ОБЛАСТЬ ТЕХНИКИ
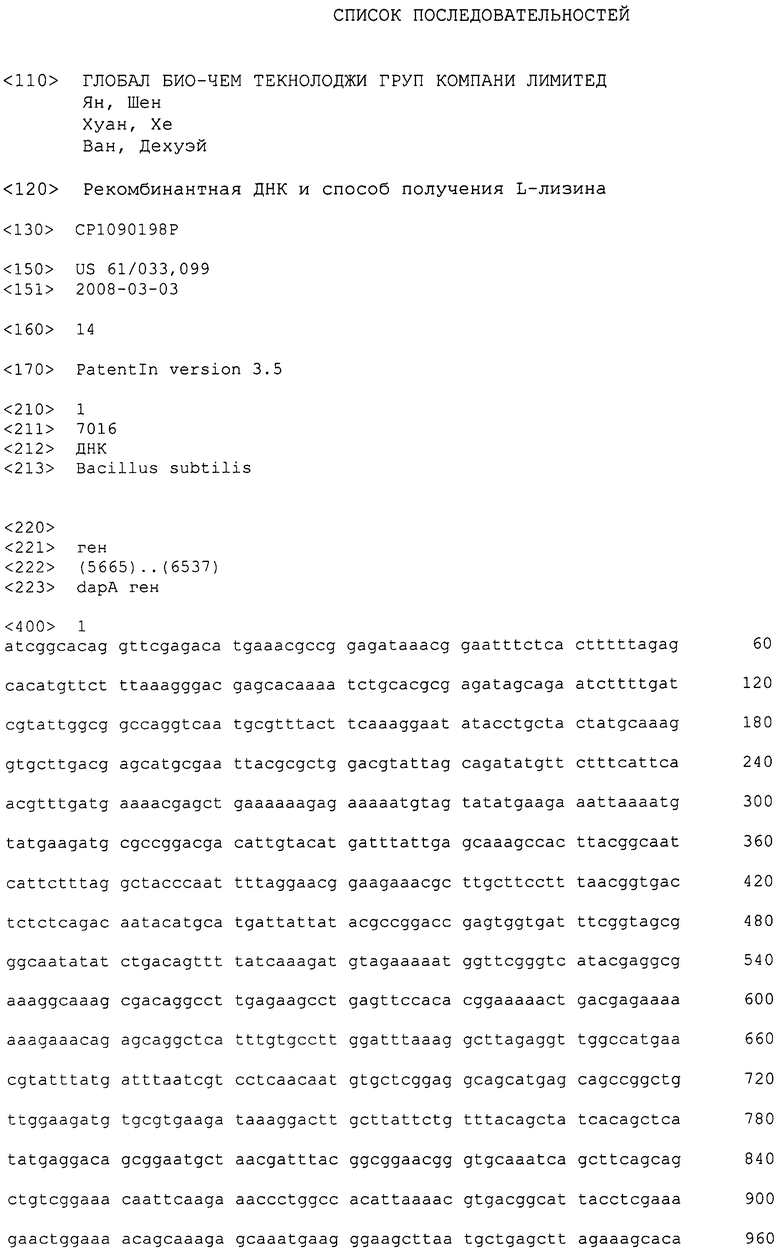
Настоящее изобретение относится к области биотехнологий. Изобретение состоит в разработке методов получения L-лизина путем выращивания трансформированной бактерии кишечной палочки, или эшерихии (Escherichia), в частности Е. coli, которая содержит дикий (немутантный) dapA ген или вариант этого гена сенной палочки (Bacillus subtilis).
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
L-лизин - незаменимая аминокислота. Она не синтезируется животными, однако было обнаружено много штаммов бактерий дикого и мутантного типов, способных ее производить. L-лизин широко используется как пищевая добавка, лекарственный препарат, химический агент и пищевой ингредиент. Массовое производство L-лизина для этих целей основано на крупномасштабной ферментации с использованием главным образом дифтериеподобных бактерий (Coryneform bacterium) или кишечной палочки (Escherichia bacterium).
У большинства бактерий L-лизин синтезируется естественным путем из аспартата в ходе ферментативного пути, включающего девять шагов, в том числе два шага, общих для пути биосинтеза метионина и треонина (Anastassiadis, S., Recent patents on Biotechnology 2007, 1(1):11-24; Chen, N. et al., J. Biol. Chem. 1993, 268(13):9448-66). Регуляторный механизм биосинтеза лизина достаточно сложен и варьируется в достаточно широких пределах у разных видов бактерий (Chen, N. et al., J. Biol Chem. 1993, 268(13):9448-66). Например, дигидродипиколинат синтеза (DDPS), фермент, который служит катализатором для первого шага в пути биосинтеза лизина, испытывает ингибирование по типу обратной связи L-лизином у грамотрицательных бактерий (например, Е. coli. Bacillus sphaericus и Methanobacterium thermoautotrophicum), но не у грамположительных бактерий (Bacillus lichenifbrmis, Bacillus megaterium. Bacillus subtilis, Corynebacterium glutamicum, Bacillus cereus и Bacillus lactofermentum) (Dobson, R. et al., Acta Cryst. 2005, D61:1 116-24). Управление путем биосинтеза лизина у Bacillus subtilis (В. subtilis) является уникальным, поскольку включает двойной контроль посредством лизина и одного из его прекурсоров, диаминопимелата (Chen, N. et al., J. Biol. Chem. 1993, 268(13):9448-66).
