительный материал может представлять собой часть растения, семя, плод, лист, корень, ткань растения, культуру ткани растения, эксплантат растения или клетку растения.
Стабильность: как используют в рамках изобретения, термин "стабильность" или "стабильный" относится к данному компоненту или признаку растения, который наследуется и поддерживается на одном и том же уровне на протяжении множества поколений семян. Например, стабильный компонент может сохраняться в течение по меньшей мере трех поколений по существу на одном и том же уровне. В этом контексте термин "по существу один и тот же" в некоторых вариантах осуществления может относиться к компоненту, сохраненному с отличием в пределах 25% между двумя различными поколениями; в пределах 20%; в пределах 15%; в пределах 10%; в пределах 5%; в пределах 3%; в пределах 2%; и/или в пределах 1%, а также к компоненту, который сохранен абсолютно между двумя различными поколениями. В некоторых вариантах осуществления стабильный компонент может представлять собой, например, и не ограничиваясь этим, масляный компонент; белковый компонент; волокнистый компонент; пигментный компонент; глюкозинолатный компонент и лигниновый компонент. На стабильность компонента может влиять один или несколько факторов окружающей среды. Например, на стабильность масляного компонента может влиять, например, и не ограничиваясь ими: температура; место посева; стрессовые воздействия; и время посева. Ожидается, что последующие поколения растения, имеющие стабильный компонент в полевых условиях, будут продуцировать растительный компонент аналогичным образом, например, как указано выше.
Признак или фенотип: термины "признак" и "фенотип" используют в настоящем описании взаимозаменяемо.
Сорт или культивар: термины "сорт" или "культивар" относятся в настоящем описании к линии растений, используемой для промышленного производства, которая является отличимой, стабильной и единообразной по ее характеристикам при размножении. В случае гибридного сорта или культивара, родительские линии являются отличимыми, стабильными и единообразными по их характеристикам.
Если нет иных указаний, форма единственного числа, как используют в рамках изобретения, относятся по меньшей мере к одному.
IV. Зародышевая плазма канолы, обеспечивающая желаемые признаки состава семян независимо от цвета семян
В предпочтительном варианте осуществления изобретение относится к зародышевой плазме Brassica, которую можно использовать для получения растений канолы, имеющих желаемые признаки состава семян независимо от цвета семян. Также предусмотрены конкретные иллюстративные инбредные линии канолы и гибриды, содержащие эту зародышевую плазму.
Является общепризнанным, что масло канолы представляет собой очень полезное масло для потребления как человеком, так и животными. Однако мучной компонент семян канолы, который остается после экстракции масляного компонента, хуже соевой муки вследствие высокого содержания волокон и более низкой питательной ценности. В некоторых вариантах осуществления растения канолы, содержащие зародышевую плазму по изобретению, могут уменьшать или преодолевать эти недостатки и могут обеспечивать муку канолы в качестве высокопитательного и экономичного источника корма для животных. Мука канолы является побочным продуктом производства масла канолы, и, таким образом, масло канолы, обеспечиваемое настоящим изобретением, сохраняет ценные ресурсы, позволяя использование этого побочного продукта конкурентно с другими видами муки.
Ранее полагали, что желтый цвет семян канолы сам по себе является важным, поскольку полагали, что он соответствует улучшенным питательным характеристикам мучного компонента, получаемого после экстракции масла. Некоторые варианты осуществления впервые могут обеспечить зародышевую плазму для канолы с низким содержанием волокон с темными семенами (например, темными, черными и крапчатыми семенами), которая также обеспечивает улучшенное масло с высоким содержанием олеиновой кислоты и низким содержанием линоленовой кислоты и которая также обеспечивает муку канолы с улучшенными питательными характеристиками (например, улучшенные компоненты семян). В некоторых вариантах осуществления растение, содержащее зародышевую плазму по изобретению, может неожиданно дополнительно обеспечивать эти признаки в комбинации с другими ценными признаками (например, но не ограничиваясь ими, превосходный выход, высокое содержание белка, высокое содержание масла и высокое качество масла). Семена с темной оболочкой в конкретных вариантах осуществления могут иметь значительно более тонкую оболочку семян, чем семена, продуцируемые стандартными сортами канолы с темными семенами. Более тонкая оболочка семян может обеспечивать уменьшенное содержание волокон в муке и увеличенное содержание в семенах масла и белка по сравнению с уровнями масла и белка в стандартном сорте с темными семенами. Темные семена, продуцируемые растениями, содержащими зародышевую плазму по изобретению, таким образом, могут иметь более высокое содержание масла и концентрацию белка в их семенах, чем наблюдается в семенах, продуцируемых стандартным растением канолы с темными семенами.
В вариантах осуществления растение, содержащее зародышевую плазму по изобретению, не проявляет существенных агрономических ограничений и/или ограничений на семена. Например, такое растение может проявлять агрономические качества и/или качества семян (например, прорастание; ранняя сезонная активность роста; эффект удобрения семян; сбор семян и пригодность к хранению), которые по меньшей мере настолько же ценны, как качества, проявляемые стандартными сортами канолы. В конкретных вариантах осуществления растение, содержащее зародышевую плазму по изобретению, также может содержать один или несколько дополнительных благоприятных признаков, проявляемых ранее существовавшей инбредной линией канолы, например, и не ограничиваясь этим, благоприятный профиль жирных кислот.
В некоторых вариантах осуществления растение, содержащее зародышевую плазму по изобретению, может продуцировать семена, имеющие по меньшей мере одну из нескольких характеристик питательности. В конкретных вариантах осуществления семена, продуцируемые таким растением канолы, могут содержать по меньшей мере одну характеристику питательности, выбранную из группы, состоящей из: благоприятного профиля масла; высокого содержания белка; низкого содержания волокон (например, ADF и NDF (включая низкое содержание полифенольных соединений)); (низкое содержание волокон и высокое содержание белка обеспечивают более высокую метаболизируемую энергию); высокого содержания фосфора; и низкого содержания сложного эфира синаповой кислоты (SAE). В определенных вариантах осуществления "высокое" или "низкое" содержание компонента относится к сравнению между семенами, продуцированными эталонным растением, содержащим зародышевую плазму по изобретению, и семенами, продуцированными стандартными сортами канолы. Таким образом, растение, продуцирующее семена с "низким" содержанием волокон, может продуцировать семена с более низким содержанием волокон, чем наблюдается в семенах, продуцируемых стандартными сортами канолы. Более того, растение, продуцирующее семена с "высоким" содержанием белка, может продуцировать семена с более высоким содержанием белка, чем наблюдается в семенах, продуцированных стандартными сортами канолы.
В некоторых вариантах осуществления может быть получена по существу единообразная коллекция рапсовых семян, продуцированных растением канолы, содержащим по меньшей мере одну характеристику питательности, выбранную из упомянутой выше группы. Такие семена можно использовать для получения по существу единообразного поля растений рапса. Конкретные варианты осуществления предусматривают семена канолы, содержащие идентифицирующие комбинации упомянутых выше характеристик. Например, комбинированное общее содержание масла и белка в семенах может быть пригодной мерой и уникальной характеристикой семян.
Некоторые варианты осуществления предусматривают канолу (например, канолу с темными семенами), содержащую зародышевую плазму по изобретению, которая способна обеспечивать масло канолы, имеющее профиль масла типа NATREON или профиль масла типа "Omega-9". Профиль масла "типа NATREON", "подобный NATREON" или "Omega-9" может обозначать содержание олеиновой кислоты в диапазоне, например, 68-80%; 70-78%; 71-77% и 72-75%, при содержании альфа-линоленовой кислоты ниже, например, 3%. В конкретных вариантах осуществления семена, продуцируемые растением канолы, содержащим зародышевую плазму по изобретению, могут обеспечивать масло, имеющее более 68%, более 70%, более 71%, более 71,5% и/или более 72% (например, 72,4% или 72,7%) олеиновой кислоты, при содержании линоленовой кислоты менее 3%, менее 2,4%, менее 2%, менее 1,9% и/или менее 1,8% (например, 1,7%). Однако в следующих вариантах осуществления канола, содержащая зародышевую плазму по изобретению, может обеспечивать масла, имеющие, например, содержание олеиновой кислоты, превышающее 80%. В определенных вариантах осуществления масло канолы, получаемое из канолы, содержащей зародышевую плазму по изобретению, может быть природным образом стабильным (например, его не гидрогенизируют искусственно). Содержание жирных кислот в масле канолы можно без труда и стандартным путем определять известными способами.
Таким образом, некоторые варианты осуществления предусматривают семена канолы (например, темные семена канолы), содержащие фракцию масла и фракцию муки, где фракция масла может иметь содержание α-линоленовой кислоты, например, 3% или менее (относительно общего содержания жирных кислот в семенах) и содержание олеиновой кислоты, например, 68% или более (относительно общего содержания жирных кислот в семенах). По определению содержание эруковой кислоты (C22:1) в таких семенах может составлять менее 2% по массе (по сравнению с общим содержанием жирных кислот в семенах). В конкретных примерах содержание масла в семенах канолы может составлять 48-50% от массы семян.
Термин "с высоким содержанием олеиновой кислоты" относится к Brassica juncea или другим видам Brassica, в зависимости от контекста, с содержанием олеиновой кислоты, превышающим ее содержание в сорте или линии дикого типа или другом эталонном сорте или линии, более часто он указывает на композицию жирных кислот, содержащую по меньшей мере 68,0% по массе олеиновой кислоты.
"Общее содержание насыщенных жирных кислот" относится к совокупному проценту пальмитиновой (C16:0), стеариновой (C18:0), арахиновой (C20:0), бегеновой (C22:0) и тетракозановой (C24:0) жирных кислот. Концентрации жирных кислот, рассмотренные в настоящем описании, определяют по стандартным методикам, хорошо известным специалистам в данной области. Конкретные методики описаны в примерах. Концентрации жирных кислот выражают в качестве процента по массе от общего содержания жирных кислот.
Термин "стабильность" или "стабильный", как используют в настоящем описании в отношении данного генетически контролируемого компонента в виде жирной кислоты, означает, что компонент в виде жирной кислоты сохраняется из поколения в поколение на протяжении по меньшей мере двух поколений и предпочтительно по меньшей мере трех поколений по существу на одном и том же уровне, например, предпочтительно ±5%. Способы по изобретению способны обеспечить получение линий Brassica juncea с улучшенным составом жирных кислот, стабильных на уровне вплоть до ±5% из поколения в поколение. Специалисту в данной области понятно, что на описанную выше стабильность может влиять температура, область, внешние воздействия и время посева. Таким образом, сравнение профилей жирных кислот между линиями канолы следует проводить с использованием семян, полученных в сходных условиях выращивания.
Когда термин "растение Brassica" используют в контексте настоящего изобретения, он также включает любые преобразования единичного гена в этой группе. Термин "растение с преобразованием единичного гена", как используют в рамках изобретения, относится к растениям Brassica, которые разработаны с помощью способа выведения растений, называемого обратным скрещиванием, где по существу все из желаемых морфологических и физиологических характеристик сорта восстанавливаются в дополнение к единичному гену, перенесенному в сорт способом обратного скрещивания. Способы обратного скрещивания можно использовать в рамках настоящего изобретения для улучшения или внесения характеристики в сорт. Термин "обратное скрещивание", как используют в рамках изобретения, относится к повторяющемуся скрещиванию гибридного потомства обратно с рекуррентным родительским растением, т.е. обратное скрещивание один или несколько раз с рекуррентным родительским растением (обозначаемое как "BC1", "BC2" и т.д.). Родительское растение Brassica, которое передает ген для желаемой характеристики, называют "нерекуррентным" или "донорским растением". Эта терминология связана с тем фактом, что нерекуррентное родительское растение используют один раз в протоколе обратного скрещивания и, таким образом, его не используют повторно. Родительское растение Brassica, в которое переносят ген или гены из нерекуррентного растения, известно как рекуррентное родительское растение, поскольку его используют в нескольких раундах в протоколе обратного скрещивания (Poehiman & Sleper, 1994; Fehr, 1987). В типичном протоколе обратного скрещивания исходный представляющий интерес сорт (рекуррентное родительское растение) скрещивают со вторым сортом (нерекуррентное родительское растение), который содержит один представляющий интерес ген, подлежащий переносу. Полученное потомство этого скрещивания затем снова скрещивают с рекуррентным родительским растением и этот процесс повторяют до тех пор, пока не получат растение Brassica, где по существу все из желаемых морфологических и физиологических характеристик рекуррентного родительского растения восстанавливаются в преобразованном растении, в дополнение к одному перенесенному гену из нерекуррентного родительского растения, как определяют при 5% уровне значимости при выращивании в тех же условиях окружающей среды. В настоящей заявке термин "Brassica" может включать любой или все из видов, относящихся к роду Brassica, включая Brassica napus, Brassica juncea, Brassica nigra, Brassica carinata, Brassica oleracea и Brassica rapa.
Канола Brassica juncea, как используют в настоящей заявке, относится к Brassica juncea, которая продуцирует семена с качеством масла и муки, которое удовлетворяет требованиям для коммерческого обозначения как масло или мука "канолы", соответственно (т.е. к растениям вида Brassica juncea, которые имеют менее 2% эруковой кислоты (Δ13-22:1) по массе в масле семян и менее 30 микромоль глюкозинолатов на грамм не содержащей масла муки).
В одном аспекте изобретение относится к растениям Brassica, таким как растения Brassica juncea, способным продуцировать семена, имеющие эндогенное содержание жирных кислот с высоким процентом олеиновой кислоты и низким процентом линоленовой кислоты по массе. В конкретных вариантах осуществления олеиновая кислота может содержать более чем приблизительно 68,0%, 69,0%, 70,0%, 71,0%, 72,0%, 73,0%, 74,0%, 75,0%, 76,0%, 77,0%, 78,0%, 79,0%, 80,0%, 81,0%, 82,0%, 83,0%, 84,0% или 85,0%, включая все целые числа и их дробные числа или любое целое число, имеющее величину более 85%, олеиновой кислоты. В конкретных вариантах осуществления содержание линоленовой кислоты в жирных кислотах может составлять менее чем приблизительно 5%, 4%, 3%, 2,5%, 2,0%, 1,5%, 1,0%, 0,5% или 0%, и включая все целые числа и их дробные числа. В одном иллюстративном варианте осуществления растение представляет собой Brassica juncea, семена которого имеют эндогенное содержание жирных кислот с по меньшей мере 68% олеиновой кислоты по массе и менее 3% линоленовой кислоты по массе. Во всех дополнительных вариантах осуществления растение представляет собой растение Brassica juncea, семена которого имеют эндогенное содержание жирных кислот с по меньшей мере 68,0% олеиновой кислоты по массе и не более чем приблизительно 5% линоленовой кислоты по массе.
