ОБЛАСТЬ ТЕХНИКИ
Данное изобретение имеет отношение к способам в области микробиологической промышленности. В частности, данное изобретение относится к способу производства L-аминокислоты посредством ферментации и к микроорганизму, используемому в данном способе.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Аминокислоты, такие как L-лизин, L-глутаминовая кислота, L-треонин, L-лейцин, L-изолейцин, L-валин и L-фенилаланин, получают промышленным путем посредством ферментации с использованием микроорганизмов, которые относятся к родам Brevibacterium, Corynebacterium, Bacillus, Escherichia, Streptomyces, Pseudomonas, Arthrobacter, Serratia, Penicillium, Candida и им подобным. Для того чтобы повысить продуктивность, в качестве таких микроорганизмов использовали штаммы, выделенные в природе, или их искусственные мутанты. Были заявлены различные способы усиления активностей ферментов биосинтеза L-глутаминовой кислоты с использованием технологии рекомбинантной ДНК, чтобы повысить способность продуцировать L-глутаминовую кислоту.
Продуктивность при получении L-аминокислот была значительно повышена с помощью селекции микроорганизмов, таких как микроорганизмы, приведенные выше, и совершенствования способов производства. Однако чтобы удовлетворить дальнейшее увеличение потребностей в будущем, по-прежнему требуется развитие способов более эффективного производства L-аминокислот.
В качестве способов получения аминокислот ферментацией метанола, который представляет собой сырье для ферментации, доступное в больших количествах при низкой стоимости, имеются традиционно известные способы с использованием микроорганизмов, которые относятся к роду Achromobacter или Pseudomonas (заявка на патент Японии (Kokoku) No. 45-25273/1970), Protaminobacter (опубликованная заявка на патент Японии (Kokai) No. 49-125590/1974), Protaminobacter или Methanomonas (опубликованная заявка на патент Японии (Kokai) No. 50-25790/1975), Microcyclus (опубликованная заявка на патент Японии (Kokai) No. 52-18886/1977), Methylobacillus (опубликованная заявка на патент Японии (Kokai) No. 4-91793/1992), Bacillus (опубликованная заявка на патент Японии (Kokai) No. 3-505284/1991) и так далее.
Однако до настоящего времени не был известен способ получения L-аминокислот с использованием бактерий Methylophilus. Хотя способы, описанные в EP 0 035 831 A, EP 0 037 273 A и EP 0 066 994 A, были известны как способы трансформации бактерий Methylophilus с использованием рекомбинантной ДНК, применение технологии рекомбинантной ДНК для повышения продуцирования аминокислот бактериями Methylophilus не было известно.
РАСКРЫТИЕ СУТИ ИЗОБРЕТЕНИЯ
Целью согласно изобретению является предоставление новой бактерии, продуцирующей L-аминокислоту, и способа получения L-аминокислоты с использованием бактерии, продуцирующей L-аминокислоту. В результате попыток авторов изобретения, посвященных достижению упомянутой выше цели, было обнаружено, что бактерии Methylophilus пригодны для получения L-аминокислот. Кроме того, хотя традиционно считается, что трудно получить ауксотрофные мутанты бактерий Methylophilus (FEMS Microbiology Rev. 39, 235-258 (1986) и Antonie van Leeuwenhoek 53, 47-53 (1987), авторы данного изобретения достигли успеха в получении ауксотрофных мутантов указанных бактерий. Таким образом, данное изобретение было осуществлено.
То есть данное изобретение предоставляет следующее.
(1) Бактерия Methylophilus, обладающая способностью продуцировать L-аминокислоту.
(2) Бактерия Methylophilus по п.(1), где L-аминокислотой является L-лизин, L-валин, L-лейцин, L-изолейцин или L-треонин.
(3) Бактерия Methylophilus по п.(1), которая обладает резистентностью к аналогу L-аминокислоты или ауксотрофией по L-аминокислоте.
(4) Бактерия Methylophilus по п.(1), у которой повышена активность ферментов биосинтеза L-аминокислоты.
(5) Бактерия Methylophilus по п.(1), у которой повышены активность дигидродипиколинатсинтазы и активность аспартокиназы, и при этом бактерия обладает способностью продуцировать L-лизин.
(6) Бактерия Methylophilus по п.(1), у которой повышена активность дигидродипиколинатсинтазы, и при этом бактерия обладает способностью продуцировать L-лизин.
(7) Бактерия Methylophilus по п.(1), у которой повышена активность аспартокиназы, и при этом бактерия обладает способностью продуцировать L-лизин.
(8) Бактерия Methylophilus по любому из п.п. с (5) по (7), у которой активность или активности одного, двух или трех ферментов, выбранных из дегидрогеназы полуальдегида аспарагиновой кислоты, дигидродипиколинатредуктазы и диаминопимелатдекарбоксилазы, усилена/усилены.
(9) Бактерия Methylophilus по п.(5), у которой активность дигидродипиколинатсинтазы и активность аспартокиназы повышены путем трансформации посредством введения в клетки ДНК, кодирующей дигидродипиколинатсинтазу, которая не подвергается ингибированию L-лизином по принципу обратной связи, и ДНК, кодирующей аспартокиназу, которая не подвергается ингибированию L-лизином по принципу обратной связи.
(10) Бактерия по п.(1), где активности аспартокиназы, гомосериндегидрогеназы, гомосеринкиназы и треонинсинтазы повышены, и бактерия обладает способностью продуцировать L-треонин.
(11) Бактерия по любому из п.п. с (1) по (10), где бактерией Methylophilus является Methylophilus methylotrophus.
(12) Способ получения L-аминокислоты, который включает в себя культивирование бактерии Methylophilus, которая охарактеризована в любом из указанных выше п.п. с (1) по (11), в среде, чтобы продуцировать и накопить L-аминокислоту в культуре, и сбор L-аминокислоты из культуры.
(13) Способ по п.(12), при котором среда содержит метанол в качестве основного источника углерода.
(14) Способ получения бактериальных клеток бактерии Methylophilus с повышенным содержанием L-аминокислоты, который включает в себя культивирование бактерии Methylophilus, охарактеризованной в любом из указанных выше пунктов с (1) по (11), в среде, чтобы продуцировать и накопить L-аминокислоту в бактериальных клетках данной бактерии.
(15) Способ получения бактериальных клеток бактерии Methylophilus по п.14, где L-аминокислотой является L-лизин, L-валин, L-лейцин, L-изолейцин или L-треонин.
(16) ДНК, которая кодирует белок, охарактеризованный в следующих пунктах (А) или (B):
(А) белок, который имеет аминокислотную последовательность SEQ ID NO: 6, или
(В) белок, который имеет аминокислотные последовательности SEQ ID NO: 6, включая замену, делецию, инсерцию, присоединение или инверсию одной или нескольких аминокислот, и обладает активностью аспартокиназы.
(17) ДНК по п.(16), которая представляет собой ДНК, охарактеризованную в следующих пунктах (а) или (b):
(а) ДНК, которая имеет нуклеотидную последовательность, включающую в себя нуклеотидную последовательность от нуклеотида номер 510 до нуклеотида номер 1736 SEQ ID NO: 5; или
(b) ДНК, которая способна гибридизоваться с пробой, имеющей нуклеотидную последовательность от нуклеотида номер 510 до нуклеотида номер 1736 SEQ ID NO: 5, или ее часть, в жестких условиях, и кодирует белок, обладающий активностью аспартокиназы.
(18) ДНК, которая кодирует белок, охарактеризованный в следующих пунктах (С) или (D):
(С) белок, который имеет аминокислотную последовательность SEQ ID NO: 8, или
(D) белок, который имеет аминокислотные последовательности SEQ ID NO: 8, включая замену, делецию, инсерцию, присоединение или инверсию одной или нескольких аминокислот, и обладает активностью дегидрогеназы полуальдегида аспарагиновой кислоты.
(19) ДНК по п.(18), которая представляет собой ДНК, охарактеризованную в следующих пунктах (с) или (d):
(с) ДНК, которая имеет нуклеотидную последовательность, включающую в себя нуклеотидную последовательность от нуклеотида номер 98 до нуклеотида номер 1207 SEQ ID NO: 7; или
(d) ДНК, которая гибридизуется с пробой, имеющей нуклеотидную последовательность от нуклеотида номер 98 до нуклеотида номер 1207 SEQ ID NO: 7 или ее часть, в жестких условиях, и кодирует белок, обладающий активностью дегидрогеназы полуальдегида аспарагиновой кислоты.
(20) ДНК, которая кодирует белок, охарактеризованный в следующих пунктах (E) или (F):
(E) белок, который имеет аминокислотную последовательность SEQ ID NO: 10, или
(F) белок, который имеет аминокислотные последовательности SEQ ID NO: 10, включая замену, делецию, инсерцию, присоединение или инверсию одной или нескольких аминокислот, и обладает активностью дигидродипиколинатсинтазы.
(21) ДНК по п.(20), которая представляет собой ДНК, охарактеризованную в следующих пунктах (e) или (f):
(e) ДНК, которая имеет нуклеотидную последовательность, включающую в себя нуклеотидную последовательность от нуклеотида номер 1268 до нуклеотида номер 2155 SEQ ID NO: 9; или
(f) ДНК, которая гибридизуется с пробой, имеющей нуклеотидную последовательность от нуклеотида номер 1268 до нуклеотида номер 2155 SEQ ID NO: 9 или ее часть, в жестких условиях, и кодирует белок, обладающий активностью дигидродипиколинатсинтазы.
(22) ДНК, которая кодирует белок, охарактеризованный в следующих пунктах (G) или (H):
(G) белок, который имеет аминокислотную последовательность SEQ ID NO: 12, или
(H) белок, который имеет аминокислотные последовательности SEQ ID NO: 12, включая замену, делецию, инсерцию, присоединение или инверсию одной или нескольких аминокислот, и обладает активностью дигидродипиколинатредуктазы.
(23) ДНК по п.(22), которая представляет собой ДНК, охарактеризованную в следующих пунктах (g) или (h):
(g) ДНК, которая имеет нуклеотидную последовательность, включающую в себя нуклеотидную последовательность от нуклеотида номер 2080 до нуклеотида номер 2883 SEQ ID NO: 11; или
(h) ДНК, которая гибридизуется с пробой, имеющей нуклеотидную последовательность от нуклеотида номер 2080 до нуклеотида номер 2883 SEQ ID NO: 11 или ее часть, в жестких условиях, и кодирует белок, обладающий активностью дигидродипиколинатредуктазы.
(24) ДНК, которая кодирует белок, охарактеризованный в следующих пунктах (I) или (J):
(I) белок, который имеет аминокислотную последовательность SEQ ID NO: 14, или
(J) белок, который имеет аминокислотные последовательности SEQ ID NO: 14, включая замену, делецию, инсерцию, присоединение или инверсию одной или нескольких аминокислот, и обладает активностью диаминопимелатдекарбоксилазы.
(25) ДНК по п.(24), которая представляет собой ДНК, охарактеризованную в следующих пунктах (i) или (j):
(i) ДНК, которая имеет нуклеотидную последовательность, включающую в себя нуклеотидную последовательность от нуклеотида номер 751 до нуклеотида номер 1995 SEQ ID NO: 13; или
(j) ДНК, которая гибридизуется с пробой, имеющей нуклеотидную последовательность от нуклеотида номер 751 до нуклеотида номер 1995 SEQ ID NO: 13 или ее часть, в жестких условиях, и кодирует белок, обладающий активностью диаминопимелатдекарбоксилазы.
В данном описании «способность продуцировать L-аминокислоту» относится к способности накапливать значительное количество L-аминокислоты в среде или к увеличению содержания аминокислоты в бактериальных клетках, в том случае, когда микроорганизм согласно изобретению культивируют в среде.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показан способ получения плазмиды RSF24P, имеющей мутантный dapA. «dapA*24» относится к мутантному dapA, который кодирует мутантный фермент DDPS, в котором остаток гистидина 118 заменен остатком тирозина.
На фиг.2 показан способ получения плазмиды RSFD80, имеющей мутантный dapA и мутантный lysC. «lysC*80» относится к мутантному lysC, который кодирует мутантную AKIII, где остаток треонина 352 заменен остатком изолейцина.
На фиг.3 показана аспартокиназная активность трансформированных штаммов E.coli, содержащих ген ask.
На фиг.4 показана активность дегидрогеназы полуальдегида аспарагиновой кислоты трансформированных штаммов E.coli, содержащих ген asd.
На фиг.5 показана дигидродипиколинатсинтазная активность трансформированных штаммов E.coli, содержащих ген dapA.
На фиг.6 показана дигидродипиколинатредуктазная активность трансформированного штамма E.coli, содержащего ген dapB.
На фиг.7 показана диаминопимелатдекарбоксилазная активность трансформированных штаммов E.coli, содержащих ген lysA.
НАИЛУЧШИЙ СПОСОБ ВЫПОЛНЕНИЯ ИЗОБРЕТЕНИЯ
<1> Микроорганизм данного изобретения
Микроорганизмом согласно изобретению является бактерия, относящаяся к роду Methylophilus и обладающая способностью продуцировать L-аминокислоту. Бактерия Methylophilus согласно изобретению включает в себя, например, штамм Methylophilus methylotrophus AS1 (NCIMB10515) и так далее. Штамм Methylophilus methylotrophus AS1 (NCIMB10515) доступен для приобретения из национальных коллекций промышленных и морских бактерий (National Collections of Industrial and Marine Bacteria) (адрес: NCIMB Lts., Torry Research Station 135, Abbey Road, Aberdeen AB9 8DG, United Kingdom).
L-аминокислоты, полученные согласно изобретению, включают в себя L-лизин, L-глутаминовую кислоту, L-треонин, L-валин, L-лейцин, L-изолейцин, L-триптофан, L-фенилаланин, L-тирозин и так далее. Можно получать один или большее количество типов таких аминокислот.
Бактерии Methylophilus, обладающие способностью продуцировать L-аминокислоты, могут быть получены посредством придания способности продуцировать L-аминокислоты штаммам бактерий Methylophilus дикого типа. Чтобы придать способность продуцировать L-аминокислоты, могут быть использованы способы, традиционно принятые в селекции коринеформных бактерий, бактерий Escherichia и им подобных, такие как способы получения ауксотрофных мутантных штаммов, штаммов, резистентных к аналогам L-аминокислот, или мутантных штаммов в отношении контроля метаболических процессов, и способы получения рекомбинантных штаммов, в которых повышены активности ферментов биосинтеза L-аминокислот (смотри "Amino Acid Fermentation" the Japan Scientific Societies Press [Gakkai Shuppan Center], 1st Edition, опубликованная 30 мая, 1986, pp. 77-100). При селекции бактерий, продуцирующих аминокислоты, такие характеристики, как ауксотрофия, резистентность к аналогу L-аминокислоты и мутация метаболического контроля, можно придать бактериям отдельно или в комбинации из двух или большего количества характеристик. Активность ферментов биосинтеза L-аминокислот может быть усилена для каждого в отдельности или для комбинации одного или большего количества ферментов. Кроме того, придание таких характеристик, как ауксотрофия, резистентность к аналогу L-аминокислоты и мутация метаболического контроля, можно комбинировать с усилением активности ферментов биосинтеза L-аминокислот.