У штаммов бактерий различных родов наблюдается ограниченная гомология (сходство) последовательностей белка DDPS и его соответствующего гена, dapA, обусловленная различной степенью чувствительности к ингибированию DDPS L-лизином по типу обратной связи. DDPS у В. subtilis имеет последовательность аминокислот, на 43% и 40% идентичную такой последовательности у Е. coli и Corynebacterium Glutamicum (С. Glutamicum) соответственно. (Chen, N. et al., J. Biol. Chem. 1993, 268(13):9448-66). Даже в пределах одного рода разные штаммы бактерий демонстрируют гомологию в весьма скромных пределах. Так, ген dapA у Bacillus methanolicus (В. methanolicus) только на 65% идентичен ранее известному гену dapA у В. subtilis в отношении последовательности нуклеотидов или аминокислот (Патент США № 6,878,533).
Таким образом, единственный способ улучшить производство L-лизина бактерией Escherichia - преодолеть ингибирование DDPS L-лизином по типу обратной связи. Чтобы уменьшить восприимчивость DDPS к L-лизину, была предпринята попытка создать мутацию в гене dapA дикого типа у Escherichia bacterium (Патент США № 6,040,160). Кроме того, пробовали изъять ген dapA дикого типа у бактерии иного вида, чем Escherichia bacterium, у которого соответствующая DDPS не испытывает ингибирование L-лизином по типу обратной связи, и внедрить его в Escherichia bacterium, однако эта попытка не дала устойчивого результата.
Группой корейских исследователей сообщалось, что пересадка гена dapA дикого типа от штамма С. glutamicum, производящего лизин в избыточном количестве, в мутантный тип штамма Е. coli (TF1), производящего лизин в обычных количествах, приводило к двоякому эффекту: с одной стороны, увеличивалась чувствительность DDPS к лизину, а с другой - увеличивалось производство самого лизина (Oh, J. et al., Biotech. Ltrs. 1991, 13(10):727-32; Korean Pat. Pub. No. 10-1992-0008382). Однако внедрение такого же dapA гена дикого типа в два других штамма Е. coli (TF13 и TF23) не привело к увеличению выработки ими лизина. В качестве пояснения исследователи приводят довод, что у бактерий Е. coli более сложный регуляторный механизм биосинтеза лизина, чем у дифтериеподобных бактерий.
Желаемый результат может дать экспрессия чуждого dapA гена, так как высока вероятность того, что соответствующий чуждый DDPS протеин подвергается расщеплению под действием протеазы и формирует прочное внутриклеточное включение в бактерии Escherichia bacterium (Патент США № 6,040,160). Кроме того, надо полагать, что DDPS бактерий С. glutamicum (Oh, J. et al., Biotech. Ltrs., 1991, 13(10):727-32; Korean Pat. Pub. No. 15 10-1992-0008382) и В. methanolicus (Патент США № 6,878,533) не будет демонстрировать своей превосходящей активности, например, нечувствительной к лизину активности DDPS, которая приводит к высоким показателям производства лизина, в частности, у Е. coli, потому что оптимальная температура культивации для С. Glutamicum и В. methanolicus отличается от оптимальной температуры для Е. coli на 10 или более градусов.
Чрезвычайно сложная регуляция синтеза лизина наблюдалась в клетках Е. coli, в которые были экспрессированы гены, задействованные в биосинтезе лизина у В. subtilis (Shevchenko, T.N. et al., Tsitol Genet. 1984, 18(1):58-60). В частности, экспрессия этих чуждых генов, включая чуждый dapA ген, в клетки Е. coli не привела к удовлетворительному уровню увеличения производства лизина. Было выдвинуто предположение, что значительного увеличения биосинтеза лизина можно добиться, используя штаммы Е. coli или В. subtilis, имеющие мутации в природных генах, задействованных в процессе биосинтеза лизина, чтобы десенсибилизировать ингибирование по типу обратной связи, производимое лизином или диаминопимелатом.
К настоящему времени так и не предложено эффективной методики производства L-лизина на основе использования бактерий Escherichia, содержащих dapA ген дикого или модифицированного типа, взятый из В. subtilis. Учитывая стремительный рост населения во всем мире и постоянно возрастающую потребность в L-лизине, особенно для изготовления кормов для животных, назрела необходимость разработки новых, более эффективных методов получения L-лизина с использованием бактериальных штаммов Escherichia.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В соответствии с настоящим изобретением, внедрение дикого или вариантного dapA гена от бактерии В. subtilis в бактерию Escherichia улучшает производство L-лизина бактерией Escherichia до уровня, приемлемого для промышленных масштабов. Кроме того, трансформированная Escherichia может использоваться для производства L-лизин в среде культивирования с высокими результатами (не менее 25, 50, 75, 100, 125 или 150 граммов на литр).
Настоящее изобретение представляет собой рекомбинантную ДНК, автономно реплицирующуюся у бактерии Escherichia и содержащую dapA ген дикого или вариантного типа от В. subtilis. Вариантный dapA ген бактерии В. subtilis имеет последовательность, не полностью идентичную, но близкую к идентичной (например, на 90%, 95%, или 99%) последовательности dapA гена дикого типа у бактерии В. subtilis. Для внедрения dapA гена от В. subtilis в Escherichia, что способствует увеличению производства L-лизина, может быть использована рекомбинантная ДНК.