В одном аспекте изобретение относится к растениям Brassica, таким как растения Brassica juncea, способным продуцировать семена, имеющие эндогенное содержание жирных кислот с высоким процентом олеиновой кислоты и низким процентом линоленовой кислоты по массе и низким общим содержанием насыщенных жирных кислот или высоким общим содержанием насыщенных жирных кислот, которое может составлять менее чем приблизительно 5,5% общих насыщенных жирных кислот или >10% общих насыщенных жирных кислот, соответственно.
Известно, что состав масла из семян Brassica juncea отличается от состава масла из Brassica napus как компонентом в виде жирных кислот (например, более высокое содержание эруковой кислоты), эфирных масел (например, аллилизотиоцианат), так и второстепенными компонентами (например, токоферолы, металлы, танины, фенольные соединения, фосфолипиды, цветные тельца и т.п.). Было выявлено, что масла семян (включая экстрагированные семена) из Brassica juncea имеют более высокую окислительную стабильность по сравнению с маслами из Brassica napus, даже несмотря на то, что масла из Brassica juncea, как правило, имеют более высокие уровни C18:3. (C. Wijesundera et al., "Canola Quality Indian Mustard oil (Brassica juncea) is More Stable to Oxidation than Conventional Canola oil (Brassica napus)", J. Am. Oil Chem. Soc. (2008) 85:693-699).
В альтернативном аспекте изобретение относится к способам увеличения содержания олеиновой кислоты и снижения содержания линоленовой кислоты в растениях Brassica. Такие способы могут вовлекать: (a) индукцию мутагенеза по меньшей мере в некоторых клетках линии Brassica, которая имеет содержание олеиновой кислоты более 55% и содержание линоленовой кислоты менее 14%; (b) регенерацию растений из по меньшей мере одной из указанных подвергнутых мутагенезу клеток и селекцию регенерировавших растений, которые имеют содержание жирных кислот, составляющее по меньшей мере 68% олеиновой кислоты (или альтернативную пороговую концентрацию олеиновой кислоты, как указано выше) и менее 3% линоленовой кислоты (или альтернативную пороговую концентрацию линоленовой кислоты, как указано выше); и (c) получение дальнейших поколений растений из указанных регенерировавших растений, где отдельные растения указанных дальнейших поколений растений имеют содержание жирных кислот, составляющее по меньшей мере 68% олеиновой кислоты (или альтернативную пороговую концентрацию) и менее 3% линоленовой кислоты (или альтернативную пороговую концентрацию). В некоторых вариантах осуществления Brassica может представлять собой Brassica juncea. Термин "высокое содержание олеиновой кислоты" и "низкое содержание линоленовой кислоты" охватывает полный диапазон возможных величин, описанных выше. В альтернативных вариантах осуществления способы по изобретению могут дополнительно включать отбор одной или нескольких линий, регенерировавших растений и дальнейших поколений растений для снижения содержания линоленовой кислоты, как, например, до диапазона возможных величин, описанного выше. В следующих вариантах осуществления стадия (c) может вовлекать отбор и выращивание семян из регенерировавших растений стадии (b). В следующих вариантах осуществления способы по изобретению могут включать повторение указанных стадий до достижения желаемого содержания олеиновой кислоты, содержания линоленовой кислоты или обеих из них.
В альтернативных вариантах осуществления предусмотрены способы скрининга отдельных семян в отношении увеличенного содержания олеиновой кислоты и сниженного содержания линоленовой кислоты, включающие: определение одного или нескольких из содержания олеиновой кислоты; или содержания линоленовой кислоты; или содержания олеиновой кислоты и содержания линоленовой кислоты среди жирных кислот части проростка семени; сравнение одного или нескольких содержаний с эталонной величиной; и установление вероятного относительного содержания олеиновой кислоты, линоленовой кислоты, или олеиновой и линоленовой кислот в семени. В конкретных вариантах осуществления часть растения, используемая для анализа, может представлять собой часть или весь лист, семядолю, стебель, черешок, цветоножку или любую другую ткань или фрагмент ткани, такой как ткани, имеющие состав, который проявляет достоверную корреляцию с составом семени. В ряде вариантов осуществления часть проростка может быть частью листа. В определенных вариантах осуществления стадия установления состава жирных кислот в семени может включать допущение, что значительно измененный уровень данной кислоты в указанном листе отражает сходное относительное изменение уровня этой кислоты в семени. В конкретном варианте осуществления настоящего изобретения предусмотрен способ скрининга растений Brassica в отношении отдельной линии растений, семена которой имеют эндогенное содержание жирных кислот, составляющее по меньшей мере 68% олеиновой кислоты и менее 3% линоленовой кислоты по массе, путем анализа ткани листа. Кроме того, ткань листа можно анализировать в отношении состава жирных кислот с помощью газожидкостной хроматографии, где экстракцию жирных кислот можно проводить способами, такими как совокупный анализ семян или анализ половин семян.
В альтернативных вариантах осуществления изобретение относится к растениям Brassica, которые могут представлять собой растения Brassica juncea, содержащим ранее описанные аллели генов из линий Brassica juncea. В определенных вариантах осуществления растение может быть гомозиготным по локусам fad2-a и fad3-a, соответствующим мутантным аллелям. В дополнительном варианте осуществления растение Brassica juncea, клетка растения или его часть содержат аллели генов, имеющие последовательности нуклеиновых кислот из ранее описанных последовательностей, описанных в настоящем описании.
В некоторых вариантах осуществления изобретение может вовлекать отличение HOLL сорта канолы Brassica juncea по настоящему изобретению (≥68% олеиновой кислоты и ≤5% линоленовой кислоты) от Brassica juncea с низким содержанием олеиновой кислоты/высоким содержанием линоленовой кислоты (~45% олеиновой кислоты и ~14% линоленовой кислоты) путем исследования присутствия или отсутствия гена BJfad2b (для справки см. публикацию патента США № 20030221217, Yao et al.). Это отличение может вовлекать подтверждение того, что ген BJfad2a является единственным функциональным геном десатуразы олеиновой жирной кислоты в линии сорта канолы Brassica juncea, как известно в данной области.
В одном варианте осуществления линия Brassica juncea содержит гены fad2 и fad3, как описано в международной публикации № US 2006/0248611 A1, которые проиллюстрированы на фиг.1 и 3 в ней. Гены fad2 и fad3 иллюстрируются в настоящем описании с помощью SEQ ID NO:1-4. Получаемые в результате аллели кодируют белки десатуразы дельта-12 жирных кислот, которые проиллюстрированы на фиг.2 международной публикации № US 2006/0248611 A1. В других вариантах осуществления линия Brassica juncea может содержать мутации в локусах генов fad2-a и fad3-a, и полученные мутантные аллели могут кодировать одну или несколько мутаций в последовательности предсказанных белков BJFAD2-a и BJFAD3-a. Репрезентативные примеры мутантных генов и белков fad2-a и fad3-a, пригодных для применения в рамках настоящего изобретения, также включают, но не ограничиваются ими, мутантные гены и белки fad2-a и fad3-a, описанные в: международной публикации № WO 2006/079567 A2 (например, фиг.1 и 2), как, например, SEQ ID NO:8 и 9; международной публикации № WO 2007/107590 A2, как, например, SEQ ID NO:10-21; патенте США № 6967243 B2 (например, фиг.2 и 3), как, например, SEQ ID NO:22-27; и европейской публикации № 1862551 A1 (например, фиг.1-10), как, например, SEQ ID NO:28-39.
В отдельных вариантах осуществления изобретение относится к выделенным последовательностям ДНК, содержащим полные открытые рамки считывания (ORF) и/или 5′-вышележащие области ранее описанных мутантных генов fad2 и fad3. Таким образом, изобретение также относится к полипептидным последовательностям предсказанных мутантных белков, содержащим мутации из ранее описанных мутантных аллелей. Известно, что мембраносвязанные десатуразы, такие как FAD2, имеют консервативные гистидиновые боксы. Изменения аминокислотных остатков вне этих гистидиновых боксов также могут влиять на активность фермента FAD2 (Tanhuanpaa et al., Molecular Breeding 4:543-550, 1998).
В одном аспекте изобретения мутантные аллели, описанные в настоящем описании, можно использовать для выведения растений. В частности, аллели по изобретению можно использовать для выведения видов Brassica с высоким содержанием олеиновой кислоты, таких как Brassica juncea, Brassica napus, Brassica rapa, Brassica nigra и Brassica carinata. Изобретение относится к молекулярным маркерам для отличения мутантных аллелей от альтернативных последовательностей. Таким образом, изобретение относится к способам сегрегационного или селекционного анализа генетических скрещиваний, вовлекающих растения, имеющие аллели по изобретению. Таким образом, изобретение относится к способам сегрегационного и селекционного анализа потомков, полученных в результате генетических скрещиваний, вовлекающих растения, имеющие аллели по изобретению.
В альтернативных вариантах осуществления изобретение относится к способам идентификации растений Brassica, таких как растения Brassica juncea, с желаемым составом жирных кислот или желаемыми геномными характеристиками. Способы по изобретению могут вовлекать, например, определение присутствия в геноме конкретных аллелей FAD2 и/или FAD3, таких как аллели по изобретению или аллель J96D-4830/BJfad2a дикого типа. В конкретных вариантах осуществления способы могут включать идентификацию присутствия полиморфизма нуклеиновой кислоты, ассоциированного с одним из идентифицированных аллелей, или антигенной детерминанты, ассоциированной с одним из аллелей по изобретению. Такое определение может быть достигнуто, например, с помощью ряда способов, таких как амплификация способом ПЦР соответствующего фрагмента ДНК, получение отпечатков ДНК, получение отпечатков РНК, блоттинг в геле и анализ RFLP, анализы защиты от нуклеаз, секвенирование соответствующего фрагмента нуклеиновой кислоты, получение антител (моноклональных или поликлональных), или альтернативных способов, адаптированных для отличения белка, продуцированного соответствующими аллелями, от других вариантов и форм дикого типа этого белка. Это изобретение также относится к способу идентификации растений B. juncea, семена которых имеют эндогенное содержание жирных кислот, составляющее по меньшей мере 68% олеиновой кислоты по массе, путем определения присутствия мутантных аллелей по изобретению.
В альтернативных вариантах осуществления изобретение относится к растениям Brassica, содержащим кодирующие последовательности fad2 и fad3, которые кодируют мутантные белки FAD2 и FAD3. Такие мутантные белки FAD2/FAD3 могут содержать только одно аминокислотное изменение по сравнению с белком FAD2 дикого типа. В репрезентативных вариантах осуществления различные линии Brassica juncea содержат ранее описанные мутантные белки FAD2, кодируемые ранее описанными аллелями. Такие аллели могут быть выбраны так, чтобы они были эффективными в отношении обеспечения увеличенного содержания олеиновой кислоты и уменьшенного содержания линоленовой кислоты в растениях по изобретению. В конкретных вариантах осуществления желаемый аллель можно вводить в растения способами скрещивания. В альтернативных вариантах осуществления аллели по изобретению можно вводить молекулярно-биологическими способами, включая трансформацию растений. В таких вариантах осуществления растения по изобретению могут продуцировать семена, имеющие содержание жирных кислот, составляющее: по меньшей мере приблизительно 68% олеиновой кислоты по массе и менее чем приблизительно 3% линоленовой кислоты по массе, или любое другое пороговое содержание олеиновой кислоты и линоленовой кислоты, как указано выше. Растения по изобретению также могут содержать от приблизительно 68% до приблизительно 85% по массе олеиновой кислоты, от приблизительно 70% до приблизительно 78% олеиновой кислоты и от приблизительно 0,1% до приблизительно 3% линоленовой кислоты, где состав масла генетически происходит из родительской линии. Растения по изобретению также могут иметь общее содержание жирных кислот от менее 7,1% до менее 6,2% по массе. В одном варианте осуществления растение продуцирует семена, имеющие эндогенное содержание жирных кислот, составляющее по меньшей мере приблизительно 68% олеиновой кислоты и менее 3% линоленовой кислоты, где состав масла генетически происходит из родительской линии.
В отдельных вариантах осуществления изобретение относится к семенам Brassica, которые могут представлять собой семена Brassica juncea, имеющие эндогенное содержание масла с составом жирных кислот, указанным для одного или нескольких из указанных выше вариантов осуществления, и где генетические детерминанты эндогенного содержания масла происходят из мутантных аллелей по изобретению. Такие семена можно получать, например, путем самоопыления каждой из линий с мутантным аллелем по изобретению. Альтернативно такие семена можно получать, например, путем скрещивания линий с мутантным аллелем со вторым родительским растением с последующей селекцией, где второе родительское растение может представлять собой любые другие линии Brassica, такие как линия Brassica juncea, представляющая собой сорт канолы Brassica juncea или не сорт канолы Brassica juncea, или любой другой вид Brassica, такой как Brassica napus, Brassica rapa, Brassica nigra и Brassica carinata. Эти способы выведения хорошо известны специалистам в данной области.
В альтернативных вариантах осуществления изобретение относится к генетически стабильным растениям рода Brassica, таким как растения Brassica juncea, в которых развиваются зрелые семена, имеющие состав, описанный в одном или нескольких из указанных выше вариантов осуществления. Такие растения могут происходить из линий Brassica juncea, имеющих мутантные аллели по изобретению. Состав масла таких растений может генетически происходить из родительских линий.
В альтернативных вариантах осуществления изобретение относится к способам получения генетически стабильного растения Brassica, такого как растение Brassica juncea, которое продуцирует зрелые семена, имеющие эндогенное содержание жирных кислот, обеспечивающее состав, указанный для одного или нескольких из указанных выше вариантов осуществления. Способы по изобретению могут вовлекать стадии: скрещивания растения с генами Omega-9 (например, fad2a и fad3a) Brassica napus с другими растениями Brassica, такими как Brassica juncea, с получением потомков F1. Размножение потомков F1 можно осуществлять, например, способами, которые могут включать самоопыление или развитие двойных гаплоидных растений. Путем комбинирования мутантных аллелей FAD2 и мутантных аллелей FAD3 растения, имеющие двойные мутантные аллели генов (fad2 и fad3), могут иметь улучшенный профиль жирных кислот в масле относительно растений с одной мутацией. Полученные потомки можно подвергать селекции для получения генетически стабильных растений, которые продуцируют семена, имеющие состав, описанный для одного или нескольких из указанных выше вариантов осуществления. Такие семена могут иметь, например, стабилизированный профиль жирных кислот, который включает общее содержание насыщенных жирных кислот от приблизительно 7,1% до приблизительно 6,5% в общих экстрагируемых маслах. В определенных вариантах потомки сами по себе могут продуцировать семена или масло, которые имеют состав, как указано выше для альтернативных вариантов осуществления. Они имеют содержание олеиновой кислоты более чем приблизительно 68% по массе и содержание линоленовой кислоты менее чем приблизительно 3% по массе.