Например, бактерии, продуцирующие L-лизин, получены при селекции как мутанты, проявляющие ауксотрофию по L-гомосерину или L-треонину и L-метионину (заявка на патент Японии (Kokoku) No. 48-28078/1973 и 56-6499/1981), мутанты, проявляющие ауксотрофию по инозиту или уксусной кислоте (опубликованная заявка на патент Японии (Kokai) No. 55-9784/1980 и 56-8692/1981), или мутанты, которые устойчивы к оксализину, лизингидроксамату, S-(2-аминоэтил)цистеину, γ-метиллизину, α-хлоркапролактаму, DL-α-амино-ε-капролактаму, α-аминолауриллактаму, аналогу аспарагиновой кислоты, сульфамидному лекарственному препарату, хиноиду или N-лауроиллейцину.
Кроме того, бактерии, продуцирующие L-глутаминовую кислоту, могут быть получены при селекции как мутанты, проявляющие ауксотрофию по олеиновой кислоте или ей подобному. Бактерии, продуцирующие L-треонин, могут быть получены при селекции как мутанты, устойчивые к α-амино-β-гидроксивалериановой кислоте. Бактерии, продуцирующие L-гомосерин, могут быть получены при селекции как мутанты, проявляющие ауксотрофию по L-треонину, или мутанты, устойчивые к аналогам L-фенилаланина. Бактерии, продуцирующие L-фенилаланин, могут быть получены при селекции как мутанты, проявляющие ауксотрофию по L-тирозину. Бактерии, продуцирующие L-изолейцин, могут быть получены при селекции как мутанты, проявляющие ауксотрофию по L-лейцину. Бактерии, продуцирующие L-пролин, могут быть получены при селекции как мутанты, проявляющие ауксотрофию по L-изолейцину.
Более того, как указано в примерах далее, штаммы, которые продуцируют один или большее количество видов разветвленных аминокислот (L-валин, L-лейцин и L-изолейцин), могут быть получены как штаммы, проявляющие ауксотрофию по казаминовой кислоте.
Чтобы получить мутанты бактерий Methylophilus, авторы изобретения сначала изучили детали оптимальных условий мутагенеза с использованием в качестве показателя частоты появления штаммов, резистентных к стрептомицину. В результате максимальная частота появления резистентных к стрептомицину штаммов была получена в том случае, когда степень выживаемости после мутагенеза составляла примерно 0,5%, и авторы достигли цели в получении ауксотрофных штаммов при данных условиях. Авторы также достигли успеха в получении ауксотрофных штаммов, получить которые считалось трудным, посредством значительного увеличения масштаба скрининга мутантов, по сравнению с проводимым ранее для E.coli и так далее.
Как описано выше, поскольку было обнаружено, что мутанты можно получить путем мутагенеза бактерий Methylophilus в соответствующих условиях, стало возможным легко получать желаемые мутанты, выбирая такие подходящие условия, при которых степень выживаемости после мутагенеза должна соответствовать примерно 0,5%, в зависимости от способа мутагенеза.
Способы мутагенеза для получения мутантов бактерий Methylophilus включают в себя УФ-облучение и обработку мутагенными средствами, используемыми при обычных мутагенных обработках, такими как N-метил-N'-нитро-N-нитрозогуанидин (NTG) и азотистая кислота. Бактерии Methylophilus, обладающие способностью продуцировать L-аминокислоты, также можно получить путем селекции мутантов бактерий Methylophilus природного происхождения.
Мутанты, устойчивые к аналогам L-аминокислот, можно получить, например, инокулируя мутантные бактерии Methylophilus на агаризованную среду, содержащую аналог L-аминокислоты в различных концентрациях, и отбирая штаммы, которые формируют колонии.
Ауксотрофные мутанты можно получить, давая возможность бактериям Methylophilus формировать колонии на агаризованной среде, содержащей целевое питательное вещество (например, L-аминокислоту), получением реплик колоний на агаризованной среде, не содержащей указанного питательного вещества, и отбором штаммов, которые не могут расти на агаризованной среде, не содержащей питательного вещества.
Способы придания или усиления способности продуцировать L-аминокислоты посредством повышения активности ферментов биосинтеза L-аминокислот будут показаны на примерах ниже.
[L-лизин]
Способность продуцировать L-лизин можно придать, например, усиливая активность дигидродипиколинатсинтазы и/или активность аспартокиназы.
Активность дигидродипиколинатсинтазы и/или активность аспартокиназы бактерий Methylophilus можно повысить путем лигирования фрагмента гена, кодирующего дигидродипиколинатсинтазу, и/или фрагмента гена, кодирующего аспартокиназу, с вектором, который функционирует в бактериях Methylophilus, предпочтительно с вектором мультикопийного типа, чтобы создать рекомбинантную ДНК, и введением его в бактерию-хозяина Methylophilus, чтобы трансформировать хозяина. В результате увеличения числа копий гена, кодирующего дигидродипиколинатсинтазу, и/или гена, кодирующего аспартокиназу, в клетках трансформированного штамма активность или активности ферментов повысятся. Далее дигидродипиколинатсинтаза, аспартокиназа и аспартокиназа III также обозначаются сокращениями DDPS, AK и AKIII соответственно.
В качестве микроорганизма, предоставляющего ген, который кодирует DDPS, и ген, который кодирует АК, могут быть использованы любые микроорганизмы, при условии, что они имеют гены, обеспечивающие экспрессию активности DDPS и активности АК у микроорганизмов, относящихся к роду Methylophilus. Такими микроорганизмами могут быть штаммы дикого типа или полученные от них мутантные штаммы. В частности, примеры таких микроорганизмов включают в себя штамм К-12 E.coli (Escherichia coli), штамм AS1 Methylophilus methylotrophus (NCIMB10515) и так далее. Так как найдены нуклеотидные последовательности обоих генов - гена, кодирующего DDPS (dapA, Richaud, F. et al., J. Bacteriol., 297, (1986)), и гена, кодирующего AKIII (lysC, Cassan, M., Parsot, C., Cohen, G.N. and Patte, J.C., J. Biol. Chem., 261, 1052 (1986)), полученных от бактерии Escherichia, указанные гены могут быть получены ПЦР с использованием праймеров, синтезированных на основе нуклеотидных последовательностей этих генов, и хромосомной ДНК такого микроорганизма, как E.coli K-12 или ему подобного, в качестве матрицы. В качестве конкретных примеров ниже будут даны пояснения относительно dapA и lysC, полученных из E.coli. Однако гены, используемые в данном изобретении, не ограничены указанными выше генами.
Предпочтительно, чтобы DDPS и АК, используемые для согласно изобретению, не претерпевали ингибирования L-лизином по принципу обратной связи. Известно, что DDPS дикого типа, полученная из E.coli, претерпевает ингибирование L-лизином по принципу обратной связи и что AKIII дикого типа, полученная из E.coli, претерпевает супрессию и ингибирование по принципу обратной связи L-лизином. Поэтому dapA и lysC, вводимые в бактерии Methylophilus, предпочтительно кодируют DDPS и AKIII, имеющие мутацию, которая десенсибилизирует к ингибированию L-лизином по принципу обратной связи. В дальнейшем DDPS, имеющая мутацию, которая десенсибилизирует к ингибированию L-лизином по принципу обратной связи, также называется «мутантной DDPS», и ДНК, кодирующая мутантную DDPS, также именуется «мутантным dapA». AKIII, полученная из E.coli, имеющая мутацию, которая десенсибилизирует к ингибированию L-лизином по принципу обратной связи, также называется «мутантной AKIII», и ДНК, кодирующая мутантную AKIII, также именуется «мутантным lysC».
Согласно данному изобретению, не обязательно необходимо, чтобы DDPS и АК были мутантными. Известно, например, что DDPS, полученная из бактерий Corynebacterium, не подвергается ингибированию L-лизином по принципу обратной связи.
Нуклеотидная последовательность dapA дикого типа, полученного из E.coli, приведена в SEQ ID NO: 1. Аминокислотная последовательность DDPS дикого типа, кодируемая указанной нуклеотидной последовательностью, иллюстрируется SEQ ID NO: 2. Нуклеотидная последовательность lysC дикого типа, полученного из E.coli, иллюстрируется SEQ ID NO: 3. Аминокислотная последовательность ATIII дикого типа, кодируемая указанной нуклеотидной последовательностью, иллюстрируется SEQ ID NO: 4.
ДНК, кодирующая мутантную DDPS, которая не подвергается ингибированию по принципу обратной связи L-лизином, включает в себя ДНК, кодирующую DDPS, которая имеет аминокислотную последовательность, описанную в SEQ ID NO: 2, где остаток гистидина 118 заменен остатком тирозина. ДНК, кодирующая мутантную AKIII, которая не подвергается ингибированию по принципу обратной связи L-лизином, включает в себя ДНК, кодирующую AKIII, которая имеет аминокислотную последовательность, описанную в SEQ ID NO: 4, где остаток треонина 352 заменен остатком изолейцина.
Плазмидой, используемой для клонирования гена, может быть любая плазмида, при условии, что она может реплицироваться в микроорганизме, таком как бактерии Escherichia и им подобные, и в частности включая pBR322, pTWV228, pMW119, pUC19 и так далее.
Вектором, который функционирует в бактериях Methylophilus, является, например, плазмида, которая может автономно реплицироваться в бактериях Methylophilus. В частности, можно упомянуть RSF1010, который представляет собой вектор широкого спектра хозяев, и его производные, например, pAYC32 (Chistorerdov, A.Y., Tsygankov, Y.D. Plasmid, 16, 161-167, (1986)), pMFY42 (Gene, 44, 53, (1990)), pRP301, pTB70 (Nature, 287, 396, (1980)) и так далее.
Чтобы приготовить рекомбинантную ДНК лигированием dapA и lysC с вектором, который функционирует в бактериях Methylophilus, вектор переваривают ферментом рестрикции, который соответствует концу фрагмента ДНК, содержащего dapA и lysC. Лигирование обычно выполняют с использованием лигазы, такой как ДНК-лигаза Т4. dapA и lysC могут быть индивидуально включены в отдельные векторы или в один вектор.
В качестве плазмиды, содержащей мутантный dapA, кодирующий мутантную DDPS, и мутантный lysC, кодирующий мутантную AKIII, была известна плазмида RSFD80 с широким спектром хозяев (WO95/16042). Штамм JM109 E.coli, трансформированный указанной плазмидой, назван AJ12396 и помещен на хранение в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (почтовый индекс 305-8566, 1-3 Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) 28 октября 1993 г. и получил инвентарный номер FERM P-13936, и перенесен для международного депонирования по условиям Будапештского соглашения 1 ноября 1994 г. и получил инвентарный номер FERM BP-4859. RSFD80 можно получить из штамма AJ12396 известным способом.
Мутантный dapA, имеющийся в RSFD80, имеет нуклеотидную последовательность dapA дикого типа SEQ ID NO: 1, включая замену С в нуклеотиде номер 623 на Т. Поэтому кодируемая мутантная DDPS имеет аминокислотную последовательность SEQ ID NO: 2, включая замену остатка гистидина 118 на остаток тирозина. Мутантный lysC, имеющийся в RSFD80, имеет нуклеотидную последовательность lysC дикого типа SEQ ID NO: 3, включая замену С в нуклеотиде номер 1638 на Т. Поэтому кодируемая мутантная AKIII имеет аминокислотную последовательность SEQ ID NO: 4, включая замену остатка треонина 352 на остаток изолейцина.
Для того чтобы ввести рекомбинантную ДНК, полученную, как описано выше, в бактерии Methylophilus, можно использовать любой способ, при условии, что он обеспечивает достаточную эффективность трансформации. Например, можно использовать электропорацию (Canadian Journal of Microbiology, 43, 197 (1997)).
Активность DDPS и/или активность АК также может быть повышена при наличии множественных копий dapA и/или lysC в хромосомной ДНК бактерий Methylophilus. Чтобы ввести множественные копии dapA и/или lysC в хромосомную ДНК бактерий Methylophilus проводят гомологичную рекомбинацию с использованием в качестве мишени последовательности, которая представлена в хромосомной ДНК бактерий Methylophilus множеством копий. В качестве последовательности, присутствующей в хромосомной ДНК во множестве копий, можно использовать повторяющуюся ДНК, инвертированные повторы, имеющиеся на конце транспозируемого элемента, и им подобные. В альтернативном случае, как заявлено в опубликованной заявке на патент Японии (Kokai) No. 2-109985/1990, множественные копии dapA и/или lysC могут быть введены в хромосомную ДНК посредством их встраивания в транспозон, чтобы их перенести. В обоих способах в результате увеличенного количества копий dapA и/или lysC в трансформированных штаммах, активность DDPS и активность АК будут повышены.
Кроме указанной выше амплификации генов, активность DDPS и/или активность АК может быть повышена путем замены последовательности, контролирующей экспрессию, такой как промоторы dapA и/или lysC, более сильными промоторами (опубликованная заявка на патент Японии (Kokai) No. 1-215280/1989). В качестве сильных промоторов известны, например, промотор lac, промотор trp, промотор trc, промотор tac, промотор PR и промотор PL фага лямбда, промотор tet, промотор amyE, промотор spac и так далее. Замещение этими промоторами усиливает экспрессию dapA и/или lysC, и таким образом активность DDPS и активность АК повышаются. Усиление последовательностей, контролирующих экспрессию, можно комбинировать с увеличением числа копий dapA и/или lysC.
Чтобы приготовить рекомбинантную ДНК лигированием фрагмента гена и вектора, вектор переваривают ферментом рестрикции, соответствующим концу фрагмента гена. Лигирование обычно выполняют с помощью лигазы, такой как ДНК-лигаза Т4. В качестве способов переваривания, лигирования и других способов для ДНК, подготовки хромосомной ДНК, ПЦР, получения плазмидной ДНК, трансформации, конструирования олигонуклеотидов, используемых в качестве праймеров, и так далее, можно использовать стандартные способы, хорошо известные специалистам в данной области. Указанные способы описаны в Sambrook, J., Fritsch, E. F., and Maniatis, T., "Molecular Cloning: A Laboratory Manual, 2nd Edition", Cold Spring Harbor Laboratory Press, (1989) и так далее.