DapA ген бактерии В. subtilis может иметь последовательность нуклеиновых кислот, которая идентична или почти идентична (совпадение на 90%, 95% или 99%) последовательности dapA гена бактерии В. subtilis, как указано в базе данных GenBank, № последовательности L08471 (SEQ ID NO:1), и у Chen, N. et al., J. Biol. Chem. 1993, 268(13):9448-66). Вариантный dapA ген бактерии В. subtilis может содержать одну или более nucleotide modifications модификаций нуклеотидов в SEQ ID NO:1. В одном специфическом нелимитированном варианте, вариантный dapA ген бактерии В. subtilis содержит две мутации от G до Т в нуклеотидном остатке 355 и от Т до С в нуклеотидном остатке 360 SEQ ID NO:1.
Далее, dapA ген бактерии В. subtilis dapA может кодировать белок с последовательностью аминокислот, идентичной или почти идентичной (на 90%, 95% или 99%) предсказанной последовательности аминокислот dapA гена бактерии В. subtilis, как указано в базе данных GenBank, № последовательности L08471 (SEQ ID NO:2) и у Chen, N. et al., J. Biol. Chem. 1993, 268(13):9448-66). Белок может иметь последовательность аминокислот, содержащую одну или более модификаций аминокислот в SEQ ID NO:2. В одном специфическом нелимитированном варианте, вариантный dapA ген бактерии В. subtilis кодирует белок с последовательностью аминокислот, содержащей мутацию от гистидина до тирозина в аминокислотном остатке 119 SEQ ID NO:2 (вариант H119Y). DapA ген бактерии В. subtilis может кодировать белок, обладающий активностью относительно DDPS.
Данное изобретение также предоставляет бактерию Escherichia, которая содержит дикий или вариантный тип dapA гена бактерии В. subtilis, пригодную для производства L-лизина в среде для культивирования. Бактерия Escherichia способна производить L-лизин в количествах не менее 50 граммов на литр в среде для культивирования. В одном специфическом нелимитированном варианте, бактерия Escherichia, содержащая dapA ген дикого типа бактерии В. subtilis, используется для производства L-лизина. В другом специфическом нелимитированном варианте, бактерия Escherichia, содержащая вариант H119Y dapA гена бактерии В. subtilis, используется для производства L-лизина.
Настоящее изобретение далее дает методы производства L-лизина, которые состоят в выращивании бактерий Escherichia в среде для культивирования и сбора L-лизина из среды для культивирования, где бактерии содержат дикий или вариантный dapA ген бактерии В. subtilis. Сначала L-лизину дают накопиться, перед тем как собирать урожай или собирать его из среды для культивирования. Собирать L-лизин разрешается, когда его содержание в среде для культивирования достигнет по меньшей мере 25, 50, 75, 100, 125 или 150 граммов на литр, желательно не менее 50 граммов на литр. В одном специфическом нелимитированном варианте, бактерия Escherichia, содержащая dapA ген дикого типа бактерии В. subtilis, выращивается в среде для культивирования, и L-лизин собирают из среды для культивирования. В другом специфическом нелимитированном варианте, бактерия Escherichia, содержащая вариант H119Y dapA гена бактерии В. subtilis, выращивается в среде для культивирования, и L-лизин собирают из среды для культивирования.
ПОДРОБНОЕ ОПИСАНИЕ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение преимущественно в основном предлагает методы, трансформированную бактерию рода Escherichia и рекомбинантные ДНК для производства L-лизина путем ферментации. К настоящему времени открыто, что L-лизин можно производить с высоким выходом с использованием бактерии Escherichia, содержащей дикий или вариантный dapA ген бактерии В. subtilis.
Подробное описание изобретения для большей ясности можно условно разделить на следующие подразделы:
(1) рекомбинантная ДНК;
(2) трансформированная бактерия Escherichia;
(3) способ производства L-лизина.
(1) Рекомбинантная ДНК
Рекомбинантная ДНК настоящего изобретения несет в себе дикий или вариантный dapA ген штамма В. subtilis, и автономно реплицирует его в бактериальном штамме Escherichia. Рекомбинантная ДНК может быть получена путем вставки фрагмента ДНК, содержащего dapA ген бактерии В. subtilis, в экспрессионный вектор воспроизводимого (реплицируемого) бактериального штамма Escherichia.
DapA ген экспрессируют из штамма В. subtilis, и соответствующая DDPS не испытывает существенного (например, на 10%, 20%, 30%, 40%, 50%, 60% или более) ингибирования по типу обратной связи L-лизином. Выбранный штамм В. subtilis сам по себе может быть производителем L-лизина или не быть им. Предпочтительным штаммом В. subtilis является штамм W168, который не производит лизин. Данный штамм можно найти в Генетическом фонде палочковидных бактерий "Bacillus Genetic Stock Center" при Университете штата Огайо, США.
Фрагмент ДНК, содержащий dapA ген дикого типа бактерии В.subtilis (BsdapA ген) может быть получен из хромосомной ДНК штамма В.subtilis дикого типа. Хромосомная ДНК может быть получена из штамма В.subtilis при помощи общепринятых способов и технологий. Фрагмент ДНК может быть получен путем амплификации dapA гена дикого типа бактерии В. subtilis из хромосомной ДНК методом полимеразной цепной реакции (ПЦР). Подходящие ПЦР-праймеры могут быть получены способами, опубликованными ранее для последовательностей dapA гена бактерии В. subtilis или других бактериальных штаммов (Е. coli и С. glutamicum) (Chen, N. et al., J. Biol. Chem. 1993, 268(13); 9448-66). Например, можно использовать пару одноцепочечных 21-мер праймеров, DapA-F (SEQ ID NO:3) и DapA-R (SEQ ID NO:4).