В одном аспекте изобретение относится к растениям, имеющим стабильный наследуемый фенотип с высоким содержанием олеиновой кислоты и низким содержанием линоленовой кислоты. Например, фенотип с высоким содержанием олеиновой кислоты и низким содержанием линоленовой кислоты, являющийся результатом мутантных аллелей по изобретению, является генетически наследуемым через поколения M2, M3 и M4.
В альтернативных вариантах осуществления изобретение относится к растениям Brassica juncea, где активность десатуразы жирных кислот изменена, изменено содержание олеиновой кислоты или изменено содержание линоленовой кислоты относительно B. juncea дикого типа, которые использовали в эксперименте по мутагенезу. Под десатуразой жирных кислот ("FAD") подразумевают, что белок проявляет активность внесения двойной связи в биосинтезе жирных кислот. Например, ферменты FAD2/FAD3 могут быть охарактеризованы по активности внесения второй двойной связи в биосинтез линоленовой кислоты из олеиновой кислоты. Измененная активность десатуразы может включать увеличение, снижение или устранение активности десатуразы по сравнению с эталонным растением, клеткой или образцом.
В других аспектах снижение активности десатуразы может включать устранение экспрессии последовательности нуклеиновой кислоты, которая кодирует десатуразу, такой как последовательность нуклеиновой кислоты по изобретению. Под устранением экспрессии в рамках настоящего изобретения подразумевают, что функциональная аминокислотная последовательность, кодируемая последовательностью нуклеиновой кислоты, не продуцируется на поддающемся выявлению уровне. Снижение активности десатуразы может включать устранение транскрипции последовательности нуклеиновой кислоты, которая кодирует десатуразу, такой как последовательность по изобретению, кодирующая фермент FAD2 или фермент FAD3. Под устранением транскрипции в рамках настоящего изобретения подразумевают, что последовательность мРНК, кодируемая последовательностью нуклеиновой кислоты, не транскрибируется на поддающихся выявлению уровнях. Снижение активности десатуразы также может включать продуцирование укороченной аминокислотной последовательности с последовательности нуклеиновой кислоты, которая кодирует десатуразу. Под продуцированием укороченной аминокислотной последовательности в рамках настоящего изобретения подразумевают, что в аминокислотной последовательности, кодируемой последовательностью нуклеиновой кислоты, отсутствует одна или несколько аминокислот функциональной аминокислотной последовательности, кодируемой последовательностью нуклеиновой кислоты дикого типа. Кроме того, снижение активности десатуразы может включать продуцирование варианта аминокислотной последовательности десатуразы. Под продуцированием варианта аминокислотной последовательности в рамках настоящего изобретения подразумевают, что аминокислотная последовательность имеет одну или несколько аминокислот, которые отличаются от аминокислотной последовательности, кодируемой последовательностью нуклеиновой кислоты дикого типа. Как более подробно рассмотрено в настоящем описании, настоящее изобретение относится к тому, что мутантные линии по изобретению продуцируют ферменты FAD2 и FAD3 с вариантами аминокислот по сравнению с линией дикого типа J96D-4830. Различные типы мутаций можно вносить в последовательность нуклеиновой кислоты для цели снижения активности десатуразы, такие как мутации со сдвигом рамки считывания, замены и делеции.
В некоторых вариантах осуществления изобретение относится к новым полипептидным последовательностям FAD2/FAD3, которые могут быть модифицированы в соответствии с альтернативными вариантами осуществления изобретения. В данной области хорошо известно, что некоторые модификации и изменения можно вносить в структуру полипептида без существенного изменения биологической функции этого пептида, с получением биологически эквивалентного полипептида. Как используют в рамках изобретения, термин "консервативные аминокислотные замены" относится к замене одной аминокислоты другой аминокислотой в данном участке пептида, где замену можно вносить без какой-либо ощутимой утраты или приобретения функции, с получением биологически эквивалентного полипептида. При получении таких изменений замены подобных аминокислотных остатков можно вносить, исходя из относительного сходства заместителей боковой цепи, например, их размера, заряда, гидрофобности, гидрофильности и т.п., и такие замены можно анализировать в отношении их эффекта на функцию пептида путем стандартного тестирования. Напротив, как используют в рамках изобретения, термин "неконсервативные аминокислотные замены" относится к замене одной аминокислоты другой аминокислотой в данном участке пептида, где замена вызывает ощутимую утрату или приобретение функции пептидом, с получением полипептида, который не является биологически эквивалентным.
Волокна являются компонентом стенки растений, и они включают углеводные полимеры (например, целлюлозу (линейные полимерные цепи глюкозы)); гемицеллюлозу (разветвленные цепи гетерополимеров, например, галактозы, ксилозы, арабинозы, рамнозы, с присоединенными молекулами фенола); и пектины (растворимые в воде полимеры галактуроновой кислоты, ксилозы, арабинозы, с различными степенями метилирования). Волокна также включают полифенольные полимеры (например, лигнин-подобные полимеры и конденсированные танины). В теории волокно ADF состоит из целлюлозы и лигнина. Как правило, фракция ADF включает конденсированные танины, однако содержание конденсированных танинов варьирует независимо от ADF. Напротив, TDF представляет собой муку, из которой удалены белок, растворимые соединения и крахмал, и оно состоит из нерастворимых компонентов клеточной стенки (например, целлюлоза, гемицеллюлоза, полифенольные соединения и лигнин).
В конкретных вариантах осуществления семена растения канолы (например, растение канолы с темными семенами), содержащие зародышевую плазму по изобретению, могут иметь сниженное содержание ADF по сравнению с некоторым сортом канолы. В конкретных примерах содержание волокон в муке канолы (из цельных семян, после удаления масла, в расчете на массу сухого вещества) может составлять, например, и не ограничиваясь этим: менее чем приблизительно 18% ADF (например, приблизительно 18% ADF, приблизительно 17% ADF, приблизительно 16% ADF, приблизительно 15% ADF, приблизительно 14% ADF, приблизительно 13% ADF, приблизительно 12% ADF, приблизительно 11% ADF и приблизительно 10% ADF) и/или менее чем приблизительно 22% NDF (например, приблизительно 22,0% NDF, приблизительно 21% NDF, приблизительно 20% NDF, приблизительно 19% NDF, приблизительно 18% NDF и приблизительно 17% NDF).
В конкретных вариантах осуществления семена растения канолы, содержащего зародышевую плазму по изобретению, могут иметь увеличенное содержание белка по сравнению со стандартным сортом канолы с темными семенами. В конкретных примерах содержание белка в муке канолы (из цельных семян, после удаления масла, в расчете на массу сухого вещества) может составлять, например, и не ограничиваясь этим, более чем приблизительно 45% (например, приблизительно 45%, приблизительно 46%, приблизительно 47%, приблизительно 48%, приблизительно 49%, приблизительно 50%, приблизительно 51%, приблизительно 52%, приблизительно 53%, приблизительно 54%, приблизительно 55%, приблизительно 56%, приблизительно 57% и приблизительно 58%) неочищенного белка. Различные сорта канолы характеризуются конкретным содержанием белка. Содержание белка (% азот × 6,25) может быть определено с использованием различных хорошо известных и общепринятых способов анализа, например, NIR и Kjeldahl.
В некоторых вариантах осуществления содержание фосфора также можно использовать для определения семян, растений и линий сортов канолы. Такие сорта канолы могут продуцировать муку канолы (из цельных семян, после удаления масла, в расчете на массу сухого вещества), которая имеет увеличенное содержание фосфора по сравнению с мукой из стандартных сортов канолы. Например, мука канолы по изобретению может иметь содержание фосфора более 1,2%, более 1,3%, более 1,4%, более 1,5%, более 1,6%, более 1,7% и/или более 1,8%.
Различные комбинации вышеупомянутых признаков также могут быть идентифицированы в инбредных линиях канолы и гибридах, предоставленных в нескольких примерах, и они могут быть проиллюстрированы ими. Эти линии иллюстрируют, что зародышевую плазму по изобретению можно использовать для обеспечения и получения различных новых комбинаций широкого множества преимущественных характеристик и/или признаков канолы. Например, инбредную линию канолы, содержащую зародышевую плазму по изобретению, можно скрещивать с другой линией канолы, которая содержит желаемую характеристику и/или признак, для внесения желаемой характеристики состава семян в инбредную линию канолы, содержащую зародышевую плазму по изобретению. Вычисление состава семян (например, содержание волокон, содержание глюкозинолата, содержание масла и т.д.) и других признаков растений можно осуществлять с использованием способов, которые известны в данной области и приняты в промышленности. Путем выбора и размножения потомков растений от скрещивания, которые включают желаемые характеристики и/или признаки родительских сортов, можно создавать новые сорта, которые содержат желаемую комбинацию характеристик и/или признаков.
V. Мука канолы, имеющая улучшенные характеристики питательности
Некоторые варианты осуществления предусматривают муку, содержащую семена канолы, где семена канолы имеют характеристики масла и муки, рассмотренные выше. Например, некоторые варианты осуществления включают экстрагированную гексаном высушенную воздухом муку канолы (белые хлопья или WF), содержащую новую комбинацию характеристик (например, состав семян), как рассмотрено выше. Конкретные варианты осуществления включают муку, содержащую семена канолы, продуцированные растением, содержащим зародышевую плазму по изобретению, и муку, содержащую семена потомков растения, содержащего зародышевую плазму по изобретению.
Инбредные линии и гибриды канолы, содержащие зародышевую плазму по изобретению, в некоторых вариантах осуществления могут обеспечивать свойства улучшенной питательности муки при использовании непосредственно в качестве ингредиента корма или пищи и/или при использовании в качестве сырьевого материала для обработки изолятов и концентратов белка. Например, такие инбредные линии и гибриды канолы могут обеспечивать характеристики корма для животных, превосходящие характеристики стандартной муки канолы. В некоторых вариантах осуществления компоненты муки канолы (и животных кормов, содержащих ее) можно использовать для предоставления хорошего питания для моногастрального животного (например, свиней и птиц).
В некоторых вариантах осуществления компоненты муки канолы (и животных кормов, содержащих ее) можно, кроме того, использовать для обеспечения хорошего питания жвачным животным (например, животным семейства бычьих, овцам, козам и другим животным подотряда Ruminantia). Кормление жвачных животных представляет собой особые проблемы и особые возможности. Особые возможности являются результатом способности жвачных животных усваивать нерастворимое целлюлозное волокно, которое может разрушаться определенными микроорганизмами в рубце этих животных, но, как правило, не усваивается моногастральными млекопитающими, такими как свиньи. Особые проблемы являются результатом тенденции к ингибированию определенными кормами усвоения волокна в рубце и тенденции к ограничению усвоения в рубце некоторых из компонентов определенных кормов, таких как жиры и белок.
Семена Brassica с экстрагированным маслом являются потенциальным источником белка высокого качества для применения в кормах для животных. После экстракции масла продовольственная мука канолы содержит приблизительно 37% белка по сравнению с приблизительно 44-48% в соевой муке, которая в настоящее время является широко предпочтительной для кормов и пищи. Белки, содержащиеся в каноле, богаты метионином и содержат достаточные количества лизина, оба из которых являются ограниченными аминокислотами в большинстве белков злаковых и масличных растений. Однако применение муки канолы в качестве источника белка является в некоторой степени ограниченным в определенных кормах для животных, поскольку она содержит нежелательные компоненты, такие как волокна, глюкозинолаты и фенольные соединения.
Одним аспектом питательности семян рапса, из которого произошла канола, является их высокий уровень (30-55 мкмоль/г) глюкозинолатов, соединений на основе серы. Когда листья или семена канолы дробят, образуются сложные эфиры изотиоцианата под действием мирозиназы на глюкозинолаты. Эти продукты ингибируют синтез тироксина щитовидной железой и имеют другие антиметаболические эффекты. Paul et al. (1986) Theor. Appl. Genet. 72:706-9. Таким образом, для применения в пище у человека, содержание глюкозинолатов, например в белках, происходящих из муки семян рапса, должно быть снижено или устранено для обеспечения безопасности продукта.
Улучшенные семена канолы, например, с благоприятным профилем и составом масла и низким содержанием глюкозинолатов в семенах, значительно снизили бы потребность в гидрогенизации. Например, более высокое содержание олеиновой кислоты и более низкое содержание α-линоленовой кислоты в таком масле может обеспечить увеличенную окислительную стабильность, тем самым снижая необходимость в гидрогенизации и продуцировании транс-жирных кислот. Снижение уровня глюкозинолатов в семенах значительно снизило бы остаточное содержание серы в масле. Сера портит никелевый катализатор, обычно используемый для гидрогенизации. Koseoglu et al., Chapter 8, Canola and Rapeseed: Production, Chemistry, Nutrition, and Processing Technology, Ed. Shahidi, Van Nostrand Reinhold, N.Y., 1990, pp. 123-48. Кроме того, гидрогенизация масла из сорта канолы с низким содержанием глюкозинолатов в семенах была бы менее дорогостоящей.
Фенольные соединения в муке канолы придают горький привкус, и полагают, что они обязательно ассоциированы с темным цветом конечных белковых продуктов. Шелуха семян, которая присутствует в больших количествах в стандартной муке канолы, не усваивается человеком и другими моногастральными животными и также обеспечивает неприглядный гетерогенный продукт.
Состав муки в семенах, продуцированных растением канолы, содержащим зародышевую плазму по изобретению, может иметь, например, и не ограничиваясь этим: высокий уровень белка; низкий уровень волокон; высокое содержание фосфора; и/или низкое содержание SAE. Нерастворимые волокна и полифенольные соединения являются антипитательными и нарушают усвоение белка и аминокислот. Таким образом, мука канолы и корма для животных, содержащие муку канолы, имеющую по меньшей мере одну характеристику состава семян, выбранную из группы, состоящей из сниженного содержания волокон, увеличенного содержания белка, сниженного содержания полифенольных соединений и увеличенного содержания фосфора, могут быть желательными в некоторых применениях.
В конкретных примерах мука канолы (не содержащая масла, в расчете на массу сухого вещества) может иметь содержание белка по меньшей мере приблизительно 45% (например, приблизительно 45%, приблизительно 46%, приблизительно 47%, приблизительно 48%, приблизительно 49%, приблизительно 50%, приблизительно 51%, приблизительно 52%, приблизительно 53%, приблизительно 54%, приблизительно 55%, приблизительно 56%, приблизительно 57% и приблизительно 58%).