В дополнение к повышению активности DDPS и/или активности АК, также может быть повышена активность другого фермента, вовлеченного в биосинтез L-лизина. К таким ферментам относятся ферменты пути метаболизма диаминопимелата, такие как дигидродипиколинатредуктаза, диаминопимелатдекарбоксилаза, диаминопимелатдегидрогеназа (WO96/40934 для всех вышеуказанных ферментов), фосфоенолпируваткарбоксилаза (опубликованная заявка на патент Японии (Kokai) No. 60-87788/1985), аспартатаминотрансфераза (опубликованная заявка на патент Японии (Kokoku) No. 6-102028/1994), диаминопимелатэпимераза, дегидрогеназа полуальдегида аспарагиновой кислоты и так далее, или ферменты пути метаболизма аминоадипата, такие как гомоаконитатгидратаза, и так далее. Предпочтительно повышают активность по меньшей мере одного фермента из дегидрогеназы полуальдегида аспарагиновой кислоты, дегидродипиколинатредуктазы и диаминопимелатдекарбоксилазы.
Аспартокиназа, дегидрогеназа полуальдегида аспарагиновой кислоты, дигидродипиколинатсинтаза, дигидродипиколинатредуктаза и диаминопимелатдекарбоксилаза, полученные из Methylophilus methylotrophus, будут описаны далее.
Кроме того, у микроорганизмов согласно изобретению может быть снижена активность фермента, который катализирует реакцию образования другого соединения, отличного от L-лизина, посредством ответвления пути биосинтеза L-лизина, или они могут быть дефицитны по такому ферменту. Фермент, который катализирует реакцию образования не L-лизина, а другого соединения, посредством ответвления пути биосинтеза L-лизина, включает гомосериндегидрогеназу (смотри WO95/23864).
Аналогично вышеупомянутые способы повышения активности фермента, вовлеченного в биосинтез L-лизина, могут быть использованы для других аминокислот, указанных ниже.
[L-глутаминовая кислота]
Способность продуцировать L-глутаминовую кислоту можно придать бактериям Methylophilus, например, путем введения ДНК, которая кодирует любой из ферментов, включая глутаматдегидрогеназу (опубликованная заявка на патент Японии (Kokai) 61-268185/1986), глутаминсинтетазу, глутаматсинтазу, изоцитратдегидрогеназу (опубликованная заявка на патент Японии (Kokai) 62-166890/1987 и 63-214189/1988), аконитатгидратазу (опубликованная заявка на патент Японии (Kokai) 62-294086/1987), цитратсинтазу (опубликованная заявка на патент Японии (Kokai) 62-201585/1987 и 63-119688/1988), фосфоенолпируваткарбоксилазу (опубликованная заявка на патент Японии (Kokai) 60-87788/1985 и 62-55089/1987), пируватдегидрогеназу, пируваткиназу, фосфоенолпируватсинтазу, енолазу, фосфоглицеромутазу, фосфоглицераткиназу, глицеральдегид-3-фосфат-дегидрогеназу, триозофосфатизомеразу, фруктозобифосфатальдолазу, фосфофруктокиназу (опубликованная заявка на патент Японии (Kokai) 63-102692/1988), глюкозофосфатизомеразу, глутамин-оксоглутаратаминотрансферазу (WO99/07853) и так далее.
Кроме того, у микроорганизмов согласно изобретению может быть снижена активность фермента, который катализирует реакцию образования другого соединения, отличного от L-глутаминовой кислоты, посредством ответвления пути биосинтеза L-глутаминовой кислоты, или они могут быть дефицитны по такому ферменту. Фермент, который катализирует реакцию образования не L-глутаминовой кислоты, а другого соединения посредством ответвления пути биосинтеза L-глутаминовой кислоты, включает в себя α-кетоглутаратдегидрогеназу (αKGDH), изоцитратлиазу, фосфатацетилтрансферазу, ацетаткиназу, синтазу ацетооксикислоты, ацетолактатсинтазу, формиатацетилтрансферазу, лактатдегидрогеназу, глутаматдекарбоксилазу, 1-пирролин-дегидрогеназу и так далее.
[L-треонин]
Способность продуцировать L-треонин можно придать или повысить, например, усиливая активности аспартокиназы, гомосериндегидрогеназы, гомосеринкиназы и треонинсинтазы. Активности указанных ферментов можно повысить, например, путем трансформации бактерий Methylophilus с использованием рекомбинантной плазмиды, содержащей оперон треонина (опубликованная заявка на патент Японии (Kokai) No. 55-131397/1980, 59-31691/1984 и 56-15696/1981 и опубликованная заявка на патент Японии (Kohyo) No. 3-501682/1991).
Способность к продуцированию также можно придать или повысить путем амплификации или введения оперона треонина, содержащего ген, кодирующий аспартокиназу, которая десенсибилизирована к ингибированию L-треонином по принципу обратной связи (опубликованная заявка на патент Японии (Kokoku) No. 1-29559/1989), ген, кодирующий гомосериндегидрогеназу (опубликованная заявка на патент Японии (Kokai) No. 60-012995/1985), или ген, кодирующий гомосеринкиназу и гомосериндегидрогеназу (опубликованная заявка на патент Японии (Kokai) No. 61-195695/1986).
Кроме того, способность продуцировать L-треонин можно повысить путем введения ДНК, кодирующей мутантную фосфоенолпируваткарбоксилазу, имеющую мутацию десенсибилизации к ингибированию аспарагиновой кислотой по принципу обратной связи.
[L-валин]
Способность продуцировать L-валин можно придать, например, путем введения в бактерии Methylophilus гена биосинтеза L-валина, механизм регуляции которого в значительной степени десенсибилизирован. Также можно ввести мутацию, которая в значительной степени десенсибилизирует механизм регуляции гена биосинтеза L-валина, который несет микроорганизм, относящийся к роду Methylophilus.
Примеры гена биосинтеза L-валина включают в себя, например, оперон ilvGMEDA E.coli. Треониндезаминаза, кодируемая геном ilvA, катализирует реакцию дезаминирования, превращающую L-треонин в 2-кетомасляную кислоту, которая является определяющим скорость этапом биосинтеза L-изолейцина. Поэтому для того чтобы добиться эффективного усиления реакций синтеза L-валина, предпочтительно использовать оперон, который не экспрессирует активность треониндезаминазы. Примеры оперона ilvGMEDA, который не экспрессирует такой треониндезаминазной активности, включают в себя оперон ilvGMEDA, в котором в ilvA введена мутация элиминирования активности треониндезаминазы, или ilvA нарушен, и оперон, в котором ilvA делетирован.
Так как оперон ilvGMEDA претерпевает контроль экспрессии оперона (ослабление) L-валином и/или L-изолейцином и/или L-лейцином, район, необходимый для ослабления, предпочтительно удаляют или подвергают мутации, чтобы ликвидировать чувствительность к супрессии экспрессии L-валином.
Оперон ilvGMEDA, который не экспрессирует треониндезаминазную активность и который лишен чувствительности к ослаблению, как описано выше, можно получить, подвергая оперон ilvGMEDA дикого типа мутагенной обработке или модифицируя его с помощью технологии рекомбинации генов (смотри WO96/06926).
[L-лейцин]
Способность продуцировать L-лейцин можно придать или повысить, например, введением в микроорганизм, относящийся к роду Methylophilus, гена биосинтеза L-лейцина, механизм регуляции которого в значительной степени десенсибилизирован, в дополнение к указанным выше характеристикам, необходимым для продукции L-валина. Также можно ввести такую мутацию, при которой механизм регуляции гена биосинтеза L-лейцина в микроорганизме, относящемся к роду Methylophilus, будет в значительной степени элиминирован. Примеры указанного гена включают в себя, например, ген leuA, который дает фермент, ингибирование которого L-лейцином в значительной степени устранено.
[L-изолейцин]
Способность продуцировать L-изолейцин можно придать, например, введением оперона thrABC, содержащего ген thrA, кодирующий аспартокиназу I/гомосериндегидрогеназу I, полученную из E.coli, который в значительной степени десенсибилизирован к ингибированию L-треонином, и оперона ilvGMEDA, который содержит ген ilvA, кодирующий треониндезаминазу, который в значительной степени десенсибилизирован к ингибированию L-изолейцином и район которого, необходимый для ослабления, удален (опубликованная заявка на патент Японии (Kokai) No. 8-47397/1996).
[Другие аминокислоты]
Биосинтез L-триптофана, L-фенилаланина, L-тирозина, L-треонина и L-изолейцина можно усилить, повышая способность бактерий Methylophilus продуцировать фосфоенолпируват (WO97/08333).
Способность продуцировать L-фенилаланин и L-тирозин повышали путем амплификации или введения десенсибилизированного гена хоризматмутазы-префенатдегидратазы (CM-PDT) (опубликованная заявка на патент Японии (Kokai) No. 5-236947/1993 и 62-130693/1987) и десенсибилизированного гена 3-дезокси-D-арабиногептулонат-7-фосфатсинтазы (DS) (опубликованная заявка на патент Японии (Kokai) No. 5-236947/1993 и 61-124375/1986).
Способность продуцировать L-триптофан повышают путем амплификации или введения триптофанового оперона, содержащего ген, кодирующий десенсибилизированную антранилатсинтазу (опубликованная заявка на патент Японии (Kokai) No. 57-71397/1982, 62-244382/1987 и патент США No. 4371614).
В данном описании выражение «активность фермента повышена» обычно относится к тому, что внутриклеточная активность фермента выше, чем активность у штамма дикого типа, и к тому случаю, когда штамм, в котором активность фермента повышена, получают путем модификации с использованием технологии рекомбинации генов или подобным способом, при этом внутриклеточная активность фермента выше, чем активность у штамма до модификации. Выражение «активность фермента снижена» обычно относится к тому, что внутриклеточная активность фермента ниже, чем активность у штамма дикого типа, и к тому случаю, когда штамм, в котором активность фермента снижена, получают путем модификации с использованием технологии рекомбинации генов или подобным способом, при этом внутриклеточная активность фермента ниже, чем активность у штамма до модификации.
L-аминокислоты можно получить, культивируя бактерии Methylophilus, обладающие способностью продуцировать L-аминокислоты, полученные, как описано выше, в среде, для того чтобы продуцировать и накопить L-аминокислоты в культуре, и собирая L-аминокислоты из культуры.
Бактериальные клетки бактерий Methylophilus с повышенным содержанием L-аминокислоты по сравнению со штаммами бактерий Methylophilus дикого типа можно получить путем культивирования бактерий Methylophilus, обладающих способностью продуцировать L-аминокислоты, в среде, чтобы продуцировать и накопить L-аминокислоты в бактериальных клетках бактерий.
Микроорганизмы, используемые в данном изобретении, можно культивировать способами, обычно применяемыми для культивирования микроорганизмов, обладающих свойством усваивать метанол. Средой, применяемой согласно изобретению, может быть природная или синтетическая среда, при условии, что она содержит источник углерода, источник азота, неорганические ионы и другие органические микрокомпоненты при необходимости.
Используя метанол в качестве основного источника углерода, L-аминокислоты можно получать при низких затратах. В том случае, когда в качестве основного источника углерода используют метанол, его обычно добавляют в среду в количестве от 0,001 до 30%. В качестве источника азота используют сульфат аммония или подобное соединение, добавляя его в среду. Помимо указанных соединений обычно добавляют небольшие количества микрокомпонентов, таких как фосфат калия, фосфат натрия, сульфат магния, сульфат железа и сульфат марганца.
Культуру обычно содержат в аэробных условиях, получаемых, например, путем встряхивания или перемешивания для аэрации, при рН от 5 до 9 и температуре от 20 до 45°С, и обычно ее завершают в пределах от 24 до 120 часов.
Сбор L-аминокислот из культуры обычно можно осуществить путем комбинирования известных способов, таких как способы с использованием ионообменной смолы, осаждения и другие.
Кроме того, клетки бактерий Methylophilus можно отделить от среды стандартными способами отделения клеток микроорганизмов.
<2> Ген согласно данному изобретению
ДНК согласно изобретению представляет собой ген, который кодирует один из ферментов, аспартокиназу (впредь также сокращенно называемую «АК»), дегидрогеназу полуальдегида аспарагиновой кислоты (впредь также сокращенно называемую «ASD»), дигидродипиколинатсинтазу (впредь также сокращенно называемую «DDPS»), дигидродипиколинатредуктазу (впредь также сокращенно называемую «DDPR») и диаминопимелатдекарбоксилазу (впредь также сокращенно называемую «DPDC»), полученный из Methylophilus methylotrophus.
ДНК согласно изобретению может быть получена, например, путем трансформации мутантного штамма микроорганизма, дефицитного по АК, ASD, DDPS, DDPR или DPDC, с использованием библиотеки генов Methylophilus methylotrophus и отбором клона, у которого ауксотрофия компенсирована.
Библиотеку генов Methylophilus mehylotrophus можно, например, получить следующим образом. Сначала получают суммарную хромосомную ДНК из штамма Methylophilus methylotrophus дикого типа, например штамма AS1 Methylophilus mehylotrophus (NCIMB10515), методом Saito et al. (Saito, H. and Miura, K., Biochem. Biophys. Acta 72, 619-629, (1963)) или подобным способом, и частично переваривают подходящим ферментом рестрикции, например Sau3AI или AluI, чтобы получить смесь различных фрагментов. Контролируя степень переваривания посредством регулирования продолжительности реакции переваривания и так далее, можно использовать широкий круг ферментов рестрикции.
Затем переваренные фрагменты хромосомной ДНК лигируют с векторной ДНК, автономно реплицирующейся в клетках Escherichia coli, чтобы получить рекомбинантную ДНК. В частности, ферменту рестрикции, дающему такие же концевые нуклеотидные последовательности как последовательности, полученные с помощью фермента рестрикции, используемого при переваривании хромосомной ДНК, дают возможность действовать на векторную ДНК, чтобы полностью переварить и расщепить вектор. Затем смесь фрагментов хромосомной ДНК и переваренную и расщепленную векторную ДНК смешивают и дают возможность лигазе, предпочтительно ДНК-лигазе Т4, действовать на смесь, чтобы получить рекомбинантную ДНК.