Последовательности нуклеиновых кислот праймеров SEQ ID NO 3 и NO 4 представлены ниже:
Фрагмент ДНК, содержащий вариантный dapA ген бактерии В. subtilis, может быть получен аналогичным способом из хромосомной ДНК мутированного штамма В. subtilis in vivo. Вариантный dapA ген бактерии В. subtilis может быть получен также путем создания модификаций в диком dapA гене бактерии В. subtilis in vitro при помощи стандартных методов, известных специалистам в данной сфере, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез.
Последовательность результирующего фрагмента ДНК может быть определена общепринятыми методами с использованием подходящих праймеров (например, DapA-F и DapA-R).
DapA ген бактерии В. subtilis может иметь последовательность нуклеиновых кислот, идентичную или почти идентичную (на 90%, 95% или 99%) такой последовательности в DapA гене бактерии В. subtilis, как указано в базе данных GenBank, № последовательности L08471 (SEQ ID NO:1) и у Chen, N. et al., J. Biol. Chem. 1993, 268(13):9448-66). DapA ген дикого типа бактерии В. subtilis может иметь последовательность нуклеиновых кислот, идентичную SEQ ID NO:1. Вариантный DapA ген бактерии В. subtilis может содержать одну или более нуклеотидных модификаций в SEQ ID NO:1. Например, вариантный DapA ген бактерии В. subtilis может содержать две мутации от G до Т в нуклеотидном остатке 355 и от T до C в нуклеотидном остатке 360 в SEQ ID NO:1. Процентная идентичность последовательностей определяется при помощи стандартного программного обеспечения, такого как BLAST или FASTA.
Далее, dapA ген бактерии В. subtilis может кодировать белок, имеющий последовательность аминокислот, идентичную или почти идентичную (на 90%, 95% или 99%) предсказанной аминокислотной последовательности dapA гена бактерии В. subtilis, как указано в базе данных GenBank, № последовательности L08471 (SEQ ID NO:2) и у Chen, N. et al., J. Biol. Chem. 1993, 268(13):9448-66). DapA ген дикого типа бактерии В. subtilis может кодировать белок, имеющий последовательность аминокислот, идентичную SEQ ID NO:2. DapA ген бактерии В. subtilis может также кодировать белок, имеющий последовательность аминокислот, содержащую одну или более модификаций аминокислот в SEQ ID NO:2. Например, вариантный dapA ген бактерии В. subtilis может кодировать белок, имеющий последовательность аминокислот, содержащую мутацию от гистидина до тирозина в аминокислотном остатке 119 в SEQ ID NO:2 (вариант H119Y).
Модификации аминокислот включают замещения, присоединения и делеции. Модификации производятся с использованием стандартных методов, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Замещения аминокислот могут включать такие, в которых аминокислотный остаток заменяется другим аминокислотным остатком, имеющим подобную боковую цепь. Перечень семейств аминокислотных остатков, имеющих сходные боковые цепи, описан и известен профессионалам в данной сфере. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин); кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота); свободными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан); неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин); бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин).
Аминокислотные замещения также могут включать такие замещения, которые связаны с аминокислотными замещениями в dapA гене дикого типа бактерии Escherichia, которые обладают способностью десенситибилизировать соответствующую DDPS относительно L-лизина.
Фрагмент ДНК, содержащий дикий или вариантный dapA ген бактерии В. subtilis, может быть впоследствии дотирован (замкнут) с подходящим экспрессионным вектором для получения рекомбинантной ДНК, содержащей dapA ген. Подходящий экспрессионный вектор ДНК автономно реплицируется в бактериальном штамме Escherichia и содержит селектируемый генетический маркер. Селектируемый генетический маркер может продемонстрировать устойчивость к антибиотикам (например, ампициллину, тетрациклину, канамицину и неомицину), изменение цвета или любых других характеристик, по которым можно отличить трансформированные участки от нетрансформированных. Примеры подходящих векторов включают экспрессионные векторы Е. coli, такие как pTrc99A, pUC19, pUC18, pBR322, pHSG299, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219, pMW218 и pSTV28. Желательно, чтобы фрагмент ДНК внедрялся в экспрессионный вектор ДНК так, чтобы dapA ген находился под контролем сильного промотора Е. coli. Примеры подходящих промоторов включают trc, tac, lac и Т7, лучше всего trc.
DapA ген бактерии В. subtilis может кодировать белок, обладающий активностью в отношении DDPS. Присутствие рекомбинантной ДНК, содержащей dapA ген бактерии В. subtilis, может быть подтверждено возросшим уровнем активности относительно DDPS у бактериальных штаммов, трансформированных при помощи рекомбинантной ДНК, или обнаружением ауксотрофии у бактериальных штаммов, испытывающих недостаток DDPS (например, штамм Е. coli JE7627), трансформированных при помощи рекомбинантной ДНК.
(2) Трансформированная бактерия Escherichia
Бактерия Escherichia может быть трансформирована при помощи рекомбинантной ДНК, содержащей дикий или вариантный dapA ген бактерии В. subtilis, с использованием стандартных методов. Родительская (нетрансформированная) бактерия Escherichia может нести дикий или мутантный природный dapA ген, и экспрессировать соответствующую DDPS дикого или мутантного природного типа. Ферментативная активность природных DDPS может испытывать ингибирование по типу обратной связи L-лизином. Родительская бактерия Escherichia может быть производителем L-лизина. Активность другого природного фермента, участвующего в биосинтезе лизина, может быть повышена. Например, можно использовать штамм Е. coli, содержащий ДНК, кодирующую природную аспартокиназу III и имеющую мутации, для десенсибилизации к ингибированию L-лизином по типу обратной связи (Патент США №6040160). Предпочтительно использовать такой штамм бактерии Escherichia, как Е. coli. В дальнейших описаниях под предпочтительным штаммом бактерии Escherichia также подразумевается Е. coli как штамм, широко используемый для промышленного производства L-лизина.