Сорта канолы, содержащие зародышевую плазму по изобретению, могут иметь хороший выход и продуцировать семена, имеющие значительно более низкое содержание кислотных детергентных волокон (ADF), по сравнению с эталонной линией канолы. В некоторых вариантах осуществления можно использовать любые эмпирические величины, определенные для компонента семян, продуцируемого сортом растений, содержащим зародышевую плазму по изобретению, для определения растений, семян и масла сорта растений. В некоторых таких примерах можно использовать конкретные числа в качестве конечных результатов для определения диапазонов выше, ниже или между любыми из заданных величин. Иллюстративные диапазоны для характеристик масла и других компонентов семян указаны выше. Линии и семена их растений также можно определять с помощью комбинаций таких диапазонов. Например, характеристики масла, рассмотренные выше вместе с характеристикам уровней волокон, уровней полифенольных соединений, уровней глюкозинолата, уровней белка и уровней фосфора, например, можно использовать для определения конкретных линий и их семян.
Не все из упомянутых выше характеристик (например, характеристики состава семян) необходимы для определения линий и семян согласно некоторым вариантам осуществления, однако можно использовать дополнительные характеристики для определения таких линий и семян (например, и не ограничиваясь этим, метаболизируемая энергия, усвояемая энергия, биологическая энергия и суммарная энергия).
VI. Растения, содержащие зародышевую плазму, обеспечивающую желаемые признаки состава семян независимо от цвета семян
Желаемые признаки конкретных инбредных линий и гибридов канолы, содержащих зародышевую плазму по изобретению, можно переносить в другие типы Brassica (путем общепринятого выведения и т.п.), например, B. rapa и B. juncea, где полученные растения продуцируют семена с желаемыми характеристиками (например, характеристики состава семян), экспрессируемыми независимо от цвета семян. Таким образом, сорт Brassica, в который был перенесен один или несколько желаемых признаков конкретной инбредной линии или гибрида канолы, содержащих зародышевую плазму по изобретению, может продуцировать семена с желаемыми характеристиками, которые являются либо желтыми, либо темными. Мука и семена таких новых или модифицированных сортов Brassica могут иметь сниженный уровень волокон семян, увеличенный уровень белка и увеличенный уровень фосфора и/или сниженный уровень полифенольных соединений.
Некоторые варианты осуществления включают не только желтые и темные семена канолы, содержащие зародышевую плазму, как описано и проиллюстрировано в настоящем описании, но также растения, выращенные или иным образом продуцированные из таких семян, и тканевые культуры регенерирующих клеток рассматриваемых растений канолы. Проиллюстрированные линии и гибриды получали без генетической инженерии и без мутагенеза, тем самым демонстрируя применимость зародышевой плазмы для продуцирования новых и модифицированных сортов канолы.
В некоторых конкретных вариантах осуществления предусмотрены конкретные иллюстративные инбредные линии и гибриды канолы. В качестве части настоящего описания по меньшей мере 2500 семян каждого из CL065620, CL044864, CL121460H, CL166102H и CL121466H были депонированы и стали доступными обществу, они являются объектом патентных прав, но в остальном без ограничений (за исключением ограничений, четко описанных в 37 C.F.R. § 1,808(b)), в American Type Culture Collection (ATCC), Rockville, Md. 20852. Депозиты были обозначены как № депозитов ATCC PTA-11697, PTA-11696, PTA-11698, PTA-_____ и PTA-11699_, соответственно, с датой депонирования 22 февраля 2011 года для образцов с PTA11696 по PTA11699 и 21 февраля 2012 года для PTA_____. Депозиты будут храниться, как указано выше, в депозитарии ATCC, который является общественным депозитарием, в течение 30 лет, или пяти лет после последнего запроса, или в течение срока действия патента, в зависимости от того, что является более длительным, и депозит будет заменен, если он станет нежизнеспособным в ходе этого периода.
Некоторые варианты осуществления включают семена любого из сортов Brassica napus, описанных в настоящем описании. Некоторые варианты осуществления также включают растения Brassica napus, получаемые из таких семян, а также тканевые культуры регенерирующих клеток таких растений. Также включается растение Brassica napus, регенерировавшее из такой тканевой культуры. В конкретных вариантах осуществления такое растение может быть способно экспрессировать все морфологические и физиологические свойства проиллюстрированного сорта. Растения Brassica napus согласно конкретным вариантам осуществления могут иметь идентифицирующие физиологические и/или морфологические характеристики растения, выращенного из депонированных семян.
Также предусмотрены способы проведения скрещиваний с использованием зародышевой плазмы по изобретению (например, встречающейся в иллюстративных инбредных линиях и гибридах канолы, представленных в настоящем описании) в по меньшей мере одном родительском растении потомства описанных выше семян. Например, некоторые варианты осуществления включают гибридное растение F1 B. napus в качестве одного или обоих родителей любого из растений, проиллюстрированных в настоящем описании. Следующие варианты осуществления включают семена B. napus, продуцированные таким гибридом F1. В конкретных вариантах осуществления способ продуцирования семян гибрида F1 B. napus включает скрещивание проиллюстрированного растения с отличающимся инбредным родительским растением канолы и сбор полученных гибридных семян. Растение канолы по изобретению (например, родительское растение канолы и растение канолы, продуцированное таким способом для получения гибрида F1) может быть либо женским, либо мужским растением.
Характеристики растений канолы в некоторых вариантах осуществления (например, уровни и/или профили масла и белка) можно далее модифицировать и/или улучшать путем скрещивания растения по изобретению с другой линией, имеющей модифицированную характеристику (например, высокие уровни масла и белка). Аналогично, другие характеристики могут быть улучшены путем тщательного изучения родительского растения. Линии канолы, содержащие зародышевую плазму по изобретению, могут быть благоприятными для внесения путем скрещивания их желаемых характеристик состава семян в другие линии рапса или канолы независимо от цвета семян. Зародышевые плазмы по изобретению позволяют перенос этих признаков в другие растения того же вида с помощью общепринятых способов выведения растений, включая перекрестное опыление и селекцию потомков. В некоторых вариантах осуществления желаемые признаки можно переносить между видами с использованием общепринятых способов выведения растений, вовлекающих перенос пыльцы и селекцию. См., например, Brassica crops and wild allies biology and breeding, Eds. Tsunada et al., Japan Scientific Press, Tokyo (1980); Physiological Potentials for Yield Improvement of Annual Oil and Protein Crops, Eds. Diepenbrock and Becker, Blackwell Wissenschafts-Verlag Berlin, Vienna (1995); Canola and Rapeseed, Ed. Shahidi, Van Nostrand Reinhold, N.Y. (1990); и Breeding Oilseed Brassicas, Eds. Labana et al., Narosa Publishing House, New Dehli (1993).
В некоторых вариантах осуществления способ переноса по меньшей мере одной желаемой характеристики состава семян независимо от цвета семян включает после внутривидового скрещивания самоопыление представителей поколения F1 с получением семян F2. Затем можно проводить обратное скрещивание для получения линий, проявляющих желаемую характеристику(и) состава семян. Кроме того, для переноса признака из одного вида в другой можно использовать слияние протопластов и способы ядерной трансплантации. См., например, Ruesink, "Fusion of Higher Plant Protoplasts", Methods in Enzymology, Vol. LVIII, Eds. Jakoby and Pastan, Academic Press, Inc., New York, N.Y. (1979), и ссылки, цитированные в ней; и Carlson et al. (1972) Proc. Natl. Acad Sci. USA 69:2292.
После получения и продуцирования иллюстративных линий канолы, содержащих зародышевую плазму по изобретению, темный цвет оболочки семян можно без труда переносить с желаемыми характеристиками состава семян в другие виды Brassica общепринятыми способами выведения растений, как указано выше. Например, темный цвет оболочки семян можно без труда переносить с желаемыми характеристиками состава семян в коммерчески доступные сорта B. rapa, например, и не ограничиваясь ими, Tobin, Horizon и Colt. Понятно, что темный цвет семян не должен переноситься вместе с другими характеристиками семян.
При наличии одного из иллюстративных сортов в качестве исходной точки специалист в данной области может манипулировать конкретными преимуществами, обеспечиваемыми сортом, без отклонения от объема настоящего изобретения. Например, профиль масла семян в иллюстративном сорте можно переносить в другой агрономически желаемый сорт B. napus общепринятыми способами выведения растений, вовлекающими перекрестное опыление и селекцию потомков, например, где зародышевая плазма иллюстративного сорта включена в другой агрономически желаемый сорт.
Конкретные варианты осуществления могут включать иллюстративные сорта B. napus, а также по существу производные сорта, которые по существу происходят из по меньшей мере одного из проиллюстрированных сортов. Кроме того, варианты осуществления изобретения могут включать растение по меньшей мере одного из проиллюстрированных сортов, растение такого по существу производного сорта и/или растение рапса, регенерировавшее из растений или ткани (включая пыльцу, семена и клетки), продуцированных из них.
Можно выбирать материалы растений, которые способны к регенерации, например, семена, микроспоры, семяпочки, пыльцу, вегетативные части и микроспоры. Как правило, такие клетки растений можно выбирать из любого сорта Brassica, включая сорта, имеющие желаемые агрономические признаки.
Способы регенерации известны в данной области. Можно первоначально выбирать клетки, способные к регенерации (например, семена, микроспоры, семяпочки, пыльцу и вегетативные части), из выбранного растения или сорта. Эти клетки необязательно можно подвергать мутагенезу. Из клеток можно получать растение с использованием способов регенерации, оплодотворения и/или выращивания, исходя из типа клеток (и того, подвергнуты ли они мутагенезу). Манипуляции растениями или семенами, или их частями, могут приводить к созданию по существу производных сортов.
В некоторых вариантах осуществления желаемые характеристики состава семян, проявляемые растениями, содержащими зародышевую плазму по изобретению, можно вносить в растение, имеющее множество дополнительных желаемых признаков независимо от цвета семян, для получения растения как с желаемыми характеристиками состава семян, так и с множеством желаемых признаков. Способ внесения желаемых характеристик состава семян в растение, содержащее один или несколько желаемых признаков, независимо от цвета семян, называют "объединением" этих признаков. В некоторых примерах объединение желаемых характеристик состава семян с множеством желаемых признаков может приводить к дальнейшему улучшению характеристик состава семян. В некоторых примерах объединение желаемых характеристик состава семян с множеством желаемых признаков может приводить к растению канолы, имеющему желаемые характеристики состава семян в дополнение к одному или нескольким (например, всем) из множества желаемых признаков.
Примеры признаков, которые могут быть желательными для комбинирования с желаемыми характеристиками состава семян, включают, например, и не ограничиваясь этим: гены устойчивости растения к заболеваниям (см., например, Jones et al. (1994) Science 266:789 (ген Cf-9 томата для устойчивости к Cladosporium fulvum); Martin et al. (1993) Science 262:1432 (ген Pto томата для устойчивости к Pseudomonas syringae); и Mindrinos et al. (1994) Cell 78:1089 (ген RSP2 для устойчивости к Pseudomonas syringae)); ген, придающий устойчивость к паразитам; белок Bacillus thuringiensis, его производное или синтетический полипептид, смоделированный на его основе (см., например, Geiser et al. (1986) Gene 48:109 (ген Bt δ-эндотоксина; молекулы ДНК, кодирующие гены δ-эндотоксина, могут быть приобретены в American Type Culture Collection (Manassas, VA), например, под номерами доступа ATCC 40098; 67136; 31995 и 31998)); лектин (см., например, Van Damme et al. (1994) Plant Molec. Biol. 24:25 (гены связывающего маннозу лектина Clivia miniata)); связывающий витамин белок, например, авидин (см. международную публикацию PCT US93/06487 (применение авидина и гомологов авидина в качестве лаврицидов против паразитов насекомых)); ингибитор фермента; протеазу или ингибитор протеазы (см., например, Abe et al. (1987) J. Biol. Chem. 262:16793 (ингибитор цистеиновой протеазы риса); Huub et al. (1993) Plant Molec. Biol. 21:985 (ингибитор протеазы I табака); и патент США № 5494813); ингибитор амилазы (см. Sumitani et al. (1993) Biosci. Biotech. Biochem. 57:1243 (ингибитор альфа-амилазы Streptomyces nitrosporeus)); специфический для насекомых гормон или феромон, например, экдистероидный гормон или ювенильный гормон, их вариант, миметик на их основе, или их антагонист или агонист (см., например, Hammock et al. (1990) Nature 344:458 (инактиватор ювенильного гормона)); специфический для насекомых пептид или нейропептид, который нарушает физиологию паразитов (см., например, Regan (1994) J. Biol. Chem. 269:9 (рецептор диуретического гормона насекомых); Pratt et al. (1989) Biochem. Biophys. Res. Comm. 163:1243 (аллостатин из Diploptera puntata); патент США № 5266317 (специфические для насекомых паралитические нейротоксины)); специфический для насекомых яд, продуцируемый в природе змеей, пчелой или другим организмом (см., например, Pang et al. (1992) Gene 116:165 (инсектотоксический пептид скорпиона)); фермент, ответственный за избыточное накопление монотерпенов, сесквитерпенов, стероидов, гидроксамовой кислоты, фенилпропаноидного производного или другой небелковой молекулы с инсектицидной активностью; фермент, вовлеченный в модификацию, включая посттрансляционную модификацию биологически активной молекулы, например, гликолитический фермент; протеолитический фермент; липолитический фермент; нуклеазу; циклазу; трансаминазу; эстеразу; гидролазу; фосфатазу; киназу; фосфорилазу; полимеразу; эластазу; хитиназу; или глюканазу, как природные, так и синтетические (см. международную публикацию PCT WO 93/02197 (ген каллазы); молекулы ДНК, которые содержат последовательности, кодирующие хитиназу (например, от ATCC, номера доступа 39637 и 67152); Kramer et al. (1993) Insect Biochem. Molec. Biol. 23:691 (хитиназа табачного бражника); и Kawalleck et al. (1993) Plant Molec. Biol. 21:673 (ген полиубиквитина ubi4-2 петрушки); молекулу, которая стимулирует передачу сигнала (см., например, Botella et al. (1994) Plant Molec. Biol. 24:757 (кальмодулин); и Griess et al. (1994) Plant Physiol. 104:1467 (кальмодулин кукурузы); пептид с гидрофобным моментом (см., например, международную публикацию PCT WO 95/16776 (пептидные производные тахиплесина, который ингибирует грибковые патогены растений); и международную публикацию PCT WO 95/18855 (синтетические противомикробные пептиды, которые обеспечивают устойчивость к заболеванию)); мембранную пермеазу, формирователь канала или блокатор канала (см., например, Jaynes et al. (1993) Plant Sci 89:43 (аналог литического пептида цекропина-β, чтобы придать трансгенным растениям устойчивость к Pseudomonas solanacearum); вирусный инвазивный белок или комплексный токсин, происходящий из него (см., например, Beachy et al. (1990) Ann. rev. Phytopathol. 28:451 (опосредуемая белком оболочки устойчивость к вирусу мозаики люцерны, вирусу мозаики огурца, вирусу полосатости табака, вирусу X картофеля, вирусу Y картофеля, вирусу гравировки табака, вирусу погремковости табака и вирусу табачной мозаики)); специфичное к насекомым антитело или иммунотоксин, происходящий из него (см., например, Taylor et al., Abstract #497, Seventh Int'l Symposium on Molecular Plant-Microbe Interactions (Edinburgh, Scotland) (1994) (ферментативная инактивация путем продуцирования одноцепочечных фрагментов антител); специфичное к вирусу антитело (см., например, Tavladoraki et al. (1993) Nature 366:469 (гены рекомбинантных антител для защиты от вирусной атаки)); связанный со стадией развития ингибирующий белок, продуцируемый в природе патогеном или паразитом (см., например, Lamb et al. (1992) Bio/Technology 10:1436 (грибковые эндо-α-1,4-D-полигалактуроназы способствуют грибковой колонизации и высвобождению питательных веществ растений путем растворения гомо-α-1,4-D-галактуроназы клеточной стенки растения; Toubart et al. (1992) Plant J. 2:367 (ингибирующий эндополигалактуроназу белок)); и связанный со стадией развития ингибирующий белок, продуцируемый в природе растением (см., например, Logemann et al. (1992) Bio/Technology 10:305 (ген, инактивирующий рибосомы ячменя, обеспечивающий увеличенную устойчивость к грибковому заболеванию)).