Раствор библиотеки генов можно получить, трансформируя Escherichia coli, например Escherichia coli штамма JM109 или ему подобного, с использованием полученной рекомбинантной ДНК, и получением рекомбинантной ДНК из культуральной жидкости трансформанта. Указанную трансформацию можно осуществить способом D.M. Morrison (Methods in Enzymology 68, 326 (1979)), методом обработки реципиентных клеток хлоридом кальция, для того чтобы увеличить проницаемость ДНК (Mandel, M. and Higa, A., J. Mol. Biol., 53, 159 (1970)) и так далее. В упоминаемых ниже примерах использовали электропорацию.
В качестве примеров вышеупомянутого вектора могут быть названы pUC19, pUC18, pUC118, pUC119, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219, pMW218, pSTV28, pSTV29 и так далее. Также можно использовать фаговые векторы. Поскольку, например, pUC118 и pUC119 содержат ген устойчивости к ампициллину и pSTV28 и pSTV29 содержат ген устойчивости к хлорамфениколу, только трансформанты, которые несут вектор или рекомбинантную ДНК, можно вырастить с использованием среды, содержащей ампициллин или хлорамфеникол.
В качестве способа культивирования трансформантов и сбора рекомбинантной ДНК из бактериальных клеток можно назвать щелочной способ с SDS и ему подобные.
Мутантный бактериальный штамм, дефицитный по АК, ASD, DDPS, DDPR или DPDC, трансформируют, используя раствор библиотеки генов Methylophilus methylotrophus, полученный, как описано выше, и отбирают клоны, у которых ауксотрофия компенсирована.
Примеры мутантного бактериального штамма, дефицитного по АК, включают в себя GT3 E.coli, дефицитный по трем видам генов, кодирующих АК (thrA, metLM, lysC). Примеры мутантного бактериального штамма, дефицитного по ASD, включают в себя Hfr3000 U482 E.coli (штамм CGSC 5081). Примеры мутантного бактериального штамма, дефицитного по DDPS, включают в себя АТ997 E.coli (штамм CGSC 4547). Примеры мутантного бактериального штамма, дефицитного по DDPR, включают в себя АТ999 E.coli (штамм CGSC 4549). Примеры мутантного бактериального штамма, дефицитного по DPDC, включают в себя АТ2453 E.coli (штамм CGSC 4505). Указанные мутантные штаммы можно получить из Центра культур E.coli Genetic Stock Center (the Yale University, Department of Biology, Osborn Memorial Labs., P.O. Box 6666, New Haven 06511-7444, Connecticut, U.S.).
Хотя все вышеупомянутые мутантные штаммы не могут расти на минимальной среде М9, трансформированные штаммы, которые содержат ген, кодирующий АК, ASD, DDPS, DDPR или DPDC, могут расти на минимальной среде М9, так как в трансформантах функционируют указанные гены. Поэтому отбирая трансформированные штаммы, которые могут расти на минимальной среде, и получая рекомбинантную ДНК из этих штаммов, можно получить фрагменты ДНК, содержащие ген, который кодирует каждый фермент. АТ999 E.coli (штамм CGSC 4549) проявляет чрезвычайно низкую скорость роста даже в полной среде, такой как среда L, в том случае, когда диаминопимелиновая кислота не добавлена в среду. Однако нормальный рост можно наблюдать для трансформированных штаммов этих бактерий, которые содержат ген, кодирующий DDPR Methylophilus methylotrophus, благодаря функционированию гена. Поэтому трансформированный штамм, который содержит ген, кодирующий DDPR, также можно получить путем отбора трансформированного штамма, нормально растущего на среде L.
Выделив встроенный фрагмент ДНК из полученной рекомбинантной ДНК и определив его нуклеотидную последовательность, можно определить аминокислотную последовательность каждого фермента и нуклеотидную последовательность кодирующего его гена.
Ген, кодирующий АК согласно изобретению (впредь также сокращенно называемый «ask»), кодирует АК, которая имеет аминокислотную последовательность SEQ ID NO: 6, указанную в списке последовательностей. В качестве конкретного примера гена ask можно указать ДНК, имеющую нуклеотидную последовательность, которая состоит из нуклеотидов SEQ ID NO: 5. Ген ask согласно данному изобретению может иметь последовательность, в которой кодон, соответствующий каждой из аминокислот, заменен эквивалентным кодоном, при условии, что он кодирует такую же аминокислотную последовательность, как аминокислотная последовательность SEQ ID NO: 6.
Ген, который кодирует ASD согласно изобретению (впредь также сокращенно называемый "asd"), кодирует ASD, которая имеет аминокислотную последовательность SEQ ID NO: 8, указанную в списке последовательностей. В качестве конкретного примера гена asd можно указать ДНК, которая содержит нуклеотидную последовательность, состоящую из нуклеотидов с номерами нуклеотидов 98-1207 в SEQ ID NO: 7. Ген asd согласно изобретению может иметь последовательность, в которой кодон, соответствующий каждой из аминокислот, заменен эквивалентным кодоном, при условии, что он кодирует такую же аминокислотную последовательность, как аминокислотная последовательность SEQ ID NO: 8.
Ген, который кодирует DDPS согласно изобретению (впредь также сокращенно называемый "dapA"), кодирует DDPS, которая имеет аминокислотную последовательность SEQ ID NO: 10, указанную в списке последовательностей. В качестве конкретного примера гена dapA можно указать ДНК, которая имеет нуклеотидную последовательность, состоящую из нуклеотидов с номерами нуклеотидов 1268-2155 в SEQ ID NO: 9. Ген dapA согласно изобретению может иметь последовательность, в которой кодон, соответствующий каждой из аминокислот, заменен эквивалентным кодоном, при условии, что он кодирует такую же аминокислотную последовательность, как аминокислотная последовательность SEQ ID NO: 10.
Ген, который кодирует DDPR согласно изобретению (впредь также сокращенно называемый "dapB"), кодирует DDPR, которая имеет аминокислотную последовательность SEQ ID NO: 12, указанную в списке последовательностей. В качестве конкретного примера гена dapB можно указать ДНК, которая имеет нуклеотидную последовательность, состоящую из нуклеотидов с номерами нуклеотидов 2080-2883 в SEQ ID NO: 11. Ген dapB согласно изобретению может иметь последовательность, в которой кодон, соответствующий каждой из аминокислот, заменен эквивалентным кодоном, при условии, что он кодирует такую же аминокислотную последовательность, как аминокислотная последовательность SEQ ID NO: 12.
Ген, который кодирует DPDC согласно изобретению (впредь также сокращенно называемый "lysA"), кодирует DPDC, которая имеет аминокислотную последовательность SEQ ID NO: 14, указанную в списке последовательностей. В качестве конкретного примера гена lysA можно указать ДНК, которая содержит нуклеотидную последовательность, состоящую из нуклеотидов с номерами нуклеотидов 751-1995 в SEQ ID NO: 13. Ген lysA согласно изобретению может иметь последовательность, в которой кодон, соответствующий каждой из аминокислот, заменен эквивалентным кодоном, при условии, что он кодирует такую же аминокислотную последовательность, как аминокислотная последовательность SEQ ID NO: 14.
Ген любого фермента согласно изобретению может иметь аминокислотную последовательность, соответствующую любой из аминокислотных последовательностей SEQ ID NO: 6, 8, 10, 12 или 14, включая замену, делецию, инсерцию, присоединение или инверсию одной или нескольких аминокислот, и может кодировать белок, обладающий активностью AK, ASD, DDPS, DDPR или DPDC. Используемое здесь выражение «одна или несколько» предпочтительно означает количество от 1 до 10, более предпочтительно - количество от 1 до 5, более предпочтительно - количество от 1 до 2.
ДНК, которая кодирует по существу такой же белок, как AK, ASD, DDPS, DDPR или DPDC, как те, которые указаны выше, можно получить, модифицируя каждую нуклеотидную последовательность так, чтобы аминокислотная последовательность содержала замену, делецию, инсерцию, присоединение или инверсию аминокислотного остатка или остатков в конкретном сайте, например, посредством сайт-специфичного мутагенеза. Такую модифицированную ДНК, которая указана выше, также можно получить путем традиционных мутагенных воздействий. Примеры мутагенных воздействий включают в себя обработку in vitro ДНК, кодирующей AK, ASD, DDPS, DDPR или DPDC, гидроксиламином или подобными средствами, обработку микроорганизма, такого как бактерии Escherichia, содержащего ген, кодирующий AK, ASD, DDPS, DDPR или DPDC, УФ-излучением или мутагенными агентами, используемыми при обычных мутагенных обработках, такими как N-метил-N'-нитро-N-нитрозогуанидин (NTG) и азотистая кислота.
Вышеуказанная замена, делеция, инсерция, присоединение или инверсия нуклеотидов включает в себя мутации природного происхождения (мутант или вариант), такие как мутации, наблюдаемые в зависимости от различий между видами или штаммами микроорганизмов, содержащих AK, ASD, DDPS, DDPR или DPDC и так далее.
ДНК, которая кодирует по существу тот же самый белок, что и AK, ASD, DDPS, DDPR или DPDC, можно получить, создав возможность для экспрессии ДНК, имеющей такую мутацию, как указано выше, в подходящей клетке, и оценивая активность AK, ASD, DDPS, DDPR или DPDC продукта экспрессии. ДНК, которая кодирует по существу тот же самый белок, что и AK, ASD, DDPS, DDPR или DPDC, также можно получить посредством выделения из ДНК, кодирующих AK, ASD, DDPS, DDPR или DPDC, которые имеют мутации, или из клеток, содержащих каждую из ДНК, ДНК, способной гибридизоваться с пробой, содержащей нуклеотидную последовательность, которая включает в себя нуклеотидную последовательность из нуклеотидов с номерами 510-1736 SEQ ID NO: 5, нуклеотидную последовательность, включающую в себя нуклеотидную последовательность из нуклеотидов с номерами 98-1207 SEQ ID NO: 7, нуклеотидную последовательность, включающую в себя нуклеотидную последовательность из нуклеотидов с номерами 1268-2155 SEQ ID NO: 9, нуклеотидную последовательность, включающую в себя нуклеотидную последовательность из нуклеотидов с номерами 2080-2883 SEQ ID NO: 11, или нуклеотидную последовательность, включающую в себя нуклеотидную последовательность из нуклеотидов с номерами 751-1995 SEQ ID NO: 13, или часть этих нуклеотидных последовательностей, в жестких условиях, и кодирующей белок, обладающий активностью AK, ASD, DDPS, DDPR или DPDC. В данном описании иметь нуклеотидную последовательность или ее часть означает иметь нуклеотидную последовательность или ее часть, или комплементарную ей нуклеотидную последовательность.
Используемый здесь термин «жесткие условия» означает условия, которые дают возможность для образования так называемого специфичного гибрида и не позволяют образоваться неспецифичному гибриду. Эти условия могут варьировать в зависимости от нуклеотидной последовательности и длины пробы. Однако это может быть, например, условие, которое обеспечивает гибридизацию высоко гомологичной ДНК, такой как ДНК, имеющей гомологию 40% или выше, но не допускает гибридизацию ДНК с более низкой гомологией, чем указанно выше, или условие, которое обеспечивает гибридизацию в условиях промывки для традиционной Саузерн-гибридизации, при температуре 60°С и концентрациях соли, соответствующих 1 х SSC и 0,1% SDS, предпочтительно 0,1 х SSC и 0,1% SDS.
В качестве пробы также можно использовать неполную последовательность каждого гена. Такая проба может быть получена посредством ПЦР (полимеразная цепная реакция) с использованием олигонуклеотидов, полученных на основе нуклеотидной последовательности каждого гена, в качестве праймеров, и фрагмента ДНК, содержащего каждый ген, в качестве матрицы. В том случае, когда в качестве пробы используют фрагмент ДНК, имеющий длину примерно 300 п.н., условия промывки при гибридизации могут быть, например, 50°С, 2 х SSC и 0,1% SDS.
Гены, которые гибридизуются при таких условиях, которые указаны выше, также включают в себя гены, имеющие стоп-кодон, находящийся в их последовательности, и гены, кодирующие фермент, не обладающий больше своей активностью вследствие мутации активного центра. Однако такие гены можно легко исключить посредством лигирования генов в доступный для приобретения вектор, экспрессирующий активность, и измерения активности AK, ASD, DDPS, DDPR или DPDC.
Поскольку нуклеотидные последовательности генов, которые кодируют AK, ASD, DDPS, DDPR или DPDC, полученных из Methylophilus methylotrophus, выявлены данным изобретением, то последовательности ДНК, которые кодируют AK, ASD, DDPS, DDPR или DPDC, можно получить из библиотеки генов Methylophilus methylotrophus посредством гибридизации с использованием олигонуклеотидных проб, полученных на основе данных последовательностей. Кроме того, последовательности ДНК, которые кодируют указанные ферменты, также можно получить путем их амплификации из хромосомной ДНК Methylophilus methylotrophus в ходе ПЦР с использованием олигонуклеотидных праймеров, полученных на основе вышеуказанных нуклеотидных последовательностей.
Вышеупомянутые гены могут быть соответствующим образом использованы для повышения способности бактерий Methylophilus продуцировать L-лизин.
ПРИМЕРЫ
Данное изобретение далее будет описано конкретно со ссылками на следующие далее примеры.
Используемые реактивы получали из Wako Pure Chemicals или Nakarai Tesque, если не оговорено особо. Составы используемых в каждом примере сред, показаны ниже. рН доводили с помощью NaOH или HCl для всех сред.
(Среда L)
Бактотриптон (DIFCO) 10 г/л
Дрожжевой экстракт (DIFCO) 5 г/л
NaCl 5 г/л
[Стерилизация паром при 120°С в течение 20 минут]
(Агаризованная среда L)
Среда L
Бактоагар (DIFCO) 15 г/л
[стерилизация паром при 120°С в течение 20 минут]
(Среда SOC)
Бактотриптон (DIFCO) 20 г/л
Дрожжевой экстракт (DIFCO) 5 г/л
10 мМ NaCl
2,5 мМ KCl
10 мМ MgSO4
10 мМ MgCl2
20 мМ глюкоза
[Компоненты, за исключением раствора магния и глюкозы, стерилизовали паром (120°С, 20 минут), затем к ним добавляли 2 М маточный раствор магния (1 М MgSO4, 1 М MgCl2) и 2 М раствор глюкозы, растворы которых были предварительно пропущены через фильтр с диаметром пор 0,22 мкм, и смесь пропускали снова через 0,22 мкм-фильтр.]