После преобразования, бактерия Escherichia получает дикий или вариантный dapA ген бактерии В. subtilis. Наличие dapA гена бактерии В. subtilis в преобразованной бактерии определяется с помощью стандартных методов, известных специалистам в данной области. DapA ген бактерии В. subtilis может быть перенесен на плазмиду. Он также может быть интегрирован в хромосому трансформированной бактерии Escherichia. Трансформированная бактерия Escherichia производит L-лизин в достаточно больших количествах (не менее 25, 50, 75, 100, 125 или 150 г/л).
В одном варианте, штамм Е. coli B-3996 (доступен в Исследовательском институте генетики и промышленного скрещивания микроорганизмов, регистрационный номер RIA 1867) трансформируют с помощью рекомбинантной ДНК, содержащей dapA ген дикого типа (напр., pTrc99A-BsdapA), после удаления одиночной плазмиды pVIC40 (Патент США № 6,040,160), и получают трансформированный бактериальный штамм B-399/pTrc99A-BsdapA. Контрольный штамм В-399/pTrc99A получают таким же образом, с помощью трансформации соответствующей рекомбинантной ДНК без BsdapA (напр., pTrc99A). Для получения среды для культивирования стерилизованный раствор (содержащий 16 г/л ((NH4)2SO4, 1 г/л KH2PO4, 1 г/л MgSO4·7H2O, 0,01 г/л FeSO4·7H2O, 0,01 г/л MnSO4·5H2O, 2 г/л дрожжевого экстракта (Difco), 0,5 г/л L-метионина, 0,1 г/л L-треонина и 0,05 г/л L-изолейцина при рН 7,0) смешивают со стерилизованным 20%-ным раствором глюкозы в соотношении 4 к 1. Также можно добавить глюкозу для улучшения производства L-лизина.
Затем к смеси добавляют стерилизованный фармакопейный СаСО3 и разводят до конечной концентрации 30 г/л. Также можно добавить подходящие антибиотики (15 µг/мл тетрациклина и 5 µт/мл канамицина). Оба штамма культивируют, перемешивая со скоростью 114-116 об/мин при температуре 37°С в течение 48 ч. L-лизин отбирают из культивационной среды и анализируют. Как ожидается, бактериальный штамм В-399/pTrc99A-BsdapA даст не менее 10 г/л L-лизина, что значительно выше, чем может дать контрольный штамм В-399/pTrc99A.
В другом варианте, штамм бактерии Е. coli DC037, взятый в компании Global Bio-Chem Technology Group Company Limited, используется для приготовления рекомбинантной бактерии Е. coli, содержащей dapA ген дикого типа бактерии В. subtilis (DC051), рекомбинантной бактерии Е. coli, содержащей H119Y вариантный dapA ген бактерии В. subtilis (DC231) и контрольной рекомбинантной бактерии Е. coli (DC073), которые производят L-лизин в количествах 150, 180 и 20 г/л, соответственно. Процессы приготовления и тестирования этих рекомбинантных штаммов Е. coli описаны в Примерах 3 и 4.
Бактериальные штаммы DC051 и DC231 были зарегистрированы 26 февраля 2009 г. Китайским центром хранения культур общей микробиологии (China General Microbiological Culture Collection Center, CGMCC) согласно Будапештскому договору и получили номера CGMCC №2923 и CGMCC №2924, соответственно.
Активность DDPS трансформированной бактерии Escherichia может не испытывать существенного (на 10%, 20%, 30%, 40%, 50%, 60% или более) ингибирования L-лизином по типу обратной связи. Активность DDPS трансформированной бактерии Escherichia может быть уменьшена не более чем на 50% в присутствии 10 µм L-лизина.
Активность DDPS у бактерий измеряется по методу, описанному у Yugari, Y. и Gilvarg С. в J. Biol. Chem., 1965, 240(12):4710-16, или по любому другому приемлемому методу. Например, экстракт с бактериями добавляют в реакционный раствор, содержащий 50 µм имидазола-HCl (рН 7,4), 20 µм L-аспарагинового полуальдегида и 20 µм пирувата натрия, и инкубируют при 37°С в течение 10 мин. Реакционный раствор без пирувата натрия может быть использован как бланковый. Активность DDPS измеряется по количеству дигидродипиколината, образованного в ходе реакции, который отслеживается с помощью спектрофотометра при длине волны 270 нм. Различные количества L-лизина добавлялись к реакционной смеси для оценивания чувствительности к нему со стороны активности DDPS.
(3) Способ получения L-лизина
L-лизин может быть получен при выращивании бактерий Escherichia, содержащих дикий или вариантный dapA ген бактерии В. subtilis, в среде для культивирования. Желательно использовать среду, подходящую для оптимального роста бактерий Escherichia (Anastassiadis, S., Recent Patents On Biotechnology 2007, 1(1):11-24). Также желательно добавлять в среду антибиотики (например, ампициллин) для поддержания селективности и стабильности трансформированных бактерий Escherichia.