Другие примеры признаков, которые могут быть желательными для комбинирования с желаемыми характеристиками состава семян, включают, например, и не ограничиваясь ими: гены, которые придают устойчивость к гербициду (Lee et al. (1988) EMBO J. 7:1241 (мутантный фермент ALS); Miki et al. (1990) Theor. Appl. Genet. 80:449 (мутантный фермент AHAS); патенты США №№ 4940835 и 6248876 (мутантные гены 5-енолпирувилшикимат-3-фосфатсинтазы (EPSP), обеспечивающие устойчивость к глифосату); патент США № 4769061 и номер доступа ATCC 39256 (гены aroA); гены глифосатацетилтрансферазы (устойчивость к глифосату); другие фосфоносоединения из видов Streptomyces, включая Streptomyces hygroscopicus и Streptomyces viridichromogenes), такие как гены, описанные в европейской заявке № 0242246 и DeGreef et al. (1989) Bio/Technology 7:61 (гены глюфосинатфосфинотрицинацетилтрансферазы (PAT), обеспечивающие устойчивость к глифосату); пиридинокси или феноксипропионовые кислоты и циклогексоны (устойчивость к глифосату); патентная европейская заявка № 0333033 и патент США № 4975374 (гены глутаминсинтетазы, обеспечивающие устойчивость к гербицидам, таким как L-фосфинотрицин); Marshall et al. (1992) Theor. Appl. Genet. 83:435 (гены Acc1-S1, Acc1-S2 и Acc1-S3, обеспечивающие устойчивость к феноксипропионовым кислотам и циклогексонам, таким как сетоксидим и галоксифоп); WO 2005012515 (гены GAT, обеспечивающие устойивость к глифосату); WO 2005107437 (гены, обеспечивающие устойчивость к 2,4-D, fop и пиридилокси-ауксиновым гербицидам); и гербициды, которые ингибируют фотосинтез, такие как триазин (гены psbA и gs+) или бензонитрил (ген нитрилазы) (см., например, Przibila et al. (1991) Plant Cell 3:169 (мутантные гены psbA); нуклеотидные последовательности для генов нитрилазы описаны в патенте США № 4810648, и молекулы ДНК, содержащие эти гены, доступны под номерами доступа ATCC 53435, 67441 и 67442; и Hayes et al. (1992) Biochem. J. 285:173 (глутатион S-трансфераза)).
Следующие примеры признаков, которые могут быть желательными для комбинирования с желаемыми характеристиками состава семян, включают, например, и не ограничиваясь этим, гены, которые обеспечивают или вносят вклад в дополнительный ценный признак, например, модифицированный метаболизм жирных кислот (см., например, Knultzon et al. (1992) Proc. Natl. Acad. Sci. U.S.A. 89:2624 (антисмысловой ген стеарил-ACP-десатуразы для увеличения содержания стеариновой кислоты в растении)); сниженное содержание фитатов (см., например, Van Hartingsveldt et al. (1993) Gene 127:87 (ген фитазы Aspergillus niger усиливает разрушение фитата, что обеспечивает больше свободного фосфата в трансформированном растении); и Raboy et al. (1990) Maydica 35:383 (клонирование и повторное введение ДНК, ассоциированной с аллелем, ответственным за мутанты кукурузы, имеющие низкие уровни фитиновой кислоты)); и модифицированный углеводный состав, достигаемый, например, путем трансформации растений геном, кодирующим фермент, который изменяет профиль ветвления крахмала (см., например, Shiroza et al. (1988) J. Bacteol. 170:810 (мутантный ген фруктозилтрансферазы Streptococcus); Steinmetz et al. (1985) Mol. Gen. Genet. 20:220 (ген левансахаразы); Pen et al. (1992) Bio/Technology 10:292 (α-амилаза); Elliot et al. (1993) Plant Molec. Biol. 21:515 (гены инвертазы томата); Sogaard et al. (1993) J. Biol. Chem. 268:22480 (ген α-амилазы ячменя); и Fisher et al. (1993) Plant Physiol. 102:1045 (фермент ветвления крахмала эндоспермы кукурузы II)).
Ссылки, рассмотренные в настоящем описании, предоставлены только для их раскрытия до даты подачи настоящей заявки. Ничто в настоящем документе не следует истолковывать как допущение того, что это изобретение дает право на противопоставление факта создания изобретения с более ранним приоритетом.
Следующие примеры предоставлены для иллюстрации особенных конкретных признаков и/или аспектов заявленного изобретения. Эти примеры не следует истолковывать как ограничивающие раскрытие конкретных описанных признаков или аспектов.
ПРИМЕРЫ
Пример 1: Средний состав питательных веществ и ценность улучшенной муки канолы (ECM) и общепринятой муки канолы
Между 2009 и 2012 годами было проведено несколько аналитических и функциональных испытаний для оценки пищевого состава и ценности линий и гибридов ECM по настоящему изобретению. Исследование проводили на цельных необработанных семенах, частично переработанной муке и полностью переработанной муке для учета возможных эффектов обработки на состав питательных веществ и ценность. Образцы анализировали в университетах Иллинойса, Миссури, Джорджии и Манитобы. Эту информацию о составе использовали для оценки энергетической ценности улучшенной муки канолы относительно общепринятой муки канолы с использованием стандартных прогнозирующих уравнений. Биологическую оценку образцов в отношении усвоения энергии и аминокислот у птиц проводили в университетах Иллинойса и Джорджии. Биологическую оценку образцов в отношении усвоения энергии и аминокислот у свиней проводили в Университете Иллинойса. Обобщение различий пищевого состава между линиями ECM (диапазоны или среднее значение) и общепринятой мукой канолы представлено в таблице 1. Детали соответствующих методик и испытаний представлены в последующих примерах.
Средний состав питательных веществ ECM и общепринятой муки канолы
** Спрогнозировано, исходя из состава питательных веществ
Линии ECM демонстрируют несколько четких улучшений состава питательных веществ, которые обеспечивают пользу при кормлении животных. Как проиллюстрировано в таблице 1, ECM приблизительно на 7% больше содержит белка, чем общепринятая мука канолы. Кроме того, баланс незаменимых аминокислот (в качестве процента от белка) сохраняется при более высоких уровнях белка. Усвояемость аминокислот в ECM птицами и свиньями по меньшей мере настолько же высокая, как в случае общепринятой муки канолы, и ключевая аминокислота лизин, по-видимому, имеет немного более высокую усвояемость. Линии ECM показали более низкие уровни волокнистых компонентов, которые встречаются в клеточной стенке и шелухе, в частности, приблизительно на 2% более низкие уровни лигнина/полифенолов, на 1% более низкие уровни целлюлозы, на 3% более низкие уровни остатка ADF (3%) и на 5% более низкие уровни ADF.
Более высокие уровни белка и более низкие уровни волокнистых компонентов коррелируют приблизительно с 10% увеличением биологической энергии в линиях ECM. Эти линии также продемонстрировали более высокие уровни фосфора, который является дорогостоящим питательным веществом для добавления в корма для животных. Более высокое содержание белка (аминокислот), энергия и фосфор коррелировали с приблизительно 20-32% увеличением стоимостной ценности ($/т) для муки канолы в кормах свиней и птиц, которое отражается в увеличенных альтернативных ценах для кормов для выращивания бройлеров и свиней. Таблица 1.
Пример 2: Способы для муки POS white flake (WF), LT и HT
Семена ECM и общепринятые семена канолы обрабатывали в POS Pilot Plant в Saskatoon, CA по следующей методике:
МАТЕРИАЛЫ:
Приблизительно 1,5 MT исследуемой линии канолы ECM (CL44864) приобретали от POS 2 августа 2011 года. Приблизительно 3,0 MT продовольственных контрольных семян канолы приобрели от POS 3 августа 2011 года. Источники основных материалов являются следующими.
Гексан/изогексан: Univar, Saskatoon, SK.
Hyflo Super-cel Filter Aid: Manville Products Corp., Denver, CO.
Азот: Air Liquide, Saskatoon, SK.
Фильтровальная ткань, однонитевая: Porritts and Spensor, Pointe Claire, PQ.
Фильтровальная бумага, 55 lb tan style 1138-55: Porritts and Spensor, Pointe Claire, PQ.
СПОСОБЫ - ПРЕДВАРИТЕЛЬНАЯ ОБРАБОТКА РАСТЕНИЙ
Между исследованиями каждого сорта канолы все оборудование на установке для "первичной" переработки пылесосили или подметали начисто. Легковоспламеняющийся экстрактор не выключали между испытаниями. Однако цепь реакторов, Schnecken и систему восстановления растворителя поддерживали в рабочем состоянии для опустошения оборудования между исследованиями сортов канолы. Вакуум не выключали, так что все пары отводились в конденсатор, конденсировались и сбрасывались в расходный бак для растворителя. Это предупреждало конденсацию воды в Schnecken и закупоривание конвейера. Образцы канолы прессовали/экстрагировали в следующем порядке:
1. Контрольный HT,
2. Контрольный LT,
3. Исследуемая линия ECM (CL44864) LT.
ФОРМИРОВАНИЕ ХЛОПЬЕВ
Формирование хлопьев проводят для разрушения масличных клеток и получения тонкого пласта с большой площадью поверхности для тепловой обработки/предварительной обработки путем пропускания семян через ряд гладких валиков. Толщину и влажность хлопьев доводят так, чтобы минимизировать количество полученных мелких частиц. Высокие уровни мелких частиц приводят к осадку на прессе, обладающему свойством плохого пропускания растворителя.
Семена канолы преобразовывали в хлопья с использованием установки минимального зазора между валиками. Диапазон толщины хлопьев для каждой партии был следующим:
Скорость подачи контролировалась скоростью прессования, и она составляла приблизительно 133-150 кг/ч.
Устройство для получения хлопьев: Lauhoff Flakmaster Flaking Mill Model S-28, Serial No. 7801 с диаметром 14" × ширину 28" (35,6 см × 71,2 см), производимое Lauhoff Corporation.
ТЕПЛОВАЯ ОБРАБОТКА (КОНДИЦИОНИРОВАНИЕ)
Тепловую обработку проводят для дальнейшего разрушения масличных клеток, придания хлопьям гибкости и увеличения эффективности шнекового пресса путем уменьшения вязкости содержащегося масла. Тепловую обработку проводят для инактивации ферментов в семенах. Устройство для тепловой обработки предварительно нагревали до начала каждого запуска. Давление пара корректировали во время работы для поддержания желаемой температуры хлопьев.
Температуры в лотках для контрольной партии HT были следующими:
верхний лоток 60±5°C,
нижний лоток 97±3°C.
Температуры в лотках для контрольной партии LT плюс партии исследуемой линии ECM (CL44864) LT были следующими:
верхний лоток 60±5°C,
нижний лоток 93±2°C.
Устройство для тепловой обработки: использовали устройства для тепловой обработки Simon-Rosedown с двумя лотками. Каждое отделение имело высоту 36 см (рабочая высота 21 см) и диаметр 91 см и поставлялось с качающейся ручкой для перемешивания материала. Пар использовали в рубашке для сухого тепла; также для содержимого емкости можно добавлять прямой пар. Устройство для тепловой обработки было установлено над винтовым прессом для прямой подачи.
ПРЕССОВАНИЕ
Прессование удаляет приблизительно 2/3 масла и формирует жмых, пригодный для экстракции растворителем. Требуется, чтобы жмых был устойчивым к дроблению для выдерживания экстрактора и имел пористость для хорошего переноса массы и стока. Преобразованные в хлопья и подвергнутые тепловой обработке семена прессовали с использованием предварительного пресса Simon-Rosedown.
Неочищенное масло после прессования собирали в емкость.
Предварительный пресс: винтовой пресс Simon-Rosedown диаметром 9,5 см и длиной 94 см. Использовали рабочую скорость винта 17 об/мин.
ЭКСТРАКЦИЯ РАСТВОРИТЕЛЕМ И УДАЛЕНИЕ РАСТВОРИТЕЛЯ
Экстракция растворителем представляет собой контактирование жмыха с гексаном для удаления масла из массы жмыха. Выполняли два механизма: вымывание масла в растворитель и промывание выжимки (гексан-твердые вещества) постепенно более слабыми мисцеллами (гексан-масло). Экстракция обычно представляет собой непрерывный противоточный процесс.
Жмых контрольной канолы HT экстрагировали смесью изогексан/гексан с использованием общего времени нахождения приблизительно 90 минут (от входа в начало цикла до выхода из цикла), соотношения растворителя к твердому веществу приблизительно 2,5:1 (масс:масс) и температуры мисцеллы 52±5°C. (Скорость подачи жмыха канолы составляла приблизительно 90 кг/ч при времени удержания 90 минут, и скорость потока составляла 220±10 кг/ч).
Образец продовольственных белых хлопьев (WF) канолы удаляли перед удалением растворителя и сушили воздухом.
Из не очищенного масла удаляли растворитель в выпарном устройстве с восходящей пленкой и отпарной колонне.
Удаление растворителя из выжимки (гексан-твердые вещества) проводили в устройстве для удаления растворителя-сушки нагреванием с паровой рубашкой с винтом Schnecken и 2 лотками. Орошающий пар добавляли к верхнему лотку DT. Заданные температуры в лотках были следующими:
выход Schnecken: <60°C,
лоток для удаления растворителя: 102±3°C,
лоток для сушки нагреванием: 102±3°C.