(Среда 121М1)
K2HPO4 1,2 г/л
KH2PO4 0,62 г/л
NaCl 0,1 г/л
(NH4)2SO4 0,5 г/л
MgSO4•7H2O 0,2 г/л
CaCl2•6H2O 0,05 г/л
FeCl3•6H2O 1,0 мг/л
H3BO3 10 мкг/л
CuSO4•5H2O 5 мкг/л
MnSO4•5H2O 10 мкг/л
ZnSO4•7H2O 70 мкг/л
NaMoO4•2H2O 10 мкг/л
CoCl2•6H2O 5 мкг/л
Метанол 1% (об./об.), рН 7,0
[Компоненты, за исключением метанола стерилизовали паром при 121°С в течение 15 минут. После достаточного охлаждения компонентов добавляли метанол.]
(Состав среды 121 для продуцирования)
Метанол 2%
Гидрофосфат калия 0,12%
Фосфат калия 0,062%
Гексагидрат хлорида кальция 0,005%
Гептагидрат сульфата магния 0,02%
Хлорид натрия 0,01%
Гексагидрат хлорида железа 1,0 мг/л
Сульфат аммония 0,3%
Пентагидрат сульфата меди 5 мкг/л
Пентагидрат сульфата марганца 10 мкг/л
Дигидрат молибдата натрия 10 мкг/л
Борная кислота 10 мкг/л
Гептагидрат сульфата цинка 70 мкг/л
Гексагидрат хлорида кобальта 5 мкг/л
Карбонат кальция (Kanto Kagaku) 3%
(рН 7,0)
(Агаризованная среда 121М1)
Среда 121М1
Бактоагар (DIFCO) 15 г/л
[Компоненты, за исключением метанола стерилизовали паром при 121°С в течение 15 минут. После достаточного охлаждения компонентов добавляли метанол.]
(Минимальная среда М9)
Na2HPO4•12H2O 16 г/л
KH2PO4 3 г/л
NaCl 0,5 г/л
NH4Cl 1 г/л
MgSO4•7H2O 246,48 мг/л
глюкоза 2 г/л
рН 7,0
[MgSO4 и глюкозу стерилизовали отдельно (120°С, 20 минут). При необходимости добавляли соответствующее количество аминокислот и витаминов.]
(Минимальная агаризованная среда М9)
Минимальная среда М9
Бактоагар (DIFCO) 15 г/л
Пример 1
Создание бактерии, продуцирующей L-лизин (1)
(1) Введение мутантного lysC и мутантного dapA в бактерию Methylophilus.
Мутантный lysC и мутантный dapA вводили в бактерию Methylophilus, используя содержащую их известную плазмиду RSFD80 (смотри WO95/16042). RSFD80 представляет собой плазмиду pVIC40 (международная заявка на патент WO90/04636, опубликованная заявка на патент Японии (Kohyo) No. 3-501682/1991), полученную из векторной плазмиды pAYC32 с широким спектром хозяев (Chistorerdov, A.Y., Tsygankov, Y.D., Plasmid, 16, 161-167, (1986)), которая является производной RSF1010, в которой мутантный dapA и мутантный lysC, полученные из E.coli, локализованы в таком порядке ниже по течению промотора (tetP) гена устойчивости к тетрациклину плазмиды pVIC40, так что направления транскрипции генов по отношению к tetP обычны. Мутантный dapA кодировал мутантную DDPS, в которой остаток гистидина 118 заменили остатком тирозина. Мутантный lysC кодировал мутантную AKIII, в которой остаток треонина 352 заменили остатком изолейцина.
RSFD80 конструировали следующим образом. Мутантный dapA в плазмиде pdapAS24 лигировали с pVIC40 в положении ниже по течению промотора гена устойчивости к тетрациклину, чтобы получить RSF24P, как показано на фиг.1. Затем получали плазмиду RSFD80, которая имела мутантный dapA и мутантный lysC, из RSF24P и pLLC*80, содержащей мутантный lysC, как показано на фиг.2. То есть, в то время как pVIC40 содержит треониновый оперон, указанный треониновый оперон в RSFD80 замещен фрагментом ДНК, содержащим мутантный dapA, и фрагментом ДНК, содержащим мутантный lysC.
Штамм JM109 E.coli, трансформированный плазмидой RSFD80, назван AJ12396 и помещен на хранение в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (почтовый индекс 305-8566, 1-3 Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) 28 октября 1993 г. и получил инвентарный номер FERM P-13936, и перенесен для международного депонирования по условиям Будапештского соглашения 1 ноября 1994 г. и получил инвентарный номер FERM BP-4859.
Штамм AJ1239 E.coli культивировали в 30 мл среды LB, содержащей 20 мг/л стрептомицина, при 30°С в течение 12 часов, и плазмиду RSFD80 очищали из полученных клеток, используя систему для очистки ДНК Wizard® Plus Midipreps (продаваемой Promega).
Плазмиду RSFD80, полученную, как описано выше, вводили в штамм AS1 Methylophilus methylotrophus (NCIMB10515) посредством электропорации (Canadian Journal of Microbiology, 43, 197 (1997)). В качестве контроля район ДНК, кодирующий треониновый оперон, делетировали из плазмиды pVIC40, используемой для получения плазмиды RSFD80, чтобы получить плазмиду pRS, содержащую только векторный район (смотри опубликованную заявку на патент Японии (Kohyo) No. 3-501682/1991), и плазмиду pRS вводили в штамм AS1 таким же способом, который использовали для RSFD80.
(2) Активность AKIII бактерий Methylophilus, содержащих мутантный lysC и мутантный dapA, полученные из E.coli.
Получали бесклеточные экстракты из штамма AS1 Methylophilus methylotrophus, содержащего плазмиду RSFD80 (также именуемого далее «AS1/RSFD80»), и штамма AS1 Methylophilus methylotrophus, содержащего плазмиду pRS (также именуемого далее «AS1/pRS»), и измеряли активность АК. Бесклеточные экстракты (неочищенные растворы ферментов) получали следующим образом. Каждый из штаммов AS1/RSFD80 и AS1/pRS инокулировали в среду для продуцирования 121 указанного выше состава, содержащую 20 мг/мл стрептомицина, культивировали при 37°С в течение 34 часов при встряхивании и затем удаляли карбонат кальция и собирали клетки.
Бактериальные клетки, полученные как описано выше, промывали 0,2% KCl в условиях 0°С, суспендировали в 20 мМ калий-фосфатном буфере (рН 7), содержащем 10 мМ MgSO4, 0,8 М (NH4)2SO4 и 0,03 М β-меркаптоэтанола, и разрушали ультразвуком (0°С, 200 Вт, 10 минут). Суспензию обработанных ультразвуком клеток центрифугировали при 33000 об/мин в течение 30 минут в условиях 0°С и отделяли надосадок. К надосадку добавляли сульфат аммония до 80% насыщения, смесь оставляли при 0°С на 1 час и центрифугировали. Осадок растворяли в 20 мМ калий-фосфатном буфере (рН 7), содержащем 10 мМ MgSO4, 0,8 М (NH4)2SO4 и 0,03 М β-меркаптоэтанола.
Измерение активности АК выполняли методом Стадтмана (Stadtman, E.R., Cohen, G.N., LeBras, G., and Robichon-Szulmajster, H., J. Biol. Chem., 236, 2033 (1961)). А именно, реакционный раствор приведенного ниже состава инкубировали при 30°С в течение 45 минут и проявление окраски вызывали добавлением раствора FeCl3 (2,8 н HCl: 0,4 мл, 12% ТХУ: 0,4 мл, 5% FeCl3•6H2O/0,1 н HCl: 0,7 мл). Реакционный раствор центрифугировали и измеряли оптическую плотность надосадка при 540 нм. Активность представляли в единицах количества гидроксамовой кислоты, полученного в 1 минуту (1 ед. = 1 мкмоль/минуту). Коэффициент молярной экстинкции принимали равным 600. В качестве пустого контроля использовали реакционный раствор, не содержащий аспартата калия. После того как измеряли ферментативную активность, к реакционному раствору ферментов добавляли L-лизин в различных концентрациях, чтобы оценить степень ингибирования L-лизином. Результаты показаны в таблице 1.
(Состав реакционного раствора)
Реакционная смесь*1 0,3 мл
Раствор гидроксиламина*2 0,2 мл
0,1 М аспартат калия (рН 7,0) 0,2 мл
Раствор фермента 0,1 мл
Вода (остальное) общий объем 1 мл
*1: 1 М трис-HCl (рН 8,1): 9 мл, 0,3 М MgSO4: 0,5 мл и 0,2 М АТФ (рН 7,0): 5 мл
*2: 8 М раствор гидроксиламина, нейтрализованный KOH непосредственно перед использованием
активность*1)
в присутствии 5 мМ L-лизина
*1: нмоль/минуту/мг белка
*2: Степень сохранения активности в присутствии 5 мМ L-лизина.
Как показано в таблице 1 активность АК повышалась примерно в 1,7 раза при введении плазмиды RSFD80. Кроме того, подтвердилось, что АК, полученная из E.coli, которую кодировала плазмида RSFD80, была полностью десенсибилизирована к ингибированию L-лизином. Кроме того, было обнаружено, что АК, которая в исходном состоянии сохранялась у штамма AS1, не ингибировалась одним L-лизином. Авторы согласно изобретению обнаружили, что АК, полученная из штамма AS1, ингибировалась на 100% в том случае, когда в реакционном растворе присутствовали L-лизин и L-треонин по 2 мМ каждого (совместное ингибирование).
(3) Продуцирование L-лизина бактерией Methylophilus, содержащей мутантный lysC и мутантный dapA, полученные из E.coli.
Затем штамм AS1/RSFD80 и штамм AS1/pRS инокулировали в среду для продуцирования 121, содержащую 20 мг/л стрептомицина, и культивировали при 37°С в течение 34 часов при встряхивании. После окончания культивирования бактериальные клетки и карбонат кальция удаляли центрифугированием и в надосадке культуры измеряли концентрацию L-лизина в аминокислотном анализаторе (JASCO Corporation [Nihon Bunko], высоко эффективная жидкостная хроматография). Результаты показаны в таблице 2.
гидрохлорида L-лизина (г/л)
Пример 2
Создание бактерии, продуцирующей L-лизин (2)
(1) Введение промоторного района tac в вектор с широким спектром хозяев.
Чтобы получить большое количество фермента, вовлеченного в биосинтез L-лизина (Lys) у Methylophilus methylotrophus для экспрессии гена целевого фермента использовали промотор tac. Данный промотор часто используется в E.coli.
Промоторный район tac получали амплификацией посредством ПЦР с использованием в качестве матрицы ДНК pKK233-3 (Pharmacia), и в качестве праймеров фрагментов ДНК, имеющих нуклеотидные последовательности SEQ ID NO: 15 и 16, и терморезистентную ДНК-полимеразу. ПЦР выполняли с использованием цикла при 94°С в течение 20 секунд, при 60°С в течение 30 секунд и при 72°С в течение 60 секунд, который повторяли 30 раз. Затем амплифицированный фрагмент ДНК собирали и обрабатывали ферментами рестрикции EcoRI и PstI. С другой стороны, вектор pRS с широким спектром хозяев (смотри опубликованную заявку на патент Японии (Kohyo) No. 3-501682/1991) также переваривали такими же ферментами рестрикции, и вышеупомянутый фрагмент ДНК, который содержал промоторный район tac, вводили между концами, полученными при переваривании ферментами рестрикции, чтобы сконструировать pRS-tac.
(2) Получение плазмиды pRS-dapA24, экспрессирующей ген dapA (ген дигидродипиколинатсинтазы), и плазмиды pRS-lysC80, экспрессирующей ген lysC (ген аспартокиназы).
Мутантный ген (dapA*24), кодирующий дигидродипиколинатсинтазу, ферментативная активность которой была частично десенсибилизирована по отношению к ингибированию аминокислотой Lys по принципу обратной связи, вводили в плазмиду pRS-tac, которую получали способом, описанным выше в (1).
Сначала получали район гена dapA*24 амплификацией посредством ПЦР, используя в качестве матрицы ДНК RSFD80 (смотри пример 1), и фрагменты ДНК, имеющие нуклеотидные последовательности SEQ ID NO: 17 и 18, в качестве праймеров. ПЦР выполняли с использованием цикла при 94°С в течение 20 секунд, при 60°С в течение 30 секунд и при 72°С в течение 90 секунд, который повторяли 30 раз. Затем фрагмент обрабатывали ферментами рестрикции Sse8387I и XbaI, чтобы получить фрагмент гена dapA*24, имеющий соответствующие расщепленные концы. С другой стороны, вектор pRS-tac так же обрабатывали Sse8387I и частично переваривали XbaI таким же образом, как описано выше. С этой переваренной плазмидой лигировали вышеуказанный фрагмент гена dapA*24, используя лигазу Т4, чтобы получить pRS-dapA24.
Подобным образом ген (lysC*80), кодирующий аспартокиназу, ферментативная активность которой была частично десенсибилизирована по отношению к ингибированию аминокислотой Lys по принципу обратной связи, получали посредством ПЦР, используя в качестве матрицы ДНК RSFD80, и фрагменты ДНК, имеющие нуклеотидные последовательности SEQ ID NO: 19 и 20, в качестве праймеров. ПЦР выполняли с использованием цикла при 94°С в течение 20 секунд, при 60°С в течение 30 секунд и при 72°С в течение 90 секунд, который повторяли 30 раз. Затем полученный фрагмент ДНК обрабатывали ферментами рестрикции Sse8387I и SapI. С другой стороны, вектор pRS-tac так же обрабатывали Sse8387I и SapI. С этой переваренной плазмидой лигировали вышеуказанный фрагмент гена lysC*80, используя лигазу Т4, чтобы получить pRS-lysC80.
(3) Введение pRS-dapA24 или pRS-lysC80 в Methylophilus methylotrophus и оценка культуры.
Каждую из плазмид pRS-dapA24 и pRS-lysC80, полученных как описано выше, вводили в штамм AS1 Methylophilus methylotrophus (NCIMB10515) посредством электропорации, чтобы получить соответственно AS1/pRS-dapA24 и AS1/pRS-lys80. Каждый штамм инокулировали в среду для продуцирования 121, содержащую 20 мг/л стрептомицина, и культивировали при 37°С в течение 48 часов при встряхивании. В качестве контрольного штамма сходным образом также культивировали штамм AS1, несущий pRS. После завершения культивирования клетки и карбонат кальция удаляли центрифугированием и в надосадке культуры измеряли концентрацию L-лизина в аминокислотном анализаторе (JASCO Corporation [Nihon Bunko], высокоэффективная жидкостная хроматография). Результаты показаны в таблице 3.