Например, среду для культивирования можно получить путем смешивания стерилизованного раствора (содержащего 16 г/л ((NH4)2SO4, 1 г/л KH2PO4, 1 г/л MgSO4·7H2O; 0,01 г/л FeSO4·7H2O, 0,01 г/л MnSO4·5H2O, 2 г/л дрожжевого экстракта (Difco), 0,5 г/л L-метионина, 0,1 г/л L-треонина и 0,05 г/л L-изолейцина при рН 7,0) со стерилизованным 20%-ным раствором глюкозы в соотношении 4 к 1. Также можно добавить глюкозу для улучшения производства L-лизина. Затем к смеси добавляют стерилизованный фармакопейный СаСО3 и разводят до конечной концентрации 30 г/л. Также можно добавить подходящие антибиотики (15 µг/мл тетрациклина и 5 µг/мл канамицина).
Желательно соблюдать оптимальные для Escherichia условия культивирования (Anastassiadis, S., Recent Patents On Biotechnology 2007, 1(1):11-24). Культивацию предпочтительно осуществлять в аэробных условиях при температуре между 25°С и 45°С и уровне рН между 5 и 8. Концентрация L-лизина достигает максимума через 2-10 дней после начала культивации. Рост бактерий отслеживается на основании плотности клеток в среде для культивирования, которая измеряется спектрофотометром.
Перед сбором L-лизину дают накопиться в среде для культивирования трансформированной бактерии Escherichia, выращиваемой в соответствии с настоящим изобретением. Сбор L-лизина осуществляется с использованием различных методов (например, ионообменных хроматографических методов) для получения L-лизин-HCl до или после сепарации клеток путем центрифугирования и фильтрации культивационной среды; также можно собирать L-лизиновый бульон, из которого получают L-лизин сульфат в процессе пеллетизации (гранулирования) (Anastassiadis, S., Recent Patents On Biotechnology 2007, 1(1):11-24; Патент КНР №20 200410050017.4). Собирать L-лизин из среды для культивирования разрешается, когда его концентрация достигает как минимум 5-10 г/л, либо - в крупномасштабных ферментационных процессах - не менее 25, 50, 75, 100, 125 или 150 г/л, лучше всего 50 г/л. Количество полученного L-лизина определяется с использованием общепринятых аналитических методов (напр., ВЭЖХ).
ПРИМЕРЫ
Итак, изобретение в общих чертах можно считать описанным. Чтобы его было легче понять, ниже приводятся ссылки на примеры, которые включены главным образом для иллюстрации некоторых аспектов и вариантов осуществления изобретения, хотя изобретение ни в коем случае не ограничивается ими.
Пример 1. Создание Плазмиды pTrc99A-BsdapA
Плазмида pTrc99A-BsdapA была создана для включения dapA гена дикого типа бактерии В. subtilis. Хромосомная ДНК подготовлена из дикого штамма В. subtilis W168 (Генетический фонд палочковидных бактерий при Университете штата Огайо, США) с использованием общепринятого метода. Фрагмент ДНК длиной 0.88 т.п.н. получен путем амплификации дикого dapA гена в хромосомную ДНК с использованием пары праймеров, DapA-F (SEQ ID NO:3) и DapA-R (SEQ ID NO:4), имеющих последовательности нуклеиновых кислот, показанные в SEQ ID NOS:3 и 4, соответственно. Фрагмент ДНК был восстановлен, расщеплен рестрикционными эндонуклеазами EcoRI and HindIII, и связан с лигандами pTrc99A плазмиды, предварительно расщепленными теми же эндонуклеазами рестрикции для получения pTrc99A-BsdapA плазмиды. Плазмида pTrc99A-BsdapA содержит последовательность нуклеиновых кислот, как в SEQ ID NO:1. Последовательность нуклеиновых кислот кодирует белок, имеющий последовательность аминокислот, как в SEQ ID NO:2.
Пример 2. Создание Плазмиды pTrc99A-BsdapAH119Y
Плазмида pTrc99A-BsdapAH119Y была создана для включения вариантного H119Y dapA гена бактерии В. subtilis. Мутация произведена с использованием pTrc99A-BsdapA плазмиды в качестве матрицы ПЦР и амплифицирована парой праймеров bsdapa119muts (SEQ ID NO:5) и bsdapa119mutas (SEQ ID NO:6). Последовательности нуклеиновых кислот SEQ ID NOS:5 и 6 приведены ниже:
Сайты мутации выделены жирным курсивом.
Компетентная клетка Е. coli DH5α трансформирована аликвотами ПЦР продукта, перенесенного посредством DpnI. Плазмида экстрагирована из трансформантов и перенесена рестрикционной эндонуклеазой Dral для идентификации желаемой мутантной плазмиды pTrc99A-BsdapAH119Y, которая содержит последовательность нуклеиновых кислот с двумя мутациями от G до Т в нуклеотидном остатке 355 и от Т до С в нуклеотидном остатке 360 в SEQ ID NO:1. Последовательность нуклеиновых кислот кодирует белок, имеющий последовательность аминокислот, содержащую мутацию от гистидина до тирозина в аминокислотном остатке 119 в SEQ ID NO:2.