Жмых контрольной канолы LT и исследуемой линии ECM (CL44864) LT экстрагировали смесью изогексан/гексан с использованием общего времени нахождения приблизительно 110 минут (от входа в начало цикла до выхода из цикла), соотношения растворителя к твердому веществу приблизительно 2,5:1 (масс:масс) и температуры мисцеллы 52±5°C. (Скорость подачи жмыха канолы составляла приблизительно 80 кг/ч при времени удержания 110 минут, и скорость потока составляла 220±10 кг/ч).
Образец продовольственных белых хлопьев (WF) исследуемой линии ECM удаляли перед удалением растворителя и сушили воздухом.
Из не очищенного масла удаляли растворитель в выпарном устройстве с восходящей пленкой и отпарной колонне.
Удаление растворителя из выжимки (гексан-твердые вещества) проводили в устройстве для удаления растворителя-сушки нагреванием с паровой рубашкой с винтом Schnecken и 2 лотками. Орошающий пар добавляли к верхнему лотку DT. Заданные температуры в лотках были следующими:
выход Schnecken: <60°C,
лоток для удаления растворителя: 93±2°C,
лоток для сушки нагреванием: 93±2°C.
Экстрактор: изготовленный полностью из нержавеющий стали экстрактор Crown Iron Works Loop Extractor (Type II). Экстракционный слой имел ширину 20,3 см × глубину 12,7 см на длину 680 см. Кроме того, этот блок включает удаление растворителя из мисцеллы с использованием выпарного устройства с восходящей пленкой и отпарной колонны и удаление растворителя из выжимки (твердые вещества плюс растворитель) с использованием устройства для удаления растворителя-сушки нагреванием с паровой рубашкой с винтом Schnecken и 2 лотками. Удаленный растворитель собирали и использовали повторно.
ВАКУУМНАЯ СУШКА
Вакуумную сушку проводили для высушивания обезжиренной муки канолы LT до <12% влаги.
Единственной мукой канолы, для которой требовалась сушка, была контрольная партия LT. Приблизительно 225 кг обезжиренной муки помещали в сушку Littleford Reactor Dryer. Затем муку нагревали до 75±2°C в вакууме 10-15” рт.ст. Взятие образцов для анализа содержания влаги начинали при ~60°C и проводили каждые 15 минут до тех пор, пока содержание влаги не составило <12%. Затем муку сбрасывали в мешок для сыпучих материалов. Описанный выше процесс повторяли до высушивания всей муки.
Вакуумная сушилка: 600 Liter Model FKM600-D (2Z) Littleford Reactor, серийный номер #5132, Littleford Day, Florence, KY.
ИЗМЕЛЬЧЕНИЕ В МОЛОТКОВОЙ МЕЛЬНИЦЕ
Измельчение в молотковой мельнице проводили для достижения единообразного размера частиц.
Высушенную муку измельчали в молотковой мельнице с использованием сита 8/64”. Молотковую мельницу пылесосили после каждой партии муки. Муку упаковывали в фибровые барабаны и хранили при температуре окружающей среды до транспортировки.
Порядок, в котором муку канолы измельчали в молотковой мельнице, был следующим:
1. Контрольная HT.
2. Исследуемая линия ECM (CL44864) LT.
3. Контрольная LT.
Молотковая мельница: Prater Industries, Model G5HFSI, серийный номер #5075, Chicago, IL
Пример 3: Процесс получения белых хлопьев Indianapolis
Семена канолы по настоящему изобретению можно обрабатывать для получения белых хлопьев канолы с использованием методики, первоначально описанной в Bailey's Industrial Oil & Fat Products (1996), 5th Ed., Chapter 2, Wiley Interscience Publication, New York, New York.
Для экстракции масла из семян канолы семена канолы сначала преобразуют в хлопья с помощью мельницы для измельчения кофе и обрабатывают нагреванием в печи до 85°C±10°C в течение по меньшей мере 20 минут. После обработки нагреванием измельченные семена прессуют с использованием пресса Taby Press Type-20A Press (Taby Skeppsta, Orebro, Швеция). Полученный жмых из Taby Press подвергают экстракции растворителем для удаления какого-либо оставшегося остаточного масла.
Затем жмых со стадии прессования масличных семян подвергают экстракции растворителем для удаления и сбора какого-либо оставшегося остаточного масла. Жмых помещают в барабаны из нержавеющей стали, которые помещают в изготовленный на заказ экстрактор SoxhletTM от LaSalle Glassware (Guelph, ON). В качестве растворителя для экстракции можно использовать гексан, и системе для экстракции SoxhletTM позволяют работать в течение 9-10 часов. Затем жмых после экстракции растворителем удаляют из барабанов и распределяют на лоток с толщиной слоя жмыха менее одного дюйма (2,5 см). Растворителю жмыха после экстракции растворителем позволяют испариться на воздухе в течение 24 часов перед измельчением. Затем белые хлопья после удаления растворителя измельчают с использованием, например, Robot Coupe R2N Ultra B (Jackson, MS).
Пример 4: Анализ образца
Химический анализ и анализ питательных веществ ECM и общепринятых образцов канолы можно проводить различным путем с использованием способов, описанных ниже. Образцы муки канолы анализировали в отношении сухого вещества (способ 930.15; AOAC International. 2007. Official Methods. Of Analysis of AOAC Int. 18th ed. Rev. 2. W. Hortwitz и G. W. Latimer Jr., eds. Assoc. Off. Anal. Chem. Int., Gaithersburg. MD. (далее "AOAC Int., 2007")), золы (способ 942.05; AOAC Int.) и GE с помощью бомбового калориметра (Model 6300, Parr Instruments, Moline, IL). AOAC International (2007) Official Methods of Analysis of AOAC Int., 18th ed. Rev. 2., Hortwitz и Latimer, eds. Assoc. Off. Anal. Chem. Int., Gaithersburg. MD. Гидролизованный кислотой экстракт сложных эфиров (AEE) определяли путем гидролиза кислотой с использованием 3 Н HCl (Sanderson) с последующей экстракцией неочищенного жира с петролейным эфиром (способ 954.02; AOAC Int.) на автоматическом анализаторе Soxtec 2050 (FOSS North America, Eden Prairie, MN). Sanderson (1986), "A new method of analysis of feeding stuffs for the determination of crude oils and fats," Pages 77-81, Recent Advances in Animal Nutrition, Haresign and Cole, eds. Butterworths, London, U.K. Неочищенный белок измеряли по сгоранию (способ 990.03; AOAC Int.) на устройстве Elementar Rapid N-cube protein/nitrogen apparatus (Elementar Americas Inc., Mt. Laurel, NJ); аминокислоты определяли в соответствии со способом 982.30 E (A, B и C) [AOAC Int.]; неочищенные волокна определяли в соответствии со способом 978.10 (AOAC Int.); ADF и лигнин определяли в соответствии со способом 973.18 (AOAC Int.); и NDF определяли в соответствии с Holst (Holst, D. O. 1973. Holst filtration apparatus for Van Soest detergent fiber analysis. J. AOAC. 56:1352-1356). Профиль сахаров (глюкоза, фруктоза, сахароза, лактоза, мальтоза) определяли в соответствии с Churms (Churms, 1982, Carbohydrates in Handbook of Chromatography. Zweig and Sherma, eds. CRC Press, Boca Raton, FL.), и Kakehi and Honda (1989. Silyl ethers of carbohydrates. pages 43-85 в Analysis of Carbohydrates by GLC and MS. C. J. Biermann and G. D. McGinnis, eds. CRC Press, Boca Raton, FL). Олигосахариды (рафиноза, стахиоза, вербаскоза) анализировали в соответствии с Churms; минералы (Ca, P, Fe, Mg, Mn, Cu, Na, K, S, Mo, Zn, Se, Co, Cr) анализировали с помощью оптической эмиссионной спектроскопии с индуктивно связанной плазмой (ICP-OES) [способ 985.01 (A, B и C); AOAC Int.], и фитаты определяли в соответствии с Ellis et al (1977. Quantitative determination of phytate in presence of high inorganic phosphate. Anal.Biochem. 77:536-539).
Пример 5: Исходные аналитические результаты на образцах белых хлопьев ECM Indianapolis и обычной муки канолы
Состав питательных веществ полученной из пробного растения, подвергнутой сушке нагреванием муки ECM и обычной муки канолы Несколько линий ECM (44864, 121460, 121466 и 65620) обрабатывали в лаборатории Dow AgroSciences в Индианаполисе с использованием способа, сходного с коммерческой обработкой муки канолы, но без конечной стадии удаления растворителя/сушки нагреванием после экстракции растворителем масла из семян. Этот процесс и полученные образцы называются "белыми хлопьями Indianapolis". Параметры обработки приведены в примере 3. Эти образцы белых хлопьев ECM Indianapolis исследовали в университетах Иллинойса и Миссури, и результаты представлены в таблицах 2a, 2b и 2c. Контрольная мука канолы представляет собой произведенную коммерчески муку канолы, которую сушили нагреванием. Величины выражены в расчете на сухую массу, но включая масло.
Пищевой состав образцов муки канолы в виде белых хлопьев ECM Indianapolis по сравнению с обычной мукой канолы
Результаты анализа образцов белых хлопьев ECM Indianapolis из университетов Иллинойса и Миссури были сходными с результатами, полученными для анализа цельных семям в университете Манитобы. Содержание олигосахаридов было более низким, и содержание простых сахаров было более высоким в образце 44864 (2010), чем в образцах ECM, включая 44864, выращенных в 2011 году. По-видимому, в случае образца 2010 года, растущее растение катаболизировало некоторое количество сахарозы и олигосахаридов в простые сахара незадолго до времени сбора.
Более высокое содержание белка, более низкое содержание ADF и более низкое содержание лигнина и полифенолов, наблюдаемое в линиях ECM по сравнению с обычной мукой канолы, при использовании протокола для белых хлопьев Indianapolis, являются сходными с результатами, наблюдаемыми для цельных семян. Величина 33% NDF для коммерческой муки находится на верхнем конце типичного диапазона.
Аминокислотный состав (% неочищенного белка) образцов белых хлопьев ECM Indianapolis по сравнению с обычной мукой канолы
Как и в случае с цельными семенами, результаты в таблице 2b показывают, что аминокислотный состав (в качестве процента неочищенного белка) является сходным как для образцов белых хлопьев ECM Indianapolis, так и коммерческой муки канолы. Это указывает на то, что, поскольку содержание белка увеличено в линиях ECM, содержание важных аминокислот возросло пропорционально.
Минеральный состав образцов белых хлопьев ECM Indianapolis по сравнению с общепринятой мукой канолы
Минеральный состав образцов белых хлопьев ECM Indianapolis сходен с общепринятой мукой канолы с двумя исключениями: фосфор и натрий. Как в случае с результатами университета Манитобы в отношении цельных семя, уровень фосфора в линиях ECM, как оказалось, является постоянно более высоким, чем в обычной муке канолы. Избыточный натрий в обычной муке канолы без сомнений является следствием добавления натрия в ходе обработки обычной канолы.
Пример 6: Обработка ECM в POS Pilot Plant в Saskatoon, Канада, для имитации коммерческой обработки
При приготовлении ECM для оценки кормления животных было определено, что образцы канолы следует приготовить в условиях коммерческой обработки, учитывая эффект обработки на пищевую ценность. Следовательно, образцы обрабатывали в POS Pilot Plant в Saskatoon. Использовали два условия обработки: регулярная температура (HT) в устройстве для удаления растворителя/сушки нагреванием и более низкая температура (LT) для обеспечения того, чтобы условия обработки не оказывали существенного влияния на пищевую ценность. Использованные в POS условия обработки приведены в примере 2.
Состав питательных веществ ECM и обычной муки канолы, полученной в сходных условиях коммерческой обработки в POS Pilot Plant в Saskatoon, Канада. (Анализ проводили в университетах Иллинойса и Миссури)
Предварительно обработанная мука показала сходный состав с образцами цельных семян и белых хлопьев Indianapolis, и отличия между образцом ECM и обычной мукой канолы согласуются с анализом, описанным в таблицах 2a и 2b: на 7% больше белка, на 5% меньше ADF, на 4% меньше лигнина и полифенолов и на 0,35% больше фосфора.
Пример 7: Полный анализ необработанной ECM и обычных семян канолы
Пищевой состав необработанных семян канолы. Пять образцов цельных семян линий ECM, продуцированных от 2010 до 2011 года, анализировали в университете Манитобы. Их сравнивали с официальным составным образцом семян Canadian Grain Commission (CGC) для продукции 2011 года, который по определению является средним качеством текущих коммерческих сортов канолы, выращенных в западной Канаде в этот сезон. Результаты пищевого состава выражены в расчете на сухую массу без масла, и они представлены в таблицах 4a и 4b.
Пищевой состав образцов семян ECM по сравнению с обычными семенами канолы
Результаты показывают, что наибольшим отличием между ECM и общепринятой канолой является более высокое содержание белка. В ECM содержание белка на 7,2% выше (51,1% против 43,9%) в расчете на сухую массу без масла и на 6,1% выше (43,5% против 37,4%) в расчете на 3% масла, 88% сухого вещества (типичные базовые характеристики для коммерческой муки канолы). См. таблицы 4a, 4b. По-видимому, более высокое содержание белка объясняется на 2% более низким содержанием лигнина и полифенолов в ECM и на 3% более низким содержанием остатка ADF (ADF - лигнин/полифенолы - целлюлоза). Остаток ADF, вероятно, является комбинацией компонентов гликопротеина и гемицеллюлозы. Волокнистые компоненты, главным образом, встречаются в клеточных стенках и шелухе. Содержание фосфора в ECM практически на 30% выше, чем в общепринятой каноле, и, по-видимому, равномерно распределено между фитатными и нефитатными формами. Фосфор является ценным питательным веществом в кормах для животных, и хотя связанный с фитатом фосфор плохо усваивается птицами и свиньями, широкое применение фермента фитазы в кормах для животных сделает этот фосфор доступным для животных. В таблице 4b предоставлено сходное сравнение аминокислотного состава в образцах цельных семян.
Аминокислотный состав (% неочищенного белка) образцов семян ECM по сравнению с общепринятыми семенами канолы
Результаты в таблице 4b показывают, что аминокислотный состав (в качестве процента неочищенного белка) является сходным между ECM и коммерческой мукой канолы. Это указывает на то, что, поскольку содержание белка увеличено в линиях ECM, оно увеличено и для важных аминокислот.
Пример 8: Усвояемость TME и аминокислот у птиц
Анализы истинной метаболизируемой энергии (TME) и истинных доступных аминокислот (TAAA) были разработаны в 1976 и 1981 годах, соответственно, Dr. Ian Sibbald of Agriculture Canada in Ottawa. Вследствие прямой и недеструктивной природы анализов, эти анализы стали предпочтительными для выбора способами определения доступности энергии и аминокислот в ингредиентах кормов для птиц в большей части мира, включая США.