гидрохлорида L-лизина (г/л)
Пример 3
Создание бактерии, продуцирующей L-лизин (3)
Штамм AS1 Methylophilus methylotrophus (NCIMB10515) инокулировали в среду 121М1 и культивировали при 37°С в течение 15 часов. Полученные бактериальные клетки обрабатывали NTG стандартным способом (концентрация NTG: 100 мг/л, 37°С, 5 минут), и наносили на агаризованную среду 121М1, содержащую 7 г/л S-(2-аминоэтил)цистеина (AEC) и 3 г/л L-треонина. Клетки культивировали при 37°С от 2 до 8 дней и собирали образованные колонии, получая устойчивые к AEC штаммы.
Указанные выше устойчивые к AEC штаммы инокулировали в среду для продуцирования 121 и культивировали при 37°С в течение 38 часов в аэробных условиях. После окончания культивирования клетки и карбонат кальция удаляли из среды центрифугированием и в надосадке культуры измеряли концентрацию L-лизина в аминокислотном анализаторе (JASCO Corporation [Nihon Bunko], высокоэффективная жидкостная хроматография). Отобрали штамм, проявляющий повышенную способность продуцировать L-лизин по сравнению с родительским штаммом, и назвали штаммом AR-166 Methylophilus methylotrophus. Количества L-лизина, продуцируемые родительским штаммом (штамм AS1) и штаммом AR-166, показаны в таблице 4.
Штамм AR-166 Methylophilus methylotrophus получил собственный номер AJ13608, и он помещен на хранение в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (почтовый индекс 305-8566, 1-3 Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) 10 июня 1999 г. и получил инвентарный номер FERM P-17416, и перенесен для международного депонирования по условиям Будапештского соглашения 31 марта 2000 г. и получил инвентарный номер FERM BP-7112.
Пример 4
Создание бактерии, продуцирующей L-треонин.
(1) Введение плазмиды с треониновым опероном в бактерию Methylophilus.
Плазмиду pVIC40 (международная заявка на патент WO90/04636, опубликованная заявка на патент Японии (Kohyo) No. 3-501682/1992), содержащую треониновый оперон, полученный из E.coli, вводили в штамм AS1 Methylophilus methylotrophus (NCIMB10515) посредством электропорации (Canadian Journal of Microbiology, 43, 197 (1997)), чтобы получить штамм AS1/pVIC40. В качестве контроля получили pRS (опубликованная заявка на патент Японии (Kohyo) No. 3-501682/1991), имеющую только векторный район, посредством делетирования из плазмиды pVIC40 района ДНК, кодирующего треониновый оперон, и эту плазмиду вводили в штамм AS1 таким же способом, который использовали для pVIC40, получая штамм AS1/pRS.
(2) Продуцирование L-треонина бактерией Methylophilus, содержащей треониновый оперон, полученный из E.coli.
Каждый из штаммов AS1/pVIC40 и AS1/pRS инокулировали в среду для продуцирования 121, содержащую 20 мг/л стрептомицина, 1 г/л L-валина и 1 г/л L-лейцина, и культивировали при 37°С в течение 50 часов при встряхивании. После окончания культивирования клетки и карбонат кальция удаляли центрифугированием и в надосадке культуры измеряли концентрацию L-треонина в аминокислотном анализаторе (JASCO Corporation [Nihon Bunko], высокоэффективная жидкостная хроматография). Результаты показаны в таблице 5.
Пример 5
Создание бактерии, продуцирующей аминокислоты с разветвленной цепью.
Штамм AS1 Methylophilus methylotrophus (NCIMB10515) инокулировали в среду 121М1 и культивировали при 37°С в течение 15 часов. Полученные бактериальные клетки обрабатывали NTG стандартным способом (концентрация NTG: 100 мг/л, 37°С, 5 минут) и наносили на агаризованную среду 121М1, содержащую 0,5% казаминовой кислоты (DIFCO). Клетки культивировали при 37°С от 2 до 8 дней и давали возможность сформировать колонии. Сформированные колонии собирали и инокулировали в агаризованную среду 121М1 и агаризованную среду 121М1, содержащую 0,5% казаминовой кислоты. Отбирали штаммы, проявляющие более хороший рост на последней среде, по сравнению с первой средой, как штаммы, ауксотрофные по казаминовой кислоте. Таким образом, из 500 обработанных NTG штаммов получили 9 штаммов ауксотрофных по казаминовой кислоте с сохранением остаточного уровня экспрессии. Из указанных ауксотрофных по казаминовой кислоте штаммов получили один штамм, который накапливал в среде больше L-валина, L-лейцина и L-изолейцина по сравнению с его родительским штаммом. Указанный штамм был назван штаммом С138 Methylophilus methylotrophus.
Штамм С138 Methylophilus methylotrophus получил собственный номер AJ13609, и он помещен на хранение в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (почтовый индекс 305-8566, 1-3 Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Japan) 10 июня 1999 г. и получил инвентарный номер FERM P-17417, и перенесен для международного депонирования по условиям Будапештского соглашения 31 марта 2000 г. и получил инвентарный номер FERM BP-7113.
Родительский штамм (штамм AS1) и штамм С138 инокулировали в среду для продуцирования 121 и культивировали при 37°С в течение 34 часов в аэробных условиях. После окончания культивирования клетки и карбонат кальция удаляли из среды центрифугированием и в надосадке культуры измеряли концентрации L-валина, L-лейцина и L-изолейцина в аминокислотном анализаторе (JASCO Corporation [Nihon Bunko], высокоэффективная жидкостная хроматография). Результаты показаны в таблице 6.
(мг/л)
(мг/л)
(мг/л)
Пример 6
Получение библиотеки хромосомной ДНК Methylophilus methylotrophus штамма AS1.
(1) Получение хромосомной ДНК Methylophilus methylotrophus штамма AS1.
Одну платиновую петлю Methylophilus methylotrophus штамма AS1 (NCIMB10515) высевали в 5 мл среды 121М1 в пробирку и культивировали при 37°С в течение ночи при встряхивании. Полученный культуральный бульон инокулировали в 50 мл среды 121М1 в колбу Сакагуши объемом 500 мл в количестве 1% и культивировали при 37°С в течение ночи, встряхивая. Затем клетки собирали центрифугированием и суспендировали в 50 мл раствора TEN (раствор, содержащий 50 мМ трис-HCl (рН 8,0), 10 мМ ЭДТА и 20 мМ NaCl (рН 8,0)). Клетки собирали центрифугированием и снова суспендировали в 5 мл раствора TEN, содержащего 5 мг/мл лизоцима и 10 мкг/мл РНКазы А. Суспензию выдерживали при 37°С в течение 30 минут и затем к ней добавляли протеиназу К и лаурилсульфат натрия до конечных концентраций 10 мкг/мл и 0,5% (мас./об.) соответственно.
Суспензию выдерживали при 70°С в течение 2 часов и затем добавляли равное количество насыщенного раствора фенола (раствор фенола, насыщенный 10 мМ трис-HCl (рН 8,0)) и перемешивали. Суспензию центрифугировали и собирали надосадок. Добавляли равное количество раствора фенола/хлороформа (фенол:хлороформ:изоамиловый спирт = 25:24:1) и перемешивали и смесь центрифугировали. Надосадок собирали и к нему добавляли равное количество раствора хлороформа (хлороформ:изоамиловый спирт = 24:1), чтобы повторить такую же процедуру экстракции. К надосадку добавляли 1/10 объема 3 М ацетата натрия (рН 4,8) и 2,5-кратный объем этанола, чтобы осадить хромосомную ДНК. Осадки собирали центрифугированием, промывали 70% этанолом, сушили при пониженном давлении и растворяли в соответствующем количестве раствора TE (10 мМ трис-HCl, 1 мМ ЭДТА (рН 8,0)).
(2) Получение библиотеки генов.
Порции хромосомной ДНК, полученной, как описано выше в (1), объемом 50 мкл (1 мкг/мкл), 20 мкл буфера Н (500 мМ трис-HCl, 100 мМ MgCl2, 10 мМ дитиотреитол, 1000 мМ NaCl (рН 7,5)) и 8 единицам фермента рестрикции Sau3AI (Takara Shuzo) давали возможность реагировать при 37°С в течение 10 минут в общем объеме 200 мкл, и затем для того, чтобы остановить реакцию, добавляли 200 мкл раствора фенола/хлороформа и перемешивали. Реакционную смесь центрифугировали и собирали верхний слой и разделяли на 0,8% агарозном геле. ДНК, соответствующую от 2 до 5 тысячам пар нуклеотидов (далее сокращенно обозначаемым «т.п.н.»), собирали, используя систему быстрой экстракции из геля ConcertTM (набор для сбора ДНК, GIBCO BRL Co.). Таким образом, получили 50 мкл раствора ДНК фракционированного размера.
С другой стороны 2,5 мкг плазмиды pUC118 (Takara Shuzo), 2 мкл буфера К (200 мМ трис-HCl, 100 мМ MgCl2, 10 мМ дитиотреитол, 1000 мМ KCl (рН 8,5)) и 10 единицам фермента рестрикции BamHI (Takara Shuzo) давали возможность реагировать при 37°С в течение 2 часов в общем объеме 20 мкл, затем добавляли 20 единиц щелочной фосфатазы тонкого кишечника теленка (Takara Shuzo) и перемешивали, смеси давали возможность реагировать еще 30 минут. Реакционную смесь смешивали с равным количеством раствора фенола/хлороформа и смесь центрифугировали. Собирали надосадок и к нему добавляли равное количество раствора хлороформа, чтобы повторить сходную процедуру экстракции. К надосадку добавляли 1/10 объема 3 М ацетата натрия (рН 4,8) и 2,5-кратный объем этанола, чтобы осадить ДНК. ДНК собирали центрифугированием, промывали 70% этанолом, сушили при пониженном давлении и растворяли в соответствующем количестве раствора TE.
Продукт переваривания хромосомной ДНК Sau3AI, полученный, как описано выше, и продукт переваривания pUC118 рестриктазой BamHI лигировали, используя набор для лигирования, вариант 2 (Takara Shuzo). К реакционной смеси добавляли 1/10 объема 3 М ацетата натрия (рН 4,8) и 2,5-кратный объем этанола, чтобы осадить ДНК. ДНК собирали центрифугированием, промывали 70% этанолом, сушили при пониженном давлении и растворяли в растворе TE (раствор А для лигазы).
Таким же образом, как в описанной выше процедуре, лигировали фрагменты, полученные при частичном переваривании хромосомной ДНК ферментом рестрикции AluI (Takara Shuzo), и продукт переваривания SmaI плазмиды pSTV29 (Takara Shuzo) (раствор В для лигазы).
Одну платиновую петлю E.coli JM109 инокулировали в 5 мл среды L в пробирку и культивировали при 37°С в течение ночи, встряхивая. Полученный культуральный бульон инокулировали в 50 мл среды L в колбе Сакагуши объемом 500 мл в количестве, равном 1%, культивировали при 37°С до того, как OD660 культуры достигала значения от 0,5 до 0,6, и охлаждали на льду в течение 15 минут. Затем клетки собирали центрифугированием при 4°С. Клетки суспендировали в 50 мл ледяной воды и центрифугировали, чтобы промыть клетки. Эту операцию повторяли снова один раз и клетки суспендировали в 50 мл ледяного 10% раствора глицерина и центрифугировали, чтобы промыть клетки. Клетки суспендировали в 10% растворе глицерина такого же объема, как объем клеток, и делили на аликвоты по 50 мкл. К клеткам в объеме 50 мкл добавляли 1 мкл раствора А для лигазы или раствора В для лигазы, полученных, как описано выше. Затем смесь помещали в специальную кювету (с зазором 0,1 см, предварительно охлажденную на льду) для прибора для электропорации BioRad.
Параметры настройки прибора были равны 1,8 кВ и 25 мкФ, и параметры настройки системы контроля импульсов - 200 Ом. Кювету вставляли в прибор и прикладывали к ней импульсы. Сразу после воздействия импульсами в нее добавляли 1 мл ледяной среды SOC и смесь переносили в стерильную пробирку и культивировали при 37°С в течение 1 часа, встряхивая. Бульон с каждой культурой клеток наносили на агаризованную среду L, содержащую антибиотик (100 мкг/мл ампициллина в случае использования раствора А для лигазы или 20 мкг/мл хлорамфеникола в случае использования раствора В для лигазы), и инкубировали при 37°С в течение ночи. Колонии, появившиеся на каждой агаризованной среде, соскребали, инокулировали в 50 мл среды L, содержащей соответствующий антибиотик, в колбе Сакагуши объемом 500 мл и культивировали при 37°С в течение 2 часов, встряхивая. Плазмидную ДНК экстрагировали из каждого бульона с культурой щелочным способом с SDS, чтобы получить раствор А библиотеки генов и раствор В библиотеки генов соответственно.
Пример 7
Клонирование гена биосинтеза лизина Methylophilus methylotrophus штамма AS1.
(1) Клонирование гена, кодирующего аспартокиназу (АК).
Клетки E.coli GT3, дефицитные по трем генам, кодирующим АК (thrA, metLM и lysC) трансформировали раствором В библиотеки генов посредством такой же процедуры электропорации, как указано выше. К раствору для трансформации добавляли среду SOC, содержащую 20 мкг/мл диаминопимелиновой кислоты и культивировали при 37°С, встряхивая. Затем культуральный бульон наносили на среду L, содержащую 20 мкг/мл диаминопимелиновой кислоты и 20 мкг/мл хлорамфеникола, чтобы получить появляющиеся колонии. С полученной чашки как с основной матрицы получали реплику на агаризованной среде М9, содержащей 20 мкг/мл хлорамфеникола, и реплику инкубировали при 37°С от 2 до 3 дней. Хозяин не мог расти на минимальной среде М9, которая не содержала диаминопимелиновой кислоты, так как он не обладал активностью АК. Напротив, ожидалось, что трансформированный штамм, который содержал ген, кодирующий АК, полученный из Methylophilus methylotrophus, мог расти в минимальной среде М9 благодаря функционированию гена.