Пример 3. Рекобинантная Е. coli, содержащая dapA ген дикого типа бактерии В. subtilis
Рекобинантная Е. coli, содержащая dapA ген дикого типа бактерии В. subtilis (DC051), получена из штамма бактерии Е. coli DC037, взятого в "Глобал биохим технолоджи Груп". Штамм DC037 несет две различные плазмиды, каждая из которых содержит мутантный dapA ген бактерии Е. coli и ген резистентности к тетрациклину или канамицину. Эти две плазмиды изъяты, а на их место поставлены плазмиды pDCtetBSdapA и pDCkanBSdapA, каждая из которых содержит BsdapA ген и ген резистентности к терациклину или канамицину, как изложено ниже.
Сконструирована плазмида pDCtetBSdapA, содержащая BsdapA ген и ген резистентности к терациклину. BsdapA получен из pTrc99A-BsdapA с использованием пары праймеров, ptrcBSdapA1-F и ptrcBSdapA1-R, имеющих последовательности нуклеиновых кислот, описанные для SEQ ID NOS:7 и 8, соответственно (см. ниже). Фрагмент ДНК из 1.6 т.п.н. был обнаружен, перенесен рестрикционной эндонуклеазой Tth111I и SpeI, и связан с лигандами плазмиды pDCtetdapA, которая была предварительно перенесена теми же рестрикционными эндонуклеазами, для получения плазмиды pDCtetBSdapA.
Была сконструирована плазмида pDCkanBSdapA, содержащая BsdapA ген и ген резистентности к канамицину. BsdapA получен из pTrc99A-BsdapA с использованием пары праймеров, ptrcBSdapA2-F и ptrcBSdapA2-R, имеющих последовательности нуклеиновых кислот, описанные для SEQ ID NOS:9 и 10, соответственно (см. ниже). Фрагмент ДНК из 1.6 т.п.н. был обнаружен и перенесен рестрикционными эндонуклеазами NotI и PshAI, и связан с лигандами плазмиды pDCkandapA, которая была предварительно перенесена теми же рестрикционными эндонуклеазами, для получения плазмиды pDCkanBSdapA.
DC045 получен из DC037. На DC037 воздействуют 800 µг/мл ЕВ в течение 24 ч, затем процеживают с 5 µг/мл тетрациклина и 50 µг/мл канамицина, соответственно, в результате получают DC039, где pDCkandapA устранен, а pDCtetdapA оставлен.
Фрагмент длиной 2.8 т.п.н. получен путем амплификации Е. coli W3110 с использованием пары праймеров, LDCup и flDN, имеющих последовательности нуклеиновых кислот, описанные для SEQ ID NOS:11 и 12, соответственно (см. ниже). Данный фрагмент субклонирован в pMD18simple Т вектор для получения pMD18swtLDC.
Ген резистентности к апрамицину длиной 1,5 т.п.н. получен путем амплификации pIJ773 с использованием пары праймеров, FRT5 и HPA1FRT3, имеющих последовательности нуклеиновых кислот, описанные для SEQ ID NOS:13 и 14, соответственно (см. ниже). pMD18swtLDC перенесен с помощью HpaI как вектор и связан с лигандами с фрагментом гена резистентности к апрамицину длиной 1,5 т.п.н., в результате получен pMD18swtLDC-apra. Фрагмент длиной 4,5 т.п.н. изолирован путем перенесения pMD 18swtLDC-apra с PvuII и использован в качестве рекомбинантного фрагмента для трансформирования DC039(pIJ790) и получения DC043, где LDC дикого типа заменен "укороченным" LDC в геноме DC039. DC045, содержащий LDC дикого типа и плазмиду pDCtetdapA, получен после элиминации гена резистентности к апрамицину из генома DC043 с помощью плазмиды рСР20.
Затем была создана pDCamBSdapA с целью оттолкнуть плазмиду pDCtetdapA в DC045.
Плазмида pIJ773 была экстрагирована и дигерирована рестрикционными эндонуклеазами EcoRI и Clal. Фрагмент длиной 1,3 т.п.н. был изолирован. Фрагмент длиной 3.4 т.п.н. получен после дигерирования pDCtetBSdapA рестрикционными эндонуклеазами PvuII и затем связан с лигандами с плазмидой pMD18simple, которая предварительно была дигерирована теми же рестрикционными эндонуклеазами для получения pMD18s(BStet). pMD18s(BStet) дигерирована рестрикционными эндонуклеазами EcoRI и ClaI для получения фрагмента длиной 4,5 т.п.н., который будет использован в качестве вектора. Фрагмент длиной 1,3 т.п.н., полученный из pIJ773, был связан с лигандами с фрагментом вектора длиной 4,5 т.п.н. Компетентные клетки DH5α трансформированы лигандной смесью и выращены в чашках с добавлением ампициллина/апрамицина. Плазмида ДНК экстрагирована из трансформированной бактерии и дигерирована рестрикционными эндонуклеазами PvuII, и фрагмент длиной 3,5 т.п.н. был получен в виде рекомбинантного фрагмента с длинным плечом.
BW25113(pIJ790), трансформированный при помощи pDCtetBSdapA, подвергся дальнейшей трансформации рекомбинантным фрагментом с длинным плечом. Рекомбинантная плазмида pDCamBSdapA экстрагирована из трансформированной бактерии и использована для трансформирования DC045. Трансформированный штамм DC045 содержит две плазмиды, pDCtetdapA и pDCamBSdapA. В связи с отталкиванием двух источников ауторепликации, получен DC049-1, где pDCtetdapA удалена, а pDCamBSdapA оставлена. DC051 получен после трансформирования DC049-1 с помощью pDCtetBSdapA и pDCkanBSdapA.