В качестве предпочтительного для выбора экспериментального животного в Университете Иллинойса и Университете Джорджии использовали зрелых петухов породы белый леггорн с листовидным гребнем (SCWL). Хорошо известно, что птицы имеют быстрое время опустошения кишечника. Путем прекращения кормления на 24 часа можно достоверно предположить, что в пищеварительном тракте исследуемых индивидуумов отсутствуют ранее употребленные остатки пищи.
Каждую птицу (как правило, 8 индивидуумов на введение) кормят с точностью в 35 граммов исследуемого корма, помещенного непосредственно в зоб путем интубации. Ингредиенты с высоким содержанием волокон обычно дают в количестве 25 вместо 35 граммов в сходном объеме. После интубации обеспечивают доступ птиц к воде, но не к дополнительному корму, на 40 часов, и в ходе этого периода экскременты собирают с определением их количества. После сбора экскременты сушат в печи с нагнетаемым воздухом, обычно при 80°C. Затем их взвешивают и измельчают для определения суммарной энергии (GE) в анализах TME или для определения содержания аминокислот. GE и состав аминокислот в ингредиентах определяют аналогичным образом. После взвешивания образцы экскрементов, как правило, объединяют и гомогенизируют для определения по отдельности GE или аминокислот. Масса экскрементов на птицу варьирует значительно больше, чем GE или состав аминокислот в конкретных экскрементах. Это наблюдение, и расходы, и задержка во времени при определении GE и аминокислот оправдывают объединение.
Усвояемость вычисляют с использованием способов, хорошо известных в данной области для определения энергии и для каждой аминокислоты по отдельности. Оценки эндогенной утраты GE и аминокислот используют для коррекции экспериментальных артефактов.
Пример 9: Усвояемая энергия (DE), метаболизируемая энергия (ME) у свиней
DE и ME. Сорок восемь растущих боровов (исходная BW: 20 кг) будут выбраны в рандомизированное полное испытание с блочной схемой в Университете Иллинойса. Свиней будут распределять на 1 из 6 рационов, по 8 свиней на рацион. Свиней поместят в метаболические клетки, которые будут оборудованы кормушкой и сосковой поилкой, полами, сделанными полностью из планок, экранным полом и лотками для мочи. Это позволит общий, но по отдельности, сбор мочи и фекальных материалов от каждой свиньи.
Количество корма, предоставляемого каждые сутки на свинью, будут вычислять в качестве 3-кратной оцененной потребности в поддерживающей энергии (т.е. 106 ккал ME на кг0,75; NRC, 1998) для наименьшей свиньи в каждой группе и разделять на 2 равных кормления. NRC 1998, Nutrient requirements of swine, Tenth Revised Edition. National Academy Press. Washington, DC. Вода будет доступна все время. Эксперимент продлится 14 суток. Первые 5 суток будут считаться периодом адаптации к рациону, со сбором мочи и фекальных материалов в течение последующих 5 суток в соответствии со стандартными методиками с использованием подхода от маркера к маркеру (Adeola, O. 2001, Digestion and balance techniques in pigs, pages 903-916 in Swine Nutrition. 2nd ed. A. J. Lewis and L. L. Southern, ed. CRC Press, New York, NY. NRC. 1998. Nutrient Requirements of Swine. 10th rev. ed. Natl. Acad. Press, Washington DC.). Образцы мочи будут собирать в ведра для мочи с консервантом, представляющим собой 50 мл хлористоводородную кислоту. Фекальные образцы и 10% собранной мочи будут хранить при -20ºC сразу после сбора. В заключение эксперимента, образцы мочи будут размораживать и смешивать для каждого животного и рациона, и для химического анализа будут брать субобразец.
Фекальные образцы будут сушить в печи с нагнетаемым воздухом и мелко измельчать перед анализом. Образцы фекалий, мочи и корма будут анализировать в двух экземплярах в отношении DM и суммарной энергии с использованием бомбовой калориметрии (Parr Instruments, Moline, IL). После химического анализа будут вычислять общие величины усвояемости в пищеварительном тракте для определения энергии в каждом рационе с использованием методик, описанных ранее (Widmer, M. R., L. M. McGinnis, and H. H. Stein. 2007. Energy, phosphorus, and amino acid digestibility of high-protein distillers dried grains and corn germ fed to growing pigs. J. Anim. Sci. 85:2994-3003). Будут вычислять величину энергии, утраченной в фекалиях и в моче, соответственно, и будут вычислять количества DE и ME в каждом из 24 рационов (Widmer et al., 2007). DE и ME в кукурузе будут вычислять путем деления величин DE и ME для рациона кукурузы на коэффициент включения кукурузы в этот рацион. Затем эти величины будут использовать для вычисления вклада кукурузы в DE и ME в рационах мука кукурузы-канолы и в рационе мука кукурузы-сои, а затем будут вычислять DE и ME в каждом источнике муки канолы и в образце муки сои по разности, как описано ранее (Widmer et al., 2007).
Данные будут анализировать с использованием Proc Mixed Procedure в SAS (SAS Institute Inc., Cary, NC). Данные, полученные для каждого рациона и для каждого ингредиента, будут сравнивать с использованием ANOVA. Однородность отличий будут подтверждать с использованием процедуры UNIVARIATE в Proc Mixed. Рацион или ингредиенты будут фиксированным эффектом, и свинья и группа повторения будут случайными эффектами. Средние значения по методу наименьших квадратов будут вычислять с использованием LSD-теста, и средние значения будут различать с использованием инструкции pdiff в Proc Mixed. Свиньи будут находиться в экспериментальном блоке для всех вычислений, и альфа-уровень 0,05 будут использовать для оценки значимости среди средних значений.
Пример 10: Усвояемость аминокислот (AID и SID) у свиней
AID и SID у свиней анализировали в испытании в Университете Иллинойса. Двенадцати растущим боровам (исходная BW: 34,0±1,41 кг) помещали T-образную канюлю вблизи дистального отдела подвздошной кишки и распределяли согласно повторяющемуся 6×6 плану опыта по схеме латинского квадрата с 6 рационами и 6 периодами в каждом квадрате. Свиней содержали по отдельности в загонах 1,2×1,5 м в комнате с контролируемыми условиями. Загоны имели плотную обшивку, полы, сделанные полностью из планок, и в каждый из загонов была помещена кормушка и сосковая поилка.
Приготавливали шесть рационов. Пять рационов были основаны на кукурузном крахмале, сахаре и SBM или муке канолы, и SBM или мука канолы были единственным источником а.к. в этих рационах. Последний рацион представлял собой рацион без N, который использовали для оценки базальных эндогенных потерь в подвздошной кишке CP и AA. Витамины и минералы включали во все рационы, чтобы они удовлетворяли или превышали современные оценки потребности для растущих свиней (NRC, 1998). Все рационы также содержали 0,4% оксида хрома в качестве неусвояемого маркера.
Массу свиней регистрировали в начале и в конце каждого периода и также регистрировали количество корма, предоставляемого каждые сутки. Всех свиней кормили на уровне, в 2,5 раза превышающем потребность в поддерживающей энергии, и вода была доступна все время на протяжении эксперимента. Первые 5 суток каждого периода считали периодом адаптации к рациону. Образцы, переваренные в подвздошной кишке, собирали в течение 8 часов на 6 и 7 сутки с использованием стандартных методик. К цилиндру канюли был подсоединен пластмассовый мешок с использованием хомута, и собирали перевариваемую пищу, попадающую в мешок. Мешки удаляли, когда они заполнялись перевариваемой пищей, или по меньшей мере каждые 30 мин, и сразу замораживали при -20°C, чтобы предотвратить бактериальную деградацию аминокислот в перевариваемой пище. После завершения экспериментального периода животных не кормили в течение ночи и на следующее утро давали новый экспериментальный рацион.
После завершения эксперимента образцы, взятые из подвздошной кишки, размораживали, объединяли для каждого животного и рациона и брали субобразец для химического анализа. Также собирали образец каждого рациона и каждый из образцов муки канолы и SBM. Образцы перевариваемой пищи лиофилизировали и тонко измельчали перед химическим анализом. Все образцы рационов и перевариваемой пищи анализировали в отношении DM, хрома, неочищенного белка и а.к., и муку канолы и SBM анализировали в отношении неочищенного белка и а.к.
Величины кажущейся усвояемости в подвздошной кишке (AID) для а.к. в каждом рационе вычисляли с использованием уравнения [1]:
AID, (%)=[1-(AAd/AAf)×(Crf/Crd)]×100, [1],
где AID представляет собой величину кажущейся усвояемости в подвздошной кишке для а.к. (%), AAd представляет собой концентрацию этой а.к. в перевариваемой пище DM в подвздошной кишке, AAf представляет собой концентрацию а.к. относительно этой а.к. в корме DM, Crf представляет собой концентрацию хрома в корме DM, и Crd представляет собой концентрацию хрома в перевариваемой пище DM в подвздошной кишке. С использованием этого уравнения также вычисляют AID для CP.
Базальный эндогенный поток в дистальную подвздошную кишку каждой а.к. определяли на основе потока, полученного после кормления рационом без N с использованием уравнения [2]:
IAAend=AAd×(Crf/Crd) [2],
где IAAend представляет собой базальную эндогенную потерю а.к. (мг на кг DMI). Базальную эндогенную потерю CP будут определять с использованием того же уравнения.
Путем коррекции AID для IAAend каждой а.к. вычисляли стандартизованную усвояемость а.к. в подвздошной кишке с использованием уравнения [3]:
SID, (%)=AID+[(IAAend/AAf)×100] [3],
где SID представляет собой стандартизованную величину усвояемости в подвздошной кишке (%).
Данные анализировали с использованием методики Proc GLM в SAS (SAS inst. Inc., Cary, NC). 5 рационов, содержавших муку канолы или SBM, сравнивали с использованием ANOVA с источником муки канолы, свиньями и периодом в качестве главных эффектов. Для различения средних значений использовали LSD-тест. Альфа-уровень, равный 0,05, использовали для оценки значимости среди средних значений. Свиньи находились в экспериментальном блоке для всех анализов.
Пример 11: Деградируемость а.к. у молочных животных
Деградируемость аминокислот ECM будут оценивать путем инкубации in-situ образцов муки ECM у животных с канюлированным рубцом, таких как молочный скот, для оценки содержания растворимого и деградируемого белка и определения скорости деградации (Kd) деградируемой фракции.
Крупный рогатый скот будут кормить смешанным рационом в качестве общего смешанного рациона (TMR), содержащего 28,1% кукурузного силоса, 13,0% силоса люцерны, 7,4% сена люцерны, 20,4% измельченной кукурузы, 14,8% влажной пивной дробины, 5,6% цельных семян хлопчатника, 3,7% соевой шелухи и 7,0% добавок (белок, минералы, витамины). Стандартные полиэфирные мешки in situ (R510, 5 см × 10 см, размер пор 50 микрон), содержащие приблизительно 6 г сухого вещества (DM) соевой муки (SBM), обычную муку канолы (CM), или улучшенную муку канолы (ECM), будут инкубировать в рубце в течение 0, 2, 4, 8, 12, 16, 20, 24, 32, 40, 48 и 64 часов. Мешки в двух экземплярах будут удалять в каждый момент времени и промывать в проточной воде до тех пор, пока стекающая жидкость не станет прозрачной. Мешки будут сушить при 55°C в течение 3 суток, а затем осадок будут удалять и взвешивать для определения исчезновения сухого вещества (DM). Осадок будут анализировать в отношении содержания N с использованием способа сжигания Leco. Образцы нулевого момента времени не будут инкубироваться в желудке, а будут промываться и обрабатываться аналогично тому, как и инкубированные в рубце образцы.
Образцы осадка нулевого времени и осадка, оставшегося после инкубации в рубце в течение 16 ч, будут анализировать в отношении приблизительного состава (DM, неочищенные жиры, неочищенные волокна и зола) и аминокислотного (AA) состава (без триптофана). Эти параметры можно использовать для проведения оценки деградируемого в рубце белка (RDP) и не деградируемого в рубце белка (RUP), как используется в руководствах National Research Council (2001) для требований к питательным веществам для молочного крупного рогатого скота.
Процент исходного N в образце, оставшегося в каждый момент времени, можно вычислять, и полученные повторно величины для каждого момента времени будут усреднять для каждой коровы. Величины, полученные для трех коров, будут приводить в соответствие с нелинейным уравнением, описанным Ørskov and McDonald (1979). В этом подходе допускают, что исчезновение CP в рубце соответствует кинетике первого порядка, как определяют по уравнению, исчезновение CP=A+B×(1−e−Kd×t), где A представляет собой растворимую фракцию CP (% CP), B представляет собой потенциально деградируемую фракцию CP (% CP), Kd представляет собой константу скорости деградации (ч-1), и t представляет собой время инкубации в рубце (ч). Фракцию C (не деградируемую в рубце) вычисляют как фракция A минус фракция B. Уравнения будут приводить в соответствие с использованием PROC NLIN в SAS (версия 9.2; SAS Institute Inc., Cary, NC), со способом вычисления Марквардта.
Уравнения для вычисления величин RDP и RUP (в качестве процентов от CP) представляют собой: RDP=A+B[Kd/(Kd+Kp)] и RUP=B[Kp/(Kd+Kp)]+C, где Kp представляет собой скорость прохождения через рубец. Поскольку скорость прохождения через рубец не может быть вычислена непосредственно из этих данных (где вещества содержатся в рубце, и предотвращается их прохождение в нижние отделы желудочно-кишечного тракта), должна быть взята скорость для Kp. В этом испытании для Kp будут использовать величину, равную 0,07, которая сходна с величиной, вычисленной в соответствии с уравнениями в NRC (2001) для высокопродуктивных молочных коров, употребляющих типичный рацион периода лактации. Поскольку целью этого проекта является сравнение источников белка и оценка усвояемости в рубце в одних и тех же условиях, выбор скорости прохождения для определения RDP и RUP является произвольным.
Конечное уравнение для каждого образца будут получать с использованием образцов, инкубированных в течение 0, 2, 4, 8, 16, 24 и 48 ч в соответствии с рекомендациями NRC (2001). Данные для дополнительных моментов времени инкубации в этом исследовании (т.е. 12, 20, 32, 40 и 64 ч) можно использовать для подтверждения кинетики системы и обеспечения того, чтобы модифицированная мука канолы соответствовала допущениям в характеристиках NRC (2001).
Пример 12: TME и TAAA у птиц, включая сравнение истинной TME с предсказанной TME, исходя из аналитических результатов из университетов Иллинойса, Миссури и Манитобы
Оценку истинной метаболизируемой энергии (TME) у птиц в образцах ECM проводили как в Университете Иллинойса, так и в Университете Джорджии. Протоколы описаны в примере 8.