Два трансформанта примерно из 3000 трансформантов формировали колонии на среде М9. Из этих колоний, появившихся на среде М9, экстрагировали и анализировали плазмиды. В результате было подтверждено наличие в плазмиде встроенного фрагмента. Плазмиды были названы pMMASK-1 и pMMASK-2 соответственно. Используя указанные плазмиды, снова трансформировали E.coli GT3. Полученные трансформанты могли расти на минимальной среде М9. Затем трансформант, который содержал каждую из указанных плазмид, культивировали в течение ночи в среде L, содержащей 20 мкг/мл хлорамфеникола, и клетки собирали центрифугированием бульона с культурой. Бесклеточные экстракты получали, обрабатывая клетки ультразвуком, и измеряли активность АК методом Miyajima et al. (Journal of Biochemistry (Tokyo), vol. 63, 139-148 (1968)) (фиг.3: pMMASK-1, pMMASK-2). Кроме того, штамм GT3, несущий вектор pSTV29, сходным образом культивировали в среде L, содержащей 20 мкг/мл диаминопимелиновой кислоты и 20 мкг/мл хлорамфеникола, и измеряли активность АК (фиг.3: Вектор). В результате наблюдали увеличение активности АК в двух клонах, содержащих встроенные фрагменты, по сравнению с трансформантом, несущим только вектор. Поэтому было подтверждено, что ген, который удалось клонировать в pSTV29, представлял собой ген АК, полученный из Methylophilus methylotrophus. Указанный ген был обозначен ask.
Последовательность нуклеотидов ДНК гена ask определяли методом с использованием дидезоксинуклеотидов. Было обнаружено, что pMMASK-1 и pMMASK-2 содержали общий фрагмент. Нуклеотидная последовательность фрагмента ДНК, содержащего ген ask, полученный из Methylophilus methylotrophus, показана в SEQ ID NO: 5. Аминокислотная последовательность, которую может кодировать нуклеотидная последовательность, показана в SEQ ID NO: 5 и 6.
(2) Клонирование гена, кодирующего дегидрогеназу полуальдегида аспарагиновой кислоты (ASD).
Клетки E.coli Hfr3000 U482 (штамм CGSC 5081), дефицитные по гену asd, трансформировали путем электропорации, используя раствор В библиотеки генов, таким же образом, как описано выше. К раствору для трансформации добавляли среду SOC, содержащую 20 мкг/мл диаминопимелиновой кислоты, и смесь культивировали при 37°С, встряхивая. Клетки собирали центрифугированием. Клетки промывали, суспендируя их в среде L и центрифугируя суспензию. Такую же операцию промывки снова повторяли один раз, и клетки суспендировали в среде L. Затем суспензию распределяли на агаризованной среде L, содержащей 20 мкг/мл хлорамфеникола, инкубировали в течение ночи при 37°С. Хозяин показывал чрезвычайно низкий рост на среде L, не содержащей диаминопимелиновой кислоты, так как он был дефицитен по гену asd. Напротив, ожидалось, что нормальный рост наблюдался бы для трансформированного штамма, который содержал ген, кодирующий ASD, полученный из Methylophilus methylotrophus, даже в среде L благодаря функционированию гена. Кроме того, клетки хозяина E.coli не могли расти в минимальной среде М9, но ожидалось, что трансформированный штамм, который содержал ген, кодирующий ASD, полученный из Methylophilus methylotrophus, будет способен расти на минимальной среде М9 благодаря функционированию гена. Поэтому колонии трансформантов, которые нормально выросли на среде L, собрали, высевали штрихом и культивировали на агаризованной среде М9. В результате наблюдали рост. Таким образом, подтвердили, что ген, кодирующий ASD, как и ожидалось, функционировал у данных трансформантов.
Из трех трансформированных штаммов, появившихся на среде М9, экстрагировали плазмиды и подтвердили наличие в плазмидах встроенного фрагмента. Плазмиды были названы pMMASD-1, pMMASD-2 и pMMASD-3 соответственно. Когда E.coli Hfr3000 U482 снова трансформировали, используя указанные плазмиды, каждый трансформант вырастал на минимальной среде М9. Затем каждый трансформант культивировали в течение ночи в среде L, содержащей 20 мкг/мл хлорамфеникола, и клетки собирали центрифугированием бульона с культурой. Клетки обрабатывали ультразвуком, получая неочищенный раствор ферментов, и измеряли активность ASD методом Boy et al. (Journal of Bacteriology, vol. 112 (1), 84-92 (1972)) (фиг.4: pMMASD-1, pMMASD-2, pMMASD-3). Кроме того, в качестве контрольного эксперимента клетки хозяина, несущие вектор, сходным образом культивировали в среде L, содержащей 20 мкг/мл диаминопимелиновой кислоты и 20 мкг/мл хлорамфеникола, и измеряли активность ASD (фиг.4: Вектор). В результате ферментативная активность не выявлялась у трансформанта, несущего только вектор, в то время как активность ASD можно было определить в трех клонах, содержащих встроенный фрагмент. Поэтому было подтверждено, что полученный ген представлял собой ген, кодирующий ASD, полученный из Methylophilus methylotrophus (обозначен asd).
Последовательность нуклеотидов ДНК гена asd определяли методом с использованием дидезоксинуклеотидов. Было обнаружено, что все три полученных клона содержали общий фрагмент. Нуклеотидная последовательность фрагмента ДНК, содержащего ген asd, полученный из Methylophilus methylotrophus, показана в SEQ ID NO: 7. Аминокислотная последовательность, которую может кодировать нуклеотидная последовательность, показана в SEQ ID NO: 7 и 8.
(3) Клонирование гена, кодирующего дигидродипиколинатсинтазу (DDPS).
Клетки E.coli AT997 (штамм CGSC 4547), дефицитные по гену dapA, трансформировали посредством такой же процедуры электропорации, используя раствор А библиотеки генов. К раствору для трансформации добавляли среду SOC, содержащую 20 мкг/мл диаминопимелиновой кислоты, и смесь культивировали при 37°С, встряхивая. Затем бульон с культурой распределяли на среде L, содержащей 20 мкг/мл диаминопимелиновой кислоты и 100 мкг/мл ампициллина, чтобы получить появляющиеся колонии. С полученной чашки как с основной матрицы получали реплику на минимальной агаризованной среде М9, содержащей 100 мкг/мл ампициллина, и реплику инкубировали при 37°С от 2 до 3 дней. Хозяин не мог расти на минимальной среде М9, которая не содержала диаминопимелиновой кислоты, так как был дефицитен по гену dapA. Напротив, ожидалось, что трансформированный штамм, который содержал ген, кодирующий DDPS, полученный из Methylophilus methylotrophus, мог расти в минимальной среде М9 благодаря функционированию гена.
Из колоний двух штаммов, появившихся на среде М9, экстрагировали и анализировали плазмиды. В результате подтвердили наличие в плазмидах встроенного фрагмента. Плазмиды были названы pMMDAPA-1 и pMMDAPA-2 соответственно. Когда E.coli AT997 снова трансформировали, используя указанные плазмиды, каждый трансформант вырастал на минимальной среде М9. Затем каждый трансформант, содержащий каждую плазмиду, культивировали в течение ночи в среде L, содержащей 100 мкг/мл ампициллина, и клетки собирали центрифугированием бульона с культурой. Клетки обрабатывали ультразвуком, получая клеточный экстракт, и измеряли активность DDPS методом Yugari et al. (Journal of Biological Chemistry, vol. 240, and p. 4710 (1965)) (фиг.5: pMMDAPA-1, pMMDAPA-2). Кроме того, в качестве контрольного эксперимента клетки хозяина, несущие вектор, сходным образом культивировали в среде L, содержащей 20 мкг/мл диаминопимелиновой кислоты и 100 мкг/мл ампициллина, и измеряли активность DDPS (фиг.5: Вектор). В результате ферментативная активность не выявлялась у трансформанта, несущего только вектор, в то время как активность DDPS можно было определить в каждом из трансформантов, несущих плазмиды, которые содержат встроенный фрагмент. Поэтому было подтверждено, что полученный ген представлял собой ген, кодирующий DDPS, полученный из Methylophilus methylotrophus (обозначен dapA).
Последовательность нуклеотидов ДНК гена dapA определяли методом с использованием дидезоксинуклеотидов. Было обнаружено, что два встроенных фрагмента содержали общий фрагмент. Нуклеотидная последовательность фрагмента ДНК, содержащего ген dapA, полученный из Methylophilus methylotrophus, показана в SEQ ID NO: 9. Аминокислотная последовательность, которую может кодировать нуклеотидная последовательность, показана в SEQ ID NO: 9 и 10.
(4) Клонирование гена, кодирующего дигидродипиколинатредуктазу (DDPR).
Клетки E.coli AT999 (штамм CGSC 4549), дефицитные по гену dapB, трансформировали посредством такой же процедуры электропорации, как описано выше, используя раствор А библиотеки генов. К раствору для трансформации добавляли среду SOC, содержащую 20 мкг/мл диаминопимелиновой кислоты, и смесь культивировали при 37°С, встряхивая. Затем клетки собирали центрифугированием. Клетки промывали, суспендируя их в среде L и центрифугируя суспензию. Такую же операцию промывки снова повторяли один раз, и клетки суспендировали в среде L. Затем суспензию распределяли на агаризованной среде L, содержащей 100 мкг/мл ампициллина, и инкубировали в течение ночи при 37°С. Хозяин показывал чрезвычайно низкий рост на среде L, не содержащей диаминопимелиновой кислоты, так как он был дефицитен по гену dapB. Напротив, ожидалось, что нормальный рост может наблюдаться для трансформированного штамма, который содержал ген, кодирующий DDPR, полученный из Methylophilus methylotrophus, даже в среде L, благодаря функционированию гена. Кроме того, клетки хозяина E.coli не могли расти в минимальной среде М9, но ожидалось, что трансформированный штамм, который содержал ген, кодирующий DDPR, полученный из Methylophilus methylotrophus, может расти на минимальной среде М9 благодаря функционированию гена.
Поэтому колонию трансформанта, который нормально вырос на среде L, соскребали и культивировали на агаризованной среде М9. В этом случае также наблюдали рост на среде М9. Таким образом, подтвердили, что ген, кодирующий DDPR, функционировал в трансформированном штамме. Из колонии, появившейся на среде М9, экстрагировали плазмиду и подтвердили наличие в плазмиде встроенного фрагмента. Когда E.coli АТ999 снова трансформировали, используя плазмиду (pMMDAPB), трансформант вырастал на минимальной среде М9. Затем трансформант, содержащий плазмиду, культивировали в течение ночи в среде L и клетки собирали центрифугированием бульона с культурой. Клетки обрабатывали ультразвуком, получая клеточный экстракт, и измеряли активность DDPR методом Tamir et al. (Journal of Biological Chemistry, vol. 249, p. 3034 (1974)) (фиг.6: pMMDAPB). Кроме того, в качестве контрольного эксперимента клетки хозяина, несущие вектор, сходным образом культивировали в среде L, содержащей 20 мкг/мл диаминопимелиновой кислоты и 100 мкг/мл ампициллина, и измеряли активность DDPR (фиг.6: Вектор). В результате ферментативная активность не выявлялась у трансформанта, несущего только вектор, в то время как активность DDPR можно было определить у трансформанта, несущего pMMDAPB. Поэтому было подтверждено, что полученный ген представлял собой ген, кодирующий DDPR, полученный из Methylophilus methylotrophus (обозначен dapB).
Последовательность нуклеотидов ДНК гена dapB определяли методом с использованием дидезоксинуклеотидов. Нуклеотидная последовательность фрагмента ДНК, содержащего ген dapB, полученный из Methylophilus methylotrophus, показана в SEQ ID NO: 11. Аминокислотная последовательность, которую может кодировать нуклеотидная последовательность, показана в SEQ ID NO: 11 и 12.
(5) Клонирование гена, кодирующего диаминопимелатдекарбоксилазу (DPDC).
Клетки E.coli AT2453 (штамм CGSC 4505), дефицитные по гену lysA, трансформировали посредством такой же процедуры электропорации, как описано выше, используя раствор А библиотеки генов. К раствору для трансформации добавляли среду SOC, и смесь культивировали при 37°С, встряхивая. Клетки собирали центрифугированием. Клетки промывали, суспендируя их в 5 мл стерильной воды и центрифугируя суспензию. Такую же операцию промывки снова повторяли один раз и клетки суспендировали в 500 мкл стерильной воды. Затем суспензию распределяли на агаризованной минимальной среде М9, содержащей 20 мкг/мл хлорамфеникола, и инкубировали при 37°С от 2 до 3 дней. Хозяин не мог расти на минимальной среде М9, не содержащей лизина, так как он был дефицитен по гену lysA. Напротив, ожидалось, что трансформированный штамм, который содержал ген, кодирующий DPDC, полученный из Methylophilus methylotrophus, может расти в минимальной среде М9 благодаря функционированию гена.
Поэтому из трех трансформированных штаммов, появившихся на среде М9, экстрагировали и анализировали плазмиды. В результате подтвердили наличие в плазмидах встроенного фрагмента. Плазмиды были названы pMMLYSA-1, pMMLYSA-2 и pMMLYSA-3 соответственно. Когда E.coli АТ2453 снова трансформировали, используя каждую их указанных плазмид, каждый трансформант вырастал на минимальной среде М9. Затем каждый трансформант, содержащий каждую из плазмид, культивировали в течение ночи в среде L, содержащей 20 мкг/мл хлорамфеникола, и клетки собирали центрифугированием бульона с культурой. Клетки обрабатывали ультразвуком, получая клеточный экстракт, и измеряли активность DPDC методом Cremer et al. (Journal of General Microbiology, vol. 134, 3221-3229 (1988)) (фиг.7: pMMLYSA-1, pMMLYSA-2, pMMLYSA-3). Кроме того, в качестве контрольного эксперимента клетки хозяина, несущие вектор, сходным образом культивировали в среде L, содержащей 20 мкг/мл хлорамфеникола, и измеряли активность DPDC (фиг.7: Вектор). В результате ферментативная активность не выявлялась у трансформанта, несущего только вектор, в то время как активность DPDC можно было определить в трех клонах, имеющих встроенный фрагмент. Поэтому было подтверждено, что полученный ген представлял собой ген, кодирующий DPDC, полученный из Methylophilus methylotrophus (обозначен lysA).
Последовательность нуклеотидов ДНК гена lysA определяли методом с использованием дидезоксинуклеотидов. Обнаружено, что все три встроенных фрагмента содержали общий фрагмент ДНК. Нуклеотидная последовательность фрагмента ДНК, содержащего ген lysA, полученный из Methylophilus methylotrophus, показана в SEQ ID NO: 13. Аминокислотная последовательность, которую может кодировать нуклеотидная последовательность, показана в SEQ ID NO: 13 и 14.