Бактериальный штамм DC051 был зарегистрирован Китайским центром хранения культур общей микробиологии (China General Microbiological Culture Collection Center, CGMCC) 26 февраля 2009 г. согласно Будапештскому договору и получил номер CGMCC No. 2923.
Штамм DC073 с экспрессионным вектором pTrc99A получен таким же способом, как и контрольный штамм.
В отличие от DC051, DC073 не содержит pBsdapA.
Последовательности нуклеиновых кислот в SEQ ID NOS:7-14 приведены ниже:
Штаммы DC051 и DC073 выращивались в среде для культивирования с добавлением ампициллина при 37°С в течение 3 дней и произвели L-лизин в среде в количествах 150 и 20 г/л, соответственно.
Пример 4. Рекомбинантная бактерия Е. coli, содержащая вариант H119Y dapA гена бактерии В. subtilis
Рекомбинантная бактерия Е. coli, содержащая вариант H119Y dapA гена бактерии В. subtilis (DC231), получена тем же методом, что и в Примере 3, с той лишь разницей, что для замещения плазмиды pTrc99A-BsdapA была использована плазмида pTrc99A-BsdapAH119Y. Штамм DC231 получен после трансформирования DC049-1 с помощью pDCkanBSdapAH119Y и pDCtetBSdapAH119Y.
Бактериальный штамм DC231 был зарегистрирован Китайским центром хранения культур общей микробиологии (China General Microbiological Culture Collection Center, CGMCC) согласно Будапештскому договору 26 февраля 2009 г. и получил номер CGMCC No. 2924.
Штамм DC231 выращен в среде для культивирования по методу, описанному в Примере 3. L-лизин получен в среде в количестве 180 г/л.
Все публикации и патенты, упомянутые в данном описании, включены в данный документ посредством ссылки целиком и полностью, как если бы каждая отдельная публикация или патент были конкретно и индивидуально предназначены для включения в качестве ссылки. В случае спорных вопросов, следует руководствоваться настоящим приложением, включая любые определения, приведенные в нем.
Несмотря на то, что конкретные варианты изобретения обсуждены, приведенное выше описание носит иллюстративный характер и не является ограничивающим. Для специалистов очевидно, что данное изобретение может быть воплощено в нескольких вариациях, как видно из описания и формулы изобретения, приведенной ниже.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФЕРМЕНТАЦИОННЫЙ СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1994 |
|
RU2204605C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2351653C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2250266C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ Methylophilus И СПОСОБ ПОЛУЧЕНИЯ БАКТЕРИАЛЬНЫХ КЛЕТОК Methylophilus | 2000 |
|
RU2334794C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА (ВАРИАНТЫ) И РЕКОМБИНАНТНАЯ ДНК, ИСПОЛЬЗУЕМАЯ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ (ВАРИАНТЫ) | 1996 |
|
RU2197528C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ 1-БУТАНОЛА | 2006 |
|
RU2429295C2 |








Изобретение относится к области биотехнологии. Изобретение касается способа получения L-лизина с использованием бактерии Escherichia coli, содержащей варианты dapA гена бактерии В. subtilis. Изобретение позволяет получить L-лизин с высокой степенью эффективности. 3 н. и 3 з.п. ф-лы, 4 пр.
1. Рекомбинантная ДНК, автономно воспроизводящаяся в бактерии Escherichia coli, содержащая dapA ген бактерии B.subtilis, где:
dapA ген имеет последовательность нуклеиновых кислот, представленную в SEQ ID NO:1, в которой нуклеотидный остаток G в положении 355 заменен на Т и нуклеотидный остаток Т в положении 360 заменен на С;
или
dapA ген имеет последовательность нуклеиновых кислот, которая кодирует белок, имеющий последовательность аминокислот, представленную в SEQ ID NO:2, в которой аминокислотный остаток гистидин в положении 119 заменен на тирозин.
2. Бактерия Escherichia coli, содержащая dapA ген бактерии B.subtilis, продуцирующая L-лизин, где dapA ген имеет последовательность нуклеиновых кислот, которая кодирует белок, имеющий последовательность аминокислот, представленную в SEQ ID NO:2, в которой аминокислотный остаток гистидин в положении 119 заменен на тирозин.
3. Бактерия по п.2, зарегистрированная под номером CGMCC No.2923 в Китайском центре хранения культур общей микробиологии.
4. Бактерия по п.2, зарегистрированная под номером CGMCC No.2924 в Китайском центре хранения культур общей микробиологии.
5. Бактерия по п.2, которая продуцирует L-лизин в количестве минимум 50 граммов на литр в среде для культивирования.
6. Способ получения L-лизина, включающий выращивание бактерии Escherichia coli, содержащей dapA ген бактерии B.subtilis, в среде для культивирования, и сбор L-лизина из среды для культивирования, где dapA ген имеет последовательность нуклеиновых кислот, которая кодирует белок, имеющий последовательность аминокислот, представленную в SEQ ID NO:2, в которой аминокислотный остаток гистидин в положении 119 заменен на тирозин.
| ШЕВЧЕНКО Т.Н., ОКУНЕВ О.В., АЛЕКСИЕВА З.М., МАЛЮТА С.С | |||
| Экспрессия генов биосинтеза лизина Bacillus subtilis в клетках Escherichia coli | |||
| - Цитология и генетика, 1984, т.18, №1, с.58-60 | |||
| CHEN N.Y., JIANG S.Q., KLEIN D.A., PAULUS H | |||
| Organization and nucleotide sequence of the Bacillus subtilis diaminopimelate operon, a cluster of genes encoding |