Содержание TME в ECM и общепринятой муке канолы в исследованиях Университета Иллинойса и Университета Джорджии
** (SE)
*** (процентное отличие)
В случае приготовленных в POS образцов ECM и муки канолы, надлежащее сравнение проводят между двумя типами муки LT для устранения эффектов обработки. Результаты были сравнимыми в испытаниях как Университета Иллинойса, так и Университета Джорджии. TME у птиц значительно выше для ECM (LT), чем для обычной муки канолы (LT) - на 9% выше в испытании Университета Иллинойса и на 14% выше в испытании Университета Джорджии. Эти результаты подтверждают результаты прогнозирующего уравнения ниже. Таблица 4. Образцы белых хлопьев ECM и обычной муки канолы также получали в POS сразу после стадии экстракции растворителем и до стадии DT. TME у птиц для этой муки WF сравнивали в отдельном испытании в Университете Джорджии и, как и в случае образцов LT, WF ECM имели значимо более высокую TME, чем WF обычной муки канолы.
Таблица 4.
Четыре сорта ECM независимо обрабатывали в Dow AgroSciences laboratories в Индианаполисе с использованием способов получения белых хлопьев, описанных в примере 3. Затем эти образцы подвергали анализу TME у птиц в двух университетах. Не было значимых отличий в TME между исследуемыми линиями ECM, за исключением того, что линия 121460, по-видимому, имела более низкую TME, чем линии 121466 или 65620.
Выявленные величины TME в этих испытаниях согласовывались со следующим спрогнозированным содержанием метаболизируемой энергии. The National Research Council Nutrient Requirements of Poultry (NRC, 1984, Nutrient requirements of poultry. Ninth Revised Edition. National Academy Press. Washington, DC) имеет уравнение для прогнозирования ME в муке канолы (двойная нулевая рапсовая мука):
ME, ккал/кг = (32,76×CP%)+(64,96×EE%)+(13,24×NFE%).
Путем вычисления на 7% более высокий CP должен компенсироваться за счет на 7% более низкой NFE, так что суммарный коэффициент для CP должен составлять: 32,76-13,24=19,52. Это приводит к на 137 ккал/кг большей ME в ECM, чем в муке канолы (7%×19,52=137). Проблема этого уравнения состоит в том, что NFE является плохой оценкой энергетической ценности сахара и крахмала.
Альтернативным уравнением является уравнение прогнозирования EEC для ME у птиц (взрослых). (Fisher, C и J.M. McNab. 1987. Techniques for determining the ME content of poultry feeds. Haresign and D.J.A. Cole (Eds), Recent Advances in Animal Nutrition - 1987. Butterworths, London. P. 3-17):
ME, ккал/кг = (81,97×EE%)+(37,05×CP%)+(39,87 × крахмал %)+(31,08 × сахара %).
Уравнение EEC представляет собой уравнение "положительного вклада", которое дает величину усвояемых питательных веществ в муке канолы, таких как белок, жиры, крахмал и свободные сахара. Поскольку единственным аналитическим отличием между ECM и мукой канолы является белок, можно использовать коэффициент 37,05 для вычисления дополнительной энергии:
37,05×7%=259 ккал/кг. Уравнение EEC предназначено для полных кормов, которые, как правило, имеют более высокую усвояемость, чем мука канолы. Таким образом, коэффициент, равный 37,05, является слишком высоким.
Альтернативным подходом является применение первых принципов для энергетической ценности белка. Грубая оценка составляет суммарную энергию 4 калории на грамм белка × 80% усвояемость белка × 5% потерю на экскрецию азота = приблизительно 75% суммарных калорий на грамм (3 калории метаболизируемой энергии на грамм или 30 × белок %). Этот дает метаболизируемую энергию: 30×7%=210 ккал/кг дополнительной ME в ECM.
В общем, ожидается, что мука ECM будет иметь на 140-260 ккал/кг больше ME у птиц, чем обычная мука канолы. Величина 140 ккал/кг, вероятно, является существенно недооцененной, и величина 260 ккал/кг может быть немного завышенной. Вероятным является увеличение, составляющее 200-220 ккал/кг ME у птиц. Выражая это, основываясь на том, "как есть" (таблица 1), коммерческая ECM вероятно может иметь ME у птиц, составляющую 2200 ккал/кг против 2000 ккал/кг для обычной муки канолы. Это представляет собой 10% увеличение энергии.
Истинную усвояемость аминокислот у птиц (TAAA) также измеряли как в Университете Иллинойса, так и в Университете Джорджии. В этом случае анализировали только приготовленные в POS образцы муки, поскольку значительно более высокая усвояемость аминокислот белых хлопьев относительно высушенной нагреванием муки канолы не считалась коммерчески значимой. Таблица 6.
Отсутствуют статистически значимые отличия в доступности аминокислот у птиц между различными образцами муки канолы. Таблица 6.
Пример 13: Усвояемость аминокислот (AID и SID) у птиц и спрогнозированная NE
Испытания усвояемости аминокислот в подвздошной кишке свиней проводили в Университете Иллинойса. Для сравнения использовали муку, полученную в POS Pilot Plant.
Были отмечены некоторые статистически значимые отличия в усвояемости белка и аминокислот между образцам ECM и муки канолы. ECM имела более высокое содержание AID неочищенного белка, чем мука канолы, но отличие в SID белка не было значимым. Как для AID, так для и SID лизин является более усвояемым в ECM, чем в обычной муке канолы, которую подвергли такой же тепловой обработке. Таблица 7.
Для свиней общепринятые уравнения для прогнозирования DE, ME и NE представляют собой уравнения Noblet, как указано в EvaPig (2008, Version 1.0. INRA, AFZ, Ajinomoto Eurolysine), и NRC Nutrient Requirements of Swine (NRC, 1998, Nutrient requirements of swine; Tenth Revised Edition; National Academy Press. Washington, DC):
Уравнения 1-4. DE, ккал/кг = 4151-(122 × зола %)+(23×CP%)+(38×EE%)-(64×CF%).
Уравнения 1-14. NE, ккал/кг = 2790+(41,22×EE%)+(8,1 × крахмал %)-(66,5 × зола %)-(47,2×ADF%).
Уравнения Noblet являются гибридом как положительных, так и отрицательных факторов вклада: жиры, белок и крахмал имеют положительные коэффициенты, а зола, CF и ADF имеют отрицательные коэффициенты. Белок не используют в уравнении для суммарной энергии (NE), но отличия между ECM и мукой канолы могут быть установлены по отличиям в ADF. Поскольку уровень крахмала и золы является одинаковым в ECM и муке канолы, то ключевым отличием является ADF. На 5% более низкие ADF приводят к 47,2×5%=236 ккал/кг более высокой NE в ECM. Это предсказанное число является сходным с числом ME у птиц, так что вновь является вероятным увеличение суммарной энергии у свиней, составляющее 200 ккал/кг для ECM, основываясь на том, "как есть" (таблица 1). Это должно приводить приблизительно к 12% увеличению энергии.
Пример 14: Дополнительные гибриды ECM
Новый гибрид канолы CL166102H также проявлял улучшенные свойства муки (ECM). Характеристики и признаки качества, измеренные для семян этого гибрида, собранных в небольших предварительных испытаниях в 2011 году, включают масло, белок муки, ADF и общее содержание глюкозинолатов (Tgluc). См. таблицу 8.
Результаты в таблице 8 отчетливо указывают на то, что эта новая линия DAS ECM превосходит коммерческий сорт в отношении качества муки.
Агрономические характеристики линий ECM (испытания C3B03)
(%)
(%)
| название | год | авторы | номер документа |
|---|---|---|---|
| ЗАРОДЫШЕВАЯ ПЛАЗМА КАНОЛЫ, ОБЛАДАЮЩАЯ КОМПОЗИЦИОННЫМИ СВОЙСТВАМИ СЕМЯН, КОТОРЫЕ ОБЕСПЕЧИВАЮТ УВЕЛИЧЕННУЮ ПИЩЕВУЮ, ПИТАТЕЛЬНУЮ ЦЕННОСТЬ КАНОЛЫ | 2012 |
|
RU2642297C2 |
| ЗАРОДЫШЕВАЯ ПЛАЗМА КАНОЛЫ, ОБЛАДАЮЩАЯ КОМПОЗИЦИОННЫМИ СВОЙСТВАМИ СЕМЯН, КОТОРЫЕ ОБЕСПЕЧИВАЮТ УВЕЛИЧЕННУЮ ПИЩЕВУЮ, ПИТАТЕЛЬНУЮ ЦЕННОСТЬ КАНОЛЫ | 2012 |
|
RU2740712C2 |
| Brassica juncea КАЧЕСТВА ОМЕГА-9 | 2009 |
|
RU2557316C2 |
| КАНОЛА HO/LL С УСТОЙЧИВОСТЬЮ К ЗАБОЛЕВАНИЮ КИЛОЙ КРЕСТОЦВЕТНЫХ | 2012 |
|
RU2618846C2 |
| КАНОЛА HO/LL С УСТОЙЧИВОСТЬЮ К ЗАБОЛЕВАНИЮ КИЛОЙ КРЕСТОЦВЕТНЫХ | 2012 |
|
RU2711934C2 |
| СОХРАНЕНИЕ ДОКОЗАГЕКСАЕНОВОЙ КИСЛОТЫ (DHA) В ПРОЦЕССЕ ПЕРЕРАБОТКИ КАНОЛЫ | 2012 |
|
RU2621555C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЗИГОТНОСТИ ГЕНА FAD3 В КАНОЛЕ | 2012 |
|
RU2630998C2 |
| SNP-МАРКЕРЫ И ОТБОР В ОТНОШЕНИИ НИЗКОГО СОДЕРЖАНИЯ КЛЕТЧАТКИ В ПРЕДСТАВИТЕЛЯХ РОДА BRASSICA | 2019 |
|
RU2834673C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЗИГОТНОСТИ ГЕНА FAD-2 КАНОЛЫ С ИСПОЛЬЗОВАНИЕМ ПЦР С ДЕТЕКЦИЕЙ ПО КОНЕЧНОЙ ТОЧКЕ | 2012 |
|
RU2639508C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЛИПИДОВ | 2012 |
|
RU2656996C2 |

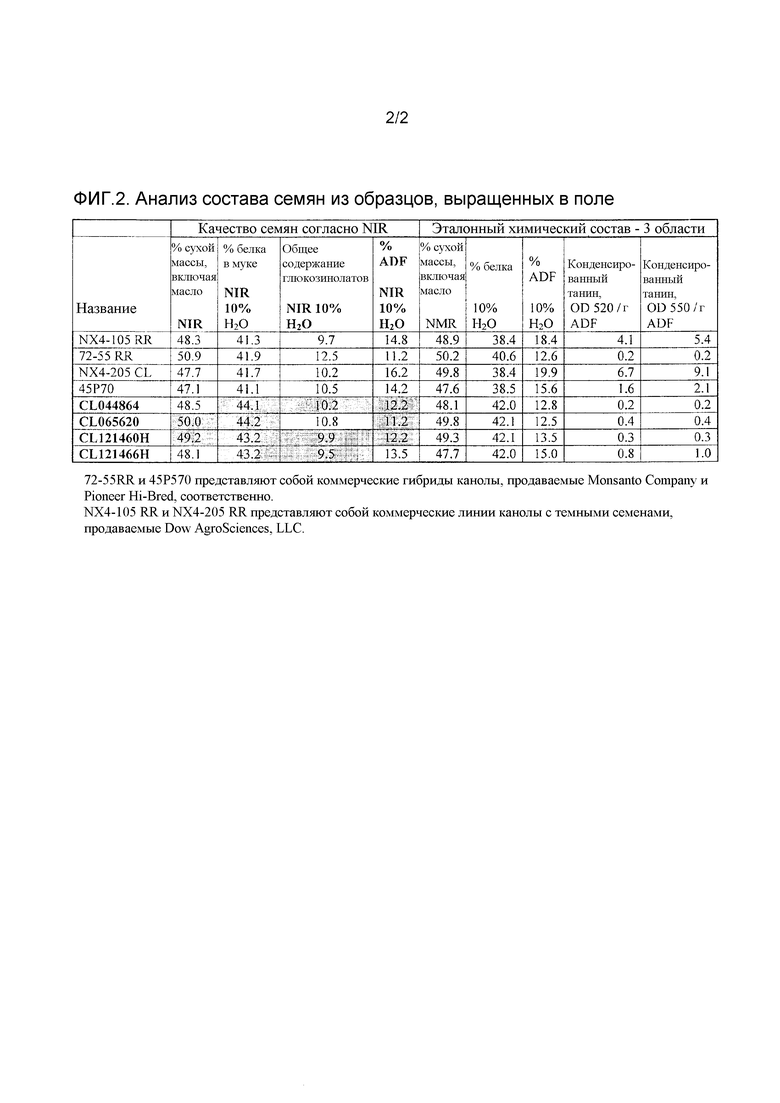
Изобретение относится к области биохимии, в частности к муке канолы, полученной из темных семян канолы, где содержание белка муки канолы составляет от 43% до 44% в расчете на 88% сухого вещества, 3% масла. Также раскрыта мука канолы, полученная из темных семян канолы, где содержание белка муки канолы составляет от 43% до 44% и содержание кислотного детергентного волокна составляет от 12% до 15% в расчете на 88% сухого вещества, 3% масла. Изобретение позволяет эффективно получать муку из темных семян канолы, обладающую благоприятными характеристиками. 2 н. и 3 з.п. ф-лы, 2 ил., 11 табл., 14 пр.
1. Мука канолы, полученная из темных семян канолы, где содержание белка муки канолы составляет от 43% до 44% в расчете на 88% сухого вещества, 3% масла.
2. Мука канолы по п. 1, где мука канолы имеет среднюю истинную метаболизируемую энергию от 2400 ккал/кг до 2635 ккал/кг.
3. Мука канолы по п. 1, где мука канолы содержит от 12% до 15% кислотного детергентного волокна в расчете на 88% сухого вещества, 3% масла.
4. Мука канолы по п. 1, где мука канолы содержит от 1,1% до 1,4% фосфора в расчете на 88% сухого вещества, 3% масла.
5. Мука канолы, полученная из темных семян канолы, где содержание белка муки канолы составляет от 43% до 44% и содержание кислотного детергентного волокна составляет от 12% до 15% в расчете на 88% сухого вещества, 3% масла.
| RAKOW, G | |||
| et al., RAPESEED GENETIC RESEARCH TO IMPROVE ITS AGRONOMIC PERFORMANCE AND SEED QUALITY, HELIA, 2007, 30, Nr | |||
| Способ изготовления звездочек для французской бороны-катка | 1922 |
|
SU46A1 |
| ПЕЧНОЙ ЖЕЛЕЗНЫЙ РУКАВ (ТРУБА) | 1920 |
|
SU199A1 |
| WO 2007016521 A2, 08.02.2007 | |||
| US 6433254 B1, 13.08.2002 | |||
| RU 2008141698 A, 27.04.2010. | |||