Промышленное применение
Согласно данному изобретению предоставлены бактерия Methylophilus, обладающая способностью продуцировать L-аминокислоту, способ получения L-аминокислоты с использованием бактерии Methylophilus, и бактериальные клетки Methylophilus с повышенным содержанием L-аминокислоты. Способом согласно изобретению предоставляется возможность получать L-аминокислоту, используя метанол в качестве сырья. Кроме того, данное изобретение предоставляет новые гены ферментов биосинтеза L-лизина, полученные из бактерий Methylophilus.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2250266C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2368659C2 |
| БАКТЕРИЯ, ПРОДУЦИРУЮЩАЯ L-АМИНОКИСЛОТУ, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2000 |
|
RU2351653C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА (ВАРИАНТЫ) И РЕКОМБИНАНТНАЯ ДНК, ИСПОЛЬЗУЕМАЯ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ (ВАРИАНТЫ) | 1996 |
|
RU2197528C2 |
| СПОСОБ ПРОДУКЦИИ АРОМАТИЧЕСКИХ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Methylophilus | 2005 |
|
RU2333950C2 |
| ФЕРМЕНТАЦИОННЫЙ СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1994 |
|
RU2204605C2 |
| ПРЕФЕНАТДЕГИДРАТАЗА-ХОРИЗМАТМУТАЗА И ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ ПРЕФЕНАТДЕГИДРАТАЗУ-ХОРИЗМАТМУТАЗУ, ИЗ БАКТЕРИИ Methylophilus methylotrophus | 2003 |
|
RU2250261C1 |
| 3-ДЕЗОКСИ-D-АРАБИНОГЕПТУЛОЗОНАТ-7-ФОСФАТСИНТАЗА И ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ 3-ДЕЗОКСИ-D-АРАБИНОГЕПТУЛОЗОНАТ-7-ФОСФАТСИНТАЗУ ИЗ METHYLOPHILUS METHYLOTROPHUS | 2000 |
|
RU2229514C2 |
| РЕКОМБИНАНТНАЯ ДНК И СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 2009 |
|
RU2451748C1 |































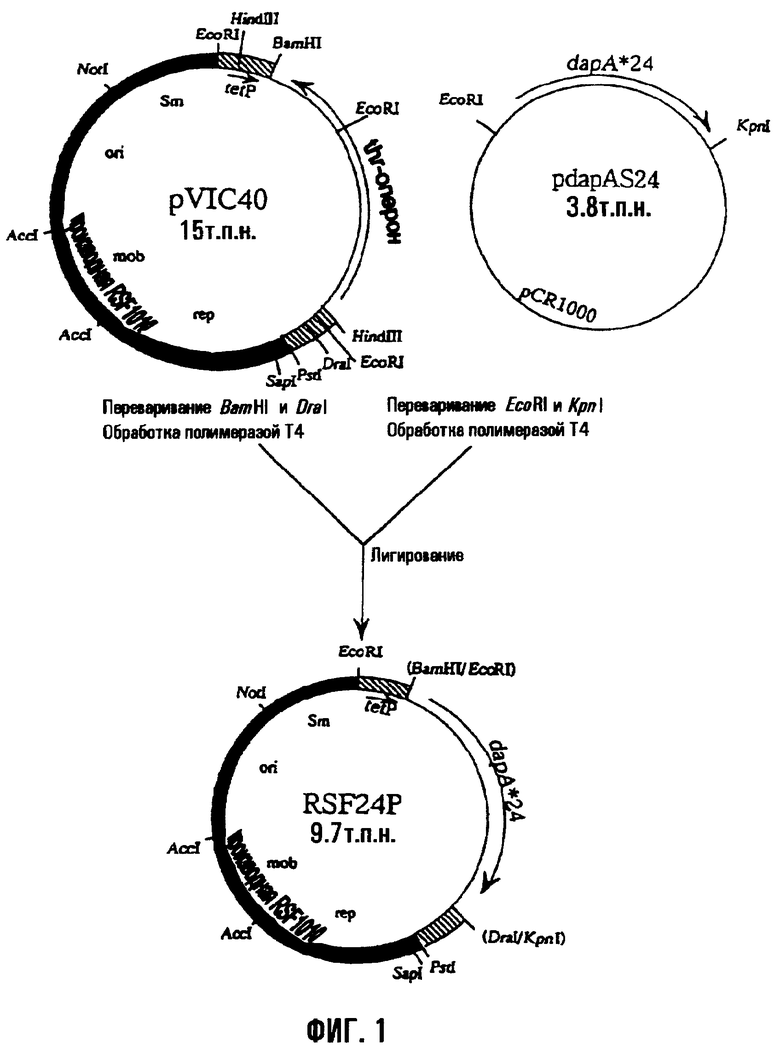
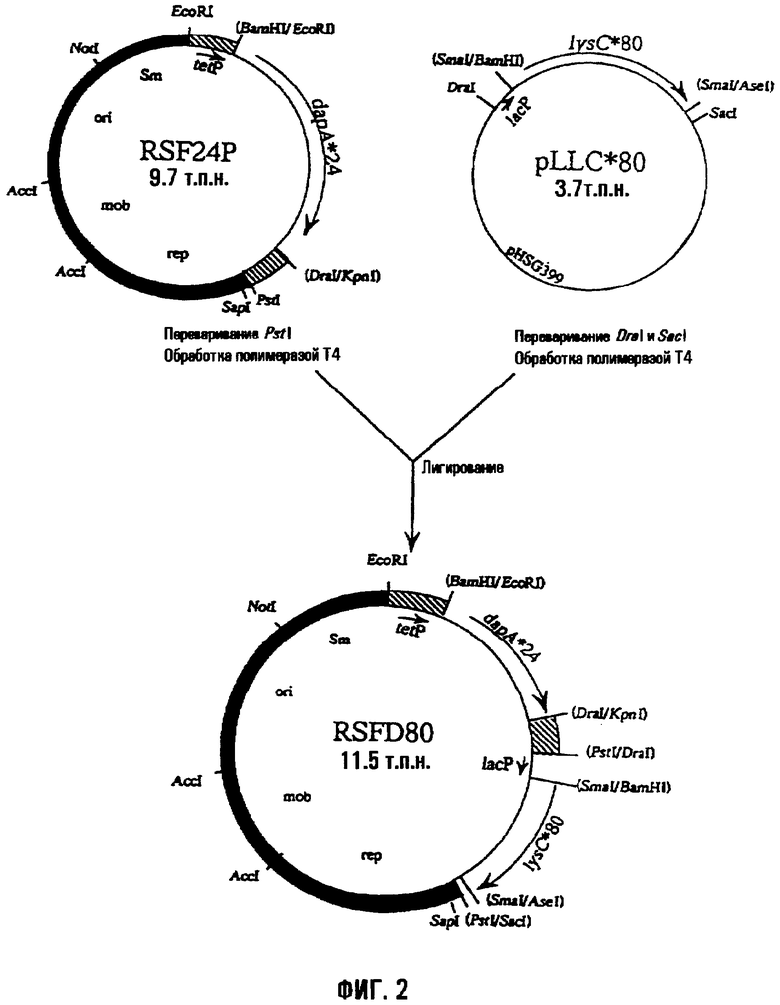
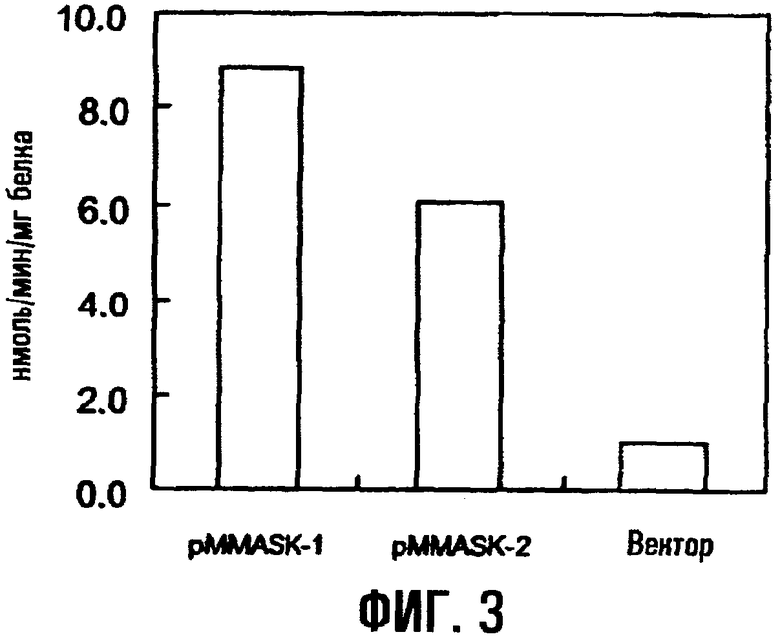
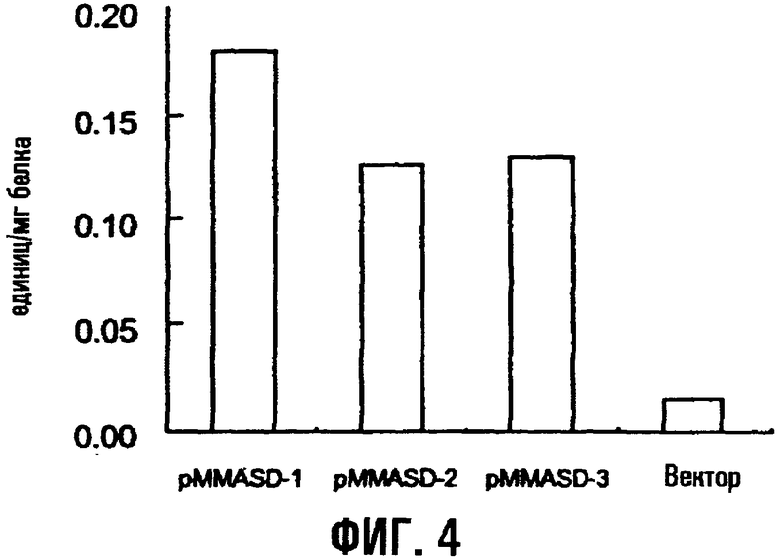

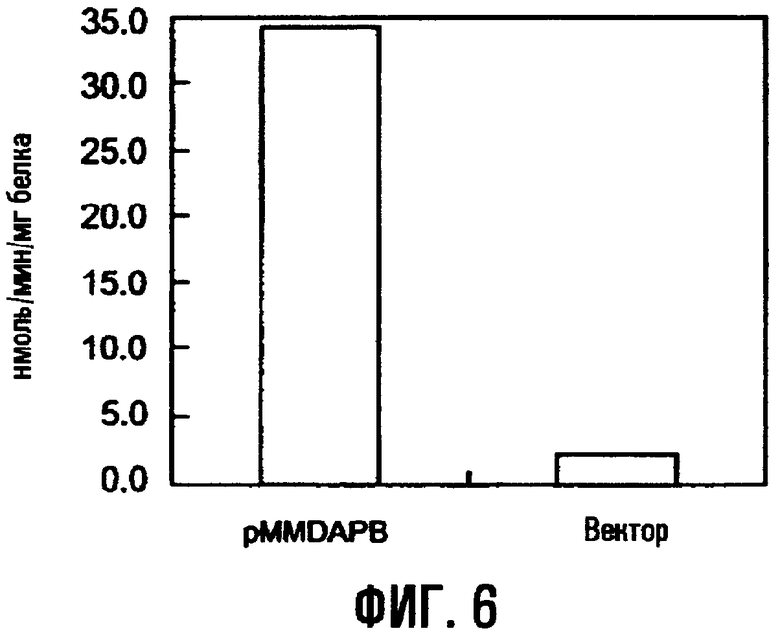
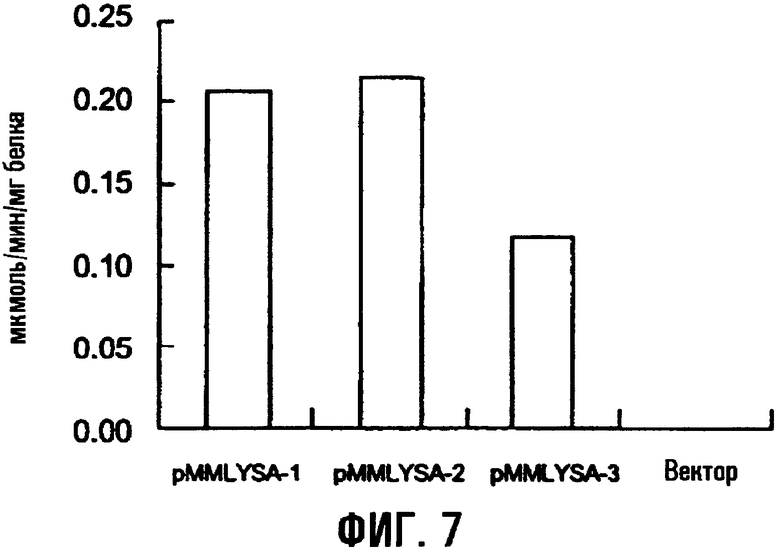
Изобретение относится к биотехнологии и представляет собой способ получения L-лизина. L-лизин получают путем культивирования бактерии Methylophilus, которая может расти, используя метанол в качестве основного источника углерода, и обладает способностью продуцировать L-лизин, и у которой активность дигидродипиколинатсинтазы повышена путем трансформации посредством введения в клетки ДНК, кодирующей дигидродипиколинатсинтазу, в среде, содержащей в качестве основного источника углерода метанол, чтобы продуцировать и накопить L-лизин в культуре, и сбором L-лизина из культуры. Изобретение позволяет получать L-лизин с высокой степенью эффективности. 4 н. и 2 з.п. ф-лы, 7 ил., 6 табл.
(Е) белок, который имеет аминокислотную последовательность SEQ ID NO: 10 и обладает активностью дигидродипиколинат/синтазы, или
(F) белок, который имеет аминокислотные последовательности SEQ ID NO: 10, включая замену, делецию, инсерцию, присоединение или инверсию одной или нескольких аминокислот, и обладает активностью дигидродипиколинатсинтазы.
(e) ДНК, которая имеет нуклеотидную последовательность, включающую в себя нуклеотидную последовательность от нуклеотида номер 1268 до нуклеотида номер 2155 SEQ ID NO: 9; или
(f) ДНК, которая гибридизуется с пробой, имеющей нуклеотидную последовательность от нуклеотида номер 1268 до нуклеотида номер 2155 SEQ ID NO: 9 или ее часть, в жестких условиях при температуре 60°С и солевых концентрациях, соответствующих 1×SSC и 0,1% SDS, и кодирует белок, обладающий активностью дигидродипиколинатсинтазы.
Приоритет по пунктам:
| WO 9641871, 27.12.1996 | |||
| JP 10215883, 18.08.1998 | |||
| Способ получения глутаматдегидрогеназы | 1982 |
|
SU1017730A1 |

